Спосіб визначення стресового стану базидіоміцетів та екологічного стану місця їх зростання за рівнем активності каталази



Формула / Реферат
Спосіб визначення стресового стану базидіоміцетів та екологічного стану місця їх зростання за рівнем активності каталази, що включає визначення каталазної активності штамів, який відрізняється тим, що визначення каталазної активності проводять в міцелії дикорослих плодових тіл їстівних лікарських базидіоміцетів Flammulina velutipes (Curt.: Fr.) Sing і Pleurotus ostreatus (Jacq.: Fr.) Kumm. з різних за екологічними умовами місць зростання та міцеліальних культур цих грибів при штучному культивуванні в оптимальних умовах і за дії температурного стресу.
Текст
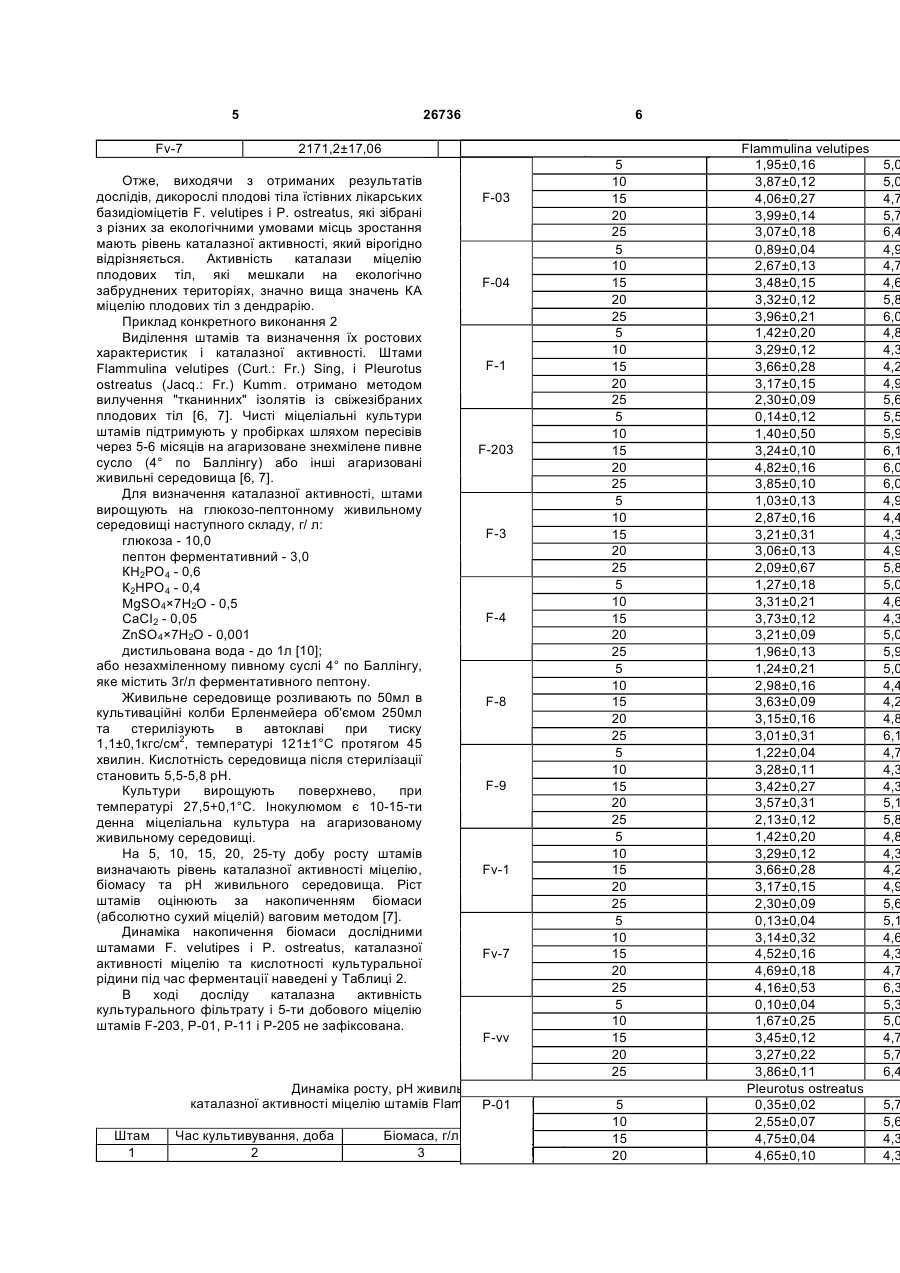
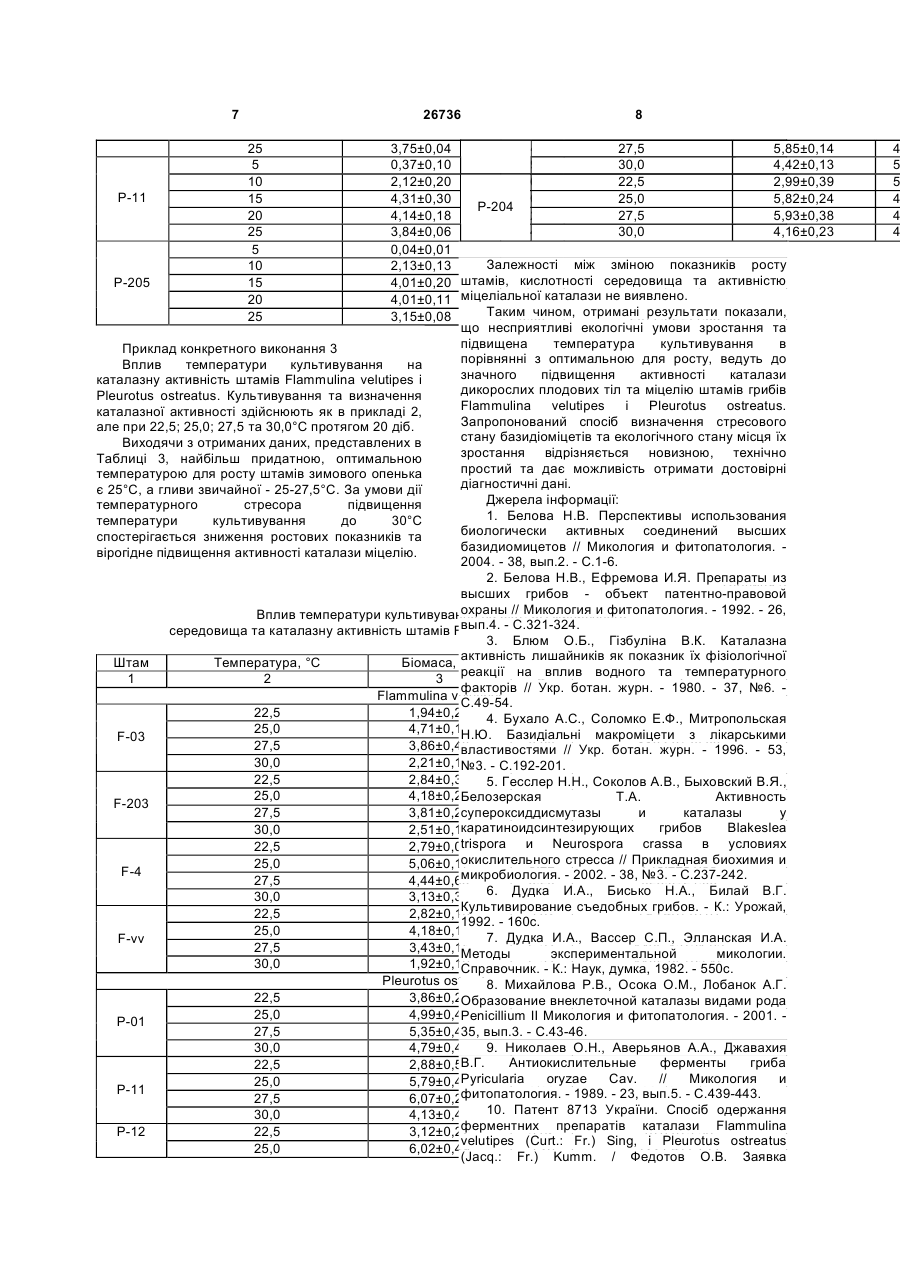
Спосіб визначення стресового стану базидіоміцетів та екологічного стану місця їх зростання за рівнем активності каталази, що 3 Cladonia mitis Sandst, Hypogymnia physodes. (L.) Nyl., Peltigera canina (L.) Willd., P. canina f. subcanina (Gyeln.) Oxn., P. aphthosa (L.) Willd. [3]; підвищення активності каталази і супероксиддисмутази в міцелії є одним з важливіших показників реакції культур каратиноїдсинтезуючих грибів Blakeslea trispora і Neurospora crassa в умовах окисного стресу [5]; антиокисні системи приймають участь в механізмі толерантності паразита як до протиінфекційних реакцій, так і до екстремальних впливів середовища [9, 13]; є результати пошуку продуцентів каталази серед дейтероміцетів Aspergillus і Реnісіllium [8] тощо. Дані щодо продуцентів каталази та зміни каталазної активності у базидіоміцетів за дії біотичних чи абіотичних стресорів не наводяться. Найбільш близький за технічною суттю і досяжності результату є спосіб одержання ферментних препаратів каталази штаму F-vv Flammulina velutipes (Curt.: Fr.) Sing, і штаму P-01 Pleurotus ostreatus (Jacq.: Fr.) Kumm., який містить визначення каталазної активності штамів та ферментних препаратів базидіоміцетів [10]. Вивчені динаміка активності каталази та ростових показників штамів, розроблено та апробовано метод виділення ферменту каталаза, не надаються дані про рівень каталазної активності дикорослих плодових тіл та міцелію грибів в залежності від дії несприятливих факторів довкілля чи умов культивування. В основу корисної моделі покладено завдання розробки біологічного способу визначення