Спосіб підвищення біосинтезу фосфогліколіпідних антибіотиків
Номер патенту: 63820
Опубліковано: 25.10.2011
Автори: Громико Олександр Миколайович, Осташ Богдан Омелянович, Уокер-Кане С'юзен, Федоренко Віктор Олександрович



Формула / Реферат
Спосіб підвищення біосинтезу фосфогліколіпідних антибіотиків, який базується на надекспресії генів плейотропних регуляторів у актиноміцетах-продуцентах цих антибіотиків, який відрізняється тим, що як регуляторний елемент використовують ген лейцил-тРНКUUА S. lividans, клонований у складі висококопійної плазміди pIJ584 у штамах S. ghanaensis ATCC14672, S. lividans ТК24, S. albus J1074.
Текст
Спосіб підвищення біосинтезу фосфогліколіпідних антибіотиків, який базується на надекспресії генів плейотропних регуляторів у актиноміцетах-продуцентах цих антибіотиків, який відрізняється тим, що як регуляторний елемент UUА використовують ген лейцил-тРНК S. lividans, клонований у складі висококопійної плазміди pIJ584 у штамах S. ghanaensis ATCC14672, S. lividans ТК24, S. albus J1074. (19) (21) u201102624 (22) 09.03.2011 (24) 25.10.2011 (46) 25.10.2011, Бюл.№ 20, 2011 р. (72) ОСТАШ БОГДАН ОМЕЛЯНОВИЧ, ФЕДОРЕНКО ВІКТОР ОЛЕКСАНДРОВИЧ, ГРОМИКО ОЛЕКСАНДР МИКОЛАЙОВИЧ, УОКЕР-КАНЕ С'ЮЗЕН, US (73) ЛЬВІВСЬКИЙ НАЦІОНАЛЬНИЙ УНІВЕРСИТЕТ ІМЕНІ ІВАНА ФРАНКА 3 впливати на трансляцію мРНК генів біосинтезу фосфогліколіпідних антибіотиків (тое-генів). В основу винаходу поставлено задачу удосконалити спосіб підвищення біосинтезу фосфогліколіпідних антибіотиків шляхом UUА надекспресії гена bldA, що кодує лейцил-тРНК S. lividans, на основі реплікону рІЛОІ, що дасть змогу забезпечити велику кількість копій цієї плазміди та підвищити продукції антибіотика. Поставлена задача вирішується так, що у способі підвищення біосинтезу фосфогліколіпідних антибіотиків, який базується на надекспресії регуляторних елементів, де як регуляторний елемент UUА використовують ген (bldA) лейцил-тРНК S. lividans, клонований у складі висококопійної плазміди pIJ584 у штамах S. ghanaensis ATCC 14672, S. lividans TK24, S. albus J1074. Фосфогліколіпідні антибіотики - природні, фосфоровмісні, високомодифіковані глікозиди, що містять ліпідний ланцюг ізопренового походження. У молярному розрахунку, фосфогліколіпіди у 1001000 разів перевищують інші відомі антибіотики за активністю проти Грам-позитивних патогенів [Ostash В., Walker S. Moenomycin family antibiotics: chemical synthesis, biosynthesis, biological activity // Nat Prod Rep. - 2010. - vol. 27. -P. 1594-1617]. Усі відомі на сьогодні фосфогліколіпідні антибіотики виявляють протибактерійну дію за рахунок прямого пригнічення пептидогліканових глікозилтрансфераз - білків, що задіяні у синтезі клітинної стінки (пептидоглікану) бактерій [Ostash В., Doud E., Fedorenko V. The molecular biology of moenomycins: towards novel antibiotics based on inhibition of bacterial peptidoglycan glycosyltransferases // Biol Chem. - 2010. -vol. 391. - P.499-504]. Природні штами-продуценти фосфогліколіпідів накопичують надзвичайно малі кількості антибіотика, що ускладнює його практичне використання та дослідження з метою отримання клінічно цінних похідних. Авторами вперше запропоновано використати ииА ген лейцил-тРНК S. lividans у складі плазміди pIJ584 [Sun J., Hesketh A., Bibb M. Functional analysis of relA and rshA, two relA/spoT homologues of Streptomyces coelicolor A3(2) // J. Bacteriol. - 2001. vol.183. - P.3488-3498] для підвищення біосинтезу фосфогліколіпідних антибіотиків. Запропонований підхід дасть змогу значно підвищити продукцію цієї групи вторинних метаболітів, оскільки наявність рідкісного кодону ТТА у кількох ключових тое генах є одним з факторів, що обмежує їхню експресію, а отже і продукцію фосфогліколіпідів [Ostash В., Saghatelian A., Walker S. A streamlined metabolic pathway for the biosynthesis of moenomycin A // Chem. Biol. - 2007.