N-ацетилглюкозамін-6-фосфатдеацетилаза людини, яка здатна взаємодіяти з субодиницею 1 і/або субодиницею 2 каспази-8, виділена послідовність днк, що її кодує, спосіб одержання даного білка (варіанти), антитіло,







































Формула / Реферат
1. N-ацетилглюкозамін-6-фосфатдеацетилаза людини, яка здатна взаємодіяти з субодиницею 1 і/або субодиницею 2 каспази-8, що має амінокислотну послідовність, приведену на Фіг. 2, або її варіанти, одержані, як розкрито в описі, здатні взаємодіяти з субодиницею 1 і/або субодиницею 2 каспази-8.
2. N-ацетилглюкозамін-6-фосфатдеацетилаза за п. 1, що розщеплюється in vitro каспазою-8.
3. N-ацетилглюкозамін-6-фосфатдеацетилаза за п. 1, що розщеплюється in vivo каспазою-8.
4. Виділена послідовність ДНК, що кодує N-ацетилглюкозамін-6-фосфатдеацетилазу людини за будь-яким з пп. 1-3.
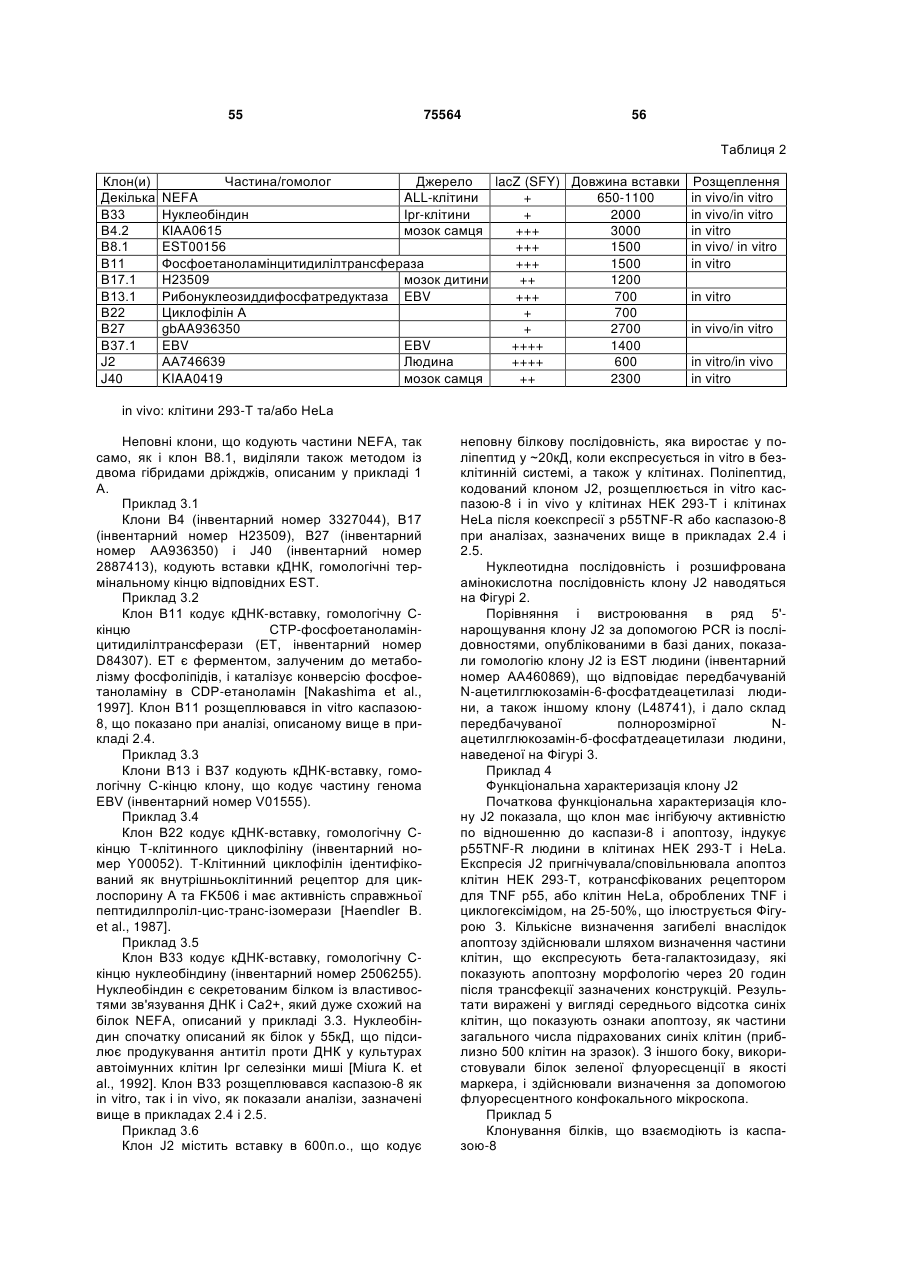
5. Виділена послідовність ДНК за п. 4, що має нуклеотидну послідовність, представлену на фіг. 3.
6. Виділена послідовність ДНК, здатна гібридизуватись з послідовністю ДНК за п. 4 в помірно суворих умовах, розкритих в описі.
7. Вектор, що містить послідовність ДНК за п. 5 або 6 і регуляторні послідовності, зокрема промотор CMV людини.
8. Еукаріотична або прокаріотична клітина-хазяїн, що містить вектор за п. 7.
9. Спосіб одержання N-ацетилглюкозамін-6-фосфатдеацетилази людини за п. 1, що включає вирощування еукаріотичної клітини-хазяїна за п. 8 в умовах, що забезпечують продукування вказаного білка і його посттрансляційні модифікації, і виділення вказаного білка.
10. Спосіб одержання N-ацетилглюкозамін-6-фосфатдеацетилази людини за п. 1, що включає вирощування прокаріотичної клітини-хазяїна за п. 8 в умовах, що забезпечують продукування вказаного білка, і виділення вказаного білка.
11. Спосіб за п. 9, згідно з яким еукаріотичною клітиною є клітина ссавця, комахи або дріжджів.
12. Спосіб за п. 11, згідно з яким клітиною ссавця є клітина HeLa або 293 Т НЕК.
13. Антитіло, направлене проти епітопу N-ацетилглюкозамін-6-фосфатдеацетилази людини за п. 1.
14. Набір для імуноаналізу, призначений для виявлення N-ацетилглюкозамін-6-фосфатдеацетилази людини, що включає як реагент антитіло за п. 13.
15. Застосування N-ацетилглюкозамін-6-фосфатдеацетилази людини за п. 1 для модуляції активності каспази-8.
16. Застосування антитіла за п. 13 для модуляції активності каспази-8.
17. Застосування N-ацетилглюкозамін-6-фосфатдеацетилази людини за п. 1 для модуляції активності, опосередкованої TNF-рецептором або Fas.
18. Застосування антитіла за п. 13 для модуляції активності, опосередкованої TNF-рецептором або Fas.
19. Застосування N-ацетилглюкозамін-6-фосфатдеацетилази людини за п. 1 для модуляції апоптозу.
20. Застосування антитіла за п. 13 для модуляції апоптозу.
21. Застосування N-ацетилглюкозамін-6-фосфатдеацетилази людини за п. 1 як лікарського засобу для терапії захворювань, пов'язаних з необхідністю модуляції активності каспази-8.
22. Застосування антитіла за п. 13 як лікарського засобу для терапії захворювань, пов'язаних з необхідністю модуляції активності каспази-8.
23. Застосування за будь-яким з пп. 21-22, де вказаним захворюванням є розсіяний склероз з первинною олігодендрогліопатією, аутоімунний увеоретиніт, діабет, вовчак, аутоімунний міокардит І, опосередкований HCV хронічний гепатит, хронічний гастрит, наприклад, гастрит типу А, змішана хвороба сполучної тканини (MCTD), хвороба Крона або неспецифічний виразковий коліт.
Текст
1. N-ацетилглюкозамін-6-фосфатдеацетилаза засобу для терапії захворювань, пов'язаних з нелюдини, яка здатна взаємодіяти з субодиницею 1 обхідністю модуляції активності каспази-8. і/або субодиницею 2 каспази-8, що має амінокис22. Застосування антитіла за п. 13 як лікарського лотну послідовність, приведену на Фіг. 2, або її засобу для терапії захворювань, пов'язаних з неваріанти, одержані, як розкрито в описі, здатні взаобхідністю модуляції активності каспази-8. ємодіяти з субодиницею 1 і/або субодиницею 2 23. Застосування за будь-яким з пп. 21-22, де вкакаспази-8. заним захворюванням є розсіяний склероз з пер2. N-ацетилглюкозамін-6-фосфатдеацетилаза за винною олігодендрогліопатією, аутоімунний увеоп. 1, що розщеплюється in vitro каспазою-8. ретиніт, діабет, вовчак, аутоімунний міокардит І, 3. N-ацетилглюкозамін-6-фосфатдеацетилаза за опосередкований HCV хронічний гепатит, хронічп. 1, що розщеплюється in vivo каспазою-8. ний гастрит, наприклад, гастрит типу А, змішана 5 75564 6 хвороба сполучної тканини (MCTD), хвороба Крона або неспецифічний виразковий коліт. Даний винахід відноситься в основному до області цистеїн-протеаз. Конкретніше, винахід відноситься до білків, що взаємодіють із каспазою-8 (МАСН) та/або таких, що модулюють її функції на шляху загибелі клітин (або при апоптозі), опосередкованій CD95 (Fas/Apo-) або CD 120а (рецептор для TNF р55). Зокрема, даний винахід відноситься до білків, що взаємодіють із каспазою-8/МАСН безпосередньо або непрямо. Винахід також відноситься до одержання і застосування білків, що взаємодіють із каспазою-8/МАСН. Передумови створення винаходу Фактор некрозу пухлини (TNF-альфа) та лімфотоксин (TNF-бета) (далі позначення TNF відноситься як до TNF-альфа, так і до TNF-бета) є багатофункціональними прозапальними цитокінами, утвореними, головним чином, мононуклеарними фагоцитами, і справляють декілька впливів на клітини (Wallach, D, (1986), у Interferon 7 (Ion Gresser, ed.), pp. 83-122, Academic Press, London; і Beutler and Cerami (1987). Як TNF-альфа, так і TNF-бета починають свій вплив через зв'язування зі специфічними рецепторами на поверхні клітини. Деякі такі впливи, мабуть, сприятливі для організму: вони руйнують, наприклад, пухлинні клітини або клітини, заражені вірусом, і підвищують антибактеріальну активність гранулоцитів. У цьому відношенні TNF вносить вклад у захист організму від пухлин і інфекційних агентів і вносить вклад у відновлення після ушкоджень. Таким чином, TNF можна використовувати в якості протипухлинного засобу, який при застосуванні зв'язується зі своїми рецепторами на поверхні пухлинних клітин і за допомогою цього ініціює події, що призводять до загибелі пухлинних клітин. TNF також можна використовувати в якості протиінфекційного засобу. Однак, як TNF-альфа, так і TNF-бета мають також руйнуючу дією. Очевидно, що надпродукування TNF-альфа може відігравати роль головного патогену при деяких захворюваннях. Наприклад, відомо, що вплив TNF-альфа, у першу чергу, на судинну мережу є основною причиною симптомів септичного шоку [Tracey et al., 1986]. При деяких захворюваннях TNF може викликати надмірну втрату ваги (кахексія), пригнічуючи активність адипоцитів і викликаючи анорексію, і тому TNF-альфа названо кахектином. Він також описаний як медіатор ушкодження тканин при ревматоїдних захворюваннях [Beutler and Cerami, 1987] і як головний медіатор ушкодження, що спостерігається при реакціях "трансплантат проти хазяїна" [Piquet et al., 1987]. Крім того, відомо, що TNF залучається до процесу запалення і до багатьох інших хворобливих станів. Два різних рецептора TNF-R p55 (CD120а) і р75 (CD120b), які експресуються незалежно і які специфічно зв'язуються як з TNF-альфа, так і з TNF-бета, ініціюють та/або опосередковують зазначену вище біологічну дію TNF. Ці два рецепто ри мають різні за структурою внутрішньоклітинні домени, причому передбачається, що вони передають сигнал по-різному [див. Hohmann et al., 1989; Engelmann et al., 1990; Brockhaus et al., 1990; Loetscher et al, 1990; Schall et al., 1990; Nophar et al., 1990; Smith et al 1990; і Holler et al., 1990]. Однак клітинні механізми, наприклад, різні білки та інші фактори, які залучаються до внутрішньоклітинної передачі сигналів CD120а і CD120b, ще не вияснені. Мова йде про внутрішньоклітинну передачу сигналу, яка відбувається, як правило, після зв'язування ліганду, тобто TNF (альфа або бета), із рецептором, що є відповідальним за початок каскаду реакцій, кінцевим результатом якого є спостережувана реакція клітини на TNF. Що стосується вищевказаної дії TNF, яка викликає загибель клітини, у більшості клітин, вивчених дотепер, то ця дія ініціюється, головним чином, CD120а. Антитіла проти позаклітинного домену (домену зв'язування ліганду) CD120а самі можуть ініціювати дію, яка викликає загибель клітин [див. ЕР 412486], яка корелює з ефективністю поперечного зшивання рецепторів антитілами і яка можливо є першою стадією при генерації процесу внутрішньоклітинної передачі сигналу. Далі, дослідження мутацій [Brakebusch et al., 1992; Tartaglia et al., 1993] показали, що біологічна функція CD120а залежить від цілісності його внутрішньоклітинного домену, і, відповідно, передбачається, що ініціація внутрішньоклітинної передачі сигналу, що призводить до дії TNF, яка викликає загибель клітини, відбувається в результаті асоціації двох або більшого числа внутрішньоклітинних доменів CD120а. Крім того, TNF (альфа і бета) зустрічається у вигляді гомотримеру, і як такий, можливо, індукує внутрішньоклітинну передачу сигналу через CD120а за рахунок своєї здатності зв'язуватися з молекулами рецептора і утворювати поперечні зшиття, тобто викликати агрегацію рецепторів. Ще одним членом суперсімейства рецепторів TNF/NGF є рецептор FAS/APO1 (CD95), який також називають FAS-антигеном, білок поверхні клітини, який експресується у різних тканинах і є гомологічним ряду рецепторів клітинної поверхні, включаючи TNF-R і NGF-R. CD95 опосередковує загибель клітини у формі апоптозу [Itoh et al., 1991] і представляється таким, що є негативним селектором аутореактивних Т-клітин, тобто під час дозрівання Т-клітин CD95 опосередковує загибель Тклітин, що розпізнають аутоантигени, внаслідок апоптозу. Також виявлено, що мутації в гені CD95 (Ірr) викликають порушення лімфопроліферації у мишей, що має подібність до автоімунної хвороби людини системним червоним вовчаком (SLE) [Watanabe-Fukunaga et al., 1992]. Виявляється, що ліганд CD95 являє собою молекулу, зв'язану з поверхнею клітини, яка міститься, серед інших, у кілерних Т-клітинах (або цитотоксичних Тлімфоцитах CTL), і, отже, коли такі CTL контактують із клітинами, що містять CD95, вони здатні 7 75564 8 індукувати апоптозну загибель клітин, що містять описується конструювання і характеризація різних CD95. Далі, отримані моноклональні антитіла, мутантних форм CD120а, що мають мутації в його специфічні для CD95, причому такі моноклональні позаклітинних, трансмембранних і внутрішньокліантитіла здатні індукувати апоптозну загибель тинних доменах. При такому способі області в меклітин, що містять CD95, включаючи мишачі клітижах вищевказаних доменів CD120а ідентифіковані ни, трансформовані кДНК, яка кодує CD95 людини як суттєві для функціонування рецептора, тобто [Itoh et al., 1991]. для зв'язування ліганду (TNF) і наступної трансдуНезважаючи на те, що цитотоксична дія лімкції сигналу і внутрішньоклітинної передачі сигнафоцитів опосередковується через взаємодію пролу, кінцевим результатом яких стає спостережувадукованого лімфоцитами ліганду з рецептором на дія TNF на клітини. Далі також описується ряд поверхні клітини CD95, який зустрічається повсюпідходів для виділення та ідентифікації білків, пепди і який має здатність ініціювати загибель клітитидів або інших факторів, здатних зв'язуватися з ни, також виявлено, що різні інші здорові клітини, різними ділянками в зазначених вище доменах крім Т-лімфоцитів, експресують CD95 на своїй CD120а, і ці білки, пептиди та інші фактори можуть поверхні і можуть бути вбиті через ініціювання залучатися до процесів регуляції або модуляції цього рецептора. Передбачається, що некерована активності TNF-R. Ряд підходів до виділення і клоіндукція такого кілінгу вносить вклад в ушкодження нування послідовностей ДНК, що кодують такі білтканин при деяких захворюваннях, наприклад, ки і пептиди; до конструювання векторів, що ексруйнування клітин печінки при гострому гепатиті. пресують, для продукування таких білків і Відповідно, пошук способів стримування цитотокпептидів; і до одержання антитіл або їхніх фрагмесичної активності CD95 може мати лікувальне знантів, що взаємодіють із CD120а або вищевказаничення. ми білками і пептидами, що зв'язуються з різними З іншого боку, оскільки також виявлено, що ділянками CD120а, також зазначені в ЕРО 368925. деякі злоякісні клітини і ВІЛ-інфіковані клітини місОднак, в ЕР 568925 не описуються ні реальні білки тять CD95 на своїй поверхні, антитіла проти CD95 і пептиди, що зв'язуються з внутрішньоклітинними або ліганд для CD95 можна використовувати для доменами TNF-R (наприклад, CD95), ні двогібридініціації опосередковуваної CD95 цитотоксичної дії ний спосіб із дріжджами для виділення та ідентив цих клітинах і за допомогою цього створити спофікації таких білків або пептидів, що зв'язуються з сіб боротьби зі злоякісними клітинами або ВІЛвнутрішньоклітинними доменами TNF-R. Подібним інфікованими клітинами [див. Itoh et al., 1991]. Почином, в ЕР 568925 не описуються білки або пепшук ще й інших способів посилення цитотоксичної тиди, здатні зв'язуватися з внутрішньоклітинним активності тому також може мати лікувальний подоменом CD95. тенціал. Таким чином, коли потрібно інгібувати дію TNF Уже тривалий час існує потреба у способі моабо ліганду CD95, варто знизити кількісно активдуляції клітинної реакції на TNF (альфа або бета) і ність TNF-R або CD95 на поверхні клітини, хоча ліганд CD95. Наприклад, у випадках зазначеної кількісне підвищення активності TNF-R або CD95 вище патології, коли TNF або ліганд CD95 експреможе знадобитися, коли домагаються посиленої суються в надлишку, бажано пригнічувати дію, яка дії TNF або CD95. З цією метою секвеновані та викликається TNF або CD95, що убиває клітини, у проаналізовані промотори як для CD120а, так і той час як в інших випадках, наприклад, при задля CD120b, і знайдений ряд ключових мотивів стосуваннях для загоєння ран, бажано підсилювапослідовностей, специфічних для різних факторів ти дію TNF, або, у випадку CD95, підсилювати регуляції транскрипції, і, по суті, експресію цих опосередковувану CD95 дію в пухлинних клітинах TNF-R можна регулювати на рівні їхніх промоторів, або у ВІЛ-інфікованих клітинах. тобто інгібувати транскрипцію від промоторів для Заявники почали спроби [див. заявки на єврозниження числа рецепторів і підсилювати транскпейський патент №№ ЕР 186833, ЕР 308378, ЕР рипцію від промоторів для збільшення числа ре398327 і ЕР 412486] регулювати згубну дію TNF за цепторів [ЕР 606869 і WO 9531206]. допомогою інгібування зв'язування TNF із його Незважаючи на те, що відомо, що рецептори рецепторами з використанням антитіл проти TNF для фактора некрозу пухлини (TNF) і структурно або з використанням розчинних рецепторів для родинний рецептор CD95 ініціюють у клітинах (пісTNF (що є, по суті, розчинними позаклітинними ля стимуляції лігандами, що продуковані лейкоцидоменами рецепторів), для того, щоб створити тами) деструктивну активність, яка приводить до конкуренцію зв'язуванню TNF із TNF-R, зв'язаним їхньої власної загибелі, механізми такого ініціюіз поверхнею клітини. Крім того, на тій підставі, що вання поки мало зрозумілі. Дослідження з мутацізв'язування TNF із його рецепторами необхідно ями показують, що в передачі цитотоксичного сигдля індукованої TNF дії в клітині, заявники почали налу в CD95 і CD120а беруть участь різні ділянки в спроби [див. наприклад ЕР 568925] модулювати межах їхніх внутрішньоклітинних доменів дію TNF за допомогою модуляції активності TNF[Brakebusch et al., 1992; Tartaglia et al., 1993; Itoh R. and Nagata, 1993]. Ці області ("домени загибелі") Наприклад, в ЕР 568925 описується спосіб мають схожі послідовності. "Домени загибелі" як модуляції сигнальної трансдукції та/або розчепCD95, так і CD120а мають тенденцію до самоагрелення в TNF-R, у результаті чого пептиди або інші гації. їхня самоагрегація, мабуть, промотує агрегамолекули можуть взаємодіяти або із самим рецепцію рецепторів, необхідну для ініціації передачі тором або з ефекторними білками, що взаємодісигналу [див. Song et al., 1994; Wallach et al., 1994; ють із рецептором, причому в такий спосіб модуBoldin et al., 1995], і при високому рівні експресії люється нормальна функція TNF-R. В ЕР 568925 рецепторів може призвести до ініціації ліганд 9 75564 10 незалежної передачі сигналу [Boldin et al., 1995]. 1996; Chinnaiyan et al., 1996], що вказує на те, що Деякі з цитотоксичних дій лімфоцитів опосеця N-кінцева область передає сигнал дії обох рередковуються через взаємодію продукованого цепторів, яка викликає загибель клітини через білімфоцитами ліганду з CD95 - рецептором поверлок-білкові взаємодії. хні клітини, що зустрічається повсюди, здатним Виконані останнім часом дослідження охопініціювати загибель клітини [див. також Nagata and люють групу цитоплазматичних тіол-протеаз, споGoldstein, 1995]; і така загибель клітини, опосередріднених по будові до протеази Caenorhabditis кована мононуклеарними фагоцитами, включає elegans CED3 і до ферменту інтерлейкіну-1-бета участь пари ліганд і рецептор - TNF і його рецепссавців, що конвертує, (ICE), на початку різних тора CD120а, спорідненої по будові до пари CD95 фізіологічних процесів загибелі клітин [огляд у і його ліганду [див. також Vandenabeele et al., роботах Kumar, 1995, і Henkart, 1996]. Є також ряд 1995]. Як і у випадку інших впливів, що індукуютьпоказань, що протеаза(и) цього сімейства можуть ся рецепторами, індукція загибелі клітин рецептобрати участь в індукованій CD95 і TNF-R цитотокрами TNF і CD95 відбувається через ряд білоксичності в клітині. Виявлено, що специфічні пептибілкових взаємодій, що ведуть від зв'язування редні інгібітори протеаз і білки, що блокують їхні фуцептора з лігандом до можливої активації ефектонкції, кодовані двома вірусами - білок вірусу рних функцій ферментів, які при дослідженнях коров'ячої віспи crmA і білок бакуловірусу р35, зароз'яснили неферментні білок-білкові взаємодії, які безпечують захист клітин від такої цитотоксичності ініціюють передачу сигналів загибелі клітин: зв'яв клітині [Enari et al., 1995; Los et al., 1995; Tewari зування тримерного TNF або молекул ліганду et al., 1995; Xue et al., 1995; Beidler et al., 1995]. CD95 із рецепторами, підсумкові взаємодії їхніх Швидке розчеплення деяких специфічних клітинвнутрішньоклітинних доменів [Brakebush et al., них білків, явно опосередковане протеазою(ами) 1992; Tartaglia et al., 1993; Itoh and Nagata, 1993], сімейства CED3/ICE, можна показати в клітинах посилені схильністю мотивів доменів загибелі до незабаром після стимуляції CD95 або TNF-R. самоагрегації [Boldin et al., 1995а], і індуковане Одна з таких протеаз, і її різні ізоформи (вклюзв'язування двох цитоплазматичних білків (які тачаючи форми, що інгібують), відома як МАСН (текож можуть зв'язуватися один з одним) із внутрішпер каспаза-8), що є білком, який зв'язує MORT-1, і ньоклітинними доменами рецепторів - MORT-1 що слугує для модуляції активності M0RT-1 і, от(або FADD) із CD95 [Boldin et al., 1995b; Chinnaiyan же, CD95 і CD120а, і яка також може діяти незалеet al., 1995; Kischkel et al., 1995] і TRADD із CD120a жно від MORT-1, недавно виділена, клонована, [Hsu et al., 1995; Hsu et al., 1996]. Три білки, що охарактеризована, і також описані її можливі зазв'язуються з внутрішньоклітинним доменом CD95 стосування, як докладно зазначено у включених у і CD120a в області "домену загибелі", залучені до даний опис у якості посилань одночасно розглянуіндукції загибелі клітини рецепторами через гететих заявках на патент Ізраїлю, що знаходяться у роасоціацію гомологічних областей, і які, незалежспільній власності, №№ IL 114615, 114986, 115319, но, здатні також ініціювати загибель клітини, іден116588 і 117932, а також у відповідному їм аналогу тифіковані за допомогою методу скринінгу з двома РСТ №PCT/US96/10521, і в останніх публікаціях гібридами дріжджів. Одним із них є білок MORT-1 авторів даного винаходу [Boldin et al., 1996]. Авто[Boldin et al., 1995b], відомий також як FADD рами даного винаходу (неопубліковані дані) і ін[Chinnaiyan et al., 1995], який специфічно зв'язушими [Fernandes-Ainemri et al., 1996; Srinivasula et ється з CD95. Другий білок TRADD [див. також Hsu al., 1996] також недавно виділена і охарактеризоet al., 1995, 1996] зв'язується з CD120a, і третій вана ще одна така протеаза і її різні ізоформи RIP [див. також Stanger et al., 1995] зв'язується як (включаючи інгібуючі форми), названа Mch4 (також із CD95, так і з CD120a. Крім зв'язування з CD95 і з називана каспазою-10). Каспаза-10 також є білком, CD 120а ці білки також здатні зв'язуватися один з що зв'язує MORT-1, слугує для модуляції активноодним, що забезпечує функціональне "кроссті MORT-1 і, отже, також, мабуть, CD95 і CD120а, спілкування" між CD95 і CD120а. Таке зв'язування і який також може діяти незалежно від MORT-1. здійснюється через консервативний мотив посліОтже, подробиці щодо всіх аспектів, особливосдовності - "модуль домену загибелі", звичайний тей, властивостей і застосування каспази-10 наводля рецепторів і асоційованих із ними білків. Крім дяться в зазначених вище публікаціях, які усі того, хоча при випробуванні з двома гібридами включені в даний опис у якості посилань. дріжджів показано, що MORT-1 у клітинах ссавців Потрібно також відзначити, що каспази - касспонтанно зв'язується з CD95, таке зв'язування паза-8 і каспаза-10, що мають подібні продомени відбувається тільки після стимуляції рецептора, [див. Boldin et al., 1996; Muzio et al., 1996; причому передбачається, що MORT-1 бере участь Fernandes-Alnemri et al., 1996; Vincent and Dixit, в ініціації подій передачі сигналу CD95. MORT-1 не 1997], взаємодіють через свої продомени з MORTмістить якого-небудь мотиву послідовності, який 1, причому така взаємодія здійснюється через характеризується ферментативною активністю, і, "мотив домену загибелі" або "ефекторний домен отже, представляється, що