стресового стану базидіоміцетів та екологічного стану місця їх зростання за рівнем активності каталази, визначенням рівня каталазної активності міцелію базидіоміцетів за дії стресорів. Поставлене завдання вирішується тим, що в способі визначення стресового стану базидіоміцетів та екологічного стану місця їх зростання, визначають каталазну активність (КА) дикорослих плодових тіл їстівних лікарських базидіоміцетів Flammulina velutipes (Curt.: Fr.) Sing, і Pleurotus ostreatus (Jacq.: Fr.) Kumm. з різних, за екологічними умовами, місць зростання та міцеліальних культур цих грибів при штучному культивуванні в оптимальних умовах і за дії температурного стресу. Пропонований спосіб є новим тому, що він не відомий з рівня техніки та об'єктів дослідження. Приклад конкретного виконання 1 Визначення каталазної активності плодових тіл. Для дослідів з визначення активності каталази, беруть дикорослі плодові тіла приблизно одного віку з яких отримують і нові чисті міцеліальні культури (ізоляти, штами). Відповідно, плодові тіла і штами мають однакові назви. Дикорослі плодові тіла їстівних лікарських базидіоміцетів F. velutipes і P. ostreatus вилучені з різних за екологічними умовами місць зростання. Плодові тіла зимового опенька, штами Fv-1, Fv-7, F-vv і F-203 зібрані у екологічно забруднених районах м.Донецька, а також в дендрарії Донецького ботанічного саду НАН України. Гриб ріс на деревині мертвих чи ушкоджених листяних дерев, переважно роду Acer 26736 4 L.: А. hyrcanum Fisch. et Mey. - штам F-3, A. pseudoplatanus L. - штами Fv-1, Fv-7, F-1, F-4 і A. negundo L. - штами F-8, F-9. Штами F-vv і F-203 виділено з плодових тіл, які росли на пні робінії (Robinia pseudoacacia L.) та ушкодженому ясені ланцетному (Fraxinus lanceolata Borkn.) відповідно. Деякі плодові тіла F. velutipes - штами F-03 і F-04 були зібрані з трухлявих пнів чи з горілих залишків дерев, встановити систематичне положення яких неможливо. Плодові тіла гливи звичайної, штами Р-01, Р204, Р-205, Р-206, Р-211 і Р-212 зібрані вздовж автомобільних трас м.