Спосіб дослідження дихання мітохондрій ацинарних клітин підшлункової залози in situ
Номер патенту: 71234
Опубліковано: 10.07.2012
Автори: Манько Богдан Олексійович, Клевець Мирон Юрійович, Манько Володимир Васильович





Формула / Реферат
Спосіб дослідження дихання мітохондрій ацинарних клітин підшлункової залози in situ, за яким отримують суспензію ізольованих ацинусів підшлункової залози, замінюють середовище виділення на середовище інкубації, пермеабілізують плазматичну мембрану клітин неіонним детергентом - дигітоніном, контролюють ступінь проникності плазматичної мембрани і полярографічно реєструють швидкість поглинання кисню, який відрізняється тим, що як основний осмотичний компонент середовища інкубації використовують сахарозу.
Текст
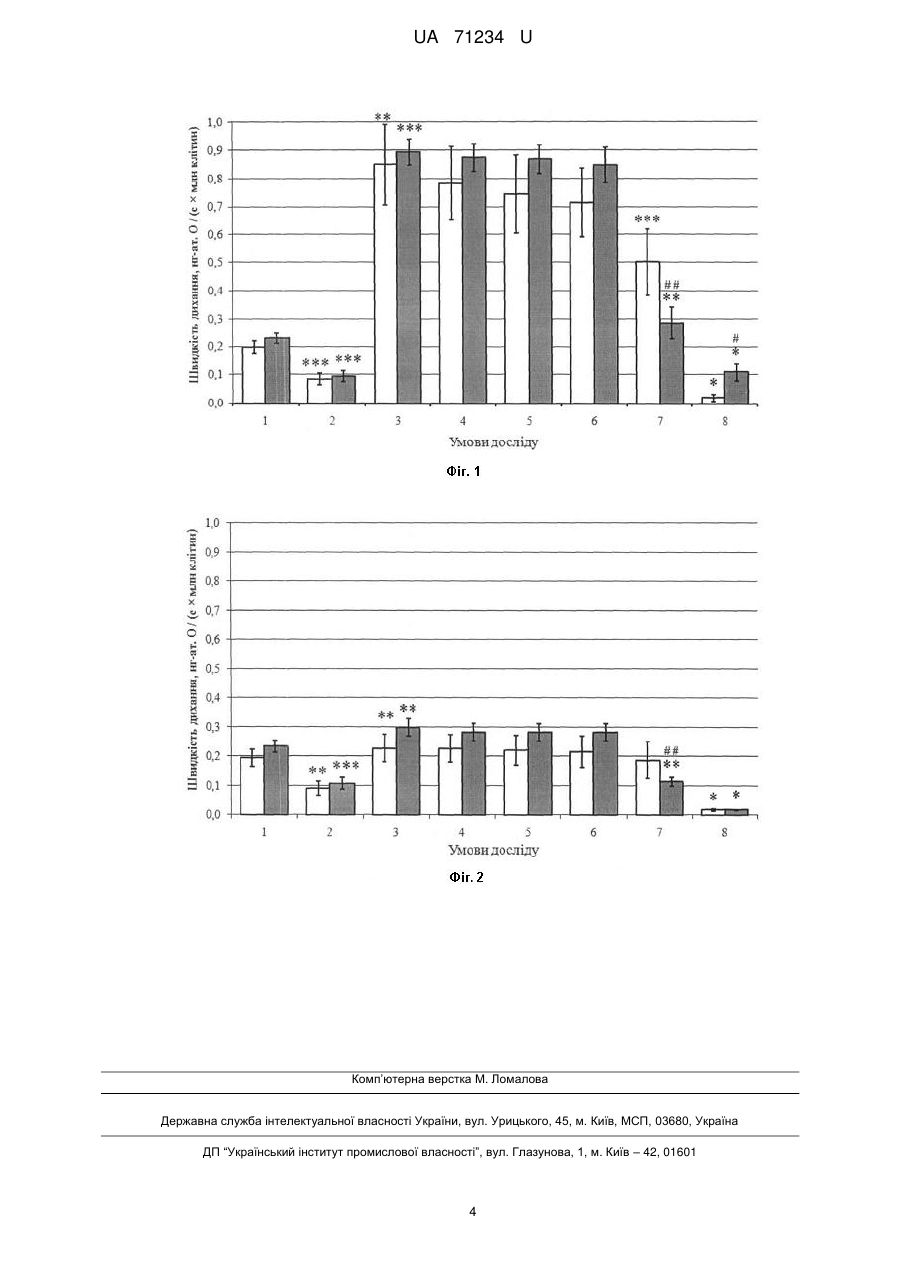
Реферат: Спосіб дослідження дихання мітохондрій ацинарних клітин підшлункової залози in situ, за яким отримують суспензію ізольованих ацинусів підшлункової залози, замінюють середовище виділення на середовище інкубації, пермеабілізують плазматичну мембрану клітин неіонним детергентом - дигітоніном, контролюють ступінь проникності плазматичної мембрани і полярографічно реєструють швидкість поглинання кисню. Як основний осмотичний компонент середовища інкубації використовують сахарозу. UA 71234 U (54) СПОСІБ ДОСЛІДЖЕННЯ ДИХАННЯ МІТОХОНДРІЙ АЦИНАРНИХ КЛІТИН ПІДШЛУНКОВОЇ ЗАЛОЗИ IN SITU UA 71234 U UA 71234 U 5 10 15 20 25 30 35 40 45 50 55 60 Корисна модель належить до галузі біології, а саме клітинної фізіології та може бути використана для дослідження процесів мітохондріального окислення ацинарних клітин в умовах, максимально наближених до фізіологічних. Відомий спосіб дослідження дихання пермеабілізованих волокон скелетних м'язів із застосуванням детергентів сапоніну чи дигітоніну (Saks V. A. et al. Permeabilised cell and skinned fiber techniques in studies of mitochondrial function in vivo // Моl. Cell. Biochem.-1998. - Vol. 184. - P. 81-100). Пермеабілізація здійснюється для забезпечення проникнення екзогенних модифікаторів мітохондріального дихання, які не проникають крізь цілісну плазматичну мембрану. Ними можуть бути субстрати чи регулятори ферментів циклу Кребса, аденілові нуклеотиди, інгібітори комплексів дихального ланцюга тощо. Цей спосіб дає змогу зберегти взаємозв'язки між мітохондріальним диханням та іншими клітинними процесами. Недоліком способу є відсутність системного підходу до визначення оптимальної концентрації детергенту, оскільки у недостатніх концентраціях дигітонін пермеабілізує плазматичну мембрану не всіх клітин, а у надмірних - може ушкоджувати мембрани мітохондрій і ендоплазматичного ретикулуму (Cook G. A. et al. Structural changes of isolated hepatocytes during treatment with digitonin // Biochim. Biophys. Acta.