Спосіб індукції синтезу нових антибіотиків у актинобактерій
Номер патенту: 94608
Опубліковано: 25.11.2014
Автори: Ющук Олександр Сергійович, Горбаль Лілія Олегівна, Осташ Богдан Омелянович, Федоренко Віктор Олександрович, Мурин Андрій Васильович, Тістечок Степан Іванович






Формула / Реферат
Спосіб індукції синтезу нових антибіотиків у актинобактерій, за яким здійснюють надекспресію регуляторних елементів, який відрізняється тим, що як регуляторний елемент використовують ген плейотропного регулятора транскрипції аdрАAT19 Actinoplanes teichomyceticus, клонований у складі реплікативної плазміди рКСІ 139.
Текст
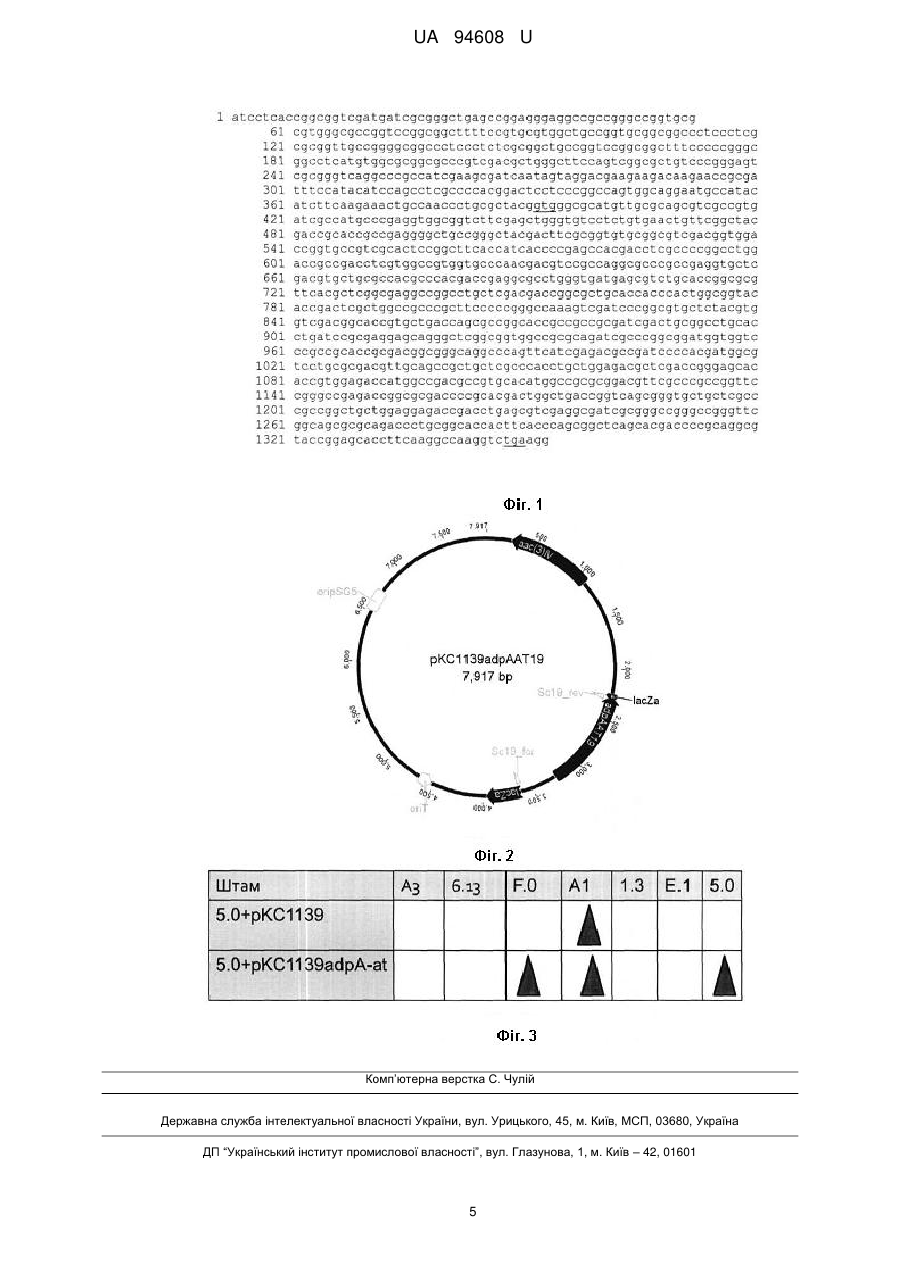
Реферат: Спосіб індукції синтезу нових антибіотиків у актинобактерій, за яким здійснюють надекспресію регуляторних елементів. Як регуляторний елемент використовують ген плейотропного регулятора транскрипції аdрАAT19 Actinoplanes teichomyceticus, клонований у складі реплікативної плазміди рКСІ 139. UA 94608 U (12) UA 94608 U UA 94608 U 5 10 15 20 25 30 35 40 45 50 55 Корисна модель належить до генетики бактерій та біотехнології і може бути використана для отримання нових біологічно активних речовин, що мають антибіотичну дію. Відомий спосіб, за яким суспензії клітин або спор актинобактерій обробляють хімічним або фізичним мутагеном і висівають на агаризовані середовища, що містять один або кілька перелічених нижче антибіотиків: стрептоміцин, рифампіцин, тіострептон. У вибірці колоній, що виростають на таких середовищах, за допомогою способів біологічного скринінгу та аналітичної хімії виявляють продукцію нових антибіотиків [Hosaka Т., Ohnishi-Kameyama Μ., Muramatsu Η., Murakami Κ., Tsurumi Y., Kodani S., Yoshida Μ., Fujie Α., Ochi К. Antibacterial discovery in actinomycetes strains with mutations in RNA polymerase or ribosomal protein S12 // Nat. Biotechnol. 2009. - Vol. 27. - P. 462-464]. Недоліком способу є трудомісткість, тривалість і потреба індивідуального підбору умов мутагенезу. Отримані мутанти часто нестабільні і потребують підтримуючої селекції. Відомий спосіб, що ґрунтується на повному секвенуванні геномів актинобактерій, комп'ютерній анотації усіх генів вторинного метаболізму та надекспресії окремих генів шляхспецифічних регуляторів, які активуватимуть транскрипцію відповідних структурних генів біосинтезу антибіотиків. Нові сполуки виявляють за допомогою біотестів та способами аналітичної хімії [Laureti L., Song L., Huang S., Corre C, Leblond P., Challis G. L., Aigle B. Identification of a bioactive 51-membered macrolide complex by activation of a silent polyketide synthase in Streptomyces ambofaciens II Proc. Natl. Acad. Sci. USA - 2011. - Vol. 108. - P. 62586263]. Недоліком цього способу є економічна невиправданість у великих масштабах, враховуючи тривалість і вартість секвенування й анотації геномів. Автоматична анотація геномів не завжди дає змогу ідентифікувати необхідні шлях-специфічні регуляторні гени, що збільшує ризик негативних результатів. Крім цього існує багато кластерів генів вторинного