Донецька, а також в дендрарії Донецького ботанічного саду НАН України. Всі плодові тіла росли на пнях тополь: штами Р-01, Р-11 і Р-12 - тополі канадської (Populus deltoides Marsh.), штами Д-29, Р-204, Р205, Р-206, Р-211 і Р-212 - тополі самаркандської {Populus bolleana Lauche). Каталазну активність визначають за методом, який заснований на здатності пероксиду водню утворювати з солями молібдену стійкий забарвлений комплекс. Інтенсивність забарвлення вимірюють на спектрофотометрі, на приклад СФ26, при довжині хвилі 410нм супроти нульової проби з дистильованою водою. Каталазну активність розраховують за формулою [10]: Е=(Ак-Ад)·V·t·k·р (мкат/ л), де Е - активність каталази (мкат/л); Ак та Ад екстинкція контрольної та дослідної проб; V - об'єм проби (культуральний фільтрат - КФ, гомогенат міцелію - МГ), що вносили (0,1мл); t - час інкубації (600с); k - коефіцієнт мілімолярної екстинкції пероксиду водню, що дорівнює 22,2·103мМ-1·см-1; р - коефіцієнт розведення. В Таблиці 1 наведено результати дослідів з КА міцелію дикорослих плодових тіл зимового опенька і гливи звичайної. Експериментальні дані піддавали дисперсійному аналізу, порівняння середніх - за методом Дункана. Отримані дані показали, що КА міцелію плодових тіл F. velutipes і Р. ostreatus, які росли в екологічно забруднених місцях м.Донецька, вірогідно вища такої активності плодових тіл цих грибів відповідно з дендрарію Донецького ботанічного саду. Різниця значень КА зимового опенька і гливи звичайної не є вірогідною. Каталазна активність міцелію дикоросли тіл грибів Flammulina velutipes і Pleurotus Плодові тіла КАМГ, мкат/л·103 Flammulіnа velutipes F-03 1771,6±35,24 F-04 1771,6±102,04 F-1 1505,2±74,16 F-203 2490,8±26,64 F-3 1300,0±50,03 F-4 1370,9±15,36 F-8 1551,8±103,48 F-9 1411,9±16,48 Fv-1 2723,9±53,59 Плодові ті F-vv Д-29 P-01 P-11 P-12 P-204 P-205 P-206 P-211 5 Fv-7 26736 2171,2±17,06 6 P-212 2704,0±65,59 Flammulina velutipes 1,95±0,16 Отже, виходячи з отриманих результатів 3,87±0,12 дослідів, дикорослі плодові тіла їстівних лікарських F-03 4,06±0,27 базидіоміцетів F. velutipes і P. ostreatus, які зібрані 3,99±0,14 з різних за екологічними умовами місць зростання 3,07±0,18 мають рівень каталазної активності, який вірогідно 0,89±0,04 відрізняється. Активність каталази міцелію 2,67±0,13 плодових тіл, які мешкали на екологічно F-04 3,48±0,15 забруднених територіях, значно вища значень КА 3,32±0,12 міцелію плодових тіл з дендрарію. 