Спосіб отримання клітини трансгенної рослини для спрямованого впливу на ген в рослинах, трансгенна рослина або рослинна тканина, насіння, отримане з трансгенної рослини
Номер патенту: 110472
Опубліковано: 12.01.2016
Автори: Майлз Ребекка Рут, Ейнлі Уілльям Майкл, Мюррей Майкл Г., Уебб Стівен Р., Корбін Девід Річард, Блю Райан С.

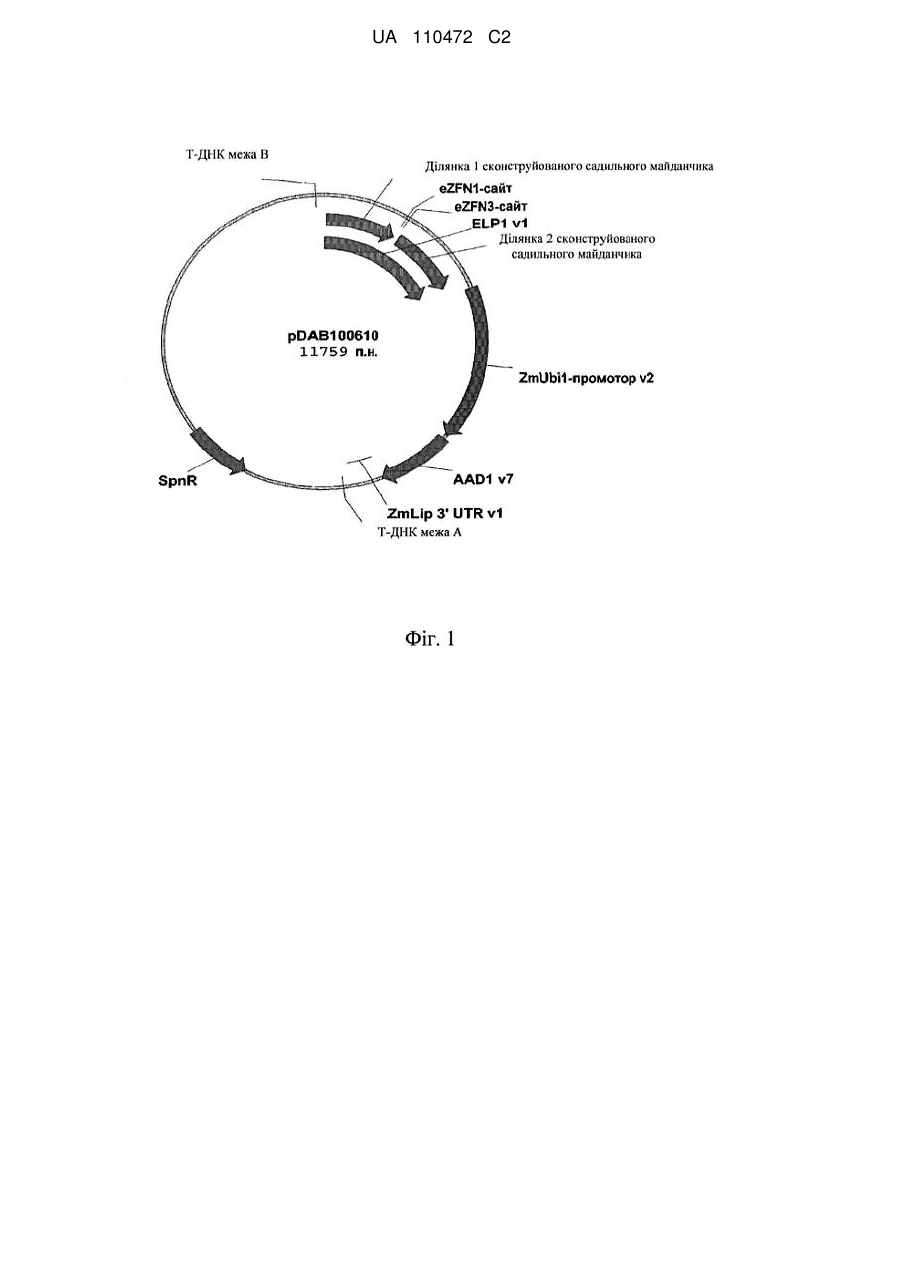






Формула / Реферат
1. Спосіб отримання клітини трансгенної рослини, причому спосіб включає:
надання молекули нуклеїнової кислоти, що включає щонайменше два полінуклеотиди, вибраних з групи, що складається з SEQ ID NO:1-10, і щонайменше один сайт розпізнавання ендонуклеази спрямованої дії, де щонайменше два полінуклотиди, вибраних з групи, що складається з SEQ ID NO:1-10, фланкують щонайменше один сайт розпізнавання ендонуклеази спрямованої дії; і
трансформування рослинної клітини молекулою нуклеїнової кислоти, де молекула нуклеїнової кислоти стабільно інтегрована в геном рослинної клітини.
2. Спосіб за п. 1, де молекула нуклеїнової кислоти додатково включає неідентичні сайти рестрикції.
3. Спосіб за п. 2, де неідентичні сайти рестрикції включають сумісні одноланцюжкові кінці, які дають можливість конкатамеризації множини нуклеїнових кислот, кожна з яких містить:
щонайменше дві ділянки послідовності нуклеїнової кислоти, які не ідентичні послідовності ДНК геному рослинної клітини, і щонайменше один сайт розпізнавання ендонуклеази спрямованої дії, де щонайменше дві ділянки послідовності нуклеїнової кислоти, які не ідентичні послідовності ДНК геному рослинної клітини, фланкують щонайменше один сайт розпізнавання ендонуклеази спрямованої дії.
4. Спосіб за п. 1, де ендонуклеаза спрямованої дії являє собою нуклеазу домену "цинкові пальці".
5. Спосіб за п. 1, де молекула нуклеїнової кислоти стабільно інтегрована в геном рослинної клітини випадковим чином.
6. Спосіб за п. 1, де молекула нуклеїнової кислоти стабільно інтегрована в один або декілька відомих сайтів-мішеней в геномі рослинної клітини.
7. Клітина трансгенної рослини, отримана способом за п. 1.
8. Спосіб отримання клітини трансгенної рослини, причому спосіб включає: надання клітини трансгенної рослини, що має трансгенну молекулу нуклеїнової кислоти, стабільно інтегровану в геном рослинної клітини, де трансгенна молекула нуклеїнової кислоти містить:
щонайменше два полінуклеотиди, вибраних з групи, що складається з SEQ ID NO:1-10, які не гібридизуються в жорстких умовах з геномною ДНК рослинної клітини, і щонайменше один сайт розпізнавання ендонуклеази спрямованої дії, де щонайменше два полінуклеотиди, вибраних з групи, що складається з SEQ ID NO:1-10, які не гібридизуються в жорстких умовах з геномною ДНК рослинної клітини, фланкують щонайменше один сайт розпізнавання ендонуклеази спрямованої дії;
надання щонайменше однієї ендонуклеази спрямованої дії або першої молекули нуклеїнової кислоти, що включає полінуклеотид, який кодує щонайменше одну ендонуклеазу спрямованої дії, де щонайменше одна ендонуклеаза спрямованої дії розпізнає щонайменше один сайт розпізнавання ендонуклеази спрямованої дії;
надання другої молекули нуклеїнової кислоти, що включає цікавлячий полінуклеотид і два додаткових полінуклеотиди, вибраних з групи, що складається з SEQ ID NO:1-10, що фланкують цікавлячий полінуклеотид, де кожний з двох додаткових полінуклеотидів гомологічний одному з двох полінуклеотидів, які не гібридизуються в жорстких умовах з геномною ДНК;
введення в рослинну клітину щонайменше однієї ендонуклеази спрямованої дії або першої молекули нуклеїнової кислоти і другої молекули нуклеїнової кислоти, де цікавлячий полінуклеоид стабільно інтегрований в геном рослинної клітини.
9. Клітина трансгенної рослини, отримана способом за п. 8.
10. Спосіб за п. 1, де молекула нуклеїнової кислоти фланкована на кожному кінці одним або декількома додатковими сайтами розпізнавання ендонуклеази спрямованої дії.
11. Спосіб за п. 10, що додатково включає введення в рослинну тканину або в рослину однієї або декількох ендонуклеаз спрямованої дії, які розпізнають один або декілька додаткових сайтів розпізнавання ендонуклеаз спрямованої дії, де молекула нуклеїнової кислоти вирізається з генома рослинної тканини або рослини.
12. Спосіб за п. 8, де ендонуклеаза спрямованої дії являє собою нуклеазу домену "цинкових пальців".
13. Спосіб отримання трансгенної рослини або рослинної тканини, що включає:
надання клітини трансгенної рослини за п. 7; і
регенерацію трансгенної рослини або рослинної тканини з клітини трансгенної рослини.
14. Тканина трансгенної рослини, отримана способом за п. 13.
15. Трансгенна рослина, отримана способом за п. 13.
16. Насіння, отримане трансгенною рослиною за п. 15, де насіння містить трансгенну молекулу нуклеїнової кислоти.
17. Спосіб отримання трансгенної рослини або рослинної тканини, що включає:
надання клітини трансгенної рослини за п. 9; і
регенерацію трансгенної рослини або рослинної тканини з клітини трансгенної рослини.
18. Тканина трансгенної рослини, отримана способом за п. 17.
19. Трансгенна рослина, отримана способом за п. 17.
20. Насіння, отримане трансгенною рослиною за п. 19, де насіння містить трансгенну молекулу нуклеїнової кислоти.
Текст
Реферат: Заявлений винахід стосується способів генерування трансгенних рослин, а саме способу введення молекули нуклеїнової кислоти в геном організму-хазяїна, що виявляє низький ступінь гомологічної рекомбінації. Розкрито способи створення і генерування "сконструйованих садильних майданчиків" (ELP) для трансформації рослини молекулою нуклеїнової кислоти, що представляє інтерес. Згідно з винаходом заявлено спосіб отримання клітини трансгенної рослини, який включає надання молекули нуклеїнової кислоти, що включає щонайменше два полінуклеотиди, які не гібридизуються в жорстких умовах з геномною ДНК рослинної клітини, і щонайменше один сайт розпізнавання ендонуклеази спрямованої дії, де щонайменше два полінуклотиди, які не гібридизуються в жорстких умовах з геномною ДНК рослинної клітини, фланкують щонайменше один сайт розпізнавання ендонуклеази спрямованої дії; і трансформування рослинної клітини молекулою нуклеїнової кислоти, де молекула нуклеїнової кислоти стабільно інтегрована в геном рослинної клітини. Також розкриті молекули нуклеїнових кислот, що використовуються у способі та генетично модифіковані рослини, що отримуються згідно зі способом, та їх насіння. UA 110472 C2 (12) UA 110472 C2 UA 110472 C2 5 10 15 20 25 30 35 40 45 50 55 60 ОПИС ПРІОРИТЕТ Дана заявка вимагає пріоритет на основі дати подачі попередньої заявки США з порядковим номером 61/297641 від 22.01.2010, "СКОНСТРУЙОВАНІ САДИЛЬНІ МАЙДАНЧИКИ ДЛЯ СПРЯМОВАНОГО ВПЛИВУ НА ГЕН В РОСЛИНАХ". ГАЛУЗЬ ТЕХНІКИ Винахід в основному стосується композицій і способів генерування трансгенних організмів, наприклад, рослин. У деяких варіантах здійснення, спосіб за винаходом дає можливість інкорпорування сконструйованих САДИЛЬНИХ МАЙДАНЧИКІВ (engineered landing pads, ELP) в геномний сайт, полегшуючи, таким чином, введення наступних цікавлячих молекул нуклеїнових кислот в певний геномний сайт, що містить один або декілька ELP. У деяких варіантах здійснення, ELP може включати ділянки нуклеотидної послідовності, що включають по суті випадкову послідовність, фланкуючу сайти зв'язування нуклеази доменів "цинкові пальці" (ZFN). РІВЕНЬ ТЕХНІКИ Багато рослин генетично трансформуються генами з інших видів для введення цільових властивостей, як наприклад, для поліпшення сільськогосподарської цінності за допомогою, наприклад, поліпшення якості поживної цінності, збільшення врожайності, надання стійкості до шкідників або захворювань, за допомогою збільшення переносимості посушливого періоду і зовнішніх впливів, за допомогою поліпшення плодівницьких якостей, таких як пігментація і ріст, і/або за допомогою надання стійкості до гербіцидів, тобто це властивості, що дають можливість виробництва промислово застосовних сполук і/або матеріалів з рослини і/або даючи можливість виробництва лікарських засобів. Введення клонованих генів в рослинні клітини і отримання стабільних плодоносних трансгенних рослин може використовуватися для створення таких модифікацій рослини і дозволяє створення генетично сконструйованих рослин (наприклад, для поліпшення культури). У цих способах чужорідну ДНК випадковим чином вводять в ядерну або в пластидну ДНК еукаріотичної рослинної клітини з подальшим виділенням клітин, що містять чужорідну ДНК, інтегровану в ДНК клітини з отриманням стабільно трансформованих рослинних клітин. Перші покоління трансгенних рослин, як правило, були генеровані за допомогою технології трансформації, опосередкованої Agrobacterium. Успіх цих методів активізував розробку інших способів введення молекули нуклеїнової кислоти, яка цікавить, в геном рослини, таких як опосередковане ПЕГ поглинання ДНК в протопластах, бомбардування мікрочастинками і трансформація, опосередкована волокнами кремнію. Однак у всіх цих методах трансформації трансгени, інкорпоровані в геном рослини, інтегрувалися випадковим чином і в непередбачуваній кількості піків. Часто, трансгени можуть бути інтегровані в формі повторів або повнорозмірного трансгена або його частин. Такий комплексний профіль інтеграції може впливати на рівень експресії трансгенів (наприклад, за допомогою порушення транскрибованою РНК за допомогою пост-транскрипційного механізму сайленсингу генів або шляхом індукування метилування введеної ДНК), здійснюючи, таким чином, негативну регуляцію транскрипції трансгена. Крім того, сайт інтегрування сам по собі може впливати на рівень експресії трансгена. Комбінація цих факторів приводить в результаті до отримання широкого спектра рівня експресії трансгенів або чужорідної ДНК, яка цікавить, серед різних трансгенних рослинних клітинних ліній і ліній рослин. Крім того, інтеграція чужорідної ДНК, яка цікавить, може мати руйнівну дію на ділянку генома, де відбувається інтеграція і може впливати на нормальне функціонування цієї ділянки, на яку спрямований вплив, або порушувати його, приводячи, таким чином, до частих небажаних побічних ефектів. Вищевикладене неминуче спричиняє за собою те, що у всіх випадках, коли досліджують ефект введення конкретної чужорідної ДНК в рослину, то з метою отримання значущих результатів генерують і аналізують велику кількість трансгенних ліній рослин. Аналогічно, при генеруванні трансгенних культур рослин, де конкретну ДНК, яка цікавить, вводять в рослини для отримання трансгенної рослини з цільовим фенотипом, то створюють велику популяцію незалежно створених трансгенних ліній рослин, щоб була можливість селекції ліній рослин з оптимальною експресією трансгенів і з мінімальними побічними ефектами, що надаються на загальний фенотип трансгенної рослини, або з їх відсутністю. Конкретно в цій галузі, доцільні більш керовані способи трансгенозу, наприклад, з точки зору обтяжливих регуляторних вимог і високих витрат, пов'язаних з повтореннями польових досліджень, що вимагаються для виключення небажаних подій трансгенозу. Крім того, очевидно, що було б корисно мати можливість вставки цільової ДНК в процесі укладання трансгена. Було розроблено декілька способів з метою контролю трансгенної вставки в рослинах. Див., наприклад, Kumar and Fladung (2001) Trends Plant Sci. 6: 155-9. Ці способи базуються на 1 UA 110472 C2 5 10 15 20 25 30 35 40 45 50 55 інтеграції трансгена на основі гомологічної рекомбінації. Дана стратегія успішно застосовувалася у прокаріотів і нижчих еукаріотів. Paszkowski et al. (1988) EMBO J. 7: 4021-6. Однак для рослин донедавна головний механізм інтеграції трансгена основувався на незаконній рекомбінації між рекомбінованими ланцюгами ДНК. Таким чином, велика проблема в цій галузі пов'язана з детектування рідких подій гомологічної рекомбінації, які маскуються набагато ефективнішою інтеграцією чужорідної ДНК за допомогою незаконної рекомбінації. Спеціально створені нуклеази домену "цинкові пальці" (ZFN) являють собою білки, створені для доставки цільових сайт-специфічних дволанцюжкових розривів в ДНК з подальшою рекомбінацією відщеплених кінців. ZFN об'єднують в собі домен неспецифічного відщеплення ендонуклеази рестрикції, таку як наприклад, FokI, разом з ДНК-зв'язувальними білковими доменами "цинкові пальці". Див., наприклад, Huang et al. (1996) J. Protein Chem. 15: 481-9; Kim et al. (1997) Proc. Natl. Acad. Sci. USA 94: 3616-20; Kim et al. (1996) Proc. Natl. Acad. Sci. USA 93: 1156-60; Kim et al. (1994) Proc Natl. Acad. Sci. USA 91: 883-7; Kim et al. (1997b) Proc. Natl. Acad. Sci. USA 94: 12875-9; Kim et al. (1997c) Gene 203: 43-9; Kim et al. (1998) Biol. Chem. 379: 489-95; Nahon and aveh (1998) Nucleic Acids Res. 26: 1233-9; Smith et al. (1999) Nucleic Acids Res. 27: 674-81. Індивідуальні мотиви "цинкові пальці" можуть бути створені для спрямованого впливу і зв'язування з широким спектром ДНК-сайтів. Канонічні мотиви "цинкових пальців" CyS 2His2, а також не канонічні CyS3His зв'язуються з ДНК шляхом вставки α-спіралі у велику борозенку подвійної спіралі. Розпізнавання ДНК за допомогою "цинкових пальців" є модульним процесом: кожний палець контактує спочатку з трьома послідовними парами основ в мішені, і декілька ключових залишків в білку опосередковують розпізнавання. Було продемонстровано, що ендонуклеаза рестрикції FokI повинна димеризуватися за допомогою нуклеазного домену з метою відщеплення ДНК, що індукує дволанцюжковий розрив. Аналогічно, ZFN також потрібна димеризація нуклеазного домену з метою розрізання ДНК. Mani et al. (2005) Biochem. Biophys. Res. Commun. 