Новий вірус рослин, позначений як томато торрадо вірус
Номер патенту: 94035
Опубліковано: 11.04.2011
Автори: Вербек Марінус, Ван ден Хевел Йоханнес Францискус Йоханна Марія, ван дер Влюгт Рене Андрієс Антоніус, Дюллеманс Аннетт Марія, Маріс Паулюс Корнеліс







Формула / Реферат
1. Вірус, названий вірусом томату торрадо (ToTV), який містить в своєму геномі щонайменше одну послідовність нуклеїнової кислоти, вибрану із групи, яка складається з SEQ ID NO:1 і SEQ ID NO:2 і послідовностей, які мають щонайменше 70 % гомологію нуклеотидної послідовності з ними.
2. Вірус за п. 1, депонований у німецькій колекції мікроорганізмів і клітинних культур під номером для посилання депозитів ToTV-E01 (DSM 16999).
3. Виділена або рекомбінантна нуклеїнова кислота або її ToTV-специфічний фрагмент, що містять послідовність нуклеїнової кислоти, вибрану із групи, яка складається з SEQ ID NO:1, SEQ ID NO:2, послідовностей, які мають щонайменше 70 % гомологію нуклеотидної послідовності з SEQ ID NO:1 або SEQ ID NO:2, комплементарних їм ланцюгів і їхніх ToTV-специфічних фрагментів.
4. Полінуклеотид, здатний гібридизуватися в жорстких умовах з нуклеїновою кислотою за п. 3.
5. Виділений або рекомбінантний поліпептид, який можна одержати з вірусу за будь-яким з пп. 1 або 2, або його ToTV-специфічний фрагмент.
6. Поліпептид за п. 5, де вказаний поліпептид вибраний з групи, яка складається з білків капсиду 23, 26 і 35кДа, гелікази, РНК-залежної РНК-полімерази, передбачуваного рухового білка (МР) ToTV і їхніх ToTV-специфічних фрагментів.
7. Антитіло, специфічно спрямоване проти антигену, що містить поліпептид за п. 5 або за п. 6.
8. Спосіб ідентифікації стійкої до ToTV рослини, що включає стадії:
a) впливу на рослину або частину рослини інфекційною дозою ToTV, і
b) ідентифікації зазначеної рослини як стійкої до ToTV, коли, після вказаного впливу, або
- симптоми захворювання в зазначеній рослині або частині рослини залишаються відсутніми, або їхній прояв затриманий, або щонайменше тяжкість зменшена, або вони є локалізованими в порівнянні із чутливою контрольною рослиною, і/або
- вірус ToTV або геномні послідовності ToTV не присутні у вказаній рослині або частині рослини, або присутність вірусу ToTV є щонайменше зменшеною кількісно в порівнянні із чутливою контрольною рослиною.
9. Спосіб за п. 8, де на стадії b) присутність ToTV у вказаній рослині або частині рослини визначають шляхом виявлення у зразку присутності вірусу ToTV або його компонента за допомогою реакції вказаного зразка з полінуклеотидом за п. 4 або антитілом за п. 7.
10. Стійка до ToTV рослина або її частина, отримана способом, що включає стадії:
a) ідентифікації стійкої до ToTV донорної рослини способом за п. 8 або 9;
b) схрещування вказаної стійкої до ToTV донорної рослини з реципієнтною рослиною, і
c) відбору з рослин-нащадків стійкої рослини способом за п. 8 або 9.
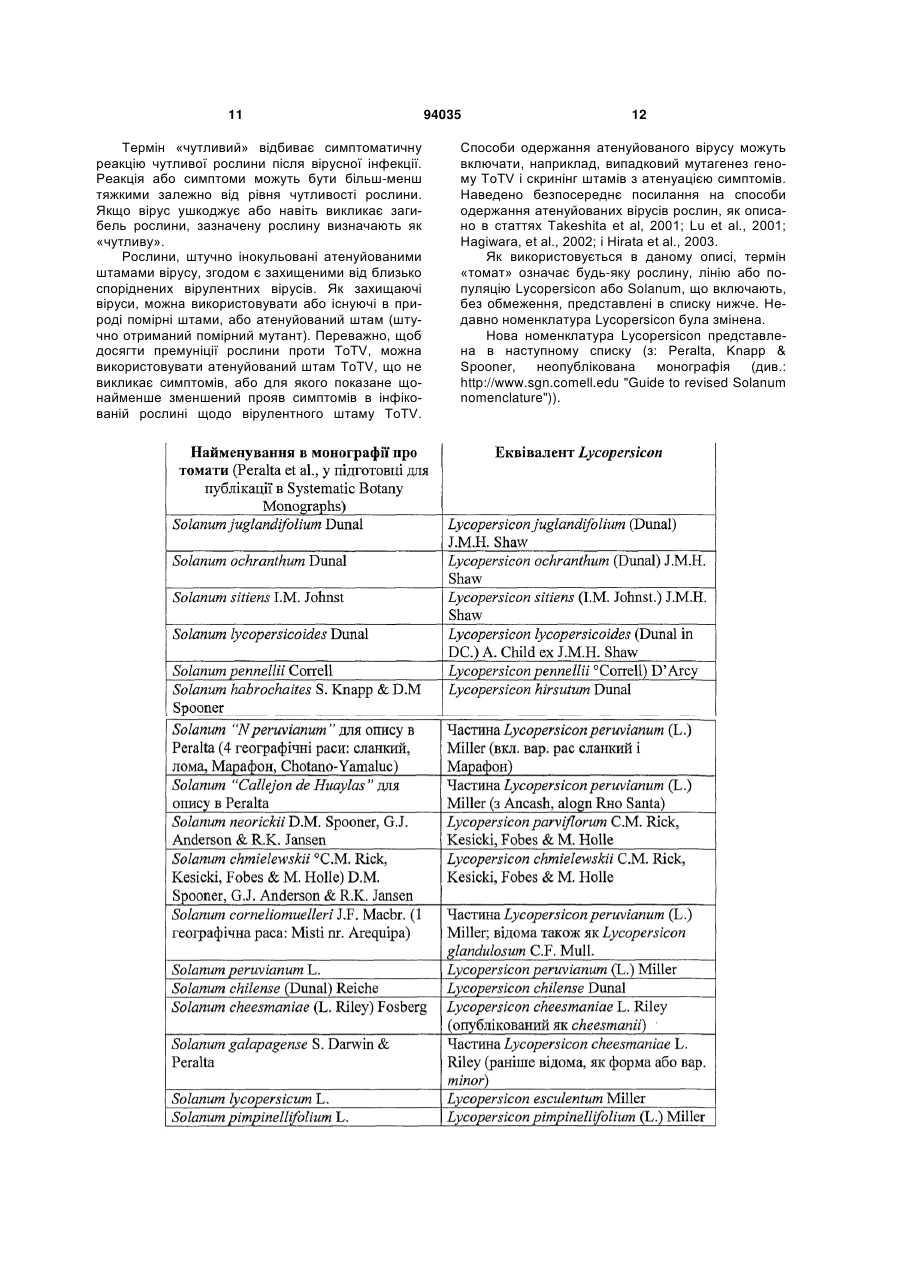
Текст
1. Вірус, названий вірусом томату торрадо (ToTV), який містить в своєму геномі щонайменше одну послідовність нуклеїнової кислоти, вибрану із групи, яка складається з SEQ ID NO:1 і SEQ ID NO:2 і послідовностей, які мають щонайменше 70 % гомологію нуклеотидної послідовності з ними. 2. Вірус за п.1, депонований у німецькій колекції мікроорганізмів і клітинних культур під номером для посилання депозитів ToTV-E01 (DSM 16999). 3. Виділена або рекомбінантна нуклеїнова кислота або її ToTV-специфічний фрагмент, що містять послідовність нуклеїнової кислоти, вибрану із групи, яка складається з SEQ ID NO:1, SEQ ID NO:2, послідовностей, які мають щонайменше 70 % гомологію нуклеотидної послідовності з SEQ ID NO:1 або SEQ ID NO:2, комплементарних їм ланцюгів і їхніх ToTV-специфічних фрагментів. 4. Полінуклеотид, здатний гібридизуватися в жорстких умовах з нуклеїновою кислотою за п.3. 5. Виділений або рекомбінантний поліпептид, який можна одержати з вірусу за будь-яким з пп.1 або 2, або його ToTV-специфічний фрагмент. 6. Поліпептид за п.5, де вказаний поліпептид вибраний з групи, яка складається з білків капсиду 23, 26 і 35кДа, гелікази, РНК-залежної РНКполімерази, передбачуваного рухового білка (МР) ToTV і їхніх ToTV-специфічних фрагментів. 7. Антитіло, специфічно спрямоване проти антигену, що містить поліпептид за п.5 або за п.6. 8. Спосіб ідентифікації стійкої до ToTV рослини, що включає стадії: a) впливу на рослину або частину рослини інфекційною дозою ToTV, і UA (21) a200709047 (22) 09.01.2006 (24) 11.04.2011 (86) PCT/NL2006/000009, 09.01.2006 (31) 05075042.1 (32) 07.01.2005 (33) EP (31) 05075725.1 (32) 29.03.2005 (33) EP (46) 11.04.2011, Бюл.№ 7, 2011 р. (72) ВАН ДЕН ХЕВЕЛ ЙОХАННЕС ФРАНЦИСКУС ЙОХАННА МАРІЯ, NL, МАРІС ПАУЛЮС КОРНЕЛІС, NL, ВЕРБЕК МАРІНУС, NL, ДЮЛЛЕМАНС АННЕТТ МАРІЯ, NL, ВАН ДЕР ВЛЮГТ РЕНЕ АНДРІЄС АНТОНІУС, NL (73) ДЕ РЕЙТЕР СІДЗ Р ЕНД Д Б.В., NL (56) MEJIA, L.: "Chocolate spot disease of tomato in Guatemala"[Online] March 2003 (2003-03), pages 1-2 DATABASE EMBL [Online] 18 July 2001 (2001-0718), "602977808F1 NIH_MGC_12 Homo sapiens cDNA clone IMAGE:5122800 5', mRNA sequence." retrieved from EBI accession no. EM_PRO:BI256469 Database accession no. BI256469 DATABASE EMBL [Online] 13 December 2004 (2004-12-13), "Maize chlorotic dwarf virus isolate M1, complete genome." retrieved from EBI accession no. EM_PRO:AY829112 Database accession no. AY829112 DATABASE EMBL [Online] 18 June 1992 (1992-0618), "Rice tungro spherical virus polyprotein gene, complete cds." retrieved from EBI accession no. EM_PRO:LERTUPOLY Database accession no. LERTUPOLY MEJIA, L.: "Chocolate spot disease of tomato in Guatemala"[Online] March 2003 (2003-03), pages 12, Retrieved from the Internet: URL:http://www.plantpath.wisc.edu/GeminivirusResist antTomatoes/CDR/Mar03/ChocSpot.htm> THOMPSON J R ET AL: "A new potato virus in a new lineage of picorna-like viruses" ARCHIVES OF VIROLOGY, vol. 149, no. 11, 23 July 2004 (2004-0723), pages 2141-2154 2 (19) 1 3 94035 4 b) ідентифікації зазначеної рослини як стійкої до ToTV, коли, після вказаного впливу, або - симптоми захворювання в зазначеній рослині або частині рослини залишаються відсутніми, або їхній прояв затриманий, або щонайменше тяжкість зменшена, або вони є локалізованими в порівнянні із чутливою контрольною рослиною, і/або - вірус ToTV або геномні послідовності ToTV не присутні у вказаній рослині або частині рослини, або присутність вірусу ToTV є щонайменше зменшеною кількісно в порівнянні із чутливою контрольною рослиною. 9. Спосіб за п.8, де на стадії b) присутність ToTV у вказаній рослині або частині рослини визначають шляхом виявлення у зразку присутності вірусу ToTV або його компонента за допомогою реакції вказаного зразка з полінуклеотидом за п.4 або антитілом за п.7. 10. Стійка до ToTV рослина або її частина, отримана способом, що включає стадії: a) ідентифікації стійкої до ToTV донорної рослини способом за п.8 або 9; b) схрещування вказаної стійкої до ToTV донорної рослини з реципієнтною рослиною, і c) відбору з рослин-нащадків стійкої рослини способом за п.8 або 9. Даний винахід належить до галузі захворювань рослин. Більш конкретно, винахід належить до нового вірусу рослин, виділеного з томату, до способів детекції зазначеного вірусу, до способів детекції стійких рослин і до способів одержання стійких рослин. Томат Solarium lycopersicum (раніше Lycopersicon esculentum) є чутливим до великої кількості видів вірусів. Деякі з найбільш відомих вірусів томату містять у собі вірус бронзовості томату (TSWV; рід Tospovirus); вірус мозаїки пепіно (PepMV; рід Potexvirus), і вірус жовтої кучерявості листя томату (TYLCV; рід Begomovirus). Ушкодження, які ці захворювання наносять рослині, знаходяться у діапазоні від знебарвлення листя і некротичних вогнищ, до серйозної втрати врожайності й загибелі рослини. Можливість одержувати стійкі рослини є вкрай важливою для комерційних селекціонерів, і для деяких з вірусів, що наносять найбільший економічний збиток, отримані стійкі варіанти рослин. Однак час від часу з'являються нові віруси, які можуть завдавати значної шкоди врожаю. В 1996р. опублікований новий вірус томату, що інфікував рослини томату в США й Італії з 1993p., і названий вірусом інфекційного хлорозутоматів (TICV; рід Crinivirus; Duffus et al., 1996). Інший новий вірус томату з того ж роду опублікований в 1998 році. Показано, що цей вірус інфікував рослини томату в США з 1989 року, і його назвали вірусом хлорозу томатів (ToCV; Wisler et al, 1998). Доведено, що обоє цих нових вірусу поширює білокрилка, комаха, що є дуже ефективним переносником захворювання. Звичайно вважають, що географічний розподіл відомих вірусів буде збільшуватися, і що будуть продовжувати з'являтися нові віруси, частково в результаті рекомбінації різних вірусів з утворенням нових штамів або нових вірусів. Розробка стійких культурних сортів може відігравати важливу роль в успішному контролі цих захворювань. Недавно на рослинах томатів з Іспанії виявили новий вірус, що викликає симптоми, які не можна приписати якому-небудь із відомих вірусів. У рослин виявляли некротичні ушкодження на листках і коричневі кільця на плодах, а також знижений ріст. Серологічні тести (ELISA) показали присутність вірусу мозаїки пепіно (PepMV). Дослідження електронною мікроскопією дійсно виявили палочковидні частинки, типові для потекс-вірусів (Potexvirus). Однак в інфікованій тканині листка виявлені також вірусні частинки сферичної форми. Авторам даного винаходу вдалося виділити новий вірус із комплексу з PepMV. Новий вірус попередньо назвали вірус томату торрадо (ToTV). Дуже важливою є можливість розпізнати захворювання, щоб простежити його джерело, здійснити моніторинг його епідеміології й попередити його можливе поширення. Тільки тоді можна вживати адекватних заходів для ізоляції рослин і починати фітосанітарну профілактику. На даний момент не існує доступних діагностичних засобів. Отже, існує необхідність розробки діагностичних засобів для даного захворювання. Більше того, у цей час невідомі рослини, що володіють специфічною стійкістю до цього нового вірусу, у той час як існує необхідність розробки таких стійких рослин. Винахід у першому аспекті належить до вірусу рослин, попередньо названому вірус томату торрадо (ToTV), депонованому в німецькій колекції мікроорганізмів і клітинних культур (DSMZ) у Брауншвейгу 24 листопада 2004р. під номером для посилання депозитів ToTV-EOl (DSM 16999). Вірус викликає симптоми захворювання в рослинах томату, так само як в інших рослинах, і може викликати симптоми самостійно, або в комплексі з іншими вірусами або захворюваннями. Перші системні симптоми складаються в некротичних плямах на верхівці рослини, починаючи з основи листочків складного листка. Некротичні плями набухають і оточені світло-зеленою або жовтою областю (див. Фіг.1). Не всі системно інфіковані листки проявляють симптоми, однак у цих листках можна виявити вірус, наприклад, за допомогою електронної мікроскопії. На плодах, інфікованих ToTV, спостерігаються некротичні кільця. Ріст інфікованих рослин може бути зниженим у порівнянні з неінфікованими рослинами. Наведений вище опис належить до рослин, заново інфікованих виділеним вірусом, і не обов'язково буде точно відбивати симптоми, що зустрічаються в польових умовах. Такі фактори, як раса 5 або варієтет рослини, стадія розвитку, вплив додаткових захворювань і абіотичні фактори (наприклад, температура й відносна вологість), будуть, в остаточному підсумку, визначати прояв і характеристики симптомів. Вірусні частинки є сферичними (ікосаедричними) за формою з діаметром приблизно 28нм (див. Фіг.2). Вірусні частинки складаються щонайменше із трьох білків капсиду приблизно по 23, 26 і 35кДа (див. Фіг.3). Після очищення вірус відображає на дисплеї щонайменше дві видимі смуги в градієнті сульфату цезію. Верхня видима смуга (верхня фракція вірусу) містить молекулу РНК приблизно 5,5т.н. (більш точно, 5,2т.н.), а нижня видима смуга (нижня фракція) містить молекулу РНК приблизно 8т.н. (більш точно, 7,7т.н.) (див. Фіг.4). Інокуляція рослин тютюну обома об'єднаними смугами приводить до інфекції. ToTV здатний механічно передаватися декільком видам Nicotiana. Загальноприйнятий буфер для інокуляції (наприклад, 0,03Μ фосфатний буфер з рН7,7) є придатним. Для розмноження ToTV переважними є N.glutinosa, N.tabacum і N.benthamina. Види тютюну N.hesperis '67A' і N.occidentalis 'Ρ1' є дуже чутливими до ToTV, і проявляють системні симптоми після 3-4 доби. У цих видів томату розвивається сильний некроз за короткий час, і, таким чином, вони є більше прийнятними для використання як індикаторна рослина, ніж як хазяїн для розмноження. N.glutinosa реагує місцевими вогнищами хлорозу, системним хлорозом і помірною деформацією листя. N.benthamiana не проявляє місцевих симптомів і реагує системним хлорозом і деформацією листя. Крім того, винахід належить до вірусу, що містить щонайменше одну послідовність нуклеїнової кислоти, вибрану із групи, яка складається з SEQ ID NO:1 і SEQ ID NO:2 і послідовностей, що володіють щонайменше 30, переважно щонайменше 40, переважно щонайменше 50%, переважно щонайменше 60%, більш переважно щонайменше 70%, більш переважно щонайменше 80%, ще більш переважно щонайменше 90%, ще більш переважно щонайменше 95%, ще більш переважно щонайменше 98% і найбільше переважно щонайменше 99% гомологією з ними. Такі віруси також охоплені терміном ToTV, як використовується в даному описі. У переважному варіанті здійснення вірусу за винаходом, що володіє вищевказаною гомологією послідовності, зазначений вірус пов'язаний із захворюваннями томату, відомими під назвами «Торрадо», «Марчитець» і/або «Захворювання шоколадної плямистості», і/або зазначений вірус, на основі чисельного таксономічного аналізу таксономічних ознак, в основному, як визначено в таблиці 1, має більше близьке споріднення з вірусом, як визначено за п.1 формули винаходу, ніж з будьяким іншим ізолятом вірусу, доступним у публічних колекціях, і зазначений вірус має необхідні характеристики, пов'язаними із захворюванням, що викликає некротичні ушкодження в томату. В іншому аспекті винахід належить до виділеної або рекомбінантної нуклеїнової кислоті, що містить послідовність нуклеїнової кислоти, вибра 94035 6 ну із групи, яка складається з SEQ ID NO:1, SEQ ID NO:2, послідовностей, що володіють щонайменше 50%, переважно щонайменше 70%, більш переважно щонайменше 80%, ще більш переважно щонайменше 90%, ще більш переважно щонайменше 95%, ще більш переважно щонайменше 98% і найбільше переважно щонайменше 99% гомологією нуклеотидної послідовності з SEQ ID NO:1 або SEQ ID NO:2, комплементарних їм ланцюгів і їхніх ToTV-специфічних фрагментів. Таку нуклеїнову кислоту можна одержати з вірусу за винаходом. В іншому аспекті винахід належить до полінуклеотиду, здатному гібридизуватися в суворих умовах з виділеною або рекомбінантною нуклеїновою кислотою за винаходом, як описано вище. В іншому аспекті винахід належить до виділеного або рекомбінантного поліпептиду, отриманого з вірусу за винаходом, або його ToTVспецифічного фрагменту. У переважному варіанті здійснення зазначений поліпептид вибраний з групи, яка складається з білків капсиду 23, 26 і 35кДа і їхній ToTV-специфічних фрагментів. В іншому аспекті винахід належить до антигену, що містить поліпептид за винаходом, або його ToTV-специфічний фрагмент. В іншому аспекті винахід належить до антитіла, специфічно спрямованого проти антигену за винаходом. В іншому аспекті винахід належить до способу одержання антитіла проти ToTV, що включає стадії: а) одержання вірусу ToTV або його (рекомбінантного) білка, або фрагмента його пептиду; Ь) імунізації прийнятного хребетного-хазяїна зазначеним вірусом, білком або фрагментом пептиду й с) збору із крові (включаючи сироватку) або спленоцитів зазначеного хребетного-хазяїна антитіл проти зазначеного вірусу, білка або фрагмента пептиду. У переважному варіанті здійснення зазначений спосіб додатково містить у собі стадії d) відбору одного продукуючого антитіло спленоциту, e) злиття зазначеного спленоциту з імморталізованою лінією клітин гібридоми й f) забезпечення можливості зазначеній злитій гібридомі продукувати моноклональні антитіла. В іншому аспекті винахід належить до антитіла, яке можна одержати способом одержання антитіла проти ToTV за винаходом. В іншому аспекті винахід належить до способу ідентифікації ізоляту вірусу як вірусу ToTV, що включає реакцію зазначеного ізоляту вірусу або його компонента з антитілом за винаходом. В іншому аспекті винахід належить до способу ідентифікації ізоляту вірусу як вірусу ToTV, що включає реакцію зазначеного ізоляту вірусу або його компонента з полінуклеотидом за винаходом. В іншому аспекті винахід належить до способу детекції присутності ToTV у зразку, що включає визначення в зазначеному зразку присутності вірусу ToTV або його компонента шляхом реакції зазначеного зразка з полінуклеотидом або з антитілом за винаходом. В іншому аспекті винахід належить до способу ідентифікації стійкої до ToTV рослини, що включає стадії: а) впливу на рослину або частину рослини інфекційної дози ToTV, і b) ідентифікації зазначе 7 ної рослини як стійкої до ToTV, коли, після зазначеного впливу, або і) симптоми захворювання в зазначеній рослині або частині рослини залишаються відсутніми, або їхній прояв затриманий, або щонайменше вага зменшена, або вони є локалізованими в порівнянні із чутливою контрольною рослиною, і/або іі) вірус ToTV або геномні послідовності ToTV не присутні в зазначеній рослині або частині рослини, або присутність вірусу ToTV є щонайменше зменшеною кількісно в порівнянні із чутливою контрольною рослиною. Стадія а) включає період інкубації досить довгої тривалості, щоб дозволити встановлення симптомів, що піддаються виявленню, захворювання в чутливих контрольних рослинах, що піддавалися впливу порівнянної інфекційної дози вірусу. За допомогою здійснення даного способу можна ідентифікувати в рослині всі форми стійкості, включаючи повну стійкість, часткову стійкість, гіперчутливість і толерантність. Щоб підтвердити толерантність, необхідно підтвердити (системну) присутність вірусу в рослині (клітинах). Стадія Ь) може включати здійснення способу детекції присутності ToTV у зразку із зазначеної рослини або частини рослини за даним винаходом, де застосовують антитіло або полінуклеотид за винаходом в загальноприйнятих способах для аналізів гібридизації нуклеотидів або імуноаналізів, добре відомих фахівцеві в даній галузі. Альтернативно, стадія b) може включати приведення в контакт частини зазначеної рослини, яка піддавалася впливу, із чутливою індикаторною рослиною. Таким чином, можна виявляти наявність системної або місцевої інфекції в зазначеній рослині, яка піддавалася впливу, через спостереження виникнення захворювання в індикаторній рослині, або навіть у додатковій контактувавшій індикаторній рослині, яка контактували із зазначеною першою контактувавшою індикаторною рослиною. В іншому аспекті винахід належить до способу одержання стійкої до ToTV рослини, що включає стадії ідентифікації стійкої до ToTV донорної рослини одним з вищевказаних способів для ідентифікації стійкої до ToTV рослини за винаходом, схрещування зазначеної стійкої до ToTV донорної рослини з реципієнтною рослиною (де реципієнтна рослина може бути або чутливою до ToTV, або стійкою до ToTV, але є, відповідно, стійкою до ToTV рослиною у випадку, якщо стійкий фенотип зумовлений рецесивним геном), і відбору з рослин-нащадків (наприклад, F1, F2, і самозапиленої рослини) стійкої рослини способом ідентифікації стійкості до ToTV у рослини, як описано вище. У випадку, якщо ознака стійкості є рецесивною ознакою, стійкі рослини можна виявити серед рослиннащадків самозапилення F1 або F2, або більше далеких поколінь. У переважних варіантах здійснення за даним аспектом зазначена стійка до ToTV донорна рослина й реципієнтна рослина являють собою рослини сімейства Solanaceae або сімейства Cucurbitaceae. У інших переважних варіантах здійснення за даним аспектом зазначена реципієнтна рослина являє собою томат, баклажан, перець, диню, кавун або огірок, більш переважно, рослину з виду Solarium lycopersicum, най 94035 8 більш переважно, лінії S.lycopersicum, що володіє комерційно бажаними характеристиками. В іншому аспекті винахід належить до стійкої до ToTV рослини, переважно, томата, баклажана, перцю, дині, кавуна або огірка, або його частини, такої як насіння, яке можна одержати способом одержання стійкої до ToTV рослини за винаходом. В іншому аспекті винахід належить до діагностичного набору для детекції присутності ToTV у зразку або для ідентифікації стійкості до ToTV у рослині, що містить вірус, полінуклеотид, поліпептид, антиген і/або антитіло за винаходом. В іншому аспекті винахід належить до застосування вірусу, полінуклеотиду, поліпептиду, антигену або антитіла за винаходом для одержання діагностичної композиції. В іншому аспекті винахід належить до діагностичної композиції, що містить вірус, полінуклеотид, поліпептид, антиген або антитіло за винаходом. В іншому аспекті винахід належить до застосування ToTV, або частин вірусного геному ToTV, як експресуючого вектора. В іншому аспекті винахід належить до застосування ToTV, або частин вірусного геному ToTV, для одержання зумовленої патогеном стійкості в рослин. В іншому аспекті винахід належить до застосування атенуйованої форми вірусу ToTV, або його геному, або його частин, для премуніції рослини. Визначення Як використовується в даному описі, термін «частина рослини» означає частину рослини, включаючи окремі клітини й клітинні тканини, такі як клітини рослини, що є інтактними в рослинах, скупчення клітин і культури тканин, з яких можна регенерувати рослини. Приклади частин рослини включають, без обмеження, окремі клітини й тканини з пилка, насінних зачатків, листя, зародків, коренів, кінчиків коренів, пиляків, квітів, плодів, відростків стебла й насіння; так само як пилок, насінні зачатки, листки, зародки, корні, кінчики коренів, пиляки, квіти, плоди, стебла, пагони, черешки, кореневища, насіння, протопласти, калюси й т.