Білий аспорогенний w1spo–мутант, резистентний до іонів міді, отриманий з paecilomyces lilacinus (thom) samson 146


Формула / Реферат
Білий аспорогенний W1Spo--мутант, резистентний до іонів міді, отриманий з Paecilomyces lilacinus (Thom) Samson 146, здатний накопичувати в своїй біомасі 104,88 мг/г іонів міді.
Текст
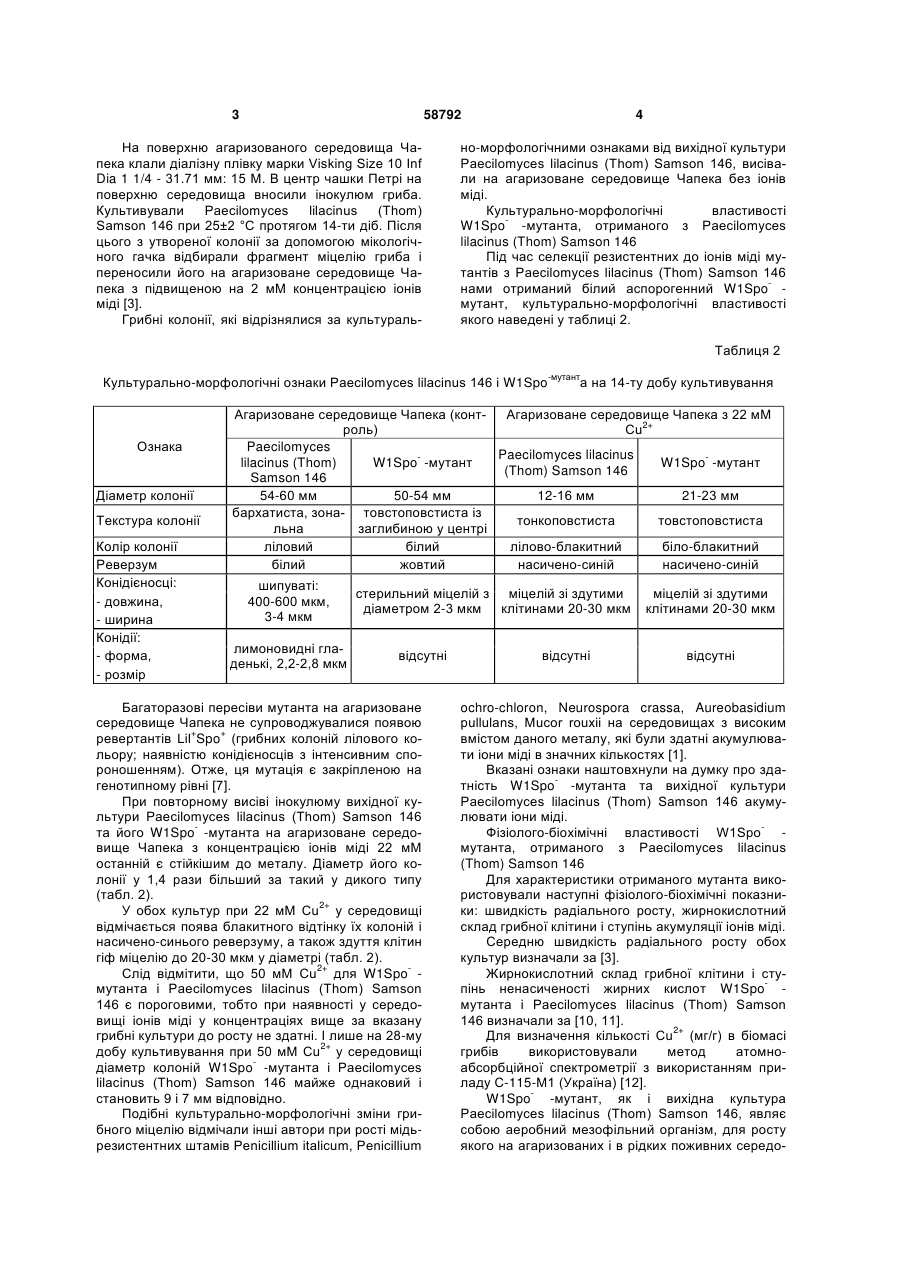
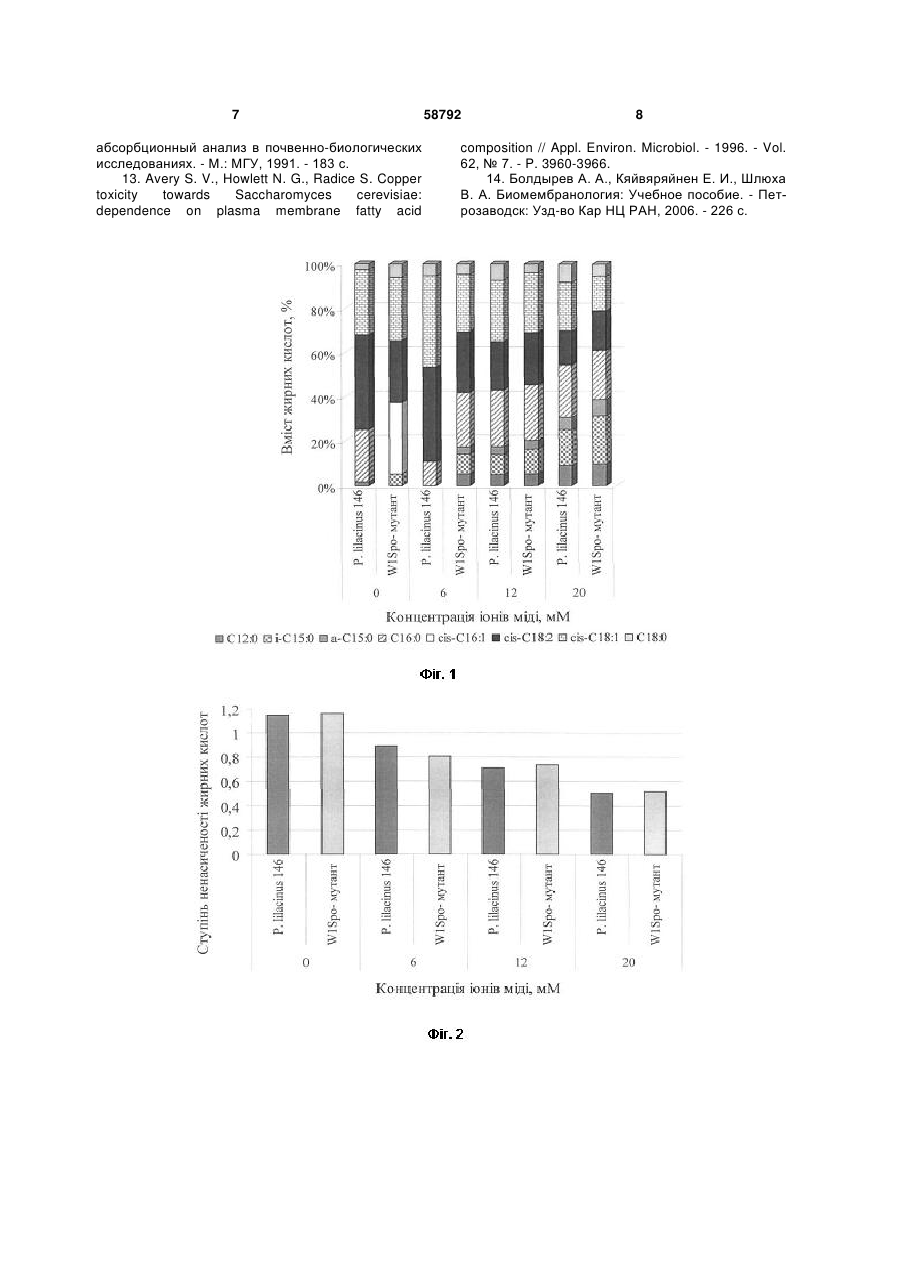
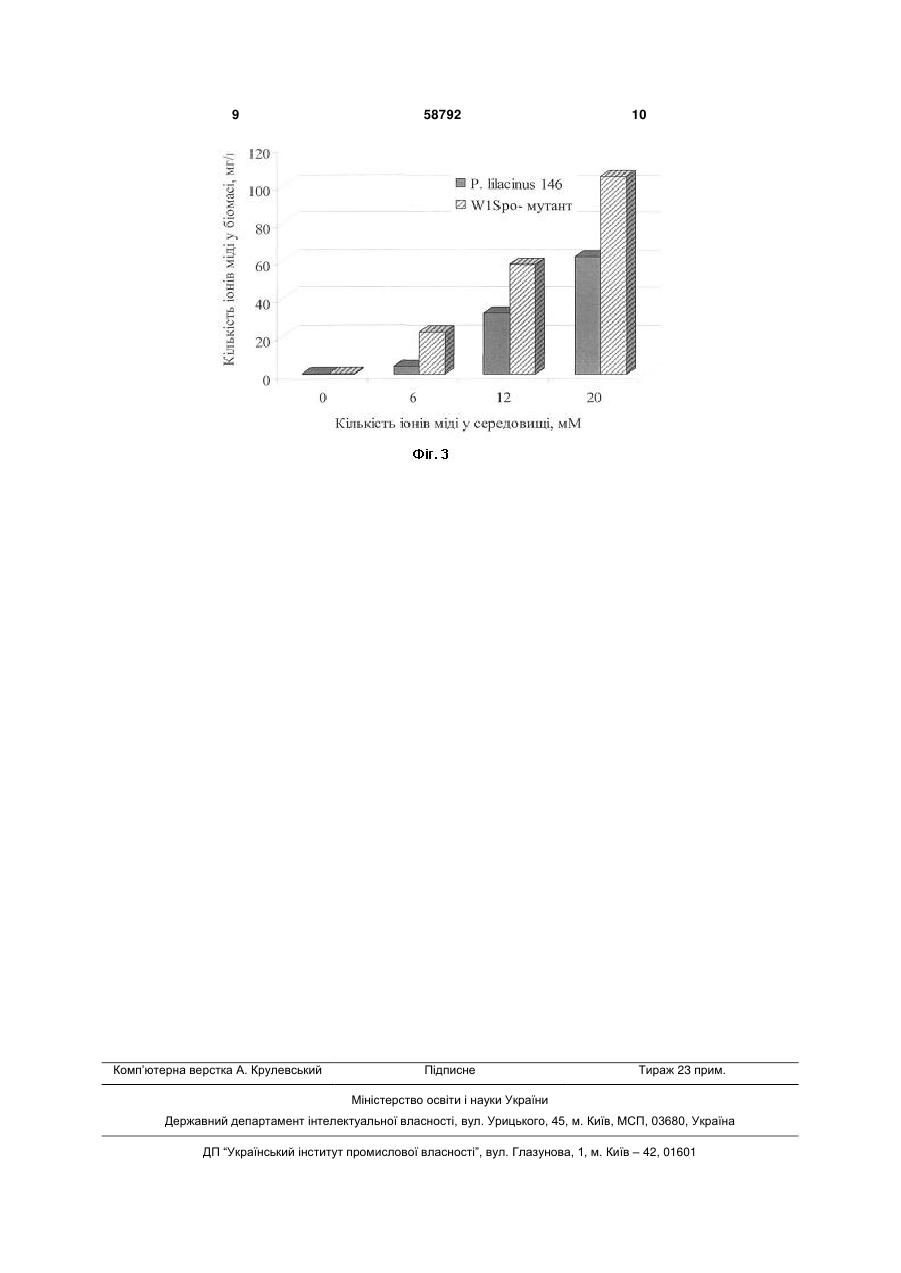
Білий аспорогенний W1Spo--мутант, резистентний до іонів міді, отриманий з Paecilomyces lilacinus (Thom) Samson 146, здатний накопичувати в своїй біомасі 104,88 мг/г іонів міді. Корисна модель стосується області біотехнології, зокрема акумуляції важких металів мікроскопічними грибами. Практично отриманий білий аспорогенний W1Spo-мутант може бути використаний для очистки стічних вод від іонів міді. В переважній більшості без втрати фізіологічної функції мікроскопічні гриби здатні рости в присутності 1-50 мкМ [1]. Найбільш близькими до запропонованого мутанта за стійкістю до іонів міді та ступенем їх акумуляції є штами грибів, наведені у таблиці 1. 62,2 1280 (20 мМ Сu2+) 104,8 Так, Aspergillus niger B-77 і Cryptococcus sp. WT здатні витримувати у 6,6 і 2,5 рази відповідно меншу концентрацію іонів міді у середовищі (а також накопичувати їх у своїй біомасі) у порівнянні з Paecilomyces lilacinus (Thom) Samson 146 і селекціонованим з нього W1Spo- -мутантом. Задача корисної моделі - одержання резистентного до іонів міді мутанта із Paecilomyces lilacinus (Thom) Samson 146, оскільки останній часто виділяється із забруднених на мідь ґрунтів [2]. Штам гриба Paecilomyces lilacinus (Thom) Samson 146 виділений методом ґрунтових розведень [3] із зразка чорноземного ґрунту мідних родовищ хутора Картамиш Луганської обл. у 2002 році. Кількість валових і рухомих форм міді у ґрунті [8] [9] перевищує гранично допустиму концентрацію в 28,5 і 147 разів відповідно [2, 4]. Ідентифікацію гриба до виду здійснено з використанням відповідних визначників [5, 6]. Даний штам гриба зберігається у колекції чистих культур відділу фізіології та систематики мікроміцетів Інституту мікробіології і вірусології ім. Д. К. Заболотного