Модулятори метаболічних шляхів внутрішньоклітинного запалення, загибелі клітин та виживання клітин








Формула / Реферат
1. Виділена або очищена молекула ДНК, що складається з послідовності ДНК, яка кодує білок В1, його ізоформи, фрагменти або аналоги, причому вказані білок В1, його ізоформи, фрагменти або аналоги здатні безпосередньо або опосередковано взаємодіяти з внутрішньоклітинними медіаторами або модуляторами каскадів запалення, клітинної загибелі або клітинного виживання, причому вказані білок В1, його ізоформи, фрагменти або аналоги є внутрішньоклітинними модуляторами вказаних внутрішньоклітинних каскадів запалення, клітинної загибелі і/або клітинного виживання, де вказана молекула ДНК вибрана з групи, що складається з:
(a) молекули кДНК, що складається з послідовності ДНК, наведеної на фігурі 3В;
(b) фрагмента послідовності (а), що кодує біологічно активний білок, здатний модулювати каскади запалення, клітинної загибелі або клітинного виживання;
(c) молекули ДНК, що складається з послідовності ДНК, здатної гібридизуватися з послідовністю (а) або (b) у помірно жорстких умовах і кодуючої біологічно активний білок В1, аналог або фрагмент, здатний модулювати внутрішньоклітинні каскади запалення, клітинної загибелі або клітинного виживання; і
(d) молекули ДНК, що складається з послідовності ДНК, генетично виродженої по відношенню до послідовностей ДНК, визначеним (а)-(с), і кодуючої біологічно активний білок В1, аналог або фрагмент, здатний модулювати каскади запалення, клітинної загибелі або клітинного виживання.
2. Вектор, що містить молекулу ДНК за п. 1.
3. Вектор за п. 2, який відрізняється тим, що здатний експресуватися в еукаріотичній клітині-хазяїні.
4. Вектор за п. 2, який відрізняється тим, що здатний експресуватися в прокаріотичній клітині-хазяїні.
5. Штам трансформованих еукаріотичних або прокаріотичних клітин-хазяїнів, що містять вектор за будь-яким з пп. 2-4.
6. Виділений або очищений білок В1, його ізоформа, фрагмент, функціональний аналог і похідне, які кодуються молекулою ДНК за п. 1, причому вказані білок, його ізоформа, фрагмент, аналог і похідне здатні безпосередньо або опосередковано модулювати внутрішньоклітинні каскади запалення, клітинної загибелі або клітинного виживання, асоціюючись з іншими внутрішньоклітинними модуляторами або медіаторами вказаних каскадів.
7. Виділений або очищений білок В1, його ізоформа, фрагмент, аналог і похідне за п. 6, який відрізняється тим, що вказані білок, ізоформа, фрагмент, аналог і похідне мають щонайменше частину амінокислотної послідовності, наведеної на фігурі 3.
8. Виділений або очищений фрагмент білка В1 за п. 6, який відрізняється тим, що є пептидом.
9. Спосіб одержання білка В1, його ізоформи, фрагмента, аналога або похідного за будь-яким з пп. 6-7, що передбачає культивування штаму трансформованих клітин-хазяїнів за п. 5 в умовах, що забезпечують експресію вказаного білка, його ізоформи, фрагмента, аналога і похідного, за необхідності проведення посттрансляційної модифікації з одержанням вказаного білка, його ізоформи, фрагмента, аналога або похідного і виділення вказаного експресованого білка, його ізоформи, фрагмента, аналога або похідного.
10. Антитіло або його активні фрагмент або похідне, специфічне у відношенні до білка В1, його ізоформи, аналога, фрагмента або похідного за будь-яким з пп. 6-7.
11. Спосіб виділення та ідентифікації білка за будь-яким з пп. 6-7, який має гомологію або здатний безпосередньо або опосередковано взаємодіяти з будь-якими білками, що містять продомен або домен рекрутингу каспази (CARD), або інші білки або ферменти, залучені до внутрішньоклітинної передачі сигналу, за допомогою кіназного або проміжного доменів, які присутні у вказаному білку за будь-яким з пп. 6 або 7, який передбачає застосування дріжджової двогібридної методики, відповідно до якої послідовність, що кодує вказаний білок із вказаними CARD-, кіназним і проміжним доменами або щонайменше з одним з цих доменів, утримується в одному гібридному векторі, а послідовність з бібліотеки кДНК або геномних ДНК утримується в другому гібридному векторі, причому вектори потім застосовуються для трансформації дріжджових клітин-хазяїнів, а позитивно трансформовані клітини виділяють з наступним виділенням вказаного другого гібридного вектора з одержанням послідовності, що кодує білок, який зв'язується із вказаним білком, що містить вказані CARD-, кіназний і/або проміжний домени.
12. Спосіб за п. 11, який відрізняється тим, що вказаний білок являє собою щонайменше одне з ізоформи, аналога, фрагмента і похідного білка В1.
13. Фармацевтична композиція для модуляції каскадів запалення, клітинної загибелі, клітинного виживання або інших каскадів у клітинах, що модулюються безпосередньо або опосередковано білком В1, яка містить як активний інгредієнт щонайменше один білок В1 за будь-яким з пп. 6-7, його біологічно активний фрагмент, аналог, похідне або їх суміші.
14. Фармацевтична композиція для модуляції каскадів запалення, клітинної загибелі, клітинного виживання або інших каскадів у клітинах, що модулюються безпосередньо або опосередковано В1, яка містить як активний інгредієнт рекомбінантний вектор на основі вірусу тварини, що кодує білок, здатний до зв'язування з рецептором на поверхні вказаних клітин, і що кодує щонайменше один білок В1 за будь-яким з пп. 6-7, його ізоформу, активний фрагмент або аналог.
15. Фармацевтична композиція для модуляції каскадів запалення, клітинної загибелі, клітинного виживання або інших каскадів у клітинах, що модулюються безпосередньо або опосередковано В1, яка містить як активний інгредієнт олігонуклеотидну послідовність, що кодує антисмислову послідовність мРНК білка В1 за п. 1.
16. Фармацевтична композиція для профілактики або лікування патологічного стану, асоційованого з регуляцією апоптозу однією або декількома молекулами, з якими безпосередньо або опосередковано зв'язується білок В1 за будь-яким з пп. 6-7, причому вказана композиція містить ефективну кількість білка В1 або молекули ДНК, що його кодує, або молекули, здатної порушувати безпосередню або опосередковану взаємодію вказаного білка В1 з однією або декількома молекулами, з якими зв'язується або взаємодіє білок В1.
17. Фармацевтична композиція для профілактики або лікування патологічного стану, асоційованого з регуляцією апоптозу однією або декількома молекулами, з якими безпосередньо або опосередковано зв'язується білок В1 за будь-яким з пп. 6-7, причому вказана композиція містить ефективну кількість білка В1, його ізоформи, фрагмента, аналога або похідного або молекули ДНК, що їх кодує, або молекули, здатної порушувати безпосередню або опосередковану взаємодію вказаного білка В1, його ізоформи, фрагмента, аналога або похідного з однією або декількома молекулами, з якими зв'язується вказаний білок В1, його ізоформа, фрагмент, аналог або похідне.
18. Фармацевтична композиція для профілактики або лікування патологічного стану, асоційованого з регуляцією апоптозу однією або декількома молекулами, з якими безпосередньо або опосередковано зв'язується білок В1 за будь-яким з пп. 6-7, причому вказана композиція містить молекулу, здатну порушувати протеїнкіназну активність В1.
19. Спосіб скринінгу ліганду, здатного зв'язуватися з білком В1 за будь-яким з пп. 6-7, який передбачає взаємодію носія для афінної хроматографії, до якого приєднаний вказаний білок, з екстрактом клітин, у результаті чого ліганд зв'язується з вказаним носієм, і елюцію, виділення й аналіз вказаного ліганду.
20. Спосіб скринінгу молекули ДНК, що складається з послідовності ДНК, що кодує ліганд, здатний зв'язуватися з білком В1 за будь-яким з пп. 6-7, який передбачає використання дріжджового двогібридного методу, відповідно до якого послідовність, що кодує вказаний білок В1, міститься в одному гібридному векторі, а послідовності з бібліотеки кДНК або геномної ДНК містяться в другому гібридному векторі, трансформацію дріжджових клітин-хазяїнів - вказаними векторами, виділення позитивних трансформованих клітин і виділення вказаного другого гібридного вектора з одержанням послідовності, що кодує вказаний ліганд.
21. Спосіб ідентифікації й одержання ліганду, здатного модулювати клітинну активність, яка модулюється або опосередковується білком В1, який передбачає:
a) скринінг ліганду, здатного зв'язуватися з поліпептидом, що містить щонайменше частину білка В1, що містить щонайменше деякі амінокислотні залишки білка В1, наведені на фіг. 3, які включають в себе по суті весь продомен (або CARD) В1;
b) ідентифікацію і характеристику ліганду, відмінного від BCL2, TRAF2 або частин рецептора із сімейства рецепторів TNF/NGF, або інших відомих білків, що містять продомен (CARD), які мають здатність до вказаного зв'язування за даними вказаної стадії скринінгу; і
c) одержання вказаного ліганду в суттєво виділеній і очищеній формі.
22. Спосіб ідентифікації й одержання ліганду, здатного модулювати клітинну активність, яка модулюється або опосередковується білком В1 за будь-яким з пп. 6-7, який передбачає:
a) скринінг ліганду, здатного зв'язуватися з поліпептидом, що містить щонайменше карбоксикінцеву частину послідовності В1, наведеної на фіг. 3, включаючи продомен (CARD);
b) ідентифікацію і характеристику ліганду, відмінного від BCL2, TRAF2 або частин рецептора із сімейства рецепторів TNF/NGF, або інших відомих білків, що містять продомен (CARD), які мають здатність до вказаного зв'язування за даними вказаної стадії скринінгу; і
c) одержання вказаного ліганду в суттєво виділеній і очищеній формі.
23. Спосіб ідентифікації й одержання ліганду, здатного модулювати клітинну активність, яка модулюється або опосередковується В1, який передбачає:
a) скринінг ліганду, здатного зв'язуватися щонайменше з амінокінцевою частиною послідовності В1, наведеної на фіг. 3, що містить по суті весь кіназний домен В1;
b) ідентифікацію і характеристику ліганду, відмінного від BCL2, TRAF2 або частин рецептора із сімейства рецепторів TNF/NGF, або інших відомих внутрішньоклітинних модуляторних білків, які мають здатність до вказаного зв'язування за даними вказаної стадії скринінгу; і
c) одержання вказаного ліганду в суттєво виділеній і очищеній формі.
24. Спосіб ідентифікації й одержання молекули, здатної безпосередньо або опосередковано модулювати клітинну активність, яка модулюється або опосередковується В1, який передбачає:
a) скринінг молекули, здатної модулювати активності, які модулюються або опосередковуються В1;
b) ідентифікацію і характеристику вказаної молекули; і
c) одержання вказаної молекули в суттєво виділеній і очищеній формі.
25. Спосіб ідентифікації й одержання молекули, здатної безпосередньо або опосередковано модулювати клітинну активність, яка модулюється або опосередковується білком за будь-яким з пп. 6-7, який передбачає:
a) скринінг молекули, здатної модулювати активності, які модулюються або опосередковуються білком за будь-яким з пп. 6-7;
b) ідентифікацію і характеристику вказаної молекули; і
c) одержання вказаної молекули в суттєво виділеній і очищеній формі.
26. Спосіб модулювання клітинної загибелі, клітинного виживання і/або запалення, який передбачає обробку клітин молекулою, здатною взаємодіяти з Е-субодиницею V-АТФази, де молекула являє собою білок В1 за будь-яким з пп. 6-7, його ізоформу, фрагмент, функціональний аналог або похідне.
Текст
Даний винахід стосується області модуляторів шляхів внутрішньоклітинної загибелі та клітинного виживання, опосередкованих, поміж іншого, рецепторами ФНП/ФРН надсімейства рецепторів та зв'язаних з ними внутрішньоклітинними адаптерними білками, каспазних та кіназних ферментів. Більш конкретно, даний винахід стосується нового білка, спочатку позначеного СВК, але зараз позначуваного В1, його ізоформ, аналогів, фрагментів та похідних, що, як здається, здатні взаємодіяти, безпосередньо чи опосередковано, з різними внутрішньоклітинними білками та ферментами, які мають відношення до шляхів клітинної загибелі, клітинного виживання та запалення, і, отже, які є модуляторами цих шляхів. Надсімейство рецепторів фактора некрозу пухлини/фактора росту нерва (ФНП/ФРН) визначається за допомогою структурної гомології між внутрішньоклітинними доменами їх членів [Bazan, 1993; Beutler та van Huffel, 1994; Smith et al., 1994]. За винятком двох рецепторів, рецептора р55 ФНП та Fas/APO1, різні члени цього сімейства рецепторів не мають ясної подібності структури в їх внутрішньоклітинних доменах. Проте, між рецепторами існує велика подібність у дії, яка свідчить про те, що вони беруть участь у звичайних сигнальних шляхах. Одним прикладом цієї подібності є здатність деяких рецепторів сімейства ФНП/ФРН активувати фактор транскрипції NF-kΒ. Цю звичайну здатність було віднесено до здатності білка цитоплазми, який активує NF-kB, Асоційований з ФНП Рецептором Фактор 2 (TRAF2), зв'язуватись із структурно дисимільованими внутрішньоклітинними доменами деяких рецепторів сімейства ФРП/ФРН. Але не відомо, якими є механізми дії TRAF2 і як координується його відповідна реакція на різні рецептори, з якими він зв'язується. TRAF2 є членом недавно описаного сімейства білків, названих TRAF, яке включає кілька білків, ідентифікованих як, наприклад, TRAF1, TRAF2 [Rothe, M., Wong, S.C., Henzel, W.G та Goeddel, D. (1994) Cell 78:681-692; опублікована заявка РСТ WO95/33051), TRAF3 (Cheng, G. et al. (1995)) та TRAF6 (див. Сао et al., 1996a]. Усім білкам, що належать до сімейства TRAF, у високому ступеню притаманна амінокислотна послідовність по їх С-кінцевим доменам, тоді як їх N-кінцеві домени можуть бути не зв'язані. Як показано на схематичній ілюстрації TRAF2 (Фіг.1), молекула містить кільцевий палець та два TFIIIA-подібні цинкові пальці в області їх С-кінців. С-кінцева половина молекули включає область, відому як «домен TRAF», що містить ефективну область лейцинового зіпера, розташовану між амінокислотами 264-358 (що називається N-TRAF), і другу частину у напрямку до карбоксикінця домену між амінокислотами 359-501 (що називається C-TRAF), яка відповідає за зв'язування TRAF з рецепторами та з іншими молекулами TRAF з утворенням гомо- чи гетеродимерів. Активація фактора транскрипції NF-kΒ є одним з проявів сигнального каскаду, ініційованого деякими рецепторами ФНП/ФРН і опосередкованого TRAF2. NF-kΒ включає члени сімейства димер-утворюючих білків з гомологією до онкогена Rel, які у їх димерній формі діють як фактори транскрипції. Ці фактори повсюдно поширені і беруть участь у регуляції експресії складних множинних генів. Хоч спочатку NF-kΒ був ідентифікований як фактор, який є структурно присутнім у В-клітинах на стадії експресії Igk легкого ланцюга, NF-kΒ є в основному відомим по його дії як індуцибельного активатора транскрипції. У більшості відомих випадків NF-kΒ веде себе як головний фактор, а саме, індукція його активності здійснюється за допомогою активації вже наявних молекул, що є присутніми у клітині у їх неактивній формі, скоріше ніж їх синтез denovo, який у свою чергу спирається на індуцибельний фактор транскрипції, що включає NF-kΒ ген. Дія NFkΒ є високо плейотропною. Більшість цих численних ефектів має спільні характеристики по швидкому індукуванню відповіді на позаклітинний подразник. Основна частина NF-kB-активуючих агентів є індукторами імунного захисту, включаючи компоненти вірусів та бактерій, цитокінів, які регулюють імунну відповідь, УФ-світло та інше. Відповідно, багато які з генів, що регулюються NF-kB, роблять внесок до імунного захисту [див. Blank et al., 1992; Grilli et al., 1993; Baeuerle and Henkel, 1994, як посилання]. Одною основною характерною ознакою NF-kB-регуляції є те, що цей фактор може існувати у цитоплазматичній формі, що не зв'язує ДНК, яка може бути індукована для перенесення до ядер, зв'язування ДНК та активації транскрипції. Ця подвійна форма NF-kΒ білків регулюється за допомогою І-kBсімейства білків, які містять повтори домену, що спочатку був розпізнаний у еритроцитному білку анкірині [Gilmore and Morin, 1993]. У незбудженому стані, NF-kΒ димер зустрічається у асоціації з І-kB молекулою, яка розміщається на місці його цитоплазматичної локалізації і перешкоджає його взаємодії з NF-kBзв'язуючою ДНК послідовністю та активації транскрипції. Дисоціація І-kB з NF-kΒ димеру включає визначальну стадію його активації за допомогою багатьох його індукуючих агентів [DiDonato et al., 1995]. Знання механізмів, які утягнені до цієї регуляції, усе ще обмежене. Також лише трохи відомо про шлях, яким визначається специфічність клітин у термінах реактивності щодо різних NF-kB-індукуючих агентів. Одним з найсильніших індукуючих агентів NF-kΒ є цитокіновий фактор некрозу пухлини (ФНП). Існують два різних рецептора ФНП, р55 та р75 рецептори (p55-R та p75-R). Їх рівень експресії незалежно варіює у різних клітинах [Vandenabeele et al., 1995]. Рецептор р75 відповідає, краще, за клітинно-зв'язану форму ФНП (ФНП експресований як у вигляді бета-трансмембранного білка, так і у вигляді розчинного білка), тоді як р55 рецептор відповідає лише за ефективне розчинення ФНП молекул [Grell et aІ., 1995]. Внутрішньоклітинні домени двох рецепторів структурно не близькі і зв'язують різні цитоплазматичні білки. Проте, принаймні частина ефектів ФНП, включаючи дію ФНП, що руйнує клітини, та індукцію NF-kΒ, може бути індукована за допомогою обох рецепторів. Ці властивості є клітинно-специфічними. Рецептор р55 є здатним індукувати дію, що руйнує клітини, чи активацію NF-kΒ в усіх клітинах, які виявляють такі ефекти у відповідь на ФНП. p75-R може мати такі ефекти лише у деяких клітинах. Інші, хоча б і такі, що експресують у високому ступеню p75-R, демонструють індукцію таких ефектів лише у відповідь на стимуляцію p55-R [Vandenabeele et al., 1995]. Крім рецепторів ФНП, різні інші рецептори сімейства рецепторів ФНП/ФРН: CD30 [McDonald et al., 1995], CD40 [Berberich et al., 1994; Lalmanach-Girard et al., 1993], бета рецептор лімфотоксину і, у деяких типах клітин, Fas/APO1 [Rensing-Ehl et al., 1995], також здатні індукувати активацію NF-kB. Рецептор ІЛ-1 І типу, також такий, що ефективно стимулює активацію NF-kΒ, бере участь у більшості ефектів рецепторів ФНП, незважаючи на те, що не є подібним до них за структурою. Активація NF-kΒ при антигенній стимуляції примірованої клітини цих різних рецепторів відбувається через індуковане фосфорилювання їх асоційованих І-kB молекул. Це фосфорилювання мітить І-kB для деградації, яка найвірогідніше протікає у протеосомі. Природа кінази, яка фосфорилює І-kB, та її механізм активації рецепторної стимуляції примірованної клітини усе ще не відомі. Однак за останні два роки був досягнутий певний прогрес у знанні, як ідентифікувати три асоційовані з рецептором білка, що, як здається, беруть участь у ініціюванні фосфорилювання [див. ілюстрацію у вигляді діаграм на фігурах 2а та 6]. Білок, що називається TRAF2, спочатку клонований D. Goeddel та його колегами [Rothe et al., 1994], як здається, відіграє центральну роль у NF-kB-активації за допомогою різних рецепторів сімейства ФНП/ФРН. Білок, який при експресії у високому ступеню може сам стимулювати активацію NF-kΒ, зв'язується з активованим р75 ΦΗΠ-R [Rothe et al., 1994], бета рецептором лімфотоксину [Mosialos et al., 1995], CD40 [Rothe et al., 1995a] та CD-30 (неопубліковані дані) і опосередковують з їх допомогою індукцію NF-kΒ. TRAF2 не приєднується ні до рецептора р55 ФНП, ні до Fas/APO1, але він може зв'язуватись з р55 рецепторасоційованим білком, що називається TRADD, і TRADD має здатність зв'язуватись з Fas/APO1асоційованим білком, що називається MORT1 [або FADD - див. Boldin et al., 1995b та 1996]. Інший рецептор-взаємодіючий білок, що називається RIP [див. (Stanger et aІ., 1995] також може взаємодіяти з TRAF2, а також з Fas/APO1, TRADD, рецептором р55 ФНП та MORT-1. Таким чином, незважаючи на те, що RIP є асоційованим з індукцією клітинної цитотоксичності (клітинна загибель), його здатність взаємодіяти з TRAF2 також утягує його до активації NF-kΒ і він також може слугувати, крім того, для посилення взаємодії FAS/APO1, MORT-1, рецептора р55 ФНП та TRADD з TRAF2 шляхом, що призводить до активації NF-kΒ. Ці асоціації, як здається, дозволяють рецептору р55 ФНП та Fas/APO1 стимулювати активацію NF-kΒ [Hsu et al., 1995; Boldin et al., 1995; Chinnaiyan et al., 1995; Varfolomeev et al., 1996; Hsu et al., 1996]. Стимуляція активації NF-kΒ за допомогою рецептора ІЛ-1 протікає незалежно від TRAF2 і може включати недавно клонований ІЛ-1 рецептор-асоційований білок-кіназу, що називається IRAK [Croston et al., 1995]. За яким механізмом діє TRAF2, є непевним. Деякі цитоплазматичні молекули, які зв'язують TRAF2, були ідентифіковані [Rothe et al., 1994; Rothe et al., 1995b]. Однак інформація про ці молекули не дає якогось ключа щодо шляху, за допомогою якого TRAF2, який сам не має якоїсь ферментної активності, викликає фосфорилювання І-kB. Також поки що немає інформації про механізми, що визначають клітинноспецифічний слід активації TRAF2 за допомогою різних рецепторів, таких як, спостерігались для індукції NFkΒ за допомогою двох ФНП рецепторів. На додаток до вищевказаного, з різних TRAF білків, необхідно також відзначити, що TRAF2 зв'язується з р55 (CD120a) та р75 (CD120b) ФНП рецепторами, а також з деякими іншими рецепторами сімейства рецепторів ФНП/ФРН, чи то безпосередньо, чи то опосередковано через інші адаптерні білки, як вказано вище, наприклад, з відсиланням до FAS/APO1 рецептора, і адаптерні білки MORT-1, TRADD та RIP. Як такий, TRAF2 є визначальним для активації NF-kΒ [див. також Wailach, 1996]. Однак, TRAF3 дійсно інгібує активацію NF-kΒ за допомогою деяких рецепторів сімейства ФНП/ФРН [див. Rothe et al., 1995a], тоді як TRAF6 є необхідним для індукції NF-kΒ за допомогою ІЛ-1 [див. Cao et al., 1996а]. Відповідно, що стосується NF-kΒ активації та її значення у підтримуванні життєздатності клітини, різні внутрішньоклітинні шляхи, включені до цієї активації, раніше не були чітко пояснені, наприклад, як включені різні TRAF білки, безпосередньо чи опосередковано. Більш того, як відомо зараз стосовно різних