- vol.14. -P.257-267]. UUА Надекспресія гена лейцил-тРНК , що декодує триплет ТТА, дасть змогу подолати це обмеження. Оскільки гени тРНК є висококонсервативними серед актиноміцетів, їх можна вільно комбінувати між різними видами без втрати функціональної активності. Авторами також вперше запропоновано використати висококопійну плазміду для надекспресії плейотропних регуляторів, що суттєво збільшує кількість копій корисного гена і його позитивний вплив на продукцію антибіотика. 63820 4 Фіг. 1 Хімічна будова моеноміцину А, основного представника фосфогліколіпідної родини. Фіг. 2 Пригнічення росту Bacillus cereus ATCC19637 унаслідок дифузії фосфогліколіпідних антибіотиків з паперових дисків в агар, де 1 - диск з екстрактом контрольного штаму S. lividans + + TK24moeno38-5 pIJ101 ; 2 - диск-негативний контроль (розчинник - вода); 3 - диск з екстрактом штаму S. lividans + + TK24moeno38-5 pIJ584 . Спосіб можна проілюструвати прикладами: Приклад із використанням штаму S. lividans + TK24moeno38-5 [Makitrynskyy R., Rebets Y., Ostash В., Zaburannyi N., Rabyk M, Walker S., FedorenkoV. Genetic factors that influence moenomycin production in streptomycetes. J Ind Microbiol Biotechnol. - 2010. - vol.37. -P.559 566]. Два інші штами актиноміцетів, S. ghanaensis та S. albus, використовуються і досліджуються аналогічно. Плазмідою pIJ584 трансформують штам Е. coli ET12567 (pUB307), який за рахунок trа-генів плазміди pUB307 забезпечує кон'югативне перенесення корезидентних плазмід [Flett F., Mersinias V., Smith C.P. High efficiency intergeneric conjugal transfer of plasmid DNA from Escherichia coli to methyl-DNA-restricting Streptomycetes II FEMS Microbiol. Lett-1997. - vol.155. -P.223-229]. Одну колонію нічної культури Escherichia coli засівають у 5 мл середовища LB з 50 мкг/мл канаміцину. Культуру вирощують до оптичної густини OD600 = 0,1, переносять у мікропробірки з об'ємом 1.5 мл і осаджують центрифугуванням при 10 тис. об./хв протягом 1 хв. Зливають супернантант та клітини ресуспендують у 50 мкл середовища LB. Отриману суспензію клітин охолоджують у льоді 5 хв, додають 1 мл 0,1 М розчину СаСl2 та інкубують у льоді 1 год. Клітини осаджують центрифугуванням при 10 тис. об./хв протягом 1 хв, зливають надосадову рідину та ресуспендують у 100 мкл ОДМ розчину СаСl2. Інкубують 1 год у льоді та додають розчин плазмідної ДНК. Інкубують 1 год у льоді, після чого клітини піддають тепловому шоку протягом 1 хв. при 40 °С, охолоджують та додають 1 мл середовища LB. Інкубують 2 год при 37 °С для індукції експресії генів стійкості та висівають на чашки з середовищем LA з 25 мкг/мл апраміцину та 50 мкг/мл канаміцину. Чашки інкубують при 37 °С 16 год, після чого трансформанти пересівають на свіже середовище LA з 25 мкг/мл апраміцину та 50 мкг/мл канаміцину. + Плазміду pIJ584 в S. lividans TK24moeno38-5 переносять шляхом міжродової кон'югації з відповідним штамом Е. coli ET12567 (pUB307). Суспензію спор штаму S. lividans + TK24moeno38-5 висівають на середовище ОМ (г/л: вівсяне борошно - 30, агар - 18, вода водопровідна - до 1 л, рН до стерилізації - 7,0; після стерилізації додають розчин хлориду магнію до кінцевої концентрації 40 мМ) та вирощують 6 діб при 28 °С для отримання спорової суспензії для кон'югаційних схрещувань. Штам Е. сої і ЕТ12567 (pUB307) з плазмідою pSOK804 5 висівають на чашку з середовищем LA з 25 мкг/мл апраміцину та 50 мкг/мл канаміцину та вирощують 