3,96±0,21 Приклад конкретного виконання 2 1,42±0,20 Виділення штамів та визначення їх ростових 3,29±0,12 характеристик і каталазної активності. Штами F-1 3,66±0,28 Flammulina velutipes (Curt.: Fr.) Sing, і Pleurotus 3,17±0,15 ostreatus (Jacq.: Fr.) Kumm. отримано методом вилучення "тканинних" ізолятів із свіжезібраних 2,30±0,09 плодових тіл [6, 7]. Чисті міцеліальні культури 0,14±0,12 штамів підтримують у пробірках шляхом пересівів 1,40±0,50 через 5-6 місяців на агаризоване знехмілене пивне F-203 3,24±0,10 сусло (4° по Баллінгу) або інші агаризовані 4,82±0,16 живильні середовища [6, 7]. 3,85±0,10 Для визначення каталазної активності, штами 1,03±0,13 вирощують на глюкозо-пептонному живильному 2,87±0,16 середовищі наступного складу, г/ л: F-3 3,21±0,31 глюкоза - 10,0 3,06±0,13 пептон ферментативний - 3,0 2,09±0,67 КН2РО4 - 0,6 1,27±0,18 К2НРО4 - 0,4 3,31±0,21 MgSO4×7H2O - 0,5 F-4 3,73±0,12 СаСІ2 - 0,05 ZnSO4×7Н2О - 0,001 3,21±0,09 дистильована вода - до 1л [10]; 1,96±0,13 або незахміленному пивному суслі 4° по Баллінгу, 1,24±0,21 яке містить 3г/л ферментативного пептону. 2,98±0,16 Живильне середовище розливають по 50мл в F-8 3,63±0,09 культиваційні колби Ерленмейера об'ємом 250мл 3,15±0,16 та стерилізують в автоклаві при тиску 3,01±0,31 2 1,1±0,1кгс/см , температурі 121±1°С протягом 45 1,22±0,04 хвилин. Кислотність середовища після стерилізації 3,28±0,11 становить 5,5-5,8 рН. F-9 3,42±0,27 Культури вирощують поверхнево, при 3,57±0,31 температурі 27,5+0,1°С. Інокулюмом є 10-15-ти 2,13±0,12 денна міцеліальна культура на агаризованому 1,42±0,20 живильному середовищі. 3,29±0,12 На 5, 10, 15, 20, 25-ту добу росту штамів визначають рівень каталазної активності міцелію, Fv-1 3,66±0,28 біомасу та рН живильного середовища. Ріст 3,17±0,15 штамів оцінюють за накопиченням біомаси 2,30±0,09 (абсолютно сухий міцелій) ваговим методом [7]. 0,13±0,04 Динаміка накопичення біомаси дослідними 3,14±0,32 штамами F. velutipes і P. ostreatus, каталазної Fv-7 4,52±0,16 активності міцелію та кислотності культуральної 4,69±0,18 рідини під час ферментації наведені у Таблиці 2. 4,16±0,53 В ході досліду каталазна активність 0,10±0,04 культурального фільтрату і 5-ти добового міцелію 1,67±0,25 штамів F-203, Р-01, Р-11 і Р-205 не зафіксована. F-vv 3,45±0,12 3,27±0,22 Таблиця 2 3,86±0,11 Pleurotus ostreatus Динаміка росту, рН живильного середовища та каталазної активності міцелію штамів Flammulina velutipes і Pleurotus ostreatus. P-01 5 0,35±0,02 10 2,55±0,07 Штам Час культивування, доба Біомаса, г/л PH КАМГ, мкат/л·103 15 4,75±0,04 1 2 3 4 5 20 4,65±0,10 5 10 15 20 25 5 10 15 20 25 5 10 15 20 25 5 10 15 20 25 5 