Атенуйований рекомбінантний вірус герпесу великої рогатої худоби типу 1 (bhv-1), вакцина, що його містить, та спосіб виявлення тварин, яким була введена рекомбінантна вірусна вакцина







Формула / Реферат
1. Атенуйований рекомбінантний вірус герпесу великої рогатої худоби типу І, який являє собою вірус герпесу великої рогатої худоби типу І, в якому частина нативної кодуючої ділянки глікопротеїну Е видалена і замінена генетичною вставкою, яка включає кодуючу ділянку стороннього гена ![]() -галактозидази і негайний ранній промотор цитомегаловірусу людини, і при цьому даний рекомбінантний вірус здатний експресувати
-галактозидази і негайний ранній промотор цитомегаловірусу людини, і при цьому даний рекомбінантний вірус здатний експресувати ![]() -галактозидазу в клітині-хазяїні.
-галактозидазу в клітині-хазяїні.
2. Вірус за п. 1, в якому генетична вставка додатково включає енхансерні послідовності зазначеного негайного раннього промотору цитомегаловірусу людини.
3. Вірус за п. 2, в якому видалено приблизно дві третини нативної кодуючої ділянки глікопротеїну Е.
4. Вірус за п. 2, в якому генетична вставка зв'язана із сайтами поліаденілування SV40.
5. Вірус за п. 1, в якому сторонній ген ![]() -галактозидази експресується як автентичний ген, що кодується вірусом герпесу великої рогатої худоби типу І.
-галактозидази експресується як автентичний ген, що кодується вірусом герпесу великої рогатої худоби типу І.
6. Вірус за п. 1, в якому відсутність частини нативної кодуючої ділянки глікопротеїну Е є генотипічним маркером.
7. Вірус за п. 1, в якому експресована в клітині-хазяїні ![]() -галактозидаза є фенотипічним маркером.
-галактозидаза є фенотипічним маркером.
8. Вірус за п. 1, в якому відсутність частини нативної кодуючої ділянки глікопротеїну Е є імунологічним маркером.
9. Вірус за п. 1, в якому кодуюча ділянка ![]() -галактозидази експресується в зазначеній клітині-хазяїні як на ранній, так і на пізній стадіях інфекції.
-галактозидази експресується в зазначеній клітині-хазяїні як на ранній, так і на пізній стадіях інфекції.
10. Вірус за п. 1, депонований під номером ATCC VR-2637.
11. Рекомбінантна вакцина, яка має активність щодо індукування антитіл, що нейтралізують вірус герпесу великої рогатої худоби типу І, яка містить атенуйований рекомбінантний вірус за будь-яким із пп. 1-10.
12. Спосіб виявлення тварин, яким була введена рекомбінантна вірусна вакцина за п. 11, який включає такі стадії:
(a) одержання від тварин зразка рідини, що містить відповідний вірус;
(б) інфікування клітин згаданим вірусом зі зразка рідини;
(в) виявлення в інфікованих клітинах вірусу з вакцини за п. 11.
13. Спосіб за п. 12, в якому стадія (в) включає аналіз на присутність ![]() -галактозидази.
-галактозидази.
14. Спосіб за п. 12, в якому стадія (в) включає аналіз на гуморальну імунну відповідь, специфічну для глікопротеїну Е.
15. Спосіб за п. 12, який включає стадію виявлення згаданих тварин, що ґрунтується на наявності гуморальної імунної відповіді, специфічної для глікопротеїну Е .
16. Спосіб за п. 12, в якому стадія (в) включає in situ гістохімічні методи виявлення активності ферменту ![]() -галактозидази.
-галактозидази.
17. Спосіб за п. 12, в яком стадія (в) включає in situ імуно-гістохімічні методи виявлення білка ![]() -галактозидази.
-галактозидази.
18. Спосіб за п. 12, в якому стадія (в) включає імуноблотинг для виявлення білка ![]() -галактозидази.
-галактозидази.
19. Спосіб за п. 12, в якому стадія (в) включає in situ гістохімічні методи виявлення глікопротеїну Е.
20. Спосіб за п. 12, в якому виявляють тварин, яким була введена рекомбінантна вірусна вакцина, що містить атенуйований вірус за п. 10.
21. Плазміда, депонована під номером АТСС 203607, яка може бути використана для одержання рекомбінантного вірусу за п. 10.
22. Спосіб імунізації великої рогатої худоби проти вірусу герпесу великої рогатої худоби типу І, який включає вакцинацію зазначеної великої рогатої худоби рекомбінантною вірусною вакциною за п. 11.
23. Спосіб за п. 22, який відрізняється тим, що рекомбінантна вірусна вакцина містить атенуйований вірус за п. 10.
