Агоніст аполіпопротеїну а-і (apoa-i), мультимірний apoa-i (варіанти), комплекс агоніста apoa-i і ліпіду та спосіб лікування дисліпідемічних порушень
Номер патенту: 71553
Опубліковано: 15.12.2004
Автори: Дюфурк Жан, Метц Гунтер, Дассе Жан-Луі, Корню Ізабелль, Буттнер Клаус, Зекуль Ренате







Формула / Реферат
1. Агоніст аполіпопротеїну A-I (АроА-І), що включає
(і) пептид або пептидний аналог, що містить від 15 до 29 амінокислотних залишків, який утворює амфіпатичну -спіраль в присутності ліпідів і який характеризується наступною формулою (І):
Z1-X1-X2-X3-X4-X5-X6-X7-X8-X9-X10-X11-X12-X13-X14-X15-X16-X17-X18-X19-X20-X21-X22-X23-Z2, (І)
або його фармацевтично прийнятна сіль, де
Х1 представлений проліном (Р), аланіном (А), гліцином (G), глутаміном (Q), аспарагіном (N), аспарагіновою кислотою (D) або D-проліном (р);
Х2 представлений аліфатичною амінокислотою;
Х3 представлений лейцином (L) або фенілаланіном (F);
Х4 представлений кислою амінокислотою;
Х5 представлений лейцином (L) або фенілаланіном (F);
Х6 представлений лейцином (L) або фенілаланіном (F);
Х7 представлений гідрофільною амінокислотою;
X8 представлений кислою або основною амінокислотою;
Х9 представлений лейцином (L) або гліцином (G);
Х10 представлений лейцином (L), триптофаном (W) або гліцином (G);
Х11 представлений гідрофільною амінокислотою;
Х12 представлений гідрофільною амінокислотою;
Х13 представлений гліцином (G) або аліфатичною амінокислотою;
Х14 представлений лейцином (L), триптофаном (W), гліцином (G) або Nal;
Х15 представлений гідрофільною амінокислотою;
Х16 представлений гідрофобною амінокислотою;
Х17 представлений гідрофобною амінокислотою;
Х18 представлений основною амінокислотою, глутаміном (Q) або аспарагіном (N);
Х19 представлений основною амінокислотою, глутаміном (Q) або аспарагіном (N);
Х20 представлений основною амінокислотою;
Х21 представлений аліфатичною амінокислотою;
Х22 представлений основною амінокислотою;
Х23 відсутній або представлений основною амінокислотою;
Z1 представлений H2N- або RC(O)NH-;
Z2 представлений -C(O)NRR, -C(O)OR або -С(O)ОН, або відповідною сіллю;
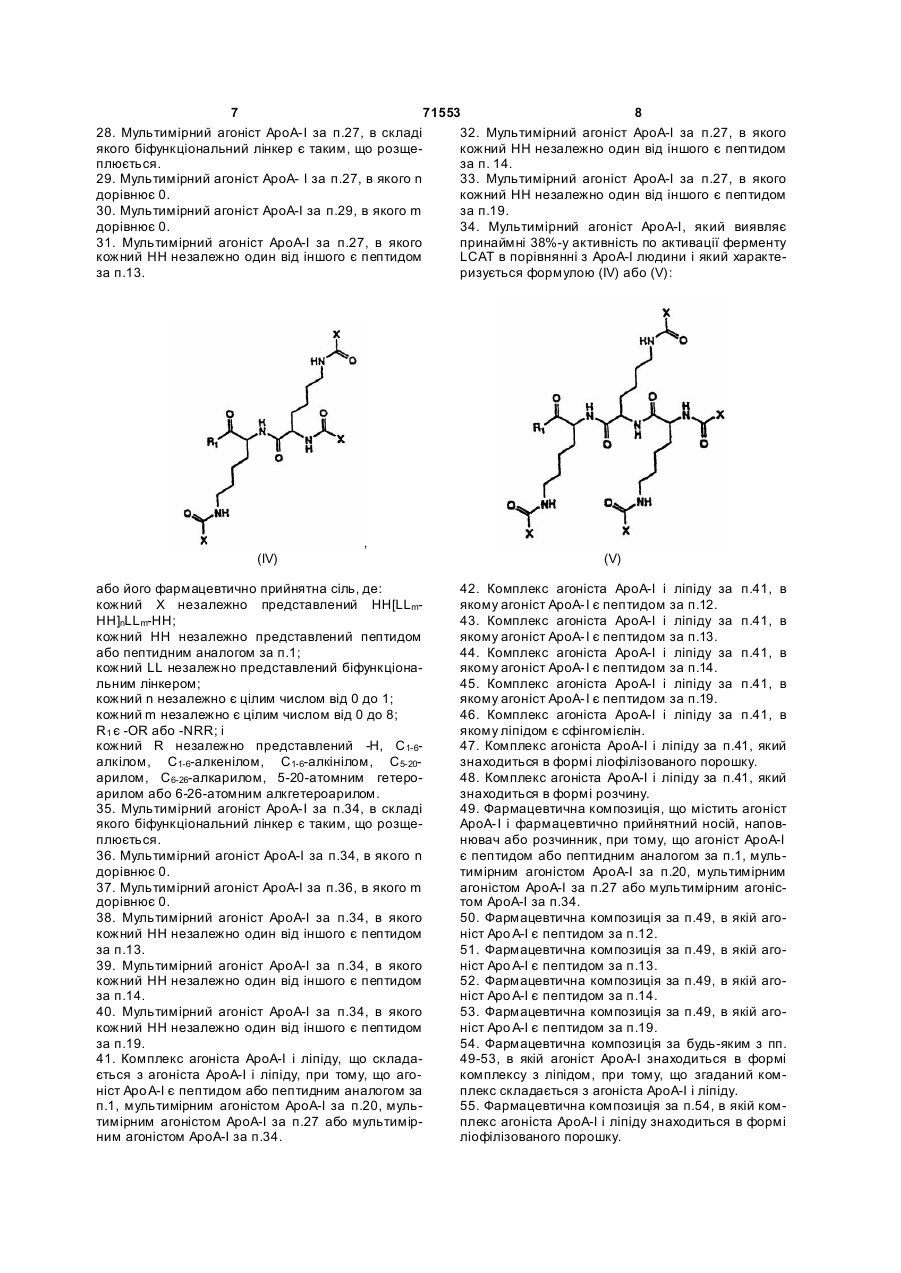
кожний R незалежно один від одного представлений -Н, С1-6-алкілом, С1-6-алкенілом, С1-6-алкінілом, С5-20-арилом, С6-26-алкарилом, 5-20-атомним гетероарилом, 6-26-атомним алкгетероарилом або 1-7-амінокислотним пептидом або пептидним аналогом;
кожний знак «-» між залишками Хn незалежно один від одного означає амідний зв'язок, заміщений амідний зв'язок, ізостер аміду або міметик аміду; або
(іі) делетовану форму формули (І), в якій принаймні один і аж до восьми залишків з Х1, Х2, Х3, Х4, Х5, Х6, Х7, X8, Х9, Х10, Х11, X12, Х13, Х14, Х15, Х16, Х17, X18, X19, Х20, Х21, Х22 делетовані; або
(ііі) змінену форму формули (І), в якій принаймні один із залишків Х1, Х2, Х3, Х4, Х5, Х6, Х7, X8, Х9, Х10, Х11, X12, Х13, Х14, Х15, Х16, Х17, X18, X19, Х20, Х21, Х22 або Х23 по консервативному типу замінений на інший амінокислотний залишок.
2. Агоніст АроА-І за п. І, який виявляє принаймні 38 %-у активність по активації ферменту LCAT в порівнянні з АроА-І людини.
3. Агоніст АроА-І за п. І, який є зміненою формою формули (І).
4. Агоніст АроА-І за п. 3, в складі якого гідрофобні залишки зафіксовані відповідно до формули (І), а принаймні один нефіксований залишок замінений по консервативному типу на інший амінокислотний залишок.
5. Агоніст АроА-І за п. 4, в якому:
Х1 представлений проліном (Р), D-проліном (р), гліцином (G) або аланіном (А);
Х2 представлений аланіном (А), лейцином (L) або валіном (V);
Х3 представлений лейцином (L) або фенілаланіном (F);
Х5 представлений лейцином (L) або фенілаланіном (F);
Х6 представлений лейцином (L) або фенілаланіном (F);
Х9 представлений лейцином (L) або гліцином (G);
Х10 представлений лейцином (L), триптофаном (W) або гліцином (G);
Х13 представлений лейцином (L), гліцином (G) або Aib;
Х14 представлений лейцином, Nal, триптофаном (W) або гліцином (G);
Х16 представлений аланіном (А), Nal, триптофаном (W), гліцином (G), лейцином (L) або фенілаланіном (F);
Х17 представлений лейцином (L), гліцином (G) або Nal;
Х21 представлений лейцином (L); і
принаймні один із залишків Х4, Х7, X8, Х11, Х12, X15, X18, X19, X20, X22 і Х23 замінений по консервативному типу на інший амінокислотний залишок.
6. Агоніст АроА-І за п. 3, в якому гідрофільні залишки зафіксовані відповідно до формули (І), а принаймні один нефіксований залишок замінений по консервативному типу на інший амінокислотний залишок.
7. Агоніст АроА-І за п. 6, в якому:
Х4 представлений аспарагіновою кислотою (D) або глутаміновою кислотою (Е);
Х7 представлений лізином (К), аргініном (R) або орнітином;
X8 представлений аспарагіновою кислотою (D) або глутаміновою кислотою (Е);
Х11 представлений аспарагіном (N) або глутаміном (Q);
Х12 представлений глутаміновою кислотою (Е) або аспарагіновою кислотою (D);
Х15 представлений аспарагіновою кислотою (D) або глутаміновою кислотою (Е);
X18 представлений глутаміном (Q), аспарагіном (N), лізином (К) або орнітином;
Х19 представлений глутаміном (Q), аспарагіном (N), лізином (К) або орнітином;
Х20 представлений лізином (К) або орнітином;
Х22 представлений лізином (К) або орнітином;
Х23 відсутній або представлений лізином (К); і
принаймні один із залишків Х1, Х2, Х3, X5, Х6, Х9, Х10, Х13, Х14, Х16, Х17 і X21 замінений по консервативному типу на інший амінокислотний залишок.
8. Агоніст АроА-І за п. 7, в складі якого Х3 представлений лейцином (L) або фенілаланіном (F), Х6 представлений фенілаланіном (F), Х9 представлений лейцином (L) або гліцином (G), Х10 представлений лейцином (L) або триптофаном (W), або гліцином (G) і принаймні один із залишків Х1, Х2, X5, Х13, Х14, Х16, Х17 і Х21 замінений по консервативному типу на інший амінокислотний залишок.
9. Агоніст АроА-І за пп. 5 або 7, в складі якого замінюючий залишок класифікується в ту ж підгрупу, що і залишок, що замінюється.
10. Агоніст АроА-І за п. 1, що є делетованою формою формули (І).
11. Агоніст АроА-І за п. 10, в складі якого один виток спіралі пептиду або пептидного аналога делетований.
12. Агоніст АроА-І за п. 1, що є пептидом, який складається з 22-23 амінокислот, або пептидним аналогом формули (І).
13. Агоніст АроА-І за п. 12, в складі якого:
знак «-» між залишками означає групу -C(O)NH-;
Z1 представлений H2N-; і
Z2 представлений -С(O)ОН або його сіллю.
14. Агоніст АроА-І за п. 13, в складі якого:
Х1 представлений проліном (Р), аланіном (А), гліцином (G), аспарагіном (N),
глутаміном (Q), аспарагіновою кислотою (D) або D-проліном (р);
Х2 представлений аланіном (А), валіном (V) або лейцином (L);
Х3 представлений лейцином (L) або фенілаланіном (F);
Х4 представлений аспарагіновою кислотою (D) або глутаміновою кислотою (Е);
Х5 представлений лейцином (L) або фенілаланіном (F);
Х6 представлений лейцином (L) або фенілаланіном (F);
Х7 представлений лізином (К), аргініном (R) або орнітином;
X8 представлений аспарагіновою кислотою (D) або глутаміновою кислотою (Е);
Х9 представлений лейцином (L) або гліцином (G);
Х10 представлений лейцином (L), триптофаном (W) або гліцином (G);
Х11 аспарагіном (N) або глутаміном (Q);
Х12 представлений глутаміновою кислотою (Е) або аспарагіновою кислотою (D);
Х13 представлений гліцином (G), лейцином (L) або Aib;
Х14 представлений лейцином (L), Nal, триптофаном (W) або гліцином (G);
Х15 представлений аспарагіновою кислотою (D) або глутаміновою кислотою (Е);
Х16 представлений аланіном (А), Nal, триптофаном (W), лейцином (L),
фенілаланіном (F) або гліцином (G);
Х17 представлений гліцином (G), лейцином (L) або Nal;
X18 представлений глутаміном (Q), аспарагіном (N), лізином (К) або орнітином;
Х19 представлений глутаміном (Q), аспарагіном (N), лізином (К) або орнітином;
Х20 представлений лізином (К) або орнітином;
Х21 представлений лейцином (L);
Х22 представлений лізином (К) або орнітином; і Х23 відсутній або представлений лізином (К).
15. Агоніст АроА-І за п. 14, в складі якого амінокислота Х23 відсутня.
16. Агоніст АроА-І за пп. 13 або 14, в складі якого один із залишків X18 або Х19 представлений глутаміном (Q) або аспарагіном (N), а при цьому інший залишок X18 або Х19 представлений лізином (К) або орнітином.
17. Агоніст АроА-І за п. 14, в складі якого кожний із залишків Х9, Х10, Х13, Х14, Х15 і Х17 не є гліцином (G).
18. Агоніст АроА-І за п. 14, в складі якого один із залишків Х9, Х10, Х13, Х14, Х15 і Х17 представлений гліцином (G), а інші не є гліцином (G).
19. Агоніст АроА-І за п. 1, який вибирають з групи, що включає:
і ацильовані по N-кінцю і (або) амідовані по С-кінцю, або етерифіковані їх форми, при тому, що Х означає Aib, Z означає Nal, a О означає орнітин.
20. Мультимірний агоніст АроА-І, який виявляє щонайменше 38 %-у активність по активації ферменту LCAT в порівнянні з АроА-І людини і який характеризується формулою (II):
HH[LLm-HH]nLLm-HH, (II)
або його фармацевтично прийнятна сіль, де:
кожний показник m незалежно один від одного є цілим числом від 0 до 1;
n є цілим числом від 0 до 10;
кожний НН незалежно представляє пептид або пептидний аналог за п. 1;
кожний LL незалежно представляє біфункціональний лінкер; і
кожний знак «-» незалежно означає ковалентний зв'язок.
