Трансгенна однодольна рослина з підвищеною ефективністю використання азоту














































Формула / Реферат
1. Трансгенна однодольна рослина або частина однодольної рослини, яка містить полінуклеотид, який кодує білок, що відповідає за утилізацію азоту, який вибирають з амінотрансферази, такої як аланінамінотрансферази або аспартатамінотрансферази, який функціонально зв'язаний з РВрr1 промотором, який містить нуклеотидну послідовність SEQ ID NO: 1.
2. Трансгенна однодольна рослина або частина однодольної рослини за п. 1, де рослиною є рис, ячмінь, пшениця або кукурудза.
3. Трансгенна однодольна рослина або частина однодольної рослини за п. 1, де рослиною є рис.
4. Трансгенна однодольна рослина або частина однодольної рослини за будь-яким із пп. 1-3, де амінотрансферазою є аланінамінотрансфераза.
5. Трансгенна однодольна рослина або частина однодольної рослини за будь-яким із пп. 1-4, де амінотрансферазою є аланінамінотрансфераза, яка містить амінокислотну послідовність SEQ ID NO: 3.
6. Насіння, одержане з трансгенної однодольної рослини за будь-яким із пп. 1-5, яке містить полінуклеотид, що кодує амінотрансферазу, таку як аланінамінотрансфераза або аспартатамінотрансфераза, який функціонально зв'язаний з РВрr1 промотором, який містить нуклеотидну послідовність SEQ ID NO: 1.
7. Клітина однодольної рослини, трансформована за допомогою полінуклеотиду, що кодує амінотрансферазу, таку як аланінамінотрансфераза або аспартатамінотрансфераза, який функціонально зв'язаний з РВрr1 промотором, який містить нуклеотидну послідовність SEQ ID NO: 1.
8. Клітина однодольної рослини за п. 7, де рослинна клітина є рослинною клітиною рису, рослинною клітиною ячменю, рослинною клітиною пшениці або рослинною клітиною кукурудзи.
9. Клітина однодольної рослини за п. 7, де рослинна клітина є рослинною клітиною рису.
10. Клітина однодольної рослини за будь-яким із пп. 7-9, в якій амінотрансферазою є аланінамінотрансфераза.
11. Рослинна клітина за будь-яким із пп. 7-9, в якій амінотрансферазою є аланінамінотрансфераза, що містить амінокислотну послідовність SEQ ID NO: 3.
12. Генетична конструкція, яка містить полінуклеотид, що кодує амінотрансферазу, таку як аланінамінотрансфераза або аспартатамінотрансфераза, який функціонально зв'язаний з РВрr1 промотором, який містить нуклеотидну послідовність SEQ ID NO: 1.
13. Генетична конструкція за п. 12, в якій амінотрансферазою є аланінамінотрансфераза.
14. Генетична конструкція за п. 12, в якій амінотрансферазою є аланінамінотрансфераза, що містить амінокислотну послідовність SEQ ID NO: 3.
15. Генетична конструкція за будь-яким із пп. 12-14, в якій конструкція додатково містить послідовність термінації транскрипції, ділянку поліаденілування або обидві з них.
16. Вектор, який містить генетичну конструкцію за будь-яким із пп. 12-15.
17. Клітина-хазяїн, яка містить генетичну конструкцію за будь-яким із пп. 12-15.
18. Спосіб одержання однодольної рослини, що має підвищену ефективність використання азоту, який включає трансформацію клітини однодольної рослини за допомогою генетичної конструкції за будь-яким із пп. 12-15, і вирощування трансформованої клітини однодольної рослини для одержання однодольної рослини, що експресує амінотрансферазу з РВрr1 промотором, що приводить до створення однодольної рослини, яка має підвищену ефективність використання азоту, де підвищена ефективність використання азоту порівнюється з ефективністю використання азоту однодольними рослинами дикого типу, вирощеними за ідентичних умов.
19. Спосіб одержання однодольної рослини, яка має підвищену біомасу, який включає трансформацію клітини однодольної рослини за допомогою генетичної конструкції за будь-яким із пп. 12-15, і вирощування трансформованої клітини однодольної рослини для одержання однодольної рослини, що експресує амінотрансферазу з РВрr1 промотором, що приводить до створення однодольної рослини, яка має підвищену біомасу, де підвищена біомаса порівнюється з біомасою рослин дикого типу, вирощених за ідентичних умов.
20. Спосіб одержання однодольної рослини, яка має підвищену врожайність насіння, який включає трансформацію клітини однодольної рослини за допомогою генетичної конструкції за будь-яким із пп. 12-15, і вирощування трансформованої клітини однодольної рослини для одержання однодольної рослини, що експресує амінотрансферазу з РВрr1 промотором, що приводить до створення однодольної рослини, яка має підвищену врожайність насіння, де підвищена врожайність насіння порівнюється з врожайністю насіння рослин дикого типу, вирощених за ідентичних умов.
21. Застосування генетичної конструкції за будь-яким із пп. 12-15 для підвищення ефективності використання азоту, біомаси або врожайності насіння в однодольних рослин.
Текст
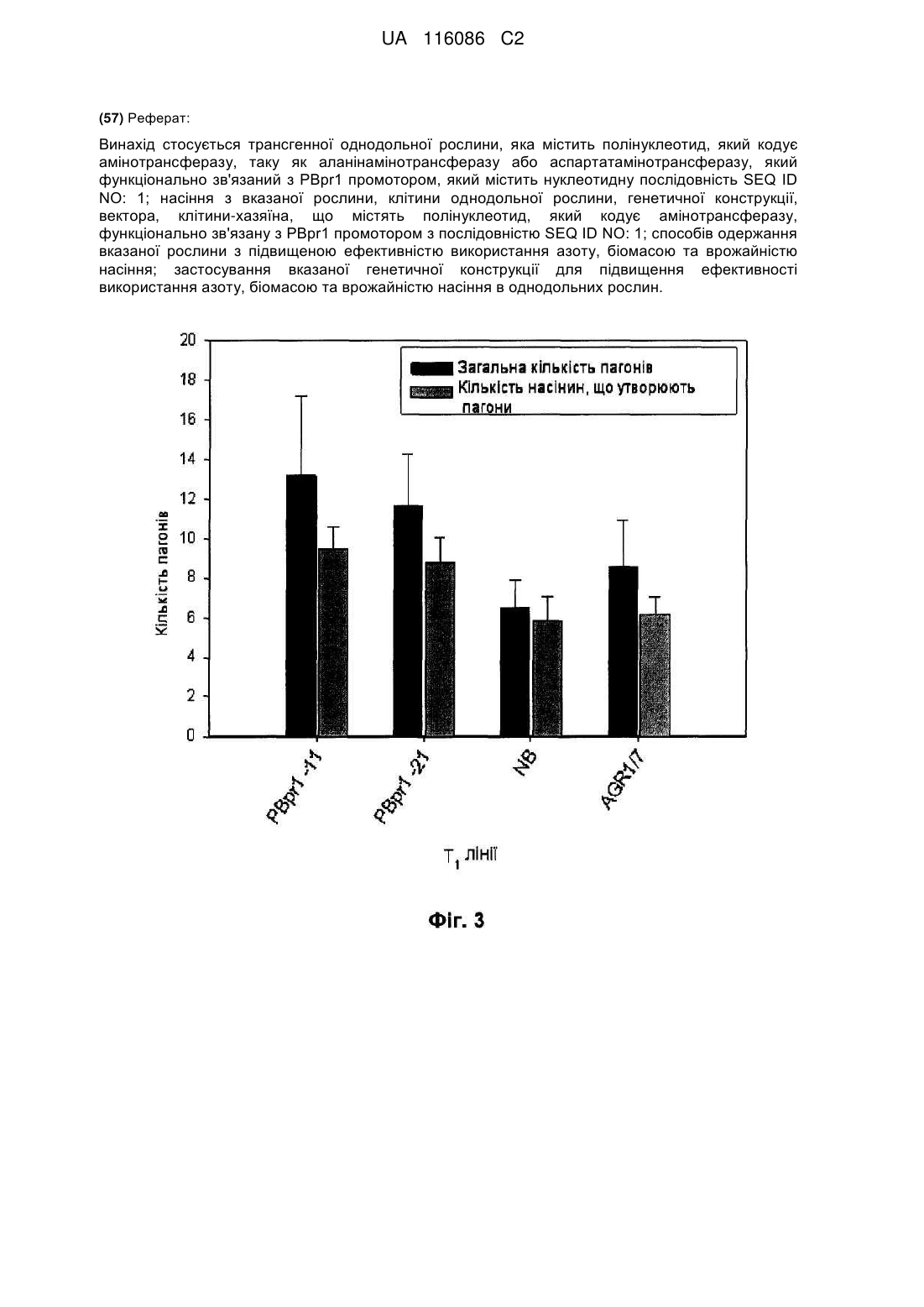
Реферат: Винахід стосується трансгенної однодольної рослини, яка містить полінуклеотид, який кодує амінотрансферазу, таку як аланінамінотрансферазу або аспартатамінотрансферазу, який функціонально зв'язаний з РВрr1 промотором, який містить нуклеотидну послідовність SEQ ID NO: 1; насіння з вказаної рослини, клітини однодольної рослини, генетичної конструкції, вектора, клітини-хазяїна, що містять полінуклеотид, який кодує амінотрансферазу, функціонально зв'язану з РВрr1 промотором з послідовністю SEQ ID NO: 1; способів одержання вказаної рослини з підвищеною ефективністю використання азоту, біомасою та врожайністю насіння; застосування вказаної генетичної конструкції для підвищення ефективності використання азоту, біомасою та врожайністю насіння в однодольних рослин. UA 116086 C2 5 10 15 20 25 30 35 40 45 50 55 60 ОБЛАСТЬ ТЕХНІКИ, ДО ЯКОЇ НАЛЕЖИТЬ ВИНАХІД Винахід належить до однодольних рослин, які мають підвищену урожайність і/або ефективність використання азоту (nitrogen utilization efficiency-NUE), способів підвищення урожайності та NUE у однодольних рослин, та способів підвищення біомаси і врожаю насіння у однодольних рослин, наприклад, при вирощуванні за умов азотного дефіциту. РІВЕНЬ ТЕХНІКИ Продуктивність рослин у більшості природних та сільськогосподарських екосистем лімітується трьома основними поживними елементами: азотом, фосфором і калієм. Загалом, азот є найважливішим серед трьох лімітуючих поживних елементів і основним компонентом добрив. Оскільки азот, як правило, є елементом, що лімітує швидкість росту у рослин, урожайність більшості сільськогосподарських культур значною мірою залежить від внесення неорганічних азотних добрив. Джерелом азоту в добриві, як правило, є нітрат амонію, нітрат калію або сечовина. Підвищена ефективність використання азоту рослинами має низку переваг, обумовлюючи, наприклад, прискорений ріст та підвищений врожай у порівнянні з традиційними рослинами, що вирощуються на ґрунтах з низьким вмістом азоту, і знижена потреба у внесенні азотних добрив під сільськогосподарські культури. Витрати на добрива складають істотну частку коштів, пов'язаних з виробництвом сільськогосподарських культур, відповідно, використання меншої кількості добрив призвело б до зменшення витрат виробниками. Зменшення кількості добрив також призвело б до зменшення негативного впливу на середовище, який викликає надмірне застосування азотних добрив. Надмірне застосування добрива призводить до підвищеної евтрофікації, кислотних дощів, ацидифікації ґрунту і парникового ефекту. Ці загрози навколишньому середовищу обумовлюють наступні проблеми, такі як загибель риби, втрата біорізноманіття, підвищене водоростеве "цвітіння" води, втрата орних земель і прискорення глобальних змін клімату, які впливають на населення землі як у соціальному, так і в економічному масштабі. Однодольні складають значний відсоток сільськогосподарських культур, які вирощуються у світі, при цьому у 2007 році пшеницею було засіяно приблизно 217 мільйонів гектарів і 158 мільйонів гектарів кукурудзою та рисом. У глобальному масштабі приблизно половина калорій і потреби у протеїні походять від пшениці, рису та кукурудзи. Рис здебільшого використовують як модельну сільськогосподарську культуру для генетичних та фізіологічних досліджень у інших однодольних культур, включаючи кукурудзу, пшеницю, цукрову тростину, ячмінь, сорго, жито та інші злаки. Рис має невеликий диплоїдний геном, який добре зберігається і відтворюється серед однодольних. Промотори – це послідовності нуклеїнової кислоти, що регулюють транскрипцію гена чи нуклеотидної послідовності. Промотори можуть використовуватись для конститутивної експресії, такі як добре відомий промотор віруса мозаїки цвітної капусти CaMV 35S, індуцибельної експресії, такий як промотор rd29A, що індукується стресом (Pino et al., 2007), тканиноспецифічної експресії, такий як коренеспецифічний промотор OsANT1 (Публікація заявки на Патент США № 2009/0288224) і стадієспецифічної експресії, такі як IPT промотори, що індуються старінням (Ma et al., 2009). Промотори також можуть бути слабкими або сильними, залежно від того, коли і де вони індукуються, вони можуть використовуватись для експресії прикріпленого гена або нуклеотидної послідовності на різних рівнях. Якщо промотор зливається з 5" кінцем гена або нуклеотидної послідовності, він може регулювати експресію гена або нуклеотидної послідовності. Разом з тим, не всі промотори можуть успішно експресувати всі гени або всі нуклеотидні послідовності у рослин всіх типів. Наприклад, промотор дводольних може функціонувати у інший спосіб при введенні в систему однодольних і навпаки. Аналогічно, промотор однодольних може функціонувати по-іншому при введенні у різні роди однодольних, так як, наприклад, промотор рису – у пшеницю. Для трансгенних досліджень існують різні типи промоторів, які можуть використовуватись, залежно від мети експерименту. Промотори часто класифікуються як конститутивні, тканиноспецифічні і/або індуцибельні. Багато трансгенних досліджень наразі використовують універсальні конститутивні промотори, такі як вірус мозаїки цвітної капусти (CAMV35S) і убіхітиновий промотор 1 кукурудзи (ubi-1), для того щоб викликати надекспресію гена інтересу у рослин. Це може бути недоліком, тому що це було б енергетично несприятливим для рослин експресувати ген протягом усього часу - і призвело б до аномального розвитку, оскільки рівні експресії трансгена не регулюються (Shelton et al., 2002). Наприклад, конститутивна надекспресія гена целюлозосинтази типу CslF6 промотором глобуліну вівса ProASGL часто призводить до зниженого проростання насіння чи росту сіянців та появи некрозів на кінчиках листків та у серйозних випадках обумовлює загибель (Burton et al., 2011). 