Текст
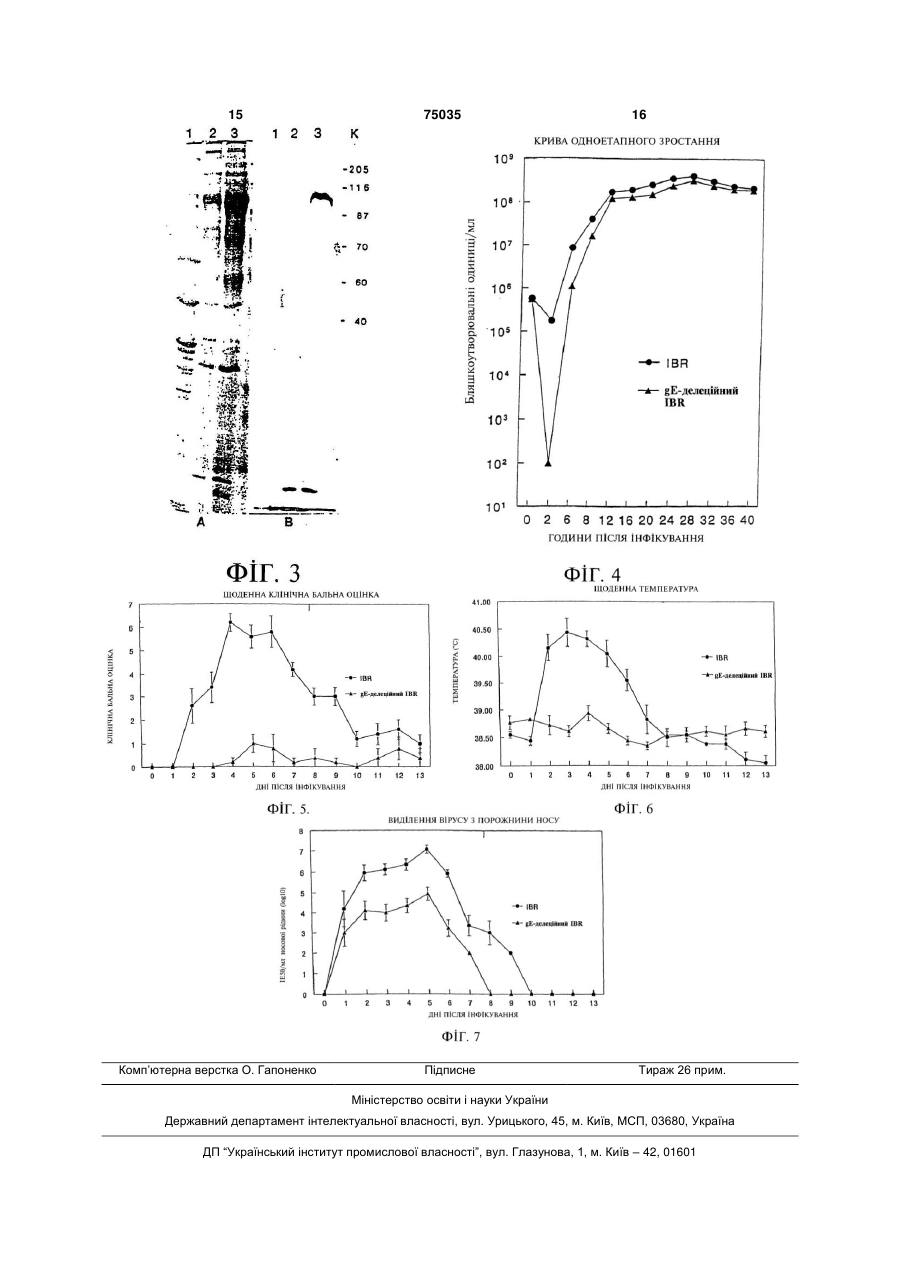
1. Атенуйований рекомбінантний вірус герпесу великої рогатої худоби типу І, який являє собою вірус герпесу великої рогатої худоби типу І, в якому частина нативної кодуючої ділянки глікопротеїну Е видалена і замінена генетичною вставкою, яка включає кодуючу ділянку стороннього гена галактозидази і негайний ранній промотор цитомегаловірусу людини, і при цьому даний рекомбінантний вірус здатний експресувати -галактозидазу в клітині-хазяїні. 2. Вірус за п.1, в якому генетична вставка додатково включає енхансерні послідовності зазначеного негайного раннього промотору цитомегаловірусу людини. 3. Вірус за п. 2, в якому видалено приблизно дві третини нативної кодуючої ділянки глікопротеїну Е. 4. Вірус за п. 2, в якому генетична вставка зв'язана із сайтами поліаденілування SV40. 5. Вірус за п. 1, в якому сторонній ген галактозидази експресується як автентичний ген, що кодується вірусом герпесу великої рогатої худоби типу І. 2 (19) 1 3 75035 4 17. Спосіб за п.12, в яком стадія (в) включає in situ 21. Плазміда, депонована під номером АТСС 203607, яка може бути використана для одержанімуно-гістохімічні методи виявлення білка ня рекомбінантного вірусу за п.10. галактозидази. 22. Спосіб імунізації великої рогатої худоби проти 18. Спосіб за п.12, в якому стадія (в) включає вірусу герпесу великої рогатої худоби типу І, який імуноблотинг для виявлення білка включає вакцинацію зазначеної великої рогатої галактозидази. худоби рекомбінантною вірусною вакциною за 19. Спосіб за п.12, в якому стадія (в) включає in п.11. situ гістохімічні методи виявлення глікопротеїну Е. 23. Спосіб за п. 22, який відрізняється тим, що ре20. Спосіб за п.12, в якому виявляють тварин, комбінантна вірусна вакцина містить атенуйований яким була введена рекомбінантна вірусна вакцивірус за п.10. на, що містить атенуйований вірус за п.10. Цей винахід в широкому значенні стосується вакцин на основі рекомбінантного вірусу герпесу великої рогатої худоби і відповідних способів. Зокрема, цей винахід відповідно до переважного варіанту стосується конструювання інфекційного рекомбінантного вірусу герпесу великої рогатої худоби типу 1 (BHV-1), який має видалену частину нативної кодуючої області глікопротеїду Ε (gE) і функціональний ген -галактозидази ( -gal), вставлений на її місце на локусі gE. Зазначена делеція нативної кодуючої області gE спричиняє ослаблення вірулентності зазначеного вірусу і служить як генотипічний або імунологічний маркер, який дозволяє відрізнити інфекцію gE-делеційного рекомбінантного вірусу від інфекції, викликаної вірусом дикого типу. У доповнення до цього, інсерція зазначеного -gal гена забезпечує отримання фенотипічного методу аналізу на присутність інфекції, зумовленої gE-делеційним рекомбінантним вірусом, за допомогою експресії -gal активності в клітинах-хазяях. Вірус герпесу великої рогатої худоби типу 1 (BHV-1), відомий також як вірус інфекційного ринотрахеїту великої рогатої худоби (IBRV), є пов'язаний з різними клінічними захворюваннями, в тому числі з ринотрахеїтом, кон'юнктивітом, інфекціями статевих шляхів і, іноді, абортами, ентеритами, енцефалітом і генералізованими інфекціями великої рогатої худоби. Геном BHV-1 включає лінійну дволанцюжкову молекулу ДНК довжиною біля 140тис.п.н. Вона складається з унікальної довгої (UL) ділянки і унікальної короткої (US) ділянки, які фланкуються внутрішнім і кінцевим інвертованим повторами (IR і TR, відповідно). Геном BHV-1 кодує приблизно 70 білків [Micpa (Misra) і інші, Proteins Specified Bovine Herpesvirus 1 (Infectious Bovine Rhinotracheitis, 40 J. Virol. 367-378 (1981)]. Подібно до деяких інших вірусів герпесу тварин, геном BHV-1 кодує ген глікопротеїду (g) gE. Є повідомлення про послідовності гена gE BHV-1, який кодує 575 амінокислотних залишків, для двох різних штамів [Ланг-Тек П. (Leung-Taek P.) і інші, The Complete DNA Sequence and the Genetic Organization of the Short Unique Region (US) of the Bovine Herpesvirus Type 1 Strain (ST strain), 199 Virology 409-421 (1994); Ребордоза Kc. (Rebordosa X.) і інші, Mapping, Cloning and Sequencing of a Glycoprotein-Encoding Gene From Bovine Herpesvirus Type 1 Homologous to the gE Gene From HSV-1, 149 Gene 203-209 (1994)]. Передбачені gE амінокислоти включають ділянки гідрофобних амінокислот на N-кінцевій області (передбачувана сигнальна послідовність) і біля С-кінцевої області (трансмембранна послідовність), що є типовим для інтегральних білків мембрани класу І. Було показано, що gE BHV-1 і його гомологи у інших вірусів герпесу не є обов'язковими для реплікації in vitro, однак наслідком делеції всієї кодуючої послідовності gE генома вірусу псевдосказу (PRV) є як знижена вірулентність живих вакцинних штамів Norgen і Bartha [Петровскіс Е.А. (Petrovskis E.A.) і інші, Deletion in Vaccine Strains of Pseudorabies Virus and Their Effect on Synthesis of Glycoprotein gp 63, 60 J. Virol. 1166-1169 (1986)], так і зміна нейроінвазивності [Кард Дж.П. (Card J.P.) і інші, Pseudorabies Virus Envelope Glycoprotein gl Influences Both Neurotropism and Virulence During Infection of the Rat Visual System, 66 J. Virol. 3032-3041 (1992)]. Таким чином, експресія зазначеного гена gE є необхідною для повного патогенного потенціалу вірусів у тварин, але не потрібна для зростання в культурі клітин тканини [Критас (Kritas) і інші, Invasion and Spread of Single Glycoprotein Deleted Mutants of Aujeszky's Disease Virus (ADV) in the Trigeminal Nervous Pathway of Pigs After Intranasal Inoculation, 50 Vet. Micobiol. 