21. Мультимірний агоніст АроА-І за п. 20 в складі якого біфункціональний лінкер є таким, що розщеплюється.
22. Мультимірний агоніст АроА-І за п. 20, в якого n дорівнює 0.
23. Мультимірний агоніст АроА-І за п. 22, в якого m дорівнює 0.
24. Мультимірний агоніст АроА-І за п. 20, в якого кожний НН незалежно один від іншого є пептидом за п. 13.
25. Мультимірний агоніст АроА-І за п. 20, в якого кожний НН незалежно один від іншого є пептидом за п. 14.
26. Мультимірний агоніст АроА-І за п. 20, в якого кожний НН незалежно один від іншого є пептидом за п. 19.
27. Мультимірний агоніст АроА-І, який виявляє принаймні 38 % - у активність по активації ферменту LCAT в порівнянні з АроА-І людини і який характеризується формулою (III):
Х-Nya-Х(уа-1)-(Nyb-X(yb-1))p, (III)
або його фармацевтично прийнятна сіль, де:
кожний Х незалежно представлений НН[LLm-HH]nLLm-HH;
кожний НН незалежно представлений базовим пептидом формули (І) або його аналогом, або мутованим, укороченим, делетованим по внутрішньому положенню або добудованим варіантом відповідно до описаного в даному тексті;
кожний LL незалежно представлений біфункціональним лінкером;
кожний m незалежно є цілим числом від 0 до 1;
кожний n незалежно є цілим числом від 0 до 8;
Nya і Nyb незалежно один від одного є багатофункціональною зв'язуючою складовою, де yа і уb представляють число функціональних груп, відповідно, в Nya і Nyb;
кожний yа або уb незалежно є цілим числом від 3 до 8;
р є цілим числом від 0 до 7; і
кожний знак «-» незалежно означає ковалентний зв'язок.
28. Мультимірний агоніст АроА-І за п. 27, в складі якого біфункціональний лінкер є таким, що розщеплюється.
29. Мультимірний агоніст АроА- І за п. 27, в якого n дорівнює 0.
30. Мультимірний агоніст АроА-І за п. 29, в якого m дорівнює 0.
31. Мультимірний агоніст АроА-І за п. 27, в якого кожний НН незалежно один від іншого є пептидом за п. 13.
32. Мультимірний агоніст АроА-І за п. 27, в якого кожний НН незалежно один від іншого є пептидом за п. 14.
33. Мультимірний агоніст АроА-І за п. 27, в якого кожний НН незалежно один від іншого є пептидом за п. 19.
34. Мультимірний агоніст АроА-І, який виявляє принаймні 38 %-у активність по активації ферменту LCAT в порівнянні з АроА-І людини і який характеризується формулою (IV) або (V):
,
(IV)
(V)
або його фармацевтично прийнятна сіль, де:
кожний Х незалежно представлений НН[LLm-HH]nLLm-HH;
кожний НН незалежно представлений пептидом або пептидним аналогом за п. 1;
кожний LL незалежно представлений біфункціональним лінкером;
кожний n незалежно є цілим числом від 0 до 1;
кожний m незалежно є цілим числом від 0 до 8;
R1 є -OR або -NRR; і
кожний R незалежно представлений -Н, С1-6-алкілом, С1-6-алкенілом, С1-6-алкінілом, С5-20-арилом, С6-26-алкарилом, 5-20-атомним гетероарилом або 6-26-атомним алкгетероарилом.
35. Мультимірний агоніст АроА-І за п. 34, в складі якого біфункціональний лінкер є таким, що розщеплюється.
36. Мультимірний агоніст АроА-І за п. 34, в якого n дорівнює 0.
37. Мультимірний агоніст АроА-І за п. 36, в якого m дорівнює 0.
38. Мультимірний агоніст АроА-І за п. 34, в якого кожний НН незалежно один від іншого є пептидом за п. 13.
39. Мультимірний агоніст АроА-І за п. 34, в якого кожний НН незалежно один від іншого є пептидом за п. 14.
40. Мультимірний агоніст АроА-І за п. 34, в якого кожний НН незалежно один від іншого є пептидом за п. 19.
41. Комплекс агоніста АроА-І і ліпіду, що складається з агоніста АроА-І і ліпіду, при тому, що агоніст АроА-І є пептидом або пептидним аналогом за п. 1, мультимірним агоністом АроА-І за п. 20, мультимірним агоністом АроА-І за п. 27 або мультимірним агоністом АроА-І за п. 34.
42. Комплекс агоніста АроА-І і ліпіду за п. 41, в якому агоніст АроА-І є пептидом за п. 12.
43. Комплекс агоніста АроА-І і ліпіду за п. 41, в якому агоніст АроА-І є пептидом за п. 13.
44. Комплекс агоніста АроА-І і ліпіду за п. 41, в якому агоніст АроА-І є пептидом за п. 14.
45. Комплекс агоніста АроА-І і ліпіду за п. 41, в якому агоніст АроА-І є пептидом за п. 19.
46. Комплекс агоніста АроА-І і ліпіду за п. 41, в якому ліпідом є сфінгомієлін.
47. Комплекс агоніста АроА-І і ліпіду за п. 41, який знаходиться в формі ліофілізованого порошку.
48. Комплекс агоніста АроА-І і ліпіду за п. 41, який знаходиться в формі розчину.
49. Фармацевтична композиція, що містить агоніст АроА-І і фармацевтично прийнятний носій, наповнювач або розчинник, при тому, що агоніст АроА-І є пептидом або пептидним аналогом за п. 1, мультимірним агоністом АроА-І за п. 20, мультимірним агоністом АроА-І за п. 27 або мультимірним агоністом АроА-І за п. 34.
50. Фармацевтична композиція за п. 49, в якій агоніст АроА-І є пептидом за п. 12.
51. Фармацевтична композиція за п. 49, в якій агоніст АроА-І є пептидом за п. 13.
52. Фармацевтична композиція за п. 49, в якій агоніст АроА-І є пептидом за п. 14.
53. Фармацевтична композиція за п. 49, в якій агоніст АроА-І є пептидом за п. 19.
54. Фармацевтична композиція за будь-яким з пп. 49-53, в якій агоніст АроА-І знаходиться в формі комплексу з ліпідом, при тому, що згаданий комплекс складається з агоніста АроА-І і ліпіду.
55. Фармацевтична композиція за п. 54, в якій комплекс агоніста АроА-І і ліпіду знаходиться в формі ліофілізованого порошку.
56. Спосіб лікування пацієнта, що страждає від захворювання, пов'язаного з дисліпідемією, при тому, що згаданий спосіб включає етап введення такому пацієнту ефективної кількості агоніста АроА-І за п. 1.
57. Спосіб за п. 56, при тому, що згаданим пацієнтом є людина.
58. Спосіб за п. 56, при тому, що згаданому пацієнту вводять від приблизно 0,5 мг/кг до приблизно 100 мг/кг агоніста АроА-І.
59. Спосіб за п. 56, при тому, що агоніст АроА-І знаходиться в формі фармацевтичної композиції, при тому, що згадана композиція містить агоніст АроА-І і фармацевтично прийнятний носій, наповнювач або розчинник.
60. Спосіб за п. 56, при тому, що агоніст АроА-І знаходиться в формі комплексу агоніста АроА-І і ліпіду, при тому, що згаданий комплекс складається з агоніста АроА-І і ліпіду.
61. Спосіб за п. 56, при тому, що пов'язаним з дисліпідемією захворюванням є гіперхолестеринемія.
62. Спосіб за п. 56, при тому, що пов'язаним з дисліпідемією захворюванням є серцево-судинне захворювання.
63. Спосіб за п. 56, при тому, що пов'язаним з дисліпідемією захворюванням є атеросклероз.
64. Спосіб за п. 56, при тому, що пов'язаним з дисліпідемією захворюванням є рестеноз.
65. Спосіб за п. 56, при тому, що пов'язаним з дисліпідемією захворюванням є дефіцит HDL або АроА-І.
66. Спосіб за п. 56, при тому, що пов'язаним з дисліпідемією захворюванням є гіпертригліцеридемія.
67. Спосіб за п. 56, при тому, що пов'язаним з дисліпідемією захворюванням є метаболічний синдром.
68. Спосіб лікування пацієнта, що страждає від септичного шоку, при тому, що згаданий спосіб включає етап введення такому пацієнту ефективної кількості агоніста АроА-І за п. 1.
69. Спосіб за п. 68, при тому, що згаданим пацієнтом є людина.
70. Спосіб за п. 68, при тому, що згаданому пацієнту вводять від приблизно 0,5 мг/кг до приблизно 100 мг/кг агоніста АроА-І.
Текст
1. Агоніст аполіпопротеїну A-I (Аро А-І), що включає (і) пептид або пептидний аналог, що містить від 15 до 29 амінокислотних залишків, який утворює амфіпатичну a -спіраль в присутності ліпідів і який характеризується наступною формулою (І): Z1-X1-X2-X3-X4-X5-X6-X7-X8-X9-X10-X11-X12-X13-X14X15-X16-X17-X18-X19-X20-X21-X22-X23-Z2, (І) або його фармацевтично прийнятна сіль, де Х1 представлений проліном (Р), аланіном (А), гліцином (G), глутаміном (Q), аспарагіном (N), аспарагіновою кислотою (D) або D-проліном (р); Х2 представлений аліфатичною амінокислотою; Х3 представлений лейцином (L) або фенілаланіном (F); Х4 представлений кислою амінокислотою; Х5 представлений лейцином (L) або фенілаланіном (F); Х6 представлений лейцином (L) або фенілаланіном (F); Х7 представлений гідрофільною амінокислотою; X8 представлений кислою або основною амінокислотою; Х9 представлений лейцином (L) або гліцином (G); Х10 представлений лейцином (L), триптофаном (W) або гліцином (G); 2 (19) 1 3 71553 4 принаймні один нефіксований залишок замінений 9. Агоніст Аро А-І за пп.5 або 7, в складі якого заміпо консервативному типу на інший амінокислотний нюючий залишок класифікується в ту ж підгрупу, залишок. що і залишок, що замінюється. 5. Агоніст АроА-І за п.4, в якому: 10. Агоніст АроА-І за п.1, що є делетованою форХ1 представлений проліном (Р), D-проліном (р), мою формули (І). гліцином (G) або аланіном (А); 11. Агоніст АроА-І за п.10, в складі якого один виХ2 представлений аланіном (А), лейцином (L) або ток спіралі пептиду або пептидного аналога делеваліном (V); тований. Х3 представлений лейцином (L) або фенілалані12. Агоніст АроА-І за п.1, що є пептидом, який ном (F); складається з 22-23 амінокислот, або пептидним Х5 представлений лейцином (L) або фенілаланіаналогом формули (І). ном (F); 13. Агоніст Аро А-І за п.12, в складі якого: Х6 представлений лейцином (L) або фенілаланізнак «-» між залишками означає групу -C(O)NH-; ном (F); Z1 представлений H2N-; і Х9 представлений лейцином (L) або гліцином (G); Z2 представлений -С(O)ОН або його сіллю. Х10 представлений лейцином (L), триптофаном (W) 14. Агоніст Аро А-І за п.13, в складі якого: або гліцином (G); Х1 представлений проліном (Р), аланіном (А), гліХ13 представлений лейцином (L), гліцином (G) або цином (G), аспарагіном (N), Aib; глутаміном (Q), аспарагіновою кислотою (D) або DХ14 представлений лейцином, Nal, триптофаном проліном (р); (W) або гліцином (G); Х2 представлений аланіном (А), валіном (V) або Х16 представлений аланіном (А), Nal, триптофаном лейцином (L); (W), гліцином (G), лейцином (L) або фенілаланіном Х3 представлений лейцином (L) або фенілалані(F); ном (F); Х17 представлений лейцином (L), гліцином (G) або Х4 представлений аспарагіновою кислотою (D) або Nal; глутаміновою кислотою (Е); Х21 представлений лейцином (L); і Х5 представлений лейцином (L) або фенілаланіпринаймні один із залишків Х4 , Х7, X8, Х11 , Х12, X15, ном (F); X18, X19 , X20, X22 і Х23 замінений по консервативноХ6 представлений лейцином (L) або фенілаланіму типу на інший амінокислотний залишок. ном (F); 6. Агоніст Аро А-І за п.3, в якому гідрофільні залиХ7 представлений лізином (К), аргініном (R) або шки зафіксовані відповідно до формули (І), а приорнітином; наймні один нефіксований залишок замінений по X8 представлений аспарагіновою кислотою (D) або консервативному типу на інший амінокислотний глутаміновою кислотою (Е); залишок. Х9 представлений лейцином (L) або гліцином (G); 7. Агоніст АроА-І за п.6, в якому: Х10 представлений лейцином (L), триптофаном (W) Х4 представлений аспарагіновою кислотою (D) або або гліцином (G); глутаміновою кислотою (Е); Х11 аспарагіном (N) або глутаміном (Q); Х7 представлений лізином (К), аргініном (R) або Х12 представлений глутаміновою кислотою (Е) або орнітином; аспарагіновою кислотою (D); X8 представлений аспарагіновою кислотою (D) або Х13 представлений гліцином (G), лейцином (L) або глутаміновою кислотою (Е); Aib; Х11 представлений аспарагіном (N) або глутаміном Х14 представлений лейцином (L), Nal, триптофа(Q); ном (W) або гліцином (G); Х12 представлений глутаміновою кислотою (Е) або Х15 представлений аспарагіновою кислотою (D) аспарагіновою кислотою (D); або глутаміновою кислотою (Е); Х15 представлений аспарагіновою кислотою (D) Х16 представлений аланіном (А), Nal, триптофаном або глутаміновою кислотою (Е); (W), лейцином (L), X18 представлений глутаміном (Q), аспарагіном фенілаланіном (F) або гліцином (G); (N), лізином (К) або орнітином; Х17 представлений гліцином (G), лейцином (L) або Х19 представлений глутаміном (Q), аспарагіном Nal; (N), лізином (К) або орнітином; X18 представлений глутаміном (Q), аспарагіном Х20 представлений лізином (К) або орнітином; (N), лізином (К) або орнітином; Х22 представлений лізином (К) або орнітином; Х19 представлений глутаміном (Q), аспарагіном Х23 відсутній або представлений лізином (К); і (N), лізином (К) або орнітином; принаймні один із залишків Х1, Х2, Х3, X5 , Х6, Х9, Х20 представлений лізином (К) або орнітином; Х10, Х13, Х14, Х16, Х17 і X21 замінений по консерватиХ21 представлений лейцином (L); вному типу на інший амінокислотний залишок. Х22 представлений лізином (К) або орнітином; і Х23 8. Агоніст АроА-І за п.7, в складі якого Х3 представідсутній або представлений лізином (К). влений лейцином (L) або фенілаланіном (F), Х6 15. Агоніст АроА-І за п.14, в складі якого амінокиспредставлений фенілаланіном (F), Х9 представлелота Х23 відсутня. ний лейцином (L) або гліцином (G), Х10 представ16. Агоніст АроА-І за пп.13 або 14, в складі якого лений лейцином (L) або триптофаном (W), або один із залишків X18 або Х19 представлений глутагліцином (G) і принаймні один із залишків Х1, Х2, міном (Q) або аспарагіном (N), а при цьому інший X5 , Х13, Х14 , Х16, Х17 і Х21 замінений по консерватизалишок X18 або Х19 представлений лізином (К) вному типу на інший амінокислотний залишок. або орнітином. 