НАН України. Селекцію мутантів, резистентних до іонів міді, здійснювали наступним чином. Вихідну культуру гриба Paecilomyces lilacinus (Thom) Samson 146 (дикий тип) пересівали на агаризоване середовище Чапека поступово збільшуючи в ньому концентрацію іонів міді з 2 до 22 мМ Сu2+, збільшуючи концентрацію металу кожен раз на 2 мМ [7]. 58792 1280 (20 мМ Сu2+) Посилання (11) Концентрація іонів міді у біомасі, мг/г 24,2 51,9 UA Cryptococcus sp. WT Aspergillus niger B-77 Paecilomyces lilacinus (Thom) Samson 146 білий аспорогенний W1Spo- мутант Концентрація іонів міді в середовищі, мг/л 500 ( 8 mM Cu2+) 200 ( 3 mM Cu2+) (19) Назва гриба (13) Порівняльна характеристика штамів мікроскопічних грибів щодо їх стійкості та акумуляції іонів міді U Таблиця 1 3 58792 На поверхню агаризованого середовища Чапека клали діалізну плівку марки Visking Size 10 Inf Dia 1 1/4 - 31.71 мм: 15 M. В центр чашки Петрі на поверхню середовища вносили інокулюм гриба. Культивували Paecilomyces lilacinus (Thom) Samson 146 при 25±2 °С протягом 14-ти діб. Після цього з утвореної колонії за допомогою мікологічного гачка відбирали фрагмент міцелію гриба і переносили його на агаризоване середовище Чапека з підвищеною на 2 мМ концентрацією іонів міді [3]. Грибні колонії, які відрізнялися за культураль 4 но-морфологічними ознаками від вихідної культури Paecilomyces lilacinus (Thom) Samson 146, висівали на агаризоване середовище Чапека без іонів міді. Культурально-морфологічні властивості W1Spo- -мутанта, отриманого з Paecilomyces lilacinus (Thom) Samson 146 Під час селекції резистентних до іонів міді мутантів з Paecilomyces lilacinus (Thom) Samson 146 нами отриманий білий аспорогенний W1Spo- мутант, культурально-морфологічні властивості якого наведені у таблиці 2. Таблиця 2 Культурально-морфологічні ознаки Paecilomyces lilacinus 146 і W1Spo-мутанта на 14-ту добу культивування Ознака Діаметр колонії Текстура колонії Колір колонії Реверзум Конідієносці: - довжина, - ширина Конідії: - форма, - розмір Агаризоване середовище Чапека (конт- Агаризоване середовище Чапека з 22 мМ роль) Сu2+ Paecilomyces Paecilomyces lilacinus lilacinus (Thom) W1Spo- -мутант