членів сімейства рецепторів ФНП/ФРН та їх асоційованих внутрішньоклітинних сигнальних шляхів, включаючи різні адаптерні медіатор/модуляторні білки [див. короткі огляди та посилання, наприклад, у паралельних ізраїльських патентних заявках тих самих заявників 114615, 114986, 115319, 116588], ФНП та FAS/APO1 ліганд, наприклад, можуть чинити як корисну, так і пагубну дію на клітини. ФНП, наприклад, робить внесок до імунного захисту організму проти пухлин та інфекційних агентів і сприяє видужанню від уражень шляхом індукування лізису пухлинних клітин та клітин, інфікованих вірусом, посилюючи антибактеріальну активність гранулоцитів і, таким чином, у цих випадках бажаним є ФНП-індукований лізис клітин. Однак, надлишок ФНП може бути шкідливим і відомо, що як такий ФНП відіграє основну патогенну роль у ряді захворювань, таких як септичний шок, анорексія, ревматичні захворювання, запалення та реакції гість-проти-хазяїна. У таких випадках ФНП-індукований лізис клітин є небажаним. Ліганд FAS/APO1, наприклад, також має бажані та шкідливі ефекти. Цей ліганд FAS/APO1 індукує через його рецептор лізинг аутореактивних Τ клітин, які розпізнають аутоантигени у процесі їх розвитку і тому попереджають аутоімунні захворювання. Далі, різні злоякісні клітини та ВІЧ інфіковані клітини несуть рецептор FAS/APO1 на їх поверхні і можуть, тому, бути зруйновані активацією цього рецептора його лігандом чи специфічними до нього антитілами, і внаслідок цього активацією внутрішньоклітинних шляхів клітинної загибелі (апоптоз), опосередкованих цим рецептором. Однак рецептор FAS/APO1 може опосередковувати шкідливі ефекти, наприклад, неконтрольований лізис тканини, який спостерігається при деяких захворюваннях, таких як гострий гепатит, що супроводжується руйнуванням клітин печінки. З точки зору вищевказаного, а саме того, що рецептори сімейства ФНП/ФРН можуть індукувати шляхи клітинної загибелі з одного боку і можуть індукувати шляхи виживання клітин (через індукцію NF-kΒ) з іншого боку, як здається, існує добра рівновага, внутрішньоклітинно, між цими двома протилежними шляхами. Наприклад, коли бажано досягнути максимальної деструкції ракових клітин чи інших інфікованих або хворих клітин, було б бажаним мати ФНП та/або FAS/APO1 ліганд, що індукує лише шлях клітинної загибелі без індукування NF-kΒ. Отже, якщо бажано захистити клітини, такі як, наприклад, при запаленні, реакціях «гість проти хазяїна», гострому гепатиті, було б бажано блокувати індукцію клітинного лізису ФНП та/або FAS/APO1 ліганду та підвищити, замість цього, їх індукцію NF-kΒ. Подібним чином, у деяких патологічних випадках, було б бажаним блокувати внутрішньоклітинні сигнальні шляхи, опосередковані за допомогою рецептора р75 ФНП та рецептора ІЛ-1, тоді як у інших випадках було б бажаним посилити ці внутрішньоклітинні шляхи. Недавно, заявниками була виділена кіназа, що називається NIK [патентні заявки Ізраїлю 117800, 119133 та WO97/37016], яка здатна до зв'язування з TRAF2 і безпосередньо утягнена до реакцій фосфорилювання, що призводять до індукції активації NF-kB. Крім того, недавно був виділений ряд каспаз (каспази) у великій кількості досліджень (включаючи даних заявників [див. одночасно подану ізраїльську патентну заявку тих самих заявників IL 120759)], які взаємодіють з вище відзначеними адаптерними білками (наприклад, MORT-1/FADD) або з комплексами між адаптерними білками та різними рецепторами сімейства рецепторів ФНП/ФРН і які здійснюють протеолітичні реакції, що призводять до апоптозної клітинної загибелі. Таким чином, пряма модуляція цих каспаз була б бажаною у ситуаціях, відзначених вище, коли бажано інгібувати чи посилити клітинну загибель, наприклад, коли бажано інгібувати клітинну загибель, було б бажано інгібувати активність цих каспаз. Стосовно цього було повідомлено [див. огляд у Hofmann et aІ., 1997], що існує область, що називається продоменом, у багатьох цих каспаз, яка також представлена у ряді адаптерних білків, таких як, наприклад, RAIDD (які взаємодіють з RIP, TRADD і, таким чином, з MORT-1/FADD, p55-0H(1-R та FAS/APO1), адаптерний білок шляху клітинної загибелі; і С-ІАР1, С-ІАР2, два білки, що, як здається, є інгібіторами апоптози і які самі взаємодіють з TRAF2, і, таким чином, можуть бути інгібіторами каспаз або, навпаки, можуть стимулювати включення TRAF2 до шляху клітинного виживання, що призводить до індукції NF-kΒ активації. Як такий, цей продомен був також позначений як CARD «капсазний рекрутинговий домен» [див. Hofmann et al., 1997]. Даний продомен (CARD), отже, представляє іншу мету для модуляції внутрішньоклітинних сигнальних шляхів, асоційованих з індукцією клітинної загибелі. Крім того, недавно було описано [див. огляд Yang та Korsmeyer, 1996] інше сімейство білків, назване сімейством білків BCL2, у якому білки BCL2, їх гомолог BCL-X, включаючи дві його форми, якими є BCL-XL та альтернативно сплайсовані BCL-XS, MCL1, А1, ВАК, BAD, BAG1, ΒΑΧ, аденовірус E1B-19k та білок Caenorhabditis elegans (С. elegans) CED-9 - усі є членами. Поміж цих білків спостерігалось, що BCL2, BCLXL, E1B-19k та CED-9 діють з метою інгібування апоптозу, або для захисту проти апоптозу, індукованого за допомогою різних внутрішньоклітинних сигнальних шляхів [див. Yang та Korsmeyer, 1996]. BCL2 та BCL-XL є також, напевно, внутрішньоклітинними мембранно-зв'язаними білками, локалізованими по відношенню до мітохондрій, а також до гладкої ендоплазматичної сітки, і перинуклеарної мембрани, при цьому С-кінці цих білків, мають сигнальну якірну послідовність, що відповідає за їх націлювання і проникнення до зовнішньої мітохондріальної мембрани та інших вказаних вище внутрішньоклітинних мембран. Після того, як заякорені у різних внутрішньоклітинних мембранах білки BCL2 та BCL-XL стануть доступними по відношенню до цитозолю, вони можуть взаємодіяти з різними іншими внутрішньоклітинними білками. Усе ще не зовсім зрозуміло, як BCL2, BCL-XL, E1B-19k та CED-9 захищають клітини, але ясно, що їх дія, напевно, вище ефекторів клітинної загибелі, які є різними каспазами, вказаними вище, такими як, наприклад, ICE та ІСЕ-подібні протеази сімейства ICE/CED-3, включаючи CPP/32Yama, ICE-LAP3 (Мсh3), ІСН-1 та інші. Дійсно, було виявлено, що CED-9 є специфічним інгібітором С.elegans протезних ефекторів загибелі CED-3 та CED-4, a BCL2, напевно, є інгібітором ІСН-1 (що називається також NEDD2), зокрема, форми ІСН-1L, яка сприяє клітинній загибелі. Таким чином, доки точний механізм інгібування апоптозу за допомогою BCL2, BCL-XL, CED-9 та Е1В-19k не зрозумілий, він, напевно, вище ICE-CED-3 протеаз, які є ефекторами загибелі [див. огляд Yang та Korsmeyer, 1996, а також Chinnaiyan et al., 1996]. Що стосується інших членів сімейства BCL2, вказаних вище, то ΒΑΧ є промотером клітинної загибелі. ΒΑΧ зв'язується сам з собою і у формі таких ΒΑΧ гомодимерів він сприяє апоптозу. ΒΑΧ також зв'язується з BCL2 та BCL-XL, і такі гетеродимери зв'язані із захисною дією BCL2 проти апоптозу. Таким чином, рівновага між кількостями гомодимерів ВАХ/ВАХ та гетеродимерів BAX/BCL2 визначає, чи будуть клітини сприйнятливими до апоптозу або ж вони будуть захищені проти апоптозу. ΒΑΧ, напевно, є також внутрішньоклітинним мембранно-зв'язаним білком, також локалізованим у великому ступені до зовнішньої мітохондріальної мембрани [стосовно вищевказаного ΒΑΧ див. також огляд Yang та Korsmeyer, 1996]. Далі, вказані вище білки ВАК та BAD також діють як негативні регулятори активності BACL2 та BCL-XL, а саме, вони пригнічують здатність BCL2 та BCL-XL захищати клітину від апоптозу. Вважають, що і ВАК, і BAD зв'язують BCL2 та BCL-XL і таким чином боронять ΒΑΧ від зв'язування з BCL2 та BCL-XL, що призводить до збільшення кількості гомодимерів ВАХ/ВАХ, і, отже, збільшення клітинної загибелі [див. огляд (Yang та Korsmeyer, 1996]. У цьому відношенні також зрозуміло, що дії ВАК по блокуванню пригнічуючої загибель активності BCL2 та BCL-XL безпосередньо у вигляді гетеродимерів BAK/BCL2 та BAK/BCL-XL не властива здатність захищати клітини від апоптозу. Здається, що BAD діє більше як конкуруючий інгібітор для зв'язування ΒΑΧ з BCL2 та BCL-XL, тоді як BAD може замінити ΒΑΧ з гетеродимерів BAX/BCL2 та BAX/BCLXL, таким чином забезпечуючи збільшення кількості гомодимерів ВАХ/ВАХ, що сприяють загибелі. Хоч, ВАК також, як здається, є внутрішньоклітинним мембранним зв'язуючим білком, локалізованим по відношенню, крім інших, до мітохондріальних зовнішніх мембран, BAD, однак, напевно позбавлений мембранної якірної послідовності і по суті не є білком, зв'язуючим мембрани [див. огляд Yang та Korsmeyer, 1996]. Іншим з вищевказаних членів сімейства BCL2 є BAG1 [див. Yang та Korsmeyer, 1996], який є позитивним модулятором активності BCL2, що призводить до посилення захисної активності BCL2 проти апоптозу і навіть забезпечує для BCL2 захисну активність проти апоптозу у клітинах, індукованих до дії апоптозу за допомогою сигналів, звичайно не пригнічуваних за допомогою BCL2. Можна також відзначити, що вищевказана альтернативно сплайсована форма BCL-XL, а саме, білок BCL-Xs, також є антагоністом активності BCL-XL та BCL2 і блокує їх захисну активність проти апоптозу [див. також огляд [Yang та Korsmeyer, 1996]. У світлі вищевказаного зрозуміло, що сімейство білків BCL2 відіграє роль у регулюванні шляхів клітинної загибелі чи клітинного виживання внутрішньоклітинно, а зсув у рівновазі від білків цього сімейства, які активно блокують апоптоз, до тих, які сприяють апоптозу чи інгібують протиапоптозну активність, може призводити до посилення клітинної загибелі і, подібним чином, зсув у рівновазі іншого шляху може призводити до посилення клітинного виживання. Отже, коли треба посилити клітинну загибель шляхом посилення апоптозу у клітинах за умов, вказаних вище, було б бажаним блокувати активність BCL2, BCL-XL та інших членів цього сімейства, які пригнічують чи інгібують апоптоз, або посилити активність ΒΑΧ, ВАК, BAD, BCL-Xs та інших членів цього сімейства, які сприяють апоптозу чи інгібують анти-апоптозну активність BCL2 чи BCL-XL. Аналогічно, коли бажано збільшити клітинне виживання у клітинах шляхом зменшення апоптозу, було б бажано збільшити активність BCL2, BCL-XL та інших членів цього сімейства, які пригнічують чи інгібують апоптоз, або зменшити активність апоптозних промотерів цього сімейства, як вказано вище. Об'єктом даного винаходу є нові білки, включаючи їх ізоформи, аналоги, фрагменти чи похідні, які здатні модулювати внутрішньоклітинні сигнальні шляхи, що ведуть до запалення, клітинної загибелі або клітинного виживання, і ця модуляція можлива через продомен (CARD) різних каспаз або через кіназні домени різних кіназ, утягнених до NF-kΒ активації. Тому такі нові білки за даним винаходом могли б, ймовірно, бути прямими модуляторами каспазної активності (шлях клітинної загибелі) та/або NF-kΒ активації через кіназну активність (шлях клітинного виживання). Аналогічно, нові білки за винаходом, ймовірно, є опосередкованими модуляторами внутрішньоклітинної біологічної активності безлічі інших білків, утягнених до запалення, шляху клітинної загибелі чи клітинного виживання (наприклад, FAS/APO1, р55 ΦΗΠ-R, р75 ΦΗΠ-R, IЛ-1-R, MORT-1, TRADD, RIP, TRAF2, NIK та інші). Аналогічно, ця модуляція, ймовірно, може здійснюватись шляхом прямої чи опосередкованої взаємодії з членами сімейства білків BCL2, нові білки за винаходом можуть модулювати активність BCL2 чи інших білків цього сімейства і, у цьому сенсі, нові білки за винаходом можуть бути опосередкованими модуляторами різних каспаз, які, у свою чергу, модульовані членами сімейства білків BCL2. Іншим об'єктом винаходу є антагоністи (наприклад, антитіла, пептиди, органічні сполуки чи навіть деякі ізоформи) по відношенню до вищевказаних нових білків, включаючи їх ізоформи, аналоги, фрагменти та похідні, які можуть бути використані для інгібування запалення, клітинної загибелі чи сигнальних процесів виживання, коли це бажано. Наступним об'єктом винаходу є застосування вищевказаних нових білків, їх ізоформ, аналогів, фрагментів та похідних для виділення та характеристики додаткових білків чи факторів, які можуть бути утягнені до регулювання запалення, шляхів клітинної загибелі чи клітинного виживання і впливу на їх активність, та/або для виділення та ідентифікації інших рецепторів чи інших клітинних білків у сигнальних процесі(ах) раніше чи пізніше, з якими ці нові білки, їх аналоги, фрагменти і похідні зв'язані і, отже, до дії яких вони також утягнені. Ще одним об'єктом винаходу є інгібітори, які можуть бути введені до клітин для зв'язування чи взаємодії з новими білками та їх можливими ізоформами, які можуть діяти для пригнічення запалення, процесів клітинної загибелі чи виживання, коли це бажано. Більш того, об'єктом даного винаходу є використання вищевказаних нових білків, їх ізоформ та аналогів, фрагментів та похідних як антигенів для одержання поліклональних та/або моноклональних антитіл. Антитіла, у свою чергу, можуть бути використані, наприклад, для очищення нових білків з різних джерел, таких як клітинні екстракти чи трансформовані клітинні лінії. Далі, ці антитіла можуть бути використані для діагностичних цілей, наприклад, для ідентифікації можливих захворювань, зв'язаних з невірним функціонуванням клітинних ефектів, опосередкованих безпосередньо за допомогою каспаз, кіназ, білків, які належать до сімейства BCL2, або білків TRAF, або опосередкованих за допомогою р55 ФНП рецептора, FAS/APO1 рецептора, або інших споріднених рецепторів та асоційованих з ними клітинних білків (наприклад, RAIDD, MORT-1, TRADD, RIP), які діють прямо чи опосередковано для модуляції/опосередковування внутрішньоклітинних процесів через взаємодію з білками TRAF, каспазами, кіназами чи членами сімейства білків BCL2. Наступним об'єктом винаходу є фармацевтичні композиції, що містять вищевказані нові білки, їх ізоформи чи аналоги, фрагменти чи похідні, а також фармацевтичні композиції, що містять вищевказані антитіла чи інші антагоністи. Згідно з даним винаходом був виділений новий білок, позначений В1 (спочатку позначений СВК «с-ІАРзв'язуюча кіназа», завдяки наявності деякої гомології з с-ІАР, див. приклад 1 нижче, але який тут та надалі називається «В1»), який має область продомену (CARD), кіназний домен та проміжну область між вищевказаними доменами CARD та кінази, і, отже, можливо, утягнений до модуляції запалення, шляхів клітинної загибелі та клітинного виживання, як детально описано далі. Як також пояснено тут нижче, модуляція за допомогою В1 шляхів клітинної загибелі чи виживання може бути позитивною (наростаючою/посилюючою) чи негативною (інгібіторною), у залежності від типу внутрішньоклітинних білків, з якими він взаємодіє. Відповідно, даний винахід стосується ДНК послідовності, що кодує білок В1, його ізоформи, фрагменти чи аналоги, де вищевказаний В1, його ізоформи, фрагменти чи аналоги здатні взаємодіяти з внутрішньоклітинними медіаторами чи модуляторами запалення, шляхів клітинної загибелі чи шляхів клітинного виживання прямо чи опосередковано, і де вказані В1, його ізоформи, фрагменти чи аналоги є внутрішньоклітинними модуляторами вказаного внутрішньоклітинного запалення, шляхів клітинної загибелі та/або клітинного виживання. Приклади вищевказаної послідовності ДНК за винаходом включають: (і) послідовність ДНК, обрану з групи, що включає: (a) послідовність кДНК, похідну кодуючої області нативного білка В1; (b) фрагмент послідовності (а), який кодує біологічно активний білок, здатний до модуляції запалення, шляхів клітинної загибелі чи клітинного виживання, або того та іншого; (c) послідовність ДНК, здатну гібридизувати послідовність (а) чи (b) у середньо-суворих умовах і яка кодує біологічно активний білок, аналог чи фрагмент, здатні до модуляції внутрішньоклітинного запалення, шляхів клітинної загибелі чи клітинного виживання, або того та іншого; (d) послідовність ДНК, яка є виродженою в результаті генетичного коду на послідовності ДНК, визначеній у (а)-(с), і яка кодує біологічно активний білок В1, аналог чи фрагмент, здатні до модуляції внутрішньоклітинного запалення, шляхів клітинної загибелі чи клітинного виживання, або того та іншого; (іі) послідовність ДНК, як вказано вище, що містить принаймні частину послідовності, зображеної на Фіг.3, і кодує принаймні один активний білок В1, ізоформу, аналог чи фрагмент. (ііі) послідовність ДНК, як вказано вище, що кодує білок В1, ізоформу, аналог чи фрагмент, яка має принаймні частину амінокислотної послідовності, зображеної на Фіг.3. За іншим аспектом, винахід стосується вектора, що містить будь-яку з вищевказаних послідовностей ДНК за винаходом, який може бути експресований до клітин-хазяїв, обраних з прокаріотидних та еукаріотидних клітин; і трансформованих прокаріотидних та еукаріотидних клітин, що містять вказаний вектор. Виходячи з іншого аспекту винаходу, запропонований білок В1, його ізоформи, фрагменти, функціональні аналоги та похідні, кодовані послідовністю ДНК за винаходом, як вказано вище, де вказані білок, його ізоформи, фрагменти, аналоги та похідні, ймовірно, здатні модулювати внутрішньоклітинне запалення, шляхи клітинної загибелі чи клітинного виживання, або то та інше, безпосередньо чи опосередковано, шляхом асоціації з іншими внутрішньоклітинними модуляторами чи медіаторами цих шляхів. Прикладом білка за винаходом є білок В1, його ізоформа, фрагмент, аналоги та Похідні, де вказаний білок, його ізоформа, аналоги, фрагменти та похідні, мають принаймні частину амінокислотної послідовності, зображеної на Фіг.3. Винахід також стосується способу одержання білка В1, його ізоформи, фрагмента, аналога чи похідного, як вказано вище, який включає вирощування вищевказаних трансформованих клітин-хазяїв за умов, придатних для експресії вказаного білка, його ізоформи, фрагмента, аналога чи похідного, здійснення пост-трансляційної модифікації, якщо треба, для одержання вказаного білка, його ізоформи, фрагмента, аналога чи похідного, і виділення вказаного експресованого білка, його ізоформи, фрагмента, аналога чи похідного. За наступним аспектом, винахід стосується антитіл або їх активних фрагментів чи похідних, специфічних щодо білка В1, його ізоформи, аналога, фрагмента чи похідного за винаходом. За наступним аспектом, винахід стосується різних способів модуляції внутрішньоклітинних сигнальних шляхів, наприклад, таких: (і) спосіб модуляції чи медіації у клітинах активності запалення, шляхів внутрішньоклітинної загибелі чи клітинного виживання, або будь-якої іншої сигнальної активності, модульованої чи опосередкованої безпосередньо чи опосередковано за допомогою В1 чи інших молекул, до яких приєднуються чи взаємодіє іншим чином безпосередньо чи опосередковано білок В1, його ізоформа, аналог, фрагмент, чи похідне за винаходом, де вказаний спосіб включає обробку вказаних клітин шляхом введення до вказаних клітин одного чи кількох із вказаних білка В1, його ізоформи, аналога, фрагмента чи його похідного у вигляді, прийнятному для їх внутрішньоклітинного введення, або шляхом введення до вказаних клітин послідовності ДНК, що кодує вказані один чи кілька з білка В1, його ізоформи, аналога, фрагмента чи його похідного у вигляді придатного вектора, що несе вказану послідовність, де вказаний вектор є здатним здійснити інсерцію вказаної послідовності до вказаних клітин шляхом, за яким вказана послідовність експресована до вказаних клітин. (іі) спосіб, як вказано вище, де вказана обробка клітин включає введення до вказаних клітин послідовності ДНК, що кодує вказаний білок В1, ізоформу, аналог, фрагмент чи похідне у вигляді придатного вектора, що несе вказану послідовність, де вказаний вектор є здатним здійснювати інсерцію вказаної послідовності до вказаних клітин шляхом, за яким вказана послідовність експресована до вказаних клітин. (ііі) спосіб, як вказано вище, де вказана обробка вказаних клітин здійснюється трансфекцією вказаних клітин рекомбінантним вектором вірусу тварини, що включає такі стадії: (а) конструювання рекомбінантного вектора вірусу тварини, що несе послідовність, яка кодує вірусний поверхневий білок (ліганд), який є здатним зв'язуватись із специфічним рецептором поверхні клітини на поверхні вказаних клітин, які піддають обробці, і другу послідовність, що кодує білок, обраний з вказаних білка В1, ізоформ, аналогів, фрагментів та похідних, як вказано вище, яка, коли вона експресована до вказаних клітин, є здатною до модуляції/опосередковуванню активності запалення, шляхів клітинної загибелі чи клітинного виживання, безпосередньо чи опосередковано, або будь-якої іншої внутрішньоклітинної сигнальної активності, модульованої/опосередкованої іншими внутрішньоклітинними молекулами, з якими вказаний білок В1, ізоформи, аналоги, фрагменти та похідні взаємодіють безпосередньо чи опосередковано; і (b) інфікування вказаних клітин вказаним вектором (а). (iv) спосіб модуляції запалення, шляхів клітинної загибелі чи клітинного виживання, які модулюються безпосередньо чи опосередковано за допомогою В1, який включає обробку вказаних клітин антитілами чи їх активними фрагментами чи похідними, як вказано вище, де вказаною обробкою є використання придатної композиції, що містить вказані антитіла, їх активні фрагменти або похідні, щодо вказаних клітин, де, коли білок В1 або його частини вказаних клітин експоновані на зовнішньоклітинній поверхні, вказану композицію готують для зовнішньоклітинного нанесення, і, коли вказані білки В1 є внутрішньоклітинними, вказану композицію готують для внутрішньоклітинного використання. (ν) спосіб модуляції запалення, шляхів клітинної загибелі чи клітинного виживання у клітинах, які модулюються безпосередньо чи опосередковано за допомогою В1, що включає обробку вказаних клітин олігонуклеотидною послідовністю, яка кодує антисмислову послідовність для, принаймні, частини послідовності ДНК, що кодує білок В1 за винаходом, де вказана олігонуклєотидна послідовність є здатною блокувати експресію білка В1. (vi) спосіб, як вказано вище, де вказану олігонуклеотидну послідовність вводять до вказаних клітин за допомогою вірусу, вказаного