Застосування глибинної морської води із сірководневої зони чорного моря як середовища культивування морських водоростей
Номер патенту: 78444
Опубліковано: 15.03.2007
Автори: Полікарпов Геннадій Григорович, Лазоренко Галина Євдокимівна, Терещенко Наталія Миколаївна




Формула / Реферат
Застосування глибинної води Чорного моря, яка містить сірководень та окислену сірководневу воду, як середовища культивування морських водоростей.
Текст
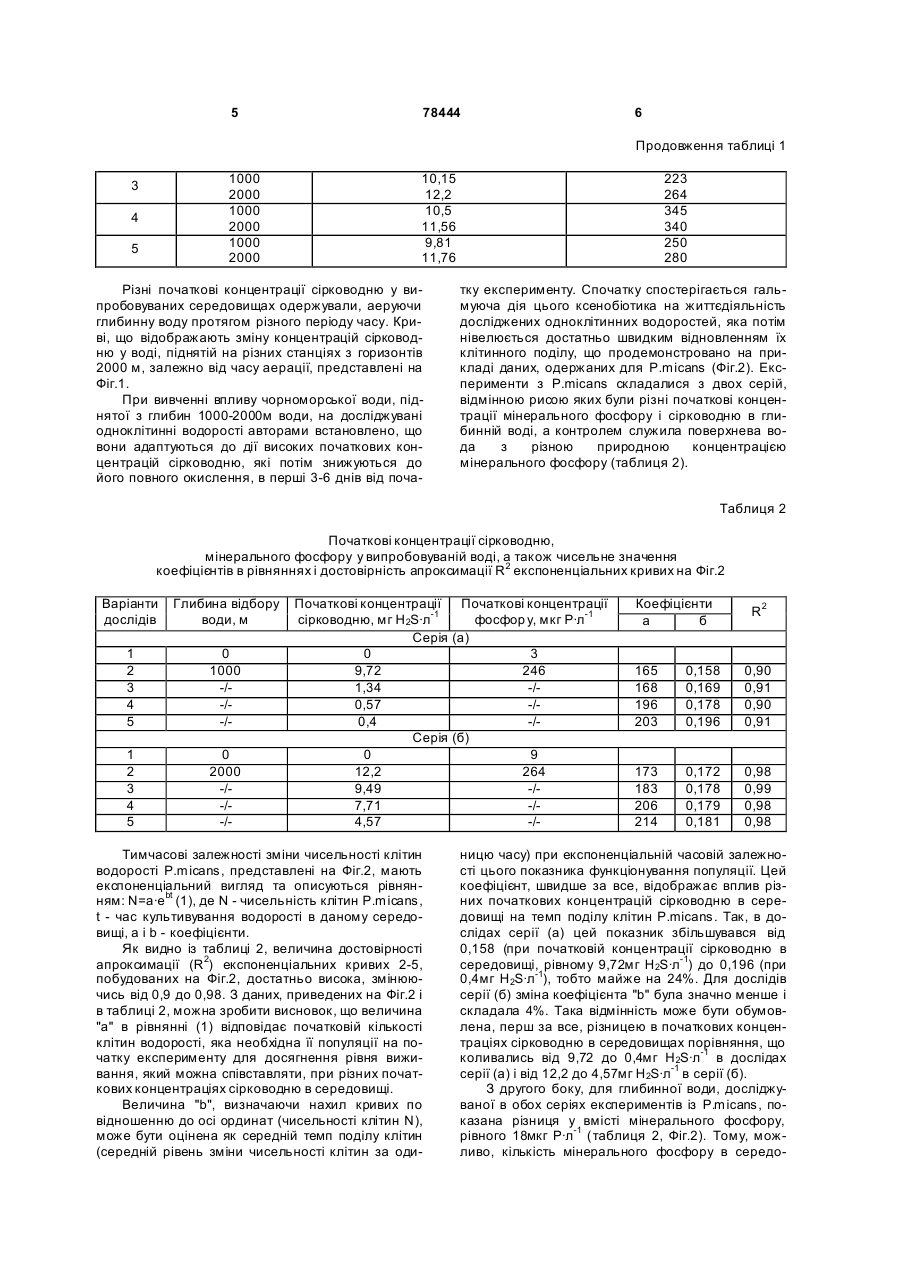
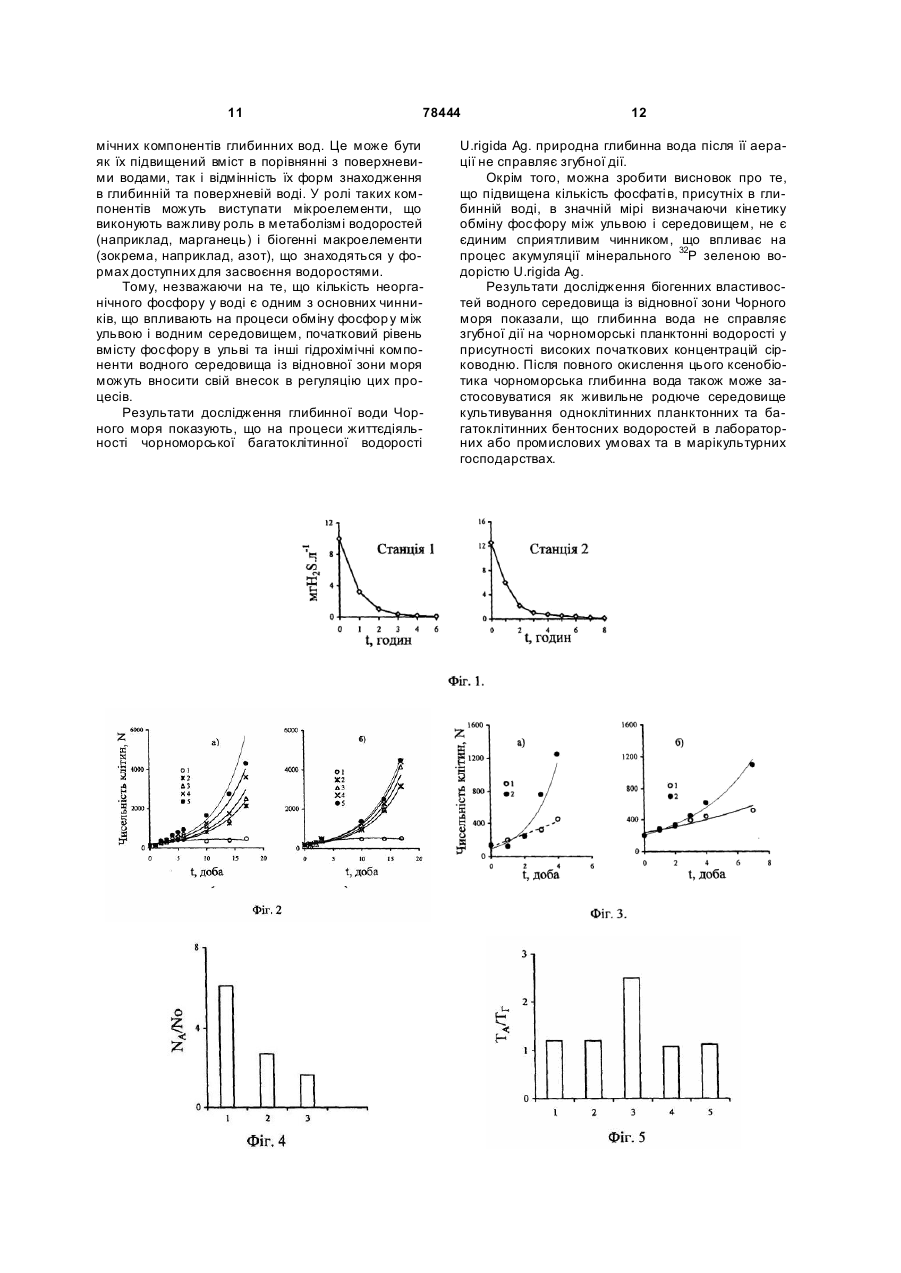
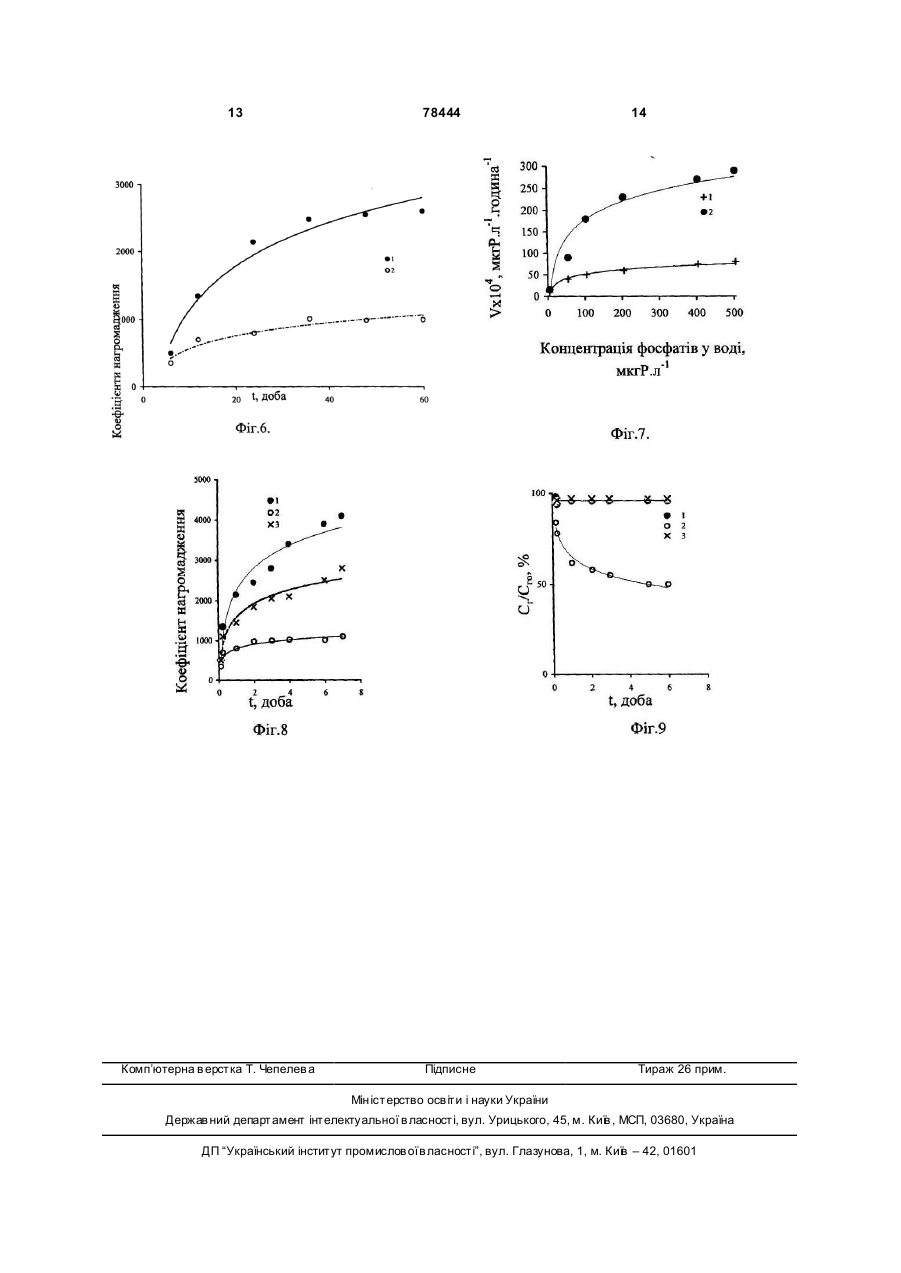
Застосування глибинної води Чорного моря, яка містить сірководень та окислену сірководневу воду, як середовища культивування морських водоростей. (19) (21) a200510066 (22) 25.10.2005 (24) 15.03.2007 (46) 15.03.2007, Бюл. №3, 2007р. (72) Полікарпов Геннадій Григорович, Лазоренко Галина Євдокимівна, Терещенко Наталія Миколаївна (73) ІНСТИТУТ БІОЛОГІЇ ПІВДЕННИХ МОРІВ ІМ. О.О.КОВАЛЕВСЬКОГО НАН УКРАЇНИ 3 78444 дінофлагеллят в глибинній воді (NA) з початковою концентрацією сірководню, рівною 12,2мгH2S·л-1, і поверхневій воді (No) через 17 діб від початку експериментів; Фіг.5 - Відношення темпів поділу чорноморських одноклітинних водоростей в аерованній воді, відібраній з глибини 2000м (ТА), і середовищі Гольдберга (ТГ); Фіг.6 - Зміна в часі Кн 32 Р в ульві в глибинній (1) і поверхневій (2) воді; Фіг.7. - Залежність швидкості поглинання (V) 32Р ульвою від концентрації фосфатів у воді, після попередньої адаптації ульви протягом 72 годин в глибинній аерованній (1) і в поверхневій воді (2); Фіг.8 - Залежність від часу Кн 32Р для водорості, культивованої в глибинній воді (1), поверхневій воді (2) і поверхневій воді з добавкою мінерального фосфор у (3); Фіг.9 - Виведення 32Р з ульви в середовища порівняння: глибинну воду (1), поверхневу воду (2) і поверхневу воду з добавкою мінерального фосфору (3). Вибір об'єктів пов'язаний з їх роллю в екосистемі Чорного моря. Так одноклітинні водорості були вибрані як первинні продуценти органічної речовини в морі. Вони є ланкою харчового ланцюга для інших об'єктів марікультури і від їх реакції на якість середовища залежить функціонування вищи х рівнів трофічних ланцюгів в екосистемі, а також вони самі можуть використовуватися як цінні продукти марікультури. Вибір макроводорості ульви (Ulva rigida Ag.) для дослідження глибинної води обумовлений належністю цього макрофіта до мезосапробних гідробіонтів, широко поширених в прибережних районах моря, рекомендованих для культивування. При дослідженні реакції чорноморських одноклітинних водоростей досліди проводили з альгологічно чистими культурами дінофлагеллят (Prorocentrum micans Ehr., Peridinium trochoideum (Stein) Lammermann, Gymnodinium kovalevskii Pitz., Gymnodinium sp.) і діатомових (Ditylum brightwellii (West), Coscinodiscus granii, Sceletonema costatum). Головним показником їх стану були темпи поділу і чисельність клітин. Середовищем випробування для них була вода, відібрана в Чорному морі із глибин 1000-2000м. Середовищами порівняння були та ж вода, активно аерована протягом 6-8 годин до повного окислення в ній сірководню, поверхнева вода, взята на тій же станції, що і глибинна, а також середовище Гольдберга, традиційно використовуване в лабораторних умовах для культивування одноклітинних водоростей. Експерименти з одноклітинними водоростями проводили в трьох повторностях. В кожну склянку вносили рівну початкову кількість клітин досліджу 4 ваних водоростей. Через 1-17 діб