Спосіб детекції нуклеїновокислотної послідовності-мішені в аналізі із розщепленням та подовженням рто







Формула / Реферат
1. Спосіб детекції нуклеїновокислотної послідовності-мішені з ДНК або суміші нуклеїнових кислот в аналізі з РТОСЕ (розщепленням та подовженням РТО) в рідкій фазі, який включає:
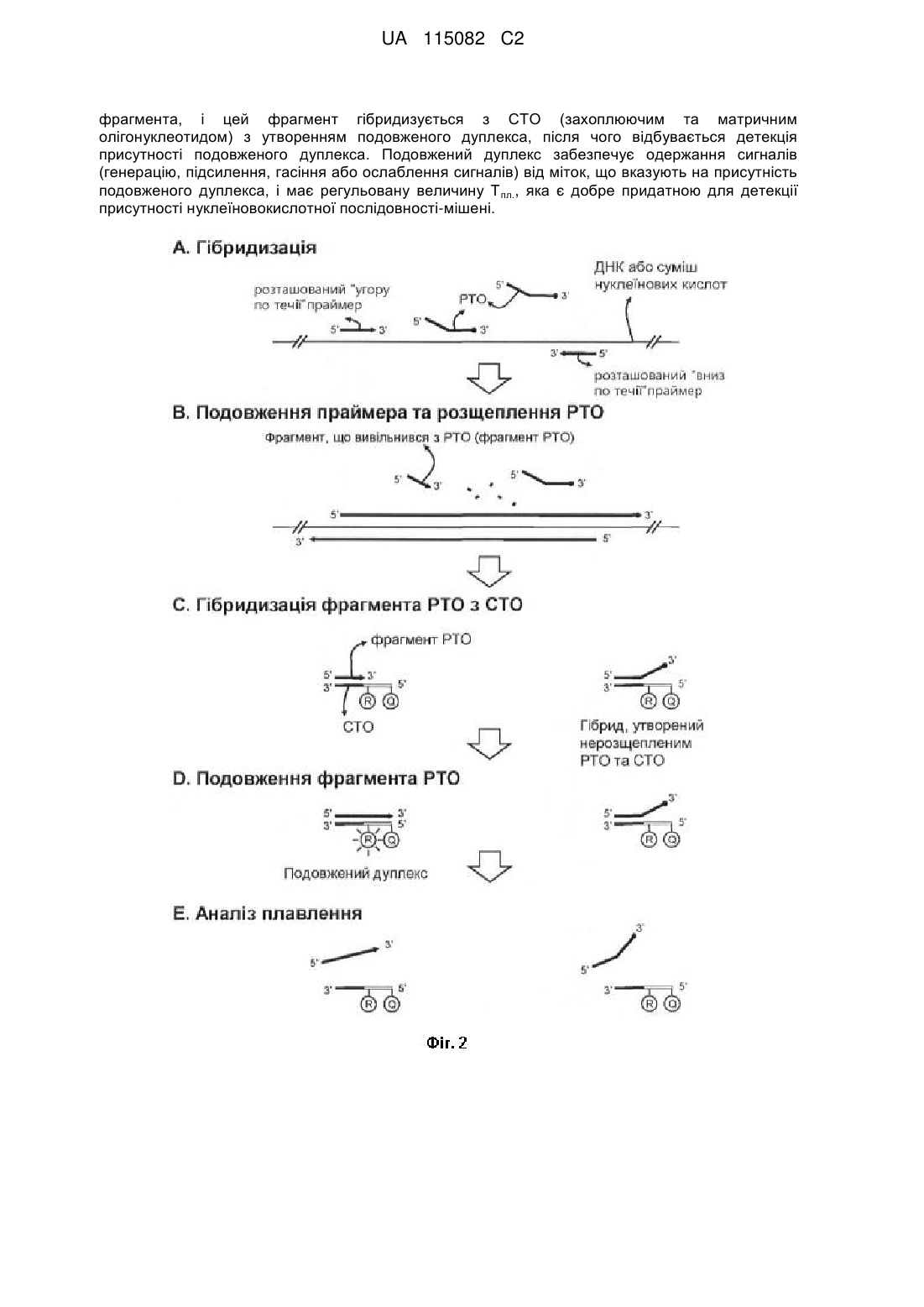
(a) гібридизацію нуклеїновокислотної послідовності-мішені з розташованими "угору по течії" олігонуклеотидом та РТО (олігонуклеотидом, що зондує та мітить); при цьому розташований "угору по течії" олігонуклеотид містить нуклеотидну послідовність, що гібридизується, комплементарну нуклеїновокислотній послідовності-мішені; РТО містить (1) 3'-кінцеву ділянку, що впізнає мішень, яка містить нуклеотидну послідовність, що гібридизується, комплементарну нуклеїновокислотній послідовності-мішені, та (2) 5'-кінцеву ділянку, що мітить, яка містить нуклеотидну послідовність, некомплементарну нуклеїновокислотній послідовності-мішені; при цьому 3'-кінцева ділянка, що впізнає мішень, гібридизується з нуклеїновокислотною послідовністю-мішенню, а 5'-кінцева ділянка, що мітить, не гібридизується з нуклеїновокислотною послідовністю-мішенню; розташований "угору по течії" олігонуклеотид локалізований "угору по течії" відносно РТО;
(b) приведення в контакт продукту зі стадії (а) з ферментом, що виявляє 5'-нуклеазну активність, в умовах, придатних для розщеплення РТО; при цьому розташований "угору по течії" олігонуклеотид або його подовжений ланцюг індукує розщеплення РТО ферментом, що виявляє 5'-нуклеазну активність, так що в результаті розщеплення вивільняється фрагмент, який містить 5'-кінцеву ділянку, що мітить, або частину 5'-кінцевої ділянки РТО, що мітить;
(c) гібридизацію фрагмента, вивільненого з РТО, з СТО (захоплюючим та матричним олігонуклеотидом); при цьому СТО містить в напрямку 3'®5' (1) захоплюючу ділянку, що містить нуклеотидну послідовність, комплементарну 5'-кінцевій ділянці, що мітить, або частині 5'-кінцевої ділянки РТО, що мітить, і (2) матричну ділянку, що містить нуклеотидну послідовність, некомплементарну 5'-кінцевій ділянці, що мітить, та 3'-кінцевій ділянці РТО, що впізнає мішень; при цьому фрагмент, вивільнений з РТО, гібридизується із захоплюючою ділянкою СТО;
(d) проведення реакції подовження з використанням продукту зі стадії (с) та матричної полімерази нуклеїнових кислот; при цьому фрагмент, гібридизований із захоплюючою ділянкою СТО, подовжується з утворенням подовженого дуплекса; причому подовжений дуплекс має величину Тпл., регульовану (1) послідовністю та/або довжиною цього фрагмента, (2) послідовністю та/або довжиною СТО, або (3) послідовністю та/або довжиною фрагмента, та послідовністю та/або довжиною СТО; при цьому подовжений дуплекс забезпечує одержання сигналу від мішені за допомогою (1) принаймні однієї мітки, з'єднаної з фрагментом та/або СТО, (2) мітки, вбудовуваної в подовжений дуплекс під час реакції подовження, (3) принаймні однієї мітки, з'єднаної з фрагментом та/або СТО, та мітки, вбудовуваної в подовжений дуплекс під час реакції подовження, або (4) інтеркалюючої мітки;
(e) детекцію подовженого дуплекса шляхом вимірювання сигналу від мішені в рідкій фазі при попередньо заданій температурі, при якій подовжений дуплекс зберігає свою дволанцюгову форму; тим самим присутність подовженого дуплекса вказує на присутність нуклеїновокислотної послідовності-мішені, при цьому подовжений дуплекс не іммобілізований на твердій підкладці.
2. Спосіб детекції нуклеїновокислотної послідовності-мішені з ДНК або суміші нуклеїнових кислот в аналізі з РТОСЕ (розщепленням та подовженням РТО) на твердій фазі, який включає:
(a) гібридизацію нуклеїновокислотної послідовності-мішені з розташованими "угору по течії" олігонуклеотидом та РТО (олігонуклеотидом, що зондує та мітить); при цьому розташований "угору по течії" олігонуклеотид містить нуклеотидну послідовність, що гібридизується, комплементарну нуклеїновокислотній послідовності-мішені; РТО містить (1) 3'-кінцеву ділянку, що впізнає мішень, яка містить нуклеотидну послідовність, що гібридизується, комплементарну нуклеїновокислотній послідовності-мішені, та (2) 5'-кінцеву ділянку, що мітить, яка містить нуклеотидну послідовність, некомплементарну нуклеїновокислотній послідовності-мішені; при цьому 3'-кінцева ділянка, що впізнає мішень, гібридизується з нуклеїновокислотною послідовністю-мішенню, а 5'-кінцева ділянка, що мітить, не гібридизується з нуклеїновокислотною послідовністю-мішенню; розташований "угору по течії" олігонуклеотид локалізований "угору по течії" відносно РТО;
(b) приведення в контакт продукту зі стадії (а) з ферментом, що виявляє 5'-нуклеазну активність, в умовах, придатних для розщеплення РТО; при цьому розташований "угору по течії" олігонуклеотид або його подовжений ланцюг індукує розщеплення РТО ферментом, що виявляє 5'-нуклеазну активність, так що в результаті розщеплення вивільняється фрагмент, який містить 5'-кінцеву ділянку, що мітить, або частину 5'-кінцевої ділянки РТО, що мітить;
(c) гібридизацію фрагмента, вивільненого з РТО, з СТО (захоплюючим та матричним олігонуклеотидом); при цьому СТО іммобілізований на твердій підкладці своїм 5'-кінцем або 3'-кінцем; при цьому СТО містить в напрямку 3'®5' (1) захоплюючу ділянку, що містить нуклеотидну послідовність, комплементарну 5'-кінцевій ділянці, що мітить, або частині 5'-кінцевої ділянки РТО, що мітить, і (2) матричну ділянку, що містить нуклеотидну послідовність, некомплементарну 5'-кінцевій ділянці, що мітить, та 3'-кінцевій ділянці РТО, що впізнає мішень; при цьому фрагмент, вивільнений з РТО, гібридизується із захоплюючою ділянкою СТО;
(d) проведення реакції подовження з використанням продукту зі стадії (с) та матричної полімерази нуклеїнових кислот; при цьому фрагмент, гібридизований із захоплюючою ділянкою СТО, подовжується з утворенням подовженого дуплекса; причому подовжений дуплекс має величину Тпл., регульовану (1) послідовністю та/або довжиною цього фрагмента, (2) послідовністю та/або довжиною СТО, або (3) послідовністю та/або довжиною фрагмента та послідовністю та/або довжиною СТО; при цьому подовжений дуплекс забезпечує одержання сигналу від мішені за допомогою (1) принаймні однієї мітки, з'єднаної з фрагментом та/або СТО, (2) мітки, вбудовуваної в подовжений дуплекс під час реакції подовження, (3) принаймні однієї мітки, з'єднаної з фрагментом та/або СТО, та мітки, вбудовуваної в подовжений дуплекс під час реакції подовження, або (4) інтеркалюючої мітки; і
(є) детекцію подовженого дуплекса шляхом вимірювання сигналу від мішені на твердій фазі при попередньо заданій температурі, при якій подовжений дуплекс зберігає свою дволанцюгову форму; тим самим присутність подовженого дуплекса вказує на присутність нуклеїновокислотної послідовності-мішені, при цьому подовжений дуплекс іммобілізований на твердій підкладці за допомогою СТО.
3. Спосіб за п. 1 або 2-, де сигнал від мішені, забезпечуваний подовженим дуплексом, доставляється під час стадії подовження (d); при цьому гібрид, утворений нерозщепленим РТО та СТО, не дає сигналу, який не належить до мішені.
4. Спосіб за п. 1 або 2, де сигнал від мішені, забезпечуваний подовженим дуплексом, доставляється в результаті гібридизації фрагмента та СТО на стадії (с), і утворення подовженого дуплекса зберігає сигнал від мішені на стадії (d); при цьому гібрид, утворений нерозщепленим РТО та СТО, дає сигнал, який не належить до мішені; при цьому попередньо задана температура є достатньою для розщеплення гібриду для видалення сигналу, який не належить до мішені.
5. Спосіб за п. 1 або 2, у якому одержання сигналу від мішені забезпечується за допомогою принаймні однієї мітки, з'єднаної з фрагментом та/або СТО.
6. Спосіб за п. 5, у якому фрагмент містить систему двох взаємодіючих міток, яка складається з репортерної молекули та молекули-гасника; при цьому гібридизація фрагмента та СТО на стадії (с) індукує зміну сигналу від системи двох взаємодіючих міток з одержанням сигналу від мішені, а подовжений дуплекс зберігає сигнал від мішені.
7. Спосіб за п. 5, у якому СТО містить систему двох взаємодіючих міток, яка складається з репортерної молекули та молекули-гасника; при цьому гібридизація фрагмента та СТО на стадії (с) індукує зміну сигналу від системи двох взаємодіючих міток з одержанням сигналу від мішені, а подовжений дуплекс зберігає сигнал від мішені.
8. Спосіб за п. 5, у якому СТО містить систему двох взаємодіючих міток, що складається з репортерної молекули та молекули-гасника; при цьому подовження фрагмента на стадії (d) індукує зміну сигналу від системи двох взаємодіючих міток з одержанням сигналу від мішені.
9. Спосіб за п. 5, у якому фрагмент містить одну з двох взаємодіючих міток, що включають репортерну молекулу та молекулу-гасник, а СТО містить іншу з двох взаємодіючих міток; при цьому гібридизація фрагмента та СТО на стадії (с) індукує зміну сигналу від системи двох взаємодіючих міток з одержанням сигналу від мішені, а подовжений дуплекс зберігає сигнал від мішені.
10. Спосіб за п. 5, у якому фрагмент або СТО містить одиночну мітку, і гібридизація фрагмента та СТО на стадії (с) індукує зміну сигналу від одиночної мітки з одержанням сигналу від мішені, а подовжений дуплекс зберігає сигнал від мішені.
11. Спосіб за п. 5, у якому СТО містить одиночну мітку, а подовження фрагмента на стадії (d) індукує зміну сигналу від цієї одиночної мітки з одержанням сигналу від мішені.
12. Спосіб за п. 5, у якому мітки розташовані таким чином, що, якщо утворюється гібрид, який складається з нерозщепленого РТО та СТО, то цей гібрид не дає сигналу, який не належить до мішені, на стадії (d).
13. Спосіб за п. 5, у якому мітки розташовані таким чином, що, якщо утворюється гібрид, який складається з нерозщепленого РТО та СТО, то цей гібрид дає сигнал, який не належить до мішені, на стадії (d); при цьому величина Тпл. для подовженого дуплекса перевищує таку для гібриду, утвореного нерозщепленим РТО та СТО.
14. Спосіб за п. 1, у якому одержання сигналу від мішені забезпечується за допомогою одиночної мітки, вбудовуваної в подовжений дуплекс під час реакції подовження; причому вбудовувана одиночна мітка з'єднана з нуклеотидом, вбудовуваним під час реакції подовження; при цьому подовження фрагмента на стадії (d) індукує зміну сигналу від одиночної мітки з одержанням сигналу від мішені на стадії (d).
15. Спосіб за п. 2, у якому одержання сигналу від мішені забезпечується за допомогою одиночної мітки, з'єднаної з фрагментом, або за допомогою одиночної мітки, вбудовуваної в подовжений дуплекс під час реакції подовження.
16. Спосіб за п. 14 або 15, у якому нуклеотид, вбудовуваний під час реакції подовження, містить першу неприродну основу, а СТО містить нуклеотид, що містить другу неприродну основу з афінністю специфічного зв'язування з першою неприродною основою.
17. Спосіб за п. 1 або 2, у якому одержання сигналу від мішені забезпечується за допомогою мітки, вбудовуваної в подовжений дуплекс під час реакції подовження, та мітки, з'єднаної з фрагментом та/або СТО; причому вбудовувана мітка з'єднана з нуклеотидом, вбудовуваним під час реакції подовження; причому ці дві мітки є системою двох взаємодіючих міток, яка складається з репортерної молекули та молекули-гасника; при цьому подовження фрагмента на стадії (d) індукує зміну сигналу від цієї системи двох взаємодіючих міток з одержанням сигналу від мішені.
18. Спосіб за п. 17, у якому нуклеотид, вбудовуваний під час реакції подовження, містить першу неприродну основу, а СТО містить нуклеотид, що містить другу неприродну основу з афінністю специфічного зв'язування з першою неприродною основою.
19. Спосіб за п. 1 або 2, у якому розташований "угору по течії" олігонуклеотид є розташованим "угору по течії" праймером або розташованим "угору по течії" зондом.
20. Спосіб за п. 1 або 2, який додатково включає повторення стадій (a)-(b), (a)-(d) або (а)-(е) з денатурацією між повторюваними циклами.
21. Спосіб за п. 1 або 2, у якому стадії (а)-(е) здійснюють в реакційній посудині або в окремих реакційних посудинах.
22. Спосіб за п. 1 або 2, який здійснюють з метою детекції принаймні двох типів нуклеїновокислотних послідовностей-мішеней; причому розташований "угору по течії" олігонуклеотид включає принаймні два типи олігонуклеотидів, РТО включає принаймні два типи РТО, а СТО включає принаймні один тип СТО.
23. Спосіб за будь-яким з пп. 1-22, який здійснюють в присутності розташованого "вниз по течії" праймера.