-1983. - Vol. 763. - P. 356-367). Найближчим за технічною суттю - прототипом - є "Спосіб дослідження дихання мітохондрій in situ" (Патент UA 60528 Манько В. В., Манько Б. О., Мерлавський В. М., Великопольська О. Ю. Спосіб дослідження дихання мітохондрій in situ). За цим способом отримують суспензію ізольованих клітин, замінюють середовище виділення на середовище інкубації, близьке за іонним складом до цитозолю, після чого пермеабілізують плазматичну мембрану клітин неіонним детергентом - дигітоніном. При цьому пермеабілізацію здійснюють оптимальною кількістю детергенту, яка знаходиться у сталому співвідношенні з кількістю досліджуваних клітин, яке для кожної тканини і для кожного типу клітин визначають емпірично. Опісля контролюють ступінь проникності плазматичної мембрани і полярографічно реєструють швидкість поглинання кисню. Недоліком способу є низький ступінь спряження між процесами дихання та окисного ˉ фосфорилювання, що є наслідком використання середовища із високою концентрацією Сl . В основу корисної моделі поставлено задачу удосконалити спосіб дослідження дихання мітохондрій ацинарних клітин підшлункової залози in situ шляхом використання оптимального середовища інкубації, що дасть змогу підвищити ступінь наближеності умов досліду до фізіологічних та досягнути кращого спряження між диханням і окисним фосфорилюванням. Поставлена задача вирішується так, що у способі дослідження дихання мітохондрій ацинарних клітин підшлункової залози in situ, за яким отримують суспензію ізольованих ацинусів підшлункової залози, замінюють середовище виділення на середовище інкубації, пермеабілізують плазматичну мембрану клітин неіонним детергентом - дигітоніном, контролюють ступінь проникності плазматичної мембрани і полярографічно реєструють швидкість поглинання кисню, де як основний осмотичний компонент середовища інкубації використовують сахарозу. При дослідженні дихання ізольованих мітохондрій різних тканин, в тому числі й підшлункової залози, здебільшого використовують середовища, основним осмотичним компонентом яких є сахароза (Wilson J. S., Korsten Μ. A. and Lieber C. S. The isolation and properties of mitochondria from rat pancreas // Biochem. and Biophys. Research Communications.-1984. - Vol. 121, № 2. - P. 545-551). Натомість, пермеабілізовані ацинарні панкреацити зазвичай інкубують у сольових ˉ середовищах з основним компонентом KСl, а концентрація Сl становить, в середньому, 150 2+ ммоль/л (Krause E., Gobel Α., Schulz I. Cell side-specific sensitivities of intracellular Ca stores for inositol 1,4,5-trisphosphate, cyclic ADP-ribose, and nicotinic acid adenine dinucleotide phosphate in permeabilized pancreatic acinar cells from mouse // J Biol. Chem.-2002. - Vol. 277, № 14.- P. 1169611702; Gerasimenko Yu., Lura G., Sherwood M. W. et al. Pancreatic protease activation by alcohol 2+ metabolite depends on Ca release via acid store IP3 receptors // PNAS-2009. - Vol. 106, № 26. - P. ˉ 10758-10763). Це значно більше, ніж [Сl ] у цитозолі ацинарних клітин підшлункової залози в стані спокою, що становить, за різними даними, 19 ммоль/кг мокрої ваги (Sasaki S., Nakagaki I., Hori S., Kondo H. Changes in element concentrations induced by agonist in pig pancreatic acinar cells // Pflügers Archiv Europ. J. of Physiol.