у (іі) вище, де вказана друга послідовність вказаного вірусу кодує вказану олігонуклеотидну послідовність. (vii) спосіб модуляції запалення, шляхів клітинної загибелі, клітинного виживання чи інших, які модулюються безпосередньо чи опосередковано за допомогою В1, який включає використання рибозимного методу, за яким вектор, що кодує рибозимну послідовність, є здатним взаємодіяти з клітинною послідовністю мРНК, що кодує білок В1 за винаходом, вводять до вказаних клітин у такому вигляді, який дозволяє експресію вказаної рибозимної послідовності до вказаних клітин, і де. Коли вказана рибозимна послідовність експресована до вказаних клітин, вона взаємодіє з вказаною клітинною послідовністю мРНК, призводячи до інгібування експресії вказаного білка В1 у вказаних клітинах. За іншим аспектом, даний винахід стосується способу виділення та ідентифікації білків за винаходом, що мають гомологію з, або є здатними до прямої чи опосередкованої взаємодії з будь-якими білками, що мають продомен чи каспазний рекрутментний домен (CARD), або з іншими білками чи ферментами, утягненими до внутрішньоклітинної сигналізації, за допомогою кінази чи проміжних доменів, присутніх у білках за винаходом, що включає використання дріжджового двогібридного методу, за яким послідовність, що кодує вказаний білок вказаним CARD, кіназою та проміжними доменами, або принаймні з одним з цих доменів, переносять одним гібридним вектором, а послідовність з бібліотеки кДНК чи геномної ДНК переносять другим гібридним вектором, вектори потім використовують для трансформування дріжджових клітин-хазяїв і позитивні трансформовані клітини виділяють з подальшою екстракцією вказаного другого гібридного вектора з одержанням послідовності, що кодує білок, який зв'язується з вказаним CARD-, кіназута/або проміжний домен-вмісним білком. У ще одному аспекті даний винахід стосується фармацевтичної композиції для модуляції запалення, шляхів клітинної загибелі чи клітинного виживання, або інших, які модулюються безпосередньо чи опосередковано за допомогою В1, яка містить як активний інгредієнт принаймні один білок В1 за винаходом, його біологічно активні фрагменти, аналоги, похідні чи їх суміші. Прикладом втілення вищевказаної фармацевтичної композиції є композиція для модуляції запалення, шляхів клітинної загибелі, клітинного виживання або інших у клітинах, які модулюються безпосередньо чи опосередковано за допомогою В1, яка містить як активний інгредієнт рекомбінантний вектор вірусу тварини, що кодує білок, здатний до зв'язування з рецептором поверхні клітини, і який кодує принаймні один білок В1, ізоформу, активні фрагменти чи аналоги. Іншим прикладом втілення вищевказаної фармацевтичної композиції є композиція для модуляції запалення, шляхів клітинної загибелі, клітинного виживання чи інших у клітинах, які модулюються безпосередньо чи опосередковано за допомогою В1, що містить як активний інгредієнт олігонуклеотидну послідовність, що кодує антисмислову послідовність мРНК білка В1. Наступним прикладом втілення вищевказаної фармацевтичної композиції є композиція для профілактики або лікування патологічного стану, асоційованого з регуляцією апоптозу за допомогою однієї чи кількох молекул, до яких приєднаний білок В1, безпосередньо чи опосередковано, де вказана композиція містить ефективну кількість білка В1 чи молекули ДНК, що кодує її, або молекули, здатної перервати пряму чи опосередковану взаємодію вказаного білка В1 з однією чи кількома молекулами, з якими білок В1 зв'язується чи з якими він взаємодіє. Ще одним прикладом втілення вищевказаної фармацевтичної композиції є композиція для профілактики або лікування патологічного стану, асоційованого з регуляцією апоптозу за допомогою однієї чи кількох молекул, з якими білок В1 зв'язується безпосередньо чи опосередковано, де вказана композиція містить ефективну кількість білка В1, його ізоформи, фрагмента, аналога чи похідного, чи молекули ДНК, що кодує його, або молекули, здатної перервати безпосередню чи опосередковану взаємодію вказаного білка В1, його ізоформи, фрагмента, аналога чи похідного, з однією чи кількома молекулами, з якими зв'язується вказаний білок В1, ізоформа, фрагмент, аналог чи похідне. Додатковим прикладом втілення вищевказаної фармацевтичної композиції є композиція для профілактики чи лікування патологічного стану, асоційованого з регуляцією апоптозу за допомогою однієї чи кількох молекул, з якими зв'язується білок В1 безпосередньо чи опосередковано, де вказана композиція містить молекулу,здатну впливати на активність білкової кінази В1. В іншому аспекті за винаходом запропоновані такі терапевтичні способи: (і) спосіб профілактики чи лікування патологічного стану, асоційованого з регуляцією апоптозу за допомогою однієї чи кількох молекул, з якими зв'язується білок В1, безпосередньо чи опосередковано, де вказаний спосіб включає введення пацієнту, у разі потреби, ефективної кількості білка чи його ізоформи, фрагмента, аналога та похідного або суміші будь-чого з них, або молекули ДНК, що кодує його, або молекули, здатної перервати пряму чи опосередковану взаємодію вказаного білка В1, або його ізоформи, фрагмента, аналога чи похідного або суміші будь-чого з них, з однією чи кількома молекулами, з якими білок В1, його ізоформа, фрагмент, аналог чи його похідна або суміш будь-чого з них, зв'язується безпосередньо чи опосередковано. (іі) Спосіб модуляції процесів запалення, процесів апоптозу чи процесів програмованої клітинної загибелі (шляхи клітинної загибелі), до яких прямо чи опосередковано утягнений білок В1, що включає обробку вказаних клітин одним чи кількома білками В1, ізоформами, аналогами, фрагментами чи похідними, де вказана обробка вказаних клітин полягає у введенні до вказаних клітин вказаного одного чи кількох білків В1, ізоформ, аналогів, фрагментів чи похідних у вигляді, прийнятному для їх внутрішньоклітинного введення, або у введенні до вказаних клітин послідовності ДНК, що кодує вказані один чи кілька білків В1, ізоформ, аналогів, фрагментів чи похідних у вигляді придатного вектора, що несе вказану послідовність, де вказаний вектор є здатним здійснювати інсерцію вказаної послідовності до вказаних клітин так, що вказана послідовність експресується у вказаних клітинах. (ііі) Спосіб модуляції процесів клітинного виживання, до яких прямо чи опосередковано утягнений білок В1, який включає обробку вказаних клітин одним чи кількома білками В1, ізоформами, аналогами, фрагментами чи похідними, де вказана обробка клітин полягає у введенні до вказаних клітин вказаного одного чи кількох білків В1, ізоформ, аналогів, фрагментів чи похідних у вигляді, прийнятному для внутрішньоклітинного введення, або у введенні до вказаних клітин послідовності ДНК, що кодує вказані один чи кілька білків В1, ізоформ, аналогів, фрагментів чи похідних у вигляді придатного вектора. Що несе вказану послідовність, де вказаний вектор є здатним здійснювати інсерцію вказаної послідовності до вказаних клітин так, що вказана послідовність експресується у вказаних клітинах. Ще один аспект даного винаходу стосується таких методів скринінгу та способів ідентифікації і одержання різних лігандів: (і) Метод скринінгу ліганду, здатного до зв'язування з білком В1, який включає контактування афінної хроматографічної матриці, до якої приєднаний вказаний білок, з клітинним екстрактом, внаслідок чого ліганд приєднується до вказаної матриці, і елюювання, виділення та аналіз вказаного ліганду. (іі) Метод скринінгу ДНК послідовності, що кодує ліганд, здатний до зв'язування з білком В1, який включає використання дріжджового двогібридного методу, за яким послідовність, що кодує вказаний білок В1, переносять одним гібридним вектором, а послідовності з бібліотеки кДНК чи геномної ДНК переносять другим гібридним вектором, трансформацію дріжджових клітин-хазяїв вказаними векторами, виділення позитивних трансформованих клітин і екстракцію вказаного другого гібридного вектора з одержанням послідовності, яка кодує вказаний ліганд. (ііі) Спосіб ідентифікації та одержання ліганду, здатного до модуляції клітинної активності, модульованої/опосередкованої В1, що включає: а) скринінг ліганду, здатного до зв'язування з поліпептидом, що містить принаймні частину В1, яка має принаймні кілька амінокислотних залишків В1, зображених на Фіг.3, які включають по суті усі з продоменів (або CARD) B1; b) ідентифікацію та характеристику ліганду, іншого, ніж BCL2, TRAF2, чи частин рецептора сімейства ФНП/ФРН рецепторів або інші відомі білки, що мають продомен (CARD), як виявлено за допомогою вказаної стадії скринінгу, який має здатність до вказаного зв'язування; і c) одержання вказаного ліганду у по суті виділеній та очищеній формі. (iv) Спосіб ідентифікації та одержання ліганду, здатного до модуляції клітинної активності, модульованої чи опосередкованої білком В1, що включає: a) скринінг ліганду, здатного до зв'язування з поліпептидом, що містить принаймні карбоксикінцеву частину послідовності В1, зображену на Фіг.3, включаючи яродомен (CARD); b) ідентифікацію та характеристику ліганду, іншого, ніж BCL2, TRAF2, чи частини рецептора сімейства ФНП/ФРН рецепторів, або інші відомі білки, що мають продомен (CARD), як виявлено за допомогою вказаної стадії скринінгу, який має здатність до вказаного зв'язування; і c) одержання вказаного ліганду у по суті виділеній та очищеній формі. (ν) Спосіб ідентифікації та одержання ліганду, здатного до модуляції клітинної активності, модульованої чи опосередкованої білком В1, який включає: a) скринінг ліганду, здатного до зв'язування з принаймні N-кінцевою частиною послідовності В1, зображеної на Фіг.3, включаючи по суті увесь кіназний домен В1; b) ідентифікацію та характеристику ліганду, іншого, ніж BCL2, TRAF2, чи частини рецептора сімейства ФНП/ФРН рецепторів, або інших відомих білків, як виявлено за допомогою вказаної стадії скринінгу, який має здатність до вказаного зв'язування; і c) одержання вказаного ліганду у по суті виділеній та очищеній формі. (vi) Спосіб ідентифікації та одержання молекули, здатної до безпосередньої чи опосередкованої модуляції клітинної активності, модульованої/опосередкованої В1, який включає: a) скринінг молекули, здатної до модуляції активності, модульованої/опосередкованої В1; b) ідентифікацію та характеристику вказаної молекули; і c) одержання вказаної молекули у по суті виділеній та очищеній формі. (vii) Спосіб ідентифікації та одержання молекули, здатної до безпосередньої чи опосередкованої модуляції клітинної активності, модульованої/опосередкованої білком за винаходом, який включає: a) скринінг молекули, здатної до модуляції активності, модульованої/опосередкованої білком за винаходом; b) ідентифікацію та характеристику молекули; і c) одержання вказаної молекули у по суті виділеній та очищеній формі. Інші аспекти винаходу стануть зрозумілими з подальшого детального опису винаходу. На Фіг.1 показана діаграма, яка ілюструє структуру молекули TRAF2; На Фіг.2 показана схематична діаграма, яка ілюструє деякі білки, утягнені до запалення, шляхів клітинної загибелі та клітинного виживання (NF-kΒ активація); На Фіг.3(А,В) схематично показана визначена амінокислотна послідовність (А) білка В1 за даним винаходом і його визначена кодуюча нуклеотидна послідовність (В), де у амінокислотній послідовності показано кіназний домен В1 (область у рамці на N-кінці) та CARD-домен В1 (підкреслена область на Скінці). На Фіг.4 показано Northern аналіз експресії В1 у різних тканинах людини, який показує, що В1 експресований у більшості типів тканин людини; і На Фіг.5 схематично показані різні конструкції В1, досліджені на активність NF-kB, потенціювання клітинної загибелі та активацію JNK. На Фіг.6 показані результати визначення NF-kΒ активації, здійснені з різними конструкціями, представленими на Фіг.6 та у прикладі 3; На Фіг.7 показані результати визначення активації JNK1, здійснені з деякими з вищевказаних конструкцій; і На Фіг.8 показано, що В1 ауто-асоціюється і зв'язує TRAF1 in vivo. Даний винахід стосується, в одному аспекті, нового білка В1, який має продомен чи CARD домен (каспазний рекрутментний домен) і який містить білковий кіназний домен, подібний RIP-кіназному домену. Як такий білок В1 за даним винаходом ймовірно, є здатним взаємодіяти з деякими внутрішньоклітинними білками, утягненими до запалення, шляхи клітинної загибелі (апоптоз) та клітинного виживання (NF-kΒ активація). Ця взаємодія може бути шляхом зв'язування різних білків або, з іншого боку, взаємодією з ними через продомен (CARD), або воно може бути шляхом активності кіназного домену, або у той самий час може проходити взаємодія обох типів. Наприклад, В1 може бути здатним рекрутувати ряд білків, що мають продомени (CARD) і потім фосфорилювати їх через кіназний домен. Подібним чином, В1 може правити за «причалюючий» чи рекрутментний білок за допомогою його продомену (CARD) для різних інших продоменвмісних білків, які можуть не бути субстратом для кіназного домену В1, або В1 може взаємодіяти з різними білками тільки через його кіназний домен, а не через його CARD домен. Крім того, як тут детально описано, результати аналізу зв'язування вказують, що новий білок В1 за винаходом ймовірно є здатним зв'язувати білок BCL2. Ця знахідка збільшує вірогідність того, що білок В1 може бути регулятором активності BCL2, особливо стосовно регуляції апоптозу. У початкових аналізах біологічної активності збільшується також можливість того, що білок В1 може інгібувати захисний ефект BCL2 щодо апоптозу. Це за спостереженнями відбувається через те, що білок В1 сам по собі не викликає клітинної загибелі, але діє для посилення клітинної загибелі при доданні до клітин з іншими індукторами клітинної загибелі, такими як, наприклад, FAS-R, р55 ΦΗΠ-R та RIP (вказане додання до клітин шляхом спів-трансформації з векторами, здатними експресувати до клітин В1, FAS-R, р55 ΦΗΠ-R чи RIP, див. у прикладі 2 нижче). Отже, зростає ймовірність того, що В1 може не діяти аналогічним шляхом на ΒΑΧ чи ВАК, які зі свого боку, у вигляді гомодимерів можуть викликати клітинну загибель (див. розділ «Передумови винаходу» вище), але скоріше, В1 ймовірно може діяти аналогічним шляхом на BAD, який служить для негативної регуляції BCL2 шляхом зв'язування BCL2 та попередження його зв'язування з ΒΑΧ або ВАК, таким чином призводячи до більш вільної ΒΑΧ та/або ВАК, які, у свою чергу, викликають зниження клітинної загибелі (див. розділ «Передумови винаходу» вище). Більш того, що стосується вищевказаної NF-kΒ активації і клітинного виживання, В1 ймовірно може також досягати помітної активності у збільшенні клітинної загибелі шляхом, який, можливо, викликає зменшення NF-kΒ активації, може бути шляхом кіназної активності В1, яка, ймовірно, може слугувати для модуляції різних білків (наприклад, NIK), необхідних для індукції активації NF-kΒ, з тим результатом, що буде протікати зменшена NF-kΒ активація, отже, буде зменшене клітинне виживання. У цьому відношенні цікаво відзначити, що коли В1 додають з індукторами клітинної загибелі, такими як FAS-R, р55 ΦΗΠ-R чи RIP, він збільшує їх активність клітинного лізису. Відомо, що як р55 ΦΗΠ-R, так і FAS-R, і, можливо, також RIP, крім індукування шляхів клітинної загибелі, при якому кульмінаційним моментом є підвищення каспазної активності, також індукують NF-kΒ активацію, яка, у певній області зводить нанівець індуковану клітинну загибель. У деяких клітинах спостерігалось навіть, що ФНП не вбиває клітини, що притаманно індукції активації NF-kΒ за допомогою рецепторів ФНП, а не рахунок порушення цими рецепторами коіндукованих шляхів клітинної загибелі, таким чином, що у цих клітинах NF-kB опосередковані шляхи виживання клітин є явно більш активними, ніж шляхи клітинної загибелі. Таким чином, блокуванням NF-kΒ індукції можна було б збільшити клітинний лізис, опосередкований, наприклад, FAS-R, р55 ФНП, RIP, а білок В1 за винаходом, ймовірно, може виконувати цю функцію і викликати збільшення клітинної загибелі, що спостерігається при його доданні до FAS-R, р55-ФНП-Я чи RIP. З точки зору вищевказаного, ясно, що В1, ймовірно, може різним шляхом регулювати процеси запалення, шляхів клітинної загибелі чи клітинного виживання, і може навіть робити це одночасно. Наприклад, В1, ймовірно, може інгібувати NF-kB активацію, або В1, ймовірно, може навіть діяти на інші внутрішньоклітинні білки, утягнені до шляху клітинної загибелі чи клітинного виживання, незалежно від його можливої дії на NF-kΒ або на додаток до його можливої дії на NF-kB. Отже, виявляється, що В1 може мати здатність модулювати широку область внутрішньоклітинних білків, зокрема, таких, які утягнені до запалення, шляхів клітинної загибелі та клітинного виживання. Як детально описано далі, ряд відомих внутрішньоклітинних білків мають продомени (CARD), такі як, наприклад, різні каспазні ферменти, утягнені до протеолітичної деструкції клітин (шлях клітинної загибелі), включаючи ICE, ICH-1, Mch6 та інші, а також різні адаптерні білки, також утягнені до шляхів клітинної загибелі, включаючи RAIDD, с-ІАР1, С-ІАР2 та інші. Таким чином В1, ймовірно, може взаємодіяти безпосередньо чи опосередковано з різними каспазами через їх спільні CARD і тому, ймовірно, модулювати їх активність. Ця модуляція може бути позитивною, а саме, В1 може, ймовірно слугувати для концентрування різних каспаз і тому підвищує їх протеолітичну активність, що призводить до збільшення клітинної загибелі. Далі, В1 був виділений, використовуючи послідовність С-ІАР1, і В1 поділяє гомологію з С-ІАР1, який сам по собі є інгібітором апоптозу. С-ІАР1 має продомен (CARD) і може інгібувати апоптоз шляхом рекрутментних каспаз, попереджуючи таким чином їх активність. В1 може, ймовірно, тому взаємодіяти безпосередньо чи опосередковано з С-ІАР1 і приводити до пригнічення інгібування їм апоптозу, призводячи таким чином до посилення клітинної загибелі. Більш того, В1, ймовірно, може опосередковано чи безпосередньо взаємодіяти з різними каспазами через їх CARD, може також моделювати їх шляхом фосфорилювання через його кіназний домен, і таким шляхом В1 може посилювати каспазну активність. Більш опосередкованим чином, В1, ймовірно, є здатним взаємодіяти опосередковано чи безпосередньо з адаптерними білками, такими як RAIDD, С-ІАР1, С-ІАР2 та іншими, що мають CARD, тому, ймовірно, може взаємодіяти з іншими білками на більш початкових стадіях запалення, шляхів клітинної загибелі та клітинного виживання. Наприклад, RAIDD взаємодіє з іншими внутрішньоклітинними білками, такими як RIP та TRADD через спільні загибельні домени, які, у свою чергу, взаємодіють з білками, такими як MORT-1, p55-ФHП-R та FAS-R. Таким чином, ймовірно, взаємодіючи з RAIDD, B1 таким чином, ймовірно, опосередковано приєднаний до цих загибель-ефекторних рецепторів та білків. Подібним чином С-ІАР1 та С-ІАР2 взаємодіють з білком TRAF2, який, у свою чергу, взаємодіє з p75-ФHП-R та з MORT-1, p55-ФHП-R та FAS-R через взаємодію між TRAF2 та RIP, а також TRADD. Відповідно, В1, ймовірно, може бути опосередкованим модулятором процесів клітинної загибелі, будучи опосередковано приєднаним до вищевказаних адаптерних білків, ефекторних білків та рецепторів. Ця опосередкована модуляція може бути позитивною, наприклад, вона може приводити до посилення клітинної загибелі. Більш того, завдяки тому, що В1, ймовірно, є здатним взаємодіяти, принаймні, опосередковано (через С-ІАР1) з TRAF2, зростає ймовірність утягнення В1 до шляху клітинного виживання, який зв'язаний з індукцією NF-kΒ активації. Зараз відомо, що TRAF2 зв'язується безпосередньо з NIK [Malinin et aІ., 1997], який безпосередньо утягнений до індукції NF-kΒ активації і, таким чином, клітинного виживання. Відповідно, через здатність модулювати TRAF2 опосередковано, В1 може бути також здатним модулювати шляхи клітинного виживання. Далі, завдяки своєму кіназному домену В1, можливо, є здатним навіть більш безпосередньо бути утягненим до МАР кіназного шляху (яким слідує NIK), що призводить до індукції NF-kΒ активації та клітинного виживання. Однак, як вказано вище, з точки зору того факту, що В1 приводить до посилення клітинної загибелі, може бути, що В1 відіграє негативну роль у модуляції процесу клітинного виживання, а саме, В1, імовірно, може модулювати TRAF2, або В1, ймовірно, може бути безпосередньо утягненим до МАР кіназного шляху, але, таким чином, що приводить до зменшення NF-kΒ активації. Також можливо, що В1 відіграє центральну роль у модуляції внутрішньоклітинних сигнальних шляхів, зокрема, запалення, шляхів клітинної загибелі та клітинного виживання, і як такий В1 може слугувати для модуляції цього таким чином, що може зсунути рівновагу від індукції клітинного виживання до індукції клітинної загибелі і відповідності з посилюючою дією В1 на індукцію клітинної загибелі, що спостерігається (див. приклад 2). Тому В1 може бути розглянутий як «модулятор внутрішньоклітинної сигнальної» активності безпосередньо чи опосередковано на різних компонентних білках, що створюють ці шляхи. Тому, розглядаючи можливі різні терапевтичні застосування В1, важливо зрозуміти, що у всіх випадках В1 може відігравати різноманітні ролі, а саме, він може посилити процеси клітинної загибелі, і у той самий час, ймовірно, може активно інгібувати індукцію NF-kΒ і, отже, інгібувати шлях клітинного виживання, або, у залежності від дійсних білків/ферментів, з якими зв'язується В1, та їх відносної кількості у клітині, В1 може, ймовірно, у деяких клітинах діяти як інгібітор шляхів клітинного виживання і, у інших, може, ймовірно, діяти як посилювач шляхів клітинної загибелі шляхом пригнічення інгібіторів клітинної загибелі. Отже, звичайно, як буде ясно з подальшого, коли бажано посилити клітинну загибель, наприклад, у пухлинах, ВІЧ інфікованих клітинах і тому подібне, можливо, використання В1 для досягнення цієї мети. Наприклад, В1 може бути введений до клітин безпосередньо чи для підвищення експресії В1 до клітин може бути введена молекула ДНК, що кодує В1. Подібним способом, у ситуаціях, коли бажано захистити клітини від клітинної загибелі, індукованої