W1Spo- -мутант (Thom) Samson 146 Samson 146 54-60 мм 50-54 мм 12-16 мм 21-23 мм бархатиста, зона- товстоповстиста із тонкоповстиста товстоповстиста льна заглибиною у центрі ліловий білий лілово-блакитний біло-блакитний білий жовтий насичено-синій насичено-синій шипуваті: 400-600 мкм, 3-4 мкм лимоновидні гладенькі, 2,2-2,8 мкм стерильний міцелій з міцелій зі здутими діаметром 2-3 мкм клітинами 20-30 мкм відсутні Багаторазові пересіви мутанта на агаризоване середовище Чапека не супроводжувалися появою ревертантів Lil+Spo+ (грибних колоній лілового кольору; наявністю конідієносців з інтенсивним спороношенням). Отже, ця мутація є закріпленою на генотипному рівні [7]. При повторному висіві інокулюму вихідної культури Paecilomyces lilacinus (Thom) Samson 146 та його W1Spo- -мутанта на агаризоване середовище Чапека з концентрацією іонів міді 22 мМ останній є стійкішим до металу. Діаметр його колонії у 1,4 рази більший за такий у дикого типу (табл. 2). У обох культур при 22 мМ Сu2+ у середовищі відмічається поява блакитного відтінку їх колоній і насичено-синього реверзуму, а також здуття клітин гіф міцелію до 20-30 мкм у діаметрі (табл. 2). Слід відмітити, що 50 мМ Сu2+ для W1Spo- мутанта і Paecilomyces lilacinus (Thom) Samson 146 є пороговими, тобто при наявності у середовищі іонів міді у концентраціях вище за вказану грибні культури до росту не здатні. І лише на 28-му добу культивування при 50 мМ Сu2+ у середовищі діаметр колоній W1Spo- -мутанта і Paecilomyces lilacinus (Thom) Samson 146 майже однаковий і становить 9 і 7 мм відповідно. Подібні культурально-морфологічні зміни грибного міцелію відмічали інші автори при рості мідьрезистентних штамів Penicillium italicum, Penicillium відсутні міцелій зі здутими клітинами 20-30 мкм відсутні ochro-chloron, Neurospora crassa, Aureobasidium pullulans, Mucor rouxii на середовищах з високим вмістом даного металу, які були здатні акумулювати іони міді в значних кількостях [1]. Вказані ознаки наштовхнули на думку про здатність W1Spo- -мутанта та вихідної культури Paecilomyces lilacinus (Thom) Samson 146 акумулювати іони міді. Фізіолого-біохімічні властивості W1Spo- мутанта, отриманого з