323-334 (1994); Критас (Kritas) і інші, Role of Envelope glycoproteins gl, gp63 and glll in the Invasion and Spread of Aujeszky's Disease Virus in the Olfactory Nervous Pathway of the Pig, 75 J. General Virol. 2319-2327 (1994)]. Недавно мутанти PRV і IBR, делеційні по гену gE, викликали інтерес завдяки своїй придатності як диференційні маркерні вакцини. У цей час gEделеційна маркерна вакцина використовується при викорінюванні IBR в Європі. Однак у зазначеного gE-делеційного вакцинного штаму в Європі відсутній -gal маркер, який забезпечує можливість застосування in situ гістохімічних методів для виявлення активності -gal ферменту і in situ гістохімічних методів або методів імуноблотингу для виявлення -gal білка. Зазначена кодуюча область -gal служить також як генотипічний маркер зазначеного рекомбінантного вірусу. Зазначений вірус може легко виявлятися саузерн-блотгібридизацією, і шляхом полімеразно-ланцюгової реакції (PCR) також можна легко відрізнити генетично чистий вакцинний вірус від вірусу дикого 5 75035 6 типу. Таким чином, є необхідним авірулентний gEвірусу і відповідає результатам, отриманим для делеційний штам IBRV, до складу якого входив би інших вірусів герпесу. придатний фенотипічУ експериментах на тваринах телята, інфіконий/гістохімічний/генотипічний -gal маркер. вані gE 3.1IBR , протягом періоду виділення віруСконструювали рекомбінантний вірус BHV-1 су виділяли приблизно в 100 разів менше вірусу в порівнянні з телятами, інфікованими батьківським (gE 3.1IBR ), у якого відкриті рамки зчитування штамом Cooper. Тривалість виділення вірусу була (ORF's), які включають частину послідовностей, також на два дні коротша у телят, інфікованих що кодують ген gE, були видалені і на їхнє місце вставлений химерний (рекомбінантний) репортерgE 3.1IBR . У той час як телята, інфіковані віруний/маркерний ген. Зазначений вставлений ген сом gE 3.1IBR , залишалися здоровими, у телят, інфікованих батьківським штамом Cooper, спостегалактозидази ( -gal) не відіграє регуляторної ролі рігалися типові для IBR симптоми і ураження. Рев реплікації зазначеного вірусу, однак служить як зультати реакції сироваткової нейтралізації покафенотипічний маркер для вірусу gE 3.1IBR . зали, що у телят, інфікованих як вірусом дикого Для конструювання зазначеного рекомбінанттипу, так і gE-делеційним вірусом IBR, розвивалиного BHV-1 клонували кодуючу область гена gE ся порівнянні сироваткові титри BHV-1 нейтраліBHV-1 і фланкуючі послідовності 3'-5' і 5'-3' дирекзуючих антитіл. Раніше повідомлялося, що IBRV, ційного типу. Для створення делеції в зазначеній делеційний по гену тимідинкінази (ТК), ріс як in кодуючій області гена gE, вищезазначену клоноvitro, так і in vivo зі значно більш низьким титром вану ДНК розщеплювали за допомогою придатних [Чоудхарі С.І. (Chowdhury S.I.), Construction and ферментів для виділення двох третин амінокінцеCharacterization of an Attenuated Bovine Herpesvirus вої послідовності цієї області і лігування із зазнаType 1 (BHV-1) Recombinant Virus, 52 Vet. ченим -gal геном. Отриману плазмідну ДНК котMicrobiol. 13-23 (1996)]. Ці результати в узагальнерансфектували з ДНК первісного вірусу IBR дикого ному вигляді вказують на те, що навіть незважаютипу (штам Cooper) в клітини MDBK (культура клічи на те, що зазначений рекомбінантний вірус тин нирок великої рогатої худоби лінії MadingE 3.1IBR росте відносно добре в порівнянні із Darby). Рекомбінантні віруси, експресовані -gal ТК-делеційним вірусом IBR, вірус gE 3.1IBR був (сині бляшки), очищали з вірусними нуклеїновими фактично авірулентним для телят. Атенуйовані кислотами в бляшках і піддавали додатковому властивості, які демонструються рекомбінантним аналізу шляхом блот-гібридизації для визначення вірусом gE 3.1IBR , нагадують властивості, отригенетичних характеристик і шляхом імуноблотингу мані: 1) делецією ТК у BHV-1, як було показано для визначення реакційної здатності проти BHV-1 [Чоудхарі С.І. (Chowdhury S.I.) в статті gE-специфічних кролячих поліклональних антитіл. "Construction and Characterization of an Attenuated Один із рекомбінантних вірусів, gE 3.1IBR , хараBovine Herpesvirus Type 1 (BHV-1) Recombinant ктеризували in vitro відносно його ростових власVirus" (52 Vet. Microbiol. 13-23 (1996))]; 2) делецією тивостей і in vivo на телятах відносно його патоgE у PRV, як було показано [Крітас (Kritas) і іншигенних властивостей. Здатність зазначеного ми в статті "Invasion and Spread of Single рекомбінантного вірусу до індукування BHV-1 нейGlycoprotein Deleted Mutants of Aujeszky's Disease тралізуючих антитіл у інфікованих телят досліджуVirus (ADV) in the Trigeminal Nervous Pathway of вали за допомогою реакції придушення бляшкоутPigs After Intranasal Inoculation" (50 Vet. Micobiol. ворення. 323-334 (1994)) і Крітас (Kritas) і іншими в статті Регуляція і експресія зазначеного химерного "Role of Envelope Glycoproteins gl, gp63 and gІll in гена -gal унікальна для цього рекомбінантного the Invasion and jSpread of Aujeszky's Disease Virus вірусу BHV-1 в двох відношеннях. Перший унікаin the Olfactory Nervous Pathway of the Pig" (75 J. льний аспект цього рекомбінантного вірусу поляGeneral Virol. 