його здатність ініціюзагибелі" DED, що присутній у N-кінцевій частині вати загибель клітини не викликає справжньої акMORT-1 і присутній у двох копіях в каспазі-8 і кастивності самого MORT-1, а скоріше активує якийсь пазі-10 [див. Boldin et al., 1995b; Chinnaiyan et al., інший білок або білки, що зв'язуються з MORT-1, і 1995]. діє далі в каскаді передачі сигналу. Показано, що Такі протеази, відомі в даний час як каспази експресія в клітинах мутантів MORT-1, позбавле(цистеїн-аспартатспецифічні протеази), відносятьних N-кінцевої частини молекули, блокує індукцію ся до сімейства цистеїн-протеаз, які сприяють росцитотоксичності CD95 або CD120a [Hsu et al., ту і які мають загальні ознаки. Виявлено, що біль 11 75564 12 шість таких каспаз беруть участь в ініціації і здійсблизькість із N-кінцем довгої субодиниці для того, ненні запрограмованої загибелі клітин або апоптощоб утворити правильно складений і активний зі, у той час як виявляється, що інші беруть участь фермент [Rotonda et al., 1996; Mitti et al., 1997; у продукуванні прозапальних цитокінів [Nicholson Srinivasula et al., 1998]. D.W. et al., 1997; Salvesen G.S. et al., 1997; Cohen N-ацетилглюкозамін-6-фосфатдеацетилаза є G.M., 1997]. Вони синтезуються як каталітично внутрішньоклітинним ферментом, що бере участь, неактивні попередники і, як правило, активуються як відомо, у внутрішньоклітинному метаболізмі за допомогою розчеплення після специфічних внуглюкозаміну. Фрагмент геномної ДНК, що містить трішніх аспартатових залишків, що присутні у міжN-ацетилглюкозамін-6-фосфатдеацетилазу, клодоменних лінкерах. Сайти розчеплення каспаз нований із бактерії, що продукує хітиназу, Vibrio визначаються тетрапептидною послідовністю (X-Xcholerae [Yamano et al., Biosci. Biotechnol. X-D), і розчеплення завжди відбувається після Biochem., 61, p.1349-53, 1997]. аспарагінової кислоти. В результаті деякі зрілі акГен nagA, що кодує N-ацетилглюкозамін-6тивні каспази можуть процесувати і активувати як фосфатдеацетилазу Ε. coli, також відомий [див., свої власні, так і інші, неактивні попередники наприклад. Peri et al., January 1990, 68(1), pp.123[Fernandes-Alnemri Т. et al., 1996; Srinivasula S.M. 137]. Про клонування і секвенування Net al., 1996]. ацетилглюкозамін-6-фосфатдеацетилази людини Активація процесу запрограмованої загибелі поки не повідомляється. клітин, як правило, специфічна і включає послідоВинахід відноситься до білка, що взаємодіє з вний процесінг нижніх каспаз, названих "виконуюкаспазою-8, або до його ізоформи, алельного вачими" каспазами, верхніми каспазами, названими ріанта, фрагмента, функціонального аналога, му"ініціюючими" каспазами. Функціональні характетанта або похідного, здатного взаємодіяти із субористики двох класів каспаз також відбиваються в диницею 1 та/або субодиницею 2 каспази-8. їхній будові. Дійсно, "ініціюючі" каспази містять Винахід також відноситься до Nбільш довгу продоменну область, у порівнянні з ацетилглюкозамін-6-фосфатдеацетилази людини "виконуючими" каспазами [Salvesen G.S. et al., або її ізоформи, алельного варіанта, фрагмента, 1997; Cohen G.M., 1997]. Довгий продомен дозвофункціонального аналога, мутанта або похідного. ляє ініціюючим або "апікальним" каспазам активуВинахід також відноситься до білка, що місватися шляхом ініціації рецепторів загибелі сімейтить амінокислотну послідовність, подану на Фіг.2, ства рецепторів для TNF. Після індукованої 3, 5В або 6. лігандом тримеризації рецепторів загибелі, ініціюВинахід також відноситься до білка, що місючі каспази набираються через їхній довгий Nтить амінокислотну послідовність білка Тір-60, крім кінцевий продомен для взаємодії зі специфічними амінокислот з 94 по 145. адаптерними молекулами з утворенням комплекТакож в обсяг винаходу входить білок, що міссу, що передає сигнал індукування загибелі [Cohen тить амінокислотну послідовність, кодовану клоG.M., 1997; Kischkel F.C., et al., 1995]. Наприклад, нами Р27, Р70, Р79, L7, L12, М26, В4, В17, J40, каспаза-8/МАСН і, мабуть, каспаза-10, які містять В13, В37, В33 або Р74, і його сплайсировані варіадва ефекторні домени загибелі (DED) або FADDнти Р16 і Р43, як описано нижче. домену, включаються в рецепторний комплекс за Винахід також відноситься до описаного вище допомогою адаптерних молекул FADD/MORT-1, у білка, що розщеплюється каспазою-8 in vitro або in той час як каспаза-2 включається за допомогою vivo. CRADD/RAIDD і RIP [Nagata S. et al., 1997; Винахід також відноситься до виділеної посліMcFarlane M. et al., 1997; Ahmad M. et al. , 1997; довності ДНК, що кодує білок винаходу. До обсягу Duan H. et al., 1997]. Внаслідок тримерної природи винаходу входить послідовність ДНК, подана на активованого рецепторного комплексу думають, Фіг.2 і Фіг.3. До обсягу винаходу також входить що щонайменше дві молекули каспази приходять виділена ДНК, здатна гібридизуватися із зазначеу безпосередню близькість одна з одною, що приною послідовністю ДНК у помірно суворих умовах. зводить, таким чином, їх до активації шляхом ауВинахід також відноситься до вектора, що містокаталітичного процесування [Yang et al., 1998; тить послідовність ДНК, визначення якого дано Muzio et al., 1998]. вище. Каспази синтезуються як проферменти, що Винахід також відноситься до еукаріотичної складаються з трьох основних субодиниць: Nабо прокаріотичної клітини-хазяїна, що містить кінцевого продомену і двох субодиниць, іноді розвектор винаходу. ділених лінкерним пептидом. Ці дві субодиниці Винахід також відноситься до способу одерназвані "довгою" або субодиницею 1, що містить жання білка, ізоформи, алельного варіанта, фрагактивну ділянку ферменту, і "короткою" або субомента, функціонального аналога, мутанта або подиницею 2. Для повної активації ферменту продохідного білка згідно з винаходом, що взаємодіє з мен і дві субодиниці розщеплюються. Дві відщипкаспазою-8, який включає вирощування клітинилені субодиниці утворюють гетеродимер, хазяїна згідно з винаходом в умовах, що допускавідповідно до чого довгий домен утворюється з Nють продукування зазначеного білка, вплив на кінця, а коротка субодиниця утворюється з Спосттрансляційні модифікації, який необхідний для кінцевої області попередника каспази. Виходячи з одержання зазначених білка, ізоформи, алельного установленої тривимірної структури каспази-3 виваріанта, фрагмента, функціонального аналога, являється, що С-кінець довгого домену так само, мутанта або похідного, і виділення зазначених як і N-кінець короткого субдомену, вільні, і С-кінець білка, ізоформи, алельного варіанта, фрагмента, короткої субодиниці приходить у безпосередню функціонального аналога, мутанта або похідного. 13 75564 14 Винахід також відноситься до зазначеного ка, що взаємодіє з каспазою-8, ізоформи, алельноспособу, де клітиною є прокаріотична клітина. го варіанта, фрагмента, функціонального аналога, Винахід також відноситься до зазначеного мутанта або похідного, зазначених рибозиму, анспособу, де клітиною є еукаріотична клітина. тисмислового олігонуклеотиду або антитіла для Винахід також відноситься до зазначеного застосування при модуляції активності каспази-8. способу, де клітиною є клітина ссавця, комахи або Винахід також відноситься до зазначених білдріжджів. ка, що взаємодіє з каспазою-8, ізоформи, алельноВинахід також відноситься до зазначеного го варіанта, фрагмента, функціонального аналога, способу, де клітиною є клітина HeLa або 293 Τ мутанта або похідного, зазначених рибозиму, анНЕК. тисмислового олігонуклеотиду або антитіла для Винахід також відноситься до зазначеного застосування при модуляції дій, опосередкованих способу, де в якості промотору використовують рецептором для TNF або Fas. промотор CMV людини. Винахід також ще відноситься до зазначених Ще винахід також відноситься до пептиду, що білка, що взаємодіє з каспазою-8, ізоформи, алевзаємодіє з каспазою-8, який містить щонайменше льного варіанта, фрагмента, функціонального 4 послідовних амінокислоти білка винаходу. аналога, мутанта або похідного, зазначених рибоПохідне зазначеного пептиду також входить до зиму, антисмислового олігонуклеотиду або антитіобсягу винаходу. Також до обсягу винаходу вхола для застосування при модуляції апоптозу. дить похідне зазначеного пептиду, здатне до утвоВинахід також відноситься до зазначених білрення ковалентного зв'язку з каспазою-8 після конка, що взаємодіє з каспазою-8, ізоформи, алельнотактування із зазначеною каспазою-8. го варіанта, фрагмента, функціонального аналога, Винахід також відноситься до рибозиму, спемутанта або похідного, зазначених рибозиму, анцифічному для нуклеотидної послідовності, що тисмислового олігонуклеотиду або антитіла для відповідає послідовності ДНК винаходу. застосування в якості лікувального засобу. Винахід також відноситься до антисмислового Винахід також відноситься до зазначених білолігонуклеотиду, що містить щонайменше 9 нукка, що взаємодіє з каспазою-8, ізоформи, алельнолеотидів послідовності, що відповідає послідовного варіанта, фрагмента, функціонального аналога, сті ДНК винаходу. мутанта або похідного, зазначених рибозиму, анВинахід також відноситься до антитіла проти тисмислового олігонуклеотиду або антитіла для епітопу білка винаходу. застосування в якості лікувального засобу при Винахід також відноситься до імуноаналізу для лікуванні розсіяного склерозу з первинною олїговиявлення білка, що взаємодіє з каспазою-8, за дендрогліопатїєю, автоімунного увеоретиніту, діаучастю антитіла винаходу. бету, вовчака, автоімунного міокардиту І, опосереВинахід також відноситься до імуноаналізу для дкованого HCV хронічного гепатиту, хронічного виявлення каспази-8 за участю пептиду винаходу. гастриту, наприклад, гастриту типу А, змішаної Винахід також відноситься до імуноаналізу для хвороби сполучної тканини (MCTD), хвороби Кровиявлення каспази-8 за участю білка винаходу. на або неспецифічного виразкового коліту. Винахід також відноситься до способу ідентиВинахід також відноситься до олігонуклеотиду, фікації білка, що взаємодіє з каспазою-8, що вклющо утворює триплет, здатному зв'язуватися з бачає стадії гатою пурином послідовністю в промоторній облаа) надання дріжджової клітини, що має репорсті гена, що взаємодіє з каспазою-8. Олігонуклеотерний ген, зв'язаний із промотором, що містить тид, що утворює триплет, може містити хімічні мотив послідовності ДНК; модифікації, такі як міжнуклеозидні фосфатні зв'яa) експресії в зазначеній дріжджовій клітині сузки, модифіковані катіоном Ν,Νбодиниці р20 зазначеної каспази-8; діетилетилендіаміну, залишки уридину, бензоb) експресії в зазначеній дріжджовій клітині [g]хіназолін-2,4-діон-(1Н,3Н)-діону або бензлитого білка з домену зв'язування ДНК і субодизо[f]хіназолін-2,4-діон-(1Н,3Н)-діону замість тиміниці р10 та/або р20 зазначеної каспази-8, де задинових залишків. Олігонуклеотид, що утворює значений домен зв'язування ДНК здатний до зв'ятриплет, також може ковалентно зв'язуватися з зування із зазначеним мотивом послідовності ДНК; хімічними речовинами, афінними до ДНК, переваc) необов'язково, експресії в зазначеній дріжжно, такими, які можна вбудувати в ДНК, такими як джовій клітині незлитої субодиниці р10 або р20 акридин і псорален. зазначеної каспази-8; Термін "взаємодіючий" у контексті даного опиd) трансформації культури зазначеної дріжсу, що відноситься до взаємодії білка винаходу з джової клітини бібліотекою, що складається з екскаспазою-8, призначений для позначення форм пресуючого вектора, який керує експресією злитопрямої взаємодії, таких як зв'язування і розчепго білка та який складається з бібліотеки кДНК і лення, і непрямої взаємодії, наприклад, через активатора транскрипції; адаптерні білки. Взаємодія може привести, необоe) скринінгу культури трансформованих дріжв'язково, до модуляції сигналу, опосередковуваноджових клітин на дріжджові клітини, в яких активого каспазою-8. ваний репортерний ген; і Термін "зв'язування" у контексті даної заявки, f) виділення дріжджової клітини зі стадії e) і коли відноситься до зв'язування білків, що взаєнаступного виділення білка, який взаємодіє з касмодіють із каспазою-8, призначений для позначенпазою-8, та який експресований в захопленому ня фізичного зв'язку білка, що взаємодіє з каспанею векторі. зою-8, із каспазою-8. Цей фізичний зв'язок можна Винахід також відноситься до зазначених білвиміряти при імуноаналізах, таких як аналізи 15 75564 16 ELISA або РІА, при двогібридному дослідженні, гу, описаному в прикладі 1В. аналізах із імунопреципітацією або аналізах на Фігура 2 представляє попередній неповний нуоснові розділення за розміром, таких як електроклеотид (Послід. №1) і розшифровану амінокислофорез у гелі, що не денатурує, або гельтну послідовність (Послід. №2) клону 32, що кодує хроматографія, суміші каспази-8 або її субодиниці білок, що взаємодіє з каспазою-8, отриманий із і білка винаходу. При використанні двогібридного клону кДНК. дослідження мається на увазі, що каспаза-8 або її Фігура 3 показує передбачувану повнорозмірсубодиниця експресуються у вигляді гібрида дону послідовність нуклеотиду людини (Послід. №3) і мену активації ДНК, а білок, що взаємодіє з каспарозшифровану амінокислотну послідовність (Посзою-8, експресується у вигляді гібрида домену лід. №4) N-ацетилглюкозамін-6зв'язування ДНК, або навпаки. фосфатдеацетилази, складену з нарощування по Термін "двогібридне дослідження" відноситься 5' клону J2 клону EST АА460869 і клону пастки до двогібридного аналізу, коли білок-білкові взаєекзону L48741. модії можна виміряти за допомогою введення в Фігура 4А показує функціональну активність дріжджові клітини першого експресуючого вектора, експресованого клону J2 у вигляді відсотка апопщо кодує гібрид першого білкa і домену зв'язувантозних клітин після трансфекції клітин НЕК-293Т ня ДНК, і другого експресуючого вектора, що кодує або одним рецептором для TNF р55 (позначеним гібрид другого білка, і домену активації ДНК. Дріжр55 TNFR), або рецептором для TNF р55 разом із джові клітини повинні містити щонайменше один р35 - бакуловірусним інгібітором каспаз (р55+р35), репортерний ген, керований промотором, що місабо з клоном J2 (p55+J2), або рецептором для тить мотив послідовності ДНК, розпізнаваний заTNF р55 разом із нерозщеплюваним мутантом J2 значеним доменом зв'язування ДНК. Як правило, (позначеним J*), що містить заміну Asp на Glu у використовують два репортерних гени, наприклад, позиції 346 на Фігурі 3 (p55+J2*). Цей мутант не гістидин-синтетазу і бета-галактозидазу. Це доможна розщепити ні in vitro, ні in vivo. зволяє дослідникам уникнути помилкових позитиФігура 4В показує відсоток апоптозних клітин вних результатів, які можуть з'явитися від мутації. HeLa, одних або після трансфекції р35, або J2, або Цей метод модифікований і удосконалений автоJ2*, оброблених TNF і циклогексімидом. рами даної заявки для застосування при скринінгу, Фігура 5А показує 1725 кодуючих пар основ в виділенні і випробуванні білків, що опосередкову5'-кінці клону Р74 (Послід. №5). ють сигнали TNF-R і Fas, див., наприклад, WO Фігура 5В показує послідовність 574 амінокис97/03998 і приведені там посилання. Двогібридне лот (Послід. №6), розшифровану з послідовності дослідження спирається на локалізацію обох зликлону Р74 показаної на Фігурі 5А. тих білків у ядрі клітини. Фігура 6 показує послідовність 1428 амінокисТермін "двогібридне дослідження", використолот (Послід. №7) відкритої рамки зчитування, вуваний тут, також включає модифікацію двогіботриману з розшифрованої амінокислотної посліридного методу, коли використовують білки, що довності клону РАС, інвентарний номер RPCI5подають сигнал росту клітини, такі як ras і sos 1057120. [Broder et al, Curr. Biol., 1998, 8, p.1121-4; Aronheim Фігура 7 показує розташовані в ряд 574 аміноet al., Nucleic Acids Res., 1997, 25, p.3373-4, і закислоти відкритої рамки зчитування, отриманої з значені в цих роботах посилання]. Для того, щоб розшифрованої амінокислотної послідовності клобути функціональними, цим білкам потрібно перену р74 (позначеної "клонована"), у порівнянні з міщення в клітинну мембрану. Це досягається за 1428 амінокислотами відкритої рамки зчитування, допомогою експресії білка, що подає сигнал росту отриманої з розшифрованої амінокислотної посліклітини, у вигляді гібрида з першим білком, і ексдовності клону РАС, RPCI5-1057120 (позначеної пресії сигналу локалізації в клітинній мембрані, "розшифрована"). такого як сигнальна послідовність міристилюванФігура 8 показує розташовану в ряд відкриту ня, у вигляді гібрида з другим білком. Взаємодія рамку зчитування розшифрованої амінокислотної між першим і другим білком буде переміщати біпослідовності клону RPCI5-1057I20 (верхня послілок, що подає сигнал росту клітини, у клітинну довність) із послідовністю гістондеацетилази А мембрану, і в такий спосіб ініціювати ріст клітини. (нижня послідовність, інвентарний номер Genbank Потім відбирають клітини, що ростуть агресивно, NP-006028.1) (Послід. №9). для подальшого дослідження. Ця система відрізФігура 9А показує ауторадіографію Bid-білка і няється від описаної вище двогібридної системи, білків, кодованих клонами кДНК J2, або Ρ16, або оскільки не заснована на локалізації обох гібридів Р43, або Р70, або Р74, або Р79, отриманих в лізаті у ядрі клітини. ретикулоцитів у присутності 35S-метіоніну, роздіТермін "наживка" відноситься до білка у вищелених методом SDS-PAGE. вказаному двогібридному дослідженні, який ексФігура 9В показує ауторадіографію Bid-білка і пресується у вигляді злитого білки з доменом зв'ябілків, кодованих клонами кДНК J2, або Р16, або зування ДНК. Р43, або Р70, або Р74, або Р79, отриманих так Термін "здобич" відноситься до білка у вищесамо, як у випадку Фігури 9А, при аналізі на зв'язувказаному двогібридному дослідженні, який ексвання з каспазою-8, експресованої у бактеріях у пресується у вигляді злитого білка з доменом аквигляді злитого білка двох її субодиниць, гібридитивації ДНК. зованих із GST. Положення маркерів молекулярної Фігура 1 показує схематичне зображення одмаси показано на гелі зліва. ноланцюгової конструкції каспази-8, використовуФігура 10 показує результати розчеплення білваної в якості наживки при двогібридному скринінка, кодованого неповним клоном кДНК Р43, каспа 17 75564 18 зою-8 або каспазою-10, або каспазою-3, або касp.22995-9, 1997]. Для виявлення відповідних винапазою-9, або мутантами каспази-3, або каспази-9, ходу білків, які взаємодіють із каспазою-8, необабо каспази-10. Об'єми усіх використаних бактеріхідно, щоб дві субодиниці каспази-8 експресувалиальних лізатів рекомбінантних каспаз, експресовася незалежно. З цією метою дві субодиниці них в Е. соІі, показані у відносних одиницях (RU). каспази-8 експресують, переважно, окремо під Положення маркерів молекулярної маси показано контролем різних промоторів. У кращому варіанті на гелі зліва. Білки, що представляють інтерес, втілення винаходу субодиниця каспази-8 р10 (від позначені зірочками: стрілки з заштрихованими серину 375 до аспарагінової кислоти 479) експревістрями показують повнорозмірний білок Р43, а сується під контролем слабкого промотору, який стрілки з незаштрихованими вістрями показують можна використовувати у дріжджах, у рамці з допродукти розчеплення. меном зв'язування ДНК. Переважно, слабкий проФігура 11 показує функціональну активність мотор являє собою промотор дріжджів ADH, а білбілків, кодованих клонами кДНК, ідентифікованими ком, що зв'язується з ДНК, є домен зв'язування за допомогою двогібридного скринінгу, виражену у ДНК дріжджового активатора транскрипції Gal-4 вигляді відсотка клітин, що зазнають апоптоз, пісабо бактеріальний білок Lex. ADH-промотор і доля котрансфекції клітин НЕК-293Т рецептором для мен зв'язування ДНК Gal4 є, наприклад, у доступTNF р55 і білком зеленої флуоресценції (позначеному комерційно pGBD, Пало-Альто, США. Однак ним PC), без або зі вставками кДНК клонів J2, або для фахівців у цій області техніки буде очевидно, Ρ16, або Р27, або Р43, або Р79, або Р74, або Р70. що можна використовувати інші промотори, доти, Фігура 12 показує інгібуючу активність у відпоки промотор ефективний у дріжджових клітинах. ношенні загибелі клітин дикого типу і нерозщепПо цій же ознаці можна використовувати інші долюваного мутанта Тір60 у клітинах НЕК-293Т, котмени зв'язування ДНК, доти, поки такі домени зв'ярансфікованих рецептором для TNF р55 і білком зування ДНК не мають функції активатора транскрипції. зеленої флуоресценції, а також дія 32-Тір60, позДовга активна субодиниця каспази-8 р20 (від бавленого перших 32 N-кінцевих амінокислот. Консерину 217 до аспарагінової кислоти 374) експретрольні клітини трансфіковані одним вектором сується у вигляді окремого білка під контролем pCGN. індуцибельного промотору, який можна викорисБагато методів молекулярної біології тут дотовувати для дріжджових клітин. Переважно, можкладно не описуються, тому що вони добре відомі на використовувати репресований промотор метіфахівцям у цій області техніки. До таких методів онін Met25, як описано в зазначеній вище відносяться сайтспрямований мутагенез, клонупублікації Tirode et al. Використання індуцибельновання методом PCR, скринінг фагової бібліотеки з го промотору спрощує виявлення комплексу р10використанням олігонуклеотидних або кДНКр20 за допомогою імунопреципітації і електрофозондів, експресія кДНК, аналіз рекомбінантних білрезу в поліакриламідному гелі. Індуцибельний ків, трансформація бактеріальних і дріжджових промотор має також перевагу в тому, що він клітин, трансфекція клітин ссавців і т.