Текст
Реферат: Винахід стосується детекції нуклеїновокислотної послідовності-мішені в аналізі з РТОСЕ (розщепленням та подовженням РТО), при цьому РТО (олігонуклеотид, що зондує та мітить), гібридизований з нуклеїновокислотною послідовністю-мішенню, розщеплюється з вивільненням UA 115082 C2 (12) UA 115082 C2 фрагмента, і цей фрагмент гібридизується з СТО (захоплюючим та матричним олігонуклеотидом) з утворенням подовженого дуплекса, після чого відбувається детекція присутності подовженого дуплекса. Подовжений дуплекс забезпечує одержання сигналів (генерацію, підсилення, гасіння або ослаблення сигналів) від міток, що вказують на присутність подовженого дуплекса, і має регульовану величину Т пл., яка є добре придатною для детекції присутності нуклеїновокислотної послідовності-мішені. UA 115082 C2 5 10 15 20 25 30 35 40 45 50 55 60 Даний винахід стосується детекції нуклеїновокислотної послідовності-мішені в аналізі з РТОСЕ (розщепленням та подовженням РТО). Гібридизація ДНК є фундаментальним процесом в молекулярній біології, і на нього впливають іонна сила, склад основ, довжина фрагмента, до якого вкорочено нуклеїнову кислоту, ступінь помилкового спарювання та присутність денатуруючих агентів. Основані на гібридизації ДНК-технології були б надзвичайно корисним засобом у визначенні конкретної нуклеїновокислотної послідовності та були б, безперечно, необхідні в області клінічної діагностики, генетичних досліджень та лабораторної судово-медичної експертизи. Однак, в традиційних методах та способах, основаних головним чином на гібридизації, з великою вірогідністю отримують псевдопозитивні результати внаслідок неспецифічної гібридизації між зондами та послідовностями, що не є мішенями. Зважаючи на це, залишаються проблеми, які потребують вирішення з метою підвищення надійності цих способів. Крім гібридизаційних способів з використанням зондів були запропоновані різні підходи з використанням додаткових ферментативних реакцій, наприклад спосіб із застосуванням ТаqМаn™-зондів. У способі із застосуванням TaqMan™-зoндiв мічений зонд, гібридизований з нуклеїновокислотною послідовністю-мішенню, розщеплюється під дією 5'-нуклеазної активності ДНК-полімерази, залежної від розташованого "угору по течії" праймера, генеруючи сигнал, що вказує на присутність послідовності-мішені (патенти США №№ 5210015, 5538848 та 6326145). Відповідно до способу із застосуванням TaqMan™-зондів передбачаються два підходи для генерації сигналу: залежне від полімеризації розщеплення та незалежне від полімеризації розщеплення. При залежному від полімеризації розщепленні подовження розташованого "угору по течії" праймера повинно відбуватися до того, як полімераза нуклеїнових кислот зустрінеться з 5'-кінцем міченого зонда. У міру проходження реакції подовження полімераза поступово розщеплює 5'-кінець міченого зонда. При незалежному від полімеризації розщепленні розташований "угору по течії" праймер та мічений зонд гібридизуються з нуклеїновокислотною послідовністю-мішенню в безпосередній близькості, так що зв'язування полімерази нуклеїнових кислот з 3'-кінцем розташованого "угору по течії" праймера приводить її у контакт з 5'-кінцем міченого зонда, результатом чого є вивільнення мітки. Крім того, у способі із застосуванням TaqMan™-зондів описується, що мічений зонд, який має на своєму 5'-кінці 5'-хвостову ділянку, що не гібридизується з послідовністю-мішенню, також розщеплюється з утворенням фрагмента, що містить цю 5'-хвостову ділянку. Повідомлялося про деякі способи, у яких зонд, що має 5'-хвостову ділянку, некомлементарну послідовності-мішені, розщеплюється під дією 5'-нуклеази з вивільненням фрагмента, який містить цю 5'-хвостову ділянку. Наприклад, в патенті США № 5691142 описується розщеплювана структура, яка повинна переварюватися під дією 5'-нуклеазної активності ДНК-полімерази. Наводиться приклад даної розщеплюваної структури, у якій олігонуклеотид, що містить 5'-ділянку, некомплементарну до матриці, та 3'-ділянку, комплементарну до матриці, гібридизується з даною матрицею, і в безпосередній близькості з цією матрицею гібридизується розташований "угору по течії" олігонуклеотид. Така розщеплювана структура розщеплюється ДНК-полімеразою, яка виявляє 5'-нуклеазну активність, або модифікованою ДНК-полімеразою зі зниженою здатністю до синтезу з вивільненням 5'-ділянки, некомплементарної даній матриці. Вивільнена 5'-ділянка далі гібридизується з олігонуклеотидом, що має шпильчасту структуру, з утворенням розщеплюваної структури, індукуючи тим самим прогресуючі реакції розщеплення, необхідні для детекції послідовності-мішені. В патенті США № 7381532 описується спосіб, у якому розщеплювана структура, що містить розташований "угору по течії" олігонуклеотид з блокованим 3'-кінцем, розщеплюється ДНКполімеразою, яка виявляє 5'-нуклеазну активність, або нуклеазою FEN (флеп-ендонуклеазою; від англ. Flap EndoNuclease) з вивільненням некомплементарної 5'-кінцевої одноланцюгової "звисаючої" ділянки (флепа), і детекцію цієї вивільненої 5'-кінцевої "звисаючої" ділянки здійснюють, використовуючи аналіз за розміром або систему двох взаємодіючих міток. В патенті США № 6893819 описується, що детектовані вивільнені "звисаючі" ділянки утворюються в результаті застосування залежного від синтезу нуклеїнової кислоти, флеп-опосередкованого способу ампліфікації послідовностей. В цьому способі вивільнена з першої розщеплюваної структури "звисаюча" ділянка опосередковує у залежний від синтезу нуклеїнової кислоти спосіб розщеплення другої розщеплюваної структури, що необхідно для вивільнення "звисаючої"" ділянки з другої розщеплюваної структури, і вивільнені "звисаючі" ділянки детектують. Використовуючи гібридизацію мічених флуоресцентною міткою зондів в рідкій фазі, можна здійснити одночасну детекцію множини нуклеїновокислотних послідовностей-мішеней із 1 UA 115082 C2 5 10 15 20 25 30 35 40 45 50 55 застосуванням флуоресцентної мітки навіть одного типу шляхом аналізу кривої плавлення. Однак, традиційні технології детекції послідовностей-мішеней із застосуванням 5'-нуклеазаопосередкованого розщеплення зондів, що містять систему двох взаємодіючих міток, потребують наявності флуоресцентних міток різних типів для різних послідовностей-мішеней при детекції численних мішеней, що обмежує число детектованих послідовностей-мішеней внаслідок обмеженої кількості типів флуоресцентних міток. В публікації заявки на патент США 2008/0241838 описується спосіб детекції мішені з використанням розщеплення зонда, що містить 5'-кінцеву ділянку, некомплементарну нуклеїновокислотній послідовності-мішені, та гібридизації зонда захоплення. Мітка розташована на некомплементарній 5'-кінцевій ділянці. Мічений зонд, гібридизований з послідовністюмішенню, розщеплюється з вивільненням фрагмента, після чого цей фрагмент далі гібридизується із зондом захоплення для детекції присутності послідовності-мішені. В цьому способі необхідно, щоб нерозщеплений/інтактний зонд не гібридизувався із зондом захоплення. Для цього зонд захоплення, який має меншу довжину, має бути іммобілізованим на твердій підкладці. Однак таке обмеження приводить до зниження ефективності гібридизації на твердій підкладці, а також до ускладнень при оптимізації реакційних умов. Таким чином, в даній області залишається давно назріла необхідність в розробці нових підходів до детекції послідовності-мішені, краще, численних послідовностей-мішеней, в рідкій фазі та на твердій фазі, не тільки шляхом гібридизації, але також з використанням ферментативних реакцій, таких як 5'-нуклеолітична реакція, у більш зручний, надійний та відтворюваний спосіб. Крім того, в даній області також існує необхідність в новому способі детекції мішені, не обмеженому кількістю типів міток (зокрема, флуоресцентними мітками). По всій цій заявці зроблені посилання на різні патенти та публікації, і згадки про них наведені у круглих дужках. Тим самим опис цих патентів та публікацій в усій своїй повноті включений до даної заявки шляхом посилання з метою більш повного опису даного винаходу та стану області, до якої цей винахід має відношення. Автори даного винаходу провели інтенсивні дослідження з метою розробки нових підходів для детекції послідовностей-мішеней, які характеризуються більш високою точністю та зручністю, крім іншого, в режимі множинної детекції. В результаті автори винаходу розробили нові протоколи для детекції послідовностей-мішеней, у яких детекція мішені здійснюється шляхом гібридизації зонда, ферментативного розщеплення зонда, подовження та детекції подовженого дуплекса. Протоколи за даним винаходом добре адаптовані як до реакцій у рідкій фазі, так і до реакцій на твердій фазі, і дають можливість здійснювати детекцію численних послідовностей-мішеней з більш високою точністю та більш зручно. Відповідно, даним винаходом вирішується задача розробки способу детекції нуклеїновокислотної послідовності-мішені з ДНК або суміші нуклеїнових кислот в аналізі з РТОСЕ (розщепленням та подовженням РТО). Іншою задачею даного винаходу є розробка набору для детекції нуклеїновокислотної послідовності-мішені з ДНК або суміші нуклеїнових кислот в аналізі з РТОСЕ (розщепленням та подовженням РТО). Інші задачі та переваги даного винаходу будуть очевидні з наведеного далі детального опису у поєднанні з прикладеною формулою винаходу та графічними матеріалами. На Фіг. 1 схематично показані структури РТО (олігонуклеотиду, що зондує та мітить (probing and tagging oligonucleotide)) та СТО (захоплюючого та матричного олігонуклеотиду (capturing and templating oligonucleotide)), використовуваних в аналізі із розщепленням та подовженням РТО (РТОСЕ-аналізі). Краще, щоб 3'-кінці РТО та СТО були блоковані, щоб заважати до подовження. На Фіг. 2 схематично представлений РТОСЕ-аналіз, який включає аналіз плавлення. СТО містить репортерну молекулу та молекулу-гасник на своїй матричній ділянці (templating portion). На Фіг. 3 схематично представлений РТОСЕ-аналіз, який включає аналіз плавлення. СТО містить репортерну молекулу та молекулу-гасник на своїй матричній ділянці. Репортерна молекула повинна демонструвати різну інтенсивність сигналу в залежності від її присутності на одноланцюговій формі або на дволанцюговій формі. На Фіг. 4 схематично представлений РТОСЕ-аналіз, який включає аналіз плавлення. СТО містить остаток ізо-dC (ізодезоксицитозину) та репортерну молекулу на своїй матричній ділянці. Структура гасник-ізо-dGTP (дезоксигуанозинтрифосфат) вбудовується в подовжений дуплекс під час реакції подовження. На Фіг. 5 схематично представлений РТОСЕ-аналіз, який включає аналіз плавлення. РТО містить репортерну молекулу на своїй 5'-кінцевій ділянці, що мітить (5'-tagging portion), а СТО 2 UA 115082 C2 5 10 15 20 25 30 35 40 45 50 55 містить залишок ізо-dC на своїй матричній ділянці. Структура гасник-ізо-dGTP вбудовується в подовжений дуплекс під час реакції подовження. На Фіг. 6 схематично представлений РТОСЕ-аналіз, який включає аналіз плавлення. РТО містить репортерну молекулу та молекулу-гасник на своїй 5'-кінцевій ділянці, що мітить. На Фіг. 7 схематично представлений РТОСЕ-аналіз, який включає аналіз плавлення. РТО містить репортерну молекулу на своій 5'-кінцевій ділянці, що мітить. Репортерна молекула повинна демонструвати різну інтенсивність сигналу в залежності від її присутності на одноланцюговій формі або дволанцюговій формі. На Фіг. 8 схематично представлений РТОСЕ-аналіз, який включає аналіз плавлення. РТО містить молекулу-гасник на своій 5'-кінцевій ділянці, що мітить, а СТО містить репортерну молекулу на своій захоплюючій ділянці (capturing portion). На Фіг. 9 схематично представлений РТОСЕ-аналіз, який включає детекцію при попередньо заданій температурі. СТО містить репортерну молекулу та молекулу-гасник на своій матричній ділянці. СТО іммобілізований на твердій підкладці своїм 3'-кінцем. На Фіг. 10 схематично представлений РТОСЕ-аналіз, який включає детекцію при попередньо заданій температурі. Мічений репортерною молекулою dATP (дезоксіаденозинтрифосфат) вбудовується в подовжений дуплекс під час реакції подовження. СТО іммобілізований на твердій підкладці своїм 3'-кінцем. На Фіг. 11 схематично представлений РТОСЕ-аналіз, який включає детекцію при попередньо заданій температурі. СТО містить залишок ізо-dC на своій матричній ділянці, а структура репортер-ізо-dGTP вбудовується в подовжений дуплекс під час реакції подовження. СТО іммобілізований на твердій підкладці своїм 3'-кінцем. На Фіг. 12 схематично представлений РТОСЕ-аналіз, який включає детекцію при попередньо заданій температурі. РТО містить репортерну молекулу на своій 5'-кінцевій ділянці, що мітить. СТО іммобілізований на твердій підкладці своїм 5'-кінцем. На Фіг. 13 схематично представлений РТОСЕ-аналіз, який включає детекцію при попередньо заданій температурі з використанням інтеркалюючого барвника. СТО іммобілізований на твердій підкладці своїм 5'-кінцем. На Фіг. 14 показані результати детекції гена Neisseria gonorrhoeae з використанням РТОСЕаналізу, який включає аналіз плавлення. СТО містить репортерну молекулу та молекулу-гасник на своій матричній ділянці. На Фіг. 15 показані результати детекції гена Neisseria gonorrhoeae з використанням РТОСЕаналізу, який включає аналіз плавлення. РТО містить молекулу-гасник на своєму 5'-кінці, а СТО містить репортерну молекулу на своєму 3'-кінці. На Фіг. 16 показані результати, які свідчать про те, що величини Т пл. для подовжених дуплексів узгоджуються з послідовностями СТО. На Фіг. 17 показані результати детекції гена Neisseria gonorrhoeae шляхом РТОСЕ-аналізу з використанням ПЛР-ампліфікації. СТО містить репортерну молекулу та молекулу-гасник на своій матричній ділянці. На Фіг. 17А показані результати РТОСЕ-аналізу, який включає детекцію з використанням ПЛР в режимі реального часу, а на Фіг. 17В показані результати РТОСЕаналізу, який включає аналіз плавлення після проведення ПЛР. На Фіг. 18 показані результати детекції гена Neisseria gonorrhoeae шляхом РТОСЕ-аналізу з використанням ПЛР-ампліфікації. СТО містить залишок ізо-dC та репортерну молекулу на своєму 5'-кінці. Структура гасник-ізо-dGTP вбудовується в подовжений дуплекс під час реакції подовження. На Фіг. 18А показані результати РТОСЕ-аналізу, який включає детекцію з використанням ПЛР в режимі реального часу, а на Фіг. 18В показані результати РТОСЕ-аналізу, який включає аналіз плавлення після проведення ПЛР. На Фіг. 19 показані результати детекції гена Neisseria gonorrhoeae шляхом РТОСЕ-аналізу з використанням ПЛР-ампліфікації. РТО містить молекулу-гасник на своєму 5'-кінці, а СТО містить репортерну молекулу на своєму 3'-кінці. На Фіг. 19А показані результати РТОСЕ-аналізу, який включає детекцію з використанням ПЛР в режимі реального часу, а на Фіг. 19В показані результати РТОСЕ-аналізу, який включає аналіз плавлення після проведення ПЛР. На Фіг. 20 показані результати одночасної детекції гена Neisseria gonorrhoeae (NG) та гена Staphylococcus aureus (SA) з використанням РТОСЕ-аналізу, який включає аналіз плавлення після проведення ПЛР. СТО містить репортерну молекулу та молекулу-гасник на своїй матричній ділянці. На Фіг. 21 показані результати детекції гена Neisseria gonorrhoeae з використанням РТОСЕаналізу, який включає аналіз плавлення на мікрочипі. СТО іммобілізований своїм 5'-кінцем. РТО містить репортерну молекулу на своій 5'-кінцевій ділянці, що мітить. 3 UA 115082 C2 5 10 15 20 25 30 35 40 45 50 55 60 На Фіг. 22 показані результати детекції гена Neisseria gonorrhoeae з використанням РТОСЕаналізу, який включає детекцію в режимі реального часу при попередньо заданій температурі на мікрочипі. СТО іммобілізований своїм 5'-кінцем. РТО містить репортерну молекулу на своій 5'-кінцевій ділянці, що мітить. На Фіг. 23 показані результати детекції гена Neisseria gonorrhoeae з використанням РТОСЕаналізу, який включає детекцію в режимі реального часу при попередньо заданій температурі на мікрочипі. СТО іммобілізований своїм 3'-кінцем і містить репортерну молекулу та молекулугасник на своій матричній ділянці. На Фіг. 24 показані результати детекції однієї мішені та численних мішеней з використанням РТОСЕ-аналізу, який включає детекцію в режимі кінцевої точки при попередньо заданій температурі на мікрочипі. СТО іммобілізований своїм 5'-кінцем. РТО містить репортерну молекулу на своій 5'-кінцевій ділянці, що мітить. Як нуклеїновокислотні послідовності-мішені використовували ген Neisseria gonorrhoeae (NG) та ген Staphylococcus aureus (SA). Даний винахід стосується нового способу детекції нуклеїновокислотної послідовності-мішені в аналізі з РТОСЕ (розщепленням та подовженням РТО) та набору для детекції нуклеїновокислотної послідовності-мішені в РТОСЕ-аналізі. Даний винахід включає не тільки реакції гібридизації, але також й ферментативні реакції, перебіг яких залежить від присутності нуклеїновокислотної послідовності-мішені. І. Спосіб детекції мішені в РТОСЕ-аналізі, який включає аналіз плавлення В одному з аспектів даного винаходу запропонований спосіб детекції нуклеїновокислотної послідовності-мішені з ДНК або суміші нуклеїнових кислот в аналізі з РТОСЕ (розщепленням та подовженням РТО), який включає: (а) гібридизацію нуклеїновокислотної послідовності-мішені з розташованими "угору по течії" олігонуклеотидом та РТО (олігонуклеотидом, що зондує та мітить); при цьому розташований "угору по течії" олігонуклеотид містить нуклеотидну послідовність, що гібридизується, комплементарну нуклеїновокислотній послідовності-мішені; РТО містить (1) 3'-кінцеву ділянку, що впізнає мішень (3'-targeting portion), яка включає нуклеотидну послідовність, що гібридизується, комплементарну нуклеїновокислотній послідовності-мішені, та (2) 5'-кінцеву ділянку, що мітить, яка включає нуклеотидну послідовність, некомплементарну нуклеїновокислотній послідовності-мішені; при цьому 3'-кінцева ділянка, що впізнає мішень, гібридизується з нуклеїновокислотною послідовністю-мішенню, а 5'-кінцева ділянка, що мітить, не гібридизується з нуклеїновокислотною послідовністю-мішенню; розташований "угору по течії" олігонуклеотид локалізований "угору по течії" відносно РТО; (b) приведення в контакт продукту зі стадії (а) з ферментом, що виявляє 5'-нуклеазну активність, в умовах, придатних для розщеплення РТО; при цьому розташований "угору по течії" олігонуклеотид або його подовжений ланцюг індукує розщеплення РТО ферментом, що виявляє 5'-нуклеазну активність, так що в результаті розщеплення вивільняється фрагмент, який містить 5'-кінцеву ділянку, що мітить, або частину 5'-кінцевої ділянки РТО, що мітить; (c) гібридизацію фрагмента, вивільненого з РТО, із СТО (захоплюючим та матричним олігонуклеотидом); при цьому СТО містить в напрямку 3'→5' (1) захоплюючу ділянку, яка містить нуклеотидну послідовність, комплементарну 5'-кінцевій ділянці, що мітить, або частині 5'-кінцевій ділянки РТО, що мітить, та (2) матричну ділянку, яка містить нуклеотидну послідовність, некомплементарну 5'-кінцевій ділянці, що мітить, та 3'-кінцевій ділянці РТО, що впізнає мішень; при цьому фрагмент, вивільнений з РТО, гібридизується із захоплюючою ділянкою СТО; (d) проведення реакції подовження з використанням продукту зі стадії (с) та матричної полімерази нуклеїнових кислот; при цьому фрагмент, гібридизований із захоплюючою ділянкою СТО, подовжується з утворенням подовженого дуплекса; причому подовжений дуплекс має величину Т пл., регульовану (1) послідовністю та/або довжиною цього фрагмента, (2) послідовністю та/або довжиною СТО, або (3) послідовністю та/або довжиною фрагмента і послідовністю та/або довжиною СТО; (е) плавлення подовженого дуплекса в діапазоні температур з одержанням сигналу від мішені (target signal), що вказує на присутність подовженого дуплекса; при цьому сигнал від мішені забезпечується за допомогою (1) принаймні однієї мітки, з'єднаної з фрагментом та/або СТО, (2) мітки, вбудовуваної в подовжений дуплекс під час реакції подовження, (3) мітки, вбудовуваної в подовжений дуплекс під час реакції подовження, та мітки, з'єднаної з фрагментом та/або СТО, або (4) інтеркалюючої мітки; і (f) детекцію подовженого дуплекса шляхом вимірювання сигналу від мішені; тим самим присутність подовженого дуплекса вказує на присутність нуклеїновокислотної послідовностімішені. 