Ліпополісахариди з escherichia coli
Номер патенту: 75068
Опубліковано: 15.03.2006
Автори: Пропперт Ханс, Малінка Йюрген, Улмер Артур, Церінгер Ульріх, Шульц Йюрген, Зонненборн Ульріх, Рітшель Ернст Теодор







Формула / Реферат
1. Ліпополісахарид (ЛПС), який відрізняється тим, що належить до ліпополісахаридів бактерій Е. соlі штаму DSM 6601, а зазначений ліпополісахарид включає 8 фосфатних залишків та 0,5 моль Etn-P, що відповідає структурній формулі, зображеній на фіг.7.
2. ЛПС за п. 1, який відрізняється тим, що містить 8 фосфатних залишків на 1 молекулу ЛПС.
3. ЛПС за пп. 1 чи 2, який відрізняється тим, що містить 0,5 моль Etn-P на 1 молекулу ЛПС.
4. Спосіб одержання ЛПС за будь-яким з пп. 1-3, який відрізняється тим, що вимиту і висушену ліпополісахаридну бактеріальну масу Е. соlі штаму DSM 6601 піддавали відомим способом екстракції в суміші фенол-вода й отриманий у такий спосіб екстракт обробляли РНКазою, ДНКазою та протеїназою К.
Текст
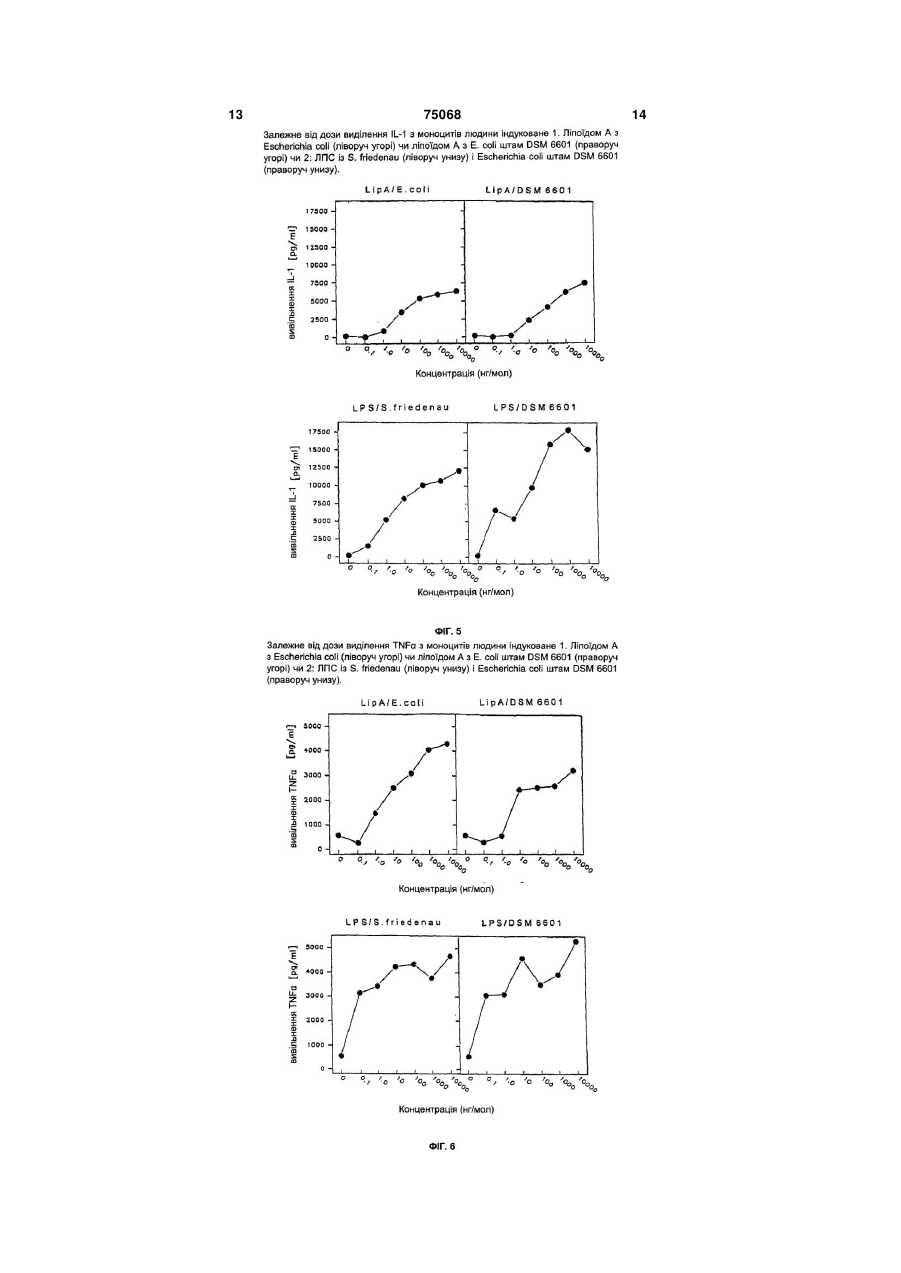
1. Ліпополісахарид (ЛПС), який відрізняється тим, що належить до ліпополісахаридів бактерій Е. соlі штаму DSM 6601, а зазначений ліпополісахарид включає 8 фосфатних залишків та 0,5 моль Etn-P, що відповідає структурній формулі, зображеній на фіг.7. 2. ЛПС за п. 1, який відрізняється тим, що містить 8 фосфатних залишків на 1 молекулу ЛПС. 3. ЛПС за пп. 1 чи 2, який відрізняється тим, що містить 0,5 моль Etn-P на 1 молекулу ЛПС. 4. Спосіб одержання ЛПС за будь-яким з пп. 1-3, який відрізняється тим, що вимиту і висушену ліпополісахаридну бактеріальну масу Е. соlі штаму DSM 6601 піддавали відомим способом екстракції в суміші фенол-вода й отриманий у такий спосіб екстракт обробляли РНКазою, ДНКазою та протеїназою К. Винахід стосується нових ліпополісахаридів з Е. соli. Під ендотоксинами розуміють бактеріальні компоненти структури, що на відміну від екзотоксинів не виділяються живими бактеріями, а звільняються переважно за допомогою аутоліза. У відношенні так званих класичних ендотоксинів мова йде про стійкі стосовно високих температур ліпополісахариди, що надалі називаються ЛПС, із зовнішніх клітинних мембран грам-негативних бактерій. ЛПС складаються з так названого ліпоїда А, ядерного олігосахарида, а також специфічного Оланцюга, причому ліпоїд А відповідальний за токсичну дію ЛПС. Ендотоксини стимулюють у макроорганізмі виробництво медіаторів імунної системи, таких як інтерлеукін 1, скорочено IL-1, і пухлинний некрофактор, скорочено TNF . За складом ендотоксинів ентеробактерій і зокрема Е. соlі уже проведений цілий ряд досліджень, причому встановлено, що S/R-мутанти як правило містять тільки повторюваність, тобто "елемент повторюваності" їхнього О-ланцюга (Фіг.1). Вихо дять з того, що в цих випадках ген, що кодований для полімеризованого ензима О-ланцюга, дефектний і тому на ядерний олігосахарид переноситься тільки один "елемент повторюваності". ЛПСструктури аналогічного типу, але іншої структури часто виявляються також у людських патогенних мікробах, таких як Neisseria, Vibrio, Campylobacter, Helicobacter і т.п. Ці мікроби володіють ЛПС, що дозволяє їм, завдяки молекулярної мімікрії, і, зокрема, завдяки наявності сіалинової кислоти і олігосахаридів, що містять сіалінову кислоту і нагадують глікопротеїни і гліколіпіди ссавців, уникати імунного захисту хазяїна. Для Е. coli DSM 6601 установлений O6-серотип. Цю структуру досліджували і опублікували [П.