ФНП чи FAS-лігандом, наприклад, при різних запаленнях, аутоімунних захворюваннях, реакціях «трансплантат проти хазяїна» і тому подібне, і замість цього промотувати клітинне виживання, тоді для досягнення цієї мети, ймовірно, можуть бути використані антагоністи В1. Наприклад, можуть бути введені антагоністи В1, такі як анти-В1 антитіла, олігонуклеотиди, що мають антисмислові послідовності В1, рибозими з В1 послідовностями, або різні пептиди чи органічні молекули, призначені спеціально для інгібування активності В1. Отже, коли тут вказуються призначення В1, вони можуть бути у термінах модуляторної дії В1 на різні внутрішньоклітинні процеси чи захворювання, і слід урахувати з точки зору вищевказаного, що ця модуляція може бути позитивною (посилювач), як у випадку, коли розглядаються шляхи клітинної загибелі, або негативною (інгібітор), як у випадку, коли розглядаються шляхи виживання клітин. Даний винахід також стосується ДНК послідовностей, що кодують біологічно активні білки В1, а також ДНК послідовностей, що кодують їх біологічно активні аналоги, фрагменти та похідні, і білків В1, аналогів, фрагментів та похідних білків які кодовані послідовностями ДНК. Одержання таких аналогів, фрагментів та похідних здійснюють звичайними способами [див., наприклад, Sambrook et al., 1989], за якими у ДНК кодуючих послідовностях один чи кілька кодонів можуть бути видалені, додані чи заміщені на інші, з одержанням кодованих аналогів, що мають принаймні одну заміну амінокислотного залишку по відношенню до нативного білка. Прийнятними аналогами є такі, що зберігають, принаймні, продомен (CARD) чи кіназний домен В1 або, принаймні, активні частини будь-якого чи обох цих доменів, разом або без опосередковування будь-якого іншого зв'язування чи ферментної активності, наприклад, не зв'язуються чи іншим способом не взаємодіють, безпосередньо чи опосередковано, з наступним білком чи іншим фактором далі, або не каталізують сигнал-залежну реакцію (наприклад, кіназну реакцію). Таким чином, можуть бути одержані аналоги, які мають так званий домінант-негативний ефект, а саме, аналог, який є дефективним чи то при зв'язуванні, чи при взаємодії іншим способом з іншими білками через продомен, або у подальшому сигналі (ймовірно, також кіназна активність), що супроводжує таке зв'язування, як вказано вище. Такі аналоги можуть бути використані, наприклад, для модуляції запалення, шляхів клітинної загибелі чи виживання, як вказано вище, шляхом конкурування з природними білками В1. Подібним способом, можуть бути одержані так звані домінант-позитивні аналоги, які могли б правити для посилення дії В1. Вони могли б мати ті ж самі чи поліпшені притаманні В1 зв'язувальні властивості по відношенню до інших білків і ті ж самі чи поліпшені сигнальні властивості чи кіназну активність природних білків В1. Аналогічним способом біологічно активні фрагменти клонів за винаходом можуть бути одержані, як вказано вище, стосовно одержання аналогів. Придатними фрагментами послідовностей ДНК за винаходом є такі, що кодують білок чи поліпептид, зберігаючи здатність В1 зв'язуватись з іншими білками, або які можуть опосередковувати будь-яке інше зв'язування чи ферментну (кіназну) активність, як вказано вище. Відповідно, можуть бути одержані фрагменти кодованих білків за винаходом, які мають домінантнегативною чи домінант-позитивною дією, як вказано вище стосовно аналогів. Подібним способом, похідні можуть бути одержані звичайними модифікаціями бокових груп одного чи кількох амінокислотних залишків білків, їх аналогів чи фрагментів, або кон'югацією білків, їх аналогів чи фрагментів з іншою молекулою, наприклад, антитілом, ферментом, рецептором і т.д., як це добре відомо у цій області. Що стосується вищевказаних послідовностей ДНК за винаходом, які кодують білок В1, ізоформу, аналог, фрагмент чи похідне, як приклад втілення за винаходом включені також послідовності ДНК, здатні до гібридизації з послідовністю кДНК, одержаною з кодуючої області нативного білка В1, де така гібридизація здійснюється у середньо жорстких умовах, і де послідовності ДНК, що гібридизуються, кодують біологічно активний білок В1. Отже, ці послідовності ДНК, що гібридизуються, включають послідовності ДНК, які мають відносно високу гомологію по відношенню до нативної кДНК послідовності В1, і як такі представляють В1-подібні послідовності, які можуть бути, наприклад, похідними природних послідовностей, що кодують різні ізоформи білка В1, або поширеними у природі послідовностями, які кодують білки, що належать до групи В1-подібних послідовностей, які кодують білок, що має активність білків В1. Далі, ці послідовності можуть також, наприклад, включати не природні, одержані синтетичним шляхом послідовності, які є подібними до кДНК послідовності нативного білка В1, але включають ряд бажаних модифікацій. Такі синтетичні послідовності, отже, включають усі можливі послідовності, що кодують аналоги, фрагменти та похідні білків В1, які усі мають активність білка В1. Для одержання різних вищевказаних природних білок В1-подібних послідовностей, можуть бути використані стандартні методи скринінгу та виділення зразків природно-похідних ДНК чи РНК з різних тканин, використовуючи кДНК природного білка В1 або її частину як зонд [див., наприклад, стандартні методи, представлені Sambrook et al., 1989]. Таким саме чином для одержання вищевказаних різних синтетичних білок В1-подібних послідовностей, що кодують аналоги, фрагменти чи похідні білка В1, може бути використаний ряд стандартних методів, як це детально описано далі стосовно одержання таких аналогів, фрагментів та похідних. Поліпептид чи білок, «які по суті відповідають» білку В1, включають не лише білок В1, а також поліпептиди чи білки, які є аналогами білка В1. Аналогами, які по суті відповідають білку В1, є такі поліпептиди, у яких одна чи кілька з амінокислот амінокислотної послідовності білка В1 замінена іншою амінокислотою, видалена та/або включена, за умови, що одержаний білок виявляє по суті таку ж чи більшу біологічну активність як білок В1, якому він відповідає. З метою відповідності по суті білку В1, зміни у послідовності білків В1, такі як ізоформи, є звичайно відносно незначними. Хоч ряд змін може бути більше, ніж десять, краще є не більше десяти змін, ще краще, не більше п'яти, і найкраще, не більше трьох таких змін. Оскільки можуть бути використані будь-які методи для виявлення потенційно біологічно активних білків, які по суті відповідають білкам В1, одним таким методом є використання загальновідомих способів мутагенезу на ДНК, що кодує білок, які призводять до невеликих модифікацій. Білки, експресовані такими клонами, можуть потім бути піддані скринінгу на їх здатність приєднуватись до різних інших білків, що мають, наприклад, продомени (CARD), сайти, що зв'язують кіназу, або до самого В1, і модулювати активність цих інших білків чи самих В1 при модуляції/опосередковуванні вищевказаних внутрішньоклітинних шляхів. «Консервативними» змінами є такі зміни, від яких не можна очікувати зміни активності білка, і звичайно є першими для цілей скринінгу, оскільки вони не передбачають по суті зміни розміру, заряду чи конфігурації білка, і тому не передбачають зміни їх біологічних властивостей. Консервативні замісники білків В1 включають аналог, де принаймні один амінокислотний залишок у поліпептиді консервативно замінений іншою амінокислотою. Такі заміщення краще здійснюють у відповідності до такого переліку, як представлено у таблиці ІА, де замісники можуть бути визначені звичайним експериментом для забезпечення модифікованих структурних та функціональних властивостей синтезованої поліпептидної молекули, при збереженні характеристик біологічної активності білка В1. Таблиця ІА Початковий залишок Ala Arg Asn Asp Cys GІn Glu Gly His lІe Leu Lys Met Phe Ser Thr Trp Tyr Val Приклад заміщення Gly; Ser Lys GІn; His Glu Ser Asn Asp Ala; Pro Asn; GІn Leu; Val lІe; Val Arg; GІn; Glu Leu; Tyr; lІe met; Leu; Tyr Thr Ser Tyr Trp; Phe He; Leu Альтернативно, інша група заміщень у білку В1 представляє собою таку, у якій принаймні один амінокислотний залишок у поліпептиді видаляли і на його місце вставляли інший залишок у відповідності з наступною таблицею IB. Типи заміщень, які можуть бути зроблені у поліпептиді, можуть бути основані на аналізі частоти амінокислотних замін у гомологічному білку різних видів, таких як представлені у таблиці 1-2 Schulz et al., G.E., Principles of Protein Structure, Springer-Verlag, New York, NY, 1798 та Creighton, T.F., Proteins: Structure and Molecular Properties, W.H. Freeman & Co., San Francisco, CA, 1983. Базуючись на такому аналізі, альтернативні консервативні заміщення визначені тут як заміни усередині однієї з таких п'яти груп: ТАБЛИЦЯ ІВ 1. 2. 3. 4. 5. Невеликі аліфатичні, неполярні чи частково полярні залишки: Ala, Ser, Thr (Pro, Gly) Полярні, негативно заряджені залишки та їх аміди: Asp, Asn, Glu, GІn Полярні, позитивно заряджені залишки: His, Arg, Lys Великі аліфатичні неполярні залишки: Met, Leu, His, Val (Cys) Великі ароматичні залишки: Phe, Tyr, Trp Три амінокислотні залишки у круглих дужках, вказані вище, мають спеціальне призначення у білковій архітектоніці. Gly є єдиним залишком, що не має ніякого бокового ланцюга, і тому надає ланцюгу гнучкості. Він, однак, сприяє утворенню вторинної структури, іншої, ніж ab-спіраль. Pro, через свою незвичайну геометрію, сильно стримує ланцюг і, звичайно, сприяє утворенню структури складчастого b-шару, незважаючи на те, що у деяких випадках Cys може бути здатним до участі у формуванні дисульфідного зв'язку, який є важливим в укладанні білка. Відзначимо, що Schulz et al., supra, об'єднав би групи 1 та 2, вказані вище. Відзначимо також, що Туг, через його здатність утворювати водневі зв'язки, має значну спорідненість до Ser та Thr, і тому подібне. Консервативні амінокислотні заміщення, згідно даному винаходу, наприклад, вказані вище, є відомими у своїй області, і, як можна припустити, після амінокислотних заміщень зберігають біологічні та структурні властивості поліпептиду. Більшість делецій та заміщень, у відповідності до даного винаходу, є такими, що не чинять радикальних замін у характеристиках білка чи поліпептидної молекули. Термін «характеристики» править для визначення неспецифічним способом обох змін у вторинній структурі, наприклад, a-спіралі чи b-шару, а також змін у біологічній активності, наприклад, зв'язування з іншими білками з продоменами (CARD) чи кіназної активності та/або модуляції шляхів клітинної загибелі чи виживання, як вказано нижче. Приклади здійснення амінокислотних заміщень у білках, які можуть бути використані для одержання аналогів білків В1 для використання за даним винаходом, включають будь-які відомі стадії методу, такі як представлені у патентах США RE 33653, 4959314, 4588585 та 4737462, Mark et al., 5116943, Koths et al., 4965195, Namen et al.; 4879111, Chong et al.; і 5017691, Lee et al.; і заміщені лізином білки представлені у патенті США No. 4904584 (Shaw et al.). Крім консервативних заміщень, розглянутих вище, які не могли б значно змінити активність білка В1, до області даного винаходу належать чи то консервативні заміщення, чи менш консервативні і більш випадкові зміни, які ведуть до збільшення біологічної активності аналогів білка В1. Якщо повинен бути визначений точний ефект заміщення чи делеції, фахівцю у цій галузі зрозуміло, що дія заміщення (заміщень), делеції (делецій) і тому подібне, буде оцінена звичайними аналізами зв'язування та клітинної загибелі. Скринінг, що використовує таке стандартне дослідження, не включає надмірного експериментування. На генетичному рівні ці аналоги звичайно одержують шляхом сайт-спрямованого мутагенезу нуклеотидів ДНК, які кодують В1 білок, внаслідок чого одержують ДНК, яка кодує аналог, і після чого синтезують ДНК та експресують поліпептид у рекомбінантних клітинних культурах. Аналоги звичайно виявляють ту ж саму чи підвищену якісну біологічну активність, що й природні білки, Ausubel et al., Current Protocols in Molecular Biology, Greene Publications and Wiley Interscience, New York, NY, 1987-1995; Sambrook et al., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor, NY, 1989. Одержання В1 білка у відповідності з вказаним тут, або альтернативної нуклеотидної послідовності, що кодує той же поліпептид, але який відрізняється від природної послідовності завдяки змінам, обумовленим відомим виродженням генетичного коду, може бути здійснено за допомогою сайт-специфічного мутагенезу ДНК, яка кодує раніше одержаний аналог чи нативну версію білка В1. Сайт-специфічний мутагенез дозволяє одержати аналоги шляхом використання специфічних олігонуклеотидних послідовностей, які кодують послідовність ДНК бажаної мутації, а також достатнє число суміжних нуклеотидів, для забезпечення праймерної послідовності достатнього розміру та варіабельності послідовностей для утворення стабільного дуплекса на обох сайтах з'єднання делецій, які перетинають. Звичайно, праймер, що містить від біля 20 до 25 нуклеотидів у довжину, є кращим, при цьому він має від біля 5 до 10 комплементуючих нуклеотидів на кожній послідовності, що змінюється. Звичайно методика сайтспецифічного мутагенезу є добре відомою у цій області, як показано на прикладах у публікаціях, таких як Adelman et al., DNA 2:183 (1983), опис якої включено сюди як посилання. Як буде зрозуміло, у методі сайт-специфічного мутагенезу звичайно використовується вектор фага, який існує як у одноланцюговому і дволанцюговому вигляді. Типові вектори, що використовуються у сайтспрямованому мутагенезі, включають вектори, такі як фаг М13, наприклад, як описано Messing et al., Third Qleveland Symposium on Macromolecules and Recombinant DNA, Editor A. Walton, Elsevier, Amsterdam (1981), вміст якої включено сюди як посилання. Ці фаги комерційно легко доступні і їх використання звичайно широко відомо фахівцю у цій області. Альтернативно, плазмідні вектори, які містять одноланцюговий фаговий початок реплікації [Veira et al, Meth. Enzymol., 153:3. 1987] можуть бути використані для одержання одноланцюгової ДНК. Звичайно, сайт-спрямований мутагенез у відповідності до вказаного тут, здійснюють одержанням спочатку одноланцюгового вектора, який включає до своєї послідовності послідовність ДНК, яка кодує релевантний поліпептид. Олігонуклеотидний праймер, що несе бажану мутовану послідовність, одержують синтетично за допомогою автоматизованого синтезу ДНК/олігонуклеотиду. Цей праймер потім відпалюють одноланцюговим вектором, що містить білкову послідовність, і піддають дії ДНК-полімеризуючих ферментів, таких як фрагмент полімерази І Klenow, E.coli для завершення синтезу ланцюга, що несе мутацію. Тому, мутована послідовність та другий ланцюг несуть бажану мутацію. Цей гетеродуплексний вектор використовують потім для трансформації придатних клітин, таких як клітини E.coli JM101, і вибирають клони, які включають рекомбінантні вектори, що несуть мутовану структуру послідовності. Після вибору такого клону, мутована послідовність білка В1 може бути видалена і поміщена до відповідного вектора, звичайно вектора переносу чи експресії такого виду, який може бути використаний для трансфекції відповідного хазяїна. Відповідно, ген чи нуклеїнова кислота, що кодують білок В1, також можуть бути встановлені, одержані, та/або модифіковані in vitro, in situ та/або in vivo, при використанні відомих методів ампліфікації ДНК чи РНК, таких як PCR та олігонуклеотидний хімічний синтез. PCR підходить для ампліфікації (підвищення числа) конкретних послідовностей ДНК за допомогою повторних реакцій полімеразних ДНК. Ця реакція може бути використана як заміщення при клонуванні; усе, що треба - це знання послідовності нуклеїнових кислот. З метою проведення PCR, конструюють праймери, які є комплементарними по відношенню до цікавлячої послідовності. Праймери потім генерують за допомогою автоматизованого синтезу ДНК. Оскільки праймери можуть бути сконструйовані для гібридизації у будь-якій частині гена, можуть бути створені такі умови, що невідповідності у комплементарному основному спарюванні можуть бути толерантними. Ампліфікація цих непридатних областей може призводити до синтезу продуктів мутагенезу, що в результаті веде до створення пептиду з новими властивостями (наприклад, сайтспрямований мутагенез). Див. також, наприклад, Ausubel, supra, Ch.16. Також, шляхом зв'язуючого комплементарного синтезу ДНК (кДНК), використовуючи зворотну транскриптазу, з PCR, як вихідний матеріал може бути використана РНК для синтезу позаклітинного домену пролактинового рецептора без клонування. Далі, PCR праймери можуть бути призначені для включення нових сайтів рестрикції чи інших ознак, таких як кінцеві кодони на кінцях генного сегмента, що піддається ампліфікації. Це розміщення сайтів рестрикції на кінцях 5' та 3' ампліфікованої генної послідовності дозволяє генним сегментам, що кодують білок В1 чи його фрагмент, бути спеціально сконструйованими для лігування інших послідовностей та/або клонуючих сайтів у векторах. PCR та інші способи ампліфікації РНК та/або ДНК добре відомі у цій області і можуть використовуватись згідно з даним винаходом без надмірного експериментування, базуючись на дослідженнях та описі, представленому тут. Відомі способи ампліфікації ДНК чи РНК включають, але не обмежуються цим, полімеразно-ланцюгову реакцію (PCR) та споріднені процеси ампліфікації [див., наприклад, патенти США №№ 4683195, 4683202, 4800159, 4965188, Mullis et al.; 4795699 та 4921794, Tabor et al.; 5142033, Innis; 5122464, Wilson et aІ.; 5091310, Innis; 5066584, Gyllensten et al.; 4889818, Gelfand et al.; 4994370, Silver et al.; 4766067, Biswas; 4656134, Ringold; і Innis et al., eds., PCR Protocols: A Guide to Method and Applications] та РНК опосередковану ампліфікацію, при якій використовується антисмислова РНК по відношенню до цільової послідовності як матриця для синтезу дволанцюгової ДНК [патент США №5130238, Malek et al., з торговою маркою NASBA]; і імуно-PCR, яка об'єднує використання ампліфікації ДНК з міченням антитіла [Ruzicka et al., Science 260:487 (1993); Sano et al., Science 258:120 (1992); Sano et al., Bioiechniques 9:1378 (1991)], повний вміст цих патентів та посилань включено сюди як посилання. Аналогічним способом, біологічно активні фрагменти В1 або його ізоформи можуть бути одержані, як вказано вище, стосовно аналогів білків В1. Придатними фрагментами білків В1 є такі, які принаймні зберігають зв'язувальну здатність, що відноситься до продомену, або кіназну активність, і які можуть прямо чи опосередковано опосередковувати біологічну активність різних інших білків чи внутрішньоклітинних шляхів, асоційованих з В1 білками. Відповідно, можуть бути одержані фрагменти білка В1, які мають домінант-негативний чи домінант-позитивний ефект, як вказано вище, щодо аналогів. Слід відзначити, що ці фрагменти представляють особливий клас аналогів за винаходом, а саме, вони представляють собою частини білків В1, одержані з повної послідовності білка В1, де кожна така частина чи фрагмент виявляє яку-небудь вищевказану активність. Таким фрагментом, наприклад, може бути пептид. Аналогічно, похідні можуть бути одержані стандартними модифікаціями бокових груп одного чи кількох амінокислотних залишків білка В1, його аналогів чи фрагментів, або кон'югацією білка В1, його аналогів чи фрагментів, з іншою молекулою, наприклад, антитілом, ферментом, рецептором і т.