4 UA 115082 C2 5 10 15 20 25 30 35 40 45 50 55 60 Автори даного винаходу провели інтенсивні дослідження з метою розробки нових підходів для детекції послідовностей-мішеней, які характеризуються більш високою точністю та зручністю, крім іншого, в режимі множинної детекції. В результаті автори винаходу розробили нові протоколи для детекції послідовностей-мішеней, у яких детекція мішені здійснюється шляхом гібридизації зонда, ферментативного розщеплення зонда, подовження та детекції подовженого дуплекса. Протоколи за даним винаходом добре адаптовані як до реакцій в рідкій фазі, так і до реакцій на твердій фазі, та дають можливість здійснювати детекцію численних послідовностей-мішеней з більш високою точністю та більш зручно. В даному винаході реалізуються почергові події в такій послідовності: гібридизація зонда; розщеплення РТО (олігонуклеотиду, що зондує та мітить) і подовження; утворення мішеньзалежного подовженого дуплекса; та детекція подовженого дуплекса. Тому спосіб називається аналізом РТОСЕ (із розщепленням та подовженням РТО). В даному винаході подовжений дуплекс характеризується присутністю мітки (міток), яка забезпечує (які забезпечують) одержання сигналу, що вказує на присутність подовженого дуплекса, в результаті аналізу плавлення або детекції при попередньо заданій температурі. При цьому даний подовжений дуплекс характеризується регульованою величиною Т пл., що відіграє вирішальну роль при детекції численних мішеней або для того, щоб провести відмінність від сигналу, не пов'язаного з мішенню (non-target signal). Оскільки подовжений дуплекс утворюється тільки тоді, коли є нуклеїнова кислота-мішень, присутність подовженого дуплекса вказує на присутність цієї нуклеїнової кислоти-мішені. РТОСЕ-аналіз, який включає аналіз плавлення, далі буде описаний більш детально. Стадія (а). Гібридизація розташованого "угору по течії" олігонуклеотиду та РТО з нуклеїновокислотною послідовністю-мішенню Згідно з даним винаходом, нуклеїновокислотна послідовність-мішень спочатку гібридизується з розташованими "угору по течії" олігонуклеотидом та РТО (олігонуклеотидом, що зондує та мітить). Використовуваний в даному описі термін "нуклеїнова кислота-мішень", " нуклеїновокислотна послідовність-мішень" або "послідовність-мішень" стосується нуклеїновокислотної послідовності, що представляє інтерес для детекції, на якій відпалюють праймер, або яку гібридизують з праймером в умовах гібридизації, відпалу або ампліфікації. Використовуваний в даному описі термін "зонд" стосується молекули одноланцюгової нуклеїнової кислоти, яка містить ділянку або ділянки, по суті комплементарну (комплементарні) нуклеїновокислотній послідовності-мішені. Термін "праймер", як він використовується в даному описі, стосується олігонуклеотиду, який є здатним діяти як точки ініціації синтезу у випадку його уміщування в умови, при яких індукується синтез продукту подовження праймера, комплементарного ланцюга нуклеїнової кислоти (матриці), тобто, в присутності нуклеотидів і агента для полімеризації, такого як ДНКполімераза, та при придатних значеннях температури та рН. Краще, щоб зонд та праймер були молекулами одноланцюгових дезоксирибонуклеотидів. Зонди та праймери, використовувані в даному винаході, можуть містити природний dNMP (дезоксинуклеозид-монофосфат) (тобто dAMP (дезоксіаденозин-монофосфат), dGMP (дезоксигуанозин-монофосфат), dCMP (дезоксицитидин-монофосфат) та dTMP (дезокситимідин-монофосфат)), модифікований нуклеотид або неприродний нуклеотид. Зонди та праймери також можуть включати рибонуклеотиди. Праймер повинен мати достатню довжину, щоб праймувати синтез продуктів подовження в присутності агента для полімеризації. Точна довжина праймерів буде залежати від багатьох факторів, включаючи температуру, спосіб застосування та джерело праймера. Термін "відпал" або "праймування", як він використовується в даному описі, стосується приєднання олігодезоксинуклеотиду або нуклеїнової кислоти до нуклеїнової кислоти, яка є матрицею, при цьому таке приєднання дозволяє полімеразі здійснювати полімеризацію нуклеотидів з утворенням молекули нуклеїнової кислоти, що є комплементарною до нуклеїнової кислоти, яка є матрицею, або її частини. Термін "гібридизація", використовуваний в даному описі, стосується утворення дволанцюгової нуклеїнової кислоти з комплементарних одноланцюгових нуклеїнових кислот. Гібридизація може здійснюватися між двома ланцюгами нуклеїнової кислоти, повністю спареними або по суті спареними з певним числом помилкових спарювань. Комплементарність, необхідна для гібридизації, може залежати від умов гібридизації, зокрема, від температури. Гібридизація нуклеїновокислотної послідовності-мішені з розташованими "угору по течії" олігонуклеотидом та РТО може бути здійснена в придатних умовах гібридизації, визначених у встановленому порядку шляхом оптимізації методик. Такі умови, як температура, концентрація 5 UA 115082 C2 5 10 15 20 25 30 35 40 45 50 55 60 компонентів, тривалість гібридизації та промивання, компоненти буферів, їх рН та іонна сила, можуть бути змінені в залежності від різних факторів, включаючи довжину та GC-склад олігонуклеотиду (розташованого "угору по течії" олігонуклеотиду та РТО) та нуклеотидної послідовності-мішені. Наприклад, коли використовується відносно короткий олігонуклеотид, краще, щоб були прийняті умови низької жорсткості. Детальні умови гібридизації можна знайти у Sambrook et al., Molecular Cloning, A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. (2001); та M.L.M. Anderson, Nucleic Acid Hybridization, Springer-Verlag New York Inc. N.Y. (1999). He передбачається різниці між термінами "відпал" та "гібридизація", і ці терміни будуть використовуватися взаємозамінно. Розташований "угору по течії" олігонуклеотид та РТО містять нуклеотидні послідовності, що гібридизуються, комплементарні нуклеїновокислотній послідовності-мішені. Термін "комплементарний" використовується в даному описі для вказівки на те, що праймери або зонди є в достатньому ступені комплементарними, щоб селективно гібридизуватися з нуклеїновокислотною послідовністю-мішенню в намічених умовах відпалу або в жорстких умовах, охоплюючи терміни "по суті комплементарний" та "повністю комплементарний", краще, "повністю комплементарний". 5'-Кінцева ділянка РТО, що мітить, має нуклеотидну послідовність, некомплементарну нуклеїновокислотній послідовності-мішені. Матрична ділянка СТО (захоплюючого та матричного олігонуклеотиду) має нуклеотидну послідовність, некомплементарну 5'-кінцевій ділянці, що мітить, та 3'-кінцевій ділянці РТО, що впізнає мішень. Термін "некомплементарний" використовується в даному описі для вказівки на те, що праймери або зонди є в достатньому ступені некомплементарними, щоб селективно гібридизуватися з нуклеїновокислотною послідовністю-мішенню в намічених умовах відпалу або в жорстких умовах, охоплюючи терміни "по суті некомплементарний" та "повністю некомплементарний", краще, "повністю некомлементарний". Використовуваний в даному описі термін "РТО (олігонуклеотид, що зондує та мітить)" означає олігонуклеотид, який містить (1) 3'-кінцеву ділянку, що впізнає мішень, яка служить зондом, та (2) 5'-кінцеву ділянку, що мітить, з нуклеотидною послідовністю, некомплементарною нуклеїновокислотній послідовності-мішені, яка вивільняється в результаті нуклеолітичної реакції з РТО після гібридизації з нуклеїновокислотною послідовністю-мішенню. 5'-Кінцева ділянка, що мітить, та 3'-кінцева ділянка, що впізнає мішень, в РТО повинні розташовуватися в напрямку 5'→3'. РТО схематично показаний на Фіг. 1. Гібридизацію на стадії (а) краще проводять в жорстких умовах, так що 3'-кінцева ділянка, що впізнає мішень, гібридизується з нуклеїновокислотною послідовністю-мішенню, а 5'-кінцева ділянка, що мітить, не гібридизується з нуклеїновокислотною послідовністю-мішенню. Для РТО не передбачається якої-небудь конкретної довжини. Наприклад, довжина РТО може складати 15-150 нуклеотидів, 15-100 нуклеотидів, 15-80 нуклеотидів, 15-60 нуклеотидів, 15-40 нуклеотидів, 20-150 нуклеотидів, 20-100 нуклеотидів, 20-80 нуклеотидів, 20-60 нуклеотидів, 20-50 нуклеотидів, 30-150 нуклеотидів, 30-100 нуклеотидів, 30-80 нуклеотидів, 3060 нуклеотидів, 30-50 нуклеотидів, 35-100 нуклеотидів, 35-80 нуклеотидів, 35-60 нуклеотидів або 35-50 нуклеотидів. 3'-Кінцева ділянка РТО, що впізнає мішень, може бути будь-якої довжини за умови, що вона специфічно гібридизується з нуклеїновокислотними послідовностями-мішенями. Наприклад, довжина 3'-кінцевої ділянки РТО, що впізнає мішень, може складати 10-100 нуклеотидів, 10-80 нуклеотидів, 10-50 нуклеотидів, 10-40 нуклеотидів, 10-30 нуклеотидів, 15-100 нуклеотидів, 15-80 нуклеотидів, 15-50 нуклеотидів, 15-40 нуклеотидів, 15-30 нуклеотидів, 20-100 нуклеотидів, 20-80 нуклеотидів, 20-50 нуклеотидів, 20-40 нуклеотидів або 20-30 нуклеотидів. 5'Кінцева ділянка, що мітить, може бути будь-якої довжини за умови, що вона специфічно гібридизується з матричною ділянкою СТО і потім подовжується. Наприклад, довжина 5'кінцевої ділянки РТО, що мітить, може складати 5-50 нуклеотидів, 5-40 нуклеотидів, 5-30 нуклеотидів, 5-20 нуклеотидів, 10-50 нуклеотидів, 10-40 нуклеотидів, 10-30 нуклеотидів, 10-20 нуклеотидів, 15-50 нуклеотидів, 15^0 нуклеотидів, 15-30 нуклеотидів або 15-20 нуклеотидів. На 3'-кінці РТО може знаходитися група 3'-ОН. Краще, 3'-кінець РТО "блокований", щоб не допустити його подовження. Блокування може бути досягнуто традиційними методами. Наприклад, блокування можна здійснити шляхом додавання до 3'-гідроксильної групи останнього нуклеотиду хімічного угрупування, такого як біотин, мітка, фосфатна група, алкільна група, ненуклеотидний лінкер, фосфоротіонат або алкандіол. Альтернативно, блокування може бути здійснене шляхом видалення 3'-гідроксильної групи останнього нуклеотиду або шляхом використання нуклеотиду, позбавленого 3'-гідроксильної групи, такого як дидезоксинуклеотид. 6 UA 115082 C2 5 10 15 20 25 30 35 40 45 50 55 60 Альтернативно, РТО може бути сконструований так, щоб приймати шпильчасту структуру. Відсутність гібридизації між 5'-кінцевою ділянкою РТО, що мітить, та нуклеїновокислотною послідовністю-мішенню означає відсутність утворення з них стабільної дволанцюгової структури у визначених умовах гібридизації. Відповідно до кращого втілення, 5'-кінцева ділянка РТО, що мітить, не залучена до гібридизації з нуклеїновокислотною послідовністю-мішенню, утворює одноланцюгову структуру. Розташований "угору по течії"" олігонуклеотид локалізований "угору по течії"" відносно РТО. Крім цього, розташований "угору по течії" олігонуклеотид або його подовжений ланцюг, гібридизована з нуклеїновокислотною послідовністю-мішенню, індукує розщеплення РТО ферментом, що виявляє 5'-нуклеазну активність. Індукція розщеплення РТО за допомогою розташованого "угору по течії" олігонуклеотиду може відбуватися двома шляхами: (1) як індукція розщеплення, незалежна від подовження розташованого "угору по течії" олігонуклеотиду; та (2) як індукція розщеплення, залежна від подовження розташованого "угору по течії"" олігонуклеотиду. В тому випадку, коли розташований "угору по течії" олігонуклеотид локалізований досить близько від РТО, щоб індукувати розщеплення РТО ферментом, який виявляє 5'-нуклеазну активність, фермент, що зв'язався із розташованим "угору по течії" олігонуклеотидом, розщеплює РТО без реакції подовження. На відміну від цього, коли розташований "угору по течії"" олігонуклеотид локалізований у віддаленні від РТО, фермент, що виявляє полімеразну активність, (наприклад, матрична полімераза) каталізує подовження розташованого "угору по течії" олігонуклеотиду (наприклад, розташованого "угору по течії" праймера), а фермент, що виявляє 5'-нуклеазну активність, який зв'язався з подовженим продуктом, розщеплює РТО. Отже, розташований "угору по течії" олігонуклеотид може бути локалізований відносно РТО двома способами. Розташований "угору по течії" олігонуклеотид може бути локалізований досить близко від РТО, щоб індукувати розщеплення РТО у незалежний від подовження спосіб. Альтернативно, розташований "угору по течії"" олігонуклеотид може бути локалізований досить далеко від РТО, щоб індукувати розщеплення РТО у залежний від подовження спосіб. Використовуваний в даному описі термін "прилеглий" відносно до розташувань або локалізацій означає, що розташований "угору по течії" олігонуклеотид локалізований поблизу 3'кінцевої ділянки РТО, що впізнає мішень, з утворенням "ніка" (одноланцюгового розриву). Крім того, цей термін означає, що розташований "угору по течії"" олігонуклеотид локалізований на і відстані 1-30 нуклеотидів, 1-20 нуклеотидів або 1-15 нуклеотидів, від 3 -кінцевої ділянки РТО, що впізнає мішень. Використовуваний в даному описі термін "віддалений" відносно до розташувань або локалізацій, включає будь-які розташування або локалізації, достатні для забезпечення проходження реакцій подовження. Відповідно до кращого втілення, розташований "угору по течії" олігонуклеотид локалізований на достатньому віддаленні від РТО, щоб індукувати розщеплення РТО у залежний від подовження спосіб. Відповідно до кращого втілення, розташований "угору по течії" олігонуклеотид є розташованим "угору по течії" праймером або розташованим "угору по течії" зондом. Розташований "угору по течії" праймер є придатним для індукції незалежного від подовження розщеплення або для індукції залежного від подовження розщеплення, а розташований "угору по течії" зонд є придатним для індукції незалежного від подовження розщеплення. Альтернативно, розташований "угору по течії" олігонуклеотид може мати послідовність, яка частково перекривається з 5'-кінцевою частиною 3'-кінцевої ділянки РТО, що впізнає мішень. Краще, щоб довжина перекривної послідовності складала 1-10 нуклеотидів, ще краще, 1-5 нуклеотидів, навіть ще краще, 1-3 нуклеотиди. В тому випадку, коли розташований "угору по течії" олігонуклеотид має послідовність, яка частково перекривається з 5'-кінцевою частиною 3'кінцевої ділянки РТО, що впізнає мішень, ця 3'-кінцева ділянка, що впізнає мішень, частково переварюється разом з 5'-кінцевою ділянкою, що мітить, в реакції розщеплення на стадії (b). Крім цього, присутність перекривної послідовності дозволяє розщеплювати бажаний сайт 3'кінцевої ділянки, що впізнає мішень. Відповідно до кращого втілення, розташований "угору по течії" праймер індукує за допомогою свого подовженого ланцюга розщеплення РТО ферментом, що виявляє 5'-нуклеазну активність. В даному винаході можуть бути застосовані традиційні технології для реакцій розщеплення з використанням розташованих "угору по течії" олігонуклеотидів за умови, що розташований "угору по течії" олігонуклеотид індукує розщеплення РТО, гібридизованого з нуклеїновокислотною послідовністю-мішенню з вивільненням фрагмента, який містить 5'-кінцеву 7 UA 115082 C2 5 10 15 20 25 30 35 40 45 50 55 60 ділянку, що мітить, або частину 5'-кінцевої ділянки РТО, що мітить. Наприклад, в даному винаході можуть бути застосовані патенти США №№ 5210015, 5487972, 5691142, 5994069 та 7381532 і публікація заявки на патент США № 2008/0241838. Відповідно до кращого втілення, виконання даного способу здійснюють в присутності розташованого "вниз по течії" праймера. Розташований "вниз по течії" праймер додатково генерує нуклеїновокислотну послідовність-мішень, яка буде гібридизуватися з РТО, підвищуючи чутливість детекції мішені. Відповідно до кращого втілення, коли використовують розташований "угору по течії" праймер та розташований "вниз по течії" праймер, додатково для подовження цих праймерів застосовують матричну полімеразу нуклеїнових кислот. Відповідно до кращого втілення, розташований "угору по течії" олігонуклеотид (розташований "угору по течії" праймер або розташований "угору по течії" зонд), розташований "вниз по течії" праймер та/або 5'-кінцева ділянка РТО, що мітить, мають структуру олігонуклеотиду з подвійним праймуванням (DPO), розроблену авторами даного винаходу. Олігонуклеотиди, що мають структуру DPO, демонструють значно покращену специфічність до мішені у порівнянні з традиційними праймерами та зондами (див. WO 2006/095981; Chun et. al., Dual priming oligonucleotide system for the multiplex detection of respiratory viruses and SNP genotyping of CYP2C19 gene, Nucleic Acid Research, 35: 6e40(2007)). Відповідно до кращого втілення, 3'-кінцева ділянка РТО, що впізнає мішень, має структуру модифікованого олігонуклеотиду з подвійною специфічністю (mDSO), розроблену авторами даного винаходу. Модифікований олігонуклеотид з подвійною специфічністю (mDSO) демонструє значно покращену специфічність до мішені у порівнянні з традиційними зондами (див. WO 2011/028041). Стадія (b). Вивільнення фрагмента з РТО Далі, продукт зі стадії (а) приводять у контакт з ферментом, що виявляє 5'-нуклеазну активність, в умовах, придатних для розщеплення РТО. РТО, гібридизований з нуклеїновокислотною послідовністю-мішенню, переварюється ферментом, що виявляє 5'нуклеазну активність, вивільняючи фрагмент, який містить 5'-кінцеву ділянку, що мітить, або частину 5'-кінцевої ділянки РТО, що мітить. Використовуваний в даному описі термін "умови для розщеплення РТО" означає умови, достатні для перетравлювання РТО, гібридизованого з нуклеїновокислотною послідовністюмішенню, ферментом, що виявляє 5'-нуклеазну активність, такі як температура, рН, іонна сила, буфер, довжина та послідовність олігонуклеотидів, і ферменти. Наприклад, коли як фермент, що виявляє 5'-нуклеазну активність, використовують ДНК-полімеразу Taq, умови для розщеплення РТО включають буфер трис-НСl, КСl, MgCl2 та температуру. і Коли РТО гібридизується з нуклеїновокислотною послідовністю-мішенню, його 3 '-кінцева ділянка, що впізнає мішень, залучена у гібридизацію, а 5'-кінцева ділянка, що мітить, утворює одноланцюгову структуру без гібридизації з нуклеїновокислотною послідовністю-мішенню (див. Фіг. 2). По суті, олігонуклеотид, що містить як одноланцюгову, так і дволанцюгову структури, може бути переварений з використанням ферменту, що виявляє 5'-нуклеазну активність, із застосуванням різноманітних технологій, відомих фахівцю в даній області. Сайти розщеплення РТО варіюють в залежності від типу розташованих "угору по течії" олігонуклеотидів (розташованого "угору по течії" зонда або розташованого "угору по течії" праймера), сайтів гібридизації розташованих "угору по течії" олігонуклеотидів та умов розщеплення (див. патенти США №№ 5210015, 5487972, 5691142, 5994069 та 7381532 і публікацію заявки на патент США № 2008/0241838). Велике число традиційних технологій можуть бути застосовані для проведення реакції розщеплення РТО, що приводить до вивільнення фрагмента, який містить 5'-кінцеву ділянку, що мітить, або частину 5'-кінцевої ділянки, що мітить. Стисло, на стадії (b) можуть бути присутніми три сайти розщеплення. По-перше, сайтом розщеплення є з'єднувальний сайт (junction site) між ділянкою гібридизації РТО (3'-кінцевою ділянкою, що впізнає мішень) та ділянкою, що не гібридизується (5'-кінцевою ділянкою, що мітить). Другим сайтом розщеплення є сайт локалізації декількох нуклеотидів в 3'-напрямку від і 3'-кінця 5'-кінцевої ділянки РТО, що мітить. Другий сайт розщеплення локалізований в 5 -кінцевій частині '-кінцевої ділянки РТО, що впізнає мішень. Третій сайт розщеплення є сайтом локалізації декількох нуклеотидів в 5'-напрямку від 3'-кінця 5'-кінцевої ділянки РТО, що мітить. Відповідно до кращого втілення, початковим сайтом розщеплення РТО матричною полімеразою, що виявляє 5'-нуклеазну активність, після подовження розташованого "угору по течії" праймера, є вихідна точка подвійного ланцюга між РТО та нуклеїновокислотною послідовністю-мішенню або сайт на відстані 1-3 нуклеотидів від цієї вихідної точки. 