-1996. - Vol. 432, № 3. - P. 538-545), 65 ммоль/л (Zhao H., + + ˉ Muallem Sh. Na , K , and Сl transport in resting pancreatic acinar cells // J. Gen. Physiol.-1995. - Vol. 106. - P. 1225-1242), або 53,7±5 ммоль/л (Preissler M., Williams J. A. Pancreatic acinar cell function: measurement of intracellular ions and pH and their relation to secretion // J. Physiol.-1981 - Vol. 321. ˉ P. 437-448). Підвищена концентрація Сl створює неадекватні умови для перебігу фізіологічних процесів та негативно впливає на мітохондріальні окисні процеси. Доведено, що мітохондрії, ізольовані у середовищі з KСl, мають нижчий вміст кардіоліпіну, нижчий рівень спряження між 1 UA 71234 U 5 10 15 20 25 30 35 40 45 50 55 диханням та окисним фосфорилюванням та схильні до патологічного набрякання (Hogeboom G. H., Schneider W. C., Palade G. E. Cytochemical studies of mammalian tissues; isolation of intactmitochondria from rat liver; some biochemical properties of mitochondria and submicroscopic particulate material // J. Biol. Chem.-1948. - Vol. 172. - P. 619-635; Corcelli Α., Saponetti M. S., Zaccagnino P., Lopalco P., Mastrodonato M., Liquori G. E., Lorusso M. Mitochondria isolated in nearly isotonic KCl buffer: Focus on cardiolipin and organelle morphology // Biochimica et Biophysica Acta.2010. - Vol. 1798. - P. 681-687). Використання сахарози, на відміну від KСl, дозволяє уникнути ˉ негативного впливу високої концентрації Сl . Автори корисної моделі вперше запропонували використовувати сахарозу як основний компонент середовища інкубації при дослідженні дихання мітохондрій ацинарних клітин підшлункової залози in situ. Це дало змогу наблизити умови досліду до фізіологічних умом та досягнути кращого спряження між диханням і окисним фосфорилюванням. Фіг. 1. Вплив середовища інкубації на кінетику поглинання кисню пермеабілізованими ацинарними панкреацитами за окислення сукцинату у середовищі з KСl (□) або з сахарозою ( ) за умов: 1 - дихання інтактних клітин, 2 - додано дигітонін, 3 - додано сукцинат, 4 - додано 0,1 ммоль/л АДФ, 5 - додано 0,5 ммоль/л АДФ, 6 - додано 1 ммоль/л АДФ, 7 - додано олігоміцин, 8 додано TTFA; де: * - статистично вірогідна різниця порівняно з попереднім показником у межах однієї дослідної групи (Р0,05); ** - статистично вірогідна різниця порівняно з попереднім показником у межах однієї дослідної групи (Р0,01); *** - статистично вірогідна різниця порівняно з попереднім показником у межах однієї дослідної групи (Р0,001); # - статистично вірогідна різниця порівняно із аналогічним показником за використання середовища із КСl (Р0,05); ## - статистично вірогідна різниця порівняно із аналогічним показником за