Насінина трансгенної кукурудзи з підвищеним вмістом лізину
Номер патенту: 102367
Опубліковано: 10.07.2013
Автори: Малвар Томас М., Хуанг Шихших, Люті Міхель Х.








Формула / Реферат
1. Насінина трансгенної кукурудзи, що містить більше ніж 4000 частин на мільйон (ppm) вільного лізину, де насінина кукурудзи містить нативну ДНК і одну або декілька екзогенних касет для експресії в рослинах, що містять молекулу ДНК, що кодує дигідродипіколінатсинтазу, в значній мірі стійку до інгібування вільним L-лізином за принципом зворотного зв'язку, молекулу ДНК, що транскрибується з отриманням молекули РНК, що пригнічує лізинкетоглутаратредуктазу/сахаропіндегідрогеназу, і молекулу ДНК, що кодує аспартаткіназу, в значній мірі стійку до дії лізин-опосередкованого зворотного зв'язку; де молекули ДНК функціонально пов'язані з однією або декількома промоторними молекулами таким чином, що відбувається транскрипція однієї або декількох молекул РНК в основному в ендоспермі кукурудзяної насінини, де, необов'язково, насінина кукурудзи містить екзогенний ген, що надає стійкості до гербіцидів.
2. Кукурудзяне борошно, одержане з насінини кукурудзи за п. 1, в яке не доданий лізин.
3. Насінина трансгенної кукурудзи за п. 1, яка містить екзогенний ген, що надає стійкості до гербіцидів.
4. Насінина трансгенної кукурудзи за п. 3, яка є стійкою до гербіциду гліфосат.
5. Застосування насінини трансгенної кукурудзи за п. 1 для одержання корму.
6. Застосування насінини трансгенної кукурудзи за п. 1 для одержання борошна грубого помелу.
7. Застосування насінини трансгенної кукурудзи за п. 1 для одержання білкового концентрату або ізоляту.
8. Спосіб одержання кукурудзяного борошна, який включає одержання насінини трансгенної кукурудзи, яка має вміст лізину щонайменше 4000 частин на мільйон, де насінина кукурудзи містить нативну ДНК і одну або декілька екзогенних касет для експресії в рослинах, що містять молекулу ДНК, що кодує дигідродипіколінатсинтазу, в значній мірі стійку до інгібування вільним L-лізином за принципом зворотного зв'язку, молекулу ДНК, що транскрибується з отриманням молекули РНК, що пригнічує лізинкетоглутаратредуктазу/сахаропіндегідрогеназу, і молекулу ДНК, що кодує аспартаткіназу, в значній мірі стійку до дії лізин-опосередкованого зворотного зв'язку; де молекули ДНК функціонально пов'язані з однією або декількома промоторними молекулами таким чином, що відбувається транскрипція однієї або декількох молекул РНК в основному в ендоспермі кукурудзяної насінини, де, необов'язково, насінина кукурудзи містить екзогенний ген, що надає стійкості до гербіцидів, без додавання лізину, і переробку насінини кукурудзи з одержанням кукурудзяного борошна.
9. Спосіб за п. 8, де вказана насінина трансгенної кукурудзи містить екзогенний ген, що надає стійкості до гербіцидів.
10. Спосіб за п. 9, де насінина трансгенної кукурудзи є стійкою до гербіциду гліфосат.
11. Застосування насінини трансгенної кукурудзи за п. 1 для вживання людиною чи твариною.
Текст
Реферат: Винахід належить до насінини трансгенної кукурудзи, що містить більше ніж 4000 частин на мільйон (ppm) вільного лізину, де насінина кукурудзи містить нативну ДНК і одну або декілька екзогенних касет для експресії в рослинах, що містять молекулу ДНК, що кодує дигідродипіколінатсинтазу, в значній мірі стійку до інгібування вільним L-лізином за принципом зворотного зв'язку, молекулу ДНК, що транскрибується з отриманням молекули РНК, що пригнічує лізинкетоглутаратредуктазу/сахаропіндегідрогеназу, і молекулу ДНК, що кодує аспартаткіназу, в значній мірі стійку до дії лізин-опосередкованого зворотного зв'язку; де молекули ДНК функціонально пов'язані з однією або декількома промоторними молекулами таким чином, що відбувається транскрипція однієї або декількох молекул РНК в основному в ендоспермі кукурудзяної насінини. UA 102367 C2 (12) UA 102367 C2 UA 102367 C2 5 10 15 20 25 30 35 40 45 50 55 60 Пріоритет, який вимагається За даною заявкою вимагається пріоритет попередньої заявки США з серійним номером 60/723178, поданої 3 жовтня 2005 р. і повністю включеної до даного опису за допомогою посилання. Попередній рівень техніки 1. Галузь техніки, до якої належить винахід У даному описі розкриті ДНК-конструкції, придатні для отримування трансгенних рослин з частинами, які мають підвищений вміст лізину, а також способи використання таких ДНКконструкцій для отримування трансгенних рослин і насіння, зібраного з рослин матеріалів, а також борошна і харчових продуктів, отриманих із вищепереліченої сировини. Такі ДНКконструкції придатні для отримання трансгенних рослин підвищеної харчової цінності, зумовленої підвищенням накопичення лізину в насінні. Також розкриті полінуклеотидні молекули і способи, придатні для отримання трансгенних рослин з підвищеним вмістом лізину в насінні. 2. Опис рівня техніки Zea mays, звичайно називається маїсом або кукурудзою, являє собою зернову культуру, що широко використовується як їжа для людини і тваринний корм. Зерно, в даному описі, яке називається також насінням, є джерелом білка, крохмалю і олії для багатьох нижчих тварин, в тому числі для свиней, м'ясної і молочної худоби, риби і домашньої птиці. У деяких країнах, таких як Мексика, більше 70% зібраної кукурудзи споживається людьми, і кукурудза і продукти, що отримуються з неї, такі як мамалига і плоскі маїсові коржики, є харчовими продуктами масового споживання. Основна частина амінокислотного складу зерна визначається кількістю і типом амінокислот, які містяться в поліпептидах. Тільки відносно невелика частина, до 10%, доступних амінокислот в зерні знаходиться у вигляді вільних амінокислот; інші містяться в різних білках. Більшість поліпептидів кукурудзи, що знаходяться в зерні, є запасними білками насіння, або зеїнами. Ці запасні білки насіння синтезуються в ході розвитку зерна і використовуються як джерело енергії в ході проростання і росту останнього. Однак ці запасні білки насіння містять мало лізину, або взагалі не містять цієї амінокислоти. З десяти амінокислот, які вважаються незамінними в змішаних зернових кормах (аргінін, гістидин, ізолейцин, лейцин, лізин, метіонін або цистеїн, фенілаланін або тирозин, треонін, триптофан і валін), не тільки лізин, але також треонін і метіонін практично відсутні в кукурудзяному зерні. Недостача цих незамінних амінокислот, особливо лізину, вимагає додавання цих поживних речовин до харчового кукурудзяного зерна, що часто досягається шляхом додаткового введення соєвого борошна або синтетичного лізину. Збільшення вмісту лізину в насінні кукурудзи з метою підвищення поживної цінності останнього, як їжа або кормове зерно може бути корисним в даній галузі техніки. Додаткова перевага може бути реалізована в тому випадку, якщо до зерна треба буде додавати лише малі кількості лізину, або не додавати його взагалі. Відносно харчування людини, має місце погана тенденція до вживання їжі, відносно багатої недорогими крохмалевмісними продуктами і відносно бідної високоякісним білком. Квашіоркор (дитяча пелагра) - форма порушення харчування, викликана споживанням білків неадекватної якості при загальній високій енергетичній цінності (загальної калорійності) споживаної їжі. Ранні симптоми є дуже загальними і включають втому, дратівливість і сонливість. При тривалому дефіциті білка спостерігається відставання в рості, втрата м’язової маси, генералізований набряк (едема) і зниження імунітету. Звичайним є здуття живота. Часто спостерігаються шкірні прояви (такі як дерматит, зміна пігментації, потоншання волосся і вітиліго). Смерті передують шок і кома. Згідно з однією з урядових оцінок, не менше 50% літніх людей в будинках для людей похилого віку США страждають від порушення харчування, пов'язаного з низьким рівнем білкової калорійності їжі. Таким чином, продукти, отримані з кукурудзи з поліпшеною в результаті підвищеного вмісту лізину якістю білкового складу, могли б значно поліпшити не тільки поживну цінність харчових продуктів, але і загальний рівень здоров'я. Молекулярно-біологічний підхід до підвищення вмісту лізину включає ідентифікацію і введення в кукурудзу генів з метою надання впливу на вміст лізину в зерні, що не супроводжується негативним впливом на агрономічні властивості. Лізин продукується з аспартату, метаболізм якого, в свою чергу, взаємно пов'язаний з біохімічними шляхами, в яких беруть участь треонін, метіонін і ізолейцин. Сам лізин служить кінцевим продуктом, що модифікує і регулює ферменти, які беруть участь в його власному біосинтезі. Ключовими ферментами, які беруть участь в метаболізмі лізину, є аспартаткіназа (AK) і дигідродипіколінатсинтаза (DHDPS), в той час як лізинкетоглутаратредуктаза (LKR, біфункціональний фермент, також відомий як цукропіндегідрогеназа, SDH), мабуть, відіграє 1 UA 102367 C2 5 10 15 20 25 30 35 40 45 50 55 60 ключову роль в катаболізмі лізину. AK і DHDPS формують петлю зворотного зв'язку, регульовану лізином; в міру підвищення вмісту лізину він придушує активність цих ферментів. Таким чином, для підвищення вмісту лізину переважно оперувати з варіантами AK або DHDPS, нечутливими до зворотного зв'язку. Були описані нечутливі до лізину варіанти бактерійного, а також рослинного генів AK (див. Falco з співавторами, патент США № 5773691, включений до даного опису за допомогою посилання). Нечутливі до лізину форми AK були ідентифіковані в ячмені, кукурудзі і тютюні. Було показано, що бактерійний ген DHDPS, виділений з E. coli, щонайменше в 20 разів менш чутливий до підвищення вмісту лізину (Glassman з співавторами, патент США № 5258300; Galili з співавторами, патент США № 5367110). У публікаціях Falcо з співавторами (патенти США № 5773691 і 6459019, публікація заявки на патент США № 2003/0056242) і Dizigan з співавторами (патентна публікація № 2005/0132437), кожна з яких повністю включена до даного опису за допомогою посилання, описане виділення і використання нечутливої до лізин-опосередкованого зворотного зв'язку аспартаткінази (ген AK, відомий як lysС) з E. coli, DHDPS з E. coli, а також DHDPS з Corynebacterium (який відомий і означається в даному описі як CORgl. dapА), для отримування трансгенних рослин - рапсу, тютюну, кукурудзи і сої, з підвищеним вмістом лізину в насінні. Даний винахід належить до ДНК-конструкцій, які, будучи експресованими в рослинних клітинах, забезпечують підвищення поживної цінності рослин, зокрема, підвищений вміст лізину в насінні рослини. Короткий опис суті винаходу Даний винахід належить до ДНК-конструкцій, які при функціонуванні в геномі трансгенної рослини, забезпечують підвищення вмісту лізину в насінні рослини. Зібране насіння з високим вмістом лізину може бути корисне для виробництва харчової і кормової продукції, такої як кукурудзяне борошно (у тому числі борошно, відоме як «маса Харіна»), кукурудзяне борошно і борошняні вироби, в які треба додавати лише невеликі кількості лізину, або не треба додавати