Е. Янссон і ін., Carbohydr. Res. 131 (1984) с. 277-283]. Структура відповідає формулі, представленій на Фіг.2. Ліпоїд А соlі-бактерій також досліджували різні групи досліджувачів, причому було встановлено, що структура ліпоїда А як правило існує в гексаациловій формі і збігається для всіх серотипів Е. соlі (Фіг.3). Структура гексаацилового зв'язку опублікована в 1984 p. [T. Рітшелем і ін., Structure and (19) UA (11) 75068 (13) (21) 2002108256 (22) 20.03.2001 (24) 15.03.2006 (86) PCT/EP01/03153, 20.03.2001 (31) 100 13 539.0 (32) 20.03.2000 (33) DE (46) 15.03.2006, Бюл. № 3, 2006 р. (72) Пропперт Ханс , DE, Малінка Йюрген , DE, Шульц Йюрген , DE, Зонненборн Ульріх , DE, Церінгер Ульріх , DE, Улмер Артур , DE, Рітшель Ернст Теодор, DE (73) ФАРМА-ЦЕНТРАЛЕ ГМБХ, DE (56) P.-E. JANSSON ET AL.: "Structural studies of the Escherichia coli O-antigen 6". Carbohydr Res. 1984 Aug 15;131(2):277-83. J.A. YETHON ET AL.: "Involvement of waaY, waaQ, and waaP in the modification of Escherichia coli lipopolysaccharide and their role in the formation of a stable outer membrane". THE JOURNAL OF C2 1 3 75068 4 conformation of the lipid A component of ті" ("repeating unit") серотипа О6, приєднана до lipopolysaccharides; Handbook of Endotoxins ядерного олігосахариду через -глікозиди й у та(Proctor, R., ed.). том 1, Chemistry of Endotoxin (E. кий спосіб відрізняється іншим з'єднанням, ніж Th. Rietsche), ed.). Ельзевір, Амстердам (1984), с. усередині О-ланцюга ( -глікозидне). Ядерний олі187-220], і відповідає зображенню на Фіг.3. госахарид має R1-структуру, це результат хімічноСпецифічні О-ланцюги і ліпоїд А з'єднані один го аналізу, підтверджений серологичними досліз одним через ядерний олігосахарид. В даний час дженнями зі специфічними R1-антитілами. відомі п'ять різних ядерних олігосахаридів у Е. соlі; Компонента ліпоїда А має характерну для Е. соlі ми вказуємо на [О. Хольста й ін., Chemical ліпоїда А визначену хімічну структуру. structure of the core region of lipopolysaccharides, y: ЛПС Е. соlі штаму DSM 6601 виявляє дивну Bacteria! Endotoxic Lipopolysaccharides, том. 1, Мооднорідність. Була встановлена тільки гетерогенррісон Д.Ц. і Ріан, Дж. Л. і ін., Бока Ратон, Флориність у відношенні заступників фосфатів (РР і Pда, США (1992) с. 135-170] (Фіг. 4). Etn vs. Ρ і Ρ), що у цій формі описані вперше. За При дослідженнях ЛПС Е. соlі штам DSM 6601 допомогою комплексних NMR-аналізів однозначно було встановлено, що ліпоїд А у своєму складі визначений P-Etn-заступник у ядерному олігосахагексаацилової форми також відповідає ліпоїду А, риді, зокрема в R1-ядерному олігосахариді в позиописаному для Е. соlі в іншому місці. ції 2 другої гептози (Hep"). Результати досліджень, що стосуються вивіПовна структура ЛПС штаму DSM 6601 предльнення IL-1 і TNF в моноцитах людини підтверставлена на Фіг.7. джують, що ліпоїд А виявляє однакову активність і Надалі винахід більш докладно пояснюється тому з максимальною імовірністю відповідає відоза допомогою прикладів. мій структурі Е. соlі ліпоїда А (Фіг.5, Фіг.6). Це приПриклад 1 пущення підтверджується