Paecilomyces lilacinus (Thom) Samson 146 Для характеристики отриманого мутанта використовували наступні фізіолого-біохімічні показники: швидкість радіального росту, жирнокислотний склад грибної клітини і ступінь акумуляції іонів міді. Середню швидкість радіального росту обох культур визначали за [3]. Жирнокислотний склад грибної клітини і ступінь ненасиченості жирних кислот W1Spo- мутанта і Paecilomyces lilacinus (Thom) Samson 146 визначали за [10, 11]. Для визначення кількості Сu2+ (мг/г) в біомасі грибів використовували метод атомноабсорбційної спектрометрії з використанням приладу С-115-М1 (Україна) [12]. W1Spo- -мутант, як і вихідна культура Paecilomyces lilacinus (Thom) Samson 146, являє собою аеробний мезофільний організм, для росту якого на агаризованих і в рідких поживних середо 5 вищах оптимальною температурою є 25±2 °С і рН 5,5. При культивуванні на агаризованому середовищі Чапека W1Spo- -мутанта і Paecilomyces lilacinus (Thom) Samson 146 середня швидкість радіального росту становить 0,083 і 0,056 мм/год відповідно. З додаванням 50 мМ Сu2+ у поживне середовище швидкість радіального росту обох культур знижується у 6 і 5 разів відповідно і становить 0,014 і 0,011 мм/год. Максимальною концентрацією іонів міді, за якої W1Spo- -мутант і Paecilomyces lilacinus (Thom) Samson 146 здатні накопичувати достатню для проведення досліджень кількість біомаси при рості на агаризованому середовищі Чапека, є 20 мМ Сu2+. Відомо, що іони важких металів посилюють перекисне окиснення ліпідів мембран, збільшуючи таким чином їх проникність, в результаті чого кількість іонів міді у грибних клітинах може підвищуватися [13, 14]. Якісний і кількісний склад жирних кислот в грибних клітинах W1Spo- -мутанта і Paecilomyces lilacinus (Thom) Samson 146 чітко відрізняється і залежить від концентрації іонів міді у середовищі (Фіг. 1). У обох культур значну кількість (15,0-43 %) становлять ненасичені цис-9,12-октадеценова (цис-С 18:2) і цис-9-октадеценова (цис-С 18:1) жирні кислоти. Лише при культивуванні W1Spo- мутанта на середовищі Чапека за відсутності іонів міді у клітинах гриба виявляється цисгексадеценова кислота у кількості 31,87 % і