Ізольоване антитіло або його фрагмент, здатне до зв’язування з людським nogo
Номер патенту: 96279
Опубліковано: 25.10.2011
Автори: Прінйоха Рабіндер Кумар, Елліс Джонатан Генрі, Коспідас Ґеорґ, Макадам Рус, Кліґґ Стефанія Джейн, Амблі Пол Ендрю, ермащевскі Волкер






Формула / Реферат
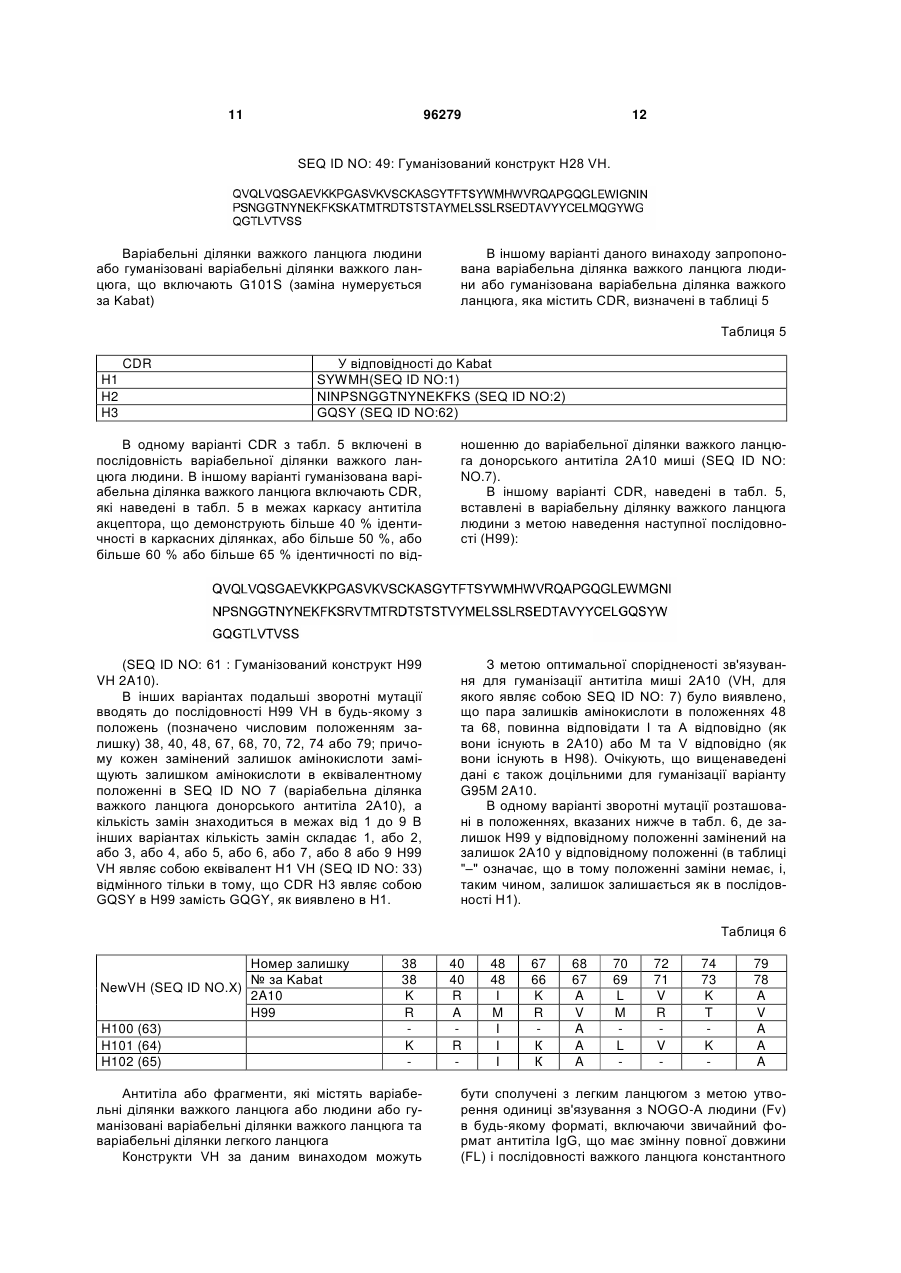
1. Ізольоване антитіло або його фрагмент, здатне до зв'язування з людським NOGO-A, що включає варіабельну ділянку важкого ланцюга, яка має амінокислотну послідовність, представлену у SEQ ID NO:49, та варіабельну ділянку легкого ланцюга, яка має амінокислотну послідовність, представлену у SEQ ID NO:14.
2. Ізольоване антитіло згідно з пунктом 1, що включає важкий ланцюг, який має амінокислотну послідовність, представлену у SEQ ID NO:55, та легкий ланцюг, який має амінокислотну послідовність, представлену у SEQ ID NO:18.
3. Експресійний вектор, який включає полінуклеотид, що кодує послідовності антитіла згідно з пунктом 1 або 2.
4. Клітина-хазяїн , що включає експресійний вектор згідно з пунктом 3.
5. Клітина-хазяїн згідно з пунктом 4, де клітина включає перший вектор, що кодує легкий ланцюг, та другий вектор, що кодує важкий ланцюг.
6. Спосіб одержання антитіла, здатного до зв'язування з людським NOGO-A, де спосіб включає етапи: трансфекції клітини-хазяїна за допомогою експресійного вектора, що включає перший полінуклеотид, який кодує варіабельну ділянку важкого ланцюга, що має амінокислотну послідовність, представлену у SEQ ID NO:49, та експресійного вектора, що включає другий полінуклеотид, який кодує варіабельну ділянку легкого ланцюга, що має амінокислотну послідовність, представлену у SEQ ID NO:14, та культивування клітини-хазяїна за умов, прийнятних для секреції антитіла з вказаної клітини-хазяїна у вказане культуральне середовище.
7. Спосіб згідно з пунктом 6, що додатково включає етап відновлення секретованого антитіла з культурального середовища.
8. Спосіб згідно з пунктом 6 або 7, де перший та другий полінуклеотиди містяться в одному експресійному векторі.
9. Фармацевтична композиція, що включає анти-NOGO антитіло або його фрагмент згідно з пунктом 1 або 2 разом з фармацевтично прийнятним розріджувачем або носієм.
10. Застосування анти-NOGO aнтитіла або його фрагмента згідно з пунктом 1 або 2 у приготуванні лікарського засобу для лікування або профілактики інсульту та інших неврологічних захворювань/розладів або для лікування пацієнта, що страждає від механічної травми центральної або периферичної нервової системи.
11. Анти-NOGO антитіло згідно з пунктом 1 або 2 для застосування у лікуванні або профілактиці інсульту та інших неврологічних захворювань/розладів або для лікування пацієнта, що страждає від механічної травми центральної або периферичної нервової системи.
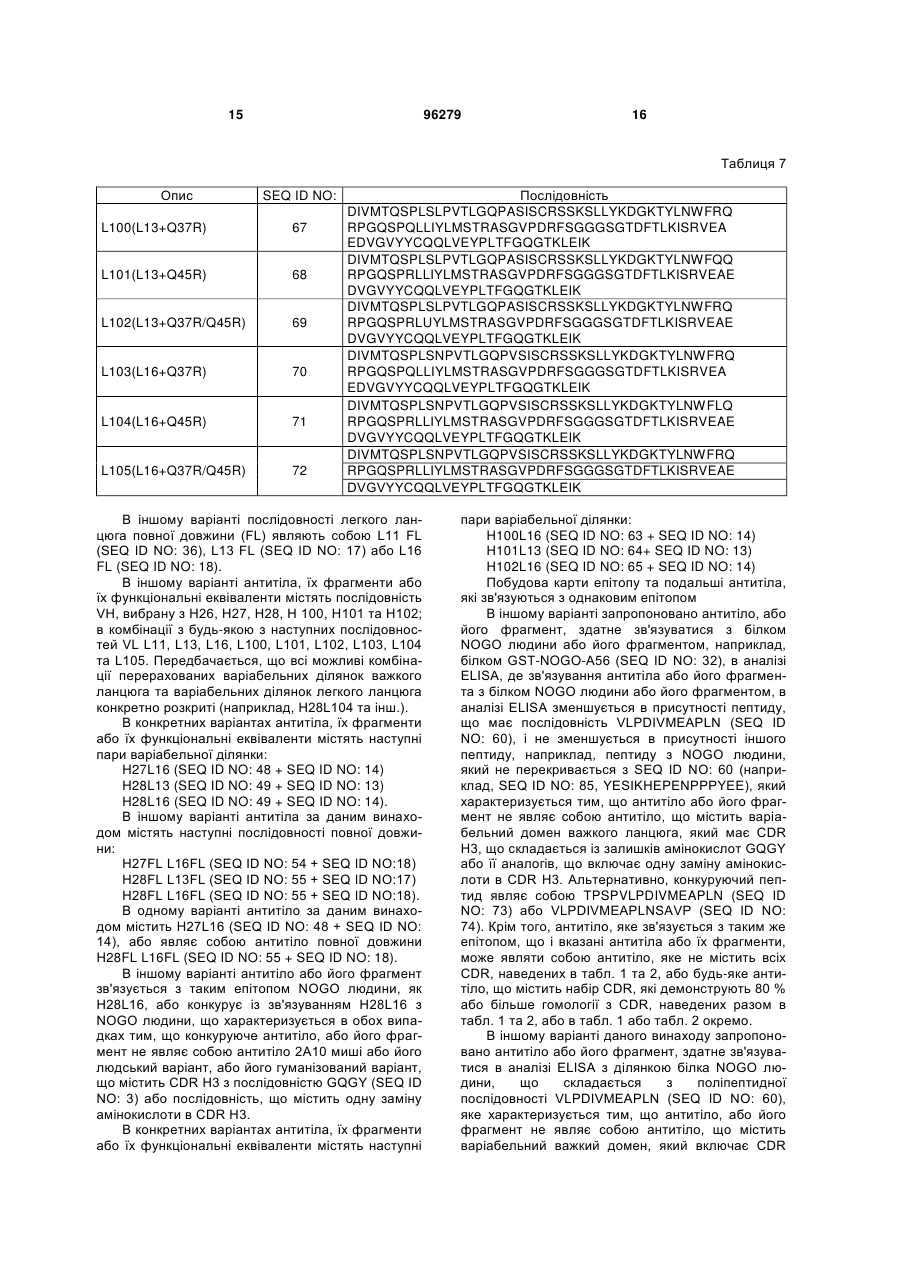
Текст
1. Ізольоване антитіло або його фрагмент, здатне до зв'язування з людським NOGO-A, що включає варіабельну ділянку важкого ланцюга, яка має амінокислотну послідовність, представлену у SEQ ID NO:49, та варіабельну ділянку легкого ланцюга, яка має амінокислотну послідовність, представлену у SEQ ID NO:14. 2. Ізольоване антитіло згідно з пунктом 1, що включає важкий ланцюг, який має амінокислотну послідовність, представлену у SEQ ID NO:55, та легкий ланцюг, який має амінокислотну послідовність, представлену у SEQ ID NO:18. 3. Експресійний вектор, який включає полінуклеотид, що кодує послідовності антитіла згідно з пунктом 1 або 2. 4. Клітина-хазяїн , що включає експресійний вектор згідно з пунктом 3. 5. Клітина-хазяїн згідно з пунктом 4, де клітина включає перший вектор, що кодує легкий ланцюг, та другий вектор, що кодує важкий ланцюг. 6. Спосіб одержання антитіла, здатного до зв'язування з людським NOGO-A, де спосіб включає етапи: трансфекції клітини-хазяїна за допомогою експресійного вектора, що включає перший полінуклеотид, який кодує варіабельну ділянку важкого ланцюга, що має амінокислотну послідовність, представлену у SEQ ID NO:49, та експресійного вектора, що включає другий полінуклеотид, який кодує варіабельну ділянку легкого ланцюга, що має амінокислотну послідовність, представлену у SEQ ID NO:14, та культивування клітини-хазяїна за умов, прийнятних для секреції антитіла з вказаної клітини-хазяїна у вказане культуральне середовище. 7. Спосіб згідно з пунктом 6, що додатково включає етап відновлення секретованого антитіла з культурального середовища. UA (21) a200807663 (22) 14.12.2006 (24) 25.10.2011 (86) PCT/EP2006/069737, 14.12.2006 (31) 0525662.3 (32) 16.12.2005 (33) GB (46) 25.10.2011, Бюл.№ 20, 2011 р. (72) КЛІҐҐ СТЕФАНІЯ ДЖЕЙН, GB, ЕЛЛІС ДЖОНАТАН ГЕНРІ, GB, ҐЕРМАЩЕВСКІ ВОЛКЕР, GB, АМБЛІ ПОЛ ЕНДРЮ, GB, КОСПІДАС ҐЕОРҐ, AU, МАКАДАМ РУС, GB, ПРІНЙОХА РАБІНДЕР КУМАР, GB (73) ҐЛАКСО ҐРУП ЛІМІТЕД, GB (56) WO 2005061544 A, 07.07.2005. US 2005215770 A1, 29.09.2005. BANDTLOW CHRISTINE ET AL: "The Escherichia coli-derived Fab fragment of the IgM/kappa antibody IN-1 recognizes and neutralizes myelin-associated inhibitors of neurite growth" EUROPEAN JOURNAL OF BIOCHEMISTRY, BERLIN, DE, vol. 241, no. 2, 1996, pages 468-475, XP000991434 ISSN: 00142956. PAPADOPOULOS C M ET AL: "FUNCTIONAL RECOVERY ANF NEUROANATOMICAL PLASTICITY FOLLOWING MIDDLE CEREBRAL ARTERY OCCLUSION AND IN-1 ANTIBODY TREATMENT IN THE ADULT RAT" ANNALS OF NEUROLOGY, BOSTON, US, vol. 51, no. 4, April 2002 (2002-04), pages 433-441, XP008034394 ISSN: 0364-5134. BROESAMLE C ET AL: "Regeneration of lesioned corticospinal tract fibers in the adult rat induced by a recombinant, humanized IN-1 antibody fragment" JOURNAL OF NEUROSCIENCE, NEW YORK, NY, US, vol. 20, no. 21, 1 November 2000 (2000-11-01), pages 8061-8068, XP002238449 ISSN: 0270-6474. UA 2002086629 A, 15.10.2002. UA 200602444 A, 15.06.2006. UA 200502127 A, 15.06.2005. 2 (19) 1 3 96279 4 8. Спосіб згідно з пунктом 6 або 7, де перший та другий полінуклеотиди містяться в одному експресійному векторі. 9. Фармацевтична композиція, що включає антиNOGO антитіло або його фрагмент згідно з пунктом 1 або 2 разом з фармацевтично прийнятним розріджувачем або носієм. 10. Застосування анти-NOGO aнтитіла або його фрагмента згідно з пунктом 1 або 2 у приготуванні лікарського засобу для лікування або профілакти ки інсульту та інших неврологічних захворювань/розладів або для лікування пацієнта, що страждає від механічної травми центральної або периферичної нервової системи. 11. Анти-NOGO антитіло згідно з пунктом 1 або 2 для застосування у лікуванні або профілактиці інсульту та інших неврологічних захворювань/розладів або для лікування пацієнта, що страждає від механічної травми центральної або периферичної нервової системи. ГАЛУЗЬ ТЕХНІКИ Даний винахід стосується імуноглобулінів, особливо антитіл, які зв'язуються з NOGO та нейтралізують його активність, полінуклеотидів, що кодують такі антитіла, фармацевтичних рецептур, що містять вказані антитіла, і застосування даних антитіл в лікуванні та/або профілактиці неврологічних захворювань. Інші аспекти, об'єкти і переваги даного винаходу стануть очевидними з опису, наведеного нижче. РІВЕНЬ ТЕХНІКИ Інсульт являє собою провідну причину смертності та інвалідності на Заході. На даний час не існує прийнятної терапії для лікування інсульту, крім тканинного плазміногену (t-PA), який необхідно вводити протягом 3 годин з початку нападу з наступним скануванням за допомогою комп'ютерної томографії (СТ) з метою виключення кровотечі. На сьогоднішній день більшість терапевтичних засобів, спрямованих на лікування гострого інсульту (тобто нейропротекцію), в основному включають цільові рецептори глютамату і, відомо, що їх сигнальні шляхи нижнього потоку залучені до гострої загибелі клітин. Однак, на сьогоднішній день дані стратегії виявилися невдалими в клінічних випробуваннях і часто супроводжуються побічними ефектами, що обмежують дозу (Hill & Hachinski, The Lancet, 352: (suppl II) 10-14 (1998)). Таким чином, існує потреба в розробці нових підходів, спрямованих на зменшення інтенсивності загибелі клітин після припинення кровопостачання. Нейропротекція полягає у здатності терапії запобігти або зменшити втрату нейронних клітин у відповідь на інсульт або захворювання. Цього можна досягти спрямуванням терапії на нейрони безпосередньо або непрямо, запобігаючи втраті гліальних клітин (включаючи олігодендроцит). Після початку інсульту спостерігають деяку ступінь мимовільного функціонального відновлення у багатьох пацієнтів, припускаючи, що мозок має (хоча і обмежену) здатність до відновлення та/або реконструювання після пошкодження. Засоби з потенціалом підсилення такого відновлення, можуть, таким чином, здійснювати свій вплив набагато пізніше (потенційно, через дні) після початку церебральної ішемії. Засоби, які можна запропонувати як для екстреного нейрозахисту, так і для підсилення функціонального відновлення, можуть забезпечити значні переваги над існуючими потенційними стратегіями нейрозахисту. Хвороба Альцгеймера (AD) характеризується наявністю двох діагностичних особливостей пато логії. Дана патологія являє собою амілоїдні бляшки та нейрофібрилярні сплетення, що складаються з агрегованого β-амілоїдного пептиду (Ар40 та А(342)) і гіперфосфорильованої т, відповідно (Dawbarn & Allen 2001 Neurobiology of Alzheimer's Disease OUP). Всебічне вивчення показало виражений зв'язок у пацієнтів між акумуляцією β-амілоїда та послаблення когнітивних процесів (Naslund et al., JAMA, March 22/29, 2000, Vol. 283, No: 12, page 1571-1577). Даний факт узгоджується з генетичними і епідеміологічними дослідженнями, які свідчать, що деякі мутації в генах білку амілоїдного прекурсора і генах пресеніліну можуть провокувати ранній розвиток AD, мутації яких також збільшують рівні пептидів Aβ40 і Aβ42, включаючи їх співвідношення. Розщеплення трансмембранного білку амілоїдного прекурсора (АРР) типу І двома різними протеазами, позначеними як бета- і гама-секретаза, є необхідним для утворення β-амілоїдного пептиду. Підтверджена молекулярна ідентичність бетасекретази як аспартил-протеази Asp2/BACE1 (Hussain et al. Mol.Cell.NeuroSci. 16, 609-619 (2000); Vassar et al., Science (1999), Oct.22; 286 (5440):735-741). Природа гама-секретази залишається джерелом дебатів і, ймовірно, що вона складається з комплексу з високомолекулярною масою, що містить як мінімум наступні білки: пресеніліни, Aph1, Реn2 і нікастрин (розглянутий в Medina & Dotti Cell Signalling 2003 15(9):829-41). Обробка АРР в ЦНС, ймовірно, відбувається в межах ряду типів клітин, включаючи нейрони, олігодендроцити, астроцити та мікроглію. На загальну швидкість обробки АРР в даних клітинах буде впливати відносний рівень експресії АРР, BACE1/Asp2, пресеніліну-1 та -2, Aph1, Pen2 і нікастрину. До того ж, додаткові фактори, що регулюють субклітинне розташування АРР, можуть також впливати на його обробку, як показано виявленням того факту, що мутація мотиву YENP в цитоплазматичному домені АРР, який блокує його ендоцитоз, зменшує продукування β-амілоїду (Perez et al. 1999 J Biol Chem 274 (27) 18851-6). Утримання АРР-бета-CTF в ER за допомогою додавання мотиву утримання KKQN є достатнім для зменшення продукування ß-амілоїду в трансфікованих клітинах (Maltese et al. 2001 J Biol Chem 276 (23) 20267-20279). І навпаки, підвищення ендоцитозу надмірною експресією Rab5 є достатнім для збільшення секреції амілоїду трансфікованими кліти 5 нами (Grbovic et al. 2003 J Biol Chem 278 (33) 31261-31268). Узгоджені з одержаними даними подальші дослідження показали, що зменшення клітинних рівнів холестерину (добре відомий фактор ризику AD) зменшує утворення β-амілоїду. Дана зміна була залежною від зміненого ендоцитозу, як продемонстровано застосуванням домінуючих мутантів з відсутністю динаміну (K44А) і надмірною експресією білку RN-Tre, що активізує Rab5 GТРазу (Ehehalt et al. 2003 J Cell Bid 160 (1)113-123). Багаті на холестерин мікродомени або маси також являють собою важливу клітинну ділянку продукування β-амілоїду і АРР, показано, що ВАСЕ1 та компоненти комплексу гама-секретази тимчасово знаходяться в межах мас. Перехресне зв'язування АРР і ВАСЕ1 антитіл у напрямку мас, багатих на холестерин, могло збільшувати продукування β-амілоїду (Ehehalt et al. 2003 J Cell Biol 160 (1) 113-123). Експресія GPI-якірного BACE1, який виключно спрямований на ліпідні маси, і може аналогічно збільшувати розщеплення АРР та продукування β-амілоїду (Cordy et al. 2003 PNAS 100(20) 11735-11740). На цей час невідомі механізми, що лежать в основі функціонального відновлення після інсульту або іншого нейропошкоджуючого явища або захворювання. Відростки пошкоджених або непошкоджених аксонів запропоновано як один з можливих механізмів. Однак, хоча дослідження in vivo продемонстрували, що лікування пошкодження спинного мозку або інсульту нейротрофічними факторами приводить до підсилення функціонального відновлення і збільшення відростків аксону, вказані факти не доводять прямого зв'язку між збільшенням відростків аксону та ступенем функціонального відновлення (Jakeman, et al. 1998, Exp. Neurol. 154: 170-184, Kawamata et al. 1997, Proc Natl Acad. Sсi. USA., 94:8179-8184, Ribotta, et al. 2000, J Neurosci. 20: 5144-5152). До того ж, відростки аксону вимагають життєздатного нейрона. При таких захворюваннях, як інсульт, що супроводжуються обширною загибеллю клітин, підсилення функціонального відновлення, запропонованого даним засобом, після інсульту, може, таким чином, відбуватися через інші механізми, ніж відростки аксону, наприклад, через диференціацію ендогенних стовбурових клітин, активацію відновлювальних шляхів, зміни в розповсюдженні рецептору або збудливості нейронів або глії (Fawcett & Asher, 1999, Brain Res. Bulletin, 49: 377-391, Homer & Gage, 2000, Nature 407 963-970). Припускають, що обмежена здатність центральної нервової системи (ЦНС) до відновлення після пошкодження залежить частково від молекул в середовищі ЦНС, що здійснюють інгібувальний вплив на відростки аксону (нейритний відросток). Вважають, що мієлін ЦНС містить інгібуючі молекули (Schwab ME та Caroni Ρ (1988) J. Neurosci. 8, 2381-2193). Два білки мієліну, глікопротеїн, що асоціюється з мієліном (MAG), і NOGO, клонували та ідентифікували як передбачувані інгібітори нейритного відростка (Sato S. et al. (1989) Biochem. Biophys. Res. Comm. 163,1473-1480; McKerracher L et al. (1994) Neuron 13, 805-811; Mukhopadhyay G 96279 6 et al. (1994) Neuron 13, 757-767; Torigoe К and Lundborg G (1997) Exp. Neurology 150, 254-262; Schaferetal (1996) Neuron 16, 1107-1113; W09522344; WO9701352; Prinjha R et a.l (2000) Nature 403, 383-384; Chen MS et al. (2000) Nature 403, 434-439; GrandPre Τ et al. (2000) Nature 403, 439-444; US005250414A; WO200005364A1; WO0031235). Ідентифіковані три форми NOGO людини: NOGO-A, що містить залишки 1192 амінокислот (номер доступу GenBank AJ251383); NOGO-B, зрощений варіант, в якому відсутні залишки 186– 1004 в передбачуваному позаклітинному домені (номер доступу GenBank AJ251384) і коротший зрощений варіант, NOGO-C, в якому також відсутні залишки 186–1004, а також менший, альтернативний аміно-кінцевий домен (номер доступу GenBank AJ251385) (Prinjha et al (2000), див. вище). Інгібування білків, що інгібують ЦНС, наприклад NOGO, може дати терапевтичний засіб для зменшення пошкодження нейронів та забезпечити відновлення і ріст нейронів, потенційно сприяючи одужанню від пошкодження нейронів, наприклад пошкодження, що відбувається при інсульті. Прикладами таких інгібіторів NOGO можуть бути низькомолекулярні пептиди і антитіла. Існують повідомлення, що мишаче моноклональне антитіло, IN-1, що зростало проти NI220/250, мієліновий білок, що являє собою потужний інгібітор росту нейриту (і згодом показано, що він є фрагментом NOGO-A), приводить до регенерації аксону (Caroni, P and Schwab, ME (1988) Neuron 1 85-96; Schnell, L and Schwab, ME (1990) Nature 343 269-272; Bregman, BS et al. (1995) Nature 378 498-501 and Thallmair, M et al. (1998) Nature Neuroscience 1 124-131). Також повідомляють, що антиген NOGO-A являє собою антиген для IN-1 (Chen et al. (2000) Nature 403 434-439). Введення фрагменту IN-1 Fab або гуманізованого IN-1 щурам, яким робили поперечний розтин спинного мозку, підсилює одужання (Fiedler, M et al. (2002) Protein Eng 15 931-941; Brosamie, С et al. (2000) J. Neuroscience 20 8061-8068). Моноклональні антитіла, що зв'язуються з NOGO, описані у WO 04/052932 та WO2005028508. У WO 04/052932 розглядається мишаче антитіло 11C7, яке зв'язується з певними формами NOGO людини з високою спорідненістю. У патентній заявці WO05/061544 також описані моноклональні антитіла з високою спорідненістю, включаючи мишаче моноклональне антитіло 2A10, і загалом розкриті його гуманізовані варіанти, наприклад, H1L11 (послідовності H1 та L11 наведені в SEQ ID NO: 33 і 34, відповідно (тільки послідовності VH або VL)). Розкриті антитіла зв'язуються з NOGO-A людини з високою спорідненістю. Мишаче антитіло 2A10 (і його CDR-трансплантовані гуманізовані варіанти) характеризується наступними послідовностями ділянок визначення комплементарності (CDR) (що визначено із застосуванням методології Kabat (Kabat et al. (1991) "Sequences of proteins of immunological interest"; Fifth Edition; US Department of Health and Human Services; NIH publication No 91-3242)) в межах варіабельних ділянок легкого та важкого ланцюга: 7 96279 Таблиця 1 CDR легкого ланцюга антитіла 2A10 CDR L1 L2 L3 Послідовність RSSKSLLYKDGKTYLN (SEQ ID NО:4) LMSTRAS (SEQ ID NО:5) QQLVEYPLT (SEQ ID NО:6) Таблиця 2 CDR важкого ланцюга антитіла 2A10 CDR H1 H2 H3 Послідовність SYWMH(SEQ ID NO:1) NINPSNGGTNYNEKFKS (SEQ ID NО:2) GQGY (SEQ ID NО:3) Крім того, у WO05/061544 розкриті "аналоги" антитіл, які включають CDR, наведені в табл. 1 та 2 вище, такі "аналоги" мають однакову специфічність зв'язування з антигеном та/або таку здатність до нейтралізації, як антитіло-донор, з якого вони були одержані. Незважаючи на то, що в даній галузі відомі анти-NOGO антитіла з високим ступенем спорідненості, бажаною метою все ще залишається виділити та розробити альтернативу, або удосконалити терапевтично придатні моноклональні антитіла, які зв'язуються з та інгібують активність NOGO людини. Процес нейродегенерації лежить в основі багатьох неврологічних захворювань/розладів, в тому числі, не обмежуючись ними, гострі захворювання, наприклад, інсульт (ішемічний або геморагічний), травматичне пошкодження мозку та пошкодження спинного мозку, а також хронічні захворювання, включаючи хворобу Альцгеймера, лобно-скроневі недоумства (тауопатії), периферичну невропатію, хворобу Паркінсона, хворобу Крейтцфельда-Якоба (CJD - спастичний псевдосклероз), шизофренію, аміотрофний латеральний склероз (ALS - хвороба Шарко), розсіяний склероз, хворобу Гантінгтона (Huntington) та міозит з тільцями включення. Таким чином, моноклональні анти-NOGO антитіла та подібні до них за даним винаходом можуть бути придатними для лікування вказаних захворювань/розладів. В даному винаході запропоновані та детально описані нижче антитіла для лікування вищезгаданих захворювань/розладів. КОРОТКИЙ ОПИС ВИНАХОДУ У даному винаході пропонуються специфічні варіабельні ділянки важкого ланцюга та антитіла або їх фрагменти, що складаються з вказаних специфічних варіабельних ділянок важкого ланцюга і варіабельної ділянки легкого ланцюга, що дозволяє димеру Fv при сполученні з варіабельними ділянками важкого ланцюга зв'язуватися з NOGOA людини з високою спорідненістю, і, таким чином, нейтралізувати активність NOGO-A людини. Варіабельним ділянкам важкого ланцюга за даним винаходом можна надавати формату разом з варіабельними ділянками легкого ланцюга для 8 того, щоб дозволити зв'язування з NOGO-A людини у звичайний для імуноглобуліну спосіб (наприклад, людського IgG, IgA, IgM, і т п.) або в будьякому іншому його фрагменті або "подібному" до антитіла форматі, який зв'язується з NOGO-A людини (наприклад, одноланцюговий Fv, димерні антитіла, Tandabs™ та інш. (огляд альтернативних форматів "антитіла" див у Holliger and Hudson, Nature Biotechnology, 2005, Vol. 23, No 9, 11261136)). Варіабельна ділянка важкого ланцюга, яка включає третій CDR, що складається по суті із залишків GQGY амінокислот, де CDR містить як мінімум одну заміну в межах основної послідовності GQGY, вказані заміни вибрані з наступних замін: де G в першому положенні замінюють на R, І, W або M; Q в другому положенні замінюють на D, І, А, L, V або S; G в третьому положенні замінюють на W, N, Y, S, L або F; та Y в четвертому положенні замінюють на W В іншому варіанті тільки третій важкий ланцюг CDR (CDR H3) містить одну заміну з метою одержання наступних послідовностей CDR H3: RQGY (SEQ ID NO: NO.75),….. В іншому варіанті варіабельні ділянки важкого ланцюга, крім зазначених вище, додатково містять інші CDR, які наведені в таблиці 2, тобто CDR H1 (SEQ ID NO: 1) та CDR H2 (SEQ ID NO: 2). Антитіла за даним винаходом, або їх фрагменти зберігають активність зв'язування з NOGO людини, властиву антитілам, що містять CDR H3: GQGY, з точки зору їх активності, яку вимірювали в експериментах ELISA та Biacore, а в деяких випадках активність в даних експериментах збільшується. Варіабельні ділянки важкого ланцюга людини або гуманізовані варіабельні ділянки важкого ланцюга, що містять G95M (нумерація замін за Kabat) В одному варіанті даного винаходу варіабельні ділянки важкого ланцюга за даним винаходом містять CDR, визначені в таблиці 3 (як визначається за допомогою Kabat): Таблиця 3 CDR H1 H2 H3 Послідовність SYWMH (SEQ ID NО:1) NINPSNGGTNYNEKFKS (SEQ ID NО:2) MQGY (SEQ ID NО:45) В одному варіанті даного винаходу запропонована варіабельна ділянка важкого ланцюга людини або гуманізована варіабельна ділянка важкого ланцюга, що містить кожний з CDR, наведених в табл. 3. В іншому варіанті даного винаходу запропонована гуманізована варіабельна ділянка важ 9 96279 кого ланцюга, що містить CDR, наведені в табл. 3, в межах більшої послідовності варіабельної ділянки важкого ланцюга людини. Ще в одному варіанті гуманізована варіабельна ділянка важкого ланцюга містить CDR, наведені в табл. 3, в межах каркасу антитіла акцептора, що демонструє більше 40 % ідентичності в каркасних ділянках, або більше 50 %, або більше 60 %, або більше 65 % ідентичності по відношенню до варіабельної ділянки важкого ланцюга донорського антитіла 2A10 миші (SEQ ID NO: 7). Якщо CDR, наведені в табл. 3, застосовують цілком, в одному варіанті послідовність варіабельної ділянки важкого ланцюга являє собою послідовність H98, запропоновану як SEQ ID NO: 66 (H98 VH являє собою еквівалент H1 VH (SEQ ID NO: 33), яка відрізняється тільки в тому, що CDR H3 являє собою MQGY в H98, замість GQGY, як визначено в H1). В одному аспекті за даним винаходом антитіла містять варіабельну ділянку важкого ланцюга, що має послідовність амінокислот SEQ ID NO: 66 (варіабельна ділянка H98), що додатково містить цілий ряд замін в одному або більше положень 38, 40, 48, 67, 68, 70, 72, 74 та 79; причому кожен замінений залишок амінокислоти заміняють на залишок амінокислоти в еквівалентному положенні SEQ ID NO: 7 (варіабельна ділянка важкого ланцюга донорського антитіла 2A10), а кількість замін становить від 1 до 9 В інших варіантах кількість замін становить 1, або 2, або 3, або 4, або 5, або 6, або 7, або 8 або 9. В даному контексті заміни, які описані, є еквівалентними з точки зору "зворотних мутацій", де каркасні залишки амінокислот людини в конкретних положеннях в межах послідовності H98 видозмінюються зворотним чином по відношенню до 10 залишків амінокислоти в еквівалентному положенні в межах послідовності донорського антитіла 2A10. Якщо тільки конкретно не вказано інше, ніж заявлено в даному описі, коли числове положення залишку амінокислоти, знайденого в межах конкретної послідовності, згадано в даному документі, наприклад, "положення 12", то це означає, що кваліфікований фахівець повинен надати першій амінокислоті в послідовності положення номер "1" і рахує від положення один та ідентифікує амінокислоту, яка знаходиться в бажаному положенні, в даному прикладі залишок дванадцятої амінокислоти в послідовності Кваліфікований фахівець помітить, що дана система нумерації не відповідає системі нумерації Kabat, яку часто застосовують для визначення положення амінокислоти в межах послідовностей антитіла. З метою оптимальної спорідненості зв'язування для гуманізації антитіла миші 2A10 (VH, для якого являє собою SEQ ID NO: 7), було виявлено, що пара залишків амінокислот в положеннях 48 та 68, повинна становити І та А, відповідно (як вони існують в 2A10) або M та V відповідно (як вони існують в H98). Очікується, що вищезазначені дані є також доцільними для гуманізації варіанту G95M 2A10. В наступній таблиці наведено деталі трьох різних варіабельних ділянок важкого ланцюга (VH), які можуть утворювати частину антитіл за даним винаходом. Кожна з розкритих VH базується на H98 VH (SEQ ID NO: 66), що додатково містить заміни, згадані в таблиці (табл. 4), де залишок H98 у відповідному положенні замінений на залишок 2A10 в тому положенні ("–" в таблиці означає, що в тому положенні заміни немає, і, таким чином, залишок залишається як в послідовності H98): Таблиця 4 Номер залишку № за Kabat NewVH (SEQ ID NO. X) 2A10 H98 H26 (47) H27 (48) H28 (49) Таким чином, в одному варіанті даного винаходу варіабельні ділянки важкого ланцюга (VH) за даним винаходом являють собою H26 VH (SEQ ID NO: 47), H27 VH (SEQ ID NO: 48) та H28 VH (SEQ 38 38 K R K 40 40 R A R 48 48 І M І І І 67 66 K R K K 68 67 A V A A A 70 69 L M L 72 71 V R V 74 73 K T K 79 78 A V A A A ID NO: 49). SEQ ID NO: 47: Гуманізований конструкт H26 VH. SEQ ID 48: Гуманізований конструкт H27 VH. 11 96279 12 SEQ ID NO: 49: Гуманізований конструкт H28 VH. Варіабельні ділянки важкого ланцюга людини або гуманізовані варіабельні ділянки важкого ланцюга, що включають G101S (заміна нумерується за Kabat) В іншому варіанті даного винаходу запропонована варіабельна ділянка важкого ланцюга людини або гуманізована варіабельна ділянка важкого ланцюга, яка містить CDR, визначені в таблиці 5 Таблиця 5 CDR H1 H2 H3 У відповідності до Kabat SYWMH(SEQ ID NO:1) NINPSNGGTNYNEKFKS (SEQ ID NО:2) GQSY (SEQ ID NО:62) В одному варіанті CDR з табл. 5 включені в послідовність варіабельної ділянки важкого ланцюга людини. В іншому варіанті гуманізована варіабельна ділянка важкого ланцюга включають CDR, які наведені в табл. 5 в межах каркасу антитіла акцептора, що демонструють більше 40 % ідентичності в каркасних ділянках, або більше 50 %, або більше 60 % або більше 65 % ідентичності по від ношенню до варіабельної ділянки важкого ланцюга донорського антитіла 2A10 миші (SEQ ID NO: NO.7). В іншому варіанті CDR, наведені в табл. 5, вставлені в варіабельну ділянку важкого ланцюга людини з метою наведення наступної послідовності (H99): (SEQ ID NO: 61 : Гуманізований конструкт H99 VH 2A10). В інших варіантах подальші зворотні мутації вводять до послідовності H99 VH в будь-якому з положень (позначено числовим положенням залишку) 38, 40, 48, 67, 68, 70, 72, 74 або 79; причому кожен замінений залишок амінокислоти заміщують залишком амінокислоти в еквівалентному положенні в SEQ ID NO 7 (варіабельна ділянка важкого ланцюга донорського антитіла 2A10), а кількість замін знаходиться в межах від 1 до 9 В інших варіантах кількість замін складає 1, або 2, або 3, або 4, або 5, або 6, або 7, або 8 або 9 H99 VH являє собою еквівалент H1 VH (SEQ ID NO: 33) відмінного тільки в тому, що CDR H3 являє собою GQSY в H99 замість GQGY, як виявлено в H1. З метою оптимальної спорідненості зв'язування для гуманізації антитіла миші 2A10 (VH, для якого являє собою SEQ ID NO: 7) було виявлено, що пара залишків амінокислоти в положеннях 48 та 68, повинна відповідати І та А відповідно (як вони існують в 2A10) або M та V відповідно (як вони існують в H98). Очікують, що вищенаведені дані є також доцільними для гуманізації варіанту G95M 2A10. В одному варіанті зворотні мутації розташовані в положеннях, вказаних нижче в табл. 6, де залишок H99 у відповідному положенні замінений на залишок 2A10 у відповідному положенні (в таблиці "–" означає, що в тому положенні заміни немає, і, таким чином, залишок залишається як в послідовності H1). Таблиця 6 Номер залишку № за Kabat NewVH (SEQ ID NO.X) 2A10 H99 H100 (63) H101 (64) H102 (65) 38 38 K R K Антитіла або фрагменти, які містять варіабельні ділянки важкого ланцюга або людини або гуманізовані варіабельні ділянки важкого ланцюга та варіабельні ділянки легкого ланцюга Конструкти VH за даним винаходом можуть 40 40 R A R 48 48 І M І І І 67 66 K R К К 68 67 A V A A A 70 69 L M L 72 71 V R V 74 73 K T K 79 78 A V A A A бути сполучені з легким ланцюгом з метою утворення одиниці зв'язування з NOGO-A людини (Fv) в будь-якому форматі, включаючи звичайний формат антитіла IgG, що має змінну повної довжини (FL) і послідовності важкого ланцюга константного 13 96279 домену. Приклади послідовностей важкого ланцюга IgG1 з повною довжиною (FL), що містять конструкти VH за даним винаходом та інактивовані мутації в положеннях 235 та 237 (нумерація за 14 індексом ЄО) для передачі нелізованих антитіл, являють собою SEQ ID NO: 53, 54 та 55. SEQ ID NO: 53: Гуманізований конструкт H26 важкого ланцюга. SEQ ID NO: 54: Гуманізований конструкт H27 важкого ланцюга. SEQ ID NO: 55: Гуманізований конструкт H28 важкого ланцюга. Послідовність варіабельної ділянки легкого ланцюга, яка утворює Fv з послідовностями варіабельної ділянки важкого ланцюга за даним винаходом, може являти собою будь-яку послідовність, що дозволяє Fv зв'язуватися з NOGO-A людини. В одному варіанті даного винаходу варіабельна ділянка легкого ланцюга являє собою легкий ланцюг (див. WO 05/061544), варіабельна ділянка легкого ланцюга якого наведена в даному описі як SEQ ID NO: 8 або його гуманізовані варіанти. Гуманізовані варіанти 2A10 легкого ланцюга перева жно містять всю варіабельну ділянку легкого ланцюга CDR, яка описана в табл. 1, трансплантованій на каркас акцептора варіабельної ділянки легкого ланцюга людини. В одному варіанті гуманізовані варіабельні ділянки легкого ланцюга являють собою L11 (SEQ ID NO: 34), L13 (SEQ ID NO: 13 ) або L16 (SEQ ID NO: 14). Альтернативні варіабельні ділянки легкого ланцюга, які базуються на L13 та L16, що містять специфічні заміни в положеннях Kabat 37 та/або 45, наведені в табл. 7. 15 96279 16 Таблиця 7 Опис SEQ ID NO: L100(L13+Q37R) 67 L101(L13+Q45R) 68 L102(L13+Q37R/Q45R) 69 L103(L16+Q37R) 70 L104(L16+Q45R) 71 L105(L16+Q37R/Q45R) 72 Послідовність DIVMTQSPLSLPVTLGQPASISCRSSKSLLYKDGKTYLNWFRQ RPGQSPQLLIYLMSTRASGVPDRFSGGGSGTDFTLKISRVEA EDVGVYYCQQLVEYPLTFGQGTKLEIK DIVMTQSPLSLPVTLGQPASISCRSSKSLLYKDGKTYLNWFQQ RPGQSPRLLIYLMSTRASGVPDRFSGGGSGTDFTLKISRVEAE DVGVYYCQQLVEYPLTFGQGTKLEIK DIVMTQSPLSLPVTLGQPASISCRSSKSLLYKDGKTYLNWFRQ RPGQSPRLUYLMSTRASGVPDRFSGGGSGTDFTLKISRVEAE DVGVYYCQQLVEYPLTFGQGTKLEIK DIVMTQSPLSNPVTLGQPVSISCRSSKSLLYKDGKTYLNWFRQ RPGQSPQLLIYLMSTRASGVPDRFSGGGSGTDFTLKISRVEA EDVGVYYCQQLVEYPLTFGQGTKLEIK DIVMTQSPLSNPVTLGQPVSISCRSSKSLLYKDGKTYLNWFLQ RPGQSPRLLIYLMSTRASGVPDRFSGGGSGTDFTLKISRVEAE DVGVYYCQQLVEYPLTFGQGTKLEIK DIVMTQSPLSNPVTLGQPVSISCRSSKSLLYKDGKTYLNWFRQ RPGQSPRLLIYLMSTRASGVPDRFSGGGSGTDFTLKISRVEAE DVGVYYCQQLVEYPLTFGQGTKLΕΙΚ В іншому варіанті послідовності легкого ланцюга повної довжини (FL) являють собою L11 FL (SEQ ID NO: 36), L13 FL (SEQ ID NO: 17) або L16 FL (SEQ ID NO: 18). В іншому варіанті антитіла, їх фрагменти або їх функціональні еквіваленти містять послідовність VH, вибрану з H26, H27, H28, H 100, H101 та H102; в комбінації з будь-якою з наступних послідовностей VL L11, L13, L16, L100, L101, L102, L103, L104 та L105. Передбачається, що всі можливі комбінації перерахованих варіабельних ділянок важкого ланцюга та варіабельних ділянок легкого ланцюга конкретно розкриті (наприклад, H28L104 та інш.). В конкретних варіантах антитіла, їх фрагменти або їх функціональні еквіваленти містять наступні пари варіабельної ділянки: H27L16 (SEQ ID NO: 48 + SEQ ID NO: 14) H28L13 (SEQ ID NO: 49 + SEQ ID NO: 13) H28L16 (SEQ ID NO: 49 + SEQ ID NO: 14). В іншому варіанті антитіла за даним винаходом містять наступні послідовності повної довжини: H27FL L16FL (SEQ ID NO: 54 + SEQ ID NО:18) H28FL L13FL (SEQ ID NO: 55 + SEQ ID NО:17) H28FL L16FL (SEQ ID NO: 55 + SEQ ID NО:18). В одному варіанті антитіло за даним винаходом містить H27L16 (SEQ ID NO: 48 + SEQ ID NO: 14), або являє собою антитіло повної довжини H28FL L16FL (SEQ ID NO: 55 + SEQ ID NO: 18). В іншому варіанті антитіло або його фрагмент зв'язується з таким епітопом NOGO людини, як H28L16, або конкурує із зв'язуванням H28L16 з NOGO людини, що характеризується в обох випадках тим, що конкуруюче антитіло, або його фрагмент не являє собою антитіло 2A10 миші або його людський варіант, або його гуманізований варіант, що містить CDR H3 з послідовністю GQGY (SEQ ID NO: 3) або послідовність, що містить одну заміну амінокислоти в CDR H3. В конкретних варіантах антитіла, їх фрагменти або їх функціональні еквіваленти містять наступні пари варіабельної ділянки: H100L16 (SEQ ID NO: 63 + SEQ ID NO: 14) H101L13 (SEQ ID NO: 64+ SEQ ID NO: 13) H102L16 (SEQ ID NO: 65 + SEQ ID NO: 14) Побудова карти епітопу та подальші антитіла, які зв'язуються з однаковим епітопом В іншому варіанті запропоновано антитіло, або його фрагмент, здатне зв'язуватися з білком NOGO людини або його фрагментом, наприклад, білком GST-NOGO-A56 (SEQ ID NO: 32), в аналізі ELISA, де зв'язування