метаболізму, що не містять шлях-специфічних регуляторів, і для них цей підхід не працюватиме. Відомий спосіб, за яким повністю секвенують геноми актинобактерій, ідентифікують кластери генів вторинного метаболізму та спрямовано делетують (нокаутують) гени репресорного типу, що приводить до синтезу нових сполук [Metsa-Ketela Μ., Ylihonko К., Mantsala P. Partial activation of a silent angucycline-type gene cluster from a rubromycin beta producing Streptomyces sp. PGA64 // J. Antibiot. - 2004. - Vol. 57. - P. 502-510]. Проте, крім високої вартості такого підходу, цей спосіб потребує розробки методів генетичних маніпуляцій геномами досліджуваних штамів, що не завжди можна досягнути для певних видів актинобактерій. Відомий спосіб, за яким частково секвенують геноми актинобактерій, передбачають біоінформатично усі кластери генів вторинного метаболізму; вирощують актинобактерії, що, згідно з даними біоінформатики, містять нові кластери генів, у великому наборі поживних середовищ; вичерпно аналізують спектр синтезованих сполук за різних умов культивування. Сполуки ідентифікують способами біологічних тестів і порівняння мас-спектрів з комерційно доступними хімічними базами даних [McAlpine J.B., Bachmann В.О., Piraee M., Tremblay S., Alarco A.M., Zazopoulos E., Farnet CM. Microbial genomics as a guide to drug discovery and structural elucidation: ECO-02301, a novel antifungal agent, as an example // J. Nat. Prod. - 2005. Vol. 68. - P. 493-496]. Недоліком способу є трудомісткість, він вимагає складної апаратури і доступу до комерційно-доступних хімічних баз даних. Найближчим за технічною суттю - прототипом є спосіб, що ґрунтується на надекспресії гена фосфатази absAl, що походить з Streptomyces coelicolor, у клітинах актинобактерій [McKenzie N. L., Thaker Μ., Koteva К., Hughes D. W., Wright G.D., Nodwell J. R. Induction of antimicrobial activities in heterologous streptomycetes using alleles of the Streptomyces coelicolor gene absAlll J. Antibiot. 2010. - Vol. 63. - P. 177-182]. Ген absAl клонують в інтегративний вектор pSET152 [Kieser Т., Bibb M.J., Buttner M.J., Chater K.F., Hopwood D.A. Practical Streptomyces genetics // John Innes Foundation, Norwich, United Kingdom. - 2000. - 613 pages] під контролем сильного конститутивного промотора гена стійкості до еритроміцину еrmЕр. Сконструйовану плазміду за допомогою міжродової кон'югації [Flett F., Mersinias V., Smith C.P. High efficiency intergeneric conjugal transfer of plasmid DNA from Escherichia coli to methyl-DNA-restricting Streptomycetes II FEMS Microbiol. Lett.-1997. - Vol. 155. - P.223-229] уводять у клітини досліджуваних штамів. Порівнюють спектри синтезованих вторинних метаболітів вихідним і рекомбінантним штамами за допомогою біотестів і мас-спектрометрії. Відбирають ті штами, що мають змінений спектр синтезу, очищують нові сполуки і встановлюють їхню структуру за допомогою загальновживаних способів аналітичної хімії. 