334: 1191-7; Smith et al. (2000) Nucleic Acids Res. 28: 3361-9. Димеризація ZFN полегшується двома сусідніми, протилежно орієнтованими сайтами зв'язування. СУТЬ ВИНАХОДУ У даному документі розкриті способи введення молекули нуклеїнової кислоти в геном організму-хазяїна, що виявляє низьку міру гомологічної рекомбінації, наприклад, що являє собою види рослин або тварин. У конкретних прикладах, використовуються "сконструйовані садильні майданчики" (ELP), де ELP можуть включати ділянки нуклеотидної послідовності, що включає нуклеотидні послідовності, по суті позбавлені гомології з геномом організму-хазяїна (наприклад, випадково генеровані послідовності), фланкуючі сайти зв'язування для ДНКзв'язувальних доменів (наприклад, білки з доменами "цинкові пальці" (ZFP), мегануклеази або лейцинові застібки). ДНК-зв'язувальні домени, які спрямовано впливають на сайти зв'язування ELP, можуть в природному стані включати функціональні домени ДНК-розщеплення або можуть бути частиною химерних білків, які додатково включають функціональний домен, наприклад, домен ендонуклеазного розщеплення або половину домену розщеплення (наприклад, ZFN, рекомбіназа, транспозаза або ендонуклеаза спрямованого впливу, що містять модифікований ДНК-зв'язувальний домен). Цей клас ДНК-зв'язувальних і розщеплювальних білків колективно позначається в даному документі як "ендонуклеази спрямованої дії". У конкретних прикладах, також може використовуватися не рандомізована нуклеотидна послідовність з біологічних джерел, відмінних від організму-хазяїна; при цьому відсутність гомології з геномом організмумішені є прийнятною. Таким чином, в деяких прикладах, нуклеотидні послідовності можуть бути сконструйовані для гарантії по суті відсутності гомології з ділянками будь-яких послідовностей генома рослини-мішені. У деяких прикладах, ELP можуть забезпечувати ділянки гомології і сайти зв'язування для високоактивних ендонуклеаз спрямованої дії (наприклад, ZFN) для спрямованого впливу на гени, пов'язаного з гомологією. Отже, ELP можуть зменшувати або виключати необхідність використання інших нуклеотидних послідовностей, зовнішніх або внутрішніх, у вставці нуклеїнової кислоти. ELP можуть бути сконструйовані так, щоб вони були позбавлені помилкових відкритих рамок зчитування і містили або не містили сайти ферментів рестрикції, переважні для конструкції векторів або аналізу рослин, трансформованою молекулою нуклеїнової кислоти, яка цікавить. У деяких прикладах, ELP можуть бути фланковані неідентичними сайтами рестрикції, які генерують сумісні кінці при гідролізі ферментом рестрикції, але гібридні ліговані сайти не розщеплюються ніяким з ферментів рестрикції. У цих і інших прикладах, це дає можливість конкатенації ELP у великі конгломерати, кожний з унікальними ділянками гомології і з сайтами зв'язування ендонуклеаз спрямованого впливу (наприклад, сайти зв'язування ZFN). 2 UA 110472 C2 5 10 15 20 25 30 35 40 45 50 55 60 У деяких прикладах, ELP можуть бути інкорпоровані випадковим чином в ген-мішень або в геномні сайти-мішені, які, як було продемонстровано, розміщують трансгенні вставки без якогонебудь шкідливого впливу або з прийнятним шкідливим впливом на отримані в результаті трансгенні рослини. Ділянки гомології в геномному сайт-мішені можуть бути ідентичні ділянкам гомології в цільовій донорній молекулі нуклеїновій кислоті, полегшуючи, таким чином, спрямовану гомологічну рекомбінацію після дволанцюжкового розщеплення ендонуклеазами спрямованого впливу (наприклад, ZFN). Відповідно, розкриті способи створення і генерування ELP для трансформації рослини молекулою нуклеїнової кислоти, яка цікавить. Також розкриті молекули нуклеїнових кислот, що застосовуються за винаходом, і генетично модифіковані рослини, що отримуються згідно з винаходом. Все вищеописане і інші ознаки винаходу стануть очевидні на основі наступного докладного опису деяких варіантів здійснення, які починаються з посилань на супроводжуючі їх креслення. КОРОТКИЙ ОПИС КРЕСЛЕНЬ Фіг. 1 включає зображення типового вектора, що включає ELP, pDAB100610. Фіг. 2 включає зображення типового вектора, що включає ELP, pDAB100611. Фіг. 3 включає зображення типового вектора, що включає ELP, pDAB100640. Фіг. 4 включає зображення типового вектора, що включає ELP, pDAB100641. Фіг. 5 включає зображення типового вектора, що включає донорний фрагмент, інтегрований в ELP, pDAB105955. Фіг. 6 включає зображення типового вектора, що включає донорний фрагмент, інтегрований в ELP, pDAB105956. Фіг. 7 включає зображення типового вектора, що включає донорний фрагмент, інтегрований в ELP, pDAB105979. Фіг. 8 включає зображення типового вектора, що включає донорний фрагмент, інтегрований в ELP, pDAB105980. Фіг. 9 являє собою схему: a) ELP 1 в маїсі; b) донорну конструкцію для переорієнтації ELP1; c) Переорієнтований локус ELP1. Фіг. 10 включає зображення типового вектора pDAB104132. Фіг. 11 включає зображення типового ДНК-фрагмента Сайту Множинного Клонування, який фланкований сконструйованими сайтами зв'язування з доменом "цинкові пальці" і буферними послідовностями. Фіг. 12 включає зображення типового вектора pDAB104126. Фіг. 13 включає зображення типового вектора pDAB104136. Фіг. 14 включає зображення типового вектора pDAB104138. Фіг. 15 включає зображення типового вектора pDAB104140. Фіг. 16 включає зображення типового вектора pDAB104142. Фіг. 17 включає зображення типового вектора pDAB104133. Фіг. 18 включає зображення типового вектора pDAB104134. Фіг. 19 включає зображення типового вектора pDAB104135. Фіг. 20 включає зображення типового вектора pDAB104137. Фіг. 21 включає зображення типового вектора pDAB104139. Фіг. 22 включає зображення типового вектора pDAB104141. Фіг. 23 включає зображення типового вектора pDAB104143. ДОКЛАДНИЙ ОПИС I. Огляд деяких варіантів здійснення У деяких варіантах здійснення, пропонуються способи для введення молекул нуклеїнових кислот, що включають "сконструйовані садильні майданчики" (ELP), в організм-хазяїн, так що полегшується інтеграція подальших молекул нуклеїнових кислот, які цікавлять, в організмхазяїн. У варіантах здійснення, ELP можуть включати ділянки послідовності нуклеїнових кислот, які не гомологічні нативним послідовностям організму-хазяїна (наприклад, по суті, випадково генеровані послідовності нуклеїнових кислот, які можуть бути потім відібрані для вставки на основі певних цільових критеріїв, описаних в даному документі), і фланкуючі сайти розпізнавання ендонуклеаз спрямованої дії (наприклад, сайти розпізнавання ZFN). Сайти інтеграції ELP можуть бути випадковими, а можуть бути визначатися шляхом ідентифікації, наприклад, структурного гена всередині генома-хазяїна, який має цільовий рівень експресії молекули нуклеїнової кислоти, яка цікавить, введеної в сайт ELP, або можуть визначатися за допомогою ідентифікації локалізації всередині генома-хазяїна, яка не чинить метаболічного, функціонального, агрономічного (у випадку рослини) або іншого небажаного ефекту трансформованому організму-хазяїну при введенні ELP в сайт. ELP можуть бути введені в 3 UA 110472 C2 5 10 15 20 25 30 35 40 45 50 55 60 геном в тандемі, так що, наприклад, ELP присутні як конкатамери в організмі, що отримується згідно зі способами за винаходом. У деяких варіантах здійснення, інтеграція молекули нуклеїнової кислоти, яка цікавить, здійснюється в сайті ELP. У варіантах здійснення, молекула нуклеїнової кислоти, яка цікавить, що вводиться в сайт ELP, асоційована з однією або декількома ендонуклеазами спрямованої дії, представленими у вигляді поліпептидів або експресованими з РНК або ДНК, що вводиться. Крім того, молекула нуклеїнової кислоти, що вводиться, може включати ELP, відмінну від вже інкорпорованої в ген-хазяїн. Фахівцям в даній галузі зрозуміло, що послідовність нуклеїнової кислоти, що вводиться в ген-хазяїн в сайт, де експресується нативна послідовність нуклеїнової кислоти в організмі дикого типу, буде, як очікується, експресуватися аналогічно нативній послідовності нуклеїнової кислоти. Наприклад, якщо нативна послідовність експресується в організмі дикого типу під контролем регуляторних елементів (наприклад, таких що послідовність нуклеїнової кислоти експресується тканиноспецифічним чином або залежно від стадії розвитку), то очікується, що послідовність нуклеїнової кислоти, що вводиться, буде піддаватися впливу такого ж або подібного регуляторного контролю. Таким чином, ELP можуть полегшувати цільову часову і/або місцеву експресію молекули нуклеїнової кислоти, яка цікавить, введеної по сайту ELP. II. Скорочення тІФА твердофазний імуноферментний аналіз ELP сконструйований садильний майданчик ZFN нуклеаза домену "цинкові пальці" III. Терміни Експресія гена: процес, за допомогою якого кодована інформація траскрипційної одиниці нуклеїнової кислоти (що включає, наприклад, геномну ДНК) перетворюється в діючу, не діючу або в структурну частину клітини, часто включаючи в себе синтез білка. На експресію гена можуть впливати зовнішні сигнали; наприклад, експонування клітини, тканини або організму з агентом, який збільшує або зменшує експресію гена. Експресія гена також може регулюватися на будь-якому рівні ланцюжка ДНК-РНК-білків. Регуляція експресії гена відбувається, наприклад, за допомогою контролів, що впливають на транскрипцію, трансляцію, РНКтранспорт і процесинг, деградацію проміжних молекул, таких як мРНК, або за допомогою активації, інактивації, компартменталізації або деградації специфічних білкових молекул після того, як вони були синтезовані, або шляхом комбінації перерахованого. Експресія гена може бути виміряна на рівні РНК або на рівні білка за допомогою будь-якого методів, відомих в даній галузі, що включають, зокрема, Нозерн-блот, ОТ-ПЛР, Вестерн-блот або аналізи активності білка in vitro, in situ, або in vivo. Гібридизація: Олігонуклеотиди і їх аналоги гібридизуються за допомогою водневого зв'язку, що включає взаємодію Уотсона-Кріка, зв'язування по Хугстену або зворотне Хугстенівське утворення водневого зв'язку між комплементарними основами. Як правило, молекули нуклеїнових кислот складаються з азотистих основ, які являють собою або піримідинові основи (цитозин (С), урацил (U), і тимідин (Т)), або пуринові (аденін (А) і гуанін (G)). Ці азотисті основи утворюють водневі зв'язки між піримідином і пурином, і зв'язування піримідину з пурином позначається як "спаровування основ". Більш конкретно, А буде утворювати водневий зв'язок з Т або U, і G буде зв'язуватися з С. "Комплементарність" означає спаровування основ, що відбувається між двома окремими послідовностями нуклеїнових кислот або між двома окремими ділянками однієї послідовності нуклеїнової кислоти. "Здатні специфічно гібридизуватися" і "специфічно комплементарні" це терміни, які вказують на достатню міру комплементарності, так що відбувається стабільне і специфічне зв'язування між олігонуклеотидом і ДНК- або РНК-мішенню. Для специфічної гібридизації олігонуклеотид необов'язково повинен бути на 100 % комплементарний його послідовності-мішені. Олігонуклеотид здатний специфічно гібридизуватися, коли зв'язування олігонуклеотиду з молекулами його ДНК- або РНК-мішеней перешкоджає нормальному функціонуванню ДНК- або РНК-мішені і є достатня міра комплементарності, щоб уникнути неспецифічного зв'язування олігонуклеотиду з послідовностями, що не є мішенями, при умовах, де доцільне специфічне зв'язування, наприклад, при фізіологічних умовах у випадку in vivo-аналізів або систем. Таке зв'язування позначається як специфічна гібридизація. Умови гібридизація, що приводять до конкретного ступеня жорсткість, будуть варіюватися залежно від характеру вибраного способу гібридизації і від складу і довжини молекул нуклеїнових кислот, що гібридизуються. Як правило, температура гібридизація і іонна сила + 2+ (особливо концентрація Na і/або Mg ) гібридизаційного буфера будуть сприяти жорсткості гібридизації, хоча кількість промивань також впливає на жорсткість. Розрахунки, що стосуються 4 UA 110472 C2 5 10 15 20 25 30 35 40 45 50 55 60 умов гібридизація, необхідні для досягнення конкретного ступеня жорсткості, розкриті в nd Sambrook et al. (ed.), Molecular Cloning: А Laboratory Manual, 2 ed., vol. 1-3, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, 1989, chs. 9 and 11. Для цілей даного опису, термін "жорсткі умови" охоплює умови, при яких гібридизація буде відбуватися, якщо має місце менш ніж 25 % незбігів між молекулою, що гібридизується, і послідовністю-мішенню. "Жорсткі умови" можуть додатково розподілятися по конкретних рівнях жорсткості. Таким чином, при використанні в даному документі, умови "помірної жорсткості" це умови, при яких молекули, що мають більше ніж 25 % незбігів, не будуть гібридизуватися. Умови "середньої жорсткості" це ті, при яких молекули, що мають більше ніж 15 % незбігів, не будуть гібридизуватися, і умови "високої жорсткості" це ті, при яких послідовність, що має більш ніж 10 % незбігів, не буде гібридизуватися. Умови "дуже високої жорсткості" це ті, при яких послідовності, що мають більше ніж 6 % незбігів, не будуть гібридизуватися. У конкретних варіантах здійснення, жорсткі умови можуть включати гібридизацію при 65 °C з подальшим послідовним відмиванням при 65 °C за допомогою 0,1 х SSC/0,1 % SDS протягом 40 хвилин. Виділений: "Виділений" біологічний компонент (такий як нуклеїнова кислота або білок) по суті відділений від, отриманий незалежно від або очищений від інших біологічних компонентів в клітині організму, в якій компонент існує в природному вигляді, тобто, від інших хромосомних або позахромосомних ДНК, РНК і білків. Молекули нуклеїнових кислот і білки, які "виділені", включають молекули нуклеїнових кислот і білки, очищені стандартними методами очищення. Термін також охоплює нуклеїнові кислоти і білки, отримані за допомогою рекомбінантної експресії в клітині-хазяїні, а також хімічно синтезовані молекули нуклеїнових кислот, білки і пептиди. Молекула нуклеїнової кислоти: полімерна форма нуклеотидів, яка може включати як смисловий, так і антисмисловий ланцюги РНК, кДНК, геномної ДНК і їх синтетичні форми і змішані полімери. Нуклеотид являє собою рибонуклеотид, дезоксинуклеотид або модифіковану форму будь-якого типу нуклеотиду. При використанні в даному документі термін "молекула нуклеїнової кислоти" є синонімом "нуклеїнової кислоти" і "полінуклеотиду". Термін включає одноланцюжкову і дволанцюжкову форми ДНК. Молекула нуклеїнової кислоти може включати кожний або обидва типи нуклеотидів, а саме природні і модифіковані нуклеотиди, зв'язані разом за допомогою природних або синтетичних нуклеотидних зв'язків. Молекули нуклеїнової кислоти можуть бути модифіковані хімічно або біохімічно або можуть містити синтетичні або модифіковані нуклеотидні основи, які відомі фахівцям в даній галузі. Такі модифікації включають, наприклад, введення мітки, метилування і заміну одного або декількох природних нуклеотидів на їх аналоги. Інші модифікації включають міжнуклеотидні модифікації, такі як незаряджені зв'язки (наприклад, метилфосфонати, фосфотриефіри, фосфорамідати, карбамати і т. д.), заряджені зв'язки (наприклад, фосфоротіоати, фосфородитіоати і т. д.), приєднані компоненти (наприклад, пептиди), інтеркалятори (наприклад, акридин, псорален і т. д.), хелатори, алкілатори і модифіковані зв'язки (наприклад, альфа-аномери нуклеїнових кислот і т. д.). Термін "молекула нуклеїнової кислоти" також включає будь-яку топологічну конформацію, що включає одноланцюжкову, дволанцюжкову, частково подвійну, потрійну, у