2319-2327 (1994))]; і 3) європейським гає в тому, що зазначений -gal ген регулюється gE-делеційним IBRV вакцинним ізолятом, як покаактивним негайним раннім промотором цитомегазали [Каашек (Kaashoek) і інші в статті "An ловірусу людини (HCMV-IE) (а не регуляторною Inactivated Vaccine Based on a Glycoprotein Εпослідовністю, отриманою від BHV-1). Другий уніNegative Strain of Bovine Herpesvirus 1 Induces кальний аспект полягає в тому, що зазначений ген Prospective Immunity and Allows Serological експресувався як BHV-1 ген, який кодується як на Differentiation" (13 Vaccine 342-346 (1995)) і Ван ранній, так і на пізній стадії інфекції. Властивості in Енгленбург (Van Englenburg) і інші в статті "А vitro і in vivo зазначеного gE-делеційного рекомбіGlycoprotein Ε Deletion Mutant of Bovine Herpesvirus нантного вірусу аналізувалися за допомогою порі1 Infects the Same Limited Number of Tissues in вняння його з батьківським штамом IBRV Cooper. Calves as Wild-Type Virus, but for a Shorter Period" У експериментах із культивуванням на кліти(76 J. Gen. Virol. 2387-2392 (1994))]. нах тканин вірус gE 3.1IBR на початковій стадії Результати цього дослідження також показапісля інфікування ріс із більш низьким титром, ніж ли, що делеція зазначених gE ORF послідовносвірус дикого типу (батьківський штам Cooper), одтей і інсерція функціонального -gal гена в gE лонак на більш пізніх стадіях після інфікування закус зазначеного вірусу стабільно атенуювала цей значений рекомбінантний вірус ріс майже з таким вірус. Практичне застосування цього вірусу поляже титром, що і штам дикого типу. Зазначений гає в його використанні як безпечної живої вакцирекомбінантний вірус, як правило, утворював знани, спрямованої проти IBR. Делеція зазначеного чно менші бляшки в порівнянні із бляшками батьgE гена буде служити як імунологічний маркер для ківського штаму дикого типу Cooper Це може поясрозрізнення вакцинованих і інфікованих тварин. У нюватися відсутністю міжклітинного поширення 7 75035 8 Приклад 1 доповнення до цього, продукування -gal забезпеМатеріали і методи чувало б можливість легкої оцінки реплікації вірусу У цьому прикладі приведений опис отримання gE 3.1IBR в епітелії порожнини носа вакциноваі характеризації рекомбінантного IBRV, делеційноних тварин в порівнянні з gE-делеційним вакцинго по гену gE і такого, що експресує ним штамом, який використовується в цей час в галактозидазу. До складу зазначеного сконструйоЄвропі, у якого відсутній цей фенотипічний -gal ваного рекомбінантного вірусу входить химерний маркер, а також для відрізнення її від інфекції, ген (довжиною 4,5тис.п.н.), який заміщає gE кодувикликаної вірусом IBR дикого типу. ючу область BHV-1. Зазначений химерний ген в На Фіг.1 представлена схема послідовності переважному варіанті включає HCMV-IE промотор операцій конструювання gE-делеційного вірусу і його енхансерні послідовності, зв'язані з кодуюBHV-1 (gE 3.1IBR ) і інсерційної/делеційної векточими послідовностями гена -gal, які зв'язані з рної плазміди, яка включає функціональний -gal сайтами поліаденілування SV40. Зазначені -gal ген, що заміщає дві третини амінокінцевої ділянки кодуючі послідовності в переважному варіанті є зазначеної кодуючої області gE; бактерійними, вважають, однак, що придатними На Фіг.2 представлено відтворення саузернбудуть будь-які кодуючі послідовності -gal гена. блот-гібридизації для призначеної делеції кодуюДля конструювання і характеризації рекомбічої області gE і інсерції -gal послідовностей в gE нантного BHV-1 з Американської колекції типових локус; культур (American Type Culture Collection) був На Фіг.3 показаний BHV-1 gE білок, виявлений отриманий штам IBRV Cooper (Colorado-1). Віруси у батьківського штаму дикого типу Cooper, але не розмножали і титрували в клітинах нирок великої виявлений у gE-делеційного рекомбінантного вірурогатої худоби лінії Madin-Darby за методом [Чоусу (gE 3.1IBR ); дхарі С.І. (Chowdhury S.I.), Molecular Basis of Фіг.4 являє собою графік, який ілюструє реAntigenic Variation Between Glycoprotein С (gC) of зультати одноетапного експеримента з вирощуRespiratory Bovine Herpesvirus 1 (BHV-1) and ванням, на якому порівнюється швидкість зросNeurovirulent BHV-5, 213 Virology 558-568 (1995)], тання gE-делеційного вірусу IBR зі швидкістю який включений до цього опису як посилання. Візростання вірусу IBR дикого типу в клітинах MDBK; русну ДНК виділялиза допомогою додецилсулькожна експериментальна точка представляє серефату натрію і лизису білка К, екстрагування суміднє значення результатів, отриманих в кожній грушшю фенолу/хлороформу і осадження етанолом пі телят; згідно з описом методу, приведеним в статті [ЧоуФіг.5 являє собою графік, на якому порівнюдхарі (Chowdhury) і інших, Equine Herpesvirus Type ються щоденні клінічні бальні показники телят, 1 (EHV-1) Induced Abortions and Paralysis in a інфікованих gE-делеційним вірусом IBR, зі щоденLipizzanner Stud: A Contribution to the Classification ними клінічними бальними показниками телят, of Equine Herpesiruses, 90 Arch. Virol. 273-288 інфікованих вірусом IBR дикого типу; кожна експе(1986)], який включений до цього опису як посириментальна точка представляє середнє значення лання. результатів, отриманих в кожній групі телят; BHV-1 gE-специфічну антипептидну кролячу Фіг.6 являє собою графік, на якому порівнюполіклональну сироватку синтезували, виходячи з ються щоденні ректальні температури телят, інфіпередбаченої регіональної гідропатичності і антикованих gE-делеційним вірусом IBR, зі щоденними генності. Синтезували зазначений пептид gE, який ректальними температурами телят, інфікованих включає залишки 378-398, як описали [Ланг-Тек П. вірусом IBR дикого типу; кожна експериментальна (Leung-Taek P.) і інші, The Complete DNA Sequence точка представляє середнє значення результатів, and the Genetic Organization of the Short Unique отриманих в кожній групі телят; і Region (US) of the Bovine Herpesvirus Type 1 Strain Фіг.7 являє собою графік, на