1 UA 116086 C2 5 10 15 20 25 30 35 40 45 50 55 60 Використання індуцибельних або тканиноспецифічних промоторів є кращим вибором, для того щоб викликати експресію трансгена. Індуцибельні промотори активують експресію гена лише у тому випадку, коли застосовуються специфічні фізичні, біологічні або хімічні стимули чи чинники навколишнього середовища. Промотор гена Hvhsp17 пшениці, який індукується теплом, може бути використаний для забезпечення високого рівня експресії гена інтересу, коли рослини зазнають впливу температури від 38 до 40 °C протягом 1-2 годин (Freeman et al., 2011). Це стимулює експресію гена чи нуклеотида протягом короткого проміжку часу і дає змогу контролювати особливості розвитку, однак тут існують обмеження щодо тканин, які не зазнають суттєвих температурних змін. Надекспресія білка NAC (TaNAC69) у Triticum aestivum, який кодує транскрипційний фактор, що бере участь у водному стресі, через індукцію промотора HvDhn4s, який ініціюється посухою, забезпечує рослини пшениці з істотно вищою біомасою пагонів на ранніх стадіях вегетативного розвитку за умов слабкого сольового стресу та водного дефіциту порівняно з рослинами дикого типу. На противагу, конститутивний промотор HvDhn8s, який індукує TaNAC69, не виявив істотних відмінностей порівняно з нетрансформованими контрольними рослинами (Xue et. al., 2011). При використанні промотора rdA29, який індукується водним та холодовим стресом, для того щоб викликати надекспресію DREB1A гена, що кодує фактор транскрипції, залучений до стійкості до стресу у рослин Arabidopsis, утворюються нормальні рослини, в той час як конститутивна експресія, викликана CAMV35S промотором, за нормальних умов культивування призводить до затримки росту (Kasuga et al., 1999). Тканиноспецифічні промотори залучаються при органоспецифічній і стадієспецифічній експресії у трансгенних рослин. Наприклад, у картоплі, StRCAp вбудовується в листки як захисний механізм проти комах-шкідників, однак не експресується у їстівних частинах рослини, що продукує цей токсин (Weber, 2003; Park and Jones, 2008). Використання 35S промотора викликає також проблему продовольчої безпеки, у тому випадку, коли токсин, що продукується у "нецільових" органах, може зробити бульби картоплі небезпечними для споживання. Крім того, у метаболічному відношенні це є значним навантаженням для рослини, оскільки змушує її постійно продукувати вторинні метаболіти, незалежно від стадії розвитку органа, що призводить до ослаблення рослини та ризику зниження врожаїв. У випадку конструювання NUE рослини, тканиноспецифічна експресія генів може підвищити ефективність поглинання N, його використання та транспорт у рослині. Навпаки, використання конститутивного промотора може підтвердити витрати енергії, тому що надекспресія нелімітуючих швидкість ферментів у деяких органах може не призвести до прояву будь-якого фенотипу, або може навіть знизити врожай. Отримання NUE рослини може також включати трансформацію множинних генів або "генне пакетування", для того щоб отримати задовільний NUE фенотип, оскільки метаболізм і транспорт азоту є надзвичайно складними процесами. До того ж, використання CAMV35S промотора може не призвести до будь-якого фенотипу, тому що експресія гена чи експресія білка були недостатніми у специфічному органі чи на певній стадії розвитку. Також, рослини можуть "вимикати" експресію трансгена, коли виявлється, що це є несприятливим у енергетичному відношенні. Якщо HvAlaAT індукується CAMV35S промотором, він не виявляє ніякого NUE фенотипу. Однак, при використанні коренеспецифічного промотора btg26, продукуються рослини, яким притаманні вищі значення ефективності використання азоту (NUE) (Good et al., 2007). Подібно до надекспресії генів інтересу, при використанні у різних видів промотори можуть не відтворювати паттерн експресії їх природних видів. Насіннєспецифічні промотори ячменю (B-hor і D-hor) та пшениці (HMW-Glu) не індукують насіннєспецифічну експресію у рису; натомість промотори індукують високий рівень експресії в листках, пагонах і материнських тканинах насінин у рослин рису (Wu et al., 1998; Qu and Takaiwa, 2004; Furtado et al., 2008). PBpr1 промотори – походять з генів, що кодують метилмалонат семіальдегід дегідрогеназу (MMSDH). PBpr1 промотор розташований вище OsALDH6 гена, який кодує ген метилмалонат семіальдегід дегідрогенази у рису (Інвентарний номер: ген: AK 121280.1 і мРНК: AF045770.1). AK121280.1 і AF045770.1 являють собою сплайсингові варіанти один одного, причому AF045770 є коротшим, ніж AK121280.1. AK121280.1, очевидно, є гіпотетичним білком, сконструйованим GenBank, тоді як було описано AF045770.1 (Oguchi et al (2004). OsALDH6 є гомологічним до гена ALDH6B2 у Arabidopsis, який також кодує метилмалонат семіальдегід дегідрогеназу. Ген OsALDH6 високо експресується в молодих коренях та стеблах (Gao and Han, 2009). Метилмалонат семіальдегід дегідрогеназа (MMSDH) каталізує незворотнє окисне декарбоксилювання малонату семіальдегіду до ацетил-CoA і метилмалонат семіальдегіду до пропіоніл-CoA в дистальних частинах катаболічного шляху валіну та піримідину. Оскільки MMSDH генерує ацетил-CoA, це є важливим чинником у гліоксилатному циклі, циклі 2 UA 116086 C2 5 10 15 20 25 30 35 40 45 50 55 60 трикарбонових кислот (ТКК) і утворенні жирних кислот. MMSDH даун-регулюється протягом окиснювального стресу завдяки рестрикції в циклі трикарбонових кислот і утворення ATФ (Sweetlove et al., 2002). У двотижневих рослин рису були виявлені високі рівні мРНК MMSDH у коренях та піхвах листків, тоді як накопичення білка було найвищим у коренях, після чого йдуть листкові пластинки (Oguchi et al…, 2004). При додаванні ауксину, рівні MMSDH у коренях зростають разом зі зростанням укорінення. Tanaka et al. (2005) висловили припущення, що MMSDH бере участь у рості коренів, диференціації тканин та у рості в товщину завдяки його експресії в кореневому чохлику, бічних коренях та кореневих волосках. Було виявлено, що MMSDH локалізується у мітохондріальному матриксі Arabidopsis, рису, людини, бика і щура, що дає підстави висловити припущення про подібність функцій MMSDH у цих організмів. Ця інформація стосовно передумов винаходу забезпечується з метою ознайомлення з інформацією, яка, на думку заявника, може мати відношення до даного винаходу. Не існує жодної потреби в отриманні дозволу і не повинно тлумачитись, що будь-яка попередня інформація містить прототип цього винаходу. СТИСЛИЙ ОПИС ВИНАХОДУ Мета цього винаходу полягала у тому, щоб забезпечити рослини, які мають підвищену ефективність використання азоту. Згідно з одним аспектом цього винаходу забезпечується трансгенна рослина або частина рослини, яка включає полінуклеотид, що кодує білок, який відповідає за утилізацію азоту, функціонально пов'язаний з PBpr1 промотором. Згідно з іншим аспектом, забезпечується насіння, отримане від трансгенної рослини винаходу. Згідно з іншим аспектом винаходу, забезпечується рослинна клітина, трансформована за допомогою полінуклеотиду, що кодує білок, який відповідає за утилізацію азоту, функціонально пов'язаний з PBpr1 промотором. Згідно з іншим аспектом винаходу, забезпечується генетична конструкція, яка включає полінуклеотид, що кодує білок, який відповідає за утилізацію азоту, функціонально пов'язаний з PBpr1 промотором. Згідно з іншим аспектом забезпечується вектор, який включає генетичну конструкцію винаходу. Згідно з іншим аспектом забезпечується клітина-господар, яка включає генетичну конструкцію або вектор згідно з винаходом. Згідно з іншим аспектом винаходу, забезпечується спосіб отримання рослини, що має підвищену ефективність використання азоту, який включає трансформовану рослинну клітину з генетичною конструкцією або вектором винаходу і культивування трансформованої рослинної клітини для отримання рослини, що експресує білок, який відповідає за утилізацію азоту, з PBpr1 промотора, що призводить до отримання рослини, яка має підвищену ефективність використання азоту, де підвищена ефективність використання азоту порівнюється з ефективністю використання азоту рослинами дикого типу, вирощеними за ідентичних умов. Згідно з іншим аспектом винаходу, забезпечується спосіб отримання рослини, що має підвищену біомасу, який включає: трасформацію рослинної клітини за допомогою генетичної конструкції чи вектора винаходу, вирощування трансформованої рослинної клітини для отримання рослини, яка експресує білок, що відповідає за утилізацію азоту, за допомогою PBpr1 промотора, що призводить до отримання рослини, яка має підвищену біомасу, де підвищена біомаса порівнюється з біомасою рослини дикого типу, вирощеної за ідентичних умов. Згідно з іншим аспектом винаходу, забезпечується спосіб отримання рослини, що має підвищену врожайність насіння, який включає; трансформацію рослинної клітини за допомогою генетичної конструкції чи вектора винаходу, вирощування трансформованої рослинної клітини для отримання рослини, що експресує білок, який відповідає за утилізацію азоту, за допомогою PBpr1 промотора, що призводить до отримання рослини, яка має підвищену врожайність насіння, де підвищена врожайність насіння порівнюється з врожайністю насіння рослин дикого типу, вирощених за ідентичних умов. Згідно з іншим аспектом винаходу, забезпечується спосіб отримання рослини, що має підвищену ефективність використання азоту, підвищену біомасу, підвищену врожайність насіння, або їх комбінацію, який включає: (a) забезпечення рослини, частини рослини або насіння, який включає генетичну конструкцію або вектор винаходу, і (b) вирощування рослини, частини рослини або насінини, внаслідок чого отримують рослину, яка має підвищену здатність до поглинання азоту, підвищену біомасу, підвищену врожайність насіння або комбінацію цих ознак, де поглинання азоту, біомаса