5 71553 6 17. Агоніст АроА-І за п.14, в складі якого кожний із залишків Х9, Х10, Х13, Х14 , Х15 і Х17 не є гліцином (G). 18. Агоніст АроА-І за п.14, в складі якого один із залишків Х9 , Х10 , Х13, Х14, Х15 і Х17 представлений гліцином (G), а інші не є гліцином (G). 19. Агоніст АроА-І за п.1, який вибирають з групи, і ацильовані по N-кінцю і (або) амідовані по Сщо включає: кінцю, або етерифіковані їх форми, при тому, що Х означає Aib, Z означає Nal, a О означає орнітин. 20. Мультимірний агоніст АроА-І, який виявляє щонайменше 38%-у активність по активації ферменту LCAT в порівнянні з АроА-І людини і який характеризується формулою (II): HH[LLm-HH]nLLm-HH, (II) або його фармацевтично прийнятна сіль, де: кожний показник m незалежно один від одного є цілим числом від 0 до 1; n є цілим числом від 0 до 10; кожний НН незалежно представляє пептид або пептидний аналог за п. 1; кожний LL незалежно представляє біфункціональний лінкер; і кожний знак «-» незалежно означає ковалентний зв'язок. 21. Мультимірний агоніст АроА-І за п.20 в складі якого біфункціональний лінкер є таким, що розщеплюється. 22. Мультимірний агоніст АроА-І за п. 20, в якого n дорівнює 0. 23. Мультимірний агоніст АроА-І за п. 22, в якого m дорівнює 0. 24. Мультимірний агоніст АроА-І за п.20, в якого кожний НН незалежно один від іншого є пептидом за п. 13. 25. Мультимірний агоніст АроА-І за п. 20, в якого кожний НН незалежно один від іншого є пептидом за п. 14. 26. Мультимірний агоніст АроА-І за п. 20, в якого кожний НН незалежно один від іншого є пептидом за п.19. 27. Мультимірний агоніст АроА-І, який виявляє принаймні 38% - у активність по активації ферменту LCAT в порівнянні з АроА-І людини і який характеризується формулою (III): Х-N ya-Х( уа-1)-(N yb-X( yb-1)) p, (III) або його фармацевтично прийнятна сіль, де: кожний Х незалежно представлений НН[LLmHH]nLLm-HH; кожний НН незалежно представлений базовим пептидом формули (І) або його аналогом, або мутованим, укороченим, делетованим по внутрішньому положенню або добудованим варіантом відповідно до описаного в даному тексті; кожний LL незалежно представлений біфункціональним лінкером; кожний m незалежно є цілим числом від 0 до 1; кожний n незалежно є цілим числом від 0 до 8; Nya і N yb незалежно один від одного є багатофункціональною зв'язуючою складовою, де yа і уb представляють число функціональних груп, відповідно, в Nya і N yb ; кожний yа або уb незалежно є цілим числом від 3 до 8; р є цілим числом від 0 до 7; і кожний знак «-» незалежно означає ковалентний зв'язок. 7 71553 8 28. Мультимірний агоніст АроА-І за п.27, в складі 32. Мультимірний агоніст АроА-І за п.27, в якого якого біфункціональний лінкер є таким, що розщекожний НН незалежно один від іншого є пептидом плюється. за п. 14. 29. Мультимірний агоніст АроА- І за п.27, в якого n 33. Мультимірний агоніст АроА-І за п.27, в якого дорівнює 0. кожний НН незалежно один від іншого є пептидом 30. Мультимірний агоніст АроА-І за п.29, в якого m за п.19. дорівнює 0. 34. Мультимірний агоніст АроА-І, який виявляє 31. Мультимірний агоніст АроА-І за п.27, в якого принаймні 38%-у активність по активації ферменту кожний НН незалежно один від іншого є пептидом LCAT в порівнянні з АроА-І людини і який характеза п.13. ризується формулою (IV) або (V): , (IV) або його фармацевтично прийнятна сіль, де: кожний Х незалежно представлений НН[LLmHH]nLLm-HH; кожний НН незалежно представлений пептидом або пептидним аналогом за п.1; кожний LL незалежно представлений біфункціональним лінкером; кожний n незалежно є цілим числом від 0 до 1; кожний m незалежно є цілим числом від 0 до 8; R1 є -OR або -NRR; і кожний R незалежно представлений -Н, С1-6алкілом, С1-6-алкенілом, С1-6-алкінілом, С5-20арилом, С6-26-алкарилом, 5-20-атомним гетероарилом або 6-26-атомним алкгетероарилом. 35. Мультимірний агоніст АроА-І за п.34, в складі якого біфункціональний лінкер є таким, що розщеплюється. 36. Мультимірний агоніст АроА-І за п.34, в якого n дорівнює 0. 37. Мультимірний агоніст АроА-І за п.36, в якого m дорівнює 0. 38. Мультимірний агоніст АроА-І за п.34, в якого кожний НН незалежно один від іншого є пептидом за п.13. 39. Мультимірний агоніст АроА-І за п.34, в якого кожний НН незалежно один від іншого є пептидом за п.14. 40. Мультимірний агоніст АроА-І за п.34, в якого кожний НН незалежно один від іншого є пептидом за п.19. 41. Комплекс агоніста АроА-І і ліпіду, що складається з агоніста АроА-І і ліпіду, при тому, що агоніст Аро А-І є пептидом або пептидним аналогом за п.1, мультимірним агоністом АроА-І за п.20, мультимірним агоністом АроА-І за п.27 або мультимірним агоністом АроА-І за п.34. (V) 42. Комплекс агоніста АроА-І і ліпіду за п.41, в якому агоніст АроА-І є пептидом за п.12. 43. Комплекс агоніста АроА-І і ліпіду за п.41, в якому агоніст АроА-І є пептидом за п.13. 44. Комплекс агоніста АроА-І і ліпіду за п.41, в якому агоніст АроА-І є пептидом за п.14. 45. Комплекс агоніста АроА-І і ліпіду за п.41, в якому агоніст АроА-І є пептидом за п.19. 46. Комплекс агоніста АроА-І і ліпіду за п.41, в якому ліпідом є сфінгомієлін. 47. Комплекс агоніста АроА-І і ліпіду за п.41, який знаходиться в формі ліофілізованого порошку. 48. Комплекс агоніста АроА-І і ліпіду за п.41, який знаходиться в формі розчину. 49. Фармацевтична композиція, що містить агоніст АроА-І і фармацевтично прийнятний носій, наповнювач або розчинник, при тому, що агоніст АроА-І є пептидом або пептидним аналогом за п.1, мультимірним агоністом АроА-І за п.20, мультимірним агоністом АроА-І за п.27 або мультимірним агоністом АроА-І за п.34. 50. Фармацевтична композиція за п.49, в якій агоніст Аро А-І є пептидом за п.12. 51. Фармацевтична композиція за п.49, в якій агоніст Аро А-І є пептидом за п.13. 52. Фармацевтична композиція за п.49, в якій агоніст Аро А-І є пептидом за п.14. 53. Фармацевтична композиція за п.49, в якій агоніст Аро А-І є пептидом за п.19. 54. Фармацевтична композиція за будь-яким з пп. 49-53, в якій агоніст АроА-І знаходиться в формі комплексу з ліпідом, при тому, що згаданий комплекс складається з агоніста АроА-І і ліпіду. 55. Фармацевтична композиція за п.54, в якій комплекс агоніста АроА-І і ліпіду знаходиться в формі ліофілізованого порошку. 9 71553 10 56. Спосіб лікування пацієнта, що страждає від 63. Спосіб за п.56, при тому, що пов'язаним з дисзахворювання, пов'язаного з дисліпідемією, при ліпідемією захворюванням є атеросклероз. тому, що згаданий спосіб включає етап введення 64. Спосіб за п.56, при тому, що пов'язаним з дистакому пацієнту е фективної кількості агоніста ліпідемією захворюванням є рестеноз. АроА-І за п.1. 65. Спосіб за п.56, при тому, що пов'язаним з дис57. Спосіб за п.56, при тому, що згаданим пацієнліпідемією захворюванням є дефіцит HDL або том є людина. АроА-І. 58. Спосіб за п.56, при тому, що згаданому пацієн66. Спосіб за п.56, при тому, що пов'язаним з дисту вводять від приблизно 0,5мг/кг до приблизно ліпідемією захворюванням є гіпертригліцеридемія. 100мг/кг агоніста АроА-І. 67. Спосіб за п.56, при тому, що пов'язаним з дис59. Спосіб за п.56, при тому, що агоніст АроА-І ліпідемією захворюванням є метаболічний синдзнаходиться в формі фармацевтичної композиції, ром. при тому, що згадана композиція містить агоніст 68. Спосіб лікування пацієнта, що страждає від АроА-І і фармацевтично прийнятний носій, наповсептичного шоку, при тому, що згаданий спосіб нювач або розчинник. включає етап введення такому пацієнту ефектив60. Спосіб за п.56, при тому, що агоніст АроА-І ної кількості агоніста АроА-І за п.1. знаходиться в формі комплексу агоніста АроА-І і 69. Спосіб за п.68, при тому, що згаданим пацієнліпіду, при тому, що згаданий комплекс складаєтьтом є людина. ся з агоніста АроА-І і ліпіду. 70. Спосіб за п.68, при тому, що згаданому пацієн61. Спосіб за п.56, при тому, що пов'язаним з дисту вводять від приблизно 0,5мг/кг до приблизно ліпідемією захворюванням є гіперхолестеринемія. 100мг/кг агоніста АроА-І. 62. Спосіб за п.56, при тому, що пов'язаним з дисліпідемією захворюванням є серцево-судинне захворювання. 1. Вступ Даний винахід стосується використання композицій агоністів аполіпопротеїна А-І (АроА-І) для лікування захворювань, асоційованих з дисліпопротеїнемією, включаючи гіперхолестеринемію, серцево-судинні захворювання, атеросклероз, рестеноз і інші захворювання, такі як септичний шок. 2. Передумови для винаходу Циркулюючий в організмі холестерин переноситься плазматичними ліпопротеїнами - комплексними частками, що складаються з білка і ліпіда, які забезпечують транспорт ліпідів в крові. Низькомолекулярні ліпопротеїни (LDL) і високомолекулярні ліпопротеїни (HDL) є основними переносниками холестерину. Вважається, що LDL є критичним чинником доставки холестерину з печінки (де він синтезується або з'являється з харчових джерел) у «позапечінкові» тканині тіла. Термін «оборотний транспорт холестерину» характеризує транспорт холестерину з «позапечінкових» тканин зворотно в печінку, де він катаболізується і елімінується. Вважається, що частки плазматичних HDL грають визначальну роль в процесі оборотного транспорту, діючи по типу «прибиральника» тканинного холестерину. Доказів зв'язку між підвищенням рівня сироваткового холестерину з ішемічною хворобою серця безліч. Наприклад, атеросклероз є повільно прогресуючим захворюванням, що характеризується накопиченням холестерину в тканині стінки судини. Зіставлення наявних доказів підтверджує концепцію, відповідно до якої ті, що відкладаються в атеросклеротичних бляшках ліпіди є первинними продуктами плазматичних LDL: це пояснює популярне позначення LDL як «поганого холестерину». З іншого боку, рівень си роваткових HDL зворотно пропорційно корелює з ішемічною хворобою серця - дійсно, високий рівень сироваткових HDL розглядається як її негативний чинник ризику. Передбачається, що високий рівень плазматичного HDL є не тільки чинником захисту від ішемічної хвороби серця, але ще і може зумовлювати розсмоктування атеросклеротичних бляшок (наприклад, див. Badimon et al., 1992, Circulation, 86, suppl. I11, 86-94). Таким чином, традиційно HDL називають «хорошим холестерином». 2.1. Транспорт холестерину Система транспорту ліпідів може бути поділена на дві підсистеми (механізми): екзогенна підсистема, призначена для холестерину і тригліцеридів, що всмоктуються через стінки кишечнику, і ендогенна підсистема, призначена для холестерину і тригліцеридів, що попадає в кров'яне русло з печінки і інших «позапечінкових» тканин. В екзогенній підсистемі харчові ліпіди упаковуються в ліпопротеїнові частки, які називаються хіломікронами, які попадають в кров'яне русло і переносять ті, що містяться в них тригліцериди в жирову тканину (для накопичення) і до м'язів (для окислення з виділенням енергії). Залишки хіломікронів, що містять ефір холестерину, видаляються з крові за рахунок активності специфічних рецепторів, що знаходяться на поверхні гепатоцитів. Такий холестерин потім стає доступним знову для клітинного метаболізму або для повторного перенесення у «позапечінкові» тканині у вигляді плазматичних ліпопротеїнів. В ендогенній підсистемі печінка секретує велику частку екстранизькомолекулярного ліпопротеїна (VLDL) в кров'яне русло. Основу часток VLDL складають в основному тригліцериди, що 11 71553 12 синтезуються в печінці при наявності невеликої кількість холестерину вистачала для покриття покількості ефіру холестерину (або синтезованих в треб метаболізму, але без його перевантаження печінці, або відновлених з хіломікронів). Двома (як огляд, див. Brown & Goldstein, 1990, "In Pharпереважаючими білками, що знаходяться на поmacological Basis of Therapeutics", 8th ed., Goodверхні часток VLDL, є апопротеїн В-100 і апопроman & Gilman, Pergamon Press, NY, Chapter 36, теїн Е. Коли VLDL досягає капілярів жирової ткаpp.874-896). нини і м'язів, тригліцериди, що містяться в ньому 2.2. Оборотний транспорт холестерину екстрагуються з утворенням нового типу часток, Загалом, периферичні (непечінкові) клітини що характеризуються зменшеним розміром і збаотримують холестерин за рахунок поєднання гачених ефіром холестерину, але що зберігають місцевого синтезу і поглинання заздалегідь обидва «своїх» апопротеїна ці частки називають підготовленого стеролу з VLDL і LDL. З іншого босередньомолекулярними ліпопротеїнами (IDL). ку оборотний транспорт холестерину (ОТХ) є меУ людини приблизно половина часток IDL ханізмом, по якому холестерин периферичних швидко вилучається з циркулюючої крові (протяклітин може бути повернений в печінку для погом 2-6 годин після їх утворення), оскільки вони вторного перенесення у «позапечінкові» тканині функціонально тісно пов'язані з клітинами печінки, або