1 UA 94608 U 5 10 15 20 25 30 35 40 45 50 55 Проте, цей підхід є вузьким, бо базується на дефосфорилюванні репресора AbsA2 фосфатазою AbsAl, a AbsA2 у вихідному стані має невелике коло операторних ділянок. Коло генів вторинного метаболізму, який він контролює, не є широким. Неможливо індукувати синтез тих нових сполук, які не підлягають репресії з боку AbsA2 В основу корисної моделі поставлено задачу удосконалити спосіб індукції синтезу нових антибіотиків у актинобактерії шляхом надекспресії гена adpAAT19, що кодує плейотропний регулятор транскрипції Actinoplanes teichomyceticus, на основі олігокопійної реплікативної плазміди рКС1139, що дасть змогу індукувати продукцію нових антибіотиків. Поставлене завдання вирішується так, що у способі індукції синтезу нових антибіотиків у актинобактерій, за яким здійснюють надекспресію регуляторних елементів, як регуляторний елемент використовують ген плейотропного регулятора транскрипції adpAAT19 Actinoplanes teichomyceticus, клонований у складі реплікативної плазміди рКСІ 139. Актинобактерії - одні із найважливіших продуцентів медично-важливих антибіотиків, які виявляли переважно за допомогою традиційних способів мікробіологічного скринінгу за біологічною активністю штамів. Потенціал актинобактерій щодо продукції біоактивних сполук за сучасними даними геноміки виглядає значно вищим, ніж це випливає з даних мікробіологічного скринінгу. А саме, типовий геном актинобактерії розміром 8-10 млн. п.н. може містити 20-30 кластерів генів біосинтезу вторинних метаболітів, з яких лише 3-5 мають вияв на рівні продукції відповідної сполуки. Кластери генів вторинного метаболізму, що не експресуються або експресуються дуже слабо названі криптичними, або мовчазними. З літературних джерел відомо, що активація мовчазної частини вторинного метаболому може привести до ідентифікації нових класів сполук, які не вдається виявити традиційними способами скринінгу. Активації мовчазних кластерів синтезу вторинних метаболітів можна досягти за рахунок різноманітних маніпуляцій геномами актинобактерій з подальшим високочутливим хімічним аналізом вторинного метаболізму. Авторами вперше запропоновано використати ген плейотропного регулятора транскрипції adpAAT19 Actinoplanes teichomyceticus для індукції синтезу нових антибіотиків у актинобактерії. Запропонований підхід має низку переваг порівняно із відомими способами активації мовчазних кластерів генів. Біоінформатичний аналіз імовірного продукту трансляції гена adpAAT19 вказує на його гомологічність до білків родини AdpA, які виявлено у роді Streptomyces [Ostash В., Zaburannyi N., Horbal L., Luzhetskyy Α., Shulga S., Fedorenko V. Homologues of pleiotropic regulatory gene adpA in Actinoplanes: analysis in silico II Visn. Lviv Univ. Ser. Biol. - 2012. - Vol. 60. P. 156-162]. Білок AdpA розпізнає дегенеровану декануклеотидну послідовність 5TGGCSNGWWY-3', яка, згідно розрахунків, зустрічається близько 40000 разів у хромосомі модельного об'єкта, продуцента стрептоміцину Streptomyces griseus. Експериментальні дані свідчать, що AdpA контролює експресію більше 500 генів - це найбільший регулон із описаних