Спосіб визначення стресового стану базидіоміцетів та екологічного стану місця їх зростання



Формула / Реферат
Спосіб визначення стресового стану базидіоміцетів та екологічного стану місця їх зростання включає визначення пероксидазної активності штамів, який відрізняється тим, що включає визначення пероксидазної активності дикорослих плодових тіл їстівних базидіоміцетів Flammulina velutipes (Curt.: Fr.) Sing. і Pleurotus ostreatus (Jacq.: Fr.) Kumm з різних за екологічними умовами місць зростання та міцеліальних культур цих грибів при штучному культивуванні в оптимальних умовах і за дії стресорів.
Текст
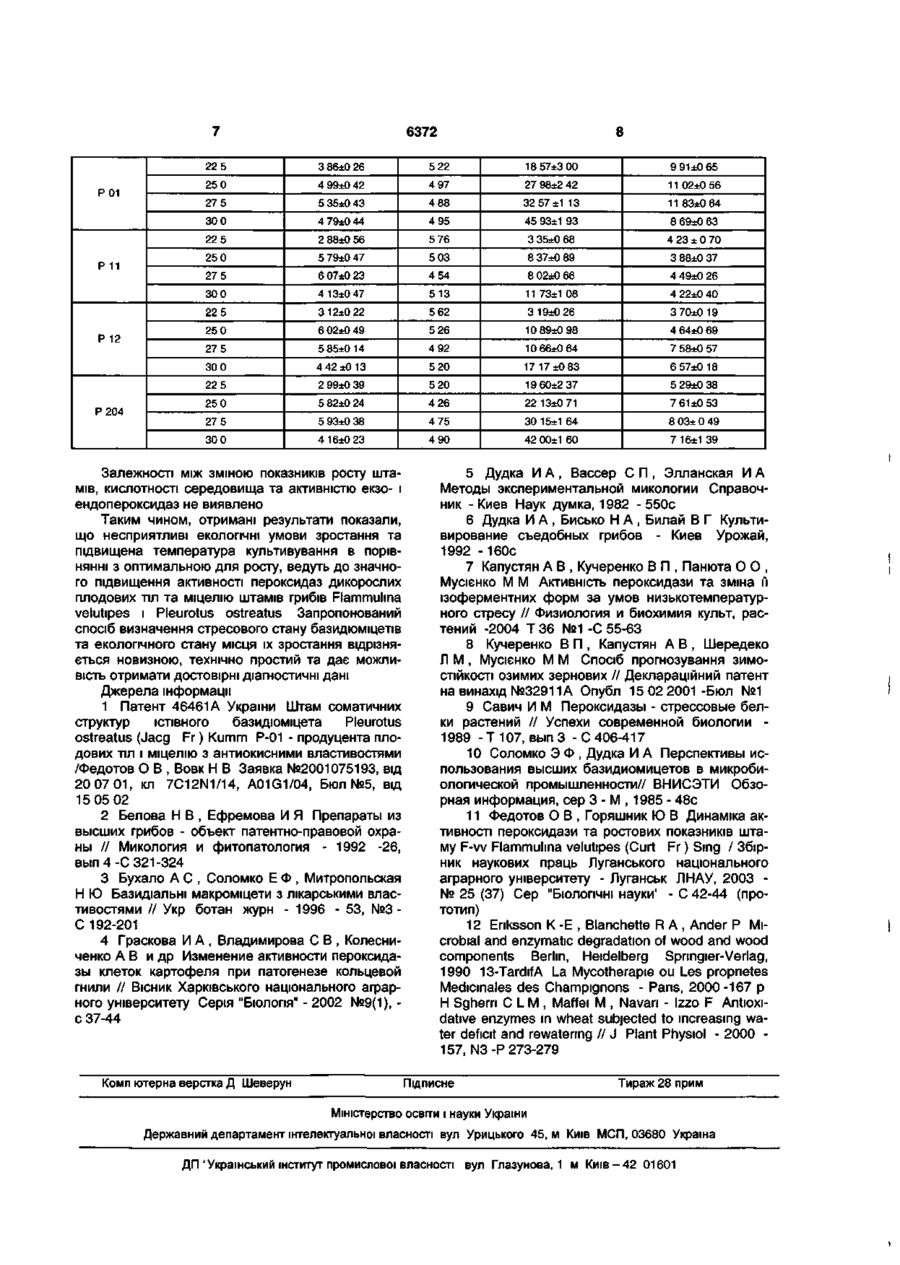
Спосіб визначення стресового стану базидіоміцетів та екологічного стану місця їх зростання включає визначення пероксидазної активності штамів, який відрізняється тим, що включає визначення пероксидазної активності дикорослих плодових тіл їстівних базидіоміцетів Flammuhna velutipes (Curt Fr) Sing і Pleurotus ostreatus (Jacq Fr) Kumm з різних за екологічними умовами місць зростання та міцеліальних культур цих грибів при штучному культивуванні в оптимальних умовах і за ди стресорів Корисна модель відноситься до мікологи, екологи і бютехнологн та може бути використана як індикатор стресового стану базидіоміцетів, у діагностиці екологічного стану довкілля, мікробіологічному виробництві, фармацевтичній та інших галузях промисловості Для більшості базидіоміцетів характерним субстратом є деревина та різноманітні рослинні відходи, що визначає провідну роль ксилотрофів в деструкції лігноцелюлозних субстратів [12] Як наслідок, їх ферментативний комплекс та бюсинтетичні процеси достатньо розвинені і лабільні Доведено ДОЦІЛЬНІСТЬ використання ксилотрофів перспективних об'єктів бютехнологм для отримання різноманітних БАР [2, 10, 13] Ксилотроф Flammuhna velutipes - зимовий опеньок має антибактеріальні, антивірусні, антифунгальні, протипухлинні, імуномодулюючі, гіпоглікемічні, тромболітичні властивості і представляє інтерес для грибівництва, фармакологічної і харчової промисловостей [3] Встановлено, що за вмістом активних речовин протипухлинної дії Pleurotus ostreatus глива звичайна стоїть на третьому МІСЦІ ПІСЛЯ ШИитаке й опенька літнього, має антивірусну, антифунгальну, радюпротекторну та імуномодулюючу дію, покращує роботу м язів [3] Джерелом фармакологічних речовин, в тому числі і ферментів, можуть бути не лише плодові тіла - дикорослі, або вирощені на щільних субстратах, але й міцеліальна біомаса, яку отримують сучасними бютехнолопчними методами культивування [5, 6] Переважна КІЛЬКІСТЬ ВИДІВ макромщетів на території Донбасу є малодослідженими Для створення штамового різноманіття необхідна їх інтродукція, що дасть змогу здійснити скринінг і виявити перспективні штами для бютехнолопчного використання Несприятливі умови ЖИТТЄДІЯЛЬНОСТІ спричиняють розширення пластичності функцій та властивостей усіх головних метаболічних систем макромщетів Адаптаційні перебудови в першу чергу стосуються ферментативних систем, вони призводять до утворення ферментів, здатних активно функціонувати за дії несприятливих