екскреції в кишечник у вигляді жовчі, як в моякі екстрагують холестерин з утворенням нових дифікованій формі, так і в окисленій формі (у виVLDL і жовчних кислот. Частки IDL, які не гляді жовчних кислот). Ме ханізм ОТХ представляє відбираються печінкою, залишаються в кров'яному єдиний шлях видалення холестерину з більшості руслі довше. У цей час апопротеїн Ε «позапечінкових» тканин і є критичним для відщеплюється від циркулюючих часток, перетвопідтримки структури і функцій більшості клітин рюючи їх в тип часток LDL, до складу яких входить тіла. єдиний білок -апопротеїн В-100. Ме ханізм ОТХ включає три базових етапи: (1) Насамперед, печінка відбирає і розщеплює вихід холестерину, тобто початкове виділення велику частину холестерину в жовчні кислоти, які є холестерину з різних груп периферичних клітин; кінцевими продуктами в метаболізмі холестерину. (2) етерифікація холестерину внаслідок активності Поглинання часток, що містять холестерин, опосеферменту лецитин/холестеринацилтрансферази редується LDL-рецепторами, які присутні на по(LCAT), що запобігає попаданню холестерину, що верхні гепатоцитів у великій кількості. Рецептор вийшов з клітин зворотно в ці клітини; і (3) поглиLDL зв'язує обидва апопротеїна -Ε і В-100, - і нання і доставка HDL-асоційованого ефіру холевідповідає за скріплення і вилучення і IDL, і LDL з стерину в гепатоцити. Механізм ОТХ опосерекрові. Однак, афінність апопротеїна Ε відносно дується частками HDL. Термін «HDL» означає LDL-рецептора вище в порівнянні з афінністю аполіпопротеїнові частки, які характеризуються висопротеїна В-100. Отже, частки LDL характеризуютькою молекулярною масою. Основними ліпідними ся більшою тривалістю життя в циркулюючій крові компонентами HDL-комплексів є різні фосв порівнянні з частками IDL - LDL знаходяться в фоліпіди, холестерин (ефір) і тригліцериди. Осній в середньому 2,5 дні перед скріпленням на новними аполіпопротеїновими компонентами є А-І LDL-рецепторах в печінці і інших тканинах. і А-ІІ, які визначають функціональні характеристиВисокі рівні сироваткового LDL («поганий хоки HDL; також є невеликі кількості аполіпопротеїнів лестерин») позитивно корелюють з ішемічною С-І, С-ІІ, С-ІІІ, D, Е, J і інші. Розміри HDL можуть в хворобою серця. Наприклад, при атеросклерозі істотній мірі варіюватися і характеризуватися похідний від циркулюючих LDL холестерин нагрорізним складом згаданих вище компонентів, що маджується в стінках артерій, що приводить до залежить від характеру участі в метаболічному утворення бляшок, які порушують протік крові аж каскаді процесу ОТХ. навіть до утворення тромбів, внаслідок чого Ключовим ферментом, залученим до мепросвіт артерії закривається, зумовлюючи серцеханізму ОТХ, є фермент LCAT. Він в основному вий приступ або удар. виробляється в печінці і циркулює в плазмі крові Зрештою кількість внутрішньоклітинного холеасоційовано з фракцією HDL. стерину, що вивільняється з LDL, є чинником конФермент LCAT конвертує клітинний холестетролю метаболізму клітинного холестерину. Накорин в ефір холестерину, який виявляється в HDL, пичення клітинного холестерину, похідного від призначених для елімінації. Внесок в подальше часток VLDL і LDL, контролює такі процеси: поперетворення популяції циркулюючих HDL вносять перше, воно зумовлює зниження інтенсивності білки СЕТР (білок-переносник ефіру холестерину) і синтезу клітинного холестерину за рахунок переPLTP (білок-переносник фосфоліпідів). СЕТР може микання синтезу HMG-Ko A-редуктази, що є клюперенести ефір холестерину, утворений внаслідок човим ферментом в біосинтезі холестерину. Поактивності LCAT, в інші ліпопротеїнові комплекси, друге, притік похідного від LDL-часток холестерину зокрема в АроВ-включаючі ліпопротеїни, такі як забезпечує запасання холестерину з участю актиVLDL і LDL. Білок PLTP забезпечує HDL лецитивуючої АСАТ - клітинного ферменту, контролююном. Ті, що входять в склад HDL тригліцериди мочого конверсію холестерину в ефір холестерину, жуть катаболізуватися з участю позаклітинної який і відкладається у вигляді жирових крапель. печінкової тригліцеридліпази, а вхідний в склад По-третє, накопичення холестерину в клітинах ліпопротеїнів холестерин видаляється в печінці по забезпечує роботу механізму по типу зворотного різних механізмах. зв'язку, який придушує клітинний синтез нових Кожна частка HDL включає принаймні одну LDL-рецепторів. Отже, клітини контролюють копію (а звичайно від двох до чотирьох копій) кількість своїх LDL-рецепторів на такому рівні, щоб АроА-І. АроА-І синтезується печінкою і тонким ки 13 71553 14 шечником у вигляді препропослідовності, яка секцитів. Раніше вже передбачалося існування HDLретується у вигляді прополіпептиду, який швидко рецептора, і недавно рецептор-помічник SR-BI був зазнає процесинг з утворенням зрілого ідентифікований як такий HDL-рецептор (Acton et поліпептиду, що складається з 243 амінокислотних al., 1996, Science, 271, 518-520; Xu et al., 1997, залишків. Аро А-І звичайно включає від 6 до 8 Lipid Res., 38, 1289-1298). Рецептор SR-BI в різних 22-амінокислотних повторів, розділених найбільшій кількості експресований в стелінкером, в ролі якого часто виступає пролін, а в роїдогенних тканинах (наприклад, в наднирках) і в деяких випадках включає своєрідний мотив, склапечінці (Landshuiz et al., 1996, J.Clin. Invest., 98, дений декількома залишками. АроА-І разом з 984-995; Rigotti et al., 1996, J.Biol. Chem., 271, ліпідами утворить три базових типи комплексів: 33545-33549). невеликі комплекси з низьким вмістом ліпідів, що Як передбачається, білок-переносник СЕТР не позначаються як «пре-β1-HDL», сплощені дискограє істотної ролі в процесі ОТХ, але при цьому подібні частки, що включають полярні ліпіди залучений до метаболізму ліпідів, похідних від (фосфоліпіди і холестерин), що позначаються як VLDL і LDL. Однак, зміни активності СЕТР або йо«пре-β2-HDL»; і сферичні частки, що включають і го акцепторів - VLDL і LDL - грають роль в процеполярні, і неполярні ліпіди, що позначаються як сах перетворення популяції HDL. Наприклад, у сферичні, або зрілі HDL (HDL3 і HDL2). Більшість відсутність СЕТР частки HDL перетворюються в HDL в циркулюючій популяції включають і АроА-І, і укрупнені частки, що не підлягають видаленню АроА-ІІ (другий основний білок складу HDL): в да(див. огляди по ОТХ і HDL: Fielding & Fielding, ному тексті для них прийняте позначення «фракції 1995, J.Lipid Res., 36, 211-228; Barrans et al, 1996, AI/AII-HDL» в спільній популяції HDL. Однак, Biochem. Biophys. Acta, 1300, 73-85; Hirano et al, фракція HDL, в складі якої є тільки АроА-І (в дано1997, Arteriosclerosis, Thrombosis & Vascular Biol, му тексті означається як «фракція AI-HDL»), ма17, 1053-1059). буть, є більш ефективною з точки зору механізму 2.3. Сучасні способи лікування ОТХ. Деякі епідеміологічні дослідження дисліпопротеїнемій підтверджують гіпотезу, відповідно до якої фракція В цей час доступними є ряд способів AI-HDL є «антиатерогенною» (Parra et al., 1992, лікування, націлених на зниження рівнів сироватArteriosclerosis & Thrombosis, 12, 701-707; Decossin кових холестерину і тригліцеридів (див, наприклад, et al., 1997, Eur. J.Clin. Invest., 27, 299-307). Brown & Goldstein, цит. вище). Однак, для кожного Хоча механізм перенесення холестерину з з цих способів характерні «свої» обмеження і клітинної поверхні (тобто «вихід холестерину з складності з точки зору їх ефективності, виникненклітин») невідомий, вважається, що комплекс з ня побічних ефектів і придатності для конкретної низьким змістом ліпідів, тобто «пре-β1-HDL», є групи пацієнтів. переважним акцептором для холестерину, переЗв'язуючі жовчну кислоту полімери представносимого від периферичних тканин в механізмі ляють клас лікарських засобів, які переривають ОТХ (див. Davidson et al., 1994, J. Biol. Chem., 269, повторне повернення жовчних кислот з тонкого 22975-22982; Bielicki et al., 1992, J.Lipid Res., 33, кишечнику в печінку -наприклад, холестирамін 1699-1709; Rothblad et al., 1992, J.Lipid Res., 33, (Questran Light®, Bristol-Myers Squibb) і холестипо1091-1097; Kawano et al., 1993, Biochemistry, 32, ла гідрохлорид (Colestid®, Upjohn Co.). При перо5025-5028; Kawano et al, 1997, Biochemistry, 36, ральному введенні ці полімери, що характеризу9816-9825). У ході процесу вилучення холестерину ються позитивним зарядом, зв'язуються в з клітинних поверхонь пре-β1-HDL-частки швидко кишечнику з негативно зарядженими жовчними конвертуються в частки пре-β2-HDL. Білок PLTP кислотами. Оскільки такі полімери не можуть може збільшувати швидкість утворення дисковсмоктуватися з просвіту кишечнику, то вони виподібних часток пре-β2-HDL, однак дані про роль водяться з організму разом з пов'язаними на них цього білка-переносника в ОТХ відсутні. Фермент жовчними кислотами. Однак, використання таких LCAT переважно активний у відношенні дискополімерів щонайбільше знижує рівень сироваткоподібних і сферичних HDL, переносячи 2-ацильну вого холестерину до приблизно 20% і при цьому груп у лецитина або інших фосфоліпідів на вільний асоціюється з виникненням побічних ефектів в гідроксил в молекулі холестерину з утворенням травному тракті, включаючи запори і виникнення складних ефірів холестерину (що залишаються в дефіциту по деяких вітамінам. Більш того оскільки складі HDL) і лізолецитина. У реакції, що контроці полімери зв'язують і інші лікарські препарати, люється LCAT, в якості активатора необхідна інші медикаменти, що вводяться перорально поучасть АроА-І: тобто Аро А-І є природним кофактовинні вводитися не менш ніж за 1 годину або ж ром ферменту LCAT. Конверсія холестерину в через 4-6 годин після введення такого полімеру: його складний ефір, що залишається в HDL, затаким чином, ускладнюється режим прийому ліків побігає зворотному попаданню холестерину в пацієнтами-сердечниками. клітини, внаслідок чого ефір холестерину признаПонижуючими рівень холестерину засобами є чається до видалення. Ефіри холестерину в зрілих статини: ці сполуки блокують синтез холестерину частках HDL у фракції AI-HDL (тобто включаючи внаслідок придушення активності HMG-KoAАроА-І, але не маючи АроА-ІІ) віддаляються редуктази ключового ферменту в печінкою і процесуються в жовч більш ефективно, біосинтетичному шляху холестерину. Статини, ніж той матеріал, який міститься в HDL-фракції, що наприклад, ловастатин (Mevacor, Merck & Зі. Inc.) і включає і АроА-І, і Аро А-ІІ. Це, принаймні часткоправастатин (Pravacholo, Bristol-Myers Squibb Co.), во, може пояснюватися більш ефективним іноді використовуються в поєднанні з полімерами, скріпленням часток AI-HDL на мембранах гепатозв'язуючими жовчні кислоти. Статини в істотній 15 71553 16 мірі знижують рівень сироваткового холестерину і спостерігалася в групі чоловіків у віці 40-55 років, в вмісту LDL в сироватці крові, забезпечуючи знияких ішемічної хвороби не було і немає в момент ження прогресії ішемічної хвороби серця. Однак, дослідження, то незрозуміло, в якій мірі можна рівень сироваткових HDL збільшується лише в екстраполювати ці дані на інші групи пацієнтів (насередній мірі. Механізм ефекту зниження кількості приклад, на жінок і більш молодих чоловіків). LDL може залучати і зниження концентрації VLDL, Дійсно, не було досягнуто скільки-небудь значущоі індукцію експресії клітками LDL-рецептора, го ефекту відносно групи пацієнтів з ішемічною внаслідок чого знижується вироблення і (або) похворобою серця. Застосування фібратів супросилюється катаболізм LDL. З використанням цих воджується серйозними побічними ефектами лікарських засобів асоційовані такі побічні ефекти, наприклад, токсичністю, пов'язаною із злоякісними як дисфункція печінки і нирок (Physicians Desk пухлинами (особливо такими як рак травного тракRef., Med. Economics Co. Inc., Montvale, NJ, 1997). ту), захворюваннями жовчного міхура і Нещодавно федеральне агентство США предстазбільшенням рівня смертності від причин, не вило на ринок препарат аторвастатин (інгібітор пов'язаних з серцевою діяльністю. - Ці лікарські HMG-Ko A-редуктази, розроблений фірмою Parkeзасоби не застосовуються в лікуванні пацієнтів, в Davis, Warner Lambert), призначений для лікування яких єдиною аномалією метаболізму жирів є рідкісних, але вимагаючих інтенсивного втручання підвищений вміст LDL або знижений вміст HDL випадків сімейної гіперхолестеринемії(1995, Scrip, (Physicians Desk Ref., Med. Economics Co. Inc., 20 [19], 10). Montvale, NJ, 1997). Нікотинова кислота (ніацин) є водорозчинним Терапія на основі перорального заміщення есВ-вітамінним комплексом, що використовується як трогенів може бути застосована у випадках харчова добавка і антигіперліпідемічний засіб. гіперхолестеринемії середнього ступеня у жінок в Ніацин зумовлює придушення вироблення VLDL і постклімактеричний період. Однак, збільшення ефективний в зниженні LDL. У деяких випадках він вмісту HDL може супроводжуватися