вигляді шпильки, циклічну і блоковану конформації. Функціонально пов'язаний: Перша послідовність нуклеїнової кислоти функціонально пов'язана з другою послідовністю нуклеїнової кислоти, коли перша послідовність нуклеїнової кислоти знаходиться в функціональному зв'язку з другою послідовністю нуклеїнової кислоти. Наприклад, промотор функціонально пов'язаний з кодуючою послідовністю, коли промотор впливає на транскрипцію або експресію кодуючої послідовності. При рекомбінантному отриманні, функціонально пов'язані послідовності нуклеїнових кислот, як правило, є зв'язаними, а також там, де необхідно з'єднувати дві ділянки, кодуючі білки, в одній рамці зчитування. Однак для того, щоб бути функціонально пов'язаними, елементи необов'язково повинні бути зв'язаними. Промотор: Ділянка ДНК, яка, як правило, локалізована вище (відносно 5'-області гена), і необхідна для транскрипції. Промотори дають можливість коректної активації або репресії гена, який вони контролюють. Промотор містить специфічні послідовності, які розпізнаються транскрипційними факторами. Ці фактори зв'язуються з промоторними послідовностями ДНК і приводять в результаті до залучення РНК-полімерази, ферменту, який синтезує РНК з кодуючої ділянки гена. У деяких варіантах здійснення, використовуються тканиноспецифічні промотори. Тканиноспецифічний промотор це послідовність ДНК, яка керує вищим рівнем транскрипції асоційованого гена в тканині, для якої промотор є специфічним, відносно іншої тканини організму. Приклади тканиноспецифічних промоторів включають промотори, специфічні для 5 UA 110472 C2 5 10 15 20 25 30 35 40 45 50 55 60 тапетуму; промотори, специфічні для пильовика; промотори, специфічні для пилку (див., наприклад, патент США No. 7141424, і міжнародну публікацію PCT No. WO 99/042587); промотори, специфічні для насінного зачатку; (див., наприклад, патентну заявку США No. 2001/047525 Al); промотори, специфічні для плоду (див., наприклад, патент США No. 4943674, і 5753475); і промотори, специфічні для насіння (див., наприклад, патент США No. 5420034 і 5608152). У деяких варіантах здійснення також використовуються промотори, специфічні для стадій розвитку, наприклад, промотор, активний в пізній стадії розвитку. Трансформований: Вірус або вектор "трансформує" або "трансдукує" клітину, коли він переносить молекули нуклеїнової кислоти в клітину. Клітина є "трансформованою" молекулою нуклеїнової кислоти, трансдукованою в клітину, коли молекула нуклеїнової кислоти починає стабільно реплікуватися клітиною, або шляхом інкорпорування молекули нуклеїнової кислоти в клітинний геном, або за допомогою епісомної реплікації. При використанні в даному документі, термін "трансформація" охоплює всі методи, за допомогою яких молекула нуклеїнової кислоти може бути введена в клітину. Приклади включають, зокрема, трансфекцію вірусним векторами, трансформацію плазмідними векторами, електропорацію (Fromm et al. (1986) Nature 319: 791-3), ліпофекцію (Feigner et al. (1987) Proc. Natl. Acad. Sci. USA 84: 7413-7), мікроін'єкцію (Mueller et al. (1978) Cell 15: 579-85), перенесення, опосередковане Agrobacterium (Fraley et al. (1983) Proc. Natl. Acad. Sci. USA 80: 4803-7), пряме поглинання ДНК і бомбардування мікрочастинками (Klein et al. (1987) Nature 327: 70). Трансген: Екзогенна послідовність нуклеїнової кислоти. У одному варіанті здійснення, трансген являє собою послідовність гена (наприклад, гена стійкості до гербіциду), що кодує промислово або фармацевтично використовувану сполуку або гена, що кодує цільову сільськогосподарську властивість. Ще в одному варіанті здійснення, трансген являє собою антисмислову послідовність нуклеїнової кислоти, де експресія антисмислової послідовності нуклеїнової кислоти інгібує експресію послідовності нуклеїнової кислоти-мішені. Трансген може містити регуляторні послідовності, функціонально пов'язані з трансгеном (наприклад, промотор). У деяких варіантах здійснення, молекула нуклеїнової кислоти, яка цікавить, що вводиться за допомогою ELP-спрямованої рекомбінації, є трансгеном. Однак в інших варіантах здійснення, молекула нуклеїнової кислоти, яка цікавить, являє собою ендогенну послідовність нуклеїнової кислоти, де доцільні додаткові геномні копії ендогенної послідовності нуклеїнової кислоти, або являє собою молекулу нуклеїнової кислоти, яка в організмі хазяїна знаходиться в антисмисловій орієнтації відносно молекули нуклеїнової кислоти-мішені. Вектор: Молекула нуклеїнової кислоти, що вводиться в клітину, з отриманням трансформованої клітини. Вектор може включати послідовності нуклеїнової кислоти, які дають йому можливість реплікації в клітині-хазяїні, такі як послідовність початку реплікації. Приклади включають, зокрема, плазміду, косміду, бактеріофаг або вірус, які переносять екзогенну ДНК в клітину. Вектор також може включати один або декілька генів, антисмислових молекул і/або генів маркерів селекції і інших генетичних елементів, відомих в даній галузі. Вектор може трансдукувати, трансформувати або інфікувати клітину, викликаючи, таким чином, в клітині експресію молекул нуклеїнової кислоти і/або білків, що кодуються вектором. Вектор необов'язково включає матеріали, що допомагають в досягненні проникнення молекули нуклеїнової кислоти в клітину (наприклад, ліпосоми, кодовані білки і т. д.). IV. Сконструйовані садильні для спрямованого впливу на ген в рослинах А. Короткий опис У деяких варіантах здійснення, ELP вводяться в рослини, наприклад, для полегшення введення однієї або декількох додаткових молекул нуклеїнової кислоти, які цікавлять. Додаткові молекули нуклеїнової кислоти, які цікавлять, можуть включати, наприклад, будь-яку послідовність нуклеїнової кислоти, що кодує білок, який експресується в організмі-хазяїні. У додаткових варіантах здійснення, ELP використовуються для полегшення введення молекул нуклеїнової кислоти невідомої функції для з'ясування їх функції на основі ектопічної експресії гена. Ділянки, фланкуючі ELP, можуть бути гомологічні геномній послідовності нуклеїнової кислоти рослини-хазяїна, так що ELP інтегруються в геном рослини-хазяїна сайт-специфічним чином. ELP можуть використовуватися для інкорпорування молекул нуклеїнових кислот різних розмірів. У деяких варіантах здійснення, молекула нуклеїнової кислоти, спрямована до трансгенної ELP-мішені, може містити другу ELP для можливості продовження додавання генів в цільовий сайт. Одна або декілька ендонуклеаз спрямованої дії (наприклад, ZFN) можуть в даному процесі використовуватися послідовно або одночасно. Якщо доцільно, то один або декілька сайтів зв'язування з ендонуклеазою спрямованої дії (наприклад, сайт зв'язування з ZFN) можуть додатково додаватися до ділянок ELP, які гомологічні геномній послідовності нуклеїнової 6 UA 110472 C2 5 10 15 20 25 30 35 40 45 50 55 60 кислоти рослини-хазяїна. У деяких варіантах здійснення, сайти зв'язування ендонуклеаз спрямованої дії можуть бути додані так, щоб вони фланкували ELP. У даному останньому і в наступних варіантах здійснення, експресія ендонуклеаз спрямованої дії (наприклад, ZFN), які розщеплюють в доданих сайтах зв'язування, приводить до вирізання ELP з генома рослини. У деяких варіантах здійснення, ділянки, гомологічні геномній послідовності нуклеїнової кислоти рослини-хазяїна, також можуть використовуватися в комбінації з іншими фланкуючими ДНК, щоб була можливість вставки молекул нуклеїнової кислоти по сусідству з молекулою нуклеїнової кислоти, яка цікавить, введеною по сайту-мішені ELP, таких як, наприклад, елемент експресії гена, який цікавить, або ген, який цікавить. ДНК-розщеплення може використовуватися для посилення гомологічної рекомбінації по цьому сайту. У деяких варіантах здійснення, ділянки гомології в ELP, які гомологічні геномній послідовності нуклеїнової кислоти, також можуть використовуватися для спрямованої вставки молекул нуклеїнових кислот, яка полегшується за допомогою ферментів, які розщеплюють ДНК, таких як ZFN, мегануклеази або ендонуклеази спрямованої дії. У деяких варіантах здійснення, ELP можуть бути інкорпоровані в модифіковані хромосомні ділянки, такі які можуть бути генеровані за допомогою процесів ДНК-ампліфікації, або в мініхромосоми. У деяких варіантах здійснення, ділянки гомології і відповідно розташовані ендонуклеази спрямованої дії (наприклад, ZFN) використовуються для полегшення модифікації послідовностей, внутрішніх відносно фланкуючих ділянок, гомологічних геномній послідовності нуклеїнової кислоти рослини-хазяїна. В. Дизайн ELP У деяких варіантах здійснення, ELP можуть бути сконструйовані для полегшення гомологічної рекомбінації в хромосомних локалізаціях рослини. ELP можуть мати нейтральний вплив на навколишні їх гени або послідовності ДНК. ELP можуть характеризувати унікальні послідовності, здатні в геномі рослини піддаватися спрямованому впливу. У деяких варіантах здійснення, розмір (наприклад, 1 т. п. н.) для кожної 5'- і 3'-ділянки гомології з цільовою хромосомною локалізацією вибирають так, щоб він відповідав мінімальному розміру, який приблизно доцільний для полегшення спрямованої гомологічної рекомбінації. У деяких варіантах здійснення, розмір для кожної 5'- і 3'-ділянки гомології становить приблизно 50 п. н.; 100 п. н.; 200 п. н.; 400 п. н.; 600 п. н.; 700 п. н.; 750 п. н.; 800 п. н.; 1 т. п. н.; 1,2 т. п. н.; 1,4 т. п. н.; 1,6 т. п. н.; 1,8 т. п. н.; 2 т. п. н.; або 3 т. п. н. Немає необхідності, щоб в конкретних варіантах здійснення 5'- і 3'-ділянки гомології мали однаковий розмір. ELP-послідовності можуть бути отримані, наприклад, з використанням комп'ютерної програми для генерації випадкових чисел. ELP-послідовності, отримані, наприклад, за допомогою генерація випадкових послідовностей, потім можуть бути відібрані для використання як ELP на основі цільових характеристик, що включають зокрема послідовності, по суті або повністю позбавлені гомології з нативними геномними послідовностями організму-мішені. Відібрані ELP також можуть бути модифікованими, як описано детально нижче. У деяких варіантах здійснення, критерії для конструювання ELP і/або модифікації відібраних ELP можуть включати один або декілька з наступних: видалення сайтів рестрикції, що звичайно використовуються для клонування; зменшення кількості потенційних сайтів метилування (наприклад, CG і CNG); і відсутність будь-якої відкритої рамки зчитування більше ніж 300 п. н. Крім того, під час конструювання ELP, можуть бути видалені інші сайти в послідовностях нуклеїнових кислот, що включають, наприклад: сполуку екзон:інтрон (5'- або 3'-); сигнали додавання poly А; сигнали термінації РНК-полімерази; і високостабільні внутрішньолонцюжкові вторинні структури. Послідовності також можуть бути проаналізовані і модифіковані для зменшення частоти пар TA або CG. Блоки послідовності, які містять більше ніж приблизно шість послідовних залишків [G+С] або [А+Т], також можуть бути видалені з послідовності під час конструювання ELP. У варіантах здійснення, ELP можуть включати, наприклад, нуклеотидні послідовності формули X1-Y-X2, де Y являє собою щонайменше один сайт зв'язування ендонуклеази спрямованої дії (наприклад, сайт зв'язування ZEN), де X 1 являє собою відібрану нуклеотидну послідовність, позбавлену гомології з геномом організму-хазяїна і розташовану в 5'-області відносно Y, і де X2 являє собою відібрану нуклеотидну послідовність, позбавлену гомології з геномом організму-хазяїна, і яка відмінна від X1. Типові нуклеотидні послідовності X1 і X2 незалежно можуть бути вибрані з наступних послідовностей: SEQ ID NO:1; SEQ ID NO:2; SEQ ID NO:3; SEQ ID NO:4; SEQ ID NO:5; SEQ ID NO:6; SEQ ID NO:7; SEQ ID NO:8; SEQ ID NO:9; і SEQ ID NO:10. Додаткові модифікації, делеції або додавання до цих послідовностей (наприклад, видалення або додавання сайтів рестрикції) можуть бути зроблені в деяких варіантах здійснення для забезпечення доповненої або відмінної 7 UA 110472 C2 5 10 15 20 25 30 35 40 45 50 55 60 функціональності. Наприклад, в деяких варіантах здійснення: SEQ ID NO:1 може бути модифікована з отриманням SEQ ID NO:11; SEQ ID NO:3 може бути модифікована з отриманням SEQ ID NO:12; SEQ ID NO:2 може бути модифікована з отриманням SEQ ID NO:13; SEQ ID NO:4 може бути модифікована з отриманням SEQ ID NO:14; SEQ ID NO:5 може бути модифікована з отриманням SEQ ID NO:17; і SEQ ID NO:6 може бути модифікована з отриманням SEQ ID NO:18. У конкретних варіантах здійснення, типові ELP включають, зокрема, наступні послідовності: SEQ ID NO:15; SEQ ID NO:16; і SEQ ID NO:19. Сайти ферментів рестрикції (наприклад, сайти ферменту FseI) можуть бути введені, фланкуючи ELP з можливістю клонування ELP у відповідний вектор. Також можуть вводитися такі сайти ферментів рестрикції, фланкуючи ELP, які виробляють сумісні кінці при гідролізі ферментами рестрикції (наприклад, сайти BglII і BamHI), щоб була можливість зчеплення ELP разом, наприклад, у вигляді конкатамеру, в геномі рослини-хазяїна. Також сайти ферментів рестрикції можуть вводитися для можливості аналізу цікавлячих послідовностей нуклеїнових кислот в рослині-хазяїні, які потім спрямовуються до ELP за допомогою рекомбінації. Два або більше сайтів рестрикції можуть бути введені по флангах однієї ELP. Наприклад, сайти BglII і BamHI можуть бути включені на внутрішній ділянки відносно сайтів FseI. Також сайти ферментів рестрикції можуть вводитися для можливості аналізу послідовностей нуклеїнових кислот, які цікавлять, в рослини-хазяї, які спрямовуються до ELP для вставки за допомогою рекомбінації. Наприклад, сайти PmeI можуть бути введені по флангах відносно сайту вставки. С. Доставка молекул нуклеїнових кислот в рослинні клітини, що містять ELP Для можливості інтеграції, опосередкованої ендонуклеазами спрямованої дії, молекули нуклеїнової кислоти, яка цікавить, (наприклад, ELP і/або екзогенних молекул нуклеїнових кислот, спрямованих до раніше інтегрованих ELP) в геном рослини за допомогою спрямованої інтеграції, необхідна доставка ендонуклеаз спрямованої дії або молекул нуклеїнових кислот, що кодує ендонуклеази спрямованої дії, з подальшою експресією функціонального білка ендонуклеази спрямованої дії в геномі рослини-хазяїна. Молекула нуклеїнової кислоти, яка цікавить, також повинна бути присутньою в рослинній клітині в той момент, коли туди доставлена або коли там експресується ендонуклеаза спрямованої дії, так що функціональний білок ендонуклеази спрямованої дії може індукувати дволанцюжкові розриви в сайтах-мішенях, які потім піддаються репарації за допомогою спрямованої гомологічної інтеграції