факторів [14] Одним із таких надзвичайно функціонально лабільних ферментів, що реагує на порушення гомеостазу клітинного метаболізму при дії різноманітних стресорів, є пероксидаза ( 1 1 1 1 7 ) Слід ВІДМІТИТИ, що біосинтез та властивості грибних пероксидаз практично не вивчені, а літературні дані стосуються таких ферментів рослинного, мікробного і тваринного походження Відзначаються зміни як у наборі молекулярних форм ферменту, так і в їх активності за дії біотичних та абіотичних чинників Отже, розробка способів оцінки стресового стану макроміцетів на несприятливі умови культивування чи навколишнього середовища, бюіндикацн екологічного стану довкілля є актуальними проблемами розвитку мікологи, бютехнологн та екологи Відомо, ЩО спостерігається достовірне збільшення пероксидазної активності клітин картоплі при патогенезі Clavibacter michiganensis subsp sepedonicus (Cms) [4], озимих зернових під час водного дефіциту [8], озимих злаків за дії температурного стресу [7] тощо Літературні дані стосуються рослинних організмів Дані щодо зміни пероксидазної активності макроміцетів за ди біотичних чи абіотичних стресорів не наводяться Найбільш близький за технічною суттю і досяжності результату є штам F-w Flammuhna velutipes CM со (О О) 6372 (Curt.: Fr.) Sing. - продуцент плодових тіл і міцелію з антиокисними властивостями та пероксидаз [11]. Вивчені динаміка активності пероксидази та ростових показників штаму F-vv, не надаються дані про рівень пероксидазної активності плодових тіл та культури в залежності від дії несприятливих факторів довкілля чи умов культивування. В основу корисної моделі покладено завдання розробки біологічного способу діагностики стресового стану базидіоміцетів та екологічного стану місця їх зростання, який відрізняється від прототипу визначенням рівня активності екзо- та ендогенних пероксидаз макроміцетів за дії стресорів. Поставлене завдання вирішується тим, що в способі визначення стресового стану базидіоміцетів та екологічного стану місця їх зростання, визначають пероксидазну активність (ПА) дикорослих плодових тіл їстівних базидіоміцетів Flammulina velutipes (Curt.: Fr.) Sing, і Pleurotus ostreatus (Jacq.: Fr.) Kumm. з різних, за екологічними умовами, місць зростання та міцеліальних культур цих грибів при штучному культивуванні в оптимальних умовах і за дії температурних стресорів. Пропонований спосіб є новим тому, що він не відомий з рівня техніки та об'єктів дослідження. Приклад конкретного виконання 1 Визначення пероксидазної активності плодових тіл. Для дослідів з ПА, беруть плодові тіла приблизно одного віку з яких отримують і нові чисті міцеліальні культури (ізоляти, штами). Відповідно, плодові тіла і штами мають однакові назви. Дикорослі плодові тіла їстівних базидіоміцетів Flammulina velutipes (Curt.: Fr.) Sing, і Pleurotus ostreatus (Jacq.: Fr.) Kumm. з різних за екологічними умовами місць зростання. Плодові тіла зимового опенька, штами Fv-1, Fv-7, F-w і F-203 зібрані у екологічно забруднених районах м. Донецька, а також в дендрарії Донецького ботанічного саду ПАН України, Гриб ріс на