збільшенням застосовується разом з полімерами, зв'язуючими рівня тригліцеридів. Лікування естрогенами, звижовчні кислоти. Ніацин при використанні його в чайно, обмежене конкретною групою пацієнтів адекватних дозах може збільшувати вміст HDL, (жінки після клімаксу) і пов'язане з виявом серйозоднак його застосування обмежується сильними них побічних ефектів, включаючи індукцію злопобічними виявами при призначенні таких високих якісних новоутворень, захворювання жовчного доз. міхура, тромбоемболію, аденому печінки, Фібрати являють собою клас лікарських запідвищений артеріальний тиск, непереносимість собів, що забезпечують зниження рівня ліпідів, глюкози і гіперкальціємію. призначених для лікування різних форм Таким чином, є необхідність в розробці більш гіперліпідемій (наприклад, випадків підвищення безпечних лікарських засобів, які б були ефективні рівня сироваткових тригліцеридів), які також мов зниженні рівня сироваткового холестерину, жуть бути асоційовані з гіперхолестеринемією. збільшенні вмісту HDL в сироватці крові, Вважається, що фібрати зумовлюють зниження профілактиці ішемічної хвороби серця і (або) вмісту фракції VLDL і істотне підвищення HDL лікуванні такої хвороби, що вже є, а особливо атеоднак, вплив цих препаратів на рівень сироваткоросклерозу. вого холестерину виявляється непостійним. У 2.4. Аро А-І, як мішень Сполучених Штатах Америки фібрати дозволені Жоден з доступних в цей час лікарських задля використання в якості антиліпідемічних засобів, призначених для зниження рівня холестерисобів, однак, як засоби проти гіперхолестеринемії, ну, не забезпечує безпечного підвищення рівня вони не сертифіковані. Наприклад, клофібрат HDL і стимулювання ОТХ - тобто не працює на (Atromid-S®, Wyeth-Ayerst Lab.) є антиліпідемічним рівні механізму транспорту холестерину, модулюзасобом, який зумовлює (по поки не встановленовання його надходження зїжею, повторного вклюму механізму) зниження сироваткових чення в цикл, синтезу холестерину і утворення тригліцеридів за рахунок придушення фракції популяції VLDL. VLDL. Хоча рівень сироваткового холестерину Оскільки бажаним є створити лікарські засоби, може бути знижений у деяких груп пацієнтів, які б стимулювали вихід холестерину і його видабіохімічна реакція на цей препарат виявляється лення, а також через наявність декількох порізною, і при цьому не завжди можна передбачити, тенційних мішеней в системі ОТХ - таких як LCAT, ким з пацієнтів будуть досягнуті найбільш ефекHDL і їх різних компонентів (АроА-І, АроА-ІІ і фостивні результати. Як було показано, препарат фоліпіди), PLTP і СЕТР, то невідомо, яка з цих Atromid-S® не виявляє ефективності відносно мішеней буде найбільш ефективною з точки зору ішемічної хвороби серця. Споріднений по хімічних досягнення бажаного складу ліпопротеїнів і захисі фармакологічних власти востях лікарський засіб ного ефекту. Зміна будь-якого одного компонента гемфіброзил (Lopid®, Parke-Davis), - є препаратом, в механізмі ОТХ неминуче приведе до зміни скларегулюючим зміст ліпідів, який в середній мірі ду циркулюючи х ліпопротеїнів і, відповідно, вплине збільшує пов'язаний з HDL холестерин - тобто на ефективність ОТХ. збільшується вміст фракцій HDL2 і HDL3, рівно як і Деякі доказові дані, що засновуються на мофракції, що включає і Аро А-І, і АроА-ІІ (тобто делях in vivo , вказують на HDL і на його базовий фракції AI/AII-HDL). Однак, «ліпідна відповідь» компонент, яким є Аро А-І, як на чинники завиявляється різноманітною, особливо при побігання атеросклеротичним пошкодженням і порівнянні різних груп пацієнтів. Більше за те, потенційного розсмоктування бляшок, тобто робоскільки профілактика ішемічної хвороби серця лять їх привабливими мішенями для терапевтич 17 71553 18 ного втручання. По-перше, є зворотна кореляційна ливу роль АроА-І в активації LCAT і стимуляції залежність між вмістом в сироватці АроА-І (HDL) і механізму ОТХ, додаткові дослідження вказують процесів атерогенезу у чоловіків (Gordon & Rifkind, на наявність більш складного сценарію: основними 1989, New England J.Med., 321, 1311-1316; Gordon компонентами, які забезпечують зміну виходу et al., 1989, Circulation, 79, 8-15). Дійсно, був встаклітинного холестерину, є фосфоліпіди (Fournier et новлений взаємозв'язок між окремими варіантами al., 1996, J.Lipid Res., 37, 1704-1711). HDL і ризиком розвитку атеросклерозу у людини З точки зору вірогідної ролі HDL, а по суті і са(Miller, 1987, Amer. Heart, 113, 589-597; Cheung et мого АроА-І, і асоційованого з ним фосфоліпіда, в al., 1991, Lipid Res., 32, 383-394;- Fruchart & Ailзахисті від атеросклерозу, були здійснені клінічні haud, 1992, Clin, Chem., 38, 79). випробування з використанням рекомбінантного По-друге, дослідження на тваринних моделях АроА-І, які були припинені і потім відновлені в підтверджують захисну роль ApoA-I (HDL). UCB, Бельгія (Pharmacoprojects, 27 жовтня 1995p.; Лікування кроликів, раціон яких включав холестеIMS R&D Focus, 30 червня 1997p.; Drug Status Upрин, шляхом введення АроА-І або HDL приводило date, 1997, Atherosclerosis, 2, 261-265; також див. у таких тварин до придушення розвитку і прогреM.Eriksson, "The role of HDL in disease prevention". сування бляшок (жирового нашарування) (Koizumi Congress, 7-9 Nov., 1996, Fort Worth; Lacko & Miller, et al., 1988, J.Lipid Res., 29, 1405-1415; Badimon et 1997, J.Lipid Res., 38, 1267-1273; і міжнародна паal., 1989, Lab. Invest, 60, 455-461; Badimon et al., тентна заявка WO 94/13819), а також проводилися 1990, J.Clin. Invest, 85, 1234-1241). Однак, ефекі були потім припинені Bio-Tech (Pharmacoprojects, тивність такого лікування варіювалася в залеж7 квітня 1989p.). Також робилися випробування, в ності від джерела HDL (Beitz et al., 1992, Prosяких АроА-І використовували для лікування сепtaglandins, Leukotrienes & Essential Fatty Acids, 47, тичного шоку (Opal, "Reconstituted HDL as a Treat149-152; Mezdour et al., 1995, Atherosclerosis, 113, ment Strategy for Sepsis", IBC's 7th Intern. Conf. on 237-246). Sepsis, April 28-30, 1997, Washington, DC; Gouni et По-третє, прямий доказ ролі АроА-І був отриal., 1993, J.Lipid Res., 94, 139-146; Levine, маний в експериментах на трансгенних тваринах. міжнародна патентна заявка WO 96/04916). Однак, Експресія людського гена, що кодує АроА-І, перевсе це зв'язане з існуванням різних «пасток», несеного мишам, генетично схильних до атеропов'язаних з отриманням і застосуванням АроА-І, склерозу, що індукується харчовими компонентаякі роблять його, м'яко кажучи, неідеальним ми, забезпечувала захист від пошкоджень аорти лікарським засобом: наприклад, АроА-І є великим (Rubin et al., 1991, Nature, 353, 265-267). Трансген білком, отримання якого трудомістке і недешеве; АроА-І також, як було показано, придушує атероповинні бути подолані значні проблеми виробницсклероз у мишей, що характеризуються дефіцитом тва і відновлювальності в зв'язку із забезпеченням апопротеїна Е, і у мишей, трансгенних по Аро(а) стабільності в процесі його зберігання, доставки (Paszty et al., 1994, J.Clin. Invest, 94, 899-903; активного компонента і часу напівжиття in vivo. Plump et al., 1994, Proc. Natl. Acad. Sci. USA, 91, Беручи до уваги ці проблеми, і були зроблені 9607-9611; Liu et al, 1994, J.Lipid Res, 35, 2263спроби сформувати пептиди, які б імітували ак2266). Схожі результати були отримані при вивтивність АроА-І. Оскільки ключовою для вияву акченні трансгенних кроликів, експресуючих АроА-І тивності Аро А-І є наявність множинних повторів людини (Duverger, 1996, Circulation, 94, 713-717; унікальної вторинної організації цього білка Duverger et al, 1996, Arteriosclerosis, Thrombosis & амфіпатичної α-спіралі Α-класу (Segrest, 1974, Vascular Biol, 16, 1424-1429), і трансгенних FEBS Lett., 38, 247-253), - то основні зусилля при пацюків, в яких підвищені рівні людського АроА-І конструюванні пептидів, які б імітували активність забезпечували захист від атеросклерозу і придуАроА-І, були зосереджені на створенні молекул, шення рестеноза, можливого внаслідок проведенякі б і утворювали такі амфіпатичні α-спіралі Аня судинопластики (Burkey et al, 1992, Circulation, класу. suppl. I, 86, 1-472, Abstr. 1876; Burkey et al, 1995, Амфіпатичні α-спіралі Α-класу є унікальними в J.Lipid Res, 36, 1463-1473). тому, що залишки позитивно заряджених У механізмі ОТХ більш ефективним представамінокислот кластеризовані в зоні гідрофобноляється AI-HDL в порівнянні з фракцією AI/AII-HDL. гідрофільного «інтерфейсу» (тобто зіткнення двох Дослідження, проведені на трансгенних мишах, типів поверхонь білка), а залишки негативно занесучих гени АроА-І або АроА-І+АроА-ІІ (АІ/АІІ) ряджених амінокислот сконцентровані на поверхні людини, показали, що білковий склад HDL в гідрофільного сегмента. Далі, пептиди, які мають істотній мірі впливає на його вияви - при цьому α-спіраль Α-класу характеризуються величиною більш істотний антиатерогенний ефект був харакгідрофобного кута, меншою 180° (Segrest et al., терний для AI-HDL в порівнянні з AI/AII-HDL 1990, Proteins: Structure, Function & Genetics, 8, (Schultz et al., 1993, Nature, 365, 762-764). Пара103-117). Початкові нові стратегії конструювання лельні дослідження з використанням трансгенних імітаторів АроА-І не засновувалися на первинних мишей, експресуючих ген LCAT людини, показали, амінокислотних послідовностях що зустрічаються що деяке зростання активності LCAT значною в природі аполіпопротеїнів, але базувалися на мірою змінює рівні пов'язаного з ліпопротеїнами включенні цих унікальних спіралей Α-класу до холестерину і що LCAT володіє виразною переваскладу послідовності пептидних аналогів, рівно як і гою по відношенню до фракції HDL, що включає деяких інших властивостей доменів, що є в складі АроА-І (Francone et al., 1995, J.Сіn. Invest, 96, АроА-І (див., наприклад, Davidson et al., 1996, 1440-1448; Berard et al., 1997, Nature Med., 3 [7], Proc. Natl. Acad. Sci. USA, 93, 13605-13610; Rogers 744-749). При тому, що ці дані підтверджують важet al., 1997, Biochemistry, 36, 288-300; Lins et al., 19 71553 20 1993, Biochim. Biophys. Ada, Biomembranes, 1151, амінокислот є мінімально необхідною для забез137-142; Ji & Jonas, 1995, J.Biol. Chem., 270, печення властивостей по скріпленню з ліпідами 11290-11297; Collet et al., 1997, J.Lipid Res., 38, при моделюванні амфіпатичних пептидів. 634-644; Sparrow & Gotto, 1980, Ann. NY Acad- Sci., Дослідження, проведені на матеріалі мутант348, 187-211; Saprrow & Gotto, 1982, CRC Crit. Rev. них варіантів LAP20, в яких залишок проліну зайBiochem., 13, 87-107; Sorci-Thomas et al., 1993, мав в амінокислотній послідовності різні положенJ.Biol. Chem., 268, 21403-21409; Wang et al., 1996, ня, показали, що є прямий взаємозв'язок між Biochem. Biophys. Acta, 174-184; Minnich et al., скріпленням ліпідів і активацією LCAT, але при 1992, J.Biol. Chem., 267, 16553-16560; Holvoet et цьому ще і те, що сам по собі спіральний поal., 1995, Biochemistry, 34, 13334-13342; Sorciтенціал такого пептиду не викликає активації ферThomas et al., 1997, J.Biol. Chem, 272, 7278-7284; менту LCAT (Ponsin et al., 1986, J.Biol. Chem, 261, Frank et al., 1997, Biochemistry, 36, 1798-1806). 