10 15 20 25 5 10 15 20 25 5 10 15 20 25 5 10 15 20 25 5 10 15 20 25 5 10 15 20 25 5 10 15 20 25 5,0 5,0 4,7 5,7 6,4 4,9 4,7 4,6 5,8 6,0 4,8 4,3 4,2 4,9 5,6 5,5 5,9 6,1 6,0 6,0 4,9 4,4 4,3 4,9 5,8 5,0 4,6 4,3 5,0 5,9 5,0 4,4 4,2 4,8 6,1 4,7 4,3 4,3 5,1 5,8 4,8 4,3 4,2 4,9 5,6 5,1 4,6 4,3 4,7 6,3 5,3 5,0 4,7 5,7 6,4 5,7 5,6 4,3 4,3 7 26736 25 5 10 15 20 25 5 10 15 20 25 8 3,75±0,04 4,47 27,5 2317,68±16,15 5,85±0,14 0,37±0,10 5,90 30,0 4,42±0,13 2,12±0,20 5,65 22,5 2082,36±27,01 2,99±0,39 P-11 4,31±0,30 4,05 25,0 2086,80±11,75 5,82±0,24 P-204 4,14±0,18 4,20 27,5 3123,64±24,01 5,93±0,38 3,84±0,06 5,42 30,0 3023,64±24,01 4,16±0,23 0,04±0,01 6,54 Залежності між зміною показників росту 2,13±0,13 6,53 2064,60±30,76 кислотності середовища та активністю P-205 4,01±0,20 штамів, 4,99 2064,60±7,69 4,01±0,11 міцеліальної каталази не виявлено. 5,46 3163,50±17,62 Таким чином, отримані результати показали, 3,15±0,08 5,95 2977,02±11,53 що несприятливі екологічні умови зростання та підвищена температура культивування в Приклад конкретного виконання 3 порівнянні з оптимальною для росту, ведуть до Вплив температури культивування на значного підвищення активності каталази каталазну активність штамів Flammulina velutipes і дикорослих плодових тіл та міцелію штамів грибів Pleurotus ostreatus. Культивування та визначення Flammulina velutipes і Pleurotus ostreatus. каталазної активності здійснюють як в прикладі 2, Запропонований спосіб визначення стресового але при 22,5; 25,0; 27,5 та 30,0°С протягом 20 діб. стану базидіоміцетів та екологічного стану місця їх Виходячи з отриманих даних, представлених в зростання відрізняється новизною, технічно Таблиці 3, найбільш придатною, оптимальною простий та дає можливість отримати достовірні температурою для росту штамів зимового опенька діагностичні дані. є 25°С, а гливи звичайної - 25-27,5°С. За умови дії Джерела інформації: температурного стресора підвищення 1. Белова Н.В. Перспективы использования температури культивування до 30°С биологически активных соединений высших спостерігається зниження ростових показників та базидиомицетов // Микология и фитопатология. вірогідне підвищення активності каталази міцелію. 2004. - 38, вып.2. - С.1-6. 2. Белова Н.В., Ефремова И.