деревині мертвих чи ушкоджених листяних дерев, переважно роду Acer L: A. hyrcanum Fisch. et Mey. - штам F-3, A. pseudoplatanus L. - штами Fv-1, Fv-7, F-1, F-4 і А. negundo L - штами F-8, F-9. Штами F-w і F-203 виділено з плодових тіл, які росли на пні робінії (Robinia pseudoacacia L.) та ушкодженому ясені ланцетному (Fraxinus lanceolata Borkn.) відповідно. Деякі плодові тіла F.velutipes - штами F-03 і F-04 були зібрані з трухлявих пнів чи з горілих залишків дерев, встановити систематичне положення яких неможливо. Плодові тіла гливи звичайної, штами Р-01, Р204, Р-205, Р-206, Р-211 і Р-212 зібрані вздовж автомобільних трас м.Донецька, а також в дендрарії Донецького ботанічного саду ПАН України. Всі плодові тіла росли на пнях тополь: штами Р01, Р-11 і Р-12 - тополі канадської (Populus deltoides Marsh.), штами Д-29, Р-204, Р-205, Р-206, Р211 і Р-212 - тополі самаркандської (Populus bolleana Lauche). Визначають пероксидазну активність плодових тіл їстівних базидіоміцетів Flammulina velutipes (Curt.: Fr.) Sing, і Pleurotus ostreatus (Jacq.: Fr.) Kumm. за методом, який базується на визначенні за допомогою фотоелектроколориметра інтенсивності забарвлення продукту окислення одіанізидину перекисом водню, який утворився при дії пероксидази. Одиниця пероксидази відповідає кількості ферменту, що каталізує перетворення 1 мкмоля Н2О2 за одну хвилину в оптимальних умовах. Розрахунки проводять за формулою [5]: Х= РE-|-K-V2 t де, X - активність ферменту пероксидази; Е екстинкція; Vi - об'єм забарвленої проби; V2 - об'єм гомогенату міцелію (МГ) або культурального фільтрату (КФ); Еі - коефіцієнт мікромолярної екстинкції (0,0128); К - коефіцієнт для перерахунку з міліметрів у літри (1000); t - час інкубації (5хв.); р розведення. В табл.1 наведено результати дослідів з ПА міцелію дикорослих плодових тіл гливи звичайної і зимового опенька. Експериментальні дані піддавали дисперсійному аналізу, порівняння середніх за методом Дункана. Отримані дані показали, що ПА плодових тіл зимового опенька, які росли в екологічно забруднених місцях м.Донецька, вірогідно вища такої активності плодових тіл Flammulina velutipes і Pleurotus ostreatus з дендрарію Донецького ботанічного саду. ПА плодових тіл гливи звичайної різних місць зростання також вірогідно відрізняється. Є вірогідною різниця ПА зимового опенька і гливи звичайної, які росли в м. Донецьку. Різниця між пероксидазною активністю плодових тіл зимового опенька з дендрарію Донецького ботанічного саду і гливи звичайної не є вірогідною. Таблиця 1 Пєроксидазна активність міцелію дикорослих плодових тіл грибів Flammulina velutipes і Pleurotus ostreatus 1 2 3 4 Плодові тіла ПАМГ, »10 2 ум. од. Плодові тіла ПАМГ, • 10 2 ум. од. Flammulina velutipes F-w 42,77±2,87 F-03 19,03 ±2,04 Pleurotus ostreatus F-04 22,87±1,19 Д-29 9,73±0,66 F-1 12,27±0,97 P-01 26,47±1,47 F-203 67,03±4,29 P-11 9,03±0,81 F-3 27,37±0,61 P-12 11,73±0,83 F-4 27,13±0,93 P-204 21,07±0,60 F-8 15,97±0,85 P-205 31,10+1,60 6372 F-9 11,17±0,45 P-206 19,23±1,16 Fv-1 50,73±1,72 P-211 24,87±1,70 Fv-7 53,30±1,83 P-212 25,77±1,59 Отже, виходячи з