антитіла або його фрагмента з білком NOGO людини або його фрагментом, в аналізі ELISA зменшується в присутності пептиду, що має послідовність VLPDIVMEAPLN (SEQ ID NO: 60), і не зменшується в присутності іншого пептиду, наприклад, пептиду з NOGO людини, який не перекривається з SEQ ID NO: 60 (наприклад, SEQ ID NO: 85, YESIKHEPENPPPYEE), який характеризується тим, що антитіло або його фрагмент не являє собою антитіло, що містить варіабельний домен важкого ланцюга, який має CDR H3, що складається із залишків амінокислот GQGY або її аналогів, що включає одну заміну амінокислоти в CDR H3. Альтернативно, конкуруючий пептид являє собою TPSPVLPDIVMEAPLN (SEQ ID NO: 73) або VLPDIVMEAPLNSAVP (SEQ ID NO: 74). Крім того, антитіло, яке зв'язується з таким же епітопом, що і вказані антитіла або їх фрагменти, може являти собою антитіло, яке не містить всіх CDR, наведених в табл. 1 та 2, або будь-яке антитіло, що містить набір CDR, які демонструють 80 % або більше гомології з CDR, наведених разом в табл. 1 та 2, або в табл. 1 або табл. 2 окремо. В іншому варіанті даного винаходу запропоновано антитіло або його фрагмент, здатне зв'язуватися в аналізі ELISA з ділянкою білка NOGO людини, що складається з поліпептидної послідовності VLPDIVMEAPLN (SEQ ID NO: 60), яке характеризується тим, що антитіло, або його фрагмент не являє собою антитіло, що містить варіабельний важкий домен, який включає CDR 17 H3, що складається із залишків амінокислоти GQGY, або його аналогів, що включають одну заміну амінокислоти в CDR H3. Альтернативно, антитіло або його фрагмент здатне зв'язуватися з TPSPVLPDIVMEAPLN (SEQ ID NO: 73) або VLPDIVMEAPLNSAVP (SEQ ID NO: 74). Крім того, антитіло, яке зв'язується з таким же епітопом, що й антитіла або їх фрагменти, може являти собою антитіло, яке не містить всіх CDR, наведених в табл. 1 та 2, або будь-яке антитіло, що містить набір CDR, які мають 80 % або більше гомології з CDR, наведених разом в табл. 1 та 2, або в табл. 1 або табл. 2 окремо. В іншому варіанті даного винаходу запропоновано спосіб одержання антитіла, або його фрагмента зв'язування, що зв'язується з епітопом VLPDIVMEAPLN (SEQ ID NO: 60) NOGO людини, що включає імунізацію ссавця вказаним пептидом та виділення клітин, здатних продукувати антитіло, яке зв'язується з вказаним пептидом. В іншому варіанті даного винаходу запропоновано спосіб одержання ізольованого антитіла або його фрагменту зв'язування, що зв'язується з епітопом VLPDIVMEAPLN (SEQ ID NO: 60) NOGO людини, що включає скринінг бібліотеки, яка складається з численних антитіл або їх фрагментів зв'язування, кожне з них може бути виділене з бібліотеки разом з послідовністю нуклеотиду, яка кодує антитіло або його фрагмент зв'язування, здатні до зв’язування антитіла, або його фрагмент зв'язування з епітопом VLPDIVMEAPLN (SEQ ID NO: 60) NOGO. Фармацевтична композиція Подальший аспект винаходу пропонує фармацевтичну композицію, що містить анти-NOGO антитіло за даним винаходом або його функціональний фрагмент, або його еквівалент разом з фармацевтично прийнятним розчинником або носієм. У подальшому аспекті даного винаходу запропоновано спосіб лікування або профілактики інсульту (особливо ішемічного інсульту) та інших неврологічних захворювань у людини, зокрема хвороби Альцгеймера, і лікування пацієнта, що зазнав механічної травми ЦНС (наприклад, пошкодження хорди), де вказаний спосіб включає введення людині, яка потребує допомоги, ефективної кількості анти-NOGO антитіла за даним винаходом або його функціональних фрагментів. В іншому аспекті даного винаходу запропоновано застосування анти-NOGO антитіла за даним винаходом або його функціонального фрагмента в одержанні лікарського засобу для лікування або профілактики інсульту (особливо ішемічного інсульту) та інших неврологічних захворювань, зокрема хвороби Альцгеймера, і лікування пацієнта, що зазнав механічної травми ЦНС (наприклад, пошкодження хорди). У подальшому аспекті даного винаходу запропоновано спосіб інгібувальної нейродегенерації та/або прискорення функціонального відновлення пацієнта-людини, ураженого або з ризиком розвитку інсульту (особливо ішемічного інсульту) або іншого неврологічного захворювання, зокрема хвороби Альцгеймера, і лікування пацієнта, що 96279 18 зазнав механічної травми ЦНС (наприклад, пошкодження хорди), який включає введення вказаній людині, яка потребує допомоги, ефективної кількості анти-NOGO антитіла за даним винаходом або його функціонального фрагменту. В ще подальшому аспекті даного винаходу запропоновано застосування анти-NOGO антитіла за даним винаходом або його функціонального фрагмента в одержанні лікарського засобу для інгібувальної нейродегенерації та/або прискорення функціонального відновлення пацієнта-людини, ураженого або з ризиком розвитку інсульту та іншого неврологічного захворювання, зокрема хвороби Альцгеймера, і лікування пацієнта, що зазнав механічної травми ЦНС (наприклад, пошкодження хорди). Інші аспекти та переваги за даним винаходом описані далі в детальному опису даного винаходу та його переважних варіантах. ДЕТАЛЬНИЙ ОПИС ВИНАХОДУ Варіабельні ділянки важкого ланцюга за даним винаходом можуть бути форматовані в структуру природного антитіла або його функціонального фрагмента або його еквівалента. Таким чином, антитіло може містити ділянки VH даного винаходу, форматовані в антитіло з повною довжиною, фрагмент (Fab)2, фрагмент Fab, або його еквівалент (наприклад, scFV, бі- три- або тетра-тіла, Tandabs та інш.) при спарюванні з відповідним легким ланцюгом. Антитіло може являти собою IgG1, ІgG2, ІgG3, або ІgG4; або IgM; IgA, IgE або IgD або його модифікований варіант. Відповідно можна вибрати константну ділянку антитіла важкого ланцюга. Константна ділянка легкого ланцюга може являти собою константну ділянку капа або лямбда. До того ж, антитіло може містити модифікації всіх класів, наприклад, димери IgG, мутанти Fc, які більше не зв'язуються з рецепторами Fc або проміжне зв'язування Clq. Антитіло може також являти собою химерне антитіло типу, описаного в WO86/01533, який містить ділянку зв'язування антигену та ділянку не-імуноглобуліну. Константну ділянку вибирають відповідно необхідній функціональності Звичайно IgG1 буде демонструвати здатність до лізису через зв'язування з комплементом та/або буде опосередковувати ADCC (клітинна цитотоксичність, залежна від антитіла). Переважним буде ІgG4, якщо потрібне не цитотоксичне блокуюче антитіло. Однак, антитіла ІgG4 можуть демонструвати нестабільність в продукуванні, і, таким чином, може бути більш переважною модифікацією загалом більш стійкого IgG1. Запропоновані модифікації описані в EP0307434, переважні модифікації вводять в положення 235 та 237. Таким чином, в даному винаході запропонована літична або нелітична форма антитіла відповідно даного винаходу. У переважних формах, таким чином, антитіло даного винаходу являє собою нелітичне антитіло IgG1 з повною довжиною (тобто, тетрамер H2L2), що містить варіабельні ділянки важкого ланцюга, описані в даному описі. У подальшому аспекті даного винаходу запропоновано полінуклеотиди, що кодують варіабельні ділянки важкого ланцюга, як описано в даному 19 описі. "NOGO" означає будь-який поліпептид NOGO, включаючи варіантні форми. Сюди входить, не обмежуючись ним, NOGO-A, що має залишки 1192 амінокислот (номер доступу GenBank AJ251383); NOGO-B, варіант зрощування, у якого немає залишків 186 - 1004 у передбачуваному позаклітинному домені (номер доступу GenBank AJ251384) і коротший варіант зрощування, NOGO-C, у якого також немає залишків 186 – 1004, він також має менший, альтернативний аміно кінцевий домен (номер доступу GenBank AJ251385) (Prinjha et al. (2000) див. вище). Мається на увазі, що всі посилання на "NOGO" в даному описі включають будьякі та всі варіантні форми NOGO, наприклад, NOGO-A та описані варіанти зрощування, якщо тільки не вказана специфічна форма. "Нейтралізація" та її граматичні варіації означає інгібування, або повне, або часткове, функцій NOGO, включаючи його зв'язування з нейронами та інгібування росту нейриту. Терміни Fv, Fc, Fd, Fab, або F(ab)2 використовують в їх стандартних значеннях (див., наприклад, Harlow et al., Antibodies A Laboratory Manual, Cold Spring Harbor Laboratory, (1988)). "Химерне антитіло" означає тип сконструйованого антитіла, яке містить природну варіабельну ділянку (легкий ланцюг і важкі ланцюги), що походить з донорського антитіла в асоціації з константними ділянками легкого та важкого ланцюга, що походять з антитіла-акцептора. "Гуманізоване антитіло" означає тип сконструйованого антитіла, що містить власні CDR, одержані з нелюдського донорського імуноглобуліну, одержані з імуноглобуліну частини молекули, що залишилися, будучи одержаними з одного (або більше) імуноглобуліну(ів) людини. Крім того, залишки підтримки каркасу можна змінити з метою збереження спорідненості зв'язування (див., наприклад, Queen et al., Proc Natl Acad Sci USA, 86:10029-10032 (1989), Hodgson et al., Bio/Technology, 9:421 (1991)). Придатне антитілоакцептор людини може являти собою одне з вибраних із звичайної бази даних, наприклад, база даних KABAT®, база даних Los Alamos і швейцарська база даних білків, по гомології з нуклеотидом і послідовностями амінокислот донорського антитіла (в даному випадку донорське антитіло 2A10 миші). Антитіло людини, що характеризується гомологією з каркасними ділянками донорського антитіла (на основі амінокислоти), може бути придатним з метою наведення константної ділянки важкого ланцюга та/або варіабельної каркасної ділянки важкого ланцюга для вставки донорських CDR (див. табл. 1 для 2A10 CDR для вставки в каркас акцептора). В подібній манері можна вибрати придатне антитіло-акцептор, константною ділянкою легкого ланцюга або варіабельною каркасною ділянкою якого можна пожертвувати Слід зазначити, що не потрібно, щоб важкі і легкі ланцюги антитіла-акцептора походили з того ж самого антитіла-акцептора Існують попередні повідомлення, що описують декілька шляхів продукування таких гуманізованих антитіл – див., наприклад, EPA-0239400 та EP-A-054951. 96279 20 Термін "антитіло-донор" означає нелюдське антитіло, яке робить внесок в послідовності амінокислоти варіабельних ділянок, CDR, або інших функціональних фрагментів або їх аналогів в гуманізованому антитілі, що забезпечуює гуманізоване антитіло антигенною специфічністю та нейтралізуючою активністю, характерними для донорського антитіла. Термін "антитіло-акцептор" означає антитіло, гетерологічне донорському антитілу, яке надає послідовності амінокислоти каркасних ділянок важкого та/або легкого ланцюга та/або константних ділянок важкого та/або легкого ланцюга гуманізованому антитілу Антитіло-акцептор можна одержати з будь-якого ссавця за умови, що воно є не імуногенним для людей Переважно антитілоакцептор являє собою антитіло людини. Альтернативно, гуманізацію можна забезпечити способом "маскування". Статистичний аналіз варіабельних ділянок важкого і легкого ланцюга унікального імуноглобуліну людини та миші виявив, що точні зразки виділених залишків різні в антитілах людини та миші, і найбільш індивідуальні поверхневі положення набагато більш переважні, ніж маленька кількість різних залишків (див. Padlan E.A et al.; (1991) Mol.lmmunol.28, 489-498 and Pedersen J.T et al (1994) J Mol Biol 235; 959973). Таким чином, можливо скоротити імуногенність нелюдського Fv, замінюючи виділені залишки в його каркасних ділянках, які відрізняються від тих залишків, що звичайно знаходяться в антитілах людини. Оскільки антигенність білка можна корелювати з поверхневою доступністю, може бути достатнім замінити поверхневі залишки, щоб зробити варіабельну ділянку миші "невидимою" для імунної системи людини (див також Mark G.E et al. (1994) в Handbook of Experimental Pharmacology vol. 113: The pharmacology of monoclonal Antibodies, Springer-Verlag, pp.105-134). Дана процедура гуманізації означає "маскування", тому що змінена тільки поверхня антитіла, залишки підтримки залишаються недоторканими. Інший альтернативний підхід описаний у WO04/006955. "CDR" визначені як послідовності амінокислоти антитіла на ділянці визначення комплементарності, вони являють собою гіперваріабельні ділянки важких і легких ланцюгів імуноглобуліну. Див., наприклад, Kabat et al., Sequences of Proteins of Immunological Interest, 4th Ed., U.S Department of Health and Human Services, National Institutes of Health (1987). У варіабельній частині імуноглобуліну існує три CDR важкого ланцюга і три CDR легкого ланцюга (або ділянки CDR). Таким чином, "CDR", як їх використовують в даному описі, означають всі три CDR важкого ланцюга, або всі три CDR легкого ланцюга (або як всі CDR важкого, так і всі CDR легкого ланцюга, якщо потрібно). Структура і складання білка антитіла, може означати, що інші залишки вважаються частиною ділянки зв'язування антигену, та фахівець буде вважати їх таким. Див., наприклад, Chothia et al., (1989) Conformations of immunoglobulin hypervariable regions; Nature 342, p.877-883. Біспецифічне антитіло являє собою антитіло, що володіє специфічністю зв'язування як мінімум з 21 двома різними епітопами. Способи виготовлення таких антитіл відомі в даній галузі. Традиційно, рекомбінантне продукування біспецифічних антитіл базується на співекспресії двох ланцюгових пар ланцюга L імуноглобуліну H, де два ланцюги H мають різну специфічність зв'язування, див Millstein et al., Nature 305 537-539 (1983), WO93/08829 and Traunecker et al. EMBO, 10, 1991, 3655-3659. Через випадковий асортимент ланцюгів H та L продукується потенційна суміш десяти різних структур антитіла, з яких тільки одна має бажану специфічність зв'язування. Альтернативний підхід включає злиття варіабельних ділянок з бажаною специфічністю зв'язування з константною ділянкою важкого ланцюга, що складається з як мінімум частини ділянки кріплення, ділянок CH2 та CH3. Переважно мати ділянку CH1, що містить ділянку, необхідну для зв'язування легкого ланцюга, присутнього в як мінімум в одному із продуктів злиття ДНК, що кодує дане злиття, і при бажанні ланцюг L, вставляють в окремі вектори експресії, а потім здійснюють співтрансфекцію в придатний організм-хазяїна. Хоча можливо вставити послідовності кодування для двох або всіх трьох ланцюгів в один вектор експресії. В одному переважному підході біспецифічне антитіло складається з ланцюга H з першою специфічністю зв'язування в одній гілці і ланцюговій парі H-L, що дає другу специфічність зв'язування в іншій гілці, див WO94/04690. Див. також Suresh et al. Methods in Enzymology 121, 210, 1986. В одному варіанті даного винаходу запропоновано біспецифічне терапевтичне антитіло, де як мінімум одна специфічність зв'язування вказаного антитіла зв'язується з NOGO людини в епітопі, описаному в SEQ ID NO: 60. В іншому варіанті даного винаходу біспецифічне антитіло містить послідовність варіабельної ділянки CDR H3 важкого ланцюга MQGY (SEQ ID NO: 45). В іншому варіанті біспецифічне антитіло містить наступні пари варіабельних ділянок важкого та легкого ланцюга: H27L16 (SEQ ID NO: 48 + SEQ ID NO: 14), H28L13 (SEQ ID NO: 49 + SEQ ID NO: 13) або H28L16 (SEQ ID NO: 49 + SEQ ID NO: 14). Антитіла за даним винаходом можна продукувати шляхом трансфекції клітини-хазяїна з вектором експресії, що включає послідовність кодування для антитіл за даним винаходом. Вектор експресії або рекомбінантну плазміду продукують, розміщуючи вказані кодуючі послідовності для антитіла в оперативній асоціації із звичайними регуляторними контрольними послідовностями, здатними контролювати реплікацію та експресію в, та/або секрецією з клітини-хазяїна. Регуляторні послідовності включають послідовності промотору, наприклад, промотор ЦМВ, і сигнальні послідовності, які можуть походити з інших відомих антитіл. Аналогічно, може бути продукований другий вектор експресії, що має послідовність ДНК, яка кодує легкий або важкий ланцюг комплементарного антитіла. Переважно даний другий вектор експресії є ідентичним першому, крім інзофара, стосовно послідовності кодування і маркерів, які можна вибрати, таким чином, щоб гарантувати в міру можливості, що 96279 22 кожен ланцюг поліпептиду функціонально експресований. Альтернативно, послідовності кодування важкого та легкого ланцюга для зміненого антитіла можуть знаходитися на єдиному векторі. Відібрана клітина-хазяїн одержана спільною трансфекцію за допомогою звичайних методів як з першим, так і другим векторами (або просто трансфікована єдиним вектором) з метою створення трансфікованої клітини-хазяїна даного винаходу, що містить рекомбінантні або синтетичні легкі ланцюги та важкі ланцюги. Далі вирощують трансфіковану клітину за допомогою звичайних методів з метою продукування сконструйованого антитіла за даним винаходом. Проводили скринінг антитіла, яке включає поєднання обох рекомбінантних важкого ланцюга та/або легкого ланцюга, з культури за допомогою відповідного аналізу, наприклад, ELISA або RIA. Подібні звичайні методи можна застосовувати для конструювання інших модифікованих антитіл і молекул. Одна придатна система експресії являє собою систему глутаматсинтетази (наприклад ту, що продає Lonza Biologies), особливо, де клітинахазяїн являє собою CHO або NSO (див нижче). Полінуклеотид, що кодує антитіло, легко ізолюється і розбивається на послідовності, із застосуванням звичайних процедур (наприклад, олігонуклеотидні зонди). Вектори, які можна застосовувати, включають плазміду, вірус, фаг, транспосони, мініхромсоми, з яких плазміда представляє типовий варіант. Звичайно такі вектори додатково включають сигнальну послідовність, походження реплікації, один або більше маркерних генів, посилюючий елемент, промотор і послідовності закінчення транскрипції, функціонально зв'язані з полінуклеотидом легкого та/або важкого ланцюга з метою полегшення експресії. Полінуклеотид, що кодує легкі та важкі ланцюги, можна вставляти в окремі вектори і вводити (наприклад, за допомогою електропорації) в одну клітину-хазяїн або при бажанні, як важкий ланцюг, так і легкий ланцюг можна вставляти в однаковий вектор для трансфекції в клітинухазяїн. Таким чином, відповідно до одного варіанта за даним винаходом запропоновано спосіб конструювання вектора, що кодує легкі та/або важкі ланцюги терапевтичного антитіла або його фрагмента зв'язування з антигеном за даним винаходом, вказаний спосіб включає вставку у вектор полінуклеотиду, що кодує легкий ланцюг та/або важкий ланцюг терапевтичного антитіла за даним винаходом. В іншому варіанті запропоновано полінуклеотид, що кодує гуманізовану варіабельну ділянку важкого ланцюга, що має послідовність, сформульовану як SEQ ID NO: 47, 48 або 49. В іншому варіанті запропоновано полінуклеотид, що кодує гуманізований важкий ланцюг, що має послідовність, сформульовану як SEQ ID NO: 53, 54 або 55. Для фахівців даної галузі буде очевидним, що завдяки надмірності генетичного коду, альтернативні полінуклеотиди, крім тих, що розкриті в даному описі, також доступні і будуть кодувати поліпептиди даного винаходу. 