8 UA 115082 C2 5 10 15 20 25 30 35 40 45 50 55 60 У зв'язку з цим, використовуваний в даному описі термін "фрагмент, який містить 5'-кінцеву ділянку, що мітить, або частину 5'-кінцевої ділянки РТО, що мітить" відносно до розщеплення РТО ферментом, який виявляє 5'-нуклеазну активність, застосовується для позначення (1) 5'кінцевої ділянки, що мітить, (2) 5'-кінцевої ділянки, що мітить, та 5'-кінцевої частини 3'-кінцевої ділянки, що впізнає мішень, і (3) частини 5'-кінцевої ділянки, що мітить. В даній заявці термін "фрагмент, який містить 5'-кінцеву ділянку, що мітить, або частину 5'-кінцевої ділянки РТО, що мітить", також можна описати як "фрагмент РТО". Термін "частина", використовуваний відносно до РТО або СТО, як частини 5'-кінцевої ділянки РТО, що мітить, 5'-кінцевої частини 3'-кінцевої ділянки РТО, що впізнає мішень, та 5'кінцевої частини захоплюючої ділянки СТО, стосується нуклеотидної послідовності, що містить 1-40,1-30, 1-20, 1-15, 1-10 або 1-5 нуклеотидів, краще 1, 2, 3 або 4 нуклеотиди. Відповідно до кращого втілення, фермент, що виявляє 5'-нуклеазну активність, є ДНКполімеразою, яка виявляє 5'-нуклеазну активність, або нуклеазою FEN, ще краще, термостабільною ДНК-полімеразою, що виявляє 5'-нуклеазну активність, або нуклеазою FEN. Придатною ДНК-полімеразою, що виявляє 5'-нуклеазну активність, в даному винаході є термостабільна ДНК-полімераза, одержана з ряду бактеріальних видів, включаючи Thermus aquaticus (Taq), Thermus thermophilus (Tth), Thermus filiformis, Thermus flavus, Thermococcus literalis, Thermus antranikianii, Thermus caldophilus, Thermus chliarophilus, Thermus igniterrae, Thermus lacteus, Thermus oshimai, Thermus ruber, Thermus rubens, Thermus scotoductus, Thermus silvanus, Thermus species Z05, Thermus species sps 17, Thermus thermophilus, Thermotoga maritima, Thermotoga neapolitana, Thermosipho africanus, Thermococcus litoralis, Thermococcus barossi, Thermococcus gorgonarius, Thermotoga maritima, Thermotoga neapolitana, Thermosiphoafricanus, Pyrococcus woesei, Pyrococcus horikoshii, Pyrococcus abyssi, Pyrodictium occultum, Aquifex pyrophilus та. Aquifex aeolieus. Найкраще, щоб термостабільна ДНКполімераза була Taq-полімеразою. Альтернативно, в даному винаході можна використовувати ДНК-полімерази, що виявляють 5'-нуклеазну активність, модифіковані з метою зниження полімеразної активності. Використовувана нуклеаза FEN (флеп-ендонуклеаза) є флеп-специфічною 5'-нуклеазою. Придатна для даного винаходу нуклеаза FEN включає нуклеази FEN, одержані з ряду бактеріальних видів, включаючи Sulfolobus solfataricus, Pyrobaculum aerophilum, Thermococcus litoralis, Archaeaglobus veneficus, Archaeaglobus profundus, Acidianus brierlyi, Acidianus ambivalens, Desulfurococcus amylolyticus, Desulfurococcus mobilis, Pyrodictium brockii, Thermococcus gorgonarius, Thermococcus zilligii, Methanopyrus kandleri, Methanococcus igneus, Pyrococcus horikoshii, Aeropyrum pernix та Archaeaglobus veneficus. Коли розташований "угору по течії" праймер використовують на стадії (а), краще, щоб умови для розщеплення РТО були умовами для здійснення реакції подовження розташованого "угору по течії" праймера. Відповідно до кращого втілення, на стадії (а) використовують розташований "угору по течії"" праймер, для подовження розташованого "угору по течії" праймера використовують матричну полімеразу, і ця матрична полімераза є ідентичною ферменту, що виявляє 5'-нуклеазну активність. В деяких випадках на стадії (а) використовують розташований "угору по течії" праймер, для подовження розташованого "угору по течії" праймера використовують матричну полімеразу, і ця матрична полімераза відрізняється від ферменту, що виявляє 5'-нуклеазну активність. Стадія (с). Гібридизація фрагмента, вивільненого з РТО, з СТО Фрагмент, вивільнений з РТО, гібридизується з СТО (захоплюючим та матричним олігонуклеотидом). СТО містить в напрямку 3'→5' (1) захоплюючу ділянку, яка містить нуклеотидну послідовність, комплементарну 5'-кінцевій ділянці, що мітить, або частині 5'-кінцевої ділянки РТО, що мітить, та (2) матричну ділянку, що містить нуклеотидну послідовність, некомплементарну 5'-кінцевій ділянці, що мітить, та 3'-кінцевій ділянці РТО, що впізнає мішень. СТО діє як матриця для подовження фрагмента, вивільненого з РТО. Цей фрагмент, що служить як праймер, гібридизується з СТО та подовжується з утворенням подовженого дуплекса. Матрична ділянка може містити будь-яку послідовність за умови, що вона не комплементарна 5'-кінцевій ділянці, що мітить, та 3'-кінцевій ділянці РТО, що впізнає мішень. Крім того, матрична ділянка може містити будь-яку послідовність за умови, що вона може діяти як матриця для подовження фрагмента, вивільненого з РТО. Як описано вище, коли вивільняється фрагмент, який містить 5'-кінцеву ділянку РТО, що мітить, краще, щоб захоплююча ділянка СТО була сконструйована так, щоб вона містила 9 UA 115082 C2 5 10 15 20 25 30 35 40 45 50 55 нуклеотидну послідовність, комплементарну 5'-кінцевій ділянці, що мітить. Коли вивільняється фрагмент, який містить 5'-кінцеву ділянку, що мітить, та 5'-кінцеву частину 3'-кінцевої ділянки, що впізнає мішень, краще, щоб захоплююча ділянка СТО була сконструйована так, щоб вона і містила нуклеотидну послідовність, комплементарну 5'-кінцевій ділянці, що мітить, та 5 -кінцевій частині 3'-кінцевої ділянки, що впізнає мішень. Коли вивільняється фрагмент, який містить частину 5'-кінцевої ділянки РТО, що мітить, краще, щоб захоплююча ділянка СТО була сконструйована так, щоб вона містила нуклеотидну послідовність, комплементарну части 5кінцевої ділянки, що мітить. Крім того, можливе конструювання захоплюючої ділянки СТО з передбачуваними сайтами розщеплення РТО. Наприклад, якщо захоплююча ділянка СТО сконструйована так, щоб вона містила нуклеотидну послідовність, комплементарну 5'-кінцевій ділянці, що мітить, тоді або фрагмент, який містить частину 5'-кінцевої ділянки, що мітить, або фрагмент, який містить 5'кінцеву ділянку, що мітить, міг би гібридизуватися із захоплюючою ділянкою і потім подовжуватися. Коли вивільняється фрагмент, який містить 5'-кінцеву ділянку, що мітить, та 5'кінцеву частину 3'-кінцевої ділянки, що впізнає мішень, він може гібридизуватися із захоплюючою ділянкою СТО, сконструйованою так, щоб вона містила нуклеотидну послідовність, комплементарну 5'-кінцевій ділянці, що мітить, і потім успішно подовжуватися, незважаючи на присутність помилково спарених нуклеотидів на 3'-кінцевій ділянці фрагмента. Це можливо тому, що праймери можуть подовжуватися в залежності від реакційних умов, незважаючи на те, що їх 3'-кінець містить деяку кількість помилково спарених нуклеотидів (наприклад, 1-3 помилково спарених нуклеотидів). Коли вивільняється фрагмент, який містить 5'-кінцеву ділянку, що мітить, та 5'-кінцеву частину 3'-кінцевої ділянки, що впізнає мішень, 5'-кінцева частина захоплюючої ділянки СТО може бути сконструйована так, щоб вона містила нуклеотидну послідовність, комплементарну розщеплюваній 5'-кінцевій частині 3'-кінцевої ділянки, що впізнає мішень, в результаті чого долаються проблеми, зв'язані з помилково спареними нуклеотидами (див. Фіг. 1). Краще, нуклеотидна послідовність 5'-кінцевої частини захоплюючої ділянки СТО, комплементарна розщеплюваній 5'-кінцевій частині 3'-кінцевої ділянки, що впізнає мішень, може бути вибрана в залежності від передбачуваних сайтів розщеплення на 3'-кінцевій ділянці РТО, що впізнає мішень. Краще, щоб нуклеотидна послідовність 5'-кінцевої частини захоплюючої ділянки СТО, комплементарна розщеплюваній 5'-кінцевій частині 3'-кінцевої ділянки, що впізнає мішень, складала 1-10 нуклеотидів, ще краще 1-5 нуклеотидів, навіть ще краще, 1-3 нуклеотиди. На 3'-кінці СТО можуть міститися додаткові нуклеотиди, не залучені в гібридизацію з фрагментом. Крім того, захоплююча ділянка СТО може містити нуклеотидну послідовність, комплементарну тільки частині фрагмента (наприклад, частині фрагмента, що містить його 3'кінцеву ділянку), за умови, що вона стабільно гібридизується з цим фрагментом. Використовуваний термін "захоплююча ділянка, що містить нуклеотидну послідовність, комплементарну 5'-кінцевій ділянці, що мітить, або частині 5'-кінцевої ділянки, що мітить", охоплює в даному описі різні конструкції та склади захоплюючої ділянки СТО, як обговорювалося вище. СТО може бути сконструйований так, щоб приймати шпильчасту структуру. Довжина СТО може варіюватися в широких межах. Наприклад, довжина СТО складає 71000 нуклеотидів, 7-500 нуклеотидів, 7-300 нуклеотидів, 7-100 нуклеотидів, 7-80 нуклеотидів, 760 нуклеотидів, 7-40 нуклеотидів, 15-1000 нуклеотидів, 15-500 нуклеотидів, 15-300 нуклеотидів, 15-100 нуклеотидів, 15-80 нуклеотидів, 15-60 нуклеотидів, 15-40 нуклеотидів, 20-1000 нуклеотидів, 20-500 нуклеотидів, 20-300 нуклеотидів, 20-100 нуклеотидів, 20-80 нуклеотидів, 2060 нуклеотидів, 20-40 нуклеотидів, 30-1000 нуклеотидів, 30-500 нуклеотидів, 30-300 нуклеотидів, 30-100 нуклеотидів, 30-80 нуклеотидів, 30-60 нуклеотидів або 30-40 нуклеотидів. Захоплююча ділянка СТО може мати будь-яку довжину за умови, що вона специфічно гібридизується з фрагментом, вивільненим з РТО. Наприклад, довжина захоплюючої ділянки СТО складає 5-100 нуклеотидів, 5-60 нуклеотидів, 5-40 нуклеотидів, 5-30 нуклеотидів, 5-20 нуклеотидів, 10-100 нуклеотидів, 10-60 нуклеотидів, 10-40 нуклеотидів, 10-30 нуклеотидів, 10-20 нуклеотидів, 15-100 нуклеотидів, 15-60 нуклеотидів, 15-40 нуклеотидів, 15-30 нуклеотидів або 15-20 нуклеотидів. Матрична ділянка СТО може мати будь-яку довжину за умови, що вона може діяти як матриця при подовженні фрагмента, вивільненого з РТО. Наприклад, довжина матричної ділянки СТО складає 2-900 нуклеотидів, 2-400 нуклеотидів, 2-300 нуклеотидів, 2-100 нуклеотидів, 2-80 нуклеотидів, 2-60 нуклеотидів, 2-40 нуклеотидів, 2-20 нуклеотидів, 5-900 нуклеотидів, 5-400 нуклеотидів, 5-300 нуклеотидів, 5-100 нуклеотидів, 5-80 нуклеотидів, 5-60 нуклеотидів, 5-40 нуклеотидів, 5-30 нуклеотидів, 10-900 нуклеотидів, 10-400 нуклеотидів, 10-300 нуклеотидів, 15 10 UA 115082 C2 5 10 15 20 25 30 35 40 45 50 55 900 нуклеотидів, 15-100 нуклеотидів, 15-80 нуклеотидів, 15-60 нуклеотидів, 15-40 нуклеотидів або 15-20 нуклеотидів. На 3'-кінці СТО може знаходитися група 3'-ОН. Краще, 3'-кінець СТО блокований, щоб не допустити його подовження. Блокування СТО, яке не допускає подовження, може бути досягнуто традиційними методами. Наприклад, блокування можна здійснити шляхом додавання хімічного угрупування, такого як біотин, мітка, фосфатна група, алкільна група, ненуклеотидний лінкер, фосфоротіонат або алкандіол, до 3'-гідроксильної групи останнього нуклеотиду СТО. Альтернативно, блокування може бути здійснене шляхом видалення 3'-гідроксильної групи останнього нуклеотиду або шляхом використання нуклеотиду, позбавленого 3'-гідроксильної групи, такого як дидезоксинуклеотид. Фрагмент, вивільнений з РТО, гібридизується з СТО, в результаті чого утворюється структура, придатна для подовження фрагмента. Незважаючи на те, що непереварений РТО також гібридизується із захоплюючою ділянкою СТО через свою 5'-кінцеву ділянку, що мітить, його 3'-кінцева ділянка, що впізнає мішень, не гібридизується з СТО, що перешкоджає утворенню подовженого дуплекса. Гібридизація стадії (с) може бути описана більш детально з посиланням на описи, наведені на стадії (а). Стадія (d). Подовження фрагмента Реакцію подовження проводять, використовуючи продукт зі стадії (с) та матричну полімеразу нуклеїнових кислот. Фрагмент, гібридизований із захоплюючою ділянкою СТО, подовжується з утворенням подовженого дуплекса. На відміну від цього, нерозщеплений РТО, гібридизований із захоплюючою ділянкою СТО, не подовжується, і в результаті цього подовжений дуплекс не утворюється. Використовуваний в даному описі термін "подовжений дуплекс" позначає дуплекс, утворений шляхом реакції подовження, у якій фрагмент, гібридизований із захоплюючою ділянкою СТО, подовжується з використанням матричної ділянки СТО як матриці та матричної полімерази нуклеїнових кислот. Подовжений дуплекс має величину Т пл, що відрізняється від такої для гібрида, утвореного нерозщепленим РТО та СТО. Краще, щоб подовжений дуплекс мав більш високу величину Т пл, ніж гібрид, утворений нерозщепленим РТО та СТО. Величина Тпл такого подовженого дуплекса регулюється в залежності від (1) послідовності та/або довжини фрагмента, (2) послідовності та/або довжини СТО, або (3) послідовності та/або довжини фрагмента та послідовності та/або довжини СТО. Дивовижною ознакою даного винаходу є те, що можливість регулювання величини Т пл. подовженого дуплекса використовується для одержання сигналу від мішені, що вказує на присутність подовженого дуплекса, в результаті плавлення подовженого дуплекса на стадії (е). Використовуваний в даному описі термін "Т пл." стосується температури плавлення, при якій половина популяції дволанцюгових молекул нуклеїнової кислоти дисоціює до одноланцюгових молекул. Величина Тпл. визначається довжиною та вмістом G/C-пар в гібридизованих нуклеотидах. Величина Тпл може бути розрахована традиційними методами, такими як правило Wallace (R.B. Wallace et al, Nucleic Acids Research, 6: 3543-3547(1979)) та метод найближчих сусідів (SantaLucia J. Jr., et al., Biochemistry, 35: 3555-3562(1996)); Sugimoto N… et al., Nucleic Acids Res., 24: 4501-4505(1996)). Відповідно до кращого втілення, величина Т пл. стосується фактичних величин Т пл. в реакційних умовах, реально використовуваних на практиці. Матрична полімераза нуклеїнових кислот, використовувана на стадії (d), може включати будь-яку полімеразу нуклеїнових кислот, наприклад, кленовський фрагмент ДНК-полімерази І з Е. соlі, термостабільну ДНК-полімеразу та ДНК-полімеразу бактеріофага Т7. Краще, полімераза є термостабільною ДНК-полімеразою, яка може бути одержана з ряду бактеріальних видів, включаючи Thermus aquaticus (Taq), Thermus thermophilus (Tth), Thermusfiliformis, Thermis flavus, Thermococcus literalis, Thermus antranikianii, Thermus caldophilus, Thermus chliarophilus, Thermus flavus, Thermus igniterrae, Thermus lacteus, Thermus oshimai, Thermus ruber, Thermus rubens, Thermus scotoductus, Thermus silvanus, Thermus species Z05, Thermus species sps 17, Thermus thermophilus, Thermotoga maritima, Thermotoga neapolitana, Thermosipho africanus, Thermococcus litoralis, Thermococcus barossi, Thermococcus gorgonarius, Thermotoga maritima, Thermotoga neapolitana, Thermosiphoafricanus, Pyrococcus furiosus (Pfu), Pyrococcus woesei, Pyrococcus horikoshii, Pyrococcus abyssi, Pyrodictium occultum, Aquifex pyrophilus та Aquifex aeolieus. Найкраще, матрична полімераза нуклеїнових кислот є полімеразою Taq. Найкраще, щоб матрична полімераза нуклеїнових кислот була полімеразою Taq. 11 UA 115082 C2 5 10 15 20 25 30 35 40 45 50 55 60 Відповідно до кращого втілення, фермент, що виявляє 5'-нуклеазну активність, використовуваний на стадії (b), є ідентичним матричній полімеразі нуклеїнових кислот, використовуваній на стадії (d). ще краще, щоб фермент, що виявляє 5'-нуклеазну активність, використовуваний на стадії (b), матрична полімераза нуклеїнових кислот, використовувана для подовження розташованого "угору по течії" праймера, та матрична полімераза нуклеїнових кислот, використовувана на стадії (d), були ідентичні один одному. Подовжений дуплекс містить мітку, що походить з (1) принаймні однієї мітки, з'єднаної з фрагментом РТО та/або з СТО, (2) мітки, вбудовуваної в подовжений дуплекс під час реакції подовження, (3) мітки, вбудовуваної в подовжений дуплекс під час реакції подовження, та мітки, з'єднаної з фрагментом РТО та/або з СТО, або (4) інтеркалюючої мітки. Присутність подовженого дуплекса може вказувати на присутність нуклеїновокислотної послідовності-мішені, тому що подовжений дуплекс утворюється, коли присутня нуклеїновокислотна послідовність-мішень. Щоб детекцію присутності подовженого дуплекса проводити безпосередньо, утворення подовженого дуплекса, що містить мітку, яка забезпечує одержання детектованого сигналу, здійснюють на стадії (d). Мітка, використовувана в подовженому дуплексі, забезпечує реєстрацію зміни сигналу в залежності від того, чи є подовжений дуплекс дволанцюговою структурою або одноланцюговою структурою, в кінцевому підсумку надаючи сигнал від мішені, який вказує на присутність подовженого дуплекса, в результаті плавлення цього подовженого дуплекса. Стадія (e). Плавлення подовженого дуплекса Після закінчення реакції подовження подовжений дуплекс піддають плавленню в діапазоні температур з одержанням сигналу від мішені, що вказує на присутність подовженого дуплекса. Сигнал від мішені забезпечується за допомогою (1) принаймні однієї мітки, з'єднаної з фрагментом та/або СТО, (2) мітки, вбудовуваної в подовжений дуплекс під час реакції подовження, (3) мітки, вбудовуваної в подовжений дуплекс під час реакції подовження, та мітки, з'єднаної з фрагментом та/або СТО, або (4) інтеркалюючої мітки. Використовуваний в даному описі термін "сигнал від мішені" означає будь-який сигнал, здатний вказати на присутність подовженого дуплекса. Наприклад, термін "сигнал від мішені" включає сигнал від міток (генерацію або гасіння сигналу), зміну сигналу від міток (підсилення або ослаблення сигналу), криву плавлення, картину плавлення та температуру плавлення (або величину Тпл.). Відповідно до кращого втілення, сигнал від мішені є зміною сигналу від мітки на подовженому дуплексі на стадії плавлення. Зміна сигналу може бути одержана шляхом вимірювання сигналів не менш ніж при двох різних температурах. Альтернативно, сигнал від мішені є кривою плавлення, картиною плавлення та температурою плавлення (або величиною Тпл), одержаними шляхом вимірювання сигналів від мітки на подовженому дуплексі в діапазоні температур. Краще, щоб діапазон температур був діапазоном температур для аналізу кривої плавлення або був температурою, близькою до величини Т пл подовженого дуплекса. Подовжений дуплекс має більш високу величину Т пл, ніж гібрид, утворений нерозщепленим РТО та СТО. Зважаючи на це, подовжений дуплекс та гібрид демонструють відмінні одну від одної картини плавлення. Такі різні картини плавлення дозволяють провести відмінність сигналу від мішені від сигналів, що не належать до мішені. Інша картина плавлення або температура плавлення описує сигнал від мішені спільно з придатною системою міток. Плавлення може бути здійснене з використанням традиційних технологій, включаючи, без обмеження, нагрівання, обробку лугом, формамідом, сечовиною та гліоксалем, ферментативні методи (наприклад, дія гелікази) та застосування зв'язуючих білків, але без обмеження ними. Наприклад, плавлення можна досягти шляхом нагрівання при температурі, що змінюється в діапазоні від 80 °C до 105 °C. Загальні способи здійснення такої обробки наведені в Joseph Sambrook, et al., Molecular Cloning, A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. (2001). Придатні системи міток, використовувані в цьому винаході, відрізняються з погляду їх типів, розташування та способу генерації сигналу. Системи міток, корисні в цьому винаході, будуть обговорені більш детально нижче. (1) Мітка, з'єднана з фрагментом та/або СТО Відповідно до кращого втілення, одержання сигналу від мішені забезпечується принаймні однією міткою, з'єднаною з фрагментом та/або СТО. Як тільки відбудеться утворення подовженого дуплекса між фрагментом РТО та СТО, на подовженому дуплексі з'явиться мітка або від фрагмента РТО, або від СТО, яка забезпечує сигнал від мішені на стадії плавлення. Така мітка включає систему двох взаємодіючих міток та одиночну мітку. (1-1) Система двох взаємодіючих міток 12 UA 115082 C2 5 10 15 20 25 30 35 40 45 50 55 60 Система взаємодіючих міток є генеруючою сигнал системою, у якій енергія передається від донорної молекули акцепторній молекулі без участі радіоактивності. Як репрезентативна система взаємодіючих міток, система міток при FRET (резонансному перенесенні енергії флуоресценції) включає флуоресцентну репортерну молекулу (донорну молекулу) та молекулугасник (акцепторну молекулу). При FRET донор енергії є флуоресцентним, а акцептор енергії може бути флуоресцентним або не бути флуоресцентним. Для іншої форми систем взаємодіючих міток донор енергії не є флуоресцентним, наприклад є хромофором, а акцептор енергії є флуоресцентним. Для ще однієї іншої форми систем взаємодіючих міток донор енергії є люмінесцентним, наприклад, біолюмінесцентним, хемілюмінесцентним, електрохемілюмінесцентним, а акцептор є флуоресцентним. Донорна молекула та акцепторна молекула можуть бути описані в даному винаході як репортерна молекула та молекула-гасник, відповідно. Краще, щоб сигнал, який вказує на присутність подовженого дуплекса (тобто, на