використання середовища із KСl (Р0,01). Фіг. 2. Вплив середовища інкубації на кінетику поглинання кисню пермеабілізованими ацинарними панкреацитами за окислення суміші малату з піруватом у середовищі з KСl (□) або з сахарозою ( ) за умов: 1 - дихання інтактних клітин, 2 - додано дигітонін, 3 - додано піруват і малат, 4 - додано 0,1 ммоль/л АДФ, 5 - додано 0,5 ммоль/л АДФ, 6 - додано 1 ммоль/л АДФ, 7 додано олігоміцин, 8 - додано ротенон; де умовні позначення як на Фіг. 1. Запропонований спосіб здійснюється так. Дослідження проводять на суспензії ізольованих панкреатичних ацинусів щурів, яку отримують способом Вільямса (Williams J. Α., Korc M., Dormer R. L. Action of secretagogues on a new preparation of functionally intact, isolated pancreatic acini // Am. J. Physiol.-1978. - Vol. 235, № 5. - P. 517-24). Ацинуси ізолюють у середовищі виділення, що має такий склад, ммоль/л: NaCl 140,0; KСl - 4,7; СаСl2 - 1,3; MgCl2 - 1,0; HEPES - 10,0; глюкоза - 10,0; рН 7,4. Крім того, середовище містить бичачий сироватковий альбумін (2,5 мг/мл) та соєвий інгібітор трипсину (0,1 мг/мл). Для перевірки цілісності плазматичних мембран клітини фарбують 0,1 % розчином трипанового синього. Кількість клітин з цілісними плазматичними мембранами повинна становити не менше 95 %. Підрахунок ацинарних панкреацитів здійснюють за допомогою камери Горяєва. Суспензію ізольованих ацинусів центрифугують, надосадову рідину замінюють на середовище інкубації пермеабілізованих ацинарних панкреатитів, що містить, ммоль/л: 2+ сахароза - 250,0; КН2РО4 - 2,0; MgCl2 - 1,0; ЕГТА - 1,0; СаСl2 - 0,28 (концентрація вільного Са 100 нмоль/л); HEPES - 10,0; соєвий інгібітор трипсину - 0,1 мг/мл; рН 7,2. Швидкість поглинання кисню визначають полярографічним методом. Величину дифузного струму реєструють за допомогою полярографічної установки, зібраної на базі закритого електрода Кларка, полярографа YSI 5300, самописця КСП-4, магнітної мішалки для розмішування суспензії та скляної термостатованої закритої комірки об'ємом 1,6 мл. Температура інкубації становить 37 °C. Після внесення клітин у полярографічну комірку під час реєстрації поглинання кисню 6 почергово додають наступні речовини: дигітонін у концентрації 50 мкг у розрахунку на 10 клітин, субстрат ФАД-залежної дегідрогенази сукцинат 5 ммоль/л або суміш субстратів НАДзалежних дегідногеназ піруват і малат по 5 ммоль/л, АДФ у зростаючих концентраціях 0,1, 0,5 і + 1 ммоль/л, блокатор Н -АТФ-синтази олігоміцин 6 мкмоль/л та блокатор комплексів дихального ланцюга ротенон 10 мкмоль/л або теноїлтрифлуорацетон (TTFA) 20 мкмоль/л. Значення коефіцієнтів дихального контролю розраховують як відношення швидкості дихання, стимульованого субстратами, і АДФ до швидкості дихання, заблокованого олігоміцином. 