і врожайність насіння є підвищеними у порівнянні з поглинанням азоту, біомасою та врожайністю насіння у рослин дикого типу, вирощених за 3 UA 116086 C2 5 10 15 20 25 30 35 40 45 50 55 60 ідентичних умов. Згідно з іншим аспектом, забезпечується використання трансгенних рослин винаходу для отримання потомства. Згідно з іншим аспектом винаходу, забезпечується використання генетичної конструкції або вектора винаходу для підвищення ефективності використання азоту, біомаси або врожайності насіння у рослин. Згідно з іншим аспектом винаходу, забезпечується процес для підвищення ефективності використання азоту, біомаси або врожайності насіння у рослин, або комбінації цих ознак, у рослини, який включає: забезпечення насіння, що включає генетичну конструкцію або вектор винаходу, застосування гербіциду, інсектициду, добрива або їх комбінації щодо насіння, вирощування рослини з насінини, де рослина має підвищену здатність до використання азоту, підвищену біомасу, підвищену врожайність насіння або комбінацію цих ознак, і де поглинання азоту, біомаса і врожайність насіння є підвищеними у порівнянні з поглинанням азоту, біомасою та врожайністю насіння у рослин дикого типу, вирощених за ідентичних умов. У деяких втіленнях, цей винахід забезпечує спосіб створення рослини з підвищеною ефективністю використання азоту, який включає, введення нуклеїнової кислоти, що кодує один або більше білків, які відповідають за утилізацію азоту, функціонально пов'язаної зі специфічним промотором, активним у епідермісі кореня, і отримання рослини, що включає підвищені рівні одного або більше білків, що відповідають за утилізацію азоту, а саме тих, що локалізовані в епідермісі кореня. У деяких втіленнях, цей винахід забезпечує спосіб для спрямованої тканиноспецифічної експресії гена-мішені або нуклеотидної послідовності у рослин, включаючи отримання рослини з трансформованої рослинної клітини, так що тканиноспецифічна експресія гена-мішені або нуклеотидної послідовності відбувається в межах обраної тканини рослини, де трансформована клітинна рослина містить ген-мішень або нуклеотидну послідовність, яка функціонально пов'язана з елементом промотора PBpr1. В одному втіленні тканиноспецифічна експресія має місце у коренях рослини. У деяких втіленнях, цей винахід забезпечує спосіб для спрямованої тканиноспецифічної або стадієспецифічно-регульованої експресії, або експресії, що регулюється чинниками оточуючого середовища, гена-мішені або нуклеотидної послідовності у рослини, включаючи отримання рослини з трансформованої рослинної клітини, так що тканиноспецифічна, або регульована у процесі розвитку чи під впливом чинників оточуючого середовища, експресія гена чи нуклеотидної послідовності відбувається в межах обраної тканини, де трансформована рослинна клітина містить ген-мішень або нуклеотидну послідовність у оперативному зв'язку з PBpr1 промоторним елементом. В одному втіленні, тканиноспецифічна експресія має місце у коренях чи листках рослини і експресія, що регулюється умовами середовища або стадією розвитку, відбувається за умов стресу – недостатньої кількості поживних речовин. У деяких втіленнях, цей винахід забезпечує спосіб для підвищення біомаси рослини, що вирощується за умов впливу одного або більше несприятливих чинників навколишнього середовища, який включає; трансформацію рослини за допомогою гена-мішені або нуклеотидної послідовності, оперативно зв'язаних з PBpr1 елементом промотора для отримання трансформованих рослин, ген-мішень або нуклеотидну послідовність, які кодують фермент, який бере участь в асиміляції та метаболізмі азоту; і вирощування трансформованих рослин. В одному втіленні ферментом є аланінамінотрансфераза (AlaAT). У деяких втіленнях, цей винахід забезпечує спосіб для підвищення біомаси рослини, що вирощується за умов низького вмісту азоту, який включає; трансформацію рослини за допомогою гена-мішені або нуклеотидної послідовності, оперативно зв'язаних з елементом промотора PBpr1, для отримання трансформованої рослини, ген-мішень або нуклеотидну послідовність, які кодують фермент, що бере участь в асиміляції азоту; і вирощування трансформованої рослини. В одному втіленні, ферментом є AlaAT. У деяких втіленнях, цей винахід забезпечує спосіб для підвищення біомаси рослини, яка росте під дією одного або більше несприятливих чинників зовнішнього середовища, який включає; трансформацію рослини за допомогою гена або нуклеотидної послідовності, що кодують AlaAT, оперативно пов'язані з елементом промотора для отримання трансформованої рослини; і вирощування трансформованої рослини. В одному втіленні, промотором є PBpr1. У деяких втіленнях, цей винахід забезпечує спосіб для збільшення врожайності насіння, який включає; трансформацію рослини за допомогою гена-мішені або нуклеотидної послідовності або нуклеотидної послідовності, оперативно пов'язаних з елементом промотора PBpr1, для отримання трансформованої рослини, ген-мішень або нуклеотидну послідовність або нуклеотидну послідовність, які кодують фермент, що бере участь в асиміляції або 4 UA 116086 C2 5 10 15 20 25 30 35 40 45 50 55 60 метаболізмі азоту; і вирощування трансформованої рослини. В одному втіленні, ферментом є AlaAT. У деяких втіленнях, цей винахід забезпечує спосіб для збільшення врожаю насіння рослини, яка вирощується за умов низького вмісту азоту, що включає; трансформацію рослини за допомогою гена-мішені або нуклеотидної послідовності або нуклеотидну послідовність, оперативно пов'язану з елементом промотора PBpr1 для отримання трансформованої рослини, ген-мішень або нуклеотидну послідовність чи нуклеотидну послідовність, які кодують фермент, що бере участь в асиміляції азоту; і вирощування трансформованої рослини. Переважно, ферментом є AlaAT. У деяких втіленнях, цей винахід забезпечує спосіб для збільшення урожайності насіння рослини, яка вирощується за умов високого вмісту азоту, який включає; трансформацію рослини за допомогою гена-мішені або нуклеотидної послідовності, або нуклеотидної послідовності, оперативно пов'язаної з елементом промотора PBpr1, для отримання трансформованої рослини, ген-мішень або нуклеотидну послідовність або нуклеотидну послідовність, які кодують фермент, що бере участь в асиміляції азоту; і вирощування трансформованої рослини. В одному втіленні, ферментом є AlaAT. Цей короткий опис винаходу не охоплює обов'язково всі особливості винаходу і жодним чином не є обмежуючим. КОРОТКИЙ ОПИС КРЕСЛЕНЬ Ці та інші особливості винаходу стануть більш очевидними з наступних описів, в яких зроблено посилання на креслення, що додаються. На ФІГ. 1 представлена карта конструкції PBpr1: HvAlaAT в pCAMBIA1300. HvAlaAT контролюється PBpr1 промотором і визначається nos-термінатором. Селективним маркером для добору плазміди є ген стійкості до канаміцину та гігроміцину. На ФІГ. 2 представлено порівняння загальної надземної біомаси (г) і врожайності насіння (г) T1 PBpr1 ліній, NB і AGR1/7 при дозріванні. Кожна величина була отримана з 5-7 повторностей, а планки похибок – це стандартне відхилення. Чорні стовпчики: загальна надземна біомаса (г); сірі стовпчики: загальна врожайність насіння (г). На ФІГ. 3 представлено кількість пагонів та репродуктивних пагонів, що утворились на T1 PBpr1 рослинах, у порівнянні з NB і AGR1/7 при дозріванні. Кожне вимірювання було отримано як середнє значення, щонайменше, із 4-ох повторностей, а планки похибок відображають стандартне відхилення (СВ). Чорні стовпчики: загальна кількість пагонів; сірі стовпчики: кількість репродуктивних пагонів (що несуть насіння). На ФІГ. 4 представлено порівняння між T 2 PBpr1 рослинами, NB і AGR1/7 щодо надземної біомаси (г) і врожайності насіння (г) при дозріванні. Величини з кожної лінії були отримані із 7 повторностей, а планки похибок відображають СВ. Чорні стовпчики: загальна надземна біомаса (г); сірі стопвчики: загальна врожайність насіння (г). На ФІГ. 5 представлена: (A) суха надземна біомаса T 3 PBpr1 рослин (г) на 52 день після проростання; кожна лінія була представлена 5 рослинами, а планки похибок відображають стандартне відхилення СВ, і (B) сира маса (г) трьох PBpr1:AlaAT надекспресуючих ліній рису у порівнянні з диким типом (ДК) (NB; Nipponbare) при дозріванні при вирощуванні у ґрунтових умовах. На ФІГ. 6 представлена активність пагоноспецифічної AlaAT у мкмM NADH/хв.мг білка T 1 PBpr1 ліній, вирощених у ґрунті, на 40 день після проростання у порівнянні з NB і AGR1/7. Всі лінії, помічені буквою N, − це "нульові сегреганти". Кожна точка даних на графіку для трансгенних рослин була отримана в середньому у трьох повторностях; планки похибок означають стандартне відхилення (СВ) при трьох повторностях. ФІГ. 7 відображає активність аланінамінотрансферази (мкмM NADH/хв*мг білка) у ліній ДТ (NB), AGR 1/7 (OsANT1:AlaAT) і трьох незалежних PBpr1:AlaAT надекспресуючих T 3 ліній рису (PBpr1-11, PBpr1-12, PBpr1-21) у п'яти (5) різних періодів росту протягом вирощування у ґрунтових умовах при високому рівні N. Планки похибки означають стандартне відхилення (СВ). На ФІГ. 8 представлено AlaAT активність пагонів T 2 PBpr1 ліній в мкмM NADH/хв.мг білка у 28-денних та 52-денних рослин, вирощених в умовах гідропонної культури. PBpr1-21-2 і PBpr121-3 є потомками однієї й тієї ж лінії. Кожна точка даних на графіку базується на 5 повторностях, а планки похибок означають стандартне відхилення (СВ). На ФІГ. 9 представлена AlaAT активність у коренях T 2 PBpr1 ліній у 28-денних та 52-денних рослин, вирощених в умовах гідропонної культури. Кожна точка даних на графіку базується на 5 повторностях, а планки похибок означають стандартне відхилення (СВ). PBpr1-21-2 і PBpr1-21-3 є потомками однієї й тієї ж лінії. Чорні стовпчики: 28-денні рослини; сірі стовпчики: 52-денні рослини. 