п. До літерауможливлює експресію в дріжджах білка, який мотури, де описуються такі методи, відносяться, наже бути токсичним для дріжджових клітин, внасліприклад, видання [Sambrook et al., "Molecular док можливості обмеження періоду, у межах якого Cloning A Laboratory Manual", Cold Spring Harbor екс пресується потенційно токсичний білок. Laboratory; ISBN: 0879693096, 1989, "Current Білки, які скриніються згідно зі способом винаProtocols in Molecular Biology", by F.M. Ausubel, ходу, переважно, представляють у формі бібліотеISBN: 047150338X, 1988, і "Short Protocols in ки кДНК. Однак можна використовувати також геMolecular Biology", by F.M. Ausubel et al. (eds.), 3rd номні бібліотеки або комбіновані бібліотеки. ed., Jhon Wiley & Sons; ISBN: 0471137812, 1995]. Ці Бібліотеку клонують по С-кінцю домену активації публікації включені до даного опису у якості поситранскрипції, операбельного в дріжджах. Перевалань. жно використовують дріжджовий домен активації Для того, щоб методом двогібридного скринінтранскрипції Gal-4, однак можна використовувати гу ідентифікувати білки, що взаємодіють із каспабагато інших активаторів транскрипції. Переважно, зою-8, можна використовувати систему з двома для клонування бібліотеки використовують вектор гібридами або трьома гібридами. pGAD GH, доступний від Clontech. Систему з двома гібридами у способі винаходу Штам дріжджів, використовуваний для скринінвикористовують, по суті, у тому вигляді, у якому гу, повинен містити селективний маркер, такий як вона описана [Fields and Song (Nature, 340, p.245, гістидинсинтетаза, під контролем промотору, що 1989)]. Переважно, окремі вектори, дріжджові містить послідовність ДНК, з якою специфічно штами і бібліотеки можна одержати від Clontech зв'язується вищевказаний домен зв'язування ДНК. (Пало-Альто, США)], як компоненти двогібридної Переважно, дріжджова клітина також містить ресистеми Matchmaker (#PT1265-1). портерний ген під контролем промотору, що місКращий варіант системи з двома дріжджовими тить послідовність ДНК, з якою специфічно зв'язугібридами, використовуваної в способі винаходу, ється вищевказаний домен зв'язування ДНК. Штам описаний [Boldin et al., Cell, 85, p.803-15, 1996]. дріжджів HF7c, доступний від Clontech, можна виСистема з двома дріжджовими гібридами також користовувати для скринінгу з гібридами домену описана в [патенті США 5580736, Brent et al]. Ці зв'язування Gal-4; штам L40 можна використовупублікації включені до даного опису у якості посивати, коли використовують домен зв'язування ДНК лань. lexA. Тригібридна система використовується, по суПісля трансформації дріжджові клітини поміті, так, як описано [Tirode et al., J. Biol. Chem., 272, 19 75564 20 щають в умови, селективні для активного висіванподвійних трансформантів, тобто дріжджових кліня в середовища, що позбавлені деяких амінокистин, трансформованих знову ідентифікованим лот, які потрібні для стійкості введених у них плазклоном і субодиницею каспази-8 - або р10 або р20, мід. де зазначена субодиниця злита з доменом зв'язуСередовище є селективним для дріжджових вання ДНК. Зв'язування з комплексом субодиниць клітин, в яких активується ген для вищевказаного р10 і р20 також можна легко оцінити за допомогою селективного маркера. Переважно, селективний визначення lacZ-активності потрійних трансформаркер являє собою ген гістидинсинтетази. Клітимантів, де одна із субодиниць каспази-8 експресуни дріжджів, що експресують цей ген, можна відібється у вигляді злитого білки з доменом зв'язуванрати для культивування в середовищі, позбавленя ДНК, а інша є окремим білком. Очевидно, що ному гістидину. Перевагою такої системи є вищевказана тригібридна система, таким чином, можливість додавання в середовище для росту також корисна для визначення характеристик зв'яінгібітору гістидинсинтетази 3-амінотриазолу. Тазування клонів, знайдених за допомогою двогібриким чином, можливо пригнічувати ріст дріжджових дної системи, тому що індуцибельний промотор, клітин, в яких гістидинсинтетаза експресується в використовуваний у ній, дає можливість швидкої невеликій кількості, що викликано витоком промоідентифікації клонів, що взаємодіють тільки із сутору, який містить послідовність, з якою специфічбодиницею 10 або з комплексом субодиниць р10но зв'язується вищевказаний домен зв'язування р20. ДНК. У деяких клонах слабка неспецифічна взаєКлони, здатні рости в середовищі, позбавлемодія між субодиницею каспази-8 р10 та/або р20 і ному гістидину, і такі, що експресують lacZзазначеним клоном може викликати гадану актиактивність, потім відбирають для подальшого досвацію зазначеного промотору. Таким чином, підлідження. Спочатку білки, кодовані зазначеними вищуючи концентрацію зазначеного інгібітору в клонами, перевіряють на їхню здатність зв'язувасередовищі, використовуваному для селекції взатися з нерелевантними білками, такими як ламін. ємодіючих клонів, можна відібрати тільки такі клоЯк правило, клони, що зв'язуються з ламіном, відни, що взаємодіють із певною мінімальною силою. кидають. Концентрація 3-амінотриазолу становить, переваКлони, знайдені для специфічної взаємодії із жно, 7,5мМ. каспазою-8, потім аналізують далі. Такий аналіз Клони, ідентифіковані по їхній здатності рости здійснюють із неповними клонами, отриманими в середовищі, позбавленому гістидину, потім анабезпосередньо при описаних вище способах скрилізують шляхом кількісного визначення активності нінгу, а також із повнорозмірними клонами, отриїхнього репортерного гена. Переважно в якості маними на основі послідовності зазначених непорепортерного гена використовують ген lacZ. Кільвних клонів. Для того, щоб одержати кісне визначення активності lacZ здійснюють, пеповнорозмірні клони, одержують послідовність реважно, у рідкій культурі, як описано [Boldin et al., неповного клону за допомогою екстракції ДНК заJ. Biol. Chem., 270, 7795-8, 1995]. значеного клону з дріжджових клітин способами, Описаний вище спосіб скринінгу можна здійсвідомими фахівцям у цій області техніки. Потім нити подібним чином із використанням двогібридДНК переносять у бактерії, для того, щоб одержаного дослідження. Істотна відмінність полягає в ти велику кількість очищеної ДНК, яку можна викотому, що дві субодиниці каспази-8 експресуються ристовувати для секвенування. З іншого боку, для у вигляді єдиного пептидного ланцюга і являють цілей секвенування вставку у векторі, що предстасобою злитий білок із зазначеним вище доменом вляє собою, переважно, вищевказаний вектор зв'язування ДНК. Субодиницю р20 мутують у її pGBD, можна вирізати з використанням рестриктаз активному сайті (цистеїн 360 - серин 360) так, як і клонувати в інший вектор, такий як pBluescript, описано нижче. доступний від Stratagene. Секвенування здійснюПереважно, субодиницю р10 відокремлюють ють методом термінації ланцюга, переважно, із від субодиниці р20 за допомогою лінкера, що, певикористанням ферменту секвенази-2, доступної із реважно, має довжину від 10 до 50 амінокислот. набору для секвенування від United States Зазначений лінкер містить, переважно, невеликі Biochemicals. незаряджені амінокислоти, такі як гліцин, серин, Потім отриману в такий спосіб послідовність треонін, аланін і валін. Краще, лінкер складається можна ввести в базу даних програми пошуку і за із залишків серину і гліцину. Найбільш переважно, допомогою комп'ютерного пошуку ідентифікувати коли співвідношення між залишками гліцину і запослідовності, що перекриваються. Використовулишками серину в зазначеному лінкері складає вані програми добре відомі фахівцям у цій області приблизно від 3:1 до 4:1. техніки і включають, наприклад, пакет GCG Одержання клонів із використанням двогібри(genetics computer group). Переважно використодної системи з описаною вище наживкою каспазивують сервісну програму пошуку, таку як Basic 8 подібно описаному вище застосуванню тригібриLocal Aligment Search Tool (BLAST), доступну з дної системи, за винятком того, що клони, що зв'ясерверу EMBL (наприклад, http://dove.emblзуються тільки з однією із субодиниць, важко відріheidelberg.de/Blast2/). Команди Blastn можна викознити від клонів, що зв'язуються з обома ристовувати для пошуку нуклеотидних послідовсубодиницями або вимагають для зв'язування ностей, які перекривають або які схожі на ідентикомплекс із двох субодиниць. Однак будь-який фіковані клони. клон, виявлений при двогібридному способі, можБілок, ідентифікований згідно зі способом вина легко перевірити на зв'язування з двома субонаходу, представляють у вигляді злитого білка з диницями за допомогою оцінки lacZ-активності доменом зв'язування ДНК. Отже, рамка, в якій по 21 75564 22 винна транслюватися нуклеотидна послідовність, Qiagen Gmb, Хілден, Німеччина. Метод імунопревідома, тому що вона повинна бути в рамці з коципітації описаний у зазначеній вище роботі дуючою послідовністю домену зв'язування ДНК. Boldin et al., Cell, 1996]. Реагенти і вказівки до імуПослідовність ДНК клону, ідентифікованого згідно нопреципітації також доступні у формі набору від з винаходом, можна, отже, однозначно транслюBoehringer Mannheim, Маннгейм, Німеччина. вати в амінокислотну послідовність. Потім для Потім лізат цілих клітин очищеного білка розідентифікації білкових послідовностей, що перекділяють за розміром за допомогою електрофорезу риваються, або подібних білків можна використов поліакриламідному гелі з додаванням SDS. вувати програму Blastp, доступну на вищевказаДосліджуваний білок або його мічені фрагменному сервері EMBL. ти тепер можна візуалізувати методом вестернЗ іншого боку, або крім вищевказаних способів блотінгу з використанням антитіл проти мітки. пошуку в базі даних, для того, щоб ідентифікувати Кращою міткою є гістидинова мітка в сполученні з повні клони, можна скриніювати бібліотеку, таку як антитілом проти полігістидину. Однак можна викогеномна бібліотека або бібліотека кДНК. Такі спористовувати інші сполучення послідовності-мітки соби скринінгу описані в зазначених вище виданта антитіла, специфічного до неї, доти, поки антинях [Sambrook et al. і Ausubel et al.]. З іншого боку, тіло залишається специфічним до послідовностіабо крім того, можна використовувати методи кломітки, тобто не впізнає інші білки в лізаті цілих клінування на основі PCR, такі як швидка ампліфікатин. Специфічність розчеплення можна перевірити ція кінців кДНК [5' and 3' RACE, Graham et al., шляхом проведення контрольних реакцій, при яких Biochem. Biophys. Res. Commun., 177, p.8-16,1991, до культури клітин ссавця в зазначений вище пеі зазначені в цій роботі посилання]. ріод часу добавляють специфічний інгібітор каспаНеповні клони, ідентифіковані при скринінгзи. Кращий інгібітор являє собою інгібітор, обраний аналізі згідно з винаходом, або повнорозмірні клосеред zVAD-fmk, zDEVD-fmk, zIETD-fmk [див. ни, отримані будь-яким з описаних вище способів, Keppler-Hafkemeyer et al., Biochemistry, 37, потім досліджують далі. Це здійснюють, наприp.16934-42, 1998]. Більш кращим інгібітором є клад, шляхом дослідження чутливості цих клонів zVAD-fmk. Інші інгібітори, які можна використовудо протеолітичного розчеплення активною каспавати, являють собою білки, такі як Всіх або білок зою-8. Цей аналіз можна здійснювати in vivo. Для р35, що діють як клітинні інгібітори каспаз. Пригніцієї мети лінію клітин ссавця трансфікують експречення розчеплення, коли в ході аналізу коекспресуючим вектором, який продукує білок, кодований суються такі білки, показує, що розчеплення спедосліджуваним клоном, і експресуючим вектором, цифічно для каспаз. який кодує другий білок, експресія якого буде індуІншим аналізом з метою дослідження, чи може кувати активність каспази-8. Експресуючі вектори, досліджуваний білок розщеплюватися каспазою-8, переважно, містять сильний промотор для експреє аналіз in vitro, при якому рекомбінантно продукосії клону і другого білка, такий як вірус саркоми вана каспаза-8 використовується у ферментативРауса [RSV, Yamamoto et al., Cell, 22, p.787-97, ній реакції разом із міченим досліджуваним білком. 1980], вірус мієлопроліферативної саркоми [MPSV, Досліджуваний білок можна одержати, як описано Artelt P. et al.. Gene, 68, p.213-9, 1988], цитомегавище, за допомогою клонування його кодуючої ловірус [CMV, Thomsen et al., PNAS, 81, p.659-63, послідовності в експресуючий вектор, який містить 1984], або подібні промотори вірусного або клітинсильний промотор, і трансфекції в клітину ссавця. ного походження. Вигідно мітити досліджуваний білок, як описано Другий білок, експресія якого індукує активвище, із тим, щоб його можна було виділити в чисність каспази-8, вибирають серед внутрішньоклітому вигляді з екстракту клітин ссавця за допомотинного домену Fas, внутрішньоклітинного домену гою антитіл проти мітки або інших агентів, здатних CD120a, Mort-1, каспази-8, або еквівалента білка, специфічно зв'язуватися з послідовністю-міткою. З здатного індукувати активність каспази-8. З іншого іншого боку, досліджуваний білок можна одержати боку, активність каспази-8 можна індукувати у кліin vitro із використанням системи трансляції in vitro. тинах за допомогою обробки TNF або лігандом Метод трансляції in vitro добре відомий фахівцям у CD95. Подробиці експерименту, можливий мехацій області техніки, і всі реагенти для нього і донізм і інші технічні подробиці аналізу такого типу кладні протоколи доступні, наприклад, від описані в зазначеній вище роботі [Boldin et al., Cell, Stratagene, La Jolla, USA. 1996]. З іншого боку, досліджуваний білок можна поПісля введення в клітину ссавця зазначеного значити, наприклад, із використанням радіоізотовище експресуючого вектора, який кодує другий пу. Вигідно, при міченні ізотопами, експресувати білок, і досліджуваного білка, вирощують культуру досліджуваний білок in vitro, і під час реакції транклітин протягом часу, достатнього для здійснення сляції in vitro мічену ізотопами амінокислоту добаекспресії білків, активації або експресії каспази-8 і вляти разом із неміченою кислотою. Переважно розчеплення досліджуваного білка. Зазначений ізотоп являє собою S35. Також переважно, щоб період часу звичайно становить від 4 до 72 годин, мічена амінокислота являла собою S35-метіонін, і переважно - від 16 до 30 годин, і найбільш переспіввідношення між міченою і неміченою амінокисважно - від 20 до 24 годин. Для того, щоб визначилотою становило від 1:1 до приблизно 1:1000. ти ступінь розчеплення, одержують лізат цілих Потім отриманий рекомбінантно досліджуваклітин. З іншого боку, можна очистити мічений біний білок і отриманий рекомбінантно активний лок із використанням антитіл проти мітки або хробілок каспазу-8 з'єднують у відповідному буфері і матографії з нікелем і нітрилооцтовою кислотою, дають проходити реакції розчеплення протягом реагенти і докладні протоколи яких доступні від достатнього для цього часу. Кращий буфер і інші 23 75564 24 кращі параметри аналізу описані в зазначеній висті кДНК нативного білка, що взаємодіє з каспаще роботі Boldin et al., Cell, 1996]. Кращий період зою-8, але які містять ряд необхідних модифікацій. часу, як правило, становить від 10 хвилин до декіОтже, такі синтетичні послідовності включають усі лькох годин, переважно - від 30 хвилин до 1 годиможливі послідовності, що кодують аналоги, франи. гменти і похідні білка, що взаємодіє з каспазою-8, Після здійснення розчеплення реакційну суміш які всі мають активність білка, що взаємодіє з касрозділяють за розміром молекул методом електопазою-8. рофорезу в поліакриламідному гелі в присутності Використовувані тут суворі умови є функцією SDS. Якщо використане мічення ізотопами, гель температури, використовуваної при експерименможна висушити, і виявити ізотоп за допомогою тах по гібридизації, молярності одновалентних фотоплівки або люмінофорів (Fuji). Досліджуваний катіонів і процентного вмісту формаміду в розчині білок мітять, і виявити його можна з використандля гібридизації. Для того, щоб визначити ступінь ням специфічних до мітки антитіл при вестернстрогості для будь-якого даного набору умов, виблотінзі. користовують, у першу чергу, рівняння Мейнкота Поява додаткових низькомолекулярних смуг у (Meinkoth) et al. (1984) для визначення стійкості реакційних сумішах, у які доданий білок каспаза-8, гібридів 100% ідентичності, виражене, у вигляді у порівнянні з контрольними реакційними сумішатемператури плавлення Тm гібрида ДНК-ДНК: ми без каспази-8, вказує на розчеплення досліТm=81,5°С+16,6(LogM)+0,41(%GC)джуваного білка каспазою-8. Крім того, розмір ни0,61(%form)-500/L, де М являє собою молярність зькомолекулярних смуг вказує приблизне одновалентних катіонів, %GC являє собою % нукрозташування сайту розчеплення. леотидів G і С в ДНК, %form являє собою проценДаний винахід відноситься до послідовності тний вміст формаміду в розчині для гібридизації, і ДНК, що кодує білки, які взаємодіють із каспазоюL являє собою довжину гібрида в парах основ. На 8. кожний 1°С, коли Тm знижується, виходячи з велиКрім того, даний винахід також відноситься до чини, обчисленої для 100% ідентичного гібрида, послідовностей ДНК, які кодують біологічно активкількість допустимих помилкових спарювань зросні ізоформу, алельний варіант, функціональний тає приблизно на 1%. Таким чином, якщо Тm, вианалог, мутант або похідне білка, що взаємодіє з користовувана для будь-якого даного експерименкаспазою-8, і до кодованого ними білка, ізоформи, ту по гібридизації з певної сіллю і концентраціями алельного варіанту, функціонального аналогу, формаміду, на 10°С нижче Тm, обчисленої для мутанту або похідного. Одержання таких аналогів, 100% гібрида відповідно до рівняння Мейнкота, фрагментів, мутантів і похідних є стандартною гібридизація буде відбуватися, навіть якщо є припроцедурою [див., приклад, Sambrook et al., 1989], близно до 10% помилкових спарювань. при якій у послідовностях ДНК, що кодують білок, "Помірно суворими умовами" є умови, що заякий взаємодіє з каспазою-8, один або декілька безпечують Тm, яка не більше, ніж на 20°С нижче кодонів можна делетувати, додати або замінити Тm, що повинна бути для зробленого дуплекса з іншими і одержати аналоги, що мають щонайменнаміченою послідовністю, чи обчислена вона за ше одну заміну амінокислотного залишку щодо зазначеною вище формулою або виміряна фактинативного білка. чно. Без обмежень, у помірно суворих (на 15-20°С Серед послідовностей ДНК згідно з винахонижче обчисленої або виміряної Тm, гібрида) умодом, які кодують білок, що взаємодіє з каспазою-8, вах використовують розчин для промивання 2 X ізоформу, алельний варіант, функціональний анаSSC (стандартний сольовий розчин цитрату) і лог, мутант або похідне, також є, як варіант вті0,5% SDS (натрійдодецилсульфат) при відповідній лення винаходу, послідовності ДНК, здатні до гібтемпературі нижче обчисленої Тm гібрида. Остаторидизації з послідовністю кДНК, отриманою з чна строгість умов зумовлена в першу чергу умокодуючої області нативного білка, який взаємодіє з вами промивання, особливо, якщо використовувакаспазою-8, і така гібридизація здійснюється в поні умови гібридизації такі, при яких поряд із мірно суворих умовах, і здатні до гібридизації посстійкими гібридами утворюються менш стійкі гіблідовності ДНК кодують біологічно активний білок, риди. У такому випадку більш суворі умови промищо взаємодіє з каспазою-8. Отже, до таких здатвання дозволяють видалити менш стійкі гібриди. них до гібридизації послідовностей ДНК відносятьЗвичайною умовою гібридизації, яку можна викося послідовності ДНК, що мають відносно високу ристовувати з помірно суворими умовами промигомологію з послідовністю кДНК нативного білка, вання, описаними вище, є гібридизація в розчині 6 який взаємодіє з каспазою-8, і, як такі, представх SSC (або 6 X SSPE (стандартний сольовий фоляють послідовності, схожі на білок, що взаємодіє сфат-ЕДТК розчин)), 5 X розчин Денхардта, 0,5% з каспазою-8, що можуть являти собою, наприSDS і 100мкг/мл денатурованої фрагментованої клад, послідовності природного походження, які ДНК сперми лосося при температурі приблизно на кодують різні ізоформи білка, що взаємодіє з кас20-25°С нижче Тm. Якщо використовують змішані пазою-8, або послідовності, що трапляються в зонди, переважно використовувати тетраметилаприроді, які кодують білки, що належать до групі монійхлорид (ТМАС) замість SSC (Ausubel, 1987, послідовностей, схожих на білок, що взаємодіє з 1999). каспазою-8, та які кодують білок, що має активДля того, щоб одержати зазначені вище подіність білка, що взаємодіє з каспазою-8. Крім того, бні до білка, що взаємодіє з каспазою-8, послідовдо таких послідовностей також можуть відноситиності, що трапляються в природі, можна викорисся, наприклад, послідовності, що не трапляються в товувати стандартні процедури скринінгу і природі, отримані синтетично, подібні послідовновиділення зразків ДНК або РНК природного похо 25 75564 26 дження з різних тканин із використанням у якості Gly Ala;Pro зонда кДНК природного білка, що взаємодіє з касHis Asn; Gln пазою-8, або її частини [див., наприклад, стандарlle Leu;Val тні процедури, описані в Sambrook et al., 1989]. Leu Ile;Val Винахід відноситься до білка, що взаємодіє з Lys Arg;Gln;Glu каспазою-8, який можна ідентифікувати за допомоMet Leu;Tyr;Ile гою описаного вище скринінг-аналізу. Винахід таPhe Met;Leu;Tyr кож відноситься до поліпептиду або білка який, по Ser Thr суті, відповідає білку, що взаємодіє з каспазою-8. Thr Ser Термін "який, по суті, відповідає" включає не тільки Trp Tyr білок, що взаємодіє з каспазою-8, але також поліТуг Trp;Phe пептиди або білки, що є його аналогами. Val Ile;Leu Аналоги, які, по суті, відповідають білку, що взаємодіє з каспазою-8, являють собою поліпепЗ іншого боку, іншу групу замін білка, що взаєтиди, у яких одна або декілька амінокислот аміномодіє з каспазою-8, складають заміни, при яких кислотної послідовності білка, що взаємодіє з касщонайменше один амінокислотний залишок у попазою-8, замінені іншими амінокислотами, ліпептиді видаляють і на його місце вбудовують делетовані та/або вбудовані, за умови, що отриінший залишок відповідно до приведеної далі маний білок виявляє, по суті, таку ж або більш витабл.