Я. Препараты из Таблиця 3 высших грибов - объект патентно-правовой охраны // Микология и фитопатология. - 1992. - 26, Вплив температури культивування на ріст, рН живильного вып.4. - С.321-324. середовища та каталазну активність штамів Flammulina velutipes і Pleurotus ostreatus. 3. Блюм О.Б., Гізбуліна В.К. Каталазна активність лишайників як КАМГ, мкат/л·103 показник їх фізіологічної Штам Температура, °С Біомаса, г/л рН реакції на вплив водного та температурного 1 2 3 4 5 факторів // Укр. ботан. журн. - 1980. - 37, №6. Flammulina velutipes С.49-54. 22,5 1,94±0,24 4. Бухало А.С., Соломко Е.Ф., Митропольская 5,77 405,8±12,41 25,0 4,71±0,11 6,44 900,3±21,34 Н.Ю. Базидіальні макроміцети з лікарськими F-03 27,5 3,86±0,40 6,20 1167,2±70,83 властивостями // Укр. ботан. журн. - 1996. - 53, 30,0 2,21±0,14 6,36 1003,9±34,10 №3. - С.192-201. 22,5 2,84±0,32 5. Гесслер Н.Н., Соколов501,3±9,31 5,85 А.В., Быховский В.Я., 25,0 4,18±0,26 6,52 1124,7±16,34 Белозерская Т.А. Активность F-203 супероксиддисмутазы и каталазы у 27,5 3,81±0,29 6,02 1507,4±101,04 каратиноидсинтезирующих 1011,1±67,39 Blakeslea грибов 30,0 2,51±0,14 6,73 trispora и Neurospora 329,9±16,10 условиях crassa в 22,5 2,79±0,08 5,93 окислительного стресса // Прикладная биохимия и 25,0 5,06±0,17 6,22 748,2±31,85 F-4 микробиология. - 2002. - 38, 928,6±24,05 №3. - С.237-242. 27,5 4,44±0,63 6,29 6. Дудка И.А., Бисько Н.А., Билай В.Г. 30,0 3,13±0,39 6,21 753,8±18,17 Культивирование съедобных грибов. - К.: Урожай, 22,5 2,82±0,18 5,82 793,40±16,01 1992. - 160с. 25,0 4,18±0,11 6,18 1285,38±26,15 7. Дудка И.А., Вассер С.П., Элланская И.А. F-vv 27,5 3,43±0,15 6,27 1406,43±21,02 Методы экспериментальной микологии. 30,0 1,92±0,19 6,37 1185,06±93,15 Справочник. - К.: Наук, думка, 1982. - 550с. Pleurotus оstreatus Михайлова Р.В., Осока О.М., Лобанок А.Г. 8. 22,5 3,86±0,26 5,22 Образование внеклеточной1198,5±38,54 каталазы видами рода 25,0 4,99±0,42 2063,7±7,38 Penicillium 4,97 II Микология и фитопатология. - 2001. P-01 27,5 5,35±0,43 вып.3. - С.43-46. 4,88 2690,6±6,71 35, 30,0 4,79±0,44 9. Николаев О.Н., Аверьянов А.А., Джавахия 4,95 2015,9±63,33 В.Г. Антиокислительные 438,7±3,16 ферменты гриба 22,5 2,88±0,56 5,76 Pyricularia 5,03 oryzae Cav. 961,0±21,81 // Микология и 25,0 5,79±0,47 P-11 фитопатология. - 1989. - 23,1421,3±52,05 вып.5. - С.439-443. 