його взагалі. Переважно, трансгенна рослина і насіння являє собою рослину і зерно сільськогосподарських культур, переважно - однодольну рослину і насіння і найбільш переважно - насіння і рослину кукурудзи. Даний винахід належить до трансгенних рослин і насіння, які містять ДНК-конструкції з підвищеним вмістом лізину в насінні. ДНК-конструкції за даним винаходом включають молекулу, яка, будучи експресованою в клітинах кукурудзи, інгібує продукцію лізин-деградуючого поліпептиду, а також молекулу, яка, будучи експресованою в клітинах кукурудзи, забезпечує продукцію нечутливих до лізину поліпептиду дигідродипіколінатсинтази і аспартаткінази. Навіть в тому випадку, якщо зібране зерно є нежиттєздатним і, таким чином, не може бути використане як насіння, воно проте містить підвищений вміст лізину і характеризується підвищеною якістю білка згідно з даним винаходом. Подібне зібране зерно може бути перероблене тим же чином, що і звичайне зерно. Відповідно, в іншому втіленні даний винахід належить до корму для тварин з підвищеним вмістом лізину, що отримується шляхом вирощування трансгенної рослини за даним винаходом, і збору насіння з підвищеним вмістом лізину, або шляхом переробки за даним винаходом, або його частини. У іншому втіленні даний винахід належить до борошна або борошняних виробів, що отримуються шляхом вирощування трансгенної рослини за даним винаходом, і збору насіння з підвищеним вмістом лізину, або шляхом переробки насіння за даним винаходом, або його частини. В одному з втілень даного винаходу ДНК-конструкція включає нові полінуклеотидні композиції, вдосконалені для експресії в рослинних клітинах, особливо в клітинах кукурудзи. Один з нових полінуклеотидів (SEQ ID NO:5) кодує поліпептид дигідродипіколінатсинтази, інший новий полінуклеотид (SEQ ID NO:9) є гомологічним і комплементарним частини полінуклеотидної послідовності, що кодує лізинкетоглутаратредуктазу. Експресія цієї ДНКконструкції в рослинній клітині, особливо в клітині кукурудзи, переважно - в клітині кукурудзяного насіння приводить до підвищення вмісту лізину в кукурудзяному насінні. ДНК-конструкція включає полінуклеотиди, наприклад, таку послідовність, як SEQ ID NO:1, що кодує нечутливу до лізину дигідродипіколінатсинтазу, а також таку послідовність, як SEQ ID NO:2, що кодує лізинкетоглутаратредуктазу і використовується як матриця для конструювання інгібіторних молекул РНК, або таку послідовність, як SEQ ID NO:3, що кодує аспартаткіназу. ДНКконструкція переважно містить молекулу промотору, вдосконалену для керування транскрипцією функціонально пов'язаного полінуклеотиду в насінні кукурудзи. Крім цього, ДНКконструкція може містити нову молекулу ДНК (SEQ ID NO:10), яка містить молекулу для придушення гену, мішенню якої є ген лізинкетоглутаратредуктази, де молекула, що придушує ген, вміщена в інтрон ДНК-конструкції. 2 UA 102367 C2 5 10 15 20 25 30 35 40 45 50 55 60 У іншому втіленні даний винахід належить до способу отримання кукурудзяного насіння з підвищеним вмістом лізину, за допомогою схрещування інбредних батьківських рослин з подальшим отримуванням гібридного насіння, при якому кожне з батьківських рослин містить в своєму геномі одну або декілька ДНК-конструкцій, які складають предмет даного винаходу, в складі касет для експресії в рослинах, причому в кожній з батьківських рослин експресійні касети різні, і жодна з батьківських рослин не містить всіх трьох; а також подальшим збиранням насіння, що має вміст лізину вище, ніж в кожному з батьківських рослин. У іншому втіленні даний винахід належить до способу отримання продуктів харчування людини або кормових продуктів для тварин, які містять підвищений вміст лізину, що включає стадію переробки насіння або його частини, отриманих з трансгенних рослин, геном яких містить екзогенну ДНК-конструкцію або набір конструкцій згідно з даним винаходом. Крім цього, даний винахід належить до способу отримання продуктів харчування людини або кормових продуктів для тварин, які мають підвищений вміст лізину, що включає стадію вирощування трансгенної рослини з подальшим отриманням насіння, геном яких містить екзогенну ДНКконструкцію за даним винаходом, з подальшим збиранням насіння. Короткий опис графічного матеріалу Фіг. 1. Карта плазміди pMON66649 Фіг. 2. Карта плазміди pMON80003 Фіг. 3. Карта плазміди pMON79465 Фіг. 4. Карта плазміди pMON93092 Фіг. 5. Карта плазміди pMON93093 Фіг. 6. Карта плазміди pMON80378 Фіг. 7. Вміст вільного лізину в зернах, отриманих з об'єктів, що містять pMON93092. Фіг. 8. Вміст вільного лізину в зернах, отриманих з об'єктів, що містять pMON93093. Докладний опис винаходу Трансгенна рослина або насіння, що виявляє бажану властивість, наприклад, підвищений вміст лізину за винаходом, містить специфічну екзогенну ДНК, вбудовану в геном трансгенної рослини і яка обумовлює наявність бажаної властивості. Дана властивість може бути виражена як зміна природної властивості контрольної рослини, що вимірюється, наприклад, рослини або насіння практично того ж генотипу, позбавленого специфічної екзогенної ДНК. Переважно, зміна оцінюється шляхом зіставлення кількісного вираження даної властивості в трансгенній рослині або насінні, що має специфічну екзогенну ДНК, і кількісного вираження тієї ж властивості в контрольній рослині або насінні. Таким чином «маїс з підвищеним вмістом лізину» являє собою рослину кукурудзи (маїсу) з підвищеним вмістом лізину в будь-якій частині рослини, переважно в насінні, яке в даному описі також може бути назване зерном (у тому числі і в агрегованій формі). Даний винахід належить до ДНК-конструкцій і насіння, які містять в своєму геномі принаймні одну з ДНК-конструкцій в складі касет, експресії в рослинах за даним винаходом, а також підвищеним вмістом лізину, що характеризується в порівнянні з насінням, яке не містить конструкції. Підвищений вміст лізину може виявлятися рослиною шляхом накопичення підвищених кількостей даної амінокислоти в зерні, і може бути виміряне за допомогою будь-якого відповідного способу, такого як мас-спектрофотометрія або високоефективна рідинна хроматографія відповідним чином отриманого тканинного екстракту. Трансгенне кукурудзяне зерно з підвищеним вмістом лізину за даним винаходом особливо корисне як харчовий продукт або корм, а також борошно або борошняні вироби, як сировина для отримання очищених кукурудзяних білкових продуктів, або як сировина для отримання інших продуктів, що отримуються із зерна, яке містить підвищений вміст лізину в порівнянні з нетрансгенним зерном подібного сорту. Загальний вміст лізину в кукурудзяному зерні, який складає приблизно 4000 частин на мільйон (ppm), є достатнім для того, щоб в харчові продукти, які отримуються з цього зерна або корми не треба було додавати лізин з додаткових джерел. Продукція і накопичення лізину в кукурудзі з підвищеним вмістом цієї амінокислоти за даним винаходом, досягає вищезгаданого рівня. Даний винахід належить до способів і композицій, значно поліпшених в порівнянні з раніше описаними способами і композиціями для підвищення вмісту лізину в кукурудзяному зерні (див., наприклад, патент США № 5773691). Способи підвищення вмісту амінокислот в трансгенних рослинах включають експресію рекомбінантних молекул ДНК, що кодують білки, залучені в шляхи синтезу амінокислот, на більш високому рівні в порівнянні з експресією нативних генів. Одним зі способів підвищення вмісту лізину в кукурудзі є експресія бактерійної дигідродипіколінатсинтази (DHDPS), згідно з розкритим в патентах США № 5288300, 6459019, а також в публікації патентної заявки США № 3 UA 102367 C2 5 10 15 20 25 30 35 40 45 50 55 60 2003/0056242А1, і придушування ендогенної LKR (публікація патентної заявки США № 2005/0193444); кожна з вищенаведених в даному абзаці публікацій повністю включена до даного опису за допомогою посилання. Будь-яка з рослин або їх частин за даним винаходом може бути перероблена для отримання корму, борошна тонкого і грубого помелу, білкових продуктів або олії. Особливо переважною для цих цілей частиною рослин є насіння. У одному з переважних втілень даного винаходу корм, борошно тонкого і грубого помелу, білкові продукти або олія використовуються для годування сільськогосподарських тварин (худоби). Способи отримання корму, борошна тонкого і грубого помелу, білкових продуктів або олії відомі в даній галузі техніки, див., наприклад, патенти США № 4957748, 5100679, 5219596, 5936069, 6005076, 6146669 і 6156227, повністю включених до даного опису за допомогою посилання. У переважному втіленні даного винаходу білкові продукти являють собою продукти з високим вмістом білка, такі як концентрати або ізоляти. Продукт з високим вмістом білка має вміст білка, що перевищує 5% мас./об., переважно більше 10% мас./об., ще більш переважно 15% мас./об. У іншому втіленні борошно за даним винаходом може бути змішане з іншими борошняними продуктами. У переважному втіленні даного винаходу борошно, отримане з рослин або насіння за даним винаходом, або за допомогою способу за даним винаходом, складає більш ніж приблизно 0,5%, приблизно 1%, приблизно 5%, приблизно 10%, приблизно 25%, приблизно 50%, приблизно 75% або приблизно 95% за об'ємом або масою борошняного компонента будьякого продукту. У іншому втіленні даного винаходу борошняний продукт може бути змішаний і складати більше приблизно 10%, більше приблизно 25%, більше приблизно 35%, більше приблизно 50% або більше приблизно 75% від суміші за об'ємом. Даний винахід також належить до способу посилення інтенсивності росту людини або нижчої тварини, що включає вживання людиною або нижчою твариною їжі, принаймні частина якої являє собою кукурудзу з високим вмістом лізину, що містить одну або декілька касет, для експресії в рослинах, за даним винаходом. У даному описі термін «нижчі тварини» належить до птахів, риб, а також ссавців. Рекомбінантні ДНК-конструкції Даний винахід належить до ДНК-конструкцій, які містять функціонально пов'язані полінуклеотиди, здатні забезпечити підвищений вміст лізину в зернах кукурудзи. Термін «ДНКконструкція» включає без обмеження полінуклеотидні молекули, які керують транскрипцією пов'язаних полінуклеотидних молекул в клітині рослини. Щонайменше частина ДНК-конструкції за даним винаходом може використовуватися в способі трансформації рослинної клітини, що має на увазі стабільне вбудовування цієї частини ДНК-конструкції в геном даної клітини. Ця частина обумовлює наявність фенотипу, який характеризується високим вмістом лізину в зернах кукурудзи або борошняних продуктах, виготовлених з таких зерен, і є аспектом даного винаходу. ДНК-конструкції pMON93092 і pMON09093 були депоновані Monsanto Technology LLC згідно з Будапештською угодою в Американській колекції типових культур (АТСС) під номерами доступу PTA-6733 і PTA-6734, відповідно. Дані депозити становлять частину розкриття даного винаходу, і хоч послідовності, приведені в даному