результатами хімічних Виготовлення ЛПС аналізів. ЛПС одержували з промитої і висушеної бакСтруктура специфічного О-антигену Е. соlі теріальної маси методом модифікованої фенольDSM 6601 приголомшує тим фактом, що свідчить но-водяної екстракції; оскільки вказують на [О. про те, що в ланцюзі очевидно регулярно існує Вестфаль і ін., Бактеріальні ліпополісахариди, тільки один єдиний "елемент повторюваності" Extraction with Phenol-Water and Further (Фіг.1), що дозволяє зробити висновок про те, що у Applications of the Procedure, Meth. Carbohydr. випадку штаму DSM 6601 йдеться про S/RChem., Том. V (1965) с. 83-91]. мутантів, що є надзвичайно незвичайним для 47г висушених заморожуванням бактерій, що людської субстанції. Структура цього "елемента попередньо двічі промивали дистильованою воповторюваності" відповідає однак, як показує седою, екстрагували за методом Вестфаля і Джанна рологічний аналіз, основному зразку О-ланцюга Е. відповідно модифікованому розпорядженню. Мосоlі О6. дифікація полягала в наступній обробці ферменту Регіон ядра в штамі DSM 6601 відповідає уже (DNA, RNA, Proteinase К) водяного екстракту, що відомій структурі-R1. Структурні особливості поляслужить для того, щоб видалити можливі сторонні гають, зрозуміло, у тім, що аналітично встановлепротеїни і складові частини DNA/RNA. но 8 фосфатних залишків на молекулу ЛПС, приДля цього водяну фазу (близько 1,2л) при кімчому на ліпоїд А приходиться як правило тільки 2 натній температурі змішують з 20мг RNA фосфатних залишки. Крім того виявився нестехіо(Ribonuclease A, bovine pancreas, Sigma) і 20мг метричний зміст пірофосфо-етаноламіна. DNA (Dnase 1, bovine pancreas, II ступеня, Sigma) і Підсумовуючи, можна тому установити, що суміш при кімнатній температурі помішують протяЛПС штаму DSM 6601 значно відрізняється від гом 30 годин. Потім додають 20мг Proteinase К дотепер відомого ЛПС із Е. Соlі, зокрема фосфо(Tritirachium album, Борінгер, Маннгейм) і помішурилованою кількістю цукру ядра і ступенем поліють протягом наступних 12 годин. Потім протягом меризації О-ланцюга. Ліпоїд А структурно і біологі24 годин при температурі 4 С тричі проводять діачно відповідає звичайному для Е. соlі типу, що ліз з 15 літрами дистильованої води і за допомопідкреслює роль цієї субстанції у відношенні біологою цього висушують способом заморожування. гічної ефективності. Описаний ЛПС призначений Оброблений ензимом екстракт знову поміщають у не тільки для ідентифікації несущого його штаму дистильовану воду, так що кінцева концентрація соlі, але і додає йому також знижену патогенність, досягає 50мг/мол. Цю суспензію в холодному призберігаючи його імунодопустимий ефект. Той міщенні тричі піддають ультрацентрифугуванню факт, що О-ланцюг з'єднаний через -глікозиди (155,000 г, 4 С, 4 години). Осад (ЛПС) поміщазамість -глікозидів, зміг стати вперше однозначно ють у 150мол. дистильованої води і знову проводоведеним на прикладі S/R-мутантів DSM 6601 дять діаліз протягом трьох днів водою, а за доподля Е. соlі. могою цього ліофілізують (вихід ЛПС: 1,45г, 3,1% У відношенні ліпополісахарида (ЛПС) Е. coli м/м). DSM 6601 йдеться про нову однорідно-грубу струПриклад 2 ктуру (smooth-rough) (S/R), що з одного боку склаАналіз ЛПС із Е. соlі штам DSM 6601 дається з відомих дотепер структур (специфічного Гексозамін (HexN) (тут означає Глюкозамін + О-ланцюга, ядерного олігосахарида і ліпоїда А), а Галактозамін, GlcN+GalN) визначали за допомоз другого боку вперше і цілком структурно охаракгою модифікованого тесту Моргана-Ельсона теризована в представленій тут комплексній формі [Штромінгер, Дж.