9202-9205). Більше того присутність такого «брейВ одному з досліджень Фукусіма з співавт. кера спіралі» (пролін) поблизу від середини даного (Fukushima et al.) синтезували 22-амінокислотний пептиду знижує його афінність стосовно фоспептид, що повністю складається із залишків Glu, фоліпідів, рівно як і здатність активувати LCAT. Lys і Leu, взаєморозташованих таким чином, щоб При тому, що деякі LAP-пептиди, як було показаутворювати амфіпатичну α-спіраль з рівними но, зв'язуються з фосфоліпідами (Sparrow et al., гідрофільною і гідрофобною поверхнями (тобто цит. вище), проте залишається спірним питання «ELK-пептид») (Fukushima et al., 1979, J.Amer. про ту міру, з якою LAP-пептиди зберігають Chem. Soc, 101, 3703-3704; Fukushima et al., 1980, спіральну організацію в присутності ліпідів (Buchko J.Biol. Chem, 255, 10651-10657). ELK-пептид хаet al., 1996, J.Biol. Chem., 271, 3039-3045; Zhong et рактеризується 41%-ою гомологією з ділянкою al., 1994, Peptide Res., 7, 99-106). амінокислот 198-219 в складі поліпептиду АроА-l. Segrest з співавт. синтезували пептиди, що При проведенні аналізу методом кількісної ультскладаються з 18-24 амінокислот, які не виявлярафільтрації, гель-хроматографії і кругового дихють гомології зі спіральними ділянками АроА-І роїзму для цих ELK-пептидів було продемонстро(Kannelis et al., 1980, J.Biol. Chem., 255, 11464ване ефективне асоціювання з фосфоліпідами і 11472; Segrest et al., 1983, J.Biol. Chem., 258, 2290забезпечення імітації деяких фізичних і хімічних 2295). Конкретні послідовності були сконструйовластивостей Аро А-І (Kaiser et al., 1983, Proc. Natl. вані таким чином, щоб імітувати амфіпатичні Acad. Sci. USA, 80, 1137-1140; Kaiser et al., 1984, спіральні домени Α-класу аполіпопротеїнів по таScience, 223, 249-255; Fukushima et al., 1980, цит. ких параметрах, як гідрофобний момент (Eisenberg вище; Nakagawa et al., 1985, J.Amer. Chem. Soc, et al., 1982, Nature, 299, 371-374) і розподіл заряду 107, 7087-7092). За результатами такого аналізу (Segrest et al., 1990, Proteins, 8, 103-117; патент Іокояма з співавт. дійшли висновку, що критичним США №4643988). Один з 18-амінокислотних пепчинником забезпечення активності ферменту тидів, позначений як «пептид-18А», був сконструйLCAT є проста присутність великої амфіпатичної ований як модель α-спіралі А-класу (Segrest et al., структури (Yoko yama et al, 1980, J.Biol. Chem, 255, 1990, цит. вище). Дослідження на матеріалі таких 7333-7339). Пізніше було виявлене, що димер тапептидів і пептидів, що характеризуються протикого 22-амінокислотного пептиду в більшій мірі лежними параметрами заряду, таких як «пептидімітує властивості Аро А-І в порівнянні з 18К», виразно показали, що розподіл заряду є кривідповідним мономером: ґрунтуючись на отриматичним для функціональності параметром: пептиних даних було підтверджено, що такий 44ди з ревертованим розподілом заряду виявляють амінокислотний пептид, який уривається в своїй знижену афінність відносно пептидів в порівнянні з середині «брейкером спіралі» (або гліцином, або імітаторами 18А Α-класу і меншу спіральну склапроліном), являє собою мінімальний дову в присутності ліпідів (Kanellis et al., 1980, функціональний домен білка АроА-І (Nakagawa et J.Biol. Chem., 255, 11464-11473; Anantharamaiah et al., 1985, цит. вище). al., 1985, J.Biol. Chem., 260, 10248-10255; Chung et В інших дослідженнях як модель використали al., 1985, J.Biol. Chem., 260, 10256-10262; Epand et амфіпатичні пептиди, названі «LAP-пептидами» al., 1987, J.Biol. Chem., 262, 9389-9396; Ananthara(Pownall et al., 1980, Proc. Natl. Acad. Sci. USA, 77, maiah et al., 1991, Adv. Exp. Med. Biol, 285, 1313154-3158; Sparrow et al., 1981, "In Peptides: Syn140). thesis-Structure-Function", eds. Roch & Gross, Pierce Інші синтетичні пептиди, що не мають схожості Chem. Co, Rockford, IL, pp.253-256). Ґрунтуючись з амінокислотною послідовністю аполіпопротеїнів, на параметрах скріплення ліпідів з фрагментами які були представлені і мали «менший успіх», нативних аполіпопротеїнів, були сконструйовані включають димери і тримери пептиду 18А (Ananдекілька LAP-пептидів, позначених LAP-16, LAP-20 tharamaiah et al., 1986, Proteins Biol. Fluids, 34, 63і LAP-24 (включають, відповідно, 16, 20 і 24 66), пептиди GALA і EALA (Subbarao et al., 1988, амінокислотних залишків). Ці модельні Proteins: Structure, Function & Genet, 3, 187-198) і амфіпатичні пептиди не виявляють схожості пептиди ID (Labeur et al., 1997, Arteriosclerosis, послідовності з аполіпопротеїнами і були сконстThrombosis & Vascular Biol, 17, 580-588) і пептид руйовані так, щоб мати гідрофільні поверхні, ор18АМ4 (Brasseur et al., 1993, Biochim. Biophys. ганізовані відмінно від таких поверхонь, що є в Acta, 1170, 1-7). складі амфіпатичних спіралей Α-класу в Також на базі амінокислотної послідовності аполіпопротеїнів (Segrest et al., 1992, J.Lipid Res, спіралей АроА-І людини був сконструйований 33, 141-166). Виходячи з даних цих досліджень, «консенсусний» пептид, що складається з 22 автори роботи дійшли висновку, що наявність 20 амінокислот (Anantharamaiah et al., 1980, Arterio 21 71553 22 sclerosis, 10, 95-105; Venkatachalapathi et al., 1991, активності (активації, що визначаються Моl. Conform, and Biol. Interact., Ind. Acad. Sci., ser. відношенням LCAT до одиниці молекулярної маси) У, 585-596). Пептид був сконструйований з урахувідповідали або перевищували такі показники наванням ідентифікації найбільш переважного зативної молекули. Зокрема, агоністами АроА-І по лишку по кожному положенню гіпотетичних даному винаходу є пептиди або пептидні аналоги, спіралей в складі АроА-І. Як і у випадку з описаниякі: утворюють амфіпатичні спіралі (в присутності ми вище пептидами, спіраль, що утворюється даліпідів), зв'язують ліпіди, утворюють пре-β-подібні ним пептидом, характеризується наявністю клаабо HDL-подібні комплекси, активують LCAT, зустера позитивно заряджених амінокислот по мовлюють підвищення сироваткових рівнів центру гідрофільної поверхні і величиною фракцій HDL і сприяють виходу холестерину з гідрофобного кута менше за 180°. При тому, що клітин. димер такого пептиду в деякій мірі ефективний по Даний винахід засновується, зокрема, на виактивації LCAT, відповідний мономер виявляє конаних заявниками дослідженнях і відкриттях слабкі властивості по скріпленню ліпідів пептидів, які імітують функції АроА-І. Пептиди по (Venkatachalapathi et al., 1991, цит. вище). даному винаходу були сконструйовані на основі Ґрунтуючись насамперед на дослідженнях in передбачуваної спіральної структури і vitro, проведених на матеріалі описаних вище пепамфіпатичних параметрів 22-амінокислотної контидів, були сформульовані деякі «правила» констсенсусної послідовності, яка є похідною від руювання пептидів, які б імітували функції білка спіральних повторів в складі АроА-І. До подиву АроА-І. Істотним є те, що наявність амфіпатичної пептиди по даному винаходу володіють спеα-спіралі, що має позитивно заряджені залишки, цифічною активністю, яка перевершує активність кластеризовані в області гідрофобнопохідних від Аро А-І пептидів, описаних в науковій гідрофільного «інтерфейсу», і негативно заряджені літературі. Дійсно, деякі варіанти по даному винаамінокислоти, кластеризовані по центру ходу забезпечують 100% активності нативного гідрофільної поверхні, необхідно для вияву АроА-І, при тому, що деякі суперагоністи, предліпідної афінності і активації LCAT (Venkatachalaставлені тут, перевершують специфічну активність pathi et al., 1991, цит. вище). Також Anantharamaiah нативного АроА-І. з співавт. встановили, що негативно заряджені Даний винахід проілюстрований експеримензалишки глутамінової кислоти в 13-му положенні в тальними прикладами, в яких описана структура, складі консенсусного 22-амінокислотного пептиду, приготування і застосування конкретних який знаходиться в межах гідрофобної поверхні αамфіпатичних пептидів, які утворять спіралі (в спіралі, грає важливу роль в активації LCAT присутності ліпідів), зв'язують ліпіди, утворюють (Venkatachalapathi et al., 1991, цит. вище). Більш комплекси і підвищують активність LCAT. того Brasseur показав, що гідрофобний кут (кут Ґрунтуючись на структурі і активності варіантів, що «pho») менше за 180° є необхідною умовою для наводяться в прикладах, заявники розробили «звід оптимальної стабільності ліпіду і аполіпопротеїна, правил», який може бути використаний для конста також для утворення дископодібних часток, в руювання змінених або мутованих форм, які також яких пептиди розташовані навколо краю ліпідного входять в обсяг даного винаходу. бі-шару (Brasseur, 1991, J.Biol. Chem., 66, 16120Також даний винахід стосується фармацев16127). Rosseneu з співавт. також продемонструтичних композицій, що містять такі агоністи АроА-І вали, що гідрофобний кут менше за 180° не(або у вигляді самих пептидів, або у вигляді обхідний для активації ферменту LCAT (заявка ліпопротеїнових комплексів) як активний компоWO 93/25581). нент, рівно як і способів приготування таких комОднак, незважаючи на визначення даних позицій і їх застосування для лікування захворю«правил», не вдалося сконструювати і виробити вань, асоційованих з дисліпопротеїнемією пептид, який був би так же активний, як і АроА-І - в (наприклад, з серцево-судинними захворюваннянайкращому варіанті була відзначена менш ніж ми, атеросклерозом, метаболічним синдромом), 40%-а від нативного АроА-І активність, що було рестенозом або появою в крові ендотоксинів (навизначено в описаному в даній заявці тесті на акприклад, як при септичному шоці). тивацію LCAT. Жоден з описаних в науковій 3.1. Скорочення літературі пептидних міметиків не підходив для По використанню в даному тексті прийняті назастосування його в якості лікарського засобу. ступні стандартні скорочення для генетично деЗ урахуванням викладеного вище, є нетермінованих L-амінокислот: обхідність розробки стабільного агоніста АроА-І, який би імітував активність АроА-І і який би міг Стандартна Амінокислота Однобуквений бути зроблений за допомогою відносно простої і символ абревіатура недорогої процедури. Однак, «правила» констАланін А Ala руювання ефективних імітаторів АроА-І не були Аргінін R Arg встановлені, а також залишилися невідомими Аспарагін N Asn принципи конструювання органічних молекул, що Аспарагінова D Asp володіють функціями АроА-І. кислота 3. Резюме винаходу Цистеїн С Cys Даний винахід стосується агоністів АроА-І, Глутамін Q Gin здатних утворювати амфіпатичні α-спіралі, які заГлутамінова Ε Glu безпечують імітацію активності АроА-І, при тому, кислота щоб конкретні рівні активності, наприклад, одиниці 23 71553 24 якому один з атомів водню, приєднаних до кінцевого атома вуглецю, заміщений арильною Стандартна Амінокислота Однобуквений групою. Типовими алкарильними групами, є, тим символ абревіатура самим не вичерпуючись, бензил, бензилиден, бенГліцин G Gly зилидин, бензолбензил, нафтенбензил і подібне. У Гистидин Η His переважних варіантах алкарильною групою є С 6-26 Ізолейцин I Ile алкарил, тобто алкільною, алкенільною або Лейцин L Leu алкінільною складовою алкарильної групи є С 1-6, а Лізин К Lys арильною складовою є С 5-20-арил. У більш переМетіонін Μ Met важних варіантах алкарильною групою є С 6-13 алФенілаланін F Phe карил, тобто алкільною, алкенільною або Пролін Ρ Pro алкінільною складовою алкарильної групи є С 1-3Серин S Ser група, а арильною складовою є С5-10-арил. Треонін Τ Thr «Гетероарил»: означає арильну складову, при Триптофан W Trp тому, що один або декілька атомів вуглецю Тирозин Υ Tyr заміщені іншим атомом, таким як N, Р, О, S, As, Валін V Val Se, Si, Те і т.п. Типовими гетероарильними групами є, тим самим не обмежуючись, акридарсин, Скорочення, що використовуються для познаакридин, арсантридин, арсиндол, арсиндолін, карчення D-енантіомерів генетично детермінованих базол, (3-карболін, хромей, цинолін, фуран, амінокислот, є рядковими аналогами однобуквеімідазол, індазол, індол, індолізин, ізоарсиндол, них позначень. Наприклад, позначення «R» ізоарсинолін, ізобензофуран, ізохромен, ізоіндол, відповідає L-аргініну, а позначення «r» - D-аргініну. ізофосфоіндол, ізофосфінолін, ізохінолін, 3.2. Визначення термінів ізотіазол, ізоксазол, нафтиридин, піримідин, феПо використанню в даному тексті наступні нантридин, фенантролін, феназин, фосфоіндол, терміни повинні прийматися в наступних значенфосфінолін, фталазин, птеридин, пурин, піран, нях: піразин, піразол, піридазин, піридин, піримідин, «Алкіл»: означає насичений розгалужений, непірол, піролізин, хіназолін, хінолін, хінолізин, розгалужений або циклічний вуглеводневий радихіноксалін, селенофен, телурофен, тіофен і ксанкал. Типовими алкільними групами, тим самим не тен. У переважних варіантах гетероарильною груобмежуючись, є метил, етил, пропіл, ізопропіл, пою є 5-20-атомний гетероарил, а більш переважбутил, ізобутил, трет-бутил, пентил, ізопентил, ним є 5-10-атомний гетероарил. гексил і подібне. У переважних варіантах «Алкгетероарил»: означає алкільну, алалкільною групою є Сі_6-алкіл. кенільну або алкінільну групи з нерозгалуженим «Алкеніл»: означає ненасичений розгалужеланцюгом, в якому один з атомів водню, ний, нерозгалужений або циклічний вуглеводневий приєднаних до кінцевого атома вуглецю, радикал, в складі якого є принаймні один заміщений гетероарильною групою. У переважних міжвуглеводневий подвійний зв'язок. Цей радикал варіантах алкгетероарильна група представлена може характеризуватися або цис-, або транс6-26-членним алкгетероарилом, тобто в його конформацією відносно подвійного зв'язку. Типоскладі алкільна, алкенільна або алкінільна складовими алкенільними групами, тим самим не обмева алкгетероарила є С 1-6-групою, а гетероарил є 5жуючись, є етеніл, пропеніл, ізопропеніл, бутеніл, 20-членним гетероарилом. У конкретних переважізобутеніл, трет-бутеніл, пентеніл, гексеніл і них варіантах алкгетероарилом є 6-13-членний подібне. У переважних варіантах алкенільною груалкгетероарил, тобто алкільна, алкенільна або пою є С 1-6-алкеніл. алкінільна складова сполучена з 5-10-атомним «Алкініл»: означає ненасичений розгалужений, гетероарилом. нерозгалужений або циклічний вуглеводневий ра«Заміщені алкіл, алкеніл, алкініл, арил, алкадикал, в рил, гетероарил. або алкгетероарил»: означає складі якого є принаймні один алкільну, алкенільну, алкінільну, арильну, алкаміжвуглеводневий потрійний зв'язок. Типовими рильну, гетероарильну або алкгетероарильну груалкенільними групами, тим самим не обмежуюпу, в якій один або декілька атомів водню заміщені чись, є