молекули нуклеїнової кислоти, яка цікавить, в цільовий локус. Фахівець в даній галузі може представити, що експресія функціонального білка ендонуклеази спрямованої дії може бути досягнута декількома методами, що включають, зокрема, трансгеноз конструкції, що кодує ендонуклеазу спрямованої дії, або транзиторну експресію конструкції, що кодує ендонуклеазу спрямованої дії. У обох випадках, експресія функціонального білка ендонуклеази спрямованої дії і доставка донорної ДНК в рослинну клітину можуть досягатися одночасно з метою керування спрямованою інтеграцією. Таким чином, в конкретних варіантах здійснення, ендонуклеази спрямованої дії (наприклад, ZFN) експресуються з молекул нуклеїнових кислот в трансформованих рослинах для введення дволанцюжкових розривів в геномі трансформованих рослин, наприклад, в одному або в декількох ELP, раніше введених в рослину. Можуть використовуватися такі ендонуклеази спрямованої дії, які націлені на одну або декілька послідовностей розпізнавання, які, згідно з конструкцією, присутні в ELP. Дизайн і вибір способів для конструювання ELP за винаходом можуть, таким чином, починатися з визначення однієї або декількох послідовностей нуклеїнових кислот, які розпізнаються ендонуклеазами спрямованої дії, що використовуються згодом для введення молекули нуклеїнової кислоти, яка цікавить, в ELP. Гнучкість і специфічність систем ZFN забезпечує рівень контролю, раніше не досяжний за допомогою відомих стратегій вирізання генів, опоседкованих рекомбіназою. Як приклад, ZFN легко можуть бути сконструйовані, наприклад, для розпізнавання специфічних послідовностей нуклеїнових кислот (наприклад, ELP). Wu et al. (2007) Cell. Mol. Life Sci. 64: 2933-44. Рандомізація кодонів для залишків розпізнавання "цинкових пальців" дає можливість селекції нових "цинкових пальців", які мають високу афінність до довільно вибраних послідовностей ДНК. Крім того, "цинкові пальці" являють собою природні ДНК-зв'язувальні молекули, і сконструйовані "цинкові пальці", як було продемонстровано, впливають на їх сконструйовані мішені в живих клітинах. Таким чином, нуклеази на основі "цинкових пальців" здатні спрямовуватися до специфічних, але довільних сайтів. Вимога димеризації доменів розщеплення химерних нуклеаз доменів "цинкові пальці" забезпечує високий рівень специфічності послідовності. Оскільки кожний набір з трьох "цинкових пальців" зв'язується з дев'ятьма послідовними парами основ, то дві химерні нуклеази 8 UA 110472 C2 5 10 15 20 25 30 35 40 45 50 55 ефективно справляються з мішенню 18 п. н., якщо кожний домен "цинковий палець" має ідеальну специфічність. Прогнозується, що будь-яка дана послідовність даного розміру є 9 унікальною в єдиному геномі (передбачається приблизно 10 п. н.). Bibikova et al. (2001) Mol. Cell. Biol. 21(l): 289-97; Wu et al. (2007) (вище). Крім того, додаткові "цинкові пальці" забезпечують підвищену специфічність, Beerli et al. (1998) Proc. Natl. Acad. Sci. USA 95: 1462833; Kim and Pabo (1998) Proc. Natl. Acad. Sci. USA 95: 2812-7; Liu et al. (1997) Proc. Natl. Acad. Sci. USA 94: 5525-30, так що кількість "цинкових пальців" в кожному ДНК-зв'язувальному домені може бути збільшена з отриманням додаткової специфічності. Наприклад, специфічність може бути додатково збільшена шляхом використання 4-пальцевих ZFN, які розпізнають послідовність 24 п. н. Urnov et al. (2005) Nature 435: 646-51. Таким чином, ZFN можуть використовуватися так, щоб послідовність розпізнавання в ELP, що вводиться в геном рослинихазяїна, була унікальною в геномі. Молекула нуклеїнової кислоти, яка цікавить, яку вводять за допомогою спрямованої рекомбінації в ELP, може бути функціонально пов'язана з одним або декількома рослинними промоторами, що керують експресією гена, в кількості, достатній для надання цільової властивості або фенотипа. Промотори, прийнятні для даного і для інших застосувань, добре відомі в даній галузі. Необмежувальні приклади, що описують такі промотори, включають патент США No. 6437217 (маїсовий RS81-промотор); 5641876 (рисовий промотор актину); 6426446 (маїсовий RS324-промотор); 6429362 (маїсовий PR-1-промотор); 6232526 (маїсовий A3-промотор); 6177611 (конститутивні промотори маїсу); 5322938, 5352605, 5359142, і 5530196 (35S-промотор); 6433252 (маїсовий промотор L3 олеосину); 6429357 (рисовий промотор актину 2, і рисовий інтрон актину 2); 5837848 (промотор, специфічний для коріння); 6294714 (промотори, що індукуються світлом); 6140078 (промотори, що індукуються сіллю); 6252138 (промотори, що індукуються патогеном); 6175060 (промотори, що індукуються дефіцитом фосфору); 6388170 (двоспрямовані промотори); 6635806 (промотор гамма-коїксину); і патентна заявка США з порядковим номером No. 09/757089 (маїсовий промотор адолази хлоропластів). Додаткові промотори включають промотор нопалін-синтази (NOS) (Ebert et al. (1987) Proc. Natl. Acad. Sci. USA 84(16): 5745-9); промотор октопін-синтази (OCS) (який переноситься на пухлиноіндукуючих плазмідах Agrobacterim tumefaciens); колимовірусні промотори, такі як 19Sпромотор вірусу мозаїки цвітної капусти (CaMV) (Lawton et al. (1987) Plant Mol. Biol. 9: 315-24); промотор CaMV 35S (Odell et al. (1985) Nature 313: 810-2; 35S-промотор вірусу мозаїки ранника (Walker et al. (1987) Proc. Natl. Acad. Sci. USA 84(19): 6624-8); промотор синтази сахарози (Yang and Russell (1990) Proc. Natl. Acad. Sci. USA 87: 4144-8); комплексний промотор гена R (Chandler et al. (1989) Plant Cell 1: 1175-83); промотор гена, що кодує білок, що зв'язує хлорофіл а/b; CaMV35S (патент США No. 5322938, 5352605, 5359142 і 5530196); FMV35S (патент США No. 6051753 і 5378619); PC1SV-промотор (патент США No. 5850019); SCP1-промотор (патент США No. 6677503); і промотори AGRtu.nos (реєстраційний номер GenBank No. V00087; Depicker et al. (1982) J. Mol. Appl. Genet. 1: 561-73; Bevan et al. (1983) Nature 304: 184-7), і так далі. Додаткові генетичні елементи, які необов'язково можуть бути функціонально пов'язані з молекулою нуклеїнової кислоти, яка цікавить, включають послідовності, що кодують транзитну пептиди. Наприклад, інкорпорування прийнятного транзитного пептиду хлоропластів, таких як А. thaliana EPSPS CTP (Klee et al. (1987) Mol. Gen. Genet. 210: 437-42), і Petunia hybrida EPSPS CTP (della-Cioppa et al. (1986) Proc. Natl. Acad. Sci. USA 83: 6873-7), як було продемонстровано, спрямовує гетерологічні послідовності білка EPSPS до хлоропластів в трансгенних рослинах. Монооксигеназа Dicamba (DMO) також може спрямовуватися до хлоропластів, як описано в Міжнародній публікації PCT No. WO 2008/105890. Додаткові генетичні елементи, які необов'язково можуть бути функціонально пов'язані з молекулою нуклеїнової кислоти, яка цікавить, також включають 5'UTR, розташовані між промоторною послідовністю і кодуючою послідовністю, яка функціонує як лідерна послідовність трансляції. Лідерна послідовність трансляції присутня в повністю процесованій мРНК в 5'області відносно послідовності старту трансляції. Лідерна послідовність трансляції може впливати на процесинг первинного транскрипту в мРНК, на стабільність мРНК і/або на ефективність трансляції. Приклади лідерних послідовностей трансляції включають лідери білка теплового шоку маїсу і петунії (патент США No. 5362865), лідери білків оболонки вірусів рослин, лідери рибулозобіфосфаткарбоксилази/оксигенази рослин і інші. Див., наприклад, Turner and Foster (1995) Molecular Biotech. 3(3): 225-36. Необмежувальні приклади 5'UTR включають GmHsp (патент США No. 5659122); PhDnaK (патент США No. 5362865); AtAntl; TEV (Carrington and Freed (1990) J. Virol. 64: 1590-7); і AGRtunos (реєстраційний номер GenBank No. V00087; і Bevan et al. (1983) Nature 304: 184-7). 9 UA 110472 C2 5 10 15 20 25 30 35 40 45 50 55 60 Додаткові генетичні елементи, які необов'язково можуть бути функціонально пов'язані з молекулою нуклеїнової кислоти, яка цікавить, також включають 3'-нетрансльовані послідовності, 3'-ділянки термінації транскрипції або ділянки поліаденілування. Це генетичні елементи, локалізовані в 3'-області полінуклеотидної молекули, і включають полінуклеотиди, які забезпечують сигнал поліаденілування і/або інші регуляторні сигнали, здатний впливати на транскрипцію, процесинг мРНК або на експресію гена. Сигнал поліаденілування функціонує в рослинах для того, щоб індукувати додавання поліаденілатних нуклеотидів до 3'-кінця мРНКпопередника. Послідовність поліаденілування може бути виділена з натурального гена, з множини рослинних генів або з генів Т-ДНК. Необмежувальним прикладом 3'-ділянки термінації транскрипції є 3'-ділянка нопалін-синтази (nos 3'; Fraley et al. (1983) Proc. Natl. Acad. Sci. USA 80: 4803-7). Приклад застосування різних не трансльованих 3'-ділянок пропонується в Ingelbrecht et al., (1989) Plant Cell 1: 671-80. Необмежувальні приклади сигналів поліаденілування включають сигнал з гена bcS2 Pisum sativum (Ps.RbcS2-E9; Coruzzi et al. (1984) EMBO J. 3: 1671-9) і AGRtu.nos (реєстраційний номер GenBank No. E01312). D. Трансформація клітин-хазяїнів молекулами нуклеїнової кислоти Для отримання трансформованої рослини згідно з винаходом може використовуватися будь-який з методів, відомих в даній галузі, для введення молекул нуклеїнових кислот в рослини, наприклад, для введення однієї або декількох ELP в геном рослини-хазяїна і/або для додаткового введення молекули нуклеїнової кислоти, яка цікавить. Прийнятні методи трансформації рослин включають будь-який метод, за допомогою якого ДНК може бути введена в клітину, такий як: електропорація, як проілюстровано в патенті США No. 5384253; бомбардування мікрочастинками, як проілюстровано в патентах США No. 5015580, 5550318, 5538880, 6160208, 6399861, і 6403865; трансформація, опосередкована Agrobacterium, як проілюстровано в патентах США No. 5635055, 5824877, 5591616; 5981840, і 6384301; і трансформація протопластів, представлена в патенті США No. 5508184. За допомогою застосування методів, таких як ці, віртуально, клітини будь-якого виду рослини можуть бути стабільно трансформовані, і ці клітини можуть бути регенеровані в трансгенні рослини за допомогою методів, відомих фахівцям в даній галузі. Наприклад, методи, які можуть бути конкретно застосовні в контексті трансформації бавовни, розкриті в патентах США No. 5846797, 5159135, 5004863, і 6624344; методи трансформації рослин Brassica конкретно розкриті, наприклад, в патенті США No. 5750871; методи трансформації сої розкриті, наприклад, в патенті США No. 6384301; і методи трансформації кукурудзи розкриті, наприклад, в патенті США No. 7060876, патенті США No. 5591616 і в Міжнародній публікації PCT WO 95/06722. Після здійснення доставки екзогенної ДНК клітинам-реципієнтам трансформовані клітини, як правило, ідентифікують для подальшого культивування і регенерації рослини. Для поліпшення здатності ідентифікації трансформантів, фахівцеві може бути доцільно застосувати селектований і скринований ген маркера разом з вектором трансформації, що використовується для генерування трансформанта. У цьому випадку, потенційно трансформована популяція клітин може бути проаналізована за допомогою експонування клітин з селективним агентом або агентами, або клітини можуть бути скриновані на присутність властивості, властивої цільовому гену маркера. Клітини, які життєздатні при експонуванні з селективним агентом, або клітини, які оцінили як позитивні в скринінговому аналізі, можуть культивуватися в середовищі, яке підтримує регенерацію рослин. У деяких варіантах здійснення, будь-яке прийнятне середовище для культивування тканин рослин (наприклад, середовище MS і N6) може бути модифіковане шляхом включення додаткових речовин, таких як регулятори росту. Тканина може підтримуватися на основному середовищі, що містить регуляторів росту доти, доки не буде доступно достатньо тканини для того, щоб почати робити зусилля для регенерації рослини, або після повторних раундів ручної селекції доти, доки морфологія тканини не буде прийнятною для регенерації (наприклад, щонайменше 2 тижня), потім тканину переносять в середовище, сприятливе для утворення пагонів. Культури переносять періодично доти, доки не відбувається достатнє утворення пагонів. Після того, як утворилися пагони, їх переносять в середовище, сприятливе для утворення коріння. Після утворення достатньої кількості коріння, рослини можуть бути перенесені в ґрунт для подальшого зростання і дозрівання. Для підтвердження присутності молекули нуклеїнової кислоти, яка цікавить, в регенерованій рослині може бути здійснена множина аналізів. Такі аналізи включають, наприклад: молекулярно-білогічні аналізи, такі як Саузерн- і Нозерн-блотинг і ПЛР; біохімічні аналізи, такі як детектування присутності білкового продукту. наприклад, за допомогою імунологічний методів (тІФА і/або Вестерн-блот) або за допомогою ферментативної функції; аналізи частин рослини, такі як аналізи листя або коріння; і аналіз фенотипу суцільної регенерованої рослини. 