письмовому розкритті, на думку авторів, є точними, при необхідності полінуклеотидні послідовності молекул ДНК, що містять регуляторні елементи, кодуючі і некодуючі молекули, можуть бути визначені за допомогою цих депозитів і використані для корекції послідовностей, розкритих в даному винаході. Полінуклеотидні молекули пов'язані в рекомбінантних ДНК-конструкціях за допомогою способів, відомих фахівцям в даній галузі. Прикладом придатної технології створення ДНКконструкцій може служити технологія клонування GATEWAY™ (Invitrogen Life Technologies, Carlsbad, California), в якій застосовується сайт-специфічна реакція клонування LR-рекомбінази в системі Integrase/att бактеріофагу лямбда для створення векторної конструкції без використовування ендонуклеаз і лигаз. У посібнику по використанню технології клонування GATEWAY™ компанії Invitrogen також приводяться чіткі інструкції по стандартному клонуванню будь-якого бажаного полінуклеотиду у вектор, що містить діючі елементи для експресії в клітинах рослин. Додаткові способи розкриті в Molecular Cloning: А Laboratory Manual, 3-є видання, томи 1, 2 і 3. J. F. Sambrook, D. W. Russell і N. Irwin, Cold Spring Harbor Laboratory Press, 2000 (в даному тексті посилання на дане джерело приводяться як «Sambrook з співавторами»). Використовуваний в даному описі термін «екзогенна ДНК або виділена ДНК» належить до молекули ДНК, яка в нормі не присутня в геномі клітини-хазяя, або ж до ДНК, яка в нормі не присутня в геномі клітини-хазяя в ідентичному контексті, або ж до двох пов'язаних прилеглих одна до одної послідовностей, які в нормі або в природі не пов'язані одна з одною. Екзогенна ДНК може включати полінуклеотидну молекулу ДНК або РНК, яка є нативною для геному клітини-хазяя, або ж може містити нативний полінуклеотид, змінений шляхом приєднування або 4 UA 102367 C2 5 10 15 20 25 30 35 40 45 50 55 60 делеції одного або декількох полінуклеотидів, або ж шляхом модифікацій регуляторних елементів асоційованого полінуклеотиду або іншої полінуклеотидної послідовності згідно з описаним в даному описі. У доповнення до регуляторних елементів екзогенна ДНК може містити кодуючу послідовність, продуктом якої є білок або поліпептид, або ж некодуючу послідовність, продуктом якої є небілковий продукт, такий як, наприклад, молекула РНК, що впливає на транскрипцію або трансляцію іншої послідовності ДНК. ДНК-конструкція за даним винаходом містить кодуючу і некодуючу послідовності. Використовувані для трансформації рослинних клітин екзогенні ДНК-конструкції містять кодуючу послідовність, яка являє собою інтерес, а також звичайно інші регуляторні елементи згідно з описаним в даному описі, такі як (без обмеження) промотори, інтрони, 5'- і 3'нетрансльовані області, лідерні послідовності, послідовності сигналів локалізації і транзитних сигналів, а також енхансери, функціональне з'єднання яких обумовлює експресію РНК в клітинах рослин. Подібні ДНК-конструкції в контексті даного винаходу називаються касетами для гетерологічної експресії в рослинах. ДНК-конструкції, перераховані в таблиці 1, ілюструють зв'язки регуляторних елементів в експресійних касетах, які експресують кодуючі і некодуючі послідовності; крім цього також вказаний середній рівень лізину в трансгенному насінні маїсу, яке містить кожну конструкцію. Прикладами молекул промоторів ДНК екзогенної ДНК-конструкції за даним винаходом можуть служити промотори, що посилюються в насінні або насінневоспецифічні промотори, що включають, наприклад, промотори, використовувані в однодольних сільськогосподарських культурах, такі як кукурудзяний L3-промотор (промотор, описаний в патенті США № 6433252, включений до даного опису за допомогою посилання, або його гомологи; в контексті даного винаходу - P-Zm. L3), кукурудзяний B32-промотор (послідовність SEQ ID NO:4 або її гомологи; в контексті даного винаходу - P-Zm. B32), промотор кукурудзяного гамма-коїксину (промотор, описаний в патенті США № 6635806, включений до даного опису за допомогою посилання, або його гомологи; в контексті даного винаходу - P-Zm. Gcx), а також будь-який з промоторів, використовуваних в ДНК-конструкціях за даним винаходом, або інші промотори, якііндукують підвищену транскрипцію в насінні кукурудзи; всі подібні промотори в контексті даного винаходу означаються як P-Zm. seed. Приклади додаткових промоторів, які функціонують в насінні, включають без обмеження промотори, що індукують експресію запасних білків насіння, таких як рисовий глутелін і проламінові білки Triticeae, а також промотори, які контролюють експресію ферментів, залучених до синтезу крохмалю в насінні, таких як промотори, що активують експресію АДФ-глюкоза-крохмаль-глікозилтрансферази однодольних рослин. Вибраний промотор насіння функціонально пов'язаний з кодуючою областю гену DHDPS (наприклад, послідовність SEQ ID NO:1, в контексті даного винаходу що означається як CORgl. dapA), або з кодуючою областю гену аспартаткінази (наприклад, послідовність SEQ ID NO:3). Додаткові промотори, що посилюються в насінні або насіннево-специфічні промотори, відомі в даній галузі і можуть бути вибрані і протестовані у функціональному зв'язку з кодуючими і некодуючими молекулами ДНК за даним винаходом, за допомогою способів, описаних в контексті даного винаходу, або подібних способів, відомих фахівцям в галузі молекулярної біології рослин. Промотор, що посилюється в насінні, первинно експресується у всьому насінні або його частині, хоч може бути вторинно експресований в інших клітинах, тканинах або органах рослини. Частини насіння включають без обмеження зародок, ендосперм, насіннєву оболонку, колеоптиль, сім’ядолю, підсім’ядольне коліно, алейроновий шар, перикарпій і скутелум. Додаткові регуляторні елементи, що містяться в ДНК-конструкціях, включають без обмеження інтрон (I-), транзитні сигнали (TS-), 5'-нетрансльовані лідерні послідовності (L-), 3'-області поліаденілювання (Т-) або некодуючі антисмислові області. Також можуть бути включені експресійні касети, які містять селектований або кількісно оцінюваний маркер для селекції або ідентифікації трансформованих рослинних клітин і тканин. Конструкція за даним винаходом необов'язково містить експресійну касету, яка додає кукурудзяній рослині стійкість до гліфосату і може бути використана як селектований маркер. Використовуваний термін в даному описі «трансген» належить до екзогенної ДНК, вбудованої в геном хазяя або здібної до автономної реплікації в клітині-хазяї, що експресує один або декілька клітинних продуктів. За допомогою подібних трансгенів можна отримати клітину-хазяя, рослини, отримувані з неї або частини цих рослин, з новим в порівнянні з нетрансформованою клітиною або рослиною фенотипом, наприклад, з підвищеним вмістом лізину в кукурудзяному зерні. Трансгени можуть бути безпосередньо введені в рослини за допомогою генетичної трансформації і переважно є стабільними і успадкованими таким чином, що вони можуть бути успадковані від рослин будь-якого попереднього покоління, які були трансформовані екзогенною ДНК. Використовуваний в даному описі термін «ген», або «кодуюча послідовність» належить до 5 UA 102367 C2 5 10 15 20 25 30 35 40 45 50 55 60 молекули ДНК, з якої транскрибується молекула РНК. Молекула ДНК містить область (CR-), що кодує мРНК, яка, в свою чергу, кодує білковий продукт, або РНК, яка функціонує як молекула, що придушує ген, або структурні молекули РНК, такі як тРНК, рРНК, snРНК або інші молекули РНК. Використовуваний термін в даному описі «експресія» належить до комбінації внутрішньоклітинних процесів, що включають транскрипцію і трансляцію, за допомогою яких молекула ДНК, така як ген, продукує поліпептид або молекулу РНК. Прикладом кодуючої послідовності є ген дигідродипіколінатсинтази Corynebacterium (DHDPS; Bonnassie з співавторами, 1990; Richaud з співавторами, 1986), придатний для отримування кукурудзяних зерен з підвищеним вмістом лізину. Кукурудзяна рослина, трансформована таким чином, що вона містить і експресує ген DHDPS Corynebacterium, або будь-який інший ген, експресія якого приводить до підвищення вмісту лізину в тканині зерна, в контексті даного винаходу також називається кукурудзяною рослиною з підвищеним вмістом лізину. Кодуюча область гену DHDPS Corynebacterium згідно з даним винаходом додатково модифікується для підвищення експресії в рослинних клітинах, переважно в клітинах однодольних рослин, і найбільш переважно в клітинах кукурудзи. Модифікована кодуюча область гену DHDPS Corynebacterium, використовувана в контексті даного винаходу, відповідає послідовності SEQ ID NO:5, що означається в даному описі як CORgl. dapA. nno. Використовуваний в даному описі термін «промотор» належить до області послідовності ДНК, яка ініціює транскрипцію РНК з ДНК. Промотори розташовані з 5'-кінця смислового ланцюга транскрибованої молекули ДНК і містять області, що служать, як сайти зв’язування для РНК-полімерази, а також області, залучені у взаємодії з іншими чинниками, які індукують транскрипцію РНК. Більш конкретно, базальні промотори рослин містять канонічні області, асоційовані з ініціацією транскрипції, такі як CAAT- і ТАТА-бокси. У контексті даного винаходу переважні молекули промоторів і 5'-нетрансльовані молекули індукують транскрипцію в клітинах насіння або тканинах на більш високому рівні, ніж в інших клітинах або тканинах рослини. Молекули промоторів можуть бути вибрані з кількості молекул, відомих в даній області, наприклад, з промоторів, описаних в патенті США № 6437217 (кукурудзяний RS81-промотор), в патенті США № 5641876 (промотор рисового актину), в патенті США № 6426446 (кукурудзяний RS324-промотор), в патенті США № 6429362 (кукурудзяний PR-1-промотор), в патенті США № 6232526 (кукурудзяний A3-промотор), в патенті США № 6177611 (конститутивний кукурудзяний промотор), в патентах США № 5322938, 5352605, 5359142 і 5530196 (35S-промотор), в патенті США № 6433252 (кукурудзяний L3-промотор олеозину, P-Zm. L3), в патенті США № 6429357 (промотор рисового актину 2, а також інтрон рисового актину 2), в патенті США № 5837848 (специфічний для коріння промотор), в патенті США № 6294714 (світлоіндукований промотор), в патенті США № 6140078 (солеіндукований промотор), в патенті США № 6252138 (патогеніндукований промотор), в патенті США № 6175060 (промотор, індукований недоліком фосфору), в патенті США № 6635806 (промотор гамма-коїксину, P-Cl. Gcx) і в патентній заявці США № 09/757089 (промотор хлоропластової альдолази кукурудзи); всі вищеперелічені публікації включені до даного опису за допомогою посилання. Крім цього можуть бути сконструйовані химерні промоторні молекули, що містять цис-елементи, отримані з гетерологічних джерел, які додатково поліпшують експресійний профіль промотору, використовуваного в контексті даного винаходу. Наприклад, було показано, що елементи промоторів рослинних вірусів підвищують ефективність промоторних елементів рослин, функціонально пов'язаних вірусними промоторами (патент США № 6660911, повністю включений до даного опису за допомогою посилання). Крім промотору, ДНК-конструкції, використовувані для трансформації рослин, звичайно також містять інші регуляторні елементи, такі як, без обмеження, 3'-нетрансльовані області (які містять сайти поліаденілювання), транзитні або сигнальні пептиди для направлення білка в певні клітинні органели рослин, зокрема, в хлоропласти, лейкопласти і інші пластидні органели. Фахівцям в даній галузі відомі різні інтрони, енхансери, транзитні пептиди, що націлюють сигнальні послідовності. 