якому порівню(ST strain), 199 Virology 409-421 (1994)], послідовється виділення з порожнини носа gE-делеційного ність і генетична організація якого включена до вірусу IBR із виділенням із порожнини носа вірусу цього опису як посилання. Синтезування зазначеIBR дикого типу; кожна експериментальна точка ного пептиду gE було здійснене за методом із випредставляє середнє значення результатів, отрикористанням 9-флуоренілметоксикарбонілу маних в кожній групі телят. (FMOC), опис якого приведений в статті [АбдельУ представлених далі прикладах приведений магід (Abdelmagid) і інших, Fine Mapping of Bovine опис конструювання інфекційного рекомбінантного Herpesvirus-1 (BHV-1) Glycoprotein D (gD) BHV-1, який має видалену нативну кодуючу обNeutralizing Epitopes by Type-Specific Monoclonal ласть gE, що ослабляє вірулентність зазначеного Antibodies and Sequence Comparison With BHV-5 вірусу, і який має функціональний -gal ген, вставgD, 206 Virology 242-253 (1995)], і включено до лений в локус gE, способу імунізації тварин проти цього опису як посилання. Для полегшення кон'юзахворювань, які викликаються BHV-1, із викорисгації з гемоціаніном лімфи равлика (KLH), до Станням зазначеного рекомбінантного BHV-1 як кінцевої області зазначеного пептиду був доданий вакцини, а також способів виявлення і розрізнення додатковий сторонній цистеїн (С) (помічений значяк генотипічно, так і фенотипічно, інфекції, викликом *). Зазначений 17-членний пептид [H]каної у тварин зазначеним рекомбінантним віруTSDRLVRAVTDHTRPEC*-[OH] приєднували до сом і вірусом дикого типу. Ці приклади приведені гемоціаніну лімфи равлика і антисироватки отрилише з ілюстративними цілями, і їх не треба приймували за описом методу, який був приведений в мати як обмеження загального обсягу цього винастатті [Кайт Дж. (Kyte J.), Дулітл Р.Ф. (Doolittle ходу. R.F.), A Simple Method for Displaying the 9 75035 10 Hydropathic Character of Protein, 157 J. Mol. Biol. (інвентарний номер американської колекції типо105-132 (1982)], і який включений до цього опису вих культур (АТСС) VR-2637) і первісну ДНК IBRV як посилання. дикого типу (штам Cooper) котрансфектували в Електрофорез в поліакриламідному гелі в прикультуру клітин нирок великої рогатої худоби лінії сутності додецилсульфату натрію і вестерн-блот Madin-Darby (MDBK). Коректна інсерція -gal в псевдо- і вірус-інфікованих клітинних білків здійслокус gE первісного IBRV і подальша делеція часнювали в відновлювальних умовах, як описано в тини gE-кодуючої області IBRV дикого типу зумовстаттях [Чоудхарі С.І. (Chowdhury S.I.), Molecular лені гомологічною рекомбінацією специфічних Basis of Antigenic Variation Between Glycoprotein С фланкуючих послідовностей BHV-1 в плазміді з (gC) of Respiratory Bovine Herpesvirus 1 (BHV-1) реплікацією вірусною ДНК. Зазначена вірусспециand Neurovirulent BHV-5, 213 Virology 558-568 фічна фланкуюча послідовність включається в (1995) і Лемлі У.К. (Laemli U.K.), Cleavage of новосинтезовану вірусну ДНК, створюючи делецію Structural Proteins During the Assembly of the Head в кодуючій області, а також інсерцію в тій же самій of Bacteriophage T4, 227 Nature (London) 680-685 області. Рекомбінантні віруси, експресовані -gal, (1970)]; обидва зазначених методи включені до тричі зазнавали очищення з вірусними нуклеїноцього опису як посилання. вими кислотами в бляшках шляхом відбору синіх Конструювання рекомбінантних плазмід було бляшок під покривним шаром Bluo-Gal за методом, здійснене за допомогою плазмід pBHV1HK і описаним в статті [Чоудхарі С.І. (Chowdhury S.I.), pBHV1HF, отриманих від доктора В. Лоуренса (W. Construction and Characterization of an Attenuated Lawrence) (U. Pennsylvania, Філадельфія, США). Bovine Herpesvirus Type 1 (BHV-1) Recombinant Зазначені плазміди включали, відповідно, фрагVirus, 52 Vet. Microbiol. 13-23 (1996)], який включементи Hindlll-K і HindIII-F ДНК BHV-1. Субфрагмент ний до цього опису як посилання. Декілька рекомXhoI/Hindlll (4,4тис.п.н.) плазміди pBHV1HK, який бінантних ізолятів були додатково характеризовані містить gD, gl в повному об'ємі, і частину, в перешляхом блот-гібридизації і імуноблотингу з антиважному варіанті дві третини, амінокінцевої ділянBHV-1 gE-специфічною антипептидною кролячою ки gE-кодуючої області, субклонували на сайтах поліклональною сироваткою. Був використаний XhoI/HindllI плазміди pGEM7Z (pBHVlgE5'). Далі, метод блот-гібридизації [Чоудхарі С.І. (Chowdhury фрагмент HindIII/Bsu36I (1,15тис.п.н.) (з "тупими" S.I.), Construction and Characterization of an (дефосфорилованими) кінцями) pBHV1HF, який Attenuated Bovine Herpesvirus Type 1 (BHV-1) включає карбоксикінцеву третину gE-кодуючої обRecombinant Virus, 52 Vet. Microbiol. 13-23 (1996)], ласті і кодуючу послідовність US9 ORF в повному який включений до цього опису як посилання. Був об'ємі, субклонували на сайтах Hindlll/HincII плазвикористаний метод імуноблотингу [Чоудхарі С.І. міди pBluescript KS (pBHV1gE3'). І, нарешті, для (Chowdhury S.I.), Molecular Basis of Antigenic збирання кодуючої області гена gE в повному Variation Between Glycoprotein С (gC) of Respiratory об'ємі і фланкуючих послідовностей, фрагмент Bovine Herpesvirus 1 (BHV-1) and Neurovirulent (4,4тис.п.