п. Термін «зразок» містить у собі зразок з рослини, із частини рослини або з переносника, або зразок ґрунту, води або повітря. Термін «переносник», як використовується в даному описі, належить до агента, що поширює захворювання, або речовини. Переносники ToTV у польових умовах можуть включати, без обмеження, тварин, таких як Arthropoda (зокрема, із класів Insecta і Arachnida), Nematoda (зокрема, із класу Adenophorea), а також більших тварин, таких як, наприклад, птахи, кролики й миші, гриби (тобто тип Еumусоtа, зокрема, гриби класу Phycomycota), (паразитичні) рослини (включаючи члени сімейства Cuscutaceae), пилок, насіння, воду, тверді частинки й навіть руки людини, устаткування й взуття. Термін рослина-«нащадок» належить до будьякої рослини, отриманої як потомство завдяки вегетативному або статевому розмноженню від однієї або декількох батьківських рослин або їхніх нащадків. Наприклад, рослину-нащадок можна 9 одержати клонуванням або самозапиленням батьківської рослини, або схрещуванням двох батьківських рослин, і вона містить у собі рослини, отримані самозапиленням, так само як F1 або F2, або більш далекі покоління. F1 являє собою потомство першого покоління, отримане від батьків, щонайменше один з яких застосовували перший раз як донор ознаки, тоді як потомство другого покоління (F2) або наступних поколінь (F3, F4 і т.д.) являє собою екземпляри, отримані від самозапилень F1, F2 і т.д. F1 може, таким чином, бути (і звичайно є) гібридом, отриманим від схрещування між двома батьками з розведення гомозигот (гомозиготи з розведення є гомозиготними по ознаці), тоді як F2 може бути (і звичайно є) потомством від самозапилення зазначених гібридів F1. Термін «стійкий», як використовується в даному описі, належить до рослини, здатної опиратися розмноженню вірусу в його клітинах і/або (системному) пересуванню вірусу до інших клітин і/або розвитку симптомів захворювання після інфекції зазначеним вірусом, де вірус здатний до інфікування й розмноження у відповідних нестійких або чутливих варієтетів зазначеної рослини. Термін використовують для включення таких окремо ідентифікованих форм стійкості, як «повна стійкість», «імунітет», «часткова стійкість», «гіперчутливість» і «толерантність». «Повна стійкість» означає повну нездатність вірусу розвиватися після інфекції, і може бути або результатом нездатності вірусу входити в клітину (відсутність початкової інфекції) або може бути результатом нездатності вірусу розмножуватися в клітині й інфікувати наступні клітини (відсутність підпорогової інфекції, відсутність поширення). Наявність повної стійкості можна визначити встановленням відсутності вірусних частинок або вірусної РНК у клітинах рослини, так само як відсутності яких-небудь симптомів захворювання в зазначеній рослині після впливу на зазначену рослину інфекційної дози вірусу (тобто після «інфекції»). Серед селекціонерів даний фенотип часто позначають як «імунний». «Імунітет», як використовується в даному описі, таким чином, належить до форми стійкості, яка характеризується відсутністю вірусної реплікації, навіть коли вірус активно переносять у клітини, наприклад, за допомогою електропорації. «Інфекційну дозу» визначають як дозу вірусних частинок або нуклеїнової кислоти вірусу, здатну інфікувати рослину, де доза може розрізнятися між рослинами й між тестованими ізолятами ToTV. Теоретично, кількість від приблизно 1-10 до приблизно 500-5000 вірусних частинок зазначеного вірусу або його нуклеїнових кислот буде достатньою. Інфекції, таким чином, можна досягати механічною інокуляцією очищених частинок вірусу або нуклеїнової кислоти вірусу в рослини. «Часткова стійкість» означає зменшене розмноження вірусу в клітині, зменшене (системне) пересування вірусу, і/або зменшений розвиток симптомів після інфекції. Наявність часткової стійкості можна визначити встановленням системної присутності низьких титрів вірусних частинок або вірусної РНК у рослині й присутності зменшених або відстрочених симптомів захворювання в за 94035 10 значеній рослині після впливу на зазначену рослину інфекційної дози вірусу. Титри вірусу можна визначити з використанням способу кількісної детекції (наприклад, способу ELISA або кількісної полімеразної ланцюгової реакції зі зворотною транскрипцією [RT-PCR]). Серед селекціонерів даний фенотип часто позначають як «проміжно стійкий». Термін «гіперчутливий» належить до форми стійкості, при якій інфекція залишається місцевою й не поширюється системно, наприклад, через місцевий некроз інфікованої тканини, або відсутність системного пересування за межі інокульованої тканини. У гіперчутливих рослинах виявляють місцеві, але тяжкі симптоми захворювання, і в таких рослинах можна встановити місцеву присутність вірусу. Термін «толерантний» застосовують у даному описі для позначення фенотипу рослини, при якому симптоми захворювання залишаються відсутніми після впливу на зазначену рослину інфекційної дози вірусу, при якому можна встановити присутність системної або місцевої вірусної інфекції, розмноження вірусу, щонайменше присутність вірусних геномних послідовностей у клітинах зазначеної рослини і/або їхню інтеграцію в геном. Таким чином, толерантні рослини є стійкими для прояву симптомів, однак безсимптомні рослини є носіями вірусу. Іноді вірусні послідовності можуть бути присутніми або навіть розмножуватися в рослинах, не викликаючи симптомів захворювання. Цей феномен відомий також як «латентна інфекція». Деякі ДНК- і РНК-віруси можуть ставати такими, що не підлягають виявленню після первинної інфекції, але з'являтися знову пізніше й приводити до гострого захворювання. При латентній інфекції вірус може існувати в істинно латентній неінфекційній схованій формі, можливо, у вигляді інтегрованого геному або епісомального агента (так що вірусні частинки не можна виявити в цитоплазмі, у той час як способами PCR можна показати присутність послідовностей вірусної нуклеїнової кислоти) або у вигляді інфекційного й який постійно реплікується агента. Реактивований вірус може поширюватися й викликати епідемію серед контактуючих чутливих рослин. Наявність «латентної інфекції» не можна відрізнити від наявності «толерантного» фенотипу в рослини. Термін «чутливий» використовують у даному описі для позначення рослини, що не володіє стійкістю до вірусу, що приводить до проникнення вірусу в клітини рослини, розмноження й системного поширення вірусу, викликаючи симптоми захворювання. Термін «чутливий», таким чином, є еквівалентним «нестійкому». У чутливій рослині виявляють нормальні титри вірусу в клітинах після інфекції. Таким чином, чутливість можна визначити за допомогою встановлення нормальних (тобто щодо інших вірусних інфекцій у рослин) титрів вірусних частинок або вірусної РНК у клітинах рослини й присутності нормальних симптомів захворювання (тобто щодо симптомів захворювання, як описано тут для рослини, з якої спочатку виділений ToTV) у зазначеній рослині після впливу на зазначену рослину інфекційної дози вірусу. 11 Термін «чутливий» відбиває симптоматичну реакцію чутливої рослини після вірусної інфекції. Реакція або симптоми можуть бути більш-менш тяжкими залежно від рівня чутливості рослини. Якщо вірус ушкоджує або навіть викликає загибель рослини, зазначену рослину визначають як «чутливу». Рослини, штучно інокульовані атенуйованими штамами вірусу, згодом є захищеними від близько споріднених вірулентних вірусів. Як захищаючі віруси, можна використовувати або існуючі в природі помірні штами, або атенуйований штам (штучно отриманий помірний мутант). Переважно, щоб досягти премуніції рослини проти ToTV, можна використовувати атенуйований штам ToTV, що не викликає симптомів, або для якого показане щонайменше зменшений прояв симптомів в інфікованій рослині щодо вірулентного штаму ToTV. 94035 12 Способи одержання атенуйованого вірусу можуть включати, наприклад, випадковий мутагенез геному ToTV і скринінг штамів з атенуацією симптомів. Наведено безпосереднє посилання на способи одержання атенуйованих вірусів рослин, як описано в статтях Takeshita et al, 2001; Lu et al., 2001; Hagiwara, et al., 2002; і Hirata et al., 2003. Як використовується в даному описі, термін «томат» означає будь-яку рослину, лінію або популяцію Lycopersicon або Solanum, що включають, без обмеження, представлені в списку нижче. Недавно номенклатура Lycopersicon була змінена. Нова номенклатура Lycopersicon представлена в наступному списку (з: Peralta, Knapp & Spooner, неопублікована монографія (див.: http://www.sgn.comell.edu "Guide to revised Solanum nomenclature")). 13 «Експресуючий вектор» визначають як молекулу нуклеїнової кислоти, що містить ген, звичайно гетерологічний ген, який експресується в клітиніхазяїні. Як правило, цей ген містить послідовність, що кодує білок. Експресію гена завжди вміщують під контроль промотору, і такий ген, як говорять, є «функціонально пов'язаним з» промотором. Термін «гетерологічний» належить до молекули ДНК, або до популяції молекул ДНК, що у природних умовах не існує в даної клітині-хазяїні. Термін «полінуклеотид», як використовується в даному описі, є взаємозамінним з терміном «нуклеїнова кислота» і належить до мультимеру нуклеотидів або полімерної форми нуклеотидів, що володіє будь-яким числом нуклеотидів, наприклад, дезоксирибонуклеотидів або рибонуклеотидів, або до отриманих синтетично сполук (наприклад, PNA, як описано в патенті США №5948902 і процитованих там посиланнях), і може бути або дво-, або одноланцюжкових. Полінуклеотид може гібридизуватися з існуючими в природі полінуклеотидами специфічним до послідовності чином, аналогічним гібридизації двох існуючих у природі полінуклеотидів, наприклад, може брати участь у взаємодіях Уотсон-Кріковського спарювання основ. Термін також містить у собі модифіковані, наприклад, метилюванням і/або кепіюванням, і немодифіковані форми полінуклеотиду. Терміни «рибонуклеїнова кислота» і «РНК», як використовується в даному описі, означають полімер, що складається з рибонуклеотидів. Терміни «дезоксирибонуклеїнова кислота» і «ДНК», як використовується в даному описі, означають полімер, що складається з дезоксирибонуклеотидів. Термін «олігонуклеотид» означає коротку послідовність мономерів нуклеотидів (звичайно від 6 до 100 нуклеотидів), з'єднаних фосфорними зв'язками (наприклад, фосфодіефірним, алкіл- і арилфосфатним, фосфоротіоатним), або нефосфорними зв'язками (наприклад, пептидним, сульфаматним і іншими). Олігонуклеотид може містити модифіковані нуклеотиди, що мають модифіковані основи (наприклад, 5-метилцитозин) і модифіковані групи цукрів (наприклад, 2'-Ометилрибозил, 2'-О-метоксіетилрибозил, 2'фторорибозил, 2'-амінорибозил і т.п.). Олігонуклеотиди можуть являти собою природні або синтетичні молекули дво- і одноланцюжкової ДНК і дво- і одноланцюжкової РНК кільцевої, розгалуженої або лінійної форми й, не обов'язково, містять домени, здатні формувати вторинні структури (наприклад, стебло-петля, псевдовузли й структури дотичних петель). Термін «гомологія нуклеотидної послідовності», як використовується в даному описі, означає наявність гомології між двома полінуклеотидами. Полінуклеотиди володіють «гомологічними» послідовностями, якщо послідовність нуклеотидів у двох послідовностях є однаковою при порівнянні по максимальній відповідності. Порівняння послідовності між двома або більше полінуклеотидами, як правило, проводять порівнянням частин двох послідовностей у вікні порівняння для ідентифіка 94035 14 ції й порівняння локальних областей подібності послідовності. Вікно порівняння звичайно становить приблизно від 20 до 200 безперервних нуклеотидів. «Відсоток гомології послідовності» полінуклеотидів, наприклад, 50, 60, 70, 80, 90, 95, 98, 99 або 100 відсотків гомології послідовності, можна визначити порівнянням двох оптимально вирівняних послідовностей у вікні порівняння, де частина послідовності полінуклеотиду у вікні порівняння може містити додавання або делеції (тобто пропуски) у порівнянні з контрольною послідовністю (яка не містить додавань або делецій) для оптимального вирівнювання двох послідовностей. Відсоток обчислюють за допомогою (а) визначення числа положень, у яких ідентичні основи нуклеїнової кислоти присутні в обох послідовностях, для одержання числа співпадаючих положень; (b) розподілу числа співпадаючих положень на загальне число положень у вікні порівняння; і °С) множення результату на 100 для одержання відсотка гомології послідовності. Оптимальне вирівнювання послідовностей для порівняння можна проводити комп'ютеризованим виконанням відомих алгоритмів, або візуальним контролем. Легко доступними алгоритмами порівняння послідовностей і множинного вирівнювання послідовностей є, відповідно, основна програма пошуку локального вирівнювання (BLAST) (Altschul et al., 1990; Altschul et al., 1997) і програма Clustal, обидві доступні через Інтернет. Інші прийнятні програми включають, без обмеження, GAP, BestFit, PlotSimilarity і FASTA у пакеті програмного забезпечення Wisconsin Genetics (Genetics Computer Group (GCG), Madison, WI, USA) (Devereux et al., 1984). Як використовується в даному описі, «в основному комплементарні» означає, що дві послідовності нуклеїнової кислоти володіють щонайменше приблизно 65%, переважно приблизно 70%, більш переважно приблизно 80%, навіть більш переважно 90% і найбільше переважно приблизно 98% комплементарністю послідовностей один з одним. Це означає, що праймери й зонди повинні володіти достатньою комплементарністю з їхньою матрицею й нуклеїновою кислотою-мішенню, відповідно, щоб гібридизуватися в суворих умовах. Таким чином, послідовності праймера й зонда не повинні відбивати точну комплементарну послідовність єднальної області на матриці, і можна застосовувати вироджені праймери. Наприклад, некомплементарний нуклеотидний фрагмент можна приєднувати до 5'-кінця праймера, причому інша послідовність праймера є комплементарною ланцюгу. Альтернативно, у праймер можна вставляти некомплементарні основи або більш довгі послідовності, за умови, що праймер володіє достатньою комплементарністю з послідовністю одним з підлягаючих ампліфікації ланцюгів для гібридизації з нею й, таким чином, формування структури дуплекса, яку можна добудувати способами полімеризації. Некомплементарні нуклеотидні послідовності праймерів можуть включати ділянки для рестрикційних ферментів. Додавання ділянки для рестрикційного ферменту на кінець (кінці) послідовності-мішені може бути особливо корисним для 15 клонування послідовності-мішені. В основному комплементарна послідовність праймера являє собою послідовність, що володіє достатньою комплементарністю послідовності з матрицею для ампліфікації, щоб приводити до зв'язування праймера й синтезу другого ланцюга. Фахівцям у даній галузі відомо, що праймери повинні володіти достатньою комплементарністю послідовності з матрицею для ампліфікації. Термін «гібрид» у контексті нуклеїнових кислот належить до дволанцюжкової молекули нуклеїнової кислоти, або дуплексу, утвореної за допомогою водневих зв'язків між комплементарними нуклеотидними основами. Терміни «гібридизуватися» або «відпалюватися» належить до процесу, за допомогою якого окремі ланцюги послідовності нуклеїнової кислоти утворюють двоспіральні відрізки шляхом водневого зв'язування між комплементарними основами. Термін «гібрид» у контексті селекції рослин належить до рослини, що є нащадком генетично різнорідних батьків, отриманим шляхом схрещування рослин з різних ліній, порід або видів. Термін «зонд» належить до одноланцюжкової олігонуклеотидної послідовності, що буде розпізнавати комплементарну послідовність у послідовності-мішені аналіту нуклеїнової кислоти або похідній від її кДНК і формувати з нею дуплекс із водневими зв'язками. Термін «праймер», як використовується в даному описі, належить до олігонуклеотиду, здатному відпалюватися з мішенню для ампліфікації, дозволяючи приєднання ДНК-полімерази, таким чином, виконуючи функцію точки ініціації синтезу ДНК при вміщенні в умови, у яких індукується синтез продукту подовження праймерів, тобто в присутності нуклеотидів і засобу для полімеризації, такого як ДНК-полімераза, і при прийнятній температурі й рН. Праймер (для ампліфікації) переважно є одноланцюжковим для максимальної ефективності ампліфікації. Переважно, праймер являє собою олігодезоксирибонуклеотид. Праймер повинен бути досить довгим для праймування синтезу продуктів подовження в присутності засобу для полімеризації. Точні довжини праймерів будуть залежати від багатьох факторів, включаючи температуру й склад (вміст А/Т і G/C) праймера. Пара двосторонніх праймерів складається з одного прямого й одного зворотного праймера, як загальноприйнято в сфері ампліфікації ДНК, такій як ампліфікація PCR. Зрозуміло, що «праймер», як використовується в даному описі, може належати до більш ніж одного праймера, зокрема, у випадку, де існує деяка невизначеність інформації щодо кінцевої послідовності (послідовностей) області-мішені, що підлягає ампліфікації. Таким чином, «праймер» включає набір олігонуклеотидів-праймерів, що містить послідовності, які представляють можливі варіанти послідовності, або включає нуклеотиди, що дозволяють типове спарювання основ. Олігонуклеотидні праймери можна одержувати будь-яким прийнятним способом. Способи одержання олігонуклеотидів конкретної послідовності відомі в даній галузі й містять у собі, наприклад, 94035 16 клонування й рестрикцію прийнятних послідовностей, і прямий хімічний синтез. Способи хімічного синтезу можуть включати, наприклад, фосфо диабо три-ефірний спосіб, діетилфосфорамідатний спосіб і твердофазний спосіб, описані, наприклад, у патенті США №4458066. Праймери, якщо бажано, можна мітити шляхом введення засобів, здатних до виявлення, наприклад, спектроскопічними, флуоресцентними, фотохімічними, біохімічними, імунохімічними або хімічними способами. Залежне від матриці подовження олігонуклеотидного праймера (праймерів) каталізує полімеризуючий засіб у присутності прийнятних кількостей чотирьох дезоксирибонуклеотидтрифосфатів (dATP, dGTP, dCTP і dTTP, тобто dNTP) або аналогів, у реакційному середовищі, що складається з прийнятних солей, катіонів металів, і системи для забуферювання рН. Прийнятними полімеризуючими засобами є ферменти, відомі як каталізуючі залежний від праймера й матриці синтез ДНК. Відомі ДНК-полімерази включають, наприклад, ДНКполімеразу І Е.