отриманих результатів дослідів, дикорослі плодові тіла їстівних базидіоміцетів Flammulina velutipes (Curt.: Fr.) Sing, і Pleurotus ostreatus (Jacq.: Fr.) Kumm., які зібрані з різних за екологічними умовами місць зростання мають рівень пероксидазної активності, що вірогідно відрізняється. ПА плодових тіл, які мешкали на екологічно забруднених територіях, значно вища значень пероксидазної активності міцелію плодових тіл з дендрарію. Приклад конкретного виконання 2 Виділення чистих культур, їх пересіви та культивування. Зібрані плодові тіла, які росли в різних екологічних умовах, звільняють від залишків субстрату. Для подальших досліджень використовують бажано одновікові зразки. Штами їстівних базидіоміцетів Flammulina velutipes та Pleurotus ostreatus отримано за методами, які звичайно застосовуються для одержання чистих культур макроміцетів, а саме вилученням "тканинних" ізолятів із свіже-зібраних плодових тіл [5]. Чисту міцеліальну культуру штамів підтримують шляхом пересівів через 5-6 місяців на агаризоване знехмілене пивне сусло (4° по Баллінгу), або інші агаризовані живильні середовища [6]. Для вивчення росту і визначення пероксидазної активності, штами вирощують на рідкому глюкозо-пептонному середовищі [5] або знехміленому пивному суслі (4° по Баллінгу), яке містить Зг/л ферментативного пептону. Вихідна кислотність середовища після стерилізації становить від 5,5 до 5,8рН. Культури вирощують поверхнево в колбах Ерленмейера місткістю 250мл з об'ємом середовища 50мл. Посівним матеріалом є 10-15-денна культура гриба, яка росла на агаризованому живильному середовищу. Термін культивування до ЗО діб. Для отримання плодових тіл, штам вирощують за інтенсивною технологією [6]. Приклад конкретного виконання З Вплив температури культивування на пероксидазну активність штамів Flammulina velutipes і Pleurotus ostreatus. Штами культивують в колбах Ерленмейєра на рідкому глюкозо-пептонному середовищі при 22,5; 25,0; 27,5 та 30,0°С протягом 20 діб. Пероксидазну активність міцелію та культурального фільтрату визначають по інтенсивності забарвлення продукту окислення о-діанізидину перекисом водню, який утворився при дії пероксидази [5]. Виходячи з отриманих даних (табл.2), найбільш придатною, оптимальною температурою для росту штамів зимового опенька є 25°С, а гливи звичайної - 25-27,5°С. За умови дії температурного стресора підвищення температури культивування до 30°С спостерігається зниження ростових показників та вірогідне підвищення ПА міцелію. Щодо ПА культурального фільтрату, то зафіксовано її достовірне підвищення для всіх штамів при 27,5°С культивування. З підвищенням температури до 30,0°С відбувається збільшення пероксидазної активності КФ штамів F-203, F-w та зниження активності пероксидаз інших штамів, яке не є вірогідним. Таблиця 2 Вплив температури культивування на ріст, рН живильного середовища та пероксидазну активність штамів Flammulina velutipes і Pleurotus ostreatus 1 2 3 4 5 6 Штам Температура, °С Біомаса, г/л pH ПАМГ, • 10 2 ум. од. ПАКФ, • 10 2 ум. од. 1 2 3 4 5 6 Flammulina velutipes 22,5 F-03 1,94±0,24 5,77 8,38±0,71 