н.) векторного сайту Hindlll/Xbal BHV-5, 213 Virology 558-568 (1995)], який включеpBHVlgE5', який включає фрагмент Hindlll/Xhol, ний до цього опису як посилання. клонували на сайтах Hindlll/Xbal плазміди Результати pBHV1gE3'. Отриманий клон отримав позначення Результати аналізу ДНК двох рекомбінантних pBHVlgE5'3'. вірусів, gE 3.1IBR (інвентарний номер АТСС VRДля видалення gE-кодуючої області, ДНК 2367) і gE 3.5IBR , шляхом саузерн-блотpBHVlgE5'3' частково гідролізували AsuII з подагібридизації на призначену делецію і інсерцію -gal льшим повторним гідролізом до завершення з допослідовностей в gE локусі, представлені на Фіг.2. помогою Hindlll. Більш великий фрагмент очищали Відсутність послідовностей фрагмента Asull-HindllI методом імуносорбції в лунках з гелем і лігували із (1тис.п.н.), які кодують перші 372 амінокислоти на фрагментом PstI (4,5тис.п.н.) (з "тупими" (дефосамінокінцевій ділянці зазначеного gE гена і присуфорилованими) кінцями, отриманим за допомогою тність -gal послідовностей в ізолятах gE 3.1IBR і Т4 полімерази) pCMV [отриманої від компанії gE 3.3IBR демонструє, що призначена рекомбіClontech, Palo Alto, штат Каліфорнія, США], до нація в цих ізолятах здійснилася сайтспецифічним складу якої входили -gal послідовності, регульоспособом. Відповідно до зазначеного, у батьківсьвані раннім промотором CMV. Отримана gEкого штаму Cooper дикого типу за допомогою BHVдeлeцiйнa/ -gal-iнcepцiйнa плазміда, pBHV1gEA , 1 gE-специфічної антипептидної кролячої полікломала делецію послідовностей ДНК (1тис.п.н.) BHVнальної сироватки був виявлений gE білок (921, які включають послідовності гена, що кодують 95кДа) BHV-1, який був відсутній у gE-делеційного перші 372 амінокислоти, і інсерцію -gal гена під рекомбінантного вірусу gE 3.1IBR . Отриманий регуляцією CMV промотору. Зазначений -gal ген результат показаний на Фіг.3 і зазначений ізолят фланкований вірусспецифічними послідовностями був використаний для додаткових досліджень. У 3'-5' дирекційного типу (3,32тис.п.н.) (які включадоповнення до цього, gE 3.1IBR може реверсують послідовності генів gD і gl в повному об'ємі і ватися у вірус IBR дикого типу шляхом котрансфеgE-промоторні послідовності) і послідовностями 5'кції плазмідою, яка включає фланкуючі послідов3' дирекційного типу (які включають карбоксикінності і початкову gE-послідовність IBRV дикого цеву третину gE-кодуючої області і послідовності типу. Це по суті реверсує процес котрансфекції, гена US9 в повному об'ємі), які необхідні для ревикористаний для отримання gE 3.1IBR . комбінації з вірусною ДНК. Кінетику експресії -gal в інфікованих вірусом Для отримання gE-делеційного рекомбінантклітинах MDBK визначали гістохімічними засобами ного вірусу IBR, ДНК лінеаризованого pBHV1gE 11 75035 12 через 3год., 6 год., 12год. і 24год. після інфікуванноса і рота, кон'юнктивіт і порушення дихання. Кожний зазначений параметр індивідуально оціня, -gal активність виявляли вже через 3год. і ще нювали в балах і щоденна клінічна бальна оцінка через 24год. після інфікування. Це свідчить про те, для кожного теляти, а також середня щоденна що регульований CMV-IE промотором ген клінічна бальна оцінка для кожної групи обчислюгалактозидази вірусу gE 3.1IBR експресується як валися шляхом додавання всіх бальних оцінок по на ранніх, так і на пізніх етапах. Більш того, незвакожному параметру. У балах оцінювалися такі пажаючи на те, що зазначений химерний ген регулюраметри: ється стороннім HCMV-IE промотором вірусу герВиділення з порожнини носа: норма=0; помірпесу людини і включає невірусні -gal но серозні=1; сильно серозні=2; слабо мікропурупослідовності, а також енхансери, сторонні як для лентні=2; помірно мікропурулентні=3; сильно мік-gal, так і для BHV-1, він регулюється і експресуропурулентні=4. ється як аутентичний ген BHV-1. Таким чином, Поведінка (депресія): відсутність=0; слабка gal є стороннім (відсутнім у BHV-1 в природному ступінь=1; помірна ступінь=2; важка ступінь=3. стані) BHV-1 і HCMV-IE, і його енхансерні послідоГіперемованість/почервоніння слизової оболовності є сторонніми як для BHV-1, так і для -gal, нки порожнини носа=1. які ними регулюються/стимулюються. Покриття виразками слизової оболонки пороДелеція зазначеного gE гена буде служити як жнини носа=2. імунологічний маркер для відрізнення вакциноваКон'юнктивіт=2. них і інфікованих тварин. При антитілогенезі, індуКашель=2. кованому вакцинним штамом, будуть відсутніми Задишка=2. gE-специфічні антитіла, внаслідок чого він буде Щоденна ректальна температура: 39,7відрізнятися від антитілогенезу, індукованого ін39,99°С=1; 40,0-40,5°С=2; 40,6-41,0°С=3; більше фекцією BHV-1 дикого типу, при якому будуть приніж 41,0°С=4. сутні gE-специфічні антитіла. Завдяки цьому у ваРезультати кцинованому стаді можна буде ідентифікувати і У телят групи В, інфікованих батьківським вибракувати інфікованих тварин. Цей серологічний штамом IBRV Cooper (IBRV дикого типу), спостерімаркер важливий для ліквідації BHV-1 в стаді. галися типові симптоми інфекції: депресія, погірЗазначена плазміда (pBHV1gE ), використашення апетиту, виділення з очей і порожнини нона для отримання рекомбінантного вірусу, і отриса, покриття виразками слизової оболонки маний рекомбінантний вірус, gE 3.1IBR , були порожнини носа/бляшки в порожнині носа і кадепоновані в Американській колекції типових кульшель. Наслідком цих клінічних симптомів була тур (АТСС). Зазначеній плазміді (pBHV1gE ) був висока щоденна клінічна бальна оцінка протягом присвоєний інвентарний номер АТСС 203607; задекількох днів, як показано на Фіг.5. Однак у телят, значеному рекомбінантному вірусу (gE 3/1IBR ) інфікованих рекомбінантним вірусом gE 3.1IBR був присвоєний інвентарний номер АТСС VR-2637. (група А), були повністю відсутні виявлювані клініПриклад 2 чні симптоми (що також показано на Фіг.5), і їхня Матеріали і методи поведінка і апетит залишалися в межах норми. На У цьому експерименті