від початку експериментів з кожної склянки відбирали по 0,1мл суспензії клітин, в якій визначали їх кількість. Для оцінки можливого впливу глибинної окисленої води на процеси життєдіяльності макрофітів досліджували кінетику обміну фосфатів між ульвою і водним середовищем за допомогою методу мічених атомів з використанням 32Р. Визначуваними параметрами кінетики фосфорного обміну ульви з середовищем були: коефіцієнт накопичення (Кн) 32Р, швидкість поглинання мінерального фосфору ульвою з середовища (V, мкгР·л-1час-1), частка 32Р, виведеного із водорості у водне середовище. У всіх дослідах з ульвою використовували воду, підняту з глибини 1000м, яку перед початком експериментів активно аерували протягом 6 годин. Контроль складався із двох варіантів, середовищем порівняння в яких була поверхнева вода з природним вмістом фосфатів, (варіант 1), і поверхнева вода з добавкою фосфатів в такій кількості, що в сумі концентрація фосфатів в середовищі була рівною такій в глибинній воді (варіант 2). Вибір подвійного контролю в дослідах був обумовлений тим, що параметри кінетики обміну 32 Р ульвою залежать від концентрації розчиненого мінерального фосфору у водному середовищі. Тому наявність другого контролю - поверхневої води з добавкою фосфатів - дозволяла виключити цю залежність і виявити можливий вплив інших гідрохімічних компонентів окисленої глибинної води на значення параметрів кінетики фосфорного обміну між водоростю і водним середовищем. Експерименти з ульвою проводили на фрагментах, вирізаних із центральної частини її таллома, діаметр яких складав 15мм, а початкова біомаса - 10-12мг з розрахунку на сиру масу. В кожну з скляних посудин місткістю 1-2л поміщали 10 таких фрагментів. Температура води в акваріумахскладала 14-15°С. Похибка радіометричних вимірювань не .перевищувала 5%. В кожній часовій точці відбирали 8-10 фрагментів і 3-5 проб води. Для кожної вибірки в одній часовій точці розраховували середнє значення і визначали довірчий інтервал середнього значення з 95% надійністю. У всіх вибірках ширина довірчого інтервалу не перевищувала 10% від значення середнього. Всі експерименти виконані в трьох повторностях. Для визначення концентрацій сірководню та фосфа тів в поверхневій і глибинній воді використовували стандартні гідрохімічні методики. Результати експериментів. Таблиця 1 Початкові концентрації сірководню і фосфатів в чорноморській воді, відібраній відновної зони Чорного моря для експериментів з водоростями № станції 1 2 Горизонт, м 2000 1000 1900 Концентрація сірководню, мг Н2S·л-1 9,98 9,72 11,95 Концентрація фосфатів, мкг Р·л-1 300 246 336 5 78444 6 Продовження таблиці 1 1000 2000 1000 2000 1000 2000 3 4 5 10,15 12,2 10,5 11,56 9,81 11,76 Різні початкові концентрації сірководню у випробовуваних середовищах одержували, аеруючи глибинну воду протягом різного періоду часу. Криві, що відображають зміну концентрацій сірководню у воді, піднятій на різних станціях з горизонтів 2000 м, залежно від часу аерації, представлені на Фіг.1. При вивченні впливу чорноморської води, піднятої з глибин 1000-2000м води, на досліджувані одноклітинні водорості авторами встановлено, що вони адаптуються до дії високих початкових концентрацій сірководню, які потім знижуються до його повного окислення, в перші 3-6 днів від поча 223 264 345 340 250 280 тку експерименту. Спочатку спостерігається гальмуюча дія цього ксенобіотика на життєдіяльність досліджених одноклітинних водоростей, яка потім нівелюється достатньо швидким