Л., Парк, Дж.Т. Томсон, P.E. Дж. (Фіг.7). Специфічний О-ланцюг, що складається Biol. Chem. 234, 3263-3268 (у 1959)], але альтертільки з одного єдиного "елемента повторюваноснативно також за допомогою HPLC (PICO-TAG, 5 75068 6 Waters). На противагу тесту Моргана-Ельсона при (ν/ν/ν) очищали далі. Окремі фракції олігосахарида цьому методі аналізу можна роздільно визначати і (Група А, В, С і D) аналізували за допомогою GCкількісно ідентифікувати не тільки GlcN і GalN, але MS і NMR-спектроскопії. Головну фракцію (Група і крім того можна також ще і паралельно встановА, #28-41; 49,05мг), що містить як компоненти цуклювати присутність GlcN-фосфата, 2-етаноламіна ру О-ланцюга (Man, GalNac), так також і компонен(Etn) і 2-етаноламінфосфата (Etn-P), що часто є ти цукру ядерного олігосахарида (Hep, Kdo), очиприсутнім у ЛПС. Газо-рідинну хроматографію щали далі. Iнші фракції містили моносахариди, (GC) проводили на газохроматографі типу Varian більш детально не досліджувані утворення Kdo 3700 GC чи Hewlett Packard (HP 5890 серія II) на (ангідро- і лактоні) і нарешті сіль. Головна фракція поділу на TSK знайшла як в аналізі GC-MS так капілярній колоні (fused-silica SPB-5 , 30м, також і в аналізі NMR усі компоненти ядерного Supelco). Комбіновану газо-рідинну хроматограолігосахарида (Kdo, Gal, Hep) і О-ланцюга (man, фію/масспектрометрію (GC-MS) проводили на маGalNAc) і тому оброблялася далі. сспектрометрі (HP модель 5989), що був оснащеДля цього насамперед перевіряли, чи придатний капілярною колоною НР-1 (30м, Hewlett на аналітична аніонообмінна хроматографія висоPackard). GC- і відповідно GC-MS-аналізи застосокого тиску ("high pressure anion exchange вували для визначення нейтральних цукрів (GIC, chromatography, HPAEC"), щоб очищати олігосаGal, Hep, Man) у якості їх алдитолацетатів [Савархариди до стану однорідності. Для цього застосодекер, Дж.С, Слонекер, Дж.Х. Джианс, А. Anal. вували специфічну методику HPLC для аналізу Chem. 37, 1602-1604 (у 1967)], а також для визнакомплексних структур цукру (DIONEX-система) з чення і квантифіціювання жирних кислот у якості їх похідних метилових ефірів жирних кислот за доаналітичною колоною CarboPac PA1; (4,6мм помогою інтенсивного метанолізу (2М HCl/MeOH, 250мм) і лінійним соляним градієнтом (5 хвилин при 0, потім протягом 50 хвилин до 0,5Μ NaOAc) 120 C, 16 годин.) [Волленвебер, Х.-В. і Рітшель, при нормі плинності 1мол/хв. Элюат детектували Е.Т., Аналіз ліпополісахаридів (ліпід А) жирних за допомогою амперометричного детектора (PAD) кислот, Дж. Мікробіологічні методи 11, (1990) 195до еквівалентів відновлення (молекули цукру). У 211] і екстракції хлороформом. При обох GCтакий спосіб одержали чотири фракції олігосахаметодах аналізу процес починали при 150 С (ізоридів, які за допомогою напівпрепаративної метотермічно на 3 хвилини) і за допомогою лінійних дики НРАЕС очищали далі аналогічним способом. температурних градієнтів підвищували від 5 С/хв. Напівпрепаративну методику НРАЕС застосодо 320 С. Фосфат визначали за методом Lowry і вували за допомогою колони CarboPac PA1 [(9мм ін. [Lowry, O.H., Roberts, N.R., Leiner, Κ.