п., добре відомими у цій області. Відповідно, термін «похідні», як тут використовується, охоплює похідні, які можуть бути одержані з функціональних груп, які представлені у вигляді бокових ланцюгів на залишках або на N- чи с-кінцевих групах за допомогою засобів, відомих у цій області способом, включені до даного винаходу. Похідні можуть мати хімічні групи, такі, як вуглеводні чи фосфатні залишки, за умови, що така фракція має таку ж чи більш високу біологічну активність, ніжу білка В1. Наприклад, похідні можуть включати аліфатичні складні ефіри карбоксильних груп, аміди карбоксильних груп взаємодією з аміаком чи з первинними або вторинними амінами, N-ацильними похідними чи вільними аміногрупами амінокислотних залишків, утворених з ацильними групами (наприклад, алаканоїльні чи карбоциклічні ароїльні групи) або О-ацильними похідними вільних гідроксильних груп (наприклад, їх серильні або треонільні залишки), утворені з ацильними групами. Термін «похідне» призначений для включення лише таких похідних, які не міняють одну амінокислоту на іншу з двадцяти основних амінокислот, що звичайно зустрічаються. Білок В1 представляє собою білок чи поліпептид, тобто послідовність амінокислотних залишків. Припускається, що поліпептид, який складається з більшої послідовності, яка включає повну послідовність білка В1 у відповідності з даним тут визначенням, вважається таким поліпептидом доти, доки ці доповнення не впливають на основні чи нові характеристики винаходу, тобто якщо вони чи то зберігають, чи то підвищують біологічну активність білка В1, або може бути розщеплений для видалення білка або поліпептиду, який має біологічну активність білка В1. Так, наприклад, даний винахід передбачає включення гібридних білків білка В1 з іншими амінокислотами чи пептидами. Нові білки В1, їх аналоги, фрагменти та похідні мають ряд можливих застосувань, як вказано вище і далі, наприклад: (і) Вони можуть бути використані для модуляції шляхів клітинного виживання за допомогою прямої чи опосередкованої модуляції внутрішньоклітинних білків, з якими вони зв'язуються. В ситуації, де посилена активність цих шляхів є небажаною, тобто бажано інгібувати їх у бік посилення шляхів клітинної загибелі, наприклад, так, як при протипухлинному чи імуностимулювальному застосуванні, тоді бажано, щоб ця модуляція за допомогою В1, його ізоформ, аналогів, фрагментів чи похідних була інгібіторною. У цьому випадку, білки за винаходом, їх аналоги, фрагменти чи похідні, коли вони є інгібіторами шляхів клітинного виживання, можуть бути введені до клітин стандартними способами, відомими per se. Наприклад, оскільки білки, кодовані клонами ДНК за винаходом, є внутрішньоклітинними, і їх слід вводити лише до клітин, де це треба, необхідна система для специфічного введення цих білків до клітин. Одним шляхом здійснення цього є створення рекомбінантного вірусу тварини, наприклад, вірусу, похідного вірусу коров'ячої віспи, в ДНК якого будуть введені два такі гени: ген, що кодує ліганд, який зв'язується з білками на поверхні клітин, що специфічно експресовані клітинами, наприклад, такі, як білок gp 120 вірусу СНІД (ВІЧ), який специфічно зв'язується з деякими клітинами (CD4 лімфоцити і зв'язані з ними лейкози), або який-небудь інший ліганд, який специфічно зв'язується з клітинами, що несуть відомий рецептор, такий, щоб рекомбінований вірусний вектор міг зв'язувати такі клітини; і ген, що кодує білки за даним винаходом. Таким чином, експресія поверхнево-клітинно-зв'язуючого білка на поверхні вірусу буде метою вірусу, специфічного по відношенню до пухлинної клітини чи іншої клітини, яка несе рецептор, після чого білки, які кодують послідовності, будуть введені до клітини за допомогою вірусу, і після експресії у клітинах, будуть інгібувати шляхи клітинного виживання, що призводять до бажаної клітинної загибелі чи імуностимулювального ефекту у цих клітинах. Конструювання такого рекомбінантного вірусу тварини здійснюють звичайними способами [дивись, наприклад, Sambrook et al., 1989]. Іншою можливістю є введення послідовностей кодованих білків у вигляді олігонуклеотидів, які можуть бути абсорбовані клітинами і там експресовані. Аналогічно, якщо білки В1, ізоформи, аналоги, фрагменти чи похідні стимулюють або інакше посилюють процеси клітинної загибелі, тоді вони також можуть бути введені до клітин, як вказано вище, для посилення протипухлинної, імуностимулювальної чи іншої активності клітинної загибелі. (іі) Вони можуть бути використані для посилення чи додання шляхів клітинного виживання або, наприклад, у випадках, таких як ушкодження тканин при СНІДі, септичному шоку або реакції відторгнення «трансплантат проти хазяїна», у яких бажано блокувати шляхи клітинної загибелі або стимулювати шляхи клітинного виживання. В цій ситуації можливо у випадку, коли білки В1 дійсно інгібують процеси клітинного виживання або стимулюють, або інакше збільшують шляхи клітинної загибелі, наприклад, введення до клітин стандартними способами олігонуклеотидів, які мають антисмислову кодуючу послідовність для білків В1 за винаходом, які могли б ефективно блокувати трансляцію мРНК, що кодують білки і таким чином блокують їх експресію і ведуть до інгібування небажаного ефекту (клітинна загибель). Такі олігонуклеотиди можуть бути введені до клітин з використанням вищевказаних рекомбінантних вірусних підходів, при цьому друга послідовність, яку несе вірус, є олігонуклеотидною послідовністю. Друга можливість представляє собою використання антитіл, специфічних для білків за винаходом, для інгібування їх внутрішньоклітинної сигнальної активності. Ще одним шляхом інгібування небажаного ефекту є недавно розроблений рибозимний підхід. Рибозими є каталітичними РНК молекулами, які специфічно розщеплюють РНК. Рибозими можуть бути спроектовані для розщеплення мішеневої РНК на вибір, наприклад мРНК, що кодує білки В1 за винаходом. Такі рибозими мали б мати послідовність, специфічну для мРНК білків і могли б взаємодіяти з ними (комплементарне зв'язування) з подальшим розщепленням мРНК, в результаті одержуючи зменшення (чи повну втрату) експресії білків, при цьому рівень зниженої експресії залежить від рівня експресії рибозимів у клітині-мішені. Для введення рибозимів до клітин за вибором (наприклад, такі, які несуть послідовність білків В1), може бути використаний будь-який придатний вектор, наприклад, плазмідний вектор, вектори вірусу тварини (ретровірус), які звичайно використовуються з цією метою (дивись також (і) вище, де вірус містить як другу послідовність кДНК, що кодує рибозимну послідовність за вибором). [Огляди, методи і т.д. стосовно рибозим, дивись Chen et al., 1992; Zhao and Pick, 1993]. (iii) Вони можуть бути використані для виділення, ідентифікації та клонування інших білків, які здатні до зв'язування з ними, наприклад, інших білків, утягнених до внутрішньоклітинного запалення, шляхів клітинної загибелі чи клітинного виживання. Наприклад, ДНК послідовності, що кодують білки за винаходом, можуть бути використані у дріжджовій двогібридній системі, у якій кодовані білки будуть використовуватись як «принада» для виділення, клонування та ідентифікації з бібліотек кДНК чи геномних ДНК інших послідовностей («preys»), що кодують білки, які можуть зв'язуватись з клонами білків. Таким же чином, може бути також визначено, чи можуть білки за винаходом зв'язуватись з іншими клітинними білками, наприклад, іншими рецепторами ФНПФРН надсімейства рецепторів, або іншими членами сімейства BCL2. (iv) Кодовані білки, їх аналоги, фрагменти чи похідні також можуть бути використані для виділення, ідентифікації та клонування інших білків того ж класу, тобто, які мають продомени (CARD) або кіназні домени, або для функціонально споріднених білків, і утягнених до внутрішньоклітинних сигнальних процесів. У цій заявці може бути використана вищевказана дріжджова двогібридна система або може бути використана недавно розроблена система, що використовує не точну Southern гібридизацію з подальшим клонуванням PCR [Wilks et al., 1989]. (ν) Ще одним підходом до використання кодованих білків за винаходом, їх аналогів, фрагментів чи похідних є використання їх у методах афінної хроматографії, для виділення та ідентифікації інших білків чи факторів, з якими вони можуть зв'язуватись, наприклад, білків, що мають відношення до білків В1, або інших білків, або факторів, утягнених до внутрішньоклітинного сигнального процесу. У цій заявці білки, їх аналоги, фрагменти чи похідні за винаходом можуть бути індивідуально приєднані до матриксів афінної хроматографії і потім приведені до контакту з клітинними екстрактами чи виділеними білками або факторами, які гадано утягнені до внутрішньоклітинного сигнального процесу. У відповідності з методом афінної хроматографії інші білки чи фактори, які зв'язуються з білками, їх аналогами, фрагментами чи похідними за винаходом, можуть бути елюйовані, виділені та охарактеризовані. (vi) Як вказано вище, білки, їх аналоги, фрагменти чи похідні за винаходом можуть бути також використані як імуногени (антигени) для одержання специфічних до них антитіл. Ці антитіла можуть бути також використані для очищення білків за винаходом чи то від клітинних екстрактів, чи то від трансформованих клітинних ліній, що продукують їх, їх аналоги чи фрагменти. Далі ці антитіла можуть бути використані для діагностичних цілей при ідентифікації захворювань, пов'язаних з патологічним функціонуванням рецепторних систем чи запалення, шляхів клітинної загибелі чи виживання, у яких вони функціонують. Таким чином, такі захворювання повинні бути зв'язані із збоями у роботі сигнальної системи, що включає білки за винаходом, такі антитіла могли б слугувати важливим діагностичним інструментом. Термін «антитіло» припускає включення поліклональних антитіл, моноклональних антитіл (mAb), химерних антитіл, анти-ідіотипних (анти Id) антитіл до антитіл, у яких може бути введена мітка, в розчиненому чи зв'язаному вигляді, а також їх фрагментів, таких як, наприклад, Fab та F(ab')2 фрагменти, позбавлені Fc фрагмента інтактного антитіла, які здатні до зв'язування антигену. (vii) Антитіла, включаючи фрагменти антитіл, використані у винаході, можуть бути використані для кількісного чи якісного визначення клонів за винаходом у зразку або для визначення наявності клітин, які експресують клони за винаходом. Це може бути здійснено імунофлуоресцентними методами, використовуючи флуоресцентно мічене антитіло, зв'язане з визначенням методами світлової мікроскопії, цитометрії потоку чи флуорометричного визначення. Антитіла (або їх фрагменти), використані за винаходом, можуть бути застосовані гістологічно, як у імунофлуоресцентній чи імуноелектронній мікроскопії для in situ визначення клонів за винаходом. In situ визначення може бути виконано шляхом відбору гістологічного зразка у пацієнта і введення таким чином міченого антитіла за винаходом. Антитіло (чи фрагмент) краще вводять шляхом нанесення чи перекривання міченого антитіла (або фрагмента) біологічного зразка. Шляхом застосування такого способу, можливо визначити не лише присутність клонів, але також їх розподіл на досліджуваній тканині. При використанні даного винаходу фахівець у цій області легко визначить, що будь-який з великої різноманітності гістологічних способів (такий, як спосіб забарвлення) може бути модифікований для досягнення такого in situ визначення. Такі аналізи для клонів за винаходом звичайно включають інкубацію біологічного зразка, такого як біологічна рідина, екстракт тканини, свіже харвестовані клітини, такі як лімфоцити чи лейкоцити, або клітини, які були інкубовані у культурі тканини, у присутності визначаємо міченого антитіла, здатного до ідентифікації кодованих білків, і визначення антитіла будь-якими методами, добре відомими у цій області. (viii) Кодовані білки за винаходом також можуть біти використані як опосередковані модулятори ряду інших білків завдяки їх здатності зв'язуватись з рештою внутрішньоклітинних білків, де інші внутрішньоклітинні білки прямо зв'язуються з ще іншими внутрішньоклітинним білками або ще іншим внутрішньоклітинним доменом трансмембранного білка. З метою модуляції цих інших внутрішньоклітинних білків або внутрішньоклітинних доменів трансмембранних білків, білки за винаходом можуть бути введені до клітин різними шляхами, як вказано тут вище у (і) та (іі). Слід також відзначити, що виділення, ідентифікація та характеристика білків за винаходом можуть бути виконані будь яким, добре відомим стандартним скринінговим методом. Наприклад, один з цих скринінгових методів, дріжджовий двогібридний метод, був використаний для ідентифікації білків за винаходом. Аналогічно, можуть бути використані інші методи, такі, як афінна хроматографія, методи ДНК гібридизації і т.п., а також добре відомі в цій області, для виділення, ідентифікації та характеристики білків за винаходом, або для виділення, ідентифікації та характеристики додаткових білків, факторів, рецепторів і т.п., які здатні зв'язуватись з білками за винаходом. Більш того, білки, що зв'язуються з білками за винаходом, самі можуть бути використані аналогічним способом, шляхом, за яким використовували білки за винаходом, як вказано вище та далі, для виділення, ідентифікації та характеристики інших білків, факторів і тому подібне, які здатні до зв'язування з білками за даним винаходом, що зв'язуються, і які можуть представляти фактори, утягнені далі до асоційованих сигнальних процесів, або які можуть мати їх сигнальну активність і, отже, могли б представляти білки, утягнені до окремих сигнальних процесів. ДНК послідовності та кодовані білки за винаходом можуть бути одержані будь-яким стандартним способом рекомбінантної ДНК [дивись, наприклад, Sambrook, et al., 1989], у якому придатні еукаріотичні чи прокаріотичні клітини-хазяї трансформовані відповідними еукаріотичними чи прокаріотичними векторами, що містять кодуючі білки послідовності. Відповідно, даний винахід також стосується таких векторів експресії і трансформованих клітин-хазяїв для одержання білків за винаходом. Як вказано вище, ці білки включають також їх біологічно активні аналоги, фрагменти та похідні, і, таким чином, вектори, що кодують їх, включають також вектори, що кодують аналоги та фрагменти цих білків, і трансформовані клітини-хазяї включають такі, що продукують такі аналоги та фрагменти. Похідні цих білків представляють собою похідні, одержані стандартною модифікацією білків або їх аналогів, або фрагментів, продуковані трансформованими клітинами-хазяями. Даний винахід стосується також фармацевтичних композицій для модуляції дій, опосередкованих В1. Фармацевтичні композиції, що містять як активний інгредієнт будь-яке одне чи кілька з таких: (і) одну чи кілька послідовностей ДНК за винаходом, або їх частини, субклоновані до відповідного вектора експресії, (іі) білок за винаходом, його біологічно активні фрагменти, аналоги, похідні або їх суміш, (ііі) рекомбінантний вектор вірусу тварини, що кодує білок за винаходом, його біологічно активні фрагменти, аналоги чи похідне. Фармацевтичні композиції застосовують у відповідності до захворювання, яке піддають лікуванню, і у кількості найкращій для пацієнта, в залежності від ваги тіла та інших параметрів, які визначає лікуючий лікар. Як вказано вище, В1 може бути, ймовірно, опосередкованим модулятором TRAF2, і як такий, він, ймовірно, може бути утягнений до NF-kΒ активації через TRAF2-NIK взаємодію. Таким чином, В1, ймовірно, відіграє роль у шляхах клітинного виживання таким чином, що TRAF2 діє незалежно або у поєднанні з іншими білками (наприклад, р55 ФНП та р75 ФНП рецепторами, FAS/APO1 рецептором, MORT-1, RIP та TRADD). У цьому відношенні виявлена важливість створення ліків, які можуть посилювати чи інгібувати TRAF2-NIK взаємодію, як бажано. Наприклад, якщо необхідно збільшити клітинну цитотоксичність, індуковану ФНП, було б бажаним інгібувати NF-kΒ індукцію шляхом інгібування TRAF2-NIK взаємодії або шляхом вибіркового інгібування TRAF2 та/або NIK. Аналогічно, наприклад, якщо бажано інгібувати клітинну цитотоксичність, індуковану ФНП, було б бажаним збільшити NF-kB Індукцію шляхом посилення TRAF2-NIK взаємодії або шляхом посилення TRAF2-та/або NIK-специфічної NF-kΒ індукції. Існує багато захворювань, при яких такі ліки можуть бути дуже корисні. Поміж інших, (дивись також обговорення вище), гострі гепатити, при яких гостре ураження печінки, мабуть, відображає FAS/APO1 рецептор опосередковану загибель клітин печінки з подальшою індукцією Fas лігандом; аутоімунно індуковану клітинну загибель, таку як загибель b-клітин Лангерганса підшлункової залози в результаті діабету; клітинну загибель при відторгненні трансплантата (наприклад, нирки, серце та легені); загибель олігодендроцитів у мозку при розсіяному склерозі; і СНІД-інгібоване Т-клітинне «самогубство», яке є причиною проліферації вірусу СНІДу і, отже, захворювання СНІДом. У таких випадках було б бажаним інгібувати опосередкований FAS/APO1 рецептором шлях клітинної цитотоксичності (апоптоз) і збільшити опосередковану FAS/APO1 рецептором індукцію NF-kΒ за допомогою TRAF2 та TRAF2-NIK взаємодії. Одним із шляхів здійснення цього могло б бути збільшення кількості NIK у клітинах або підвищення кількості TRAF2 та NIK так, що NIK- чи TRAF2-NIK опосередкована індукція NF-kΒ активації збільшила б забезпечення більш високих рівнів NF-kB активації і, отже, клітинного виживання; або так, щоб пряма чи непряма взаємодія між FAS/APO1 рецептором та TRAF2 (або TRAF2-NIK) посилилось би, призводячи до зменшення FAS/APO1 рецепторної взаємодії з медіаторами клітинної цитотоксичності (наприклад, МАСН, дивись схему на Фіг.2) для забезпечення підвищення індукції NF-kΒ активації та клітинного виживання. Навпаки, у випадках, наприклад, пухлин та інфікованих клітин (дивись також обговорення вище) було б бажаним підвищити опосередковану FAS/APO1 рецептором клітинну цитотоксичність для підвищення клітинної загибелі. У цьому випадку було б бажаним інгібувати взаємодію FAS/APO1 рецептор-ЛТЧАР2 (або -TRAF2-NIK) та/або прямо інгібувати NIK, і, таким чином, знизити індукцію NF-kB активності. Оскільки білок В1 за винаходом, ймовірно, може взаємодіяти з TRAF2, то можна посилити чи блокувати цю взаємодію і, таким чином, посилити чи інгібувати активність TRAF2, зокрема, взаємодію TRAF2 з NIK та асоційовану індукцію NF-kB активації. Посилення чи інгібування взаємодії між В1 та TRAF2, ймовірно, може бути прямим або через інші білки (наприклад, С-ІАР1, С-ІАР2), які зв'язуються з TRAF2 і які, можливо, взаємодіють з В1 прямо чи опосередковано. Таким чином, зосередив увагу на білку В1 і модулюючи його можливу взаємодію (пряму чи опосередковану) з TRAF2, можливо також модулювати активність TRAF2 і, таким чином, ефекти FAS/APO1 (FAS-R), а також p55-ФHП-R, як вказано вище. Так само, як вказано вище, В1 ймовірно може діяти прямо на медіатори клітинної загибелі, а саме на різні каспазні ферменти, протеолітична активність яких веде до клітинної загибелі. Відповідно, вищевказані ефекти FAS/APO1 (FAS-R) або p55-ФHП-R можуть бути модульовані прямо чи опосередковано В1 через В1 можливу модуляцію каспаз (наприклад, МАСН та інші), які асоційовані з р55 ΦΗΠ-R, FAS-R чи зв'язуючим його білком MORT-1 і які, напевно, здійснюють апоптозні реакції, опосередковані таким чином. Так, якщо В1 взаємодіє з такими каспазами шляхом, який посилює їх активність, то таку взаємодію слід збільшити, якщо бажана клітинна загибель, як вказано вище, або слід інгібувати, якщо клітинна загибель небажана, як вказано вище. Таким чином, з точки зору вищевказаного, різні речовини, такі як пептиди, органічні сполуки, антитіла і т.п. можуть бути відібрані для одержання конкретних ліків, які здатні інгібувати можливу взаємодію між В1 та різними іншими білками, коли така взаємодія небажана. Такими ліками, ймовірно, були б такі, що специфічно розпізнають продомен (CARD) B1, наприклад, пептиди, органічні молекули, антитіла чи фрагменти антитіл, які специфічно зв'язуються з В1 CARD і перешкоджають його взаємодії з іншими CARDвмісними білками. Навпаки, якщо така взаємодія між В1 та іншими білками є бажаною, тоді вона може бути посилена збільшенням кількості В1 у клітинах стандартними методами, вказаними у (і) вище. Тут також можна підібрати різні конкретні ліки, які можуть посилювати активність В1 внутрішньоклітинно, або посилити його взаємодію з іншими білками. Крім того, як вказано вище, В1 також має кіназний домен, який може бути утягнений до його модуляторних ефектів запалення, шляхи клітинної загибелі чи виживання. Відповідно, цей кіназний домен може слугувати для зв'язування та фосфорилювання різних білків і у зв'язку з цим підвищувати чи знижати їх активність, і таким чином підвищувати чи знижати активність запалення, шляхів клітинної загибелі чи клітинного виживання, в залежності від ситуації. Відповідно, різні білки, органічні сполуки, антитіла і т.п., можуть бути підібрані для одержання конкретних ліків, які можуть інгібувати кіназну активність В1, коли це бажано, або для інгібування, або для посилення запалення, шляхів клітинної загибелі чи виживання. Не обмежувальні приклади того, як пептидні інгібітори В1 взаємодіють з іншими білками через його продомен чи кіназний домен, як вказано вище, розроблені і підібрані на основі попередніх досліджень з пептидних інгібіторів ICE чи І СЕ-подібних протеаз, на субстратній специфічності ICE і стратегіях для епітопного аналізу з використанням пептидного синтезу. Як виявлено, мінімальною вимогою до ефективного розщеплення пептиду за допомогою ICE, має бути включення чотирьох амінокислот зліва від сайта розщеплення при сильній перевазі аспарагінової кислоти у Р1 положенні і з метиламіном, розміщеним справа відносно Р, положення [Sleath et зі, 1990; Howard et al., 1991; Thornberry et al., 1992]. Крім того, пептид - флуорогенний субстрат - (тетрапептид) ацетил-Аsp-GІu-\/аІ-АSр-а-(4-метил-кумарил-7-амід), скорочено Ac-DEVD-AMC, відповідає послідовності у полі(АДФ-рибоза)полімеразі (PARP), як виявлено, розщеплюється у клітинах зразу після FAS-R стимуляції, а також інших апоптозних процесів [Kaufmann, 1989; Kaufmann et al., 1993; Lazebnik et al., 1994], і ефективно розщеплюється за допомогою СРР32 (член GED3/ICE сімейства протеаз) та МАСH протеаз. Тоді як, поява Asp у положенні Р1 субстрату є важливим, тетрапептиди, що мають Asp як четвертий амінокислотний залишок, і різні комбінації амінокислотних залишків у перших трьох положеннях, можуть бути швидко відібрані для зв'язування з активним сайтом протеаз, використовуючи, наприклад, метод, розроблений Гейсеном [Geysen, 1985; Geysen et al., 1987], де величезна кількість пептидів на твердих основах була відібрана для специфічної взаємодії з антитілами. Зв'язування МАСH протеаз із специфічними пептидами може бути визначено різними методами, добре відомими фахівцям у цій області, такі, як радіоізотопне мічення, і т.