23 Придатні вектори для стадій клонування та субклонування, що використовують в способах і конструкціях композицій за даним винаходом, можуть бути вибрані фахівцями в даній галузі. Наприклад, можна використовувати звичайні серії pUC векторів клонування. Один вектор, pUC19, є комерційно доступним від таких постачальників, як Amersham (Buckinghamshire, Великобританія) або Pharmacia (Uppsala, Швеція). Додатково, для клонування можна використовувати будь-який вектор, який здатний легко відтворюватися, має достатньо ділянок клонування і генів, що можна вибирати (наприклад, антибіотична резистентність) та легко маніпулювати ними. Таким чином, відбір вектора клонування не є фактором обмеження в даному винаході. Інші переважні послідовності вектора включають сигнальну послідовність полі-A, наприклад, з телячого гормону росту (BGH) і послідовність промотору бетаглобіну (бетаглопро). Вектори експресії, придатні в даному описі, можна синтезувати методами, добре відомими фахівцям в даної галузі. Типові селекційні гени кодують білки, які (a) забезпечують резистентність до антибіотиків або інші антитоксини, наприклад, резистентність до ампіциліну, неоміцину, метотрексату або тетрацикліну, або (b) компенсують недостачу аукситрофних речовин, або постачають поживні речовини, недоступні в комплексних середовищах. В схему відбору може бути задіяний спосіб, що припиняє ріст клітини-хазяїна. Клітини, які успішно перетворені генами, що кодують терапевтичне антитіло за даним винаходом, виживають завдяки, наприклад, резистентності до лікарського засобу, забезпеченої селекційним маркером. Інший приклад - це так званий селекційний маркер DHFR, де трансформанти вирощують у присутності метотрексату. Клітини CHO являють собою особливо придатну лінію клітини для селекції DHFR. Способи селекції перетворених клітин-хазяїна та збільшення кількості копій клітин трансгену включають застосовування системи DHFR див., Kaufman R.J et a.l J Mol Biol (1982) 159, 601-621, для огляду, див., Werner RG, NoeW, Kopp K.Schluter M," Appropriate mammalian expression systems for biopharmaceuticals", Arzneimittel-Forschung 48(8):870-80, 1998 Aug. Ще один приклад являє собою систему експресії глутаматсинтетази (Lonza Biologies). Придатний селекційний ген для застосування в дріжджах являє собою ген trp1; див., Stinchcomb et al. Nature 282, 38, 1979. Компоненти таких векторів, наприклад, реплікони, селекційні гени, підсилювачі, промотори, сигнальні послідовності і т п., можна одержати з комерційних або природних джерел, або синтезувати за допомогою відомих процедур, що застосовують для спрямування експресії та/або секреції продукту рекомбінантної ДНК у відібраному хазяїні. Для даної цілі можна також відібрати інші відповідні вектори експресії, численні типи яких відомі в даній галузі для експресії клітин ссавців, бактерій, комах, дріжджів і грибків. У даному винаході також розглядають лінію клітин, трансфіковану рекомбінантною плазмідою, що містить послідовності кодування антитіл або 96279 24 еквіваленти за даним винаходом. Також звичайним є клітини-хазяїв, придатні для клонування та інші маніпуляції даних векторів клонування Однак, більш бажано використовувати клітини з різних штамів E. coli для реплікації векторів клонування та інших стадій в конструюванні змінених антитіл за даним винаходом. Придатні клітини-хазяї або лінії клітин для експресії антитіла за даним винаходом являють собою переважно клітини ссавця, наприклад, NSO, Sp2/0, CHO (наприклад, DG44), COS, фібробласт (наприклад, 3T3) і клітини мієломи, більш переважна, CHO або клітини мієломи. Можна використовувати клітини людини, таким чином, дозволяючи модифікацію молекулі в моделях глікозилювання у людини. Альтернативно, можна задіяти інші еукаріотні лінії клітини. В даній галузі відомі відбір придатних клітин-хазяїна ссавця і способів для перетворення, культивування, розведення, скринінга і продукування продукту та очищення. Див., наприклад, Sambrook et al., процитовані вище. Бактеріальні клітини можуть виявитися придатними як клітини-хазяїна, придатні для експресії антитіл або їх фрагментів (наприклад, рекомбінантні Fab або ScFv) за даним винаходом (див., наприклад, Pluckthun, A., Immunol Rev., 130:151-188 (1992)). Однак, завдяки тенденції, що білки, експресовані в бактеріальних клітинах, знаходяться в не звитій або неналежно звитій формі, або в неглікозилованій формі, будь-який рекомбінантний фрагмент, продукований в бактеріальній клітині, необхідно піддавати скринінгу, щоб гарантувати здатність до зв'язування з антигеном. Якщо молекула, експресована бактеріальною клітиною, була продукована в належним чином звитій формі, то бактеріальна клітина була би бажаним хазяїном. Наприклад, різні штами E. coli, що використовують для експресії, відомі як клітини-хазяї в сфері біотехнології. Різні штами B. subtilis, Streptomyces, інших бацил та їм подібних можна також задіяти в даному способі. При бажанні штами клітин дріжджів, відомих фахівцям в даній галузі, також доступних як клітини-хазяї, а також для даної цілі підходять клітини комах, наприклад, Drosophila і Lepidoptera та вірусні системи експресі.ї Див., наприклад, Miller et al., Genetic Engineering, 8:277298, Plenum Press (1986) і посилання, процитовані в даному описі. Загальні способи, за якими можна конструювати вектори, способи трансфекції, необхідні для продукування клітин-хазяїв за даним винаходом, і способи культивування, необхідні для продукування антитіла за даним винаходом від такої клітинихазяїна, являють собою цілком звичайні методи. Звичайно, спосіб культивування за даним винаходом являє собою спосіб культивування, вільний від сироватки, звичайно за допомогою культивування в суспензії клітин, вільних від сироватки. Таким же чином, продуковані антитіла за даним винаходом можна очистити з культури клітини за допомогою стандартних процедур даної галузі, включаючи преципітацію амонію сульфатом, афінну хроматографію, колонкову хроматографію, електрофорез на гелі і т. п. Подібні методи знаходяться в компетенції фахівців даної галузі і не 25 обмежують даний винахід. Наприклад, одержання антитіл описаний у WO 99/58679 та WO 96/16990. Ще в іншому способі експресії антитіл можна використовувати експресію у трансгенній тварині, наприклад, як описано в патенті США №4,873,316. Це стосується системи експресії, що застосовує промотор казеїну тварини, який, при трансгенному введенні ссавцю дозволяє секретувати бажаний рекомбінантний білок в молоці самки. У подальшому аспекті даного винаходу запропоновано спосіб продукування антитіла за даним винаходом, вказаний спосіб включає стадію культивування клітини-хазяїна, перетвореної або трансфікованої вектором, що кодує легкий та/або важкий ланцюг антитіла за даним винаходом і відновлює таким чином продуковане антитіло. Придатні клітини-хазяї для векторів клонування або експресії, що кодують антитіла за даним винаходом, являють собою прокароїтні, дріжджові або вищі еукаріотні клітини. Відповідні прокаріотні клітини включають еубактерії, наприклад, ентеробактерії, такі як Escherichia, наприклад, E. coli (наприклад, ATCC 31,446; 31,537; 27,325), Enterobacter, Erwinia, Klebsiella Proteus, Salmonella, наприклад, Salmonella typhimurium, Serratia, наприклад, Serratia marcescans та Shigella, а також бацили, такі як B. subtilis та B. licheniformis (див. DD 266 710), Pseudomonas, такі як P. aeruginosa та Streptomyces. Також розглядають дріжджові клітини-хазяї Saccharomyces cerevisiae, Schizosaccharomyces pombe, Kluyveromyces (наприклад, ATCC 16,045; 12,424; 24178; 56,500), yarrowia (EP402, 226), Pichia Pastoris (EP183, 070, див. також Peng et al. J.Biotechnol 108 (2004) 185192), Candida, Trichoderma reesia (EP244, 234J, Penicillin, Tolypocladium та хазяї Aspergillus, наприклад, A nidulans та A nigerare. Хоча у даному винаході конкретно розглядаються прокаріотні та дріжджові клітини-хазяї, однак звичайно клітини-хазяї за даним винаходом являють собою клітини хребетних тварин. Придатні клітини-хазяї хребетних включають клітини ссавця, наприклад, COS-1 (ATCC, NO: CRL 1650) COS-7 (ATCC CRL 1651), ембріональну ниркову лінію 293 людини, ниркові клітини дитинчати хом'яка (BHK) (ATCC CRL.1632), BHK570 (ATCC, NO: CRL 10314), 293 (ATCC NO: CRL 1573), клітини яєчника китайського хом'яка CHO (наприклад, CHO-K1, ATCC, NO: CCL 61, лінія клітини DHFRCHO така, як DG44 (див Urlaub et al., (1986) Somatic Cell Mol Genet 12, 555-556)), особливо ті лінії клітин CHO, що пристосовані для культури суспензії, клітини Сертолі миші, ниркові клітини мавпи, ниркові клітини африканської зеленої мавпи (ATCC CRL-1587), клітини HELA, ниркові клітини собаки (ATCC CCL 34), клітини легень людини (ATCC CCL 75), Hep G2 і клітини мієломи або лімфоми, наприклад, NSO (див. США 5,807,715), Sp2/0, Y0. Таким чином, в одному варіанті даного винаходу запропоновано стабільно перетворену клітину-хазяїна, яка містить вектор, що кодує важкий ланцюг та/або легкий ланцюг терапевтичного антитіла або його фрагмент зв'язування антигену, як описано в даному описі. Звичайно такі клітини 96279 26 хазяїни містять перший вектор, що кодує легкий ланцюг і другий вектор, що кодує вказаний важкий ланцюг. Клітини-хазяї, перетворені векторами, що кодують терапевтичні антитіла за даним винаходом або їх фрагменти зв'язування з антигеном, можна вирощувати за допомогою будь-якого способу, відомого фахівцям в даній галузі. Клітини-хазяї можна вирощувати в шейкерних колбах, ролерних пляшках або системах порожнього волокна, але переважним для широкомасштабного виробництва є резервуарних реакторах з перемішуванням, які переважно застосовують для суспендованих культур. Звичайно резервуари з перемішуванням пристосовані для аерації, застосовуючи, наприклад, розбризкувачі, турбулізатори потоку або роторні мішалки низької частоти. Для колонок міхурців і повітряних реакторів можна застосовувати пряму аерацію повітрям або міхурцями кисню. Якщо клітини-хазяї вирощують у культурі середовища, вільній від сироватки, переважно додавати у середовище агент, який захищає клітини, наприклад, плюронік F-68, з метою запобігання пошкодження клітин в результаті процесу аерації. В залежності від характеристик клітини-хазяїна можна використовувати або мікроносії як субстрати росту для ліній клітин, залежних від закріплення, або клітини можна адаптувати до культури суспензії (що є типовим). При вирощуванні клітин-хазяїнів, особливо клітин-хазяїнів хребетних можна застосовувати різноманітні оперативні методи, наприклад, насичення партії, повторна обробка партії (див. Drapeau et al. (1994) Cytotechnology 15: 103109), розширена обробка партії або культура перфузії. Хоча рекомбінантно перетворені клітинихазяї ссавця можна вирощувати в середовищах, що містять сироватку, такі середовища містять ембріональну телячу сироватку (FCS), переважним є вирощування таких клітин-хазяїнів в синтетичних вільних від сироватки середовищах, наприклад, як розкрито у Keen et al. (1995) Cytotechnology 17:153-163, або комерційно доступні середовища, такі як ProCHO-CDM або UltraCHO™ (Cambrex NJ, США), поповнені, якщо необхідно, джерелом енергії, наприклад, глюкозою та синтетичними факторами росту таким, як рекомбінантний інсулін. При вирощуванні клітинхазяїнів, вільних від сироватки, може знадобитися, щоб такі клітини адаптували для вирощування в умовах, вільних від сироватки. Один підхід адаптації являє собою вирощування таких клітинхазяїнів в сироватці, що містить середовища, і повторно обмінює 80 % середовища культури на середовища, вільні від сироватки, таким чином, що клітини-хазяїни вчаться адаптуватися в умовах, вільних від сироватки (див., наприклад, Scharfenberg K et al. (1995) in Animal Cell technology: Developments towards the 21st century (Beuvery E.C. et al. eds), pp.619-623, Kluwer Academic publishers). Антитіла за даним винаходом, секретовані в середовища, можна добути та очистити від середовища, застосовуючи різноманітні методи, з метою забезпечення ступеня очищення, придатного для передбаченого застосування. Наприклад, за 27 стосування терапевтичних антитіл за даним винаходом для лікування пацієнтів-людей звичайно вимагає як мінімум 95 % чистоти, більш традиційно, 98 % або 99 % чистоти в порівнянні з середовищем культури, що містить терапевтичні антитіла. В першу чергу, сторонні речовини клітин з середовища культури звичайно видаляють, застосовуючи центрифугування, після якого відбувається стадія очищення супернатанта, з використанням, наприклад, мікрофільтрації, ультрафільтрації та/або фільтрації глибини. Існують численні інші методи, такі як діаліз і електрофорез на гелі, а також хроматографічні методи, такі як гідроксиапатит (HA), афінна хроматографія (необов'язково, залучення системи афінного зв’язування, наприклад, полігістидину) та/або хроматографії гідрофобної взаємодії (HIC, див США 5, 429,746). В одному варіанті, антитіла за даним винаходом, проходячи різні стадії очищення, захоплюються із застосуванням протеїну білок А або афінної хроматографії G, далі ідуть подальші стадії хроматографії, такі як іонообмінна хроматографія та/або хроматографія HA, обмін аніонів або катіонів, ексклюзійна хроматографія та осадження амонію сульфатом. Звичайно, також використовують різні стадії видалення вірусу (наприклад, нанофільтрацію за допомогою, наприклад, фільтру DV-20). Слідуючи вказаним різним стадіям, запропоновано очищений (звичайно, моноклональний) препарат, що містить як мінімум 75 мг/мл або більше, наприклад, 100 мг/мл або більше антитіла за даним винаходом або його фрагмента зв'язування з антигеном, і, таким чином, утворено варіант за даним винаходом. Відповідно, такі препарати є істотною мірою вільними від агрегатних форм антитіл за даним винаходом. Відповідно до даного винаходу запропоновано спосіб продукування анти-NOGO антитіла за даним винаходом, яке специфічно зв'язується та нейтралізує активність NOGO-A людини, де вказаний спосіб включає стадії: (a) забезпечення першого вектора, що кодує важкий ланцюг антитіла; (b) забезпечення другого вектора, що кодує легкий ланцюг антитіла; (c) трансформація клітини-хазяїна ссавця (наприклад, CHO) вказаними першим і другим векторами; (d) вирощування клітини-хазяїна із стадії (c) в умовах, сприяючих секреції антитіла вказаною клітиною-хазяїном у вказану культуру середовища; (e) добування секретованого виділеного антитіла із стадії (d). Експресоване цільовим