присутність нуклеїновокислотної послідовності-мішені), генерувався системами взаємодіючих міток, ще краще, системою FRET-міток (тобто, системою двох взаємодіючих міток). Перше втілення (внутрішньоланцюгова система двох взаємодіючих міток) В першому втіленні системи двох взаємодіючих міток фрагмент або СТО містить систему двох взаємодіючих міток, яка складається з репортерної молекули та молекули-гасника; при цьому плавлення подовженого дуплекса на стадії (е) індукує зміну сигналу від системи двох взаємодіючих міток з одержанням сигналу від мішені на стадії (е). Перше втілення такої системи двох взаємодіючих міток проілюстроване на Фіг. 2, 6 та 9. Перше втілення називається внутрішньоланцюговою системою двох взаємодіючих міток. Перше втілення, наведене на Фіг 2 (внутрішньоланцюгова система двох взаємодіючих міток) Пояснене прикладом втілення описане з посиланням на Фіг. 2. Матрична ділянка СТО містить репортерну молекулу та молекулу-гасник. РТО, гібридизований з нуклеїновокислотною послідовністю-мішенню, переварюється, вивільняючи фрагмент, цей фрагмент гібридизується із захоплюючою ділянкою СТО та подовжується з утворенням подовженого дуплекса. Якщо на стадії (d) утворюється подовжений дуплекс, то репортерна молекула та молекулагасник на СТО конформаційно розділяються, що не дозволяє молекулі-гаснику гасити сигнал від репортерної молекули; при цьому, якщо на стадії (є) цей подовжений дуплекс піддають плавленню, то репортерна молекула та молекула-гасник конформаційно розташовуються близько одна до одної, що дозволяє молекулі-гаснику гасити сигнал від репортерної молекули, так що доставляється сигнал від мішені, що вказує на присутність подовженого дуплекса на стадії (е). Використовуваний в даному описі вираз "репортерна молекула та молекула-гасник конформаційно розташовуються близько одна до одної" означає, що репортерна молекула та молекула-гасник просторово наближені одна до одної завдяки конформаційній структурі фрагмента або СТО, такої як випадкова спіраль та шпильчаста структура. Використовуваний в даному описі вираз "репортерна молекула та молекула-гасник конформаційно розділяються" означає, що репортерна молекула та молекула-гасник просторово розділяються в результаті зміни конформаційної структури фрагмента або СТО після утворення подвійного ланцюга Краще, щоб сигнал від мішені, що доставляється на стадії (е), був кривою плавлення, картиною плавлення або величиною Т пл. одержуваною шляхом вимірювання зміни сигналу флуоресценції, генерованого на стадії (d). Відповідно до кращого втілення, репортерна молекула та молекула-гасник можуть бути розташовані в будь-якому місці на СТО, за умови, що гасіння та негасіння сигналу від репортерної молекули здійснюється в залежності від плавлення подовженого дуплекса. Відповідно до кращого втілення, репортерна молекула та молекула-гасник обидві з'єднані з матричною ділянкою або із захоплюючою ділянкою СТО. Відповідно до кращого втілення, репортерна молекула та молекула-гасник розташовані на 5'-кінці та 3'-кінці СТО. Відповідно до кращого втілення, одна з молекул на СТО, репортерна молекула або молекула-гасник, розташована на його 5'-кінці або на відстані 1-5 нуклеотидів від його 5'-кінця, а інша розташована так, щоб гасіння та негасіння сигналу від репортерної молекули здійснювалося в залежності від конформації СТО. Відповідно до кращого втілення, одна з молекул на СТО, репортерна молекула або молекула-гасник, розташована на його 3'-кінці або на відстані 1-5 нуклеотидів від його 3'-кінця, а інша розташована так, щоб гасіння та негасіння сигналу від репортерної молекули здійснювалося в залежності від конформації СТО. 13 UA 115082 C2 5 10 15 20 25 30 35 40 45 50 55 60 Відповідно до кращого втілення, репортерна молекула та молекула-гасник розташовані на відстані не більше 80 нуклеотидів, ще краще, не більше 60 нуклеотидів, навіть ще краще, не більше 30 нуклеотидів, ще істотно краще, не більше 25 нуклеотидів одна від одної. Відповідно до кращого втілення, репортерна молекула та молекула-гасник розділені принаймні 4 нуклеотидами, ще краще, принаймні 6 нуклеотидами, навіть ще краще, принаймні 10 нуклеотидами, ще істотно краще, принаймні 15 нуклеотидами. В даному винаході можливе одержання гібрида, утвореного нерозщепленим РТО та СТО. Якщо матрична ділянка СТО помічена з використанням системи двох взаємодіючих міток, як показано на Фіг. 2, то індукції зміни сигналу від мітки на гібриді, утвореному нерозщепленим РТО та СТО, не відбувається. Внаслідок цього гібрид не дає сигналу, який не відноситься до мішені. Якщо захоплююча ділянка СТО помічена з використанням системи двох взаємодіючих міток, то гібрид, утворений нерозщепленим РТО та СТО, забезпечує одержання сигналу, який не відноситься до мішені, на стадії плавлення. У цьому випадку різниця у величинах Т пл для подовженого дуплекса та гібрида дозволяє відрізнити сигнал від подовженого дуплекса, що відноситься до мішені, від сигналу гібриду, який не належить до мішені. Перше втілення, наведене на Фіг 6 (внутрішньоланцюгова система двох взаємодіючих міток) Пояснене прикладом втілення описане з посиланням на Фіг. 6. 5'-Кінцева ділянка РТО, що містить репортерну молекулу та молекулу-гасник. РТО, гібридизований з нуклеїновокислотною послідовністю-мішенню, переварюється, вивільняючи фрагмент, який містить 5'-кінцеву ділянку, що мітить, з репортерною молекулою та молекулою-гасником. Цей фрагмент гібридизується із захоплюючою ділянкою СТО. Якщо на стадії (d) утворюється подовжений дуплекс, то репортерна молекула та молекулагасник на фрагменті конформаційно розділяються, що не дозволяє молекулі-гаснику гасити сигнал від репортерної молекули; при цьому, якщо на стадії (е) цей подовжений дуплекс піддають плавленню, то репортерна молекула та молекула-гасник конформаційно розташовуються близько одна до одної, що дозволяє молекулі-гаснику гасити сигнал від репортерної молекули, так що доставляється сигнал від мішені, що вказує на присутність подовженого дуплекса на стадії (е). Відповідно до кращого втілення, репортерна молекула та молекула-гасник можуть бути розташовані в будь-якому місці на фрагменті, за умови, що гасіння та негасіння сигналу від репортерної молекули здійснюється в залежності від плавлення подовженого дуплекса. Відповідно до кращого втілення, одна з молекул на фрагменті, репортерна молекула або молекула-гасник, розташована на його 5'-кінці або на відстані 1-5 нуклеотидів від його 5'-кінця, а інша розташована так, щоб гасіння та негасіння сигналу від репортерної молекули здійснювалося в залежності від конформації фрагмента. Відповідно до кращого втілення, репортерна молекула та молекула-гасник розташовані на відстані не більше 50 нуклеотидів, ще краще, не більше 40 нуклеотидів, навіть ще краще, не більше 30 нуклеотидів, ще істотно краще, не більше 20 нуклеотидів одна від одної. Відповідно до кращого втілення, репортерна молекула та молекула-гасник розділені принаймні 4 нуклеотидами, ще краще, принаймні 6 нуклеотидами, навіть ще краще, принаймні 10 нуклеотидами, ще істотно краще, принаймні 15 нуклеотидами. Як представлено на Фіг. 6, гібрид, утворений нерозщепленим РТО та СТО, забезпечує одержання сигналу, який не належить до мішені, на стадії плавлення. У цьому випадку різниця у величинах Т пл. для подовженого дуплекса та гібриду дозволяє відрізнити сигнал від подовженого дуплекса, що належить до мішені, від сигналу гібриду, який не належить до мішені. Друге втілення (міжланцюгова система двох взаємодіючих міток) В другому втіленні системи взаємодіючих міток фрагмент містить одну з двох взаємодіючих міток, що включають репортерну молекулу та молекулу-гасник, а СТО містить іншу з двох взаємодіючих міток; при цьому плавлення подовженого дуплекса на стадії (е) індукує зміну сигналу від системи двох взаємодіючих міток з одержанням сигналу від мішені на стадії (е). Пояснене прикладом втілення описане з посиланням на Фіг. 8. Якщо на стадії (d) утворюється подовжений дуплекс, то сигнал від репортерної молекули, з'єднаної з СТО, гаситься молекулою-гасником, з'єднаною з РТО. Якщо на стадії (е) подовжений дуплекс піддають плавленню, то репортерна молекула та молекула-гасник розділяються, що не дозволяє молекулі-гаснику гасити сигнал від репортерної молекули, так що доставляється сигнал від мішені, що вказує на присутність подовженого дуплекса на стадії (е). Краще, щоб сигнал від мішені, що доставляється на стадії (е), був кривою плавлення, картиною плавлення або величиною Т пл., одержаними шляхом вимірювання зміни сигналу флуоресценції від системи двох взаємодіючих міток. 14 UA 115082 C2 5 10 15 20 25 30 35 40 45 50 55 60 Репортерна молекула та молекула-гасник можуть бути розташовані в будь-якому місці фрагмента РТО та СТО, за умови, що сигнал від репортерної молекули гаситься молекулоюгасником в подовженому дуплексі. Відповідно до кращого втілення, репортерна молекула або молекула-гасник на фрагменті РТО розташована на 5'-кінці 5'-кінцевої ділянки, що мітить. Відповідно до кращого втілення, репортерна молекула або молекула-гасник на СТО розташована на його 3'-кінці. Як представлено на Фіг. 8, гібрид, утворений нерозщепленим РТО та СТО, забезпечує одержання сигналу, який не належить до мішені, на стадії плавлення. У цьому випадку різниця у величинах Т пл. для подовженого дуплекса та гібриду дозволяє відрізнити сигнал від подовженого дуплекса, що належить до мішені, від сигналу гібриду, який не належить до мішені. Репортерна молекула та молекула-гасник, корисні в даному винаході, можуть включати будь-які молекули, відомі в даній області техніки. Їх прикладами є такі: Су2™ (506), YO-PRO™-1 (509), YOYO™-1 (509), кальцеїн (517), FITC (флуоресцеїнізотіоціанат) (518), FluorX™ (519), Alexa™ (520), родамін ПО (520), Oregon Green™ (орегон зелений) 500 (522), Oregon Green™ 488 (524), RiboGreen™ (525), Rhodamine Green™ (родаміновий зелений) (527), родамін 123 (529), Magnesium Green™ (531), Calcium Green™ (533), TO-PRO™-1 (533), TOTO1 (533), JOE (2'7'-диметокси4'-5'-дихлорофлуоресцеїн) (548), BODIPY530/550 (550), Dil (565), BODIPY TMR (568), BODIPY558/568 (568), BODIPY564/570 (570), Cy3™ (570), Alexa™ 546 (570), TRITC (тетраметилродамін-ізотіоціанат) (572), Magnesium Orange™ (575), фікоеритрин R&B (575), родамін-фалоїдин (575), Calcium Orange™ (576), піронін Y (580), родамін В (580), TAMRA (тетраметилродамін) (582), Rhodamine Red™ (родамін червоний) (590), СуЗ.5™ (596), ROX (6карбокси-Х-родамін) (608), Calcium Crimson™ (615), Alexa™ 594 (615), техаський червоний (615), нільський червоний (628), YO-PRO™-3 (631), YOYO™-3 (631), R-фікоціанін (642), Cфікоціанін (648), TO-PRO™-3 (660), TOTO3 (660), DiD DilC(5) (665), Cy5™ (670), тіадикарбоціанін (671) та Су5.5 (694), HEX (556), ТЕТ (536), Biosearch Blue (447), CAL Fluor Gold 540 (544), CAL Fluor Orange 560 (559), CAL Fluor Red 590 (591), CAL Fluor Red 610 (610), CAL Fluor Red 635 (637), FAM (карбоксифлуоресцеїн) (520), флуоресцеїн (520), флуоресцеїн-СЗ (520), Pulsar 650 (566), Quasar 570 (667), Quasar 670 (705) та Quasar 705 (610). Числа в дужках є довжиною хвилі, що відповідає максимуму випромінювання, в нанометрах. Краще, щоб репортерна молекула та молекула-гасник були JOE, FAM, TAMRA, ROX та іншою міткою на основі флуоресцеїну. Придатні пари репортер-гасник описані в ряді публікацій, які вказані нижче: Pesce et al, editors, Fluorescence Spectroscopy (Marcel Dekker, New York, 1971); White et al., Fluorescence Analysis: A Practical Approach (Marcel Dekker, New York, 1970); Berlman, Handbook of nd Fluorescence Spectra of Aromatic Molecules, 2 Edition (Academic Press, New York, 1971); Griffiths, Color and Constitution of Organic Molecules (Academic Press, New York, 1976); Bishop, editor, Indicators (Pergamon Press, Oxford, 1972); Haugland, Handbook of Fluorescent Probes and Research Chemicals (Molecular Probes, Eugene, 1992); Pringsheim, Fluorescence and Phosphorescence (Interscience Publishers, New York, 1949); Haugland, R. P., Handbook of th Fluorescent Probes and Research Chemicals, 6 Edition, Molecular Probes, Eugene, Oreg., 1996; патенти США №№ 3996345 та 4351760. Варто відзначити, що в даному винаході може бути використана нефлуоресцентна "чорна" молекула-гасник, здатна гасити флуоресценцію в широкому діапазоні довжин хвиль або на конкретній довжині хвилі. Прикладами таких гасників є BHQ (Black Hole 1) та DABCYL (4-((4(диметиламіно)феніл)азо)бензойна кислота). Для FRET-міток, адаптованих до СТО, термін "репортерна молекула" включає донора FRET, а термін "гасник" включає іншого партнера (акцептора) FRET. Наприклад, як репортерну молекулу використовують флуоресцентний барвник, а як гасник - родаміновий барвник. (1-2) Одиночна мітка Даний винахід також чудово здійснюється з використанням систем із одиночною міткою для одержання сигналів, що вказують на присутність нуклеїновокислотних послідовностей-мішеней. Відповідно до кращого втілення, фрагмент або СТО містить одиночну мітку, а плавлення подовженого дуплекса на стадії (є) індукує зміну сигналу від цієї одиночної мітки з одержанням сигналу від мішені на стадії (є). Перше втілення, наведене на Фіг. 3 (система з одиночною міткою) Пояснене прикладом втілення описане з посиланням на Фіг. 3. Матрична ділянка СТО містить одиночну флуоресцентну мітку. РТО, гібридизований з нуклеїновокислотною послідовністю-мішенню, переварюється, вивільняючи фрагмент. Цей фрагмент гібридизується із захоплюючою ділянкою СТО та подовжується з утворенням подовженого дуплекса. В 15 UA 115082 C2 5 10 15 20 25 30 35 40 45 50 55 60 результаті утворення подовженого дуплекса інтенсивність флуоресценції від одиночної флуоресцентної мітки збільшується. Якщо на стадії (е) подовжений дуплекс піддають плавленню, то інтенсивність флуоресценції від одиночної флуоресцентної мітки зменшується, так що доставляється сигнал від мішені, що вказує наприсушість подовженого дуплекса на стадії (е). Відповідно до кращого втілення, одиночна мітка може бути розташована в будь-якому місці СТО, за умови, що рівень сигналу від одиночної мітки змінюється в залежності від плавлення подовженого дуплекса. Відповідно до кращого втілення, одиночна мітка з'єднана з матричною ділянкою або із захоплюючою ділянкою СТО. Якщо матрична ділянка СТО помічена одиночною міткою, як показано на Фіг.3, то індукції зміни сигналу від мітки на гібриді, утвореному нерозщепленим РТО та СТО, не відбувається. Внаслідок цього гібрид не дає сигналу, який не належить до мішені. Якщо захоплююча ділянка СТО помічена одиночною міткою, то гібрид, утворений нерозщепленим РТО та СТО, забезпечує одержання сигналу, який не належить до мішені, на стадії плавлення. У цьому випадку різниця у величинах Т пл для подовженого дуплекса та гібрида дозволяє відрізнити сигнал від подовженого дуплекса, що належить до мішені, від сигналу гібриду, який не належить до мішені. Друге втілення, наведене на Фіг. 7 (система з одиночною міткою) Пояснене прикладом втілення описане з посиланням на Фіг. 7. 5'-Кінцева ділянка РТО, що мітить, одиночну флуоресцентну мітку. РТО, гібридизований з нуклеїновокислотною послідовністю-мішенню, переварюється, вивільняючи фрагмент, який містить 5'-кінцеву ділянку, що мітить, з одиночною флуоресцентною міткою. В результаті гібридизації інтенсивність сигналу від одиночної флуоресцентної мітки на 5'-кінцевій ділянці, що мітить, збільшується. Якщо на стадії (е) подовжений дуплекс піддають плавленню, то інтенсивність сигналу від одиночної флуоресцентної мітки зменшується, так що доставляється сигнал від мішені, що вказує на присутність подовженого дуплекса на стадії (е). Відповідно до кращого втілення, одиночна мітка може бути розташована в будь-якому місці фрагмента РТО, за умови, що рівень сигналу від одиночної мітки змінюється в залежності від плавлення подовженого дуплекса. Як представлено на Фіг. 7, гібрид, утворений нерозщепленим РТО та СТО, забезпечує одержання сигналу, який не належить до мішені, на стадії плавлення. У цьому випадку різниця у величинах Т пл. для подовженого дуплекса та гібриду дозволяє відрізнити сигнал від подовженого дуплекса, що належить до мішені, від сигналу гібриду, який не належить до мішені. Використання в даному винаході одиночної мітки має забезпечити одержання різних сигналів в залежності від її присутності на дволанцюговій або одноланцюговій структурі. Одиночна мітка є флуоресцентною міткою, люмінесцентною міткою, хемілюмінесцентною міткою, електрохімічною міткою та металічною міткою. Краще, щоб одиночна мітка була флуоресцентною міткою. Типи та кращі сайти зв'язування одиночних флуоресцентних міток, використовуваних в даному винаході, описані в патентах США №№ 7537886 та 7348141, ідеї яких включені до даного опису шляхом посилання в усій своїй повноті. Краще, щоб одиночна флуоресцентна мітка була JOE, FAM, TAMRA, ROX та іншою міткою на основі флуоресцеїну. Мічений нуклеотидний залишок краще розташований в позиції внутрішнього нуклеотидного залишку в межах олігонуклеотиду, а не на 5'-кінці або 3'-кінці. Одиночна мітка, використовувана в даному винаході, може бути описана з посиланням на вказані вище описи для репортерної молекули та молекули-гасника. Зокрема, якщо даний винахід здійснюють на твердій фазі із застосуванням одиночної мітки, то можна використовувати звичайну флуоресцентну мітку, і не потрібно спеціальної флуоресцентної мітки, здатної забезпечити одержання сигналів флуоресценції з різними інтенсивностями в залежності від її присутності на дволанцюговій або одноланцюговій структурі. Сигнал від мішені, створюваний на твердій підкладці, вимірюють. Таке втілення системи з одиночною міткою разом з іммобілізованим СТО проілюстроване на Фіг. 12. В тому випадку, коли застосовують СТО, іммобілізований на твердій підкладці, можна використовувати хімічні мітки (наприклад, біотин) або ферментативні мітки (наприклад, лужну фосфатазу, пероксидазу, -галактозидазу та -глюкозидазу). В системах з міткою, що використовують "мітку, з'єднану з фрагментом та/або СТО", ці мітки можуть бути розташовані таким чином, що, якщо утворюється гібрид, який складається з нерозщепленого РТО та СТО, то цей гібрид на стадії (е) не дає сигналу, який не належить до мішені. Альтернативно, мітки можуть бути розташовані таким чином, що, якщо утворюється 16 UA 115082 C2 5 10 15 20 25 30 35 40 45 50 55 гібрид, який складається з нерозщепленого РТО та СТО, то цей гібрид на стадії (е) дає сигнал, який не належить до мішені; при цьому величина Т пл. для подовженого дуплекса більше такої для гібриду, утвореного нерозщепленим РТО та СТО. Зокрема, якщо мітки розташовані таким чином, що гібрид, утворений нерозщепленим РТО та СТО, не дає сигналу, який не належить до мішені, то для вибору величини Т пл. подовженого дуплекса з метою детекції нуклеїновокислотної послідовності-мішені можна використовувати діапазон, що включає величину Т пл. для гібриду. (2) Мітка, вбудовувана в подовжений дуплекс В даному винаході можна застосовувати мітку, вбудовувану в подовжений дуплекс під час реакції подовження, для одержання сигналу від мішені, що вказує на присутність подовженого дуплекса. Незважаючи на те, що фрагмент РТО або СТО не містить жодної мітки, успішно використовується мітка, вбудовувана в подовжений дуплекс під час реакції подовження, що дає можливість зробити подовжений дуплекс міченим. На Фіг. 10 та 11 проілюстроване втілення, у якому нуклеотид з одиночною міткою вбудовується в подовжений дуплекс під час реакції подовження (див. С та D на Фіг. 10 та 11). Це втілення також є застосовним до інших втілень з використанням аналізу плавлення. Відповідно до кращого втілення, одержання сигналу від мішені забезпечується за допомогою одиночної мітки, вбудовуваної в подовжений дуплекс під час реакції подовження; причому вбудовувана одиночна мітка з'єднана з нуклеотидом, вбудовуваним під час реакції подовження; при цьому плавлення подовженого дуплекса на стадії (е) індукує зміну сигналу від одиночної мітки з одержанням сигналу від мішені на стадії (е). Пояснене прикладом втілення описане з посиланням на Фіг. 10. РТО, гібридизований з нуклеїновокислотною послідовністю-мішенню, переварюється, вивільняючи фрагмент. Цей фрагмент гібридизується із захоплюючою ділянкою СТО, іммобілізованого на твердій підкладці, та подовжується в присутності нуклеотидів, мічених одиночною флуоресцентною міткою, з утворенням подовженого дуплекса. Флуоресцентний сигнал від подовженого дуплекса можна детектувати у плямі на твердій підкладці з іммобілізованим СТО. Коли подовжений дуплекс піддають плавленню, ланцюг, що містить флуоресцентну мітку, вивільняється, та флуоресцентний сигнал в цій плямі більше не детектується (на Фіг. 10 не показано). Таким чином, зміна сигналу у плямі може бути одержана шляхом плавлення подовженого дуплекса. При цьому доставляється сигнал від мішені, що вказує на присутність подовженого дуплекса на стадії (е). Сигнал від мішені, що доставляється на стадії (е), є кривою плавлення, картиною плавлення або величиною Тпл., одержаними шляхом вимірювання зміни сигналу флуоресценції у плямі з іммобілізованим СТО. Відповідно до кращого втілення, нуклеотид, вбудовуваний під час реакції подовження, містить першу неприродну основу, а СТО містить нуклеотид, що містить другу неприродну основу з афінністю специфічного зв'язування (specific binding affinity) з першою неприродною основою, як показано на Фіг. 11. Нуклеотид, що містить другу неприродну основу, краще розташований в будь-якому сайті на матричній ділянці СТО. Використовуваний в даному описі термін "неприродна основа" стосується похідних природних основ, таких як аденін (А), гуанін (G), тимін (Т), цитозин (С) та урацил (U), які здатні утворювати зв'язані водневими зв'язками пари основ. Використовуваний в даному описі термін "неприродна основа" включає основи, які характеризуються іншими картинами утворення пар основ у порівнянні з природними основами як вихідними сполуками (mother compounds), як описано, наприклад, в патентах США №№ 5432272, 5965364, 6001983 та 6037120. Утворення пар основ між неприродними основами полягає в утворенні двох або трьох водневих зв'язків, як і у природних основ. Утворення пар основ між неприродними основами також відбувається з урахуванням специфічності. Конкретні приклади неприродних основ включають такі основи в комбінаціях пар основ ізоС/ізо-G, ізо-dС/ізо-dG, К/Х, H/J та M/N (див. патент США № 7422850). Пояснене прикладом втілення описане з посиланням на Фіг. 11. Фрагмент гібридизується з СТО, що містить нуклеотид, який має другу неприродну основу (наприклад, ізо-dC) з афінністю специфічного зв'язування з першою неприродною основою (наприклад, ізо-dG). Подовження здійснюють в присутності нуклеотиду, що містить першу неприродну основу, мічену одиночною флуоресцентною міткою, з утворенням подовженого дуплекса. В реакції подовження нуклеотид, що містить першу неприродну основу, вбудовується в сайт, розташований навпроти нуклеотиду, що містить другу неприродну основу. 