п. Показано, що цим методом Гейсена можна дослідити, принаймні, 4000 пептидів кожен робочий день. Аналогічним способом точна зв'язуюча область чи область гомології, яка визначає взаємодію між В1 та іншими білками, може бути пояснена і потім можуть бути відібрані пептиди, які можуть слугувати для блокування цієї взаємодіє, наприклад, пептиди, що синтезуються, які мають послідовність, подібну до зв'язуючої області або комплементарну до неї, яка може змагатися з природним В1 для зв'язування, або іншої взаємодії з іншими білками шляхів клітинної загибелі чи клітинного виживання через CARD чи кіназні домени, або, навіть, інтермедіаторний домен В1 між його CARD та кіназними доменами. Оскільки може бути зручним вказати пептидні інгібітори, які селективно інгібують В1 взаємодії без втручання фізіологічних процесів клітинної загибелі чи виживання, до яких утягнені інші члени внутрішньоклітинних сигнальних шляхів, пул пептидів, що зв'язуються з В1 у дослідженні, такий, як описаний вище, може бути потім синтезований у вигляді пептид-флуорогенного субстрату - для тестування селективного зв'язування В1 з такими іншими білками для відбору лише тих, що є специфічними по відношенню до В1. Пептиди, які визначені як специфічні для, наприклад, CARD чи кіназного домену В1, потім можуть бути модифіковані для посилення клітинної проникності і модуляції запалення, процесів клітинної загибелі чи клітинного виживання оборотно чи необоротно. Thornberry et al. (1994) повідомили, що тетрапептид (ацилокси)метилкетон Ac-Tyr-Val-Ala-Asp-CH2OC(O)-[2,6-(CF3)2]Ph є сильним інактиватором ICE. Аналогічно, Milligan et al. (1995) повідомили, що тетрапептидні інгібітори, що мають хлорметил кетонові (необоротно) чи альдегідні (оборотно) групи, інгібували ICE. Крім того, було показано, що бензилоксикарбоніл-Аsр-СН2ОС(О)-2,6-дихлорбензол (ДХБ) інгібує ICE [Mashima et al., 1995]. Відповідно, аналогічним способом, тетрапептиди, які селективно зв'язуються, наприклад, з CARD чи кіназним доменом В1, можуть бути модифіковані, наприклад, альдегідною групою, хлорметилкетоном, (ацилокси)метилкетоном чи СН2ОС(О)-ДХБ групою для створення пептиду-модулятора В1 активності. Далі, для поліпшення проникності, пептиди можуть бути, наприклад, хімічно модифіковані чи перетворені для посилення їх проникності скрізь клітинну мембрану та полегшення транспорту таких пептидів скрізь мембрану та до цитоплазми. Muranishi et al. (1991) повідомили про перетворюючий тиреотропін-рілізінг гормоні з лауриновою кислотою для утворення ліпофільного лауроїльного похідного з добрими характеристиками проникності скрізь клітинні мембрани. Zacharia et al. (1991) також повідомили про окиснення метіоніну до сульфоксиду та заміни пептидного зв'язку його кетометиленовим ізоефіром (СОСН2) для полегшення транспорту пептидів скрізь клітинну мембрану. Це лише кілька відомих модифікацій та похідних, які добре відомі фахівцям у цій області. Далі, лікарські чи пептидні інгібітори, які здатні до інгібуючої дії на, наприклад, шляхів клітинної загибелі чи виживання, перешкоджаючи можливій взаємодіє між В1 та будь-яким з білків, з якими він зв'язується через CARD, кіназний чи інтермедіаторний домени, можуть бути кон'юговані чи утворювати комплекс з молекулами, що полегшує вхід до клітини. У патенті США 5149782 описана кон'югація молекули для перенесення скрізь клітинну мембрану за допомогою мембранозв'язувального агента, такого як злиті поліпептиди, поліпептиди, що утворюють іонні канали, інші мембранні поліпептиди та довгий ланцюг жирних кислот, наприклад, міристинова кислота, пальмітинова кислота. Ці мембранозв'язувальні агенти вбудовуються до молекулярних кон'югатів у Ліпідний бішар клітинних мембран і полегшують їх вхід до цитоплазми. Low et al., патент США 5108921, дали огляд доступних способів для трансмембранної доставки молекул, таких як, але не обмежених цим, білки та нуклеїнові кислоти, за допомогою механізму рецепторної передачі ендоцитотичної активності. Ці рецепторні системи включають розпізнаючі галактозу, манозу, 6фосфат манозу, трансферин, азіалоглікопротеїн, транскобаломін (вітамін В12), a-2 макроглобуліни, інсулін та інші пептидні фактори росту, такі як епідермальний фактор росту (ЕФР). Low et al. показують, що живильні рецептори, такі як рецептори для біотину та фолату можуть бути успішно використані для посилення транспорту скрізь клітинну мембрану, завдяки положенню та множині біотинових та фолатних рецепторів на поверхні мембран більшості клітин та асоційованих з рецепторною передачею трансмембранних транспортних процесів. Таким чином, комплекс, що утворився між сполукою, яку треба доставити до цитоплазми, та лігандом, таким як біотин чи фолат, входить до контакту з клітинними мембраннозв'язаними біотиновими чи фолатними рецепторами для ініціації опосередкованого рецептором механізму трансмембранного транспорту і, таким чином, дає можливість для входу бажаної сполуки до клітини. ICE, як відомо, має здатність допускати вільні заміщення у положенні Р2 і ця допустимість до вільних заміщень була використана для розвитку сильного та високоселективного афінного мічення, що містить біотинову мітку (Thornberry et al., 1994). Отже, положення Р2, а також, ймовірно, N-кінець тетрапептидного інгібітора, можуть бути модифіковані чи перетворені, наприклад, доданням молекули біотину, для посилення проникності цих пептидів скрізь клітинну мембрану. Крім того, у цій області відомо, що поєднання бажаних пептидних послідовностей з провідною/сигнальною пептидною послідовністю для створення «химерного пептиду» допустить такий «химерний пептид» до переміщення скрізь клітинну мембрану до цитоплазми. Як зрозуміло фахівцю в області пептидів, пептидні інгібітори взаємодії В1 з іншими білками, як вказано вище, згідно з даним винаходом, припускають включення пептидоміметичних ліків чи інгібіторів, які можуть також бути швидко підібрані для зв'язування з CARD, кіназним чи інтермедіаторним доменом В1 для створення можливо більш стабільних інгібіторів. Також зрозуміло, що одні й ті самі засоби для полегшення чи посилення транспорту пептидних інгібіторів скрізь клітинні мембрани, як обговорювалось вище, також є застосовними до аналогів, фрагментів чи ізоформ В1, а також інших В1-апецифічних пептидів та білків (включаючи гібридні білки), які діють внутрішньоклітинно. Стосовно антитіл, вказаних тут, термін «антитіло» припускає включення поліклональних антитіл, моноклональних антитіл (mAb), химерних антитіл, антиідіотипних (анти-Id) антитіл до антитіл, які можуть бути помічені у вільній чи зв'язаній формі, а також фрагментів, одержаних будь-яким відомим способом, таким як, але не обмеженим, ферментативним розщепленням, пептидним синтезом чи рекомбінантними способами. Полїклональні антитіла представляють собою гетерогенні популяції молекул антитіл, одержаних з сироватки тварин, імунізованих антигеном. Моноклональні антитіла містять, по суті, гомогенну популяцію антитіл, специфічних до антигенів, популяції яких містять, в основному, епітоп зв'язуючі сайти. Mab можуть бути одержані відомими у цій області методами. Дивись, наприклад, Kohler та Milstein, Nature, 256:495-497 (1975); патент США №4376110; Ausubel et al., eds., Harlow та Lane ANTIBODIES: A LABORATORY MANUAL, Cold Spring Harbor Laboratory (1988); і Colligan et al., eds., Current Protocols in Immunology, Greene Publishing Assoc. та Wiley interscience N.Y., (1992-1996), вміст цих посилань включено тут повністю як посилання. Такі антитіла можуть бути антитілами будь-якого класу імуноглобулінів, включаючи IgG, IgM, IgE, IgA, GILD та будь-який їх підклас. Гібридома, що продукує mAb за даним винаходом, може бути культивована in vitro, in situ чи in vivo. Одержання високих титрів MAb in vivo чи in situ, робить кращим цей спосіб одержання у даний час. Химерні антитіла представляють собою молекули, різні частини яких одержують у різних видів тварин, такі, що мають варіабельну область, одержану з мишачого mAb і з константною областю імуноглобуліну людини. Химерні антитіла краще використовуються для зменшення імуногенності при застосуванні і для збільшення виходу при одержанні, наприклад, коли мишачі mAb мають більш високий вихід з гібридоми, але імуногенність більше у людських, так, що використовують людський/мишачий химерні mAb. Химерні антитіла та способи їх одержання відомі у цій області [Cabilly et al., Proc. Natl. Acad. Sci. США 81:3273-3277 (1984); Morrison et a!., Proc. Natl. Acad. Sci. США 81:6851-6855 (1984); Boulianne et al., Nature 312:643-646 (|984); Cabilly et al., європейська патентна заявка 125023 (опублікована 14 листопада, 1984); Neuberger et al., Nature 314:268-270 (1985); Taniguchi et al., європейська патентна заявка 171496 (опублікована 19 лютого, 1985); Morrison et al., європейська патентна заявка 173494 (опублікована 5 березня, 1986); Neuberger et al., заявка РСТ WO8601533 (опублікована 13 березня, 1986); Kudo et al., європейська патентна заявка 184187 (опублікована 11 червня, 1986); Sahagan et al., J. Immunol. 137:1066-1074 (1986); Robinson et al., міжнародна патентна заявка No. WO8702671 (опублікована 7 травня, 1987); Liu et al., Proc. Natl. Acad. Sci. США 84:3439-3443 (1987); Sun et al., Proc. Natl. Acad. Sci. США 84:214-218 (1987); Better et al., Science 240:1041-1043 (1988)]; і Harlow та Lane, ANTIBODIES: A LABORATORY MANUAL, supra. Повний перелік рефератів наведено тут як посилання. Антиідіотипне (анти-Id) антитіло представляє собою антитіло, яке розпізнає унікальні сайти зв'язування, звичайно, асоційовані з антиген-зв'язуючим сайтом антитіла. Id антитіло може бути одержано імунізацією тварин одного й того самого виду та генетичного типу (наприклад, мишача лінія), як джерело mAb, для якого готують анти-Id. Імунізовані тварини розпізнають і реагують на ідіотипні детермінанти імунизуючих антитіл продукуванням антитіла до цих ідіотипних детермінантів (анти-Id антитіло). Дивись, наприклад, патент США No. 4699880, який повністю включений тут як посилання. Анти-Id антитіло також може бути використано як «імуноген» для індукції Імунної відповіді у ще однієї іншої тварини, продукуючи так зване анти-анти-ld антитіло. Анти-анти-ld може бути епітопічно ідентично початковому mAb, яке індукує анти-Id. Тому, використовуючи антитіла до ідіотипних детермінантів mAb, можливо ідентифікувати інші клони, що експресують антитіла ідентичної специфічності. Відповідно, mAb, вироблені проти В1 білків, їх аналогів, фрагментів чи похідних даного винаходу, можуть бути використані для індукції анти-Id антитіл у придатних тварин, таких як BALB/c миші. Клітини селезінки мишей, імунізованих таким чином, використовують для одержання анти-Id гібридом, що секретують анти-Id mAb. Далі, анти-Id mAb можуть бути з'єднані з носієм, таким, як гемоціанін лімфи равлика (KLH), і використані для додаткової імунізації мишей BALB/c. Сироватка цих мишей буде містити анти-анти-ld антитіла, які мають зв'язувальні властивості вихідного mAb, специфічні для епітопа вищевказаного В1 білка, або його аналогів, фрагментів та похідних. Таким чином, анти-Id mAb мають власні ідіотипні епітопи, або «ідіотопи», структурно східні з епітопом, який оцінюють, таким, як GRB білок-а. Термін «антитіло» також заключає у собі обидві інтактні молекули, а також їх фрагменти, такі, як, наприклад, Fab та F(ab')2, які здатні зв'язувати антиген. Fab та F(ab')2 фрагменти, позбавлені фрагмента Fc інтактного антитіла, евакуюються швидше, ніж циркулюють, і можуть мати менше неспецифічного тканинного зв'язування, ніж інтактне антитіло [Wahl et al., J. Nucl. Med., 24:316-325 (1983)]. Слід враховувати, що Fab та F(ab')2 і інші фрагменти антитіл, використаних у даному винаході, можуть бути використані для виявлення та кількісного визначення білка В1, відповідними методами, описаними тут для інтактних молекул антитіл. Такі фрагменти звичайно одержують протеолітичним розщепленням, використовуючи ферменти, такі, як папаїн (для одержання Fab фрагментів) або пепсин (для одержання F(ab')2 фрагментів). Говорять, що антитіло «є здатним до зв'язування» молекули, якщо здатне до специфічної взаємодії з молекулою, таким чином зв'язувати молекулу з антитілом. Термін «епітоп» має відношення до тієї частини будь-якої молекули, яка може бути зв'язана антитілом, яка також може розпізнаватись цим антитілом. Епітопи чи «антигенні детермінанти», звичайно складаються з хімічно активних згрупованих на поверхні молекул, таких, як амінокислоти чи бокові ланцюги цукрів і мають тривимірні структурні характеристики, а також конкретні характеристики заряду. «Антиген» представляє собою молекулу чи частину молекули, здатну зв'язуватись антитілом, яке до того ж має здатність викликати у тварин вироблення антитіл, здатних до зв'язування з епітопом цього антигену. Антиген може мати один чи більше епітопів. Специфічні реакції, описані вище, показують, що антиген буде взаємодіяти високовибірковим способом з відповідним антитілом, а не з множиною Інших антитіл, які можуть бути викликані іншими антигенами. Антитіла, включаючи фрагменти, використані у даному винаході, можуть використовуватись для кількісного чи якісного визначення В1 білка у зразку або виявлення наявності клітин, які експресують В1 білок за даним винаходом. Це може бути виконано методом імунофлуоресценції з використанням флуоресцентно міченого антитіла (див. далі), у поєднанні із світловим мікроскопом, потоковою цитометрією чи флуорометричним визначенням. Антитіла (або їх фрагменти), що використовуються у даному винаході, можуть бути застосовані гістологічно, як у імунофлуоресцентній або імуноелектронній мікроскопії, для in situ визначення білка В1 за даним винаходом. In situ визначення іцоже бути здійснено взяттям у пацієнта гістологічного зразка, і наданням йому міченого антитіла даного винаходу. Антитіло (або фрагмент) краще використовувати накладенням чи перекриванням міченого антитіла (або фрагмента) з біологічним зразком. Використанням цього способу можна визначити не лише наявність В1 білка, але також його поширення у піддослідній тканині. Використовуючи даний винахід, середньому фахівцю зрозуміло, що будь-який з широкого різноманіття гістологічних методів (таких, як методи забарвлювання), може бути модифікований для досягнення такого виявлення in situ. Такі дослідження білка В1 за даним винаходом звичайно включають інкубування біологічного зразка, такого, як біологічна рідина, екстракт тканини, свіжозібрані клітини, такі, як лімфоцити чи лейкоцити, або клітини, які були інкубовані у культурі тканини, у присутності детектовано міченого антитіла, здатного ідентифікувати В1 білок, і визначення антитіла будь-яким із способів, добре відомих у цій області. Біологічний зразок може бути оброблений на твердофазовій основі чи носії, таких як нітроцелюлоза чи інша тверда основа чи носій, які можуть фіксувати клітини, клітинні частинки чи розчинні білки. Основа чи носій потім можуть бути промити придатними буферами, після чого йде обробка детектовано міченим антитілом згідно з даним винаходом, як вказано вище. Твердофазова основа чи носій після цього може бути промита буфером удруге для видалення незв'язаних антитіл. Кількість зв'язаної мітки на вищевказаній основі чи носії потім може бути визначене звичайними способами. «Твердофазова основа», «твердофазовий носій», «тверда основа», «твердий носій», «основа» чи «носій» позначають будь-яку основу чи носій, здатний зв'язувати антиген чи антитіла. Добре відомі основи чи носії включають скло, полістирол, поліпропілен, поліетилен, декстран, поліамідні амілази, природну та модифіковану целюлозу, поліакриламіди, габроз та магнетит. Носій за природою може бути чи то розчинним у певному ступеню або нерозчинним для цілей даного винаходу. Матеріал основи може мати будь-яку можливу структурну конфігурацію так, щоб зв'язана молекула була здатною зв'язувати антиген чи антитіло. Тому конфігурація основи чи носія може бути сферичною, як у кульки, циліндричною, як у внутрішньої поверхні пробірки, або зовнішньої поверхні стрижня. Альтернативно, поверхня може бути пласкою, такою як пластинка, аналіз на смужках (test strip), і так далі. Кращі основи чи носії включають кульки полістиролу. Фахівцям у цій області можуть бути відомі інші придатні носії для зв'язування антитіла чи антигену, або можна вияснити це звичайним експериментуванням. Зв'язувальна активність великої кількості даних антитіл за винаходом, як вказано вище, може бути визначена у відповідності з добре відомими способами. Фахівці у даній області визначать діючі оптимальні умови дослідження для кожного визначення звичайним експериментуванням. Інші стадії, такі як промивання, перемішування, струшування, фільтрація та подібні можуть бути додані у дослідження як звичайно або за потребою у окремих випадках. Один із шляхів, у якому антитіло згідно з даним винаходом може бути визначаємо помічене, представляє сполучення з ферментом і використання у імуноферментному аналізі (ІФА). Цей фермент, у свою чергу, при обробці далі відповідного субстрату, буде взаємодіяти з субстратом у такий самий спосіб, що одержать хімічну групу, яка може бути визначена, наприклад, спектрофотометрично, флуорометрично або візуальними способами. Ферменти, які можуть бути використані для визначаємо міченого антитіла, включають, але не обмежені цим, малатдегідрогеназу, нуклеазу стафілокока, дельта-5-стероїд ізомеразу, дріжджову алкогольдегідрогеназу, альфа-гліцерофосфат дегідрогеназу, триозофосфатізомеразу, пероксидазу звичайного хрону, лужну фосфатазу, аспарагіназу, оксидазу глюкози, бета-галактозидазу, рибонуклеазу, уреазу, каталазу, глюкозо-6-фосфат дегідрогеназу, глюкоамілазу та ацетилхолін естеразу. Виявлення може бути здійснене колориметричними методами, у яких використовують хромогенний субстрат для ферменту. Виявлення також можна здійснити візуальним порівнянням ступеню ферментативної реакції субстрату у порівнянні із стандартом, приготовленим подібним способом. Виявлення може бути виконано з використанням будь-якого з інших імунодосліджень. Наприклад, радіоактивним міченням антитіл чи фрагментів антитіл, можна виявити R-РТРазу, використовуючи радіоімунний аналіз (РІА). Детальний опис РІА можна знайти у Laboratory Techniques and Biochemistry in Molecular Biology, by Work, T.S. et al., North Holland Publishing Company, NY (1978) з посиланням, зокрема, до глави, під назвою «An Introduction to Radioimmune Assay and Related Techniques», написану Chard, Т., приведену тут як посилання. Радіоактивний ізотоп може бути виявлений використанням такого методу, як gпідрахунок чи сцинтиляційний підрахунок, або авторадіографією. Також можна мітити антитіло, згідно з даним винаходом, флуоресцентною сполукою. Коли флуоресцентно мічене антитіло піддають дії світла певної довжини хвилі, завдяки флуоресценції можна виявити його присутність. Поміж сполук, що найчастіше використовуються для флуоресцентного січення, є такі, як флуоресцин ізотіоціанат, родамін, пікоеритрин, пікоціанін, алофікоціанін, о-фталдегід та флуоресціамін. Антитіло може також бути визначаємо помічено з використанням металів, що випромінюють флуоресценцію, таких, як 152Е, або інші з ряду лантанідів. Ці метали можуть бути приєднані до антитіла з використанням таких метал хелатуючих груп, як діетилентриамін пентаоцтова кислота (ЕТРА). Антитіло також може бути визначаємо помічено сполученням з хемолюмінесцентною сполукою. Присутність хемолюмінесцентно міченого антитіла визначають за допомогою виявлення люмінесценції, яка виявляється в ході протікання хімічної реакції. Прикладами особливо корисних хімілюмінесцентно мічених сполук є: люмінол, ізолюмінол, імідазол, сіль та оксалат складного ефіру акридинію. Також можуть бути використані біолюмінесцентні сполуки для мічення антитіла за даним винаходом. Біолюмінесценція представляє собою тип люмінесценції, знайдений у біологічних системах, у яких білоккаталізатор збільшує ефективність хемілюмінесцентної реакції. Наявність біолюмінесцентного білка визначають виявленням люмінесценції. Важливими біолюмінесцентними сполуками для мічення є люциферин, люцифераза та екворин. Молекула антитіла за даним винаходом може бути застосована для використання в імунометричному аналізі, відомому також як «двосторонній» чи «сендвіч» аналіз. У звичайному імунометричному аналізі множину немічених антитіл (або фрагментів антитіл) зв'язано з твердою основою чи носієм і додають кількість визначаємо мічених розчинених антитіл для проведення визначення та/або кількісного визначення потрійного комплексу, утвореного між твердофазовим антитілом, антигеном та міченим антитілом. Звичайний, і кращий, імунометричний аналіз включає у себе «форвардні» дослідження, у яких антитіло, зв'язане з твердою фазою, спочатку контактує із зразком, що тестується, для видобування антигену із зразка шляхом утворення бінарного твердофазового комплексу антиген-антитіло. Після відповідного періоду інкубації, тверду основу чи носій промивають для видалення залишків рідкого зразка, включаючи непрореагований антиген, якщо вони є, і потім вводять у контакт з розчином, який містить невідому кількість міченого антитіла (яке функціонує як «молекула-репортер»). Після другого періоду інкубації, для можливості утворення комплексу міченого антитіла з антигеном, зв'язаним з твердою основою чи носієм через немічене антитіло, тверду основу чи носій промивають другий раз для видалення міченого антитіла, яке не провзаємодіяло. Другий тип «сендвіч» аналізу, який також може бути використаний з антигенами за даним винаходом, так звані «simultaneous» (одночасні) та «reverse» (обернені) дослідження. Одночасний аналіз включає одинарну стадію інкубації, оскільки антитіло, зв'язане з твердою основою чи носієм, і мічене антитіло, обидва одночасно додають до зразка, що тестується. Після завершення інкубації тверду основу чи носій промивають для видалення залишків рідкого зразка і міченого антитіла, що не утворило комплексу. Присутність міченого антитіла, асоційованого з твердою основою чи носієм, потім визначають як у звичайному «форвардному» сендвіч аналізі. У «reverse» аналізі, здійснюють постадійне додання спочатку розчину мічених антитіл до рідкого зразка, слідом за цим додають немічені антитіла, зв'язані з твердою основою чи носієм після відповідного періоду інкубації. Після другої інкубації, тверду фазу промивають звичайним способом для звільнення від залишків зразка, що тестується, і розчину мічених антитіл, які не провзаємодіяли. Визначення мічених антитіл, асоційованих з твердою основою чи носієм, потім визначають як у «simultaneous», так і у «forward» дослідженнях. Як вказано вище, даний винахід також стосується фармацевтичних композицій, що включають рекомбінантні вектори вірусу тварини, що кодують білки В1, при цьому такий вектор також кодує білки на поверхні вірусу, здатні до зв'язування поверхневих білків специфічних клітин-мішеней (наприклад, ракових клітин) для прямого включення послідовності білка В1 до клітин. Далі, фармацевтичні композиції за винаходом включають як активний інгредієнт (а) олігонуклеотидну послідовність, що кодує антисмислову послідовність послідовності білка В1, або (b) ліки, які блокують взаємодію В1 з іншими білками. Фармацевтичні композиції згідно з даним винаходом містять достатню кількість активного інгредієнта для досягнення поставленої мети. До того ж, фармацевтичні композиції можуть містити придатні фармацевтично прийнятні носії, включаючи наповнювачі та допоміжні речовини, які сприяють одержанню з активних сполук препаратів, які можуть бути використані фармацевтично і які можуть стабілізувати такі препарати при введенні суб'єкту, при необхідності у них, як добре відомо фахівцям у цій області. Гадають, що білок В1 та його ізоформи чи ізотипи, можливо, експресуються у різних тканинах на розрізняємо різних рівнях і, мабуть, також з різними структурами ізотипів аналогічним способом для експресії різних інших білків, утягнених