способом, антитіло далі досліджують щодо активності in vitro, застосовуючи відповідний аналіз. На сьогоднішній час застосовують звичайні формати аналізу ELISA з метою оцінки якісного та кількісного зв'язування антитіла з NOGO. Додатково можна використовувати інші аналізи in vitro з метою перевірки нейтралізуючої ефективності до проведення подальших клінічних досліджень на людях, які виконують для оцінки персистенції антитіла в організмі, незважаючи на звичайні механізми очищення. Інші модифікації антитіл за даним винаходом 96279 28 включають варіанти глікозилювання антитіл за даним винаходом. Відомо, що глікозилювання антитіл в консервативаних положеннях константних ділянок здійснює виражений вплив на функцію антитіла, особливо функціонування ефектору, наприклад, як описано вище, див., наприклад, Boyd et al. (1996), Mol. Immunol 32,1311-1318. Розглядають варіанти глікозилювання терапевтичних антитіл або їх фрагментів зв'язування з антигеном за даним винаходом, де один фрагмент вуглеводу або більше додають, заміняють, видаляють або модифікують. Введення мотиву аспарагін-X-серіну або аспарагін-X-треоніну створює потенційну ділянку для ферментного прикріплення вуглеводу, і, таким чином, може використовуватися з метою маніпуляції глікозилюванням антитіла. У Raju et al. (2001) Biochemistry 40, 8868-8876 збільшували кінцеве сіалування імуноадгезину TNFR-IgG за допомогою способу повторного галактозилування та/або повторного сіалування, застосовуючи бета1,4-галактозилтрансферазу та/або альфа, 2,3 сіалілтрансферазу. Вважають, що збільшення кінцевого сіалування збільшує період напіввиведення імуноглобуліну. Антитіла, в сукупності з більшістю глікопротеїнів, звичайно продукуються в природі як суміш глікоформ. Дана суміш особливо очевидна, якщо антитіла продукуються в еукаріотних клітинах, особливо в клітинах ссавця. Були розроблені різноманітні способи виробництва певних глікоформ, див. Zhang et al. Science (2004), 303, 371, Sears et al., Science, (2001) 291, 2344, Wacker et al. (2002) Science, 298 1790, Davis et al. (2002) Chem Rev 102, 579, Hang et al. (2001) Acc Chem Res 34, 727. Таким чином, даний винахід стосується множини терапевтичних (звичайно моноклональних) антитіл (які можуть бути IgG ізотипу, наприклад, IgG1), як описано в даному описі, що включає певну кількість (наприклад 7 або менше, наприклад 5 або менше, наприклад дві або одну) глікоформ вказаних антитіл або їх фрагментів зв'язування антигену. Терапевтичні засоби за даним винаходом можна вводити як профілактичні, або як лікування після інсульту на початку клінічних симптомів, або при необхідності. Доза і тривалість лікування пов'язані з відносною тривалістю циркуляції молекул за даним винаходом в організмі людини, і фахівець може коригувати її в залежності від стану, що лікують, і загального стану здоров'я пацієнта. Передбачається, що для досягнення максимальної терапевтичної ефективності може знадобитися повторна доза (наприклад, один раз на тиждень або один раз кожні два тижні) протягом тривалого періоду часу (наприклад, від чотирьох до шести місяців). Спосіб введення терапевтичного засобу за даним винаходом може являти собою будь-який придатний спосіб введення засобу хазяїну. Антитіла та фармацевтичні композиції за даним винаходом є особливо придатними для парентерального введення, тобто, підшкірно (п/ш), інтратекально, внутрішньочеревинно, внутрішньом'язово (в/м), внутрішньовенно (в/в) або інтраназально (і/н). Терапевтичні засоби даного винаходу можна приготувати як фармацевтичні композиції, що міс 29 тять ефективну кількість антитіла за даним винаходом як активний інгредієнт у фармацевтично прийнятному носії. Як профілактичний засіб даного винаходу переважною є водна суспензія або розчин, що містить сконструйоване антитіло, переважно, буферизовані до фізіологічних значень pH, у формі, готовій для ін'єкції. Композиції для парентерального введення будуть звичайно містити розчин антитіла за даним винаходом або його коктейль, розчинений у фармацевтично прийнятному носії, переважно водному носії. Можна використовувати різноманітні водні носії, наприклад, 0,9 % сольовий розчин, 0,3 % розчин гліцину і т. п. Вказані розчини є стерильними і звичайно вільні від твердих частинок. Ці розчини можна стерилізувати звичайними, добре відомими методами стерилізації (наприклад, фільтрацією). Композиції можуть містити фармацевтично прийнятні допоміжні речовини, як необхідно для наближення фізіологічних умов, наприклад корекції pH та вмісту буферизуючих агентів і т. п. Концентрація антитіла за даним винаходом в такій фармацевтичної рецептурі може широко варіювати, тобто від менш, ніж приблизно 0,5 %, звичайно вона складає 1 % або як мінімум приблизно 1 %, до 15 або 20 % (мас.), її вибирають перш за все, базуючись на об'ємах рідини, в'язкості і т. п., у відповідності до вибраного конкретного способу введення. Таким чином, фармацевтичну композицію за даним винаходом для внутрішньом'язової ін'єкції можна було би приготувати так, щоб вона містила 1 мл стерильної буферизованої води, в межах від приблизно 1 нг до приблизно 100 мг, наприклад, від приблизно 50 нг до приблизно 30 мг або, більш переважно, від приблизно 5 мг до приблизно 25 мг, та антитіла за даним винаходом. Аналогічно, фармацевтичну композицію за даним винаходом для внутрішньовенної інфузії можна було би приготувати так, щоб вона містила приблизно 250 мл стерильного розчину Рингера, і від приблизно 1 до приблизно 30, а переважно 5 мг до приблизно 25 мг сконструйованого антитіла за даним винаходом на мл розчину Рингера. Способи одержання композицій, що вводяться парентерально, добре відомі або будуть очевидними для фахівців в даній галузі, вони описані більш детально, наприклад, у Remington's Pharmaceutical Science, 15th ed., Mack Publishing Company, Easton, Pennsylvania. Для одержання внутрішньовенно препаратів антитіла за даним винаходом, що вводять внутрішньовенно, див. Lasmar U and Parkins D "The formulation of Biopharmaceutical products", Pharma Sci Tech rd Today, page 129-137, Vol.3 (3 April 2000), Wang, W "Instability, stabilisation and formulation of liquid protein pharmaceuticals", Int J Pharm 185 (1999) 129-188, Stability of Protein Pharmaceuticals Part A and В ed Ahem T.J., Manning M.C., New York, NY: Plenum Press (1992), Akers M.J "Excipient-Drug interactions in Parenteral Formulations", J Pharm Sci 91 (2002) 2283-2300, Imamura, K et al. "Effects of types of sugar on stabilization of Protein in the dried state", J Pharm Sci 92 (2003) 266-274, Izutsu, Kkojima, S. "Excipient crystalinity and its proteinstructure-stabilizing effect during freeze-drying", J Pharm Pharmacol, 54 (2002) 1033-1039, Johnson, R, 96279 30 "Mannitol-sucrose mixtures-versatile formulations for protein lyophilization", J. Pharm Sci, 91 (2002) 914922. Читач конкретно може звертатися до роботи Ha, E Wang W, Wang Y.J. "Peroxide formation in polysorbate 80 and protein stability", J. Pharm Sci, 91, 2252-2264,(2002), повний зміст, якої включений в даний опис шляхом посилання. Переважно, терапевтичний агент за даним винаходом, якщо знаходиться у фармацевтичному препараті, присутній у формах одиниці на дозу. Відповідна терапевтично ефективна доза буде легко визначена фахівцем в даній галузі. З метою ефективного лікування інсульту та інших неврологічних захворювань у людини, передбачається, що одну дозу антитіла в межах інтервалу від 700 до 3500 мг на 70 кг маси тіла за даним винаходом будуть вводити парентерально, переважно підшкірно, внутрішньовенно або внутрішньом'язово. Таку дозу можна, при необхідності, повторювати у відповідні проміжки часу, вибрані на розсуд лікаря в кожному конкретному випадку. Антитіла, описані в даному описі, можуть бути ліофілізовані для зберігання і розведені в придатному носії до застосування. Показано, що дана техніка ефективна із звичайними імуноглобулінами, і можна задіяти відомі в даній галузі методи ліофілізації та відновлення. В іншому аспекті даного винаходу пропонується фармацевтична композиція, що містить антиNOGO антитіло за даним винаходом або його функціональний фрагмент і фармацевтично прийнятний носій для лікування або профілактики інсульту та інших неврологічних захворювань. В іншому аспекті даного винаходу пропонується фармацевтична композиція, що містить антиNOGO антитіло за даним винаходом або його функціональний фрагмент і фармацевтично прийнятний носій для інгібування нейродегенерації та/або прискорення функціонального відновлення пацієнта-людини, ураженого або з ризиком розвитку інсульту або іншого неврологічного захворювання. В даному винаході додатково пропонується спосіб лікування або профілактики інсульту (особливо ішемічного інсульту) та інших неврологічних захворювань/розладів у людини, зокрема хвороби Альцгеймера, де вказаний спосіб включає введення людині, яка потребує допомоги, ефективної кількості анти-NOGO антитіла за даним винаходом або його функціонального фрагмента. Антитіла даного винаходу можна застосовувати в способах лікування з метою уповільнення або зупинки прогресування та/або початку хвороби Альцгеймера на додаток до (або як альтернатива) лікування встановленого захворювання у пацієнта-людини. Далі в даному винаході пропонується застосування анти-NOGO антитіла за даним винаходом, або його функціонального фрагмента в одержанні лікарського засобу для лікування або профілактики інсульту та інших неврологічних захворювань/розладів, зокрема хвороби Альцгеймера. У даному винаході також пропонується спосіб інгібування нейродегенерації та/або прискорення функціонального відновлення пацієнта-людини, ураженого або з ризиком розвитку інсульту або 31 іншого неврологічного захворювання/розладу, зокрема хвороби Альцгеймера, де вказаний спосіб включає введення людині, яка потребує допомоги, ефективної кількості анти-NOGO антитіла за даним винаходом або його функціонального фрагмента. Крім того, у даному винаході пропонується застосування анти-NOGO антитіла за даним винаходом або його функціонального фрагмента в одержанні лікарського засобу інгібування нейродегенерації та/або прискорення функціонального відновлення пацієнта-людини, ураженого або з ризиком розвитку інсульту, та іншого неврологічного захворювання/розладу, зокрема, хвороби Альцгеймера. Далі у даному винаході пропонується спосіб лікування або профілактики інсульту або іншого неврологічного захворювання/розладу, зокрема, хвороби Альцгеймера, у людини, включаючи стадію парентерального введення терапевтично ефективної кількості анти-NOGO антитіла за даним винаходом. Переважно вказане анти-NOGO антитіло вводять внутрішньовенно. Неврологічні захворювання або розлади, як застосовується вище, включають, не обмежуючись ними, травматичні пошкодження мозку, пошкодження спинного мозку, лобно-скроневі недоумства (тауопатії), периферичну невропатію, хворобу Паркінсона, хворобу Гантінгтона, і особливо хворобу Альцгеймера, розсіяний склероз або аміотрофний латеральний склероз (ALS). В даному винаході також пропонується спосіб прискорення росту відростків аксону, що включає стадію контакту людського аксону з анти-NOGO антитілом за даним винаходом. Даний спосіб можна виконувати in-vitro або in-vivo, переважно спосіб виконують in-vivo. Таким чином, у подальшому аспекті пропонується спосіб лікування інсульту (особливо ішемічного інсульту), пошкодження мозку, пошкодження спинного мозку, лобно-скроневих недоумств (тауопатії), периферичної невропатії, хвороби Паркінсона, хвороби Гантінгтона, розсіяного склерозу, і особливо хвороби Альцгеймера, у пацієнталюдини, де вказаний спосіб включає внутрішньовенне введення терапевтично ефективної кількості анти-NOGO антитіла за даним винаходом. У подальшому аспекті даного винаходу пропонується спосіб прискорення росту відростків аксону нейронів в межах центральної нервової системи людини (наприклад, пацієнта), де вказаний спосіб включає введення (наприклад, внутрішньовенне введення) терапевтично ефективної кількості анти-NOGO антитіла за даним винаходом. У подальшому аспекті даного винаходу пропонується застосування анти-NOGO антитіла за даним винаходом (наприклад, анти-NOGO антитіло, що включає CDR, розглянуті в даному описі) у виробництві лікарського засобу, що вводять внутрішньовенно, для лікування інсульту (особливо ішемічного інсульту), пошкодження мозку, пошкодження спинного мозку, лобно-скроневих недоумств (тауопатій), периферичної невропатії, хвороби Паркінсона, хвороби Гантінгтона, і особливо хвороби Альцгеймера, розсіяного склерозу 96279 32 або аміотрофного латерального склерозу (ALS) у пацієнта-людини. У подальшому аспекті даного винаходу пропонується спосіб регенерації процесів аксону в нейронах центральної нервової системи у пацієнталюдини, ураженого (або сприйнятливого) інсультом (особливо ішемічним інсультом), пошкодженням мозку, пошкодженням спинного мозку, лобноскроневими недоумствами (тауопатіями), периферичною невропатією, хворобою Паркінсона, хворобою Гантінгтона, розсіяним склерозом, і зокрема, хворобою Альцгеймера, де вказаний спосіб включає стадію введення (наприклад, внутрішньовенно) терапевтично ефективної кількості антитіла анти-NOGO за даним винаходом. У подальшому аспекті даного винаходу пропонується застосування анти-NOGO антитіла за даним винаходом у виробництві фармацевтичної композиції, що вводиться внутрішньовенно, для регенерації процесів аксону в нейронах центральної нервової системи у пацієнта-людини, ураженого (або сприйнятливого) інсультом (особливо ішемічним інсультом), пошкодженням мозку, пошкодженням спинного мозку, лобно-скроневими недоумствами (тауопатіями), периферичною невропатією, хворобою Паркінсона, хворобою Гантінгтона, розсіяним склерозом, і, зокрема, хворобою Альцгеймера. У подальшому аспекті даного винаходу пропонується спосіб модуляції продукування амілоїдогенного пептиду, що включає контакт клітини, яка експресує прекурсор, з якого походить амілоїдогенний пептид і поліпептид NOGO (наприклад, NOGO-A людини) з анти-NOGO антитілом за даним винаходом. У типових варіантах прекурсор являє собою APP. У подальших типових варіантах амілоїдогенний пептид являє собою Aβ, більш переважно Aβ40, Aβ42 або комбінація обох. Термін "функціональне відновлення", як його використовують в даному описі, означає моторне та/або сенсорне та/або поведінкове покращення у суб'єкта після, наприклад, ішемічного інсульту або пошкодження або початку клінічних симптомів. Функціональне відновлення у людей можна оцінити пристосуваннями, розробленими для виміру елементарних неврологічних функцій, наприклад рухової сили, сприйняття і координації, когнітивних функцій таких, як пам'ять, мова і здатність додержуватися вказівок, і функціональними можливостями, наприклад, основними видами діяльності повсякденного життя або інструментальною активністю. Відновлення елементарної неврологічної функції можна виміряти за допомогою пристроїв таких, як шкала виявлення інсульту NIH (NIHSS), відновлення пізнавальної функції можна виміряти за допомогою нейропсихологічних тестів, наприклад, Бостонський тест назв, тести діагностування слідів, і Каліфорнійський вербальний тест навчання, а види повсякденної діяльності можна виміряти з використанням таких інструментів, як шкала ADCS/ADL. (Клінічне дослідження хвороби Альцгеймера/Види діяльності повсякденного життя) або Бристольська шкала видів діяльності повсякденного життя, всі вказані тести та шкали, відомі в даній галузі. 