соlі або її фрагмент Кленова, ДНКполімеразу Т4, і ДНК-полімеразу Taq. Умови реакції для каталізу синтезу ДНК цими ДНКполімеразами відомі в даній галузі. Продукти синтезу являють собою молекули дуплекса, що складає з ланцюгів матриці й ланцюгів після подовження праймера, що містять послідовність-мішень. Ці продукти, у свою чергу, служать матрицею для іншого циклу реплікації. У другому циклі реплікації ланцюг після подовження праймера з першого циклу відпалюється з комплементарним їй праймером; у результаті синтезу одержують «короткий» продукт, що як на 5'-, так і на 3'-кінці обмежений послідовностями праймерів або комплементарними їм. Повторювані цикли денатурації, відпалу праймерів і добудування приводять до експонентного нагромадження областімішені, зумовленої праймерами. Проводять достатньо циклів для досягнення бажаної кількості полінуклеотиду, що містить область-мішень нуклеїнової кислоти. Бажана кількість може, змінюватися й визначається призначенням, для якого повинен служити полінуклеотидний продукт. Спосіб PCR добре описаний у керівництвах і відомий фахівцям у даній галузі. Після ампліфікації PCR полінуклеотиди можна виявляти гібридизацією з полінуклеотидомзондом, що утворює стабільний гібрид з послідовністю-мішенню в умовах гібридизації й відмивання від суворих до помірковано суворих. Якщо очікують, що зонди будуть по суті повністю комплементарними (тобто приблизно на 99% або більше) послідовності-мішені, то застосовують суворі умови. Якщо очікують деякі невідповідності, наприклад, очікують, що варіанти штамів будуть приводити до неповної, комплементарності зондів, суворість гібридизації можна зменшувати. Однак вибирають умови, які виключають неспецифічне/випадкове зв'язування. Умови, що впливають на гібридизацію, які вибирають проти неспецифічного зв'язування, відомі в даній галузі й описані, наприклад, в Sambrook et al., (2001). Як правило, більш низька концентрація солі й більш висока температура збільшують суворість зв'язування. 17 Наприклад, звичайно вважають, що суворі умови являють собою інкубації в розчинах, що містять приблизно 0,1×SSC, 0,1% SDS, при температурі інкубації/відмивання приблизно 65°С, а помірковано суворі умови являють собою інкубації в розчинах, що містять приблизно 1-2×SSC, 0,1% SDS, при температурі інкубації/відмивання приблизно 50°-65°С. Умови низької суворості являють собою 2×SSC і приблизно 30°-50°С. Терміни «суворість» або «суворі умови гібридизації» належить до умов гібридизації, що впливає на стабільність гібридів, наприклад, температури, концентрації солі, рН,; концентрації формаміду й т.п.Ці умови емпірично оптимізують для максимізації специфічного зв'язування й мінімізації неспецифічного зв'язування праймера або зонда з іпослідовністю-мішенню нуклеїнової кислоти. Терміни, як використовується, містять у собі позначення умов, при яких зонд або праймер буде гібридизуватися з його послідовністю-мішенню більшою мірою, ніж з іншими послідовностями, що можна виявити (наприклад, щонайменше в 2 рази вище фону). Суворість умов залежить від послідовності й буде розрізнятися в різних обставинах. Більш довгі послідовності специфічно гібридизуються при більш високих температурах. Як правило, вибирають умови суворості приблизно на 5°С нижче, ніж температурна точка плавлення (Тm) для конкретної послідовності при певній іонній силі й рН. Тm являє собою температуру (при певних іонній силі й рН), при якій 50% комплементарної послідовності-мішені гібридизується з точно співпадаючим зондом або праймером. Як правило, суворими умовами є такі, при яких концентрація солі становить менше ніж приблизно 1,0Μ іона Na+, звичайно, концентрація іона Na+ (або інших солей) становить приблизно 0,01-1,0Μ при рН7,08,3, а температура становить щонайменше приблизно 30°С для коротких зондів або праймерів (наприклад, 10-50 нуклеотидів) і щонайменше приблизно 60°С для довгих зондів або праймерів (наприклад, більше ніж 50 нуклеотидів). Суворих умов можна досягати також додаванням дестабілізуючих засобів, таких як формамід. Характерні умови з низькою суворістю або «умови зниженої суворості» містять у собі гібридизацію в буферному розчині 30% формаміду, 1М NaCl, 1% SDS при 37°С і відмивання в 2×SSC при 40°С. Характерні умови з високою суворістю містять у собі гібридизацію в 50% формаміді, 1М NaCl, 1% SDS при 37°С і відмивання в 0,1×SSC при 60°С. Способи гібридизації добре відомі в даній галузі й описані, наприклад, в Ausubel et al., 1998 і Sambrook et al, 2001. Термін «антиген» належить до речовини, здатної запускати імунна відповідь у хребетного, що приводить до продукування антитіла як частини захисту проти зазначеної речовини. Антигени можуть являти собою білки вірусу, які можуть провокувати продукування антитіла, наприклад, у клітинах крові, клітинах лімфатичних вузлів і селезінці хребетних. Термін «антитіло» включає антигензв'язувальні пептиди й належить до антитіл, моноклональних антитіл, повнорозмірному імуноглобуліну або ан 94035 18 титіла, або будь-якого функціонального фрагмента молекули імуноглобуліну. Приклади таких пептидів включають повні молекули антитіла, фрагменти антитіл, такі як Fab, F(ab')2, що визначають комплементарність області (CDR), VL (варіабельну область легкого ланцюга), VH (варіабельну область важкого ланцюга) і будь-яке їхнє сполучення або будь-яку іншу функціональну частину пептиду антитіла. Термін «антитіло» належить до поліпептиду, який в основному кодується геном імуноглобуліну або генами імуноглобулінів, або його фрагментів, які специфічно зв'язують і впізнають аналіт (антиген). Однак у той час як різні фрагменти антитіл можна визначити виходячи з розщеплення інтактного антитіла, фахівець у даній галузі буде брати до уваги, що такі фрагменти можна синтезувати de novo, або хімічно, або з використанням способів рекомбінантної ДНК. Таким чином, термін антитіло, як використовується в даному описі, містить у собі також фрагменти антитіл, такі як одноланцюжковий Fv, химерні антитіла (тобто які містять константні й варіабельні області з різних видів), гуманізовані антитіла (тобто які містять визначаючу комплементарність область (CDR) з джерела, яке не належить до людини) і гетерокон'юговані антитіла (наприклад, біспецифичні антитіла). Терміни «в основному чистий» і «виділений» використовуються взаємозамінно й описують білок, пептид або нуклеїнову кислоту, які є в основному відділеними від інших (суб)клітинних компонентів, що супроводжують їх у природі. Термін охоплює нуклеїнову кислоту або білок, видалені з їхнього природного оточення, і містить у собі рекомбінантні або клоновані ізоляти ДНК і хімічно синтезовані аналоги, або аналоги, біологічно синтезовані в гетерологічних системах. Як правило, термін належить до очищених білка й нуклеїнової кислоти, які володіють чистотою щонайменше приблизно 75%, наприклад, 85%, 95% або 98% по масі. Незначні зміни або хімічні модифікації звичайно розділяють одну і ту ж поліпептидну або нуклеотидну послідовність. В основному чистий білок або нуклеїнова кислота звичайно будуть містити приблизно 85-100% (мас/мас.) зразка білка або нуклеїнової кислоти, більш звичайно, приблизно 95%, і переважно будуть володіти більш ніж приблизно 99% чистотою. Чистоту або гомогенність білка або нуклеїнової кислоти можна показати рядом способів, добре відомих у даній галузі, таких як електрофорез зразка білка в поліакриламідному гелі з наступною візуалізацією окремої смуги поліпептиду після фарбування, або електрофорез зразка нуклеїнової кислоти в агарозному гелі з наступною візуалізацією окремої смуги полінуклеотиду в агарозному гелі після фарбування. «Фарбування» або може позначати застосування неспецифічних для пептиду або нуклеїнової кислоти барвників, таких як барвники зі сріблом і Кумасі, або барвники бромід етидію й SYBR®, або може позначати застосування специфічного для пептиду або нуклеїнової кислоти фарбування, такого як контактування пептиду з антитілом і візуалізація антитіла з використанням міченого вторинного антитіла (наприклад, кон'югованого з лужною 19 фосфатазою) у випадку білків або пептидів, або контактування нуклеїнової кислоти з комплементарним зондом, міченим для візуалізації наявності гібридизації між нуклеїнової кислотою й зондом. Для конкретних цілей можна забезпечити більше високе розрізнення з використанням для очищення високоефективної рідинної хроматографії (ВЕРХ) або подібних способів. Такі способи знаходяться в сфері загальновідомих загальних знань (див., наприклад, Katz, et al., 1998). Ідентифікація й таксономія Винахід належить до виділеного вірусу, що містить щонайменше одну послідовність нуклеїнової кислоти згідно з SEQ ID NO:1 і/або SEQ ID NO:2 і послідовності, що володіють щонайменше 30%, 35%, 40%, 45%, 50%, 55%, 60% або переважно 65% гомологією нуклеотидної послідовності з ними. Як зазначено вище, полінуклеотидиі володіють «гомологічними» послідовностями, якщо послідовність нуклеотидів у двох послідовностях є однаковою при вирівнюванні для максимальної відповідності. Пошуками BLAST з використанням нуклеотидних послідовностей, отриманих з ізоляту вірусу ToTV, не виявили значної гомології з якийнебудь із відомих^ вірусних або невірусних послідовностей у базах даних GenBank і EMBL. Найбільший відсоток гомології, виявлений таким чином, становив 46,5% між мотивами гелікази в ORF1 SEQ ID NO:2, що відповідають RNA1 ToTV, з мотивами гелікази інших вірусів рослин (див. приклади нижче). Варто розуміти, що гомологія може бути великою,: коли дві послідовності порівнюють у невеликому вікні порівняння, оскільки часто можна знайти локальні області подібності послідовності при порівнянні двох довгих нуклеотидних послідовностей. Однак фахівець у даній галузі усвідомлює, що гомологія послідовності вимагає встановлення загальних мотивів між послідовностями, серед яких ідентичність послідовності локально може бути настільки високою,·як від 35 до 100%, однак може бути настільки низькою, як 10-20% в інших частинах геномної послідовності iз тієї ж самою ORF. Таким чином, коли в даному описі зазначено, що послідовність має гомологію нуклеотидної послідовності щонайменше 50% з SEQ ID NO:1 або SEQ ID NO:2, це може належати до гомології послідовності між областями серед загальних моти 94035 20 вів, де гомологія є найбільшою, а також між кодуючою послідовністю двох гомологічних білків. Необхідно відзначити, що гомологія послідовності може розрізнятися між різними генами в геномі (див. приклади). Як виявлення споріднення між знову ідентифікованим ізолятом вірусу ToTV і іншими вірусами (включаючи віруси, що підлягають порівнянню з ним) можна звичайно провести філогенетичний аналіз на основі (частини) інформації про геномної послідовності вірусів. Декілька таких аналізів представлені в прикладах нижче. Інформація, отримана таким чином, далі вказує на те, що ToTV має найвищий рівень гомології з вірусами з родів Sequivirus і Waikavirus (Sequiviridae) і родів Cheravirus і Sadwavirus. Віруси із цих родів розрізняють на основі числа вірусних РНК - Sequiviridae володіє 1 РНК, у той час як Cheravirus і Sadwavirus володіють двома РНК; і числом білків капсиду - Sadwavirus володіє двома СР, Cheravirus - трьома СР. Ці критерії дозволяють припускати, що вірус томату торрадо найбільш ймовірно входить у групу роду Cheravirus. Однак філогенетичні аналізи з використанням декількох різних мотивів з можливих білків RdRp і гелікази, розташовують ToTV чітко окремо від родів Cheravirus і Sadwavirus і фактично дозволяють припускати, що він є більш близько спорідненим з Sequiviridae. На жаль, у цей час доступні повні послідовності тільки одного Cheravirus (CRLV) і двох Sadwavirus (SDV і SMoV). Попередні дані про переносника дозволяють припускати, що ToTV можуть переносити білокрилки, у той час як природні переносники CRLV і Sadwavirus невідомі. На основі доступної в цей час інформації, включаючи інформацію про послідовності, так само як додаткову таксономічну інформацію, як представлено в таблиці 1 нижче, знову виявлений вірус, найбільше імовірно, являє собою новий рід. Таблиця 1: Таксономічні описи для недавно виявленого вірусу ToTV (як перераховано в міжнародно прийнятих способах з керівництва "Matthew's Plant Virology" ("Matthew's Plant Virology", Fourth Edition, Roger Hull (ed.) Academic Press, San Diego, p 15, таблиця 2.1) (Нумерацію з таблиці в Matthews дотримують у таблиці 1 нижче). Таблиця 1 І Властивості віріонів А. Морфологічні властивості віріонів 1. Розмір 28нм 2. Сферичний (ікосаедричний) за формою 3. Відсутність зовнішньої оболонки В. Фізичні властивості віріонів (не досліджували) С. Властивості геному 1. Нуклеїнова кислота РНК-типу 2. Одноланцюжкова РНК 3. Лінійна РНК 4. Позитивне смислове кодування 5. Щонайменше 2 фрагменти (РНК 2; SEQ ID NO:1 і РНК 1; SEQ ID NO:2) 21 94035 22 Продовження таблиці 1 6. 5,5т.н. (більш точно, 5,2т.н. або 5389 нуклеотидів, крім полі(А)хвоста), і 8т.н. (більш точно, 7,7т.н. або 7793 нуклеотидів, крім полі(А)хвоста) 7. 5'-кінцевий кеп невідомий 8. Ковалентно приєднаний до 5'-кінця поліпептид невідомий 9. Полі(А)хвіст присутній в обох фрагментах 10. Порівняння нуклеотидної послідовності: - РНК 1 містить ORF1 з мотивами, типовими для гелікази (Неl), протеази й РНК-залежної РНК-полімерази (RdRp). Рівень гомології з мотивами гелікази й RdRp інших вірусів докладно описаний у прикладах. - РНК 2 містить дві ORF, одна з яких має на N-кінці мотив, типовий для можливого рухового білка (МР) і містить на С-кінці послідовності для трьох білків капсиду (оболонки) (СР). Рівень гомології з білками капсиду інших вірусів докладно описаний у прикладах. D. Властивості білків 1. 3×СР; Неl; RdRp; протеаза; можливий МР 2.СР: 35, 26 і 23кДа 3. (Види функціональної активності див. вище й у прикладах нижче) 4. (Порівняння амінокислотної послідовності див. у прикладах нижче) II. Організація й реплікація геному 1. Організацію геному див. на Фіг.7 6. Цитопатологія: щонайменше епідерміс, або мезофільні клітини, або сусідні клітини флоеми III. Антигенні властивості 1. Немає відомого серологічного споріднення IV. Біологічні властивості 1а. Природне коло хазяїв: томат; 1b. Експериментальне коло хазяїв: Протестовані альтернативні рослини-хазяї для ToTV Місцеві симптоми/ системні симптоми Chenopodium quinoa -lGomphrena globosa -lNicotiana benthamiana -lс Nicotiana clevelandii -lс Nicotiana glutinosa -lс Nicotiana hesperis 67A nl/c, n, mf Nicotiana occidentalis P1 nl/c, n, mf Nicotiana rustica -l-(la) Nicotiana tabacum 'White Burley' -l-(la) Physalis floridana nl c, n, mf, do с-хлороз; n=некроз; І=вогнища; mf=деформація; do=відмирання; la=латентна інфекція, -=відсутність симптомів. 2. Патогенність Симптоми в природній рослині-хазяїні: Некротичні ушкодження, що закінчуються опікоподібним, повним некрозом тканини рослини й загибеллю рослини; концентричні кільця некротичних тканин на плодах. Зв'язок із захворюванням: Торрадо; можливо також шоколадна плямистість; можливо також Марчитець 3. Трофічний вплив на тканини: щонайменше на епідерміс або мезофільні клітини, або сусідні клітини флоеми 4. Способи передачі в природі невідомі 5. Зв'язок з переносником: приблизно білокрилка 6. Географічний розподіл: Середземномор'я; Америка (Центральна Америка, Мексика й південна частина Північної Америки) Автори даного винаходу недавно виявили, що геномні послідовності вірусу, пов'язаного із захворюванням томату в Центральній Америці (наприклад, у Мексиці й Гватемалі) за назвою «Захворювання шоколадної плямистості», «Шоколад» або «Марчитець», є ідентичними послідовностям описаного тут ToTV (див. приклад 2). Таким чином, збудник захворювання, пов'язаний з даним захворюванням, є аспектом даного винаходу. У міру того як більше вірусних послідовностей стануть доступними, як з вірусів, які не належать до ToTV, які можуть бути або можуть не бути близько спорідненими з ним, так і з вірусів, близько споріднених з вірусом ToTV або істотно нагадують його, філогенетичний аналіз буде служити корисним способом визначення об'єму таксону або кладу ToTV на основі філогенетичного споріднення між ізолятами. Філогенетичне споріднення можна визначити, наприклад, на основі будь-який або всіх нуклеотидних послідовностей RNA 1 і/або РНК 2 з вірусного геному або за даними послідовності (гена) білка капсиду. Філогенетичні аналізи добре відомі фахівцеві в даній галузі й можуть, наприклад, містити в собі аналіз шляхом реконструкції дерев на основі відстані (наприклад, від сусідніх вузлів), способа 23 ми максимальної правдоподібності або економії з використанням таких програм, як Clustal (Thompson et al., 1997), PAUP (Swofford, 2000) або PHYLIP (Felsenstein, 1989). Щоб провести аналіз філогенетичного споріднення між новим ізолятом, послідовністю ToTV, як представлено в даному описі, і контрольними послідовностями з вірусних штамів, наприклад, з баз даних GenBank, EMBL або DDBJ, геномну РНК із зазначеного нового ізоляту можна виділити безпосередньо з інфікованих рослин, і геномні послідовності можна ампліфікувати з неї. Способи ампліфікації PCR зі зворотною транскрипцією (RT-PCR) можна виконувати, наприклад, з використанням вироджених олігонуклеотидних праймерів, де такі праймери, наприклад, здатні діяти як праймер для ампліфікації послідовностей нуклеїнової кислоти з геномів дивергентних ізолятів ToTV, так само як з геномів близько споріднених видів вірусів. Переважно, але не обов'язково, у такий спосіб можна одержати продукти ампліфікації повнорозмірного геному з контрольних штамів (наприклад, дивергентних ізолятів), тестованих ланцюгів (можливих ізолятів ToTV) і близько споріднених видів. Переважно, для порівняння ампліфікують конкретні цікавлячі генетичні області. Продукти ампліфікації (ДНК) потім можна секвенувати, наприклад, прямим секвенуванням дволанцюжкових нуклеотидів з використанням флуоресцентно мічених зупиняючих дидезоксинуклеотидів (Smith et al., 1986) за допомогою вироджених олігонуклеотидних праймерів, використовуваних для RT-PCR. Редагування, аналіз, необов'язкове передбачування амінокислотної послідовності й вирівнювання нуклеотидної послідовності можна проводити за допомогою доступних пакетів програм, таких як пакет для аналізу послідовності LaserGene версії 5 (DNASTAR, Inc., Madison, Wis.) і програмне забезпечення IntelliGenetics Gene Works версії 2.5.1 (IntelliGenetics, Mountain View, Calif.). Потім філогенетичні аналізи можна доповнити філогенетичним аналізом з використанням, наприклад, програмного забезпечення для пошуку максимальної економії (PAUP) за допомогою алгоритму сусідніх вузлів з використанням абсолютних відстаней з наступним евристичним пошуком і 1000 незалежними повторами, і можна одержати філогенетичне дерево аналізом максимальної економії вирівнюваних безперервних нуклеотидних або амінокислотних послідовностей, при якому в таких деревах номера, як правило, являють собою рівні достовірності після повторів. Після 1000 повторів рівень достовірності вищий 60%, переважно вищий 70%, більш переважно вищий 80%, 90%, 95% або 98% у філогенетичному дереві вважають достатнім доказом правильного філогенетичного висновку (вміщення ізолятів у конкретний клад), за умови, що дерево є досить розгалуженим, у силу чого, не обов'язково, розгалуження можна поліпшити утворенням кореня з прийнятними видами поза групами. Таким чином, можна визначити, які ізоляти є найбільш близько спорідненими з послідовностями ToTV, як представлено в даному описі. Споріднення, як правило, виражають у переведенні на 94035 24 відсоток подібності послідовностей, позначений тут гомологією послідовності. Хоча філогенетичні аналізи надають прийнятний спосіб ідентифікації вірусу у випадку достатньої гомології нуклеїнових кислот, для відомих вірусів у даному описі також представлено декілька інших, можливо, більш прямих, хоча декілька більш грубих способів для ідентифікації зазначеного вірусу або вірусних білків, або нуклеїнових кислот із зазначеного вірусу. Як емпіричне правило, вірус ToTV можна ідентифікувати по відсотках гомології підлягаючих ідентифікації вірусних білків або нуклеїнових кислот у порівнянні з вірусними білками або нуклеїновими кислотами, ідентифікованими відповідно до винаходу по послідовності. Загальновідомо, що види вірусів, особливо види РНК-вірусів, часто складаються із псевдовидів, де для кластерів зазначених вірусів показана гетерогенність серед їхніх членів. Таким чином, очікують, що кожний ізолят може володіти певною мірою різним відсотком гомології з послідовностями ізоляту, представленого в даному описі. Таким чином, інші ізоляти вірусу, що володіють достатньою гомологією послідовності з ToTV (наприклад, більш ніж 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 98%, або 99% гомологією