наразі в біології [Higo Α., Нага Η., Horinouchi S., Ohnishi Υ. Genome-wide distribution of AdpA, a global regulator for secondary metabolism and morphological differentiation in Streptomyces, revealed the extent and complexity of the AdpA regulatory network // DNA Res. - 2012. - Vol. 19. - P. 259-273]. Серед AdpA-залежного регулону виявлено велику кількість генів вторинного метаболізму, і в абсолютній більшості описаних випадків AdpA є позитивним регулятором цих генів [Makitrinskyy R., Ostash В., Tsypik О., Rebets Υ., Doud Ε., Meredith Т., Luzhetskyy Α., Bechthold Α., Walker S., Fedorenko V. Pleiotropic regulatory genes bldA, adpA and absB are implicated in production of phosphoglycolipid antibiotic moenomycin // Open Biol. - 2013. -Vol. 3. - P. 1-13]. Відтак, з усіх описаних на сьогодні плейотропних регуляторів саме білки родини AdpA - найперспективніші для активації мовчазних генів, оскільки мають велику кількість операторних послідовностей і їхнє використання базуватиметься на надекспресії, а не на технічно складніших генних нокаутах. Ген adpAAT19, що походить з Actinoplanes, на відміну від його гомологів зі Streptomyces, не містить у своїй послідовності рідкісного кодона ТТА. Цей кодон у стрептоміцетах декодується UUA виключно однією лейцил-тРНК , яка визначається геном bldA. Наявність рідкісного кодону ТТА у актиноміцетних генах суттєво обмежує їхню експресію і ставить у залежність від появи UUA зрілої лейцил-тРНК [Chater K.F., Chandra G. The use of the rare UUA codon to define "expression space" for genes involved in secondary metabolism, development and environmental adaptation in streptomyces // J. Microbiol. - 2008. - Vol. 46. - P. 1-11]. Такий різновид трансляційної регуляції експресії обмежує застосування генів родини adpA стрептоміцетного походження з метою індукції мовчазного металобому. Ген adpA AT19 Actinoplanes teichomyceticus позбавлений цього недоліку. Оскільки ген adpA AT19 походить з рідкісної актиноміцетної культури, то це підвищує імовірність того, що він матиме унікальну структуру і механізм дії, що відрізняється від таких у його гомологів зі стрептоміцетів. Це дасть змогу активувати експресію тих генів, які 2 UA 94608 U 5 10 15 20 25 30 35 40 45 50 55 60 непідконтрольні типовим білкам AdpA, кодованим геномами стрептоміцетів і індукувати продукцію нових антибіотиків. Фіг. 1 Нуклеотидна послідовність гена adpA AT19 разом з промоторною ділянкою; старт - і стоп - кодони відкритої рамки зчитування adpA AT19 підкреслено. Фіг. 2 Схема плазміди pKCl 139adpA AT19, де: aac(3)IV~ ген стійкості до апраміцину, оrіТ- ділянка кон'югаційного переносу плазміди RK2, adpAAT19 -ген adpA AT19, клонований з хромосоми Actinoplanes teichomyceticus. -pSG5 ori - ділянка ініціації реплікації плазміди pSG5 Фіг. 3 Активація синтезу нових антибіотичних сполук у ґрунтових ізолятах за умов експресії в їхніх клітинах гена adpA AT19, де у затіненому сірим кольором ряді перераховано тестерні штами актиноміцетів, у колонці – два штами, що тестували. Трикутники позначають ті тестерні штами, ріст яких пригнічувався штамами, що тестували. Спосіб можна проілюструвати прикладами: Для надекспресії гена adpA AT19 конструюють плазміду pKC1139adpA AT19. За допомогою полімеразної ланцюгової реакції з використанням полімерази Pfu ампліфікують з хромосомної ДНК Actinoplanes teichomyceticus послідовність ДНК, розміром 1,4 т.