33 96279 Наступні приклади ілюструють, не обмежуючи, даний винахід. Приклад 1 Побудова та експресія гуманізованих антитіл VH та VL Гуманізовані конструкти VH та VL приготували de novo шляхом побудови перекриття олігонуклеотидів, включаючи ділянки обмеження для клонування в вектори експресії Rid та RIn ссавця (або будь-який інший придатний вектор експресії для експресії білків в ссавцевих клітинах), а також в сигнальну послідовність людини. Ділянки обмеження Hind III та Spe I вводили з метою обрамлення ділянки VH, що містить сигнальну послідовність CAMPATH-1 H для клонування в Rid, що містить видозмінену константну ділянку γ1 людини для запобігання активності ADCC і CDC (L235A та G237A - Європейська система нумерації Індекс). Ділянки обмеження Hind III та BsiWI вводили з метою обрамлення ділянки VL, що містить сигнальну послідовність CAMPATH-1 H для клонування в Rin, 34 що містить константну ділянку капа людини. Сигнальна послідовність CAMPATH-1 H: MGWSCIILFLVATATGVHS (SEQ ID NO:31) Продуковані плазміди, що кодують послідовності амінокислот важкого ланцюга IgG людини, де CDR являла собою ту ділянку, що описана в табл. 2. Плазміди, що кодують послідовності амінокислот важкого ланцюга IgG людини, де CDR являли собою, продуковані з існуючих раніше плазмід шляхом введення мутацій єдиної точки, G95M (нумерація замін за Kabat), застосовуючи набір Quickchange (Stratagene). В наступній таблиці наведено, які послідовності білка важкого ланцюга з повною довжиною були зроблені у векторах плазміду та які послідовності були спарені, в значенні того, що єдиною різницею в послідовностях амінокислот спарених послідовностей важкого ланцюга з повною довжиною (FL) була заміна в G95M (нумерація замін за Кabat) в межах CDR H3 варіабельної ділянки: Таблиця 8 CDR, як визначено в табл. 2 H1 FL (SEQ ID NO: 35) H6 FL (SEQ ID NO: 15) H1 6FL(SEQ ID NO: 16) H20 FL(SEQ ID NO: 42) Заміна G95M (для утворення CDR H3 табл. 3) Немає H26 FL (SEQ ID NO: 53) H27 FL (SEQ ID NO: 54) H28 FL (SEQ ID NO: 55) Плазміди, що кодують важкі ланцюги далі були співтрансфіковані в клітини CHO (для більш конкретної інформації див. приклад 2) з однією з наступних послідовностей легкого ланцюга з повною довжиною: L11 FL (SEQ ID NO: 36), L13 FL (SEQ ID NO: 17), або L16 FL (SEQ ID NO: 18). Паралельно продукували химеру, названу HcLc (яка являє собою химеру 2A10 (SEQ ID NO: 9 та 10 - ланцюги з повною довжиною, що містять 2A10 мишачий VH (SEQ ID NO: 7) та VL (SEQ ID NO: 8) і константні ділянки людського IgG)). Приклад 2 Експресія антитіл в клітинах CHO Плазміди Rid та RIn (або інші вектори, придатні для застосування в клітинах ссавця), що кодують важкі та легкі ланцюги відповідно, були тимчасово co-трансфіковані в клітини CHO та експресували в маленькому масштабі або широкомасштабно з метою продукування антитіл. Альтернативно ті ж самі плазміди були співтрансфіковані в клітини DHFR-CHO шляхом електропорації, а стабільну поліклональну популяцію клітин, що експресують відповідне антитіло, відбирали, застосовуючи середовище, вільне від нуклеозидів (Rid містить ген DHFR, RIn містить селекційний маркер неоміцину. У деяких аналізах антитіла оцінювали безпосередньо з супернатанту тканинної культури. В інших аналізах рекомбінантне антитіло добували та очищали за допомогою афінної хроматографії на сефарозі/протеїні А. Приклад 3 Гуманізоване анти-NOGO антитіло зв'язується з NOGO Планшети Nunc immunosorp (100 мкл на лунку) вкривали GST-людським NOGO-A56 (див. приклад 5) в концентрації 0,05-1 мкг/мл в фосфатному буферному розчину протягом всієї ночі при 4 C. Лунки споліскували один раз TBS + 0,05 % Твіну (TBST), а потім інкубували з 2 % сироватки телячого альбуміну в TBST з метою блокування неспецифічних ділянок зв'язування при кімнатній температурі протягом 1 години. Антитіла розбавляли TBST + 2 % сироватки телячого альбуміну та доводили концентрацію до 10 мкг/мл з одержанням розведень ½. Антитіла двічі додавали в лунки та інкубували при кімнатній температурі протягом 1 години. Лунки промивали тричі TBST, а потім інкубували з кон'югатом проти людської пероксидази капа (1:2000) протягом 1 години. Лунки промивали тричі TBST, а потім інкубували з 100 мкл субстрату пероксидази OPD (Sigma) на лунку протягом 10 хвилин. Кольорову реакцію зупиняли, додаючи 25 мкл концентрованої H2SO4. Оптичну густину на довжині хвилі 490 нм вимірювали, застосовуючи показання планшета. Показання значень фону з лунок без антитіл віднімали. Фіг. 1-4 ілюструють залежне від дози зв'язування гуманізованих антитіл в порівнянні з химерою (названою HcLc, яка являє собою химеру 2A10 (включаючи 2A10 мишачий VH (SEQ ID NO: 7) та VL (SEQ ID NO: 8) і константні ділянки IgG людини)) з GST-людським NOGO-A56 (див. Приклад 5 для більш конкретної інформації) в аналізі ELISA. Вісь ординат демонструє виміряну оптичну густину (OD) в 490 нм, кількісна міра антитіла, захопленого в лунках. Вісь абсцис демонструє концентрацію антитіла, що використали (мкг/мл) на лунку в кожній точці даних. Матеріал антитіл, використаний на фіг 1-4, очищують, антитіло генерують або поліклональною системою експресії, або широкомасштабними тимчасовими трансфекціями. В даних випадках кількість IgG визначали за допомогою аналізу ELISA та вимірювання оптичної 35 96279 густини. Результати експериментів, показаних на фіг. 1-4, демонструють, що введення мутації G95M збільшує ефективність антитіла. Єдиним виключенням є H27L16, показаний на фіг. 3 і 4, яка виконувалася дуже слабо. Ми вважаємо, що вказані факти відбулися через неідентифіковану технічну проблему з аналізом H27L16, оскільки в інших аналізах H27L16 постійно демонструвала непогані результати (в ELISA, показаний на фіг, 1-2, в аналізах BIAcore (табл. 9, 10)). Також показано, що H27L16 добре працює в експериментах, виконаних пізніше (див. фіг. 11 та 12). Приклад 4 Протокол кількісного визначення антитіл Планшети Nunc lmmunosorp вкривали козячим антитілом захоплення проти людського ланцюга IgG (Sigma #13382) з концентрацією 2 мкг/мл в бікарбонатному буфері (Sigma #C3041) та інкубували протягом всієї ночі при 4 C. Планшети промивали двічі розчином буферу, що містить 0,05 % Твін 20 (TBST), і блокували 200 мкл TBST, що містить 2 % (або 1–3 %) сироватки телячого альбуміну (блокуючий буфер), протягом 1 години при кімнатній температурі. Планшети промивали двічі TBST. Супернатанти культури тканин, що містять антитіло, титрували через пластину в стадіях 2порядкового розведення в блокуючому буфері та інкубували при кімнатній температурі протягом 1 години. Планшети промивали тричі TBST. Кон'юговане антитіло H23 HRP (козяче проти людського ланцюга капа, Sigma #A7164) розбавляли TBST 1:2000 та додавали 100 мкл до кожної лунки. Планшети інкубували при кімнатній температурі протягом 1 години. Планшети промивали тричі TBST і проявляли 100 мкл субстрату Fast-OPD (Sigma #P9187). Колір проявлявся протягом 5–10 хвилин, після цього часу ELISA зупиняли за допомогою 25 мкл 3M H2SO4. Визначали поглинання на довжині хвилі 490 нм, концентрацію антитіла на планшеті визначали шляхом посилання на стандартну криву. Приклад 5 Продукування фрагмента NOGO-A (NOGO-A56 SEQ ID NO:32) Створили послідовність кДНК, що кодує поліпептид, який включає амінокислоти 586- 785 і мітку GST (SEQ ID NO:32) NOGO-A людини, шляхом клонування кДНК, що кодує амінокислоти 586-785 NOGO-A людини в ділянці BamHI-Xhol pGEX-6P1, з метою генерації злитого білка з міткою GST, що позначає GST-людський-NOGO-A56. Плазміду експресували в клітинах BL21 в середовищі 2XTY з 100 мкг/мл ампіциліну після індукування з IPTG до 0,5 мM при 37 C протягом 3 годин. Гранули 36 клітин піддавали лізису шляхом обробки ультразвуком, а злитий білок очищали, застосовуючи глутатіон-сепарозу (Amersham Pharmacia), дотримуючись інструкцій виробників. Здійснювали злиття очищеного білка, використовуючи відновлений глутатіон, і піддавали екстенсивному діалізу проти розчину фосфатного буферу, проводили кількісне визначення, застосовуючи стандарти сироватки телячого альбуміну та аналіз білка, що базується на BioRad coomassie, а потім зберігали цілими частинами при -80C. Приклад 6 Аналіз BiaCore гуманізованих моноклональних анти-NOGO антитіл Проаналізовано кінетику зв'язування моноклонального анти-NOGO антитіла (mAb) відносно рекомбінантно експресованого GST-людського антиNOGO, застосовуючи біодатчик Biacore3000 або BIAcore T100. Мікрочіп hNOGO-A готували, як вказано нижче: Метод GST-людський NOGO-A56 фіксували на мікрочіпі CM5 за допомогою сполучення з первинним аміном, застосовуючи програму Biacore Wizard, розроблену для цільових рівнів іммобілізації. Поверхню датчика CM5 активували, пропускаючи розчин 50 мМ N-гідрокси-сукциніміду (NHS) і 200 мM N-етил-N'-диметиламінопропілкарбонілу (EDC). Далі пропускали над мікрочіпом GSTлюдський NOGO-A56 в буфері натрію ацетату, pH 5,0 або pH 4,5, та фіксували. Після завершення іммобілізації будь-які активовані ефіри блокували шляхом ін'єкції 1 M етаноламіну гідрохлориду, pH 8,5. Анти-NOGO mAb розбавляли в HBS-EP (10 мM HEPES, pH 7,4, 150 мM NaCl, 3 мM ЕДТА та 0,005 % поверхнево-активної речовини P-20) у випадку T100, і дослідження зв'язування виконували в інтервалі певних концентрацій антитіл. Всі цикли реєстрували проти порожньої поверхні сенсору (такої, яка була активована та блокована, як описано раніше, але без додавання ліганду). Аналіз зв'язування здійснювали, використовуючи BIA оцінку програмної версії 4.1 кінетичного аналізу для BIAcore 3000 та програмну версію 1.0 кінетичного аналізу T100. Аналіз Biacore інших антитіл за даним винаходом в основному відбувався за таким же протоколом, як описано в даному описі. Якщо не було конкретних вказівок, експерименти BIAcore виконували при 25 C. В наступному розділі «Результати» кожна таблиця даних представляє результати, одержані в результаті індивідуального експерименту. Результати Таблиця 9 Антитіло HcLc H27L13 H26L13 H28L13 H27L16 H28L16 ka (1/мс) 3,19E6 6,2E6 3,23E6 7,26E6 6,24E6 7,25E6 kd (1/с) 2,49E-3 1,8E-3 3,11E-3 3,3E-3 1,21E-3 2,14E-3 KD (пМ) 779 291 963 454 194 296 37 96279 38 Результати Таблиця 10 Антитіло HcLc (25°C) HcLc (37°C) H16L16(25°C) H16L16(37°C) H20L16 (25°C) H20L16(37°C) H27L16(25°C) H27L16(37°C) H27L13(25°C) H27L13(37°C) H28L16(25°C) H28L16 (37°C) H28L13(25°C) H28L13(37°C) ka (1/мс) 2,66E6 5,08E6 3,43E6 5,31 E6 4,69E6 7,17E6 3,94E6 7,18E6 3,50E6 6,58E6 4,33E6 7,73E6 4,16E6 7,43E6 kd (1/с) 3,13E-3 7,74E-3 3,72E-3 6,16E-3 5,42E-3 1,08E-3 1,50E-3 3,06E-3 2,13E-3 4,22E-3 2,64E-3 5,24E-3 3,89E-3 7,59E-3 KD (нM) 1,18 1,46 1,08 1,16 1,16 1,51 0,380 0,426 0,606 0,641 0,610 0,678 0,936 1,02 Результати Таблиця 11 Антитіло HcLc H26L13 H27L13 H28L13 H28L16 ka (1/мс) 3,17E6 3,45E6 6,58E6 6,97E6 6,89E6 Приклад 7 Аналіз BiaCore гуманізованих моноклональних анти-NOGO антитіл, застосовуючи ранжування розв'язування Мікрочіп GST-людського NOGO-A56 готували щодо кінетичного аналізу. Супернатанти клітин брали безпосередньо з тимчасово трансфікованих клітин CHO-K1. Їх безпосередньо пропускали через поверхню сенсору та вимірювали взаємодію. Супернатант фальшиво трансфікованих клітин застосовували двічі з метою видалення будь-яких артефактів, що з'являлися в середовищі культури тканини. Всі цикли реєстрували проти порожньої поверхні сенсору (такої, яка була активована та блокована, як описано раніше, але без додавання ліганду). Аналіз зв'язування здійснювали, використовуючи оцінку програмної версії BIA 4.1 кінетичного аналізу. Приклад 8 Побудова карти пептидів Були одержані 47 перекритих пептидів, що охоплюють частину NOGO-A56 GST-людської ділянки (SEQ ID NO: 32) NOGO-A56, (з Mimotope™). Пептиди являють собою 16 амінокислот по довжині з дванадцятьма перекриттями амінокислот з суміжним пептидом (кожен пептид додатково містить послідовність біотин-SGSG на N-кінці) за винятком першого пептиду, який містить мітку GSGбіоцитин на C-кінці. Пептиди використовували для побудови карти епітопу ділянки зв'язування 2A10 та H28L16. Спосіб побудови карти епітопу: Планшети Nunc immunosorp (100 мкл на лунку) вкривали стрептавідином 5 мкг/мл в стерилізованій воді протягом всієї ночі при 37 C. Планшети kd (1/с) 2,33E-3 2,88E-3 1,83E-3 3,17E-3 1,95E-3 KD (нM) 0,74 0,87 0,28 0,45 0,28 тричі споліскували розчином фосфатного буферу, що містить 0,05 % Твіну (PBST), а потім блокували 3 % сироваткою телячого альбуміну в PBST при 4 C протягом всієї ночі. Планшети промивали тричі TBST. Далі до лунок додавали пептиди в концентрації приблизно 10 мкг/мл (розбавлені 3 % сироваткою телячого альбуміну в PBST) та інкубували при кімнатній температурі протягом 1 години. Планшети промивали тричі PBST, потім інкубували протягом 1 години з анти-NOGO антитілами, доведеними до 5 мкг/мл в 3 % сироватці телячого альбуміну в PBST. Планшети промивали тричі PBST, потім інкубували з кон'югатом проти людської пероксидази капа (1:1000, розбавленого в 3 % сироватці телячого альбуміну в PBST) протягом 1 години. Планшети промивали тричі PBST, а потім інкубували з 100 мкл субстрату пероксидази OPD (Sigma) на лунку протягом 10 хвилин. Кольорову реакцію зупиняли, додаючи 50 мкл 3 М H2SO4. Визначали поглинання на довжині хвилі 490 нМ, застосовуючи показання планшета. Результати показані на фіг. 5 (побудова карти епітопу, застосовуючи H28L16), фіг. 6 (побудова карти епітопу, застосовуючи 2A10). Фіг. 5 та 6 демонструють результати побудови карти епітопу 2A10 та H28L16 відповідно. Показані дані вказують на то, що 2A10 та H28L16 зв'язуються з пептидами 6 та 7, специфічна частина NOGO яких наведана в SEQ ID NO: 73 та SEQ ID NO: 74, відповідно, вони обидва містять послідовність VLPDIVMEAPLN. Дані результати вказують, що VLPDIVMEAPLN (SEQ ID NO: 60) містить епітоп зв'язування 2A10 та H28L16. 39 96279 Приклад 9 Порівняння HcLc та HcLc що містять мутацію G95M CDR H3 Модифікований варіант HcLc сконструювали з існуючої експресійної плазміди, вводячи одну точкову мутацію, G95M (нумерація замін за Kabat), застосовуючи набір Quikchange (Stratagene). Послідовність варіабельної важкої ділянки білка Hc (G95M) наведена в SEQ ID NO: 59. Hc(G95M) Lc експресували в клітинах CHO, як 40 описано раніш. Кількість антитіла визначали, як описано в Прикладі 4 Фіг. 7 та 8 демонструють порівняння активності зв'язування Hc(G95M)Lc та HcLc, як визначено, застосовуючи ELISA зв'язування NOGO-A людини, коли планшети Nunc immunosorp вкривали NOGO в 0,05 (фіг. 7) та 1 мкг/мл (фіг. 8). В таблиці нижче продемонстровано порівняння спорідненості зв'язування для Hc(G95M) Lc та HcLc. Таблиця 12 Дисоціація, виміряна за допомогою ранжування Biacore, що базується на одному експерименті Антитіло H6L13 H6(G95M)L13 HcLc Hc(G95M)Lc Послідовність ID варіабельної ділянки важкого ланцюга 11 47 7 59 Дані демонструють, що заміна G95M в межах CDR H3 не тільки збільшує активність зв'язування гуманізованих антитіл (H6L13), але також і мишачого донорського антитіла 2A10 (HcLc). Приклад 10 Конструювання та перевірка NOGO антитіл, що містять заміни в CDR H3 Панель 90 варіабельних ділянок важкого ланцюга була створена мутаціями єдиної точки в залишках, що містяться в CDR H3, або попередньому лейцині. Конкретно, були одержані вектори, що кодують важкий ланцюг (базуючись на H6FL, SEQ ID NO: 15), шляхом кодування варіабельних ділянок важкого ланцюга, де кожен залишок амінокислоти в CDR H3 і попередньому лейцині заміняли (використовуючи набір Quikchange (Stratagene)) на Дисоціяація kd (1/с) 1,38E-2 4,31 E-3 2,66E-3 6,14E-4 всі інші амінокислоти, що зустрічаються у природі, за винятком цистеїну, і експресували, з'єднуючи з легким ланцюгом (L13FL, SEQ ID NO: 17) з метою одержання 90 різних антитіл. Дані антитіла були проаналізовані щодо їх зв'язування з NOGO в експериментах ELISA та Biacore. Фіг. 9 та 10 демонструють порівняння активності зв'язування варіантів H6FL та H6FL L13FL. Табл. 14 та 15 демонструють порівняння кінетики розв'язування, що виміряно за допомогою Biacore, - показані тільки результати для тих антитіл, які показали вимірне розв'язування в аналізі Biacore і порівнянну активність зв'язування з H6L13 в ELISA. Таблиця 14 Батьківське антитіло H6L13 HcLc H6L13 H6L13 H6L13 H6L13 H6L13 H6L13 H6L13 H6L13 H6L13 H6L13 H6L13 H6L13 H6L13 H6L13 H6L13 H6L13 H6L13 Висновки Результати вказують, що антитіла, які зберігають властивості зв'язування мишачого 2A10, і GQGY, що містить антитіло H6L13, являють собою антитіла, що містять наступні CDR H3: RQGY, VH CDR3 MQGY GQGY GQNY GQLY IQGY RQGY GQSY GQGY GSGY GDGY GQGW GIGY GQYY GQFY WQGY GAGY GLGY GVGY GQWY kd (1/с) 4,85E-03 5,58E-03 9,66E-03 1,32E-02 172E-02 1,75E-02 1,86E-02 1,98E-02 2,07E-02 2,12E-02 2,16E-02 2,57E-02 3,28E-02 3,35E-02 1,98E-02 3,15E-02 1,90E-02 1,78E-02 1,77E-02 IQGY, MQGY, GDGY, GIGY, GSGY, GQNY, GQYY, GQSY, GQLY, GQFY, GQGW, WQGY, GAGY, GLGY, GVGY, GQWY. Приклад 11 Порівняння GQGY, що містить mab (H20L16) з G95M варіантних mabs (H27L16 та 41 96279 H28L13 та H28L16) Антитіла, наведені в табл. 15, одержували, як описано вище. Таблиця 15 Гуманізовані анти-Nogo-A антитіла 2A10, що дають загальну кількість зворотних мутацій для суцільного антитіла (2x важких ланцюга + 2x легких ланцюга) Антитіло H20L16 H28L16 H28L13 H27L16 Загальна кількість зворотних мутацій на суцільне антитіло/тетрамер 22 22 16 32 Характеристики зв'язування in vitro У спробі ранжування антитіл досліджували їх властивості зв'язування в низці аналізів, включаючи ELISA, зворотний формат ELISA, конкурентний метод ELISA, Biacore та протокову цитометрію. 