18 год при 37 °С. Готують суспензію клітин у 4 мл середовища LB. Одночасно готують суспензію + спор штаму S. lividans TK24moeno38-5 у 4 мл стерильного фізіологічного розчину. Суспензію клітин та спор стрептоміцетів осаджують центрифугуванням 2 хв при 10000 об/хв., зливають надосадову рідину, розчиняють у 100 мкл середовища LB, змішують між собою та висівають на чашки з середовищем ОМ. Чашки інкубують 18-20 год при 28 °С та заливають 1 мл водного розчину 25 мкг апраміцину та 50 мкг налідиксової кислоти. Перевіряють рівень продукції фосфогліколіпідних антибіотиків транскон'югантом + + S. lividans TK24moeno38-5 pIJ584 та копійність плазміни у клітинах. Штам висівають у 30 мл середовища TSB та вирощують протягом 5 діб при 30 °С. Біомасу осаджують за допомогою центрифугування, промивають водою та екстрагують 7 мл метанолу протягом 14 год. Одночасно, на чашку із середовищем LA висівають репортерний штам Bacillus cereus ATCC19637 для отримання нічної культури. Екстракт випаровують і сухий залишок Комп’ютерна верстка Д. Шеверун 63820 6 розчиняють у 100 мкл води. Одночасно на чашку із середовищем LA висівають суспензію клітин нічної культури В. cereus. На паперові диски Whatman діаметром 5 мм наносять 20 мкл водного розчину і потім накладають на щойно засіяний газон репортерного штаму. Чашки з дисками інкубують при 37 °С протягом 24 год. Кількість моеноміцинів у зразках визначають вимірюванням діаметра зон пригнічення екстрактом росту В. cereus та порівнянням із діаметрами пригнічення росту з використанням дисків із відомою кількістю моеноміцину А. Транскон'юганти продукують у 3 рази більше моеноміцинів (6±1 мг/л ферментаційного середовища) ніж контрольний штам (2±0,3 мг/л ферментаційного середовища), як відображено на Фіг.2. Також, виділення плазмідної ДНК із клітин транскон'юганта та її спектрофотометричний аналіз підтверджує її присутність у 100-400 копія у перерахунку на одну хромосому. Висока копійність плазміди та підвищений рівень синтезу антибіотика підтверджують отримання передбачуваного результату. Підписне Тираж 23 прим. Державна служба інтелектуальної власності України, вул. Урицького, 45, м. Київ, МСП, 03680, Україна ДП “Український інститут промислової власності”, вул. Глазунова, 1, м. Київ – 42, 01601
ДивитисяДодаткова інформація
Назва патенту англійськоюMethod for enhancing phosphoglycolipid antibiotics biosynthesis
Автори англійськоюOstash Bohdan Omelianovych, Fedorenko Viktor Oleksandrovych, Hromyko Oleksandr Mykolaiovych, Woker-Kane Syuzen
Назва патенту російськоюСпособ повышения биосинтеза фосфогликолипидных антибиотиков
Автори російськоюОсташ Богдан Емельянович, Федоренко Виктор Александрович, Громыко Александр Николаевич, Уокер-Кане С'юзен
МПК / Мітки
МПК: C12Q 1/02, C12Q 1/04, C12Q 1/00
Мітки: біосинтезу, фосфогліколіпідних, підвищення, спосіб, антибіотиків
Код посилання
<a href="https://ua.patents.su/3-63820-sposib-pidvishhennya-biosintezu-fosfoglikolipidnikh-antibiotikiv.html" target="_blank" rel="follow" title="База патентів України">Спосіб підвищення біосинтезу фосфогліколіпідних антибіотиків</a>
Спосіб підвищення специфічної активності антибіотиків

Номер патенту: 34206
Опубліковано: 11.08.2008
Автори: Манський Олександр Анатолійович, Дикий Ігор Леонідович, Домарьов Анатолій Павлович, Філімонова Наталія Ігорівна
МПК: A61L 2/08
Мітки: спосіб, підвищення, специфічно, активності, антибіотиків
Формула / Реферат:
Спосіб підвищення специфічної активності антибіотиків шляхом обробки останніх γ-опромінюванням, який відрізняється тим, що опромінювання проводять протягом понад 1,0 до 1,5 сек. до одержання дози понад 10 до 15 Грей.