етиніл, пропініл, бутиніл, ізобутиніл, пеніншим заступником. Переважними заступниками є теніл, гексиніл і подібне. У переважних варіантах групи -OR, -SR, -NRR, -NO2, -CN, галоген, -C(O)R, алкінільною групою є С1-6-алкиніл. C(O)OR і -C(O)NR, де кожний R представлений Η, «Арил»: означає ненасичений вуглеводневий алкілом, алкенілом, алкінілом, арилом, алкарилом, радикал, що характеризується кон'югованою елекгетероарилом або алкгетероарилом. тронною π-системою. Типовими алкенільними гру4. Короткий опис креслень пами, тим самим не обмежуючись, є пента-2,4На фігурі 1А показана кругова схема структури дієн, феніл, нафтил, антрацил, азуленіл, хризеніл, спіралі Шиффера-Едмундсона та ідеалізованої короненіл, флуорантеніл, індаценіл, іденіл, оваамфіпатичної α-спіралі, в якій незафарбовані леніл, периленіл, феналеніл, фенантреніл, піценіл, кільця означають гідрофільні амінокислотні заплейаденіл, піреніл, пірантреніл, рубіценіл і лишки, а затінені кільця представляють гідрофобні подібне. У переважних варіантах арильною групою амінокислотні залишки. є С5-20-арил, а більш переважним є С 5-10-аріл. На фігурі 1В показана схема ідеалізованої «Алкарил»: означає алкільну, алкенільну або спіралі по Фіг.1А у вигляді решітки зі спіраллю. алкінільну групи з нерозгалуженим ланцюгом, в 25 71553 26 На фігурі 1С показана схема ідеалізованої (SEQ ID NO 4) і для консенсусного 22спіралі по Фіг.1А у вигляді циліндра зі спіраллю. амінокислотного пептиду по Segrest (SEQ ID NO На фігурі 2А кругова схема структури спіралі 75). Шиффера-Едмундсона для базового пептиду На фігурі 8В приведений графік, що формули (І), ілюструюча амфіпатичні властивості відображає відмінності між виявленим хімічним спіралі (незафарбовані кільця означають зсувом для протона в складі аміду і хімічним зсугідрофільні амінокислотні залишки, затінені кільця вом для протона в складі аміду, табульованим для представляють гідрофобні амінокислотні залишки, випадкових спіралей, визначених для пептиду 4 а кільця з похилою штриховкою означають або (SEQ ID NO 4) і для консенсусного 22гідрофільні, або гідрофобні амінокислотні залишамінокислотного пептиду по Segrest (SEQ ID NO ки). 75). На фігурі 2В показана схема у вигляді решітки На фігурі 8С показана схема, що відображає зі спіраллю для базового пептиду формули (І), що періодичні відносини протонного хімічного зсуву ілюструє гідрофобну поверхню даної спіралі. На для пептиду 4 (SEQ ID NO 4) і його α-спіральної На фігурі 2С показана схема у вигляді решітки конформації. зі спіраллю для базового пептиду формули (І), що На фігурах 9 A-9D показані гель-хроматограми ілюструє гідрофільну поверхню даної спіралі. виділених HDL людини, що інкубувалися протягом На фігурі 3А показана схема у вигляді решітки 2 годин при 37°С в буфері, з вимірами рівнів позі спіраллю, ілюструюча гідрофільну поверхню, глинання (-), і в присутності 14С-поміченого пептивизначену в роботі Segrest консенсусної 22ду 4 з вимірами рівнів поглинання (---) або амінокислотної послідовності (PVLDEрадіометричного тестування по 14С (♦). ХроматоFREKLNEELEALKQKLK; SEQ ID N O 75). грами були отримані при співвідношеннях маси На фігурі 3В показана схема у вигляді решітки пептиду і HDL, рівних 1:15 (Фіг.9А), 1:10 (Фіг.9В), зі спіраллю, ілюструюча гідрофільну поверхню, що 1:5 (Фіг.9С) і 1:3 (Фіг.9D). є прикладом базового пептиду 4 (PVLDLFRELLНа фігурі 9Е показані результати контрольної 14 NELLEALKQKLK; SEQ ID NO 4). гель-фільтрації вільного незв'язаного СНа фігурі 4А показана схема у вигляді решітки поміченого пептиду 4 за даними вимірювання зі спіраллю, ілюструюча гідрофобну поверхню вирівня поглинання (---) і радіометричного тестуванзначеної в роботі Segrest консенсусної 22ня по 14С (♦). амінокислотної послідовності (SEQ ID NO 75). На фігурі 9F показані гель-фільтраційні хромаНа фігурі 4В показана схема у вигляді решітки тограми, що ілюструють відмінності між параметзі спіраллю, ілюструюча гідрофобну поверхню барами поглинання HDL-пептидних комплексів, зового пептиду 4 що є прикладом (SEQ ID NO 4). представленими на Фіг.9А (---), 9В (-), 9С (---) і 9D На фігурі 5А показана кругова схема спіралі (-) і контрольної хроматограми, показаної на Фіг. Шиффера-Едмундсона для визначеного в роботі 9Е (позитивні значення різниці вказують на більш Segrest консенсусного 22-амінокислотного пептиду високий рівень поглинання в зразках, оброблених (SEQ ID NO 75). білком, що тестується; негативні значення На фігурі 5В показана кругова схема спіралі відповідають більш потужному рівню поглинання в Шиффера-Едмундсона для базового пептиду 4, контрольному зразку). що є прикладом (SEQ ID NO 4). На фігурі 10 показаний графік, що відображає На фігурі 6А представлена комп'ютерна москлад ліпопротеїнів у кроликів, яким ін'єктували дель двох пептидів 4 (SEQ ID NO 4), показаних в 10мг/кг пептиду 4 (SEQ ID NO 4) (у вигляді комантипаралельній орієнтації, в яких залишки Glu-8 і плексів DPPC). Gln-19 виділені, щоб проілюструвати здатність цих На фігурі ПА показана схема, що відображає двох пептидів утворювати міжмолекулярні водневі різні стани агрегації і ліпопептидні комплекси, які зв'язки в процесі скріплення з ліпідами. можуть бути отримані з використанням агоністів На фігурі 6В представлена комп'ютерна моАроА-І по даному винаходу. Зліва: процес мультидель двох пептидів 102 меризації пептидів, що зумовлюється взаємодією (PVLDLFRELLNLXLEALKEKLK; SEQ ID NO 102), декількох пептидних спіралей і що приводить до показаних в антипаралельній орієнтації, в яких утворення олігомерів в умовах точно встановленої залишки Glu-8 і Gln-19 виділені, щоб концентрації білка, рН і іонної сили розчину. У проілюструвати нездатність цих двох пептидів центрі: взаємодія пептидів (в будь-якому стані агутворювати міжмолекулярні водневі зв'язки в прорегації) з ліпідним компонентом (таким як SUV), цесі скріплення з ліпідами. що приводить до реорганізації ліпідів. Праворуч: На фігурі 7А показана схема даного винаходу шляхом зміни молярного відношення ліпіду і пепз розгалуженнями третього порядку. тиду можуть бути отримані різні типи На фігурі 7В показана схема даного винаходу ліпопептидних комплексів - від ліпопептидних з розгалуженнями четвертого порядку. коміцел при низькому співвідношенні На фігурі 7С показана схема даного винаходу «ліпід/пептид» до дископодібних часток і нарешті з розгалуженнями змішаних порядків. до великим мультимерних комплексів при На фігурі 7D показаний приклад схеми даного збільшенні співвідношення «ліпід/пептид». винаходу з розгалуженнями по залишках лізину. На фігурі 11В показана базова модель дискоНа фігурі 8А приведений графік, що подібних ліпопептидних комплексів, що утворювідображає відмінності між виявленим хімічним ються при точно встановлених співвідношеннях зсувом Нα і хімічним зсувом Нα, табульованим для «ліпід/пептид». Кожний пептид, оточуючий край випадкових спіралей, визначених для пептиду 4 27 71553 28 диска, знаходиться в тісному контакті з двома його і далі вона позначається як «консенсусна 22-мірна найближчими сусідами. послідовність Segrest» або «консенсусний 225. Докладний опис винаходу мірний пептид»), який приблизно є критичним по Агоністи Аро А-І по даному винаходу імітують активності синтетичних пептидів, забезпечує синфункціональну активність АроА-І. Вони утворюють тетичним пептидам активність, рівну, а в деяких амфіпатичні спіралі (в присутності ліпідів), здатні експериментах і перевищуючу активність нативнозв'язувати ліпіди, утворюють пре-β-подібні або го Аро А-І. Зокрема, заявники встановили, що HDL-подібні комплекси, активують фермент LCAT, заміна трьох заряджених амінокислотних залишків зумовлюють підвищення концентрації HDL і в консенсусному 22-мірному пептиді Segrest (Gluсприяють виходу холестерину з клітин. Біологічні 5, Lys-9 і Glu-13) на гідрофобний залишок лейцину функції пептидів корелюють з їх спіральною струкпризводить до отримання пептидів, які імітують турою або з конверсією в спіральні структури, що структурні і функціональні параметри АроА-І в мірі, відбувається в присутності ліпідів. яка є безпрецедентною для даної області техніки. Агоністи Аро А-І по даному винаходу можуть Поза зв'язком з якими-небудь теоретичними бути отримані у вигляді стабільних стандартних побудовами, вважається, що спіраль, що утводоз, наприклад, у вигляді ліофілізованих прорюється агоністами АроА-І по даному винаходу, в дуктів, які можуть бути знову відновлені перед більш вираженій мірі імітує стр уктурні і безпосереднім застосуванням in vivo або включені функціональні параметри ділянки амфіпатичної в складу нового препарату. Даний винахід включає спіралі в складі нативного АроА-І, що дуже важлифармацевтичні композиції і застосування таких во з точки зору ефективного скріплення ліпідів, композицій для лікування гіперліпідемії, виходу холестерину з клітин і активації LCAT, в гіперхолестеринемії, ішемічної хвороби серця, порівнянні з тим, що характерно для α-спіралей, атеросклерозу і інших станів, таких як поява ендощо утворюються пептидами-міметиками АроА-І, генних токсинів, що зумовлюється септичним шораніше описаними в науковій літературі, тим саком. мим визначаючи в результаті пептиди, які харакДаний винахід ілюструється експериментальтеризуються істотно більш високою АроА-Іними прикладами, які показують, як агоністи АроАподібною активністю в порівнянні з цими раніше І по даному винаходу взаємодіють з HDLохарактеризованими пептидами. Дійсно, при тому, компонентом плазми крові і можуть збільшува ти що більшість агоністів АроА-І по даному винаходу концентрацію HDL і пре-β-часток. Агоністи АроА-І виявляють рівну, а в деяких варіантах навіть пепо даному винаходу з умовлюють інтенсифікацію ревищуючу активність нативного АроА-І, кращі за виходу (вивільнення з рецепторів) клітинного хохарактеристиками з описаних в літературі лестерину. Також ці агоністи виключно ефективні з міметиків АроА-І пептид 1RAM4 (EWточки зору активації ферменту LCAT, що тим саLEAFYKKVLEKLKELF; SEQ ID NO 246) (Corinjn et мим приводить до сприяння процесу ОТХ. al., 1993, Biochim. Biophys. Acta, 1170, 8-16; Labeur Застосування агоністів Аро А-І по даному виet al., 1994, Arteriosclerosis: Abstracts 186 & 187) і находу в тваринних моделях in vivo з умовлює N-ацетильований С-амідований пептид 18АМ4 підвищення концентрації сироваткового HDL. (SEQ ID NO 239) (Brasseur, 1993, Biochim. Biophys. Більш детально даний винахід описаний в окActa, 1170, 1-7) - виявляють менш ніж 4% і 11% ремих розділах нижче, включаючи: склад і струкактивності, відповідно, від активності нативного туру пептидних агоністів АроА-І, структурні і АроА-І, на що вказують дані вимірів в тесті на акфункціональні характеристики, способи отримання тивацію ферменту LCAT, описаному в даній зазагальних і стандартних доз і способи їх застосуявці. вання. Загалом, базові пептиди (або їх аналоги), які 5.1. Структура і функції пептиду складають агоністи АроА-І по даному винаходу, Агоністи Аро А-І по даному винаходу звичайно характеризуються наступною формулою: є пептидами або їх аналогами, які здатні утворюX1-X2-X3-X4-X5-X6-X7-X8-X9-X10-X11-X12-X13-X14вати амфіпатичні α-спіралі в присутності ліпідів і, X15-X16-X17-X18-X19-X20-X21-X22 , які імітують активність АроА-І. Агоністи включають де в якості своїх основних компонентів базовий Χ1 - пролін (Ρ), аланін (А), гліцин (G), глутамін («серцевинний») пептид, що складається з 15-29 (Q), аспарагін (Ν), аспарагінова кислота (D) або Dамінокислотних залишків, переважно з 22 пролін (р); амінокислотних залишків, або вони є аналогами Х2 - аліфа тична амінокислота; таких пептидів, при тому, що принаймні один Х3 - лейцин (L) або фенілаланін (F); амідний зв'язок в пептиді замінений на заміщений Х4 - кисла амінокислота; амід, на ізостери аміду або міметики аміду. Х5 - лейцин (L) або фенілаланін (F); Агоністи АроА-І по даному винаходу частково Х6 - лейцин (L) або фенілаланін (F); засновані на результатах досліджень, що провоХ7 - гідрофільна амінокислота; дилися заявниками, які із здивуванням встановиX8 - кисла або основна амінокислота; ли, що зміна декількох амінокислотних залишків в Х9 - лейцин (L) або гліцин (G); первинній (амінокислотній) послідовності консенХ10 - лейцин (L), триптофан (W) або гліцин (G); сусного пептиду що складається з 22 Χ11 - гідрофільна амінокислота; амінокислотних залишків, визначеній в роботах Х12 - гідрофільна амінокислота; Venkatachalapathi et al., 1991 (Моl. Conform. & Biol. Х13 - гліцин (G) або аліфатична амінокислота; Interactions, Ind. Acad. Sci., ser. У, 585-596 Х14 - лейцин (L), триптофан (W), гліцин (G) або (PVLDEFREKLNEELEALKQKLK; SEQ ID NO 75: тут нафтилаланін-Nal; 29 71553 30 Х15 - гідрофільна амінокислота; 4404 (група -CH(OH)CH2-); і Hruby, 1982, Life Sci., Х16 - гідрофобна амінокислота; 31, 1890199 (група -CH2-S-). Х17 - гідрофобна амінокислота; Крім того, один або декілька