ІВ. Типи замін, що можна зробити в поліпепсоку біологічну активність, ніж білок, що взаємодіє тиді, можуть грунтуватися на аналізі частоти стріз каспазою-8, якому він відповідає. вальності амінокислотних замін у гомологічних Для відповідності, по суті, білку, що взаємодіє білках різного виду, таких як заміни, подані [в з каспазою-8, зміни в послідовності білків, що взатабл.1-2 у роботі Schuiz et al., G.E., "Principles of ємодіють із каспазою-8, такі як ізоформи, є, як Protein Structure", Springer-Verlag, New York, NY, правило, незначними. Хоча число змін може пере1978, і на Фіг.3-9 у роботі Creighton, Т.Е. "Proteins: вищувати 10, краще, щоб змін було не більш 10, Structure and Molecular Properties", W.H. Freeman & краще - не більше 5, і найбільш переважно - не Co., San Francisco, CA, 1983]. На підставі такого більше 3 таких змін. Можна використовувати багааналізу альтернативні консервативні заміни вито методів для пошуку потенційно біологічно актизначені тут як заміни в межах однієї із зазначених вних білків, які, по суті, відповідають білкам, що далі груп. взаємодіють із каспазою-8, і одним із таких методів є застосування звичайного методу мутагенезу для ДНК, яка кодує білок, що призводить до декількох модифікацій. Потім білки, експресовані клонами, можна скриніювати на їхню здатність зв'язуватися з каспазою-8 і модулювати активність каспази-8 при модуляції/посередництві в зазначених вище внутрішньоклітинних каскадах реакцій. "Консервативними" змінами є зміни, в результаті яких не очікують зміни активності білка, і, як правило, спочатку скриніються як зміни, в результаті яких не очікують змін розміру, заряду або конфігурації білка, і не очікують зміни його біологічних властивостей. До консервативних замін білків, що взаємодіють із каспазою-8, відноситься аналог, де щонайменше один амінокислотний залишок у поліпептиді консервативно замінений іншою амінокислотою. Такі заміни, переважно, здійснюють у відповідності зі списком, поданим у таблІА, і такі заміни можна визначити шляхом звичайного експерименту і забезпечити модифікацію структурних і функціональних властивостей молекули синтезованого поліпептиду при збереженні властивостей біологічної активності білка, що взаємодіє з каспазою-8. Таблиця ΙΑ Вихідний залишок Ala Arg Asn Asp Cys Gln Glu Приклад заміни Gly;Ser Lys Gln;His Glu Ser Asn Asp Таблиця IB 1 2 3 4 5 Невеликі аліфатичні неполярні або слабкополярні залишки Полярні негативно заряджені залишки і їхні аміди Полярні позитивно заряджені залишки Великі аліфатичні неполярні залишки Великі ароматичні залишки Ala, Ser, Thr, (Pro, Gly) Asp, Asn, Glu, Gln His, Arg, Lys Met, Leu, He, Val, (Cys) Phe, Туr, Тrр Три амінокислотні залишки, зазначені вище в дужках, грають особливу роль у будові білка. Gly є єдиним залишком, позбавленим якогось бічного ланцюга, і, таким чином, додає ланцюгу гнучкості. Однак ця обставина викликає тенденцію промотування утворення вторинної структури іншого типу, ніж а-спіраль. Pro, у силу своєї незвичайної геометрії, жорстко стримує ланцюг і, як правило, має схильність промотувати структури, подібні бетаповороту, хоча в деяких випадках Cys може бути здатний брати участь в утворенні дисульфідного зв'язку, який важливий для структури білка. Schuiz et al., цит. вище, змогли з'єднати зазначені вище групи 1 і 2. Потрібно також відзначити, що Туr, у силу його можливості утворювати водневий зв'язок, має суттєву подібність до Ser і Thr, і т.п. Консервативні заміни амінокислот відповідно до даного винаходу, наприклад, такі, які зазначені вище, відомі в техніці, і очікується, що після заміни амінокислот біологічні і структурні властивості поліпептиду зберігаються. Більшість делецій і замін відповідно до даного винаходу є такими, що не 27 75564 28 викликають радикальних змін у характеристиках 1989]. білкової або пептидної молекули. "ХарактеристиОдержання в такий спосіб білка, що взаємодіє ки" не є винятковим визначенням як замін у втоз каспазою-8, або альтернативної нуклеотидної ринній структурі, наприклад, а-спіралі на бетапослідовності, що кодує той же поліпептид, але що складку, а також змін біологічної активності, навідрізняється від природної послідовності внасліприклад, зв'язування з каспазою-8, так та/або оподок замін, дозволених відомим виродженням генесередкування дії каспази-8 на загибель клітин. тичного коду, можна досягти за допомогою сайтсПриклади одержання замін амінокислот у білпрямованого мутагенезу ДНК, яка кодує ках, які можна використовувати для одержання отриманий раніше аналог або нативний варіант аналогів білків, що взаємодіють із каспазою-8, для білка, що взаємодіє з каспазою-8. Сайтспецифічвикористання в даному винаході включають стадії ний мутагенез дозволяє одержати аналоги за добудь-яких відомих способів, наприклад, подані [в помогою застосування специфічних олігонуклеопатентах США RE 33653, 4959314, 4588585 і тидних послідовностей, що кодують послідовність 4737462, Mark et al.; 5116943, Koths et al.; 4965195, ДНК потрібної мутації, а також достатнє число суNamen et al.; 4879111, Chong et al.; і 5017691, Lee сідніх нуклеотидів, щоб одержати послідовністьet al.]; а [в патенті США №4904584 (Show et al.)] затравку достатнього розміру і складності послідоподані лізинзаміщені білки. вності для утворення стійкого дуплекса по обидва Мається на увазі, що до обсягу винаходу, крім боки пересічної області делеції. Як правило, краописаних вище консервативних замін, що, по суті, щим є праймер довжиною приблизно в 20-25 нукне змінюють активність білка, що взаємодіє з каслеотидів, приблизно з 5-10 комплементуючими пазою-8, входять або консервативні зміни або нуклеотидами з кожного боку змінюваної послідоменш консервативні і більш випадкові заміни, що вності. Взагалі, метод сайтспецифічного мутагенепризводять до зростання біологічної активності зу добре відомий у техніці, наприклад, із таких аналогів білків, що взаємодіють із каспазою-8. публікацій, як [Adelman et al., DNA, 2:183 (1983)], Коли фахівець у даній області техніки підтвервключеної в даний опис в якості посилання. джує точний вплив заміни або делецій, варто мати Як буде зрозуміло, у методі сайтспецифічного на увазі, що дія заміни(замін), делеції(ій) і т.п. буде мутагенезу, як правило, використовується фагооцінюватися за допомогою звичайних аналізів вий вектор, що існує як в одноланцюговій, так і в зв'язування і загибелі клітин. Скринінг із викорисдволанцюговій формі. Типові вектори, корисні при танням такого стандартного дослідження не вклюсайтспрямованому мутагенезі, включають такі векчає унікальних експериментів. тори, як фаг Μ13, наприклад, як описано в роботі Прийнятними аналогами, що взаємодіють із [Messing et al., Third Cleveland Symposium on каспазою-8, є аналоги, що зберігають щонайменMacromolecules and Recombinant DNA, Editor A. ше здатність взаємодіяти з каспазою-8 і, у силу Walton, Elsevier, Amsterdam (1981)], включеної в цього, опосередковувати активність каспази-8 у даний опис у якості посилання. Такі фаги доступні внутрішньоклітинному метаболізмі, або модулювакомерційно, і їхнє застосування добре відомо фати активність самої каспази-8. При цьому можна хівцям у цій області техніки. З іншого боку, для одержати аналоги, що мають так звану домінантодержання одноланцюгової ДНК можна викориснегативну дію, а саме, аналог, що дефектний по товувати плазмідні вектори, що містять одноланвідношенню до зв'язування з каспазою-8 або нацюговий фаговий оріджин реплікації [Veria et al., ступної передачі сигналу, або по відношенню до Meth. Enzymol., 153:3,1987]. іншої активності після такого зв'язування. Такі Взагалі, сайтспрямований мутагенез відповіданалоги можна використовувати, наприклад, для но до описаного вище здійснюють за допомогою інгібування цитотоксичної дії каспази-8 або для її одержання спочатку одноланцюгового вектора, що посилення, у залежності від того, потрібно підсивключає у свою послідовність ДНК, що кодує релити загибель клітин або їхню виживаність, і в залевантний поліпептид. Олігонуклеотидний прайлежності від того, яка з цих активностей є головмер, що містить потрібну мутовану послідовність, ною, що модулюється взаємодією білка, який одержують синтетично за допомогою автоматизовзаємодіє з каспазою-8, і каспазою-8 (див. вище), і ваного синтезу ДНК/олігонуклеотидів. Потім цей це за допомогою таких аналогів, які конкурують із праймер відпалюють з одноланцюговим вектором, природним білком, що взаємодіє з каспазою-8, за що містить послідовність білка, і піддають впливу зв'язування або взаємодію з каспазою-8. ферментів, які полімеризують ДНК, таких як фрагНа генетичному рівні такі аналоги одержують, мент Кленова полімерази І Е. соlі, для завершення як правило, за допомогою сайтспрямованого мутасинтезу ланцюга, що містить мутацію. Таким чигенезу нуклеотидів у ДНК, що кодує білок, який ном, мутована послідовність і другий ланцюг місвзаємодіє з каспазою-8, причому за допомогою тять потрібну мутацію. Потім цей гетеродуплексцього продукується ДНК, що кодує аналог, і потім у ний вектор використовують для трансформації рекомбінантній клітинній культурі синтезується відповідних клітин, таких як клітини Е. соlі JM101, і ДНК і експресується поліпептид. Аналоги, як правідбирають клони, що включають рекомбінантні вило, виявляють таку ж або якісно підвищену біовектори, що містять порядок мутованої послідовлогічну активність у порівнянні з білком, що зустріності. чається в природі, [Ausubel et al, "Current Protocols Після того, як такий клон відібрали, послідовin Molecular Biology", Greene Publications and Viley ність білка, що взаємодіє з каспазою-8, можна виInterscience, New York, NY, 1987-1995; Sambrook et тягти і помістити у відповідний вектор, як правило, al., "Molecular Cloning: A Laboratory Manual", Cold вектор переносу або експресуючий вектор типу, Spring Harbor Laboratory, Cold Spring Harbor, NY, який можна використовувати для трансфекції від 29 75564 30 повідного хазяїна. фрагменти будь-яких білків, що взаємодіють із Відповідно, ген або нуклеїнову кислоту, що кокаспазою-8, або їхніх ізоформ), можна одержати дує білок, який взаємодіє з каспазою-8, також мотак, як зазначено вище у відношенні аналогів білжна виявити, отриманий він та/або модифікований ка, що взаємодіє з каспазою-8. Відповідними фраin vitro, in situ та/або in vivo, шляхом використання гментами білка, що взаємодіє з каспазою-8, є фравідомих методів ампліфікації ДНК або РНК, таких гменти, що зберігають здатність білка, що як PCR і хімічний синтез олігонуклеотидів. PCR взаємодіє з каспазою-8, і можуть опосередковувадає можливість ампліфікації послідовностей ДНК ти біологічну активність каспази-8 або інших білків, за допомогою повторюваних полімеразних реакцій безпосередньо або непрямо зв'язаних із каспазоюДНК. Таку реакцію можна використовувати як за8. Відповідно, можна одержати фрагменти білка, міну клонування; усе, що потрібно - це знання нукщо взаємодіє з каспазою-8, які мають домінантнелеотидної послідовності. Для того, щоб здійснити гативну або домінантпозитивну дію, як вказувалоPCR, створюють праймери, комплементарні до ся вище по відношенню до аналогів. Слід зазначипослідовності, що представляє інтерес. Потім ти, що такі фрагменти представляють особливий праймери одержують за допомогою автоматизоклас аналогів згідно з винаходом, а саме, вони ваного синтезу ДНК. Оскільки можна створити визначаються як частини білків, що взаємодіють із праймери для гібридизації будь-якої частини гена, каспазою-8, утворені від повної послідовності білумови можна створити такі, що можна допустити ка, що взаємодіє з каспазою-8 (наприклад, від попомилки при спарюванні комплементарних основ. слідовності будь-якого з білків, що взаємодіють із Ампліфікація таких помилкових областей може каспазою-8, або їхніх ізоформ), причому кожна призвести до синтезу мутагенного продукту, і притака частина або фрагмент має будь-яку із зазначому результатом є генерація пептиду з новими чених вище активностей. Такий фрагмент може властивостями (тобто сайтспрямований мутагеявляти собою, наприклад, пептид. нез). Див. також Ausubel, цит. вище, гл.16. Крім Подібним чином, похідні можна одержати того, за допомогою сполучення синтезу комплемешляхом стандартних модифікацій бічних груп однтарних ДНК (кДНК) із використанням зворотної ного або декількох амінокислотних залишків білка, транскриптази з PCR можна використовувати РНК що взаємодіє з каспазою-8, його аналогів або фрау якості вихідної речовини для синтезу позаклітингментів, або за допомогою кон'югації білка, що ного домену рецептора пролактину без клонуванвзаємодіє з каспазою-8, його аналогів або фрагня. ментів, з іншою молекулою, наприклад, антитілом, Крім того, можна створити праймери PCR для ферментом, рецептором і т.п., що добре відомо в внесення нових сайтів рестрикції або інших ознак, техніці. Відповідно, використовуваний тут термін таких як кодони термінації, у кінці ампліфікованого "похідні" охоплює похідні, що можна одержати з сегмента гена. Таке розміщення сайтів рестрикції функціональних груп, що зустрічаються в бічних по 5' і 3'-кінцям ампліфікованої послідовності гена ланцюгах залишків, або N- або С-кінцевих груп, дозволяє створити сегменти гена, що кодує білок, способами, відомими в техніці, і усі вони входять що взаємодіє з каспазою-8, або його фрагмент, до обсягу винаходу. Похідні можуть мати хімічні для лігування інших послідовностей та/або сайтів групи, такі як вуглеводні або фосфатні залишки, за клонування з векторами. умови, що така фракція має таку ж або більш виPCR та інші способи ампліфікації РНК та/або соку біологічну активність, як білки, що взаємодіДНК добре відомі в техніці і можуть застосовувають із каспазою-8. тися відповідно до даного винаходу без зайвого Наприклад, похідними можуть бути аліфатичні експериментування на основі положень і вказівок, ефіри, отримані по карбоксильних групах, аміди, приведених тут. До відомих способів ампліфікації отримані по карбоксильних групах за допомогою ДНК або РНК відносяться, але не обмежуються взаємодії з аміаком або первинними або вторинперерахованим, полімеразна ланцюгова реакція ними амінами, N-ацильні похідні вільних аміногруп (PCR) і родинні способи ампліфікації [див., наприамінокислотних залишків, утворені за допомогою клад, патенти США №№ 4683195, 4683202, ацильних груп (наприклад, алканоїльних або кар4800159, 4965188, Mullis et al.; 4795699 і 4921794, боциклічних ароїльних груп), або О-ацильні похідні Tabor et al.; 5142033, Innis; 5122464, Wilson et al.; вільних гідроксильних груп (наприклад, серильних 5091310, Innis; 5066584, Gyllensten et al.; 4889818, або треонільних залишків), утворені за допомогою Gelfand et al.; 4994370, Silver et al.; 4766067, ацильних груп. Biswas; 4656134, Ringold; і Innis et al., eds., PCR Термін "похідні" призначений для позначення Protocols: A Guide to Method and Applications], і тільки таких похідних, в яких немає заміни однієї ампліфікація, опосередковувана РНК, при якій у амінокислоти на іншу з числа двадцятьох амінокиякості матриці для синтезу дволанцюгової ДНК слот, що зустрічаються звичайно в природі. використовується антисмислова РНК до послідовЯк описано вище, аналізи з розчепленням моності-мішені [патент США №5130238, Maiek et al., жна використовувати для визначення того, чи роторгова назва NASBA]; та імуно-PCR, що з'єднує зщеплюється білок, що взаємодіє з каспазою-8, застосування ампліфікації ДНК із міченням антитіл каспазою-8. Розділення за розміром розщеплених [Ruzicka et al., Science, 260:487 (1993); Sano et al., фрагментів приблизно вказує місце розташування Science, 258:120 (1992); Sano et al., Biotechniques, сайта розчеплення. 9:1378 (1991)], що включені в даний опис у якості Сайт розчеплення також можна визначити за посилань. допомогою одержання делеційних мутантів досліАналогічним чином, біологічно активні фрагджуваного білка і дослідженням кожного делеційменти білків, що взаємодіють із каспазою-8 (тобто ного мутанта на його схильність до розчеплення 31 75564 32 каспазою-8, як описано вище. Делеційні мутанти розчеплення каспазою-8, потім можна досліджуваможна сконструювати за допомогою PCRти, повторивши ті ж прийоми, але відбираючи клонування потрібних фрагментів досліджуваного більш дрібні зони, обрані з послідовності ідентифібілка з використанням у якості матриці послідовкованого пептиду. ності ДНК клону, що кодує зазначений досліджуФактичний сайт розчеплення пептидів повинен ваний білок. Фрагменти, ампліфіковані методом відповідати послідовності розчеплення каспазою PCR, потім можна клонувати у експресуючі вектоXXXD [див. Boldin et al., Cell, 1996, і Nicholson et ри за рахунок чого повинні забезпечуватися старal., Killer caspases. Trends in Biochem. Sci., 22, 299товий кодон ATG і, переважно, послідовність Козак 306, 1997]. Внесок кожної амінокислоти в пептид [Kozak, M., Nucleic Acids Res., 12, p.857-72, 1984]. можна оцінити за допомогою одержання пептидів, Інші подробиці експресування білків можна знайти мутованих по одній з амінокислот, і дослідженням в зазначених вище зведеннях від Qiagen, що відтаких мутованих пептидів на схильність до розченосяться до мічених гістидином білків, а також плення каспазою-8. Мутовану амінокислоту, перевзагалі до експресії білків. Іншим посиланням, що важно, заміняють амінокислотою, обраною з групи містить дані про експресію білків, є зазначені вище заряджених неполярних амінокислот [див. Current Protocols, і особливо глава 16 в них. Lehninger, Biochemistry, Worth/ NY, 1979, chapter Таким чином, сайт розчеплення досліджувано4], і найбільш переважно, обраних серед гліцину го білка можна визначити за допомогою одержанабо аланіну. ня його делеційних мутантів і визначення найШляхом мутації критичних амінокислот можна меншого такого делеційного мутанта, який одержати пептиди, що зв'язуються з каспазою-8, відщеплюється каспазою-8. але несприйнятливі до розчеплення нею. Зв'язуПри іншому способі ідентифікації сайта розчевання можна перевірити за допомогою розділення плення використовують пептиди, що утворюються за розміром комплексів пептиду і каспази-8 в умовідповідно до передбачуваної білкової послідоввах без денатурації з використанням електрофоності досліджуваного клону. Пептиди можна синрезу в акриламідному гелі. тезувати хімічним способом, наприклад, так, як Також можна сконструювати модифіковані педокладно описано в [Bodanszky and Bodanszky, птиди, здатні до взаємодії з активним цистеїном "The practice of peptide synthesis", Springer, New каспази-8, при цьому вони ковалентно зв'язуються York, ISBN 0-387-13471-9, і в Bodanszky, "The із зазначеним цистеїном. Для цієї мети можна виprinciples of peptide synthesis". Springer, New York, користовувати реагенти, що взаємодіють із тіольISBN 0-387-12359-4]. Синтез потрібного пептиду ними групами, відомі фахівцям в області хімії. Натакож доступний від деяких комерційних компаній, приклад, із SH-групою активного цистеїну каспазинаприклад, SynPep Corp., Dublin, CA, USA, і 8 можуть реагувати реагенти, що містять тіольную California Peptide Research, Inc., Napa, CA, USA. групу. Ковалентний зв'язок S-S, що утворився, Пептиди також можна одержати або у вигляді гібможна розщепити за допомогою відновлення, наридів з іншими білками або окремо за допомогою приклад, у фізіологічних умовах у цитозолі або з експресії рекомбінантної ДНК, що кодує їх, що довикористанням реагенту, що відновлює S-S-групи, кладно описано в цитованій вище главі 16 Current такого як дитіотреїтол (DTT). Реагенти, що взаєProtocols. модіють із тіольними групами, можна також незвоДля того, щоб використовувати пептиди для ротньо приєднати до каспази-8. Такі реагенти мокартування сайта розчеплення досліджуваного жна легко знайти серед крос-лінкерів, здатних білка, передбачувану амінокислотну послідовність взаємодіяти з тіольними групами, відомих у технізазначеного білка розділяють на зони, і синтезуці, таких як описані на с 0-90 і далі в каталозі ють пептид, що відповідає кожній зоні. Крім того, PIERCE Life 3ciences (PIERCE, Rockford, IL, U3A). синтезують пептиди, що містять приблизно полоВідповідними групами, що реагують із тіольвину амінокислот однієї із зон і містять на стику ними групами, є, наприклад, піридилдитіо-, йодатакож приблизно половину амінокислот безпосецетамідо- або малеімідогрупи. Ці групи можна редньо прилягаючої області, для того, щоб перекприєднати до пептиду через лінкери, що містять рити границю між двома зонами. Зони містять від ланцюги необов'язково ненасичених аліфатичних 5 до 100 амінокислот, переважно - від 9 до 40 амівуглеводнів, -О-, -S-, -NH- або ароматичні групи. нокислот, і найбільш переважно - від 20 до 30 аміЛінкери, необов'язково, можуть містити замісники. нокислот. Потім повний набір пептидів випробують Тіолреактивні групи можна приєднати до пептак, як описано вище, на схильність до розчеплентиду шляхом хімічного синтезу, як відомо в техніці; ня каспазою-8. Отримані пептиди можна очистити і функціональні групи пептиду можна ввести до взаодержати у великій кількості. Після реакції розчепємодії з відповідними функціональними групами лення їх можна потім проаналізувати безпосередмолекул лінкера з тіолреактивною групою. Наприньо методом електрофорезу в поліакриламідному клад, залишок лізину, що присутній в пептидній гелі в присутності SDS і Уф-детекції або візуалізапослідовності або введений до неї з метою ствоції за допомогою забарвлювання, наприклад, із рення відповідної функціональної групи, якою, у використанням кумасі синього. З іншого боку, для випадку лізину, є епсилон-аміногрупа, можна ввесбільш легкого виявлення пептиди можна позначити до взаємодії з гетеробіфункціональним крости, наприклад, шляхом ізотопного мічення кінців лінкером, таким як N-гамма[див., наприклад, Shevchenko A. et al.. Rapid малеімідобутирилоксисукцинімідефір, і створити Commun. Mass Spectrum, 11, p.1015-24, 1997]. пептид, де зазначена епсилон-аміногрупа реагує з Після завершення скринінгу пептидів, описаноN-гідроксисукцинімідною групою крос-лінкера, у го вище, пептид, що, як тепер відомо, містить сайт той час як малеімідогрупа крос-лінкера не вступає 33 75564 34 в реакцію і може, після