до внутрішньоклітинних шляхів передачі сигналу, як показано у вищевказаних одночасно поданих патентних заявках, які одночасно розглядаються. Ці відмінності, ймовірно, можуть сприяти тканинноспецифічним особливостям відповіді на FAS/APO1 ліганд та ФНП. У випадку інших CED3/ICE гомологів [Wang et at., 1994; Alnemri et al., 1995], заявники раніше показали (у вищевказаних патентних заявках), що МАСH ізоформи, які містять неповні CED3/ICE області (наприклад, МАСНa3), як виявлено, мають інгібуючу дію на активність ко-експресованих МАСНa1 чи МАСНa2 молекул; вони також, як виявлено, блокують загибель, індуковану Fas/APO1 та p55-R (p55-ФHП-R). Експресія таких інгібіторних ізоформ у клітинах може складати механізм клітинного самозахисту проти Fas/APO1- та ФНПопосередкованої цитотоксичності. Широка різноманітність МАСH ізоформ, яке значно перевищує те, що спостерігається у будь-яких інших протеаз сімейства CED3/ICE, дозволяє, зокрема, добре регулювати функціональну активність МАСH ізоформ. Відомо також, що BCL2, BCL-XL та інші члени сімейства BCL2, експресуються і активні у тому чи іншому ступеню у різних типах клітин, викликаючи зміну чутливості різних клітин до індукованого апоптозу, наприклад, виживаність деяких клітин більш ймовірна, ніж інших [див. вищевказаний огляд Yang та Korsmeyer, 1996]. Як вказано вище, білки В1 або можливі ізоформи, ймовірно можуть мати різні ефекти у різних тканинах. Наприклад, такі різні ефекти, ймовірно, стосуються їх взаємодії з іншими білками запалення, шляхів клітинної загибелі чи клітинного виживання, та їх впливу, отже, на активність цих шляхів, зокрема на рівновагу між ними і чи не буде ця рівновага змінена тим чи іншим шляхом. Також ймовірно, що деякі з можливих ізоформ білка В1 виконують інші функції. Наприклад, В1, їх аналоги, або ізоформи, також можуть діяти як докінг-сайти для молекул, які утягнені до інших внутрішньоклітинних шляхів, не зв'язаних з вищевказаними шляхами клітинної загибелі чи виживання. Завдяки унікальній можливості Fas/APO1 та ФНП рецепторів викликати клітинну загибель, а також здатності рецепторів ФНП запускати інші процеси ураження тканин, відхилення у функціонуванні цих рецепторів могли бути надзвичайно шкідливими для організму. Дійсно, і надмірне, і недостатнє функціонування цих рецепторів, як було показано, сприяють патологічному прояву різних захворювань [Vassalli, 1992; Nagata and Golstein, 1995]. Ідентифікація молекул, які беруть участь у сигнальній активності рецепторів, і находження шляхів модуляції активності цих молекул, могли б направити нові терапевтичні підходи. З точки зору передбачуваної важливої ролі TRAP білків, і, отже, білка В1, який, ймовірно, може взаємодіяти з ними прямо чи опосередковано, або передбачуваної взаємодії між В1 та різними каспазами, що здається надзвичайно важливим для розробки ліків, які можуть впливати чи модулювати взаємодію між В1 та іншими білками, з якими він взаємодіє, і таким чином посилювати чи інгібувати клітинну загибель чи клітинне виживання, як треба. Даний винахід також стосується білків чи інших лігандів, які можуть зв'язуватись з білками В1 за винаходом і, таким чином, модулювати/опосередковувати активність білків В1. Такі білки чи ліганди можуть бути відібрані, виділені та одержані будь-яким з вищевказаних способів. Наприклад, може бути виділена деяка кількість нових лігандів, включаючи білки, здатні зв'язуватись з білками В1 за винаходом. Як детально описано вище, такі нові В1-зв'язуючі білки/ліганди можуть слугувати, наприклад, як інгібітори чи енхансери В1-медіаторної активності, і тому відігравати важливу роль у різних патологічних та інших станах, як детально описано вище. Іншою функцією таких В1-зв'язуючих білків/лігандів може бути їх використання як специфічних агентів для очищення білків В1, наприклад, за допомогою афінної хроматографії нові зв'язуючі білки/ліганди прикріплюються до придатних хроматографічних матриць для утворення твердого чи афінного носій/матриця розчину, скрізь який пропускають екстракт, або подібне, що містить білки В1, і, таким чином, полегшують їх очищення. Такі методи афінної хроматографії добре відомі і, як правило, є звичайними стандартними методиками у цій області. Більш того, усі вищевказані білки В1, аналоги, фрагменти,ізоформи та похідні за даним винаходом можуть бути використані для очищення за допомогою афінної хроматографії різних білків запалення, шляхів клітинної загибелі чи виживання, з якими вони зв'язуються. Наприклад, білки В1 і аналоги, фрагменти та їх мутеїни можуть бути використані для очищення В1-зв'язуючих білків афінною хроматографією. Такий спосіб для ідентифікації та одержання цих В1-зв'язуючих білків, включає стадію скринінгу, на якій білок В1, або, принаймні, його конкретна частина використовується як субстрат чи «затравка» для одержання білків чи будь-яких інших лігандів, здатних їх зв'язувати; потім йде стадія ідентифікації та характеристики таких білків/лігандів, одержаних таким способом; і одержання потім таких білків/лігандів у по суті виділеному та очищеному вигляді. Усі ці стадії добре відомі фахівцям у цій області і детально розглянуті тут вище та далі. Винахід далі буде описаний більш детально у подальших необмежувальних прикладах та у супроводі рисунків: Слід також відзначити, що методи: і) двогібридний скринінг та двогібридний аналіз експресії b галактозидази; (іі) індукована експресія, метаболічне нанесення мітки та імунопреципітація білків; (iii) in vitro зв'язування; (iv) визначення цитотоксичності; і (ν) аналізи Нозерн та послідовності, а також інші методи, використані у подальших прикладах, були детально розглянуті у попередніх публікаціях даних заявників по відношенню до інших внутрішньоклітинних сигнальних білків та шляхів [дивись, наприклад, Boldin et al., 1995а, 1995b, та Boldin et al., 1996]. Ці методи також представлені детально у ізраїльських заявках тих самих заявників, що одночасно розглядаються, №№ 114615, 114986, 115319, 116588, 117932 та 120367, а також у відповідній заявці РСТ No. РСТ US 96/10521). Відповідно, повний опис цих публікацій та патентних заявок включені сюди повністю, принаймні, що стосується експериментальних методик. Що стосується NIK білка та його ролі у активації NFkΒ і, отже, клітинного виживання та роль TRAF2 у цьому шляху клітинного виживання, наприклад, взаємодія між TRAF2 та p55-R, FAS-R, RIP та іншими білками, було детально розглянуто заявниками у вищевказаних ізраїльських заявках тих самих заявників, що одночасно розглядаються, та РСТ заявках і у Malinin et al, 1997. Приклад 1: Виділення, секвенування та часткова характеристика нового В1 білка З використанням різних методів, описаних у патентах тих самих заявників, вказаних вище, була виділена, секвенована та частково охарактеризована нова клонована послідовність ДНК. Ця послідовність ДНК кодує новий білок, спочатку позначений як с-ІАР, зв'язуючий кіназу (СВК) на основі гомології з с-ІАР білками та присутності кіназного домену, але тепер позначений як В1. Стисло, щоб у подальшому пояснити внутрішньоклітинну активність недавно знайдених гомологів СІАР1 та С-lАР2 інгібітора клітинного апоптозу (ІАР) [див. Rothe et al., 1995; Uren et al., 1996; Hofmann et al., 1997] і з яким внутрішньоклітинні білки взаємодіють, послідовності с-ІАР використовувались для скринінгу інших можливих гомологічних чи іншим способом зв'язаних послідовностей у різних базах даних, включаючи ті, які мають не характеризований (і не повністю секвеновані) мітки послідовності, що експресуються (ests). Таким чином, була визначена часткова послідовність нового клону, яка мала високу гомологію з С-ІАР1. З використанням цієї частинної послідовності, попередньо не охарактеризованої, були одержані PCR праймери для PCR клонування повнорозмірної послідовності ДНК цього нового клону за допомогою як матриці бібліотек ДНК, кДНК, одержаних комерційно. В результаті була одержана нова повнорозмірна ДНК, що кодує раніше невідомий білок, а саме, новий білок, позначений В1. Послідовність спочатку була визначена для В1 (ДНК та амінокислота). Подальший аналіз та визначення початкової послідовності В1 показали деякі відмінності в N-кінцевій частині амінокислотної послідовності (5'-кінець нуклеотидної послідовності), яка торкається перших 19 виведених амінокислотних залишків. Це подальше визначення послідовності та аналіз далі виведену В1 амінокислотну послідовність та її кодуючу нуклеотидну послідовність, як показано на Фіг.3А та В, відповідно. З аналізу амінокислотної послідовності на Фіг.3 випливає, що існує кіназний мотив на N-кінці білка, який кодується першими приблизно 1000 нуклеотидами відкритої рамки зчитування (ORF) нуклеотидної послідовності на Фіг.3. Далі, на С-кінці нуклеотидної послідовності знаходиться структура продомену (CARD), яка є спільною у ряду внутрішньоклітинних білків, утягнених до апоптичного сигнального шляху, наприклад, С-ІАР1, RAIDD[див. Duan and Dixit, 1997] та інші каспази, такі як ICE та ІСН-1. В амінокислотній послідовності В1, описаній на Фіг.3, показано N-кінець кіназного домену (область у рамці) та С-кінець CARD (підкреслена область). Між цими двома доменами знаходиться проміжний домен В1 білка. Вищевказаний кіназний домен В1 має високу гомологію (чи подібність) з відомими кіназами RAF-типу та RIP-кіназним доменом. Вищевказаний продомен В1 був також недавно позначений як CARD для «каспазного рекрутментного домену» [див. Hofmann et al., 1997] і призначений, щоб слугувати як область, через яку різні білки взаємодіють протягом апоптозної сигналізації внутрішньоклітинно. Наприклад, р55 ΦΗΠ-R, який не має, продомен (або CARD) взаємодіє з іншим внутрішньоклітинним білком TRADD (адаптерний білок) через загибельну область домену, представлену на обох цих білках. У свою чергу, TRADD може взаємодіяти з RIP та з RAIDD [такий додатковий адаптерний білок, див. також Hofmann та інші, 1997; Duan and Dixit, 1997; Wallach, 1997] усі з яких мають загибельні домени, такі, що, через загибельну область домену p55-ФHП-R можуть бути зв'язані у комплекс з RAIDD безпосередньо чи опосередковано. RAIDD має продомен (або CARD) який може взаємодіяти чи зв'язуватись з одним чи кількома каспазами, наприклад, ІСН-1 (каспаза2), і, можливо, іншими, і внаслідок цього може зв'язувати p55-ФHП-R з такими каспазами і викликати апоптоз через дію каспаз. Аналогічно, p75-ФHП-R може взаємодіяти з TRAF2 та TRAF1 білками за допомогою спільних мотивів, і TRAF білки можуть взаємодіяти з С-ІАР1 та С-ІАР2. Подібними методами [див. також Malinin et al., 1997, WO97/37016), на основі здатності FAS-R (Fas/APO1) взаємодіяти з MORT1 (FADD), який, у свою чергу, взаємодіє з TRADD (усі через їх спільні загибельні домени), і здатності TRADD взаємодіяти з TRAF2, MORT1 може тому бути зв'язаний з С-ІАР1, С-ІАР2 (через TRAF2) і внаслідок цього до ICE, Мch6 чи інших каспаз, або бути зв'язаним з ІСН-1, FLICE/MACH чи іншими каспазами (через TRADD-RIP-RAIDD взаємодії, вказані вище). Треба також вказати, що р55 ΦΗΠ-R може також бути зв'язаний з ICE, Мch6 та іншими такими каспазами через вищевказані відзначені TRADD-TRAF2-clAP1, C-IAP2-ICE, Мch6 взаємодії, також на основі здатності Р55 ΦΗΠ-R взаємодіяти з TRADD. Крім того, відомо, що TRAF2 також утягнений до внутрішньоклітинного шляху (або більш ніж одного шляху), що сприяє клітинному виживанню за допомогою індукції NF-kΒ активації. По цьому(цим) шляху(ам) здається, що NIK безпосередньо утягнений до фосфорилювання І-kB, що веде до І-kB дисоціації з NF-kΒ і внаслідок цього активації NF-kΒ, тим самим NF-kΒ може проникати до ядра та ініціювати транскрипцію різних генів, експресія яких пов'язана з клітинним виживанням (див. також розділ «Передумови винаходу» вище). Таким чином, TRAF2 утягнений до обох шляхів клітинної загибелі клітини та виживання, в залежності від яких білки переважно взаємодіють з TRAF2 у будь-який даний період у відповідь на різні зовнішні стимули (наприклад, ліганди зв'язують різні рецептори), клітина може піддаватися індукції клітинної загибелі чи клітинного виживання. Зрозуміло, що існує добра рівновага між різними внутрішньоклітинними сигнальними білками, яка може бути зсунута чи то у бік клітинної загибелі, чи то до шляху виживання клітин, і, як здається, TRAF2 є одним з ключових білків, що підтримує цю рівновагу, і є відповідальним за будь-яке зміщення у рівновазі одним шляхом чи іншим. На Фіг.1 показана схематично структура білка TRAF2 з його різними доменами, і на Фіг.2 схематично показані деякі можливі взаємодії між різними клітинними рецепторами та внутрішньоклітинними сигнальними білками та їх участь у шляхах клітинної загибелі чи клітинного виживання (NF-kΒ активація). Відповідно, виникає можливість того, що новий білок В1 за даним винаходом може мати важливу модуляторну роль у запаленні, шляхах клітинної загибелі та клітинного виживання. В1 має продомен (або CARD домен), який може, ймовірно, взаємодіяти, навіть опосередковано, з продоменом С-ІАР1, С-ІАР2, RAIDD та різними каспазами (ICE, ICH-1 і т.д.) і внаслідок цього може взаємодіяти навіть опосередковано з TRAF2 та різними білками, які взаємодіють безпосередньо чи опосередковано з TRAF2, включаючи RIP, TRADD, р75 ΦΗΠ-R, р55 ΦΗΠ-R, MORT-1 та FAS-R. В1 також має кіназний домен і як такий може, ймовірно, бути утягненим безпосередньо чи опосередковано до МАР-кіназного шляху, у якому, як здається, NIK є його членом, і внаслідок цього може бути також утягненим до шляху NF-kΒ активації. Крім того, В1 на основі гомології до С-ІАР1 може бути, ймовірно, модулятором активності С-ІАР1 (та СІАР2), шляхом модуляції С-ІАР1 біологічної активності чи шляхом модуляції зв'язування С-ІАР1 з іншими білками. У цьому відношенні (див. також приклад 2 нижче), В1може, ймовірно, діяти по збільшенню апоптозу, взаємодіючи навіть опосередковано з білками С-ІАР (С-ІАР1, С-ІАР2) і перериваючи чи іншим чином зменшуючи їх здатності до рекрутування каспаз та обмеження їх протеолітичної активності, так що у підсумку більша кількість каспаз буде вільно діяти протеолітично. Інша можливість - це те, що В1 через вищевказану можливу здатність взаємодіяти з різними медіаторами клітинної загибелі, безпосередньо чи опосередковано, включаючи TRAF-1 та TRAF2, RAIDD, RIP, TRADD, p55-ФHП-R, Р75-ΦΗΠ-R, MORT-1 та FAS-R; і з різними каспазами, може, ймовірно, слугувати для зв'язування цих білків з каспазами, і внаслідок цього, ймовірно, може слугувати як проміжний агент у шляху(ах) клітинної загибелі, яким(ому) належать ці білки. Також, В1 може бути важливим медіатором апоптозу. Наступна можливість полягає у ймовірній взаємодії (навіть опосередковано) В1 з вищевказаними білками с-ІАР, В1 може, ймовірно, попереджувати С-ІАР зв'язування чи взаємодію з TRAF2, і внаслідок цього може, ймовірно, блокувати TRAF2 активність по відношенню до МАР-кіназного шляху, наприклад, TRAF2-C-IAP взаємодії можуть бути важливими для TRAF2 взаємодій з NIK, і, якщо це відвернено шляхом В1 взаємодії з с-ІАР, тоді TRAF2-опосередкованa NF-kΒ активація може бути блокована, призводячи до меншого підвищення клітинного виживання і, ймовірно, збільшення клітинної загибелі. Ще одна подальша можливість - те, що В1 може діяти більш прямим способом при модуляції активності різних каспаз. Тому через взаємодії, безпосередні чи опосередковані, між продоменами (CARD домени) В1 та різними каспазами, В1 може, можливо, вести до збільшення активності цих ферментів і, внаслідок цього збільшувати цитотоксичність цих ферментів. Таким чином, В1 може бути прямим маніпулятором апоптозу шляхом рекрутування чи іншим способом активуючи каспази (див. також приклад 2 нижче). Додаткова можливість - це те, що В1 може діяти, щоб модулювати внутрішньоклітинні сигнальні шляхи, опосередковую чи клітинну загибель чи клітинне виживання шляхом зв'язування чи взаємодії з іншими доки ще невідомими білками. Цікаво відзначити (див. вище), що В1 має кіназний домен, подібний до RIP-кінази. RIP - також центральний білок, утягнений до рівноваги між шляхами клітинної загибелі та клітинного виживання на основі його здатності зв'язувати медіатори клітинної загибелі (наприклад, р55 ΦΗΠ-R, FAS-R, MORT-1, TRADD) та TRAF-2, і за допомогою цього NF-kΒ активацію та клітинне виживання (див. Фіг.2). RIP-кіназна активність може також бути фактором у цій точній рівновазі, в залежності від того, що є субстратом для цієї кінази, наприклад, які білки фосфорильовані за допомогою RIP, і чи впливає це на їх активність, спрямовану на збільшення апоптропної активності, зменшену апоптропну активність, збільшену NF-kΒ активацію чи зменшену NF-kB активацію. За аналогією, В1 може, ймовірно, також відігравати таку центральну роль, у якій його кіназна активність може бути важливою, в залежності від того, які білки є субстратами для такої кіназної активності. Приклад 2: Аналіз біологічної активності В1 білка (і) Аналіз початкового зв'язування для визначення, який відомий білок може зв'язуватись з В1 Використовуючи способи з методів по WO97/37016 для одержання та експресії ДНК конструкцій та аналіз дріжджового двогібридного зв'язування, була застосована конструкція В1, з якого був видалений кіназний домен, наприклад, зрізаний В1, що має лише проміжну область та С-кінцеву CARD область, для того, щоб дослідити його здатність зв'язувати різні відомі білки, утягнені до внутрішньоклітинних сигнальних шляхів (шляхи клітинної загибелі та виживання). Початкові, попередні результати (не показані), здається, вказують, що цей зрізаний В1 зв'язується з BCL2. (іі) Аналіз цитотоксичності клітин для визначення дії В1 на клітинну загибель чи клітинне виживання Використовуючи способи з методів за WO97/37016 для одержання ДНК конструкцій та трансфекції/трансформування клітин та визначення дії на клітинну загибель чи клітинне виживання експресованих продуктів цих конструкцій, ДНК конструкція, що кодує повнорозмірний білок В1, була використана для трансфекції клітини у культуру. Далі, у іншій серії експериментів, В1-кодуюча конструкція була використана для ко-трансфекції клітин з іншими конструкціями, що кодують, поміж інших, FAS-R, р55 ΦΗΠ-R та RIP. Результати, одержані з цих трансфекцій (не показані) вказують, що екепресований білок В1 сам по собі не викликає клітинну загибель. Однак, якщо В1 репресований разом з FAS-R, р55 ΦΗΠ-R чи RIP, він розширює рівень клітинної загибелі, індукованої цими відомими збудниками клітинної загибелі. Ці результати, які приймаються з урахуванням того, що, як вказано вище, (і) В1 може зв'язуватись з BGL2, підвищують можливість того, що В1 може слугувати як інгібітор BCL2 активності, наприклад, що В1 може попереджувати BCL2 активність до захисту клітин проти апоптозу (див. розділ «Передумови винаходу» вище), і також В1, напевно, є здатним до розширення шляхів клітинної загибелі, індукованих FAS-R, р55 ΦΗΠ-R та RIP і, можливо, іншими збудниками клітинної загибелі (як також вищевказані у розділі «Передумови винаходу»). У цьому відношенні, В1 може, ймовірно, діяти аналогічним способом на BAD білок, член BCL2 сімейства, який зв'язується з BCL2 та BCL-XL і, внаслідок цього призводить до підвищених рівнів ΒΑΧ та ВАК, які, як відомо, будуть безпосередньо утягнені до викликання клітинної загибелі. Інша можливість - це те, що В1, завдяки його кіназному домену, може фосфорилювати BCL2 на ділянках фосфорилювання, присутніх на BCL2, і, таким чином, може впливати на активність BCL2 до захисту клітин проти апоптозу, і, у кінцевому результаті, як це спостерігалось, В1 чинить посилюючу дію на індукцію клітинної загибелі. Крім того, можливо також, що В1 може, на додаток до або, незалежно від його можливої взаємодії з BCL2, здійснювати індукцію активації NF-κΒ, що відбувається через кіназну активність В12, яка діє по шляху, що веде до активації NF-kB, наприклад, В1 може можливо взаємодіяти з NIK чи іншими кіназами по шляху, частиною якого є NIK, або він може діяти на інші адаптерні білки, що беруть участь у цьому процесі, наприклад, TRAF2, способом, який призводить до зменшення активації NF-kΒ, і, у кінцевому підсумку, до зниженого клітинного виживання та підвищеної клітинної загибелі. Отже, стисло, виявляється, що В1 грає роль у модуляції внутрішньоклітинних сигнальних шляхів, незалежно від того, чи ведуть вони до запалення, клітинної загибелі чи клітинного виживання. В1 може тому розглядатись як «модулятор внутрішньоклітинної сигналізації», оскільки він напевно має здатність впливати на запалення, шляхи клітинної загибелі та клітинного виживання різними способами, наприклад, прямо (рекрутмент різних білків та їх активація чи інгібування, або через кіназну активність) чи опосередковано (через взаємодію з різними іншими проміжними ланками, наприклад, BCL2, і, можливо, також с-ІАР, і, внаслідок цього, з TRAF2, і т.д.; або RAIDD та внаслідок цього, з RIP, TRADD, і т.д.). Приклад 3: Додатковий аналіз біологічної активності В1 NF-kΒ активність, аналіз клітинної загибелі, Нозерн аналіз та аналіз JNK активності були проведені з використанням таких В1 та В1 мутантних конструкцій (див. Фіг.6). В1 (див. приклад 1) B1 mut, мутант В1, у якому лізин в положенні 47 був замінений на аланін. DCARD, B1 без CARD домену, одержаний в результаті PCR та клонування до векторів експресії DXba, B1 без CARD домену, але коротший на його 3'-кінці порівняно з DCARD, створений з використанням сайта рестрикції та клонування до векторів експресії DВат, подібний до DXba, і створений тим самим способом, використовуючи фермент рестрикції Ват, DNde, що містить частину кіназного домену та CARD домен, створений PCR і клонуванням до векторів експресії, і DΚ, створений PCR з використанням таких праймерів 1. 5'-СAGААTТСCAGAGTGTTTCAAGTGCCATTC; 2. 