зовсім відсутня гексадеканова кислота (Фіг. 1). Дані щодо зниження ступеня ненасиченості жирних кислот у W1Spo- -мутанта і Paecilomyces lilacinus (Thom) Samson 146 при підвищенні іонів міді у середовищі є непрямим доказом перекисного окиснення ліпідів мембран грибів, яке виникає в результаті дії металу. Зокрема, при наявності 20 2+ мМ Сu у середовищі ступінь ненасиченості жирних кислот у W1Spo- -мутанта і Paecilomyces lilacinus (Thom) Samson 146 у 2 рази менший за контроль (Фіг. 2). Отриманий в результаті селекції W1Spo- мутант має вищу акумуляційну здатність до іонів міді. Так, при наявності у середовищі 20 мМ Сu2+ ступінь акумуляції металу W1Spo- -мутантом у 1,7 разів вищий, ніж у дикого типу Paecilomyces lilacinus (Thom) Samson 146 і становить 104,88 мг/г. Слід зазначити, що з підвищенням концентрації іонів міді у середовищі ступінь акумуляції обох культур поступово підвищується і завжди є вищим у W1Spo- -мутанта, ніж у Paecilomyces lilacinus (Thom) Samson 146 (Фіг. 3). Для довгострокового зберігання W1Spo- мутанта і Paecilomyces lilacinus (Thom) Samson 146 використовуються біологічні пробірки з Сусловим агаром, що має скошену поверхню, під ватномарлевими пробками з чарунковою покришкою. На 14-ту добу культивування грибів пробірки вміщують у холодильник при 4 °С. Пересівати культури грибів на свіже поживне середовище достатньо 2 рази на рік. При цьому обидві культури не втрачають свої культурально-морфологічні ознаки та 58792 6 фізіолого-біохімічні властивості. Наведені вище дані підтверджують суть корисної моделі, яка полягає в отриманні резистентного до іонів міді білого аспорогенного W1Spo- мутанта з Paecilomyces lilacinus (Thom) Samson 146. Основною перевагою W1Spo- -мутанта перед Paecilomyces lilacinus (Thom) Samson 146 є його вищий в 1,7 разів ступінь акумуляції іонів міді, ніж у дикого типу. Перелік фігур креслення Фіг. 1. Жирнокислотний склад W1Spo- -мутанта і Paecilomyces lilacinus (Thom) Samson 146 при рості на середовищі Чапека з різними концентраціями іонів міді Фіг. 2. Ступінь ненасиченості жирних кислот W1Spo- -мутанта і Paecilomyces lilacinus (Thom) Samson 146 при рості на середовищі Чапека з різними