п.н., яку зображено на фіг. 1, і що містить досліджуваний ген. Усі маніпуляції виконують згідно стандартних методик [Sambrook J., Russell D. W. Molecular cloning, a laboratory manual. 3rd ed. // Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y. - 2001. - 450 P.]. Для цього використовують пару праймерів Scl9_for (ATCCTC ACCGGCGGTCGATGA) та Sc 19_rev (CCTTCAGACCTTGGCCTTGAAG). Амплікон елююють з агарозного гелю відповідно до стандартних методик [Sambrook J., Russell D. W. Molecular cloning, a laboratory manual. 3rd ed. // Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y. - 2001. - 450 P.]. Далі вектор рКС1139 [Kieser Т., Bibb M.J., Buttner M.J., Chater K.F., Hopwood D.A. Practical Streptomyces genetics // John Innes Foundation, Norwich, United Kingdom. 2000. - 613 pages] обробляють ендонуклеазою рестрикції EcoRV, що залишає тупі кінці. Отримані лінійні фрагменти вектора та досліджуваного гена лігують між собою, далі отриманою сумішшю трансформують компетентні клітини Е. соlі і проводять селекцію трансформантів за стійкістю до апраміцину у концентрації 25 мкг/мл згідно з стандартними методиками [Sambrook J., Russell D. W. Molecular cloning, a laboratory manual. 3rd ed. // Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y. - 2001. - 450 P.]. У новоствореній плазміді pKC1139adpAAT19 транскрипція гена adpAAT19 перебуває під контролем власного промотора, як зображено на фіг. 2. Плазмідою pKC1139adpAAT19 трансформують штам Е. соlі ЕТ12567 (pUB307), який за рахунок /га-генів плазміди pUB307 забезпечує кон'югативне перенесення корезидентних плазмід [Flett F., Mersinias V., Smith C.P. High efficiency intergeneric conjugal transfer of plasmid DNA from Escherichia coli to methyl-DNA-restricting Streptomycetes II FEMS Microbiol. Lett.-1997. Vol.155. - P.223-229]. Одну колонію нічної культури Escherichia соlі засівають у 5 мл середовища LB з 50 мкг/мл канаміцину. Культуру вирощують до оптичної густини OD600 = 0,1, переносять у мікропробірки з об'ємом 1.5 мл і осаджують центрифугуванням при 10 тис. об./хв. протягом 1 хв. Зливають супернантант та клітини ресуспендують у 50 мкл середовища LB. Отриману суспензію клітин охолоджують у льоді 5 хв., додають 1 мл 0,1 Μ розчину СаС12 та інкубують у льоді 1 год. Клітини осаджують центрифугуванням при 10 тис. об./хв.. протягом 1 хв., зливають надосадову рідину та ресуспендують у 100 мкл 0,1М розчину СаСl2. Інкубують 1 год. у льоді та додають розчин плазмідної ДНК. Інкубують 1 год. у льоді, після чого клітини піддають тепловому шоку протягом 1 хв. при 40 °С, охолоджують та додають 1 мл середовища LB. Інкубують 2 год. при 37°С для індукції експресії генів стійкості та висівають на чашки з