відновленням їх клітинного поділу, що продемонстровано на прикладі даних, одержаних для Р.micans (Фіг.2). Експерименти з P.micans складалися з двох серій, відмінною рисою яких були різні початкові концентрації мінерального фосфору і сірководню в глибинній воді, а контролем служила поверхнева вода з різною природною концентрацією мінерального фосфору (таблиця 2). Таблиця 2 Початкові концентрації сірководню, мінерального фосфору у випробовуваній воді, а також чисельне значення коефіцієнтів в рівняннях і достовірність апроксимації R2 експоненціальних кривих на Фіг.2 Варіанти дослідів Глибина відбору води, м 1 2 3 4 5 0 1000 -/-/-/ 1 2 3 4 5 0 2000 -/-/-/ Початкові концентрації Початкові концентрації сірководню, мг Н2S·л-1 фосфор у, мкг Р·л-1 Серія (а) 0 3 9,72 246 1,34 -/0,57 -/0,4 -/Серія (б) 0 9 12,2 264 9,49 -/7,71 -/4,57 -/ Тимчасові залежності зміни чисельності клітин водорості P.micans, представлені на Фіг.2, мають експоненціальний вигляд та описуються рівнянням: N=a·e bt (1), де N - чисельність клітин P.micans, t - час культивування водорості в даному середовищі, а і b - коефіцієнти. Як видно із таблиці 2, величина достовірності апроксимації (R2) експоненціальних кривих 2-5, побудованих на Фіг.2, достатньо висока, змінюючись від 0,9 до 0,98. З даних, приведених на Фіг.2 і в таблиці 2, можна зробити висновок, що величина "а" в рівнянні (1) відповідає початковій кількості клітин водорості, яка необхідна її популяції на початку експерименту для досягнення рівня виживання, який можна співставляти, при різних початкових концентраціях сірководню в середовищі. Величина "b", визначаючи нахил кривих по відношенню до осі ординат (чисельності клітин N), може бути оцінена як середній темп поділу клітин (середній рівень зміни чисельності клітин за оди Коефіцієнти a б R2 165 168 196 203 0,158 0,169 0,178 0,196 0,90 0,91 0,90 0,91 173 183 206 214 0,172 0,178 0,179 0,181 0,98 0,99 0,98 0,98 ницю часу) при експоненціальній часовій залежності цього показника функціонування популяції. Цей коефіцієнт, швидше за все, відображає вплив різних початкових концентрацій сірководню в середовищі на темп поділу клітин P.micans. Так, в дослідах серії (а) цей показник збільшувався від 0,158 (при початковій концентрації сірководню в середовищі, рівному 9,72мг Н 2S·л-1) до 0,196 (при 0,4мг Н 2S·л-1), тобто майже на 24%. Для дослідів серії (б) зміна коефіцієнта "b" була значно менше і складала 4%. Така відмінність може бути обумовлена, перш за все, різницею в початкових концентраціях сірководню в середовищах порівняння, що коливались від 9,72 до 0,4мг Н2S·л-1 в дослідах серії (а) і від 12,2 до 4,57мг Н2S·л-1 в серії (б). З другого боку, для глибинної води, досліджуваної в обох серіях експериментів із P.micans, показана різниця у вмісті мінерального фосфору, рівного 18мкг Р·л-1 (таблиця 2, Фіг.2). Тому, можливо, кількість мінерального фосфору в середо 7 78444 вищі є другим чинником, що впливає на чисельне значення коефіцієнта "b". Додатковим, безпосередньо неврахованим, в наших дослідах чинником середовища, вплив якого на популяцію досліджуваної планктонної водорості також відображається на коефіцієнті 'b", може бути вміст у випробовуваній воді інших мікро- і макроелементів або органічних сполук, необхідних для оптимального розвитку популяції P.micans. Криві 1 (Фіг.2 а), б)) побудовані за результатами дослідів з поверхневою водою, є поліноміальними лініями, що відповідають залежностям типу: N=-сх2+d x+k, де N - чисельність клітин в