концентраціями іонів міді Фіг. 3. Кількість іонів міді в біомасі (мг/г) W1Spo- -мутанта і Paecilomyces lilacinus (Thom) Samson 146 при їх культивуванні на середовищі Чапека з різними концентраціями іонів міді Джерела інформації: 1. Cervantes С., Gutierrez-Corona F. Copper resistance mechanism in bacteria and fungi // FEMS Microbiol. Rev. - 1994. - 14, № 2. - P. 121-137. 2. Олишевская С. В., Маничев В. И., Захарченко В. А., Артышкова Л. В., Наконечная Л. Т., Жданова Н. Н. Влияние тяжелых металлов на микобиоту почв некоторых промышленных регионов Украины // Микол, и фитопатол. - 2006. - Т. 40, вып.2. - С. 133-141. 3. Методы экспериментальной микологии / Под ред. В. И. Билай - М.: Наук, думка, 1982. 432с. 4. Земельні ресурси України / Під ред. В. В. Медведева, Т. М. Лактіонової - К.: Агронаука, 1998. - 148 с. 5. Domsh K. H., Gams W. Compendium of soil fungi. V. 1. Lnd.etc: Acad. Press, 1980. - 839 p. 6. List of cultures. Fungi (filamentous fungi and yeasts). Bacteria. Plasmids.Phages. 35th edition. Baarn: Institute of the Royal Netherlandes Academy of Arts and Sciences, 2001. - 365 p. 7. Tomsett A. B. Genetics and molecular biology of metal tolerance in fungi// Stress tolerance of fungi / Ed. by D. H. Jennings. - England, University of Liverpool, 1995. - P. 97-125. 8. Лозова О. Г. Дріжджі - сорбенти деяких іонів важких металів: Автореф. дис. ... канд. біол. наук. Київ, 2004. - 20 с 9. Tsekova К., Todorova D. Copper accumulation and superoxide dismutase activity during growth of Aspergillus niger B-77 // Z. Naturforsch. - 2002. - 57. P. 319-322. 10. Brian B. L., Gardner E. W. Preparation of bacterial fatty acid methyl esters for rapid characterization by gas-liquid chromatography // Appl. Microbiol. - 1967. - 15, № 6. - P. 1499-1500. 11. Феофилова Е. П., Горнова И. Б., Меморская A. C., Гарибова Л. В. Липидный состав плодовых тел и глубинного мицелия Lentinus edodes (Berk.) sing [Lentinula edodes (Berk.) Pegler] // Микробиология. - 1998. - 67, № 5. - С. 655-659. 