середовищем LA з 25 мкг/мл апраміцину та 50 мкг/мл канаміцину. Чашки інкубують 16 год при 37°С, після чого трансформанти пересівають на свіже середовище LA з 25 мкг/мл апраміцину та 50 мкг/мл канаміцину. Плазміду pKC1139adpA AT19 переносять у ґрунтові актиноміцетні ізоляти, виділені з ґрунтосуміші «Універсальна» [Мурин А. Штами актиноміцетів з ґрунтосуміші «Універсальна» (компанія «Зелендар»): антибіотична активність та екологічні взаємодії / А. Мурин, С. Тістечок, Т. Грень, Б. Осташ, В. Федоренко // Біорізноманіття. Екологія. Адаптація. Еволюція: VI Міжнар. конф. мол. вч., присв. 150-річчю від дня нар. видати, ботаніка В.І. Липського, 13 - 17 травня 2013 р. - Одеса, 2013. - С. 102] шляхом міжродової кон'югації з відповідним штамом Е. coli ET12567 (pUB307). Суспензію спор актиноміцетного ізоляту висівають на вівсяне середовище (г/л: вівсяне борошно - 30, агар - 18, вода водопровідна - до 1 л, рН до стерилізації - 7,0; після стерилізації додають розчин хлориду магнію до кінцевої концентрації 40 мМ) і вирощують 7 діб при 37 °С 3 UA 94608 U 5 10 15 20 25 30 для отримання спорової суспензії для кон'югаційних схрещувань. Штам Е. coli ET12567 (pUB307) з плазмідою pKCl 139 adpA AT19 висівають на чашку з середовищем LA з 25 мкг/мл апраміцину та 50 мкг/мл канаміцину та вирощують 18 год. при 37°С. Готують суспензію клітин у 4 мл середовища LB. Одночасно готують суспензію спор актиноміцету у 7 мл стерильного фізіологічного розчину та проводять тепловий шок при 50 °С протягом 10 хв. Суспензію клітин та спор стрептоміцетів осаджують центрифугуванням 2 хв. при 10000 об/хв., зливають надосадову рідину, розчиняють у 100 мкл середовища LB, змішують між собою та висівають на чашки з вівсяним середовищем. Чашки інкубують 12-16 год. при 28 °С та заливають 1 мл водного розчину 25 мкг апраміцину та 50 мкг налідиксової кислоти. При проведенні досліду використовують: для визначення температури - ртутний термометр з похибкою показів ± 0,5 °С; об'єму рідин - мірні циліндри з похибкою показів ± 0,5 мл та лабораторні дозатори з похибкою показів ± 0,05-0,5 мкл; маси речовин - аналітичну вагу "Sartorius" з похибкою показів ± 0,00005 г і лабораторну вагу AXIS з похибкою показів ± 0,05 г; час -електронним годинником з похибкою показів ± 0,5 сек. Краплинним способом перевіряють спектр продукції антибіотиків транскон'югантами, що містять pKC1139adpA AT19· Спосіб демонструють на прикладі одного штаму, Streptomyces sp. 5.0; інші штами перевіряють аналогічно. Сім штамів актиноміцетів, що слугуватимуть тестерами продукції нових антибіотиків, і плазмідвмісний штам S. sp. 5.0 (pKC1139adpA AT19) висівають на вівсяне агаризоване середовище (г: овес мелений - 15, агар - 18, вода водопровідна - до 400 мл, рН до стерилізації -8.0). Чашки інкубують протягом 140 год, при 30 °С. Шпателем Дригальського з газону штамів збирають спори у стерильні мікропробірки. Спорову суспензію концентрують за допомогою центрифугування (10 тис.об/хв., 3 хв.) до титру спор не менше 8 5810 . 