1,83±0,35 25,0 4,71±0,11 6,44 13,93±0,68 2,27±0,46 4,64± 0,22 6,20 19,60±1,85 2,21 ±0,14 6,36 29,30±1,16 3,82±0,37 2,84±0,32 5,85 20,53±2,74 2,58±0,32 25,0 4,18±0,26 6,52 47,27±3,72 2,10±0,07 27,5 3,81±0,29 6,02 73,30±2,19 3,82±0,23 30,0 2,51±0,14 6,73 76,60±1,19 4,37±0,35 22,5 2,79±0,08 5,93 14,83±0,44 1,84±0,23 25,0 5,06±0,17 6,22 14,73 ±2,59 2,86±0,36 27,5 4,44±0,63 6,29 27,20 ± 2,40 7,52±0,73 30,0 F-4 3,86±0,40 22,5 F-203 27,5 30,0 3,13±0,39 6,21 26,99±1,10 5,44± 0,25 22,5 F-w 2,82±0,18 5,82 20,70±2,12 3,69 ± 0,62 25,0 4,18±0,11 6,18 25,87±2,58 4,25±0,31 27,5 3,43 ±0,15 6,27 35,99 ±3,29 7,07±0,64 30,0 1,92 ±0,19 6,37 41,00±1,30 7,39±0,41 Pleurotus ostreatus 6372 22 5 11 02±0 56 27 5 5 35±0 43 4 88 32 57±1 13 11 83±0 64 4 79±0 44 4 95 45 93±1 93 8 69±0 63 2 88±0 56 5 76 3 35±0 68 4 23 ± 0 70 25 0 5 79±0 47 5 03 8 37±0 89 3 88+0 37 27 5 6 07±0 23 4 54 8 02±0 66 4 49±0 26 4 13±0 47 5 13 11 73±1 08 4 22±0 40 22 5 3 12±0 22 5 62 3 19±0 26 3 70±0 19 25 0 6 02±0 49 5 26 10 89±0 98 4 64±0 69 27 5 5 85±0 14 4 92 10 66±0 64 7 58±0 57 30 0 4 42±0 13 5 20 17 17 ±0 83 6 57±0 18 22 5 2 99±0 39 5 20 19 60±2 37 5 29±0 38 25 0 5 82±0 24 4 26 22 13±0 71 7 61 ±0 53 27 5 5 93±0 38 4 75 30 15±1 64 8 03± 0 49 30 0 P204 9 91±0 65 27 98±2 42 30 0 P 12 18 57±3 00 4 97 22 5 p 11 5 22 4 99±0 42 30 0 P01 3 86±0 26 25 0 4 16±0 23 4 90 42 00±1 60 7 16±1 39 Залежності між зміною показників росту штамів, кислотності середовища та активністю екзо- і ендопероксидаз не виявлено Таким чином, отримані результати показали, що несприятливі екологічні умови зростання та підвищена температура культивування в порівнянні з оптимальною для росту, ведуть до значного підвищення активності пероксидаз дикорослих плодових тіл та міцелію штамів грибів Flammulma velutipes і Pleurotus ostreatus Запропонований спосіб визначення стресового стану базидіоміцетів та екологічного стану місця їх зростання відрізняється новизною, технічно простий та дає можливість отримати достовірні діагностичні дані Джерела інформації 1 Патент 46461А України Штам соматичних структур їстівного базидюмщета Pleurotus ostreatus (Jacg Fr) Kumm P-01 - продуцента плодових тіл і міцелію з антиокисними властивостями /Федотов О В , Вовк Н В Заявка №2001075193, від 2007 01, кл 7C12N1/14, A01G1/04, Бюл №5, від 15 05 02 2 Белова Н В , Ефремова И Я Препараты из высших грибов - объект патентно-правовой охраны // Микология и фитопатология - 1992 -26, вып 4 -С 321-324 3 Бухало А С , Соломко Е Ф , Митропольская Н Ю Базидіальні макроміцети з лікарськими властивостями // Укр ботан журн - 1996 - 53, №3 С 192-201 4 Праскова И А , Владимирова С В , Колесниченко А В и др Изменение активности пероксидазы клеток картофеля при патогенезе кольцевой гнили // Вісник Харківського національного аграрного університету Серія "БІОЛОГІЯ" - 2002 №9(1), с 37-44 Комп ютерна верстка Д Шеверун 5 Дудка И А , Вассер С П , Элланская И А Методы экспериментальной микологии Справочник - Киев Наук думка, 1982 - 550с 6 Дудка И А , Бисько Н А , Билай В Г Культивирование съедобных грибов - Киев Урожай, 1992 -160с 7 Капустян А В , Кучеренко