популяції, с і d - коефіцієнти, що визначають нахил кривих по відношенню до осі ординат, а величина k, можливо, відповідає тій початковій кількості клітин, яку необхідно враховувати в конкретних порівняльних експериментах. Результати вивчення відгуку інших видів чорноморських планктонних водоростей, зокрема, G.kovalevskii і D.brightwellii, на склад води із відновної зони моря представлені на Фіг.3, з якого слідує, що рівняння, що характеризують зміни чисельності клітин в популяціях водоростей в часі, мають експоненціальний вигляд. У досліді з глибинною водою при величині достовірності апроксимації кривої R2=0,9, рівняння, що описує зміну чисельності клітин G.kovalevskii з часом, мало вигляд: N=94е0,624t. Чисельність клітин цієї ж водорості в поверхневій воді змінювалася з часом також по експоненціальному закону і виражена рівнянням N=140e0,29t при величині R2=0,98. Аналогічно для D.brightwellii рівняння мали вигляд: N=207е0,247t при R2=0,98 (для варіанту досвіду з глибинною водою) та N=239е0,129tb при R2=0,88 (для поверхневої води). Як видно, коефіцієнти "а" і "b" в рівняннях, що описують закономірності зміни чисельності популяцій G.kovalevskii і D.brightwellii залежно від часу експериментів, несуть таке ж змістовне навантаження, як і у рівняннях для P.micans. Із представлених на Фіг.2, 3 даних видно, що період адаптації для досліджених чорноморських одноклітинних водоростей залежить від їх видової належності, початкової концентрації сірководню в середовищі та часу, протягом якого відбувається природне окислення ксенобіотика в дослідних акваріумах. Відношення темпів поділу чорноморських дінофлагеллят в глибинній і поверхневій воді (Фіг.4) свідчить про те, що, не дивлячись на високі початкові концентрації сірководню, глибинна вода не викликає токсичної дії на досліджені види водоростей. Навпаки, при окисленні сірководню в середовищі з відновної зони Чорного моря, чисельність і темпи ділення клітин в ній вищі, ніж в поверхневій воді моря (Фіг.2-4). Відомо, що поверхнева вода Чорного моря не містить біогенні елементи в кількостях, достатніх для тривалого і повноцінного культивування одноклітинних водоростей. В першу чергу це відноситься до фосфору, концентрації якого в пробах води, узятих в різних районах Чорного моря для експериментів з одноклітинними водоростями, були в межах 3-9мкг Р·л-1. В той же час у воді з 8 глибин 1000-2000м, вміст фосфатів був вищим в 30-130 разів. Окрім фосфору, на чисельність одноклітинних водоростей в середовищі культивування впливають і інші елементи. Тому в наступній серії експериментів якість окисленої глибинної води з відновної зони Чорного моря порівнювали з традиційно використовуваним в лабораторному культивуванні мікроводоростей середовищем Гольдберга, основою приготування якого служить поверхнева вода відкритої частини моря. Вміст основних елементів в живильному середовищі Гольдберга вищий, ніж в глибинній воді. Виняток склав лише марганець (таблиця 3). Таблиця 3 Концентрації найважливіших хімічних елементів (мг-атом·л-1) в глибинній воді Чорного моря і середовищі Гольдберга. Хімічний еле- Вода з глибини Середовище Гомент 1000м льдберга N 82 2014 Co 0,1 102 Р 7 20 Fe 0,5 1 Мn 5,3 0,8 Наші експерименти показали, що у порівнянні із середовищем Гольдберга, темпи поділу і чисельність популяцій чорноморських мікроводоростей в глибинній воді після повного окислення в ній сірководню достовірно вищі (Фіг.5). Це обумовлено особливостями сольового, зокрема мікроелементного