5'- і 3'-нетрансльовані області (UTR), корисні для конструювання ефективних векторів для експресії в рослинах, подібних розкритим, наприклад, в публікації патентної заявки США № 2003/01403641 (включеної до даного опису за допомогою посилання). 5'-UTR, що функціонує як трансляційна лідерна послідовність, являє собою генетичний ДНКелемент, локалізований між промоторною і кодуючою послідовностями гену. Трансляційна лідерна послідовність присутня в повністю процесованій мРНК з 5'-кінця від послідовності початку трансляції. Трансляційна лідерна послідовність може впливати на процесинг первинного транскрипту в мРНК, або на стабільність мРНК або ефективність трансляції. Приклади трансляційних лідерних послідовностей включають лідерні послідовності генів білків теплового шоку кукурудзи і петунії (патент США № 5362865), лідерні послідовності генів 6 UA 102367 C2 5 10 15 20 25 30 35 40 45 50 55 60 оболонкових білків вірусів рослин, лідерні послідовності рослинних генів рибулозабісфосфаткарбоксилази/оксигенази (Rubisco) і т. д. (Turner і Foster, 1995). Прикладами лідерних молекул, в природному контексті асоційованих з промоторними молекулами, описаними в контексті даного винаходу, можуть служити лідерна послідовність B32 кукурудзи (L-Zm. B32), лідерна послідовність рисового актину 1 (L-Os. Act1) і лідерна послідовність гаммакоїксину (L-Cl. Gcx). 3'-нетрансльована область (3' UTR) або 3'-область поліаденілювання являє собою молекулу ДНК, пов'язану зі структурною полінуклеотидною молекулою і локалізовану з 3'-кінця від останньої, і містить полінуклеотиди, які містять сигнал поліаденілювання і інші регуляторні сигнали, що впливають на транскрипцію, процесинг мРНК або експресію генів. Функція сигналу поліаденілювання в рослинах складається із запуску поліаденілювання, тобто додавання поліаденілатнуклеотиду до 3'-кінця попередника мРНК. Послідовність поліаденілювання може бути отримана з природного гену, з множини рослинних генів або з Т-ДНК генів агробактерій. Прикладами 3'-UTR можуть служити 3'-область нопалинсинтази (nos 3'; Fraley з співавторами, 1983), 3'-UTR-послідовності гену hsp17 пшениці (T-Ta. Hsp17) і T-Ps. RbcS2:E9 (мала субодиниця рибулозабісфосфаткарбоксилази/оксигенази (rubisco) гороху), 3'-UTRпослідовності, розкриті в патенті № WO0011200A2, а також інші відомі в даній галузі техніки 3'UTR-послідовності, які можуть бути протестовані і використані в комбінації з кодуючими областями DHDPS або AK і в даному описі означаються як Т-3'UTR. Під терміном «транзитні сигнали» загалом розуміють пептидні молекули, які, будучи пов'язаними з представляючим інтерес білком, направляють останній в ту або іншу тканину, клітину, внутрішньоклітинний компартмент або клітинну органелу. Приклади подібних пептидів включають без обмеження хлоропласт-транзитні пептиди, сигнали ядерної локалізації і вакуолярні сигнали. У контексті даного винаходу особливо застосовними є пластид-транзитні пептиди, що направляють експресію ферменту DHDPS в пластиди клітин насіння. Хлоропласттранзитний пептид (CTP) може бути отриманий у вигляді химерної конструкції, в якій CTP злитий з N-кінцем білка, призначеного для транспорту в хлоропласти. Множина білків, локалізованих в хлоропластах, експресується ядерними генами у вигляді попередників і далі прямує в хлоропласти за рахунок CTP, який видаляється в ході процесів, пов'язаних з імпортом білка в хлоропласти. Приклади хлоропластних білків включають малу субодиницю (RbcS2) рибулозу-1,5-бісфосфат-карбоксилази, фередоксин, фередоксин-оксидоредуктазу, білки 1 і 2 світлозбирального комплексу і тиоредоксин F. Було продемонстровано, як in vivo, так і in vitro, що нехлоропластні білки можуть бути перенаправлені в хлоропласти шляхом використання химерних СТР-злитих білків, а також що наявність CTP є достатньою умовою для направлення білка в хлоропласти. Було показано, що вбудовування відповідного хлоропласт-транзитного пептиду, такого як CTP генів EPSPS Arabidopsis thaliana (Klee з співавторами, 1987), і Petunia hybridа (della-Cioppa з співавторами, 1986), приводить до направлення гетерологічних білків в хлоропласти трансгенних рослин. CTP гену DapA Zea mays (SEQ ID NO:6, в даному описі, що означається як TS-Zm. DapA) згідно з даним винаходом може бути застосований для напрямку пов'язаного з ним поліпептиду DHDPS в пластиди насіння. Фахівцям в даній галузі буде зрозуміло, що можуть бути отримані різні химерні конструкції, в яких використовується функціональність специфічного CTP для імпорту гетерологічного поліпептиду DHDPS в пластиди рослинної клітини. Опис використання хлоропласт-транзитних пептидів приведений в патенті США № 5188642, повністю включеного до даного опису за допомогою посилання. В таблиці 1 приведені комбінації промоторів і інших регуляторних елементів, функціонально пов'язаних з молекулами, які придушують ген LKR, і з кодуючими молекулами DHDPS, а також відповідні ефекти відносно вмісту лізину в рослинах і зернах кукурудзи. Фахівцеві в даній галузі буде ясно, що екзогенні молекули ДНК зі схожими властивостями можуть бути замінені молекулами, перерахованими в таблиці 1, і протестовані за допомогою способів, описаних в даному описі, а також інших схожих способах, придатних для визначення рівня лізину в рослинних тканинах, зернах, а також отриманій з них продукції. ДНК-конструкції за даним винаходом можуть необов'язково містити селектовану або маркерну молекулу, яка кількісно оцінюється. Особливо важливими є селектовані маркерні молекули, які також додають рослинам цінну агрономічну властивість, наприклад, стійкість до гербіцидів. Гербіциди, стійкість до яких була продемонстрована для трансгенних рослин, і до яких можуть бути застосовані способи за даним винаходом, включають, без обмеження, гліфосат, глюфосинат, сульфонілсечовини, імідазолінони, бромоксиніл, далапон, циклогександіон, інгібітори протопорфіриногеноксидази, а також ізоксафлютолові гербіциди. Полінуклеотидні молекули, які кодують білки, залучені до контролю стійкості до гербіцидів, добре відомі в даній галузі і включають, без обмеження, полінуклеотидні молекули, що кодують 7 UA 102367 C2 5 10 15 20 25 30 35 40 45 50 55 60 5-енолпірувілшикімат-3-фосфатсинтазу (EPSPS, патенти США № 5627061; 5633435; 6040497; Padgette з співавторами, 1996; Penaloza-Vazquez з співавторами, 1995; і aroA (патент США № 5094945), стійкість до гліфосату; бромоксинілнітрилази (Bxn), стійкість до бромоксинілу (патент США № 4810648); фітоендесатурази (crtI, Misawa з співавторами, 1993 і 1994); стійкість до норфлуразону, ацетогідроксиацидсинтазу (AHAS, aka ALS, Sathasiivan з співавторами, 1990); і ген bar для стійкості до глюфосинату і біалафосу (DeBlock з співавторами, 1987). Стійкість до гербіцидів є бажаним фенотипом для сільськогосподарських культур. ДНКконструкції за даним винаходом переважно містять експресійну касету, наявність якої надає трансформованій кукурудзяній рослині стійкість до гліфосату. Також, експресійна касета дозволяє проводити селекцію трансформованих клітин кукурудзи в гліфосатовмісному середовищі. Експресійна касета переважно містить промотор, що активує транскрипцію гена, який кодує стійку до гліфосату EPSPS, у всіх клітинах трансформованої рослини. Зокрема, експресійна касета, яка обумовлює наявність стійкості до гліфосату, містить промотор і інтрон рисового актину 1 (P-Os. Act1, I-Os. Act1, патент США № 5641876, включений до даного опису за допомогою посилання), функціонально пов'язані з полінуклеотидом, що кодує хлоропласттранзитний пептид, а також з полінуклеотидом, що кодує стійку до дії гліфосату EPSPS штаму CP4 Agrobacterium (в даному описі означається як aroA:CP4 EPSPS, див. патент США № 5633435, включений до даного опису за допомогою посилання). N-фосфонометилгліцин, також відомий як гліфосат, є добре відомим гербіцидом з фітотоксичною активністю, направленою ® проти широкого спектра рослинних видів. Гліфосат є активним інгредієнтом гербіциду Roundup (Monsanto Co., St. Louis, MO), безпечного гербіциду з коротким часом напівжиття в навколишньому середовищі. При нанесенні на поверхню рослини гліфосат системно проникає в рослину. Токсичність гліфосату по відношенню до рослин зумовлена блокадою шикіматного метаболічного шляху, пов'язаного з синтезом попередника ароматичних амінокислот. Більш конкретно, гліфосат порушує конверсію фосфоенолпірувату і 3-фосфошикімової кислоти в 5енолпірувілшикімат-3-фосфат шляхом блокування ферменту 5-енолпірувілшикімат-3фосфатсинтази (що далі означається як EPSP-синтаза або EPSPS). У контексті даного винаходу, термін «гліфосат» належить до будь-якої гербіцидно-ефективної форми Nфосфонометилгліцину (включаючи будь-які його солі), а також до будь-яких інших речовин, що перетворюються в гліфосат-аніон in planta. Гліфосат і N-фосфонометилгліцин, а також його солі використовуються як компоненти культуральних середовищ для селекції бактерійних і рослинних клітин, стійких до гліфосату, або використовуються для біохімічних аналізів стійкості до цих речовин відповідних ферментів in vitro. Приклади комерційних препаратів гліфосату включають без обмеження гербіцидні препарати, які продаються Monsanto Company під ® ® ® ® торгівельними марками ROUNDUP , ROUNDUP ULTRA, ROUNDUP ULTRAMAX, ROUNDUP ® ® ® ® ® CT, ROUNDUP EXTRA, ROUNDUP BIACTIVE, ROUNDUP BIOFORCE, RODEO , POLARIS , ® ® SPARK і ACCORD , що містять гліфосат у вигляді ізопропіламонієвої солі; гербіцидні ® препарати, які продаються Monsanto Company під торгівельними марками ROUNDUP DRY і ® RIVAL , що містять гліфосат у вигляді амонієвої солі; препарат, який продається Monsanto ® Company під торгівельною маркою ROUNDUP GEOFORCE, що містить гліфосат у вигляді натрієвої солі; препарат, який продається Monsanto Company під торгівельною маркою ® ROUNDUP WEATHERMAX, що містить гліфосат у вигляді калієвої солі; а також препарат, який ® продається Zeneca Limited під торгівельною маркою TOUCHDOWN , що містить гліфосат у вигляді триметилсульфонієвої солі. Гліфосатовмісні гербіцидні препарати можуть бути безпечно нанесені з повітря на стійкі до гліфосату сільськогосподарські культури для контролю чисельності бур'янів на полях в кількостях від 8 до 64 унцій/акра. У експериментальних умовах гліфосат може бути нанесений на стійкі до дії гліфосату сільськогосподарські