Стабілізовані поліпептиди інсуліноподібного фактора росту

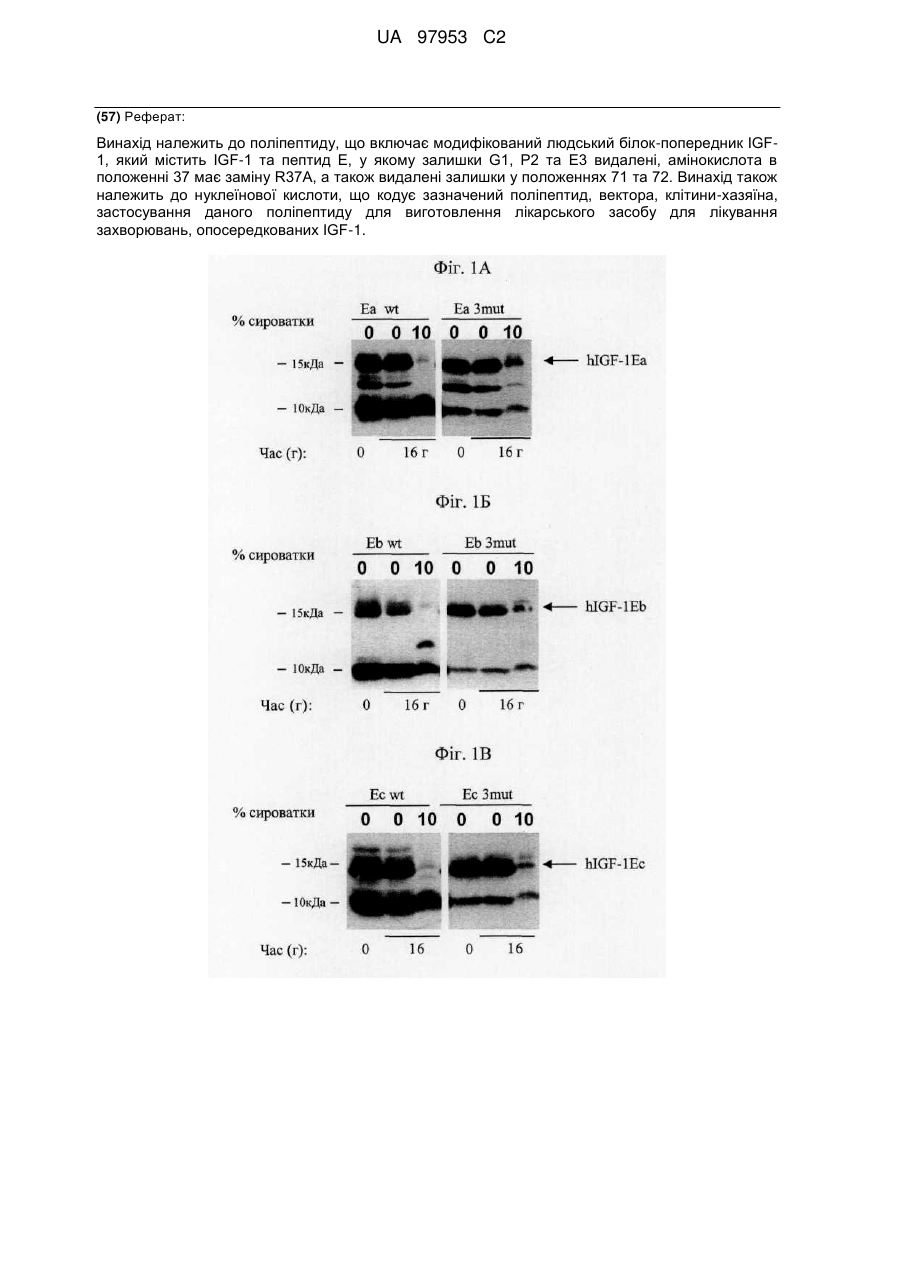






Формула / Реферат
1. Поліпептид, що включає модифікований людський білок-попередник IGF-1, який містить IGF-1 та пептид Е, у якому залишки G1, P2 та E3 видалені, та амінокислота в положенні 37 має заміну R37А, а також видалені амінокислоти у положеннях 71 і 72.
2. Поліпептид за п. 1, у якому білок-попередник містить Ea-пептид.
3. Поліпептид за п. 1, у якому білок-попередник містить Eb-пептид.
4. Поліпептид за п. 3, у якому останні сім C-кінцевих амінокислот Eb видалені.
5. Поліпептид за п. 1, у якому білок-попередник містить Ec-пептид.
6. Поліпептид за будь-яким з пп. 1-5, який додатково містить NXS/T консенсусну послідовність N-зв'язаного глікозилювання.
7. Поліпептид за п. 3, який додатково містить амінокислоти 93-102 з Ea, вставлені між амінокислотами N95 та T96 з Eb.
8. Поліпептид за будь-яким з пп. 1-7, який додатково містить олігосахарид, ковалентно зв'язаний з бічним ланцюгом амінокислоти білка-попередника.
9. Поліпептид за п. 8, у якому олігосахарид ковалентно зв'язаний з бічним ланцюгом аргініну білка-попередника.
10. Поліпептид за будь-яким з пп. 1-9, у якому залишок білка-попередника замінений на неприродну амінокислоту.
11. Поліпептид за п. 10, у якому неприродна амінокислота містить ацетиленову або азидогрупу.
12. Поліпептид, що містить людський білок-попередник IGF-1 та білок Еа, який представлений в SEQ ID NO: 8.
13. Поліпептид, що містить людський білок-попередник IGF-1 та білок Еа, який представлений в SEQ ID NO: 53.
14. Поліпептид, що включає модифікований людський білок-попередник IGF-1, який містить IGF-1 та пептид Еа, у якому видалені амінокислота Е3 та амінокислоти R71 та S72, а амінокислота R37 мутована на аланін.
15. Поліпептид за будь-яким з пп. 1-14, який додатково включає залишок полі(етиленгліколю), ковалентно приєднаний до бічного ланцюга білка-попередника.
16. Поліпептид за п. 15, у якому залишок полі(етиленгліколю) ковалентно приєднаний до бічного ланцюга білка-попередника за п. 14.
17. Поліпептид за будь-яким з пп. 1-16 для застосування у лікуванні захворювання кістково-м'язової системи, діабету, загибелі нейронів, хронічного обструктивного захворювання легенів, опікового ушкодження або анемії.
18. Спосіб лікування захворювання кістково-м'язової системи, діабету, загибелі нейронів, хронічного обструктивного захворювання легенів, опікового ушкодження або анемії, у якому суб'єкту вводять терапевтично ефективну кількість поліпептиду за будь-яким з пп. 1-16.
19. Застосування поліпептиду за будь-яким з пп. 1-16 для виготовлення лікарського засобу для лікування захворювання кістково-м'язової системи, діабету, загибелі нейронів, хронічного обструктивного захворювання легенів, опікового ушкодження або анемії.
20. Нуклеїнова кислота, що кодує поліпептид за будь-яким з пп. 1-16.
21. Вектор, що містить нуклеїнову кислоту за п. 20.
22. Клітина, трансфекована вектором за п. 21.
Текст
Реферат: Винахід належить до поліпептиду, що включає модифікований людський білок-попередник IGF1, який містить IGF-1 та пептид Е, у якому залишки G1, P2 та E3 видалені, амінокислота в положенні 37 має заміну R37A, а також видалені залишки у положеннях 71 та 72. Винахід також належить до нуклеїнової кислоти, що кодує зазначений поліпептид, вектора, клітини-хазяїна, застосування даного поліпептиду для виготовлення лікарського засобу для лікування захворювань, опосередкованих IGF-1. UA 97953 C2 5 10 15 20 25 30 35 40 45 50 55 60 Інсуліноподібні фактори росту (IGF) є частиною комплексної системи, яку клітини використовують для зв'язування з їхнім фізіологічним оточенням. Ця комплексна система (яку часто називають віссю інсуліноподібних факторів росту) складається із двох розташованих на клітинній поверхні рецепторів (IGF-1R та IGF-2R), двох лігандів (IGF-1 та IGF-2), сімейства, що складається із шести маючих високу афінність до зв'язування IGF білків (IGFBP 1-6), та асоційованих з IGFBP ферментів, що розщеплюють (протеаз). Ця система важлива не тільки для регуляції нормальної фізіологічної функції, вона бере участь також у деяких патологічних станах (Glass, Nat Cell Biol 5, 2003, стор. 87-90). Встановлено, що вісь IGF відіграє роль у посиленні клітинної проліферації та інгібуванні клітинної загибелі (апоптоз). IGF-1 секретується, головним чином, печінкою в результаті стимуляції людським гормоном росту (hGH). IGF-1 робить вплив майже на всі клітини в людському організмі, насамперед на клітини в м'язах, хрящах, костях, печінці, нирках, нервах, шкірі та легенях. Крім інсуліноподібної дії, IGF-1 може регулювати також ріст клітин. IGF-1 та IGF-2 регулюються сімейством генних продуктів, які відомі як IGF-єднальні білки. Ці білки сприяють модуляції дії IGF складними шляхами, які включають як інгібування дії IGF у результаті запобігання зв'язування з IGF-рецепторами, так і посилення дії IGF завдяки «допомозі» у переносі до рецепторів та подовження часу напівжиття IGF у кровотоці. Існує щонайменше шість маючих здатність до зв'язування з IGF білків (IGFBP 1-6). У своїй зрілій формі людський IGF-1 (gpetlcgaelvdalqfvcgdrgfyfnkptgygsssrrapqtgivdeccfrscdlrrlemycaplkpaksa; SEQ ID NO:1), що називають також соматомедіном, являє собою невеликий білок, що складається з 70 амінокислот, що, як встановлено, стимулює ріст широкої розмаїтості клітин у культурі. Кодування зрілого білку ініціюється трьома відомими отриманими в результаті сплайсингу варіантами мРНК. Відкрита рамка зчитування кожної мРНК кодує білок-попередник, що містить 70 амінокислот IGF-1 та конкретний Е-пептид на С-кінці, залежно від конкретної мPHK IGF-1. Ці Е-пептиди, які позначені як Еа-пептид (rsvraqrhtdmpktqkevhlknasrgsagnknyrm; SEQ ID NO:2), Ebпептид (rsvraqrhtdmpktqkyqppstnkntksqrrkgwpkthpggeqkegteaslqirgkkkeqrreigsrnaecrgkkgk; SEQ ID NO:3) та Ес-пептид (rsvraqrhtdmpktqkyqppstnkntksqrrkgstfeerk; SEQ ID NO:4), складаються з 3387 амінокислот та містять загальну послідовність N-кінці та варіабельну послідовність на Скінці. Наприклад, відкрита рамка зчитування дикого типу IGF-1-Ea кодує поліпептид, що складається з 105 амінокислот (gpetlcgaelvdalqfvcgdrgfyfnkptgygsssrrapqtgivdeccfrscdlrrlemycaplkpaksarsvraqrhtdmpktqkevhlknasr gsagnknyrm; SEQ ID NО:5). При експресії у фізіологічних умовах Е-пептиди відщеплюються від попередника за допомогою ендогенних протеаз із утворенням зрілого IGF-1, що складається з 70 амінокислот, як відомо, має біологічну активність. Встановлено, що в певних ситуаціях 1-3 Nкінцевих амінокислоти IGF-1 відщеплюються у фізіологічних умовах з утворенням активного IGF-1, що складається з 67-70 амінокислот. Експресія та процесинг гену IGF-2 характеризуються подібними особливостями за винятком наявності тільки одного Е-пептиду (rdvstpptvlpdnfprypvgkffqydtwkqstqrlrrglpallrarrghvlakeleafreakrhrplialptqdpahggappemasnrk; SEQ ID NO:6), що був ідентифікований у попереднику, що складається з 156 амінокислот, IGF-2 (ayrpsetlcggelvdtlqfvcgdrgfyfsφasrvsrrsrgiveeccfrscdlalletycatpakserdvstpptvlpdnfprypvgkffqy dtwkqstqrlrrglpallrarrghvlakeleafreakrhrplialptqdpahggappemasnrk; SEQ ID NO:7). Як IGF-1, так і IGF-2, очевидно, мають незадовільні властивості як лікарські засоби-кандидати, оскільки ці білки швидко розщеплюються ендогенними протеазами в сироватці пацієнтів. Однією з можливих стратегій є стабілізація IGF-1 з одержанням лікарського засобу шляхом утворення комплексу з одним з білків, що зв'язуються з ним. Короткий опис суті винаходу Даний винахід оснований на відкритті того факту, що білок-попередник IGF-1 або IGF-2, що містить практично відповідний йому Е-пептид, є біологічно активним та стабілізованим у присутності сироватки, що приводить до утворення поліпептиду IGF-1 або IGF-2, який можна застосовувати як фармацевтичний засіб. У композиціях, запропонованих у винаході, не відбувається (нормальне) відшіпяення Е-пептиду від IGF-1, що має місце у природних умовах, наприклад, у результаті мутації або делеції або аргініну в положенні 1, або серіну в положенні 2 Е-пептидів (що відповідає положенням 71 та 72 у попереднику IGF-1 дикого типу). В IGF-2 відщіплення не відбувається, наприклад, у результаті мутації або делеції або аргініну в положенні 1, або аспарагінової кислоти в положенні 2 Е-пептидів (що відповідає положенням 68 та 69 у попереднику IGF-2 дикого типу). Інші модифікації в білку-попереднику IGF можуть привести до анулювання або зниження зазначеного розщеплення. Крім того, додаткові модифікації амінокислотної послідовності попереднику IGF-1 можуть надавати додаткових цінних фармацевтичних властивостей. Наприклад, поліпептиди, 1 UA 97953 C2 5 10 15 20 25 30 35 40 45 50 55 запропоновані у винаході можуть мати підвищену афінність до рецептору IGF-1 або знижену здатність до зв'язування з інгібуючим IGF-1 або IGF-2 білком, що зв'язується. Для ясності та однаковості нумерація амінокислотних залишків у попередниках або зрілих білках IGF-1 або IGF-2 у контексті даного опису та у формулі винаходу основана на нумерації в послідовності білку-попереднику дикого типу без сигнального пептиду. Таким чином, винахід відноситься до поліпептиду, що містить людський білок-попередник IGF-1, у якому відщіплення Е-пептиду від IGF-1 протеазою знижене в результаті модифікації білку-попереднику. Е-пептид може являти собою Еа-, Еb- або Ес-пептид. На N-кінці попереднику амінокислоти G1, P2 або Е3 білку-попереднику можуть бути вилучені в результаті делеції або змінені в результаті мутації, наприклад, можуть являти собою R36 (наприклад, R36A) та R37 (наприклад, R37A). Білок-попередник може додатково включати консенсусну послідовність N-зв'язаного глікозилювання NXS/T, наприклад, у результаті вбудовування амінокислот 93-102, характерних для Еа-пептиду, між амінокислотами N95 та Т96 Еb-пептиду. В цілому, білок-попередник може включати олігосахарид, ковалентно пов'язаний з бічним ланцюгом амінокислоти білкупопереднику, такий як бічний ланцюг аргініну білку-попереднику. Крім того, залишки білку-попереднику можна заміняти на амінокислоти, що не зустрічаються в природних умовах (наприклад, на амінокислоти, які містять ацетиленову групу або азидогруппу). Такі амінокислоти, що не зустрічаються в природних умовах, можуть полегшувати зв'язування полі(етиленглікольної) ланки з бічним ланцюгом білку-попереднику, хоча в даній галузі добре відомі загальноприйняті стратегії пегілювання білків. Білок-попередник може включати також один або кілька додаткових Е-пептидів, зчеплених з С-кінцем білку-попереднику. Наприклад, поліпептид може включати в напрямку від N-кінця до Скінця (1) білок-попередник IGF-1, що несе перший Еb-пептид, у якому G1, Р1 та Е1 вилучені в результаті делеції, або R36, або R37, або обидві амінокислоти змінені в результаті мутації, R71 та S72 вилучені в результаті делеції та останні сім С-кінцевих амінокислот першого Eb-пептиду вилучені в результаті делеції; (2) другий Еb-пептид, при цьому, R71, S72 та останні сім Скінцевих амінокислот другого Eb-пептиду вилучені в результаті делеції; (3) третій Еb-пептид, при цьому R71, S72 та останні сім С-кінцевих амінокислот третього Eb-пептиду вилучені в результаті делеції; та (4) четвертий Еb-пептид, у якому R71 та S72 вилучені в результаті делеції. Ефективним шляхом попередження відщіплення Е-пептиду від IGF-1 є делеція або мутація R71 або S72. Аналогічно цьому, винахід відноситься до людського білку-попереднику IGF-2, у якому відщіплення Е-пептиду від IGF-2 протеазою знижують шляхом модифікації білку-попереднику. Зокрема, делеція або мутація R68 або D69 можуть являти собою ефективні шляхи анулювання протеазного розщеплення білку-попереднику GF-2. Крім того, будь-який Е-пептид IGF-1 можна поєднувати з IGF-2 та будь-який Е-пептид IGF-2 можна поєднувати з IGF-1 з одержанням зазначених у даному описі сприятливих дій. Винахід відноситься також до способу лікування захворювань кістково-м'язової системи, діабету, загибелі нейронів, який полягає в тому, що у ефективній кількості вводять поліпептид, запропонований у винаході. Винахід відноситься також до застосування поліпептиду, запропонованого у винаході, для готування лікарського засобу, що призначено для лікування захворювань кістково-м'язової системи, діабету, загибелі нейронів або анемії. Іншим варіантом здійснення винаходу є пегільований IGF-1, що не містить Е-пептид, але в який інтродукована амінокислота, що не зустрічається в природних умовах, як сайт пегілювання. Під обсяг винаходу підпадає також будь-який модифікований, пегільований IGF-1, що містить амінокислоту, що не зустрічається в природних умовах, зазначену в даному описі, без Е-пептиду. У винаході описані також застосовувані у ветеринарії способи та варіанти введення поліпептиду, запропонованого у винаході, в ефективній кількості для одержання необхідної дії. Застосування у ветеринарії включає (І) збільшення швидкості росту та/або розміру тварини, (II) підвищення ефективності перетворення корму в тканині організму, (III) збільшення виробництва молока у тварин на стадії лактації, (ІV) лікування тварини, що має симптоми, пов'язані з кахексією, травмою або іншими пов'язаними з виснаженням захворюваннями, та (V) лікування тварин, що знаходяться на стадії лактації, для поліпшення здоров'я немовляти. Всі процитовані посилання або документи включені в даний опис як посилання. 