10 UA 110472 C2 5 10 15 20 25 30 35 40 45 50 55 60 Події спрямованої інтеграції можуть бути скриновані, наприклад, за допомогою ПЛРампліфікації з використанням, наприклад, олігонуклеотидних праймерів, специфічних для молекул нуклеїнової кислоти, які цікавлять. Зрозуміло, що ПЛР-генотипування включає зокрема ампліфікацію з використанням полімеразної ланцюгової реакції (ПЛР) геномної ДНК, отриманої з виділеної калюсної тканини рослини-хазяїна, яка приблизно містить цікавлячу молекулу нуклеїнової кислоти, інтегровану в геном, з подальшим стандартним клонуванням і аналізом послідовності продуктів ПЛР-ампліфікації. Методи ПЛР-генотипування добре описані (наприклад, Rios, G. et al. (2002) Plant J. 32: 243-53) і можуть застосовуватися до геномної ДНК, отриманої з будь-якого виду рослин або типу тканини, включаючи клітинні культури. Комбінації олігонуклеотидних праймерів, які зв'язуються як з послідовністю-мішенню, так і з введеною послідовністю, можуть використовуватися послідовно або одночасно в реакціях ПЛРампліфікації. Можливе створення олігонуклеотидних праймерів, сконструйованих для відпалу в сайті-мішені введених послідовностей нуклеїнової кислоти, і/або їх комбінація. Таким чином, стратегії ПЛР-генотипування можуть включати (але не обмежуючись цим) ампліфікацію специфічних послідовностей в геномі рослини, ампліфікацію множину специфічних послідовностей в геномі рослини, ампліфікацію неспецифічних послідовностей в геномі рослини або їх комбінацію. Фахівець в даній галузі може винайти додаткові комбінації праймерів і реакцій ампліфікації для перевірки генома. Наприклад, може бути сконструйований набір прямих і зворотних олігонуклеотидних праймерів для відпалу послідовностей нуклеїнових кислот, специфічних для мішеней за межами введеної послідовності нуклеїнової кислоти. Прямі і зворотні олігонуклеотидні праймери можуть бути сконструйовані для специфічного відпалу на введеній молекулі нуклеїнової кислоти, яка цікавить, наприклад, на послідовності, що відповідає кодуючій області всередині молекули нуклеїнової кислоти, яка цікавить, або відповіднає іншим частинам молекули нуклеїнової кислоти, яка цікавить. Ці праймери можуть використовуватися спільно з праймерами, описаними вище. Олігонуклеотидні праймери можуть бути синтезовані згідно з цільовою послідовністю і є комерційно доступними (наприклад, з Integrated DNA Technologies, Inc., Коралвілл, Айова). За ампліфікацією може йти клонування і секвенування або прямий аналіз продуктів ампліфікації. Фахівцеві в даній галузі очевидні альтернативні методи аналізу продуктів ампліфікації, отриманих в процесі ПЛР-генотипування. У одному варіанті здійснення, олігонуклеотидні праймери, специфічні для гена-мішені, застосовуються в ПЛР-ампліфікаціях. Е. Культивування і застосування трансгенних рослин Трансгенна рослина, що включає одну або декілька ELP і/або молекула нуклеїнової кислоти, яка цікавить, вставлена за допомогою спрямованої рекомбінації в ELP-сайт згідно з даним винаходом, може мати одну або декілька цільових властивостей. Такі властивості можуть включати, наприклад: стійкість до комах, інших шкідників і до агентів, що викликають захворювання; стійкість до гербіцидів; підвищену стабільність, врожайність або термін придатності; стійкість до впливу навколишнього середовища; виробництво лікарського засобу; виробництво промислового продукту; і підвищення поживних властивостей. Цільові властивості можуть додаватися однією або декількома молекулами нуклеїнової кислоти, які цікавлять, вставленими за допомогою спрямованої рекомбінації по сайту ELP, якими експресуються в рослині, що демонструє цільові властивості. Таким чином, в деяких варіантах здійснення, цільові властивості можуть виникнути завдяки присутності трансгена (трансгенів) в рослині, який вводиться в геном рослини по сайту ELP. У додаткових варіантах здійснення, цільова властивість може бути отримана за допомогою традиційного розведення, причому цільова властивість може додаватися за допомогою однією або декількох молекул нуклеїнової кислоти, вставлених за допомогою спрямованої рекомбінації по сайту ELP. Трансгенна рослина згідно з винаходом може являти собою будь-яку рослину, здатну до трансформації з використанням молекул нуклеїнової кислоти за винаходом. Відповідно, рослина може бути дводольною або однодольною. Необмежувальні приклади дводольних рослин, що використовуються в способах за даним винаходом, включають люцерну, квасолю, броколі, капусту, моркву, цвітну капусту, селеру, китайську капусту, бавовну, огірок, баклажан, салат, диню, горох, перець, арахіс, картоплю, гарбуз, хрін, рапс, шпинат, сою, гарбуз крупноплодовий, цукровий буряк, соняшник, тютюн, томат і кавун. Необмежувальні приклади однодольних рослин, що використовуються в способах за даним винаходом, включають кукурудзу, лук, рис, сорго, пшеницю, жито, просо, цукрову тростину, овес, тритикалє, просо прутоподібне, і газонну траву. Трансгенні рослини згідно з винаходом можуть використовуватися або культивуватися будьяким способом, де доцільна присутність ELP і/або молекул нуклеїнової кислоти, які цікавлять. Відповідно, серед іншого, можуть бути створені трансгенні рослини, що мають одну або 11 UA 110472 C2 5 10 15 20 25 30 35 40 45 50 55 декілька цільових властивостей, за допомогою трансформації молекулами нуклеїнової кислоти згідно з винаходом, і вони можуть оброблятися і культивуватися за допомогою будь-якого методу, відомого фахівцеві в даній галузі. ПРИКЛАДИ Наступні приклади включені для ілюстрації варіантів здійснення винаходу. Фахівцям в даній галузі зрозуміло, що методи, розкриті в прикладах, являють собою методи, розроблені авторами винаходу для успішного практичного функціонування винаходу. Однак фахівцям в даній галузі в світлі даного опису буде зрозуміло, що може бути зроблена множина змін в конкретних варіантах здійснення, які розкриті в даному документі, і, проте, будуть отримані подібні або аналогічні результати, не виходячи при цьому за рамки винаходу. Більш конкретно, очевидно, що існують певні агенти, які як хімічно, так і фізіологічно можуть замінювати агенти, описані в даному документі, і при цьому будуть досягатися такі ж або подібні результати. Передбачається, що як такі, подібні заміни і модифікації, очевидні фахівцям в даній галузі, знаходяться в рамках винаходу, визначеного за допомогою прикладеної формули винаходу. Приклад 1: Конструкція вектора Синтезували дві різні ELP, які складаються з 5'-ділянки гомології розміром 1 т. п. н. ("сконструйована ділянка садильного майданчика 1" (SEQ ID NO:11) в pDABl00610/pDAB 100640 і "сконструйована синтетична гомологічна ділянка 3" (SEQ ID NO:12) в pDAB10061l/pDAB100641) і 3'-ділянка гомології розміром 1 т. п. н. ("сконструйована ділянка садильного майданчика 2" (SEQ ID NO:13) в pDAB100610 і "сконструйована синтетична гомологічна ділянка 4" (SEQ ID NO:14) в pDAB100611/pDAB100641). Ці ділянки гомології розділені двома різними ТОЧНИМИ сайтами зв'язування Нуклеази домену "цинкові пальці" (eZFN). Дві ELP, що використовуються для pDAB100610 і pDAB100611, позначені ELP1 (SEQ ID NO:15) і ELP2 (SEQ ID NO:16), відповідно, переносили з використанням реакції за допомогою GATEWAY® LR-клонази (Invitrogen, Карлсбад, Каліфорнія) в pDAB100704, вектор подвійного призначення, який отриманий з pSB11 (WO94/00977 і WO95/06722). pDAB100704 містить промотор ZmUbil (промотор, 5'-нетрансльована ділянка (UTR) і інтрон, отримана з промотору убіквітину 1 Zea mays (Christensen et al, (1992) Plant Molecular Biology, 18(4); 675-89)), ген маркера селекції AAD-1 (синтетичний, оптимізований для рослин варіант гена арилоксіалканоат діоксигенази з Sphingobium herbicidovorans (ATCC® 700291), що кодує фермент з альфакетоглутатарт-залежною діоксигеназною активністю, який надає стійкості до арилоксифеноксипропіонатних гербіцидів (WO 2005/107437, WO 2008/141154A2, і US 2009/0093366, кожний з яких включений в даний документ посиланням), і ZmLip 3'UTR (3'нетрансльована ділянка (UTR), що включає термінатор транскрипції і сайт поліаденілування гена LIP Zea mays; реєстраційний номер GenBank L35913). Дві ELP, що використовуються для pDAB 100640 і pDAB 100641, позначені ELP1 (SEQ ID NO:15) і ELP2 (SEQ ID NO:16), відповідно, переносили з використанням реакції за допомогою GATEWAY® LR-клонази (Invitrogen, Карлсбад, Каліфорнія) в pDAB101849, бінарний вектор, pDAB101849 містить промотор OsActl (промотор і інтрон, отриманий з рисового промотору актину (патент США No. 5641876)), ген маркера селекції pat (ген фосфінотрицинацетилтрансферази; Wohlleben et al, (1988) Gene 70: 25-37), і ZmLip 3'UTR (3'-нетрансльована ділянка (UTR), що включає термінатор транскрипції і сайт поліаденілування гена LIP Zea mays; реєстраційний номер GenBank L35913). Отримані в результаті клони, pDAB100610 (фіг. 1), pDAB100611 (фіг. 2), pDAB100640 (фіг. 3), і pDAB100641 (фіг. 4) підтверджували секвенуванням. Вектори pDAB100610 і pDAB100611 переносили в Agrobacterium tumefaciens, штам LBA4404, що несе бінарну плазміду pSBl, і Тланцюжкову ділянку інтегрували в pSBl за допомогою гомологічної рекомбінації. Вектори pDAB100640 і pDAB100641 переносили в Agrobacterium tumefaciens, штам LBA4404. Структуру кожної плазміди підтверджували за допомогою гідролізу ферментами рестрикції. Вектори, що містять молекули нуклеїнової кислоти, які цікавлять, (донорна ДНК) збирали шляхом вставки експресуючої касети YFP і або PAT- або AAD-1-експресуючої касети між укороченими варіантами 5'- і 3'-ділянок гомології (позначені як варіанти 2 (v2)) для ELP1 і ELP2. Експресуюча касета PAT містить промотор OsActl (рисовий промотор актину; McElroy et al, (1990) Plant Cell 2: 163-171), який використовували для експресії гена pat (генів фосфінотрицинацетилтрансферази; Wohlleben et al, (1988) Gene 70: 25-37), який фланкований 3'нетрансльованою ділянкою ZmLip. Експресуюча касета AAD-1 містить промотор ZmUbi 1 для керування геном aad-l і ZmPer5 3'UTR (патент США No. US 6384207). Експресуюча касета YFP складається з промотору ZmUbil, який використовується для керування експресії гена PhiYFP 12 UA 110472 C2 5 10 15 20 25 30 35 40 45 50 55 60 (жовтий флуоресцентний білок (патент Заявка США US2007/0298412)) і фланкований ZmPer5 3'UTR (патент США No. US 6384207). Експресуючі касети PAT і YFP клонували між укороченими варіантами "сконструйованого садильного майданчика 1" і "сконструйованим садильним майданчиком 2" в ELP1 (pDAB105955, фіг. 5). Альтернативно, експресуючі касети PAT і YFP клонували між укороченими варіантами "сконструйованого садильного майданчика 3" і "сконструйованим садильним майданчиком 4" в ELP2 (pDAB 105956, фіг. 6). Аналогічно, експресуючі касети AAD-1 і YFP клонували між укороченими варіантами "сконструйованого садильного майданчика 1" і "сконструйованим садильним майданчиком 2" в ELP1 (pDAB105979, фіг. 7). Альтернативно, експресуючі касети AAD-1 і YFP клонували між укороченими варіантами "сконструйованого садильного майданчика 3" і "сконструйованим садильним майданчиком 4" в ELP2 (pDAB105980, фіг. 8). Донорну ДНК клонували в бінарний вектор для біолістичної трансформації, для трансформації, опосередкованої волокнами карбіду кремнію або Agrobacterium-опосередкованої трансформації. Крім того, конструкції ДНК можуть бути проклоновані у висококопійну плазміду (наприклад, похідні pBR322, що несуть послідовність початку реплікації ColEl) для генерування плазмідною ДНК, що використовується для прямої доставки ДНК, а саме для трансформації. Приклад 2: Доставка ДНК, опосередкована волокнами карбіду кремнію Ембріогенні клітинні культури Hi-II маїсу отримували, як описано в патенті США No. 7179902, і використовували як джерело живих рослинних клітин, в яких здійснювали як приклад спрямовану інтеграцію. Фрагменти, що містять касету з маркером селекції рослин AAD-1 а також ELPl і ELP2, відповідно, з pDAB100610 і pDAB 100611, і фрагменти, що містять касету з маркером селекції рослин PAT, а також ELPl і ELP2, відповідно, з pDAB100640 і pDAB100641, використовували для генерування трансгенних подій. Трансгенні події виділяли і охарактеризували. Ці події потім ставали мішенню з використанням спрямованої гомологічної рекомбінації, де цікавлячу молекулу нуклеїнової кислоти можна було інтегрувати всередині сконструйованого садильного майданчика. 12 мл об'єму осаджених клітин (PCV) із збереженої раніше за допомогою кріоконсервації клітинної лінії разом з 28 мл кондиціонованого середовища пересівали в 80 мл рідкого середовища GN6 (N6 середовище (Chu et al, (1975) Sci Sin. 18: 659-668), 2 мг/л 2,4-D, 30 г/л сахарози, pH 5,8) в 500 мл конічної колби Ерленмейєра, і поміщували на шейкер при 125 об/хв при 28 °C. Цю стадію повторювали два рази з використанням тієї ж клітинної лінії, так щоб в цілому 36 мл PCV було розподілено по трьох колбах. Через 24 години, рідке середовище GN6 видаляли і замінювали на 72 мл GN6 S/M осмотичного середовища (середовище N6, 2 мг/л 2,4D, 30 г/л сахарози, 45,5 г/л сорбіту, 45,5 г/л маніту, 100 мг/л міоїнозиту, pH 6). Колбу інкубували в темряві протягом 30-35 хвилин при 28 °C з помірним перемішуванням (125 об/хв). Протягом періоду інкубації отримували 50 мг/мл суспензії волокон карбіду кремнію (Advanced Composite Materials, LLC, Грір, Південна Кароліна) шляхом додавання 8,1 мл GN6 S/M рідкого середовища до 405 мг стерильних волокон карбіду кремнію. Після інкубації в осмотичному середовищі GN6 S/M, вміст кожної колби об'єднували в 250мл центрифугальну пробірку. Після того, як всі клітини в колбі осідали на дно, вміст об'єму в надлишку, що становить приблизно 14 мл рідини GN6 S/M, витягували і збирали в стерильну 1л колбу для майбутнього використання. Попередньо зволожену суспензію волокон змішували при максимальній швидкості на вортексі протягом 60 сек, і потім додавали в центрифугальну пробірку. 170 мкг очищеного фрагмента або з pDAB100610 або з pDAB100611 плазмідної ДНК додавали в кожну пробірку. Після додавання ДНК пробірку негайно вміщували в модифікований комерційний змішувач для фарби Red Devil 5400 (Red Devil Equipment Co., Плімут, Міннесота), і перемішували протягом 10 сек. Після перемішування коктейль клітин, середовище, волокна і ДНК додавали до вмісту 1-л колби разом з 125 мл свіжого рідкого середовища GN6 для зниження осмотичної дії. Клітини можна було витягнути на шейкері, встановленому на 125 об/хв протягом 2 годин. Шість мілілітрів диспергованої суспензії фільтрували на фільтрувальному папері Ватман #4 (5,5 см) з використанням скляного елемента колектора клітин, з'єднаного з центральною вакуумною лінією, так що на одну пробірку отримували 60 фільтрів. Фільтри поміщували на чашки 60×20 мм твердого середовища GN6 (таке ж, як рідке середовище GN6 за винятком 2,5 г/л гелеутворювального агента Гелрит (Gelrite)) і культивували при 28 °C в умовах темряви протягом 1 тижня. Приклад 3: Ідентифікація і виділення передбачуваних трансгенних подій 13 UA 110472 C2 5 10 15 20 25 30 35 40 45 50 55 60 Через один тиждень після доставки ДНК, фільтрувальний папір переносили на чашки 60×20 мм селекційного середовища GN6 (1H) (N6 середовище, 2 мг/л 2,4-D, 30 г/л сахарози, 100 мг/л міо-інозиту, 2,5 г/л Гелриту, pH 5,8), що містить відповідний селективний агент. Ці селекційні чашки інкубували при 28 °C протягом одного тижня в темряві. Через 1 тиждень після селекції в темряві, тканину впроваджували на свіже середовище шляхом зішкрібання ½ клітин з кожної чашки в пробірку, що містить 3 мл агарозного середовища GN6, що підтримується при 37-38 °C ® (середовище N6, 2 мг/л 2,4-D, 30 г/л сахарози, 100 мг/л міо-інозиту, 7 г/л SEAPLAQUE агарози, pH 5,8, автоклавована протягом 10 хвилин при 121 °C). Суміш агарози/тканини руйнували за допомогою шпателя і потім рівномірно розподіляли 3 мл суміші агароза/тканина на поверхні чашки петрі 100×15 мм, що містить середовище GN6 (1H). Цей процес повторювали для обох половин кожної чашки. Після впровадження всієї ® тканини чашки індивідуально герметично закривали за допомогою NESCOFILM або ® PARAFILM M, і культивували при 28 °C в умовах темряви протягом терміну до 10 тижнів. Приблизно трансформовані ізоляти, які росли в