контакту з цистеїном касрні елементи, які можуть підсилити транскрипцію пази-8, який зустрічається, коли зазначений пепвід вищевказаних промоторів. тид специфічно зв'язується із зазначеною каспаЧасто інтронні послідовності також містять зою-8, реагувати з тіольною групою зазначеного транскрипційні або транскрипційні контрольні посцистеїну і за допомогою цього інактивувати зазналідовності, які додають експресії тканиноспецифічену каспазу-8. чність. Отже, коли білок винаходу потрібно ексПозицію в пептиді, використовувану для взаєпресувати специфічно до тканини, може бути модії з ним тіолреактивного реагенту, можна вибвигідним використання таких інтронних послідоврати так, щоб вона була поблизу амінокислоти (як ностей. Прикладом інтрону, що містить тканиносправило, аспарагінової кислоти), де відбувається пецифічні енхансерні елементи, є еритроїдспецирозчеплення каспазою-8. Лінкер, за допомогою фічний енхансер, розташований в інтроні 8 гена 5якого тіолреактивна група зв'язується з пептидом, амінолевулинатсинтази 2 людини [Surinya et al., J. може змінюватися по своїй довжині, наявності поBiol. Chem., 273, p.16798-809, 1998], а обговорення лярних груп, таких як гідрокси, заряджених груп, принципів посилення продукування білків із викотаких як нітро- і сульфогрупи, і ароматичних груп, ристанням інтронних послідовностей, поряд із таких як феніленовий або фенільний залишок. Такі прикладами інтронних послідовностей, описані варіації в будові лінкера дозволять одержати реа[Huang et al., Nucleic Acids Res., 18, p.937-47, гент, що містить пептид і зв'язану з ним ковалент1990]. но тіолреактивну групу, що специфічно зв'язується Транскрипційні термінаційні послідовності і сиз каспазою-8 і ефективно реагує з тіольною групою гнали поліаденілювання можна додати до 3'-кінця її активного цистеїну. ДНК, що кодує білок, який потрібно експресувати. Досліджуваний білок або його пептидний фраТакі послідовності можна знайти в багатьох або гмент, можна також охарактеризувати за допомонавіть у більшості генів. Вигідно використовувати гою введення зазначеного білка або пептиду в сигнал поліаденілювання SV40 [Schek et al., Моl. клітину ссавця і вимірювання його дії на реагенти, Cell. Biol., p.5386-93, і приведені там посилання]. що викликають апоптоз в зазначеній клітині. Кращим вектором експресії білка в клітині ссаЕкспресію білка або пептиду в клітині ссавця вця є вектор pcDNAHis (Invitrogen), що містить можна здійснити шляхом вбудовування ДНК, що промотор CMV для управління експресією гена, кодує досліджуваний білок, у вектор, що містить який кодує потрібний білок. Іншими векторами, що промотор, необов'язково, інтрону послідовність і можна використовувати, є вектори pCDNA3 або донорні/акцепторні сигнали сплайсингу, і також, pMPSVEH. Ці вектори містять промотори CMV і такий, що необов'язково містить послідовність MPSV, відповідно. термінації. Ці методи в цілому описані в зазначеВикористовуючи рекомбінантну експресію доних вище [Current Protocols, глава 16]. сліджуваного білка, зазначений білок тепер можна Вищевказані промоторна, інтронна і термінаоцінити по його дії на сигнал апоптозу, опосередційна послідовності операбельні в клітинах ссавця. ковуваний каспазою-8. З цією метою можна індуПромотор, переважно, є сильним промотором, кувати апоптоз або шляхом надекспресії білка, що таким як зазначені вище промотори RSV, CMV або індукує апоптоз, такого як внутрішньоклітинний MPSV. Промотор також може представляти собою домен CD120а, внутрішньоклітинний домен CD95, ранній промотор SV40 [Everett et al., Nucleic Acids білок Mort-1, каспаза-8 або їхні еквіваленти, або Res., 11, p.2447-64, 1983, і зазначені там посиланшляхом активації сигналу апоптозу за допомогою ня] або клітинний промотор, такий як промотор ініціації CD120а, CD95, TRAMP/DR3 або еквівалебета-актин або промотор ELF-1 [Tokushige et al., J. нтного рецептора. Активації рецептора можна доVirol. Methods, 64, p.73-80, 1997]. Можна також могтися або за допомогою приведення рецепторів використовувати гібридний промотор, такий як у контакт із лігандом, або за допомогою поперечгібрид lac-оператора і ELF-1 альфа-промотору ного зшивання рецепторів з антитілами, переважлюдини, описаний [Edamatsu et al. (Gene, 187, но - поліклональними антитілами [див. Engelmann p.289-94, 1997)], гібридний промотор CMV-бетаet al., J. Biol. Chem., 265, p.14497-504, 1990]. актин, описаний [Akagi et al., Kidney Int., 51, p.1265Незважаючи на те, що, взагалі, ініціація реце9, 1997], або гібрид tet-операторних послідовносптора, подібного CD120а, вимагає додавання інгітей і промотору CMV [Furth et al., PNAS/ 91, 9302бітору синтезу білка, подібного до циклогексіміду, 6,1994, і зазначені там посилання]. для того, щоб домогтися сильного сигналу для Інтронні послідовності, що можна вбудовувати апоптозу, надекспресії рецепторних внутрішньоку вигляді повних послідовностей, тобто такі, що літинних доменів або білків, залучених до трансвключають сайти донора і акцептора сплайсувандукції сигналу апоптозу, не відбувається [див. ня, можна вбудовувати у кодуючу послідовність Boldin et al., Cell, 85, p.803, 1996]. Детекція апоптобілка, який потрібно експресувати. Вбудовування зу, час інкубації та інші подробиці і параметри цьотаких інтронних послідовностей може підсилити го аналізу описані в цитованій вище роботі Boldin стійкість РНК і, таким чином, збільшити продукуet al. вання потрібного білка. Незважаючи на те, що, в Загибель клітин, що експресують досліджувапринципі, прийнятні інтронні послідовності можна ний білок, у порівнянні з клітинами, які не експревибрати серед будь-яких інтронів, що містять ген, сують білок, можна оцінити будь-яким із багатьох кращими інтронними послідовностями є інтрон способів, таким як спосіб на основі фрагментації бета-актину, інтрон SV40 та інтрон рецептора до ДНК або виявлення апоптозспецифічних антигенів TNF р55. та епітопів, а реагенти і протоколи для виявлення Інтронна послідовність може містити енхансеапоптозу у формі набору доступні від вищевказа 35 75564 36 ної компанії Boehringer Mannheim і від інших комЯкщо білок або пептид експресований у випаній. гляді злитого білка, бажано відщепити партнера по Загибель клітин можна також визначити шлягібридизації перед застосуванням білка для одерхом оцінки морфологічного стану клітин. Загибель жання антитіл проти партнера по гібридизації. Роклітин внаслідок апоптозу характеризується хвизчеплення партнерів по гібридизації і виділення лястою мембраною в клітин і зморщуванням клітин потрібного білка описано в цитованих вище при відсутності лізису. [Current Protocols in Molecular Biology, глава 16]. Вигідно експресувати у клітині ссавця репорВектори, протоколи і реагенти для експресії та терний ген для того, щоб забезпечити маркер для очищення рекомбінантних білків, злитих із білком успішної трансфекції. Тому що сама процедура зв'язування мальтози, також доступні комерційно. трансфекції призводить до загибелі деяких клітин, При одержанні пептиду винаходу може бути у тому числі клітин, які не є трансфікованими, вигібажано не видаляти партнера по гібридизації, тодно оцінювати тільки ті клітини, що є трансфіковаму що злитий білок може стимулювати продукуними. Кращим репортерним геном для цієї мети є вання антитіл проти пептиду. Як правило, це буде ген lacZ, який легко виявляється шляхом інкубації доречно, коли одержують антитіла з пептидів, дотрансфікованих клітин із Xgal або подібним реагевжина яких менше 50 амінокислот. нтом, що вказує на активну бета-галактозидазу. Як відзначено також вище, пептид також можОднак можна використовувати будь-який інший на синтезувати хімічними способами, відомими в репортерний ген, переважно ген, продукти якого хімії. легко виявляються з використанням простої реакОдержання поліклональних антитіл проти білції забарвлювання, результати якої можна оцінити ків [описане в главі 2 Current Protocols in з використанням мікроскопа. Наприклад, для безImmunology, Wiley and Sons Inc.]. При одержанні посередньої детекції без реакції фарбування можантитіл проти пептидів можуть знадобитися деякі на використовувати білок зеленої флуоресценції. зміни в протоколі, як правило, внаслідок більш Цей репортерний ген вимагає застосування флуонизької антигенності пептидів у порівнянні з білкаресцентного мікроскопа. ми. Одержання поліклональних антитіл проти пепТаким чином, розглядаючи тільки клітини, що є тидів описано в цитованих вище [Current Protocols трансфікованими, тобто які експресують репортеin Immunology, глава 9]. рний ген, і підраховуючи відсоток клітин, що деМоноклональні антитіла можна одержати з Вмонструють морфологію, характерну для апоптоклітин, узятих із селезінки або лімфовузлів імунізу, можна оцінити дію певного трансфікованого зованих тварин, зокрема, пацюків або мишей, клону і експресованого ним білка на апоптоз. шляхом злиття з іморталізованими В-клітинами в Клітини ссавця являють собою, переважно, умовах, сприятливих для росту гібридних клітин. HeLa або ембріональні клітини нирки людини Для злиття з В-клітинами мишей кращою є клітин(НЕК) 293-Т. Трансфекцію, переважно, здійснюють на лінія Ag-8. кальційфосфатним способом, як описано в цитоМетоди одержання моноклональних антитіл ваних вище Current Protocols. Морфологію клітин описані в багатьох статтях і книгах, таких як цитооцінюють через 1-150 годин після трансфекції, вані вище [Current Protocols in Immunology]. У главі переважно - через 4-35 годин, і найбільш перева9 цього видання описується імунізація тварин за жно - через 20 годин після трансфекції. допомогою пептидів. Клітини селезінки або лімфоОдержання антитіл вузлів цих тварин можна використовувати для Поліклональні антитіла можна одержати в одержання моноклональних антитіл так само, як кроликах, курках, мишах, пацюках, вівцях або поклітини селезінки або лімфовузлів тварин, імунізодібних ссавцях. Для одержання антитіл проти білваних білками, як це описано в цьому виданні в ка або пептиду винаходу в клітинах ссавця одерглаві 2. жують білок або пептид, як описано вище, за Методи, використовувані при одержанні монодопомогою технології рекомбінантних ДНК. Білок клональних антитіл, також описані в [Kohler and також можна одержати в клітинах бактерій або Milstein, Nature, 256,495-497, і в USP 4376110]. комах, як докладно описано в цитованих вище Для одержання антитіл із банку генів антитіл [Current Protocols, глава 16]. людини гіперваріабельні області генів заміняють Білок або пептид очищають від клітин, в яких майже випадковими послідовностями, як описано його було продуковано. Способи очищення білків в USP 5840479. Такі антитіла є кращими, якщо відомі фахівцям у цій області техніки та описані імунізацію тварини даним пептидом або білком докладно, наприклад, у цитованих вище [Current важко зробити. Деякі структури є слабкоімуногенProtocols in Molecular Biology, глава 16, і в Current ними і можуть залишатися такими, незважаючи на Protocols in Protein Science,Wiley and Sons Inc., додавання ад'ювантів і зв'язок з іншими білками в глави 5 і 6]. Білок можна вигідно одержати у вигібридні конструкції. Антитіла, описані в USP гляді гібрида з іншим білком, таким як глутатіон-S5840479, також є кращими, якщо потрібно викоритрансфераза, або подібним білком, або послідовстання антитіл із структурою, подібною до антитіл ністю-міткою, такою як послідовність гістидинової людини, наприклад, коли потрібні антитіла з низьмітки. Використання гібридних або мічених білків кою імуногенністью у людей. спрощує процедуру очищення, що докладно опиЯк тільки відповідні антитіла ідентифіковані, сано в цитованих вище [Current Protocols in може знадобитися зміна їхніх властивостей. НаMolecular Biology, глава 16], і в інструкціях для заприклад, при одержанні можна досягти більш визначеного вище набору для експресії та очищення соких виходів із химерними антитілами. Химерні білків із гістидиновою міткою від Qiaqen. антитіла, де константні області замінені на конста 37 75564 38 нтні області антитіл людини, також можуть знадочеплення з використанням ферментів, таких як битися, коли бажано, щоб антитіла мали низьку папаїн (для одержання Fab-фрагментів) або пепімуногенність у людей. Одержання химерних антисин (для одержання Р(аb')2-фрагментів). тіл описано в багатьох публікаціях, наприклад, у Про антитіло говорять, що воно "здатне до [Cabilly et al., PNAS, 81, p.3273, 1984; Morrison et зв'язування" із молекулою, якщо воно здатне спеal., PNAS, 81, p.6851, 1984; Boulianne et al., Nature, цифічно взаємодіяти з молекулою, і за допомогою 312, p.643, 1984; EP 125023, EP 171496, EP цього молекула зв'язується з антитілом. Термін 173494, EP 184187, WO 86/01533, WO 87/02671 і в "епітоп" відноситься до тієї частини будь-якої моHarlow and Lane, "Antibodies: A Laboratory Manual", лекули, здатної зв'язуватися антитілом, яку це Cold Spring Harbor Laboratory, 1988]. антитіло може впізнати. Епітопи або "антигенні Іншим типом антитіл є антиідіотипічні антитіла. детермінанти", як правило, складаються із хімічно Антиідіотипічні (анти-Id) антитіла є антитілами, які активних груп молекул на поверхні, таких як амінорозпізнають унікальні детермінанти, як правило, кислоти або бічні ланцюги цукрів, і мають специфіасоційовані із антигензв'язуючим сайтом антитіла. чні тривимірні структурні характеристики, так само Антитіла проти Id можна одержати за допомогою як і специфічні характеристики за зарядом. імунізації тварини того ж виду і генетичного типу "Антиген" являє собою молекулу або частину (наприклад, штаму мишей), що і джерело mAb, до молекули, здатну зв'язуватися антитілом, яке таякого одержують анти-Id. Імунізована тварина букож здатна індукувати тварина до продукування де впізнавати і реагувати на ідіотипічні детермінаантитіл, здатних до зв'язування з епітопом цього нти імунізуючого антитіла шляхом продукування антигену. Антиген може мати один або декілька антитіл до цих ідіотипічних детермінантів (анти-Id епітопів. Специфічна взаємодія, згадане вище, антитіла). Див., наприклад, патент СІЛА № означає, що антиген буде взаємодіяти з високою 4699880, включений у даний опис у якості посиселективністю зі своїм відповідним антитілом, а не лання. з багатьма іншими антитілами, які можуть виклиАнти-Id антитіла також можна використовувати катися іншими антигенами. в якості "імуногена" для індукції імунної реакції ще Антитіла, у тому числі, фрагменти антитіл, кой у іншої тварини, продукуючи так звані анти-антирисні в даному винаході, можна використовувати Id антитіла. Анти-анти-Id можуть бути епітопно для якісної і кількісної детекції білка даного винаідентичні до вихідних mAb, які індукували анти-Id. ходу, що взаємодіє з каспазою-8. Це можна здійсТаким чином, використовуючи антитіла до ідіонити імунофлуоресцентними методами з використипічним детермінантів mAb, можна ідентифікуватанням флуоресцентно помічених антитіл (див. ти інші клони, що екс пресують антитіла ідентичної нижче) разом із детекцією оптичними методами, специфічності. методом потокової цитометрії або флюорометриВідповідно, mAb, отримані проти білка, що чних методів. взаємодіє з каспазою-8, його аналогів, фрагментів Антитіла (або їхні фрагменти), корисні в даноабо похідних даного винаходу можна використовуму винаході, можна використовувати для гістології вати для індукції анти-Id антитіл у відповідних тваз методами імунофлуоресценції або імуноелектрин, таких як миші BALB/c. Клітини селезінки таких ронної мікроскопії для детекції in situ білка даного імунізованих мишей використовують для одервинаходу, що взаємодіє з каспазою-8. Детекцію in жання анти-Id гібридом, що секретує анти-Id mAb. situ можна виконати за допомогою узяття гістологіДалі, анти-Id mAb можна поєднувати з носієм, тачного зразка в пацієнта і надання міченого антитіким як гемоціанін лімфи равлика (KLH) і викорисла даного винаходу до такого зразка. Антитіла товувати для імунізації інших мишей BALB/c. Си(або їхні фрагменти), переважно, надають за дороватка цих мишей буде містити анти-анти-Id помогою нанесення або нашарування мічених анантитіла, які мають властивість зв'язуватися з вититіл (або фрагментів) на біологічний зразок. Західними mAb, специфічними для епітопу вищевкастосовуючи таку процедуру, можна визначити не заного білка, що взаємодіє з каспазою-8, або його тільки наявність білка, що взаємодіє з каспазою-8, аналогів, фрагментів або похідних. але також його розподіл у тканині, що перевіряТаким чином, анти-Id mAb мають свої власні ється. Застосовуючи даний винахід, фахівці легко ідіотипічні епітопи або "ідіотопи", схожі за будовою представлять, що будь-який із численних гістолодо оцінюваних епітопів. гічних методів (таких як процедури забарвлюванТермін "антитіло" також означає як інтактні ня) можна модифікувати для того, щоб домогтися молекули, і їхні фрагменти, такі, як, наприклад. такої детекції in situ. Fab і F(ab')2, здатні зв'язуватися з антигеном. ФраТакі аналізи на білок даного винаходу, що взагменти Fab і F(ab')2, позбавлені Fc-фрагмента інємодіє з каспазою-8, як правило, включають інкутактного антитіла, швидко виводяться з кровообігу бацію біологічного зразка, такого як біологічна ріі можуть мати менше нетканиноспецифічного зв'ядина, тканинний екстракт, свіжезібрані клітини, такі зування, ніж інтактне антитіло [Wahl et al., J. Nucl. як лімфоцити або лейкоцити, або клітини, інкубоMed., 24:316-325 (1983)]. вані в культурі тканини, в присутності мічених анВарто мати на увазі, що Fab і F(ab')2 і інші титіл, що виявляються, здатних ідентифікувати фрагменти антитіл, корисні в даному винаході, білок, що взаємодіє з каспазою-8, і детекцію антиможна використовувати для детекції і кількісного тіл будь-яким із багатьох способів, відомих у техвизначення білка, що взаємодіє з каспазою-8, у ніці. відповідності зі способами, описаними тут для моБіологічний зразок можна обробити за допомолекул інтактних антитіл. Такі фрагменти, як правигою твердофазної основи або носія, таких як нітло, одержують за допомогою протеолітичного розроцелюлоза, або інших, твердої основи або носія, 39 75564 40 здатних іммобілізувати клітини, частинки клітин порівняння з отриманими таким же способом стаабо розчинні білки. Потім основу або носій можна ндартами. промити відповідними буферами з наступною обДетекцію можна здійснити з використанням робкою міченими антитілами, що детектуються, багатьох інших імуноаналізов. Наприклад, за довідповідно до даного винаходу, як зазначено випомогою мічення антитіл або фрагментів антитіл ще. Потім твердофазну основу або носій можна радіоізотопами можна виявити R-РТРазу, застосопромити буфером другий раз і видалити незв'язані вуючи радіоімуноаналіз (RIA). Гарний опис RIA антитіла. Потім звичайними способами можна виможна знайти в ["Laboratory Techniques and значити кількість зв'язаної мітки на зазначеній осBiochemistry in Molecular Biology", Work, T.S., et al.. нові або носії. North Holland Publishing Company, NY (1978), осоПід "твердофазною основою", "твердофазним бливо в главі за назвою "An Introduction to носієм", "твердою основою", "твердим носієм", Radioimmune Assay and Related Techniques", "основою" або "носієм" маються на увазі будь-якої Chard, Т., включених у даний опис у якості посиоснови або носії, здатні зв'язувати антиген або лань]. Радіоактивні ізотопи можна виявити за доантитіла. Добре відомими основами або носіями є помогою таких засобів, як лічильник гамма-квантів скло, полістирол, поліпропілен, поліетилен, декабо сцинтиляційний лічильник, або методом аутостран, найлонамілази, природні і модифіковані радіографії. целюлози, поліакриламіди, габро і магнетит. Носій Антитіла відповідно до даного винаходу також для цілей даного винаходу за природою може бути можна помітити флуоресцентною сполукою. Коли або розчинним до деякоїміри або нерозчинним. на антитіла з флуоресцентними мітками діє світло Матеріал основи фактично може мати будь-яку з відповідною довжиною хвилі, їхню присутність можливу структурну конфігурацію доти, поки приєможна виявити внаслідок флуоресценції. Серед днана молекула здатна зв'язуватися з антигеном найбільш часто використовуваних сполук для або антитілом. Так, конфігурація основи або носія флуоресцентного мічення знаходяться ізотіоціаможе бути сферичною, як у гранул, циліндричною, нат, родамін, фікоеритрин, пікоціанін, алофікоціаяк внутрішня поверхня пробірки або як зовнішня нін, о-фтальдегид і флуорескамін. поверхня стрижня. З іншого боку, поверхня може Антитіла також можна позначити для детекції з бути плоскою, такою як лист, тест-пластина і т.п. використанням металів, що флуресціюють, таких Кращими основами або носіями є полістирольні як 152Е, або інших металів ряду лантаноїдів. Ці гранули. Фахівцям у цій області техніки відомі баметали можна приєднати до антитіл із викорисгато інших відповідних носіїв для зв'язування антанням таких груп, що утворюють комплекси з метитіл або антигенів, або в цьому можна засвідчиталами, як залишок диетилентриамінпентаоцтової тися за допомогою простих експериментів. кислоти (ЕТРА). Зв'язуючу активність отриманих антитіл, згідно Антитіла також можна помітити для детекції за з винаходом, як зазначено вище, можна визначити допомогою сполучення їх із хемілюмінесцентною відповідно до добре відомих способів. Фахівці в сполукою. Присутність мічених хемілюмінесцентцій області техніки зможуть визначити робочі та ною сполукою антитіл потім визначають шляхом оптимальні умови аналізу у випадку кожного видетекції наявності люмінесценції, що з'являється в значення, використовуючи звичайні експерименти. ході хімічної реакції. Прикладами сполук, особливо Інші стадії, такі як промивання, перемішуванкорисних для хемілюмінесцентного мічення, є люня, струшування, фільтрація і т.