визначали патогенність Фіг.6 порівнюються середні ректальні температури вірусу gE 3/HBR на телятах і порівнювали отрителят кожної групи (А і В). Високі ректальні темпемані результати з патогенністю батьківського штаратури в межах від 39,7°С до 40,5°С реєструвалиму IBRV Cooper. Десять шестимісячних телят голся протягом декількох днів у телят, інфікованих штинської породи, вільних від IBRV і вірусу батьківським штамом Cooper (група В). Ректальні вірусної діареї великої рогатої худоби, довільно температури телят, інфікованих gEA3.1IBRß, не розділили на дві групи (А і В) по п'ять голів в кожперевищували 38,9°С (група А). ній. Обидві зазначені групи утримували в окремих Приклад 3 стійлах в однакових умовах. Всі телята перед проМатеріали і методи веденням експеримента були здоровими і до поУ цьому експерименті проводили порівняння чатку експеримента мали серонегативну реакцію. кінетики зростання вірусу gE 3.1IBR в клітинах Кожному теляті в групі А інтраназально вводиMDBK і батьківського штаму вірусу IBR Cooper в ли рекомбінантний gE-делеційний IBRV. Кожному клітинах MDBK. Ряд реплікованих культур клітин теляті в групі В інтраназально вводили батьківсьMDBK роздільно інфікували 5 бляшкотвірними 7 кий штам IBRV Cooper. Інокулят включав 1 10 одиницями (РFU)/клітину рекомбінантного вірусу TCID50 (ІЕ50) відповідного вірусу на теля. ІнокуляgE 3.1IBR або батьківського штаму Cooper. Інфіцію здійснювали шляхом аерозолювання за допоковані культури збирали через послідовні проміжки могою аерозольного інгалятора DEVILBIS 50 мочасу після інфікування і штами вірусу готували для делі компанії Delvis Co., Somerset, штат титрування. Пенсільванія (2мл інокуляту на ніздрю протягом Результати від 30с. до 1хв.). Результати одноетапних експериментів із виІнтенсивні клінічні спостереження за всіма терощуванням представлені на Фіг.4. Криві зростанлятами здійснювалися щодня протягом 14 днів ня вірусу показують, що зростання потомства було після дії вірусу (інокуляції). Щодня реєструвалася однаковим як у gE 3.1IBR , так і у батьківського ректальна температура. Особливу увагу приділяли штаму Cooper. Однак урожай ί рекомбінантного таким параметрам: поведінка (депресія), апетит, вірусу на ранніх стадіях після інфікування був кашель, виділення з очей і порожнини носа, гіпеменшим. ремія або ураження слизової оболонки порожнини 13 75035 14 Приклад 4 рожнини носа представлене на Фіг.7. Виділення Матеріали і методи вірусу gE 3.1IBR було нижчим за виділення BHVУ цьому експерименті виділяли і проводили кі1 (батьківський штам Cooper) дикого типу приблилькісну оцінку вірусу від кожної тварини в групах А зно в 15-550 разів. Тривалість виділення вірусу у і В (з вищенаведеного прикладу 3). У кожну носову телят, інфікованих вірусом gE 3.1IBR , також бупорожнину вводили простий ватний тампон на ла на 2 дні коротшою, ніж тривалість виділення паличці. Тампон притискували до слизової оболовірусу у телят, інфікованих вірусом дикого типу. нки і тричі провертали. Вірус елюювали в 3мл сеПриклад 5 редовища для зразка (мінімальне підтримувальне Матеріали і методи середовище, до складу якого включали 100мкг/мл У цьому прикладі визначали титри нейтралігентаміцину, 3% (в об'ємному відношенні) забуфезуючих антитіл BHV-1 в сироватці, яка відбиралася реного фосфатом фізіологічного розчину, амфоу телят в групах А і В (з вищенаведених прикладів терицину В в дозі 25мкг/мл) протягом 1год. при 3 і 4). У кожного теляти з обох груп А і В через 13 кімнатній температурі. Зразки прояснили центриднів після інфікування відбирали проби крові. Сифугуванням при 1000g протягом 5хв. і зберігали роватковий титр BHV-1 нейтралізуючих антитіл при температурі -70°С. Виділення вірусу на клітивизначали за методом редукції бляшок на 12нах MDBK здійснювали за допомогою 100мкл тамлуночних планшетах за методом, описаним [Чоудпонної суспензії в розведенні 1:10. Віруспозитивні харі С.І. (Chowdhury S.I.) і іншими, Molecular зразки титрували на титраційних мікропланшетах. Biological Characterization of Equine Herpesvirus 1 Готували серійне десятиразове розведення в ку(EHV-1) Isolated From Ruminant Hosts, 11 Virus льтуральному середовищі і по 50мкл кожного розRes. 127-139 (1988)], який включений до цього ведення додавали в кожну з восьми лунок 96опису як посилання. луночного планшета), в яких знаходилося 1,5 105 Результати клітин MDBK. Через 5 днів при температурі 37°С Як вірус дикого типу (батьківський штам планшети за допомогою мікроскопа прочитували Cooper), так і gE-делеційний IBRV (gE 3.1IBR ) на цитопатичну дію (СРЕ). Титри вірусу обчислюіндукували BHV-1 нейтралізуючі антитіла у телят. вали в TCID50 (ІЕ50) за методом [Ріда Л.Дж. (Reed Однак телята, інфіковані вірусом дикого типу, маL.J.) і Менша Г. (Muench Η.), 27 Am. J. Hyg. 493 ли трохи більш високі сироваткові титри нейтралі(1938)], який включений до цього опису як посизуючих антитіл (1:20-1:35 зі середнім титром 1:30), лання. в порівнянні з титрами нейтралізуючих антитіл gEРезультати делеційного вірусу (1:11-1:30 зі середнім титром Кількість вірусу, виділена з носових тампонів, 1:16). показала, що вірус gE 3.1IBR в клітинах епітелію порожнини носа ріс із меншою ефективністю, ніж батьківський штам Cooper. Виділення вірусу з по 15 Комп’ютерна верстка О. Гапоненко 75035 Підписне 16 Тираж 26 прим. Міністерство освіти і науки України Державний департамент інтелектуальної власності, вул. Урицького, 45, м. Київ, МСП, 03680, Україна ДП “Український інститут промислової власності”, вул. Глазунова, 1, м. Київ – 42, 01601
ДивитисяДодаткова інформація