Υ., Wu, 250мм) Dionex система] з однаковим сольовим M.KL. Farr, A.L, Дж. Віоl. Chem. 207, 1-17 (1954)], а градієнтом як при аналітичній методиці НРАЕС (5 2-кето-3-дезокси-D-манно-октулозонову кислоту хвилин при 0, потім протягом 50 хвилин до 0,5Μ (Kdo) за допомогою тесту на тіобарбітурову кислоNaOAc) і при нормі плинності 4мол/хв. Накладення ту [Waravdekar, V.C. Saslaw, L.D. J. Biol. Chem. олігосахарида (42мг; група А з колони TSK) на на234, 1945-1950 (1959)]. півпрепаративної НРАЕС відбувалося в два анаОдержання й очищення вільного ліпоїда А і логічних ходи НРАЕС. Елюати по фракціях збираядерного олігосахарида ли протягом однієї хвилини кожну, і кожний окремо ЛПС (250,8мг) суспендували у 25 мол 0,1Μ досліджували за допомогою аналітичної НРАЕС. У NaOAc/HOAc (pH 4,4) і при температурі 100 С протакий спосіб за допомогою напівпрепаративної тягом 1 години піддавали м'якому кислотному гідНРАЕС одержували дві головних фракції (фракція ролізу. Потім ліпофільну частину (ліпоїд А) тричі I, період утримання tR~12 хвилин і фракція II, tR~15 екстрагують з гідролізата з 25 мол хлороформу хвилин). Обидві фракції НРАЕС перед аналізом (вихід 23,2мг). Ліпоїд А з органічної фази очищуMALDI-TOF-MS і NMR опрісняли за допомогою вали далі за допомогою препаративної багатошаколони G-10 (2,5 120см) (вихід: фракція I рової хроматографії (PSC) (2мм PSC силікагель 60 4,68мг; фракція II - 4,39мг). пластин, Е. Мерк, Дармштадт), причому процес Період протікання процесу матричнохроматографування проводили із сумішшю хлодопоміжної лазерної десорбції/ іонізації при массроформ-метанол-вода у співвідношенні 100:75:15 пектрометрії (ν/ν/ν), а проявлення здійснювали шляхом зануПеріод протікання процесу матричнорення в дистильовану воду. У такий спосіб одердопоміжної лазерної десорбції/ іонізації при массжували 6 фракцій, і з головної з цих фракцій пектрометрії знімали за допомогою рефлексного (Rf 0,4) одержували очищений дифосфориловаспектрометра часу протікання процесу Брукера ний гексаацил-ліпоїд А (DPHLA-Еc6601). Очищений (Методи аналізу Брукера-Францена, Бремен) виDPHLA-Еc6601 (вихід 2,06мг) розчиняли у суміші нятково в лінійній конфігурації і у негативному рехлороформ-метанол у співвідношенні 8:2 (ν/ν) і жимі при потенціалі, що прискорює, 20квт і "запізперед MALDI-TOF-MS обробляли іонообмінниками нілої екстракції іона". Проби розчиняли спочатку в (Амберліт IRA 120, Н+-форма). Аліквот (250мг) хлороформі (Ліпоїд А) чи в дистильованій воді очищеного DPHLA-Еc6601 застосовували для екс(фракції-олігосахарида) при концентрації 10 г/ л, периментів з біології. а 2 л аліквота з цих проб розчиняли з 2 л матриВодяну фазу екстракції хлороформу висушучного розчину, що складався з 0,5Μ 2,4,6ють способом заморожування (вихід: 272мг), а тригідроксіацетофенона (Альдрих, Штайнхайм) у олігосахарид за допомогою колони-TSK [3,5 метанолі. Аліквот (0,5 л) цієї суміші поміщали на 90см, TSK HW-40(S), Ε. Мерк] у суміші піридинметалевий тримач і сушили феном. оцтова кислота-вода у співвідношенні 