11.1 Зв'язування з рекомбінантним NOGO-A людини в аналізі ELISA Досліджена здатність антитіл зв'язуватися з рекомбінантним NOGO-A людини (GST- людський NOGO-A 56) за допомогою різних аналізів, подібних ELISA (що виконували за схожим, але з невеликою різницею, протоколом, як той, що описано в Прикладі 3). У першому аналізі рекомбінантним NOGO-A безпосередньо вкривають планшетах в різних концентраціях антигену. Результати прямого зв'язування ELISA, якщо антиген навантажують 1 мкг/мл або 0,05 мкг/мл, показані у фіг. 11A та фіг. 11B відповідно. Дані підтверджують, що всі антитіла демонструють порівнянну активність зв'язування з рекомбінантним NOGO-A людини при порівнянні з химерною формою батьківського антитіла (HcLc). У вищих концентраціях антигену покриття всі антитіла дають подібне значення EC50. На відміну від цього, при нижчій концентрації антигену покриття аналіз міг відрізнятися серед антитіл. Хоча не одержували криві насичення, аналіз тенденції на лініях виявив наступний ранг порядку: H27L16>H28L16, H28L13, H20L16. У паралельному експерименті формат аналізу був зворотним. В даному форматі антитіло захоплюють на планшет і визначають зв'язування рекомбінантного NOGO-A людини (GST-людський NOGO-A-56), застосовуючи мітку GST. Результати 42 зворотного формату ELISA показані на фіг. 12. Дані підтверджують, що всі антитіла демонструють порівнянну активність зв'язування з рекомбінантним NOGO-A людини при порівнянні з химерною формою батьківського антитіла (HcLc). Даний формат зв'язування ELISA не відрізнявся між антитілами. 11.2 Конкурентний метод ELISA Застосовуючи конкурентний метод ELISA, оцінили здатність антитіл конкурувати безпосередньо з батьківським антитілом для однакового епітопу NOGO-A людини. Планшети вкривали рекомбінантним NOGO-A людини (GST-людський NOGO-A 56 людини). Батьківське антитіло 2A10 та гуманізовані антитіла попередньо перемішують до додавання на планшети. Кількість зв'язування 2A10 визначали, застосовуючи кон'югат анти-мишачого IgGHRP (Dakocytomation, #P0260). Результати, показані у фіг. 13, підтверджують, що всі чотири антитіла можуть конкурувати з 2A10. Даний факт свідчить, що гуманізовані антитіла та батьківське антитіло розпізнають епітоп перекривання на NOGO-A людини. До того ж, активність гуманізованих антитіл порівнянна або краще, ніж химери HcLc. Результати вказують, що H27L16, H28L16 та H28L13 більш могутні, ніж H20L16. 11.3 Вимірювання спорідненості за допомогою Biacore Biacore застосовували з метою визначення спорідненості та ранжування антитіл, застосовуючи дві різні методології. У першому підході рекомбінантний NOGO-A сполучали з поверхнею мікрочіпа, NOGO-A антитіла пропускали через дану поверхню. У другому підході протеюн. A використовували з метою захоплення антитіла на поверхні мікрочіпа, над яким пропускали рекомбінантний GST-людський Nogo-A56. Результати, показані в табл. 16, були одержані шляхом сполучення антигену з поверхнею, і підтверджено, що всі чотири антитіла демонструють порівнянну/кращу спорідненість, ніж батьківське антитіло (HcLc). Базуючись в середньому на шести незалежних циклах, антитіла ранжовані в наступному порядку з точки зору загальної спорідненості: H27L16 > H28L16 > H28L13 > H20L16, що узгоджується з порядком ранжування прямого зв'язування ELISA (фіг. 11B). У випадку H27L16 та H28L16 гуманізовані антитіла демонструють в два-три рази вищу спорідненість, ніж у батьківському антитілі (HcLc). 43 96279 44 Таблиця 16 Кінетика зв'язування гуманізованих анти-NOGO-A антитіл з рекомбінантним NOGO-A людини (GSTлюдський NOGO-A 56), як визначено, застосовуючи Biacore Т100. Антиген був зв'язаний з мікрочіпом CMS за допомогою первинного сполучення аміну. Антитіла витікали при різних концентраціях (0,125- 8 нM). Показники демонструють середнє та стандартне відхилення (у дужках) 6 незалежних циклів, кожний цикл, зроблений двічі, а кожен набір даних проаналізований незалежно до обчислення середнього та стандарт** ного відхилення. Проаналізовано тільки 11 наборів даних для H20L16, оскільки один набір проаналізувати не змогли. Антитіло H20L16** H27L16 H28L13 H28L16 HcLc Ka 5,37E6 (7,65E5) 3,96E6 (9,93E5) 8,13E6 (1,35E6) 6,97E6 (6,62E5) 3,80E6 (7,11E5) У подібний спосіб ELISA кінетику зв'язування антитіл з рекомбінантним NOGO-A людини (GSTлюдський NOGO-A 56) також оцінювали в зворотному форматі (див. Приклад 11.1). У даному аналізі гуманізовані антитіла захоплювали на мікрочіп CM5 з протеїном А. Середні результати для шести kd 9,70E-3 (2,65E-3) 2,30E-3 (1,11E-3) 9,10E-3 (2,65E-3) 4,43E-3 (1,18E-3) 7,09E-3 (2,22E-3) KD(нM) 1,80 (0,31) 0,56 (0,15) 1,11 (0,18) 0,64 (0,15) 1,86 (0,32) незалежних циклів показані в табл. 17. Узгоджено із зворотним форматом ELISA, всі гуманізовані антитіла демонструють подібну кінетику зв'язування з химерою (HcLc) в зворотному форматі Biacore. Таблиця 17 Кінетика зв'язування зворотного формату гуманізованих анти-NOGO-A антитіл з рекомбінантним NOGO-A людини (GST NOGO-A 5+6), як визначено, застосовуючи Biacore T100. Протеїн А фіксували з приблизно 4000 ОР за допомогою первинного аміну та використовували для захоплення приблизно 200-300 ОР зразкових антитіл. Рекомбінантний NOGO-A людини проходив в різних концентраціях (0,125–8 нM). Показники демонструють середнє та стандартне відхилення (у дужках). З незалежних циклів, кожний цикл, зроблений двічі, а кожен набір даних проаналізований незалежно до обчислення середнього та стандартного відхилення. Антитіло H20L16 H27L16 H28L13 H28L16 HcLc Ka 1,01E6 (1,35E5) 9,93E5 (2,02E4) 1,12E6 (1,21E5) 1,18E6 (8,32E4) 1,38E6 (3,70E5) 11.4 Зв'язування з природним NOGO людини З метою демонстрації того, що гуманізовані антитіла зв'язуються з природним NOGO-A людини з профілем, порівнянним з батьківським антитілом, розробили два аналізи, що базуються на протоковій цитометрії. В першому аналізі генерували лінію клітини, що базується на CHO-K1, що експресує позаклітинний домен NOGO-A людини на поверхні клітини. Зв'язування гуманізованих антитіл оцінювали за допомогою протокової цитометрії, використовуючи PE-мічений анти-людський IgG (Sigma, #P8047). Фіг 14 нижче демонструє типовий профіль для анти-NOGO-A антитіл на лінії клітини CHO- NOGO-A. Хоча аналіз є недостатньо чутливим для визначення різниці між антитілами, результати підтверджують, що всі чотири антитіла можуть розпізнавати поверхню клітини, експресуючої NOGO-A людини, на рівнях, порівнянних з рівнями химери. Жодне з антитіл не розпізнало батьківську лінію клітин (CHO-K1, дані не показані). У другому аналізі здатність гуманізованих ан kd 3,13E-4 (2,79E-5) 3,04E-4 (1,83E-5) 3,84E-4 (3,24E-5) 4,01E-4 (2,48E-5) 5,69E-4 (1,54E-4) KD(нM) 0,31 (0,036) 0,31 (0,019) 0,34 (0,015) 0,34 (0,032) 0,41 (0,062) титіл зв'язуватися з природним NOGO-A оцінювали, застосовуючи лінію клітин нейробластоми людини, IMR32. Дана лінія клітин характеризується високими внутрішньоклітинними/низькими зовнішніми рівнями білка поверхні клітини NOGO-A. У спробі збільшити сигнал зв'язування був розроблений аналіз визначення внутрішньоклітинного NOGO-A (ER-резидента). Клітини IMR32 просочували і фіксували перед забарвленням гуманізованими анти-NOGO-A антитілами. Зв'язування антитіл з NOGO-A визначали, застосовуючи антилюдський IgG-PE, мічений вторинно (Sigma, #P8047). Результати, показані на фіг. 15 нижче, підтверджують, що всі антитіла зв'язуються з внутрішньоклітинним NOGO-A на рівнях, порівнянних або вище, ніж батьківське антитіло HcLc. Вказані дані разом з результатами для лінії клітин CHONOGO-A підтверджують, що гуманізовані антитіла можуть розпізнавати більш природну форму білка NOGO-A на рівнях, порівнянних або краще, ніж химера, HcLc. Дані аналізи недостатньо чутливі для їх ранжування до панелі антитіл. 45 11.5 Аналізи нейритного відростка Гуманізовані антитіла перевіряли на їх здатність до нейтралізації інгібуючої нейритний відросток (NO) активності NOGO-A в аналізі, що базується на визначенні кількості NO, як описано раніше. Антитіла, перевірені в даному аналізі, були вибрані на підставі їх кінетики зв'язування з NOGO-A. Гуманізовані антитіла з високою спорідненістю, а саме, H28L16, H27L16, H20L16, і, як довідкові, їх батьківські антитіла 2A10 (мишачі моноклональні) та HcLc (людська химера миші) перевірили на нейтралізацію NOGO-A. Для порівняння антитіло 11C7 (див. Приклад 13) також перевіряли за допомогою даного аналізу. З метою перевірки активності нейтралізації відібраних гуманізованих антитіл, лунки вкривали рекомбінантним GST-людським Nogo-A56 людини і обробляли варіюючими концентраціями антитіл при 37 C протягом 1 години перед додаванням гранульованих нейронів мізочка (CGN). Контрольні лунки обробляли HBSS Середню довжину нейриту на нейрит вимірювали для кожної лунки. Фіг 16 демонструє результати для гуманізованих антитіл, перевірених в даному аналізі. Панель контрольних антитіл (контрольний IgG, очищений IgG миші; Campath та інші не споріднені гуманізовані антитіла), використовували для підтвердження специфічності активності. Як додатковий контроль однакові гуманізовані антитіла титрували на планшетах, вкритих GST. Результати підтверджують, що зворотне NOGO-A - опосередковане інгібування нейритного відростка за допомогою H28L16, H27L16 та H20L16 до подібного ступеня спостерігали для батьківських антитіл (2A10 та HcLc). Ймовірно, ефекти були сильними та стійкими і спостерігалися з H28L16 у восьми з одинадцяти незалежних експериментів з нейритним відростком. Навпаки, гуманізовані антитіла не збільшують відросток нейриту на планшетах, вкритих GST, а панелі контрольних антитіл не демонструють будь-якої дозозалежної зміни інгібування, підтверджуючи, що вплив гуманізованих антитіл ж специфічним по відношенню до опосередкованого NOGO-A інгібування. Дані, представлені для нейритного відростка, вибрані з низки експериментів, що повторювалися. Оскільки ряд повторів виявилися різними за природою, вважають, що продемонстровані дані відображають справжню активність антитіл за даним винаходом при зменшенні інгібуючого ефекту NOGO в аналізах з нейритним відростком. Приклад 12 Подальша характеристика H28L16 12.1 Зв'язування з рекомбінантним NOGO-A з повною довжиною Досліджена здатність антитіл до зв'язування з рекомбінантним NOGO-A людини позаклітинного домену з повною довжиною (GST-людський NogoA-ECD) за допомогою прямого аналізу зв'язування ELISA. В даному випадку ECD був варіантом з'єднання, що знаходився в межах ділянки приблизно в положенні 186-1004 NOGO А людини (частина, що починається з DETFAL (SEQ ID NO: 95) і закінчується з ELSKTS (SEQ ID NO: 96)). Планшети безпосередньо вкривали 1 мкл/мл рекомбінантного GST-людського NOGO-A-ECD. Дані, показані на фіг. 17, підтверджують, що 96279 46 H28L16 може розпізнавати GST-людський Nogo-AECD на рівнях, порівнянних або краще, ніж батьківські (HcLc) або H20L16. 12.2 Інгібування функціональності Fc З метою покращення профілю безпеки кандидата залишки L235 та G237 в межах домену CH2 константної ділянки важкого ланцюга (Європейська система Індекс) були видозмінені до залишків аланіну, зменшуючи, таким чином, ймовірність запуску опосередкованих антитілом імунологічних ефекторних функцій. Зменшене зв'язування з C1q людини застосовували як сурогат для інгібування функціональності Fc. Фіг 18 нижче демонструє, що H28L16 істотно знижував активність зв'язування C1q, порівнянну з активністю Campath-lgG1 (дикий тип) і порівнянну з активністю конструкта Campath IgG1, що піддається таким же мутаціям (Fcмутоване антитіло (Fc-)) та Campath lgG4 Вказані дані свідчать, що мутації домену CH2 присутні в H28L16, будуть значно зменшувати ймовірність запуску опосередкованих ефекторних функцій Fc. 12.3 Зв'язування ортологів Для підтвердження того, що H28L16 демонструє активність зв'язування з різними ортологами NOGO-A, порівнянну з активністю батьківського антитіла (HcLc), виконували серії аналізів зв'язування. Фіг. 19 A-D нижче демонструють результати прямого зв'язування ELISA з рекомбінантним NOGO (GST-людським NOGO-A 56) щура (SEQ ID NO: 94), яванської макаки (SEQ ID NO: 92), мавпочки (SEQ ID NO: 93) і біличої мавпи (SEQ ID NO: 91), відповідно. У всіх випадках H28L16 демонструє активність, порівнянну або кращу, ніж активність химерного антитіла (HcLc). Обчислені значення EC50 дуже подібні до значень, обчислених для зв'язування з рекомбінантним NOGO-A людини. Кінетику зв'язування H28L16 з різними ортологами NOGO-A в порівнянні з HcLc та 11C7 визначали, застосовуючи Biacore. Табл. 18 та 19, наведені нижче, демонструють кінетику зв'язування в двох різних форматах аналізу. Якщо рекомбінантний NOGO-A сполучений безпосередньо з мікрочіпом CM5 (табл. 18), кінетика зв'язування для щура, яванської макаки, біличої мавпи і мавпочки дуже подібна до кінетики людини (інтервал = 0,330,67 нM). Якщо формат аналізу був зворотнім і антитіла захоплюють на мікрочіп, застосовуючи протеїн А (табл. 19), спорідненість зв'язування H28L16 з NOGO-A щурів є приблизно на 4 порядки нижчою, ніж у NOGO-A людини. Подібну тенденцію спостерігають для NOGO-A яванської макаки (на 8,5 порядки нижча спорідненість, ніж у людей) та ортологів інших приматів (на 12-17 порядків нижча спорідненість, ніж у людей). Химерне антитіло HcLc демонструє подібний профіль зв'язування з ортологами NOGO-A в обох орієнтаціях аналізу. Оскільки не є зрозумілим, який формат аналізу краще всього представляє ситуацію in vivo, первинні висновки, які можна зробити з даного дослідження, такі: 1) H28L16 зберігає профіль перехресної реактивності ортологу, асоційований з химерним антитілом HcLc і 2) спорідненість HcLc для NOGO-A щура і яванської макаки знаходиться в межах 4 - 8,5 порядків спорідненості NOGO-A 47 96279 людини і за певних умов може бути дуже подіб 48 ною. Таблиця 18 Кінетика зв'язування H28L16, 11C7 та HcLc рекомбінантними ортологами NOGO-A людини, як визначено за допомогою Biacore T100. Приблизно 140-180 ОР різних ортологів NOGO-A захоплені на мікрочіпі CM5 сполученням з первинним аміном. Антитіла витікали при різних концентраціях (0,125-8 нM). Показники демонструють середнє та стандартне відхилення (у дужках) 1-2 незалежних циклів, кожний цикл, зроблений двічі, а кожен набір даних проаналізований незалежно до обчислення середнього та стандартного відхилення. * Один набір кривих відкинули через неможливість інтерпретації кривих для антитіла 11C7. Ортолог Ka H28L16 Kd Яванська макака (2 цикли) Щур (2 цикли) Мавпочка (1 цикл) Мавпа Squirrel (1 цикл) Людина 4,65E6 (7,47E5) 11C7 KD Ka Kd (нM) 3,07E-3 0,67 1,47E6 3,40E-4 (2,37E-4) (0,06) (1,67E5) (4,45E-5) 4,64 E6 (2,34E5) 4,2E6 (2,47E4) 4,46E6 (6,08E4) 6,97E6 (6,62E5) 1,54E-3 (3,06E-5) 3,02E-3 (5,09E-5) 2,73E-3 (4,95E-6) 4,43E-3 (1,18E-3) 0,33 (0,01) 0,626 (0,000) 0,61 (0,000) 0,64 (0,15) 8,36E5 (5,58E5) 1,16E6 (5,37E4) 1,10E6 (3,25E4) 1,58E6 (6,42E5) KD (нM) 0,23 (0,01) 1,20E-4 0,11 (2,14E-5) (0,03) 2,80E-4 0,24 (6,15E-6) (0,006) 2,86E-4 0,26 (1,87E-5) (0,010) 2,64 E-4 0,19 (5,57E-5) (7,96E-2) HcLc Kd Ka 2,94E6 4,78E-3 (7,13E5) (6,34E-4) KD (нM) 1,68 (0,35) 2,53E6 (5,32E4) 3,13E6 (2,76E4) 3,04E6 (1,64E5) 3,80E6 (7,11E5) 1,12 (0,03) 1,419 (0,03) 1,54 (0,15) 1,86 (0,32) 2,83E-3 (2,30E-5) 4,44E-3 (1,41 E-4) 4,68E-3 (2,11 E-4) 7,09E-3 (2,22E-3) Таблиця 19 Кінетика зв'язування зворотного формату H28L16, 11C7 та HcLc з рекомбінантними ортологами NOGO-A людини, як визначено за допомогою Biacore T100. Білок А фіксували на поверхні приблизно з 4000 ОР та антитіла анти-NOGO-A захопили приблизно з 300-400 ОР. Рекомбінантні білки (GST- NOGO-A56) витікали при різних концентраціях (0,125- 64 нM) в залежності від конструкта. Всі цикли здійснювали в подвійному повторенні. Показники демонструють середнє та стандартне відхилення (у дужках) 1-3 незалежних циклів, кожний цикл, зроблений двічі, а кожен набір даних проаналізований незалежно до обчислення середнього та стандартного відхилення. H28L16 ортолог Cynomolgus (3 цикли) Щур (3 цикли) Мавпочка (1 цикл) Мавпа Squirrel (1 цикл) Людина (1 цикл) Ka Kd 3,26E5 (4,06E3) 3,80E5 (5,68E3) 2,22E5 (3,61 E3) 1,57E5 (2,69E3) 1,20E6 (8,49E4) 1,11E-3 (2,23E-5) 6,69E-4 (1,24E-5) 1,09E-3 (7,35E-5) 1,08E-3 (5,02E-5) 4,75E-4 (9,97E-6) 11C7 KD (нM) 3,41 (0,05) 1,76 (0,03) 4,89 (0,25) 6,86 (0,20) 0,40 (0,02) 12.4 Фізичні властивості Оцінювали фізико-хімічні властивості H28L16 та H20L16 за допомогою ЕК-ВЕРХ та SDS-PAGE. ЕК-ВЕРХ здійснювали в 1,0 мл/хв., використовуючи 100 мM натрію фосфату, 400 мM натрію хлориду pH 6,8 і колонку з неіржавіючої сталі TSK G3000 SW 130 см 7,8 мм з детектуванням на довжині Ka 4,02E5 (6,85E4) 2,83E5 (4,66E4) 1,91E5 (2,90E3) 1,03E5 (2,12E3) 2,64E5 (3,32E3) KD Kd (нM) 2,97E-4 0,76 (1,11E-5) (0,12) 1,77E-4 0,64 (1,34E-5) (0,09) 2,54E-4 1,33 (3,46E-6) (0,00) 2,78E-4 2,69 (3,61 E-6) (0,02) 1,49E-4 0,57 (1,61E-5) (0,07) HcLc Ka Kd 3,03E5 (4,58E3) 5,47E5 (1,20E4) 3,02E5 (9,90E2) 1,74E5 (2,19E3) 1,32E6 (2,71 E5) 1,41E-3 (2,84E-5) 1,10E-3 (2,86E-5) 1,36E-3 (7,92E-5) 1,29E-3 (7,64E-5) 7,00E-4 (3,18E-5) KD (нM) 4,66 (0,08) 2,01 (0,07) 4,51 (0,28) 7,45 (0,34) 0,54 (0,09) хвилі 214 нм та 280 нм SDS-PAGE виконували на 4-20 % гелі Novex Трис-HCL, завантажуючи 10 мкг продукту та забарвлюючи Sypro Ruby. C-IEF виконували на Beckman MDQ, застосовуючи pH 3,5 -10 амфоліній. Були