5 UA 116086 C2 5 10 15 20 25 30 35 40 45 50 55 60 На ФІГ. 10 представлені результати Вестерн-блот аналізу білкових екстрактів 52-денних o рослин T3 PBpr1 ліній, детекція яких була проведена з використанням HvAlaAT-2 специфічної 1 імунної сироватки. Доріжка 1: AGR1/7, доріжка 2: дикий-тип Nipponbare, доріжка 3: PBpr1-10, доріжка 4: PBpr1-11, доріжка 5: PBpr1-11-2N, доріжка 6: PBpr1-12 і доріжка 7: PBpr1-21. Концентрацію білка стандартизували поперек доріжок за допомогою методу кількісного аналізу білка Бредфорда. На ФІГ. 11 представлені результати Вестерн-блот аналізу білкового екстракту коренів рослин T2, вирощених за умов гідропонної культури (A), і пагонів (B) 28-денних рослин ліній рису o за допомогою Hv AlaAT 1 детекції антитіл, для того щоб виявити кількість HvAlaAT білка у кожному зразку. Доріжка 1: PBpr1-11, доріжка 2: PBpr1-11-2N, доріжка 3: PBpr1-21-2, доріжка 4: PBpr1-21-3, доріжка 5: PBpr1-21-3 повторність, доріжка 6: PBpr1-21-N, доріжка 7: AGR1/7 і доріжка 8: дикий тип Nipponbare. Концентрацію білка стандартизували поперек доріжок за допомогою методу кількісного аналізу білка Бредфорда. На ФІГ. 12 представлена Log2 відносного кількісного аналізу (ddCT) PBpr1 пагонів, T 2 PBpr1 пагонів і T2 PBpr1 коренів, AGR1/7 і "нульових сегрегантів" PBpr1 ліній у порівнянні з NB рослинами. 7 транскриптів (barley HvAlaAT: AlaAT; GRP: білок, багатий на гліцин, LRR: часті потори лейцину) були визначені за допомогою полімеразної ланцюгової реакції в режимі реального часу (qRT-PCR). Рослини вирощували за умов гідропонної культури і збирали через 52 дні після проростання. Використовувались ті ж зразки тканини, що й при дослідженні AlaAT активності, результати представлені на Фігурі 9. (2-кратні зміни представляє 1 ddCT. Log22=1). На ФІГ. 13 представлена нуклеотидна послідовність PBpr1 промотора, яка включає ATG старт-кодон та початкові нуклеотиди кодуючої послідовності (SEQ ID NO:6) і представляє сайти рестрикції EcoRI і SacI, які використовуються для клонування. На ФІГ. 14 представлена (A) нуклеотидна послідовність гена AlaAT ячменю (Hordeum vulgare) (SEQ ID NO:2) і (B) амінокислотна послідовність білка AlaAT (SEQ ID NO:3). На ФІГ. 15 представлено GUS забарвлювання (A) 3-денного PBpr1:GUSplus (PBpr1G лінія) сіянця, (B) 3-денних OsANT1:GUSplus (OsANT1:GUS лінії) сіянців як позитивний контроль. (C) до (M) – це GUS-забарвлені тканини сіянців віком від 7 до 10 днів PBpr1G і OsANT1:GUS ліній. (C) стебло PBpr1G, (D), (G), і (H) базальна ділянка пагона /межа розподілу пагін/корінь, що містить залишки насінини PBpr1G, (E) базальна ділянка пагона/межа розподілу пагін/корінь, що містить залишки насінини OsANT1GUS, (F) стебло OsANT1GUS, (I) межа розподілу між листковою піхвою і листковою пластинкою OsANT1GUS, (J) листкова пластинка і жилки PBpr1G, (K) листкова пластинка і жилки OsANT1GUS, (L) корені та кореневі волоски PBpr1G, і (M) корені та кореневі волоски OsANT1GUS. ФІГУРА 16 представляє таблицю, в якій наведено концентрацію амінокислоти у листках сорту дикого типу (культивар Nipponbare) рису у порівняні з barley AlaAT надекспресуючими трансгенними лініями рису; AGR 1/7 (OsANT1:HvAlaAT) і PBpr1-11, PBpr1-12, PBpr1-21 (OsPBpr1:HvAlaAT). Концентрація амінокислоти (нM/г СМ) отримана із чотирьох повторностей. На ФІГ. 17 представлений (A) PBpr2 промотор, який контролює експресію AlaAT у рослин рису, T1 покоління, вирощеного у грунті в умовах кліматичної камери; порівняння висоти рослини, загальної біомаси і ваги насіння з культиваром рису дикого типу cv. Nipponbare (NB), і (B) PBpr2 промотор, який ініціює експресію AlaAT у рослин рису, T2 покоління, вирощеного у грунті в умовах кліматичної камери; порівняння загальної біомаси і ваги насіння з культиваром рису дикого типу cv. Nipponbare (NB). На ФІГ. 18 представлена (A) нуклеотидна послідовність PBpr1 промотора (SEQ ID NO:1), і (B) нуклеотидна послідовність PBpr2 промотора (SEQ ID NO:4). На ФІГ. 19 представлено вирівнювання нуклеотидних послідовностей OsANT1 промотора (SEQ ID NO:5), PBpr1 промотора (SEQ ID NO:1) і PBpr2 промотора (SEQ ID NO:4). ДОКЛАДНИЙ ОПИС ВИНАХОДУ Даний винахід стосується однодольних рослин, які мають підвищену врожайність і/або ефективність використання азоту (NUE), способів підвищення врожайності і NUE у однодольних рослин, способів підвищення біомаси та врожайності насіння у однодольних рослин. Цей винахід пов'язаний з PBpr1 промоторами, що є промоторами з генів, які кодують метилмалонат семіальдегід дегідрогеназу (MMSDH). Беручи до уваги світові потреби в однодольних рослинах і зменшення родючості існуючих орних земель, бажано отримувати однодольні рослини, які здатні рости за субоптимальних умов живлення. Одним із засобів досягнення цієї мети є отримання однодольних рослин, які можуть більш ефективно використовувати азот. Такі однодольні рослини мають перевагу, оскільки здатні рости на ґрунтах з низьким вмістом азоту, а, отже, здатні більш ефективно використовувати наявний азот без втрати врожайності. Крім того, такі однодольні рослини 6 UA 116086 C2 5 10 15 20 25 30 35 40 45 50 55 60 можуть також демонструвати підвищену врожайність на ґрунтах, що мають нормальні рівні азоту. Ефективність використання азоту рослинами є результатом двох складових (підкомпонентів); ефективності поглинання N і ефективності використання N. Рослина, яка фенотипово виявляє ефективність використання азоту, може мати вдосконалення щодо її здатності до поглинання азоту з грунту, що є бажаною ознакою у рослин, які вирощуються на ґрунтах з низьким рівнем поживних речовин. Інакше, NUE рослини можуть мати пристосування стосовно їх здатності використовувати N, який поглинається, внаслідок чого наявний N (незалежно від того, низький його рівень чи високий) ефективно включається у субклітинні компоненти (такі як нуклеїнові кислоти, білки, запасаючі речовини, тощо), транспортуються до потрібних тканин і накопичуються на певній стадії розвитку в насінині. Або існує інша можливість, коли NUE рослина має пристосування як до покращеного поглинання N, так і до його утилізації. Будь-яка з-поміж цих можливостей забезпечила б підвищену врожайність NUE сільськогосподарських культур при вирощуванні за нормального вмісту азоту, тому що ці рослини були б здатні поглинати необмежену кількість азоту і були б здатні використовувати азот для підвищення біомаси та врожайності насіння, або внаслідок збільшення кількості насінин, або внаслідок збільшення ваги насінин чи внаслідок обох чинників. Певні втілення винаходу забезпечують трансгенні рослини, що мають покращену ефективність використання азоту у порівнянні з рослинами дикого типу. Протягом росту та розвитку рослини зазнають впливу цілої низки неоптимальних умов навколишнього середовиаща. Такі умови можуть включати водний дефіцит, надлишкове засолення, лужні або кислі ґрунти, зараження шкідниками, захворювання чи температурний стрес, будь-який з-поміж яких окремо взятий може істотно негативно відбитись на рості та/або врожайності сільськогосподарської культури. Деякі втілення даного винаходу забезпечують способи, за допомогою яких можуть бути сконструйовані рослини та їх насіння, здатні рости та добре розвиватись за змінних умов навколишнього середовища, які, зазвичай, непридатні для розвитку рослин. В деяких втіленнях забезпечуються способи отримання рослин, які можуть підтримувати або нарощувати свою біомасу і врожайність, зростаючи за умов, що не є оптимальними, з точки зору застосування добрив, як описується. У деяких втіленнях даний винахід переважно дозволяє користувачеві отримувати рослини, переваги яких для оточуючого середовища полягають у тому, що користувачі можуть підтримувати врожайність, зменшуючи при цьому потребу у підтриманні високого рівня внесення добрив. У деяких втіленнях, цей винахід дозволяє користувачу отримувати рослини, які за умов високих рівнів поживних речовин мають здатність до покращеного поглинання поживних елементів, що дає змогу рослинам екстрагувати більше поживних речовин з їх середовища протягом проміжку часу, коли поживні речовини є у достатній кількості. Використовуючи способи та композиції певних втілень винаходу, рослини можуть бути вдосконалені для вирощування за умов навколишнього середовища, що є зазвичай непридатними для вирощування рослин. В деяких втіленнях, способи та композиції винаходу дають змогу генетично конструювати рослину, для того щоб змінити одну або більше характерних особливостей рослини лише в обраних тканинах рослини. Якщо не визначено щось інше, всі технічні та наукові терміни, які тут використовуються, мають те ж саме значення, що зазвичай зрозуміле будь-якому фахівцеві зі звичайним досвідом роботи в галузі, до якої цей винахід належить. Як тут використовується, терміни "який включає", "що має", "включаючи" і "що містить" та граматичні їх варіанти, є інклузивні або неостаточні і не виключають додаткових, неописаних елементів і/або етапів способу. Термін "складається істотно з" при використанні тут у зв'язку з композицією, застосуванням або способом, свідчить, що можуть бути присутні додаткові елементи і/або етапи способу, однак ці доповнення по суті не впливають на характер функціонування викладеної композиції, способу або застосування. Термін "складається з" при використанні тут у зв'язку з композицією, застосуванням або способом, виключає присутність додаткових елементів і/або етапів способу. Композиція, застосування або спосіб, тут описані, які включають деякі елементи і/або етапи можуть також, у деяких втіленнях складатись по суті з тих же елементів і/або етапів, і в інших втіленнях складатись з тих елементів і/або етапів, яких ці втілення конкретно стосуються чи ні. До того ж, застосування слів у однині, включає й множину, і "або" означає "і/або", якщо не стверджується інше. Як тут використовується, термін "близько" стосується приблизно +/-10 % варіювання від заданої величини. Зрозуміло, що таке варіювання завжди включається в будь-яке наведене значення величини, незалежно від наявності чи відсутності спеціального посилання на це. Термін "значною мірою ідентичний", як тут використовується стосовно нуклеотидної або 7 UA 116086 C2 5 10 15 20 25 30 35 40 45 50 55 60 амінокислотної послідовності, свідчить, що при оптимальному вирівнюванні, наприклад з використанням методів, наведених нижче, нуклеотидна або амінокислотна послідовності поділяють, щонайменше 70 %, щонайменше 75 %, щонайменше 80 %, щонайменше 85 %, щонайменше 90 %, щонайменше 95 %, щонайменше 96 %, щонайменше 97 %, щонайменше 98 % або щонайменше 99 % ідентичності послідовності порівняно з іншою визначеною нуклеотидною або амінокислотною