Спосіб підвищення та відновлення чутливості патогенної мікрофлори до антибіотиків

Номер патенту: 47879
Опубліковано: 15.07.2002
Автори: Жалдаченко Вікторія Петрівна, Негара Вікторія Анатоліївна, Мартинець Петро Андрійович
МПК: A61K 31/715
Мітки: мікрофлори, антибіотиків, відновлення, чутливості, патогенної, підвищення, спосіб
Формула / Реферат:
Спосіб підвищення та відновлення чутливості патогенної мікрофлори до антибіотиків шляхом безпосереднього впливу медикаментозних препаратів на мікрофлору, який відрізняється тим, що хворому додатково проводять 5-7 інгаляцій натрієвої солі гепарину в дозі 5000 Од у 1мл 0,85% розчину хлориду натрію один раз на добу протягом 15 хвилин.
Спосіб виявлення антибіотиків тіострептонового ряду

Номер патенту: 91725
Опубліковано: 25.08.2010
Автори: Федоренко Віктор Олександрович, Осташ Богдан Омелянович, Мироновський Максим Леонідович, Осташ Ірина Степанівна
МПК: C12Q 1/04, C12Q 1/02, C12Q 1/00
Мітки: спосіб, тіострептонового, виявлення, ряду, антибіотиків
Формула / Реферат:
Спосіб виявлення антибіотиків тіострептонового ряду, що базується на індукції експресії гена канаміциностійкості nео у Streptomyces lividans, який відрізняється тим, що як векторну молекулу ДНК використовують інтегративну плазміду рIJ6902.
Спосіб підвищення специфічної активності антибіотиків
Номер патенту: 73447
Опубліковано: 15.07.2005