одержані наступні результати 49 96279 50 Таблиця 20 Аналіз антитіл анти-NOGO-А за допомогою ексклюзійної (ЕК) ВЕРХ Показані значення являють собою відсотковий вміст антитіла, призначеного для кожного з трьох різних видів Антитіло H28L16 H20L16 Агрегат % 0,50 14,21 Мономер % 99,50 85,75 Фрагмент % 0,00 0,05 Таблиця 21 Аналіз SDS-PAGE антитіл анти-NOGO-А Показані значення являють собою відсотковий вміст антитіла, знайденого в основних смугах Антитіло Невідновлені H28L16 82,4 % H20L16 84,6 % Дані ЕК-ВЕРХ свідчать, що H20L16 є більш сприйнятливим до агрегації, ніж H28L16 (H28L16). Якщо дані, про які повідомляють в даному опису, необхідно буде відтворити у великому масштабі, це вплине на здатність виробничого способу продукувати матеріал прийнятної якості для клінічного застосування (> 95 % мономеру). Дані SDS-PAGE показують, що обидва кандидати прийнятні, демонструючи типовий профіль. Приклад 13: Порівняння H28L16 з 11C7 Мишаче анти-NOGO-A антитіло, позначене як 11C7, яке підвищилося до епітопу пептиду, описано в WO 2004052932. Химерне 11C7 створено, базуючись на інформації послідовності, запропонованій у WO 2004052932. З метою порівняння епітопів зв'язування 2A10 та 11C7 було здійснено конкурентний метод ELISA для того, щоб дослідити, чи розпізнають 11C7 та 2A10 накладення епітопу на NOGO-A. Як показано на фіг. 20 нижче, HcLc (химерна форма 2A10) могла конкурувати з 2A10 за зв'язування з рекомбінантним NOGO-A людини, тоді як 11C7 не демонструвало конкуренції з 2A10, навіть в концентраціях до 100 мкг/мл. Приклад 14: Конкуренція ELISA з метою демонстрації здатності пептидів безпосередньо конкурувати з NOGO-5+6 людини за зв'язування з NOGO H28L16 Спосіб конкурентного методу ELISA Застосовуючи конкурентний метод ELISA, оцінили здатність пептидів конкурувати безпосередньо з NOGO-A (GST-людським Nogo-A56) за зв'язування з NOGO H28L16. Анти-людським IgG кролика (Sigma, #1-9764) при 5 г/мл в бікарбонатному буфері вкривали планшети Nunc immunosorp (100 мкл на лунку) протягом ночі при 4 °C. Планшети промивали тричі буферним розчином, що містить 0,05 % Твін (TBST), потім блокували 1 % сироваткою телячого альбуміну в TBST при кімнатній температурі протягом 1 години. Далі H28L16 наносили на планшет (1 мкг/мл, розбавлені в 1 % сироватці телячого альбуміну в TBST, 50 мкл на Відновлені HC:67,2 % LC:27,7 % H+L: 94,9 % HC: 69,3 % LC: 26,4 % H+L: 95,7 % лунку) при кімнатній температурі протягом 1 години. Планшети промивали тричі TBST Пептиди (з 0 до 100 г/мл) та GST-людський NOGO-A56 в концентрації 1 мкг/мл (розбавлені в 1 % сироватці телячого альбуміну в TBST) попередньо перемішували до того, як додати в лунки, та інкубували при кімнатній температурі протягом 1 години. Планшети промивали тричі TBST, потім інкубували протягом 1 години з кон'югатом анти-GST пероксидази кролика (Sigma, #A7340, 1:2000, розбавлений в 1 % сироватці телячого альбуміну в TBST) протягом 1 години. Планшети промивали тричі TBST, а потім інкубували з субстратом пероксидази 50 I OPD (Sigma) на лунку протягом 10 хвилин. Кольорову реакцію зупиняли, додаючи 25 I концентрованої H2SO4. Визначали поглинання на довжині хвилі 490 нМ, застосовуючи показання планшета. Результати, показані на фіг. 21, підтверджують, що пептиди 6 та 7, які були позитивними в побудові карти епітопів за допомогою ELISA (Приклад 8), можуть конкурувати з GST-людським NOGO-A56, зв'язаним з H28L16. Це свідчить про той факт, що пептиди, які були позитивні при вивченні побудови карти епітопів, містять епітоп для зв'язування з H28L16. Пептиди 16 та 17 (які містять пептиди NOGO, але не перекриваються з пептидами 6 або 7), які не містять запропонованого епітопу, не конкурують з NOGO-5+6. Приклад 15: Аналіз ELISA гуманізованого моноклонального анти-NOGO антитіла на базі варіанту NOGO антитіла G101S/O37R G101S (також відомий як H100 (SEQ ID NO: 63)), модифікований варіант варіабельної ділянки H6 важкого ланцюга (SEQ ID NO: 11) генерували шляхом введення єдиної заміни G101S (за нумерацією Kabat) в CDR H3, як описано вище. Аналогічно, Q37R, модифікований варіант варіабельної ділянки L13 легкого ланцюга (SEQ ID NO: 13) генерували шляхом введення єдиної заміни (за нумерацією Kabat Q37R) в каркасну ділянку (з метою утворення L100). Послідовність білка варіабельної 51 96279 легкої ділянки Q37R наведена в SEQ ID NO: 67. Гени, що кодують версії з повною довжиною важких і легких ланцюгів, що містять заміни G101S/Q37R, експресували в клітинах CHO, як описано раніш, та аналізували при прямому зв'язуванні ELISA, як описано раніш. У фіг. 22 показані результати прямого зв'язування ELISA, коли антиген завантажують в конце 52 нтрації 0,05 мкг/мл. Дані підтверджують, що антитіло H100L100 демонструє порівнянну активність зв'язування з рекомбінантним GST-людського NOGO-A56 відносно H27L16, і що H100L100 має покращений профіль зв'язування в порівнянні з H6L13. Відповідні значення EC50 показані в таблиці нижче: Таблиця 22 Вимірювання EC50 для варіанту G101S/Q37R в порівнянні з H6L13 та H27L16 Антитіло H6L13 H27L16 H100/L100 Приклад 16: Аналіз BiaCore гуманізованих моноклональних анти-NOGO антитіл, базуючись на варіанті G101S CDR H3 H 100, модифікований варіант варіабельної ділянки важкого ланцюга H6 (SEQ ID NO: 11) генерували шляхом введення єдиної заміни G101 S (за нумерацією Kabat) в CDR H3. Послідовність білка H100 варіабельної ділянки важкого ланцюга наведена в SEQ ID NO: 63. Аналогічно, L100 та L101, модифіковані варіанти варіабельної ділянки легкого ланцюга L13 (SEQ ID NO: 13) генерували шляхом введення єдиної заміни (за нумерацією Kabat Q37R та Q45R відповідно) в каркасну ділянку. Послідовності білка варіабельних легких ділянок біл Показник EC50 0,086 0,052 0,048 ків L100 та L101 дані в SEQ ID NO: 67 та SEQ ID NO: 68 відповідно. Версії повної довжини H100L100 та H100L101 експресували в клітинах CHO, як описано раніше. Табл. 23 демонструє порівняння спорідненості зв'язування H6L13 з H100L100 та 00L101, і показує, що H100L100 та 0L101 мають покращену спорідненість зв'язування в порівнянні з H6L13. В даному прикладі спосіб здійснювали в основному, як описано в Прикладі 6, де осколок CM5 був активізований, пропускаючи розчини NHS та EDC над даним осколком в концентрації 5 мкг/мл протягом 7 хвилин, а NOGO суспендували в буфері, що містить 10 нM натрію ацетату (pH 4,5) до проходження над осколком. Таблиця 23 Вимірювання Biacore для варіантів G101S варіабельного важкого ланцюга H6 в комбінації з варіантами варіабельного легкого ланцюга L13 в порівнянні з H6L13 Антитіло H6L13 H100L100 H100L101 Зв'язування ka (1/мс) 1,04E+06 1,28E+07 1,30E+07 Дисоціація kd (1/с) 7,22E-03 5,07E-03 4,29E-03 Спорідненість (KD, нM) 6,97 0,396 0,329 53 96279 54 Таблиця 24 Короткий опис послідовностей NOGO антитіла Опис 2A10,CDR-H1 2A10, CDR-H2 2A10, CDR-H3 2A10, CDR-L1 2A10, CDR-L2 2A10, CDR-L3 2A10,VH (мишачий) 2A10,VL (мишачий) Химерний важкий ланцюг Hс Химерний легкий ланцюг Lc Гуманізований конструкт H6 VH 2A10 Гуманізований конструкт H16 VH 2A10 Гуманізований конструкт L13 VL 2A10 Гуманізований конструкт L16 VL 2A10 Гуманізований конструкт важкого ланцюга H6 2A10 Гуманізований конструкт важкого ланцюга H16 2A10 Гуманізований конструкт легкого ланцюга L13 2A10 Гуманізований конструкт легкого ланцюга L16 2A10 Лідерна послідовність Campath Амінокислоти 586-785 NOGO A людини (NOGO-A56), злитого з GST Гуманізований конструкт H1 VH 2A10 Гуманізований конструкт L11 VL 2A10 Гуманізований конструкт важкого ланцюга H1 2A10 Гуманізований конструкт легкого ланцюга L11 2A10 Гуманізований конструкт H20 VH 2A10 Гуманізований конструкт важкого ланцюга H20 2A10 2A10, CDR-H3 (G95M) Послідовність фрагменту NOGO-A мавпи Гуманізований конструкт H26 VH Гуманізований конструкт H27 VH Гуманізований конструкт H28 VH Гуманізований конструкт важкого ланцюга H26 Гуманізований конструкт важкого ланцюга H27 Гуманізований конструкт важкого ланцюга H28 Химерний важкий ланцюг Hс (G95M) Епітоп Гуманізований конструкт H99 VH 2A10 CDR (G101S) Гуманізований конструкт H100 VH Гуманізований конструкт H101 VH Гуманізований конструкт H102 VH Гуманізований конструкт H98 VH L100(L13+Q37R) L101 (L13+Q45R) L102(L13+Q37R/Q45R) L103(L16+Q37R) L104(L16+Q45R) L105(L16+Q37R/Q45R) Пептид Пептид Аналог CDR H3 Аналог CDR H3 Аналог CDR H3 Ідентифікатор послідовності (SEQ ID NO) Послідовність аміно- Послідовність полікислот нуклеотидів 1 2 3 4 5 6 7 19 8 20 9 21 10 22 11 23 12 24 13 25 14 26 15 27 16 28 17 29 18 30 31 32 33 34 35 36 41 42 45 46 47 48 49 53 54 55 59 60 61 62 63 64 65 66 67 68 69 70 71 72 73 74 75 76 77 37 38 39 40 43 44 50 51 52 56 57 58 55 96279 56 Таблиця 24 Короткий опис послідовностей NOGO антитіла Ідентифікатор послідовності (SEQ ID NO) Послідовність аміно- Послідовність полікислот нуклеотидів 78 79 80 8182 83 84 85 86 87 88 89 90 91 92 93 94 95 96 Опис Аналог CDR H3 Аналог CDR H3 Аналог CDR H3 Аналог CDR H3 Аналог CDR H3 Аналог CDR H3 Аналог CDR H3 Пептид NOGO Аналог CDR H3 Аналог CDR H3 Аналог CDR H3 Аналог CDR H3 Аналог CDR H3 NOGO (A56) біличої мавпи плюс мітка GST NOGO (A56) яванської макаки плюс мітка GST NOGO (A56) мавпи плюс мітка GST NOGO (A56) щура плюс мітка GST Пептид NOGO людини Пептид NOGO людини Послідовності SEQ ID NO: 7: 2A10, VH (мишачий) SEQ ID NO: 8: 2A10, VL (мишачий) SEQ ID NO: 9: Химерний важкий ланцюг Hc 57 96279 58 SEQ ID NO: 10: Химерний легкий ланцюг Lc SEQ ID NO: 11: Гуманізований конструкт H6 VH 2A10 SEQ ID NO: 12: Гуманізований конструкт H16 VH 2A10 SEQ ID NO: 13: Гуманізований конструкт L13 VL 2A10 SEQ ID NO: 14: Гуманізований конструкт L16 VL 2A10 SEQ ID NO: 15: Гуманізований конструкт важкого ланцюга H6 2A10 SEQ ID NO: 16: Гуманізований конструкт важкого ланцюга H16 2A10 SEQ ID NO: 17: Гуманізований конструкт легкого ланцюга L13 2A10 59 96279 60 SEQ ID NO: 18: Гуманізований конструкт легкого ланцюга L16 2A10 SEQ ID NO: 19: Послідовність нуклеотидів, що кодує 2A10, VH (мишачий)SEQ ID: 7 SEQ ID NO: 20: Послідовність нуклеотидів, що кодує 2A10, VL (мишачий)SEQ ID: 8 SEQ ID NO: 21: Послідовність нуклеотидів, що кодує химерний важкий ланцюг Hc SEQ ID: 9 SEQ ID NO: 22: Послідовність нуклеотидів, що кодує химерний легкий ланцюг Lc SEQ ID: 10
ДивитисяДодаткова інформація
Назва патенту англійськоюIsolated antibody or fragment thereof that is capable of binding to human nogo
Автори англійськоюClegg Stephanie Jane, Ellis Jonathan Henry, Germaschewski Volker, Hamblin, Paul, Andrew, Kopsidas George, Mcadam Ruth, Prinjha Rabinder Kumar
Назва патенту російськоюИзолированное антитело или его фрагмент, способное к связыванию с человеческим nogo
Автори російськоюКлигг Стефания Джейн, Эллис Джонатан Генри, Гермащевски Волкер, Амбли Пол Эндрю, Коспидас Георг, Макадам Рус, Принйоха Рабиндер Кумар
МПК / Мітки
МПК: A61K 39/395, C12N 5/10, C07K 16/18, C12N 15/63, C12N 15/13, A61P 25/00, C12P 21/08
Мітки: людським, антитіло, ізольоване, фрагмент, здатне, зв'язування
Код посилання
<a href="https://ua.patents.su/54-96279-izolovane-antitilo-abo-jjogo-fragment-zdatne-do-zvyazuvannya-z-lyudskim-nogo.html" target="_blank" rel="follow" title="База патентів України">Ізольоване антитіло або його фрагмент, здатне до зв’язування з людським nogo</a>
Моноклональне антитіло, яке спричиняє апоптоз кров’яних клітин, що містять ядро та інтегрин-пов’язаний протеїн людини (іпп), фрагмент моноклонального антитіла, гібридома, яка продукує моноклональне антитіло (ва

Номер патенту: 72192
Опубліковано: 15.02.2005
Автори: Фукушіма Наоші, Уно Шінсуке
МПК: C12N 5/20, C12P 21/08, C07K 16/28
Мітки: моноклональне, клітин, яке, інтегрин-пов'язаний, фрагмент, апоптоз, гібридома, ва, яка, антитіла, ядро, кров'яних, містять, продукує, моноклонального, людини, спричиняє, іпп, протеїн, антитіло
Формула / Реферат:
1. Моноклональне антитіло, яке зв’язується із інтегрин-пов’язаним протеїном людини (ІПП) і спричиняє апоптоз кров'яних клітин, що містять ядро та ІПП.2. Моноклональне антитіло за п.1, яке відрізняється тим, що антитіло отримують способом, що включає:а) імунізацію тварин з використанням ІПП людини як сенсибілізуючого антигена,б) одержання гібридом;в) відбір антитіл, що розпізнають ІПП людини та спричиняють апоптоз...
Химерне антитіло проти cd40 людини, молекула нуклеїнової кислоти, вектор експресії, фармацевтична композиція для лікування захворювань, опосередкованих т-клітинами, яка містить химерне антитіло

Номер патенту: 71909
Опубліковано: 17.01.2005
Автори: Херріс Лінда Дж., Торн Барбара А., Бейорет Юрген, Беррі Карен К., Холленбау Даєн, Хьюз Уільям Д., Сайдек Ентоні В., Уоткінс Джеффрі Д., Аруффо Аледжандро А., Ву Херрен
МПК: A61P 29/00, C12N 15/13, A61P 37/06, A61P 43/00, C07K 16/28, C12N 15/09, C07K 14/725, C07K 14/46, A61P 37/02, A61P 19/02, A61K 39/395, C12N 1/21
Мітки: людини, т-клітинами, вектор, опосередкованих, кислоти, експресії, молекула, антитіло, яка, композиція, нуклеїнової, химерне, лікування, фармацевтична, захворювань, містить
Формула / Реферат:
1. Варіабельна ділянка легкого ланцюга химерного антитіла, що зв’язується з CD40 людини, яка містить амінокислотну послідовність SEQ ID NО:1 (Фіг. 4а).2. Варіабельна ділянка важкого ланцюга химерного антитіла, що зв’язується з CD40 людини, яка містить амінокислотну послідовність SEQ ID NО:2 (Фіг. 4b).3. Химерне антитіло, що зв'язується з CD40 людини, яке містить легкий і важкий ланцюг, причому зазначений легкий ланцюг містить ...
Моноклональне антитіло, химерне антитіло, повністю гуманізоване антитіло, антитіло до антиідіотипічного антитіла або його фрагмент до рецептора ліпопротеїнів низької щільності (лпнщ) людини, спосіб їх одержання

Номер патенту: 78489
Опубліковано: 10.04.2007
Автори: Бельзер Ілана, Суісса Дані, Дреано Мішель, Смоларскі Моше, Йонах Начум, Антонетті Франческо
МПК: C12P 21/08, C07K 16/28, C12N 5/20, G01N 33/577, G01N 33/68
Мітки: фрагмент, лпнщ, одержання, людини, антиідіотипічного, гуманізоване, моноклональне, низької, антитіло, повністю, щільності, антитіла, химерне, рецептора, спосіб, ліпопротеїнів
Формула / Реферат:
1. Моноклональне антитіло, химерне антитіло, повністю гуманізоване антитіло, антитіло до антиідіотипічного антитіла або його фрагмент, що специфічно розпізнає і зв'язує розчинний рецептор ліпопротеїнів низької щільності (ЛПНЩ)людини і його фрагменти, а також здатне інгібувати реплікацію вірусу гепатиту С, і яке одержують шляхом імунізації тварин формою +291 розчинного рецептора ЛПНЩ людини, тобто формою розчинного рецептора людини, що містить...
Химерне антитіло, що специфічно зв’язується з людським il-6, та фармацевтична композиція, що його містить

Номер патенту: 85995
Опубліковано: 25.03.2009
Автори: Джайлз-Комар Джилл, Тріха Мохіт, Перітт Девід, Найт Девід
МПК: C07K 16/24, A61P 35/00, C07K 16/46, A61P 37/00, A61K 39/395
Мітки: фармацевтична, специфічно, людським, композиція, химерне, антитіло, зв'язується, il-6, містить
Формула / Реферат:
1. Химерне антитіло або його фрагмент, що специфічно зв'язується з людським IL-6 з високою афінністю зв'язування, яке містить послідовність варіабельної ділянки важкого ланцюга SEQ ID NO: 7 та послідовність варіабельної ділянки легкого ланцюга SEQ ID NO: 8, а також послідовність константної ділянки людського антитіла.2. Антитіло або його фрагмент за п. 1, яке відрізняється тим, що зв'язується з людським IL-6 з афінністю (Kd) 10-11...
Виділене антитіло або фрагмент антитіла, що специфічно зв’язуються з фрагментом поліпептиду rg1, імунокон’югат, який включає виділене антитіло або фрагмент антитіла, способи вибіркового руйнування клітини, ліку

Номер патенту: 85527
Опубліковано: 10.02.2009
Автори: Перрі Гордон, Паркес Дебора, Харкінс Річард, Штейнбрехер Ренате, Шнайдер Дуглас В.
МПК: A61P 13/08, C07K 16/18, C12N 1/19, A61P 35/04, G01N 33/53, C07K 14/47, C12N 5/10, A61K 39/395, C12N 15/09
Мітки: включає, фрагментом, антитіло, специфічно, антитіла, імунокон'югат, руйнування, поліпептиду, виділене, способи, фрагмент, вибіркового, зв'язуються, ліку, клітині
Формула / Реферат:
1. Виділене антитіло або фрагмент антитіла, що специфічно зв'язуються з фрагментом поліпептиду RG1, вибраним із групи, яка складається з:(а) біологічно або імунологічно активного фрагменту поліпептиду, що має амінокислотну послідовність, яка відповідає частині, але не всій амінокислотній послідовності, представленій на фіг. 2 (SEQ ID NO: 2);(б) фрагменту поліпептиду, який складається з амінокислот 28-46 послідовності,...
Попередній патент: Застосування флібансерину для лікування розладів статевого потягу в передклімактеричний період
Наступний патент: Похідні бензамідазолу та їх застосування для модуляції рецепторного комплексу гамкa
Випадковий патент: Спосіб виготовлення просочених нітридкремнієвих виробів