2 UA 97953 C2 5 10 15 20 25 30 35 40 45 50 55 60 Короткий опис креслень На кресленнях показано: на фіг. 1А-1В – результати Вестерн-блотингу поліпептидів, запропонованих у винаході, та попередника IGF-1 дикого типу після відсутності інкубації (0 год.) або 16-часової інкубації в присутності 10% людської сироватки або без неї при 37°С. Експресійними векторами, що кодують різні конструкції IGF-1, трансфектували клітини лінії Cos7 та одержували кондиціоноване середовище. Позначення «3mut» відноситься до попередника, що являє собою hIGF-1-Е-пептид, що має наступні три набори модифікацій: делецію G1, Р2 та Е3; мутацію, що приводить до заміни Arg 37 на Ala (R37A); та делецію R71 та S72. На фіг. 1А показані результати Вестерн-блотингу (з використанням антитіла до IGF-1) поліпептиду дикого типу та 3mut-попереднику, що містить Еа. На фіг. 1Б показані результати Вестерн-блотингу (з використанням антитіла до hIGF-1) поліпептиду дикого типу та 3mut-попереднику, що містить Еb. На фіг. 1B показані результати Вестерн-блотингу (з використанням антитіла до hIGF-1) поліпептиду дикого типу та 3mut-попереднику, що містить Ес; на фіг. 2А-2Г - лінійні графіки, що демонструють біологічну активність різних поліпептидів IGF-1 («ліганди»). Біологічну активність оцінювали шляхом стимуляції міобластів С2С12 поліпептидами, що експресуються Соs7-клітинами. Потім у стимульованих С2С12-клітинах аналізували відносні кількості загальної АКТ та фосфорильованої АКТ. Long-R3-IGF-1 являє собою наявний у продажу реагент (фірма Sigma, продукт № І-1271), що складається зі зрілої людської амінокислотної послідовності IGF-1 з мутацією E3R та з додаванням подовжуючого Nкінцевого пептиду, що складається з 13 амінокислот. На фіг. 2А показані дані про активність IGF-1-Ea3mut. На фіг. 2Б показані дані про активність IGF-1-Eb3mut. На фіг. 2В наведені дані про активність IGF-1-Eab3mut, що представляє собою несучу 3mut конструкцію, у якій амінокислоти 93-102 Еа убудовані між амінокислотами 95 та 96 Еb. На фіг. 2Г показані дані про активність IGF-1-Ec3mut; на фіг. 3А-3Г та 4А-4Г - лінійні графіки, що дозволяють визначати чи зберігають поліпептиди-попередники IGF-1, запропоновані у винаході, селективність стосовно відповідного рецептору при аналізі фосфорилювання рецептору у відповідь на зв'язування з лігандом. На фіг. 3А та 3Б представлені результати оцінки селективності до рецептору IGF-1-Еа3mut у порівнянні з рецептором IGF-1 (фіг. 3А) та інсуліновим рецептором (фіг. 3Б). На фіг. 3В та 3Г представлені результати оцінки селективності до рецептору IGF-1-Eb3mut у порівнянні з рецептором IGF-1 (фіг. 3В) та інсуліновим рецептором (фіг. 3Г). На фіг. 4А та 4Б представлені результати оцінки селективності до рецепторау IGF-1-Ec3mut у порівнянні з рецептором IGF-1 (фіг. 4А) та інсуліновим рецептором (фіг. 4Б). На фіг. 4В та 4Г представлені результати оцінки селективності до рецептору IGF-1-Eab3mut у порівнянні з рецептором IGF-1 (фіг. 4В) та інсуліновим рецептором (фіг. 4Г). «IGF1-R3» позначає описаний вище Long-R3-IGF-1. Поліпептид, позначений як «IGFlEab», являє собою конструкцію, у якій амінокислоти 93-102 Еа вбудовані між амінокислотами 95 та 96 Еb; на фіг. 5 - результати Вестерн-блотингу, що демонструють відносне фосфорилювання АКТ після стимуляції м'язових трубочок лінії С2С12 (у результаті диференціювання протягом 3-4 днів міоцитів С2С12) різними лігандами. Поняття «мультимери IGF-1Eb» використовують для позначення конструкції, зображеної схематично на фіг. 6А; на фіг. 6А и 6Б - схематичні зображення двох поліпептидів, запропонованих у винаході. На фіг. 6А показаний поліпептид-попередник IGF-1-Eb з 4 наборами модифікацій: делеція G1, P2 та Е3; мутація, що приводить до заміни R37 на А; делеція R71 та S72; та делеція останніх семи Скінцевих амінокислот. Крім того, поліпептид подовжений шляхом додавання ще двох Ebпептидів (але без R71 та S72 та без останніх семи С-кінцевих амінокислот) та додавання останного Еb-пептиду (але без R71 та S72) на С-кінці поліпептиду. Цю конструкцію частопозначають як мультимер IGF-1-Eb. фіг. 6Б показаний поліпептид-попередник IGF-1-Eab з 4 наборами модифікацій: делеція G1, P2 та Е3; мутація, що приводить до заміни R37 на А; делеція R71 та S72; та інсерція амінокислот 93-102 Еа між амінокислотами 95 та 96 Еb; на фіг. 7А - порівняльний аналіз послідовності людського IGF-1 (SEQ ID NO:1) і послідовності IGF-1 відповідної тварини. Представлено всі проаналізовані види тварин та реєстраційні номери в GenBank відповідних їм послідовностей. G1, P2, Е3 є консервативними для всіх проаналізованих видів крім стерляді (у послідовності цього виду S2 замінений на Р2). R36 та R37 є консервативними для всіх проаналізованих видів; на фіг. 7Б - графік, що демонструє філогенію проаналізованих амінокислотних послідовностей щодо людського IGF-1 (SEQ ID NO:1). Під філогенетичним деревом зображена шкала, на якій зазначена кількість «амінокислотних замін» на 100 залишків білкових послідовноетей. Дня розрахунку величин відстаней використовують призначену для цієї мети 3 UA 97953 C2 5 10 15 20 25 30 35 40 45 50 55 60 формулу Кімура (Kimura), отриману на основі даних про кількість помилкових спарювань, що не входять у пролом, та скоректовану з урахуванням замін, що мовчать. Отримані за допомогою комп'ютерної обробки величини являють собою середнє значення розходжень на сайт та знаходяться у діапазоні від 0 до 1. Нуль позначає повну ідентичність, а 1 позначає відсутність ідентичності. На шкалі філогенетичного дерева ці величини помножені на 100; на фіг. 8А - порівняльний аналіз послідовностей людського Еа-пептиду (SEQ ID NO:2) та Еапептидів різних тварин. Представлено всі проаналізовані види тварин та реєстраційні номери в GenBank відповідних ним послідовностей. R71 та S72 є консервативними для всіх проаналізованих видів; на фіг. 8Б - графік, що демонструє філогенію проаналізованих амінокислотних послідовностей щодо людського Еа-пептиду IGF-1 (SEQ ID NO:2); на фіг. 9А - порівняльний аналіз послідовностей людського Eb-пептиду (SEQ ID NO:3) та Еbпептидів різних тварин. Представлено всі проаналізовані види тварин та реєстраційні номери в GenBank відповідних ним послідовностей. R71 та S72 є консервативними для всіх проаналізованих видів; на фіг. 9Б - графік, що демонструє філогенію проаналізованих амінокислотних послідовностей щодо людського Eb-пептиду IGF-1 (SEQ ID NO:3); на фіг.10А - порівняльний аналіз послідовностей людського Ес-пептиду (SEQ ID NO:4) та Еспептидів різних тварин. Представлено всі проаналізовані види тварин та реєстраційні номери в GenBank відповідних ним послідовностей R71 та S72 є консервативними для всіх проаналізованих видів; на фіг. 10Б - графік, що демонструє філогенію проаналізованих амінокислотних послідовностей щодо людського Ес-пептиду IGF-1 (SEQ ID NO:4); на фіг. 11А - порівняльний аналіз послідовності людського IGF-2 (SEQ ID NO:7) та відповідних послідовностей IGF-2 різних видів тварин. Представлено всі проаналізовані види тварин та реєстраційні номери в GenBank відповідних ним послідовностей. R68 є консервативними для всіх проаналізованих видів; D69 є консервативним для всіх видів крім шимпанзе, у послідовності цього виду в цьому положенні знаходиться залишок гістидину; на фіг. 11Б - графік, що демонструє філогенію проаналізованих амінокислотних послідовностей щодо людського IGF-2 (SEQ ID NO:7); на фіг. 12А - порівняльний аналіз послідовності людського Е-пептиду IGF-2 (SEQ ID NO:6) та Е-пептидів IGF-2 різних видів тварин. Представлено всі проаналізовані види тварин та реєстраційні номери в GenBank відповідних ним послідовностей. R68 є консервативними для всіх проаналізованих видів; D69 є консервативним для всіх видів крім шимпанзе, у послідовності цього виду в цьому положенні знаходиться залишок гістидину; на фіг. 12Б - графік, що демонструє філогенію проаналізованих амінокислотних послідовностей щодо людського Е-пептиду IGF-2 (SEQ ID NO:6). Докладний опис винаходу Даний винахід відноситься до нових поліпептидів-попередників IGF-1 та IGF-2, які містять практично відповідний ним Е-пептид, модифікований для попередження, зниження або анулювання типового протеазного розщеплення, відповідального за відділення активної форми IGF-1 або IGF-2 від її Е-пептидів. Можливість застосування поліпептидів, запропонованих у винаході, основана на неочікувано встановленому при створенні винаходу факті, що такі поліпептиди-попередники мають біологічну активність, стабільність та сприятливу дію як фармацевтичні засоби. Скринінг поліпептидів-попередників IGF відносно біологічної активності Можливість застосування будь-якого поліпептиду, запропонованого у винаході, можна оцінювати за допомогою наступних аналізів. Стабільність Поліпептид, запропонований у винаході, повинен мати достатню стабільність у присутності ендогенних протеаз, таких як протеази, що входять до складу людської сироватки, для того, щоб являти собою ефективний лікарський засіб. Для оцінки стабільності експресійним вектором, що кодує поліпептид, можна трансфектувати клітини лінії Cos7 (ATCC) у середовищі DMEM, що містить 10% фетальної телячої сироватки, 100 од./мл пеніциліну та 100 мкг/мл стрептоміцину. Культуральне середовище, яке містить поліпептиди, що секретуються, можна застосовувати для додаткових аналізів, або в іншому варіанті експресійний вектор може кодувати легко доступні мітки, такі як гекса-гістидинову мітку, разом з поліпептидом для полегшення ефективного очищення екепресуємих поліпептидів у культурах Cos7-клітин. Поза залежністю від шляху одержання зразок поліпептиду інкубують у сироватці здорової людини (фірма Sigma) або в ЗФР протягом різних проміжків часу (наприклад, 0, 1, 5, 10 та 16 год.), 4 UA 97953 C2 5 10 15 20 25 30 35 40 45 50 55 60 аналізують за допомогою електрофорезу в поліакриламідному гелі, блотгують на нітроцелюлозу та відповідні білки візуалізують за допомогою первинного антитіла до IGF-1 або IGF-2 та вторинного антитіла, наприклад, кон'югованого з пероксидазою із хрону. Можна використовувати будь-які численні аналогічні методи блоттингу та виявлення, у деяких з яких використовуються флуоресцентні барвники або навіть радіонукліди. Інтенсивність смуги, що відповідає попередникові, у порівнянні з інтенсивністю смуга, що відповідає IGF-1 або IGF-2, повинна свідчити про ступінь розщеплення попередника в різних умовах. Поліпептид, запропонований у винаході, оброблений людською сироваткою протягом 16 год. при 37°С, може характеризуватися співвідношенням нерозщепленого попередника та розщепленого зрілого IGF, що становить приблизно від 1:2 до 1:0,1, наприклад, приблизно від 1:1 до 1:0,5, зокрема співвідношенням приблизно 1:1 або сшввідношенням приблизно 1:0,5. Як правило, попередник повинен бути присутнім у співвідношенні, що становить щонайменше 1:1. Фосфорилювання АКТ Поліпептид, запропонований у винаході, повинен зберігати здатність передавати сигнал через рецептор IGF-1 (сигнали та IGF-1, та IGF-2 передаються через рецептор IGF-1). Для визначення зазначеної здатності здійснювати передачу сигналу можна оцінювати фосфорилювання передачі, що знаходиться по ходу сигналу внутрішньоклітинної мішені, тобто АКТ, у відповідь на зв'язування ліганду на поверхні клітини. Для аналізу фосфорилювання АКТ біобласти лінії С2С12 вирощують в умовах «голодування» у безсироватковому середовищі, а потім стимулюють різними лігандами. Клітини лізують та освітлюють центрифугуванням. Фосфорилювання АКТ та рівні загальної АКТ аналізують за допомогою ELISA з використанням набору для двовалентного (сендвіч) ELISA PathScan phospho АКТ (Ser473) та набору для двовалентного ELISA PathScan АКТ (фірма Cell Signaling) відповідно. Специфічність відносно рецептору IGF-1 Поліпептид, запропонований у винаході, переважно зберігає специфічність відносно рецептору IGF-1 та повинен зв'язуватися із спорідненим інсуліновим рецептором з низькою афінністю. Для оцінки специфічності відносно рецептору зразки поліпептиду додають до вирощених в безсироватковому середовищі NIH3T3клітин, що надекспресують рецептор IGF-1 або інсуліновий рецептор, та рівні фосфорилювання рецептору IGF-1 або фосфорилювання інсулінового рецептору визначають, лізуючи клітини та піддаючи лізати аналізу за допомогою ELISA з використанням людського фоефорильованого IGF-1-рецептору та ELISA-набору для інсулінового рецептору (фірма R&D Systems). Оцінка гіпертрофії in vivo на мишачих моделях Для вирішення питання про те, чи може поліпептид, запропонований у винаході, підвищувати масу скелетних м'язів в умовах, які вже приводять до м'язової гіпертрофії, можна піддавати оброблених та необроблених тварин фізичному навантаженню та визначати чи буде у оброблених поліпептидом тварин утворюватися більше м'язової маси у порівнянні з необробленими тваринами. Моделі фізичного навантаження Одна з відомих у даній галузі моделей основана на застосуванні обертового колеса, рух якого визначається тим навантаженням, що прикладає до нього тварина без додаткового примусу (див., наприклад, Konhilas та ін., Am J Physiol Heart Circ Physiol 289, 2005, стор. 455465). Колесо, яке знаходиться в клітці, що тварина обертає без додаткового примусу, дозволяє усувати інсульти, пов'язані з фізичним та психологічним станом, які характерні для моделей із примусовим навантаженням, і тому є прийнятною моделлю для оцінки лікарських засобівкандидатів, які можна застосовувати для відносно здорових особин, для яких підвищення м'язової маси є бажаним. Можна застосовувати будь-яку