послідовністю (або "референс-послідовністю"). "Істотна ідентичність" може використовуватись при посиланні на різні типи і довжину послідовностей, таких як повна послідовність, функціональний домен, кодуючі і/або регуляторні послідовності, промотори і геномні послідовності. Відсоток ідентичності між двома амінокислотними або нуклеотидними послідовностями може бути визначений у різний спосіб, що знаходяться в межах компетенції фахівців у цій галузі, наприклад, з використанням загальнодоступного комп'ютерного програмного забезпечення, такого як Smith Waterman Alignment (Smith, T. F. and M. S. Waterman (1981) J Mol Biol 147:195-7); "BestFit" (Smith and Waterman, Advances in Applied Mathematics, 482-489 10 (1981)), включеного в GeneMatcher Plus™, Schwarz and Dayhof (1979) Atlas of Protein Sequence and Structure, Dayhof, M. O., Ed pp 353-358; BLAST програми (Basic Local Alignment Search Tool (Altschul, S. F., W. Gish, et al. (1990) J Mol Biol 215: 403-10), і її видозмін, включаючи BLAST-2, BLAST-P, BLAST-N, BLAST-X, WU-BLAST-2, ALIGN, ALIGN-2, CLUSTAL, і програмне забезпечення Megalign (DNASTAR). Крім того, ці досвідчені фахівці можуть застосовувати прийнятні параметри для вимірювання вирівнювання, включаючи алгоритми, необхідні для досягнення максимального вирівнювання уздовж всієї довжини послідовностей, що порівнюються. Загалом, для амінокислотних послідовностей, довжина послідовностей, що порівнюються, повинна становити, щонайменше, 10 амінокислот. Будь-який фахівець з досвідом роботи в галузі має розуміти, що дійсна довжина буде залежати від загальної довжини послідовностей, що порівнюються, і може становити, щонайменше 20, щонайменше 30, щонайменше 40, щонайменше 50, щонайменше 60, щонайменше 70, щонайменше 80, щонайменше 90, щонайменше 100, щонайменше 110, щонайменше 120, щонайменше 130, щонайменше 140, щонайменше 150, або щонайменше 200 амінокислот, або це може бути повна (непроцесована) амінокислотна послідовність. Для нуклеотидних послідовностей, довжина послідовностей, що беруться для порівняння, як правило, становлять щонайменше 25 нуклеотидів, однак можуть становити, щонайменше 50, щонайменше 100, щонайменше 125, щонайменше 150, щонайменше 200, щонайменше 250, щонайменше 300, щонайменше 350, щонайменше 400, щонайменше 450, або щонайменше 500, або це може бути повна (непроцесована) нуклеотидна послідовність. Термін "тканиноспецифічна експресія" нуклеотидної послідовності-мішені відомий з рівня техніки і включає експресію нуклеотидних послідовностей-мішеней переважно в обраних тканинах. Тканиноспецифічна експресія може призводити до експресії нуклеотидної послідовності-мішені лише в обраній тканині, у випадку якої нуклеотидна послідовність-мішень може бути присутня у багатьох тканинах, але експресуватись лише у підсукупності (підмножині) цих тканин. Альтернативно, тканиноспецифічна експресія може призводити до вищих рівнів експресії нуклеотидної послідовності-мішені в обраних тканинах, якщо експресія може також спостерігатись в інших тканинах, то на нижчому рівні, ніж у тканині(ах)-мішені(ях), наприклад, на 50 % нижче, на 40 % нижче, на 30 % нижче, на 25 % нижче або на 20 % нижче. Така селективна експресія можлива завдяки впливу одного або більше регуляторних генетичних елементів, наприклад, але не обмежуючись ними, елементів промоторів, елементів репресора, елементів енхансора або інших регуляторних факторів, які можуть взаємодіяти з ДНК та РНК. Терміни "ген-мішень" або "нуклеотидна послідовність-мішень" є загальновизнаними в галузі та включають нуклеотидні послідовності, які бажано експресувати в одній або більше обраних рослинних тканинах. Необмежуючі приклади генів-мішеней або нуклеотидних послідовностей, які можуть бути використані у поєднанні зі способами винаходу, включають гени або нуклеотидні послідовності, що беруть участь в асиміляції і/або утилізації азоту, гени або нуклеотидні послідовності, які беруть участь у стійкості до стресу, гени або нуклеотидні послідовності, які беруть участь у стійкості до захворювань та шкідників, і гени або нуклеотидні послідовності, які беруть участь у поглинанні та утилізації поживних речовин. Такі гени або нуклеотидні послідовності добре відомі кожному фахівцеві в цій галузі. Терміни "оперативний зв'язок", "оперативно зв'язаний" і "функціонально зв'язаний" використовуються тут поперемінно, при посиланні на розташування нуклеотидної послідовностімішені відносно регуляторної послідовності нуклеїнової кислоти, внаслідок чого експресія нуклеотидної послідовності-мішені контролюється регуляторною послідовністю. Ця регуляторна послідовність може виявляти позитивний вплив (підвищувати) на експресію гена-мішені або нуклеотидної послідовності (напр., регуляторна послідовність – це промотор або енхансерний 8 UA 116086 C2 5 10 15 20 25 30 35 40 45 50 55 60 елемент), або регуляторна послідовність може зменшувати експресію гена-мішені або нуклеотидної послідовності (напр., регуляторна послідовність – це елемент репресора). Регуляторна послідовність може фізично бути розташована на 5" або 3" кінцях гена-мішені або нуклеотидної послідовності, може бути всередині кодуючої послідовності гена-мішені або нуклеотидної послідовності, або може міститись на інтроні в межах гена-мішені або нуклеотидної послідовності. Термін "нульові лінії" є загальновизнаним у галузі і включає рослини, які були вирощені в культурі тканин, але не несуть трансгена або селективного маркера. Тут описується отримання рослин, які експресують один або більше генів-мішеней або нуклеотидних послідовностей у тканиноспецифічний спосіб. У деяких втіленнях, винахід забезпечує насіння, яке містить один або більше генів-мішеней або нуклеотидних послідовностей під контролем промоторного елемента, який специфічно ініціює тканиноспецифічну експресію гена або нуклеотидної послідовності. У деяких втіленнях, способи винаходу дозволяють отримання рослин, які мають одну або більше бажаних ознак або властивостей у обраних тканинах; напр., здатність специфічно змінювати генетичні і/або фізіологічні властивості плодів або коренів рослин. Деякі втілення винаходу, крім того, забезпечують способи отримання рослин, які виявляють коренеспецифічну або листкоспецифічну експресію однієї або більше бажаних нуклеотидних послідовностей, з використанням елемента промотора PBpr1. Способи винаходу для отримання рослин, які мають тканиноспецифічну експресію одного або більше генів-мішеней або нуклеотидних послідовностей, втілюються через застосування генетичного регуляторного елемента, який ініціює тканиноспецифічну експресію гена(ів)мішені(ей). Активність регуляторних елементів може бути негативною або позитивною: тканиноспецифічний промотор рослини або енхансерний елемент дозволяють експресію генамішені або нуклеотидної послідовності(тей) в одній або більше специфічних тканин, тоді як тканиноспецифічний репресор рослини пригнічує експресію генів-мішеней або нуклеотидних послідовностей в одній або більше специфічних тканин; натомість експресія в іншій тканині (ах) не припиняється. З метою впровадження винаходу, слід розуміти, що послідовності промотора утворюють кращі генетичні регуляторні елементи винаходу. Тканиноспецифічні промотори, що використовуються у цьому винаході, можуть бути гомологічні або гетерологічні стосовно рослини, до якої вони використовуються. Промотори, які ініціюють експресію оперативно зв'язаного гена або нуклеотидної послідовності в одній або більше рослинній тканині, однак виключають можливість експресії зчепленого гена або нуклеотидної послідовності в одній або більше рослинній тканині, можуть бути використані у способах і конструкціях винаходу. Прийнятні рослинні клітини включають, але не обмежені ними, напр., корінь, листок, пелюстку, чашолисток, тичинку, пиляк, приймочку, зав'язь, стовпчик, епідерміс, флоему, ксилему, кору, серцевину, камбій, стебло або стовбур, кореневі волоски, черешок, плід і бульбу. Переважно, промотор − PBpr1 промотор. Згідно з деякими втіленнями винаходу, промотор є PBpr1 промотором, що має нуклеотидну послідовність, яка є значною мірою ідентичною до послідовності, що представлена у SEQ ID NO:1 (Фігура 13). Будь-якому фахівцеві з досвідом роботи в галузі має бути зрозуміло, що можуть бути зроблені модифікації промоторів, які використовуються у способах та конструкціях винаходу, для того щоб оптимізувати або змінити активність промотора. Численні копії відібраного промотора можуть бути оперативно зв'язані з єдиним геном-мішенню або нуклеотидною послідовністю, що призводить до зміни рівня експресії зчепленого гена або нуклеотидної послідовності, або відібраний промотор може бути оперативно зв'язаний з одним або більше генів-мішеней або нуклеотидних послідовностей, в результаті чого експресія кожного генамішені або нуклеотидної послідовності узгоджено регулюється. Промотор може мати будь-який розмір, прийнятний для забезпечення тканиноспецифічного функціонування промотора. Промотор може бути модифікований (напр., шляхом мутагенезу, делеції, інсерції або внаслідок усічення), для того щоб змінити ступінь, до якого оперативно зв'язаний ген або нуклеотидна послідовність експресуються в обраній тканині, або для того щоб змінити специфічність експресії в тканині, ініційованої промотором. Далі, розміщення промотора по відношенню до оперативно зв'язаного гена-мішені або нуклеотидної послідовності, може бути змінене (напр., переміщене далі, або розташоване ближче), для того щоб досягти бажаного рівня промоторспрямованої експресії. Промотор може контролювати експресію оперативно зчепленого гена або нуклеотидної послідовності в одній або більше відібраних тканин рослини, а також ініціювати експресію гена або нуклеотидної послідовності у відповідь на специфічні фізіологічні умови або умови 9 UA 116086 C2 5 10 15 20 25 30 35 40 45 50 55 60 навколишнього середовища. Наприклад, промотори можуть бути активовані за умов стресу (напр., водного стресу, сольового стресу, температурного стресу, окиснювального стресу, pHстресу, або стресу, викликаного важкими металами), за умов дефіциту поживних речовин, за умов впливу захворювань та шкідників. В іншому прикладі, промотори можуть бути активовані під впливом