27,5 6,07±0,23 4,54 30,0 4,13±0,47 10. Патент 8713 України. Спосіб одержання 5,13 980,9±43,33 ферментних препаратів каталази Flammulina P-12 22,5 3,12±0,22 5,62 678,0±9,98 velutipes (Curt.: Fr.) Sing, і Pleurotus ostreatus 25,0 6,02±0,49 5,26 1221,4±73,11 (Jacq.: Fr.) Kumm. / Федотов O.B. Заявка 4 5 5 4 4 4 9 №200501341, від 14.02.2005, кл.7 C12N9/58, A612K38/00, Бюл. №8, від 15.08.05. (прототип) 11. Соломко Э.Ф., Дудка И.А. Перспективы использования высших базидиомицетов в микробиологической промышленности// ВНИСЭТИ: Обзорная информация, сер.3. - М., 1985. - 48с. 12. Tardif A. La Mycotherapie ou Les proprietes Medicinales des Champignons. - Paris, 2000. - 167p. 13. Sgherri C.L.M., Maffei M., Navari - Izzo F. Antioxidative enzymes in wheat subjected to increasing water deficit and rewatering // J. Plant Physiol. - 2000. - 157, N3. - P.273-279. 26736 10
ДивитисяДодаткова інформація
Назва патенту англійськоюMethod for determining stress state of basidiomycetes and ecological state of place of growth thereof according to catalase activity level
Автори англійськоюFedotov Oleh Valeriiovych
Назва патенту російськоюСпособ определения стрессового состояния базидиомицетов и экологического состояния места их роста по уровню активности каталазы
Автори російськоюФедотов Олег Валериевич
МПК / Мітки
МПК: A01H 15/00
Мітки: зростання, екологічного, спосіб, базидіоміцетів, каталази, визначення, стану, стресового, рівнем, активності, місця
Код посилання
<a href="https://ua.patents.su/5-26736-sposib-viznachennya-stresovogo-stanu-bazidiomicetiv-ta-ekologichnogo-stanu-miscya-kh-zrostannya-za-rivnem-aktivnosti-katalazi.html" target="_blank" rel="follow" title="База патентів України">Спосіб визначення стресового стану базидіоміцетів та екологічного стану місця їх зростання за рівнем активності каталази</a>
Спосіб визначення стресового стану базидіоміцетів та екологічного стану місця їх зростання

Номер патенту: 6372
Опубліковано: 16.05.2005
Автор: Федотов Олег Валерійович
Мітки: стресового, базидіоміцетів, екологічного, стану, зростання, визначення, спосіб, місця
Формула / Реферат:
Спосіб визначення стресового стану базидіоміцетів та екологічного стану місця їх зростання включає визначення пероксидазної активності штамів, який відрізняється тим, що включає визначення пероксидазної активності дикорослих плодових тіл їстівних базидіоміцетів Flammulina velutipes (Curt.: Fr.) Sing. і Pleurotus ostreatus (Jacq.: Fr.) Kumm з різних за екологічними умовами місць зростання та міцеліальних культур цих грибів при штучному...