придатну лінію мишей. Наприклад, можна довільно розділяти самців мишей лінії C57B1/6J на експериментальну (наприклад, що одержує поліпептидпопередник IGF) та контрольну групи. Тварин поміщають окремо в клітку з колесом для фізичних вправ; контрольних тварин, що ведуть «малорухомий спосіб життя», поміщають в ідентичні клітці без колеса. Колесо для фізичних вправ описане в Allen та ін., J Appl Physiol 90, 2001. cтop. 1900-1908. У цілому, система складається з колеса діаметром 11,5 см з поверхнею для пробігу шириною 5,0 см (модель 6208, фірма Petsmart, Фенікс, шт. Аризона), що забезпечена цифровим магнітним лічильником (модель ВС 600, фірма Sigma Sport, Олні, шт. Іллінойс), що активується при обертанні колеса. Крім того, кожне колесо забезпечене механізмом, що забезпечує опір, що дозволяє регулювати навантаження. Для цієї мети приєднують дротик з нержавіючої сталі до верхньої частини клітки та намотують дріт на нерухомий шків, що закріплюють на колесі, що знаходиться в клітці, на вісі обертання, таким чином, що це не впливає на прикладене до колеса навантаження. Дріт знову закріплюють у верхній частині клітки за допомогою пружини та гвинта. Така конструкція дозволяє здійснювати 5 UA 97953 C2 5 10 15 20 25 30 35 40 45 50 55 60 точне регулювання навантаження на колесо, що рівномірно розподіляється під час обертання колеса. Щодня реєструють тривалість фізичного навантаження та відстань, що пробігає кожна тварина, що знаходиться в умовах, які дозволяють здійснювати фізичне навантаження, протягом всього періоду витримування в зазначених умовах. Всі тварини мали вільний доступ до води та стандартного твердого корму для гризунів. У всіх групах пробіг без додаткового примусу (знаходження в клітці з колесом) можна починати з використанням мишей, вік яких становить у середньому приблизно 12 тижнів. Кожна група тварин продовжувала бігати в умовах різного опору, залежно від експериментальної групи, протягом 50 днів до досягнення тваринами віку приблизно 19 тижнів. Навантаження колеса визначали шляхом підвішування на колесо вантажів відомої маси доти, поки колесо злегка не зміщалося. Всі групи, що знаходяться в умовах фізичного навантаження, спочатку протягом першого тижня мали колесо в клітці без додаткового навантаження. Однак умови «без навантаження» фактично являли собою навантаження в 2 г, що визначали як навантаження, необхідне для підтримки в колесі сили інерції та тертя. З огляду на те, що період акліматизації до колеса становив 1 тиждень, навантаження колеса можна було змінювати з інтервалами в один тиждень за винятком більш високих навантажень, які можна було змінювати через 2 тижні. Зміна навантажень може становити від 2 г аж до 12 г. Тварин, що знаходяться в умовах навантаження, та контрольних тварин, що ведуть «малорухомий спосіб життя», умертвляли під інгаляційним наркозом шляхом зсуву шиї відразу після закінчення конкретного періоду знаходження в умовах навантаження. Визначали масу тіла та певні м'язи швидко вирізали, промивали та заморожували для гістологічного або біохімічного аналізу, які здійснювали пізніше. Фахівцям у даній галузі відомі також альтернативні моделі пов'язаної з фізичним навантаженням гіпертрофії (див., наприклад, модель фізичного навантаження з використанням педального обертового пристрою, що описане в Lerman та ін., J Аррі Physiol 92, 2002, стор. 2245-2255). Модель, створена шляхом ін'єкції кленбутеролу Кленбутерол являє собою 2-адренергічний агоніст, що має посилюючі ріст властивості, що, як встановлено, викликає збільшення м'язової маси. Точний механізм дії кленбутерола поки не відомий, хоча передбачається зниження розщеплення м'язових білків при його застосуванні. У клінічних умовах кленбутерол застосовують як протиастматичний засіб, але ним, очевидно, найбільш широко зловживають як структуроутворюючим агентом (агентом для бодібілдингу) для підвищення м'язової маси як у людей, так і у шоу-тварин. П'яти мишам вводили щодня шляхом ін'єкції кленбутеролу (3 мг/кг, підшкірно (s.c.)) протягом 3, 7 або 14 днів для індукції м'язової гіпертрофії. Миші, яким вводили ін'єкцією ЗФР, служили як негативний контроль. Тварин обстежували щодня (візуальний огляд) у відношенні будь-яких побічних реакцій (тобто неохайність вовняного покриву, сонливість) на обробку. Обробка кленбутеролом потенційно може робити мишей більш полохливими або агресивними, тому за мишами, якщо вони поміщені в клітини групами, необхідно особливо ретельно стежити з позицій бійок одна з іншою. Миші могли вільно рухатися та могли нормально їсти та пити. Мишей обстежували щодня до їх умертвіння в день 3, 7 або 14 та відбирали тканини для додаткового аналізу. Оцінка на моделях м'язової атрофії in vivo Здатність поліпептиду-попереднику IGF, запропонованого у винаході, підтримувати м'язову масу в умовах, які звичайно приводять до зниження м'язової маси, можна оцінювати на різних моделях атрофії скелетних м'язів. При використанні прикладів моделей, описаних нижче, фахівець у даній галузі легко може створювати та здійснювати контрольовані експерименти, що включають введення та застосування поліпептидів-попередників IGF для вирішення питання про те, чи можуть зазначені поліпептиди приводити до збільшення м'язової маси. Наприклад, самців мишей лінії С57В16/2 купували у фірми The Jackson Laboratories. На початку кожного експерименту миші повинні були мати вік приблизно 9 тижнів. Як правило, мишей поміщали в клітки-мікроізолятори зі звичайним кормом для гризунів. На початку кожного експерименту мишей зважували. Наприкінці кожного експерименту мишей звичайно умертвляли інгаляційним наркозом з наступним зсувом шиї та м'язові тканини збирали для додаткової обробки. Мишей зважували для визначення «кінцевої маси тіла». Скелетні м'язи, які можна одержувати, являли собою передній великоберцовий м'яз, довгий розгинач пальців стопи, камбаловидний м'яз та ікроножний м'яз. Довільно збирали інші тканини, такі як: серце, печінка, селезінка, нирки, яєчка та головний мозок. Всі м'язи та тканини повністю висікали та зважували на вагах, нижня межа яких становить 0,0001 г. Потім тканини швидко заморожували в рідкому азоті для наступної екстракції РНК та білків або швидко заморожували зануренням в ОСТ (орто-хлортолуол) на корковому диску. М'язи, заморожені на корковому диску для наступного одержання кріозрізів, 6 UA 97953 C2 5 10 15 20 25 30 35 40 45 50 55 60 занурювали в ізопентан, охолоджений за допомогою рідкого азоту до утворення густої суспензії. Всі зразки зберігали при -80°С. Обробка дексаметазоном Фармакологічний метод індукції м'язової кахексії у мишей передбачає щоденну внутріочеревинну ін'єкцію дексаметазоном у дозі 20 мг/кг. Дексаметазон є синтетичним представником класу глюкокортикоїдних гормонів. Він має протизапальну та імуносупресорну активність, що перевищує по активності гідрокортизон приблизно в 40 разів. Дексаметазон застосовують для лікування запалення та аутоімунних станів, наприклад, ревматоїдного артриту. Його дають також страждаючим раком пацієнтам, що піддаються хіміотерапії, для зниження деяких побічних дій протипухлинного лікування. Дексаметазон викликає м'язову атрофію як у мишей, так і у людей. Мишам вводили внутрішньочеревинною (і.р.) ін'єкцією дексаметазон протягом 3, 7 або 14 днів. У день закінчення досліду тварин умертвляли за допомогою СО 2, та виділяли м'язи лап. Тварин обстежували щодня (візуальний огляд) у відношенні будь-яких побічних реакцій (тобто неохайність вовняного покриву, сонливість) на обробку. Миші, як правило, були рухливими та могли нормально їсти та пити. Для створення негативного контролю мишам вводили ін'єкцією ЗФР. Іммобілізація за допомогою пов’язки Фізичне незастосування різних груп м'язів приводить до атрофії цих м'язів. Доведено, що фіксація гомілковостопного суглобу («закріплена п'ята» або накладення пов'язки) є дуже ефективним та дозволяючим одержувати відтворювані результати засобом індукції фізичної іммобілізації мускулатури задніх кінцівок щурів та мишей. Для іммобілізації мишей анестезували ізофлуораном. Гомілковостопний та колінний суглоб фіксували під кутом 90° шляхом накладення пов'язки з легкого перев'язного матеріалу (VETLITE) навколо суглобів. Матеріал замочували в теплій воді та потім обертали кінцівку, залишаючи вільними суглоби пальців стопи та тазостегновий суглоб. Суглоби підтримували в положенні під кутом 90° до висихання перев'язного матеріалу. Контралатеріальні кінцівки служили як контроль. Мишам давали відійти від наркозу та поміщали у звичайні кліткимікроізолятори. Як встановлено, перев'язка не привела до підвищеного стресу та тварини вільно пересувалися в клітці до корму та питва. Однак мишей щодня обстежували відносно побічних дій, які можуть впливати на масу тіла, активність та збудливість. Після накладення на мишей пов'язки тварин щодня обстежували для одержання гарантій того, що пов'язка зберігається на місці, оскільки миші можуть її зжовувати. Тварини могли рухатися, пити та їсти після відходу від наркозу, та вони не мали потреби в спеціальному ложі, клітках або іншій допомозі. Денервація Як правило, мишей для здійснення денервації анестезували за допомогою газу ізофлуорану. За допомогою асептичних хірургічних процедур (три промивання бетадином та кінцеве промивання етанолом) оголювали правий сідничний нерв у середині стегна та вирізали шматок розміром 2-5 мм. Контралатеріальна кінцівка служила як контроль. Більш конкретно, розріз на шкірі закривали за допомогою шкірного затискача та тварині вводили ін'єкцією одну дозу бупренорфіну перед тим як дати їм відійти від наркозу. Через 3, 7 або 14 днів після операції тварин умертвляли інгаляційним наркозом з наступним зсувом шиї та м'язи (комплекс ікроножних м'язів, передній великоберцовий м'яз, довгий розгинач пальців стопи, камбаловидний м'яз) виділяли для гістологічних та біохімічних аналізів. Оскільки сідничний нерв був розсічений, то відповідна кінцівка, іннервована ним, ставала нерухомою, індукуючи атрофію скелетних м'язів. Незважаючи на це тварини могли рухатися, пити та їсти після відходу від наркозу, та вони не мали потреби в спеціальному ложі, клітках або іншій допомозі. Проте, тварин обстежували відразу після операції та їх відходу від наркозу (1-2 год.). Крім того, області розрізу та загальний стан здоров'я тварин оцінювали протягом 3 днів після операції. Шкірний затискач видаляли через 7-10 днів після операції. Генетичні моделі Як моделі м'язової атрофії можна використовувати також генетично змінених трансгенних мишей. Наприклад, так називані міні-миші (Mini Mice) (фірма The Jackson Laboratory, лінія № 003258) містять мутацію, що виключає («knock оut»-мутацію) у гені IGF-1, що приводить до аномально зниженого післянатального росту, а також низької маси тіла та росту. Додаткову інформацію можна почерпнути в Powell-Braxton та ін., Genes Dev 7, 1993, стор. 2609-2617. Крім того, так звані міді-миші (Midi Mice) (фірма The Jackson Laboratory, лінія Xa 003259) містять іншу мутацію в гені, що приводить до одержання гіпоморфного гену, що обумовлює низьку масу 7 UA 97953 C2 5 10 15 20 25 30 35 40 45 50 55 60 дорослих особин та інші серцево-судинні фенотипи. Додаткову інформацію можна почерпнути в Lembo та ін., J Clin Invest 98, 1996, стор. 2648-2655. Мутації або модифікації попередників IGF. що мають вирішальне значення та є необов'язковими Мутації, що мають вирішальне значення Даний винахід оснований, зокрема, на отриманих даних про те, що поліпептид-попередник IGF, що містить практично відповідний йому Е-пептид, зберігає біологічну активність та стабільність у присутності сироватки. Для гарантії того, що Е-пептид не відщеплюється ендогенними протеазами, мішенню яких є двоосновний протеазний сайт, звичайно кожну із двох N-кінцевих двоосновних амінокислот Е-пептиду в попереднику видаляють шляхом делеції, змінюють шляхом мутації або маскують іншим способом. У випадку hIGF-1 ці дві амінокислоти являють собою R71 та S72, а у випадку hIGF-2, ці перші дві амінокислоти являють собою R68 тa D69. Зазначене розщеплення можуть попереджати різні модифікації: (1) делеція одного або обох двоосновних залишків, (2) мутація одного або обох двоосновних залишків, що приводить до заміни на неосновну амінокислоту, таку як аланін, (3) інсерція однієї або двох неосновних амінокислот між двухосновними залишками, (4) вбудовування сайту глікозилювання біля двоосновних залишків, що є достатнім для маскування протеазного сайту, (5) сайтнаправлене пегілювання із застосуванням заміни кожного із двоосновних залишків або вбудовування поблизу або між двоосновними залишками з використанням амінокислоти, що не зустрічається в природних умовах, як буде описано нижче. Крім того, залишки К68 та К65, імовірно, відіграють роль у розщепленні IGF-1/E-пептиду; таким чином, мутації або делеції зазначених залишків можна застосовувати в сполученні з будь-якою зазначеною вище тактикою, застосовуваною у відношенні двоосновних амінокислот. Мутації на N-кінці зрілого IGF У деяких варіантах здійснення винаходу поліпептиди-попередники IGF мають делеції або мутації декількох перших N-кінцевих амінокислот. У випадку IGF-1, будь-які перші три N-кінцеві амінокислоти можна вилучати шляхом делеції або піддавати мутації, у той час як у випадку IGF2, будь-які перші шість N-кінцевих амінокислот можна вилучати шляхом делеції або піддавати мутації. Було встановлено, що певні N-кінцеві амінокислоти відщеплюються в