складу, характерного для води із відновної зони Чорного моря, та її сприятлива дія на мікрофіти викликана не тільки більш широким спектром різноманітності хімічних елементів, але і природною збалансованістю складу хімічних елементів та форм їх фізико-хімічного стану, співвідношенням основних біогенних макроелементів, а також наявністю інших біологічно активних речовин, доступніших для споживання їх водоростями, в глибинній воді, що позитивно впливає на темп поділу клітин. Отже, вода із відновної зони Чорного моря навіть при найвищих природних початкових концентраціях в ній сірководню не проявляє згубної дії на популяції чорноморських одноклітинних водоростей. Більше того, після повного окислення сірководню, біологічна якість води із відновної зони Чорного моря перевищує таку не тільки в поверхневій воді, але навіть в культуральному живильному середовищі Гольдберга. При дослідженні кінетики накопичення 32Р багатоклітинною зеленою чорноморською водорістю U. rigida Ag. у глибинній аерованій воді встановлено, що, у порівнянні із поверхневою водою (із концентрацією фосфатів, рівною 3мкг Р·л-1), акумуляція 32Р фрагментами талому ульви відбувається інтенсивніше в глибинній воді, вміст фосфатів в якій складав 230мкг Р·л-1 (Фіг.6). Зміна Кн 32Р ульвою в часі протягом перших 2,5 діб експерименту описується рівнянням вигляду Кн=a lnt-b, 9 78444 де а і b - коефіцієнти. Зокрема, для варіанту з глибинною водою при величині достовірності апроксимації, рівній 0,96 (крива 1) рівняння має вигляд: Кн=939,7lnt-1039,5, а для варіанту в поверхневій воді, при R2=0,93, крива 2 описується рівнянням: Кн=281,5lnt-83,2. Як видно, чисельні значення коефіцієнтів "а" і "b" в рівняннях для кривих 1 та 2 (Фіг.6), значно розрізнялися. Так, для коефіцієнта "а" ця різниця складала 3,3 раза, а для коефіцієнта "b" - 12,5 раза. До кінця експерименту, результати якого приведені на Фіг.6, коефіцієнти нагромадження міченого фосфору водорістю в «глибинній воді майже в 6 разів вищі, ніж у поверхневій воді, що може бути обумовлене, як однією із причин, різним вмістом фосфатів в середовища х порівняння. Для уточнення наявності можливих чинників, регулюючи х фосфорний обмін ульви, нами були проведені експерименти по вивченню впливу різних добавок мінерального фосфору у поверхневій воді на швидкість поглинання 32P ульвою (Фіг.7). Перед початком дослідів по акумулюванню 32Р водорості витримували в середовищах порівняння (глибинна аерована та поверхнева вода) 72 години з метою можливої оцінки впливу преадаптації водоростей до різної кількості фосфаті в в середовищі і формування різного рівня внутріклітинного вмісту фосфору в дослідних зразках ульви. З приведених на Фіг.7 результатів видно, що при добавках мінерального фосфору до поверхневої води приблизно до 10-20мкг Р·л-1 швидкість поглинання 32P ульвою, практично, не залежить від умов попередньої адаптації водорості. При подальшому збільшенні концентрації мінерального фосфор у, що вноситься в чорноморську поверхневу воду у вигляді добавок фосфатів, швидкість поглинання 32Р ульвою майже в 3,6 рази нижче для водоростей, що проходили преадаптацію в глибинній аерованій воді (крива 1), яка містить 230мкг Р·л-1, ніж для водоростей, що культивувалися перед початком експерименту в поверхневій воді (крива 2). Для варіанту досліду, в якому в період адаптації водорості культивували в глибинній аерованій воді (Фіг.7, крива 1) при величині достовірності апроксимації, рівній 0,94, залежність швидкості поглинання (V) ульвою 