послідовності), вважають приналежними до того ж самого вірусу. Таким чином, вірус ToTV за даним винаходом являє собою вірус, що володіє щонайменше 50 або 60% гомологією, переважно щонайменше 70%, більш переважно щонайменше 80%, ще більш переважно щонайменше 90%, ще більш переважно щонайменше 95%, ще більш переважно щонайменше 98% і найбільше переважно щонайменше 99% гомологією з послідовностями білка або нуклеїнової кислоти, представленими у даному описі, і викликаючий симптоми індукованого ToTV захворювання в Solanaceae, більш конкретно, в Solarium і Nicotiana, де симптоми захворювання можуть бути або можуть не бути подібними до симптомів, описаних у даному документі. В Cucurbitaceae вірус, очевидно, не викликає видимих симптомів, хоча вірус здатний до розмноження в рослинах. Якщо бажано порівняти виділений ізолят вірусу з послідовностями білка або нуклеїнової кислоти, описаними в даному документі, винахід надає виділений вірус (ToTV), який можна ідентифікувати як філогенетично відповідний ToTV шляхом визначення послідовності білка або нуклеїнової кислоти зазначеного виділеного ізоляту вірусу й визначення того, що послідовність зазначеного білка або нуклеїнової кислоти має процентну гомологію послідовності щонайменше 60%, переважно щонайменше, 70%, більш переважно щонайменше 80%, ще більш переважно щонайменше 90%, ще більш переважно щонайменше 95%, ще більш переважно щонайменше 98% і найбільше переважно щонайменше 99% з послідовностями, перерахованими в даному описі. Ізоляти вірусу, що мають окремі білки або нуклеїнові кислоти з більш високою гомологією, ніж ці зазначені максимальні значення, вважають філогенетично відповідними й, таким чином, таксономічно відповідними вірусу ToTV, і як правило, білки 25 будуть кодувати послідовність нуклеїнової кислоти, структурно відповідну послідовності, зазначеної в даному описі. При цьому винахід належить до вірусу, філогенетично відповідному виділеному вірусу, послідовності якого перераховані в даному описі. Варто відмітити, що, подібно до інших вірусів, можна чекати, що буде виявлений певний ступінь мінливості між вірусами ToTV, виділеними з різних джерел. Крім того, нуклеотидні або амінокислотні послідовності вірусу ToTV або його фрагментів, як наведено в даному описі, проявляють, наприклад, менше ніж 95%, переважно менше ніж 90%, більш переважно менше ніж 80%, більш переважно менше ніж 70% і найбільше переважно менше ніж 60% гомологію нуклеотидної послідовності, або менше ніж 95%, переважно менше ніж 90%, більш переважно менше ніж 80%, більш переважно менше ніж 70% і найбільше переважно менше ніж 60% гомологію амінокислотної послідовності з відповідною нуклеотидною або амінокислотною послідовністю будь-якого, що не належить до вірусу ToTV або найближчого спорідненого йому. Відхилення послідовності штамів ToTV у світі можуть бути певною мірою вищі, за аналогією з іншими вірусами рослин. Винахід належить до виділеного вірусу (ToTV), який можна ідентифікувати як філогенетично відповідний цьому шляхом визначення послідовності нуклеїнової кислоти прийнятного фрагмента геному зазначеного вірусу й тестування у філогенетичних аналізах, де одержують дерева з максимальною правдоподібністю з використанням, наприклад, 1000 повторів, як описано вище, і виявлення того, що він є філогенетично більш близько спорідненим з ізолятом вірусу, що містить послідовності з SEQ ID NO:1 або SEQ ID NO:2, як перераховано в даному описі для ToTV, ніж спорідненим з ізолятом вірусу з контрольного штаму, що не належить до ToTV, або найближчого спорідненого йому. Прийнятними фрагментами нуклеїновокислотного геному, кожний з яких можна застосовувати для таких філогенетичних аналізів, є, наприклад, будь-яка частина послідовності нуклеїнової кислоти із фрагментів РНК 5,5т.н. або 8т.н. (відповідно позначених тут РНК 2 і РНК 1), як описано в прикладі. Із забезпеченням інформації про послідовності даного вірусу ToTV, винахід надає діагностичні засоби й способи для використання при детекції вірусу ToTV у зразку. Переважно, детекцію вірусу ToTV проводять за допомогою реагентів, які є найбільш специфічними для вірусу ToTV. Це в жодному разі, однак, не виключає можливості використовувати замість цього менш специфічні, але в міру перехресно реактивні реагенти, наприклад, тому що вони є більш легко доступними й підходять для розв'язуваного завдання. Винахід належить, наприклад, до способу детекції присутності ToTV у рослинах, переважно в рослинах томату, більш переважно в рослинах S.lycopersicum. Зазначений спосіб може, наприклад, включати визначення в зазначеному зразку 94035 26 присутності вірусу ToTV або його компонента за допомогою реакції зазначеного зразка зі специфічною для ToTV нуклеїновою кислотою або антитілом за винаходом. Хоча контактна інфекція індикаторної рослини також є прийнятним способом для детекції присутності вірусу в тестованій рослині. Винахід належить до часткової нуклеотидної послідовності нового виділеного вірусу (який називається тут також вірус ToTV) і до специфічних для вірусу ToTV компонентів або їхніх синтетичних аналогів. Додаткові геномні послідовності вірусу ToTV, крім представлених у даному описі, можна визначити способами секвенування, відомими фахівцям у даній галузі. Тепер, коли за даним винаходом наданий вірус ToTV, так само як його часткові геномні послідовності, визначення геномної послідовності легко доступно фахівцеві в даній галузі. Ці способи містять у собі, наприклад, способи, описані в прикладі нижче. Як правило, такі способи секвенування містять у собі виділення нуклеїнових кислот з вірусного геному способами виділення нуклеїнової кислоти й визначення нуклеотидної послідовності виділеної нуклеїнової кислоти, наприклад, способами дидезокси-зупинки ланцюга (Sanger et al., 1977), не обов'язково, які випереджають зворотною транскрипцією РНК у ДНК. Винахід, серед -іншого-, належить до виділеної або рекомбінантнаї нуклеїнової кислоти або її специфічного для вірусу функціонального фрагменту, які можна одержати з вірусу за винаходом. Виділені або рекомбінантні нуклеїнові кислоти містять у собі послідовності, перераховані в даному описі, або послідовності гомологів, здатні гібридизуватися з ними в суворих умовах. Зокрема, винахід належить до праймерів і/або зондів, придатних для ідентифікації нуклеїнової кислоти вірусу ToTV. Додаткові зонди й праймери, здатні гібридизуватися з послідовністю нуклеїнової кислоти вірусу ToTV, можна розробляти способами, відомими фахівцеві в даній галузі. Експресуючі вектори й експресія кодуючих генів вірусу Крім того, винахід належить до експресуючого вектора, що містить нуклеїнову кислоту за винаходом. По-перше, експресуючі вектори, такі як плазмідні вектори, що містять (частини) дволанцюжкової послідовності вірусного геному ToTV, вірусні вектори, що містять (частини) геному ToTV (наприклад, без обмеження, вірус осповакцини, ретровіруси, бакуловірус), або вірус ToTV, що містить (частини) геному інших вірусів або інших патогенів, є частиною даного винаходу. Експресуючий вектор може містити геномну послідовність ToTV або її частину, що перебуває під контролем регуляторного елемента, такого як промотор, або є функціонально пов'язаною з ним. Фрагмент ДНК, позначений як промотор, є відповідальним за регуляцію транскрипції ДНК у мРНК. Експресуючий вектор може містити один або декілька промоторів, що підходять для експресії гена, переважно, для експресії гена, що кодує вірусний білок, у клітинах рослин, клітинах грибів, бактеріальних клітинах, клітинах дріжджів, клітинах комах 27 або інших еукаріотичних клітинах. Експресуючі вектори за винаходом дуже корисні для одержання антигенів вірусу в системах експресії гена. Винахід належить також до клітини-хазяїна, що містить нуклеїнову кислоту або експресуючий вектор за винаходом. Плазмідні або вірусні вектори, що містять нуклеїнові кислоти, які кодують білкові компоненти вірусу ToTV, можна одержати в прокаріотичних клітинах для експресії компонентів у прийнятних типах клітин (клітинах рослин, клітинах грибів, бактеріях, клітинах комах, клітинах рослин або інших еукаріотичних клітинах). Плазмідні або вірусні вектори, що містять повнорозмірні або часткові копії геному вірусу ToTV, можна одержати в прокаріотичних клітинах для експресії вірусних нуклеїнових кислот in vitro або in vivo. Способи виділення й очищення ToTV Вірус ToTV можна виділити з інфікованих рослин або інших джерел будь-яким доступним способом. Виділення може містити в собі очищення або часткове очищення вірусних частинок ToTV з прийнятного джерела. Для виділення й очищення вірусу доступний широкий ряд способів (наприклад, див. Dijkstra and De Jager, 1998). Очищення ToTV можна, наприклад, проводити з використанням загальноприйнятих способів, наприклад, для неповірусів або лютеовірусів (за допомогою органічних розчинників) (див., наприклад, Walker, 2004). Хоча такі способи можуть приводити до втрати інфекційності вірусу, ці способи ще можуть бути застосовними для одержання вірусного матеріалу для інших цілей. Переважно, щоб підтримувати цілісність вірусу, для очищення ToTV використовують спосіб м'якого очищення. Такий спосіб м'якого очищення може містити в собі, наприклад, гомогенізацію інфікованого матеріалу рослини (такого як листя) у буфері для гомогенізації (що містить, наприклад, 0,1Μ буфер Тріс-НСІ при рН приблизно 8, приблизно 20мМ сульфіт натрію [Na2SO3], приблизно 10мМ діетилдитіокарбамат натрію [Na-DIECA] і приблизно 5мМ етилендіамінтетраацетат натрію (Na-EDTA)), відділення дебриса центрифугуванням (наприклад, протягом 30хв. при 49000g), вміщення супернатанту на прийнятну подушку сахарози (наприклад, 20%) і осадження вірусних частинок (наприклад, центрифугуванням протягом 1,5 годин при 70000g). Потім осад, що містить вірусні частинки, можна ресуспендувати у прийнятному буфері (наприклад, у Тріс-НСІ, рН8), і фракцію, яка містить вірус, можна відділити від іншої суспензії з використанням центрифугування в градієнті щільності сахарози (наприклад, 10-40% сахароза в буфері для гомогенізації, центрифугування протягом 2 година при 110000g). Фракцію, яка містить вірус, можна визначити з використанням експериментів по інфекції. Додаткове очищення можна проводити з використанням центрифугування в градієнті щільності сульфату цезію (наприклад, 10-40% градієнт сульфату цезію в Тріс-НСІ, рН8, центрифугування протягом 16 годин при 125000g). Збагачені вірусом смуги можна потім збирати із градієнта й додатково концентрувати центрифугуванням або діалізувати (наприклад, проти 0,1Μ Тріс-НСІ, рН8). 94035 28 Інфекційність ToTV після очищення можна перевірити інокуляцією в чутливу рослину (наприклад, N.hesperis '67A'). Способи очищення пов'язаних з ТоТУ білків і амінокислотного секвенування Після одержання очищеного або частково очищеного вірусу ToTV можна одержати в основному чистий препарат пов'язаних з вірусом білків (наприклад, кодованих вірусом білків). Хоча в даній галузі відомі численні способи й стратегії очищення білка, найприйнятнішим є очищення вірусних білків ToTV, таких як білки вірусної оболонки, або електрофорезом з використанням, наприклад, поліакриламідного гелю з додецилсульфатом натрію (SDS-PAGE), або афінною хроматографією. Кожний із цих способів буде описаний нижче. Білки ToTV, які представляють інтерес, можна розділяти електрофорезом з використанням, наприклад, трицин-SDS-PAGE (Schagger і Von Jagow, 1987) або гліцин-SDS-PAGE (Laemmli, 1970). Можна, звичайно, застосовувати інші системи електрофорезу, здатні розділяти різні білки, які містяться в ізоляті вірусу, або транскрибовані з його геному й експресовані в прийнятній експресуючій системі, такі як не денатуруючий гельелектрофорез. Область гелю для PAGE, що містить заданий білок, можна вирізати, і задані поліпептиди можна елюювати звідти, наприклад, з використанням пристрою Elutrap® (Schleicher & Schuell, Dassel, Germany). Заданий білок можна ідентифікувати по його рухливості щодо контрольних поліпептидів у гелі. Для збільшення ступеня очищення можна проводити електрофорез елюйованого білка в другому гелі для SDS-PAGE і елюювати його другий раз. Білок або пептид, який міститься у вирізаному фрагменті гелю, можна потім знову елюювати, і він є придатним для використання при імунізації або секвенуванні білка. Білки ToTV, які представляють інтерес, можна очистити також афінною хроматографією з використанням антитіла (такого як моноклональне антитіло), що специфічно зв'язується з білком ToTV. Антитіло можна ковалентно приєднати до твердих підкладок, таких як целюлози, полістирол, поліакриламід, поперечно-зшитий декстран, гранульована агароза або скло з контрольованим розміром пор, з використанням біфункціональних зшивних засобів, які реагують із функціональними групами на підкладці й функціональних групах (тобто реакційноздатними бічними ланцюгами амінокислот) у молекулі антитіла. Такі способи легко доступні фахівцеві в даній галузі. Отриману несучу антитіло тверду фазу приводять у контакт із очищеним або частково очищеним вірусом у відновлювальних умовах з використанням рН, іонної сили, температури й тривалості обробки, що дозволяють білку, який представляє інтерес, зв'язуватися з іммобілізованим антитілом. Вірус або білок елююють з колонки пропускання елюента, що дисоціює водневі зв'язки з підкладкою. Звичайно застосовуваними елюентами є буфери з конкретним рН або більш ніж приблизно 2Μ розчини NaCl. Способи здійснення афінної хроматографії з використанням антитіл, так само як інші способи імуноафінною очищення білків (таких як білки віру 29 сного капсиду), добре відомі в даній галузі (див., наприклад, Harlow and Lane, 1988). По наданих тут поясненнях фахівець у даній галузі може виділити специфічний для вірусу ToTV білок, визначити амінокислотну послідовність, наприклад, N-кінцевої частини зазначеного білка, сконструювати набір вироджених зондів (через виродженість генетичного коду) для гібридизації із ДНК, що кодує область зазначеного білка, з використанням цих зондів для масиву генів у геномній бібліотеці, отриманої з вірусу, одержати позитивну гібридизацію й локалізувати відповідні гени. Потім фахівець у даній галузі може ідентифікувати структурну область гена й, не обов'язково, області послідовності проти ходу транскрипції й по ходу транскрипції від неї. Після цього фахівець у даній галузі може встановити правильну послідовність амінокислотних залишків, що формують білок. Одержання антитіла Антитіла, або моноклональні, або поліклональні, можна одержати до очищеного або частково очищеного білка або фрагмента пептиду вірусу ToTV великою кількістю способів, відомих фахівцям у даній галузі, включаючи ін'єкцію білка як антигена твариною, злиття гібридоми, і рекомбінантні способи, що включають бактеріальні або фагові системи (див. Marks et al, 1992a; Marks et al., 1992b; Lowman et al., 1991; Lerner et al., 1992, де в кожному із цих посилань описані прийнятні способи). Антитіла проти вірусних частинок, білків або пептидів вірусу можна одержати імунізацією прийнятного хазяїна - хребетного, переважно ссавця, наприклад, кроликів, кіз, щурів, курей і мишей, частинками, білками або пептидами самостійно або в об'єднанні з ад'ювантом. Звичайно передбачають дві або більше імунізації, і кров або селезінку збирають через декілька днів після останньої ін'єкції. У випадку поліклональної антисироватки, імуноглобуліни можна осадити, виділити й (афінно) очистити. У випадку моноклональних антитіл, спленоцити звичайно зливають із імморталізованим лімфоцитом, наприклад, мієлоїдною лінією, в умовах для селекції гібридом. Потім гібридоми можна клонувати в умовах лімітуючих розведень і проводити скринінг їх супернатантів по антитілах, що володіють бажаною специфічністю. Способи продукції (моноклональних) антитіл і способи їхнього одержання й застосування в різних способах добре відомі в літературі (див., наприклад патенти США №№4381292, 4451570 і 4618577; Harlow and Lane, 1988; Ausubel, et al, 1998; Rose et al., 1997; Coligan et al., 1997). Звичайно, антитіло, спрямоване проти пов'язаного з вірусом білка, буде володіти афінністю зв'язування щонайменше 1×1051×107М-1. Рекомбінантний білок, отриманий з вірусу ToTV, такий як може бути отриманий шляхом експресії кодуючої білок геномної послідовності вірусу в прийнятній експресуючій системі, є переважними як антиген. Однак можна використовувати також очищені білки. Антигени, що підходять для детекції антитілами, містять у собі будь-який білок ToTV, що з'єднується з будь-яким специфічним до ToTV антитілом ссавця, що піддавався впливу вірусу 94035 30 ToTV або інфікованого ним. Переважні антигени за винаходом містять у собі антигени, що викликають імунну відповідь у ссавців, які піддавалися впливу ToTV, які, таким чином, як правило, найбільше легко впізнають антитіла ссавців. Зокрема, переважні антигени містять у собі білки капсиду ToTV. Найбільш переважними є структурні білки з очищеного вірусу. Способи клонування геномних послідовностей в експресуючі вектори й з них для маніпуляції з геномними послідовностями й для експресії білка, що кодується геномною послідовністю в гетерологічному хазяїні, добре відомі, і ці способи можна використовувати для одержання експресуючих векторів, клітин-хазяїв і клонованих геномних послідовностей, що кодують антигени, де послідовності експресовані в хазяїні для одержання антитіл для застосування в діагностичних аналізах (див., наприклад, Sambrook et al., 2001 і Ausubel, et al, 1998). Для одержання антигенів ToTV можна використовувати велику кількість експресуючих систем. Наприклад, описана велика кількість експресуючих векторів, придатних для одержання білків в E.coli, B.subtilis, дріжджах, клітинах комах, клітинах рослин і клітинах ссавців, кожний з яких можна використовувати для одержання антигену ToTV, що підходить для одержання анти-ToTV антитіла або його фрагмента. Зрозуміло, власне ToTV також можна використовувати як експресуючий вектор для даної мети. Одним із застосувань антитіл за винаходом є скринінг експресуючих бібліотек кДНК для ідентифікації клонів, які містять вставки кДНК, що кодують представляючі інтерес білки або структурно споріднені, перехресно імунореактивні білки. Такі скринінги експресуючих бібліотек кДНК добре відомі в даній галузі (див., наприклад, Young and Davis, 1983), посилання на який наведена в даному контексті, так само як інші опубліковані джерела. Іншим застосуванням цих антитіл є використання в афінній хроматографії для очищення білків ToTV. Ці антитіла також застосовні для аналізу інфекції ToTV. Даний винахід, таким чином, належить до ToTV-специфічного вірусного білка або його фрагменту, надалі в цьому документі позначеному білковою молекулою. Застосовними білковими молекулами є, наприклад, похідні від будь-якої з геномних послідовностей або їхні фрагменти, які можна одержати з вірусу за винаходом. Такі білкові молекули, або їхні антигенні фрагменти, як наведено в даному описі, застосовні, наприклад, у діагностичних способах або наборах, і в діагностичних композиціях. Зокрема, застосовними є такі білкові молекули, що кодуються фрагментами рекомбінантної нуклеїнової кислоти, які ідентифіковані як викликаючі утворення специфічних для вірусу ToTV антитіл, або in vivo (наприклад, для одержання діагностичних антитіл), або in vitro (наприклад, з використанням технології фагового дисплея або іншого способу, застосовного для одержання синтетичних антитіл або їхніх частин). Також в даному документі передбачені антитіла, чи є вони природними поліклональними або 31 моноклональними, або синтетичними антитілами (наприклад, отриманими з (фагової) бібліотеки молекулами, що зв'язуються), які специфічно реагують із антигеном, що містить білкову молекулу або її специфічний для вірусу ToTV функціональний фрагмент, такий як білок капсиду за винаходом. Способи ідентифікації ізоляту вірусу як вірусу ТоТУ Крім детекції вірусу ToTV, що містить у собі діагностичні способи, даний винахід також належить до способів ідентифікації, тобто підтвердження, що даний ізолят є ToTV. Такі способи можуть бути основані на філогенетичному висновку, як описано вище, і визначенні рівня гомології нуклеотидної або амінокислотної послідовності між неідентифікованим ізолятом вірусу й одним або декількома контрольними штамами підтверджених вірусів ToTV і вірусів, які не належать до ToTV. Такі способи можуть, наприклад, містити в