культури в кількостях від 4 до більш ніж 128 унцій/акра без значного пошкодження сільськогосподарських культур. Вибір кількості гліфосатовмісного препарату, що наноситься, і складає біологічно ефективну дозу, знаходиться в галузі компетенції фахівця-агронома. Для ознайомлення з отримуванням трансгенних рослин, стійких до гербіцидів, див. публікації патентних заявок США №№ 2003/0106096A1 і 2002/0112260A1, а також патенти США №№ 5034322, 6107549 і 6376754, кожний(а) з яких включений(а) до даного опису за допомогою посилання. Даний винахід належить до кукурудзяних рослин з підвищеним вмістом лізину і опційно стійких до гліфосату. У ході трансформації екзогенна ДНК може бути вбудована в геном рослини довільно, тобто не сайт-специфічно. У деяких випадках може бути корисним направити вбудовування екзогенної ДНК в певний сайт, тобто провести сайт-специфічну інтеграцію, наприклад, замінити існуючу послідовність гену або геномної області. У деяких інших випадках може бути корисним направити вбудовування екзогенної ДНК в певний сайт, про який відомо, що в ньому має місце експресія гена(ів). Існує декілька сайт-специфічних рекомбінаційних систем, що функціонують в 8 UA 102367 C2 5 10 15 20 25 30 35 40 45 50 55 60 рослинах, в тому числі Cre/lox-система, розкрита в патенті США № 4959317 і FLP/FRT-системі, розкрита в патенті США № 5527695; вищенаведені в даному абзаці публікації включені до даного опису за допомогою посилання. Корисні маркерні селектовані гени являють собою гени, що забезпечують стійкість до антибіотиків, таких як ампіцилін, канаміцин (nptII), гігроміцин В (aph IV) і гентаміцин (aac3 і aacС4), або селектовані маркерні гени, описані в патентах США №№ 5550318, 5633435, 5780708 і 6118047, кожний з яких включений до даного опису за допомогою посилання. Також можуть бути використані скринінгові маркери, які надають можливість візуально ідентифікувати трансформанти, наприклад, гени, експресуючі кольорові або флуоресцентні білки, такі як люцифераза або зелений флуоресцентний білок (GFP), або ген, експресуючий бетаглюкоуронідазу, або генів uidA (GUS), для яких відомі різні хромогенні субстрати. Придушування генів ДНК-конструкція за даним винаходом забезпечує експресію антисмислової молекули РНК, яка придушує ендогенний ген LKR (лізинкетоглутаратредуктаза (LKR, біфункціональний рослинний фермент, також відомий як цукропіндегідрогеназа SDH) в трансгенній рослинній клітині. Подібна ДНК-конструкція містить промотор, активний в тканині, в якій планується придушити дію цільового ферменту, а також транскрибовану послідовність ДНК-елементу, комплементарну полінуклеотидній послідовності гену LKR, експресію якого планується придушити. Цільовий генний елемент, послідовність якого «копіюється» для використання як транскрибованої ДНК в конструкції для придушення експресії гену, може являти собою промоторний елемент, інтронний елемент, екзонний елемент, або 5'- або 3'-UTR-елемент. Хоч мінімальна довжина фрагмента ДНК, «копійованого» з послідовності гену, експресію якого планується придушити, становить приблизно 21 або 23 нуклеотиду, переважними є більш довгі сегменти, наприклад, аж до повнорозмірної послідовності цільового гену. ДНК-елемент може містити декілька фрагментів гену, наприклад, нуклеотиди, комплементарні безперервним або окремим генним елементам, відповідним UTR, екзону і інтрону. Подібні конструкції можуть також містити відповідно до необхідності інші регуляторні елементи, ДНК-фрагменти, що кодують транзитні пептиди, сигнальні пептиди, селекційні і скринінгові маркери. Для отримання антисмисловим чином орієнтованої РНК-завіси довжина комплементарного ДНК-елемента умовно складає не більше половини довжини антисмисловим чином орієнтованого ДНКелемента, часто не більше однієї третини від довжини антисмисловим чином орієнтованого ДНК-елемента, наприклад, не більше однієї чверті від довжини вищезазначеного антисмисловим чином орієнтованого ДНК-елемента. Загальна довжина комбінації ДНКелементів може варіюватися. Наприклад, довжина антисмисловим чином орієнтованого ДНКелемента може складати від 500 до 5000 нуклеотидів, а довжина комплементарного ДНКелемента може складати від 50 до 500 нуклеотидів. Дизайн антисмислової транскрипційної одиниці може бути проведений таким чином, щоб придушувати експресію множини генів або членів генного сімейства, при цьому ДНК містить два або більше антисмисловим чином орієнтованих елементів, отриманих з різних генів, наприклад, комплементарних щонайменше частини 5'-антисмислового елемента. Придушення експресії включає в себе добре відомі способи придушення транскрипції гену або накопичення відповідною цьому гену мРНК, перешкоджаючих трансляції транскрипту в білок. Посттрансляційне придушення експресії опосередковується транскрипцією інтегрованої рекомбінантної ДНК з подальшим формуванням дволанцюгової РНК (дцРНК), що є гомологічною гену, експресію якого планується придушити. Утворення дцРНК звичайно відбувається в результаті транскрипції інтегрованого інвертованого повтору цільового гену і є звичайною подією при використанні способів придушення експресії, відомих як антисмислове придушення, косупресія або РНК-інтерференція (RNAi). Транскрипційне придушення може бути опосередковане транскрибованою дцРНК, гомологічною промоторною послідовністю ДНК ефект, який називається транс-супресією промотору. Більш конкретно, посттрансляційне придушення експресії гену, що досягається шляхом вбудовування рекомбінантної ДНК-конструкції, яка містить антисмисловим чином орієнтований ДНК-фрагмент, для регуляції експресії гена в клітинах рослин, розкрито в патентах США № 5107065 (Shewmaker з співавторами) і 5759829 (Shewmaker з співавторами). Трансгенні рослини, трансформовані за допомогою подібних антисмисловим чином орієнтованих ДНКконструкцій, призначених для придушення транскрипції генів, можуть містити інвертований повтор, виникаючий в результаті вбудовування ДНК-конструкції в рослини за допомогою агробактеріальної трансформації, згідно з розкритим Redenbaugh з співавторами (1992). Вставки інвертованих повторів можуть містити всю Т-ДНК-конструкцію або її частину, наприклад, інвертований повтор повної транскрипційної одиниці, або інвертований повтор 9 UA 102367 C2 5 10 15 20 25 30 35 40 45 50 55 60 послідовності термінації транскрипції. Скринінг на вбудовану ДНК, що містить інвертовані повтори, може підвищити ефективність ідентифікації трансформаційних подій, ефективних для «сайленсингу» генів, при цьому трансформуюча конструкція може являти собою просту антисмислову ДНК-конструкцію, яка повинна бути вбудована в декількох копіях, або ж комплексну інвертовану ДНК-конструкцію (наприклад, RNAi-конструкцію), яка може бути вбудована в єдиній копії. ДНК-конструкція, що складає предмет даного винаходу, може містити декілька інвертованих повторів, вбудованих в регуляторний елемент, наприклад, в інтрон. Інвертований повтор містить молекулу ДНК, гомологічну і комплементарну ендогенному гену LKR рослинної клітини-хазяя. У контексті даного винаходу, молекула ДНК (SEQ ID NO:9, dsSDH bfx, також означається як LKR dsRNA bfx) є гомологічною до фрагменту ДНК, що кодує LKR/SDH Zea mays (SEQ ID NO:2) і використовується для придушення експресії нативних кукурудзяних генів LKR/SDH. Позначення bfx належить до послідовності молекули дцРНК, використовуваної для придушення експресії, яка була спеціально відібрана за допомогою аналізу цільової молекули після виключення родинних цільових послідовностей, знайдених в інших організмах. У деяких описаних в даному описі конструкціях LKR фрагмент dsRNA bfx вбудований в інтрон (Zm. LKR dsRNA bfx, вбудований в I-Zm. DnaK, SEQ ID NO:10). Додаткові полінуклеотиди, гомологічні і комплементарні послідовності гену LKR Zea mays, також можуть бути вибрані для використання в контексті даного винаходу. Функціонування гену також може бути порушене за допомогою придушення експресії, що досягається за рахунок інсерційних мутацій, які створюються мобільними генетичними елементами. Наприклад, багато які дводольні рослини можуть бути легко трансформовані агробактеріальною Т-ДНК, в результаті чого може бути швидко отримана велика кількість трансформованих рослин. Крім цього, існують лінії деяких видів, таких як Zea mays, що містять активні мобільні генетичні елементи, які можуть бути ефективно використані для генерування великої кількості інсерційних мутацій, хоч інші види позбавлені подібних можливостей. Мутантні рослини з порушеною експресією цікавлячого поліпептиду, отримані за допомогою агробактеріальної трансформації або мутагенезу, опосередкованого мобільними генетичними елементами, можуть бути ідентифіковані за допомогою полінуклеотидів, які складають предмет даного винаходу. Наприклад, численна популяція мутантних рослин може бути проскринована за допомогою полінуклеотидів, що кодують цікавлячий поліпептид, для ідентифікації мутантних рослин з вбудованим геном, що кодує цікавлячий поліпептид. Білкові молекули Білки, що складають предмет даного винаходу, являють собою повнорозмірні білки, або щонайменше фрагменти повнорозмірних білків, достатні для реалізації релевантної біологічної активності повнорозмірного білка, наприклад, підвищення вмісту лізину в трансгенному кукурудзяному зерні. Термін «білок» також включає в себе молекули, що складаються з одного або більше поліпептидних ланцюгів. Таким чином, корисний білок в контексті даного винаходу може являти собою продукт повнорозмірного гену, або один або більше функціональних фрагментів природного білку, які надають бажану агрономічну властивість згідно з даним винаходом, тобто підвищений вміст лізину. Гомологи білків, що складають предмет даного винаходу, можуть бути ідентифіковані за допомогою зіставлення амінокислотних послідовностей білків DHDPS (SEQ ID NO:7) або аспартаткінази (SEQ ID NO:8) з амінокислотними послідовностями білків тих же або інших рослинних або бактерійних організмів, наприклад, вручну, або за допомогою відомих пошукових алгоритмів, заснованих на пошуку гомології, таких як BLAST, FASTA і алгоритм Smith-Waterman. У іншому аспекті даний винахід належить до послідовностей, що кодують функціональні гомологічні білки, відмінні від білків DHDPS і аспартаткінази згідно з даним винаходом на одну або більше амінокислоту; виникнення подібних білків є результатом добре відомих консервативних амінокислотних замін, наприклад, заміна на валін є консервативною для аланіну, а треонін - консервативною заміною для серину. При експресії подібних гомологічних білків в трансгенній рослині вони будуть надавати ефекти, практично еквівалентні таким, що надаються білками DHDPS або аспартаткінази. Консервативні заміни для тієї або іншої амінокислоти в нативній послідовності білка можуть бути вибрані з інших членів класу, до якого належить дана природна амінокислота. Типові амінокислоти, що належать до різних класів, включають в себе без