природних умовах in vivo, та інтродукція зазначених мутацій або делецій мінімізує in vivo асоціації поліпептидів, запропонованих у винаході, з IGF-зв'язуючими білками (IGFBP). Взаємодія IGF-1 та IGF-2 з рецептором IGF-1 регулюється IGFBP-білками. Встановлено, що всі шість IGFB інгібують дію IGF (насамперед IGFBP5), але в деяких випадках виявлена стимулююча дія. Щонайменше 99% IGF у кровотоці в нормі пов'язане з IGFBP. IGFBP, що має найбільш високий рівень у кровотоці в післянатальний період, являє собою IGFBP3, що може зв'язуватися з однаковою афінністю та з IGF-1, та з IGF-2. Укорочений IGF-1, що зустрічається в природних умовах, (що має делецію G1, Р2 та Е3) зв'язується з IGFBP3 у кілька разів з більш низькою афінністю в порівнянні із IGF1, що зустрічається в природних умовах. Крім того, G3 є важливим для зв'язування з IGFBP, a G6 відіграє аналогічну роль в IGF-2-пептиді. Таким чином, у випадку попередника hIGF-1, кожний із залишків G1, P2 та Е3 можна вилучати шляхом делеції або піддавати мутації або індивідуально, або в комбінації. Коли потрібна мутація, то можна інтродукувати мутацію, що приводить до заміни на аланін. В іншому прикладі у випадку попереднику hIGF-2, кожний із залишків Р4, S5 та Б6 можна вилучати шляхом делеції або піддавати мутації або індивідуально, або в комбінації. Коли потрібна мутація, то можна інтродукувати мутацію, що приводить до заміни на аланін. Мутації на зашийках 36 та 37 IGF-1 можна розщеплювати за допомогою серінових протеаз, присутніх у людській сироватці. Мутація або R36, або R37, що приводить до зміни на А, може попереджати розщеплення IGF-1 у цьому передбачуваному сайті розщеплення, що знаходиться між R36 та R37. У випадку hIGF-2, R38 можна піддавати мутації або вилучати шляхом делеції для попередження цього розщеплення, що робить несприятливий вплив. Застосування глікозилювання Час напівжиття in vivo поліпептидів, запропонованих у винаході, можна підвищувати, додаючи N-зв'язані сайти глікозилювання або в IGF, або у фрагменти Е-пептиду попередника при експресії в клітинах ссавців або інших еукаріотичних клітинах, у яких може проходити Nзв'язане глікозилювання. Було встановлено in vitro, що людський IGF-1-Еа є глікозильованим на N92 та N100, оскільки ці фрагменти Еа відповідають консенсусній послідовності N-зв'язаного 8 UA 97953 C2 5 10 15 20 25 30 35 40 45 50 55 60 сайту глікозилювання N-X-S/T, де X може позначати будь-яку амінокислоту, а третя амінокислота триплету являє собою або S, або Т. Відомо також, що амінокислотний контекст, що примикає до консенсусу, повинен впливати на те, наскільки сильно глікозилюється аспарагін. Таким чином, однією зі стратегій інтродукції сайту глікозилювання в Еb або Ес є вбудовування амінокислот Еа поблизу консенсусної послідовності приблизно в ту ж ділянку Еb або Ес. Конкретне застосування зазначеної стратегії проілюстровано нижче в прикладах. У будь-якому разі, будь-яку іншу консенсусну послідовність N-зв'язаного сайту глікозилювання, включаючи навколишні контекстні амінокислоти, що відомі фахівцям у даній галузі, можна вбудовувати в поліпептид-попередник, запропонований у винаході. Крім того, можна здійснювати О-зв'язане глікозилювання поліпептиду, запропонованого у винаході, шляхом вибору конкретного хазяїна, застосовуваного для одержання поліпептиду. Наприклад, застосування певних ліній дріжджів для експресії IGF-1 приводить до додавання олігосахаридів до залишків серіну або треоніну (див., наприклад, US 5273966). Додавання полі(етиленгліколя) Доведено, що кон'югація з полі(етиленгліколем) (ПЕГ; пегілювання) робить сприятливу дію відносно подовження часу напівжиття терапевтичних лікарських засобів. Можна чекати, що пегілювання поліпептидів-попередншсів IGF, запропонованих у винаході, може мати аналогічну перевагу з позицій фармації. Методи пегілювання IGF-1 добре відомі в даній області (див,, наприклад, опубліковану заявку на патент США 2006/0154865, у якій описана сприятлива дія кон'югату лізин-монопегільований IGF-1). Таке лізин-монопегілювання можна адаптувати для поліпептидів-попередників IGF, запропонованих у винаході. Крім того, пегілювання можна здійснювати в будь-якій частині поліпептиду, запропонованого у винаході, шляхом інтродукції амінокислот, що не зустрічаються в природних умовах. Деякі амінокислоти, що не зустрічаються в природних умовах, можна інтродукувати за допомогою технології, описаної в Deiters та ін. , J Am Chem Soc 125, 2003, стор. 11782-11783; Wang та Science, Science 301, 2003, стор. 964-967; Wang та ін., Science 292, 2001, стор. 498-500; Zhang та ін., Science 303, 2004, crop. 371-373 або в US 7083970. У цілому, для деяких із зазначених систем експресії застосовували сайтнаправлений мутагенез для інтродукції нонсенсу-кодону, такого як амберчкодон TAG, у відкриту рамку зчитування, що кодує поліпептид, запропонований у винаході. Такі експресійні вектори потім інтродукують у хазяїна, що може використовувати тРНК, специфічну для інтродукованого нонсенсу-кодону, та «завантажують» обраною амінокислотою, що не зустрічається в природних умовах. Конкретні амінокислоти, що не зустрічаються в природних умовах, які мають сприятливу дію при їх застосуванні як компоненти для кон'югації з поліпептидами, запропонованими у винаході, являють собою амінокислоти з бічними ланцюгами, які несуть ацетиленові групи та азидогрупи. Поліпептиди-попередники IGF, які містять ці нові амінокислоти, потім можна пегілювати на зазначених обраних сайтах у білку. Крім того, такі пегільовані молекули IGF, що не містять Е-пептид, можна застосовувати також як терапевтичні агенти. Мультимери Е-пептидів У певних фармакологічних контекстах може вимагатися збільшувати розмір пептидного або білкового лікарського засобу для гарантії того, що лікарський засіб зберігається на одній або на іншій стороні гематоенцефалічного бар'єру. Оскільки зрілі молекули IGF являють собою відносно короткі пептиди, навіть у випадку, коли в них зберігається Е-пептид, може виявитися важливим збільшувати розмір поліпептидів, запропонованих у винаході. Одним зі шляхів досягнення цього є створення мультимерів Е-пептидів на С-кінці поліпептиду-попереднику IGF, що проілюстровано в деяких описані нижче прикладах. С-кінцева делеція Е-пептидів Можна припустити, що наявність вільного цистеїну в положенні 81 Еb може привести до гомодимеризації або іншим діям, які можуть приводити до зниження активності поліпептидів, запропонованих у винаході, як лікарські засоби. Так, делеція або мутація С81 в Еb може оптимізувати активність лікарського засобу. У конкретному варіанті сприятливу дію робить делеція останніх семи амінокислот Еb (тобто, амінокислот 81-87). Інші мутації або модифікації Додаткові мутації або модифікації IGF, яким можна піддавати поліпептиди-попередники IGF, запропоновані у винаході, описані в US 5077276; та в опублікованих заявках на патент США 2005/0287151, 2006/0211606 та 2006/0166328. Під обсяг винаходу підпадають крім людських IGF-1 та IGF-2 всі відомі або поки невідомі послідовності попередників IGF-1 або IGF-2 тварин крім людини, які містять практично відповідний їм Е-пептид, якщо відщіплення Е-пептиду, що має місце у природних умовах, анульоване або знижене в результаті модифікацій, запропонованих у даному винаході. 9 UA 97953 C2 5 10 15 20 25 30 35 40 45 50 55 60 Кращий для застосування тип IGF залежить від видів, що підлягають лікуванню. Переважно, щоб IGF відповідав виду, для лікування якого його застосовують, наприклад, коли лікуванню підлягають корови, тоді кращим типом IGF є бичачий IGF. Хоча, очевидно, всі форми IGF впливають на різних індивідуумів завдяки високій гомології їхніх послідовностей, у відповідних видах можна уникати потенційних небажаних імунологічний ускладнень, які є результатом індукції імунної відповіді на IGF з інших видів. Одним з варіантів здійснення винаходу є модифіковані послідовності попередників IGF-1 із тварин крім людини. Кращими є послідовності попередників IGF-1 із хребетних тварин, які містять практично відповідний їм Е-пептид, у яких відщіплення Е-пептиду, що має місце у природних умовах, анульоване або знижене в результаті модифікацій, запропонованих у даному винаході. Прикладами таких послідовностей є (але не обмежуючись тільки ними) послідовності мишей, щурів, корів, свиней, коней, овець, кіз, птахів, собак, кішок, риб тощо, отримані з будьякого джерела, у тому числі природного, синтетичного або рекомбінантного. Іншим варіантом здійснення винаходу є модифіковані послідовності попередників IGF-2 із тварин крім людини. Кращими є послідовності попередників IGF-2 із хребетних тварин, які містять практично відповідний їм Е-пептид, у яких відщіплення Е-пептиду, що має місце у природних умовах, анульоване або знижене в результаті модифікацій, запропонованих у даному винаході. Прикладами таких послідовностей є (але не обмежуючись тільки ними) послідовності мишей, щурів, корів, свиней, коней, овець, кіз, птахів, собак, кішок, риб тощо, отримані з будьякого джерела, у тому числі природного, синтетичного або рекомбінантного. Терапевтичне застосування поліпептидів-попередників IGF Показання Винахід відноситься також до застосування поліпептиду-попереднику IGF, запропонованого у винаході, для готування лікарського засобу, призначеного для лікування або попередження захворювань кістково-м'язової системи. Крім того, винахід відноситься також до застосування поліпептидів-попередників IGF для підвищення м'язової або кісткової маси в індивідуума, поза залежністю від того має або не має індивідуум ризику розвитку захворювання кістково-м'язової системи або має або не має захворювання кістково-м'язової системи. Зокрема, захворювання кістково-м'язової системи може являти собою м'язову атрофію. Відомо багато типів м'язової атрофії, що у тому числі є результатом лікування глюкокортикоїдами, такими як кортизол, дексаметазон, бетаметазон, преднізон, метилпреднізолон або преднізолон. М'язова атрофія може бути результатом денервації, викликаної травмою нерву або дегенеративної, метаболічної або запальної невропатії (наприклад, синдром Гійєна-Барре, периферична невропатія або вплив токсинів, що перебувають у навколишньому середовищі, або лікарських засобів). Крім того, м'язова атрофія може бути результатом захворювання моторних нейронів у дорослих, дитячої атрофії спинних м'язів, юнацької атрофії спинних м'язів, аутоімунної моторної невропатії із багатоочаговою провідниковою блокадою, паралічу, пов'язаного з «ударом» або ушкодженням спинного мозку, скелетною іммобілізацією через травму, пролонгованого перебування на постільному режимі, добровільної зниженої активності, примусової зниженої активності, метаболічного стресу або недостатності живильних речовин, раку, СНІДу, голодування, гострого некрозу скелетних м'язів, захворювання щитовидної залози, діабету, доброякісної вродженої гіпотонії, захворювання центрального ядра, немалинової (непрогресуючої вродженої ниткоподібної) міопатії, міопатії (центронуклеарної) м'язових трубочок, опікового ушкодження, хронічного обструктивного захворювання легенів, хвороби печінки, сепсису, ниркової недостатності, застійної серцевої недостатності або старіння. Захворювання кістково-м'язової системи може бути пов'язане із синдромом м'язової дистрофії, таким як симптом Дюшенна, симптом Беккера, міотонія, плече-лопатко-лицьова м'язова дистрофія, м'язова дистрофія Емері-Дрейфуса, глазофарингеальна м'язова дистрофія, лопатко-лицьова м'язова дистрофія, тазово-плечова м'язова дистрофія, вроджена м'язова дистрофія або принадбана дистальна міопатія. Захворювання кістково-м'язової системи може являти собою також остеопороз, перелом кістки, низький зріст або карликовість. Передбачається, що IGF-1 можна застосовувати для лікування нечутливого до інсуліну діабету, оскільки IGF-1 може зв'язуватися також з гетеродимерами рецептору IGF-1 та інсулінового рецептору. Таким чином, поліпептиди, запропоновані у винаході, можна застосовувати для лікування діабету. IGF-1 має нейротрофну дію та подовжує тривалість існування нейронів. Було висловлене припущення про те, що IGF-1 можна застосовувати для лікування випадків загибелі моторних 10 UA 97953 C2 5 10 15 20 25 30 35 40 45 50 55 нейронів, що характерно для аміотрофічного бічного склерозу (ALS), атрофії головного мозку, старіння та деменції. Таким чином, поліпептиди, запропоновані у винаході, можна застосовувати для лікування станів, пов'язаних із загибеллю нейронів, таких як ALS, атрофія головного мозку або деменція. IGF-1 підвищує популяції як лейкоцитів, так і еритроцитів та має аддитивну дію при введенні в сполученні з еритропоетином. Таким чином, поліпептиди, запропоновані у винаході, можна застосовувати для лікування анемії. Оскільки IGF-1 та 1GF-2 є широко розповсюдженими та основними регуляторами розподілу клітин та росту хребетних тварин, може виявитися доцільним застосовувати їх у різних ветеринарних методах для екзогенного посилення або підтримки росту тварин. Прикладами такого застосування є (але не обмежуючись тільки ними): (I) збільшення швидкості росту та/або розміру тварини, наприклад, прискорення збільшення м'язової тканини у свиней, великої рогатої худоби, домашнього птаха та риби; (II) підвищення ефективності перетворення корму в тканині організму (співвідношення худої маси та жирової маси), наприклад, у свиней, великої рогатої худоби, овець, домашнього птаха та риби; та (III) збільшення виробництва