специфічних умов розвитку (напр., під час пагоноутворення, плодоношення або утворення насіння) або у відповідь на зміну умов оточуючого середовища (напр., від умов дефіциту поживних речовин до їх надходження або від гіпоксії – до нормоксії). У деяких втіленнях, цей винахід забезпечує конструкції, що включають ген-мішень під контролем PBpr1 промотора, який при трансформації в прийнятній клітині-господарі, призводить до тканиноспецифічної експресії генного продукту. Наприклад, експресія може переважно відбуватись у коренях і/або в пагонах рослини. У деяких втіленнях, цей винахід забезпечує конструкції, які включають ген-мішень під контролем PBpr1 промотора, який при трансформації в прийнятній клітині-господарі, призводить до регульованої в процесі розвитку експресії генного продукту. Ген-мішень або нуклеотидна послідовність винаходу є ген або нуклеотидна послідовність, яку бажано експресувати в рослині. Загальні класи генів-мішеней або нуклеотидних послідовностей, які можуть переважно бути використані у способах та конструкціях винаходу, включають гени або нуклеотидні послідовності, що кодують структурні білки рослин, гени або нуклеотидні послідовності, що кодують білки, які беруть участь у транспорті і/або поглинанні поживних речовин, гени або нуклеотидні послідовності, що кодують ферменти і білки, які беруть участь в утилізації поживних речовин, гени або нуклеотидні послідовності, що кодують білки, які беруть участь у стійкості рослин до гербіцидів, гени або нуклеотидні послідовності, що кодують білки, які беруть участь у стійкості рослин до нематод, вірусів, комах або мікроорганізмів; гени або нуклеотидні послідовності, що кодують білки, які беруть участь у стійкості рослин до стресів (напр., осмотичного, температурного, pH-стресу або окиснювального стресу), гени або нуклеотидні послідовності, що кодують білки, які беруть участь у стимуляції або відновленні росту рослини, або гени чи нуклеотидні послідовності, що кодують білки, які беруть участь у фіторемедіації. Крім того, ген-мішень або нуклеотидна послідовність можуть бути нуклеотидною послідовністю, яка при транскрибуванні є антисмисловою до природної послідовності, транскрипцію та трансляцію якої бажано пригнічувати. У деяких втіленнях винаходу, гени-мішені або нуклеотидні послідовності є такими, що кодують ферменти, які беруть участь в асиміляції і/або метаболізмі азоту. Гени або нуклеотидні послідовності, що становлять інтерес, можуть включати гени або нуклеотидні послідовності, які кодують більки, які беруть участь в асиміляції амонію в амінокислотах або використанні утворених амінокислот у реакціях біосинтезу, що являє собою "білок, який відповідає за утилізацію азоту". Приклади білків, які відповідають за утилізацію азоту, включають, але не обмежені ними, нітрат амонію і транспортери амінокислот, глутамінсинтазу (GS), аспарагінсинтазу (AS), глутаматсинтазу (відому також як глутамат 2:оксоглутурат амінотрансфераза і GOGAT), аспарагіназу (ANS), глутаматдегідрогеназу (GDH), аспартатамінотрансферазу (AspAT) і аланінамінотрансферазу (AlaAT) і ті гени або нуклеотидні послідовності, які можуть бути залучені до визначення ефективності використання азоту (NUE), як описано Beatty et al. (2009), та у Патенті США № 7,589,257. Послідовності прийнятних генівмішеней і нуклеотидні послідовності є легко досяжними з публічних джерел, наприклад з генного банку GenBank, або можуть бути визначені за допомогою стандартних методик, добре відомих фахівцям у цій галузі. Ген-мішень або нуклеотидна послідовність можуть бути геном або нуклеотидною послідовністю, які природно експресуються в обраній рослині, або вони можуть бути гетерологічними щодо відібраної рослини. Ген або нуклеотидна послідовність можуть походити з цілої низки джерел, включаючи, вірусні, бактеріальні, рослинні або тваринні джерела. Деякі втілення винаходу забезпечують генетичні конструкції, які включають ген-мішень або нуклеотидну послідовність, оперативно пов'язані з PBrp1 промотором. У деяких втіленнях, ген або нуклеотидна послідовність є гетерологічними до PBrp1 промотора, з яким вони зчеплені. Ген-мішень або нуклеотидна послідовність може бути модифікована у випадку необхідності. Наприклад, ген або нуклеотидна послідовність модифікуються, для того щоб бути прийнятними для транскрипції та трансляції в рослинній системі, наприклад, за допомогою оптимізації кодону. У деяких втіленнях, ген або нуклеотидна послідовність можуть бути модифіковані таким чином, що вони містять всі необхідні послідовності поліаденилювання, сайти ініціації та термінації, які дають змогу кодуючій послідовності бути транскрибованою в інформаційній (матричній) рибонуклеїновій кислоті (мРНК) і мРНК – бути трансльованою у функціональний білок в обраній рослинній системі. 10 UA 116086 C2 5 10 15 20 25 30 35 40 45 50 55 60 Таким чином, деякі втілення винаходу забезпечують генетичні конструкції, які включають ген-мішень або нуклеотидну послідовність, функціонально пов'язані з PBrp1 промотором і також включають одну або більше послідовностей поліаденилювання, сайти ініціації і/або термінації транскрипції. Такі модифікації гена-мішені або нуклеотидної послідовності і способи, за допомогою яких вони можуть бути створені, добре відомі з рівня техніки. У деяких втіленнях, ген-мішень або нуклеотидна послідовність кодує білок, який відповідає за утилізацію азоту, наприклад, транспортер азоту, транспортер амонію, траспортер амінокислоти, глутамінсинтазу (GS), аспарагінсинтетазу (AS), глутаматсинтазу (відому також як глутамат 2:оксоглутурат амінотрансферазу і GOGAT), аспарагіназу (ANS), глутаматдегідрогеназу (GDH), аспартатамінотрансферазу (AspAT), або аланінамінотрансферазу (AlaAT). Інші приклади включають ті гени або нуклеотидні послідовності, які можуть брати участь у ефективності використання поживних речовин (NUE), як описано Beatty et al. (2009), і в Патенті США № 7,589,257. У деяких втіленнях, цей винахід забезпечує вектори, що включають генетичні конструкції. Будь-який фахівець у цій галузі розуміє, що застосування чітко визначеного вектора не є критично важливим для даного винаходу і прийнятні вектори легко можуть бути відібрані досвідченою особою. Приклади експресуючих векторів та клонуючих носіїв включають, але не обмежені ними, вірусні частинки, бакуловіруси, фаги, плазміди, фагміди, косміди, фозміди, штучні бактеріальні хромосоми, ретровірусні вектори, вірусну ДНК (наприклад, вірусу вісповакцини, аденовірусу, вірусу курячої віспи, PRV-вірусу і похідних SV40), штучні хромосоми на основі P1, дріжджові плазміди, штучні дріжджові хромосоми та інші відомі вектори, специфічні для специфічних клітин-господарів, що становлять інтерес. Деякі втілення забезпечують клітини-господарі, які включають генетичні конструкції або вектори згідно з винаходом. Генетична конструкція або вектор може бути інтродукований в прийнятну клітину-господаря за допомогою цілої низки стандартних методів. Такі методи добре відомі з рівня техніки і включають, наприклад, стабільну або транзієнтну трансфекцію, ліпофекцію, електропорацію і введення рекомбінатних вірусних векторів. Кожен фахівець з досвідом роботи у цій галузі має зрозуміти, що добір прийнятної клітини-господаря буде залежати від обраного вектора. Приклади клітин-господарів включають, але не обмежені ними, бактеріальні, дріжджові, рослинні клітини, клітини комах та ссавців. У деяких втіленнях, клітинагосподар – це рослинна клітина. Способи і генетичні конструкції, тут розкриті, можуть бути використані для створення рослини або частини рослини будь-якого виду, здатного використовувати промотор, внаслідок чого трансгенна (неприродна) рослина, має тканиноспецифічну експресію одного або більше бажаних генів або нуклеотидних послідовностей. Винахід призначений для конкретного застосування щодо, наприклад, рослин сільськогосподарських культур (особливо тих, що належать до роду Oryza), декоративних рослин, і дерев (зокрема, шпилькових та представників роду Populus). Особливо прийнятні рослини для практики в цьому винаході включають, але не обмежені ними, канолу, ячмінь, цукрову тростину, кукурудзу, тютюн, сою, бавовник, люцерну, томати, пшеницю, картоплю, осику, тополю та дерева шпилькових порід, або частини будь-яких з цих рослин, наприклад, корені, кореневі кінчики, листки, стебла, квітки, апікальні бруньки, меристематичні тканини тощо. У деяких втіленнях, рослин для застосування у винаході включають канолу, ячмінь, маїс, рис, тютюн, сою, бавовник, люцерну, томати, пшеницю, картоплю, і деякі роди дерев, включаючи шпилькові та види Populus. У деяких втіленнях, способи, використання та конструкції винаходу застосовуються щодо рослин роду Oryza та інших родів, що є близько спорідненими з Oryza, такими як Porteresia і Leersia. Приклади видів з цих родів включають, але не обмежені ними, Oryza sativa, Oryza punctata, Oryza officinalis, Oryza malapuzhanensis, Oryza latifolia, Oryza australiensis, Oryza brachyantha, Oryza granulate, Oryza longiglumis, Oryza schlechteri, Porteresia coarctata, Leersia perrieri, Leersia hexandra та Leeria tisserantti. Трансгенна (неприродна) рослина, частини рослини і насіння, отримані згідно з даним винаходом, можуть бути далі використані у селекційних програмах для отримання видів рослин, які мають більше, ніж одну бажану властивість. Наприклад, дві трансгенні рослини винаходу, кожна з-поміж яких має експресія бажаного трансгена в різних тканинах рослини, можуть бути схрещеними для отримання в потомстві трансгенних рослин, які мають тканиноспецифічну експресію обох трансгенів; або дві трансгенні рослини винаходу, кожна з яких має експресію різних бажаних трансгенів у тій же рослинній тканині, можуть бути схрещеними для того, щоб отримати у потомстві трансгенні рослини, які мають тканиноспецифічну експресію обох трансгенів. У такий спосіб можливо отримати трансгенні рослини, що мають комбінацію бажаних властивостей у обраній тканині (ах) рослини. 