собі секвеновані (частини) геному ізоляту вірусу або білка капсиду й порівняння рівня гомології даної послідовності з послідовностями, наданими тут для ToTV. Ізолят, що володіє більш ніж 50% гомологією послідовності з SEQ ID NO:1 або SEQ ID NO:2, як представлено в даному описі, вважають таксономічно відповідним ToTV або приналежним до таксона вірусу ToTV. Такий вірус є частиною даного винаходу. Щоб ідентифікувати вірус як вірус томату торрадо (ToTV), можна також використовувати таксономічні ознаки, як представлено в таблиці 1 вище, і шляхом порівняння визначити, що новий ізолят належить до можливого нового роду, типовим видом якого можна прийняти штам ToTV, ідентифікований по наданих тут послідовностях нуклеїнової кислоти SEQ ID NO:1 і 2, і депонований у німецькій колекції мікроорганізмів і клітинних культур DSM 24 листопада 2004 року під номером для посилання депозитів ToTV-EOl (DSM 16999). Таким чином, не є необхідним проводити порівняння послідовності, щоб визначити те, що вірус є ToTV. Переважніше, спосіб ідентифікації вірусу як вірусу томату торрадо (ToTV) може містити в собі стадії визначення наявності сполучення таксономічних ознак, вибраних із групи, яка складається з a) морфологічних ознак, таких як сферичні частинки віріону без зовнішньої оболонки діаметром приблизно 28нм; b) ознак властивостей геному, таких як властивість вірусу володіти одноланцюжковою лінійною позитивною смисловою РНК на основі двох фрагментів РНК, що містять полі(А)-хвости, які кодують полібілки 5,5 і 8кДа, відповідно, і містять кодуючи області або мотиви для 3 білків капсиду, гелікази, протеази, RdRP і можливого рухового білка, де фрагменти РНК і/або полібілки, і/або мотиви мають гомологію, на основі порівняння послідовності, в основному, як описано в даному документі, і c) ознак біологічних властивостей, таких як продукування некротичних ушкоджень і опікоподібних симптомів у томату, володіння колом хазяїв, зв'язком з переносником і/або географічним розподілом в основному, як описано в даному доку 94035 32 менті, і наявність зв'язку із захворюваннями рослин томату, відомими в межах певної місцевості під такими назвами, як Торрадо, Марчитець і/або шоколадна плямистість. Сполученням таксономічних ознак, що приводять до позитивної ідентифікації ізоляту вірусу як такого, що є вірусом томату торрадо (ToTV), є таке сполучення, що вказує на те, що ізолят є більш близько спорідненим (на основі численних таксономічних способів, добре відомих фахівцеві в даній галузі) з ToTV, як описано в даному документі, ніж з іншими вірусами, і де зазначений ізолят переважно викликає в томату симптоми захворювання, типові для торрадо, як описано в даному документі. Таким чином, вірус, який, на основі численних таксономічних аналізів таксономічних ознак, в основному, визначених у таблиці 1, є більш близько спорідненим з вірусом, як визначено в пункті 1 формули винаходу в даному документі, ніж з будьяким іншим вірусом, відомим до дати подачі даної заявки, і який є вірусом, пов'язаним із захворюванням, що викликає некротичні ушкодження в томату, вважають тут таким, що є ToTV і входить в об'єм даного винаходу. Таким чином, винахід належить до ізоляту вірусу, який можна ідентифікувати способом за винаходом як вірус рослин, таксономічно відповідний вірусу, ідентифікованому як можливо приналежний до таксона вірусу ToTV. Залежно від філогенетичного споріднення, або своєрідності стосовно інших вірусних таксонів, таксон вірусу ToTV може бути ізолятом, видом, родом або навіть сімейством вірусу. Альтернативно, способи ідентифікації ізоляту вірусу як вірусу ToTV можуть бути основані на симптоматології, тобто ідентифікації вірусу по пов'язаних з ним симптомах захворювання. Однак у переважному варіанті здійснення як спосіб ідентифікації ізоляту вірусу як вірусу ToTV застосовують антитіла за винаходом, за умови, що ефективно виключають перехресну реактивність таких антитіл для споріднених не стосовних до ToTV штамів. Такі способи містять у собі стадію реакції зазначеного вірусного ізоляту або його компонента з антитілом, як передбачено в даному описі. Реакцією в даному документі називають забезпечення можливості зв'язування антитілоантиген. Це можна здійснити, наприклад, з використанням очищеного або неочищеного вірусу ToTV або його частин (білків, пептидів). Переважно, інфіковані клітини або культури клітин використовують для ідентифікації вірусних антигенів із застосуванням будь-якого прийнятного імунологічного способу. Особливо корисними щодо цього є антитіла, індуковані проти білків капсиду вірусу ToTV. Інші переважні способи ідентифікації ізоляту вірусу як вірусу ToTV містять у собі реакцію зазначеного ізоляту вірусу або його компонента зі специфічним для вірусу полінуклеотидом за винаходом, де полінуклеотид здатний гібридизуватися в суворих умовах з послідовністю нуклеїнової кислоти, вибраної із групи, яка складається з SEQ ID NO:1, SEQ ID NO:2, послідовностей, що володіють 33 щонайменше 60% гомологією нуклеотидної послідовності з SEQ ID NO:1 або SEQ ID NO:2, комплементарних їм ланцюгів і їхніх ToTV-специфічних фрагментів. Таку реакцію гібридизації можна проводити в будь-якому форматі, доступному фахівцеві в даній галузі, і вона, як правило, містить у собі одержання відбитків тканин, способи дотблотингу, Саузерн/Нозерн-блотинг або гібридизацію, гібридизацію in situ, PCR, RT-PCR і т.п. Імунологічні способи детекції Способи за винаходом, якими виявляють антигени, можна, у принципі, здійснювати з використанням будь-якого імунологічного способу, такого як, наприклад, класична імунофлуоресценція (IF), імуногістохімічні способи або порівнянні формати аналізу для детекції антигену. Переважні способи детекції ToTV, основані на детекції білка вірусної оболонки, можуть, наприклад, містити в собі такі способи, як тести преципітації й аглютинації, радіоімуноаналіз (RIA), імунологічний метод мічення золотом, імуносорбційну електронну мікроскопію (ISEM), твердофазний імуноферментний аналіз (ELISA), вестерн-блотинг і імуноблотинг. Прикладами типів імунологічних аналізів, у яких можна використовувати антитіла за винаходом, є конкурентний і не конкурентний імунологічні аналізи або в прямому, або в непрямому форматі. Антитіла можна використовувати в імунологічних аналізах у рідкій фазі або пов'язаними із твердофазним носієм. Крім того, антитіла в даних імунологічних аналізах можна різними способами мітити детектованою міткою. Фахівцям у даній галузі відомі або легко зрозумілі без зайвих експериментів прийнятні формати імунологічних аналізів. Формати аналізів добре відомі в літературі й описані, наприклад, в Harlow and Lane (1988). Велику кількість форматів імунологічного аналізу можна використовувати для відбору антитіл, що специфічно реагують із конкретним поліпептидом за винаходом, таким як білки оболонки вірусу ToTV. Наприклад, для даної мети звичайно використовують твердофазні імунологічні аналізи ELISA. Див. опис форматів імунологічного аналізу й умови, які можна використовувати для визначення вибіркового зв'язування, в Harlow and Lane (1988). Антитіла можна зв'язувати з багатьма різними носіями й використовувати для детекції присутності молекул-мішеней. Альтернативно, антигени можна зв'язувати з багатьма різними носіями й використовувати для детекції присутності антитіла. Приклади добре відомих носіїв містять у собі скло, полістирол, поліпропілен, поліетилен, декстран, нейлон, амілази, природні й модифіковані целюлози, поліакриламіди, агарози й магнетит. Для цілей даного винаходу носій по своїй природі може бути або розчинним, або нерозчинним. Фахівцям у даній галузі відомі інші прийнятні носії для зв'язування антитіл, або вони здатні визначити такі носії з використанням загальноприйнятих експериментів. Аналізи вестерн-блотингом у загальних рисах описані в Harlow and Lane (1988). За даним способом вірусні білки (і інші білки в препараті вірусу) розділяють гель-електрофорезом і переносять на 94035 34 тверду фазу (тобто мембрану, таку як нітроцелюлоза). Послідовно проводять реакцію іммобілізованого антигену з антитілом і системою детекції (наприклад, кон'югованим з лужною фосфатазою вторинним антитілом). Як буде очевидно фахівцеві в даній галузі, переважним буде включення в аналіз відповідних матеріалів для негативного й позитивного контролю (таких як в основному очищений антиген або вірус ToTV). Твердофазні імуноферментні аналізи (ELISA) у загальних рисах описані в Harlow and Lane (1988). Аналіз містить у собі реакцію вірусного компонента (наприклад, білка капсиду) з антитілом. В одному з варіантів здійснення зразок може містити тканину рослини, яку перемелюють і проводять її реакцію з антитілом, яким покрита тверда фаза, така як планшет для тестування. Якщо в зразку присутній вірус, мічене ферментом специфічне антитіло буде зв'язуватися з комплексом антитіло-вірус, і це можна виявляти реакцією ферменту із субстратом з одержанням кольорової реакції. Переважними способами аналізів ELISA є ELISA із прямим сендвічем з подвійним антитілом (DAS) °Clark and Adams, 1977), непрямий ELISA з DAS (Vela et al. 1986), або TAS-ELISA. Фахівцям у даній галузі буде очевидним, що іноді в ELISA або будь-якому іншому типі аналізу буде бажаним визначення присутності більше одного білка вірусного капсиду в одній реакційній суміші, наприклад, шляхом змішування двох або більше антитіл з різними специфічностями й аналізу будь-якого або всіх разом пов'язаних із захисним капсидом білків за винаходом. Способи детекції на основі нуклеїнової кислоти ToTV складається щонайменше із двох рибонуклеїнових кислот (РНК). Існують серйозні вказівки на присутність трьох білків захисного капсиду. Способи, описані вище, спрямовані на імунологічну детекцію вірусних білків для детекції вірусу. Способом рекомбінантної ДІЖ можна одержувати зонди, які прямо або опосередковано гібридизуються з вірусними РНК (або комплементарними їм), або із кДНК, отриманими з них зворотною транскрипцією, які можна використовувати в аналізах для детекції вірусу. Способи ампліфікації нуклеїнової кислоти дозволяють ампліфікацію фрагментів вірусних нуклеїнових кислот, які можуть бути присутнім у дуже малих кількостях. Щоб розробити способи детекції на основі нуклеїнової кислоти, необхідно визначити специфічні для вірусу послідовності, для яких можна потім розробити праймери або зонди. Для детекції ToTV за допомогою ампліфікації і/або гібридизації із зондом нуклеїнової кислоти, білок капсиду ToTV можна секвенувати або, альтернативно, можна виділити вірусну геномну РІЖ із очищеного вірусу, провести зворотну транскрипцію в кДНК і прямо клонувати і/або секвенувати. Або з використанням клонованої нуклеїнової кислоти як зонда для гібридизації, або з використанням інформації про послідовності, отриманої із клону, або шляхом конструювання вироджених праймерів на основі амінокислотної послідовності білка ToTV, можна сконструювати зонди для гібридизації нуклеїнової 35 кислоти і/або праймери для ампліфікації нуклеїнової кислоти й використовувати в аналізі детекції для детекції присутності вірусу в зразку, як визначено в даному описі. Способи за винаходом, який виявляють нуклеїнові кислоти, можна, у принципі, здійснювати з використанням будь-якого способу ампліфікації нуклеїнової кислоти, такого як полімеразна ланцюгова реакція (PCR; Mullis and Faloona, 1987; патенти США №№4683195; 4683202 і 4800159), або з використанням таких реакцій ампліфікації, як лігазна ланцюгова реакція (LCR; Ваrаnу, 1991; ЕР 0320308), самопідтримувана реплікація послідовності (3SR; Guatelli et al., 1990), ампліфікація із заміщенням ланцюгів (SDA; Walker et al., 1992; патенти США №№5270184 і 5455166), система транскрипційної ампліфікації (TAS; Kwoh et al., 1989), з використанням Q-бета реплікази (Lizardi et al., 1988), ампліфікації по типу кільця, що котиться (RCA; патент США №5871921), ампліфікації на основі послідовності нуклеїнової кислоти (NASBA; Compton, 1991), поліморфізму довжини фрагментів розщеплення клевазою (патент США №5719028), ізотермічної й ініційованої химерним праймером ампліфікації нуклеїнової кислоти (ICAN), способу ампліфікації з розгалуженим подовженням (RAM; патенти США №№5719028 і 5942391) або інших прийнятних способів ампліфікації нуклеїнових кислот. Оскільки вірус є РНК-вірусом (тобто послідовності на Фіг.5 і 6 є ДНК-еквівалентами вірусного РНК-геному), прийнятний спосіб детекції може містити в собі виділення вірусних нуклеїнових кислот зі зразка, наприклад, з інфікованої рослини, з використанням способів, відомих по суті фахівцеві в даній галузі (наприклад, Chomczynski and Sacchi, 1987; Boom et al., 1990) або комерційно доступних систем (наприклад, набір для виділення загальної РНК RNeasy або набір для виділення РНК рослин RNeasy plant від QIAGEN Gmb, Hilden, Germany, або High-Pure-RNA-Isolation-Kit® (Roche Diagnostics, відділення F. Hoffmann-La Roche Ltd, Basel, Switzerland). Тотальну РНК можна, наприклад, виділити з матеріалу листка або протопластів клітин рослин, і тотальну РНК, або конкретно вірусну геномну РНК, або її частину, можна потім піддавати зворотній транскрипції в кДНК із використанням, наприклад, зворотної транскриптази вірусу мієлобластозу птаха (AMV) або зворотної транскриптази вірусу лейкемії мишей Молоні (M-MuLV). Прийнятний спосіб може, наприклад, містити в собі змішування в прийнятній водній буферній системі (наприклад, у комерційно доступному буфері для RT) прийнятної кількості тотальних РНК (наприклад, 1-5мкг), прийнятної кількості (наприклад, 10 пмоль) праймера для зворотної транскрипції, прийнятної кількості dNTP і зворотної транскриптази, денатурацію нуклеїнових кислот кип'ятінням протягом 1 хвилини й охолодженням їх на льоді з наступною зворотною транскрипцією, наприклад, при 45°С протягом 1 години, як рекомендовано для конкретної використовуваної зворотної транскриптази, для одержання кДНК-копій вірусних послідовностей. 94035 36 Як праймер для зворотної транскрипції можна використовувати полінуклеотид за даним винаходом, наприклад, 18-26-вимірний олігонуклеотид, що містить нуклеотидну послідовність, комплементарну геномній послідовності ToTV, або переважно, щонайменше здатну гібридизуватися в суворих умовах з послідовністю нуклеїнової кислоти SEQ ID NO:1 або SEQ ID NO:2, або її ToTVспецифічний фрагмент. Альтернативно, можна використовувати неспецифічний поліТ-праймер (олігоdТ-праймер), щоб починати зворотну транскрипцію з мотивів полі-Α РНК. Після RT-стадії отриману кДНК можна ампліфікувати за допомогою PCR, використовуючи, наприклад ДНК-полімерази Pfu і Taq і праймери для ампліфікації, специфічні для послідовностей вірусної геномної кДНК. Для RT-PCR можна використовувати також повні комерційно доступні системи (наприклад, системи для RT-PCR Access і AccessQuick™ від Promega [Madison WI, USA], або система для RT-PCR в одній пробірці Titan™, або системи для двоступінчастої RT-PCR, що поставляють Roche Diagnostics [відділення F. HoffmannLa Roche Ltd, Basel, Switzerland]). Щоб ампліфікувати нуклеїнову кислоту з невеликою кількістю розбіжностей з одним або декількома праймерами для ампліфікації, реакцію ампліфікації можна проводити в умовах зниженої суворості (наприклад, ампліфікація PCR з використанням температури відпалу 38°С, або в присутності 3,5мМ MgCl2). Фахівець у даній галузі буде здатний вибрати умови прийнятної суворості. Праймери в даному описі вибирають «в основному» комплементарними (тобто щонайменше на 65%, більш переважно щонайменше на 80% точно комплементарними) їхнім галузям-мішеням, які присутні на різних ланцюгах кожної конкретної послідовності, що підлягає ампліфікації. Можна використовувати послідовності праймерів, що містять, наприклад, залишки інозиту або невизначені основи, або навіть праймери, що містять одну або декілька розбіжностей у порівнянні з послідовністю-мішенню. Як правило, послідовності, що володіють щонайменше 65%, більш переважно щонайменше 80% гомологією з олігонуклеотидними послідовностями-мішенями ДНК або РНК, вважають прийнятними для використання в способі за даним винаходом. Розбіжності послідовності також не є критичними при використанні умов гібридизації низької суворості. Детекцію продуктів ампліфікації можна, у принципі, здійснювати будь-яким прийнятним способом, відомим у даній галузі. Ампліфіковані фрагменти можна забарвлювати прямо або мітити радіоактивними мітками, антитілами, люмінесцентними барвниками, флуоресцентними барвниками або ферментативними реагентами. Прямі барвники для ДНК містять у собі, наприклад, інтеркалюючі барвники, такі як барвники акридин жовтогарячий, бромід етидію, моноазид етидію або барвник Хехста. Альтернативно, фрагменти ДНК або РНК можна виявляти за допомогою включення мічених основ dNTP у синтезовані фрагменти. Мітки для детекції, які можуть бути пов'язаними з 37 нуклеотидними основами, містять у собі, наприклад, флуоресцеїн, барвник ціанін, дигоксигенін (DIG) або бромдезоксіуридин (BrdUrd). При використанні систем детекції, основаних на зондах, пгийнятний спосіб детекції для застосування за даним винаходом може, наприклад, містити в собі формат ферментного імунологічного аналізу (ЕІА) (Jacobs et al., 1997). Для здійснення детекції способом процедури ЕІА, або прямий, або зворотний праймер, використовуваний у реакції ампліфікації, може містити групу для захоплення, таку як група біотину для іммобілізації ампліконів PCR ДНК-мішені, наприклад, на покритих стрептавідином ямках планшета для мікротитрування або на покритих стрептавідином Dynabeads® (Dynal Biotech, Oslo, Norway) для наступної детекції ЕІА ампліконів ДНК-мішені. Фахівцеві в даній галузі зрозуміло, що для іммобілізації ампліконів ДНКмішені у форматі ЕІА можна використовувати інші групи. Зонди, застосовні для детекції послідовностей нуклеїнової кислоти-мішені, як описано в даному документі, переважно зв'язують тільки щонайменше частину області послідовності нуклеїнової кислоти, ампліфікованої способом ампліфікації нуклеїнової кислоти. Фахівці в даній галузі без зайвих експериментів можуть одержати прийнятні зонди для детекції на основі нуклеотидної послідовності нуклеїнової кислоти-мішені, як описано в даному документі. Також як типоспецифічні зонди для детекції в способі за винаходом, відповідно, можна використовувати комплементарні нуклеїновій кислоті-мішені нуклеотидні послідовності, або ДНК, або РНК, або хімічно синтезовані аналоги, за умови, що такий комплементарний ланцюг ампліфікують у використовуваній реакції ампліфікації. Прийнятні способи детекції для застосування в даному винаході, можуть, наприклад, містити в собі іммобілізацію ампліконів і гібридизацію із зондами послідовностей їх нуклеїнової кислоти, наприклад, за допомогою нозерн- і Саузернблотинга. Інші формати можуть містити в собі формат ЕІА, як описано вище. Для полегшення детекції зв'язування специфічні зонди для детекції амплікону можуть містити групу мітки, таку як флуорофор, хромофор, фермент або радіоактивна мітка, так щоб полегшувати моніторинг зв'язування зондів із продуктом реакції ампліфікації. Такі мітки добре відомі фахівцям у даній галузі й містять у собі, наприклад, ізотіоціанат флуоресцеїну (FITC), β-галактозидазу, пероксидазу хрону, стрептавідин, біотин, дигоксигенін, S, 4С, 32Р або 125І. Інші приклади будуть очевидні фахівцям у даній галузі. Детекцію можна проводити також шляхом так званого аналізу блота з переверненими доріжками (RLB), такого як, наприклад, описаний Van den Brule et al. (2002). Для даної мети зонди для RLB переважно синтезують із 5'-аміногрупою для наступної іммобілізації, наприклад, на покритих карбоксилом нейлонових мембранах. Перевагами формату RLB є простота системи і її швидкість, що, таким чином, дозволяє більш високу продуктивність обробки зразків. 