обмеження (1) кислі (негативно заряджені) амінокислоти, такі як аспарагінова кислота і глутамінова кислота; (2) основні (позитивно заряджені) амінокислоти, такі як аргінін, гістидин і лізин; (3) нейтральні полярні амінокислоти, такі як гліцин, серин, треонін, цистеїн, тирозин, аспарагін і глутамін; і (4) нейтральні неполярні (гідрофобні) амінокислоти, такі як аланін, лейцин, ізолейцин, валін, пролін, фенілаланін, триптофан і метіонін. Консервативні заміни для тієї або іншої амінокислоти 10 UA 102367 C2 5 10 15 20 25 30 35 40 45 50 55 60 в нативній послідовності білка можуть бути вибрані з інших членів групи, до якої належить дана природна амінокислота. Наприклад, група амінокислот з аліфатичними бічними ланцюгами включає в себе гліцин, аланін, валін, лейцин і ізолейцин; група амінокислот з бічними гідроксилвмісними аліфатичними ланцюгами включає в себе серин і треонін; група амінокислот з амідовмісними бічними ланцюгами включає в себе аспарагін і глутамін; група амінокислот з ароматичними бічними ланцюгами включає в себе фенілаланін, тирозин і триптофан; група амінокислот з основними бічними ланцюгами включає в себе лізин, аргінін і гістидин; і група амінокислот з сірковмісними бічними ланцюгами включає в себе цистеїн і метіонін. Групами, в яких відбуваються природні консервативні амінокислотні заміни, є амінокислотні пари валінлейцин, валін-ізолейцин, фенілаланін-тирозин, лізин-аргінін, аланін-валін, аспарагінова кислота - глутамінова кислота, і аспарагін-глутамін. У іншому аспекті даний винахід належить до білків, відмінних від описаних білкових послідовностей DHDPS і аспартаткінази на одну або більше амінокислоту; виникнення подібних білків є результатом інсерції або делеції однієї або більше амінокислоти в нативній послідовності. При експресії подібних гомологічних білків в трансгенній рослині вони будуть надавати ефекти, практично еквівалентним чином, наприклад, забезпечувати підвищення вмісту лізину в кукурудзяному зерні. Білки, що складають предмет даного винаходу і які є варіантами білків, до яких належить даний винахід, в загальному випадку демонструють значну ступінь ідентичності з останніми, таку як щонайменше 50% і більше, наприклад, щонайменше 60% або 70% ідентичності з білками DHDPS і аспартаткінази. Корисні білки також включають в себе білки, що демонструють ще більшу ідентичність з амінокислотами білкових сегментів білків DHDPS і аспартаткінази, наприклад, 80%, 90%, 95%, 98% або до 99% ідентичності. Способи трансформації і трансформовані рослини У контексті даного винаходу термін «кукурудза» належить до Zea mays, також відомому як кукурудза, і включає в себе всі види рослин, які можуть бути схрещені з кукурудзою, включаючи дикі сорти кукурудзи. У практичних втіленнях даного винаходу способи і композиції для трансформації рослин за допомогою вбудовування екзогенної ДНК в геном рослини можуть включати в себе будь-які з добре відомих і продемонстрованих способів. Переважними способами трансформації рослин є бомбардування мікрочастинками, описане в патентах США № 5015580, 5550318, 5538880, 6160208, 6399861 і 6403865, а також трансформація за допомогою агробактерій, описана в патентах США № 5635055, 5824877, 5591616, 5981840, 6384301 і патентної публікації 20030196219; всі вищенаведені в даному абзаці публікації повністю включені до даного опису за допомогою посилання. Трансформація за допомогою мікрочастинок належить до способу доставки ДНК, іммобілізованої на поверхні мікрочастинок, якими за допомогою різних способів бомбардується цільова тканина. Трансформація за допомогою агробактерій виробляється за допомогою генно-інженерних грунтових бактерій, що належать до роду Agrobacterium. Деякі види Agrobacterium забезпечують перенесення специфічної плазмідної ДНК, відомої як Т-ДНК, яка може бути піддана генно-інженерним маніпуляціям для доставляння будь-якого бажаного фрагмента ДНК у багато які види рослин. Використовуваний в даному описі термін «трансгенний» організм належить до організму, геном якого містить екзогенну ДНК або ізольовану ДНК. Трансгенним організмом може бути рослина, тваринна, комаха, гриб, бактерія або вірус. Використовуваний в даному описі термін «трансгенна рослина» належить до стабільно трансформованої рослини або отриманої від неї потомства будь-якого покоління, в якому ДНК рослини і її потомства містять екзогенну ДНК. Крім цього, трансгенну рослину можуть містити також послідовності, нативні для трансформованої рослини, в які була вбудована екзогенна ДНК для зміни рівня і патерну експресії гену. Використовуваний в даному описі термін «стабільно» трансформована рослина належить до рослини, в якій екзогенна ДНК успадковується. Екзогенна ДНК може успадковуватися як фрагмент ДНК, присутній в клітині рослини, при цьому не вбудований в геном хазяя. Стабільно трансформована рослина переважно містить екзогенну ДНК, вбудовану в хромосомну ДНК в ядрі, мітохондріях або хлоропластах, більш переважно - вбудовану в ядерну хромосомну ДНК. Використовуваний в даному описі термін «трансгенна рослина R 0» належить до рослини, яка була трансформована екзогенною ДНК або отримана з клітини, калюса або кластера клітин, які були трансформовані екзогенною ДНК. Використовуваний в даному описі термін «потомство» належить до подальшого покоління, включаючи насіння і отримані з них рослини, які виникли від конкретної батьківської рослини або декількох батьківських рослин; це потомство може бути високовиродженим або значною мірою гомозиготним, в залежності від сорту. Потомство трансгенної рослини даного винаходу можливо, наприклад, запліднене 11 UA 102367 C2 5 10 15 20 25 30 35 40 45 50 55 60 шляхом самоопилення, схрещено з іншою трансгенною рослиною, схрещено з нетрансгенною рослиною і/або піддано зворотному схрещуванню з батьківською рослиною. Насіння рослин, що складає предмет даного винаходу, може бути зібране з родючих трансгенних рослин і може бути використане для виведення поколінь потомства рослин, що складають предмет даного винаходу, включаючи гібридні лінії рослин, які містять в геномі екзогенну ДНК-конструкцію згідно з приведеним в таблиці 1 даного опису, яка забезпечує трансгенні продукти перевагою, що полягає в підвищенні вмісту лізину в кукурудзяному зерні і опціональному підвищенні стійкості до гліфосатних гербіцидів. Трансгенний «продукт» отримують шляхом трансформації рослинної клітини екзогенної ДНК-конструкції, подальшої регенерації рослини, отриманої в ході специфічного вбудовування екзогенної ДНК в рослинний геном і селекції рослини, що характеризується наявністю трансгенної ДНК. Кожний продукт є унікальним індивідом, кожний з яких відрізняється від інших продуктів специфічною локалізацією геномної вставки екзогенної ДНК-конструкції, що складає предмет даного винаходу. Детектування трансгенною ДНК здійснюється за допомогою будь-якої кількості способів виявлення ДНК, відомих фахівцям в даній області техніки, які можуть бути виконані з використанням рослинних тканин, насіння і оброблених трансгенних продуктів, які містять трансгенну ДНК. Звичайно трансформують декілька рослинних клітин, кожна з яких потенційно являє собою продукт інтеграції, відмінний від інших продуктів, що приводить до отримання популяції рослин, з якої далі вибирається та або інша конкретна рослина. В результаті того, що локус інтеграції в більшості випадків є стабільним протягом репродуктивних циклів рослин, що повторюються, термін «продукт» належить до первинного трансформанту R 0 і його потомства, які містять екзогенну ДНК, вбудовану в специфічний і унікальний сайт в геномі, тобто трансгенну ДНК. Потомство, отримане в результаті статевого ауткросингу, самозапліднювання або вторинного зворотного схрещування, при проведенні яких хоч би одна з використовуваних в схрещуванні рослин, є нащадком трансформанту R0 будь-якого покоління і містить ту ж трансгенну ДНК. У ході процесингу насіння і зерен, які складають предмет даного винаходу, для отримання корму і продуктів харчування, нормальні структури тканин рослини знищуються, клітини рослини руйнуються і нативна ДНК і екзогенні експресійні касети рослин можуть не залишитися повністю інтактними, особливо при оброблюванні з використанням високих температур, розчинників, що екстрагують і ним подібних. Незважаючи на це, унікальні процесовані продукти, отримані в ході практичних втілень даного винаходу, можуть бути легко ідентифіковані як такі, що містять нативну ДНК і фрагменти екзогенних касет, призначених для експресії в рослинах. Під «фрагментами» маються на увазі сегменти ДНК, які ізолюються або ампліфікуються довжиною щонайменше 20 пар основ. Такі фрагменти можуть бути секвеновані або картовані за допомогою рестрикційного аналізу для підтвердження ідентичності їх послідовностей нативної ДНК або екзогенними експресійними касетами вихідного матеріалу. Приклади Нижченаведені приклади приведені для демонстрації прикладів конкретних переважних втілень даного винаходу. Фахівцям в даній галузі техніки буде зрозуміло, що технології, розкриті в нижченаведених прикладах, являють собою підходи, які, за даними авторів даного винаходу, добре працюють в контексті практичних реалізацій даного винаходу, і таким чином можуть служити прикладами переважних практичних втілень даного винаходу. Проте, в світлі даного розкриття, фахівці в даній галузі техніки повинні розуміти, що розкриті специфічні втілення можуть бути піддані множині змін, і проте приводити до схожих результатів, без виходу за рамки суті і об'єму даного винаходу. Приклад 1 ДНК-конструкції і трансформування рослин ДНК-конструкції, що складають предмет даного винаходу, містять регуляторні молекули, функціонально пов'язані з цікавлячою ДНК. Зокрема, ДНК-конструкція містить промотор, інтрон, 5'-лідерну послідовність, кодуючу або некодуючу молекулу і 3'-нетрансльовану область. ДНКконструкція, що складає предмет даного винаходу, була отримана з використанням різних молекулярних способів і інструментів, наприклад, описаних Sambrook з співавторами; значною мірою схожі конструкції можуть бути отримані за допомогою варіантів даних способів, відомих фахівцям в області маніпуляцій з ДНК. Такі методи, як ПЦР-ампліфікація, клонування і субклонування використовувалися для функціонального зв’язування промоторів, лідерних послідовностей, інтронів, транзитних пептидів, кодуючих молекул, некодуючих молекул і 3'нетрансльованих областей, приведених в таблиці 1 в функціональних конфігураціях і далі проілюстрованих на фіг. 1-6. ДНК-конструкції були клоновані в один або більше плазмідних «скелетів», придатних для використання в агробактеріальній трансформації клітин кукурудзи, 12 UA 102367 C2 5 10 15 20 25 30 35 40 45 50 55 60 хоч можуть бути вибрані і інші плазмідні «скелети», наприклад, висококопійні плазміди, для ізоляції ДНК-фрагментів і трансформації з використанням мікрочастинок, або плазміди, сумісні з іншими бактерійними хазяями. Для