32Р від концентрації мінерального фосфору у воді (С) має логарифмічний вигляд: V=61,6lnС-106,4. При величині достовірності апроксимації, рівній 0,98, залежність швидкості поглинання міченого фосфор у водоростями від концентрації мінерального фосфору у водному середовищі, для варіанту адаптації в поверхневій воді (Фіг.7, крива 2), може бути описана рівнянням аналогічного вигляду: V=15lnC-17. Як видно із Фіг.7, на процес акумуляції 32P ульвою впливає кількість мінерального фосфору у водному середовищі та внутріклітинна концентрація фосфору, що сформувалася у водоростей. 10 Кінетичні залежності Кн 32P ульвою, отримані авторами в експериментах із різними концентраціями мінерального фосфору у воді, були передумовою для проведення порівняльного аналізу кривих, що описують нагромадження міченого фосфор у в умовах, коли середовищами порівняння були глибинна аерована вода із концентрацією фосфа тів, рівною 230мкг Р·л-1, поверхнева вода, в якій фосфати були визначені на рівні 3мкг Р·л-1 і поверхнева вода, в яку мінеральний фосфор внесли в кількості, рівній 227мкг Р·л-1, тобто в сумі в даному середовищі концентрація фосфатів відповідала такій в глибинній воді (Фіг.8). Через сім діб після початку експерименту Кн міченого фосфору склали такі величини: у глибинній аерованій воді 4000, у поверхневій - 1080, у поверхневій із добавкою фосфатів - 2900 одиниць. Отже, результати, приведені на Фіг.8, свідчать, що кількість мінерального фосфору у середовищі може виступати в ролі чинника, регулюючого процес акумуляції міченого фосфору, але не є єдиним параметром глибинної води, визначаючим інтенсивність цього процесу. Вплив вмісту фосфатів у воді на процеси фосфорного обміну ульви підтверджують і досліди по вивченню виведення 32P водоростями. Аналізуючи криві виведення 32Р ульвою (Фіг.9), можна зробити висновок про те, що мічений фосфор, накопичений ульвою у середовищах із концентрацією мінерального фосфору у воді, рівною 230мкг Р·л-1 (криві 1, 3), відображає переважне надходження міченого фосфор у до необмінних фондів фосфору, що використовуються ульвою на зростання та продукцію саме в умовах ви щи х концентрацій фосфатів у середовищі, як в поверхневій, із добавкою фосфатів, так і в глибинній аерованій воді. При цьому кінетика виведення фосфатів в природній поверхневій (без добавок фосфору - крива 2) і глибинній чорноморській воді (крива 3) достовірно відрізняється. Як видно із представлених графіків (Фіг.9), у поверхневій воді частка 32P, що виводиться на третю добу, досягає 50% і надалі збільшується, в той час, як у глибинній воді за цей же час частка виведеного фосфору складає лише 6% від початкової кількості акумульованого водоростями міченого фосфору та з часом залишається на цьому ж рівні. Виведення 32Р в глибинній і поверхневій воді із добавкою фосфатів, в яких концентрації фосфору були вирівняні, не розрізняється. Таким чином, для процесів виведення міченого фосфор у ульвою концентрація фосфатів у водному середовищі виконує провідну роль. Величини ж Кн 32P достовірно розрізняються в середовищах порівняння (Фіг.8), не зважаючи на рівні умови за вмістом фосфатів у воді (криві 1, 3). Так, у поверхневій воді із добавкою фосфатів Кн 32 P водоростями вище (крива 3), ніж в поверхневій воді із природним вмістом фосфатів (крива 2), найвищі Кн 32P макрофітоми спостерігали в глибинній воді. Ймовірно, величина Кн 32P ульвою обумовлена не тільки підвищеним вмістом фосфору у воді, але і інтенсивнішим надходженням фосфор у до