94035 38 Використання нуклеїновокислотних зондів для детекції фрагментів РНК або ДНК добре відомо в даній галузі. Більшість даних способів містять у собі гібридизацію нуклеїнової кислоти-мішені із зондом з наступними відмиваннями після гібридизації. Специфічність, як правило, є функцією відмивань після гібридизації, де критичними факторами є іонна сила й температура кінцевого розчину для відмивання. Для гібридів нуклеїнової кислоти Тm можна приблизно вивести з рівняння Meinkoth-Wahl (1984): Tm=81,5°C+16,6 (log M)+0,41(%GC)-0,61 (%form)-500/L, де М являє собою молярність моновалентних катіонів, %GC являє собою відсоток нуклеотидів гуанозину й цитозину в нуклеїновій кислоті, %form являє собою процентний вміст формаміду в розчині для гібридизації, і L являє собою довжину гібрида в парах основ. Тm являє собою температуру (при певній іонній силі й рН), при якій 50% комплементарної послідовності-мішені гібридизується з повністю співпадаючим зондом. Тm зменшують приблизно на 1°С для кожного 1% невідповідності; таким чином, умови гібридизації і/або відмивання можна регулювати для гібридизації з послідовностями бажаної ідентичності. Наприклад, при пошуку послідовностей з ідентичністю >90% Тm можна збільшити на 10°С. Як правило, суворі умови вибирають приблизно на 5°С нижче температурної точки плавлення (Тm) для конкретної послідовності й комплементарної їй при певній іонній силі й рН. Однак для надзвичайно суворих умов можна використовувати гібридизацію і/або відмивання на 1, 2, 3 або 4°С нижче температурної точки плавлення (Тm); для помірно суворих умов можна використовувати гібридизацію і/або відмивання на 6, 7, 8, 9 або 10°С нижче температурної точки плавлення (Тm); для умов з низькою суворістю можна використовувати гібридизацію і/або відмивання на 11, 12, 13, 14, 15 або 20°С нижче температурної точки плавлення (Тm). Звичайному фахівцеві в даній галузі буде зрозумілим, що з використанням рівняння, складів для гібридизації й відмивання, і бажаної Тm, по суті описують варіанти суворості гібридизації і/або розчинів для відмивання. Якщо бажаний ступінь невідповідності приводить до Тm менше ніж 45°С (розчин у воді) або 32°С (розчин у формаміді), переважним є збільшення концентрації SSC, щоб можна було використовувати більш високу температуру. Обширний посібник з гібридизації нуклеїнових кислот представлений в Tijssen, 1993; Ausubel et al., 1998. В іншому аспекті винахід належить до олігонуклеотидних зондів для детекції РНК або кДНК ToTV. Зонди для детекції тут вибирають як «в основному» . комплементарні одноланцюжковій молекулі РНК, або одному з ланцюгів дволанцюжкових нуклеїнових кислот, отриманих за допомогою реакції ампліфікації за винаходом. Переважно, зонди є в основному комплементарними, не обов'язково, іммобілізованим (наприклад, міченим біотином) антисмисловим ланцюгам ампліконів, отриманих з мішені РНК або ДНК. Для зондів для детекції за даним винаходом допустимо містити одну або декілька невідповідностей їхньої послідовності-мішені. Як правило, 39 послідовності, що володіють щонайменше 65%, більш переважно щонайменше 80% гомологією з олігонуклеотидними послідовностями-мішенями, вважають прийнятними для використання в способі за даним винаходом. Стійкі до ТоТУ рослини Крім того, винахід належить до способу ідентифікації стійкої до ToTV рослини, або її частини. Існують різні можливості ідентифікувати стійкі до ToTV рослини. У першому наборі варіантів здійснення такого способу можна застосовувати активний/інфекційний вірус або повнорозмірні інфекційні клони, тоді як в альтернативному варіанті здійснення використовують тільки засоби для детекції вірусу. Перша стадія способу ідентифікації стійкої до ToTV рослини з використанням активного/інфекційного вірусу містить у собі вплив на рослину або частину рослини, таку як фрагмент листка або стебла, інфекційної дози ToTV, метою чого є досягти інфекції. Вплив у багатьох випадках містить у собі встановлення фізичного контакту. Інфекційна доза може розрізнятися між рослинами й між тестованими ізолятами ToTV. Теоретично, кількість від приблизно 1-10 до приблизно 5005000 вірусних частинок зазначеного вірусу або його нуклеїнових кислот буде достатньою. Інфекції, таким чином, можна досягати механічною інокуляцією очищених частинок вірусу або нуклеїнової кислоти вірусу в здорові рослини. Альтернативно, інфекції можна досягати, наприклад: - вирощуванням здорового черешка на зараженому ToTV кореневищі, або навпаки; - впливом на здорову рослину переносників, що містять вірус (включаючи інфіковані рослини, наприклад, рослини-паразити, подібні до видів Cuscuta); - введенням у здорову рослину експресуючого вектора, що несе кодуючи область геному вірусу ToTV; - застосуванням агроінфекційних клонів, таких як штами Agrobacterium tumefaciens, що містять експресуючий вектор, який несе кодуючу область геному вірусу ToTV. В контексті даного винаходу способи впливу на рослину або частину рослини інфекційної дози ToTV не є обмеженими яким-небудь конкретним способом. Як зазначено, інфекція може містити в собі механічну інокуляцію вірусу в здорові рослини. Наприклад, частину ураженого захворюванням листка можна втерти безпосередньо в листок підлягаючої інфікуванню рослини. За альтернативним способом інокулят можна одержати, наприклад, перемелюванням тканини рослини, яка містить вірус, переважно, молодого листя з вираженими симптомами, за допомогою ступки й товкачика, або будь-якого іншого типу гомогенізатора, наприклад, у буфері, придатному для інокуляції (наприклад 0,03Μ фосфатний буфер, рН7,7). Після перемелювання отриманий гомогенат (сік) переважно фільтрують, наприклад, через сувору марлю. Потім сік можна інокулювати, наприклад, шляхом обережного приведення листя у контакт із 94035 40 якою-небудь кількістю соку. Листки переважно попередньо обробляють, щоб зруйнувати нижній епідерміс і підсилити проникнення вірусу. Цього можна досягати, наприклад, попереднім припудрюванням листя порошком карбіду кремнію. Переважно уникають надмірного ушкодження. Переважно використовують порошок карбіду кремнію, що володіє мікроскопічно малими частинками карбіду кремнію кутастої форми (400-500меш). Порошок карбіду кремнію можна також додавати безпосередньо в сік, у цьому випадку попередню обробку виключають. Сік можна наносити, наприклад, вказівним пальцем, тампоном із просоченого соком пінопласту або тканини, або навіть товкачиком, використовуваною для перемелювання, скляним шпателем, твердим пензлем або пульверизатором. Після інокуляції листки переважно негайно промивають водою. Друга стадія способу ідентифікації стійкої до ToTV рослини містить у собі ідентифікацію зазначеної рослини як стійкої до ToTV рослини, коли, після зазначеного впливу, або і) симптоми захворювання в зазначеній рослині або частині рослини залишаються відсутніми, або їхній прояв затриманий, або щонайменше вага зменшена, або вони є локалізованими в порівнянні зі сприйнятливим і/або чутливою контрольною рослиною, і/або іі) вірус ToTV або геномні послідовності ToTV не присутні у зазначеній рослині або частині рослини, або присутність вірусу ToTV є щонайменше зменшеним кількісно в порівнянні із чутливою контрольною рослиною. Як використовується в даному описі, термін локалізований означає обмежений інокульованим листком. Визначення розвитку симптомів індукованого ToTV захворювання в інфікованих рослинах можна здійснювати кількісними способами, наприклад, де відзначають період, необхідний для розвитку явних (наприклад, видимих) симптомів захворювання, або кількісними способами, де після припинення певного періоду рослину перевіряють на прояв симптомів і відзначають присутність або тяжкість симптомів. На додаток до визначення розвитку індукованих ToTV-симптомів захворювання або як альтернатива йому, залежно від підлягаючого детекції типу стійкості до ToTV, виявляють присутність вірусу в рослині або частині рослини. Для детекції відсутності вірусу в тестованих рослинах, у принципі, можна використовувати будь-який спосіб. Наприклад, можна застосовувати спосіб, у якому використовують специфічне для ToTV антитіло, набір праймерів або зонд за даним винаходом. Альтернативно, частину тестованої рослини можна приводити в контакт із чутливою індикаторною рослиною (наприклад, N.hesperis '67A') для визначення, є присутнім або відсутній вірус у тестованій рослині. Фахівцеві в даній галузі зрозуміло, що для таких способів є важливим дезінфікувати поверхню тестованої рослини, щоб провести розходження між переносником, толерантною тестованою рослиною й стійкою тестованою рослиною, оскільки необхідно показати тільки присутність вірусу в клітинах рослини. 41 При проведенні другої стадії способу ідентифікації стійкої до ToTV рослини можна одержати наступні результати. Якщо після успішної інокуляції (наприклад, після здійснення контакту рослини з вірусом в умовах, що приводять до інфекції в сприйнятливій і чутливій контрольній рослині): і) симптоми захворювання залишаються відсутніми; або вірусні частинки, або вірусна РНК не піддаються виявленню: рослина є стійкою; іі) прояв симптомів захворювання затриманий, або їхня вага зменшена; або можна виявити низькі системні титри вірусних частинок або вірусної РНК: рослина є частково стійкою; ііі) симптоми захворювання є важкими, але залишаються локалізованими, обмеженими інокульованим листком, і не поширюються системно за межі інокульованої тканини; або вірусні частинки, або вірусну РНК можна детектувати тільки локалізовано: рослина є гіперчутливою; iv) якщо симптоми захворювання залишаються відсутніми; і можна детектувати вірусні частинки, або вірусну РНК: рослина є толерантною; ν) якщо в рослини розвиваються симптоми захворювання, і вона має високі системні титри вірусу, тоді рослина є сприйнятливою і чутливою. Прикладами таких рослин є рослини, з яких виділений вірус за даним винаходом. Ці рослини можуть служити прийнятними контрольними рослинами в способах за даним винаходом. Для цілей одержання стійких рослин і з погляду фітосанітарії, тільки положення і), іі) і ііі) можна вважати такими, що представляють інтерес. Для цілей одержання рослин, придатних для одержання безсимптомного врожаю й продуктів, результат iv) може представляти також конкретний комерційний інтерес. В альтернативному варіанті здійснення способу для ідентифікації стійкої до ToTV рослини використовують тільки засоби для детекції вірусу. Наприклад, стійку до ToTV рослина можна ідентифікувати в польових умовах за допомогою виявлення або ідентифікації безсимптомної рослини серед рослин, що володіють симптомами, і визначення відсутності вірусу в зазначеній рослині кожним зі способів детекції вірусу за даним винаходом. Фактично, це відповідає способу ідентифікації стійкої до ToTV рослини, де стадію а) впливу на рослину або частину рослини інфекційної дози ToTV проводять пасивно (наприклад, природно). При здійсненні такого способу переважно використовувати спосіб детекції присутності ToTV у зразку за даним винаходом, де присутність вірусу ToTV або його компонента демонструють за допомогою реакції зазначеного зразка з полінуклеотидом або антитілом за даним винаходом. Переважно, для способу ідентифікації стійкої до ToTV рослини необхідні використання або вірусу за даним винаходом, або полінуклеотиду або антитіла за даним винаходом. Крім того, винахід належить до способу одержання стійкої до ToTV рослини, або її частини. Після ідентифікації стійкої до ToTV рослини ця рослина може служити донорною рослиною для генетичного матеріалу, яку можна переносити від зазначеної донорної рослини до реципієнтної рос 94035 42 лини, щоб забезпечити зазначену реципієнтну рослина генетичним матеріалом. Перенесення генетичного матеріалу від донорної рослини до реципієнтної рослини можна здійснювати будьяким прийнятним способом, відомим у даній галузі. Генетичний матеріал у більшості випадків буде геномним матеріалом. Однак є важливим переносити щонайменше одну, яка надає стійкості частину геному донорної рослини. За відсутності способів визначення, які частини геному донорної рослини надають стійкості до ToTV, перенесення можна здійснювати, відповідно, за допомогою перенесення цілих хромосом. Переважно, стійка до ToTV рослина служить чоловічим або жіночим батьком-матір'ю у схрещуванні для одержання стійких рослин-нащадків, таким чином, рослинанащадок одержує геномний матеріал від стійкого донора й діє як реципієнтна рослина. Хоча чутливий батько при схрещуванні не обов'язково є реципієнтною рослиною в буквальному значенні, такий чутливий батько тут також включений у термін реципієнтна рослина. У способі одержання стійкої до ToTV рослини можна також використовувати злиття протопластів для перенесення геномного матеріалу, що надає стійкості, від донорної рослини до реципієнтної рослини, тобто як спосіб схрещування зазначених рослин. Злиття протопластів є індукованим або спонтанним об'єднанням, таким як соматична гібридизація, між двома або більше протопластами (клітинами, клітинні стінки яких видалені ферментативною обробкою) для одержання однієї бі- або багатоядерної клітини. Злита клітина, яку можна одержати навіть із видів рослин, які не можуть схрещуватися в природі, являє собою тканину, яка культивується до гібридної рослини, що володіє бажаним сполученням ознак. Більш конкретно, перший протопласт можна одержати з рослини томату або іншої лінії рослин, що володіє стійкістю до інфекції ToTV. Наприклад, можна використовувати протопласт зі стійкої до ToTV лінії (томату, баклажана, перцю, дині, кавуна або огірка). Другий протопласт можна одержати із другої, чутливої, лінії рослин, не обов'язково, з інших видів рослин або варієтету, переважно, з тих же самих видів рослин або варієтету, що володіють комерційно бажаними характеристиками, такими як, без обмеження, стійкість до захворювання, стійкість до комах, різні характеристики плодів і т.д. Потім протопласти зливають із використанням загальноприйнятих способів злиття протопластів, відомих у даній галузі для одержання схрещування. Альтернативно, можна використовувати порятунок ембріона для перенесення геномного матеріалу, що надає стійкості, від донорної рослини реципієнтній рослині, тобто як спосіб схрещування зазначених рослин. Порятунок ембріона можна використовувати як спосіб виділення ембріонів з нащадків схрещування, коли рослини нездатні виробляти життєздатне насіння. За даним способом запліднену зав'язь або незріле насіння рослини культивують для одержання нових рослин (цей спосіб докладно описаний в Pierik, 1999). Таким чином, спосіб одержання стійкої до ToTV рослини в одному з варіантів здійснення 43 містить у собі стадії ідентифікації стійкої до ToTV донорної рослини, як описано тут вище, і схрещування зазначеної стійкої до ToTV донорної рослини з реципієнтною рослиною, як описано вище, таким чином, одержуючи рослини-нащадки. Спосіб одержання стійкої до ToTV рослини додатково містить у собі стадію відбору з рослиннащадків стійкої рослини способом ідентифікації стійкої до ToTV рослини, як описано раніше. Переважно, зазначена стійка до ToTV донорна рослина являє собою рослину сімейства Solanaceae або Cucurbitaceae, навіть більш переважно, томат, баклажан, перець, диню, кавун або огірок. Переважно, зазначена реципієнтна рослина являє собою рослину сімейства Solanaceae або Cucurbitaceae, навіть більш переважно томат, баклажан, перець, диню, кавун або огірок. Ще більш переважно, зазначена реципієнтна рослина являє собою томат видів Solarium lycopersicum, більш переважно, рослина S.lycopersicum, що володіє комерційно бажаними характеристиками. Реципієнтна рослина може бути сприйнятливою до ToTV рослиною, чутливою до ToTV рослиною або стійкою до ToTV реципієнтною рослиною. Як пояснюється вище, вибір рослини буде в першу чергу визначатися тим, чи є ознака стійкості домінантною або рецесивною. Фахівець у даній галузі буде усвідомлювати, що різні способи доступні для рішення таких завдань. Також аспектом за даним винаходом є стійка до ToTV рослина, або її частина, які можна одержати способом за винаходом. Як зазначено, переважний варіант здійснення способу для одержання стійкої до ToTV рослини містить у собі перенесення за допомогою інтрогресії зазначеної послідовності нуклеїнової кислоти, що надає стійкості, від стійкої до ToTV донорної рослини до реципієнтної рослини шляхом схрещування зазначених рослин. У стійких рослинах, отриманих за даним переважним варіантом здійснення, більшість ознак успішно можуть походити з реципієнтної рослини, а стійкість до ToTV - походити з донорної рослини. В одному зі способів, що позначається як племінне розведення, донорну рослину, що володіє стійкістю до ToTV, схрещують із реципієнтною рослиною, що переважно володіє комерційно бажаними характеристиками, такими як, без обмеження, стійкість до захворювання, стійкість до комах, корисні характеристики плодів, і т.д. Отриману популяцію рослин (які являють собою гібриди F1) потім піддають самозапиленню й забезпечують можливість утворення насіння (насіння F2). Потім рослини F2, вирощені з насіння F2, скрикують по стійкості до ToTV. Проводити скринінг популяції можна рядом різних способів, переважно, способом візуального контролю за даним винаходом. Оскільки ідентифікація стійких до ToTV рослин спочатку стала можливою тільки за даним винаходом, спосіб одержання стійкої рослини є аспектом винаходу. Також аспектом даного винаходу є стійка до ToTV рослина, або її частина, яку можна одержати способом за винаходом. 94035 44 Даний винахід належить до способів запобігання поширення інфекції ToTV у рослинах томату шляхом одержання стійких рослин томату, так само як шляхом знищення рослин, що несуть вірус ToTV. Дані міри можуть становити частину загальної стратегії поліпшення фітосанітарної профілактики стосовно вірусу ToTV. Таким чином, можна ідентифікувати й знищувати толерантні рослини, щоб знищити такі джерела вірусу ToTV. В одному з варіантів здійснення способу одержання стійкої до ToTV рослини, або її частини, даний винахід належить до способу одержання толерантної до ToTV рослини. З толерантної рослини можна одержувати повноцінний урожай, плоди й насіння, оскільки, хоча рослина може бути носієм вірусу, для неї не виявляють симптомів захворювання. Такий спосіб буде містити в собі ідентифікацію толерантних рослин і використання таких толерантних рослин як джерел або донорів бажаного генетичного матеріалу. Метою є не одержання рослини, здатної опиратися проникненню або розмноженню вірусу в клітинах, а одержання рослини, що не страждає від симптомів. Таким чином, даний винахід належить до способу ідентифікації стійкої до ToTV рослини, що включає стадії а) впливу на рослину або частину рослини інфекційної дози ToTV, і b) ідентифікації зазначеної рослини як толерантної до ToTV рослини, коли, після зазначеного впливу, симптоми захворювання в зазначеній рослині або частині рослини залишаються відсутніми, a ToTV є присутнім у зазначеній рослині або частині рослини. Визначення розвитку симптомів індукованого ToTV захворювання в інфікованих рослинах можна здійснювати кількісними способами, наприклад, де відзначають період, необхідний для розвитку явних (наприклад, видимих) симптомів захворювання, або кількісними способами, де після припинення певного періоду рослину перевіряють на відсутність прояву симптомів або відзначають зменшення тяжкості симптомів. У переважному варіанті здійснення присутність ToTV у зазначеній рослині або частині рослини визначають на стадії b) способом, що включає визначення в зазначеній рослині або частині рослини присутності вірусу ToTV або його компонентів за допомогою реакції зазначеної рослини або частини рослини з полінуклеотидом за винаходом або антитілом за винаходом. Діагностичні набори Способи й засоби, надані в даному описі, є, зокрема, застосовними в діагностичному наборі для діагностики інфекції вірусу ToTV за допомогою вірусологічного діагнозу. Такі набори або аналізи можуть, наприклад, містити вірус, нуклеїнову кислоту, білкову молекулу або її фрагмент, і/або антитіло за винаходом. Винахід належить також до діагностичного набору для діагностики інфекції ToTV, що містить вірус ToTV, специфічну для вірусу ToTV нуклеїнову кислоту, білкову молекулу або її фрагмент і/або антитіло за винаходом, і переважно, засобу для детекції зазначеного вірусу ToTV, специфічної для вірусу ToTV нуклеїнової кислоти, білкової молекули або її фрагмента, і/або антитіла, де зазначені 45 засоби, наприклад, містять у собі збуджувану групу, таку як флуорофор, або ферментативну систему детекції, застосовувану в даній галузі (приклади прийнятних форматів діагностичних наборів включають IF, ELISA, аналіз нейтралізації, аналіз RT-PCR, аналізи гібридизації). Прийнятні аналізи для детекції включають прямі й непрямі аналізи, сендвіч-аналізи, твердофазні аналізи, такі як, серед інших, аналізи з використанням планшетів або гранул, і аналізи в рідкій фазі. Прийнятні аналізи включають аналізи з використанням первинних і вторинних антитіл і аналізи з використанням