8:20:2000 NMR Спектроскопія 7 75068 8 Одномірні (1D) 1Н і 31P-NMR- j двомірні (2D) ням західних плям (Western-Blots), що розробляли NMR-спектри знімали за допомогою спектрометра за допомогою трьох різних антитіл. Брукера Avance DRX-600 (Брукер, Райнштеттен), а 1. Поліклональну анти-O6-антисиворотку (кроспектри 13С NMR за допомогою спектрометра Брулики) виготовляли за допомогою Е. соlі штам DSM 6601 (серотип О6:К5:Н1) в інституті гігієни в Гамкера АМХ-360 при температурі 300 К у 2H2О. Пебурзі (професор Бокемюль). ред кожним виміром проби двічі висушували спо2. Поліклональну анти-Е. соlі RI-антисиворотку собом заморожування дейтерированою водою (кролики, внутрішнє позначення: K299/d58) одер(2H2O). жували шляхом імунізації мутантами шорсткуватої Як зовнішній сигнал застосовували ацетон ( н форми бактеріальних колоній, котрі містять ядро 2,225 ppm, с 31/45 ppm) чи 85% Н3РО4 ( p 0 ppm). R1 (Анти-R1). Стандарт програмного забезпечення Брукера 3. Застосовували моноклональне антитіло (XWINNMR 1,3) використовували для сканування (WN1-222-5, внутрішнє позначення F 167), що шиданих-NMR. Періоди змішування для TOCSY (зароко перехресно реагує проти всіх Е. соlі ядерних гальна спектроскопія, що корелює) чи NOESY олігосахаридів, починаючи з мінімальної структури (Ядерна понадретельна посилена спектроскопія) (>Rd). складали 100 чи 500мс. Серологічні аналізи Серологічні аналізи проводили з застосуванТаблиця Аналіз компонента von E. coli ЛПС екстрагують зі штаму DSM6601 Компоненти Вуглеводи GlcNb GaIN HexNc Kdo Man Gal Glc L, D-Hep Полярні головні групи P Etn-P Жирні кислоти 12:0 14:0 14:0 (З-ОН) 16:0 а Кількість компонентів nмоль/мг (моль/ЛПС) 1. Аналіз 2. Аналіз 283(1,8) n.b. 139(0,9) n.b. 591 (3,8) 589 (2,9) 248 (1,6) 242 (1,2) 321 (2,1) 383 (1,9) 474 (3,0) 557 (2,8) 1069 (6,9) 1291 (6,4) 566 (3,6) 442 (2,2) 1188 (7,6) 85 (0,5) 1146 (5,7) n.b. 130 (0,8) 156 (1,0) 460 (3,0) Сліди 162 (0,8) 201 (1,0) 504 (2,5) Сліди а Молярне співвідношення окремих компонентів (у дужках) через присутність GalNAc і GlcNAc у Оланцюзі нормували до значення миристинової кислоти (14:0) (1,0моль 14:0/моль ЛПС). b GlcN визначають за допомогою аналізатора амінової кислоти. Значення одержують із суми GlcN і GlcN-6P. c HexN фотометрично визначають за методикою Моргана-Ельсона. n.b. не визначено. Отримані відповідно описаній методиці препарати-ЛПС разом з порівнюваними ЛПС піддавали електрофорорезу в середовищі гелю поліакриламіду (Фіг.1). Для проведення аналізу SDS-PAGEЛПС працювали з 16% гелем поліакриламіду [U.K., Laemmli, Cleavage of structural proteins during assembly of head of bacteriophage T4, Nature, 227, 680-685 (1970)]. Смуги ЛПС офарблювали за методом чуттєвої лужної методики фарбування сріблом [Ц.М. Тсаї і Фраш, Ц.Ф., чуттєвий срібний барвник для детектування ліпополісахаридів у гелі поліакриламіду, Anal. Biochem., 119, 1982,115119]. Результат досліджень представлений на Фіг.1. Приклад 3 Біологічна активність a) IL-1-активність IL-1-активність визначають за допомогою MNC-аналізу проліферації у виступаючій частині культури. Моноцити людини (MNC) виділяють з периферичної крові добровільних донорів (8 105 MNC/200 мол), переносять у склянку й одночасно змішують із субстанцією для тестування. Для тестування