необмінних фондів водорості, що, в свою чергу, може бути викликано сприятливим впливом на фосфорний обмін ульви інших гідрохі 11 78444 мічних компонентів глибинних вод. Це може бути як їх підвищений вміст в порівнянні з поверхневими водами, так і відмінність їх форм знаходження в глибинній та поверхневій воді. У ролі таких компонентів можуть виступати мікроелементи, що виконують важливу роль в метаболізмі водоростей (наприклад, марганець) і біогенні макроелементи (зокрема, наприклад, азот), що знаходяться у формах доступних для засвоєння водоростями. Тому, незважаючи на те, що кількість неорганічного фосфору у воді є одним з основних чинників, що впливають на процеси обміну фосфор у між ульвою і водним середовищем, початковий рівень вмісту фосфору в ульві та інші гідрохімічні компоненти водного середовища із відновної зони моря можуть вносити свій внесок в регуляцію цих процесів. Результати дослідження глибинної води Чорного моря показують, що на процеси життєдіяльності чорноморської багатоклітинної водорості 12 U.rigida Ag. природна глибинна вода після її аерації не справляє згубної дії. Окрім того, можна зробити висновок про те, що підвищена кількість фосфаті в, присутніх в глибинній воді, в значній мірі визначаючи кінетику обміну фосфору між ульвою і середовищем, не є єдиним сприятливим чинником, що впливає на процес акумуляції мінерального 32P зеленою водорістю U.rigida Ag. Результати дослідження біогенних властивостей водного середовища із відновної зони Чорного моря показали, що глибинна вода не справляє згубної дії на чорноморські планктонні водорості у присутності високих початкових концентрацій сірководню. Після повного окислення цього ксенобіотика чорноморська глибинна вода також може застосовуватися як живильне родюче середовище культивування одноклітинних планктонних та багатоклітинних бентосних водоростей в лабораторних або промислових умовах та в марікультурних господарствах. 13 Комп’ютерна в ерстка Т. Чепелев а 78444 Підписне 14 Тираж 26 прим. Міністерство осв іт и і науки України Держав ний департамент інтелектуальної в ласності, вул. Урицького, 45, м. Київ , МСП, 03680, Україна ДП “Український інститут промислов ої в ласності”, вул. Глазунова, 1, м. Київ – 42, 01601
ДивитисяДодаткова інформація
Назва патенту англійськоюUse of deep sea water from hydrosulphuric zone of black sea as medium for cultivation of algae
Назва патенту російськоюИспользование глубинной морской воды из сероводородной зоны черного моря в качестве среды культивирования морских водорослей
МПК / Мітки
Мітки: води, водоростей, культивування, зони, морських, глибинної, застосування, середовища, чорного, сірководневої, моря, морської
Код посилання
<a href="https://ua.patents.su/7-78444-zastosuvannya-glibinno-morsko-vodi-iz-sirkovodnevo-zoni-chornogo-morya-yak-seredovishha-kultivuvannya-morskikh-vodorostejj.html" target="_blank" rel="follow" title="База патентів України">Застосування глибинної морської води із сірководневої зони чорного моря як середовища культивування морських водоростей</a>
Установка для добування сірководню з морської води

Номер патенту: 53136
Опубліковано: 15.01.2003
Автор: Баранов Дмитро Степанович
МПК: E02D 17/00, C02F 1/20
Мітки: сірководню, установка, морської, добування, води
Формула / Реферат:
Установка для добування сірководню з морської води, яка містить добувне судно, обладнане пристроєм для піднімання розчину сірководню на поверхню судна, яка відрізняється тим, що пристрій для піднімання розчину сірководню виконаний у вигляді ланцюгового конвеєра з прикріпленими до нього ємностями, кожна з яких має редукційний клапан для її герметизації, при цьому в днищі судна установлений водяний затвор з трубою передачі сірководню на...
Спосіб культивування одноклітинних водоростей

Номер патенту: 16270
Опубліковано: 15.08.2006
Автори: Мінюк Галина Семенівна, Гордієнко Алла Павлівна, Єрохін Владислав Євстафійович
МПК: C12N 1/12, C12R 1/89, G01N 21/76
Мітки: одноклітинних, водоростей, культивування, спосіб
Формула / Реферат:
Спосіб культивування одноклітинних водоростей, що включає вирощування водоростей і контроль за розвитком культур, який відрізняється тим, що контроль здійснюють шляхом люмінесцентного спектрального аналізу, який включає попередній відбір по спектру люмінесценції культур водоростей довжин хвиль збудження, регулярну реєстрацію спектрів люмінесценції культивованих водоростей у період їхнього вирощування і визначення співвідношень інтенсивностей...
Спосіб одержання глікопротеїну з морських водоростей

Номер патенту: 62069
Опубліковано: 15.12.2003
Автори: Колоденко Володимир Олександрович, Мікуліч Діана Василівна, Анцупова Людмила Василівна, Руденко Юрій Степанович
МПК: C07K 1/14, A23L 1/0532
Мітки: глікопротеїну, спосіб, одержання, морських, водоростей
Формула / Реферат:
Спосіб одержання глікопротеїну з морських водоростів шляхом гідролізу, який відрізняється тим, що лужний екстракт глікопротеїну одержують шляхом обробки водоростів 2 %-ним розчином їдкого натру за температури 85 - 87оС протягом 2 - 2,5 годин, після чого лужний екстракт нейтралізують розчином сірчаної кислоти до рН ізоелектричної точки білка 4,0, а потім осаджують глікопротеїн з екстракту 80 %-ним етіловим спиртом.
Спосіб визначення токсичності морської води

Номер патенту: 34783
Опубліковано: 15.03.2001
Автори: Тимофєєв Віталій Анатолійович, Бондаренко Людмила Василівна, Оскольська Ольга Іллівна
МПК: A01K 61/00, G01N 33/18
Мітки: морської, токсичності, води, спосіб, визначення
Формула / Реферат:
Спосіб визначення токсичності морської води з використанням, наприклад, середземноморської мідії Mytilus galloprovincialis, який складається в дослідженні м'якунів, побудованні калібровочної прямої і порівнянні з нею даних з різних районів дослідження, який відрізняється тим, що у м'якунів Mytilus galloprovincialis досліджують морфофізичні показники, а саме площу і об'єм зяберних пелюсток, розраховують показник приведеної питомої поверхні So,...
Спосіб видобутку енергетичних газів з сірководневої води глибоководних водоймищ

Номер патенту: 28506
Опубліковано: 16.10.2000
Автори: Рязанов Аркадій Ксенофонтович, Горбенко Юрій Олександрович
МПК: E21B 43/01
Мітки: енергетичних, спосіб, газів, води, сірководневої, водоймищ, видобутку, глибоководних
Формула / Реферат:
Способ добычи энергетических газов из сероводородной воды глубоководных водоемов, включающий продувку воздухом сероводородной воды на глубине забора до полного окисления содержащихся в ней полезных компонентов, отличающийся, тем, что выделение сероводородосодержащих горючих газов осуществляют путем смешивания окисленной до рН = 5,6 сероводородной воды в пропорции 1 : 2,5 с необработанной сероводородной во дои, в следствие чего в смесителе...
Попередній патент: Спосіб одержання гранульованого металевого заліза
Наступний патент: Роторно-поршневий двигун
Випадковий патент: Сепаратор для відділення механічних домішок і рідини високої щільності від рідини низької щільності та газу