3 мкл спорової суспензії рекомбінантного штаму наносять на чашку з середовищем Чапека (г/л: сахароза - 30,0; NaNO3 - 2,0; MgSO4 - 0,5; FeSO4 - 0,01; КН2РО4 - 1,0; КСl -0,5; дистильована вода -1,0 л.). Поруч наносять 3-кмл краплю одного з тестерних штамів так, щоб проміжок між краплями становив 23 мм. Повторюють цю процедуру для інших тестерних штамів. Синтез нових антибіотиків рекомбінантним штамом S. sp. 5.0 (pKC1139adpA AT19) виявляють за здатністю пригнічувати ріст тих тестерних штамів, які не пригнічуються штамом, що містить вектор рКС1139 (S. sp. 5.0 (рКС1139). Отримані дані узагальнено в таблиці на фіг. 3, де здатність пригнічувати ріст позначено трикутником. Помітно, що контрольний штам S. sp. 5.0 (рКСІ 139) пригнічує ріст лише одного тестерного штаму - А1, у той час як рекомбінантний штам пригнічує ріст ще двох штамів - F1 та вихідного 5.0. Уведення гена adpA AT19 у клітини актинобактерій активує синтез нових антибіотиків, які за відсутності цього гена не продукуються. Це підтверджує отримання передбачуваного результату. 35 ФОРМУЛА КОРИСНОЇ МОДЕЛІ 40 Спосіб індукції синтезу нових антибіотиків у актинобактерій, за яким здійснюють надекспресію регуляторних елементів, який відрізняється тим, що як регуляторний елемент використовують ген плейотропного регулятора транскрипції аdрАAT19 Actinoplanes teichomyceticus, клонований у складі реплікативної плазміди рКСІ 139. 4 UA 94608 U Комп’ютерна верстка С. Чулій Державна служба інтелектуальної власності України, вул. Урицького, 45, м. Київ, МСП, 03680, Україна ДП “Український інститут промислової власності”, вул. Глазунова, 1, м. Київ – 42, 01601 5
ДивитисяДодаткова інформація
Автори англійськоюOstash Bohdan Omelianovych, Fedorenko Viktor Oleksandrovych
Автори російськоюОсташ Богдан Емельянович, Федоренко Виктор Александрович
МПК / Мітки
МПК: C12Q 1/04, C12Q 1/00, C12Q 1/02
Мітки: індукції, нових, антибіотиків, спосіб, актинобактерій, синтезу
Код посилання
<a href="https://ua.patents.su/7-94608-sposib-indukci-sintezu-novikh-antibiotikiv-u-aktinobakterijj.html" target="_blank" rel="follow" title="База патентів України">Спосіб індукції синтезу нових антибіотиків у актинобактерій</a>
Спосіб стимулювання ендокринної функції тимуса та індукції синтезу речовин з тимозиноподібною активністю при тимусектомії

Номер патенту: 104970
Опубліковано: 25.03.2014
Автори: Мельничук Дмитро Олексійович, Грищенко Вікторія Анатоліївна
МПК: A61P 5/00, A61K 31/683, A61K 31/07, A61K 31/355
Мітки: функції, синтезу, речовин, спосіб, ендокринної, тимозиноподібною, тимуса, активністю, стимулювання, тимусектомії, індукції
Формула / Реферат:
Спосіб стимулювання ендокринної функції тимуса та індукції синтезу речовин з тимозиноподібною активністю при тимусектомії, який включає застосування тваринам ліпосомальної форми 0,7-0,9 % розчину фосфоліпідовмісної суміші з маслянки, суміші ненасичених жирних кислот та вітамінів А і Е, який відрізняється тим, що тваринам при недостатності ендокринної функції тимуса та при його хірургічному видаленні однократно, внутрішньочеревинно вводять БАД...