В П , Панюта О О , Мусієнко М М Активність пероксидази та зміна її ізоферментних форм за умов низькотемпературного стресу // Физиология и биохимия культ, растений -2004 Т 36 №1 -С 55-63 8 Кучеренко В П , Капустян А В , Шередеко Л М , Мусієнко М М Спосіб прогнозування ЗИМОСТІЙКОСТІ озимих зернових // Деклараційний патент на винахід №32911А Опубл 15 02 2001-Бюл №1 9 Савич И М Пероксидазы - стрессовые белки растений // Успехи современной биологии 1989 -Т107, выпЗ -С 406-417 10 Соломко Э Ф , Дудка И А Перспективы использования высших базидиомицетов в микробиологической промышленности// ВНИСЭТИ Обзорная информация, сер 3 - М , 1985 - 48с 11 Федотов О В , Горяшник Ю В Динаміка активності пероксидази та ростових показників штаму F-w Flammulina velutipes (Curt Fr) Sing / Збірник наукових праць Луганського національного аграрного університету - Луганськ ЛНАУ, 2003 № 25 (37) Сер "БІОЛОГІЧНІ науки' - С 42-44 (про тотип) 12 Eriksson К -Е , Blanchette R A , Ander P Microbial and enzymatic degradation of wood and wood components Berlin, Heidelberg Spnngier-Verlag, 1990 13-TardifA La Mycotherapie ou Les propnetes Medicinales des Champignons - Pans, 2000-167 p H Sghern С L M , Maffei M , Navan - Izzo F Antioxidative enzymes in wheat subjected to increasing water deficit and rewatenng // J Plant Physiol - 2000 157, N3-P 273-279 Підписне Тираж 28 прим Міністерство освіти і науки України Державний департамент інтелектуальної власності вул Урицького 45, м Київ МСП, 03680 Україна ДП'Український інститут промислової власності вул Глазунова, 1 м К и ї в - 4 2 01601
ДивитисяДодаткова інформація
Назва патенту англійськоюMethod of determining the stress state of basidiomycetes and the ecological state of place of growing thereof
Автори англійськоюFedotov Oleh Valeriiovych
Назва патенту російськоюСпособ определения стрессового состояния базидиомицетов и экологического состояния места их произрастания
Автори російськоюФедотов Олег Валериевич
МПК / Мітки
Мітки: спосіб, зростання, місця, стану, екологічного, стресового, базидіоміцетів, визначення
Код посилання
<a href="https://ua.patents.su/4-6372-sposib-viznachennya-stresovogo-stanu-bazidiomicetiv-ta-ekologichnogo-stanu-miscya-kh-zrostannya.html" target="_blank" rel="follow" title="База патентів України">Спосіб визначення стресового стану базидіоміцетів та екологічного стану місця їх зростання</a>
Спосіб визначення каталазної активності базидіоміцетів

Номер патенту: 39243
Опубліковано: 15.06.2001
Автори: Гавриленко Галина Володимирівна, Федотов Олег Валерійович
МПК: C12N 9/50
Мітки: базидіоміцетів, активності, визначення, каталазної, спосіб
Формула / Реферат:
Спосіб визначення каталазної активності базидіоміцетів, який включає стадії утворення стійкого забарвленого комплексу перекису водню з солями молібдену, вимірювання його оптичної густини, який відрізняється тим, що включає культивування штамів, гомогенізацію соматичних структур або плодових тіл макроміцетів у буферному розчині чи дистильованій воді, ліофільне центрифугування, додавання до культурального фільтрату або надосадової рідини...