єднальне антитіло реагентів, таких як білок А. Більш того, за винаходом можна використовувати велику кількість способів детекції, включаючи колориметричний, флуоресцентний, фосфоресцентний, хемілюмінесцентний, люмінесцентний і радіоактивний способи. Щоб визначити, чи можна компонент ще не ідентифікованого вірусу або його синтетичний аналог, такий як нуклеїнова кислота, білкова молекула або її фрагмент, ідентифікувати як специфічний для вірусу ToTV, достатньо аналізувати послідовність нуклеїнової кислоти або амінокислоти зазначеного компонента, наприклад, фрагмент зазначеної нуклеїнової кислоти або амінокислоти, переважно щонайменше 10, більш переважно щонайменше 25, більш переважно щонайменше 40 нуклеотидів або амінокислот (відповідно), шляхом порівняння послідовності з наданими послідовностями вірусу ToTV і з відомими послідовностями, що не належить до вірусу ToTV (переважно використовують найближчі філогенетичні родичі ToTV), з використанням, наприклад, філогенетичних аналізів, як передбачено в даному описі. Залежно від ступеня споріднення із зазначеними вірусними послідовностями ToTV або не стосовними ToTV, можна ідентифікувати компонент або синтетичний аналог. Набір для детекції вірусу ToTV може, залежно від формату аналізу, містити одне або декілька антитіл, специфічних для білка, переважно специфічних щонайменше для одного білка капсиду ToTV, і переважно також містити в основному очищений білок ToTV або антиідіотипове антитіло для використання як позитивного контролю. Противірусні засоби Винахід належить також до способів одержання противірусного засобу, застосовного для лікування інфекції ToTV у рослин, що включають одержання культури клітин або експериментальної рослини, що містить вірус за винаходом, обробку зазначеної культури або рослини потенційним противірусним засобом і визначення ефекту зазначеного засобу на зазначений вірус або його інфекцію зазначеної культури або рослини. Приклади такого противірусного засобу містять у собі нейтралізуюче ToTV антитіло, або його функціональний компонент, як передбачено в даному описі, однак можна одержувати також противірусні засоби іншої природи. Існують різні противірусні засоби, застосовні для рослин, такі як хімічні продукти, бактерії, гриби, комахи й віруси. Більшість із них пов'язані із системною набутою стійкістю (SAR). Даний вина 94035 46 хід належить до застосування геному ToTV або його частини як індуктора системної набутої стійкості у рослини. Системна набута стійкість може бути спрямованою проти ToTV або проти інших захворювань. За даним аспектом ToTV, його геном або його частини, що надають стійкості, можна застосовувати як противірусний агент. Винахід належить також до застосування противірусного засобу за винаходом для одержання композиції для лікування, зокрема, для лікування інфекції ToTV у рослин, і належить до фармацевтичної композиції, що містить противірусний засіб за винаходом, застосовної в способі лікування або попередження інфекції вірусом ToTV, де зазначений спосіб включає введення такої композиції для лікування в окрему рослину. Винахід також належить до рослинної моделі, застосовної для тестування способів і/або композицій для лікування. Очевидно, деякі види Nicotiana можна інфікувати вірусом ToTV, завдяки чому спостерігати симптоми захворювання, що відрізняються від симптомів, виявлених у рослин томату, які страждають від вірусу ToTV. Піддаючи рослини Nicotiana противірусному лікуванню або до, або під час інфекції вірусом можна мати передбачуване значення застосування такого противірусного засобу для рослин томату. Винахід також належить до застосування ToTV, або частин геному вірусу ToTV, як експресуючого вектора, наприклад, для застосування для індукованого вірусом мовчання генів (VIGS). VTGS являє собою технологію, що використовує РНК-опосередкований механізм противірусного захисту в рослин. У рослин, інфікованих немодифікованими вірусами, механізм специфічно спрямований проти вірусного геному. З використанням вірусних експресуючих векторів, що несуть вставки, отримані з генів хазяїв, механізм можна використовувати також для атаки проти відповідних РНК рослин. VIGS широко використовували в рослинах для аналізу функції генів і адаптували для високопродуктивної функціональної геноміки. Дотепер більшість застосувань VIGS належало до Nicotiana benthamiana. Однак даний винахід належить до застосування ToTV як нових експресуючих векторних систем, що дозволяють аналізувати функцію генів в інших рослинах, таких як томат або інші види сімейства Solanaceae, такі як перець і картопля, і у видах сімейства Cucurbitaceae. Винахід далі пояснюється в прикладах без обмеження ними. Приклади Приклад 1. Виділення і характеризація ToTV з рослин томату Методи Введення Зразки рослин томату з Іспанії одержували для діагностичних досліджень. Симптоми в рослин томату проявлялися у вигляді некротичних плям і хлорозу на листках і коричневих кільцях на плодах. Серологічні тести (ELISA) указували на присутність вірусу мозаїки пепіно (PepMV), однак, беручи до уваги симптоми, очевидно, був присутній інший, ще не ідентифікований агент. 47 Електронно-мікроскопічними дослідженнями тканини інфікованого листка були виявлені сферичні вірусні частинки. Потім проводили тест на інфекцію й виявили, що сприйнятливими до ToTV були багато зразків томату, деякі з яких реагували з явними симптомами (некроз листя, що починається в основі окремих листочків (див. Фіг.1). Перенесення і розмноження вірусу ToTV виділяли, як описано нижче, з уражених хворобою рослин, отриманих з Іспанії. ToTV можна було механічно перенести до декількох видів Nicotiana. Показано, що загальноприйнятий буфер для інокуляції є придатним (наприклад, 0,03Μ фосфатний буфер, рН7,7). Скрізь у даному прикладі ToTV механічно інокулювали в N.glutinosa або N.benthamiana і розмножували в них. Очищення вірусу проводили приблизно через 14 днів після інокуляції. Очищення вірусу Виконували декілька спроб очищення ToTV відповідно до загальноприйнятих способів, наприклад, для неповірусів або лютеовірусів (за допомогою органічних розчинників). Ці способи завжди приводили до втрати інфекційності вірусу. Більш того, ToTV виявляв тенденцію до агрегації, коли на стадіях центрифугування використовували низькі температури (нижче 5°С). В остаточному підсумку, дуже м'яким способом очищення в результаті одержували прийнятно чисті препарати вірусу, на яких можна було проводити подальші експерименти. Для очищення ToTV використовували наступний спосіб (всі стадії центрифугування проводили при 6°С). Інфіковане листя N.glutinosa або N.benthamiana гомогенізували в 5 частинах 0,1Μ Тріс-НСІ (рН8) плюс 20мМ Na2SO3, 10мМ NaDIECA і 5мМ Na-EDTA (буфер для гомогенізації) і гомогенат центрифугували протягом 30 хвилин при 49000×g. Супернатант вміщували на подушку 20% сахарози й центрифугували протягом 1,5 годин при 70000×g. Осад ресуспендували в 2мл Тріс-НСІ, рН8, суспензію наносили на градієнт сахарози (10-40% сахарози в буфері для гомогенізації) і центрифугували протягом 2 годин при 110000×g. Оскільки смуга з вірусом не була видимою, градієнт аліквотували на окремі фракції й присутність вірусу в кожній фракції визначали шляхом інокуляції частини зазначеної фракції в листки N.hesperis '67A', як описано в даному документі, і спостереження наявності інфекції. Фракцію, яка містить вірус, вміщували в 10-40% градієнт сульфату цезію (у Тріс-НСІ, рН8) і центрифугували протягом 16 годин при 125000×g. Смуги з вірусом збирали й концентрували центрифугуванням або діалізували проти 0,1Μ Тріс-НСІ, рН8. Інфекційність ToTV після кожної стадії очищення перевіряли інокуляцією в N.hesperis '67А' (перед інокуляцією фракції градієнта сульфату цезію діалізували проти Тріс-НСІ, рН8). Електронна мікроскопія Суспензії вірусу «монтували» на сітці, покритій Formvar®, відомим інакше як полівінілформаль, 94035 48 забарвлювали 2% уранілацетатом і досліджували на електронному мікроскопі Philips CM12. Аналіз PAGE Вірусні білки розділяли електрофорезом в 12% денатуруючому поліакриламідному гелі ×SDS-PAGE, Laemmli, 1970) і забарвлювали сріблом. Виділення й оцінка нуклеїнової кислоти Очищений вірус концентрували центрифугуванням (2 години при 115000g). Опади піддавали екстракції РНК за способом очищення Qiagen RNeasy MinElute (Qiagen, Hilden, Germany). Концентрацію РНК визначали на УФ-спектрофотометрі (Beckman Coulter, Inc., Fullerton, USA). Цілісність РНК перевіряли електрофорезом в агарозному гелі. Після електрофорезу протягом 2 годин при 60 В РНК забарвлювали з використанням ортотолуїдину синього. Синтез і клонування кДНК кДНК синтезували з використанням системи для синтезу кДНК Invitrogen Superscript Choice (Invitrogen, Breda, The Netherlands) відповідно до інструкцій виробника. Перший ланцюг кДНК праймували з використанням або oліго-d(T), або випадкових гексамерних праймерів. Після синтезу другого ланцюга лігували адаптери для EcoRI для полегшення клонування. Після фосфорилювання кДНК із адаптерами для EcoRI, неліговані лінкери видаляли колонковою хроматографією. Отриману кДНК лігували в попередньо розщеплений EcoRI експресуючий вектор pBluescript II (Stratagene, La Jolla, USA) і лігатом трансформували компетентні клітини Тор 10 (Invitrogen). 5'-Кінець послідовності ToTV визначали з використанням системи швидкої ампліфікація кінців кДНК 5'RACE (LifeTechnologies) з використанням dCTP відповідно до інструкцій виробника. Аналіз кДНК Після трансформації в рекомбінантних клонах аналізували присутність вставок за допомогою PCR з використанням праймерів, специфічних як для Т3, так і для Т7. Розмір продуктів PCR аналізували в 1% агарозному гелі. Клони, що містять вставки приблизно по 1500 нуклеотидів, використовували для подальшого аналізу послідовності. Отримані дані про послідовності аналізували з використанням пакета програм DNASTAR. Визначення амінокислотної послідовності трьох білків капсиду ToTV Очищені частинки ToTV наносили на денатуруючий гель для PAGE і розділяли білки капсиду. Смуги з розділеними білками капсиду вирізали з гелю, обробляли трипсином і гідролізати аналізували з використанням тандемного масспектрометра (MSMS), по суті з використанням способу, описаного Kinter & Sherman, 2000. У результаті цього одержували амінокислотні (ак) послідовності малих пептидів, кожна з яких проявляла гомологію з амінокислотною послідовністю, прогнозованою з нуклеотидної послідовності RNA2 ToTV. Порівняння з іншими даними про вірусні послідовності виконували за допомогою програм з пакета PHYLIP. Результати Перенесення і розмноження вірусу 49 Для розмноження ToTV можна використовувати N.glutinosa і N.benthamina. Види тютюну N.hesperis '67A' і N.occidentalis 'Ρ1' є дуже сприйнятливими до ToTV, і проявляють симптоми після 3-4 доби. У цих видах тютюну розвивається сильний некроз за короткий час, і, таким чином, їх вважають більш прийнятними як індикаторна рослина, ніж хазяїн для розмноження. N.glutinosa і N.benthamiana реагують із місцевими вогнищами хлорозу й системним хлорозом, а також з помірною деформацією листя. Очищення вірусу Очищення віріонів з інфікованої тканини листка виявилися досить складною. ToTV є нестійким до органічних розчинників і виявляє тенденцію до агрегації при центрифугуванні при низьких температурах. Використовуваним протоколом очищення (див. 1.2.) одержали два видимі смуги, які містять віруси, в градієнті сульфату цезію. Смуги з'являлися в нижній частині градієнта, що вказує на досить високу плавучу густину в Cs2SO4, яка дорівнює або перевищує 1,4г/см2. Сульфат цезію впливав на інфекційність ToTV, однак вона не втрачалася повністю при високій концентрації вірусу у вихідному матеріалі. Фракції градієнта сульфату цезію, що містять обидві смуги з вірусом, були інфекційними. Інфекційність окремих смуг не визначали. Електронна мікроскопія Дві смуги аналізували електронною мікроскопією, і вони містили частинки вірусу діаметром приблизно 28нм (Фіг.2). Аналіз PAGE Електрофорезом у поліакриламідному гелі (PAGE) очищених фракцій вірусу з наступним фарбуванням гелю сріблом виявили три білки капсиду (СР) приблизно 23, 26 і 35кДа (див. Фіг.3, де показані очищені верхня (TAgV-T) і нижня фракції (TAgV-B) очищеного вірусу). Виділення нуклеїнової кислоти й аналіз кДНК Виділенням РНК із обох смуг з вірусом разом одержали дві РНК: приблизно 5,5т.н. і 8т.н. (див. Фіг.4). Виділенням РНК із верхньої смуги одержували тільки фрагмент РНК 5,5т.н. РНК 5,5т.н. з верхньої смуги використовували як матрицю для синтезу й клонування кДНК. Для різних клонів проводили реакції секвенування з використанням прямого й зворотного праймерів для секвенування (Т3/Т7 або M13F/M13R). Для визначення точного 5'-кінця РНК виконували швидку ампліфікацію кінців кДНК (RACE). У результаті аналізу отриманих даних про послідовності з використанням пакета програм Lasergene® (DNASTAR, Inc., Madison, WI, USA), одержали повну послідовність РНК2 вірусу (SEQ ID NO:1; див. Фіг.5). Розмір РНК становить 5389 нуклеотидів, крім полі Α-хвоста. У РНК виявили дві відкриті ра 94035 50 мки зчитування (ORF). ORF1 локалізована від нуклеотиду 182 до 742, має довжину 561 нуклеотидів і кодує білок з 187 амінокислот. Для даної послідовності не виявлено гомології в базах даних NCBI на білковому й нуклеотидному рівні. ORF2 простягається від нуклеотиду 702 до 4298, має довжину 3597 нуклеотидів і кодує білок з 1199 амінокислот. Після пошуку BLAST у базі даних NCBI виявили невелику гомологію з декількома вірусними полібілками. Для виявлених гомологів інвентарні номери з бази даних NCBI наведені разом з назвою вірусу й типом білка. Як послідовності нуклеїнової кислоти, так і похідні амінокислотні послідовності у всіх трьох рамках зчитування використовували для аналізу BLAST. Для ідентифікації ORF використовували MAPDRAW з пакета програм Lasergene®. Карти ORF на обох РНК: РНК1 РНК1 містить одну ORF (ORF1) з мотивами, типовими для гелікази, РНК-залежної РНКполімерази (RdRp). Крім цього, для амінокислот у положеннях 106-338 виявили низький рівень гомології амінокислотної (ак) послідовності з кофактором протеази (Pro-Co) вірусу слабкої мозаїки пачулі (PatMMV) (NP647592.1), з ідентичністю 22%. Типові мотиви гелікази A (GKS), В (D), С (Ν) ідентифіковані в положеннях 398-400, 444 і 495 можливого поліпептиду. Для області гелікази найбільш близьку ідентичність виявили зі сферичним вірусом рису тунгро (RTSV; 42% ідентичності в 140ак, що перекриваються), вірусом карликового хлорозу кукурудзи (MCDV; 43% ідентичності в 137ак, що перекриваються), вірусом крапчастості суниці (SMoV; 42% ідентичності в 135ак, що перекриваються) і вірус жовтої плямистості пастернаку (PYFV; 42% у 138ак, що перекриваються). У можливій області Vp не виявлено подібності послідовності з іншими вірусами. Найбільша подібність по протеазі виявлена для ак 1000-1100, де виявлено 25% ідентичності з вірусом картоплі V (PW; у 86ак протеази NIa, що перекриваються). Область RdRp між мотивами І (KDE)-VII (FLSR) виявили між ак 1303-1554 (Koonin 1991). Найбільш близьку ідентичність послідовності полібілка виявили з: сферичним вірусом рису тунгро (RTSV) з 29% ідентичності в 751ак, що перекриваються, вірусом карликового хлорозу кукурудзи (MCDV) з 28% ідентичності в 742ак, що перекриваються, вірусом жовтої плямистості пастернаку (PYFV) з 33% ідентичності в 501ак, сферичним латентним вірусом яблука (ALSV) з 32% ідентичності в 472ак, вірусом крапчастості суниці (SMoV) з 30% ідентичності в 680ак і вірусом рашпілевидності листя вишні (CRLV) з 33% в 465ак. 51 94035 52 Таблиця 2 Загальний рівень гомології (в %) між мотивами RdRp в ORF1 в RNA1 ToTV і мотивами RdRp з інших вірусів рослин NIMV PYFV RTSV SDV SMoV ToTV ALSV CRLV MCDV NIMV 100 PYFV 32,7 100 RTSV 33,9 43,7 100 SDV 88,6 33,2 34,3 100 SMoV 49,4 32,0 32,7 50,2 100 ToTV 33,1 35,2 35,1 33,1 35,7 100 ALSV 39,6 36,4 37,5 38,4 40,6 38,1 100 CRLV 39,6 36,0 35,1 38,4 38,5 38,1 79,5 100 MCD V 35,1 42,9 69,4 36,7 38,5 33,7 35,5 35,1 100 NIMV=вipyc інфекційної крапчастості апельсина Навель (Sadwa); PYFV=вipyc жовтої плямистості пастернаку (Sequivirus); RTSV=сверичний вірус рису тунгро (Waikavirus); SDV=вipyc карликовості мандарина уншіу (Sadwavirus); SMoV=вipyc крапчастості суниці (Sadwavirus); ToTV - вірус томату торрадо (приблизно, рід); ALSV=сферичний латентний вірус яблуні (Cheravirus); CRLV=вipyc рашпілевидності листя вишні (Cheravirus); MCDV=вipyc карликового хлорозу кукурудзи (Waikavirus) Таблиця 3 Загальний рівень гомології (в %) між мотивами гелікази в ORF1 в RNA1 ToTV з мотивами гелікази з інших вірусів рослин SDV SMoV ToTV ALSV CRLV MCDV PYFV RTSV SDV 100 SMoV 42,5 100 ToTV 37,2 42,5 100 ALSV 31,9 31,0 36,0 100 CRLV 33,6 31,9 34,2 86,7 100 MCDV 39,8 32,7 46,5 34,5 34,5 100 PYFV 39,8 33,6 40,4 31,0 30,1 39,8 100 RTSV 38,1 32,7 44,7 30,1 31,0 69,0 42,5 100 PYFV=вipyc жовтої плямистості пастернаку (Sequivirus); RTSV=cферичний вірус рису тунгро (Waikavirus); SDV=вipyc карликовості мандарина уншіу (Sadwavirus); SMoV=вipyc крапчастості суниці (Sadwavirus); ToTV=вірус томату торрадо (приблизно, рід); ALSV=сферичний латентний вірус яблуні (Cheravirus); CRLV=вipyc рашпілевидності листя вишні (Cheravirus); MCDV=вipyc карликового хлорозу кукурудзи (Waikavirus) РНК2: РНК2 містить дві потенційних ORF (Фіг.7). ORF1 кодує можливий білок з 187ак з молекулярною масою 20кДа. Аналізом послідовності не виявили гомології з яким-небудь білком з баз даних EMBL. Невідомо, чи кодує дана ORF реально існуючий білок. Друга ORF, що частково перекривається з ORF1, починається із трьох стартових кодонів ATG у рамці. Вона кодує можливий білок з 1198ак з передвіщеною молекулярною масою 134кДа. Ідентифікацією білків за допомогою мас-спектрометрії виділених білків оболонки чітко картували три цистрони білків оболонки на С-кінці PHK2-ORF2. N-кінцева область PHK2-ORF2 для полібілка, найбільше імовірно, кодує можливий руховий білок (МР), оскільки в положенні ак 262-267 виявлений мотив LRVPML, дуже схожий на консенсусну послідовність можливого рухового білка LxxPxL (Mushegian, 1994). Інших гомологій послідовності для N-кінця ORF2 РНК2 не виявлено. ORF2 приблизно кодує чотири білки, які повинні відщеплюватися від полібілка-попередника за допомогою протеолітичного розщеплення. Однак неможливо ідентифікувати очевидну гомологію з ділянками розщеплення відомих полібілків. Залишається необхідність визначити точне положення цих ділянок розщеплення. Для послідовності полібілка СР1 автори даного винаходу виявили тільки деяку гомологію з пареховірусом людини (HPeV) з 21% ідентичністю в 103ак. Найбільш близьку ідентичність із послідовністю полібілка СР2 виявили для вірусу звичайної черемхової попелиці (RhPV) з 25% ідентичністю в 53 94035 168ак, вірусу енцефаломієліту птахів (AEV) з 33% в 74ак, вірусу чорного маточника (BQCV) з 43% ідентичністю в 37ак і вірусу червоної вогненної мурахи (SINV-1) з 30% ідентичністю в 51ак. Не виявлено гомологій з послідовністю полібілка СР3. Нетрансльовані області (UTR) 3'-UTR РНК1 і РНК2 становлять 1210н. і 1092н., відповідно. Існує 98% ідентичність кінцевих 988 нуклеотидів обох РНК. Для підтвердження того, що 3'-області UTR обох РНК є ідентичними, проводили RT-PCR тотальної вірусної РНК із одним зворотним праймером, виведеним з ідентичної області 3'-UTR і двох специфічних для кожної РНК прямих праймерів. Отримані продукти PCR секвенували. По цих результатах зробили наступний висновок: вірус, виділений з рослин томату й попередньо названий вірусом томату торрадо (ToTV), має сферичні (ікосаедричні) частинки діаметром приблизно 28нм. При очищенні вірус проявляє на дисплеї щонайменше дві видимі смуги в градієнті сульфату цезію. Обидві об'єднані смуги є інфекційними при інокуляції рослин тютюну. Час 54 тинки вірусу, очевидно, складаються щонайменше із трьох білків капсиду приблизно 23, 26 і 35кДа. Верхня фракція вірусу із градієнта сульфату цезію містить молекулу РНК приблизно 5,5т.н. (РНК2; SEQ ID NO:1), а нижня фракція - молекулу РНК приблизно 8т.н. (РНК1; SEQ ID NO:2). У результаті синтезу й клонування кДНК із використанням РНК 5,5т.