Спосіб визначення стресового стану базидіоміцетів та екологічного стану місця їх зростання за вмістом продуктів перекисного окиснення ліпідів

Номер патенту: 12384
Опубліковано: 15.02.2006
Автор: Федотов Олег Валерійович
Мітки: базидіоміцетів, перекисного, місця, екологічного, стану, окиснення, вмістом, продуктів, стресового, визначення, ліпідів, спосіб, зростання
Формула / Реферат:
Спосіб визначення стресового стану базидіоміцетів та екологічного стану місця їх зростання за вмістом продуктів перекисного окиснення ліпідів, що включає визначення вмісту тіобарбітурової кислоти як активних продуктів в культурах грибів, який відрізняється тим, що визначення їх вмісту проводять в дикорослих плодових тілах базидіоміцетів із різних за екологічними умовами місць зростання та міцеліальних культурах цих грибів при штучному...
Спосіб визначення каталазної активності базидіоміцетів

Номер патенту: 39243
Опубліковано: 15.06.2001
Автори: Федотов Олег Валерійович, Гавриленко Галина Володимирівна
МПК: C12N 9/50
Мітки: спосіб, каталазної, базидіоміцетів, активності, визначення
Формула / Реферат:
Спосіб визначення каталазної активності базидіоміцетів, який включає стадії утворення стійкого забарвленого комплексу перекису водню з солями молібдену, вимірювання його оптичної густини, який відрізняється тим, що включає культивування штамів, гомогенізацію соматичних структур або плодових тіл макроміцетів у буферному розчині чи дистильованій воді, ліофільне центрифугування, додавання до культурального фільтрату або надосадової рідини...
Спосіб визначення антиокисної активності культур базидіоміцетів

Номер патенту: 40415
Опубліковано: 16.07.2001
Автори: Федотов Олег Валерійович, Когут Ігор Олександрович
МПК: C12N 9/50
Мітки: спосіб, культур, активності, визначення, антиокисної, базидіоміцетів
Формула / Реферат:
Спосіб визначення антиокисної активності культур базидіоміцетів, що містить підготовку суспензії ліпопротеїдів жовтка курячих яєць, який відрізняється тим, що включає культивування штамів, підготовку культурального фільтрату до аналізу, гомогенізації грибних тканин - соматичних структур чи плодових тіл макроміцетів і ліофільного центрифугування міцеліальної суспензії, а також додавання до суспензії жовткових ліпопротеїдів культурального...
Спосіб одержання ферментних препаратів каталази flammulina velutipes (curt.: fr.) sing. i pleurotus ostreatus (jacq.: fr.) kumm.

Номер патенту: 8713
Опубліковано: 15.08.2005
Автор: Федотов Олег Валерійович
МПК: C12R 1/645, C12N 1/14, A01H 15/00, C12N 9/00
Мітки: kumm, velutipes, ostreatus, jacq, одержання, pleurotus, curt, спосіб, sing, препаратів, flammulina, каталази, ферментних
Формула / Реферат:
Спосіб одержання ферментних препаратів каталази Flammulina velutipes (Curt.: Fr.) Sing. і Pleurotus ostreatus (Jacq.: Fr.) Kumm., що включає культивування продуцентів на глюкозо-пептонному живильному середовищі, який відрізняється тим, що культивування ведеться до досягнення максимальної каталазної активності вегетативного міцелію, вилучення міцелію фільтруванням, його гомогенізацію, осадження ферменту сульфатом амонію при 60-80 %...
Попередній патент: Спосіб доочистки стічних та природних вод від органічних речовин, сульфідів, двовалентного заліза та аміаку
Наступний патент: Регулятор вологості кореневого шару ґрунту
Випадковий патент: Вимірювач частоти свіп-генератора