2 UA 71234 U 5 10 15 20 25 30 35 40 Необхідні статистичні розрахунки проводять за допомогою комп'ютера з використанням пакету програм Microsoft Office Excel. Переваги способу можна продемонструвати на такому прикладі. Як альтернативу до запропонованого середовища інкубації використовували KСl-вмісне середовище такого складу, ммоль/л: KСl - 127,0; NaCl - 20,0; KН2РО4 - 2,0; MgCl2 - 1,0; ЕГТА 2+ 1,0; СаСl2 - 0,28 (концентрація вільного Са - 100 нмоль/л); HEPES - 10,0; соєвий інгібітор трипсину - 0,1 мг/мл; рН 7,2. Дихання інтактних ацинарних панкреатитів складало 0,20±0,0021 нг-ат. О / (с×млн клітин) у середовищі з KСl і не відрізнялось від дихання у середовищі із сахарозою (n=5). Пермеабілізація плазматичної мембрани дигітоніном супроводжувалась заміною іонного складу цитозолю та виходом розчинних цитозольних факторів із клітин, що й приводило до статистично вірогідного зниження швидкості дихання вже через 3-5 хв. після внесення дигітоніну (Фіг. 1, 2, n=5, Р0,001). Далі у комірку вносили сукцинат (Фіг. 1) або суміш малату з піруватом (Фіг. 2), які статистично вірогідно підвищували швидкість дихання (n=5, Р0,01). При цьому, швидкість дихання, стимульованого субстратами, не залежала від складу середовища. АДФ не стимулював дихання ні у низьких, ні у високих концентраціях (Фіг. 1, 2, n=5). Це свідчить або про наявність високих його ендогенних запасів, або про порушення спряження між диханням і окисним фосфорилюванням. + Для перевірки цих гіпотез вносили блокатор H -АТФ-синтази олігоміцин. Олігоміцин значно сильніше пригнічує дихання пермеабілізованих панкреацитів у середовищі зі сахарозою, ніж з KСl як за окислення сукцинату (Фіг. 1, n=5, Р0,01), так і суміші малату з піруватом (Фіг. 2, n=5, Р0,01). Це підтверджує гіпотезу про високий рівень ендогенного АДФ у пермеабілізованих клітинах та свідчить про краще спряження між диханням і окисним фосфорилюванням за використання сахарози. За окислення сукцинату при застосуванні середовища із KСl коефіцієнт дихального контролю становив 1,51±0,14, а при застосуванні середовища зі сахарозою 3,52±0,80 (n=5, Р0,05). Аналогічно, за окислення НАД-залежних субстратів коефіцієнт дихального контролю становив 1,34±0,15 і 2,58±0,18 за відповідних середовищ (n=5, Р0,001). Після подальшого внесення у полярографічну комірку TTFA на фоні сукцинату (Фіг. 1) або ротенону на фоні пірувату і малату (Фіг. 2) швидкість дихання у всіх дослідних пробах знизилась практично до нуля (Р0,05, n=5). Це ще раз підтверджує інтактність НАД-залежних та ФАДзалежних шляхів окислення та свідчить про мінорний вклад немітохондріального дихання у досліджені показники швидкості поглинання кисню за умов in situ. Дисперсія даних була значно меншою для дослідних проб у середовищі із сахарозою: статистично вірогідна різниця дисперсій за Фішером за окислення сукцинату (Р0,05) і за окислення суміші малату з піруватом після додавання олігоміцину (Р0,01). Оскільки при використанні сахарози як основного осмотичного компонента середовища інкубації пермеабілізованих ацинарних панкреатитів