біологічної активності в скляній судині клітки спочатку стимулюють за допомогою ЛПС (10нг/мол). Після інкубаційного періоду протягом 8 годин досліджують 150мол виступаючої частини культури на появу цитокинеза. IL-1-активність визначають за допомогою аналізу проліферації фібробластів у виступаючій частині культури. Необхідні для цього фібробласти одержували з крайньої плоті людини. Проліферацію цих фібробластів підвищували за допомогою IL-1. Шляхом порівняння кривої ефективності дози виступаючої 9 75068 10 частини культури з кривої стандарту в аналізі проб позитивний контроль служить відомий ендотоксивизначають біологічну активність у виступаючій чно активний ЛПС із Salmonella friedenau. Резульчастині культури. ЛПС із відомого ендотоксично тати графічно представлені на Фіг.6. активного штаму бактерій (Salmonella friedenau) Результати показують, що у відношенні викислужить за базову основу (позитивний контроль) і дів секрету в IL-1 і TNF не встановлено ніяких тому показаний на Фіг.5. значних розходжень між ЛПС із Salmonella b) TNF -активність friedenau, що служить як стандарт і позитивного контролю, і ЛПС зі штаму DSM 6601 (Фіг.5 і 6, знімTNF -активність у виступаючій частині культуки внизу). Це підтверджується також тим, що ліпори визначають в аналізі цитотоксичності за допоїд А штаму DSM 6601 майже конгруентно активний могою чуттєвих до TNF ліній клітки L929. Поріву порівнянні з ліпоїдом А високого очищення з Е. няння кривої ефективності дози виступаючої соlі (Фіг.5 і 6, знімки вгорі). частини культури з кривої стандарту в аналізі проб дозволяє визначити TNF-активність. Тут також як 11 75068 12 13 75068 14 15 Комп’ютерна верстка А. Крижанівський 75068 Підписне 16 Тираж 26 прим. Міністерство освіти і науки України Державний департамент інтелектуальної власності, вул. Урицького, 45, м. Київ, МСП, 03680, Україна ДП “Український інститут промислової власності”, вул. Глазунова, 1, м. Київ – 42, 01601
ДивитисяДодаткова інформація
Назва патенту англійськоюLypopolysaccharides of escherichia coli
Автори англійськоюTseringer Ulrikh
Назва патенту російськоюЛипополисахариды из escherichia coli
Автори російськоюЦерингер Ульрих
МПК / Мітки
МПК: C08B 37/00, C12P 19/04, A61P 35/00, C12P 19/26, A61K 31/715
Мітки: ліпополісахариди, escherichia
Код посилання
<a href="https://ua.patents.su/8-75068-lipopolisakharidi-z-escherichia-coli.html" target="_blank" rel="follow" title="База патентів України">Ліпополісахариди з escherichia coli</a>
Вакцинний штам escherichia coli “з-12/150″

Номер патенту: 67040
Опубліковано: 15.06.2004
Автори: Дементьєва Світлана Авксентіївна, Риженко Василь Петрович, Кравцова Оксана Леонідівна, Галка Ігор Васильович, Каменчук Павло Петрович, Акименко Лариса Іванівна
МПК: A61K 39/108
Мітки: вакцинний, escherichia, штам
Формула / Реферат:
Штам Escherichia coli, що пропонується для виробництва асоційованих інактивованих вакцин проти ешеріхіозів, сальмонельозів, анаеробної ентеротоксемії та проти маститів і пневмоентеритів тварин, а також для виготовлення антигену для імунологічних реакцій, має добре виражені морфологічні, культуральні, біохімічні, патогенні властивості та імуногенні властивості і задепонований в офіційній колекції ДНКІБШМ під номером 150.