п., можна включити мінол, ізолюмінол, theromatic ефір акридинію, імів аналізи як звичайні або необхідні в даній ситуадазол, сіль акридинію та ефір щавлевої кислоти. ції. Подібним чином, для мічення антитіл даного Одним із способів, яким антитіла відповідно до винаходу можна використовувати біолюмінесценданого винаходу можна помітити для виявлення, є тну сполуку. Біолюмінесценція є видом хемілюмізв'язування їх із ферментом і застосування імунонесценції, виявленої в біологічних системах, в яких ферментного аналізу (ЕІА). Фермент, у свою чергу, каталітичний білок підвищує ефективність хеміпри подальшому впливі відповідного субстрату люмінесцентної реакції. Наявність біолюмінесценбуде реагувати із субстратом таким чином, що тного білка визначають за допомогою детекції наутвориться хімічна група, яку можна виявити, наявності люмінесценції. Важливими приклад, спектрофотометричним, флюорометричбіолюмінесцентними сполуками для цілей мічення ним або візуальним способом. Ферменти, які можє люциферин, люцифераза та аекорин. на використовувати для детектованого мічення Молекулу антитіла даного винаходу можна антитіл, включають, але не обмежуються перераадаптувати для застосування в імунометричному хованим, малатдегідрогеназу, стафілокококову аналізі, відомому також як "двошаровий" або "саннуклеазу, дельта-5-стероїдізомеразу, дріжджову двіч"-аналіз. При типовому імунометричному анаалкогольдегідрогеназу, альфалізі деяка кількість позначених антитіл (або фраггліцерофосфатдегідрогеназу, триозофосфатізоментів антитіл) зв'язується з твердою основою або меразу, пероксидазу хріну, лужну фосфатазу, асносієм, і добавляють деяку кількість мічених для парагіназу, глюкозооксидазу, бета-галактозидазу, детекції розчинних антитіл, щоб створити можлирибонуклеазу, уреазу, каталазу, глюкозо-6вість детекції та/або кількісного визначення потфосфатдегідрогеназу, глюкоамілазу та ацетилхорійного комплексу, утвореного антитілами твердої лінестеразу. Детекцію можна здійснити колоримефази, антигеном і міченими антитілами. тричними методами, при яких використовують Типовими, і кращими, імунометричними аналіхромогенний субстрат для ферменту. Детекцію зами є "прямі" аналізи, при яких антитіла, зв'язані з також можна здійснити за допомогою візуального твердою фазою, спочатку вводять у контакт із бі 41 75564 42 нарним твердофазним комплексом антитілоНаприклад, для того, щоб управляти експресіантиген. Після відповідного періоду інкубації тверєю білка, що взаємодіє з каспазою-8, може знадоду основу або носій промивають для видалення битися одержання в клітині антисмислової РНК. залишків рідкого зразка, включаючи антиген, що Для цієї мети повну або неповну кДНК, що кодує не прореагував, якщо такий є, і потім вводять у білок, який взаємодіє з каспазою-8, вбудовують в контакт із розчином, що містить невідому кількість експресуючий вектор, який містить промотор, що міченого антитіла (яке функціонує як "репортерна також зазначено вище. За допомогою цього 3'молекула"). Після другого періоду інкубації для кінець кДНК вбудовують по сусідству з 3'-кінцем створення комплексу міченого антитіла з антигепромотору, причому 5'-кінець кДНК відділяється ном, зв'язаним із твердою основою або носієм за від 3'-кінця промотору зазначеної кДНК. Тому, пісдопомогою неміченого антитіла, тверду основу ля експресії кДНК у клітині продукується антисмиабо носій промивають вдруге для видалення мічеслова РНК, нездатна кодувати білок. Наявність них антитіл, що не прореагували. антисмислової РНК у клітині знижує експресію кліВ іншому типі "сандвіч"-аналізу, який також тинної (геномної) копії гена, що взаємодіє з каспаможна застосовувати з антигенами даного виназою-8. ходу, використовують так називані "одночасні" і Для одержання антисмислової РНК можна ви"зворотні" аналізи. Одночасний аналіз включає користовувати повну кДНК. З іншого боку, можна одну стадію інкубації, тому що антитіла, зв'язані з використовувати її фрагмент, який, переважно, твердою основою або носієм, і мічені антитіла домає довжину від 9 до 2000 нуклеотидів, краще - від бавляють до досліджуваного зразка одночасно. 15 до 500 нуклеотидів, і найбільш переважно від Після завершення інкубації тверду основу або но30 до 150 нуклеотидів. сій промивають для видалення залишків рідкого Фрагмент, переважно, відповідає області в зразка і мічених антитіл, що не увійшли до комплемежах 5'-половини кДНК, краще - 5'-області, що ксу. Потім визначають наявність мічених антитіл, містить 5'-нетрансльовану область та/або першу зв'язаних із твердою основою або носієм, так, як екзонну область, і найбільш переважно - що місце звичайно роблять при "прямому" сандвічтить сайт початку трансляції ATG. З іншого боку, аналізі. фрагмент може відповідати послідовності ДНК При "зворотному" аналізі використовують постільки 5'-нетрансльованої області. тадійне додавання до рідкого зразка спочатку розВ якості антисмислового олігонуклеотиду мочину мічених антитіл, а потім немічених антитіл, жна використовувати синтетичний олігонуклеотид. зв'язаних із твердою основою або носієм, після Олігонуклеотид, переважно, являє собою олігонувідповідного періоду інкубації. Після другої інкубаклеотид ДНК. Довжина антисмислового олігонукції тверду фазу промивають звичайним способом леотиду становить, переважно, від 9 до 150, крадля звільнення її від залишків досліджуваного зраще - від 12 до 60, і найбільш переважно - від 15 до зка і розчину мічених антитіл, що не прореагували. 50 нуклеотидів. Область, що перекривається анПотім здійснюють визначення мічених антитіл, тисмисловим олігонуклеотидом, містить, переважзв'язаних із твердою основою або носієм, як при но, 3'-нетрансльовану область кДНК, краще, коли "одночасному" і "прямому" аналізах. вона містить сигнал поліаденілювання або стопІмуноаналізи кодон трансляції, або те і інше. Здійснення імуноаналізів, таких як RIA або Огляд механізму дії антисмислової РНК і сьоELISA, описано в багатьох статтях, книгах і інших годнішній стан техніки використання антисмислопублікаціях. Дається посилання на [WO 97/03998, вих інструментів приведений у [Kumar et al., із с.48, рядок 4, по с.52, рядок 27]. Імуноаналізи Microbiol. МоІ. Biol. Rev., 62, p.1415-1434, 1998]. винаходу можуть бути двох загальних типів: поЗастосування антисмислових олігонуклеотидів при перше, можна використовувати імуноаналізи з інгібуванні синтезу рецептора BMP описане [Yeh et використанням іммобілізованого білка, що взаєaІ., J. Bone Miner. Res., 13, p.1870-9, 1998]. Застомодіє з каспазою-8, або його еквівалента для кільсування антисмислових олігонуклеотидів для інгікісного визначення каспази-8; по-друге, можна бування синтезу гена вольтзалежного калієвого використовувати імуноаналізи з використанням каналу Kvq.4 описано [Meiri et al., PNAS, 95, іммобілізованих антитіл, спрямованих проти епітоp.15037-1042, 1998]. Застосування антисмислових пу білка, що взаємодіє з каспазою-8, для кількісноолігонуклеотидів для інгібування синтезу Bcl-х го визначення білків, що взаємодіють із каспазоюописане [Kondo et al., Oncogene, 17, p.2585-91, 8. 1998]. Такі аналізи можуть знайти застосування в діаЛікувальне застосування антисмислових лігностиці, коли потрібно оцінити вміст каспази-8 і карських засобів описують [Stix. Sci. Am., 279, p.46, інших білків, залучених до апоптозного каскаду 50, 1998; Flanagan, Cancer Metastasis Rev., 17, реакцій, при багатьох порушеннях або синдромах, p.169-76, 1998; Guinot and Temsamani, Pathol. Bioi. де можлива участь таких каскадів. (Paris), 46, p.347-54, 1998; і в приведених у цих Нуклеїнові кислоти роботах посиланнях]. Очікується, що клони, отримані при скринінгу Коли створюють антисмислові олігонуклеотизгідно з винаходом, будуть неповними клонами. ди, як правило, використовують модифікації олігоОдержання повних клонів, коли це необхідно, тануклеотидів, що підсилюють потрібні властивості. кож описано вище. Послідовність ДНК повного Наприклад, замість фосфоефірних зв'язків, що клону і неповного клону, знайдена спочатку при зустрічаються в природних ДНК, використовують скринінгу згідно з винаходом, може знайти різні фосфоротіоатні зв'язки, головним чином, внаслізастосування. док того, що такі олігонуклеотиди, що містять фо 43 75564 44 сфоротіоат, менше сприйнятливі до руйнування можна ввести в клітини шляхом поглинання ДНК, клітинними ферментами. Peng et al. повідомляють, поглинання модифікованої ДНК (див. модифікації що небажані in vivo побічні дії олігонуклеотидів, що олігонуклеотидів і білків, що приводять до посилемістять фосфоротіоат, можна послабити при виконої мембранної проникності, як описано нижче) ристанні змішаного фосфодіефірфосфоротіоатноабо переносу гена, опосередкованого вірусним го скелета. Переважно, 2'-метоксирибонуклеотидні вектором, що докладно описано нижче. модифікації використовують у 60% нуклеотиду. Введення в клітини білків, що взаємодіють із Такі модифіковані олігонуклеотиди здатні виявлякаспазою-8, пептидів і ДНК ти антисмислову дію, порівняну з дією, що спостеДаний винахід відноситься до білків, що взаєрігається у випадку олігонуклеотидів, що містять модіють із каспазою-8, отриманих із них пептидів, фосфоротіоат. Peng et al. також повідомляють, що антисмислових молекул ДНК та олігонуклеотидів. аналоги олігонуклеотидів, нездатні до підтримки Застосування для лікування або досліджень таких активності рибонуклеази Н, є неактивними. інструментів вимагає їх введення у клітини живого Отже, кращий антисмисловий олігонуклеотид організму. Для цієї мети потрібно підсилити мемвинаходу має змішаний фосфодіефірфосфоротіобранну проникність для пептидів, білків та олігонуатний скелет. Найбільш переважно, 2'клеотидів. Способи поліпшення мембранної прометоксирибонуклеотидні модифікації використоникності для олігонуклеотидів описані вище. Ті ж вують у приблизно 30%-80%, найбільш переважно принципи, а саме, деривацію ліофільними струк- у приблизно 60%, олігонуклеотиду. турами, можна також використовувати при ствоВ антисмисловий олігонуклеотид можна ввесренні білків і пептидів із посиленою здатністю проти інші модифікації. Наприклад, молекулу олігонуходити крізь мембрану. Наприклад, до клеотиду можна з'єднати з групою, що містить непослідовності пептиду або білка можна додати обов'язково частково ненасичений вуглеводневий послідовність відомого мембранотропного пептиланцюг і одну або декілька полярних або заряджеду, як зазначено вище. Крім того, пептид або білок них груп, таких як карбоксильна кислотна група, можна дериватизувати за допомогою частково складноефірні групи і спиртові групи. З іншого боліофільних структур, таких як вищевказані вуглеку, олігонуклеотиди можна з'єднати з пептидными водневі ланцюга, що заміщені щонайменше одніструктурами, що, переважно, є мембранотропними єю полярною або зарядженою групою. Наприклад, пептидами. Такі модифіковані олігонуклеотиди лауроїлпохідні пептидів описані [Muranishi et al., проникають крізь мембрани легше, що є критичPharm. Research., 8, 649, 1991]. Інші модифікації ним для їхнього функціонування, і, отже, можна пептидів і білків включають окислення метіоніноістотно підсилити їхню активність. Проникнення вих залишків, за допомогою чого створюються крізь мембрани особливо бажано для антисмиссульфоксидні групи, як описано [Zacharia et al., лових лікарських засобів, яким потрібно досягти Eur. J. Pharmacol, 203, p.353, 1991]. Zacharia із головного мозку. Олігонуклеотиди, з'єднані з паспівробітниками також описують пептид або похідльмітилом, описані [Gerster et al., Anal. Biochem., ні, де відносно гідрофобний пептидний зв'язок за262, p.177-84, 1998]. Олігонуклеотиди, з'єднані з міняється її кетометиленовим ізоефіром (СОСН2). гераніолом, описані [Shoji et al., J. Drug Target, 5, Такі та інші модифікації, відомі фахівцям в області p.261-73, 1998]. Олігонуклеотиди, з'єднані з пептихімії білків і пептидів, підсилюють здатність пронидами, наприклад, мембранотропними пептидами, і кати крізь мембрану. їхнє одержання описані [Soukchareun et al., Іншим способом посилення мембранної проBioconjug. Chem., 9, p.466-75, 1998]. Модифікації никності є використання рецепторів, таких як віруантисмислових молекул або інших лікарських засні рецептори, на поверхні клітини для індукції собів, що спрямовують молекулу в певні клітини і поглинання клітиною пептиду або білка. Цей мехапідсилюють поглинання олігонуклеотиду зазначенізм часто використовується вірусами, що специними клітинами, описані [Wang, J. Controlled фічно зв'язуються з деякими молекулами поверхні Realease, 53, p.39-48, 1998]. клітини. Після зв'язування клітина пропускає вірус Рибозими всередину. Така молекула поверхні клітини назиОтримавши відому послідовність мРНК гена, вається вірусним рецептором. Наприклад, молеможна створити рибозими, що представляють сокули інтегрину CAR і Ad описані як вірусні рецепбою молекули РНК, що специфічно зв'язуються і тори для аденовірусу, [див. Hemmi et al., Hum. розщеплюються зазначеною послідовністю мРНК Gene Ther., 9, p.2363-73, 1998], і приведені там [див., наприклад, Chen et al., Ann. NY Acad. Sci., посилання. Молекули CD4, GPR1, GPR15 і 660, 271-3, 1992; Zhao and Pick, Nature, 365, p.448, STRL33 ідентифіковані як рецептори/корецептори 1993; Shore et al., Oncogene, 8, 3183, 1993; Joseph для ВІЛ, [див. Edinger et al., Virology, 249, p.367-78, and Burke, J. Biol. Chem., 24515, 1993; Shimayama 1998, і приведені там посилання]. et al., Nucleic Acids Symp., Ser. 29 p.177, 1993; Таким чином, кон'югування пептидів, білків та Cantor et al., PNAS, 90, p.10932, 1993]. олігонуклеотидів із молекулами, відомими як реВідповідно, можна створити послідовність цептори для зв'язування з поверхнею клітини, буРНК, що кодує рибозим, яка розщеплює мРНК білде підсилювати мембранну проникність для зазнака винаходу, що взаємодіє з каспазою-8. Місце чених пептидів, білків або олігонуклеотидів. розчеплення розташовується, переважно, в облаПрикладами відповідних груп для утворення кон'юсті, що кодує, або в 5'-нетрансльованій області, гатів є цукри, вітаміни, гормони, цитокіни, транскраще - у 5'-частині області, що кодує, пов'язаною ферин, азіалоглікопротеїн і подібні молекули. У зі стартовому кодону трансляції AUG. патенті США 5108921, Low et al., описується заДНК, що кодує рибозим згідно з винаходом, стосування таких молекул для цілей посилення 45 75564 46 мембранної проникності для пептидів, білків та описано, наприклад, [Teoh et al., Blood, 92, p. 4591олігонуклеотидів, і одержання зазначених кон'юга4601, 1998; Narumi et al., Am. J. Respir. Cell. МоІ. тів. Biol., 19, p.936-941, 1998; Pederson et al., J. Low із співробітниками також вказують, що Gastrointest. Surg., 2, p.283-91, 1998; Guang-Lin et молекули, такі як фолат або біотин, можна викориal., Transplant. Proc, 30, p. 2923-4, 1998, і в цитовастовувати для спрямованої доставки кон'югату до них у цій роботі посиланнях; Nishida et al., Spine, багатьох клітин організму, у силу численної і не23, p.2437-42, 1998; Schwarzenberger et al., J. специфічної експресії рецепторів для цих молекул. Immunol., 161, p.6383-9, 1998; і Cao et al., J. Описане вище застосування білків поверхні Immunol., 161, p.6238-44, 1998]. Ретровірусний клітини для посилення мембранної проникності перенос антисмислових послідовностей описаний для пептиду, білка або олігонуклеотиду винаходу [Daniel et al., J. Biomed. Sci., 5, 383-94, 1998]. можна також використовувати при спрямованій Коли в якості векторів використовують віруси, доставці зазначеного пептиду, білка або олігонукдля спрямованої доставки вірусу звичайно виколеотиду винаходу до клітин або тканин певного ристовують білки вірусної поверхні. Оскільки багатипу. Наприклад, якщо необхідно потрапити в рато вірусів, такі як вищевказаний аденовірус, скорікові клітини, краще використовувати білок поверхше неспецифічні у своєму клітинному тропізмі, ні клітини, що експресується більше на поверхні може знадобитися надання більшої специфічності таких клітин. Прикладами є фолатні рецептори, за допомогою використання промотору клітинного антигени до муцину MUC1, MUC2, MUC3, MUC4, типу або тканиноспецифічного промотору. [Griscelli MUC5AC, MUC5B і MUC7, глікопротеїнові антигени et al., Hum. Gene Ther., 9, p.1919-28, 1998], описуKSA, карциномо-ембріональний антиген, специфіють застосування промотору легкого ланцюга 2 чний для передміхурової залози мембранний анкардіального міозину, специфічного для шлуночку, тиген (PSMA) HER-2/neu та хоріонний гонадотродля специфічної спрямованої доставки в серце пін-бета людини. У цитованій вище роботі [Wang et гена, перенос якого опосередковується аденовіруal., 1998], описується застосування фолату для сом. спрямованої доставки в ракові клітини, а в [Zhang З іншого боку, можна створити методами генet al., Clin. Cancer Res., 4, p.2669-76, 1998], вказуної інженерії вірусний вектор для експресії іншого ється на відносну велику кількість кожного зінших білка на його поверхні, або білок поверхні вірусноантигенів, зазначених вище, у ракових і здорових го вектора можна замінити для введення потрібної клітинах різного типу. пептидної послідовності. Таким чином, можна Отже, білок, пептид або олігонуклеотид винастворити вірусний вектор для експресії одного або ходу можна, із використанням описаного вище декількох додаткових епітопів, пептидів, що зв'яметоду кон'югації, спрямовано доставити в певзуються з КГС класу II або можна використовувати ний, який потрібно, тип клітин. Наприклад, якщо епітопи, утворені від "хомінг"-молекул, для напряпотрібно підсилити апоптоз в клітинах лімфоцитамку вірусного вектора відповідно до вказівок винарної лінії, у ці клітини можна спрямувати білок або ходу. пептид винаходу, які позитивно модулюють каспаЗастосування описаних вище інструментів зу-8, наприклад, використовуючи молекули КГС Білки винаходу, що взаємодіють із каспазою-8, класу II, що експресуються на цих клітинах. Цього взаємодіють специфічно із субодиницями каспазиможна досягти за допомогою сполучення антитіла, 8, залученими до протеолітичної активності каспаабо його антиген-зв'язуючого сайта, спрямованого зи-8. Не заглиблюючись у теорію, автори винаходу проти константної області зазначеної молекули думають, що білки винаходу, що взаємодіють із КГС класу II, із білком або пептидом винаходу. каспазою-8, є "наступними" елементами в каскаді Далі, описані багато рецепторів клітинної поверхні передачі сигналу за участю каспази-8. Виявляєтьдля різних цитокінів та інших молекул клітинної ся, що "попередні" агенти зв'язуються через прокомунікації, і багато які з цих молекул експресудомен каспази-8, домен, що опосередковує взаєються більш-менш тканино- або клітиноспецифічмодію білок-білок. Виявляється, що екстенсивне но. Так, коли потрібна спрямована доставка підг"крос-спілкування" із багатьма засобами, що оргарупи Т-клітин, для одержання кон'югату винаходу нізують апоптозну реакцію, може бути опосередможна використовувати молекулу поверхні клітини ковано взаємодіями білок-білок за участю ефектоCD4. Молекули, що зв'язують CD4, надаються вірного домену загибелі (DED) і родинних доменів, русом ВІЛ, чий поверхневий антиген gp42 здатний таких як домени, локалізовані в продомені каспазв'язуватися з молекулою CD4. Білок винаходу, що зи-8. На відміну від таких білків, білки даного винавзаємодіє з каспазою-8, або пептид винаходу, які ходу, що взаємодіють із каспазою-8, можуть знапідсилюють апоптоз, можна спрямовано доставити ходитися серед безпосередніх виконавців процесу переважно в Т-клітини при лікуванні пацієнта, що загибелі клітин. Наприклад, один із білків винахостраждає від автоімунних реакцій на основі Тду, що взаємодіють із каспазою-8, Тір-60 є фермеклітин, таких як пацієнти з червоним вовчаком. нтом гістондеацетилазою. Отже, зазначений білок Спрямована доставка в клітини, опосередкоможе безпосередньо брати участь у змінах струквана вірусами тури хроматину. Білки, пептиди та антисмислові послідовності Взаємодія білків винаходу з каспазою-8 має винаходу можна ввести в клітини, використовуючи ряд суміжних наслідків. По-перше, відбувається вірусний вектор. Застосування вакцинного вектора модуляція активності каспази-8. Це показано тут для таких цілей докладно описано [в цитованій при аналізі in vivo, де клон J2 винаходу інгібує вище главі 16 Current Protocols in Molecular апоптоз, опосередковуваний каспазою-8. Biology]. Застосування аденовірусного вектора По-друге, білок, що взаємодіє з каспазою-8, 47 75564 48 може підвищити активність каспази-8 за допомоздатні діяти як негативні регулятори активності гою попередження руйнування каспази-8, або зникаспази-8. Отже, спочатку варто перевірити ефект зити її активність, діючи як інгібітор. використання антисмислових олігонуклеотидів і По-третє, активність білка, що взаємодіє з касантисмислової РНК та рибозимів, наприклад, в пазою-8, можна модулювати. Це показано тут чеописаному вище аналізі in vivo, перед тим, як обрез здатність каспази-8 розщеплювати білки, що говорювати застосування таких засобів для лікувзаємодіють із каспазою-8. Можливо, що деякі з вання. таких білків інактивуються шляхом розчеплення. З іншого боку, існують певні ситуації, коли моОднак також можливо, що активність білків змінюже знадобитися підвищення активності каспази-8. ється, індукуються нові активності, або що білок, Це може відбутися у випадку тих же захворювань, який взаємодіє з каспазою-8, активується шляхом зазначених вище, наприклад, при системному черозчеплення, саме як самі каспази. рвоному вовчаку. Однак типи клітин, що повинні Отже, білки, що взаємодіють із каспазою-8, стати мішенями, є іншими. Наприклад, при вовчаку пептиди, олігонуклеотиди та антитіла винаходу популяція Т-клітин може містити аутореактивні корисні при модуляції активності каспази-8. Модуклітини, що не руйнуються в тимусі. Отже, до Тляція каспази-8 по низхідній бажана в ситуаціях, клітинам варто спрямувати засіб винаходу, що де відбувається надлишкова загибель клітин внамодулює активність каспази-8 по висхідній. Песлідок апоптозу. Наприклад, передбачається, що редбачається, що при деяких захворюваннях, тапри розсіяному склерозі з первинною