даних умовах селекції, видаляли із вставлених чашок і переносили на свіже селекційне середовище в чашки 60×20 мм. Якщо стабільний ріст був очевидний через приблизно 2 тижні, то передбачали, що подія стійка до застосовуваного гербіциду (селективний агент) і аліквоту клітин потім збирали для аналізу генотипування. Приклад 4: Молекулярна характеризація подій Виділення геномної ДНК Геномну ДНК (гДНК) виділяли з виділених клітин маїсу і використовували як матрицю для експериментів ПЛР-генотипування. гДНК виділяли з приблизно 100-300 мкл об'єму осаджених клітин (PCV) калюсу Hi-II, які виділяли згідно з протоколами виробника, детально описаним в ® наборі реагентів DNEASY 96 Plant Kit (QIAGEN Inc., Валенсія, Каліфорнія). Геномну ДНК елюювали в 100 мкл буферу елюції з набору реагентів з отриманням кінцевої концентрації 20200 нг/мкл, і потім аналізували за допомогою методів генотипування на основі ПЛР. Молекулярний аналіз кількості піків Визначення кількості піків трансгена за допомогою аналізу гідролізу зонда, аналогічного ® аналізу TAQMAN , здійснювали за допомогою ПЛР в реальному часі з використанням системи ® LIGHT CYCLER480 (Roche Applied Science, Індіанаполіс, Індіана). Аналізи розробляли для генів AAD-1 і PAT і для гена внутрішнього контролю з використанням комп'ютерної програми ® LIGHTCYCLER Probe Design Software 2.0. Для ампліфікації, Майстер-мікс для зондів системи LIGHT CYCLER®480 (Roche Applied Science, Індіанаполіс, Індіана) отримували в 1x кінцевій концентрації в 10 мкл об'єми багатокомпонентної реакції, що містить 0,4 мкМ кожного праймера і 0,2 мкМ кожного зонда. Двостадійну реакцію ампліфікації здійснювали з протягуванням при 58 °C протягом 38 секунд з набуванням флуоресценції. Всі зразки проганяли по три рази і середні порогові значення для циклів (Ct) використовували для аналізу кожного зразка. Аналіз даних ПЛР в реальному часі здійснювали з використанням комп'ютерної програми ® LIGHTCYCLER версії 1.5 з використанням модуля визначення відносної кількості на основі методу ΔΔCt. Для цього зразок геномної ДНК з калібратора однієї копії і відомий 2-копійний ® контроль включали в кожен прогін (ідентично тому, як використовувалося в аналізі INVADER вище). На основі цих даних виявлену кількість піків AAD-1 або PAT потім оцінювали для кожного зразка. Дизайн праймерів для ПЛР-генотипування Олігонуклеотидні праймери синтезували (наприклад, за допомогою Integrated DNA Technologies, Inc. (Коралвілл, Айова)) при умовах стандартного знесолення і розведення за допомогою води до концентрації 100 мкМ. Олігонуклеотидний праймер конструювали для відпалу на кінцеві ділянки ДНК-вставки. Праймери тестували з використанням розведення плазмідної ДНК в присутності ДНК, виділеної з нетрансгенних організмів. Вставку pDAB100610, pDAB100611, pDAB100640 і pDAB100641 ампліфікували за допомогою ПЛР з геномною ДНК передбачуваних трансгенних подій з використанням праймерів. Отриманий в результаті фрагмент клонували в плазмідний вектор і секвенували для підтвердження того, що сконструйований садильний майданчик повністю інтегрувався в геном рослини в процесі трансформації. Саузерн-блот-аналіз Саузерн-аналіз здійснювали для підтвердження кількості піків трансгена. Для цього аналізу геномну ДНК гідролізували за допомогою відповідних ферментів рестрикції і гібридизували із зондами. Тканину маїсу ідентифікували як таку, що містить передбачувану ELP за допомогою локусспецифічної реакції ПЛР, яку адаптували для Саузерн-блот-аналізу. Для Саузерн-аналізу зразки тканини збирали в 2-мл пробірки eppendorf і ліофілізували протягом 2 днів. Мацерацію 14 UA 110472 C2 5 10 15 20 25 30 35 40 45 50 55 тканини здійснювали за допомогою гомогенізатора тканини Kleco і вольфрамових кульок (Kleco, Вісалія, Каліфорнія). Після мацерації тканини геномну ДНК виділяли з використанням набору реагентів DNeasy Plant Mini kit (Qiagen, Германтаун, Меріленд) згідно з протоколом виробника, що пропонується. Геномну ДНК оцінювали кількісно за допомогою аналітичного набору реагентів Quant-IT Pico Green DNA (Molecular Probes, Invitrogen, Карлсбад, Каліфорнія). ДНК з певною кількістю доводили до 4 мкг для Саузерн-блот-аналізу і гідролізували з використанням відповідного ферменту рестрикції протягом ночі при 37 °C. Гідролізовану ДНК очищали з використанням Quick-Precip (Edge BioSystem, Гайтерсбург, Меріленд) згідно з протоколом виробника, що пропонується. Осаджену ДНК ресуспендували в 10X барвнику і піддавали електрофорезу протягом 17 годин на 0,8 % SeaKem LE агарозному гелі (Lonza, Рокленд, Меріленд) при напрузі 40 вольт. ДНК переносили на нейлонові заряджені мембрани (Millipore, Бедфорд, Массачусетс) протягом ночі і зв'язували з мембраною з використанням лінкера UV Strata 1800 (Stratagene, ЛаХойя, Каліфорнія). Блоти передгібридизували з 20 мл PerfectHyb Plus (Sigma, Сент Луїс, Міссурі) і гібридизували з відповідним радіоактивно-міченим зондом протягом ночі. Блоти відмивали і вміщували для візуалізації на екрани люмінофор на 24 години і потім аналізували з використанням сканера STORM 860 (Molecular Dynamics). Приклад 5: Селекція трансгенних подій з вставкою ДНК Події скринували за допомогою аналізу з гідролізом зонда і за допомогою ПЛР, як описано вище для інтактної рослинної транскрипційної одиниці (PTU), що містить касети з генами AD-1 або PAT і інтактними ELP. Кількість піків далі підтверджували за допомогою Саузерн-аналізу з використанням стандартних методів з використанням зондів для генів AAD-1 і PAT і для ELP. Калюс з селектованих трансгенних подій, що несуть одну копію інтактних вставок з pDAB100610 і pDAB100611, зберігали для подальшого тестування експресії гена AAD-1. За допомогою скринінгу з використанням методу кРЧ-ПЛР (кількісний аналіз з використанням полімеразної ланцюгової реакції в режимі реального часу, qRT-PCR) для AAD-1 і тІФА (нижче) ідентифікували події експресії AAD-1. Аналіз кРЧ-ПЛР подій, експресуючих AAD-1 Кількісний аналіз ПЛР в реальному часі (кРЧ-ПЛР) використовували для кількісної оцінки мРНК-експресії гена AAD-1. Аналіз розробляли для кількісної оцінки відносної експресії мРНК AAD-1 із зразків трансгенного калюсу Hi-II за допомогою нормалізації цього рівня відносно експресії мРНК гена внутрішнього контролю. Нормалізація мРНК AAD-1 відносно мРНК гена внутрішнього контролю дає можливість порівняння експресії AAD-1 між різними зразками, і може використовуватися для ідентифікації подій, які, як виявлено, є високо-експресованими. Сумарну РНК отримували зі свіжої тканини калюсу з використанням набору реагентів Qiagen ® RNeasy 96 Kit (Qiagen, Валенсія, Каліфорнія). РНК тестували за допомогою вільної від РНК-ази ДНК-ази згідно з інструкціями набору реагентів для видалення забруднень геномної ДНК. Синтез першого ланцюга проводили згідно з інструкціями виробника для ферменту Зворотна ® транскриптаза III (Invitrogen, Карлсбад, Каліфорнія) і затравлювали синтез за допомогою випадкових гексамерів. Синтезовані ланцюги кДНК розводили у воді в співвідношеннях 1:10 і 1:50 (це забезпечує достатню кількість матриці для ПЛР-ампліфікації множини мішеней). Кожну аліквоту зберігали при -20 °C на невизначений термін. Суміші реакцій кРЧ-ПЛР складали для ампліфікації кДНК AAD-1, як указано далі: 7,5 мкл 2X LC480 Майстра-буфера для зондів (Roche Diagnostic, Індіанаполіс, Індіана), 0,3 мкл прямого праймера, специфічного для гена, з 10 мкМ концентрованого розчину, 0,3 мкл зворотного праймера, специфічного для гена, з 10 μΜ® концентрованого розчину, 0,15 мкл UPL-зонда з концентрованого розчину зондів LightCycler 480, Roche Diagnostic, Індіанаполіс, Індіана) 1,5 мкл 10 % (мас./об.) полівінілпіролідону-40 (PVP40), і 3,9 мкл води. UPL-зонд (Roche Diagnostics, Індіанаполіс, США) блокує нуклеїнову кислоту і, таким чином, має вищу Tm, ніж якщо розраховувати по-іншому. Всі компоненти вміщували назад в морозильник перед стандартними або іншими маніпуляціями. 384-ямковий мікропланшет розмічали і маркували, додаючи в кожну ямку 13,5 мкл мастер-міксу. Герметичною плівкою акуратно обгортали мікропланшет. Планшет центрифугували протягом 1 хвилини при 3000 об/хв в центрифузі Qiagen для мікропланшетів. Додавали 1,5 мкл відморожених, розведених і синтезованих кДНК-ланцюгів. Крім того, додавали в окрему ямку 1,5 мкл стандартів кількості піків плазмідної ДНК способом серійного розведення від найнижчої концентрації до найвищої, ці стандарти порівнювали з кДНК AAD-1 (синтезованою з сумарної мРНК) для оцінки кількості піків. Серії стандартів кількості піків ДНК AAD-1 отримували шляхом клонування амплікону-мішені в плазміду pCR2.1 (Invitrogen, Карлсбад, Каліфорнія) і шляхом отримання серійного розведення для оцінки кількості піків. Плівку міцно фіксували на планшет і центрифугували, як описано 15 UA 110472 C2 5 10 15 20 25 30 35 40 45 50 55 60 раніше. Задавали ПЛР-програму і ДНК ампліфікували за допомогою приладу для ПЛР в реальному часі LC480 (Roche, Індіанаполіс, Індіана) або в еквівалентному йому. Приклад 6: Визначення білка AAD-1 в тканинах маїсу за допомогою тІФА Був розроблений спосіб кількісного визначення білка AAD-1 в калюсі маїсу з використанням твердофазного імуноферментного аналізу (тІФА). Спосіб, описаний в даному документі, може використовуватися для детектування білка AAD-1 і для аналізу зразків тканин рослини з трансгенних калюсів маїсу. Білок AAD-1 виділяли із зразків маїсу з використанням калюс-специфічних буферів на основі фосфатно-сольового буферного розчину, що містить 0,05 % Tween 20 (PBST), і що можливо містить бичачий сироватковий альбумін, протеазні інгібітори або аскорбінову кислоту. Екстракт центрифугували; водну надосадову рідину збирали, розводили і аналізували з використанням AAD-1-специфічного аналізу тІФА. Одночасно в цьому аналізі застосовували формат сендвічтифу. Аліквоту розведеного зразка і біотинільоване моноклональне антитіло, зв'язуюче AAD-1, (MAb 473F 185) інкубували в ямку мікропланшета з покриттям у вигляді імобілізованого моноклонального антитіла, зв'язуючого AAD-1 (MAb 473H274). Ці антитіла, зв'язані з білком AAD-1, що експресується маїсом, в ямку з утворенням "сендвіча" з білком AAD-1, зв'язаним між розчинним і імобілізованим антитілом. Незв'язані зразки і кон'югат потім видаляли з планшета шляхом промивання за допомогою PBST. Надлишок кількості кон'югату стрептавідину і ферменту (лужна фосфатаза) додавали до ямки для інкубування. Присутність AAD-1 детектували за допомогою інкубуваня ферментного кон'югату з ферментним субстратом; генеруючи забарвлений продукт. Оскільки AAD-1 був зв'язаний в сендвічі антитіл, то рівень виявленого фарбування пов'язаний з концентрацією AAD-1 в зразку (тобто чим менша концентрація білка, тим менше виявлення фарбування). Поглинання при 405 нм вимірювали з використанням планшетного рідера. Високоекспресуючі AAD-1 події ідентифікували з селектованих трансгенних подій і зберігали для подальшого спрямованого впливу донорної ДНК. Приклад 7: Опосередкована біолістикою доставка ДНК в рослинні клітини, що містять ELP Регенерація трансгенних подій, що містять ДНК-мішень Події з pDAB100160 і pDAB100611, які, як було підтверджено, є низькокопійними і містять інтактні PTU, регенерували з отриманням незрілого ембріонального донорного матеріалу для спрямованого впливу. Здорову вирощену тканину переносили спочатку в 28+100 галоксифоп (haloxyfop) (середовище MS (середовище Мурасіге-Скуга (1962) Physiol Plant 15: 473-497), 0,025 мг/л 2,-4D, 5 мг/л BAP, 0,0362 мг/л галоксифоп, 30 г/л сахарози, 2,5 г/л гелрит (gelrite), pH 5,7) і 2 інкубували при низькому освітленні (фотоперіод 14 мкЕ/м сек 16 годин) протягом 7 днів з 2 подальшим високим освітленням (фотоперіод 89 мкЕ/м сек 16 годин) протягом наступних 7 днів. Позеленілі структури переносили в середовище галоксифоп 36+100 (те ж середовище 28+100 галоксифоп без BAP і 2,4-D) і інкубували при високому освітленні (фотоперіод 40 2 мкЕ/м сек 16 годин) доти, доки структури пагонів не утворять достатні кореневі структури для пересаджування в теплицю. Рослини вирощували до дозрівання в теплиці з використанням ґрунту у вигляді 95 % Metro-Mix 360о і 5 % глини і запилювали залежно від стану рослини. Енергійно зростаючі рослини, самозапилені або перезапилені (рослини від однієї і тієї ж події) і менш енергійно зростаючі рослини схрещували з Hi-II, A188 або з B104 для збереження ембріогенної здатності донорного матеріалу. Приклад 8: Опосередкована біолістикою доставка ДНК в рослинні клітини, що містять ELP Спрямований вплив на події, отримані з pDAB100610 і pDAB100611. Спрямований вплив донорної послідовності здійснювали з використанням двох різних протоколів трансформації. Для трансгенних подій з інтактними ELP і які містять AAD-1 як маркер селекції (події pDAB100610 і pDAB 100611), спрямований вплив здійснювали за допомогою опосередкованої біолістикою доставки ДНК в передкалюсні незрілі ембріони. Ембріони розміром 1,2-2 мм збирали через 10-13днів після запилення і поміщували на середовище N6E (N6-середовище, 2 мг/л 2,4-D, 2,8 г/л L-проліну, 100 мг/гідролізату казеїну, 100 мг/л міо-інозиту, 4,25 мг/л нітрату срібла, 30 г/л сахарози, pH 5,8) і інкубували при 28 °C протягом 2-3 днів в темряві. Набряклі ембріони переносили на середовище N60SM (те ж середовище N6E з додаванням 36,4 г/л сорбіту, 36,4 г/л маніту і L-проліну, із зменшеними концентраціями до 0,7 г/л), яка заповнювала коло діаметром 2,5 см на фільтрувальному папері Ватман # 4. Використовували екран 1000 мкm для утримування ембріонів для бомбардування, і ембріони інкубували в темряві при 28 °C протягом 4 годин перед бомбардуванням. Для покриття біолістичних частинок ДНК, 3 мг золотих частинок діаметром 0,6 мікрон промивали один раз за допомогою 100 % етанолу, два рази за допомогою стерильної дистильованої води і 16 UA 110472 C2 5 10 15 20 25 30 35 40 45 50 55 60 ресуспендували в 50 мкл води в силіконізованих мікроцентрифугальних пробірках. Всього 5 мкг окремих векторів плазмідної ДНК, що кодує нуклеазу для доменів "цинкові пальці", і фрагмент Донорної ДНК; pDAB 105955 або pDAB 105956), 20 мкл спермідину (0,1 M) і 50 мкл хлориду кальцію (2,5 M) додавали окремо до суспензії золота і змішували на вортексі. Суміш інкубували при кімнатній температурі протягом 10 хв, осаджували при 10000 об/хв в настільній мікроцентрифузі протягом 10 секунд, ресуспендували в 60 мкл холодного 100 %-етанолу і 8-9 мкл розподіляли на кожний макроносій. Бомбардування проводили з використанням Біолістичної системи Biolistic PDS-1000/HE™ (Bio-Rad Laboratories, Геркулес, Каліфорнія). Планшети, що