11 UA 116086 C2 5 10 15 20 25 30 35 40 45 50 55 60 До того ж, будь-якому фахівцеві з досвідом роботи у галузі має бути зрозуміло, що різні види рослин можуть бути більшою чи меншою мірою прийнятними для генетичних маніпуляцій загалом, і що, відповідно, передусім бажано спочатку трансформувати споріднені види бажаних рослин за допомогою способів і конструкцій винаходу, а потім вже індукувати тканиноспецифічну експресію гена-мішені або нуклеотидної послідовності у бажаних видів рослин шляхом селекційних методик. Такі методики і прийнятні споріднені види рослин добре відомі кожному досвідченому фахівцеві у цій галузі. Рослинні клітини або протопласти, які були трансформовані за допомогою генної конструкції даного винаходу, можуть бути регенеровані в диференційованих рослинах за допомогою стандартних живильних середовищ, доповнених гормонами, що індукують розвиток пагонів та коренів, використовуючи методи, загальновідомі фахівцям з досвідом роботи в галузі (див., наприклад, Патент США № 4,634,674 і представлені тут посилання). Насіння може додатково бути зібране з таких трансгенних рослин за допомогою загальновідомих методів та використане у подальшому для репродукції трансгенних рослин та гібридів винаходу. Застосування Способи і конструкції винаходу роблять можливим отримання рослин і насіння, які мають експресію одного або більше бажаних генів або нуклеотидних послідовностей, наприклад, в одній або більше відібраних тканин рослини. Таким чином, у деяких втіленнях, способи і конструкції винаходу дозволять отримати рослини, які мають одну або більше бажаних ознак, забезпечуючи у такий спосіб спрямування ознаки до тканини, яка є для неї найбільш прийнятною, або внаслідок уникнення експресії бажаного гена або нуклеотидної послідовності, якщо їх вплив є небажаним. Існує широкий спектр специфічних втілень винаходу, включаючи, але не обмежуючись цим, отримання рослин, які мають підвищену врожайність, стійкість до стресу, які мають покращену здатність до поглинання поживних речовин і/або її утилізації, які мають покращений вміст поживних речовин і/або врожай (вихід) бажаних складових, і які мають фіторемедіаційні властивості. Специфічні втілення винаходу наводяться нижче. Одне застосування винаходу полягає у отримання рослин, краще пристосованих до розвитку на бідних ґрунтах. Добре відомо з рівня техніки, що деякі види рослин, особливо рослини сільськогосподарських культур, виснажують ґрунт, споживаючи поживні речовини, необхідні для забезпечення постійного росту, такі як азот, фосфор і калій. Для того щоб відновити втрату поживних речовин, необхідно або вносити добриво в ґрунт (витратна практика, яка завдає шкоди навколишньому середовищу) або культивувати рослини, відомі своєю здатністю відкладати в ґрунт спожиті поживні речовини (напр., конюшина або соя у випадку виснаження вмісту азоту), якими можуть бути сільськогосподарські культури, що є менш прибутковими або менш поживними і, відповідно, їх вирощування є менш бажаним. Часте внесення добрива у ґрунт потребує значних витрат з точки зору вартості сільськогосподарської продукції, таких як оплата праці та пального; з цієї причини рослини сільськогосподарських культур знаходяться, як правило, або в стані надлишку поживних речовин, або їх дефіциту. Інше втілення винаходу полягає в отриманні рослин, краще пристосованих до захоплення та утилізації азоту, якщо він присутній у адекватній чи достатній кількості у ґрунті, до того, як відбувається втрата азоту через вилужування (вимивання), випаровування або мікробну деградацію. У деяких втіленнях, способи винаходу дають змогу здійснювати цільову експресію генів або нуклеотидних послідовностей, що беруть участь у поглинанні поживних речовин (напр., транспорту молекул) до тих тканин, у яких це поглинання відбувається (напр., коренів або кореневих волосків), покращуючи таким чином здатність рослини абсорбувати поживну речовину з оточуючого середовища. В деяких втіленнях винахід може бути використаний для отримання рослин, які експресують гетерологічні нуклеотидні послідовності, пов'язані зутилізацією поживних речовин, або оптимізовані (наприклад, оптимізовані для рослинної експресії) природні нуклеотидні послідовності, що відповідають за утилізацію поживних речовин, які сприяють ефективнішому використанню поживної речовини, в результаті чого зменшується потреба у поживній речовині для нормального росту і функціонування рослини. У деяких втіленнях, експресія таких послідовностей спрямовується до певних тканин (напр., коренів або листків). У деяких втіленнях, способи винаходу роблять можливою експресію генів або нуклеотидних послідовностей, що беруть участь у використанні та поглинанні поживних речовин, які зазвичай не використовуються рослиною у цих рослинних танинах, які безпосередньо відкриті для різних поживних речовин (напр., кореня і листка). У такий спосіб можуть бути отримані рослини, які здатні рости і добре розвиватись на різних джерелах поживних речовин (напр., різних джерелах азоту). Особливо цінні гени-мішені або нуклеотидні послідовності для оптимізації ефективності використання азоту рослинами включають: 12 UA 116086 C2 5 10 15 20 25 30 35 40 45 50 55 60 транспортери нітрату, транспортери амонію і траспортери амінокислот, глутамінсинтазу (GS), аспарагінсинтетазу (AS), глутаматсинтазу (відому також як глутамат 2:оксоглутуратамінотрансфераза і GOGAT), аспарагіназу (ANS), глутаматдегідрогеназу (GDH), аланіндегідрогеназу, аспартатамінотрансферазу(AspAT) і аланінамінотрансферазу (AlaAT), а також ті гени або нуклеотидні послідовності, які можуть брати участь у визначенні ефективності використання поживних речовин, описані Beatty et al. (2009) і в Патенті США № 7,589,257. У деяких втіленнях рослини, отримані за допомогою способів цього винаходу, можуть більш ефективно використовувати внесені добрива за рахунок швидкого засвоєння азоту з добрива і накопичення його протягом часу застосування, зменшуючи тим самим кількість азотних добрив, які втрачаються в результаті вимивання тощо. Це може дати змогу зменшити кількість азотних добрив, потрібних для внесення під сільськогосподарські культури, для того щоб отримати врожаї, які можна порівняти з врожаями, які можна отримати при застосуванні звичайних методів культивування, і рослин, що не були модифіковані відповідно до даного винаходу. Додаткові агрономічні переваги можуть включати швидший ріст та збір врожаю, де внесення азотних добрив підтримується на рівнях, що використовуються при загальноприйнятих способах культивування сільськогосподарських культур. Створення зернових сільськогосподарських культур Як описується нижче, трансформовані рослини Oryza sativa, що ектопічно експресують нуклеотидну послідовність, яка кодує ферменти, що беруть участь в асиміляції або метаболізмі азоту, наприклад, але не обмежені ними, аланіндегідрогеназу, глутамінсинтазу, аспарагінсинтетазу, глутаматсинтазу, аспарагін азу, глутаматдегідрогеназу, аспартатамінотрансферазу, аланінамінотрансферазу, і ті нуклеотидні послідовності, які беруть участь в ефективності використання поживних речовин, описані Beatty et al. (2009), і в Патенті США № 7,589,257, були створені за допомогою PBpr1 промотора і вирощені в лабораторних умовах, для того щоб визначити, чи може бути отриманий позитивний вплив на ріст рослин та врожайність у контрольованих умовах. Наприклад, було досліджено, чи впливає кількість доступного азоту на біомасу рослини і врожайність насіння у рослин, що експресують PBpr1/AlaAT, вирощених в лабораторних умовах, у порівнянні з подібними рослинами, які ектопічно не експресують нуклеотидну послідовність-мішень. Було досліджено врожайність насіння у контрольних і трансгенних рослин, вирощених в лабораторних умовах. Трансгенні рослини, які експресують ген або нуклеотидну послідовність, які кодують фермент, що бере участь в асиміляції або метаболізмі азоту, виявили вищу біомасу і врожайність насіння, ніж контрольні рослини, за умови наявності адекватної кількості азоту. Узагальнені результати свідчать про те, що трансгенні рослини, які ектопічно експресують фермент, що бере участь в асиміляції або метаболізмі азоту, здатні оптимізувати використання наявного азоту при різноманітних умовах навколишнього середовища, що призводить до збільшення біомаси рослини, підвищення врожайності або до обох цих показників. Таким чином, в одному аспекті, цей винахід забезпечує спосіб для збільшення врожайності насіння, який включає; трансформацію рослини за допомогою гена-мішені або нуклеотидної послідовності в оперативному зв'язку з елементом промотора PBpr1, для отримання трансформованої рослини; ген-мішень або нуклеотидну послідовність, які кодують фермент, що бере участь в асиміляції або метаболізмі азоту; і вирощування трансформованої рослини. Наведений вище опис не має на меті обмежити формулу винаходу жодним чином, тим більше, обговорена сукупність особливостей може не бути абсолютно необхідною для винахідницького рішення. Для того щоб досягти кращого розуміння винаходу, який тут описано, наведено наступні приклади. Слід зрозуміти, що ці приклади призначені для того, щоб в ілюстративному вигляді описати втілення винаходу, і не мають на меті жодним чином обмежити сферу застосування винаходу. ПРИКЛАДИ ПРИКЛАД 1: Промотор PBpr1, злитий з геном alaAT Конструювання бінарних векторів і трансформація, опосередкована Agrobacterium Був обраний PBpr1 промотор і сконструйований для клонування за допомогою GeneArt (Invitrogen, Life Technologies, Карлсбад, Каліфорнія, США). HvAlaAT кДНК (SEQ ID NO: 2) була введена в pCAMBIA1300 за допомогою PstI/HindIII зшивки, тоді як PBpr1 промотор був вставлений вище HvAlaAT для ініціювання експресії з використанням EcoRI/SacI сайтів (Фігури 1,13,14). Промотор PBpr1 був введений в pCAMBIA1305.1, щоб ініціювати GUSplus для аналізу паттерна промотора з використанням EcoRI/NcoI сайтів. Конструкції були трансформовані окремо у штамі Agrobacterium tumefaciens EHA105 за допомогою методу заморожуваннярозморожування (Weigel and Glazebrook, 2002). Калус рису (Oryza sativa c.v. Nipponbare (NB)) 13 UA 116086 C2 5 10 15 20 25 30 35 40 45 50 55 60 був трансформований з використанням обох конструкцій за допомогою системи трансформації на основі Agrobacterium, розробленої в нашій лабораторії (Shrawat and Good, 2011). Лінії, трансформовані за допомогою PBpr1:HvAlaAT, названі тут як PBpr1 лінії, натомість лінії, трансформовані за