Назва патенту англійськоюAttenuated recombinant herpes virus of cattle of type і (bhv-1), vaccine containing it and a method for the detection of animals injected with recombinant viral vaccine
Назва патенту російськоюАттенуированный рекомбинантный вирус герпеса крупного рогатого скота типа 1 (bhv-1), вакцина, содержащая его, и способ выявления животных, которым была введена рекомбинантная вирусная вакцина
МПК / Мітки
МПК: A61P 31/22, G01N 33/573, G01N 33/53, C12N 7/04, A61P 37/04, C12N 7/01, C12N 15/869, C12N 15/09, C12Q 1/34, A61K 39/245
Мітки: вірусна, виявлення, містить, яким, введена, рогатої, рекомбінантний, була, тварин, спосіб, рекомбінантна, вакцина, герпесу, великої, bhv-1, худоби, типу, атенуйований, вірус
Код посилання
<a href="https://ua.patents.su/8-75035-atenujjovanijj-rekombinantnijj-virus-gerpesu-veliko-rogato-khudobi-tipu-1-bhv-1-vakcina-shho-jjogo-mistit-ta-sposib-viyavlennya-tvarin-yakim-bula-vvedena-rekombinantna-virusna-vakc.html" target="_blank" rel="follow" title="База патентів України">Атенуйований рекомбінантний вірус герпесу великої рогатої худоби типу 1 (bhv-1), вакцина, що його містить, та спосіб виявлення тварин, яким була введена рекомбінантна вірусна вакцина</a>
Вакцина інактивована рекомбінантна проти інфекційного ринотрахеїту великої рогатої худоби

Номер патенту: 49450
Опубліковано: 16.09.2002
Автори: Кассіч Володимир Юрійович, Волосянко Олена Вікторівна
МПК: A61K 39/12
Мітки: вакцина, інактивована, ринотрахеїту, рогатої, худоби, рекомбінантна, великої, інфекційного
Формула / Реферат:
Вакцина інактивована рекомбінантна проти інфекційного ринотрахеїту (ІРТ) великої рогатої худоби, що містить мікроорганізм, який має антигени, ідентичні вірусу ІРТ, яка відрізняється тим, що як вірус використовують рекомбінантний штам Bacillus alvei-413, при цьому вакцинний матеріал вирощують на м'ясо-пептонному бульйоні з додаванням (1 %) глюкози та (5 %) сироватки крові коней.
Вакцина для великої рогатої худоби проти респіраторних патологій, спосіб вакцинації та набір для вакцинації великої рогатої худоби

Номер патенту: 70914
Опубліковано: 15.11.2004
Автори: Бушардон Аннабель, Одонне Жан-Крістоф, Бодю Філіпп, Рівьєр Мішель
МПК: C07K 14/06, A61K 39/155, C12N 15/09, A61K 39/295, C07K 14/18, A61K 39/12, A61P 31/22, C07K 14/135, C07K 14/115, C12N 15/45, C12P 21/02, A61K 39/245, A61P 31/20
Мітки: рогатої, патологій, набір, вакцина, спосіб, великої, респіраторних, худоби, вакцинації
Формула / Реферат:
1. Вакцина для великої рогатої худоби, яка включає плазміду, що містить одну або більше нуклеїнових кислот, що кодує F та/або G білок респіраторного синцитіального вірусу великої рогатої худоби.2. Вакцина за п. 1, яка відрізняється тим, що вказана плазміда містить нуклеїнову кислоту, яка кодує обидва гени F та G.3. Вакцина за п. 1, яка відрізняється тим, що вказана плазміда містить нуклеїнову кислоту, яка кодує F ген, і при...
Спосіб виявлення захворювання на лейкоз у великої рогатої худоби

Номер патенту: 44416
Опубліковано: 15.02.2002
Автори: Грицик Олександр Борисович, Бусол Володимир Олександрович, Мандигра Микола Станіславович, Рудь Олег Григорович
МПК: G01N 33/559
Мітки: виявлення, рогатої, спосіб, лейкоз, великої, захворювання, худоби
Формула / Реферат:
Спосіб виявлення захворювання на лейкоз у великої рогатої худоби, що полягає у проведенні на основі реакції імунодифузії (РІД) з лейкозним антигеном серологічних досліджень всіх тварин починаючи з 4 - 6 місячного віку, який відрізняється тим, що серологічні дослідження проводяться через кожні 10 днів, що забезпечує повну ізоляцію та швидке вилучення джерела збудника інфекції по всіх вікових групах тварин.
Бівалентна інактивована концентрат-вакцина проти інфекційного ринотрахеїту та вірусної діареї великої рогатої худоби

Номер патенту: 4626
Опубліковано: 17.01.2005
Автори: Стегній Борис Тимофійович, Кучерявенко Вікторія Вікторівна, Кучерявенко Роман Олексійович, Стеценко Володимир Іванович, Білокінь Віктор Степанович
МПК: A61K 39/265
Мітки: рогатої, концентрат-вакцина, вірусної, ринотрахеїту, інактивована, інфекційного, бівалентна, худоби, великої, діареї
Формула / Реферат:
Вакцина проти респіраторних вірусних захворювань, що містить компонент вірусу ІРТ, ад'ювант та інактиватор, яка відрізняється тим, що використовують штам "Молдавський" вірусу ІРТ, додають компонент вірусу діареї штам "ВК-1", використовують як ад'ювант масло ISA-70, a як інактиватор використовують формалін.
Штам перещеплюваних клітин нирки вівці flk-sbbl, який продукує вірус лейкозу великої рогатої худоби

Номер патенту: 55921
Опубліковано: 15.04.2003
Автори: Берус Павло Тихонович, Стеценко Володимир Іванович, Стегній Борис Тимофійович, Лаврик Олексій Анатолійович, Білокінь Віктор Степанович
Мітки: лейкозу, нирки, худоби, перещеплюваних, продукує, штам, вівці, рогатої, великої, flk-sbbl, клітин, вірус
Формула / Реферат:
Штам перещеплюваних клітин нирки вівці FLK-SBBL, який продукує вірус лейкозу великої рогатої худоби, депонований та зберігається за номером 18 у колекції клітинних культур Інституту експериментальної та клінічної ветеринарної медицини УААН, м. Харків, Україна.
Попередній патент: Трициклічні інгібітори полімерази полі(адф-рибози) для лікування раку та інгібування нейротоксичності, їх застосування та фармацевтична композиція на їх основі
Наступний патент: Спосіб радіоактивного мічення кон’югованого з комплексоном антитіла або фрагмента антитіла радіоактивним ізотопом ітрій-90 та комплект для здійснення способу
Випадковий патент: Фрез для розбурювання бурильних колон