містять ембріони, вміщували на ++ середню полицю при умовах тиску 650 фунтів на квадратний дюйм і 27 дюймів Hg і бомбардували один раз, слідуючи інструкціям виробника. Ембріони через двадцять чотири години після бомбардування переносили безпосередньо на середовище (без фільтрувального паперу) N6E (той же склад, як указано вище) для регенерації і інкубували протягом 13 днів при 28 °C в темряві. Ембріони переносили на селекційне середовище N6S (N6-середовище, 2 мг/л 2,4-D, 100 мг/л міоінозиту, 0,85 мг/л нітрату срібла, 2 мг/л біалафосу, 30 г/л сахарози, 2,5 г/л гелриту і при pH 5,8) протягом кожних 2 тижнів по 3 перенесення на селекційне середовище і інкубували при 28 °C в умовах темряви терміном до 10 тижнів. Приблизно трансформовані ізоляти, які росли в даних умовах селекції, видаляли із вставлених чашок і переносили в свіже селекційне середовище на чашки 60×20 мм. Якщо стабільний ріст був очевидний через приблизно 2 тижні, то передбачали, що подія стійка до застосовуваного гербіциду (селективний агент) і аліквоту клітин потім збирали для аналізу генотипування (аналіз описаний нижче). Спрямований вплив на події, отримані з pDAB100640 і pDAB100641. Для трансгенних подій з інтактними ELP і які містять PAT як маркер селекції (pDAB100640 і pDAB100641), спрямований вплив здійснювали за допомогою опосередкованої біолістикою доставкою ДНК в ембріогенний калюс маїсу. Через шість-вісім днів після субкультивування, ембріогенну маїсову тканину (приблизно 0,4 мл PCV клітин) розподіляли тонким шаром по окружності діаметром 2,5 см на поверхні фільтрувального паперу Ватман #4, поміщеної на чашки петрі 100×15 мм, що містять GN6 S/M середовище (N6-середовище, 2 мг/л 2, 4-D, 30 г/л сахарози, 45,5 г/л сорбіту, 45,5 г/л маніту, 100 мг/л міо-інозиту, pH 5,8) отверджуване за допомогою 2,5 г/л гелриту. Клітини інкубували в темряві протягом 4 годин. ДНК готували для бомбардування, як описано раніше, pDAB105979 і pDAB105980 використовували для спрямованого впливу. Бомбардування проводили з використанням Біолістичної системи Biolistic PDS-1000/HE™ (Bio-Rad Laboratories, Геркулес, Каліфорнія). Планшети, що містять клітини, вміщували на ++ середню полицю при умовах тиску 1100 фунтів на квадратний дюйм і 27 дюймів Hg і бомбардували один раз, слідуючи інструкціям виробника. Через двадцять чотири години після бомбардування, фільтрувальний папір, що містить клітини, переносили на тверде середовище GN6 (N6-середовище, 2 мг/л 2, 4-D, 30 г/л сахарози, 100 мг/л міо-інозиту, pH 5,8), отверджуване за допомогою 2,5 г/л Гелриту, і інкубували протягом 24 годин при 28 °C в умовах темряви. Через двадцять чотири години після періоду регенерації, фільтрувальний папір, що містить клітини, переносили на тверде середовище GN6+100 галоксифоп (N6-середовище, 2 мг/л 2,4-D, 30 г/л сахарози, 100 мг/л міо-інозиту, pH 5,8), отверджуване за допомогою 2,5 г/л Гелриту, і інкубували протягом 24 годин при 28 °C в умовах темряви. Перенесення продовжували кожні 2 тижні по 3 перенесення на селекційне середовище. Тканину інкубували протягом терміну до 12 тижнів доти, доки не почнуть з'являтися передбачувані трансгенні ізоляти, отримані в результаті інтеграції донорної ДНК. Приблизно трансформовані ізоляти, які росли в даних умовах селекції, видаляли із вставлених чашок і переносили на свіже селекційне середовище на чашки 60×20 мм. Якщо стабільний ріст був очевидний через приблизно 2 тижні, то передбачали, що подія стійка до застосовуваного гербіциду (селективний агент) і аліквоту клітин потім збирали для аналізу генотипування (аналіз описаний нижче). Приклад 9: Скринінг на предмет спрямованих інтегрованих подій за допомогою ПЛРгенотипування Спрямований вплив донорних молекул (pDAB105979, pDAB105980, pDAB105955, pDAB105956) в ELP в трансгенному маїсі аналізували з використанням комбінації: 1) локусспецифічної ПЛР; 2) донора-специфічного аналізу з TaqMan, і; 3) локус-специфічного Саузернблот-аналізу для оцінки точності вставки, правильності і кількостей піків. Очікується, що позитивні спрямовані події мають a) позитивний результат out-out ПЛР або перекривання результатів in-out ПЛР і 2) присутність донора за допомогою візуалізації за допомогою Саузернаналізу. Виділення ДНК 17 UA 110472 C2 5 10 15 20 25 Тканина з трансгенного маїсу, повторно підданого спрямованому впливу, (тканина калюсу або листя рослини) ліофілізували протягом щонайменше 2 днів в 96-ямкових колекторних мікропланшетах (Qiagen, Германтаун, Меріленд). ДНК виділяли з ліофілізованої тканини з використанням робочої станції BioSprint 96 (Qiagen, Германтаун, Меріленд), слідуючи інструкціям виробника і ресуспендували в 200 мкл води. Для руйнування тканини використовували гомогенізатор тканини моделі 2-96A Kleco (Garcia Manufacturing, Вісалія, Каліфорнія). Кількісна оцінка ДНК: Отриману в результаті геномну ДНК кількісно оцінюють з використанням аналітичного набору реагентів QUANT-IT® Pico Green DNA (Molecular Probes, Invitrogen. Карлсбад, Каліфорнія). П'ять стандартів ДНК з проведеною для них кількісною оцінкою концентрацій, які коливаються від 20 нг/мкл до 1,25 нг/мкл (серійне розведення) використовували для генерування калібрувальної кривої. Невідомі зразки спочатку розводили 1:10 або 1:20, так щоб розведення було всередині лінійного інтервалу аналізу. 5 мкл розведених зразків і стандартів змішували з 100 мкл розведеного субстрату Pico Green (1:200) і інкубували протягом десяти хвилин в темряві. Флуоресценцію потім записували з використанням планшетного рідера Synergy2 (Biotek). Концентрацію геномної ДНК оцінювали на основі калібрувальної кривої, розрахованій після корекції фонової флуоресценції. Потім ДНК нормалізували відносно концентрацій 2 нг/мкл з використанням автоматизованого рідинного дозатора Biorobot-3000 (Qiagen, Германтаун, Меріленд). Нормалізовану ДНК використовували для ПЛР і аналізу оцінки кількості піків. ПЛР-аналіз Три типи локус-специфічної ПЛР (out-out, 5' out-in, 3' out-in) здійснювали для оцінки того, чи спрямовані донорні плазміди до ELP. ПЛР-реакції здійснювали для дослідження присутність інтактної копії донорної ДНК (out-out ПЛР). Додаткові реакції ПЛР фокусували на 5'-межі між мішенню і донором і 3'-межі між донором і мішенню (in-out ПЛР). Схема з положеннями праймерів, використаних для аналізу, приведена як приклад на фіг. 9. Очікувані ПЛР-продукти для кожного донора і ELP-мішені приведені в таблиці 1. Послідовності праймерів, використаних для аналізу, представлені в таблиці 2. 18 UA 110472 C2 Таблиця 1 Очікувані розміри ампліконів для локус-специфічної ПЛР для аналізу спрямованої інтеграції донора *праймери перераховані в порядку: out-out 5'in-out 3'out-in Таблиця 2 Послідовності праймерів, використаних для аналізу спрямованої інтеграції 5 10 ПЛР-ампліфікація: Полімеразну ланцюгову реакцію (ПЛР) здійснювали для оцінки і підтвердження повторного спрямованого впливу на ELP. ПЛР-реакції готували в об'ємі 25 мкл з ДНК-полімеразою Phusion (New England Biolabs, Іпсуїч, Массачусетс) або AccuPrime (Invitrogen, Карлсбад, Каліфорнія). Для Phusion-ПЛР кожна реакція містила 1X буфер Phusion GC, 0,2 мM dNTP, 0,2 мкМ прямого і зворотного праймерів, 2,5 одиниць ДНК-полімерази Phusion і 20 нг геномної ДНК. Від десяти до двадцяти нанограмів плазмідної ДНК, сконструйованої для імітації спрямованої події, проганяли як позитивний контроль. Також проганяли контроль без матриці (вода як матриця). Умови ПЛР 19 UA 110472 C2 5 10 15 20 25 30 вказані нижче: 35 циклів 98 °C 1 хв, 98 °C 10 сек, 65 °C 20 сек, 72 °C 2,5 хв з подальшим протягуванням при 72 °C 10 хв. Двадцять п'ять мікролітрів ПЛР-реакції здійснювали за допомогою AccuPrime (Invitrogen, Карлсбад, Каліфорнія), що містить 1X буфер II, 0,2 мкM прямого і зворотного праймерів, 2,5 одиниці Taq-полімерази AccuPrime і 20 нг геномної ДНК. Позитивні і негативні контролі проганяли, як описано для Phusion. Умови для AccuPrime вказаний нижче: 35 циклів 98 °C 1 хв, 98 °C 10 сек, 65 °C 20 сек, 68 °C 2,5 хв з подальшим протягуванням при 68°C 10 хв. Ампліфіковані фрагменти вирізали з гелю і очищали згідно з інструкціями виробника. Потім очищені фрагменти клонували в плазмідний вектор і трансформували в компетентні клітини Е. coli. Індивідуальні колонії селектували і підтверджували вміст ампліфікованого ПЛР-фрагмента. Реакції дволанцюжкового секвенування плазмідних клонів здійснювали для підтвердження того, що ПЛР-ампліфікована геномна послідовність містить інтегрований донор. Події, в яких ідентифікували вміст донорного фрагмента, являють собою мішень, в якій сталася спрямована гомологічна репарація ZFN-опосередкованого дволанцюжкового розриву і спрямована інтеграція донорної ДНК в специфічний ген-мішень. TaqMan-аналіз з гідролізом зонда TaqMan-аналіз проводили для визначення кількості піків донорів в передбачуваних спрямованих подіях. Визначення кількості піків трансгена за допомогою аналізу гідролізу зонда, аналогічного аналізу TAQMAN®, здійснювали за допомогою ПЛР в реальному часі з використанням системи LIGHT CYCLER480® (Roche Applied Science, Індіанаполіс, Індіана). Аналізи розробляли для PAT і AADl і для гена внутрішнього контролю GLPl і Інвертази з використанням комп'ютерної програми LIGHTCYCLER® Probe Design Software 2.0. Для ампліфікації, LIGHTCYCLER®480 Мастер-мікс для зондів (Roche Applied Science, Індіанаполіс, Індіана) отримували в 1X кінцевій концентрації в об'ємі 10 мкл багатокомпонентної реакції, що містить 0,4 мкМ кожного праймера і 0,2 мкМ кожного зонда (таблиця 3). Двостадійну реакцію ампліфікації здійснювали з протягуванням при 58 °C протягом 38 сек для PAT/GLP1 і при 60 °C протягом 40 сек для AAD-1 і з інвертазою для отримання флуоресценції. Всі зразки проганяли по три рази і середні порогові значення для циклів (Ct) використовували для аналізу кожного зразка. Аналіз даних ПЛР в реальному часі здійснювали з використанням комп'ютерної програми LIGHTCYCLER® версії 1.5 з використанням модуля визначення відносної кількості на основі методу ΔΔCt. Для цієї мети в кожен прогон включали гДНК з калібратора однієї копії і відомий 2копійний контроль. Таблиця 3 Інформація про праймери і зонди для аналізу з гідролізом зонда pat і внутрішнього контролю (HMG) 35 20 UA 110472 C2 5 10 15 20 25 30 35 Саузерн-блот-аналіз Саузерн-аналіз здійснювали для підтвердження напрямку донорної ДНК в локус ELP. Для цього аналізу геномну ДНК гідролізували за допомогою відповідного ферменту рестрикції і гібридизували із зондом у вигляді локус-специфічного радіоактивно-міченого фрагмента (не присутнього в донорі). Використовували калюс маїсу (pDAB100640 і pDAB100641-спрямовані експерименти) або рослини маїсу T0 (pDAB100610- і pDAB100611-спрямовані експерименти). Зразки тканини збирали в 2 мл-мікроцентрифугальні пробірки і ліофілізували протягом 2 днів. Мацерацію тканини здійснювали за допомогою гомогенізатора тканини Kleco і вольфрамових кульок. Після мацерації тканини геномну ДНК виділяли з використанням набору реагентів DNeasy Plant Mini kit (Qiagen, Германтаун, Меріленд) згідно з протоколом виробника, що пропонується. Геномну ДНК оцінювали кількісно за допомогою аналітичного набору реагентів Quant-IT Pico Green DNA (Molecular Probes, Invitrogen, Карлсбад, Каліфорнія). Чотири мікрограма ДНК для кожного зразка гідролізували за допомогою відповідного ферменту рестрикції протягом ночі при 37 °C. Гідролізовану ДНК очищали з використанням Quick-Precip (Edge BioSystem, Гайтерсбург, Меріленд) згідно з протоколом виробника, що пропонується. Після електрофорезу на 0,8 % SeaKem LE агарозному гелі (Lonza, Рокленд, Меріленд), ДНК переносили на нейлонові заряджені мембрани (Millipore, Бедфорд, Массачусетс) протягом ночі і зв'язували з мембраною з використанням лінкера UV Strata 1800 (Stratagene, Ла-Хойя, Каліфорнія). Блоти передгібридизували з 20 мл PerfectHyb Plus (Sigma, Сент Луїс, Міссурі) і гібридизували з відповідним радіоактивно-міченим зондом протягом ночі. Блоти відмивали і вміщували для візуалізації на екрани люмінофор на 24 години і потім аналізували з використанням сканера STORM 860 (Molecular Dynamics). Приклад 10: Конструкція проміжного трансформуючого вектора рослин, яка акцептує ELP, маркери селекції і дає можливість вирізання незалежних елементів конструкції Сконструйовані сайти зв'язування нуклеази для доменів "цинкові пальці" (eZFN) і елементи ELP можуть бути інкорпоровані індивідуально або разом в трансформуючі вектори рослин, так що отримані в результаті трансформовані рослинні клітини, тканини або рослини будуть мати наступні характеристики: 1) можливість повторно трансформуватися додатковими трансгенами точним спрямованим чином всередині ELP по вихідному сайту вставки трансгена і 2) можливість модифікації трансгенних локусів шляхом передбачуваного і ефективного видалення трансгенів, конкретно генів маркерів селекції рослин. Серії векторів конструювали для трансформації ряду видів рослин. Цього досягали шляхом використання "проміжного трансформуючого вектора рослин" (описано нижче), вектора для впливу на інкорпорування eZFN, фланкуючих всі маркери селекції і ELP, в основні вектори, відомі як вектори первинної трансформації. 