тестування комбінацій стратегій експресії нечутливого до дії лізин-опосередкованої завіси зворотного зв'язку поліпептиду DHDPS (SEQ ID NO:7) і репресії катаболізму лізину за допомогою придушення експресії ендогенного гену LKR/SDH кукурудзи (SEQ ID NO:2), або експресії гену аспартат-кінази (SEQ ID NO:8), були отримані 35 конструкцій, приведених в таблиці 1. Ці стратегії включають в себе, наприклад: 1) експресію дцРНК гену LKR в зародку; 2) експресію поліпептиду DHDPS і дцРНК гену LKR в зародку; 3) експресію поліпептиду DHDPS і дцРНК гену LKR в ендоспермі; 4) експресію поліпептиду DHDPS в ендоспермі і дцРНК гену LKR в зародку; 5) експресію поліпептиду DHDPS і аспартат-кінази, а також експресію дцРНК гену LKR. Отримання трансгенних рослин Тканини кукурудзи (інбредні лінії LH198 або LH244), що містять незрілі зародки або калюс, для трансформації ізолювали, після чого ДНК-конструкцію, яка складає предмет даного винаходу, вбудовували в геном кукурудзи за допомогою агробактерій (див., наприклад, патенти США № 5635055, 5824877, 5591616, 5981840 і 6384301, кожний з яких повністю включений до даного опису за допомогою посилання). Агробактерії, трансформовані ДНК-конструкціями, перерахованими в таблиці 1, інокулювали на тканину кукурудзи. Після експонування тканини трансформованими бактеріями, для розмноження і селекції трансформованих клітин кукурудзи використовували стандартні протоколи трансформації з використанням середовища з гліфосатним гербіцидом. Після селекції клітин, що вижили в присутності гліфосатного гербіциду, трансгенні рослини R0 регенеровували і вирощували до зрілості. Насіння збирали з плодоносних трансгенних рослин після самозапліднювання або ауткросингу з іншою кукурудзяною рослиною. Трансгенні вставки визначали за допомогою молекулярного аналізу. Для ідентифікації трансгенних продуктів, що містять одиночну інтактну вставку ДНК-конструкції, ® або для аналізу зиготності, використовували ПЦР, Саузерн-блот-аналіз, Taqman -аналіз або інші методики, відомі фахівцям в галузі молекулярної біології. Отримане зернове потомство висаджували, і отримані з них рослини вирощували для подальшого аналізу вмісту лізину в тканинах і насінні потомства. Приклад 2 Визначення вмісту лізину в тканинах і насінні кукурудзи Різні способи є корисними для визначення вмісту лізину в тканинах рослин, їх зернах і процесованих продуктах, наприклад, в даному винаході для аналізу вмісту вільного лізину (в частинах на мільйон, ppm) в зернах кукурудзи, відповідних різним трансгенним продуктам, ідентифікованим як трансформовані за допомогою ДНК-конструкції, описаній в таблиці 1, використовувалася рідинна хромотографія-мас-спектрофотометрія/мас-спектрофотометрія (LCMS-MS); результати даного аналізу приведені в таблицях 1 і 2. Проби індивідуальних зрілих кукурудзяних зерен, відповідних кожному з трансгенних продуктів, спочатку зважували, потім перемелювали в тонкий гомогенний порошок, який далі екстраговували за допомогою розчинника, що складається з метанолу, води і мурашиної кислоти. У випадку «bulked»-зерна використовували приблизно 30 мг подрібненого порошку. Для виділення лізину з екстракту застосовували рідинну хроматографію і мас-спектрометрію з моніторингом множинних реакцій (MRM). Після відділення кількість лізину обчислювали за площею мас-спектрометричного піку відносно калібрувальної кривої, яка була отримана з використанням дейтерованого d4-лізину як внутрішнього стандарту (IS). У іншому випадку визначення вмісту лізину в зернах кукурудзи було засноване на визначенні вільних амінокислот за допомогою високоефективної рідинної хроматографії (HPLC). Окремі зерна або зернові пули кожного з трансгенних продуктів перемелювали в тонкий гомогенний порошок згідно раніше описаному, і в цьому випадку для аналізу використовували приблизно 30 мг даного порошку. Екстракцію амінокислот проводили за допомогою 5% трихлороцтової кислоти, визначення амінокислот проводили за допомогою передколончастої первинної амінної дериватизації з використанням о-фталальдегіду (OPA). При проведенні хроматографії із поверненою фазою, поділення відбувається завдяки гідрофобності R-групам кожної з амінокислот. Для стабілізації флуорофору додається тіол, такий як 2-меркаптоетанол (SHCH2CH2OH) або 3-меркаптопропіонова кислота (SHCH2CH2COOH). Рівні вільного лізину (в частинах на мільйон, ppm) в трансгенних продуктах, отриманих в ході трансформації кожної з конструкцій, були усереднені і узагальнені в таблиці 1. Також приведені покоління (F1, F2, F3 і F4) проаналізованих зерен. Результати вимірювання вмісту вільного лізину в зернах покоління R1 трансгенних кукурудзяних продуктів, що містять pMON80378, приведені в таблиці 2. Вміст лізину був несподівано високим в проаналізованих 13 UA 102367 C2 5 10 продуктах 5 і 6. Вміст лізину може бути далі відрегульований шляхом схрещування кукурудзяних продуктів, які містять різні комбінації касет, призначених для експресії в рослинах і описаних в таблиці 1, таких, що гібридне зерно, отримане від схрещування батьківських рослин, які містять одну або більше експресійних касет, буде містити бажану кількість лізину. Інбредна батьківська рослина, що містить, наприклад, експресійні касети CordapA і lysC E. coli (дикий тип або Ec. lysC. mut, SEQ ID NO:11, молекула ДНК, що кодує білок, модифікований в чотирьох позиціях - S2A, C58G, T352I і A401G), може бути схрещена з іншою батьківською рослиною, яка містить експресійну касету дцРНК LKR, в результаті чого зернове потомство, що отримується містить більшу кількість лізину, ніж сумарний вміст лізину в обох батьківських рослинах. Батьківська рослина, яка містить інші комбінації, такі як, наприклад, комбінація CordapA і дцРНК LKR, може бути схрещена з іншою батьківською рослиною, яка містить LysC, або ж батьківська рослина, яка містить LysC і дцРНК LKR, може бути схрещена з іншою батьківською рослиною, яка містить CordapA, для отримання несподівано високого вмісту лізину в гібридних зернах. 14 UA 102367 C2 15 UA 102367 C2 16 UA 102367 C2 5 10 15 20 25 30 35 40 45 Вміст вільного лізину в зернах трансгенних продуктів, які містять pMON93092, також був виміряний за допомогою тандемної мас-спектрофотометрії (LC-MS-MS) в подрібнених і екстрагованих пробах, приблизно по 20 зерен в кожній лунці. Вміст вільного лізину в частинах на мільйон (ppm) в зернах покоління R2 з ряду трансформованих продуктів продемонстрований на фіг. 7 і 8. Як показано для продуктів, що містять pMON93092, вміст вільного лізину становив приблизно 3500-5500 ppm, тоді як для продуктів, що містять pMON93093, вміст вільного лізину становив приблизно 1700-6500 ppm. У більшості випадків були проведені незалежні вимірювання в двох або більше лунках, відповідних одному і тому ж продукту; на фігурах також продемонстровані значення стандартного відхилення. Для порівняння, вміст лізину в кукурудзі дикого типу складає від 50 до 100 ppm. Розкриті вище і процитовані в формулі винаходи плазміди pMON93092 і pMON93093 були депоновані Monsanto Technology LLC за Будапештською угодою з Американською колекцією типових культур (ATCC), 10801 University Boulevard, Manassas, Va. 20110. Номери ATCC - PTA6733 і PTA-6734, відповідно. Депозит буде зберігатися в депозитарії протягом 30 років, або протягом 5 років після останнього запиту, або протягом терміну ефективного використання патенту - в залежності від того, який термін виявиться довшим, і буде при необхідності замінений протягом цього періоду. Всі публікації, патенти і патентні заявки включені до даного опису за допомогою посилання в тій же мірі, як якби кожна окрема публікація або патентна заявка була спеціально і індивідуально включена за допомогою посилання. Посилання Посилання, приведені нижче, включені до даного опису за допомогою посилання в такій мірі, щоб доповнити, пояснити, розкрити основу або описати методологію, технологію і/або композиції, використані в контексті даного винаходу. Патент США № 4810648; патент США № 4957748; патент США № 4959317; патент США № 5015580; патент США № 5034322; патент США № 5094945; патент США № 5100679; патент США № 5107065; патент США № 5188642; патент США № 5219596; патент США № 5258300; патент США № 5288300; патент США № 5322938; патент США № 5352605; патент США № 5359142; патент США № 5362865; патент США № 5367110; патент США № 5527695; патент США № 5530196; патент США № 5538880; патент США № 5550318; патент США № 5591616; патент США № 5627061; патент США № 5633435; патент США № 5635055; патент США № 5641876; патент США № 5759829; патент США № 5773691; патент США № 5780708; патент США № 5824877; патент США № 5837848; патент США № 5936069; патент США № 5981840; патент США № 6005076; патент США № 6040497; патент США № 6107549; патент США № 6118047; патент США № 6140078; патент США № 6146669; патент США № 6156227; патент США № 6160208; патент США № 6175060; патент США № 6177611; патент США № 6232526; патент США № 6252138; патент США № 6294714; патент США № 6376754; патент США № 6384301; патент США № 6399861; патент США № 6403865; патент США № 6426446; патент США № 6429357; патент США № 6429362; патент США № 6433252; патент США № 6437217; патент США № 6459019; патент США № 6635806; патент США № 6660911. Патентна заявка США № 2002/0112260; патентна заявка США № 2003/0056242; патентна заявка США № 2003/0106096; патентна заявка США № 2003/0140364; патентна заявка США № 2003/0196219; патентна заявка США № 2005/0132437; патентна заявка США № 2005/0193444. 17 UA 102367 C2 5 10 15 Патентна заявка США з серійним № 09/757089. Bonnassie з співавторами, Nucleic Acids Res., 18: 6421, 1990. DeBlock з співавторами, EMBO J., 6: 2513-2519, 1987. della-Cioppa з співавторами, Proc. Natl. Acad. Sci. USA, 83: 6873-6877, 1986. Fraley з співавторами, Proc. Natl. Acad. Sci. USA, 80: 4803-4807, 1983. Klee з співавторами, Mol. Gen. Genet, 210: 437-442, 1987. Misawa з співавторами, Plant J., 4: 833-840, 1993. Misawa з співавторами, Plant J., 6: 481-489, 1994. Padgette з співавторами, In: Herbicide Resistant Crops, Lewis Publishers, 53-85, 1996. PCT заявка WO 0011200A2. Penaloza-Vazquez з співавторами, Plant Cell Reports, 14: 482-487, 1995. TM Redenbaugh з співавторами, In: Safety Assessment of Genetically Engineered Flavr Savr Tomato, CRC Press, Inc., 1992. Richaud з співавторами, J. Bacteriol., 166: 297-300, 1986. Sambrook з співавторами, In: Molecular cloning, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, 2000. Sathasiivan з співавторами, Nucl. Acids Res., 18: 2188-2193, 1990. Turner and Foster, Molec. Biotech., 3: 225, 1995. 18 UA 102367 C2 19 UA 102367 C2 20 UA 102367 C2 21 UA 102367 C2 22 UA 102367 C2 23 UA 102367 C2 24 UA 102367 C2 25 UA 102367 C2 26 UA 102367 C2 27 UA 102367 C2 28
ДивитисяДодаткова інформація
Назва патенту англійськоюTransgenic corn seed with increased lysine content
Автори англійськоюMalvar Thomas, Huang Shihshieh, Liuti Michael X.