Автори: Дикий Ігор Леонідович, Чуєшов Владислав Іванович, Яковенко Володимир Дмитрович, Холупяк Ірина Юріївна, Шевельова Наталія Єфимівна, Стрельников Леонід Семенович, Ковальова Тетяна Іванівна, Велика Марина Марківна, Шевченко Михайло Іванович
Мітки: специфічно, спосіб, активності, антибіотиків, підвищення
Формула / Реферат:
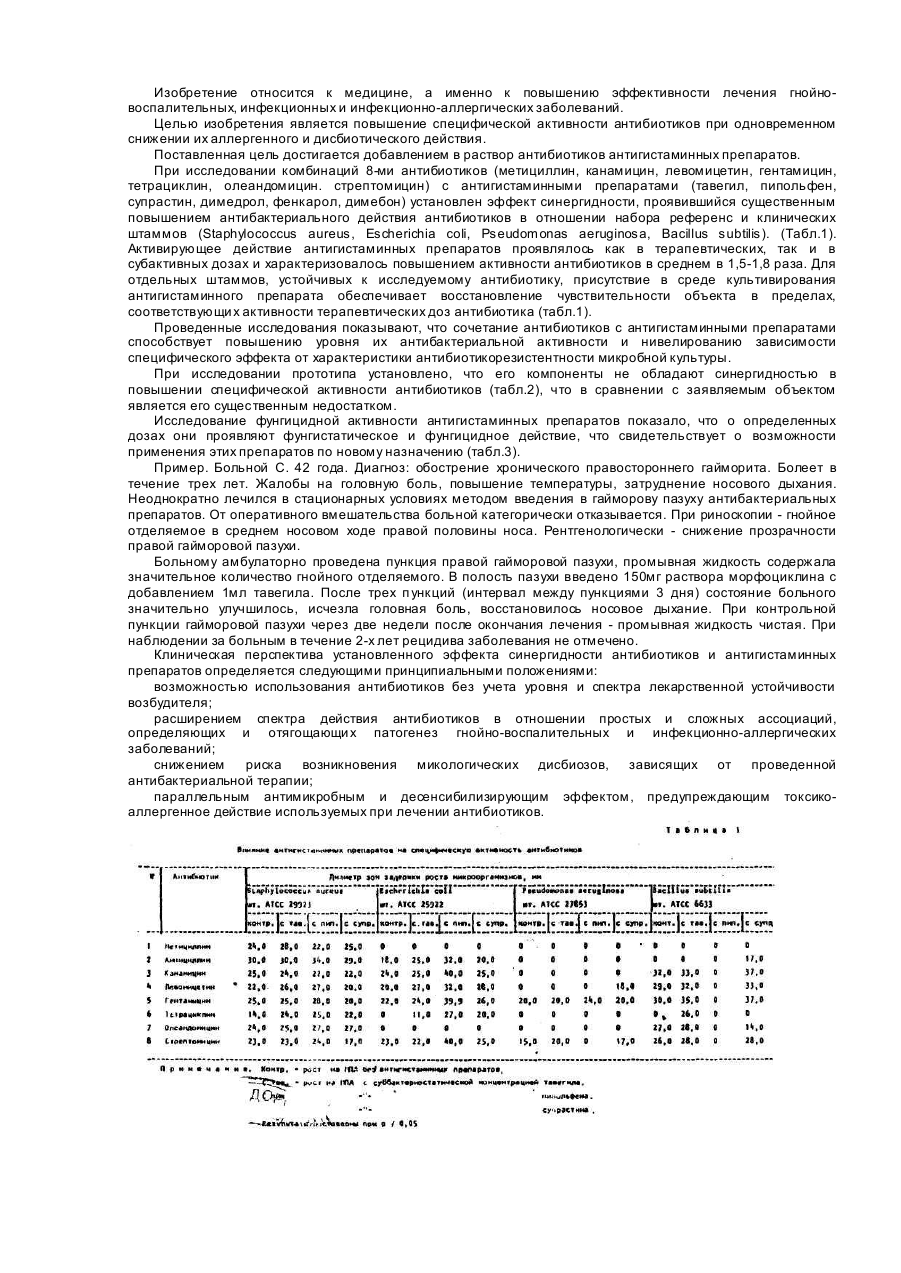
Способ повышения специфической активности антибиотиков, включающий добавление активаторов антибиотической активности, отличающийся тем, что, с целью снижения аллергенного и дисбиотического эффекта, в качестве активатора используют антигистаминные препараты.
Спосіб експрес-виявлення антибіотиків групи ландоміцинів

Номер патенту: 88383
Опубліковано: 12.10.2009
Автори: Осташ Ірина Степанівна, Бехтольд Андреас, Лужецький Андрій Миколайович, Федоренко Віктор Олександрович, Осташ Богдан Омелянович
МПК: C12Q 1/04
Мітки: групи, експрес-виявлення, спосіб, антибіотиків, ландоміцинів
Формула / Реферат:
Спосіб експрес-виявлення три-, пента- і гексаглікозильованих ландоміцинів, який базується на індукції експресії гена канаміцин-стійкості nео у стрептоміцетів, який відрізняється тим, що як регуляторний елемент використовують ген lаnК, клонований у складі плазміди рМО11с у актиноміцетів Streptomyces albus.
Попередній патент: Спосіб одержання тернарних сполук кристалічної ромбічної структури типу gd3ni5al19 з аморфних сплавів аl-пм-рзм
Наступний патент: Термоелектричний сплав
Випадковий патент: Електропривід постійного струму з модулем штучного інтелекту