5'-AACTCGAGACTTACATGCTTTTATTTTGAA. Фрагмент PCR був клонований до векторів експресії і верифіцирован за допомогою секвенування. Оцінку NF-kΒ активації здійснювали репортерним генним аналізом, як описано у WO97/37016. Стисло, клітини були ко-трансфектовані з генною плазмідою ВІЧ LTR-люциферази (1мкг) і векторами експресії В1 та мутанта В1 (3мкг). Кількість трансфектованої ДНК підтримували сталим шляхом додання «пустого» вектора. Нерез 24 години після трансфекції клітини промивали PBS та лізували. Здійснювали аналіз з використанням люциферази, як описано у Current Protocols in Molecular Biology, Ausubel et al. Результати також представлені на Фіг.6. Аналіз клітинної загибелі був проведений шляхом вирощування клітин 293-Т у модифікованому Дульбеко мінімальному основному середовищі Ігла, доповненому 10% фетальної сироватки теляти, несуттєвими амінокислотами, 100 од./мл пеніциліну та 100 мкг/мл стрептоміцину. 293-Т клітини (5x105 клітин у 6см чашках) були миттєво трансфектовані методом осадження фосфату кальцію з використанням різних конструкцій кДНК разом з вектором експресії b-галактозидази. В експериментах, результати яких представлені на Фіг.6, кожну чашку трансфікували 1мкг р55 ΦΗΠ-R, RIP-чи TRADD конструкта, 1мкг відповідного В1 чи В1 мутантного конструкта (або пустого вектора, узятого як контроль), і 1мкг pSV-b-gal (Promega). Рівень клітинної загибелі у кінці періоду інкубації оцінювали шляхом визначення експресіїта bгалактозидази, як описано Boldin et al., 1996. Нозерн аналіз здійснювали звичайними методами [див., наприклад, Boldin et al., 1995], і цей аналіз показав, що В1 є присутнім у багатьох тканинах людини (Фіг.4). JNK активацію здійснювали шляхом швидкого трансфікування 293-Т клітин (5x105 клітин у 6см чашках) з використанням методу осадження фосфату кальцію з 1,5мкг PSR-HA-JNK1 конструкта (мічений НА епітопом вектор експресії JNK-1) і 4мкг вектора експресії кожного В1 та В1 мутантного конструкта. Через 24 години клітини лізували у лізисному буфері (20мМ Hepes pH 7,6, 10мМ EGTA, 40мМ b-гліцерофосфату, 2,0мМ МgСІ 2, 1мМ DTT і 1% ΝΡ-40) і білок Ha-JNK1 імунопреципітували з анти-НА антитілами [див., наприклад, Rothe et al., 1995b] (клон 12 CAS). Кіназний аналіз здійснювали у 30мкл кіназного буфера (20мМ Hepes, pH 7,6, 40мМ b-гліцерофосфату, 2,0мМ МдСІ 2, 2мМ DTT ТТ, 3нмоль АТР та 3мкКі g-Ρ32-АТР) при 30°С протягом 20 хвилин. Бактерицидно продукований білок GST-Jun (біля 10мкг) використовували як субстрат. Реакцію зупиняли додаванням 2xSDS-навантаженого буфера, кип'ятили протягом 3 хвилин і аналізували SDS-PAGE гелевим методом. Результати представлені на Фіг.7. Результати, представлені на Фіг.6 показують, що В1 може безпосередньо індукувати MP-JCP активацію. Однак, враховуй те, що В1 повинен також індукувати NF-kΒ, ця активація, як виявилось, є незалежною від його кіназного домену, і гадають, що вона може бути пов'язана з CARD доменом, при цьому проміжний домен В1 може вносити чи не вносити свій внесок, частково чи повністю. Подальший аналіз клітинної цитотоксичності показує, що не лише В1 (див. приклад 2(іі)), але також В1 мутант, будучи експресованими разом з p55-ФHП-R, RIP чи TRADD посилюють рівень клітинної загибелі. DCARD, DNde та DΚ роблять це у меншому ступеню, тоді як інші конструкти не роблять цього. Мабуть, це вказує на те, що, принаймні, CARD домен утягнений до посилення клітинної загибелі, можливо, разом з проміжним доменом. Результати JNK активації також, видимо, вказують, що, принаймні, CARD домен утягнений до цієї активації, також, можливо, разом з проміжним доменом. На додаток до вищевказаного, проведені дослідження показали, що В1 спонтанно фосфорилює. Це доказ того, що В1 - дійсно кіназа. Далі було підтверджено, що В1 має гомологію з RIP. Комп'ютерний аналіз вказує 37%-ову тотожність двох білків на амінокислотному рівні і 47%-ову гомологію. У даний час RIP в основному розглядається як NF-DΒ модулятор, і вищевказані результати вказують, що В1 діє аналогічно. Приклад 4: Характеристики зв'язування Характеристики зв'язування В1 та його мутантів представлені далі у таблиці VI: Таблиця VI ДНК-зв'язуючий гібрид Гібрид активації LacZ В1 В1 +++ В1 +++ DΚ И1 +++ DΚ DΚ DΚ В1 TRAF2 В1 TRAF3 В1 TRAF6 TRAF2 В1 TRAF6 В1 TRAF1 В1 + В1 TANK В1 NIK NIK В1 В1 CASH CASH В1 В1 RIP В1 RAIDD В1 ICE В1 ICH-1 В1 MACHa1 (C360S) В1 MORT-1 В1 сІАР-1 СІАР-1 В1 + RIP B1 RAIDD B1 ICE B1 ICH-1 B1 B1 MACHa1 (C360S) З результатів, показаних у вищевказаній таблиці, зрозуміло, що, коли В1 функціонує, щоб стимулювати NF-kΒ активації, він може робити це незалежно від зв'язування з іншими білками, відомими як утягнені до NF-kΒ активації, такими як, наприклад, IRAK, TRAF2, NIK, TRAF6 та RIP. Тому В1 може індукувати NF-kB активацію безпосередньо чи опосередковано через взаємодію з деякими іншими білками, що формують частину цього шляху активації. Що стосується активності В1, що спостерігається, у посиленні клітинної загибелі, з вищевказаної таблиці випливає, що В1 може бути центральним білком, що бере участь у точній рівновазі між внутрішньоклітинними шляхами, які ведуть до клітинної загибелі чи до клітинного виживання. У цьому зв'язку В1, в залежності від того, з яким білком він взаємодіє, є здатним до зсуву рівноваги між клітинною загибеллю та клітинним виживанням. 293 клітини нирки людського ембріону (5Хю106; 2,5х106/на 10см чашку) були одночасно трансфектовані у відповідності до процедури, що використовує фосфат кальцію, 10мкг плазміди, що кодує НА-мічений білок В1 (НА-В1) і 10мкг будь-якої ллазміди, що кодує Flag-мічений білок В1 (FL-B1), або плазміди, що кодує Flagмічений С-ІАР1 білок (FL-IAP1), або плазміди, що кодує Flag-мічений TRAF1 (FL-TRAF1), або плазміди, що кодує Flag-мічений TRAF2 (FL-TRAF2) або з 10мкг комбінації (у співвідношенні 1:1) Flag-міченого TRAF1 та не-міченого TRAF2 (FL-TR1+TR2), або з 10мкг комбінації (1:1:1) Flag-міченого TRAF1, не-міченого TRAF2 і с-ІАР1 (FL-TR1+TR2+IAP1). Через 7 годин після трансфекції клітини промивали і через 18 годин клітини лізували у буфері, що містив 50мМ HEPES, рН 7,5, 250мМ NaCI, 0,2% NP-40, 5мМ EDTA, 1мМ фенілметилсульфонілфториду, 2,0мкг/мл апротоніну та 20мкг/мл леупептину (лізисний буфер). Імунопреципітацію здійснювали інкубацією (2год., 4°С) 1 мл аліквот лізату з анти-Flag епітопним антитілом (5мкг/аліквота) та з кульками білка G-агарози (30мкг/аліквота). Імунопреципітати промивали три рази лісним буфером і один раз PBS, розділяли на фракції SDS-PAGE (10%) і поміщали на нітроцелюлозну мембрану (Schieicher & Schuell, Dassel, Німеччина). Вестерн блот аналіз виконували з анти-НА епітопними моноклональними антитілами при розбавленні 1:1000, і з використанням набору ECL (Amersham, Бакінгемшир, Британія). З Фіг.8 видно, що В1 є здатним до самоасоціювання, а також до взаємодії з TRAF1. Рівень взаємодії виявився приблизно таким самим. Що й самоасоціації. Ніякої прямої взаємодії з TRAF-2 чи 1 АРІ не спостерігається. Приклад 5: В1 зв'язується з Ε суб'одиницею V-АТРази Два гібридні фільтри з CARD-доменом В1 як принадою одержували у процесі клонування Ε суб'одиниці V-АТРази (огляд Nelson et al., Experientia 52 (1996 pp.1101-1110). Ε суб'одиницю V-АТРази флуоресцентно мітять та інкубують із зразком CARD-домену В1 у присутності різних зразків бібліотеки органічних молекул або пептидів. Після інкубації, B1-CARD піддають імунопреципітації визначеними антитілами і вимірюють кількість флуоресценції, асоційовану з преципітатом. Молекули, що зашкоджують осадженню флуоресцентно міченого Ε-білка, потім досліджують на можливість використання сполуки як лікарського засобу, що впливає на ріст та/або дапалення клітин в результаті дії Е-суб'одиниці АТРази. При наявності повного опису даного винаходу, фахівцям буде зрозуміло, що заявлена пропозиція може бути здійснена в широкому діапазоні еквівалентних параметрів, концентрацій та умов, не відходячи від суті та обсягу винаходу і без надмірного експериментування. Хоч даний винахід описаний у вигляді його конкретних втілень, зрозуміло, що можливі й його подальші модифікації. Ця заявка охоплює будь-які модифікації, використання чи застосування винаходу, які у цілому слідують основним принципам винаходу, включаючи такі відступи від даного опису, які находяться у межах відомої чи загальноприйнятої практики в даній області, до якої належить винахід, і у відповідності З ознаками, сформульованими далі в об'ємі запропонованої формули винаходу. Усі посилання, що цитуються тут, включаючи видані журнальні статті чи резюме, або відповідні заявки США чи іноземні заявки, подані у США, або іноземні патенти, або будь-які інші посилання, повністю включені до даного опису, включаючи усі дані, таблиці, фігури та текст, представлений у посиланнях, що цитуються. Додатково включений також повний вміст посилань, що цитуються у тих посиланнях, які представлені в описі, У посиланнях на відомі стадії способів, звичайні стадії способів, відомі способи чи звичайні способи ні в якому випадку не розкритий який-небудь аспект опису чи варіант втілення даного винаходу, представленого тут, і немає ніякої вказівки чи припущення про таку можливість такого розкриття. Попередній опис кожного конкретного втілення повністю ілюструє суть винаходу, тому будь-який фахівець у цій області може, використовуючи навики у даній області (а також беручи до уваги вміст посилань, що цитуються тут), легко модифікувати та/або пристосовувати для різних застосувань таке конкретне втілення без надмірного експериментування, не відхиляючись від загальної концепції існуючого винаходу. Тому, такі застосування та модифікації мають бути у межах параметрів і діапазону еквівалентів описаних втілень, основаних на вказівках та керівництві, представлених у цій заявці. Має бути зрозуміло, що фразеологія чи термінологія, що використовується тут, служить лише для цілей опису, а не обмеження, і є такими, щоб термінологія чи фразеологія існуючої специфікації могла інтерпретуватись кваліфікованим фахівцем на основі досліджень та рекомендацій, представлених тут, у поєднанні із звичайними знаннями у цій області. Посилання 1. Adelman et al., (1983) DNA 2, 183. 2. Alnemri. E.S. et al., (1995) J. Biol. Chem. 270, 4312-4317. 3. Ausubel, F.M. et al., (1994) eds., Current Protocols in Molecular Biology. 4. Baeuerle, P.Α., and Henkel, T. (1994) Ann. Rev. Immunol. 5. Bazan, J.F. (1993). Current Biology 3, 603-606. 6. Berberich, I., Shu, G.L., and Klark, E.A. (1994) J. Immunol. 153, 4357-66. 7. Beutler, В., and van Huffel, C. (1994). Science, 264, 667-8. 8. Blank, V., Kourilsky, P., and Israel, A. (1992). Trends Biochem. Sci., 17, 135-40. 9. Boldin, M.P. et al., (1995a) J. Biol. Chem. 270, 337-341. 10. Boldin. M.P., Varfolomeev, E.E., Pancer, Z., Mett, I.L., Camonis, J.H., and Wallach, D. (1995b). J. Biol. Chem., 270, 7795-7798. 11. Boldin, M.P. et al., (1996) Cell, 85, 803-815. 12. Cao, Z. et al., (1996a) Nature, 383, 443-446. 13. Cao, Z. et al., (1996b) Science, 271, 1128-1131. 14. Chen, C.J. et al., (1992) Ann. N.Y. Acad. Sci., 660:271-273. 15. Cheng, G., Cleary, A.M., Ye, Z-s., Hong, D.I., Lederman, S. and Baltimore, D. (1995) Science, 267:14941498). 16. Cheng, G. and Baltimore, D. (1996) Genes Dev. 10, 963-973. 17. Chinnaiyan, A.M., O'Rourke, K., Tewari, M., and Dixit, V.M. (1995) Cell, 81, 505-512. 18. Chinnaiyan, A.M. et al. (1996) J. Biol. Chem. 271, 4573-4576. 19. Creighton, Т.Е., Proteins: Structure and Molecular Properties, W.H. Freeman & Co., San Francisco, Ca. 1983. 20. Croston, G.E., Cao, Z., and Goeddel, D.V., (1995). J. Biol. Chem., 270, 16514-7. 21. DiDonato, J.A., Mercurio, F. and Karin, M. (1995). МоІ. Cell. Biol., 15, 1302-11. 22. Duan, H. and Dixit, V.M., (1997) Nature, 385, 86-89. 23. Durfee, T. et al., (1993) Genes. Dev., 7:555-569. 24. Field, J. et al., (1988) МоІ. Cell Biol., 8:2159-2165. 25. Geysen, H.M., (1985) Immunol. Today, 6, 364-369. 26. Geysen, H.M. et al., (1987) J. Immunol. Meth., 102, 259-274. 27. Gilmore, T.D, and Morin, P.J. (1993). Trends Genet., 9, 427-33. 28. Gossen, M. and Bujard, M. (1992) PNAS 89:5547-5551. 29. Grell, M., Douni, E., Wajant, H., Lohden, M., Clauss, Μ., Baxeiner, В., Georgopoulos, S., Lesslauer, W. . Kollias, G., Pfizenmaier, K., and Scheurich, P. (1995). Cell 83, 793-802. 30. Grilli, M., Chiu, J.J., and Lenardo, M.J. (1993). Int RevCytoL. 31. Hanks, S.K., Quinn, A.M., and Hunter, T. (1988). Science, 241, 42-52. 32. Hofmann K. et al., (1997) TIBS May 22, 1997, p. 155-156. 33. Howard, A.D. et al., (1991) J. Immunol., 147, 2964-2969. 34. Hsu, H., Shu, H.-B., Pan. M.-G., and Goeddel. D.V. (1996). Cell 84, 299-308. 35. Hsu, H., Xiong, J., and Goeddel, D.V. (1995). Cell; 81, 495-504. 36. Kaufmann, S.H. (1989) Cancer Res. 49, 5870-5878. 37. Kaufmann, S.H. (1993) Cancer Res. 53, 3976-3985. 38. Lalmanach-Girard, A.C., Chiles, T.C., Parker, D.C., and Rothstein, T.L. (1993). J. Exp. Med. 177, 1215-1219. 39. Lazebnik, Y.А. et al. (1994) Nature, 371, 346-347. 40. Malinin, N.L. et al., (1997) Nature, 385, 540-544. 41. Mashirna, T. et al., (1995) Biochem. Biophys. Res. Commun., 209, 907-915. 42. McDonald. P.P., Cassatella. M.A., Bald, Α., Maggi, E., Romagnani, S., Gruss, H.J., and Pizzolo, G. (1995). Eur. J. Immunol. 25, 2870-6. 43. Messing et al., Third Cleveland Symposium on Macromoleculas and Recombinant DNA, Ed. A. Walton, Elsevier. Amsterdam (1981) 44. Milligan. C.E. et al., (1995) Neuron, 15, 385-393. 45. Mosialos, G., Birkenbach, M., Yalamanchili, R., VanArsdale, Т., Ware, C, and Kieff, E. (1995). Cell, 80, 389399. 46. Muranishi, S. et al., (1991) Pharm. Research, 8, 649. 47. Nagata, S. and Golstein, P. (1995) Science, 267, 1449-1456. 48. Rensing-Ehl, Α., Hess, S., Ziegler-Heitbrock, H. W. L., Riethmuller, G., and Engelmann, H. (1995). J. Inlamm. 45, 161-174. 49. Rothe. M., Pan, M.-G., Henzel, W.J., Ayres, T.M., and Goeddel, D.V. (1995b). Cell 83, 1243-1252. 50. Rothe, M., Sarma, V., Dixit, V.M., and Goeddel, D.V. (1995a). Science 269, 1424-1427. 51. Rothe, M., Wong, S.C., Henzel, W.J., and Goeddel, D.V. (1994). Cell 78, 681-692. 52. Rothe, M. et al., (1996) Proc. Natl. Acad. Sci. U.S.A. 93, 8241-8246. 53. Ruzicka et al., (1993) Science, 260, 487. 54. Sambrook et al., (1989) Molecular cloning: a laboratory manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. 55. Sano et al., (1992) Science, 258, 120. 56. Sano et al., (1991) Biotechniques, 9, 1378. 57. Schreiber, E., Matthias, P., Muller, M.M. and Schaffner, W. (1989), Nuc. Acids Res. 17:6419. 58. Schulz et al., G.E., Principles of Protein Structure, Springer-Verlag, New York, N.Y. 1798. 59. Sleath, P.R. et al., (1990) J. Biol. Chem. 265, 14526-14528. 60. Smith, C.A., Farrah, Т., and Goodwin, R.G. (1994). Cell 76, 959-962. 61. Stanger, B.Z. et al., (1995) Cell 81, 513-523. 62. Thornberry, N.A. et al., (1992) Nature, 356, 768-774. 63. Thornberry, N.A. et al., (1994) Biochemistry, 33, 3934-3940. 64. Uren, A.G. et al. (1996) Proc. Natl. Acad. Sci. USA 93, 4974-4978. 65. Vandenabeele, P., Declercq, W., Beyaert. R., and Fiers, W. (1995). Trends Cell Biol. 5, 392-400. 66. Varfolomeev, E.E., Boldin, M.P., Goncharov, T.M., and Wallach, D. (1996). J. Exp. Med. in press. 67. Vassalli, P. (1992) Ann. Rev. Immunol. 10, 411-452. 68. Veira et al., (1987) Meth. Enzymol. 153, 3. 69. Wallach, D. (1996) Eur. Cytokine Net. 7, 713-724. 70. Wallach. D. (1997) Trends Biochem. Sci. 22, 107-109. 71. Wang, L. et al., (1994) Cell 78, 739-750. 72. Wilks, A.F. et al., (1989) Proc. Natl. Acad. Sci. USA, 86:1603-1607. 73. Yang, E. and Korsmeyer, J. (1996) Blood 88(2), 386-401. 74. Zaccharia, S. et al., (1991) Eur. J. Pharmacol. 203, 353-357. 75. Zhao, J.J. and Pick, L. (1993) Nature 365: 448-451. ПЕРЕЛІК ПОСЛІДОВНОСТЕЙ (1) ЗАГАЛЬНА ІНФОРМАЦІЯ: (і) ЗАЯВНИК: (A) АДРЕСАТ: YEDA RESEARCH AND DEVELOPMENT CO. LTD. (B) ВУЛИЦЯ: WEIZMANN INSTITUTE OF SCIENCE, P.O.B. 95 (C) МІСТО: REHOVOT (E) КРАЇНА: ISRAEL (F) ІНДЕКС (ZIP): 76100 (G) ТЕЛЕФОН: 972-8-9344093 (Η) ТЕЛЕФАКС: 972-8-9470739 (A) ІМ'Я: WALLACH, DAVID (B) ВУЛИЦЯ: 24 BOROCHOV STREET (C) МІСТО: REHOVOT (Ε) КРАЇНА: ISRAEL (F) ІНДЕКС (ZIP): 76406 (A) ІМ'Я: BOLDIN, MARK (B) ВУЛИЦЯ: BEIT CLORE, WEIZMANN INSTITUTE OF SCIENCE (C) МІСТО: REHOVOT (E) КРАЇНА: ISRAEL (F) ІНДЕКС (ZIP): 76100 (A) ІМ'Я: MALININ, NIKOLAI (B) ВУЛИЦЯ: BEIT CLORE, WEIZMANN INSTITUTE OF SCIENCE (C) МІСТО: REHOVOT (E) КРАЇНА: ISRAEL (F) ІНДЕКС (ZIP): 76100 (іі) НАЗВА ВИНАХОДУ: МОДУЛЯТОРИ ВНУТРІШНЬОКЛІТИННОГО ЗАПАЛЕННЯ, ШЛЯХІВ КЛІТИННОЇ ЗАГИБЕЛІ ТА КЛІТИННОГО ВИЖИВАННЯ (ііі) ЧИСЛО ПОСЛІДОВНОСТЕЙ: 2 (iv) ЕЛЕКТРОННА ФОРМА: (A) ТИП НОСІЯ: Floppy disk (B) КОМП'ЮТЕР: IBM PC compatible (C) ОПЕРАЦІЙНА СИСТЕМА: PC-DOS/MS-DOS (D) ПРОГРАМНЕ ЗАБЕЗПЕЧЕННЯ: Patent in Release #1.0, Version #1.30 (ЕРО) (vi) ДАНІ ПО ПРИОРІТЕТНІЙ ЗАЯВЦІ: (A) НОМЕР ЗАЯВКИ: IL 121011 (B) ДАТА ПОДАННЯ: 05 ЧЕРВНЯ 1997 РОКУ (vi) ДАНІ ПО ПРИОРІТЕТНІЙ ЗАЯВЦІ: (A) НОМЕР ЗАЯВКИ: IL 121199 (B) ДАТА ПОДАННЯ: 30 ЧЕРВНЯ 1997 РОКУ (vi) ДАНІ ПО ПРИОРІТЕТНІЙ ЗАЯВЦІ: (A) НОМЕР ЗАЯВКИ: IL 121746 (B) ДАТА ПОДАННЯ: 11 ВЕРЕСНЯ 1997 РОКУ (2) ІНФОРМАЦІЯ ПРО SEQ ID NO: 1 : (і) ХАРАКТЕРИСТИКИ ПОСЛІДОВНОСТІ: (A) ДОВЖИНА: 540 амінокислот (B) ТИП: амінокислотна (C) КІЛЬКІСТЬ ЛАНЦЮГІВ: один (D) ТОПОЛОГІЯ: лінійна (іі) ТИП МОЛЕКУЛИ: білок (2) ІНФОРМАЦІЯ ПРО SEQ ID NO: 2: (і) ХАРАКТЕРИСТИКИ ПОСЛІДОВНОСТІ: (A) ДОВЖИНА: 2098 пар основ (B) ТИП: нуклеїнова кислота (C) КІЛЬКІСТЬ ЛАНЦЮГІВ: один (D) ТОПОЛОГІЯ: лінійна (іі) ТИП МОЛЕКУЛИ: кДНК
ДивитисяДодаткова інформація
Назва патенту англійськоюModulators of metabolic ways of intracellular inflammation, cells death and survival
Назва патенту російськоюМодуляторы метаболических путей внутриклеточного воспаления, гибели клеток и выживания клеток
МПК / Мітки
МПК: C07K 14/47, A61P 43/00, C12N 15/09, C12N 1/15, C12N 1/19, C12N 5/10, C07K 16/18, A61K 48/00, C12N 1/21, C12Q 1/68, A61K 31/7105
Мітки: внутрішньоклітинного, модулятори, клітин, шляхів, виживання, запалення, метаболічних, загибелі
Код посилання
<a href="https://ua.patents.su/32-72877-modulyatori-metabolichnikh-shlyakhiv-vnutrishnoklitinnogo-zapalennya-zagibeli-klitin-ta-vizhivannya-klitin.html" target="_blank" rel="follow" title="База патентів України">Модулятори метаболічних шляхів внутрішньоклітинного запалення, загибелі клітин та виживання клітин</a>
Дренажна полімерна трубка для інтраопераційної профілактики гострого інфекційного запалення сечових шляхів при відкритих та ендоскопічних втручаннях

Номер патенту: 31989
Опубліковано: 15.12.2000
Автори: Гогуленко Олег Васильович, Ухаль Михайло Іванович, Борисов Олександр Валентинович, Щегринов Микола Миколайович, Борисов Сергій Олександрович
МПК: A61M 39/00, A61B 17/00, A61M 1/00
Мітки: профілактики, відкритих, шляхів, запалення, сечових, трубка, полімерна, інтраопераційної, ендоскопічних, гострого, дренажна, інфекційного, втручаннях
Текст:
...зал т за т з в^онгоьан^і конгэйДовжина трубки 80 см., дів^егр відлов Іцав }п; o-V шкал яіарьсра. ^lajerp трьох д^інажних отворів, уозлЦцзних основи контейн^У - Т,5 ГЛЛЇ , доьж^а контейнеру - Іом і. До катетеру надається ;.шндран довж^-ною В^> CJVI. , з з сумТшоіІ ак^ідобутадіснсгірольної лластшси з ^^Х OratіільнІ вироби ііО^ЗіцвнІ до аласг*яов*х иакагів ш/моуостання дренажних грубок здійснюьалхя тактом чином: ha аэ^шому етааі було...
Спосіб запобігання загибелі та піднімання затоплених підводних кораблів

Номер патенту: 63165
Опубліковано: 15.01.2004
Автори: Вершинін Валерій Афанасійович, Зорін Андрій Микитович, Алєксєєв Станіслав Георгієвич
МПК: B63C 7/08
Мітки: кораблів, піднімання, загибелі, спосіб, запобігання, підводних, затоплених
Формула / Реферат:
Спосіб запобігання загибелі і піднімання затоплених підводних кораблів, що включає надпалубне остроплювання балонів до затопленого корабля і їх наповнення повітрям, який відрізняється тим, що балони у складеному стані розміщують між стінками внутрішнього і зовнішнього корпусів підводного корабля з урахуванням розподілу його ваги та водотонажності, причому балони з'єднують з індивідуальним джерелом стиснутого газу через керований клапан, який...
Спосіб прогнозування переходу гострого запалення внутрішніх органів і шкіри у хронічне запалення

Номер патенту: 54087
Опубліковано: 17.02.2003
Автори: Свірідова Валентина Василівна, Барінова Марія Едуардівна, Бондаренко Надія Миколаївна, Чурілов Андрій Вікторович, Баринов Едуард Федорович
МПК: A61B 10/00, A61B 5/15
Мітки: хронічне, внутрішніх, гострого, спосіб, прогнозування, шкіри, запалення, органів, переходу
Формула / Реферат:
Cпосіб прогнозування переходу гострого запалення внутрішніх органів і шкіри в хронічне запалення шляхом аналізу функціональної активності клітин крові, який відрізняється тим, що досліджують стимульовану активність НАДФН-оксидази моноцитів до і через 3-5 днів лікування, причому в контрольну пробу із суспензією моноцитів додають АТФ, а в дослідну - додатково вводять форбол мірістату ацетат, і якщо процентна різниця активності НАДФН-оксидази...
Спосіб виявлення внутрішньоклітинного кальцію

Номер патенту: 23611
Опубліковано: 02.06.1998
Автори: Слінченко Микола Зіновійович, Філіпченко Лідія Лаврентіївна, Сидоренко Ольга Миколаївна
МПК: G01N 33/48, G01N 33/84, G01N 21/76
Мітки: спосіб, кальцію, внутрішньоклітинного, виявлення
Формула / Реферат:
Спосіб виявлення внутрішньоклітинного кальцію шляхом люмінесцентної мікроскопії, який відрізняється тим, що дослідження проводять на сухих мазках клітин після експозиції їх на протязі 5 хвилин при кімнатній температурі з розчином 6-дезоксі-5-оксітетрацикліну гідрохлориду в безводнім метанолі при розведенні 1:10000.
Спосіб ідентифікації внутрішньоклітинного збудника при хронічному гаймориті
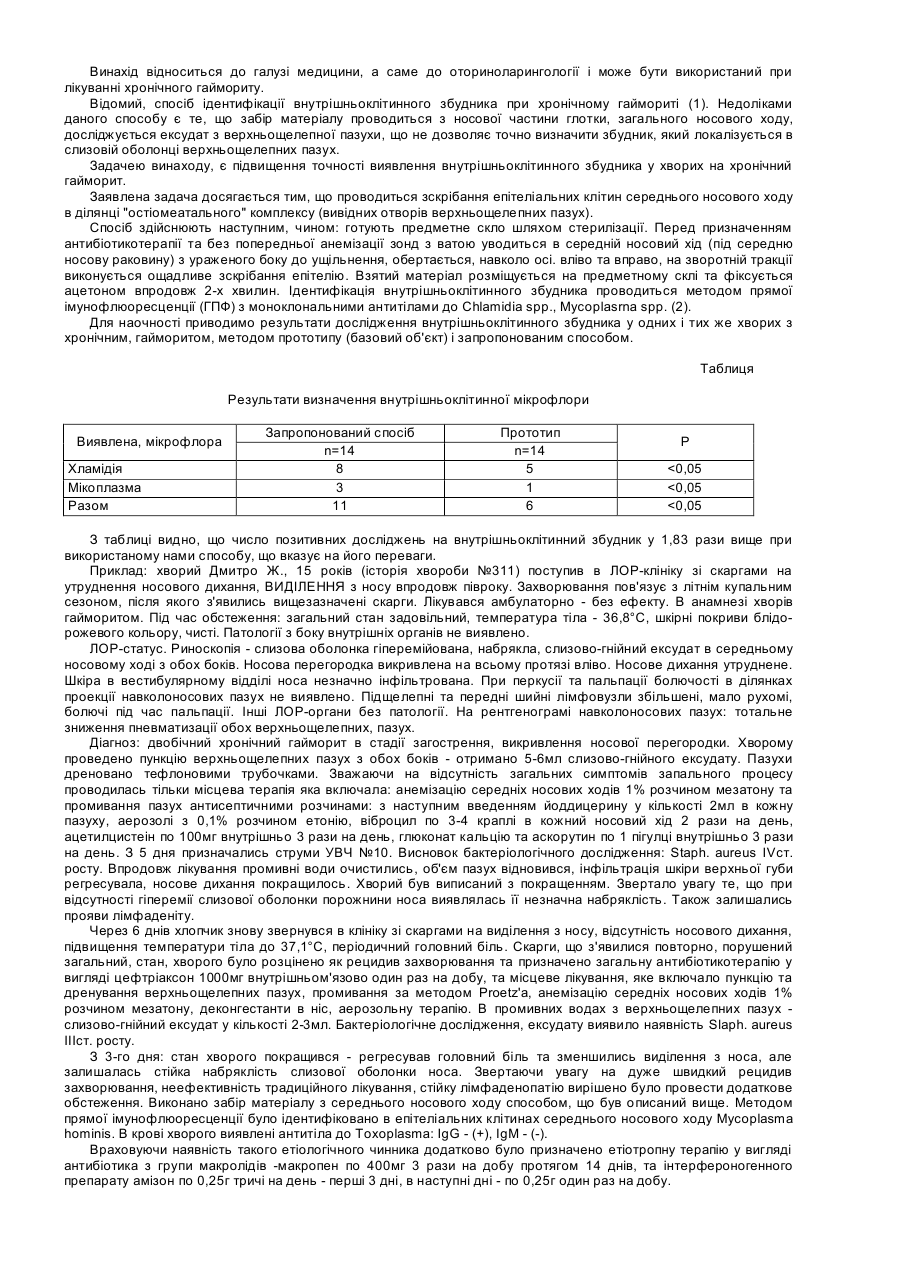
Номер патенту: 65986
Опубліковано: 15.04.2004
Автори: Лайко Андрій Афанасійович, Бредун Олександр Юрійович, Косаковський Анатолій Лук'янович
МПК: G01N 33/533, A61B 10/02
Мітки: ідентифікації, збудника, спосіб, внутрішньоклітинного, хронічному, гаймориті
Формула / Реферат:
Спосіб ідентифікації внутрішньоклітинного збудника при хронічному гаймориті, який виконується методом прямої імунофлуоресценції з моноклональними антитілами до Chlamidia spp., Mycoplasma spp., який відрізняється тим, що забір матеріалу для дослідження проводиться з середнього носового ходу в ділянці "остіомеатального" комплексу (вивідних отворів приносових пазух).
Попередній патент: Спосіб обробки металургійного шлаку
Наступний патент: Сплайсингові варіанти lag-3
Випадковий патент: Спосіб побудови частотно-часових сигнально-кодових конструкцій