спостерігається краща відтворюваність даних та вищий ступінь спряженості між процесами дихання та окисного фосфорилювання, то їх мітохондрії перебувають у кращому фізіологічному стані, ніж при застосуванні KСl. Запропонований спосіб дає змогу розв'язати поставлене технічне завдання, а саме підвищити ступінь наближеності умов дослідження дихання мітохондрій ацинарних клітин підшлункової залози in situ до фізіологічних та досягнути кращого спряження між диханням і окисним фосфорилюванням. 45 ФОРМУЛА КОРИСНОЇ МОДЕЛІ 50 Спосіб дослідження дихання мітохондрій ацинарних клітин підшлункової залози in situ, за яким отримують суспензію ізольованих ацинусів підшлункової залози, замінюють середовище виділення на середовище інкубації, пермеабілізують плазматичну мембрану клітин неіонним детергентом - дигітоніном, контролюють ступінь проникності плазматичної мембрани і полярографічно реєструють швидкість поглинання кисню, який відрізняється тим, що як основний осмотичний компонент середовища інкубації використовують сахарозу. 3 UA71234 U Комп’ютерна верстка М. Ломалова Державна служба інтелектуальної власності України, вул. Урицького, 45, м. Київ, МСП, 03680, Україна ДП “Український інститут промислової власності”, вул. Глазунова, 1, м. Київ – 42, 01601 4
ДивитисяДодаткова інформація
Назва патенту англійськоюMethod for investigation mitochondrial breath of acinar cells of pancreatic gland in situ
Автори англійськоюManko Bohdan Oleksiiovych, Manko Volodymyr Vasyliovych, Klevets Myron Yuriiovych
Назва патенту російськоюСпособ исследования дыхания митохондрий ацинарных клеток поджелудочной железы in situ
Автори російськоюМанько Богдан Алексеевич, Манько Владимир Васильевич, Клевец Мирон Юрьевич
МПК / Мітки
МПК: G01N 33/00, C12N 5/02
Мітки: спосіб, підшлункової, ацинарних, дослідження, дихання, мітохондрій, залози, клітин
Код посилання
<a href="https://ua.patents.su/6-71234-sposib-doslidzhennya-dikhannya-mitokhondrijj-acinarnikh-klitin-pidshlunkovo-zalozi-in-situ.html" target="_blank" rel="follow" title="База патентів України">Спосіб дослідження дихання мітохондрій ацинарних клітин підшлункової залози in situ</a>
Спосіб дослідження дихання мітохондрій in situ

Номер патенту: 60528
Опубліковано: 25.06.2011
Автори: Мерлавський Володимир Михайлович, Великопольська Ольга Юріївна, Манько Володимир Васильович, Манько Богдан Олексійович
МПК: G01N 33/50, A01N 61/00
Мітки: спосіб, мітохондрій, дихання, дослідження
Формула / Реферат:
Спосіб дослідження дихання мітохондрій in situ, за яким пермеабілізують плазматичну мембрану клітин неіонним детергентом-дигітоніном, контролюють ступінь проникності плазматичної мембрани і полярографічно реєструють швидкість поглинання кисню, який відрізняється тим, що пермеабілізацію здійснюють оптимальною кількістю детергенту, яка знаходиться у сталому співвідношенні з кількістю досліджуваних клітин, яке для кожної тканини і для кожного...