Спосіб визначення антиокисної активності культур базидіоміцетів

Номер патенту: 40415
Опубліковано: 16.07.2001
Автори: Федотов Олег Валерійович, Когут Ігор Олександрович
МПК: C12N 9/50
Мітки: культур, визначення, активності, базидіоміцетів, антиокисної, спосіб
Формула / Реферат:
Спосіб визначення антиокисної активності культур базидіоміцетів, що містить підготовку суспензії ліпопротеїдів жовтка курячих яєць, який відрізняється тим, що включає культивування штамів, підготовку культурального фільтрату до аналізу, гомогенізації грибних тканин - соматичних структур чи плодових тіл макроміцетів і ліофільного центрифугування міцеліальної суспензії, а також додавання до суспензії жовткових ліпопротеїдів культурального...
Живильне середовище для виділення і пересівання чистих культур вищих базидіоміцетів

Номер патенту: 63177
Опубліковано: 15.01.2004
Автори: Бугрім Євген Юрійович, Крюков Олексій Анатолійович, Федотов Олег Валерійович
МПК: A01H 15/00
Мітки: живильне, пересівання, вищих, базидіоміцетів, виділення, культур, чистих, середовище
Формула / Реферат:
Винахід відноситься до біотехнології і може бути використаний у науково-дослідних і промислових мікологічних лабораторіях для отримання з плодових тіл і пересівів чистих культур вищих дереворуйнівних базидіоміцетів-продуцентів біологічно активних речовин для різних галузей промисловості, втому числі - харчової і фармакологічної.Оптимізація умов штучного культивування та вивчення фізіологічно активних речовин базидіоміцетів -...
Спосіб культивування базидіоміцетів

Номер патенту: 26705
Опубліковано: 12.11.1999
Автори: Єжов Валерій Микитович, Осовик Анатолій Миколайович, Лупашко Валентина Олексіївна, НОСКО Інна Іванівна
МПК: C12N 9/02, A01H 15/00, C12N 1/14
Мітки: спосіб, культивування, базидіоміцетів
Формула / Реферат:
Способ культивирования базидиомицетов, предусматривающий выращивание грибного мицелия в аэробных условиях на питательной среде, содержащей сахарид в качестве источника углерода, минеральные соли и воду, отличающийся тем, что осуществляют культивирование базидиомицета Pleurotus ostreatus (Fr.) Kummer до концентрации биомассы 12 - 22г сухого вещества/л, в качестве сахарида используют крахмалсодержащее сырье, а в питательную среду дополнительно...
Спосіб визначення нітратного екологічного навантаження на організм людини

Номер патенту: 24153
Опубліковано: 07.07.1998
Автори: Нетудихатка Олег Юрійович, Доренський Володимир Сергійович, Гоженко Анатолій Іванович, Гуцель Юрій Іларіонович, Славіна Ніна Георгієвна
МПК: G01N 33/48, A61B 10/00
Мітки: екологічного, спосіб, людини, навантаження, визначення, організм, нітратного
Формула / Реферат:
Способ определения нитратной экологической нагрузки на организм человека, включающий определение концентрации нитратов в моче, отличающийся тем, что до исследования мочи, обследуемому производят нагрузку дистиллированной водой в количестве 0,3 - 0,5% от массы тела, через, 1 - 2 часа производят забор мочи, определяют концентрацию нитратов в моче и рассчитывают нитратную нагрузку по формулеЭН = (ENO3сут. - ENO3энд.) ×...
Попередній патент: Пристрій для зриву і вилучення труб
Наступний патент: Носоглотковий металевий буж
Випадковий патент: Канатний механізм натиску екскаватора