олігодендроких як розсіяний склероз, деякі клони Т-клітин грагліопатією, автоімунному увеоретиніті, діабеті, ють критичну роль у розвитку захворювання. вовчаку, аутоімунному міокардиті І, печінковій неОтже, у такі клітини можна спрямувати засіб згідно достатності, незалежно від етіології, опосередкоз винаходом, що модулює активність каспази-8 по ваному HCV хронічному гепатиті, гастриті, напривисхідній, використовуючи одне або декілька антиклад, гастриті типу А, змішаній хворобі сполучної тіл, специфічно спрямованих на варіабельну обтканини (MCTD), хворобі Крона і неспецифічному ласть Т-клітинного рецептора клонів аутореактиввиразковому коліті руйнування тканин організму них Т-клітин, для спрямування засобу винаходу, визивається сигналами апоптозу. Отже, для пацієщо модулює активність каспази-8 по висхідній, що нтів, що страждають від таких хвороб, може бути може являти собою білок, який взаємодіє з каспасприятливою модуляція активності каспази-8 по зою-8, або пептид, що відповідають винаходу. низхідній в тих клітинах, що руйнуються шляхом З огляду на вищевказане, даний винахід відапоптозу. носиться до фармацевтичних препаратів, що місНаприклад, при зазначеній вище олігодендротять активну речовину, яка складається з одного патії потрібно інгібувати активність каспази-8 в або декількох білків, що взаємодіють із каспазоюолігодендроцитах. Рецептор фосфоліпідлізофос8, пептидів, антитіл, рибозимів, антисмислових фатидної кислоти в сполученні з G-протеїном кліРНК або антисмислових олігонуклеотидів, які відтинної поверхні експресується в олігодендроцитах повідають винаходу. і в різних інших клітинах головного мозку, але не в Винахід також відноситься до фармацевтичної інших тканинах організму. Отже, пептид або білок композиції, яка містить вірусний вектор, здатний винаходу спрямовують у ці клітини. Цього можна інфікувати клітини ссавця, де зазначений вектор досягти або за допомогою сполучення зазначеного містить операбельно зв'язаний промотор і посліпептиду або білка з фосфоліпідлізофосфатидною довність ДНК винаходу, що кодує білок, який взаєкислотою або за допомогою введення послідовномодіє з каспазою-8, або пептид, рибозим, антиссті антитіла, що специфічно впізнає зазначений мислову РНК, антисмисловий олігонуклеотид або рецептор фосфоліпідлізофосфатидної кислоти, у антитіло, що відповідають винаходу. Вірусний веквірусний вектор, так що зазначений вірусний вектор може містити, необов'язково, кодуючу послітор специфічно зв'язується із зазначеним рецепдовність, операбельно зв'язану з промотором, яка тором фосфоліпідлізофосфатидної кислоти. кодує пептид або білок, розташовані на поверхні Подібним чином пептиди або білки винаходу вірусу, і яка здатна до зв'язування з білком поверможна направити в клітини іншого типу, які порухні клітини ссавця. Білок поверхні, переважно, явшуються при інших захворюваннях, перераховаляє собою білок, що може поглинати вірусний векних вище, та інших захворюваннях, при яких, як тор, і, переважно, експресується показано, надлишок апоптозних клітин опосередтканиноспецифічно або специфічно для типу кліковує ушкодження в тканині організму, яка спостетин, так що створюється можливість спрямованої рігається. доставки вірусного вектора. Подібним чином антисмислову ДНК, антисмиБілок, що взаємодіє з каспазою-8, його аналословий олігонуклеотид та рибозим винаходу також ги, фрагменти або похідні також можна використоможна спрямувати у вищевказані олігодендроцити вувати для виділення, ідентифікації та клонування або у відповідні клітини при інших захворюваннях. інших білків того ж класу, тобто білків, що зв'язуУ такому випадку пригнічується експресія білків, ються з каспазою-8, або функціонально родинних що взаємодіють із каспазою-8, а не експресія сапротеаз або білків, залучених до процесу внутрішмої каспази-8. Шляхом інгібування експресії ряду ньоклітинної передачі сигналу. При такому застобілків, що взаємодіють із каспазою-8, можна знисуванні можна використовувати зазначену вище зити апоптозну дію каспази-8. Однак зниження систему з двома гібридами дріжджів, або можна експресії певних білків, які взаємодіють із каспавикористовувати недавно розроблену систему з зою-8, може фактично підвищити дію каспази-8, використанням гібридизації по Саузерну в нестротому що деякі білки, які взаємодіють із каспазою-8, гих умовах із наступним клонуванням методом 49 75564 50 PCR [Wilks et al., 1989]. У публікації Wilks et al. тифікації та характеризації інших білків, факторів, описано ідентифікацію і клонування двох передбарецепторів і т.п., здатних зв'язуватися з білками чуваних протеїнтирозинкіназ за допомогою застовинаходу, які взаємодіють із каспазою-8, можна сування гібридизації по Саузерну в нестрогих умовикористовувати інші процедури, такі як афінна вах із наступним клонуванням методом PCR на хроматографія, методи гібридизації ДНК і т.п., дооснові відомої послідовності мотиву кінази задубре відомі в техніці. маної послідовності кінази. Такий підхід можна Приклад ІА застосувати, відповідно до даного винаходу, із Двогібридний скринінг для ідентифікації білків, використанням послідовності білка, що взаємодіє з що взаємодіють із каспазою-8 каспазою-8, для ідентифікації і клонування білків, Для скринінгу білків, що взаємодіють із каспародинних білкам, що взаємодіють із каспазою-8. зою-8, і їхніх потенційних субстратів використовуІнший підхід до застосування білка, що взаєвали модифіковану систему з двома гібридами модіє із каспазою-3, або його аналогів, фрагментів дріжджів, описану як ["Yeast-three Hybrid system" або похідних згідно з винаходом полягає у викори(Tirode F. et al., 1997)]. Окремі використані вектостанні їх у методах афінної хроматографії для вири, штами дріжджів і бібліотеки одержували від ділення й ідентифікації інших білків або факторів, Clontech (Palo Alto, USA) у вигляді компонентів залучених до процесу внутрішньоклітинної передвогібридної системи Matchmaker (#РТ1265-1). дачі сигналу. При такому застосуванні білок, що Дві субодиниці каспази-8 експресували окремо взаємодіє з каспазою-8, його аналоги, фрагменти під контролем різних промоторів. Коротку субодиабо похідні даного винаходу можна окремо приєдницю р10 (від серину 375 до аспарагінової кислоти нати до матриць для афінної хроматографії і потім 479) клонували у вектор pGBT9 (Clontech) у рамці привести у контакт із клітинними екстрактами або з доменом зв'язування ДНК дріжджового білка виділеними білками або факторами, що, як передGal4 [амінокислоти 1-147, числа в послідовностях бачається, залучені до процесу внутрішньоклітинвідповідно до Laughon et al., Molecular and Cellular ної передачі сигналу. Дотримуючись процедурі Biology, 4, 260-267, 1984]. Довгу активну субодиафінної хроматографії, можна елюювати, виділити ницю р20 (від серину 217 до аспарагінової кислоти та охарактеризувати інші білки або фактори, що 374) мутували в позиції 360, тобто цистеїн, що зв'язуються з білком, який взаємодіє із каспазою-8, знаходиться в цій позиції, заміняли на серії або його аналогами, фрагментами або похідними (C360S), надаючи протеазної активності неактиввинаходу. ному ферменту. Мутовану субодиницю C360S р20 Як вказувалося вище, білок, що взаємодіє із експресували у вигляді незлитого білка під конткаспазою-8, або його аналоги, фрагменти або поролем промотору Меt25, що позитивно регулюєтьхідні винаходу можна також використовувати в ся в середовищі, позбавленому метіоніну якості імуногенів (антигенів) для одержання спе[Sangsoda, Моl. Gen. Genet., 200, p.407-14, 1985]. цифічних антитіл до них. Такі антитіла також можМожливість регулювання активності промотору, на використовувати для цілей очищення білка, що що управляє експресією субодиниці р20, за доповзаємодіє з каспазою-8 (наприклад, Nмогою добору концентрації метіоніну в середовищі ацетилглюкозамін-6-фосфатдеацетилази людини для росту дріжджових клітин робить можливим (1) або її будь-яких ізоформ), або від клітинних екствикористання субодиниць токсичного білка в якості рактів або від трансформованих клітинних ліній, наживки і (2) регулювання залежності взаємодії які продукують білок, що взаємодіє з каспазою-8, білок-білок від третього партнера, тобто субодиабо його аналоги або фрагменти. Далі ці антитіла ниці р20. Одиниці, що експресують, р10 і р20 можможна використовувати для цілей діагностики для на локалізувати в різних векторах. їх можна локаідентифікації порушень, зв'язаних з аномальним лізувати також у одному векторі. У прикладі, що функціонуванням опосередкованих каспазою-8 описується, використовували модифікований ліганду FAS-R або системи TNF. Таким чином, pGΒΤ9-вектор pGBT9-3H [див. вище Tirode et al.]. якщо такі порушення зв'язані з порушеннями в Субодиницю р20, переважно, експресують у висистемі внутрішньоклітинної передачі сигналу за гляді гібрида із сигналом ядерної локалізації. участю білка каспази-8 або білка, що взаємодіє з Після експресії у відсутності метіоніну дві сукаспазою-8, такі антитіла могли б служити в якості бодиниці асоціюють одна з одною у дріжджовій важливого діагностичного інструмента. клітині. Асоціація двох субодиниць показана за Потрібно також відзначити, що виділення, ідедопомогою експериментів по коімунопреципітації і нтифікацію та характеризацію білка винаходу, що вестерн-блотінгу. взаємодіє з каспазою-8, можна здійснити з викориБібліотеку кДНК В-клітини, клоновану у вектор станням будь-який із добре відомих стандартних pGAD GH [Durfee et al., Genes Dev., 7, 555-569], процедур скринінгу. Наприклад, одну з таких проодержали в дарунок від д-ра S. Elledge. Вектор цедур скринінгу -процедуру з двома гібридами містить домен активації Gal4 (амінокислоти 768дріжджів, як зазначено нижче, використовували 881). Цього домену активації GAL4 достатньо, кодля ідентифікації білка, що взаємодіє з каспазою-8 ли злититий із доменом зв'язування ДНК GAL4, [див. Stanger et al., 1995], і потім різних білків видля індукції суттєвої транскрипційної активності находу, що взаємодіють із каспазою-8 (крім різних гена GAL4, [див. Ma and Ptashne, Cell, 48, 847-853 інших нових білків, зазначених вище і нижче, у (1987)]. спільних заявках, що розглядаються). Подібним Сайт активації GAL, що розташований вище чином, як указується вище і нижче, для виділення, (UAS.sub.G), як описано Keegan et al., (1986), цит. ідентифікації та характеризації білка винаходу, що вище, є присутнім у розташованій вище області взаємодіє з каспазою-8, або для виділення, іденрепортерного гена (lac) і гена, що допускає селек 51 75564 52 цію (His) у штамі дріжджів HF7c, отриманому від взаємодію з іншими білками. Це дослідження здійClontech. снювали за допомогою трансформації досліджуКультуру HF7c трансформували вищевказаваних клонів разом із нерелевантним білком, ексною каспазою-8, що містить плазміди, і трансфорпресованим у вигляді гібрида з доменом активації манти дріжджових клітин відбирали за допомогою ДНК у векторі pGAD GH. вирощування в середовищі для селекції, як описаПодвійні трансформанти, здатні зростати в но в протоколах Clontech для дріжджів, тобто позсередовищі, позбавленому гістидину, або такі, що бавленої триптофану, лейцину, метіоніну, тирозивиявляли lacZ-активність, вказували на те, що бібну. Необов'язково добавляли гомосерин у кількості ліотечний клон зв'язувався неспецифічно. 80мг/л. У якості нерелевантних білків використовуваПотім культуру зазначених трансформантів ли білки, відомі як "прилипаючі", тобто такі, що трансформували далі бібліотекою В-клітин, що взаємодіють неспецифічно з іншими білками, так містить вектор, із наступним висіванням на вищесамо, як інші нерелевантні білки. Наприклад, білки вказане середовище, позбавлене гістидину (серевипробовували на зв'язування з ламіном, RIP, довище для селекції). RAIDD, TRAF2 і MORT1. Взагалі, білки, що зв'язуНеобов'язково, до середовища для селекції ють ламін, відкидали. добавляли 3-амінотриазол, що є інгібітором ферЗазначене вище тригібридне дослідження таменту гістамінсинтетази. Додавання інгібітору кож здійснювали із субодиницею каспази-8 p10, служить для регуляції витоку з промотору, що злитою з доменом зв'язування ДНК lexA. Кращим управляє his-геном у дріжджових клітинах, які не вектором є вектор pLexA, доступний від Clontech. містять білків, які взаємодіють із каспазою-8. У Коли в якості домену зв'язування ДНК використодеяких випадках слабка неспецифічна взаємодія вують lexA, варто використовувати штам дріжджів може призвести до помилкової транскрипції від L40 або еквівалентний штам. промотору Gal4-UAG-his, що веде до дріжджових Білки, що взаємодіють із каспазою-8, ідентифіклонів, які ростуть в середовищі для селекції. кували за допомогою описаного вище методу Трансформанти, що ростуть у середовищі для скринінгу і потім аналізували з використанням селекції, відбирали та екстрагували ДНК-плазміди. аналізів на розчеплення in vivo і in vitro і аналізів Потім ДНК трансформували в бактерії НВ101, які на трансдукцію сигналів, як описано далі. допускають селекцію в середовищі, позбавленому Приклад IB лейцину. Модифікована наживка для двогібридного Після вирощування бактерій і екстракції та скринінгу для ідентифікації білків, що взаємодіють очищення від них плазмідної ДНК потім трансфоіз каспазою-8 рмували плазмідну ДНК у дріжджові клітини Модифіковану каспазу-8 використовували для SFY526. Потім здійснювали дослідження lacZіншого скринінгу білків, що взаємодіють із каспаактивності трансформантів SFY526 за допомогою зою-8, із системою з двома гібридами дріжджів висівання на середовище для селекції, що вклю[Fields and Song, 1989]. Каспазу-8 експресували з чає гістидин і містить Xgal. Потім колонії дріжджів вектора-наживки pGBT9 (Clontech) у вигляді однознімали з використанням фільтрувального паперу ланцюгового білка. Для того, щоб створити білок№50, Whatmann 3ММ [опис знімання колоній див. наживку, що мав би подібність до конформації у цит. вище, Sambrook et al.], потім поміщали приактивної каспази, видаляли пре домен, і малу сублизно на 20 секунд на алюмінієву фольгу, перебодиницю 2 (яка починається від серину 375 і заносили приблизно на 25 секунд у рідкий азот для кінчується аспарагіновою кислотою 479) гібридитого, щоб заморозити дріжджові клітини, витримузували з доменом зв'язування ДНК Gal4. С-Кінець вали приблизно 1 хвилину при кімнатній темперамалої субодиниці відокремлювали від N-кінця ветурі для того, щоб дріжджові клітини розмерзлися, ликої субодиниці 1 (яка починається від серину поміщали в чашку Петрі на фільтрувальний папір 217 і закінчується аспарагіновою кислотою 374) №1, Whatmann 3ММ, що попередньо змочували в 16-амінокислотним гліцин-сериновим лінкером Z-буфері [буфер для реакції з бета(GGGGSGGGGSGGGGSG). Таким чином, дві сугалактозидазою, див., наприклад, протоколи бодиниці активної каспази утворюються при спонClontech, цит. вище, Sambrook et al. або цит. вище танному утворенні складчастої структури однієї Current Protocols in Molecular Biology], що містить молекули. Цистеїн активного сайта в субодиниці 1 Xgal і бета-меркаптоетанол. Поява синього забармутували на серії (sub 1 C360S). Показано, що влення в колоніях вказує на активну бетанадекспресія такої одноланцюгової каспази-8 фунгалактозидазу. Білки, що взаємодіють із каспазоюкціонально схожа на надекспресію каспази-8 дико8, як правило, дають синє забарвлення при цьому го типу, що визначається по її здатності індукувати аналізі в інтервалі від хвилин до декількох годин, апоптоз в клітинах НЕК 293Т при надекспресії з переважно - від 5 хвилин до 3 годин, і найбільш вектора pcDNAHis, подібно до каспази-8. Активпереважно - в інтервалі від 5 хвилин до однієї гоність одноланцюгової каспази-8 можна блокувати дини. З іншого боку, здійснювали кількісну оцінку за допомогою коекспресії р35, що є інгібітором бета-галактозидазної активності шляхом вирощукаспаз. вання рідкої культури в середовищі, що містить Двогібридний скринінг здійснювали так, як Xgal, видалення клітин за допомогою центрифугуописано вище, за винятком того, що одноланцюговання і вимірювання поглинання середовища з ву каспазу-8, як описано вище, експресували з використанням спектрофотометру. вектора pGBT9 (Clontech). Клони кДНК, ідентифіковані в описаній вище Приклад 2 процедурі скринінгу, потім випробували далі на Ідентифікація білків, що взаємодіють із каспа 53 75564 54 зою-8 Приклад 2.5 Використовуючи модифіковану двогібридну Виявлено, що клон L5 кодує кДНК, ідентичну систему скринінгу, описану в прикладі ІА, ідентифібілку Тір60 (інвентарний номер 3024755). Тір60 кували декілька клонів, що кодують білки, які зв'я(взаємодіючий із Tat білок у 60кД) вперше описазуються із каспазою-8. Специфічність зв'язування ний як білок, що взаємодіє з клітинним трансактиклонів підтверджували при двогібридному досліватором HIV-Tat [Kamine et al., Virology, 216, 357дженні з дріжджами, у той час як виявлялася від366, 1996], а пізніше показано, що він має гістонсутність зв'язування з контрольними білками. Відіацетил-трансферазну активність [Yamamoto and брані клони кДНК виділяли та секвенували, і Horikoshi, 1997]. Біологічну роль Тір60, крім його нуклеотидні послідовності порівнювали з послідоздатності підсилювати Tat-опосередковувану активностями, знайденими в Genbank, як описано нижвацію промотору HIV, залишається визначити. че. При двогібридному дослідженні в дріжджах Приклад 2.1 виявлено, що клон L5 зв'язується з однією субодиВиявлено, що клон L1 містить неповну посліницею 2 каспази-8, а також із комплексом р10-р20. довність кДНК, ідентичну амінокислотам 690-750 Клон L5 розщеплювався каспазою-8 in vitro при Stat 1. Stat 1 ідентифікували як фактор транскрипаналізі in vitro із протеазами, описаному в прикладі ції, що зв'язується з інтерферон-стимульованим 2.4. Тір60 також розщеплювався залежно від кассприйнятливим елементом (ISRE) і з елементом пази після конадекспресії разом із рецептором для гамма-активованої послідовності (GAS) [огляд див. TNF р55 у клітинах HeLa і НЕК 293-Т. Він також у Iffic S.N., 1998]. Недавно показано, що Stat 1 зарозщеплювався в клітинах HeLa після обробки лучається до регуляції вмісту конститутивної касTNF навіть у відсутність інгібіторів синтезу білка. пази [Kuiner et al., 1997] і служить субстратом in Коротко, білок клонували у вектор pcDNAHis vitro для каспази-3 [King P. et al., 1998]. Зроблене (Invitrogen) і експресували в клітинах НЕК 293-Т припущення, що Stat 1 може грати деяку роль у або HeLa разом із білком, який індукує апоптоз регуляції самої апоптозної реакції. (наприклад, p55TNF-R або каспазою-8). Через 20 Приклад 2.2 годин після трансфекції клітини збирали і до лізаВиявлено, що клон L7 кодує неповну кДНК, що тів 0,5-1 10б цілих клітин застосовували SDSмайже цілком відповідає С-кінцевій частині клону PAGE і потім вестерн-блотінг з антитілами проти EST, знайденого в Genbank, якому функціонуванполі-His (Sigma). Результати візуалізували за доня не приписується (інвентарний номер помогою ECL. Виділили декілька клонів кДНК, що АА608733). кодують вставки, які відповідають білку Тір60. ВиПриклад 2.3 явлено, усі клони, включаючи "повнорозмірний, Виявлено, що клон L20 ідентичний амінокисдикого типу" клон Тір60, позбавлені сегмента пролотам 104-420 NEFA (інвентарний номер 462693). тягом від амінокислоти 94 (пролін) до 145 (треоNEFA є новим білком, який містить домен зв'язунін). Ця частина білка не є частиною ацетилазаквання основних амінокислот передбачуваної ДНК тивного сайта і не вважається суттєвою для із потенційним ядерним сигналом напрямку достафункціонування білка. Виявлено, що мутант Тір60, вки, дві області мотиву спіраль-петля-спіраль де залишки аспарагінової кислоти в позиціях 200 і (HLH), мотиви EF-hand, що діють спільно, кислотну 203 замінені залишками аланіну, не піддається багату амінокислотами область між EF-hand і лейрозчепленню. циновий мотив-блискавку [Barnikol-Watanabe et al., Приклад 2.6 1994]. Також виявлено, що NEFA є кальційзв'язуюВиявлено, що клон М26 кодує неповну кДНК, чим білком, і виявлено, що він локалізується як у що майже цілком відповідає клону EST людини, цитоплазмі, так і на поверхні клітини, і його також знайденому в Genbank (інвентарний номер можна виявити в культуральному середовищі. С18037). При дослідженні з 2 гібридами в дріжNEFA належить до субсімейства нуклеобіндину. джах виявлено, що клон М26 зв'язується з однією Однак біологічну роль NEFA поки не встановлено. субодиницею 2 каспази-8. Клон L20 розщеплюється каспазою-8 як in vitro, так Приклад 3 і in vivo. Виділення білків, що взаємодіють із каспазоюПриклад 2.4 8 Виявлено, що клон L12 кодує кДНК, що майже З використанням двогібридного скринінгу, опицілком відповідає клону EST людини, знайденому саного в прикладі 1В, ідентифікували декілька інв базі даних (інвентарний номер М62097). Клон ших клонів, що кодують специфічні білки, які зв'яL12 розщеплюється каспазою-8 in vitro, як показазуються з каспазою-8. Відібрані клони кДНК но при аналізі in vitro із протеазами. Коротко, синвиділяли та секвенували, і нуклеотидні послідовтезований in vitro мічений 35S білок інкубували ності порівнювали з послідовностями, знайденими протягом 30-60 хвилин у протеазному буфері в в Genbank, як описано нижче. присутності каспази-8, продукованої бактеріями. Скриніювали бібліотеку В-клітин [Durfee Т. et Білки і їхні фрагменти розділяли методом SDSal., 1993] і бібліотеку Т-клітин Юрката. В табл.2 PAGE, і результати візуалізували за допомогою наводиться початкова характеризація першої груауторадіографії або люмінофорів. Активність білка пи клонів. можна було блокувати специфічним інгібітором панкаспази z-VAD-фторметилкетоном. 