молока у тварин на стадії лактації, наприклад, молочної великої рогатої худоби, овець, кіз. Інші ветеринарні терапевтичні застосування включать (але не обмежуючись тільки ними): (IV) лікування тварини, що має симптоми, пов'язані з кахексією, травмою або іншими пов'язаними з виснаженням захворюваннями, наприклад, тварин-компаньйонів, таких як собаки, кішки та коні; та (V) лікування тварин, що знаходяться на стадії лактації, для поліпшення здоров'я немовлят, наприклад, що знаходяться на стадії лактації свиноматок для поліпшення характеристик немовлят. Способи введення Поліпептиди, запропоновані у винаході, можна вводити різними шляхами, включаючи застосування носіїв для введення генів. Для введення в терапевтичних цілях поліпептиду,запропонованого у винаході, можна застосовувати методи, відомі в даній галузі для терапевтичного введення таких агентів, як білки або нуклеїнові кислоти, наприклад, такі методи, як трансфекція клітин, генна терапія, безпосереднє введення в допомогою наповнювача, застосовуваного для введення, або фармацевтично прийнятного носія, непряме введення за допомогою рекомбінантних клітин, які містять нуклеїнову кислоту, що кодує поліпептид. У даній галузі відомі різні системи введення, та їх можна застосовувати для введення поліпептиду, запропонованого у винаході, такі, наприклад, як капсулювання в ліпосоми, мікрочастинки, мікрокапсули, рекомбінантні клітини, що мають здатність експресувати білок, опосередковуваний рецептором ендоцитоз (див., наприклад, Wu та Wu, J Biol Chem 262, 1987, crop. 4429-4432), конструювання нуклеїнових кислот як частину векторів на основі ретровірусів, адено-ассоційованих вірусів, аденовірусів, поксивірусів (наприклад, пташиного поксивірусу, насамперед, поксивірусу домашнього птаха) або інших векторів тощо. Методи інтродукції можуть бути ентеральні або парентеральні та являють собою (але не обмежуючись тільки ними) внутрішкірний, внутрім'язовий, внутріочеревшший, внутрішньовенний, підшкірний, легеневий, інтраназальний, внутріочний, епідуральний та пероральні шляхи. Поліпептиди можна вводити за допомогою будь-якого зручного шляху, наприклад, шляхом інфузії або болюсної ін'єкції, шляхом абсорбції через епітеліальні або слизово-шкірні покриви (наприклад, слизову оболонку рота, ректальну або кишкову слизову оболонку тощо) та можна вводити разом з іншими біологічно активними агентами. Введення може бути системним або місцевим. Крім того, може виявитися бажаним інтродукувати фармацевтичні композиції, запропоновані у винаході, у центральну нервову систему за допомогою будь-якого прийнятного шляху, включаючи внутрішньошлуночкову та підоболонкову ін'єкцію; внутрішньошлуночкову ін'єкцію можна полегшувати, використовуючи внутрішньошлуночковий катетер, наприклад, прикріплений до резервуара, такого як резервуар Оммайя. Легеневе введення можна здійснювати також за допомогою, наприклад, інгалятора або розпилювача та у вигляді препаративної форми, що містить пропеллент. У конкретному варіанті здійснення винаходу може вимагатися місцеве введення фармацевтичних композицій, запропонованих у винаході в область, що підлягає лікуванню; для цієї мети можна використовувати (але не обмежуючись тільки ними) місцеву інфузію в процесі хірургічної операції, місцеве нанесення, наприклад, шляхом ін'єкції, за допомогою катетера або за допомогою Імплантату, імплантат можна виготовляти з пористого, непористого або 11 UA 97953 C2 5 10 15 20 25 30 35 40 45 50 55 60 желеподібного матеріалу, включаючи мембрани, такі як сіаластичні мембрани, волокна або замінники шкіри, що поступають в продаж. В іншому варіанті здійснення винаходу діючу речовину можна вводити в пухирці, насамперед у ліпосоми (див. Langej, Science 249, 1990, стор. 1527-1533). Ще в одному варіанті здійснення винаходу діючу речовину можна вводити за допомогою системи з контрольованим вивільненням. В одному з варіантів здійснення винаходу можна використати насос. В іншому варіанті здійснення винаходу можна використовувати полімерні матеріали (див. Howard й ін., J Neurosurg71, 1989, с. 105). В іншому варіанті здійснення винаходу, у якому діюча речовина, запропонована у винаході, являє собою нуклеїнову кислоту, що кодує поліпептид, запропонований у винаході, нуклеїнову кислоту можна вводити in vivo для посилення експресії кодуємого нею білку, створюючи її як частину відповідного нуклеотидного експресійного вектору та вводячи його таким чином, щоб він ставав внутрішньоклітинним, наприклад, застосовуючи ретровірусний вектор (див., наприклад, US 4980286), або шляхом безпосередньої ін'єкції, або шляхом застосування бомбардування мікрочастинками (наприклад, за допомогою генної гармати; Biolistic, фірма Dupont), або наносячи покриття з використанням ліпідів, або за допомогою рецепторів, що знаходяться на клітинній поверхні, або за допомогою трансфектуючих агентів, або шляхом введення в сполученні з гомеобокс-подібним пептидом, для якого відома здатність проникати в ядро (див. , наприклад, Joliot й ін., Рrос. Natl. Acad. Sci. USA 88, 1991. стор. 1864-1868) і т.д. В іншому варіанті нуклеїнову кислоту можна «продукувати всередину клітини та включати в ДНК клітину-хазяїна для експресії шляхом гомологічної рекомбінації. Трансфекція клітин та генна терапія Даний винахід відноситься до застосування нуклеїнових кислот, які кодують поліпептиди, запропоновані у винаході, для трансфекції клітин in vitro та in vivo. Ці нуклеїнові кислоти можна вбудовувати в кожний з добре відомих векторів для трансфекції клітин- або організмів-мішеней. Нуклеїновими кислотами також трансфектують клітини ex vivo та in vivo за допомогою взаємодії вектору та клітини-мішені. Композиції вводять (наприклад, шляхом ін'єкції в м'яз) індивідуумові в кількості, достатній для того, щоб викликати терапевтичну відповідь. Іншим об’єктом винаходу є спосіб лікування галузі-мішені, тобто клітини- або тканинимішені, у людини або іншої тварини, що полягає в тому, що трансфектують клітину нуклеїновою кислотою, що кодує поліпептид, запропонований у винаході, де нуклеїнова кислота включає індуцибельний промотор, функціонально пов'язаний з нуклеїновою кислотою, що кодує злитий поліпептид, що забезпечує спрямований перенос. Процедури генної терапії, які застосовують для лікування або попередження хвороб людини, див., наприклад, в Van Brunt, Biotechnology 6, 1998, crop. 1149-1154. Комбіновані терапії Відповідно до декількох варіантів здійснення винаходу поліпептиди, запропоновані в даному винаході, можна вводити в сполученні з однією або декількома додатковими сполуками або видами терапії. Наприклад, кілька поліпептидів можна вводити в сполученні з однією або декількома терапевтичними сполуками. Комбінована терапія може передбачати одночасне введення або введення, що чергується. Крім того, комбінацію можна використовувати для гострого або постійного застосування. Поліпептиди, запропоновані у винаході, можна вводити в сполученні з анаболічними агентами, такими як тестостерон або специфічні модулятори андрогенного рецептору (SARM). До інших анаболічних агентів відносяться гормон росту (GH) або молекули, які індукують вивільнення GH. Грелін являє собою особливо цінний агент для комбінованої терапії кахексії, оскільки грелін може викликати посилення апетиту. Аналогічно цьому, поліпептиди, запропоновані у винаході, можна поєднувати з білковими добавками для посилення анаболізму, або поєднувати з фізіотерапією для підвищення маси тіла. Будь-яка молекула, що інгібує міостатин, також є кандидатом для комбінованої терапії. Фармацевтичні композиції Даний винахід відноситься також до фармацевтичних композицій, які містять білокпопередник IGF, запропонований у винаході, та фармацевтично прийнятний носій. Під поняттям «фармацевтично прийнятний» мають на увазі дозволений регулювальним органом Федерального або державного управління або включений у список Фармакопеї США або іншої загальноприйнятої фармакопеї як дозволений для застосування на тваринах або людині. Поняття «носій» відноситься до розріджувача, ад'юванту, ексципієнту або наповнювачу, з яким здійснюють введення терапевтичного засобу. Зазначені фармацевтично прийнятні носії можуть являти собою стерильні рідини, такі як вода та масла, включаючи вазелін, масла тваринного походження, рослинного походження або синтетичні масла, такі як арахісове масло, соєве масло, мінеральне масло, кунжутне масло тощо. Прийнятними фармацевтичними 12 UA 97953 C2 5 10 15 20 25 30 35 40 45 50 55 ексципієнтами є крохмаль, глюкоза, лактоза, сахароза, желатин, солод, рис, борошно, крейда, силікагель стеарат натрію, моностеарат гліциріну, тальк, хлорид натрію, знежирене сухе молоко, гліцерин, пропіленгліколь, вода, етанол тощо. За необхідності композиція може містити також мінорні кількості змочувальних або емульгуючих агентів або рН-забуферюючих агентів. Такі композиції можуть мати форму розчинів, суспензій, емульсії, таблеток, пігулок, капсул, порошків, форм із пролонгованим вивільненням тощо. Композицію можна одержувати у формі супозиторію з використанням традиційних зв'язувальних речовин та носіїв, таких як тригліцериди. До складу форми для перорального застосування можуть входити стандартні носії, такі як ті, що мають фармацевтичну чистоту, маніт, лактоза, крохмаль, стеарат магнію, сахарин натрію, целюлоза, карбонат магнію тощо. Приклади прийнятних фармацевтичних носіїв наведені в «Remington's Pharmaceutical Sciences» під ред. Е. W. Martin. Відповідно до деяких варіантів здійснення винаходу композицію одержують за допомогою загальноприйнятих процедур у вигляді фармацевтичної композиції, адаптованої для внутрішньоаснного введення людині. За необхідності до складу композиції можна включати солюбілізатор та місцевий анестетик, такий як лідокаїн, для ослаблення болю в місці ін'єкції. Якщо композиція підлягає введенню шляхом інфузії, її можна одержувати та випускати разом із флаконом для інфузії, що містить стерильні воду або фізіологічний розчин, що мають фармацевтичну чистоту. Коли композицію вводять шляхом ін'єкції, тоді можна поставляти ампулу зі стерильною водою для ін'єкцій або фізіологічним розчином таким чином, щоб інгредієнти можна було змішувати перед введенням. Поліпептиди, запропоновані у винаході, можна включати в препаративну форму в нейтральному вигляді або у вигляді солі. Фармацевтично прийнятні солі являють собою солі, утворені з вільними аміногрупами, що є похідними соляної, фосфорної, оцтової, щавлевої, винної кислот і т.д., та солі, утворені з вільними карбоксильними групами, що є похідними натрію, калію, амонію, кальцію, гідроксидів заліза, ізопропіламіну, триетиламіну, 2етиламіноетанолу, гістидину, прокаїну тощо. Кількість поліпептиду, запропонованого у винаході, що повинна мати ефективність при лікуванні стану або захворювання, можна визначати за стандартними клінічними методиками на основі приписання лікаря. Крім того, необов'язково можна проводити аналізи in vitro, які сприяють визначенню оптимальних діапазонів доз. Точна доза, яку слід вводити в препаративну форму, залежить також від шляху введення та серйозності стану та повинна вибиратися відповідно до рішення лікаря та залежно від індивідуального стану індивідуума. Однак прийнятні діапазони доз для внутрішньовенного введення становлять, як правило, приблизно 20-5000 мкг діючої речовини на кг маси тіла. Прийнятні діапазони доз для інтраназального введення становлять, як правило, приблизно від 0,01 пг/кг маси тіла до 1 мг/кг маси тіла. Ефективні дози можна визначати шляхом екстраполяції із кривих залежності відповіді від дози, отриманих in vitro або з використанням для оцінки змодельованих на тваринах систем. Зокрема, можлива схема прийому лікарського засобу може становити від 60 до 120 мкг/кг маси тіла шляхом підшкірної ін'єкції двічі на день. Застосування у ветеринарії Крім вищевказаних методів введення людям, можна розглядати також застосування у ветеринарії. Дози, які вводять здоровій тварині та тварині, що страждає від захворювання, можуть відрізнятися. Вибір відповідної дози легко можуть здійснювати фахівці в даній галузі на основі аналізів, відомих у даній галузі, наприклад, на основі описаного нижче аналізу проліферації міобластів (приклад 79) або аналізу епітеліальної тканини молочної залози (приклад 80). Загальні аналізи для оцінки IGF також добре відомі в даній галузі, наприклад, аналізи, описані в прикладі 81. Фахівцям у даній галузі повинно бути очевидно, що для деяких видів тварин може бути характерна сезонна фертильність, що залежить від довжини фотоперіоду. У будь-яких варіантах ветеринарного методу або його застосування необов'язково можна починати лікування в конкретний момент репродуктивного циклу тварини для того, щоб досягати необхідної дії. Фахівцям у даній галузі повинно бути очевидним, що репродуктивний статус та цикл легко визначати та при необхідності синхронізувати за допомогою відповідного режиму. При застосуванні при ветеринарних показаннях крім раніше описаних методів, застосовуваних на людях, пептид IGF-1 або IGF-2, запропонований у даному винаході, можна використовувати шляхом вливання в ротову порожнину тварини або у вигляді добавки до пероральних або твердих кормів для тварин. Нижче винахід описаний додатково за допомогою прикладів, що не обмежують його обсяг. 