12. Обухов А. И., Плеханова И. О. Атомно 7 абсорбционный анализ в почвенно-биологических исследованиях. - М.: МГУ, 1991. - 183 с. 13. Avery S. V., Howlett N. G., Radice S. Copper toxicity towards Saccharomyces cerevisiae: dependence on plasma membrane fatty acid 58792 8 composition // Appl. Environ. Microbiol. - 1996. - Vol. 62, № 7. - P. 3960-3966. 14. Болдырев А. А., Кяйвяряйнен Е. И., Шлюха В. А. Биомембранология: Учебное пособие. - Петрозаводск: Узд-во Кар НЦ РАН, 2006. - 226 с. 9 Комп’ютерна верстка А. Крулевський 58792 Підписне 10 Тираж 23 прим. Міністерство освіти і науки України Державний департамент інтелектуальної власності, вул. Урицького, 45, м. Київ, МСП, 03680, Україна ДП “Український інститут промислової власності”, вул. Глазунова, 1, м. Київ – 42, 01601
ДивитисяДодаткова інформація
Назва патенту англійськоюWhite asporogenic w1spomutant, cuprum-resistant, obtained from paecilomyces lilacinus (thom) samson 146
Автори англійськоюOlishevska Snizhana Viktorivna
Назва патенту російськоюБелый аспорогенний w1spo--мутант, резистентный к ионам меди, полученный из paecilomyces lilacinus (thom) samson 146
Автори російськоюОлишевская Снежана Викторовна
МПК / Мітки
Мітки: w1spo--мутант, іонів, lilacinus, samson, білий, отриманий, резистентний, paecilomyces, аспорогенний, міді, thom
Код посилання
<a href="https://ua.patents.su/5-58792-bilijj-asporogennijj-w1spo-mutant-rezistentnijj-do-ioniv-midi-otrimanijj-z-paecilomyces-lilacinus-thom-samson-146.html" target="_blank" rel="follow" title="База патентів України">Білий аспорогенний w1spo–мутант, резистентний до іонів міді, отриманий з paecilomyces lilacinus (thom) samson 146</a>
Спосіб очистки стічних вод від іонів міді мікроорганізмами

Номер патенту: 55717
Опубліковано: 27.12.2010
Автори: Мороз Оксана Михайлівна, Гнатуш Світлана Олексіївна, Перетятко Тарас Богданович, Гудзь Степан Петрович
МПК: C12N 1/20, C02F 11/00, C02F 3/34
Мітки: мікроорганізмами, стічних, вод, іонів, очистки, спосіб, міді
Формула / Реферат:
Спосіб очистки стічних вод від іонів міді мікроорганізмами, що містить обробку сорбентом, який відрізняється тим, що як сорбент використовують асоціацію бактерій Desulfovibrio desulfuricans Ya-11 і Psedomonas sp. (7:1), що вносять у відстійник-очисник при температурі 25-28 °С і витримують 14±2 доби за анаеробних умов.