17 UA 115082 C2 5 10 15 20 25 30 35 40 45 50 55 Флуоресцентний сигнал від подовженого дуплекса можна детектувати у плямі на твердій підкладці з іммобілізованим СТО. Коли подовжений дуплекс піддають плавленню, ланцюг, що містить флуоресцентну мітку, вивільняється, і флуоресцентний сигнал у цій плямі більше не детектується (на Фіг. 11 не показано). Таким чином, зміна сигналу у плямі може бути одержана шляхом плавлення подовженого дуплекса. При цьому доставляється сигнал від мішені, що вказує на присутність подовженого дуплекса на стадії (e). Якщо застосовується мітка, вбудовувана в подовжений дуплекс під час реакції подовження, то ця мітка не вбудовується в гібрид, утворений нерозщепленим РТО та СТО, оскільки гібрид не подовжується. Таким чином, даний гібрид не дає сигналу, який не належить до мішені. Типи та характеристики використовуваних одиночних міток можуть бути описані з посиланням на описи для системи міток, у якій використовується "мітка, з’єднана з фрагментом та/або СТО", як зазначено вище. (3) Мітка, вбудовувана в подовжений дуплекс, та мітка, з'єднана з фрагментам або СТО В даному винаході можна застосовувати систему міток, у якій використовується спільна дія мітки, вбудовуваної в подовжений дуплекс під час реакції подовження, та мітки, з'єднаної з фрагментом та/або СТО, як показано на Фіг. 4 та 5. Відповідно до кращого втілення, одержання сигналу від мішені забезпечується за допомогою мітки, вбудовуваної в подовжений дуплекс під час реакції подовження, та мітки, з'єднаної з фрагментом та/або СТО, і така вбудовувана мітка з'єднана з нуклеотидом, вбудовуваним під час реакції подовження; причому ці дві мітки є системою двох взаємодіючих міток, яка складається з репортерної молекули та молекули-гасника; при цьому плавлення подовженого дуплекса на стадії (е) індукує зміну сигналу від цієї системи двох взаємодіючих міток з одержанням сигналу від мішені на стадії (е). Краще, щоб нуклеотид, вбудовуваний під час реакції подовження, містив першу неприродну основу, а СТО містив нуклеотид, що містить другу неприродну основу з афінністю специфічного зв'язування з першою неприродною основою. Пояснене прикладом втілення описане з посиланням на Фіг. 4. Фрагмент гібридизується з СТО, що містить репортерну молекулу або молекулу-гасник та нуклеотид, який має другу неприродну основу (наприклад, ізо-dC) з афінністю специфічного зв'язування з першою неприродною основою (наприклад, ізо-dG). Подовження здійснюють в присутності нуклеотиду, що містить першу неприродну основу, мічену молекулою-гасником або репортерною молекулою, з утворенням подовженого дуплекса, у якому сигнал від репортерної молекули гаситься молекулою-гасником. В реакції подовження нуклеотид, що містить першу неприродну основу, вбудовується в сайт, розташований навпроти нуклеотиду, що містить другу неприродну основу. Якщо на стадії (е) подовжений дуплекс піддають плавленню, то репортерна молекула та молекула-гасник розділяються, що не дозволяє молекулі-гаснику гасити сигнал від репортерної молекули, так що доставляється сигнал від мішені, який вказує на присутність подовженого дуплекса на стадії (е). Краще, щоб сигнал від мішені, що доставляється на стадії (е), був кривою плавлення, картиною плавлення або величиною Т пл., одержаними шляхом вимірювання зміни сигналу флуоресценції від системи двох взаємодіючих міток. Місцезнаходження мітки на СТО та сайта вбудовування вбудовуваної мітки визначається тією мірою, у якій ці дві мітки діють як система двох взаємодіючих міток для індукції зміни сигналу на стадії плавлення. Навіть ще краще, щоб матрична ділянка СТО містила репортерну молекулу або молекулугасник та нуклеотид, який має другу неприродну основу. Реакцію подовження на стадії (d) проводять в присутності нуклеотиду, що містить молекулу-гасник або репортерну молекулу та першу неприродну основу з афінністю специфічного зв'язування з другою неприродною основою в СТО. Ці дві неприродні основи в подовженому дуплексі на стадії (d) утворюють пару основ, викликаючи гасіння сигналу від репортерної молекули за допомогою молекули-гасника, та індукуючи зміну сигналу, внаслідок чого забезпечується одержання сигналу від мішені. Альтернативно, фрагмент містить репортерну молекулу або молекулу-гасник, а матрична ділянка СТО містить нуклеотид, який має другу неприродну основу. Реакцію подовження на стадії (d) проводять в присутності нуклеотиду, що містить молекулу-гасник або репортерну молекулу та першу неприродну основу з афінністю специфічного зв'язування з другою неприродною основою в СТО. Ці дві неприродні основи в подовженому дуплексі на стадії (d) утворюють пару основ, індукуючи зміну сигналу від репортерної молекули шляхом гасіння, внаслідок чого забезпечується одержання сигналу від мішені. 18 UA 115082 C2 5 10 15 20 25 30 35 40 45 50 55 60 Інше пояснене прикладом втілення описане з посиланням на Фіг. 5. В цьому втіленні фрагмент, який містить репортерну молекулу або молекулу-гасник, гібридизується з СТО, що містить нуклеотид, який має другу неприродну основу (наприклад, ізо-dC) з афінністю специфічного зв'язування з першою неприродною основою (наприклад, ізо-dG). Подовження здійснюють в присутності нуклеотиду, що містить першу неприродну основу, мічену молекулоюгасником або репортерною молекулою, з утворенням подовженого дуплекса, у якому сигнал від репортерної молекули гаситься молекулою-гасником. В реакції подовження нуклеотид, що містить першу неприродну основу, вбудовується в сайт, розташований навпроти нуклеотиду, що містить другу неприродну основу. Якщо на стадії (d) утворюється подовжений дуплекс, то репортерна молекула та молекулагасник конформаційно розділяються, що не дозволяє молекулі-гаснику гасити сигнал від репортерної молекули; при цьому якщо на стадії (е) цей подовжений дуплекс піддають плавленню, то репортерна молекула та молекула-гасник конформаційно розташовуються близько одна до одної, що дозволяє молекулі-гаснику гасити сигнал від репортерної молекули, так що доставляється сигнал від мішені, що вказує на присутність подовженого дуплекса а стадії (е). Краще, щоб сигнал від мішені, що доставляється на стадії (е), був кривою плавлення, картиною плавлення або величиною Т пл., одержаними шляхом вимірювання зміни сигналу флуоресценції від системи двох взаємодіючих міток. Місцезнаходження мітки на РТО та сайта вбудовування вбудовуваної мітки визначається тією мірою, у якій ці дві мітки діють як система двох взаємодіючих міток для індукції зміни сигналу на стадії плавлення. Якщо застосовується мітка, вбудовувана в подовжений дуплекс під час реакції подовження, то ця мітка не вбудовується в гібрид, утворений нерозщепленим РТО та СТО, оскільки гібрид не подовжується. Таким чином, даний гібрид не дає сигналу, який не належить до мішені. (4) Інтеркалююча мітка В даному винаході можна застосовувати інтеркалюючу мітку для одержання сигналу від мішені, що вказує на присутність подовженого дуплекса. Інтеркалююча мітка більш корисна в реакції на твердій фазі з використанням іммобілізованих СТО, через те, що генерувати сигнали можуть дволанцюгові молекули нуклеїнових кислот, присутні у зразках. Приклади інтеркалюючих барвників, корисних в даному винаході, включають SYBR™ Green I, PO-PRO™-1, BO-PRO™-1, SYTO™43, SYTO™44, SYTO™45, SYTOX™Blue, POPO™-1, POPO™-3, BOBO™-1, BOBO™-3, LO-PRO™-1, JO-PRO™-1, YO-PRO™1, TO-PRO™1, SYTO™11, SYTO™13, SYTO™15, SYTO™16, SYTO™20, SYTO™23, TOTO™-3, YOYO™3, GelStar™ та триазоловий оранжевий. Ці інтеркалюючі барвники специфічно інтеркалюють в дволанцюгові молекули нуклеїнових кислот, генеруючи сигнали. На Фіг. 13 проілюстроване втілення, у якому інтеркалюючі барвники інтеркалюють між парами основ подовженого дуплекса (С та D на Фіг. 13). Дане втілення також є застосовним до іншого втілення з використанням аналізу плавлення. Пояснене прикладом втілення описане з посиланням на Фіг. 13. Фрагмент гібридизується із захоплюючою ділянкою СТО, іммобілізованого на твердій підкладці. Подовження здійснюють в присутності інтеркалюючого барвника (наприклад, SYBR™ Green) та одержують подовжений дуплекс з інтеркалюючими барвниками. Флуоресцентний сигнал від подовженого дуплекса у плямі на твердій підкладці з іммобілізованим СТО можна детектувати з використанням інтеркалюючих флуоресцентних барвників. Коли подовжений дуплекс піддають плавленню, інтеркалюючі флуоресцентні барвники вивільняються, і флуоресцентний сигнал в цій плямі більше не детектується (на Фіг. 13, не показано). При цьому доставляється сигнал від мішені, що вказує на присутність подовженого дуплекса на стадії (е). Гібрид, утворений нерозщепленим РТО та СТО, забезпечує одержання сигналу, який не належить до мішені, на стадії плавлення. У цьому випадку різниця у величинах Т пл. для подовженого дуплекса та гібриду дозволяє відрізнити сигнал від подовженого дуплекса, що належить до мішені, від сигналу гібриду, який не належить до мішені (Фіг. 13, не показано). Краще, щоб сигнал від мішені, що доставляється на стадії (е), був кривою плавлення, картиною плавлення або величиною Тпл., одержаними шляхом вимірювання зміни сигналу від системи двох взаємодіючих міток. Стадія (f). Детекція сигналу від мішені Остаточно, детекцію подовженого дуплекса здійснюють шляхом вимірювання сигналу від мішені, доставляемого на стадії (е); тим самим присутність подовженого дуплекса вказує на присутність нуклеїновокислотної послідовності-мішені. Детекція може бути здійснена різними способами в залежності від типів сигналу від мішені. 19 UA 115082 C2 5 10 15 20 25 30 35 40 45 50 55 Відповідно до кращого втілення, детекцію сигналу від мішені здійснюють з використанням аналізу плавлення. Використовуваний в даному описі термін "аналіз плавлення" означає метод, з використанням котрого сигнал від мішені, що вказує на присутність подовженого дуплекса, одержують в результаті плавлення подовженого дуплекса, включаючи метод вимірювання сигналів при двох різних температурах, аналіз кривої плавлення, аналіз картини плавлення та аналіз піків плавлення. Краще, аналіз плавлення є аналізом кривої плавлення. Відповідно до кращого втілення, за плавленням, здійснюваним на стадії (е), йде гібридизація з одержанням сигналу від мішені, що вказує на присутність подовженого дуплекса. У цьому випадку детекцію подовженого дуплекса здійснюють з використанням аналізу кривої гібридизації. Крива плавлення або крива гібридизації може бути одержана з використанням традиційних технологій, наприклад, як описано в патентах США №№ 6174670 та 5789167, Drobyshev et al., Gene 188: 45 (1997); Kochinsky and Mirzabekov, Human Mutation, 19: 343 (2002); Livehits et al., J. Biomol. Structure Dynam., 11: 783 (1994) та Howell et al., Nature Biotechnology, 17: 87 (1999). Наприклад, крива плавлення або крива гібридизації може бути графічним зображенням або відображенням зміни вихідного сигналу в залежності від зміни такого параметра, як жорсткість умов гібридизації. Можна побудувати графік залежності вихідного сигналу безпосередньо від параметра гібридизації. В типовому випадку крива плавлення або крива гібридизації будуть характеризуватися вихідним сигналом, наприклад, флуоресценцією, який вказує на відносну кількість дуплексної структури (тобто, на ступінь гібридизації), відкладеним по осі Y, та параметром гібридизації, відкладеним по осі X. РТО та СТО можуть складатися з природних dNMP. Альтернативно, РТО та СТО можуть складатися з модифікованих нуклеотидів або неприродних нуклеотидів, таких як PNA (пептидонуклеїнова кислота (peptide nucleic acid), див. публікацію РСТ № WO 92/20702) та LNA ("закрита" нуклеїнова кислота (locked nucleic acid), див. публікації РСТ №№ WO 98/22489, WO 98/39352 та WO 99/14226). РТО та СТО можуть містити універсальні основи, такі як дезоксіінозин, інозин, 1-(2'-дезокси-бета-D-рибофуранозил)-3-нітропірол та 5-нітроіндол. Термін "універсальна основа" стосується основи, здатної утворювати пари основ з кожною з природних основ ДНК/РНК з невеликою різницею між ними. Як описано вище, РТО може розщеплюватися в сайті, локалізованому в 3'-напрямку від 3'кінця 5'-кінцевої ділянки РТО, що мітить. Цей сайт розщеплення може бути розташований в 5'кінцевій частині 3'-кінцевої ділянки РТО, що впізнає мішень. В тому випадку, коли фрагмент РТО містить 5'-кінцеву частину 3нцевої ділянки РТО, що впізнає мішень, сайт СТО, гібридизований з 5'-кінцевою частиною 3'-кінцевої ділянки, що впізнає мішень, може містити універсальну основу, вироджену послідовність або їх комбінацію. Наприклад, якщо РТО розщеплюється в сайті, локалізованому на відстані одного нуклеотиду в 3'-напрямку від 3'-кінця 5'-кінцевої ділянки РТО, що мітить, то вигідно, щоб 5'-кінцева частина захоплюючої ділянки СТО містила універсальну основу для гібридизації з даним нуклеотидом. Якщо РТО розщеплюється в сайті, локалізованому на відстані двох нуклеотидів в 3'-напрямку від 3'-кінця 5'-кінцевої ділянки РТО, що мітить, то вигідно, щоб 5'-кінець захоплюючої ділянки СТО містив вироджену послідовність, а його суміжний в 3'-напрямку нуклеотид містив універсальну основу. Відповідно, в тому випадку, коли розщеплення РТО відбувається в різних сайтах 5'-кінцевої частини 3'-кінцевої ділянки, що впізнає мішень, використання універсальних основ та вироджених послідовностей в СТО є корисним. На додаток до цього, коли для скринінгу численних нуклеїновокислотних послідовностей-мішеней в умовах індукції розщеплення, залежного від подовження розташованого "угору по течії" праймера, використовують РТО, що містять одну й ту саме 5'кінцеву ділянку, що мітить, можуть бути створені фрагменти РТО, що містять різні 5'-кінцеві части 3'-кінцевої ділянки, що впізнає мішень. В таких випадках корисно використання в СТО універсальних основ та вироджених послідовностей. Стратегії з використанням універсальних основ та вироджених послідовностей в СТО дозволяють застосовувати один тип або мінімальну кількість типів СТО для скринінгу численних нуклеїновокислотних послідовностей-мішеней. Відповідно до кращого втілення, спосіб додатково включає повторення стадій (а)-(b), (а)-(d) або (a)-(f) з денатурацією між повторюваними циклами, краще, з використанням розташованого "вниз по течії" праймера. Це повторення дозволяє ампліфікувати нуклеїновокислотну послідовність-мішень та/або сигнал від мішені. Відповідно до кращого втілення, стадії (a)-(f) здійснюють в реакційній посудині або в окремих реакційних посудинах. Наприклад, стадії (a)-(b), (c)-(d) або (e)-(f) можуть бути здійснені в окремих реакційних посудинах. 20 UA 115082 C2 5 10 15 20 25 30 35 40 45 50 55 60 Відповідно до кращого втілення, стадії (а)-(b) та (c)-(f) можуть бути проведені одночасно або роздільно навіть в одній реакційній посудиш в залежності від реакційних умов (зокрема, температури). В даному винаході не потрібно, щоб нуклеїновокислотні послідовності-мішені, які підлягають детекції та/або ампліфікації, мали будь-яку визначену послідовність або довжину, включаючи будь-які молекули ДНК (гДНК та кДНК) та РНК. Якщо як вихідний матеріал використовують мРНК, перед проведенням стадії відпалу необхідна стадія зворотної транскрипції, деталі якої можна знайти в Joseph Sambrook, et al., Molecular Cloning, A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. (2001); та Noonan, K. F. et al., Nucleic Acids Res., 16: 10366 (1988)). Для зворотної транскрипції можна використовувати випадковий гексамер або олігонуклеотид dT як праймер, здатний гібридизуватися з мРНК. Нуклеїновокислотні послідовності-мішені, які можуть підлягати детекції та/або ампліфікації, включають будь-яку прокаріотичну, еукаріотичну (наприклад, з найпростіших та паразитів, грибів, дріжджів, вищих рослин, нижчих та вищих тварин, в тому числі ссавців та людину), вірусну (наприклад, з вірусу герпесу, ВІЛ (вірусу імунодефіциту людини), вірусу грипу, вірусу Епштейна-Барр, вірусу гепатиту, поліовірусу і т.д.) або віроїдну нуклеїнову кислоту природного походження. Крім того, даний винахід є корисним для детекції варіабельності нуклеотидів. Краще, нуклеїновокислотна послідовність-мішень має варіабельність нуклеотидів. Термін "варіабельність нуклеотидів", використовуваний в даному описі, стосується будь-яких одиночних або численних нуклеотидних замін, делецій або вставок в послідовності ДНК в конкретному положенні поміж безперервних сегментів ДНК, які в інших випадках є однаковими в послідовності. Такі безперервні сегменти ДНК включають ген або будь-яку іншу частину хромосоми. Такі варіабельності нуклеотидів можуть бути результатом мутації або бути варіабельностями поліморфних апелів. Наприклад, варіабельність нуклеотидів, детектована в даному винаході, включає SNP (однонуклеотидний поліморфізм), мутацію, делецію, вставку, заміну та транслокацію. Прикладом варіабельності нуклеотидів є численні варіації в геномі людини (наприклад, варіації в гені MTHFR (метилентетрагідрофолатредуктази)), варіації, які залучені у виникнення лікарської стійкості у патогенних мікроорганізмів, та приводять до онюгенезу варіації. В даному винаході для детекції варіабельності нуклеотидів в нуклеїновокислотній послідовності-мішені в тому випадку, коли використовувані праймери або зонди містять послідовність, комплементарну з урахуванням даної варіабельності нуклеотидів в нуклеїновокислотній послідовності-мішені, ця нуклеїновокислотна послідовність-мішень, що має варіабельність нуклеотидів, описується в даному винаході як порівнянна матриця (matching template). В тому випадку, коли використовувані праймери або зонди містять послідовність, некомплементарну з урахуванням даної варіабельності нуклеотидів в нуклеїновокислотній послідовності-мішені, ця нуклеїновокислотна послідовність-мішень, що має варіабельність нуклеотидів, описується в даному винаході як непорівнянна матриця (mismatching template). Для детекції варіабельностей нуклеотидів 3'-кінець розташованого "угору по течії" праймера можна сконструювати таким чином, щоб вона розташовувалася навпроти сайта з варіабельностью нуклеотидів в нуклеїновокислотній послідовності-мішені. Відповідно до кращого втілення, 3'-кінець розташованого "угору по течії" праймера містить послідовність, комплементарну з урахуванням даної варіабельності нуклеотидів в нуклеїновокислотній послідовності-мішені. 