60 13 UA 97953 C2 5 10 15 20 25 30 35 40 45 50 55 60 Приклади Приклад 1 Конструювали експресійний ДНК-вектор, що кодує поліпептид-попередник hIGF-1-Еа, що містить наступні модифікації: делеція G1, делеція Р2 та делеція Е3; мутація, що приводить до заміни R37 на А; та делеція R71 та делеція S72. Зазначені мутації іноді позначають як «3mut» у даному описі. У результаті одержували наступну білкову послідовність, що секретується: tlcgaelvdalqfvcgdrgfyfnkptgygsssraapqtgivdeccfrscdlrrlemycaplkpaksavraqrhtdmpktqkevhlknasrgs agnknyrm (SEQ ID NO:8) Клітини лінії Cos7 (отримані з АТСС) підтримували в середовищі DMEM, що містить 10% фетальної бичачої сироватки, 100 од. /мл пеніциліну та 100 мкг/мл стрептоміцину, та висівали із 6 щільністю 110 клітин на 10-сантиметровий планшет. Ці клітинні культури трансфектували 8 мкг експресійної плазміди за допомогою Fugene (фірма Roche) відповідно до інструкцій виробника. Через 24 години після трансфекції клітини одноразово промивали та культивували в безсироватковому середовищі протягом 48 годин. Супернатанти збирали та зберігали при 80°С. Для оцінки стабільності поліпептиду в людській сироватці супернатанти, зібрані після культивування клітин лінії Cos7, трансфектованих hIGF-1Ea дикого типу (wt) та hIGF-1Ea3mut, інкубували протягом 16 годин при 37°С або в присутності 10% людської сироватки (фірма Sigma), або без неї. Зразки аналізували за допомогою 18% ДСН-ПААГ та здійснювали імуноблоттинг, використовуючи козяче поліклональне антитіло до людського IGF-1. Результати, представлені на фіг. 1, свідчать про те, що hIGF-1Ea wt у значній мірі розщеплювався після інкубації із сироваткою протягом 16 годин, a hlGF-1Ea3mut виявився стабільним. Денситометричний аналіз показав, що співвідношення нерозщепленого до розщепленого IGF-1 становило приблизно 1:6,2, а у випадку hIGF-1Ea3mut зазначене співвідношення становило приблизно 1:0,68, що свідчать про те, що зазначені мутації привели до одержання стабілізованого поліпептиду. Для підтвердження того, що hIGF-1Ea3mut мав здатність передавати сигнал через IGF-1R, оцінювали фосфорилювання АКТ у клітинах при контакті з поліпептидом. Клітини лінії С2С12 одержували з АТСС та підтримували у модифікованому за способом Дульбеко середовищі Ігла (DMEM) з високим вмістом глюкози (фірма Invitrogen), що містить 10% фетальної бичачої сироватки (фірма AMIMED), 100 од. /мл пеніішліну(фірма Invitrogen), 100 мкг/мл стрептоміцину (фірма Invitrogen) та 2 мМ глутаміну (фірма Invitrogen). Для аналізу фосфорилювання АКТ 6 клітини лінії С2С12 висівали із щільністю 0,1510 клітин на лунку 6-лункового планшета та культивували в середовищі для росту протягом 72 годин. Клітини витримували протягом 4 годин в умовах «голодування» у безсироватковому середовищі та потім стимулювали різними лігандами при 37C протягом 30 хвилин. Клітини лізували за допомогою буфера PhosphoSafe (фірма Cell Signaling), що містить різні інгібітори протеаз, та освітляли центрифугуванням при 14000g протягом 15 хвил. при 4С. Фосфорилювання АКТ та загальні рівні АКТ аналізували за допомогою ELISA з використанням набору для двовалентного ELISA PathScan phospho АКТ (Ser473) та набору для двовалентного ELISA PathScan АКТ (фірма Cell Signaling), відповідно. Дані про фосфорилювання АКТ узагальнені на фіг. 2А, з якої видно, що hІGF-1Ea3mut мав здатність активувати клітинний шлях IGF-1R аналогічно застосовуваному як позитивний контроль реагенту long-R3-IGF-1 та рекомбінантному IGF-1. Крім того, дані, представлені на фіг. 5, переконливо свідчать про те, що hIGF-1Ea3mut приводить до фосфорилювання АКТ. Далі для гарантії того, що рецепторна специфічність hIGF-1Ea3mut у відношенні IGF-1R зберігалася, додавали різні ліганди в культури NІН3Т3-клітин, що надекпресують або IGF-1R, або інсуліновий рецептор (Ins). Ці клітини культивували в таких же умовах, що і описані вище для Соs7-клітин, Для аналізу фосфорилювання IGF-1R та Ins клітини ліній NIH3T3-IGF1R та 6 NIH3T3-Ins висівали із щільністю 0,2х10 клітин на лунку 6-лункового планшету та культивували в середовищі для росту протягом 24 годин. Клітини витримували протягом 16 годин в умовах «голодування» у безсироватковому середовищі та потім стимулювали різними лігандами при 37°С протягом 10 хвилин. Клітини лізували відповідно до процедури, описаної в експерименті з використанням АКТ, та рівні фосфорилювання IGF-1R та Ins аналізували за допомогою ELISA, використовуючи набір DuoSet IC human phosphor-IGF1R та -Ins ELISA (фірма R&D Systems). Результати, які узагальнені на фіг. 3А та 3Б, свідчать про те, що зазначений поліпептидпопередник IGF-1 зберігає специфічність відносно рецептору IGF-1 та може зв'язуватися із спорідненим інсуліновим рецептором з більш низькою афінністю. Приклад 2 Конструювали експресійнкй ДНК- вектор, що кодує поліпептид-попередник hlGF-1-Еb, що містить наступні мутації: делеція G1, делеція Р2 та делеція Е3; мутація, що приводить до заміни 14 UA 97953 C2 5 10 15 20 25 30 35 40 45 50 55 R37 на А; та делеція R71 та делеція S72 (тобто «3mut»). У результаті одержували наступну білкову послідовність, яка секретується: tlcgaelvdalqfvcgdrgfyfnkptgygsssarapqtgivdeccfrscdlrrlemycaplkpaksavraqrhtdmpktqkyqppstnkntk sqrrkgwpkthpggeqkegteaslqirgkkkeqrreigsrnaecrgkkgk (SEQ ID NO:9) Поліпептид аналізували за допомогою процедур, описаних вище в прикладі 1. Дані, представлені на фіг. 1Б, та отримані за допомогою денситометри демонструють, що співвідношення нерозшешіеного до розщепленого IGF-1 становило приблизно 1.9, у той час як у випадку hIGF-1Eb3mut зазначене відношення становило приблизно 1:1, що свідчить про те, що зазначені модифікації приводили до одержання стабілізованого поліпептиду. Дані, представлені на фіг. 2Б, свідчать про те, що hIGF-1Eb3mut мав здатність активувати клітинний шлях IGF-1R аналогічно застосовуваному як позитивний контроль реагенту long-R3-IGF-1 та рекомбінантному IGF-1. Крім того, дані, представлені на фіг. 5, переконливо свідчать про те, що hIGF-1Eb3mut приводив до фосфорилювання АКТ. Результати, узагальнені на фіг. 3В і 3Г, свідчать, що зазначений поліпептид-попередник IGF-1 зберігає специфічність відносно рецептору IGF-1 та може зв'язуватися із спорідненим інсуліновим рецептором з більш низькою афінністю. Приклад 3 Конструювали експресійний ДНК-вектор, що кодує пояшентид-попередник hIGF-1-Ес, що містить наступні мутації: делеція G1, делеція Р2 та делеція Е3; мутація, що приводить до заміни R37 на А; та делеція R71 та делеція S72 (тобто «3mut»). У результаті одержували наступну білкову послідовність, що секретується: tlcgaelvdalqfvcgdrgfyfnkptgygsssrapqtgivdeccfrscdlrrlemycaplkpaksavraqrhtdmpktqkyqppstnkntks qrrkgstfeerk (SEQ ID NO: 10) Поліпептид аналізували за допомогою процедур, описаних вище в прикладі 1. Дані, представлені на фіг. ЇВ, та отримані за допомогою денситометрії демонструють, що співвідношення нерозщепленого до розщепленого IGF-1 становило приблизно 1:5, у той час як у випадку hIGF-1Ec3mut зазначене відношення становило приблизно 1:0,96, що свідчить про те, що зазначені модифікації приводили до одержання стабілізованого поліпептиду. Дані, представлені на фіг. 2Г, свідчать про те, що hIGF-1Ec3mut мав здатність активувати клітинний шлях IGF-1R аналогічно застосовуваному як позитивний контроль реагенту long-R3-IGF-1 та рекомбінантному IGF-1. Крім того, дані, представлені на фіг. 5, переконливо свідчать про те, що hIGF-1Ec3mut приводив до фосфорилювання АКТ. Результати, узагальнені на фіг. 4А і 4Б, свідчать про те, що зазначений поліпептид-попередник IGF-1 зберігає специфічність відносно рецептору IGF-1 та може зв'язуватися з спорідненим інсуліновим рецептором з більш низькою афінністю. Приклад 4 Конструювали експресійний ДНК-вектор, що кодує поліпептид-попередник WGF-1-Eab, що містить наступні мутації: делеція G1, делеція Р2 та делеція Е3; мутація, що приводить до заміни R37 на А; та делеція R71 та делеція S72 (тобто «3mut»); та вставку амінокислот Еа-пептиду 93102 між амінокислотами 95 та 96 Еb-пептиду. Вставка створює можливий N-зв'язаний сигнал глікозилювання N92. У результаті одержували наступну білкову послідовність, що секретується: tlcgaelvdalqfvcgdrgfyfnkptgygsssraapqtgivdeccfrscdlrrlemycaplkpaksavraqrhtdmpktqkyqppstnkna srgsagnkntksqrrkgwpkthpggeqkegteaslqirgkkkeqrreigsrnaecrgkkgk (SEQ ID NO:11) Поліпептид аналізували за допомогою процедур, описаних вище в прикладі 1. Дані, представлені на фіг. 2В, свідчать про те, що hIGF-1Eab3mut мав здатність активувати клітинний шлях IGF-1R аналогічно застосовуваному як позитивний контроль реагенту long-R3-IGF-1 та рекомбінантному IGF-1. Результати, узагальнені на фіг. 4В и 4Г, свідчать про те, що зазначений поліпептид-попередник IGF-1 зберігає специфічність відносно рецептору IGF-1 та не активує інсуліновий рецептор. Приклад 5 Конструювали експресійний ДНК-вектор, що кодує мультимерний поліпептид-попередник hIGF-1-Eb, що містить наступні мутації: делеція G1, делеція Р2, делеція Е3, делеція R36, делеція R37, делеція R71, делеція S72, делеція останніх семи С-кінцевих амінокислот Еb; та вставка на С-кінець зазначеного попередника двох додаткових Еb-пептидів, які обидва не мали R71 та S72 та останніх семи С-кінцевих амінокислот, та четвертого та останнього Еb-пептиду, який не мав R71 та S72. На фіг. 6А представлене схематичне зображення зазначеної конструкції. У результаті одержували наступну білкову послідовність, що секретується: tlcgaelvdalqfvcgdrgfyfnkptgygsssapqtgivdeccfrscdlrrlemycaplkpaksavraqrhtdmpktqkyqppstnkntksq rrkgwpkthpggeqkegteaslqirgkkkeqrreigsrnaersvraqrhtdmpktqkyqppstnkntksqrrkgwpkthpggeqkegteaslq 15 UA 97953 C2 5 10 15 20 25 30 35 40 45 50 55 60 irgkkkeqrreigsrnaersvraqrhtdmpktqkyqppstnkntksqrrkgwpkthpggeqkegteaslqirgkkkeqrreigsrnaersvraqr htdmpktqkyqppstnkntksqrrkgwpkthpggeqkegteaslqirgkkkeqrreigsrnaecrgkkgk (SEQ ID NO: 12) Цей поліпептид оцінювали за допомогою аналізу фосфорилювання АКТ, що описаний у прикладі 1. На фіг. 5 показано, що зазначений мультимер hIGF-1-Eb має здатність здійснювати передачу сигналу через IGF-lR-шлях. Приклад 6 Можна експресувати поліпептид-попередник hIGF-1-Eb, запропонований у винаході, що схематично зображений на фіг. 6Б. Ця конструкція містила наступні модифікації: делеція G1, делеція Р2, делеція Е3, делеція R36, делеція R37, делеція R71, делеція S72; та інсерція амінокислот 93-102 Еа між амінокислотами 95 та 96 Еb, що дозволяло створювати N-зв'язаний сайт глікозилювання в положенні N92 та N100. У результаті одержували наступну білкову послідовність, що секретується: tlcgaelvdalqfvcgdrgfyfnkptgygsssapqtgivdeccfrscdlrrlemycaplkpaksavraqrhtdmpktqkyqppstnknasr gsagnkntksqrrkgwpkthpggeqkegteaslqirgkkkeqrreigsrnaecrgkkgk (SEQ ID NO: 13) Можна експресувати поліпептид-попередник hIGF-2-E, запропонований у винаході, що містив наступні модифікації: делеція Р4, делеція S5 та делеція Е6; мутація, що приводить до заміни R38 на А; та делеція R68 та делеція D69. У результаті одержували наступну білкову послідовність, що секретується: ayrtlcggelvdtlqfvcgdrgfyfsφasrvsrasrgiveeccfrscdlalletycatpaksevstpptvlpdnfprypvgkffqydtwkqstqrlr rglpallrarrghvlakeleafreakrhφlialptqdpahggappemasnrk (SEQ ID NO: 14) Приклад 8 Можна експресувати поліпептид-попередник hIGF-1-Ea, запропонований у винаході, що містив наступні мутації: делеція G1 та делеція Р2; мутація, щоприводить до заміни Е3 на X, де X позначає не амінокислоту, що пегілюють, яка зустрічається в природних умовах; мутація, що приводить до заміни R37 на А; та делеція R71 та делеція S72. У результаті одержували наступну білкову послідовність, що секретується: Xtlcgaelvdalqfvcgdrgfyfnkptgygsssraapqtgivdeccfrscdlrrlemycaplkpaksavraqrhtdmpktqkevhlknasrg sagnknyrm (SEQ ID NO: 15) Приклади 9-78 ( = делеція) 9) hIGF-1-Ea: ΔG1, ΔP2, ΔE3; R36A; ΔR71 tlcgaelvdalqfvcgdrgfyfnkptgygsssarapqtgivdeccfrscdlrrlemycaplkpaksasvraqrhtdmpktqkevhlknas rgsagnknyrm (SEQ ID NO: 16) 10) WGF-I-Ea: ΔG1, ΔP2, ΔE3; R36A; ΔS72 tlcgaelvdalqfvcgdrgfyfnkptgygsssarapqtgivdeccfrscdlrrlemycaplkpaksarvraqrhtdmpktqkevhlknas rgsagnknyrm (SEQ ID NO: 17) 10) WGF-I-Ea: ΔG1, ΔP2, ΔE3; R36A; ΔR71, ΔS72 tlcgaelvdalqfvcgdrgfyfnkptgygsssarapqtgivdeccfrscdlrrlemycaplkpaksavraqrhtdmpktqkevhlknasr gsagnknyrm (SEQ ID NO: 18) 11) hIGF-1-Ea: ΔG1, ΔP2, ΔE3; R37A; ΔR71 tlcgaelvdalqfVcgdrgfyfnkptgygsssraapqtgivdeccfrscdlrrlemycaplkpaksasvraqrhtdmpktqkevhlknas rgsagnknyrm (SEQ ID NO: 19) 12) hIGF-1-Ea: ΔG1, ΔP2, ΔE3; R37A; ΔS72 tlcgaelvdalqfvcgdrgfyfnkptgygsssraapqtgivdeccfrscdlrrlemycaplkpaksarvraqrhtdmpktqkevhlknas rgsagnknyrm (SEQ ID NO:20) 13) hIGF-1-Ea: ΔG1, ΔP2, ΔE3, ΔR37; ΔR71 tlcgaelvdalqfVcgdrgfyfnkptgygsssrapqtgivdeccfrscdlrrlemycaplkpaksasvraqrhtdmpktqkevhlknasr gsagnknyrm (SEQ ID N0:21) 14) hIGF-1-Ea: ΔG1, ΔP2, ΔE3, ΔR37; ΔS72 tlcgaelvdalqfvcgdrgfyfnkptgygsssrapqtgivdeccfrscdlrrlemycaplkpaksarvraqrhtdmpktqkevhlknasr gsagnknyrm (SEQ ID NO:22) 15) hIGF-1-Ea: ΔG1, ΔP2, ΔE3; ΔR37; ΔR71, ΔS72 tlcgaelvdalqfvcgdrgfyfnkptgygsssrapqtgivdeccfrscdlrrlemycaplkpaksavraqrhtdmpktqkevhlknasrg sagnknyrm (SEQ ID NO:23) 16) hIGF-1-Eb: ΔG1, ΔP2, ΔE3; R36A; ΔR71 tlcgaelvdalqfvcgdrgfyfnkptgygsssarapqtgivdeccfrscdlrrlemycaplkpaksasvraqrhtdmpktqkyqppstn kntksqrrkgwpkthpggeqkegteaslqirgkkkeqrreigsrnaecrgkkgk (SEQ ID NO:24) 17) hIGF-1-Eb: ΔG1, ΔP2, ΔE3; R36A; ΔS72 tlcgaelvdalqfvcgdrgfyfnkptgygsssarapqtgivdeccfrscdlrrlemycaplkpaksarvraqrhtdmpktqkyqppstn kntksqrrkgwpkthpggeqkegteaslqirgkkkeqrreigsmaecrgkkgk (SEQ ID NO:25) 18) hIGF-1-Eb: ΔG1, ΔP2, ΔE3; R37A; ΔR71 16 UA 97953 C2 5 10 15 20 25 30 35 40 45 50 55 60 