21 UA 110472 C2 5 10 15 20 25 30 35 40 45 50 55 60 Конструювали проміжний трансформуючий вектор рослин, який включав кістяк бінарного трансформуючого вектора Agrobacterium з унікальним сайтом множинного клонування (MCS), розташованого між правою і лівою межами Т-ДНК. Цей проміжний вектор являє собою pDAB104132 (фіг. 10). MCS, що використовується в pDAB104132, містить унікальні сайти ферментів рестрикції для AgeI, NotI, FseI, SwaI, XbaI, AseI, PacI, XhoI, SalI, і NsiI. У даному прикладі ці сайти використовувалися для інкорпорування ключових функціональних елементів в кінцеві первинні трансформуючі вектори. Унікальний сайт NotI може використовуватися для введення фрагмента, що несе касету вектора цільового призначення GATEWAY™, яка може потім використовуватися для розташування множини трансгенів в трансформуючому векторі з використанням системи GATEWAY™. Унікальний сайт FseI може використовуватися для введення ELP, які містять сайти FseI на своїх кінцях. Як описано раніше в даних прикладах, коли ELP присутні в трансгенній рослині, вони можуть використовуватися як сайти-мішені для вставки додаткових генів з використанням eZFN. Інші сайти в MCS можуть використовуватися для вставки інших цікавлячих генів, що включають гени маркерів селекції. З використанням унікальних сайтів рестрикції, описаних вище, можуть використовуватися проміжні траснформуючі вектори рослин, такі як pDAB104132, для перенесення разом з ним в одному трансформуючому векторі рослин будь-якої або всіх комбінацій функціональних елементів, що включає касети вектора цільового призначення GATEWAY™, ELP, модулі маркерів селекціїZFN, і будь-які інші трансгени, які можуть або не можуть бути здатні до вирізання eZFN. Приклад 11: Конструкція касет з маркером селекції рослини з можливістю їх вирізання з використанням модулів нуклеаз для доменів "цинкові пальці» У даному прикладі, сайти зв'язування eZFN використовуються для можливості делеції будьякого трансгена, що включає гени маркерів селекції, з трансформованої рослини. Див. попередню патентну заявку США No. 61/297628, яка включена в цей документ посиланням. Ця здатність досягається шляхом фланкування трансгенів за допомогою одного або декількох сайтів зв'язування eZFN і потім шляхом інкорпорування цих генних модулів, що вирізаються в трансформуючі вектори рослин, які потім використовуються для генерації трансгенних рослинних клітин. Конструкція таких модулів і векторів може бути виконана фахівцем в даній галузі з використанням стандартних методів молекулярної біології. Проміжна плазміда, що несе модуль з сайтом зв'язування з ZFN, являє собою першу стадію, що застосовується на шляху до конструювання генних касет, що вирізаються. Модуль з сайтом зв'язування з ZFN являє собою сегмент ДНК, який містить щонайменше два сайти зв'язування з eZFN, фланкуючі один або декілька сайтів зв'язування з ферментами рестрикції, по яких клонували трансген. Прикладом є ZFN-модуль 2:4-MCS-2:4, представлений за допомогою SEQ ID:20 і фіг. 11. Цей модуль містить сайт множинного клонування (MCS), що складається з сайтів рестрикції AseI, HindIII, PacI, SbfI, SphI і SalI, які фланковані парами сайтів зв'язування для eZFN2 і eZFN4. Також всередині цього ZFN-модуля є дві ідентичні копії послідовності, що складається з 100 п. н. випадкової послідовності, яка фланкує сайти зв'язування з eZFN. Крім того, пари сайтів ферментів рестрикції SpeI і XhoI фланкують суцільний функціональний модуль, даючи можливість для його клонування в трансформуючий вектор рослин. pDAB104126, плазміда, що несе модуль ZFN 2:4-MCS-2:4, подана на фіг. 12. Чотири різні гени маркерів селекції індивідуально вставляли в модуль ZFN 2:4-MCS-2:4 в pDAB104126 з використанням стандартних методів молекулярного клонування. Один ген маркера селекції - це функціональний ген PAT, розроблений для надання стійкості до глуфосинату у дводольних рослин. Цей ген PAT включає кодуючу послідовність PAT, функціонально пов'язану з CsVMV-промотором і з 5'-нетрансльованою ділянкою, виділеною з вірусу мозаїки прожилків маніокі; Verdaguer et al. (1996) Plant Molecular Biology 31 (6): 1129-1139) і AtuOR 1 3' - UTR. Касету з геном PAT вставляли у вигляді фрагмента HindIII-SphI по сайтах HindIII-SphI модуля ZFN з отриманням плазміди pDAB104136 (фіг. 13). Другий ген маркера селекції - це функціональний ген PAT, розроблений для надання стійкості до глуфосинату у однодольних рослин. Цей ген PAT включав кодуючу послідовність PAT, функціонально пов'язану з OsAct1-промотором і з ZmLip 3' UTR. Цей ген PAT однодольних вставляли у вигляді фрагмента PacI-SphI по сайтам PacI-SphI модуля ZFN з отриманням плазміди pDAB104138 (фіг. 14). Третій ген маркера селекції включав ген AAD-1, розроблений для надання стійкості до галоксифопу або до 2,4-D у однодольних рослин. Цей ген AAD-1 включав кодуючу послідовність AAD-1, функціонально пов'язану з ZmUbil-промотором і з ZmLip 3' UTR. Цей ген AAD-1 однодольних рослин вставляли у вигляді фрагмента HindIII-SbfI по сайтам HindIII-SbfI модуля ZFN з отриманням плазміди pDAB104140 (фіг. 15). 22 UA 110472 C2 5 10 15 20 25 30 35 40 45 50 55 60 Четвертий ген маркера селекції - це функціональний ген PAT, розроблений для надання стійкості до глюфосинату у рослин каноли. Цей ген PAT включав кодуючу послідовність PAT, функціонально пов'язану з CsVMV-промотором і з AtuORF 1 3' UTR, і був вміщений всередині порушеного гена ізопентенілтрансферази (ipt). Ген PAT вставляли у вигляді фрагмента AseISalI по сайтах AseI-SalI модуля ZFN з отриманням плазміди pDAB104142 (фіг. 16). Таким чином, pDAB104136, pDAB104138, pDAB104140 і pDAB104142 це проміжні плазміди, які несуть різні модулі вирізання ZFN з можливістю інкорпорування різних генів маркерів селекції в трансформуючі вектори рослин. При кінцевому інкорпоруванні в геном рослини за допомогою трансформації, модулі ZFN з маркерами селекції дають можливість подальшого видалення гена маркера селекції. Цього можна досягнути шляхом отримання білків eZFN2 або eZFN4 в рослинних клітинах або за допомогою транзиторної експресії або за допомогою стабільної експресії генів, їх що кодують. Наприклад, eZFN2 або, альтернативно, eZFN4, можуть експресуватися в раніше трансформованих рослинних клітинах за допомогою транзиторної експресії eZFN з гена eZFN2 або eZFN4. Ферменти eZFN будуть розпізнавати і зв'язуватися зі специфічними сайтами зв'язування eZFN і викликати дволанцюжкові розриви, які з'являються в геномі в цих положеннях. Дволанцюжкові розриви з обох сторін гена маркера селекції всередині модуля приведе в результаті до вирізання гена маркера селекції з геномної ДНК. Потім, дволанцюжковий розрив буде схильний до репарації або з не гомологічним з'єднанням кінців, або гомологічної одноланцюжкової репарації між повторюваними 100 п. н.-послідовностями, які включені в модуль ZFN. Цей процес приведе в результаті до отримання постійної делеції гена маркера селекції в його вихідній геномної локалізації. Приклад 12: Збирання первинних трансформуючих векторів рослин Фахівець в даній галузі здатний зібрати елементи, описані вище, покроково в комбінації з отриманням первинних трансформуючих векторів рослин для застосування в різних культурах рослин. ELPl вставляли в сайт FseI pDAB104132 з отриманням pDAB104133 (фіг. 17). Потім, касету GATEWAY™ вектора цільового призначення вставляли по сайту NotI pDAB104133 з отриманням pDAB104134 (фіг. 18) і pDAB104135 (фіг. 19), які відрізняються тільки по своїй орієнтації касети Gateway відносно послідовності ELP. Потім чотири модулі ZFN з маркерами селекції, описаними вище, клонували в pDAB104135 з отриманням чотирьох нових Трансформуючих Векторів Рослин (PTV). Це збирання може здійснити будь-який фахівець в даній галузі з використанням стандартних методів клонування ДНК, як описано нижче. Модуль PAT ZFN для дводольних рослин вирізали з pDAB104136 з використанням SpeI і клонували по сайту XbaI pDAB104135 з отриманням PTV pDAB104137 (фіг. 20). pDAB104137 являє собою бінарний трансформуючий вектор рослин, що несе ELPl і модуль ZFN з вирізуваним маркером селекції дводольних рослин PAT, а також являє собою цільовий вектор GATEWAY™, який дає можливість для GATEWAY™-клонування додаткових трансгенів. Модуль PAT ZFN для однодольних рослин вирізали з pDAB104138 з використанням SpeI і клонували по сайту XbaI pDAB104135 з отриманням PTV pDAB104139 (фіг. 21). pDAB104139 являє собою бінарний трансформуючий вектор рослин, що несе ELPl і модуль ZFN з вирізуваним маркером селекції однодольних рослин PAT, а також являє собою цільовий вектор GATEWAY™. Модуль AADl ZFN для однодольних рослин вирізали з pDAB104140 з використанням SpeI і клонували по сайту XbaI pDAB104135 з отриманням PTV pDAB104141 (фіг. 22). pDAB104141 являє собою бінарний трансформуючий вектор рослин, що несе ELPl і модуль ZFN з вирізуваним маркером селекції однодольних рослин AAD1, а також являє собою цільовий вектор GATEWAY™. Модуль PAT каноли ZFN для однодольних рослин вирізали з pDAB104142 з використанням XhoI і клонували по сайту XhoI pDAB104135 з отриманням PTV pDAB104143 (фіг. 23). pDAB104143 являє собою бінарний трансформуючий вектор рослин, що несе ELPl і модуль ZFN з вирізуваним маркером селекції PAT каноли, а також являє собою цільовий вектор GATEWAY™. Ці конструкції можуть використовуватися для трансформації культур рослин з використанням методів трансформації рослин. Отримані в результаті трансгенні події можуть бути потім піддані спрямованому впливу додаткових нових трансгенів, де новий трансген спрямовується за допомогою гомологічної рекомбінації до ELP. Крім того, експресуюча касета з маркером селекції може бути видалена або вирізана. Отримана в результаті ELP, яку отримують після вирізання касети з маркером селекції, може бути потім піддана спрямованому впливу нового трансгена за допомогою гомологічної рекомбінації з використанням методів, описаних вище. 23 UA 110472 C2 24 UA 110472 C2 25 UA 110472 C2 26 UA 110472 C2 27 UA 110472 C2 28
ДивитисяДодаткова інформація
Назва патенту англійськоюEngineered landing pads for gene targeting in plants
Автори англійськоюAinley, William, Michael, Blue, Ryan, C., Murray, Michael, G., Corbin, David, Richard, Miles, Rebecca, Ruth, Webb, Steven, R.
Автори російськоюЭйнли Уилльям Майкл, Блю Райан С., Мюррей Майкл Г., Корбин Девид Ричард, Майлз Ребекка Рут, Уэбб Стивен Р.
МПК / Мітки
МПК: C12N 15/29, A01H 5/00, C12N 15/82
Мітки: рослинна, тканина, отримання, трансгенна, спрямованого, рослини, клітині, насіння, отримане, рослинах, спосіб, впливу, рослина, ген, трансгенної
Код посилання
<a href="https://ua.patents.su/69-110472-sposib-otrimannya-klitini-transgenno-roslini-dlya-spryamovanogo-vplivu-na-gen-v-roslinakh-transgenna-roslina-abo-roslinna-tkanina-nasinnya-otrimane-z-transgenno-roslini.html" target="_blank" rel="follow" title="База патентів України">Спосіб отримання клітини трансгенної рослини для спрямованого впливу на ген в рослинах, трансгенна рослина або рослинна тканина, насіння, отримане з трансгенної рослини</a>
Спосіб одержання трансгенної рослини, послідовність днк, яка кодує поліфруктан (леван) сахаразу, рослинна клітина

Номер патенту: 40598
Опубліковано: 15.08.2001
Автори: Рьобер Мануела, Гайдер Клаус, Гайєр Гебхардт, Вілльмітцер Лотар
МПК: C12N 9/10, C12N 15/82
Мітки: яка, клітина, одержання, леван, рослини, послідовність, спосіб, сахаразу, днк, поліфруктан, трансгенної, рослинна, кодує
Формула / Реферат:
1. Способ получения трансгенного растения, характеризующегося модифицированным образованием полифруктана в хлоропластах, амилопластах, митохондриях, цитозоле и/или вакуолях, который включает следующие стадии:а) введение выделенной молекулы ДНК в растительную клетку таким образом, что она способна встраиваться в геном указанной клетки, причем указанная молекула ДНК включает(I) промотор, способный регулировать транскрипцию гена...
Плазміда pslc1-1/prd400, штам gv3101 agrobacterium tumefaciens, спосіб одержання трансгенної рослини з масловмісним насінням, спосіб виділення їстівної або неїстівної олії з масловмісного насіння рослини

Номер патенту: 49833
Опубліковано: 15.10.2002
Автори: Келлер Вілфред А., Тейлор Девід С., Зоу Джітао, Катавік Весна, Маккензі Самюел Л.
МПК: C11B 1/00, C12N 15/09, A01H 5/00, C12N 1/21, C12N 15/82, C12N 9/10, C11B 1/10, C12N 15/00, C12N 5/10
Мітки: рослини, штам, олії, tumefaciens, gv3101, спосіб, трансгенної, насіння, одержання, плазміда, agrobacterium, масловмісного, їстівної, насінням, масловмісним, виділення, неїстівної
Формула / Реферат:
1. Плазміда pSLC1-1/pRD400 (ATCC 97545).2. Штам GV3101 Agrobacterium tumefaciens, який відрізняється тим, що вищезгаданий штам модифікований введенням дріжджового гена SLC1-1, що має нуклеотидну послідовність SEQ ID NO: 1.3. Спосіб одержання трансгенної рослини з масловмісним насінням, який відрізняється тим, що в геном вищезгаданої рослини вводять дріжджовий ген SLC1-1, що здатен до експресії і який має нуклеотидну...
Спосіб індукування стійкості до вірусу, що містить послідовність потрійного блока генів 3, за умови, що це не х вірус картоплі, у клітині рослини або рослині, трансгенна рослина, стійка до вірусу

Номер патенту: 70915
Опубліковано: 15.11.2004
Автори: Річардс Кен, Жонар Жерар, Блейкастен-Гроссханс Клодін, Вейєн Гю, Бузубаа Салах, Гюйіє Убер, Лефебвр Марк
МПК: C07K 14/08, C12N 15/82
Мітки: спосіб, стійкості, генів, умови, рослина, потрійного, індукування, трансгенна, вірусу, послідовність, містить, клітині, стійка, блока, рослини, картоплі, вірус
Формула / Реферат:
1. Спосіб індукування стійкості до вірусу, що містить послідовність потрійного блока генів 3, за умови, що це не X вірус картоплі, у клітині рослини або рослині, який включає стадії:- одержання конструкції нуклеїнової кислоти, яка містить послідовність нуклеїнової кислоти, що відповідає щонайменше на 70% послідовності нуклеїнової кислоти потрійного блока генів 3 зазначеного вірусу або його відповідної кДНК, функціонально злиту з однією...
Спосіб отримання резистентної до вірусу трансгенної рослини

Номер патенту: 29438
Опубліковано: 15.11.2000
Автори: Теері Теему, Саарма Март, Труве Ерккі, Келве Мерікке
МПК: A01H 5/00, C12N 15/54, C12N 5/10, C12N 9/12, C12N 15/09, C12N 15/82
Мітки: спосіб, резистентної, трансгенної, отримання, рослини, вірусу
Текст:
...29438 Пробы листьев анализируют методом EUSA с использованием иммунодиагшстачесшго набора для PVX (Boehringer Mannheim). Концентрацию вирусов определяют по оптической плотности цветной реакции при 405 нм. Контр. = контрольный клон; R101, R102, R104, R106, R107, и R2 =трансгенные клоны. Следующие примеры иллюстрируют изобретение. Дополнительные пояснения в отношении методов молекулярной биологии, применяемых в данном изобретении,...
Спосіб збільшення розміру і кількості насіння внаслідок трансгенної надмірної експресії гена, асоційованого з ростом і/або розвитком, під час раннього ембріонального розвитку рослини

Номер патенту: 97632
Опубліковано: 12.03.2012
Автори: Нгуйен Тху, Дерочер Джей
МПК: A01H 5/00, C12N 15/82
Мітки: асоційованого, збільшення, надмірної, експресії, розвитку, рослини, розвитком, раннього, внаслідок, гена, насіння, трансгенної, ростом, розміру, спосіб, ембріонального, кількості
Формула / Реферат:
1. Генетична конструкція, яка містить послідовність нуклеїнової кислоти, що кодує REV-ген, який оперативно зв'язаний з однією або більше контролюючими послідовностями, де контролюючі послідовності містять промотор, асоційований з геном листової сім'ядолі та/або геном олеату 12-гідроксилази:десатурази.2. Генетична конструкція за п. 1, в якій геном листової сім'ядолі є ген LEC2 із виду Arabidopsis thaliana і геном олеату...
Попередній патент: 5-алкінілпіримідини
Наступний патент: Спосіб лікування раку за допомогою пухлиноасоційованого антигену, отриманого із цикліну d1
Випадковий патент: Прилад для визначення спірографічних характеристик легенів людини