3'-Кінець розташованого "угору по течії" праймера, що містить послідовність, комплементарну з урахуванням даної варіабельності нуклеотидів в нуклеїновокислотній послідовності-мішені, відпалюють з порівнянною матрицею та подовжують, щоб індукувати розщеплення РТО. Одержаний фрагмент РТО гібридизується з СТО з одержанням сигналу від мішені. На відміну від цього, коли 3'-кінець розташованого "угору по течії" праймера не відповідає варіабельності нуклеотидів в непорівнянній матриці, вона не подовжується в умовах, які необхідні для відпалу 3'-кінця праймерів та є суттєвими для подовження, навіть коли розташований "угору по течії" праймер гібридизується з непорівнянною матрицею, і внаслідок цього ніякого сигналу від мішені не генерується. Альтернативно, можна використовувати розщеплення РТО в залежності від гібридизації РТО, що містить послідовність, комплементарну з урахуванням даної варіабельності нуклеотидів в нуклеїновокислотній послідовності-мішені. Наприклад, в регульованих умовах РТО, що містить послідовність, комплементарну з урахуванням даної варіабельності нуклеотидів в нуклеїновокислотній послідовності-мішені, гібридизується з порівнянною матрицею і потім розщеплюється. Одержаний фрагмент РТО гібридизується з СТО з 21 UA 115082 C2 5 10 15 20 25 30 35 40 45 50 55 60 одержанням сигналу від мішені. Разом з тим в регульованих умовах РТО не гібридизується з непорівнянною матрицею, що містить некомплементарну послідовність в місці розташування варіабельності нуклеотидів, та не розщеплюється. У цьому випадку краще, щоб послідовність, комплементарна з урахуванням даної варіабельності нуклеотидів в РТО, була розташована в середині її 3'-кінцевої ділянки РТО, що впізнає мішень. Альтернативно, краще, щоб 5'-кінцева частина 3'-кінцевої ділянки РТО, що впізнає мішень, була орієнтована відносно до варіабельності нуклеотидів в нуклеїновокислотній послідовностімішені для детекції цієї варіабельності нуклеотидів, а 5'-кінцева частина 3'-кінцевої ділянки РТО, що впізнає мішень, містила послідовність, комплементарну з урахуванням даної варіабельності нуклеотидів в нуклеїновокислотній послідовності-мішені. В одному з втілень для детекції однонуклеотидної варіабельності 5'-кінець 3'-кінцевої ділянки РТО, що впізнає мішень, містить послідовність, комплементарну з урахуванням цієї однонуклеотидної варіабельності в нуклеїновокислотній послідовності-мішені. Як описано вище, розщеплення РТО, гібридизованого з порівнянною матрицею, може бути індуковане в сайті, безпосередньо прилеглому в 3'-напрямку до 5'-кінця 3'-кінцевої ділянки РТО, що впізнає мішень, наприклад, в умовах індукції розщеплення, залежного від подовження розташованого "угору по течії"" праймера. 3'-Кінець фрагмента РТО містить нуклеотид, комплементарний з урахуванням даної однонуклеотидної варіабельності. Фрагмент РТО гібридизується з СТО, що містить захоплюючу ділянку, яка містить послідовність, що відповідає варіабельності нуклеотидів, і потім подовжується з утворенням подовженого дуплекса, забезпечуючи одержання сигналу від мішені. Якщо з непорівнянною матрицею, що містить послідовність, ідентичну порівнянній матриці, за винятком однонуклеотидної варіабельності, гібридизується один й той самий РТО, то розщеплення РТО може відбуватися в сайті, розташованому на відстані двох нуклеотидів в 3'-напрямку від 5'-кінця 3'-кінцевої ділянки РТО, що впізнає мішень. 3'-Кінець фрагмента РТО містить ще один розщеплюваний нуклеотид, крім нуклеотиду, комплементарного з урахуванням даної однонуклеотидної варіабельності. Коли сайт СТО, гібридизованого з додатковим розщеплюваним нуклеотидом, конструюють таким, щоб він містив послідовність, некомплементарну цьому додатковому розщеплюваному нуклеотиду, 3'-кінець фрагмента РТО не гібридизується з СТО, результатом чого є відсутність якого-небудь подовження фрагмента РТО в регульованих умовах. Навіть якщо фрагмент РТО подовжується з утворенням подовженого дуплекса, даний дуплекс характеризується величиною Т пл., відмінною від такої для дуплекса, утворюваного в результаті гібридизації між РТО та непорівнянною матрицею. Відповідно до кращого втілення, сайт розщеплення РТО, що містить послідовність, комплементарну з урахуванням даної варіабельності нуклеотидів у своїй 5'-кінцевій частині 3'кінцевої ділянки, що впізнає мішень, буде різним в залежності від того, чи відбувається гібридизація з порівнянною матрицею або з непорівнянною матрицею, у зв'язку з чим фрагмент РТО, що вивільняється в результаті будь-якої з цих двох подій гібридизації, має різну послідовність, краще, у своїй 3'-кінцевій частині, ще краще, на своєму 3'-кінці. Відповідно до кращого втілення, вибір нуклеотидної послідовності в СТО з урахуванням різниці в 3'-кінцевих частинах фрагментів РТО дозволяє відрізнити порівнянну матрицю від непорівнянної матриці. Відповідно до кращого втілення, використовувана в даному винаході нуклеїновокислотна послідовність-мішень є попередньо ампліфікованою нуклеїновокислотною послідовністю. Використання попередньо ампліфікованої нуклеїновокислотної послідовності дає можливість значно підвищити чутливість та специфічність детекції мішені за даним винаходом. Відповідно до кращого втілення, спосіб здійснюють в присутності розташованого "вниз по течії" праймера. Переваги даного винаходу можуть бути особливо відзначені при одночасній (множинній) детекції принаймні двох нуклеїновокислотних послідовностей-мішеней. Відповідно до кращого втілення, спосіб здійснюють з метою детекції принаймні двох типів (краще, принаймні трьох типів, ще краще, принаймні п'яти типів) нуклеїновокислотних послідовностей-мішеней. Відповідно до кращого втілення, спосіб здійснюють з метою детекції принаймні двох типів (краще принаймні трьох типів, ще краще, принаймні п'яти типів) нуклеїновокислотних послідовностей-мішеней; причому розташований "угору по течії" олігонуклеотид включає принаймні два типи (краще, принаймні три типи, ще краще, принаймні п'ять типів) олігонуклеотидів, РТО включає принаймні два типи (краще, принаймні три типи, ще краще, принаймні п'ять типів) РТО, а СТО включає принаймні один тип (краще, принаймні два типи, ще краще, принаймні три типи, ще краще, принаймні п'ять типів) СТО; при цьому коли присутні принаймні два типи нуклеїновокислотних послідовностей-мішеней, даний спосіб забезпечує 22 UA 115082 C2 5 10 15 20 25 30 35 40 45 50 55 60 одержання принаймні двох типів сигналів від мішеней, що відповідають принаймні двом типам нуклеїновокислотних послідовностей-мішеней. 5'-Кінцеві ділянки, що мітять, принаймні двох РТО можуть мати ідентичні одна одній послідовності. Наприклад, якщо даний винахід здійснюють для скринінгу нуклеїновокислотних послідовностей-мішеней, то 5'-кінцеві ділянки РТО, що мітять, можуть мати ідентичні послідовності. Крім того, один тип СТО можна використовувати для детекції множини нуклеїновокислотних послідовностей-мішеней. Наприклад, коли для скринінгу нуклеїновокислотних послідовностеймішеней застосовують РТО, що містять ідентичну послідовність у своїх 5'-кінцевих ділянках, що мітять, можна використовувати один тип СТО. Відповідно до кращого втілення, подовжені дуплекси, що відповідають принаймні двом типам нуклеїновокислотних послідовностей-мішеней, мають відмінні одна від одної величини Тпл.. Відповідно до кращого втілення, принаймні два типи сигналів від мішеней, що відповідають принаймні двом типам нуклеїновокислотних послідовностей-мішеней, створюються за допомогою відмінних одна від одної типів міток. Відповідно до кращого втілення, принаймні два типи сигналів від мішеней, що відповідають принаймні двом типам нуклеїновокислотних послідовностей-мішеней, створюються за допомогою міток одного й того самого типу. Відповідно до кращого втілення, принаймні два типи сигналів від мішеней, що відповідають принаймні двом типам нуклеїновокислотних послідовностей-мішеней, створюються за допомогою міток одного й того самого типу; при цьому подовжені дуплекси, що відповідають принаймні двом типам нуклеїновокислотних послідовностей-мішеней, мають відмінні одна від одної величини Тпл.. Використовуваний в даному описі термін "різні типи міток" стосується міток з різними характеристиками детектованих сигналів. Наприклад, FAM та TAMRA як флуоресцентні репортерці мітки, розглядаються як різні типи міток, оскільки довжини хвиль збудження та емісії для них відрізняються одна від одної. Коли даний винахід здійснюють з метою одночасної детекції принаймні двох типів нуклеїновокислотних послідовностей-мішеней з використанням аналізу кривої плавлення, і подовжені дуплекси, що відповідають принаймні двом типам нуклеїновокислотних послідовностей-мішеней, мають відмінні одна від одної величини Т пл., можна здійснити детекцію принаймні двох типів нуклеїновокислотних послідовностей-мішеней із застосуванням мітки навіть одного типу (наприклад, FAM). Детекція мішені з використанням СТО, іммобілізованого на твердій фазі Значна перевага даного винаходу полягає в ефективній детекції нуклеїновокислотних послідовностей-мішеней, навіть на твердій фазі, такій як мікрочип. Відповідно до кращого втілення, даний винахід здійснюють на твердій фазі, і СТО іммобілізують на твердій підкладці через його 5'-кінець або 3'-кінець. У випадку використання твердої фази проводять вимірювання сигналу, створюваного на твердій підкладці. Коли застосовують іммобілізований СТО, аналіз плавлення, у якому використовуються описані вище системи міток, є застосовним до реакції на твердій фазі за даним винаходом. Відповідно до кращого втілення, одержання сигналу від мішені забезпечується за допомогою одиночної мітки, з'єднаної з фрагментом, або за допомогою одиночної мітки, вбудовуваної в подовжений дуплекс під час реакції подовження. Зокрема, якщо даний винахід здійснюють на твердій фазі із застосуванням одиночної мітки, то можна використовувати звичайну флуоресцентну мітку, і не потрібно спеціальної флуоресцентної мітки, здатної забезпечити сигнали флуоресценції з різними інтенсивностями в залежності від її присутності на дволанцюговій або одноланцюговій структурі. В тому випадку, коли застосовують СТО, іммобілізований на твердій підкладці, можна використовувати хімічні мітки (наприклад, біотин) або ферментативні мітки (наприклад, лужну фосфатазу, пероксидазу, -галактозидазу та -глюкозидазу). Для проведення реакції на твердій фазі СТО іммобілізують безпосередньо або опосередковано (краще, опосередковано) через його 5'-кінець або 3'-кінець (краще, 3'-кінець) на поверхні твердої підкладки. Крім того, іммобілізацію СТО на поверхні твердої підкладки можна проводити у ковалентний або нековалентний спосіб. В тих випадках, коли іммобілізовані СТО є СТО, іммобілізованими на поверхні твердої підкладки опосередковано, використовують придатні лінкери. Лінкери, корисні в даному винаході, можуть включати будь-які лінкери, використовувані для іммобілізації зондів на поверхні твердої підкладки. Наприклад, алкільні чи арильні сполуки з амінною функціональною групою або алкільні чи арильні сполуки з тіоловою 23 UA 115082 C2 5 10 15 20 25 30 35 40 45 50 55 60 функціональною групою служать лінкерами для іммобілізації СТО. Крім цього, лінкерами може служити полі(Т)-хвіст або полі(А)-хвіст. Відповідно до кращого втілення, тверда підкладка, використовувана в даному винаході, є мікрочипом. Мікрочип, який забезпечує дотримання умов реакції в даному винаході, може включати будь-який з мікрочипів, які відомі фахівцю в даній області. Усі процеси в даному винаході, тобто, гібридизацію з нуклеїновокислотними послідовностями-мішенями, розщеплення, подовження, плавлення та детекцію флуоресценції здійснюють на мікрочипі. Іммобілізовані на мікрочипі СТО служать гібридизованими елементами чипа. Тверда підкладка для виготовлення мікрочипа включає метали (наприклад, золото, сплав золота та міді, алюміній), оксид металу, скло, кераміку, кварц, кремній, напівпровідник, пластинку з Si/SiO2, германій, арсенід галію, вуглець, вуглецеві нанотрубки, полімери (наприклад, полістирол, поліетилен, поліпропілен та поліакриламід), сефарозу, агарозу та колоїди, без обмеження ними. Більшість використовуваних в даному винаході іммобілізованих СТО може бути іммобілізована на доступній ділянці або двох чи більше доступних ділянках на твердій підкладці, яка може містити 2-1000000 доступних ділянок. Щоб одержати мікрочип або мікрочипи для заданого застосування, іммобілізовані СТО можуть бути виготовлені з використанням традиційних технологій виготовлення, таких як фотолітографія, технологія струминного друку, механічне точкове нанесення плям та їх похідні. Згідно з даним винаходом, здійснюваним на твердій фазі, можна проводити одночасну детекцію множини нуклеїновокислотних послідовностей-мішеней з використанням мітки навіть одного типу, оскільки мітки на іммобілізованих СТО фізично відокремлені одна від одної. В цьому відношенні, кількість нуклеїновокислотних послідовностей-мішеней за даним винаходом, що підлягають детекції на твердій фазі, є необмеженою. II. Краще втілення з ампліфікацією нуклеїновокислотної послідовності-мішені Даний винахід краще здійснюють одночасно з ампліфікацією нуклеїновокислотної послідовності-мішені, використовуючи пару праймерів, яка складається з розташованого "угору по течії" праймера та розташованого "вниз по течії" праймера, здатних брати участь в синтезі даної нуклеїновокислотної послідовності-мішені. В іншому аспекті даного винаходу запропонований спосіб детекції нуклеїновокислотних послідовностей-мішеней з ДНК або суміші нуклеїнових кислот в аналізі з РТОСЕ (розщепленням та подовженням РТО), який включає: (а) гібридизацію нуклеїновокислотних послідовностей-мішеней з парою праймерів, що складається з розташованого "угору по течії" праймера та розташованого "вниз по течії" праймера, та РТО (олігонуклеотидом, що зондує та мітить); при цьому кожен з праймерів, розташований "угору по течії" праймер та розташований "вниз по течії" праймер, містить нуклеотидну послідовність, що гібридизується, комплементарну нуклеїновокислотній послідовності-мішені; РТО містить (1) 3'-кінцеву ділянку, що впізнає мішень, яка містить нуклеотидну послідовність, що гібридизується, комплементарну нуклеїновокислотній послідовності-мішені, та (2) 5'-кінцеву ділянку, що мітить, яка містить нуклеотидну послідовність, некомплементарну нуклеїновокислотній послідовності-мішені; при цьому 3'-кінцева ділянка, що впізнає мішень, гібридизується з нуклеїновокислотною послідовністю-мішенню, а 5'-кінцева ділянка, що мітить, не гібридизується з нуклеїновокислотною послідовністю-мішенню; РТО локалізований між розташованими "угору по течії" праймером та розташованими "вниз по течії" праймером; при цьому РТО блокований по його 3'-кінцю, щоб не допустити його подовження; (b) приведення в контакт продукту зі стадії (а) з матричною полімеразою нуклеїнових кислот, що виявляє 5'-нуклеазну активність, в умовах, придатних для подовження праймерів та для розщеплення РТО; при цьому, коли РТО гібридизується з нуклеїновокислотними послідовностями-мішенями, розташований "угору по течії" праймер подовжується, і подовжений ланцюг індукує розщеплення РТО матричною полімеразою нуклеїнових кислот, що виявляє 5'нуклеазну активність, так що в результаті розщеплення вивільняється фрагмент, який містить 5'-кінцеву ділянку, що мітить, або частина 5'-кінцевої ділянки РТО, що мітить; (c) гібридизацію фрагмента, вивільненого з РТО, з СТО (захоплюючим та матричним олігонуклеотидом); при цьому СТО містить в напрямку 3'→5' (1) захоплюючу ділянку, яка містить нуклеотидну послідовність, комплементарну 5'-кінцевій ділянці, що мітить, або частини 5'-кінцевої ділянки РТО, що мітить, та (2) матричну ділянку, яка містить нуклеотидну послідовність, некомплементарну 5'-кінцевій ділянці, що мітить, та 3'-кінцеву ділянку, що впізнає мішень; при цьому фрагмент, вивільнений з РТО, гібридизується із захоплюючою ділянкою СТО; (d) проведення реакції подовження з використанням продукту зі стадії (с) та матричної полімерази нуклеїнових кислот; при цьому фрагмент, гібридизований із захоплюючою ділянкою 24 UA 115082 C2 5 10 15 20 25 30 35 40 45 50 55 60 СТО, подовжується з утворенням подовженого дуплекса; причому подовжений дуплекс має величину Т пл., регульовану (1) послідовністю та/або довжиною цього фрагмента, (2) послідовністю та/або довжиною СТО, або (3) послідовністю та/або довжиною фрагмента, та послідовністю та/або довжиною СТО; (е) плавлення подовженого дуплекса в діапазоні температур з одержанням сигналу від мішені, що вказує на присутність подовженого дуплекса; при цьому сигнал від мішені забезпечується за допомогою (1) принаймні однієї мітки, з'єднаної з фрагментом та/або СТО, (2) мітки, вбудовуваної в подовжений дуплекс під час реакції подовження, (3) мітки, вбудовуваної в подовжений дуплекс під час реакції подовження, та мітки, з'єднаної з фрагментом та/або СТО, або (4) інтеркалюючої мітки; і (f) детекцію подовженого дуплекса шляхом вимірювання сигналу від мішені; тим самим присутність подовженого дуплекса вказує на присутність нуклеїновокислотної послідовностімішені. Оскільки в кращому втіленні даного винаходу застосовуються стадії способу за даним винаходом, описаного вище, спільний для них опис опущений, щоб уникнути надмірного дублювання, яке призводить до ускладнення даного опису. Відповідно до кращого втілення, спосіб додатково включає повторення стадій (а)-(b), (а)-(d) або (a)-(f) з денатурацією між повторюваними циклами. Повторення реакцій супроводжується ампліфікацією нуклеїновокислотної послідовності-мішені. Ампліфікацію краще здійснюють відповідно до ПЛР (полімеразної ланцюгової реакції), описаної в патентах США №№ 4683195, 4683202 та 4800159. Відповідно до кращого втілення, спосіб здійснюють для детекції принаймні двох типів нуклеїновокислотних послідовностей-мішеней. Відповідно до кращого втілення, принаймні два типи сигналів від мішеней, що відповідають принаймні двом типам нуклеїновокислотних послідовностей-мішеней, створюються за допомогою міток одного й того самого типу; при цьому подовжені дуплекси, що відповідають принаймні двом типам нуклеїновокислотних послідовностей-мішеней, мають відмінні одна від одної величини Тпл.. III. Спосіб детекції мішені з використанням РТОСЕ, який включає детекцію при попередньо заданій температурі. Даний винахід може бути модифікований з метою використання сигналу від мішені, генерованого спільно з утворенням подовженого дуплекса. Ще в одному аспекті даного винаходу запропонований спосіб детекції нуклеїновокислотної послідовності-мішені з ДНК або суміші нуклеїнових кислот в аналізі з РТОСЕ (розщепленням та подовженням РТО), який включає: (а) гібридизацію нуклеїновокислотної послідовності-мішені з розташованими "угору по течії" олігонуклеотидом та РТО (олігонуклеотидом, що зондує та мітить); при цьому розташований "угору по течії" олігонуклеотид містить нуклеотидну послідовність, що гібридизується, комплементарну нуклеїновокислотній послідовності-мішені; РТО містить (1) 3'-кінцеву ділянку, що впізнає мішень, яка містить нуклеотидну послідовність, що гібридизується, комплементарну нуклеїновокислотній послідовності-мішені, та (2) 5'-кінцеву ділянку, що мітить, яка містить нуклеотидну послідовність, некомплементарну нуклеїновокислотній послідовності-мішені; при цьому 3'-кінцева ділянка, що впізнає мішень, гібридизується з нуклеїновокислотною послідовністю-мішенню, а 5'-кінцева ділянка, що мітить, не гібридизується з нуклеїновокислотною послідовністю-мішенню; розташований "угору по