н. з верхньої вірусної фракції й наступного аналізу інформації про послідовності декілька контигів зібрали в SEQ ID NO:1. Два контиги явно містили поло-А-хвіст, що дозволяло припускати, що вірусна РНК має поло-А-хвіст. Аналіз BLAST нуклеотидної і похідній амінокислотної послідовності не виявив якої-небудь істотної гомології з яким-небудь відомим вірусом з бази даних EMBL. Наведена вище інформація вказує, що ToTV є новим і дотепер не описаним вірусом. Отримана дотепер інформація ще не дозволяє групувати вірус із конкретним вірусним сімейством або родом. Для цілей детекції й ідентифікації вірусу сконструйовані два набори праймерів для RT-PCR (таблиця 4) на основі послідовності SEQ ID NO:1. Таблиця 4 Праймери для RT-PCR для детекції ToTV Набір праймерів А: прямий праймер зворотний праймер Довжина продукту Оптимальна темп, відпалу Набір праймерів В: прямий праймер зворотний праймер Довжина продукту Оптимальна темп.відпалу довжина/ темп 17-мір 17-мір 493п.н. 54,8°С 24-мір 22-мір 585п.н. 54,9°С Послідовність 5' -GAGAGCCGGC АТТС АСА-3' 5'-GCACAGCTTGGCGACAC-3' SEQ ID NО SEQ ID NO:3 SEQ ID NO:4 5'-СССАТСАТСАСССТССТСТТС GTA-3' SEQ ID NO:5 5'-TTCCAGTAATGATCCAACCA АТ-3' SEQ ID NO:6 Прийнятний спосіб RT-PCR містить у собі наступне. Виділяють і очищають загальну РНК із приблизно 100мкг матеріалу інфікованого листка з використанням набору для очищення РНК, наприклад, такого як Qiagen RNA-Easy. Із цієї загальної РНК кількість приблизно 1мкг використовують в 50мкл реакційної суміші для одноступінчастої реакції Superscript RT-PCR (Invitrogen), де реакційна суміш додатково містить 25мкл 2× реакційної суміші, 1мкл (100нг) як верхнього, так і нижнього праймера, 1мкл суміші RT/Taq і 22мкл води MilliQ. Щоб провести зворотну транскрипцію РНК у кДНК і ампліфікувати цю кДНК, можна використовувати наступну програму RT-PCR: стадія 1: 30 хвилин при 50°С (реакція зворотної транскрипції); стадія 2: 3 хвилини при 94°С (активація Taq-полімерази); стадія 3: 30 секунд при 94°С; стадія 4: 30 секунд при 55°С; стадія 5: 1 хвилина при 72°С; стадія 6: повтор стадій (3-5) 40×; стадія 7: 10 хвилин при 72°С; стадія 8: 10°С так довго, як буде потрібно. Продукти PCR можна аналізувати в 1% агарозноме гелі в буфері ТАЕ або ТВЕ. 2.6. Визначення амінокислотних послідовностей трьох білків капсиду ToTV Фрагменти найбільшого білка оболонки (СР1: приблизно 35кДа) можна вирівняти із частинами області ORF2 РНК2 (ак 487-729). Фрагменти білка оболонки із середньої смуги з білком (СР2: прибл. 26кДа) можна вирівняти з областю ORF2 РНК2 між ак 730 і 983. Фрагменти найменшого білка оболонки (СР3: прибл. 23кДа) можна вирівняти з С-кінцем 0RF2 РНК2 (ак 984-1195). Із цих результатів можна зробити висновок, що кодуючи послідовності трьох білків капсиду локалізовані в ORF2 РНК2 (5,5т.н.) ToTV, і отже, що виділені молекули вірусної РНК є частиною частинок вірусу ToTV. Приклад 2. Виділення й характеризація збудника захворювання Марчитець В 2003р. виявили рослини томату із симптомами, подібними до симптомів, викликаних ToTV, вирощені в Центральній Америці (Мексика й Гватемала) Припустили, що збудник захворювання f вірусним. Захворювання в межах певної місцевості відомо під назвами «Шоколад», «Марчитець (вірус)» або «Захворювання шоколадної плямистості». Сприйнятливі рослини, вирощені в польових умовах, ставали сильно інфікованими з 2003р. і пізніше. 55 94035 Метою даного дослідження було порівняння послідовності дотепер невідомого вірусу, що викликає «Захворювання шоколадної плямистості», і послідовністю ToTV, виділеного в прикладі 1. Методи РНК виділяли із приблизно 100мкг матеріалу листка, інфікованого «Захворюванням шоколадної плямистості» з використанням системи для виділення тотальної РНК (Promega SV 96). Два мкл тотальної РНК використовували в 50мкл реакційної суміші для одноступінчастої реакції Superscript RT-PCR (Invitrogen), де реакційна суміш додатково містить 25мкл 2 реакційної суміші, 1мкл (100нг) як прямого праймера, так і зворотного праймера, 1 мкл суміші RT/Taq і 22мкл води MilliQ. Щоб прове 56 сти зворотну транскрипцію РНК у кДПК і амшііфікувати цю кДНК, використовували наступну програму RT-PCR: стадія 1: ЗО хвилин при 50°С (реакція зворотної транскрипції); стадія 2: 3 хвилини при 94°С (активація laq-полімерази); стадія 3: 30 секунд при 94°С; стадія 4: 30 секунд при 55°С; стадія 5: 1 хвилина при 72°С; стадія 6: повтор стадій (3-5) 40; стадія 7: 10 хвилин при 72°С: стадія 8: 10°С так довго, як буде потрібно. Продукти PCR аналізували в 1% агарозному гелі в буфері ТАЯ або ТВЕ. Для RT-PCR використовували різні набори праймерів, відомих як відпалювані в різних положеннях РНК1 або РНК2 з геному ToTV. Таблиця 5 Праймери для RT-PCR на основі послідовності РНК-2 ToTV, використовувані для характеризації вірусузбудника захворювання шоколадної плямистості, як застосовували в прикладі 2 і як показано на Фіг.7 Набір праймерів Набір праймерів Р1048/1049: прямий праймер зворотний праймер Довжина продукту Набір праймерів Р1056/1057: прямий праймер зворотний праймер Довжина продукту Набір праймерів Р1060/1061: прямий праймер зворотний праймер Довжина продукту Набір праймерів Р1064/1065: прямий праймер зворотний праймер Довжина продукту Послідовність SEQ ID No 5'-CAAGCCATCACGGAACCTAC-3' 5'-AGCATCTTCTTCCTCCGCT-3' Від основи 36-544=508 основ SEQ ID NO:7 SEQ ID NO:8 5'-TGCTGAGGTGCTATC ACTGG-3' 5'-CACACTTTCCACGATTTCCA-3' Від основи 2422-2955=523 основи SEQ ID NO:9 SEQ ID NO:10 5'-AAAGGAAAGAAGCAGCC АСА-3' 5'-GGAAATCTTGGTCAAGCCAG-3' Від основи 3599-4170=571 основа SEQ ID NO:11 SEQ ID NO:12 5'-GCAATGCC AGTGGTTCAGAG-3' 5'-GGTCAAATGGATACTCGGGA-3' Від основи 4820-5327=507 основ SEQ ID NO:13 SEQ ID NO:14 Результати Як очікували, використані набори праймерів забезпечували ампліфікацію частини послідовності ToTV, однак при використанні для RT-PCR чотирьох різних наборів праймерів, основаних на послідовності РНК-2 (Р1048/І049, Р1056/1057, Р1060/1061 і Р1064/1065), одержали також ампліфікацію частини геному збудника «Захворювання шоколадної плямистості». Ділянки відпалу цих наборів праймерів проілюстровані на Фіг.1. Продукти RT-PCR для чотирьох наборів праймерів, з якими ампліфікували фрагменти збудника «Захворювання шоколадної плямистості», секвенували, і нони володіли 100% ідентичністю з відповідними частинами послідовності вірусного геному вірусу, описаного в прикладі 1. На Фіг.1 показана фотографія листя рослини томату із симптомами ToTV-захворювання. На Фіг.2 показана електронна мікрофотографія очищених частинок ToTV. Частинки мають діаметр приблизно 28нм. На Фіг.3 показаний результат фарбування сріблом гелю для PAGE з очищеними верхньою (TAgV-T) і нижньою (TAgV-B) фракціями вірусу ToTV, що показує три білки капсиду приблизно 23, 26 і 35кДа. На Фіг.4 показаний результат денатуруючого гель-електрофорезу в агарозі. Розміри фрагментів зазначені в тисячах нуклеотидів (т.н.). Гель забарвлювали з використанням ортотолуїдину синього. М: стандарт розміру молекул. TAgV: 1мкг виділеної РНК ToTV. На Фіг.5 показана повна послідовність молекули РНК2 ToTV (SEQ ID NO:1). На Фіг.6 показана повна послідовність молекули РНК1 ToTV (SEQ ID NO:2). На Фіг.7 показана загальна структура вірусу ToTV з відзначеними ділянками відпалу різних наборів праймерів, як використовували в прикладі 2 для РНК 2. Література Altschul, S.F., Gish, W., Miller, W., Myers, E.W., Lipman, D.J. (1990). Basic local alignment search tool. J. Моl. Biol., 215:403-410. Altschul, S.F., Madden, T.L., Schaffer, A.A., Zhang, J., Zhang, Z., Miller, W., Lipman, D.J. (1997). Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucl. Acids Res., 25, 3389-3402. 57 Ausubel, F.M., Brent, R., Kingston, R.E., Moore, D.D., Seidman, J.G., Smith, J.A., Struhl, K. eds. (1998) Current protocols in molecular biology. V.B. Chanda, series ed. New York: John Wiley & Sons. Barany, F. (1991) Genetic disease detection and DNA amplification using cloned thermostable ligase. Proc. Natl. Acad. Set USA 88: 189-193. Boom, R., Sol, C.J.A., Salimans, M.M.M., Jansen, C.L., Wertheim-van Dillen, P.M.E., Noordaa, van der J. (1990) Rapid and simple method for purification of nucleic acids. J. Clin. Microbiol. 28: 496-503. Chomczynski, P., Sacchi, N. (1987) Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal. Bio. 162:156-159. Clark, M.F., Adams A.N. (1977) Characteristics of the microplate method of enzyme-linked immunosorbent assay for the detection plant viruses. J. Gen. Virol. 34:475-483. Coligan, J.E., Kruisbeek, A.M., Margulies, D.H., Shevach, E.M. Strober, W. (Eds.) (1997) Current Protocols in Immunology. John Wiley & Sons Inc. Baltimore. Compton, J. (1991) Nucleic acid sequence-based amplification. Nature 1991, 350:91-92. Devereux, J., Haeberli, P., Smithies, O. (1984) A comprehensive set of sequence analysis programs for the VAX. Nucleic Acids Res. 12: 387-395. Dijkstra, J., de Jager, 3 (Eds.) (1998) Practical Plant Virology - Protocols and Exercises, Springer. Duffus, J.E., Liu, H.Y., Wisler, G.C. (1996) Tomato infectious chlorosis virus -a new clostero-like virus transmitted by Trialeurodes vaporariorum. European Journal of Plant Pathology, 102(3), 219226. Felsenstein, J. 1989. PHYLIP - Phylogeny Inference Package (Version 3.2). Cladistics 5: 164166. Gruatelli, J.C., Whitfield, KM., Kwoh, D.Y., Barringer, K. J., Richman, D.D., Gingeras, T.R. (1990) Isothermal, in vitro amplification of nucleic acids by a mutienzyme reaction modeled after retroviral replication. Proc. Natl. Acad. Sci. USA 87:1874-1878. Hagiwara, K, Ichiki, T.U., Ogawa, Y., Omura, Т., Tsuda, S. (2002). A single amino acid substitution in 126-kDa protein of Pepper mild mottle virus associates with symptom attenuation in pepper; the complete nucleotide sequence of an attenuated strain, C-1421. Arch Virol 147:833-340. Harlow, E. and Lane, D. (1988) Antibodies: A Laboratory Manual. Cold Spring Harbor Laboratory, Cold Spring Harbor, NY. Hirata, H., Lu, X., Yamaji, Y., Kagiwada, S., Ugaki, M. Namba, S. (2003) A single silent substitution in the genome of Apple stem grooving virus causes symptom attenuation. J Gen Virol 84:2579-2583. Jacobs M.V., Snijders P.J., van der Brule A.J. (1997) A general primer (GP5+/GP6+)-mediated PCR-enzyme immunoassay method for rapid detection of 14 high-risk and 8 low-risk human papillomavirus genotypes in cervical scrapings. J Clin Microbiol 35:791-795. 94035 58 Katz, E., Eksteen, R., Schoenmakers, P., Miller, N. (eds.) (1998) Handbook of HPLC. Marcel Dekker, New York. Kinter, M., Sherman, N.E. (2000) Protein Sequencing and Identification Using Tandem Mass Spectrometry. Wiley Interscience. Koonin, E.V. (1991) The phylogeny of RNAdependent RNA polymerases of positive-strand RNA viruses. J. Gen Virol 72:2197-2206. Kwoh, D.Y., Davis, G.R., Whitefield, K.M., Chappelle, H.L., DiMichele, L.J., Gingeras, T.R. (1989) Transcription-Based Amplification System and Detection of Amplified Human Immunodeficiency Virus Type 1 with a Bead-Based Sandwich Hybridization Format. Proc. Natl Acad. Sci. USA, 86, 1173-1177. Laemmli, U.K. (1970) Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227, 680-685. Lerner, R.A., Kang, A.S., Bain, J.D., Burton, D.R., Barbas, C.F. (1992) Antibodies without immunization. Science 258:1313-1314. Lizardi, P.M., Guerra, C.E., Lomeli, H., TussieLuna, I., Kramer, F.R. (1988) Exponential amplification of recombinant RNA hybridization probes. Biotechnology 6, 1197-1202. Lowman, H.B., Bass, S.H., Simpson, N., Wells, J.A. (1991) Selecting high-affinity binding proteins by monovalent phage display. Biochem. 30 (45): 108328. Lu, X., Hirata, H., Yamaji, Y., Ugaki, M., Namaba, S. (2001). Random mutagenesis in a plant viral genome using a DNA repair-deficient mutator Escherichia coli strain. J. Virol Methods 94:37-43. Marks, J.D., Hoogenboom, H.R., Griffiths, A.D., Winter, G. (1992a) Molecular evolution of proteins on filamentous phage. Journal of Biological Chemistry, 267, 16007-16010. Marks, J.D., Griffiths, A.D., Malmqvist, M., Clackson, T.P., Bye, J.M. Winter, G. (1992b). By passing immunization: building high affinity human antibodies by chain shuffling. Biotechnology 10: 779:783. Meinkoth J., Wahl G. (1984) Hybridization of nucleic acids immobilized on solid supports. Anal Biochem, 138 (2): 267-284. Mullis, K.B., Faloona, F.A. (1987) Specific synthesis of DNA in vitro via a polymerasecatalyzed chain reaction. Meth. Enzymol. 155: 335-350. Pierik, R.L.M. (1999) In vitro Culture of Higher Plants, 4th edition, 360 pages, ISBN: 0-7923-5267-X. Rose, N., DeMacrio, E., Fahey, J., Friedman, H., Penn, G. (1997) Manual of Clinical Laboratory Immunology. American Soc. Microbiology Press, Washington, D.C. Sambrook, J., Russell D.W., Sambrook, J. (2001) Molecular Cloning: a Laboratory Manual. Cold Spring Harbor Laboratory Press, Plainview, N.Y. Sanger, F., Nicklen, S., Coulson, A.R. (1977) DNA sequencing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. U.S.A. 74;5463-5467. Schagger H., von Jagow, G. (1987) Tricinesodium dodecyl sulfate-polyacrylamide gel electrophoresis for the separation of proteins in the 59 range from 1 to 100kDa. Analytical Biochemistry 166, 368-379. Smith, L.M., Sanders, J.Z., Kaiser, R.J., Hughs, P., Dodd, C, Connell, C.R., Heins, C, Kent, S.B.H., Hood, L.E. (1986) Fluorescent detection in automated DNA sequence analysis. Nature 321:673-681. Swofford, D.L. (2000). PAUP*: phylogenetic analysis using parsimony (*and other methods). 4th edition. Sinauer Associates, Sunderland, Mass. Takeshita, M., Suzuki, M., Takanami, Y. (2001). Combination of amino acids in the 3 a protein and the coat protein of Cucumber mosaic virus determines symptom expression and viral spread in bottle gourd. Arch Virol 146:697-711. Thompson, J.D., Gibson, T.J., Plewniak, F., Jeanmougin, F., Higgins, D.G. (1997) The Clustal windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Research, 24:4876-4882. Thompson, J.R., Perry, K.L., De Jong, W. (2004a) A new potato virus in a new lineage of picorna-like viruses. Arch Virol. 149:2141-2154. Thompson, J.R., Perry, K.L., De Jong, W. (2004b) The characterization of a new picorna-like virus infecting tomato. Abstract book of the 23rd Annual Meeting of the American Society for Virology, Montreal, Canada, July 2004. Tijssen, P. 1993. Laboratory Techniques in Biochemistry and Molecular Biology-Hybridization 94035 60 with Nucleic Acid Probes, Part I, Chapter 2 "Overview of principles of hybridization and the strategy of nucleic acid probe assays", Elsevier. New York. Van den Bruie, A.J.C., Pol, R., FransenDaalmeijer, N., Schouls, L.M., Meijer, C. J. L.M., Snijders, P.J.F. (2002) GP5+/6+PCR followed by Reverse Line Blot Analysis Enables Rapid and HighThroughput Identification of Human Papillomavirus Genotypes. J. Clin. Microbiol. 40: 779-787. Vela, C, Cambra, M., Cortes, E., Moreno, P., Miguet, J.G., Perez de San Roman, C, Sanz, A. (1986) Production and characterization of monoclonal antibodies specific for citrus tristeza virus and their use for diagnosis. J. Gen. Virol. 67: 91-96. Walker, G.T., Fraiser, M.S., Schram, J.L., Little, M.C., Nadeau, J.G., Malinowski, D.P. (1992) Strand displacement amplification - an isothermal, in vitro DNA amplification technique Nucleic Acids Res 20:1691-1696. Walker, J.M. 2004. PCR Protocols on CD. Humana Press. Wisler, G.C., Li, R.H., Liu, H.Y., Lowry, D.S., Duffus, J.E. (1998) Tomato chlorosis virus: a new whitefly-transmitted, phloem-limited, bipartite closteroviras of tomato. Phytopathology, 88 (5), 402409. Young R.A., Davis, R.W. (1983) Efficient isolation of genes using antibody probes. Proc. Natl. Acad. Set U.S.A. 80:1194-1198.
ДивитисяДодаткова інформація
Назва патенту англійськоюNovel plant virus specified as tomato torrado virus
Автори англійськоюVan den Khevel Johannes Franciscus Johanna Maria, Maris Paulus Kornelis, Verbek Marinus, Dullemans Annette Maria, van der Wluigt Rene Andries Antonius
Назва патенту російськоюНовый вирус растений, обозначенный как томато торрадо вирус
Автори російськоюВан ден Хевел Йоханнес Францискус Йоханна Мария, Марис Паулюс Корнелис, Вербек Маринус, Дюллеманс Аннетт Мария, ван дер Влюгт Рене Андриес Антониус
МПК / Мітки
МПК: C07K 14/08, A01H 5/08, C12N 7/01, G01N 33/569
Мітки: новий, вірус, торрадо, томато, рослин, позначений
Код посилання
<a href="https://ua.patents.su/35-94035-novijj-virus-roslin-poznachenijj-yak-tomato-torrado-virus.html" target="_blank" rel="follow" title="База патентів України">Новий вірус рослин, позначений як томато торрадо вірус</a>
Спосіб консервування патологічного матеріалу, що містить вірус

Номер патенту: 71414
Опубліковано: 15.11.2004
Автори: Циганенко Анатолій Якович, Павленко Неоніла Володимирівна, Одражий Надія Костянтинівна, Герасюта Ніна Миколаївна, Цуцаєва Алла Олександрівна, Онасенко Олена Станіславна, Желтякова Ірина Олександрівна
МПК: A01N 1/02
Мітки: вірус, спосіб, консервування, містить, патологічного, матеріалу
Формула / Реферат:
Спосіб консервування патологічного матеріалу, що містить вірус, який включає заморожування і зберігання при низькій температурі, який відрізняється тим, що заморожування матеріалу здійснюють шляхом швидкого занурення у рідкий азот з наступним зберіганням при температурі -196°С.
Спосіб впливу на вірус сніду

Номер патенту: 31202
Опубліковано: 25.03.2008
Автори: Солодовников Володимир Ілліч, Капустин Володимир Олександрович, Каневський Валерій Олександрович
МПК: C12N 7/04
Мітки: впливу, спосіб, сніду, вірус
Формула / Реферат:
1. Спосіб впливу на вірус СНІДу, який включає опромінення світлом вірусного матеріалу, який відрізняється тим, що вірусний матеріал опромінюють монохромним світлом у діапазоні 382-385 нм протягом 7-20 хвилин.2. Спосіб впливу на вірус СНІДу за п. 1, який відрізняється тим, що щільність енергії монохромного світла становить 150-200 мВт/см2.3. Спосіб впливу на вірус СНІДу за п. 1, який відрізняється тим, що вплив на вірусний...
Вірус-вакцина-2 проти інфекційної бурсальної хвороби птиці

Номер патенту: 69089
Опубліковано: 16.08.2004
Автори: Герман Євген Вячеславович, Герман Ірина В'ячеславівна, Герман В'ячеслав Валентинович, Стегній Борис Тимофійович
МПК: A61K 39/12
Мітки: хвороби, птиці, бурсальної, інфекційної, вірус-вакцина-2
Формула / Реферат:
Вірус-вакцина-2 проти інфекційної бурсальної хвороби птиці (хвороби Гамборо), що містить штам УМ-93, який антигенно споріднений з епізоотичними штамами, який відрізняється тим, що вірус культивують на гетерологічній біосистемі, а імуногенну активність вірусу підтримують на курячих ембріонах, вільних від патогенів.
Молекула днк, яка кодує стійку до дії інгібітора рослинну протопорфіриногеноксидазу, спосіб одержання стійких до гербіциду рослин, спосіб боротьби з ростом небажаних рослин

Номер патенту: 70912
Опубліковано: 15.11.2004
Автори: Хейфетц Пітер Б., Волрат Сандра Л., Уорд Ерік Р., Джонсон Марі А., Поттер Шерон Л.
МПК: C12N 15/82, A01H 5/00, C12Q 1/68, C12N 9/02, C12N 5/10, G01N 33/15, C12N 9/99, C12N 15/09, C12Q 1/26, C12N 15/53
Мітки: молекула, рослинну, інгібітора, днк, ростом, стійких, небажаних, дії, спосіб, рослин, боротьби, протопорфіриногеноксидазу, яка, гербіциду, одержання, стійку, кодує
Формула / Реферат:
1. Виділена молекула ДНК, яка включає ділянку, яка кодує модифіковану протопорфіриногеноксидазу (протокс), причому немодифікований протокс являє собою протокс рослини, вибраний із групи, яка включає: SEQ ID NO: 2; 4; 6; 8; 10; 12; 16; 18; 20; 22 і 24, і має модифікацію, яка включає першу амінокислотну заміну, яка відбувається в положенні, вибраному з групи, яка включає:(а) положення, яке відповідає аланіну в положенні 164...
Спосіб виготовлення живої та інактивованої вірус-вакцини проти ньюкаслської хвороби птиці

Номер патенту: 14289
Опубліковано: 15.05.2006
Автори: Стегній Борис Тимофійович, Вовк Сергій Іванович, Гадзевич Дмитро Вікторович
МПК: A61K 39/17
Мітки: вірус-вакцини, виготовлення, спосіб, живої, інактивованої, ньюкаслської, птиці, хвороби
Формула / Реферат:
Спосіб виготовлення живої та інактивованої вірус-вакцини проти Ньюкаслської хвороби птиці, що включає накопичення вірусної сировини штаму "Ла-Сота", визначення гемаглютинуючої активності, змішування компонентів вакцини, який відрізняється тим, що культивують штам "Ла-Сота" на перещеплюваних культурах клітин свинячої нирки ембріональної версенізированої, нирки вівці, коронарних судинах телят.
Попередній патент: Аналоги рапаміцину та їх застосування у лікуванні неврологічних, проліферативних та запальних розладів
Наступний патент: Фармацевтична композиція бендамустину, призначена для ліофілізації
Випадковий патент: Спосіб підготовки кишечнику до операції