55 75564 56 Таблиця 2 Клон(и) Декілька В33 В4.2 В8.1 В11 В17.1 В13.1 В22 В27 В37.1 J2 J40 Частина/гомолог Джерело lacZ (SFY) Довжина вставки NEFA ALL-клітини + 650-1100 Нуклеобіндин Ірr-клітини + 2000 КІАА0615 мозок самця +++ 3000 EST00156 +++ 1500 Фосфоетаноламінцитидилілтрансфераза +++ 1500 Н23509 мозок дитини ++ 1200 Рибонуклеозиддифосфатредуктаза EBV +++ 700 Циклофілін А + 700 gbAA936350 + 2700 EBV EBV ++++ 1400 АА746639 Людина ++++ 600 KIAA0419 мозок самця ++ 2300 Розщеплення in vivo/in vitro in vivo/in vitro in vitro in vivo/ in vitro in vitro in vitro in vivo/in vitro in vitro/in vivo in vitro in vivo: клітини 293-Τ та/або HeLa Неповні клони, що кодують частини NEFA, так само, як і клон В8.1, виділяли також методом із двома гібридами дріжджів, описаним у прикладі 1 А. Приклад 3.1 Клони В4 (інвентарний номер 3327044), В17 (інвентарний номер Н23509), В27 (інвентарний номер АА936350) і J40 (інвентарний номер 2887413), кодують вставки кДНК, гомологічні термінальному кінцю відповідних EST. Приклад 3.2 Клон В11 кодує кДНК-вставку, гомологічну Скінцю СТР-фосфоетаноламінцитидилілтрансферази (ЕТ, інвентарний номер D84307). ЕТ є ферментом, залученим до метаболізму фосфоліпідів, і каталізує конверсію фосфоетаноламіну в CDP-етаноламін [Nakashima et al., 1997]. Клон В11 розщеплювався in vitro каспазою8, що показано при аналізі, описаному вище в прикладі 2.4. Приклад 3.3 Клони В13 і В37 кодують кДНК-вставку, гомологічну С-кінцю клону, що кодує частину генома EBV (інвентарний номер V01555). Приклад 3.4 Клон В22 кодує кДНК-вставку, гомологічну Скінцю Т-клітинного циклофіліну (інвентарний номер Y00052). Т-Клітинний циклофілін ідентифікований як внутрішньоклітинний рецептор для циклоспорину А та FK506 і має активність справжньої пептидилпроліл-цис-транс-ізомерази [Haendler В. et al., 1987]. Приклад 3.5 Клон В33 кодує кДНК-вставку, гомологічну Скінцю нуклеобіндину (інвентарний номер 2506255). Нуклеобіндин є секретованим білком із властивостями зв'язування ДНК і Са2+, який дуже схожий на білок NEFA, описаний у прикладі 3.3. Нуклеобіндин спочатку описаний як білок у 55кД, що підсилює продукування антитіл проти ДНК у культурах автоімунних клітин Ірг селезінки миші [Miura К. et al., 1992]. Клон В33 розщеплювався каспазою-8 як in vitro, так і in vivo, як показали аналізи, зазначені вище в прикладах 2.4 і 2.5. Приклад 3.6 Клон J2 містить вставку в 600п.о., що кодує неповну білкову послідовність, яка виростає у поліпептид у ~20кД, коли експресується in vitro в безклітинній системі, а також у клітинах. Поліпептид, кодований клоном J2, розщеплюється in vitro каспазою-8 і in vivo у клітинах НЕК 293-Т і клітинах HeLa після коекспресії з p55TNF-R або каспазою-8 при аналізах, зазначених вище в прикладах 2.4 і 2.5. Нуклеотидна послідовність і розшифрована амінокислотна послідовність клону J2 наводяться на Фігурі 2. Порівняння і вистроювання в ряд 5'нарощування клону J2 за допомогою PCR із послідовностями, опублікованими в базі даних, показали гомологію клону J2 із EST людини (інвентарний номер АА460869), що відповідає передбачуваній N-ацетилглюкозамін-6-фосфатдеацетилазі людини, а також іншому клону (L48741), і дало склад передбачуваної полнорозмірної Nацетилглюкозамін-б-фосфатдеацетилази людини, наведеної на Фігурі 3. Приклад 4 Функціональна характеризація клону J2 Початкова функціональна характеризація клону J2 показала, що клон має інгібуючу активністю по відношенню до каспази-8 і апоптозу, індукує p55TNF-R людини в клітинах НЕК 293-Т і HeLa. Експресія J2 пригнічувала/сповільнювала апоптоз клітин НЕК 293-Т, котрансфікованих рецептором для TNF р55, або клітин HeLa, оброблених TNF і циклогексімідом, на 25-50%, що ілюструється Фігурою 3. Кількісне визначення загибелі внаслідок апоптозу здійснювали шляхом визначення частини клітин, що експресують бета-галактозидазу, які показують апоптозну морфологію через 20 годин після трансфекції зазначених конструкцій. Результати виражені у вигляді середнього відсотка синіх клітин, що показують ознаки апоптозу, як частини загального числа підрахованих синіх клітин (приблизно 500 клітин на зразок). З іншого боку, використовували білок зеленої флуоресценції в якості маркера, і здійснювали визначення за допомогою флуоресцентного конфокального мікроскопа. Приклад 5 Клонування білків, що взаємодіють із каспазою-8 57 75564 58 Бібліотеку кДНК плаценти людини, експресонеповної кДНК клону Р74 показує гомологію з посвану з вектора рАСТ2 (доступного від Clontech, лідовністю гістондеацетилази 6 (інвентарний ноPalo Alto, USA), скриніювали методом двогібридмер NP_006035). Вистроювання в ряд амінокислоного скринінгу з використанням наживки, описаної тної послідовності повнорозмірного білка, вище в прикладі ЇВ. Використовуючи таку процерозшифрованої із послідовності РАС, із послідовдуру скринінгу, ідентифікували декілька клонів, що ністю гістондеацетилази A (Genbank, інвентарний кодують специфічні білки, які взаємодіють із касномер NP_006028.1) показано на Фігурі 8. пазою-8. Також досліджували клони Ρ16, Р27, Р43, Приклад 6 Р70, Р74 і Р79. Функціональна характеризація білків, які взаєСеквенування кДНК-вставок клонів кДНК Р43, модіють із каспазою-8 Р16 і Р74 показало, що вони беруть участь у деА) Білки, кодовані клонами кДНК J2 або Ρ16, яких гомологічних послідовностях. Клон Р74 мав або Р43, або Р70, або Р74, або Р79, ідентифікованайбільш довгу кДНК-вставку приблизно в 3000п.о. ні методом двогібридного скринінгу, експресували і, як виявилося, кодує білок із розшифрованою в лізатах ретикулоцитів у присутності 35S метіонівідкритою рамкою зчитування з 574 амінокислот і з ну в системі лізатів сполучення ретикулоцитів ΤnΤ очікуваною молекулярною масою приблизно 68Τ7 (доступної від Promega, кат. №L4610) і розділя70кД. Послідовності трьох вищевказаних кДНКли в гелі методом SDS-PAGE (див. Фігуру 9А). Такі вставок порівнювали з послідовностями, знайдеж кількості білків, експресованих у лізатах ретикуними в опублікованих базах даних. Виявлено, що лоцитів, аналізували на зв'язування з гібридним кДНК-вставки клонів Р43, Р16 і Р74 мають деяку білком двох субодиниць каспази-8 GST-S2-31 гомологію з послідовністю двох клонів EST, знай(C360S) із вищеописаного приклада 1В, злитого з дених у Genbank (W04418 і N64095). Виявилося, GST і експресованого в бактеріях. Після попередщо послідовність усіх трьох кДНК-вставок станонього очищення за допомогою 1-годинної інкубації вить 3'-кінці різних сплайсинг-ізоформ того ж білка. при 4°С тільки з гранулами GST, продуковані ТпТ Вистоювання в ряд послідовностей трьох кДНКбілки осаджували гібридним білком GST-S2-S1 вставок підтвердило відкриту рамку зчитування з каспази-8, з'єднаного з гранулами GST за допомо574 амінокислот у послідовності Р74 (показана на гою 1-годинної інкубації при 4°С з гранулами GST, Фіг.5). Подібну послідовність також знайшли в геGST-преципітати промивали, і білки, що зв'язуютьномному клоні, ідентифікованому в інший опубліся з каспазою-8, відокремлювали від гранул за кованій базі даних (RPCI5-1057120; бібліотека допомогою кип'ятіння в буфері для зразків, що Roswell Park Cancer Institute Human РАС), що ломістить SDS, і розділяли методом SDS-PAGE. Білкалізується на хромосомі людини 12q31. Відкрита ки, здатні до зв'язування з конструкцією каспази-8, рамка зчитування, розшифрована з послідовності можна потім візуалізувати методом ауторадіограцього клону РАС, мала 1428 амінокислот (показафії. Виявилося, що білок, кодований клоном Р74, на на Фіг.6), і вона містить зазначену вище відкриспецифічно зв'язується in vitro із каспазою-8 (див. ту рамку зчитування, яка має 574 амінокислоти. Фігуру 9В), якщо порівнювати зі зв'язуванням GSTФігура 7 показує вистроєну в ряд послідовність S2-S1 (C360S) з Bid- відомим проксимальним субвідкритої рамки зчитування розшифрованої аміностратом каспази-8 у каскаді передачі сигналу апокислотної послідовності кДНК-вставки клону Р74 птозу Fas. Повнорозмірні білки, отримані в лізаті (позначеної "клонована") із відкритою рамкою зчиретикулоцитів, позначені на Фігурі зірочками. тування, розшифрованою із послідовності клону При дослідженнях із двома гібридами, описаРАС (позначеною "розшифрована"). них вище в прикладі 1А, виявлено, що білок, кодоПри порівнянні розшифрованої амінокислотної ваний Р74, зв'язується з гібридним білком, кодовапослідовності клону Р74 і послідовності, розшифним двома субодиницями каспази-8, а не з малою рованої з повнорозмірного клону РАС, виявилося, субодиницею каспази-8, експресованою окремо. що повнорозмірний білок, що відповідає Р74, є Для порівняння, інші білки, зазначені в приведених довшим в 5'-кінці і може, мабуть, починатися з вище прикладах, такі як Тір60, зв'язуються з мапершого або одного з перших метіонінів послідовлою субодиницею 32 каспази-8, коли вона експреності РАС, показаної на Фігурі 6. сується одна (дані не наводяться). Також виявлено, що послідовність клону Р74 B) Білок, кодований кДНК Р43, експресували in показує суттєву гомологію з високогомологічною vitro у лізатах ретикулоцитів у присутності 35S меобластю гістондеацетилаз миші і людини, областіоніну з використанням системи лізатів сполучентю, що могла бути доменом, який містить сайт ня ретикулоцитів TnT T7 і піддавали розчепленню ферментної активності гістондеацетилази, причорекомбінантом дикого типу або мутованою каспаму передбачається, що білок, кодований Р74, мозою-3, або каспазою-9, або каспазою-10, експреже розділяти функцію цих білків. Послідовність в сованими в Е. соlі у вигляді позначених гістидином області 163-1716п.о. неповної кДНК клону Р74 гібридних білків субодиниці 2 - субодиниці 1 (S2показує приблизно 80% гомологію з гістондеацеS1). Каспазу-8 експресували у вигляді позначеного тилазою А (інвентарний номер NP_006028). Послігістидином гібридного білка субодиниці 1 - субодидовність в області 385-1707п.о. неповної кДНК ниці 2 (S1-S2). В аналізі з протеазами використоклону Р74 показує гомологію з послідовністю гісвували 1 і 4 об'єми повного бактеріального лізату, тондеацетилази 5 (інвентарний номер NP 005465). визначених як 1 і 4 відносних одиниці (RU) (Фігура Послідовність в області 418-1629п.о. неповної 10). Коротко, синтезовані in vitro мічені 35S білки кДНК клону Р74 показує гомологію з послідовністю інкубували в протеазному буфері протягом 30 гістондеацетилази mHDAl (інвентарний номер хвилин при 37°С в присутності каспази-8, продукоAAD09834), і послідовність в області 424-1551п.о. ваної бактеріями. Білки і їхні фрагменти розділяли 59 75564 60 методом SDS-PAGE і результати візуалізували Незважаючи на те, що даний винахід описаний методом ауторадіографії або за допомогою люміу зв'язку з конкретними варіантами його втілення, нофорів. Результати показали, що білок, кодоваварто мати на увазі, що можливі інші модифікації. ний кДНК Р43, використовуваний у якості субстраМається на увазі, що дана заявка перекриває ту, ефективно розщеплювався каспазою-8, слабко будь-які варіації, застосування або адаптації винарозщеплювався каспазою-10 і не розщеплювалася ходу, які випливають, узагалі, з основних полоні каспазою-3, ні каспазою-9 (Фігура 10). Таким жень винаходу і які включають такі відхилення від чином, виявилося, що білок є специфічним субданого опису, що виникають у відомій або звичайстратом каспази-8. ній практиці в області техніки, до якої відноситься C) Для аналізу дії знову клонованих білків на винахід, і які можна застосувати до суттєвих ознак, апоптозну загибель клітин, індуковану каскадом викладеним вище, як випливає з обсягу формули передачі сигналу рецептором для TNF, відібрані винаходу, що додається. кДНК клонували у експресуючі вектори pcDNA Усі посилання, цитовані тут, включаючи жур3.1/His С (доступні від Invitrogen) і нестійко котраннальні статті або реферати, опубліковані або такі, сфікували рецептором для TNF р55 у векторі що відповідають заявкам на патент США або інpcDNA 3 (Invitrogen) і білком зеленої флуоресценших країн, або будь-які інші посилання, включені в ції (GFP), експресованим з експресуючого вектора даний опис у якості посилань у всій повноті, вклюpEGFPCI (Clontech), у клітини НЕК 293-Т. Клонучаючи всі дані, таблиці, фігури і тексти, подані в цитованих посиланнях. Крім того, весь зміст цитовали кДНК Тір60 і 32-Тір60, позбавлений перших ваних тут робіт також у всій повноті включений у 32 N-кінцевих амінокислот, у векторі pCGN [описаякості посилання. ний М. Tanaka and W. Herr, Cell, 60, 375-386, 1990], Посилання на відомі стадії способів, стадії у якому гемаглутинінова (НА) мітка злита з Nтрадиційних способів, відомі способи або традикінцем кДНК, і використовували в такому ж експеційні способи ніяким чином не є визнанням факту, рименті. що якийсь аспект, опис або варіант втілення даноЧерез 24 години трансфіковані клітини перевіго винаходу розкриваються, пояснюються або перяли під флуоресцентним мікроскопом, і загибель редбачаються в родинних технічних вирішеннях. клітин підраховували шляхом визначення числа Вищевикладений опис конкретних варіантів клітин із усієї популяції флуоресціюючих клітин, що втілення винаходу буде настільки повно розкривапоказують апоптозну морфологію. Виявили, що ти загальний характер винаходу, що інші можуть, білок Р74, експресований у надлишку з неповної використовуючи знання в даній області техніки кДНК, клонованої у вектор pcDNA-His, захищає (включаючи вміст цитованих тут посилань), легко клітини НЕК 293-Т і HeLa від загибелі, викликаної модифікувати та/або адаптувати для різних застонадекспресією рецептора для TNF р55 (Фігура 11) сувань такі конкретні варіанти втілення винаходу або надекспресією злитих двох субодиниць каспабез зайвого експериментування, без відходу від зи-8 (не показане). Виявлено, що Тір60 є дикого загальної концепції даного винаходу. Отже, маєтьтипу і нездатний до розчеплення мутант Тір30, де ся на увазі, що такі адаптації і модифікації знахозалишки аспарагінової кислоти в позиціях 200 і 203 дяться в межах цілей і кола еквівалентів описаних замінені на залишки аланіну, захищають клітини варіантів втілення винаходу, заснованих на поясНЕК 293-Т від загибелі, викликаною надекспресією неннях і вказівках, викладених тут. рецептора для TNF р55 (Фігура 12), у той час як Варто мати на увазі, що фразеологія або терА32-Тір60, позбавлений перших 32 N-кінцевих мінологія використані тут для цілей опису, а не для амінокислот, не робив такої захисної дії. обмеження, так що термінологія або фразеологія в Маючи тепер повний опис даного винаходу, описі даного винаходу повинні інтепретуватися фахівцям у цій області техніки варто мати на увазі, фахівцями у світлі пояснень і вказівок, поданих що такі ж операції можна виконати із широким рятут, у сполученні зі знаннями рядового фахівця в дом еквівалентних параметрів, концентрацій і цій області техніки. умов без відходу від сутності та обсягу винаходу і без зайвого експериментування.
ДивитисяДодаткова інформація
Назва патенту англійськоюHuman n-acetylglucosamine-6-phosphate deacetylase capable to interact with subunit 1 and/or subunit 2 of caspase-8, isolated dna sequence coding thereof, method for preparation of said protein (variants), antibody directed at an epitope of human n-acetylglucosamine-6-phosphate deacetylase, set for immunoassay intended for detection thereof, use of n-acetylglucosamine-6-phosphate deacetylase as medicament for treatment diseases related to modulation of caspase-8 activity
Назва патенту російськоюN-ацетилглюказамин-6-фосфатдеацетилаза человека, способная взаимодействовать с субединицей 1 и/или субединицей 2 каспазы-8, выделенная последовательность днк, которая ее кодирует, способ получения указанного белка (варианты), антитело, направленное против эпитопа n-ацетилглюказамин-6-фосфатдеацетилазы человека, набор для иммуноанализа, предназначенный для ее выявления, применение n-ацетилглюказамин-6-фосфатдеацетилазы как лечебного средства для терапии заболеваний, связанных с необходимостью модуляции активности каспазы-8
МПК / Мітки
МПК: A61P 1/00, C12N 15/12, C12Q 1/68, C07K 16/18, C07K 14/47, A61K 38/17, A61K 39/395, C12N 5/10, A61P 3/10, G01N 33/563
Мітки: білка, послідовність, спосіб, каспази-8, днк, взаємодіяти, одержання, антитіло, варіанти, n-ацетилглюкозамін-6-фосфатдеацетилаза, кодує, субодиницею, даного, виділена, здатна, людини, яка
Код посилання
<a href="https://ua.patents.su/42-75564-n-acetilglyukozamin-6-fosfatdeacetilaza-lyudini-yaka-zdatna-vzaehmodiyati-z-subodiniceyu-1-i-abo-subodiniceyu-2-kaspazi-8-vidilena-poslidovnist-dnk-shho-kodueh-sposib-oderzhannya-d.html" target="_blank" rel="follow" title="База патентів України">N-ацетилглюкозамін-6-фосфатдеацетилаза людини, яка здатна взаємодіяти з субодиницею 1 і/або субодиницею 2 каспази-8, виділена послідовність днк, що її кодує, спосіб одержання даного білка (варіанти), антитіло,</a>
Послідовність днк, що кодує поліпептид, який має активність білка g1 і який здатний зв’язуватись або взаємодіяти прямо або опосередковано з mort-1 і/або з будь-якими білками, що зв’язуються з mort-1, і здатний

Номер патенту: 74529
Опубліковано: 16.01.2006
Автори: Ковалєнко Андрєй, Варфоломєєв Євгєній, Гольцев Юра, Валлах Девід, Бродянскій Вадім
МПК: C12N 1/19, C12N 1/15, A61P 43/00, A61P 31/18, C12Q 1/68, A61K 38/00, C12N 9/00, A61K 31/7105, A61K 48/00, G01N 33/15, C12N 5/10, G01N 33/50, C12N 15/09, C12N 9/64, A61K 35/76, A61P 35/00, C12N 15/12, C12N 1/21
Мітки: взаємодіяти, білками, білка, днк, поліпептид, зв'язуються, здатний, зв'язуватись, послідовність, mort-1, кодує, має, будь-якими, активність, опосередкованої, прямої
Формула / Реферат:
1. Послідовність ДНК, яка кодує поліпептид, що має активність білка G1, при тому, що зазначений поліпептид:(і) здатний зв'язуватися або взаємодіяти прямо або опосередковано з MORT-1 і/або з будь-якими білками, що зв’язуються з MORT-1, або з іншими внутрішньоклітинними білками-медіаторами/модуляторами; і(іі) здатний опосередковувати внутрішньоклітинний вплив, опосередкований FAS-R або p55-TNF-R, aбo інших цитотоксичних...
Виділена молекула днк, яка кодує людський еритропоетин (варіанти), біологічно функціональний кільцевий плазмідний або вірусний днк-вектор, штам еукаріотичних клітин-хазяїв (варіанти), спосіб одержання поліпепти

Номер патенту: 54363
Опубліковано: 17.03.2003
Автор: Фу-Куен Лін
МПК: C12N 15/63, A61K 38/00, C12P 21/00
Мітки: одержання, спосіб, клітин-хазяїв, людський, виділена, вірусний, функціональний, днк, молекула, яка, еукаріотичних, еритропоетин, кодує, варіанти, днк-вектор, біологічно, штам, плазмідний, кільцевий, поліпепти
Формула / Реферат:
1. Выделенная молекула ДНК, кодирующая человеческий эритропоэтин и имеющая следующую нуклеотидную последовательность:2. Выделенная молекула ДНК, кодирующая человеческий эритропоэтин и имеющая следующую нуклеотидную последовательность:3. Выделенная молекула ДНК, кодирующая человеческий эритропоэтин и имеющая следующую нуклеотидную последовательность:4. Выделенная молекула...
Плазмідний експресуючий вектор р32, кодуючий послідовність днк, відповідаючу за експресію гібридного злитного білка env-2 вірусу імунодефіциту людини другого типу( env bіл-2) в бактеріях e. coli

Номер патенту: 14843
Опубліковано: 18.02.1997
Автори: Чумак Ростислав Максимович, Чєрєпанов Пьотр Алєксєєвіч, Мартиненко Дмитро Леонідович, Міхайлова Татьяна Гавріловна
МПК: C12N 15/49, C12N 15/62, C12N 15/70
Мітки: гібридного, типу, кодуючий, вірусу, плазмідний, білка, людини, env-2, імунодефіциту, експресію, bіл-2, бактеріях, злитного, послідовність, експресуючий, другого, відповідаючу, вектор, р32, днк
Формула / Реферат:
Плазмідний експресуючий вектор р32, кодуючий послідовність ДНК, відповідаючу за експресію гібридного элитного білка Env-2 вірусу імунодефіциту людини другого типу (Env ВІЛ-2) в бактеріях Е.соlі, має розмір 5,2 т.п.н., має делецію 630 п.н. в області гена бета-галактозідази, в склад якого входять нуклеотидні послідовності, кодуючі Env-2 білки ВІЛ-2 gP110, gP36 і кодує під контролем промотора фага лямбда Рr синтез гібридного білка загальною...
Плазмідний експресуючий вектор рк1724, кодуючий послідовність днк, відповідаючу за експресію гібридного злитного білка gag вірусу імунодефіциту людини першого типу (gag biл-1) в бактеріях е.coli

Номер патенту: 14804
Опубліковано: 18.02.1997
Автори: Чумак Ростислав Максимович, Мартиненко Дмитро Леонідович, Чєрєпанов Пьотр Алєксєєвіч, Міхайлова Татьяна Гавріловна
МПК: C12N 15/62, C12N 15/49, C12N 15/70
Мітки: білка, рк1724, людини, вірусу, експресію, кодуючий, biл-1, днк, послідовність, вектор, злитного, e.coli, гібридного, експресуючий, бактеріях, типу, відповідаючу, першого, імунодефіциту, плазмідний
Формула / Реферат:
Плазмідний експресуючий вектор рК1724, кодуючий послідовність ДНК, відповідаючу за експресію гібридного элитного білка GAG вірусу імунодефіциту людини першого типу (GAG ВІЛ-1) в бактеріях E.coli, який має розмір 5,3 т.п.н., має делецію 630 т.п.н. в області гена бета-галактозидази, в склад якого входять нуклеотидні послідовності, кодуючі GAG білки ВІЛ-1 р17, р24, р15 і кодує під контролем промотора фага лямбда Рr синтез гібридного білка...
Плазмідний експресуючий вектор рк41,кодуючий послідовність днк, відповідаючу за експресію гібридного злитного білка еnv-1 вірусу імунодефіциту людини першого типу ( env віл-1) в бактеріях e.coli

Номер патенту: 14844
Опубліковано: 18.02.1997
Автори: Мартиненко Дмитро Леонідович, Чєрєпанов Пьотр Алєксєєвіч, Міхайлова Татьяна Гавріловна, Чумак Ростислав Максимович
МПК: C12N 15/70, C12N 15/62, C12N 15/49
Мітки: людини, e.coli, вірусу, злитного, першого, гібридного, білка, еnv-1, плазмідний, бактеріях, імунодефіциту, експресію, рк41,кодуючий, відповідаючу, типу, вектор, послідовність, експресуючий, віл-1, днк
Формула / Реферат:
Плазмідний експресуючий вектор рК41, кодуючий послідовність ДНК, відповідаючу за експресію гібридного элитного білка Env-1 вірусу імунодефіциту людини першого типу (Env ВІЛ-1) в бактеріях Е.соlі, що має розмір 4,9 т.п.о., має делецію 400 п.о. в області гена бета-галактозідази, в склад якого входять нуклеотидні послідовності, кодуючі Env-1 білки ВІЛ-1 gP120, gP41 і кодує під контролем промотора фага лямбда Рr синтез гібридного білка...
Випадковий патент: Дисковий ніж