течії" олігонуклеотид локалізований "угору по течії" відносно РТО; (b) приведення в контакт продукту зі стадії (а) з ферментом, що виявляє 5'-нуклеазну активність, в умовах, придатних для розщеплення РТО; при цьому розташований "угору по течії" олігонуклеотид або його подовжений ланцюг індукує розщеплення РТО ферментом, що виявляє 5'-нуклеазну активність, так що в результаті розщеплення вивільняється фрагмент, який містить 5'-кінцеву ділянку, що мітить, або частину 5'-кінцевої ділянки РТО, що мітить; (c) гібридизацію фрагмента, вивільненого з РТО, з СТО (захоплюючим та матричним олігонуклеотидом); при цьому СТО містить в напрямку 3'→5' (1) захоплюючу ділянку, що містить нуклеотидну послідовність, комплементарну 5'-кінцевій ділянці, що мітить, або частині 5'кінцевої ділянки РТО, що мітить, та (2) матричну ділянку, що містить нуклеотидну послідовність, некомплементарну 5'-кінцевій ділянці, що мітить, та 3'-кінцевій ділянці РТО, що впізнає мішень; при цьому фрагмент, вивільнений з РТО, гібридизується із захоплюючою ділянкою СТО; (d) проведення реакції подовження з використанням продукту зі стадії (с) та матричної полімерази нуклеїнових кислот; при цьому фрагмент, гібридизований із захоплюючою ділянкою СТО, подовжується з утворенням подовженого дуплекса; причому подовжений дуплекс має 25 UA 115082 C2 5 10 15 20 25 30 35 40 45 50 55 60 величину Т пл., регульовану (1) послідовністю та/або довжиною цього фрагмента, (2) послідовністю та/або довжиною СТО, або (3) послідовністю та/або довжиною фрагмента, та послідовністю та/або довжиною СТО; при цьому подовжений дуплекс забезпечує одержання сигналу від мішені за допомогою (1) принаймні однієї мітки, з'єднаної з фрагментом та/або СТО, (2) мітки, вбудовуваної в подовжений дуплекс під час реакції подовження, (3) принаймні однієї мітки, з'єднаної з фрагментом та/або СТО, та мітки, вбудовуваної в подовжений дуплекс під час реакції подовження, або (4) інтеркалюючої мітки; і (е) детекцію подовженого дуплекса шляхом вимірювання сигналу від мішені при попередньо заданій температурі, при якій подовжений дуплекс зберігає свою дволанцюгову форму; тим самим присутність подовженого дуплекса вказує на присутність нуклеїновокислотної послідовності-мішені. Оскільки в кращому втіленні даного винаходу застосовуються стадії описаного вище способу за даним винаходом, за винятком стадії плавлення, спільний для них опис опущений, щоб уникнути надмірного дублювання, яке призводить до ускладнення даного опису. В даному винаході, що використовує описаний вище аналіз плавлення, необхідна детекція сигналів від міток не менш ніж при двох різних температурах, тому що сигнал від мішені одержують у вигляді зміни сигналу, спостережуваного при плавленні подовженого дуплекса. В цьому аспекті даного винаходу малоймовірно, що подовжений дуплекс як такий видасть сигнал, за допомогою якого можна відрізнити, відбулося чи не відбулося утворення подовженого дуплекса, і цей сигнал детектують при попередньо заданій температурі, при якій подовжений дуплекс зберігає свою дволанцюгову форму; тим самим встановлюють присутність нуклеїновокислотної послідовності-мішені. Даний винахід стосується вимірювання сигналу від мішені, зв'язаного з утворенням подовженого дуплекса, для детекції присутності нуклеїновокислотної послідовності-мішені. В даному винаході подовжений дуплекс містить мітку, завдяки чому подовжений дуплекс забезпечує одержання сигналу від мішені. Краще, сигнал від мішені є сигналом (генерацією сигналу або гасінням сигналу) від мітки на подовженому дуплексі при попередньо заданій температурі. Введення мітки в даному винаході може бути здійснене у такий саме спосіб, як і для описаного вище способу з використанням аналізу плавлення. Цей аспект даного винаходу з невеликими змінами щодо детекції при попередньо заданій температурі може бути проілюстрований на Фіг. 2-13. Робочий принцип, покладений в основу одержання сигналу від мішені від подовженого дуплекса, виглядає так: (1) подовження фрагмента індукує зміну сигналу від мітки, що дає сигнал від мішені; або (2) гібридизація фрагмента та СТО індукує зміну сигналу від мітки, що дає сигнал від мішені, а подовжений дуплекс зберігає сигнал від мішені. Пояснене прикладом втілення робочого принципу (1) можна описати з посиланням на Фіг. 9. Якщо використовують іммобілізовані СТО, то в даному винаході детектують множину нуклеїновокислотних послідовностей-мішеней у суттєво більш ефективний спосіб. Матрична ділянка іммобілізованого СТО містить репортерну молекулу та молекулу-гасник. Репортерна молекула та молекула-гасник конформаційно розташовуються близько одна до одної, що дозволяє молекулі-гаснику гасити сигнал від репортерної молекули. В тому випадку, коли цей фрагмент гібридизується із захоплюючою ділянкою СТО, молекула-гасник гасить сигнал від репортерної молекули. В результаті утворення подовженого дуплекса репортерна молекула та молекула-гасник конформаційно розділяються, що не дозволяє молекулі-гаснику гасити сигнал від репортерної молекули. Сигнал від мішені доставляється на стадії подовження (С та D на Фіг. 9). Зображений на Фіг. 9 гібрид, утворений нерозщепленим РТО та СТО, не утворює подовженого дуплекса. Внаслідок цього, молекула-гасник, як і раніше, може гасити сигнал від репортерної молекули. Цей гібрид не дає сигналу, який не належить до мішені. Пояснене прикладом втілення робочого принципу (2) можна описати з посиланням на Фіг. 6. На даному малюнку проілюстрований аспект даного винаходу, а також спосіб з використанням аналізу плавлення. 5'-Кінцева ділянка РТО, що мітить, містить репортерну молекулу та молекулу-гасник. Репортерна молекула та молекула-гасник конформаційно розташовуються близько одна до одної, що дозволяє молекулі-гаснику гасити сигнал від репортерної молекули. РТО, гібридизований з нуклеїновокислотною послідовністю-мішенню, переварюється, вивільняючи фрагмент, який містить 5'-кінцеву ділянку, що мітить, з репортерною молекулою та молекулою-гасником, і цей фрагмент гібридизується із захоплюючою ділянкою СТО. В результаті гібридизації репортерна молекула та молекула-гасник конформаційно розділяються, 26 UA 115082 C2 5 10 15 20 25 30 35 40 45 50 55 60 що не дозволяє молекулі-гаснику гасити сигнал від репортерної молекули. Сигнал від мішені доставляється на стадії гібридизації фрагмента, а подовжений дуплекс зберігає сигнал від мішені (С та D на Фіг. 6). Зображений на Фіг. 6 гібрид, утворений нерозщепленим РТО та СТО, дає сигнал, який не належить до мішені (С та D на Фіг. 6), і необхідно провести дисоціацію цього гібриду, щоб виключити сигнал, який не відноситься до мішені. Внаслідок цього температура для вимірювання сигналу від мішені визначається температурою, необхідною для дисоціації гібрида. Відповідно до кращого втілення, далі цю температуру визначають з урахуванням величини Тпл. для гібриду. Відповідно до кращого втілення, детекцію подовженого дуплекса можна здійснювати при температурах, при яких гібрид дисоційований частково. Попередньо задана температура перевищує величину Т пл. для гібриду мінус 10 °C, краще, перевищує величину Тпл. для гібриду мінус 5 °C, ще краще, перевищує величину Т пл. для гібриду, і навіть ще краще, перевищує величину Т пл. для гібриду плюс 5 °C. Відповідно до кращого втілення, сигнал від мішені, забезпечуваний подовженим дуплексом, доставляється під час стадії подовження (d); при цьому гібрид, утворений нерозщепленим РТО та СТО, не дає сигналу, який не належить до мішені, як представлено на Фіг. 2-4 та 9-11. Відповідно до кращого втілення, сигнал від мішені, забезпечуваний подовженим дуплексом, доставляється в результаті гібридизації фрагмента та СТО на стадії (с), а утворення подовженого дуплекса зберігає сигнал від мішені на стадії (d); при цьому гібрид, утворений нерозщепленим РТО та СТО, дає сигнал, який не належить до мішені; при цьому попередньо задана температура перевищує величину Тпл. для гібриду, як представлено на Фіг. 5-8 та 12-13. Коли гібрид, утворений нерозщепленим РТО та СТО, дає сигнал, який не належить до мішені (панель D на Фіг. 6), необхідно провести дисоціацію цього гібриду, щоб виключити сигнал, який не належить до мішені. Внаслідок цього температура для вимірювання сигналу від мішені визначається температурою, необхідною для дисоціації гібриду. Системи міток, корисні в цьому винаході, будуть обговорені більш детально нижче. (1) Мітка, з'єднана з фрагментом та/або СТО. (1-1) Система двох взаємодіючих міток. У втіленні, яке стосується системи двох взаємодіючих міток, СТО містить систему двох взаємодіючих міток, яка складається з репортерної молекули та молекули-гасника; при цьому подовження фрагмента на стадії (d) індукує зміну сигналу від системи двох взаємодіючих міток з одержанням сигналу від мішені. Перше втілення такої системи двох взаємодіючих міток проілюстроване на Фіг. 2. Сигнал від мішені доставляється синхронізовано з генерацією сигналу при подовженні. Відповідно до кращого втілення, репортерна молекула та молекула-гасник можуть бути розташовані на матричній ділянці СТО. Відповідно до кращого втілення, одна з молекул на СТО, репортерна молекула або молекула-гасник, розташована на його 5'-кінці або на відстані 1-5 нуклеотидів від його 5'-кінця, а інша розташована так, щоб гасіння та негасіння сигналу від репортерної молекули здійснювалося в залежності від конформації СТО. У втіленні, яке належить до системи двох взаємодіючих міток, СТО містить систему двох взаємодіючих міток, яка складається з репортерної молекули та молекули-гасника; при цьому гібридизація фрагмента та СТО на стадії (с) індукує зміну сигналу від системи двох взаємодіючих міток з одержанням сигналу від мішені, а подовжений дуплекс зберігає сигнал від мішені. Відповідно до кращого втілення, репортерна молекула та молекула-гасник можуть бути розташовані на захоплюючій ділянці СТО. Відповідно до кращого втілення, одна з молекул на СТО, репортерна молекула або молекула-гасник, розташована на його 3'-кінці або на відстані 1-5 нуклеотидів від його 3'-кінця, а інша розташована так, щоб гасіння та негасіння сигналу від репортерної молекули здійснювалося в залежності від конформації СТО. В цьому втіленні гібрид, утворений нерозщепленим РТО та СТО, дає сигнал, який не належить до мішені; причому температуру для вимірювання сигналу від мішені визначають з урахуванням величини Т пл. для гібриду. У втіленні, яке належить до системи двох взаємодіючих міток, фрагмент містить систему двох взаємодіючих міток, яка складається з репортерної молекули та молекули-гасника; при цьому гібридизація фрагмента та СТО на стадії (с) індукує зміну сигналу від системи двох взаємодіючих міток з одержанням сигналу від мішені, а подовжений дуплекс зберігає сигнал від мішені. Перше втілення такої системи двох взаємодіючих міток проілюстроване на Фіг. 6. 27 UA 115082 C2 5 10 15 20 25 30 35 40 45 50 55 60 Відповідно до кращого втілення, одна з молекул на фрагменті, репортерна молекула або молекула-гасник, розташована на його 5'-кінці або на відстані 1-5 нуклеотидів від 5'-кінця фрагмента, а інша розташована так, щоб гасіння сигналу від репортерної молекули здійснювалося в залежності від конформації фрагмента. В цьому втіленні гібрид, утворений нерозщепленим РТО та СТО, дає сигнал, який не належить до мішені; причому температуру для вимірювання сигналу від мішені визначають з урахуванням величини Т пл. для гібриду. У втіленні, яке належить до системи взаємодіючих міток, фрагмент містить одну з двох взаємодіючих міток, які включають репортерну молекулу та молекулу-гасник, а СТО містить іншу з двох взаємодіючих міток; при цьому гібридизація фрагмента та СТО на стадії (с) індукує зміну сигналу від системи двох взаємодіючих міток з одержанням сигналу від мішені, а подовжений дуплекс зберігає сигнал від мішені. Таке втілення системи двох взаємодіючих міток проілюстроване на Фіг. 8. Репортерна молекула та молекула-гасник можуть бути розташовані в будь-якому місці фрагмента РТО та СТО, за умови, що сигнал від репортерної молекули гаситься молекулоюгасником. Згідно з даним втіленням, репортерна молекула або молекула-гасник на фрагменті РТО розташована краще на його 5'-кінці. Згідно з даним втіленням, репортерна молекула або молекула-гасник на СТО розташована краще на його 5'-кінці. В цьому втіленні гібрид, утворений нерозщепленим РТО та СТО, забезпечує одержання сигналу, який не належить до мішені; причому температуру для вимірювання сигналу від мішені визначають з урахуванням величини Тпл. для гібриду. (1-2) Одиночна мітка У втіленні, яке належить до системи з одиночною міткою, СТО містить одиночну мітку, і подовження фрагмента на стадії (d) індукує зміну сигналу від цієї одиночної мітки з одержанням сигналу від мішені. Таке втілення системи з одиночною міткою проілюстроване на Фіг. 3. Сигнал від мішені доставляється синхронізовано з генерацією сигналу при подовженні. Згідно з даним втіленням матрична ділянка СТО помічена одиночною міткою. У втіленні, яке належить до системи з одиночною міткою, СТО містить одиночну мітку, і гібридизація фрагмента та СТО на стадії (с) індукує зміну сигналу від системи двох взаємодіючих міток з одержанням сигналу від мішені, а подовжений дуплекс зберігає сигнал від мішені. Згідно з даним втіленням захоплююча ділянка СТО помічена одиночною міткою. В цьому втіленні гібрид, утворений нерозщепленим РТО та СТО, забезпечує одержання сигналу, який не належить до мішені; причому температуру для вимірювання сигналу від мішені визначають з урахуванням величини Т пл., для гібриду. У втіленні, яке належить до системи з одиночною міткою, фрагмент містить одиночну мітку, і гібридизація фрагмента та СТО на стадії (с) індукує зміну сигналу від системи двох взаємодіючих міток з одержанням сигналу від мішені, а подовжений дуплекс зберігає сигнал від мішені. Таке втілення системи з одиночною міткою проілюстроване на Фіг. 12. В цьому втіленні гібрид, утворений нерозщепленим РТО та СТО, забезпечує одержання сигналу, який не належить до мішені; причому температуру для вимірювання сигналу від мішені визначають з урахуванням величини Т пл. для гібриду. Використання в даному винаході одиночної мітки має забезпечити одержання різних сигналів в залежності від її присутності на дволанцюговій або одноланцюговій структурі. Одиночна мітка є флуоресцентною міткою, люмінесцентною міткою, хемілюмінесцентною міткою, електрохімічною міткою та металічною міткою. Краще, щоб одиночна мітка була флуоресцентною міткою. Типи та кращі сайти зв'язування одиночних флуоресцентних міток, використовуваних в даному винаході, описані в патентах США №№ 7537886 та 7348141, ідеї яких включені до даного опису шляхом посилання в усій своїй повноті. Краще, щоб одиночна флуоресцентна мітка була JOE, FAM, TAMRA, ROX та іншою міткою на основі флуоресцеїну. Мічений нуклеотидний залишок краще розташований в позиції внутрішнього нуклеотидного залишку в межах олігонуклеотиду, а не на 5'-кінці або 3'-кінці. Одиночна мітка, використовувана в даному винаході, може бути описана з посиланням на вказані вище описи для репортерної молекули та молекули-гасника. Зокрема, якщо даний винахід здійснюють на твердій фазі із застосуванням одиночної мітки, то можна використовувати звичайну флуоресцентну мітку, і не потрібно спеціальної флуоресцентної мітки, здатної забезпечити одержання сигналів флуоресценції з різними інтенсивностями в залежності від її присутності на дволанцюговій або одноланцюговій структурі. 28
ДивитисяДодаткова інформація
Автори англійськоюChun, Jong Yoon, Lee, Young Jo
Автори російськоюЧан Йонг Йун, Ли Йоунг Йо
МПК / Мітки
МПК: C12Q 1/48, C12Q 1/68, C12N 15/11
Мітки: спосіб, детекції, аналізі, розщепленням, подовженням, нуклеїновокислотної, рто, послідовності-мішені
Код посилання
<a href="https://ua.patents.su/80-115082-sposib-detekci-nuklenovokislotno-poslidovnosti-misheni-v-analizi-iz-rozshheplennyam-ta-podovzhennyam-rto.html" target="_blank" rel="follow" title="База патентів України">Спосіб детекції нуклеїновокислотної послідовності-мішені в аналізі із розщепленням та подовженням рто</a>
Детекція нуклеїновокислотної послідовності-мішені в аналізі із розщепленням та подовженням рто

Номер патенту: 110815
Опубліковано: 25.02.2016
Автори: Чан Йонг Йун, Лі Йоунг Йо
МПК: C12Q 1/48, C12N 15/11, C12Q 1/68
Мітки: подовженням, рто, розщепленням, детекція, аналізі, послідовності-мішені, нуклеїновокислотної
Формула / Реферат:
1. Спосіб детекції нуклеїновокислотної послідовності-мішені з ДНК або суміші нуклеїнових кислот в аналізі з РТОСЕ (розщепленням та подовженням РТО), який включає:(a) гібридизацію нуклеїновокислотної послідовності-мішені з розташованими "угору по течії″ олігонуклеотидом та РТО (олігонуклеотидом, що зондує та мітить); при цьому розташований "угору по течії" олігонуклеотид містить нуклеотидну послідовності, що...
Спосіб одержання мішені плазмовим напиленням, мішень і спосіб одержання шару з металу на підкладці шляхом розпилення мішені

Номер патенту: 106984
Опубліковано: 10.11.2014
Автор: Білльєре Домінік
МПК: C23C 14/34, C23C 14/18, C22C 27/00, C23C 4/12, C23C 4/08, B05B 7/22, H05H 1/26
Мітки: розпилення, одержання, спосіб, плазмовим, шляхом, мішень, підкладці, напиленням, металу, шару, мішені
Формула / Реферат:
1. Спосіб отримання мішені плазмовим напиленням за допомогою плазмового пальника, причому вказана мішень містить щонайменше одну сполуку, вибрану з тугоплавких металів, резистивних оксидів, летких оксидів, який відрізняється тим, що за допомогою плазмового напилення напилюють на щонайменше одну частину поверхні мішені щонайменше одну фракцію вказаної сполуки у вигляді порошкової композиції вказаної сполуки, у контрольованій атмосфері, і тим,...
Спосіб детекції генотипів соняшника, стійких до вовчка

Номер патенту: 56691
Опубліковано: 25.01.2011
Автори: Трояновська Аліна В'ячеславівна, Сиволап Юрій Михайлович, Солоденко Анжелла Євгенівна
МПК: A01H 1/04
Мітки: детекції, вовчка, соняшника, генотипів, стійких, спосіб
Формула / Реферат:
Спосіб детекції стійких до вовчка генотипів соняшника, що включає оцінку стійкості інбредних ліній і гібридів соняшника до вовчка, який відрізняється тим, що здійснюється за допомогою електрофоретичного аналізу продуктів ампліфікації ДНК за мікросателітним локусом геному соняшника ORS 1036 та ідентифікації тестованого генотипу як стійкого до вовчка рас А, В, С при наявності в спектрі ДНК алеля розміром 242 пари нуклеотидів, тобто маркера ORS...
Спосіб детекції borrelia burgdorferi sensu lato у біологічних зразках за допомогою полімеразної ланцюгової реакції

Номер патенту: 12659
Опубліковано: 15.02.2006
Автори: Пономаренко Володимир Якович, Нікіфорова Ольга Василівна, Кульшин Володимир Євгенович, Симоненко Василь Іванович
МПК: G01N 33/569, C12Q 1/04, G01N 1/00, C12Q 1/68
Мітки: зразках, ланцюгової, borrelia, burgdorferi, sensu, реакції, допомогою, детекції, полімеразної, біологічних, спосіб
Формула / Реферат:
Спосіб детекції Borrelia burgdorferi sensu lato, у біологічних зразках за допомогою полімеразної ланцюгової реакції, що включає проведення полімеразної ланцюгової реакції, підготовку буфера, ампліфікацію, детекцію ампліфікаційної ДНК, який відрізняється тим, що використовують плазмідний ген OSP і хромосомальний ген S16, що складаються з таких послідовностей пар праймерів:для виявлення ДНК Borrelia burgdorferi sensu lato...
Спосіб і пристрій для кодування мовних сигналів з розщепленням смуги

Номер патенту: 92742
Опубліковано: 10.12.2010
Автори: Вос Кон Бернард, Кандхадай Анантхападманабхан А.
МПК: G10L 21/00, G10L 19/00
Мітки: кодування, мовних, спосіб, пристрій, сигналів, розщепленням, смуги
Формула / Реферат:
1. Пристрій для обробки сигналів, який міститьперший мовний кодер, виконаний з можливістю кодування мовного сигналу смуги низьких частот;другий мовний кодер, виконаний з можливістю кодування мовного сигналу смуги високих частот; ігребінку фільтрів, що має(A) тракт обробки смуги низьких частот, виконаний з можливістю прийому широкосмугового мовного сигналу, що має частотний спектр щонайменше між 1000 і 6000 Гц, і...
Попередній патент: Дверний замок
Наступний патент: Спосіб обміну даними між електронними пристроями за допомогою жестів
Випадковий патент: Шлакоутворююча суміш