Назва патенту російськоюСемя трансгенной кукурузы с повышенным содержанием лизина
Автори російськоюМалвар Томас М., Хуанг Шихших, Люти Михель Х.
МПК / Мітки
МПК: C12N 15/82
Мітки: лізину, вмістом, трансгенної, насінина, кукурудзи, підвищеним
Код посилання
<a href="https://ua.patents.su/42-102367-nasinina-transgenno-kukurudzi-z-pidvishhenim-vmistom-lizinu.html" target="_blank" rel="follow" title="База патентів України">Насінина трансгенної кукурудзи з підвищеним вмістом лізину</a>
Спосіб створення ліній та гібридів кукурудзи з підвищеним вмістом білка та олії і поліпшеною якістю білка

Номер патенту: 21548
Опубліковано: 15.03.2007
Автори: Ніколенко Ірина Анатоліївна, Тимчук Віктор Михайлович, Тимчук Сергій Михайлович
МПК: A01H 1/04
Мітки: кукурудзи, олії, гібридів, якістю, ліній, спосіб, вмістом, білка, підвищеним, поліпшеною, створення
Формула / Реферат:
Спосіб створення ліній та гібридів кукурудзи з підвищеним вмістом білка та олії і поліпшеною якістю білка, що включає використання неалельних взаємодій між генами структури ендосперму, який відрізняється тим, що для підвищення вмісту білка та олії в зерні і незамінних амінокислот в білку використовують біохімічний ефект генної рекомбінації o2sh2.
Спосіб визначення трансгенної лінії ga21 кукурудзи за допомогою полімеразної ланцюгової реакції

Номер патенту: 61602
Опубліковано: 25.07.2011
Автори: Кучук Микола Вікторович, Банникова Марія Олександрівна, Моргун Богдан Володимирович, Сатарова Тетяна Миколаївна, Борисова Вікторія Вікторівна
МПК: C12N 15/00
Мітки: спосіб, ланцюгової, полімеразної, лінії, кукурудзи, реакції, трансгенної, допомогою, визначення
Формула / Реферат:
Спосіб детекції мутантного гена 5-енолпірувіл шікімат-3-фосфат синтази кукурудзи (Zea mays L.) у генетично модифікованій рослині методом полімеразної ланцюгової реакції, для здійснення якої проводять термальну денатурацію рослинної ДНК; циклічну ампліфікацію, де кожен цикл включає денатурацію ДНК, ренатурацію рослинної ДНК з олігонуклеотидними праймерами, синтез фрагментів цільових генів; синтез фрагментів цільових генів, для проведення...
Днк-конструкт, що забезпечує підвищений вміст лізину в рослинах кукурудзи

Номер патенту: 93654
Опубліковано: 10.03.2011
Автори: Малвар Томас, Моллой Кетлін П., Люті Міхель Ганс, Келлі Ребекка А., Войлес Дейл А., Дізіган Марк Ентоні
МПК: A01H 5/00, C12N 15/82
Мітки: рослинах, вміст, днк-конструкт, кукурудзи, забезпечує, підвищений, лізину
Формула / Реферат:
1. ДНК-конструкт, який містить: (а) перший сегмент ДНК, який містить промотор кукурудзяного глобуліну 1;(b) другий сегмент ДНК, який містить інтрон гена рисового актину 1;(с) третій сегмент ДНК, який містить молекулу ДНК, що кодує кукурудзяний хлоропластний транзитний пептид DHDPS; (d) четвертий сегмент ДНК, який містить молекулу ДНК, що кодує DHDPS Corynebacterium;...
Спосіб селекційного відбору багаторічних злакових трав з підвищеним вмістом білка і створення сортів за цією ознакою
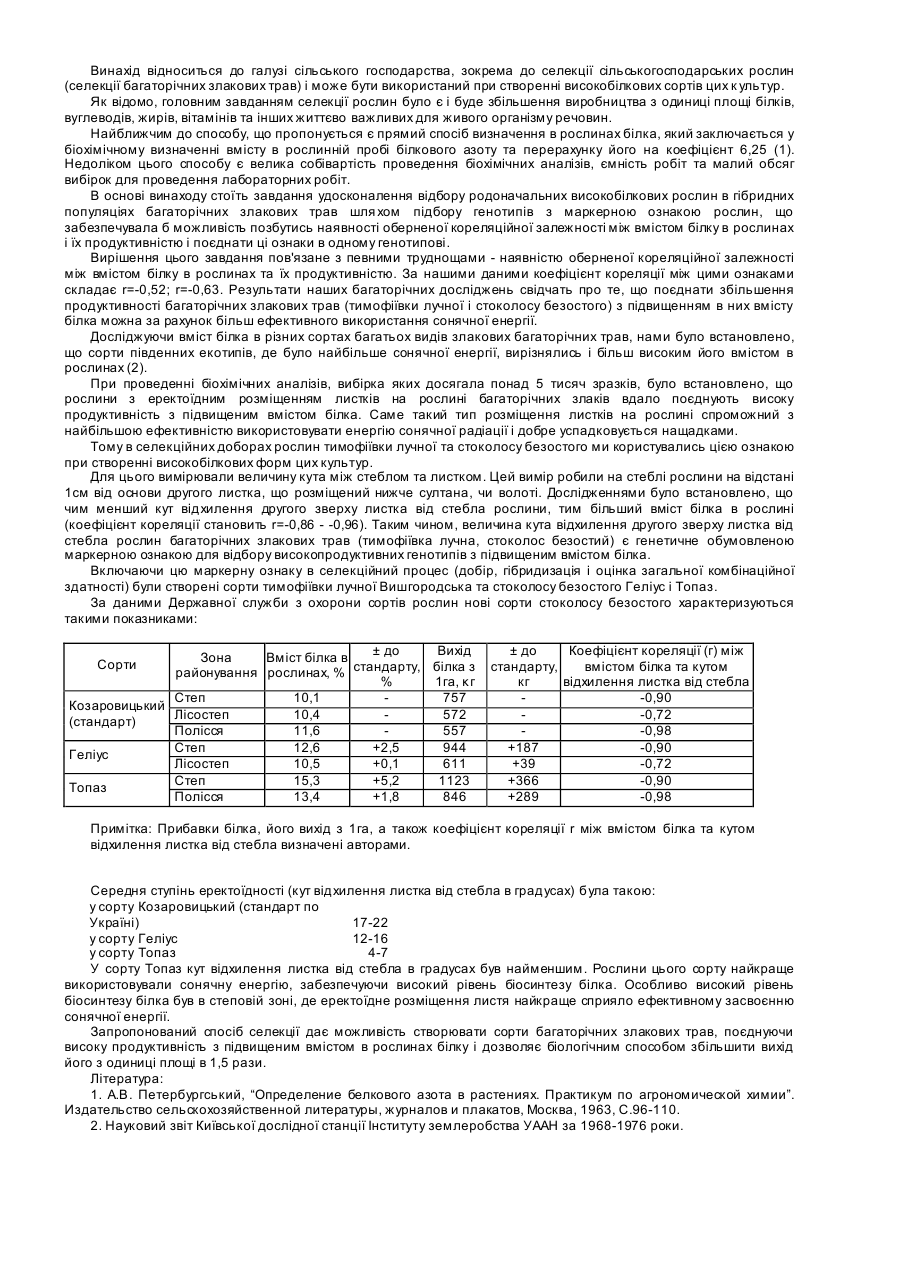
Номер патенту: 70159
Опубліковано: 15.09.2004
Автори: Сердюк Микола Арсентійович, Сердюк Олександр Миколайович
МПК: A01H 1/04
Мітки: трав, вмістом, створення, багаторічних, злакових, цією, ознакою, спосіб, відбору, селекційного, білка, підвищеним, сортів
Формула / Реферат:
Спосіб селекційного відбору багаторічних злакових трав з підвищеним вмістом білка і створення сортів за цією ознакою, згідно з яким проводять добори вихідного матеріалу різних груп генотипів, включають їх в гібридизацію, проводять оцінку за рівнем загальної комбінаційної здатності за даним показником, який відрізняється тим, що добори на високобілковість рослин проводять за генетично спадковою маркерною ознакою - еректоїдністю розміщення...
Спосіб формування смуг з підвищеним вмістом органічних матеріалів для деревних і трав’яних культур

Номер патенту: 70317
Опубліковано: 11.06.2012
Автор: Канцер Анатолій Миколайович
МПК: A01C 5/00
Мітки: трав'яних, формування, спосіб, вмістом, деревних, органічних, матеріалів, смуг, культур, підвищеним
Формула / Реферат:
Спосіб формування смуг з підвищеним вмістом органічних матеріалів для деревних і трав'яних культур, що включає посадку деревних і трав'яних культур з попереднім внесенням органічних матеріалів, який відрізняється тим, що перед посадкою деревних і трав'яних культур і внесенням органічних матеріалів уздовж них в ґрунті формують траншею, в яку послідовно засипають шар органічних матеріалів і видаленого ґрунту, після чого виконують посадку...
Попередній патент: Фрикційний планетарний редуктор
Наступний патент: Специфічні у відношенні b-амілоїду 1-42 моноклональні антитіла, які мають терапевтичні властивості
Випадковий патент: Спосіб класифікації зразків генофонду пшениці за кількісними характеристиками