Спосіб визначення ранньої стадії порушення функціональних резервів бета-клітин острівців підшлункової залози у людей літнього віку з інсулінорезистентністю

Номер патенту: 60702
Опубліковано: 25.06.2011
Автори: Коркушко Олег Васильович, Шатило Валерій Броніславович, Чижова Валентина Петрівна
МПК: G01N 33/49
Мітки: віку, порушення, ранньої, літнього, функціональних, підшлункової, визначення, спосіб, людей, інсулінорезистентністю, стадії, острівців, залози, бета-клітин, резервів
Формула / Реферат:
Спосіб визначення ранньої стадії порушення функціональних резервів b-клітин острівців підшлункової залози у людей літнього віку з інсулінорезистентністю, що включає стандартний глюкозотолерантний тест, який відрізняється тим, що одночасно з визначенням HOMA-IR визначають концентрацію імунореактивного інсуліну (ІРІ) через 2 години, якщо він перевищує 40 мкОД/мл, вважають функціональні резерви збереженими, а при встановленні менше 40 мкОД/мл -...
Спосіб дослідження зовнішньосекреторної функції підшлункової залози

Номер патенту: 21516
Опубліковано: 28.02.2000
Автори: Ковалів Юрій Михайлович, Темник Ірина Василівна, Дубовий Борис Васильович
МПК: A61B 10/00, G01N 33/50
Мітки: спосіб, функції, дослідження, підшлункової, залози, зовнішньосекреторної
Формула / Реферат:
Спосіб дослідження зовнішньосекреторної функції підшлункової залози, що включає збирання натще порції сечі, приймання ентерального подразника та збирання через 30 і 60хв. після його приймання другої і третьої порції сечі, визначення в усіх порціях активності та дебіту альфа-амілази, який відрізняється тим, що як ентеральний подразник використовують 1,5% розчин цикорію і додатково досліджують четверту порцію сечі, яку збирають через 90хв....
Спосіб встановлення ступеня токсичності комбінованих пестицидів за допомогою визначення швидкості і ефективності дихання мітохондрій печінки

Номер патенту: 41847
Опубліковано: 10.06.2009
Автори: Стегній Борис Тимофійович, Жукова Ірина Олексійовна, Малінін Олег Олексійович
МПК: G01N 27/48
Мітки: токсичності, спосіб, ефективності, пестицидів, швидкості, комбінованих, печінки, встановлення, допомогою, дихання, визначення, ступеня, мітохондрій
Формула / Реферат:
Спосіб встановлення ступеня токсичності комбінованих пестицидів за допомогою визначення швидкості і ефективності дихання мітохондрій печінки, що включає гомогенізацію, інкубацію, вимірювання до додавання АДФ (адинозиндифосфат), після додавання АДФ, перетворення його в АТФ та визначення коефіцієнтів Ларді-Вельмана, Чанса-Вільямса і фосфорилювання, який відрізняється тим, що використовують як окисник буфер-трис-HCL, використовують стандартні...
Спосіб визначення резистентності бета-клітин панкреатичних острівців підшлункової залози в експерименті

Номер патенту: 53327
Опубліковано: 11.10.2010
Автори: Іваненко Тарас Васильович, Ганчева Ольга Вікторівна, Абрамов Андрій Володимирович, Колеснік Юрій Михайлович
МПК: G09B 23/00
Мітки: острівців, підшлункової, спосіб, визначення, залози, експерименті, бета-клітин, резистентності, панкреатичних
Формула / Реферат:
Спосіб визначення резистентності бета-клітин панкреатичних острівців підшлункової залози в експерименті, який відрізняється тим, що він включає одноразове внутрішньочеревинне введення статевозрілим щурам субдіабетогенної дози стрептозотоцину 45 мг/кг ваги тварини, розведеного ех tempore в 1 мл 0,1 М цитратного буфера рН 4,5, випоювання в перший день після введення стрептозотоцину 20 % розчину глюкози, на другий день - 10 %, і визначення...
Попередній патент: Газорідинний сепаратор
Наступний патент: Спосіб отримання наноструктур zno
Випадковий патент: Метод стимуляції системи саморегуляції організму