допомогою PBpr1:GUSplus, називаються тут як PBpr1G лінії. Лінії, трансформовані за допомогою PBpr1:HvAlaAT, були раніше названі Ap лінії. Відповідність між попередньою номенклатурою та номенклатурою, яка тут використовується, представлена в Таблиці 1. Аналогічно, лінії трансформовані з використанням PBpr1:GUSplus, раніше мали назву ApG ліній. Виходячи з урожайності насіння, загальної надземної біомаси і кількості пагонів при достиганні, були відібрані дві T 0 лінії, які містять PBpr1:HvAlaAT, для отримання наступних генерацій. Тканина листка була відібрана у двотижневих рослин для визначення, чи є вони трансгенними, шляхом аналізу геномної ДНК за допомогою полімеразно-ланцюгової реакції з використанням праймерів, специфічних до гена резистентності до гігроміцину. Були відібрані, виходячи з однакового розміру та висоти, трансгенні рослини і "нульові сегреганти" для отримання наступних поколінь. Первинні трансгенні лінії були позначені, як T 0 лінії. Ґрунтові експерименти у кліматичній камері T0, T1, і T2 генерації рослин рису вирощували у безґрунтовій суміші Sunshine Mix #4 (Sun Gro Horticulture) у кліматичній камері при 28 °C, відносній вологості 70 %, тривалості фотоперіоду 14г/10г світло/темрява, щільність світлового потоку 750 мкE на рівні стелажа, як описано у Shrawat et al., 2008. У T3 покоління, три насінини були пророщені у 7-дюймовому горщику і вирощувались протягом двох тижнів. Два з-поміж трьох сіянців були відібрані, виходячи з однорідної висоти і розміру рослин, що залишались у кожному горщику. Режим підживлення був аналогічним тому, який було застосовано щодо рослин T 2 покоління у 3-тижневому віці. Всі статистичні порівняння були виконані для цих результатів експериментів у кліматичній камері з використанням Student t-тесту. Гідропонний експеримент у кліматичній камері Гідропонний експеримент було проведено в кліматичній камері при 28 °C, вологість 70 %, фотоперіод 14г/10г світло/темрява, щільність світлового потоку 750 мкмE на висоті полиці, як описано у Shrawat et al., 2008. Рослинний матеріал відбирали з 28-денних і 52-денних рослин для кількісного аналізу AlaAT і РНК експериментів. Селекція гомозиготних ліній і підтримання нульових ліній T2 насіння, отримане від T1 рослин, пророщували на середовищі MS з гігроміцином; визначали співвідношення стійкості до антибіотика, для того щоб з'ясувати співвідношення гомозиготність/гетерозиготність, як описано у Shrawat et al., 2008. "Нульові лінії" ідентифікували за допомогою ПЛР на стадії рослин T 1 з використанням праймерів, специфічних до гігроміцину. Відповідно, були відібрані та підтримувались "нульові лінії" для порівняння між NB, AGR1/7 і PBpr1 лініями. Екстракція AlaAT та імуноферментний аналіз Тканини як пагонів, так і коренів були зібрані для імуноферментного аналізу AlaAT в умовах гідропонного експерименту, тоді як у рослин, що вирощувались у горщиках, була відібрана лише тканина пагона, як це рекомендовано Muench and Good, 1994. Електрофорез білка в поліакриламідному гелі (SDS-PAGE) та Вестерн-блот аналіз Для того, щоб підтвердити результати імуноферментного аналізу AlaAT, SDS-PAGE електрофорез та Вестерн-блоттинг були виконані за допомогою протоколу Muench and Good (1994). Екстракція РНК, синтез кДНК і полімеразна ланцюгова реакція (ПЛР) у режимі реального часу Екстракція РНК, синтез кДНК і кількісна ПЛР в режимі реального часу була проведена за допомогою протоколу Beatty et. al., 2009. Генспецифічні праймери і пари зондів, попередньо використані Beatty et al. (2009), були застосовані для того, щоб виявити відмінності у генній експресії через надекспресію HvAlaAT. NB рослини були використані як негативний контроль, а 18srРНК була використана як ендогенний контроль. Біоінформаційний аналіз промотора PBpr1 нуклеотидна послідовність була обрана, ґрунтуючись на гомології з OsANT1 промотором, з використанням програм BLASTn Національного центру біотехнологічної інформації (NCBI); далі вона була досліджена за допомогою генних біоінформаційних засобів Genbank і Refseq NCBI. GUS гістохімічне забарвлювання Загалом 20 насінин T1 трансгенних ліній PBpr1:GUSplus, названих PBpr1G лінії, були 14 UA 116086 C2 -1 5 10 стерилізовані та пророщені на стерильному рідкому середовищі MS (4,4 г Л , pH 5,8) для забарвлювання. Для кожної лінії були відібрані п'ять насінин на кожному етапі відбору зразків для забарвлювання. Сіянці були відібрані на 3-ій день після проростання (ДПП) і від 7 до 10 ДПП і забарвлені для GUS експресії. РЕЗУЛЬТАТИ: Врожайність насіння гетерозиготного T0 і попередній скринінг біомаси На стадії T0, незалежні PBpr1 рослини мали вищу надземну біомасу і врожайність насіння у порівнянні з NB (Таблиця 1). Дві незалежні PBpr1 лінії були відібрані, беручи до уваги високу врожайність насіння і біомасу, для відтворення цих ознак у T 1 поколінні. При порівнянні шести ліній, відібраних з NB, вони мали значно вищу врожайність насіння (p
ДивитисяДодаткова інформація
Назва патенту англійськоюPlants having enganced nitrogen efficiency
Автори англійськоюGood, Allen, Lock, Yee Ying, Beatty, Perrin H.
Автори російськоюГуд Аллен, Лок Йи Йинг, Битти Пэррин Х.
МПК / Мітки
МПК: C12N 15/82, C12N 15/113, C12N 5/14, C12N 15/54, A01H 5/10, C12N 5/04
Мітки: підвищеною, трансгенна, рослина, азоту, ефективністю, використання, однодольна
Код посилання
<a href="https://ua.patents.su/47-116086-transgenna-odnodolna-roslina-z-pidvishhenoyu-efektivnistyu-vikoristannya-azotu.html" target="_blank" rel="follow" title="База патентів України">Трансгенна однодольна рослина з підвищеною ефективністю використання азоту</a>
Однодольна трансгенна рослина, що містить полінуклеотид, що кодує поліпептид, що обумовлює толерантність до гліфосату

Номер патенту: 96578
Опубліковано: 25.11.2011
Автори: Хекерт Мет'ю Дж., Вогт Марк Д., Лю Дунлун, Клаус Джон С., Чікойн Тімоті К., Хегстад Джеффрі М., Олсон Пол Д., Хатчісон Джеймс М., Касл Лінда А., Хейзел Крістін Б., Вілкінсон Джек К., Вон Джеймс Ф. Х., Гайда Ентоні Д., Джр., Сондерз Девід В., Піплз Кеннет А., Лу Алберт Л., Мере Вейн Дж., Маккатчен Біллі Фред, Грін Джеррі М., Мой Йорк, Чо Хьон-же
МПК: A01H 5/00, C12N 15/82
Мітки: поліпептид, кодує, толерантність, обумовлює, полінуклеотид, гліфосату, рослина, містить, однодольна, трансгенна
Формула / Реферат:
1. Однодольна трансгенна рослина, що містить полінуклеотид, який кодує поліпептид гліфосат-N-ацетилтрансферазу, де полінуклеотид функціонально зв'язаний з(а) промотором, який забезпечує експресію в рослині; та(б) принаймні однією копією енхансерної послідовності, що включає (і) нуклеотидну послідовність, представлену в SEQ ID NO: 72 або 85, або(іі) нуклеотидну послідовність, яка принаймні на 90% ідентична...
Трансгенна стійка до гербіцидів рослина сої, яка містить пакетовану подію 8264.44.06.1

Номер патенту: 115766
Опубліковано: 26.12.2017
Автори: Толедо Сандра, Цуй Юньсін, Паркхерст Дон, Чжоу Нін, Колон Джулісса, Райт Террі, Хельд Брюс, Рассел Шон, Новак Стефен Н., Хоффман Том, Секар Вайтхілінгам
МПК: C12N 5/14, A01H 5/00, C07H 21/04, C12N 15/82
Мітки: подію, трансгенна, сої, рослина, яка, 8264.44.06.1, пакетовану, гербіцидів, містить, стійка
Формула / Реферат:
1. Трансгенна клітина рослини сої, що містить геном, який містить полінуклеотидну ділянку, що має послідовність SEQ ID NO:27, де залишки 1-1494 вказаної послідовності SEQ ID NO:27 являють собою 5'-геномні фланкуючі послідовності, залишки 1495-1497 вказаної послідовності SEQ ID NO:27 являють собою вставку з трьох пар основ, залишки 1498-11774 вказаної послідовності SEQ ID NO:27 являють собою вставку Т-ланцюга pDAB8264, і залишки 11775-13659...
Трансгенна рослина кукурудзи mir604

Номер патенту: 94893
Опубліковано: 25.06.2011
Автори: Чен Ерік, Штейнер Генрі-Йорк, Мегхіджі Мьоз
МПК: C12N 15/11, A01H 5/00, C12Q 1/68, G01N 33/53
Мітки: кукурудзи, трансгенна, рослина, mir604
Формула / Реферат:
1. Виділена молекула нуклеїнової кислоти, що містить SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 3 або SEQ ID NO: 4 або комплементи SEQ ID NO: 1-4.2. Амплікон, який містить молекулу нуклеїнової кислоти за п. 1.3. Рослина кукурудзи, яка містить молекулу нуклеїнової кислоти за п. 1.4. Рослина кукурудзи за п. 3, у якій після розщеплення геномної ДНК рослини рестриктазою KрnІ при гібридизації в строгих умовах з використанням...
Трансгенна рослина зі збільшеним рівнем рослинного крохмалю

Номер патенту: 109141
Опубліковано: 27.07.2015
Автори: Рааб Р. Майкл, Бугрі Олег, Емері Йонас, Лессард Філіп А., Ланахан Майкл, Самойлов Владімір
МПК: A01H 5/00, C12N 15/82, C12N 15/52
Мітки: збільшеним, крохмалю, трансгенна, рослинного, рослина, рівнем
Формула / Реферат:
1. Трансгенна рослина, яка включає конструкцію РНКі, що включає:першу послідовність драйвера, що включає першу виділену нуклеїнову кислоту, яка має щонайменше 90 % ідентичності з еталонною послідовністю, вибраною з групи, що складається з SEQ ID NO: 7, SEQ ID NO: 8, SEQ ID NO: 11, SEQ ID NO: 12, SEQ ID NO: 37, SEQ ID NO: 44 і SEQ ID NO: 45;другу послідовність драйвера, що включає другу виділену нуклеїнову кислоту, здатну...
Трансгенна рослина сої, стійка до інгібуючих ahas гербіцидів

Номер патенту: 107923
Опубліковано: 10.03.2015
Автори: Філью Елібіу Леопольду Реш, Ульбріш Адольфу, Тань Сиюань, Лоузано Луїс, Йотсумото Тадасі, Малефіт Тім, Аріас Карлос Альберту Аррабал, Лінеманн Уте, Араган Франсіску Жозе Ліма, Карлсон Дейл, Луцці Брюс М.
МПК: A01H 5/10, C12Q 1/68, C12N 15/82
Мітки: сої, стійка, гербіцидів, трансгенна, інгібуючих, рослина
Формула / Реферат:
1. Спосіб боротьби з бур'янистими рослинами на посівній площі, який включає:застосування ефективної кількості гербіцидної композиції, яка містить інгібуючий AHAS імідазоліноновий гербіцид, на посівній площі з рослиною сої, яка містить молекулу нуклеїнової кислоти, яка містить нуклеотидну послідовність, представлену положеннями 1312-6069 SEQ ID NO:1, де ефективна кількість буде інгібувати ріст бур'янистих рослин і рослин сої дикого...
Попередній патент: Напівпорожня шина для сільськогосподарських машин, зокрема для сівалок
Наступний патент: Композиція для лікування вірусу гепатиту c
Випадковий патент: Пристрій для вимірювання вологості