Вакцинний штам escherichia coli “м-131/152″

Номер патенту: 65820
Опубліковано: 15.04.2004
Автори: Мілько Любов Степанівна, Риженко Галина Федорівна, Бондар Тетяна Олександрівна, Бєлік Світлана Миколаївна, Теплюк Ніна Анатоліївна, Кравцова Оксана Леонідівна
МПК: A61K 39/108
Мітки: escherichia, вакцинний, штам
Формула / Реферат:
Штам Escherichia coli "M-131/152", який пропонується для виробництва полівалентної вакцини "Колісан" (проти ешерихіозів тварин), асоційованих інактивованих вакцин "Вельшікол" (проти ешерихіозів, набрякової хвороби та анаеробної ентеротоксимії), "Вельшіколісальм" (проти ешерихіозів, набрякової хвороби , анаеробної ентеротоксимії та сальмонельозу), "Сердосан" (проти ешерихіозу, сальмонельозу,...
Вакцинний штам escherichia coli б-375/151 для виготовлення вакцин проти ешерихіозів сільськогосподарських тварин
Номер патенту: 65108
Опубліковано: 15.03.2004
Автори: Кучерявенко Олексій Олександрович, Мілько Любов Степанівна, Риженко Василь Петрович, Дементьєва Світлана Авксентіївна, Ображей Анатолій Федорович, Марченко Олександра Миколаївна, Каменчук Павло Петрович
МПК: A61K 39/00
Мітки: виготовлення, вакцинний, тварин, ешерихіозів, сільськогосподарських, escherichia, штам, вакцин
Формула / Реферат:
Штам Escherichia coli Б-375/151 для виробництва асоційованих інактивованих вакцин проти колібактеріозів та ешерихіозів сільськогосподарських тварин задепонований в колекції патогенних мікроорганізмів Державного науково-контрольного інституту біотехнології і штамів мікроорганізмів під №151.
Штам escherichia coli о157, продукуючий шиготоксин другого типу (stх2)

Номер патенту: 68323
Опубліковано: 15.07.2004
Автори: Волинець Леонід Кузьмич, Мазур Тетяна Василівна, Зоценко Людмила Володимирівна, Тарасюк Тетяна Іванівна, Поперечна Світлана Григорівна, Олійник Людмила Вікторівна
Мітки: о157, шиготоксин, продукуючий, stх2, escherichia, другого, типу, штам
Формула / Реферат:
Штам Escherichia coli 0157, продукуючий шиготоксин другого типу (Stx2), який задепонований в Державному науково-контрольному інституті біотехнології і штамів мікроорганізмів, реєстраційний № 264.
Вакцинний штам escherichia coli “р-165/153″, який використовується для виробництва вакцини проти ешерихіозів тварин

Номер патенту: 65811
Опубліковано: 15.04.2004
Автори: Кравцова Оксана Леонідівна, Мілько Любов Степанівна, Риженко Ірина Володимирівна, Галка Ігор Васильович, Каменчук Павло Петрович, Бондар Тетяна Олександрівна
МПК: A61K 39/108
Мітки: ешерихіозів, вакцини, тварин, штам, використовується, виробництва, вакцинний, escherichia
Формула / Реферат:
Штам Escherichia coli "P-165/153", який задепонований в офіційній колекції штамів мікроорганізмів ДНКІБШМ під номером 153, що пропонується для виробництва полівалентних та асоційованих інактивованих вакцин "Колісан", "Вельшіколісальм", "Сердосан", "Пневмомастисан", ''Поліавісан" проти колібактеріозів великої рогатої худоби, свиней та птиці і характеризується стабільними морфологічними,...
Попередній патент: Дифенілові етери, фармацевтична композиція, що їх містить, їх застосування, спосіб лікування (варіанти), спосіб їх одержання і спосіб одержання проміжних сполук
Наступний патент: Пристрій для контролю протикорозійного захисту трубопроводів
Випадковий патент: Спосіб діагностики систолічної дисфункції правого шлуночка серця