Спосіб підвищення продукції антибіотиків моеноміцинового ряду

Номер патенту: 79968
Опубліковано: 13.05.2013
Автори: Ципік Ольга Володимирівна, Осташ Богдан Омелянович, Макітринський Роман Павлович, Федоренко Віктор Олександрович
МПК: C12Q 1/00, C12Q 1/04, C12Q 1/02
Мітки: моеноміцинового, ряду, антибіотиків, продукції, спосіб, підвищення
Формула / Реферат:
Спосіб підвищення продукції антибіотиків моеноміцинового ряду, який базується на надекспресії генів плейотропних регуляторів у актиноміцетах-продуцентах цих антибіотиків, який відрізняється тим, що як регуляторний елемент використовують ген плейотропного регулятора транскрипції adpAgh S. ghanaensis, клонований у складі інтегративної плазміди pTESaadpA-exp у штамах S. ghanaensis АТСС14672, S. lividans TK24, S. coelicolor M1152, S. albus...
Спосіб підвищення біосинтезу фосфогліколіпідних антибіотиків

Номер патенту: 63820
Опубліковано: 25.10.2011
Автори: Уокер-Кане С'юзен, Федоренко Віктор Олександрович, Осташ Богдан Омелянович, Громико Олександр Миколайович
МПК: C12Q 1/00, C12Q 1/04, C12Q 1/02
Мітки: антибіотиків, фосфогліколіпідних, біосинтезу, спосіб, підвищення
Формула / Реферат:
Спосіб підвищення біосинтезу фосфогліколіпідних антибіотиків, який базується на надекспресії генів плейотропних регуляторів у актиноміцетах-продуцентах цих антибіотиків, який відрізняється тим, що як регуляторний елемент використовують ген лейцил-тРНКUUА S. lividans, клонований у складі висококопійної плазміди pIJ584 у штамах S. ghanaensis ATCC14672, S. lividans ТК24, S. albus J1074.
Спосіб виявлення антибіотиків тіострептонового ряду

Номер патенту: 91725
Опубліковано: 25.08.2010
Автори: Осташ Ірина Степанівна, Мироновський Максим Леонідович, Федоренко Віктор Олександрович, Осташ Богдан Омелянович
МПК: C12Q 1/02, C12Q 1/00, C12Q 1/04
Мітки: спосіб, антибіотиків, ряду, виявлення, тіострептонового
Формула / Реферат:
Спосіб виявлення антибіотиків тіострептонового ряду, що базується на індукції експресії гена канаміциностійкості nео у Streptomyces lividans, який відрізняється тим, що як векторну молекулу ДНК використовують інтегративну плазміду рIJ6902.
Спосіб експрес-виявлення антибіотиків групи ландоміцинів

Номер патенту: 88383
Опубліковано: 12.10.2009
Автори: Осташ Богдан Омелянович, Федоренко Віктор Олександрович, Бехтольд Андреас, Лужецький Андрій Миколайович, Осташ Ірина Степанівна
МПК: C12Q 1/04
Мітки: експрес-виявлення, ландоміцинів, антибіотиків, групи, спосіб
Формула / Реферат:
Спосіб експрес-виявлення три-, пента- і гексаглікозильованих ландоміцинів, який базується на індукції експресії гена канаміцин-стійкості nео у стрептоміцетів, який відрізняється тим, що як регуляторний елемент використовують ген lаnК, клонований у складі плазміди рМО11с у актиноміцетів Streptomyces albus.
Попередній патент: Спосіб вимірювання часу реакції людини на зовнішнє подразнення
Наступний патент: Спосіб візуалізації локальних полів у метал-діелектричних нанокомпозитних матеріалах
Випадковий патент: Розвантажувально-роз'єднувальна установка рру-21