Пристрій для електрохімічного вилучення іонів купруму (іі) з відпрацьованих розчинів травлення міді

Номер патенту: 11856
Опубліковано: 16.01.2006
Автори: Храб Ольга Валеріївна, Тевтуль Ярема Юрійович
МПК: C25D 17/00
Мітки: міді, купруму, іонів, травлення, відпрацьованих, пристрій, іі, розчинів, електрохімічного, вилучення
Формула / Реферат:
Пристрій для електрохімічного вилучення іонів купруму (ІІ) з відпрацьованих розчинів травлення міді, який включає ванну, розділену іонообмінною мембраною, при цьому одна частина ванни оснащена катодною штангою з катодом, а друга частина оснащена анодною штангою та нерозчинним анодом, який відрізняється тим, що він додатково містить встановлений під катодом піддон з неелектропровідного, кислотостійкого матеріалу, внутрішня частина якого...
Спосіб очищення стічних вод від іонів міді та шестивалентного хрому за допомогою дріжджів

Номер патенту: 85485
Опубліковано: 26.01.2009
Автори: Гойко Ірина Юріївна, Лізунов В'ячеслав В'ячеславович, Дем'яненко Ірина Володимирівна, Горобець Світлана Василівна
МПК: C02F 1/46
Мітки: дріжджів, стічних, шестивалентного, очищення, допомогою, вод, спосіб, хрому, іонів, міді
Формула / Реферат:
Спосіб очищення стічних вод від іонів міді та шестивалентного хрому за допомогою дріжджів Saccharomyces cerevisiae та внесення в розчин металевої насадки в зовнішньому постійному магнітному полі, який відрізняється тим, що для видалення дріжджів, які сорбують іони вказаних металів, використовують частки наномагнетиту, оптимальна кількість яких знаходиться в межах співвідношення дріжджів до магнетиту від 5:1 до 30:1.
Спосіб очищення стічних вод від іонів міді

Номер патенту: 10651
Опубліковано: 15.11.2005
Автори: Горобець Оксана Юріївна, Мазур Сергій Петрович, Гойко Ірина Юріївна, Горобець Світлана Василівна
МПК: C02F 1/28
Мітки: спосіб, очищення, стічних, вод, іонів, міді
Формула / Реферат:
Спосіб очищення стічних вод від іонів міді, що включає обробку сорбентом, за який використовують дріжджі Saccharomyces cerevisiae, за допомогою внесення в розчин металевої насадки та прикладення зовнішнього постійного магнітного поля, який відрізняється тим, що після очищення розчину від іонів міді проводять очищення від іонів заліза, які потрапили в розчин з металевої насадки, та отримання кристалічного осаду з магнітними властивостями -...
Склад мембрани іоноселективного електрода для визначення активності іонів міді (іі)

Номер патенту: 12567
Опубліковано: 15.02.2006
Автори: Щербаков Сергій Васильович, Тітова Наталія Юріївна
МПК: G01N 27/30
Мітки: електрода, активності, мембрани, іі, визначення, склад, іонів, іоноселективного, міді
Формула / Реферат:
Склад мембрани іоноселективного електрода для визначення активності міді (II), що містить криптанд, тетрафенілборат лужного металу, полівінілхлорид та дидецилфталат, який відрізняється тим, що як криптанд він містить 1, 3, 12, 14-тетрааза-6, 9, 17, 20, 25- пентаоксабіцикло [12, 8, 5] гептакозан-2, 13-дитіон, а як тетрафенілборат лужного металу - тетрафенілборат натрію при наступному співвідношенні вказаних компонентів, ваг. %: ...
Попередній патент: Спосіб формування анастомозу при тонкокишковій непрохідності
Наступний патент: Пристрій для очищення внутрішньої поверхні труб
Випадковий патент: Похідні 2-(1,2,3-триазол-2-іл)бензаміду та 3-(1,2,3-триазол-2-іл)піколінаміду як антагоністи рецептора орексину