tlcgaelvdalqfVcgdrgfyfnkptgygsssraapqtgivdeccfrscdlrrlemycaplkpaksasvraqrhtdmpktqkyqppstn kntksqrrkgwpkthpggeqkegteaslqirgkkkeqrreigsmaecrgkkgk (SEQ ID NO:26) 19) hIGF-1-Eb: ΔG1, ΔP2, ΔE3; R37A; ΔS72 tlcgaelvdalqfvcgdrgfyfhkptgygsssraapqtgivdeccfrscdlrrlemycaplkpaksarvraqrhtdmpktqkyqppstn kntksqrrkgwpkthpggeqkegteaslqirgkkkeqrreigsrnaecrgkkgk (SEQ ID NO:27) 20) hIGF-1-Eb: ΔG1, ΔP2, ΔE3; R37A; ΔR71, ΔS72 tlcgaelvdalqfVcgdrgfyfnkptgygsssraapqtgivdeccfrscdlrrlemycaplkpaksavraqrhtdmpktqkyqppstnk ntksqrrkgwpkthpggeqkegteaslqirgkkkeqrreigsrnaecrgkkgk (SEQ ID NO:28) 21) hIGF-1-Eb: ΔG1, ΔP2, ΔE3, ΔR37; ΔR71 tlcgaelvdalqfvcgdrgfyfnkptgygsssrapqtgivdeccfrscdlrrlemycaplkpaksasvraqrhtdmpktqkyqppstnk ntksqrrkgwpkthpggeqkegteaslqirgkkkeqrreigsrnaecrgkkgk (SEQ ID NO:29) 22) hIGF-1-Eb: ΔG1, ΔP2, ΔE3, ΔR37; ΔS72 tlcgaelvdalqfvcgdrgfyfnkptgygsssrapqtgivdeccfrscdlrrlemycaplkpaksarvraqrhtdmpktqkyqppstnk ntksqrrkgwpkthpggeqkegteaslqirgkkkeqrreigsrnaecrgkkgk (SEQ ID NO:30) 23) hIGF-1-Eb: ΔG1, ΔP2, ΔE3, ΔR37; ΔR71, ΔS72 tlcgaelvdalqfvcgdrgfyfnkptgygsssrapqtgivdeccfrscdlrrlemycaplkpaksavraqrhtdmpktqkyqppstnkn tksqrrkgwpkthpggeqkegteaslqirgkkkeqrreigsrnaecrgkkgk (SEQ ID NO:31) 24) hIGF-1-Ec: ΔG1, ΔP2, ΔE3; R36A; ΔR71 tlcgaelvdalqfVcgdrgfyfnkptgygsssarapqtgivdeccfrscdlrrlemycaplkpaksasvraqrhtdmpktqkyqppstn kntksqrrkgstfeerk (SEQ ID NO:32) 25) hIGF-1-Ec: ΔG1, ΔP2, ΔE3; R36A; ΔS72 tlcgaelvdalqfvcgdrgfyfnkptgygsssarapqtgivdeccfrscdlrrlemycaplkpaksarvraqrhtdmpktqkyqppstn kntksqrrkgstfeerk (SEQ ID NO:33) 26) hIGF-1-Ec: ΔG1, ΔP2, ΔE3; R36A; ΔR71, ΔS72 tlcgaelvdalqfvcgdrgfyfnkptgygsssarapqtgivdeccfrscdlrrlemycaplkpaksavraqrhtdmpktqkyqppstnk ntksqrrkgstfeerk (SEQ ID NO:34) 27) hIGF-1-Ec: ΔG1, ΔP2, ΔE2; R37A; ΔR71 tlcgaelvdalqfvcgdrgfyfnkptgygsssraapqtgivdeccfrscdlrrlemycaplkpaksasvraqrhtdmpktqkyqppstn kntksqrrkgstfeerk (SEQ ID NO:35) 28) hIGF-1-Ec: ΔG1, ΔP2, ΔE3; R37A; ΔS72 tlcgaelvdalqfVcgdrgfyfnkptgygsssraapqtgivdeccfrscdlrrlemycaplkpaksarvraqrhtdmpktqkyqppstn kntksqrrkgstfeerk (SEQ ID NO:36) 29) hIGF-1-Ec: ΔG1, ΔP2, ΔE3; R37A; ΔR71, ΔS72 tlcgaelvdalqfVcgdrgfyfnkptgygsssraapqtgivdeccfrscdlrrlemycaplkpaksavraqrhtdmpktqkyqppstnk ntksqrrkgstfeerk (SEQ ID NO:37) 30) hIGF-1-Ec: ΔG1, ΔP2, ΔE3, ΔR37, ΔR71 tlcgaelvdalqfvcgdrgfyfnkptgygsssrapqtgivdeccfrscdlrrlemycaplkpaksasvraqrhtdmpktqkyqppstnk ntksqrrkgstfeerk (SEQ ID NO:38) 31) hIGF-1-Ec: ΔG1, ΔP2, ΔE3, ΔR37, ΔS72 tlcgaelvdalqfvcgdrgfyfnkptgygsssrapqtgivdeccfrscdlrrlemycaplkpaksarvraqrhtdmpktqkyqppstnk ntksqrrkgstfeerk (SEQ ID NO:39) 32) hIGF-1-Eab: ΔG1, ΔP2, ΔE3; R36A; ΔR71; інерція ак 93-102 Еа між 95 and 96 of Eb (тобто "Eab") tlcgaelvdalqfvcgdrgfyfnkptgygsssarapqtgivdeccfrscdlrrlemycaplkpaksasvraqrhtdmpktqkyqppstn knasrgsagnkntksqrrkgwpkthpggeqkegteaslqirgkkkeqrreigsrnaecrgkkgk (SEQ ID NO:40) 33) hIGF-1-Eab: ΔG1, ΔP2, ΔE3; R37A; ΔR71 tlcgaelvdalqfvcgdrgfyfnkptgygsssraapqtgivdeccfrscdlrrlemycaplkpaksasvraqrhtdmpktqkyqppstn knasrgsagnkntksqrrkgwpkthpggeqkegteaslqirgkkkeqrreigsrnaecrgkkgk (SEQ ID NO:41) 34) hIGF-1-Eab: ΔG1, ΔP2, ΔE3, ΔR37, ΔR71 tlcgaelvdalqfvcgdrgfyfnkptgygsssrapqtgivdeccfrscdlrrlemycaplkpaksasvraqrhtdmpktqkyqppstnk nasrgsagnkntksqrrkgwpkthpggeqkegteaslqirgkkkeqrreigsrnaecrgkkgk (SEQ ID NO:42) 35) hlGF-1-Eab: ΔG1, ΔP2, ΔE3; R36A; ΔS72 tlcgaelvdalqfvcgdrgfyfnkptgygsssarapqtgivdeccfrscdlrrlemycaplkpaksarvraqrhtdmpktqkyqppstn knasrgsagnkntksqrrkgwpkthpggeqkegteaslqirgkkkeqrreigsrnaecrgkkgk (SEQ ID NO:43) 36) hIGF-1-Eab: ΔG1, ΔP2, ΔE3; R37A; ΔS72 tlcgaelvdalqfvcgdrgfyfnkptgygsssraapqtgivdeccfrscdlrrlemycaplkpaksarvraqrhtdmpktqkyqppstn knasrgsagnkntksqrrkgwpkthpggeqkegteaslqirgkkkeqrreigsrnaecrgkkgk (SEQ ID NO:44) 37) hIGF-1-Eab: ΔG1, ΔP2, ΔE3, ΔR37, ΔS72 tlcgaelvdalqfvcgdrgfyfnkptgygsssrapqtgivdeccfrscdlrrlemycaplkpaksarvraqrhtdmpktqkyqppstnk nasrgsagnkntksqrrkgwpkthpggeqkegteaslqirgkkkeqrreigsrnaecrgkkgk (SEQ ID NO:45) 17 UA 97953 C2 5 10 15 20 25 30 35 40 45 50 55 60 38) hIGF-1-Eab: ΔG1, ΔP2, ΔE3; R36A; ΔR71, ΔS72 tlcgaelvdalqfvcgdrgfyfnkptgygsssarapqtgivdeccfrscdlrrlemycaplkpaksavraqrhtdmpktqkyqppstnk nasrgsagnkntksqrrkgwpkthpggeqkegteaslqirgkkkeqrreigsrnaecrgkkgk (SEQ ID NO:46) 39) hIGF-1-Eab: ΔG1, ΔP2, ΔE3, ΔR37, ΔR71, ΔS72 tlcgaelvdalqfvcgdrgfyfnkptgygsssrapqtgivdeccfrscdlrrlemycaplkpaksavraqrhtdmpktqkyqppstnkn asrgsagnkntksqrrkgwpkthpggeqkegteaslqirgkkkeqrreigsrnaecrgkkgk (SEQ ID NO:47) 40) hIGF-1-Ea: ΔP2, ΔE3; R37A; ΔR71, ΔS72 gtlcgaelvdalqfvcgdrgfyfnkptgygsssraapqtgivdeccfrscdlrrlemycaplkpaksavraqrhtdmpktqkevhlkna srgsagnknyrm (SEQ ID NO:48) 41) hIGF-1-Eb: ΔP2, ΔE3; R37A; ΔR71, ΔS72 gtlcgaelvdalqfvcgdrgfyfnkptgygsssraapqtgivdeccfrscdlrrlemycaplkpaksavraqrhtdmpktqkyqppstn kntksqrrkgwpkthpggeqkegteaslqirgkkkeqrreigsrnaecrgkkgk (SEQ ID NO:49) 42) Мультимер hlGF-1-Eb: (ΔG1, ΔP2, ΔE3; R37A)-3xEb(ΔR71, ΔS72, ΔC-конц 7 aк)-Eb(ΔR71, ΔS72) tlcgaelvdalqfvcgdrgfyfnkptgygsssraapqtgivdeccfrscdlrrlemycaplkpaksavraqrhtdmpktqkyqppstnk ntksqrrkgwpkthpggeqkegteaslqirgkkkeqrreigsmaevraqrhtdmpktqkyqppstnkntksqrrkgwpkthpggeq kegteaslqirgkkkeqrreigsrnaevraqrhtdmpktqkyqppstnkntksqrrkgwpkthpggeqkegteaslqirgkkkeqrrei gsmaevraqrhtdmpktqkyqppstnkntksqrrkgwpkthpggeqkegteaslqirgkkkeqrreigsrnaecrgkkgk (SEQ ID NO:50) 43) Мультимер hIGF-1-Ebr: (ΔP2, ΔE3; R37A)-3xEb(ΔR71, ΔS72, ΔC-конц 7 aк)-Eb(ΔR71, ΔS72) gtlcgaelvdalqfvcgdrgfyfnkptgygsssraapqtgivdeccfrscdlrrlemycaplkpaksavraqrhtdmpktqkyqppstn kntksqrrkgwpkthpggeqkegteaslqirgkkkeqrreigsrnaevraqrhtdmpktqkyqppstnkntksqrrkgwpkthpgge qkegteaslqirgkkkeqrreigsrnaevraqrhtdmpktqkyqppstnkntksqrrkgwpkthpggeqkegteaslqirgkkkeqrr eigsrnaevraqrhtdmpktqkyqppstnkntksqrrkgwpkthpggeqkegteaslqirgkkkeqrreigsrnaecrgkkgk (SEQ ID NO:51) 44) hIGF-1-Ec: ΔP2, ΔE3; R37A; ΔR71, ΔS72 gtlcgaelvdalqfvcgdrgfyfhkptgygsssraapqtgivdeccfrscdlrrlemycaplkpaksavraqrhtdmpktqkyqppstn kntksqrrkgstfeerk (SEQ ID NO:52) 45) hIGF-1-Ea: ΔE3; R37A; ΔR71, ΔS72 gptlcgaelvdalqfvcgdrgfyfnkptgygsssraapqtgivdeccfrscdlrrlemycaplkpaksavraqrhtdmpktqkevhlkn asrgsagnknyrm (SEQ ID NO:53) 46) hIGF-1-Eb: ΔE3; R37A; ΔR71, ΔS72 gptlcgaelvdalqfVcgdrgfyfnkptgygsssraapqtgivdeccfrscdlrrlemycaplkpaksavraqrhtdmpktqkyqppst nkntksqrrkgwpkthpggeqkegteaslqirgkkkeqrreigsrnaecrgkkgk (SEQ ID NO:54) 47) Мультимер hIGF-1-Eb: (ΔG1, ΔP2, ΔE3; R37A)-3xEb(ΔR71, ΔS72, ΔC-конц 7 aк)-Eb(ΔR71, ΔS72) tlcgaelvdalqfvcgdrgfyfnkptgygsssraapqtgivdeccfrscdlrrlemycaplkpaksavraqrhtdmpktqkyqppstnk ntksqrrkgwpkthpggeqkegteaslqirgkkkeqrreigsrnaevraqrhtdmpktqkyqppstnkntksqrrkgwpkthpggeq kegteaslqirgkkkeqrreigsrnaevraqrhtdmpktqkyqppstnkntksqrrkgwpkthpggeqkegteaslqirgkkkeqrrei gsrnaevraqrhtdmpktqkyqppstnkntksqrrkgwpkthpggeqkegteaslqirgkkkeqrreigsrnaecrgkkgk (SEQ 1D NO:55) 48) Мультимер hIGF-1-Eb: (ΔE3; R37A)-3xEb(ΔR71, ΔS72, ΔC-конц 7 aк)-Eb(ΔR71, ΔS72) gptlcgaelvdalqfvcgdrgfyfnkptgygsssraapqtgivdeccfrscdlrrlemycaplkpaksavraqrhtdmpktqkyqppst nkntksqrrkgwpkthpggeqkegteaslqirgkkkeqrreigsrnaevraqrhtdmpktqkyqppstnkntksqrrkgwpkthpg geqkegteaslqirgkkkeqrreigsrnaevraqrhtdmpktqkyqppstnkntksqrrkgwpkthpggeqkegteaslqirgkkkeq rreigsrnaevraqrhtdmpktqkyqppstnkntksqrrkgwpkthpggeqkegteaslqirgkkkeqrreigsrnaecrgkkgk (SEQ ID NO:56) 49) hlGF-1-Ec: ΔE3; R37A; ΔR71, ΔS72 gptlcgaelvdalqfVcgdrgfyfnkptgygsssraapqtgivdeccfrscdlrrlemycaplkpaksavraqrhtdmpktqkyqppst nkntksqrrkgstfeerk (SEQ ID NO:57) 50) hIGF-1-Ea: ΔE3; R37A; ΔR71, ΔS72 gptlcgaelvdalqfvcgdrgfyfnkptgygsssraapqtgivdeccfrscdlrrlemycaplkpaksavraqrhtdmpktqkevhlkn asrgsagnknyrm (SEQ ID NO:58) 51) hIGF-1-Eb: ΔE3; R37A; ΔR71, ΔS72 gptlcgaelvdalqfvcgdrgfyfnkptgygsssraapqtgivdeccfrscdlrrlemycaplkpaksavraqrhtdmpktqkyqppst nkntksqrrkgwpkthpggeqkegteaslqirgkkkeqrreigsrnaecrgkkgk (SEQ ID NO:59) 52) hIGF-1-Ec: ΔE3; R37A; ΔR71, ΔS72 gptlcgaelvdalqfvcgdrgfyfnkptgygsssraapqtgivdeccfrscdlrrlemycaplkpaksavraqrhtdmpktqkyqppst nkntksqrrkgstfeerk (SEQ ID NO:60) 18 UA 97953 C2 5 10 15 20 25 30 35 40 45 50 55 53) hIGF-1-Eab: ΔE3; R37A; Δ R71, ΔS72 gptlcgaelvdalqfvcgdrgfyfnkptgygsssraapqtgivdeccfrscdlrrlemycaplkpaksavraqrhtdmpktqkyqppst nknasrgsagnkntksqrrkgwpkthpggeqkegteaslqirgkkkeqrreigsrnaecrgkkgk (SEQ ID NO:61) 54) Мультимер hIGF-1-Eb: (ΔE3; R37A)-3xEb(ΔR71, ΔS72, ΔC-конц. 7 aк)-Eb(ΔR71, ΔS72) gptlcgaelvdalqfVcgdrgfyfnkptgygsssraapqtgivdeccfrscdlrrlemycaplkpaksavraqrhtdmpktqkyqppst nkntksqrrkgwpkthpggeqkegteaslqirgkkkeqrreigsrnaevraqrhtdmpktqkyqppstnkntksqrrkgwpkthpg geqkegteaslqirgkkkeqrreigsrnaevraqrhtdmpktqkyqppstnkntksqrrkgwpkthpggeqkegteaslqirgkkkeq rreigsrnaevraqrhtdmpktqkyqppstnkntksqrrkgwpkthpggeqkegteaslqirgkkkeqrreigsmaecrgkkgk (SEQ ID NO:62) 55) hIGF-1-Ea: E3A; R37A; ΔR71, ΔS72 gpatlcgaelvdalqfvcgdrgfyfnkptgygsssraapqtgivdeccfrscdlrrlemycaplkpaksavraqrhtdmpktqkevhlk nasrgsagnknyrm (SEQ ID NO:63) 56) hIGF-1-Eb: E3A; R37A; ΔR71, ΔS72 gpatlcgaelvdalqfvcgdrgfyfnkptgygsssraapqtgivdeccfrscdlrrlemycaplkpaksavraqrhtdmpktqkyqpps tnkntksqrrkgwpkthpggeqkegteaslqirgkkkeqrreigsrnaecrgkkgk (SEQ ID NO:64) 57) hIGF-1-Ec: E3A; R37A; ΔR71, ΔS72 gpatlcgaelvdalqfvcgdrgfyfnkptgygsssraapqtgivdeccfrscdlrrlemycaplkpaksavraqrhtdmpktqkyqpps tnkntksqrrkgstfeerk (SEQ ID NO:65) 58) hIGF-1-Eab: E3A; R37A; ΔR71, ΔS72 gpatlcgaelvdalqfvcgdrgfyfnkptgygsssraapqtgivdeccfrscdlrrlemycaplkpaksavraqrhtdmpktqkyqpps tnknasrgsagnkntksqrrkgwpkthpggeqkegteaslqirgkkkeqrreigsrnaecrgkkgk (SEQ ID NO:66) 59) Мультимер hIGF-1-Eb: (E3A; R37A)-3xEb(ΔR71, ΔS72, ΔC-конц. 7 aк)-Eb(ΔR71, ΔS72) gpatlcgaelvdalqfvcgdrgfyfnkptgygsssraapqtgivdeccfrscdlrrlemycaplkpaksavraqrhtdmpktqkyqpps tnkntksqrrkgwpkthpggeqkegteaslqirgkkkeqrreigsrnaevraqrhtdmpktqkyqppstnkntksqrrkgwpkthpg geqkegteaslqirgkkkeqrreigsmaevraqrhtdmpktqkyqppstnkntksqrrkgwpkthpggeqkegteaslqirgkkkeq rreigsrnaevraqrhtdmpktqkyqppstnkntksqrrkgwpkthpggeqkegteaslqirgkkkeqrreigsrnaecrgkkgk (SEQ ID NO:67) 60) hIGF-1-Ea: ΔP2, ΔE3; R37A; ΔR71, ΔS72 gtlcgaelvdalqfvcgdrgfyfnkptgygsssraapqtgivdeccfrscdlrrlemycaplkpaksavraqrhtdmpktqkevhlkna srgsagnknyrm (SEQ ID NO:68) 61) hIGF-1-Eb: ΔP2, ΔE3; R37A; ΔR71, ΔS72 gtlcgaelvdalqfvcgdrgfyfnkptgygsssraapqtgivdeccfrscdlrrlemycaplkpaksavraqrhtdmpktqkyqppstn kntksqrrkgwpkthpggeqkegteaslqirgkkkeqrreigsrnaecrgkkgk (SEQ ID NO:69) 62) hIGF-1-Ec: ΔP2, ΔE3; R37A; ΔR71, ΔS72 gtlcgaelvdalqfvcgdrgfyfnkptgygsssraapqtgivdeccfrscdlrrlemycaplkpaksavraqrhtdmpktqkyqppstn kntksqrrkgstfeerk (SEQ ID NO: 181) 63) hIGF-1-Eab: ΔP2, ΔE3; R37A; ΔR71, ΔS72 gtlcgaelvdalqfvcgdrgfyfhkptgygsssraapqtgivdeccfrscdlrrlemycaplkpaksavraqrhtdmpktqkyqppstn knasrgsagnkntksqrrkgwpkthpggeqkegteaslqirgkkkeqrreigsrnaecrgkkgk (SEQ ID NO:70) 64) Мультимер hlGF-1-Eb: (ΔP2, ΔE3; R37A)-3xEb(ΔR71, ΔS72, ΔC-конц. 7 aк)-Eb(ΔR71,ΔS72) gtlcgaelvdalqfvcgdrgfyfnkptgygsssraapqtgivdeccfrscdlrrlemycaplkpaksavraqrhtdmpktqkyqppstn kntksqrrkgwpkthpggeqkegteaslqirgkkkeqrreigsrnaevraqrhtdmpktqkyqppstnkntksqrrkgwpkthpgge qkegteaslqirgkkkeqrreigsrnaevraqrhtdmpktqkyqppstnkntksqrrkgwpkthpggeqkegteaslqirgkkkeqrr eigsrnaevraqrhtdmpktqkyqppstnkntksqrrkgwpkthpggeqkegteaslqirgkkkeqrreigsrnaecrgkkgk (SEQ ID NO:71) 65) hIGF-1-Eb: ΔG1, ΔP2; E3X; R37A; ΔR71, ΔS72 Xtlcgaelvdalqfvcgdrgfyfnkptgygsssraapqtgivdeccfrscdlrrlemycaplkpaksavraqrhtdmpktqkyqppstn kntksqrrkgwpkthpggeqkegteaslqirgkkkeqrreigsrnaecrgkkgk (SEQ ID NO:72) 66) hIGF-1-Ec: ΔG1, ΔP2; E3X; R37A; ΔR71, ΔS72 Xtlcgaelvdalqfvcgdrgfyfnkptgygsssraapqtgivdeccfrscdlrrlemycaplkpaksavraqrhtdmpktqkyqppstn kntksqrrkgstfeerk (SEQ ID NO:73) 67) hlGF-1-Eab: ΔG1, ΔP2; E3X; R37A; ΔR71, ΔS72 XtlcgaelvdalqfVcgdrgfyfnkptgygsssraapqtgivdeccfrscdlrrlemycaplkpaksavraqrhtdmpktqkyqppstn knasrgsagnkntksqrrkgwpkthpggeqkegteaslqirgkkkeqrreigsrnaecrgkkgk (SEQ ID NO:74) 68) Мультимер hIGF-1-Eb: (ΔG1, ΔP2; E3X; R37A)-3xEb(ΔR71, ΔS72, ΔC-конц. 7 aк)-Eb(ΔR71, ΔS72) XtlcgaelvdalqfVcgdrgfyfnkptgygsssraapqtgivdeccfrscdlrrlemycaplkpaksavraqrhtdmpktqkyqppstn kntksqrrkgwpkthpggeqkegteaslqirgkkkeqrreigsrnaevraqrhtdmpktqkyqppstnkntksqrrkgwpkthpgge qkegteaslqirgkkkeqrreigsrnaevraqrhtdmpktqkyqppstnkntksqrrkgwpkthpggeqkegteaslqirgkkkeqrr 19 UA 97953 C2 5 10 15 20 25 30 35 40 45 50 55 60 eigsmaevraqrhtdmpktqkyqppstnkntksqrrkgwpkthpggeqkegteaslqirgkkkeqrreigsrnaecrgkkgk (SEQ ID NO:75) 69) hIGF-1-Ea: ΔG1, ΔP2, ΔE3; R37A; ΔR71; S72X tlcgaelvdalqfvcgdrgfyfnkptgygsssraapqtgivdeccfrscdlrrlemycaplkpaksaXvraqrhtdmpktqkevhlkna srgsagnknyrm (SEQ ID NO.76) 70) hIGF-1-Eb: ΔG1, ΔP2, ΔE3; R37A; ΔR71; S72X tlcgaelvdalqfVcgdrgfyfnkptgygsssraapqtgivdeccfrscdlrrlemycaplkpaksaXvraqrhtdmpktqkyqppstn kntksqrrkgwpkthpggeqkegteaslqirgkkkeqrreigsrnaecrgkkgk (SEQ ID NO.77) 71) hIGF-1-Ec: ΔG1, ΔP2, ΔE3; R37A; ΔR71; S72X tlcgaelvdalqfVcgdrgfyfnkptgygsssraapqtgivdeccfrscdlrrlemycaplkpaksaXvraqrhtdmpktqkyqppstn kntksqrrkgstfeerk (SEQ ID NO:78) 72) hIGF-1-Eab: ΔG1, ΔP2, ΔE3; R37A; ΔR71; S72X tlcgaelvdalqfvcgdrgfyfhkptgygsssraapqtgivdeccfrscdlrrlemycaplkpaksaXvraqrhtdmpktqkyqppstn knasrgsagnkntksqrrkgwpkthpggeqkegteaslqirgkkkeqrreigsrnaecrgkkgk (SEQ ID NO:79) 73) Мультимер hIGF-1-Eb: (ΔG1, ΔP2, ΔE3; R37A)-Eb(ΔR71; S72X; ΔC-конц. 7 aк)-2xEb(ΔR71, ΔS72, ΔC-конц. 7 aк)-Eb(ΔR71, ΔS72) tlcgaelvdalqfvcgdrgfyfnkptgygsssraapqtgivdeccfrscdlrrlemycaplkpakXsavraqrhtdmpktqkyqppstn kntksqrrkgwpkthpggeqkegteaslqirgkkkeqrreigsrnaevraqrhtdmpktqkyqppstnkntksqrrkgwpkthpgge qkegteaslqirgkkkeqrreigsrnaevraqrhtdmpktqkyqppstnkntksqrrkgwpkthpggeqkegteaslqirgkkkeqrr eigsrnaevraqrhtdmpktqkyqppstnkntksqrrkgwpkthpggeqkegteaslqirgkkkeqrreigsrnaecrgkkgk (SEQ ID NO:80) 74) hIGF-1-Ea: ΔG1, ΔP2, ΔE3; R37A; ΔR71, ΔS72; N92X tlcgaelvdalqfvcgdrgfyfnkptgygsssraapqtgivdeccfrscdlrrlemycaplkpaksavraqrhtdmpktqkevhlkXas rgsagnknyrm (SEQ ID NO:81) 75) hIGF-1-Eb: ΔG1, ΔP2, ΔE3; R37A; ΔR71, ΔS72; C142X tlcgaelvdalqfVcgdrgfyfnkptgygsssraapqtgivdeccfrscdlrrlemycaplkpaksavraqrhtdmpktqkyqppstnk ntksqrrkgwpkthpggeqkegteaslqirgkkkeqrreigsrnaeXrgkkgk (SEQ ID NO:82) 76) hIGF-1-Eab: ΔG1, ΔP2, ΔE3; R37A; ΔR71, ΔS72; C151X tlcgaelvdalqfvcgdrgfyfnkptgygsssraapqtgivdeccfrscdlrrlemycaplkpaksavraqrhtdmpktqkyqppstnk nasrgsagnkntksqrrkgwpkthpggeqkegteaslqirgkkkeqrreigsrnaeXrgkkgk (SEQ ID NO:83) 78) Мультимер hIGF-1-Eb (ΔG1, ΔP2, ΔE3; R37A)-3xEb(ΔR71, ΔS72, ΔC-конц. 7 aк)-Eb(ΔR71, ΔS72; C71X) tlcgaelvdalqfvcgdrgfyfnkptgygsssraapqtgivdeccfrscdlrrlemycaplkpaksavraqrhtdmpktqkyqppstnk ntksqrrkgwpkthpggeqkegteaslqirgkkkeqrreigsrnaevraqrhtdmpktqkyqppstnkntksqrrkgwpkthpggeq kegteaslqirgkkkeqrreigsrnaevraqrhtdmpktqkyqppstnkntksqrrkgwpkthpggeqkegteaslqirgkkkeqrrei gsmaevraqrhtdmpktqkyqppstnkntksqrrkgwpkthpggeqkegteaslqirgkkkeqrreigsmaeXrgkkgk (SEQ ID NO:84) Приклад 79: Аналіз проліферації міобластів Аналіз проліферації міобластів являє собою достовірний індикаторний метод оцінки in vitro активності IGF та його застосовували як модель для факторів, що роблять вплив на ембріональні міобласти та клітини-сателіти дорослих особин. Фактори, що мають активність в цій системі, проявляють свою дію так само, як і у первинних культурах міобластів. Посилення проліферації міобластів in vitro пептидом, запропонованим у даному винаході, свідчить про активність, що підвищує проліферацію міобластів, та внаслідок цього що підвищує зрештою кількість міофібрил у матці. Крім того, аналогічне посилення проліферації міобластів свідчить про те, що пептиди, запропоновані в даному винаході, можна застосовувати для підвищення м'язової гіпертрофії у дорослих осіб, наприклад, шляхом стимуляції клітинної проліферації в м'язі-сателіті. Приклад 80: Аналіз епітеліальної тканини молочної залози У тварини на стадії лактації кількість епітеліальної тканини молочної залози є лімітуючим чинником виробництва молока, оскільки ця тканина містить клітини, які продукують і секретують молоко. При використанні в системах ги vitro епітеліальні клітини, отримані з молочних залоз тварин, можна стимулювати модифікованими IGF-1 або IGF-2, запропонованими в даному винаході, відносно проліферації та виробництва підвищених кількостей компонентів молока. Можна продемонструвати також, що епітеліальні клітини молочних залоз, проліферація яких стимульована в одній з таких систем in vitro, можна повторно імплантувати в очищені скупчення жирової тканини молочної залози та стимулювати проліферацію та/або виробництво молока у тварин на стадії лактації. Приклад 81: Оцінка рівнів IGF-1 або IGF-2 у крові або інших рідинах організму Ефективна кількість введеного парентерально пептиду в дозі можна оцінювати за допомогою кривої 20 UA 97953 C2 5 10 15 20 25 30 залежності доза-відповідь. Наприклад, рівні модифікованих пептидів IGF, запропонованих у винаході, можна визначати в крові або в загальній воді організму індивідуума, що підлягає лікуванню, з метою визначення дозування. В іншому варіанті можна вводити зростаючі кількості пептиду індивідуумові та оцінювати рівні модифікованого IGF-1 та IGF-2 у сироватці індивідуума. Кількість пептиду, яку слід застосовувати, можна розраховувати в молях на основі даних про рівні модифікованого IGF-1 або IGF-2 у сироватці. Одним з методів визначення відповідного дозування пептиду передбачає оцінку пептиду IGF, запропонованого у винаході, у біологічній рідині, такій як загальна вода організму або плазма крові Оцінку зазначених рівнів можна здійснювати за будь-якими методиками, включаючи РІА та ELISA. Після визначення рівнів IGF рідину приводять у контакт із однією або декількома дозами пептиду. Після зазначеної стадії контакту в рідині повторно оцінюють рівні IGF. Якщо рівні IGF у рідині знаходяться нижче кількості, достатньої для досягнення необхідної ефективності, з метою одержання якої молекулу вводять, то дозу молекули можна регулювати для досягнення максимальної ефективності. Цей метод можна здійснювати in vitro або in vivo, Переважно зазначений метод здійснюють in vivo, тобто після екстракції рідини з організму індивідуума та оцінки рівнів IGF пептид вводять ссавцеві у вигляді однієї або декількох доз (це означає, що шляхом введення тварині досягають стадії контакту) та потім рівні IGF повторно оцінюють в екстрагованій рідині, отриманій із тварини. Інший метод визначення дозування передбачає застосування антитіл до пептиду або являє собою інший метод визначення в LIFA-форматі. Приклад 82: Фармакокінетичні характеристики hIGF-1-Ec 3mut in vivo Дорослим самцям мишей (n = 3/групу) вводили шляхом внутрішньовенної (i.v.) болюсної ін'єкції rhIGF-1 у дозі 1 мг/кг та hIGF-1-Ec3mut (описаний у прикладі 3) у дозі 1,55 мг/кг. Брали серійні зразки крові через 5, 15, 30 й 60 хвилин після введення субстанції, яку тестують. Концентрації в сироватці rhIGF-і та hIGF-1-Ec3mut визначали за допомогою ELISA. Цей аналіз є специфічним для hIGF-1. Еквімолярні дози rhIGF-і та hIGF-1-Ec3mut вводили мишам i.v. Результати свідчать про істотно більш високі рівні білку hIGF-1-Ec3mut у порівнянні з rhIGF-і в усі вивчені моменти часу, що демонструє, що hIGF-1-Ec3mut є більш стабільним відносно метаболізму, ніж IGF-1, що складається з 70 амінокислот. Час (хвил.) 5 15 30 60 IGF-l-Ec3mut(HM) 201,4 65,3 12 0,76 IGF-1 (нМ) 54,7 14,3 2,4 0,2 ФОРМУЛА ВИНАХОДУ 35 40 45 50 1. Поліпептид, що включає модифікований людський білок-попередник IGF-1, який містить IGF-1 та пептид Е, у якому залишки G1, P2 та E3 видалені, та амінокислота в положенні 37 має заміну R37А, а також видалені амінокислоти у положеннях 71 і 72. 2. Поліпептид за п. 1, у якому білок-попередник містить Ea-пептид. 3. Поліпептид за п. 1, у якому білок-попередник містить Eb-пептид. 4. Поліпептид за п. 3, у якому останні сім C-кінцевих амінокислот Eb видалені. 5. Поліпептид за п. 1, у якому білок-попередник містить Ec-пептид. 6. Поліпептид за будь-яким з пп. 1-5, який додатково містить NXS/T консенсусну послідовність N-зв'язаного глікозилювання. 7. Поліпептид за п. 3, який додатково містить амінокислоти 93-102 з Ea, вставлені між амінокислотами N95 та T96 з Eb. 8. Поліпептид за будь-яким з пп. 1-7, який додатково містить олігосахарид, ковалентно зв'язаний з бічним ланцюгом амінокислоти білка-попередника. 9. Поліпептид за п. 8, у якому олігосахарид ковалентно зв'язаний з бічним ланцюгом аргініну білка-попередника. 10. Поліпептид за будь-яким з пп. 1-9, у якому залишок білка-попередника замінений на неприродну амінокислоту. 11. Поліпептид за п. 10, у якому неприродна амінокислота містить ацетиленову або азидогрупу. 12. Поліпептид, що містить людський білок-попередник IGF-1 та білок Еа, який представлений в SEQ ID NO: 8. 21 UA 97953 C2 5 10 15 20 13. Поліпептид, що містить людський білок-попередник IGF-1 та білок Еа, який представлений в SEQ ID NO: 53. 14. Поліпептид, що включає модифікований людський білок-попередник IGF-1, який містить IGF1 та пептид Еа, у якому видалені амінокислота Е3 та амінокислоти R71 та S72, а амінокислота R37 мутована на аланін. 15. Поліпептид за будь-яким з пп. 1-14, який додатково включає залишок полі(етиленгліколю), ковалентно приєднаний до бічного ланцюга білка-попередника. 16. Поліпептид за п. 15, у якому залишок полі(етиленгліколю) ковалентно приєднаний до бічного ланцюга білка-попередника за п. 14. 17. Поліпептид за будь-яким з пп. 1-16 для застосування у лікуванні захворювання кістковом'язової системи, діабету, загибелі нейронів, хронічного обструктивного захворювання легенів, опікового ушкодження або анемії. 18. Спосіб лікування захворювання кістково-м'язової системи, діабету, загибелі нейронів, хронічного обструктивного захворювання легенів, опікового ушкодження або анемії, у якому суб'єкту вводять терапевтично ефективну кількість поліпептиду за будь-яким з пп. 1-16. 19. Застосування поліпептиду за будь-яким з пп. 1-16 для виготовлення лікарського засобу для лікування захворювання кістково-м'язової системи, діабету, загибелі нейронів, хронічного обструктивного захворювання легенів, опікового ушкодження або анемії. 20. Нуклеїнова кислота, що кодує поліпептид за будь-яким з пп. 1-16. 21. Вектор, що містить нуклеїнову кислоту за п. 20. 22. Клітина, трансфекована вектором за п. 21. 22 UA 97953 C2 23 UA 97953 C2 24 UA 97953 C2 25 UA 97953 C2 26 UA 97953 C2 27 UA 97953 C2 28
ДивитисяДодаткова інформація
Назва патенту англійськоюStabilized insulin-like growth factor polypeptides
Автори англійськоюGlass David Jonathan, Fornaro Mara
Назва патенту російськоюСтабилизированные полипептиды инсулиноподобного фактора роста
Автори російськоюГласс Девид Джонатан, Форнаро Мара
МПК / Мітки
МПК: A61K 38/18, C07K 14/65, C12N 15/27, A61P 3/10
Мітки: фактора, поліпептиди, росту, інсуліноподібного, стабілізовані
Код посилання
<a href="https://ua.patents.su/40-97953-stabilizovani-polipeptidi-insulinopodibnogo-faktora-rostu.html" target="_blank" rel="follow" title="База патентів України">Стабілізовані поліпептиди інсуліноподібного фактора росту</a>
Моноклональне антитіло людини, яке специфічно зв’язується з рецептором інсуліноподібного фактора росту і (igf-ir)

Номер патенту: 87804
Опубліковано: 25.08.2009
Автори: Кохен Брюс Д., Біб Жан, Мойєр Джеймс Д., Міллер Пенелопа Е., Галло Майкл, Корвалан Хосе Р.
МПК: A61K 39/395, C12N 15/64, C07K 16/18, A61K 49/00, C12N 15/13, C12P 21/08, A61P 35/00, G01N 33/574, C12N 5/20
Мітки: людини, рецептором, зв'язується, інсуліноподібного, яке, росту, igf-ir, антитіло, фактора, моноклональне, специфічно
Формула / Реферат:
1. Людське моноклональне антитіло або його антигензв'язувальна частина, яка специфічно зв'язується з рецептором інсуліноподібного фактора росту I (IGF-IR), де вказане антитіло містить важкий ланцюг і легкий ланцюг, де вказаний легкий ланцюг кодується геном Vκ А30 зародкової лінії людини, і де амінокислотні послідовності CDR1, CDR2 і CDR3 важкого ланцюга та амінокислотні послідовності CDR1, CDR2 і CDR3 легкого ланцюга вибрані з групи, що...
Кон’югати інсуліноподібного фактора росту-1 (іфр-1) і поліетиленгліколю

Номер патенту: 93666
Опубліковано: 10.03.2011
Автори: Шаубмар Андреас, Ланцендьорфер Мартін, Фозер Штефан, Хессе Фрідеріке, Кюнкеле Клаус-Петер, Амреін Беат, Регула Йорг, Ланг Курт, Метцгер Фрідріх
МПК: A61K 38/30, A61K 47/48, A61P 25/28
Мітки: інсуліноподібного, поліетиленгліколю, іфр-1, фактора, кон'югати, росту-1
Формула / Реферат:
1. Кон'югат, що включає варіант інсуліноподібного фактора росту І (ІФР-І) і поліетиленгліколь (ПЕГ), який відрізняється тим, що зазначений варіант ІФР-І містить заміни (заміну) амінокислот у положеннях 27, 37, 65, 68 у послідовності вихідного ІФР-I таким чином, що одна або декілька амінокислот у положеннях 37, 65, 68 є лізином (К), а амінокислота в положенні 27 є полярною амінокислотою, але не лізином, причому зазначений ПЕГ є кон'югованим...
Терапія пухлин з використанням судинного ендотеліального фактора росту і антитіла до рецептора типу 2 людського епітеліального фактора росту

Номер патенту: 95797
Опубліковано: 12.09.2011
Автори: Шойер Вернер, Хасманн Макс, Фрісс Томас
МПК: A61K 39/395, A61P 35/00
Мітки: фактора, судинного, типу, росту, пухлин, ендотеліального, терапія, антитіла, людського, використанням, епітеліального, рецептора
Формула / Реферат:
1. Застосування антитіла до HER2 та антитіла до VEGF для приготування лікарського засобу для лікування раку молочної залози, який характеризується надекспресією білка рецептора НЕR2, у пацієнта, не сприйнятливого до попередньої терапії з використанням антитіла до VEGF, що полягає в тому, що пацієнтові вводять терапевтично ефективну кількість антитіла до HER2 і антитіла до VEGF.2. Застосування за п. 1, в якому антитіло до VEGF є...
Поліпептиди фактора коагуляції vii людини

Номер патенту: 82177
Опубліковано: 25.03.2008
Автори: Перссон Егон, Ольсен Оле Вільстед
МПК: C12P 21/02, C12N 9/64, C12N 15/09, A61K 38/00, C12N 1/15, C12N 5/10, A61P 7/04, A61K 38/46, C12N 1/19, C07K 14/745
Мітки: людини, фактора, поліпептиди, коагуляції
Формула / Реферат:
1. Поліпептид фактора VII, що містить принаймні два заміщення стосовно амінокислотної послідовності SEQ ID NO:1, де вказані заміщення являють собою:(і) заміщення L305 амінокислотою, вибраною з групи, що складається з Val, Туr та Ilе, і (іі) заміщення амінокислоти К337 амінокислотою, вибраною з групи, що складається з Ala, Gly, Val, Ser, Thr, Asn, Gln, Asp та Glu;де співвідношення між активністю вказаного поліпептиду фактора VII...
Антитіла до рецептора інсулінподібного фактора росту i та їх застосування

Номер патенту: 95284
Опубліковано: 25.07.2011
Автори: Ройш Дітмар, Кюнкеле Клаус-Петер, Хансен Зільке, Шумахер Ральф
МПК: C07K 16/28
Мітки: росту, інсулінподібного, рецептора, антитіла, застосування, фактора
Формула / Реферат:
1. Антитіло, що має здатність до зв'язування з IGF-IR, яке є людським антитілом IgG1- або IgG3-типу і є глікозильованим цукровим ланцюгом на залишку Asn297, яке відрізняється тим, що кількість залишків фукози в цукровому ланцюзі складає щонайменше 99 % ("повністю фукозиловане"), і, крім того, кількість залишків NGNA складає 1 % або менше і/або кількість залишків N-кінцевої альфа-1,3-галактози складає 1 % або менше.2. Антитіло...
Попередній патент: Інгаляційний пристрій для сухого порошку для одночасного введення більше ніж одного медикаменту
Наступний патент: Тимчасовий міст
Випадковий патент: Ендопротез сегмента хребта "las"