амідних зв'язків Х18 - основна амінокислота, глутамін (Q) або можуть бути замінені імітуючими пептид або аспарагін (N), імітуючими амід складовими, які в скільки-небудь Х19 - основна амінокислота, глутамін (Q) або істотній мірі не порушують структуру і активність аспарагін (N); пептидів. Відповідними амідоміметиками є ті, які Х20 - основна амінокислота; описані, наприклад, Ольсоном з співавт. (Olson et Х21 - аліфатична амінокислота; і al., 1993, J.Med. Chem., 36, 3039-3049). Х22 - основна амінокислота. Критичною властивістю базових пептидів Базові пептиди, що мають формулу (І), частформули (І), є їх здатність утворювати в присутково визначені не конкретними амінокислотами, а ності ліпідів амфіпатичну α-спіраль. Під поняттям їх класами. Характеристики різних класів предста«амфіпатична» розуміється те, що така α-спіраль влені в даному тексті в зв'язку з описом мутантних характеризується наявністю спрямованих в різні і змінених варіантів формули (І). боки гідрофільної і гідрофобної поверхонь, У базових пептидах формули (І), символ «-» орієнтованих вздовж їх довгої осі: тобто одна поміж амінокислотними залишками Хn означає функверхня такої спіралі складена в основному цію послідовного зв'язку осьової структури пептигідрофільними бічними ланцюгами амінокислот, в ду. Отже, символ «-» відповідає пептидному зв'язтой час як протилежна поверхня складена в оску або амідному зв'язку (-C(O)NH-). Повинно бути, новному гідрофобними бічними ланцюгами. На однак, зрозуміло, що даний винахід представляє фіг.1 А і 1В показані дві схематичні версії протипептидні аналоги, в яких один або декілька амідлежних гідрофобної і гідрофільної поверхонь у них зв'язків необов'язково можуть бути замінені на взятої за приклад ідеалізованої α-спіралі. На іншій ніж з амідом зв'язок, переважно на зв'язок з Фіг.1А показана кругова діаграма спіралі по Шифізостерами аміду або із заміщеним амідом. Отже, феру-Едм ундсону (Schiffer & Edmundson, 1967, при тому, що різні залишки Хn в формулі (І) відпоBiophys. J., 7, 121-135). У цьому «колі» довга вісь відають амінокислотам, а переважні варіанти даспіралі перпендикулярна площині листа паперу, на ного винаходу представлені пептидами, для фахіякому знаходиться малюнок. Починаючи з N-кінця, вця в даній області техніки буде зрозуміло, що у подальші амінокислотні залишки (представлені у варіантах з неамідними зв'язками термін «амінокивигляді кілець) розподілені по периметру кола з слота» або «залишок» по використанню в даному інтервалом приблизно в 100°. Таким чином, тексті означає інші біфункціональні складові, що амінокислотний залишок «n+1» відхилений від включають групи, схожі по структурі з бічними лазалишку «n» на 100°, залишок «n+2» відхилений нцюгами амінокислот. на 100° від залишку «n+1» і т.д. Показник у 100° Заміщені аміди загалом включають, тим савідповідає наявності 3,6 залишків амінокислот з мим не вичерпуючись, групи формули -C(O)NH-, в розрахунку на один виток, що звичайно для якій R представлений C1-6-алкілом, заміщеним C1ідеалізованих α-спіралей. На Фіг.1А протилежні C1-6-алкенілом, заміщеним C1-6гідрофільна і гідрофобна поверхні спіралі виразно 6-алкілом, алкенілом, C1-6-алкінілом, заміщеним C1-6видні: гідрофільні амінокислоти показані у вигляді алкінілом, С5-20-арилом, заміщеним С5-20-арилом, незафарбованих кілець, а гідрофобні залишки С6-26-алкарилом, заміщеним С 6-26-алкарилом, 5-20амінокислот показані у вигляді затінених кілець. атомним гетероарилом, заміщеним 5-20-атомним На фігурі 1В показана схема ідеалізованої гетероарилом, 6-26-атомним алкгетероарилом і амфіпатичної спіралі, зображеної і на Фіг.1А, у заміщеним 6-26-атомним алкгетероарилом. вигляді молекулярної решітки (Lim, 1978, FEBS Ізостери аміду загалом включають, тим самим Lett., 89, 10-14). На звичайній схемі спіралі у вине обмежуючись, групи -CH 2NH-, -CH2S-, -СН2СН2-, гляді решітки α-спіраль зображається у вигляді -СН=СН- (цис- і транс-), -С(О)СН2-, -СН(ОН)СН2- і циліндра, який зрізаний вздовж центральної осі CH2SO-. Сполуки, що мають такі неамідні зв'язки, і своєї гідрофільної поверхні і сплощений. Таким способи отримання таких сполук добре відомі в чином, центральна частина гідрофобної поверхні, даній області техніки (див., наприклад, Spatola, що визначається по величині гідрофобного моMarch 1983, Vega Data Vol.1, Issue 3; Spatola, 1983, менту конкретної спіралі (Eisenberg et al., 1982, Peptide Backbone Modifications", In "Chemistry and Nature, 299, 371-374), знаходиться в центрі схеми і Biochemistry of Amino Acids, Peptides & Proteins", орієнтована таким чином, щоб як би підводитися ed. Weinstein, Marcel Dekker, New York, p.267 (банад поверхнею сторінки. Схема циліндра спіралі зовий огляд); Morley, 1980, Trends Pharm. Sci., 1, до «розрізання і сплощення» показана на Фіг.1С. 463-468; Hudson et al., 1979, Intern. J.Prot. Res., 14, При перетинах циліндра різними площинами мо177-185 (групи -CH2NH- і - CH2CH2-); Spatola et al., жуть бути отримані різні плани однієї і тієї ж 1986, Life Sci., 38, 1243-1249 (група-ОВД: Hann, амфіпатичної спіралі: відповідно, може бути отри1982, J.Chem. Soc. Perkin Trans., 1, 307-314 (група мана різна інформація про властивості такої -CH=CH- - цис- і транс-); Almquist et al., 1980, спіралі. J.Med. Chem., 23, 1392-1398 (група -COCH2-); Амфіпатична природа α-спіралі, що утвоJennings-White et al.. Tetrahedron Lett., 23, 2533 рюється в присутності ліпідів базовими пептидами (група -СОСН2-); європейська патентна заявка формули (І), продемонстрована на Фіг.2. На Фіг.2 45664 (1982) CA 97, 39405 (група -СН(ОН)СН2-); показана кругова схема по Шифферу-Едмундсону, Holladay et al., 1983, Tetrahedron Lett., 24, 4401а на Фіг.2В показана схема спіралі у вигляді плоскої решітки з розташованою на передньому плані 31 71553 32 гідрофобною поверхнею і на Фіг.2С показана схеамінокислотних залишків в складі даного пептиду ма у вигляді плоскої решітки з розташованою на N (тобто Ho = å Hi ), передньому плані гідрофільною поверхнею. На i¸1 кожній з Фіг.2А, 2В і 2С гідрофільні залишки предде N - число амінокислотних залишків в пепставлені незафарбованими кільцями, а гідрофобні тиді, а Ні - величина гідрофобності і-ого залишки - затіненими кільцями. Як буде обговорюамінокислотного залишку). Середня гідрофобність ватися з докладними подробицями нижче при роз() - це гідрофобність, співвіднесена до числа гляді змінених або мутованих форм пептидів форамінокислотних залишків (тобто =Ηо/Ν). Загамули (І), деякі амінокислотні залишки можуть бути лом, базові пептиди, які характеризуються середзамінені іншими амінокислотними залишками таньою величиною гідрофобності в межах від -0,050 ким чином, щоб гідрофільна і гідрофобна поверхні до -0,070 з урахуванням оцінки цього показника на спіралі, що утворюється конкретним пептидом, основі нормалізованої шкали гідрофобності за можуть і не перебувати повністю з, відповідно, Ейзенбергом (Eisenberg, 1984, J.Моl. Віоl., 179, гідрофільних і гідрофобних амінокислот. Таким 125-142), розглядаються як такі, що попадають в чином, повинно бути зрозуміло, що при описі обсяг даного винаходу, при тому, що переважним амфіпатичної а-спіралі, що утворюється пептидадіапазоном величин середньої гідрофобності є ми по даному винаходу, слова «гідрофільна подіапазон від -0,030 до -0,055. верхня» вказує на ту поверхню цієї спіралі, яка Загальна гідрофобність гідрофобної поверхні характеризується сумарною гідрофільністю. Слова (Ноpho) амфіпатичної спіралі може бути визначена «гідрофобна поверхня» означають те, що поверхяк сума величин гідрофобності гідрофобних ня пептиду характеризується сумарної амінокислотних залишків, що попадають всередигідрофобністю. ну гідрофобного кута у відповідності до визначеноНе торкаючись конкретних теоретичних побуNН дов, можна вважати, що деякі структурні і (або) го нижче (тобто i Ноpho= å Hi ) фізичні властивості амфіпатичної спіралі, що i =1 утворюється базовими пептидами формули (І), є де Nн - число гідрофобних амінокислотних заважливими для вияву активності. Ці властивості лишків в складі гідрофобної поверхні, a Hj включають рівень амфіпатичності, загальний відповідає визначеному вище). Середня рівень гідрофобності, середню величину гідрофобність гідрофобної поверхні () - це гідрофобності, гідрофобний і гідрофільний кути, відношення Н оpho/Ν, в якому Νн відповідає визнагідрофобний момент, середню величину ченому вище . Загалом, базові пептиди, які харакгідрофобного моменту і сумарний заряд α-спіралі. теризуються величиною в межах від 0,90 При тому, що зображена на Фіг.2А кругова до 1,20 з урахуванням оцінки цього показника на схема представляє стандартний підхід до основі нормалізованої шкали гідрофобності за відображення амфіпатичної природи базових пепЕйзенбергом (Eisenberg, 1984, цит. вище; тидів формули (І), рівень амфіпатичності (суть Eisenberg et al., 1982, цит. вище), розглядаються ступінь асиметрії гідрофобності) може бути станяк такі, що попадають в обсяг даного винаходу, дартним шляхом обчислений шляхом визначення при тому, що переважним діапазоном величин гідрофобного моменту (μн) конкретної спіралі. є діапазон від 0,94 до 1,10. Способи обчислення μΗ для конкретної Гідрофобний кут (кут «pho») звичайно визнаполіпептидної послідовності добре відомі в даній чається як кут або дуга, що утворюється найбільш області техніки і описані, наприклад Ейзенбергом тривалим трактом, складеним гідрофобними (Eisenberg, 1984, Ann. Rev. Biochem., 53, 595-623). амінокислотними залишками у випадку, коли такий Реальна величина μн обчислена для конкретного пептид зображений у вигляді кругової моделі за пептиду, буде залежати від загального числа Шиффером-Едмундсоном (тобто число розташоамінокислотних залишків, що складають даний ваних підряд гідрофобних залишків в такій схемі пептид. Отже, в принципі не є інформативним множиться на 20°). Гідрофільний кут (кут «phi») є порівняння показників μн для пептидів неоднакової різницею між 360° і кутом «pho» (тобто кут 360° довжини. мінус «pho»). Для фахівця в даній області техніки Амфіпатичність пептидів неоднакової довжини буде зрозуміло, що величина кутів «pho» і «phi» може бути піддана прямому порівнянню по ознаці буде, зокрема, залежати від числа амінокислотних усередненого гідрофобного моменту (). Сезалишків в конкретному поліпептиді. Наприклад, редній гідрофобний момент може бути визначений беручи до уваги зображення на фіг. 5А і 5В, можна співвіднесенням величини μΗ до загального числа бачити, що в одному колі схеми Шиффераамінокислот в складі спіралі (тобто =μн/Ν) заЕдмундсона знаходиться тільки 18 амінокислот. галом, базові пептиди, що характеризуються веЗменшення числа амінокислот приводить до утволичиною в межах від 0,45 до 0,65 з урахуванрення розриву в колі, а більша кількість ням оцінки цього показника на основі амінокислот приведе до того, що окремі сайти данормалізованої шкали гідрофобності за Ейзенберної схеми будуть зайняті більш ніж однією гом (Eisenberg, 1984, J.Моl. Віоl., 179, 125-142), амінокислотою. розглядаються як такі, що попадають в обсяг даУ тому випадку, коли пептиди, що складаютьного винаходу, при тому, що переважним ся більш ніж з 18 амінокислотних залишків, такі як діапазоном величин є діапазон 0,50-0,60. базові пептиди формули (І), поняття «безпереЗагальний або повний рівень гідрофобності рвний тракт», складений гідрофобними (Но) пептиду може бути стандартним шляхом обамінокислотними залишками, означає те, що причислений як сума величин гідрофобності кожного з 33 71553 34 наймні одна амінокислота в положеннях по перигідрофобний кластер поблизу N-кінцевої частини метру кола, що включають більше за одну даної спіралі. У центрі цього кластера знаходиться амінокислоту, є гідрофобною амінокислотою. Отзаймаючий 6-е положення залишок фенілаланіну: же, звертаючись до Фіг.5В, кут «pho» являє собою вважається, що він грає важливу роль в дугу, утворену залишками №№5, 16, 9, 2, 13, 6, 17, стабілізації гідрофобного кластера. 10, 3 і 14, незважаючи на присутність в 20-му поНе залучаючи яку-небудь конкретну теорію, ложенні гідрофільного залишку, оскільки залишок можна вважати, що гідрофобний кластер, що в положенні 2, який займає на круговій схемі те ж утворюється залишками 3, 6, 9 і 10, є істотним в положення, що і 20-й залишок пептиду, є точки зору забезпечення ефективного скріплення гідрофобним залишком. Звичайно базові пептиди, ліпідів і активації ферменту LCAT. Наприклад, при що характеризуються кутом гідрофобності в межах тому, що вибраний як приклад пептид 4 (SEQ ID від 160° до 220° розглядаються як такі, що попаNO 4) виявляє 93%-у активацію LCAT в тесті, який дають в обсяг даного винаходу, при тому, що описаний в даній заявці, похідне пептиду 4, що діапазон величин від 180° до 200° є найбільш певключає в 9-му положенні залишок лізину (К) (пепреважним. тид 33: SEQ ID NO 33), який руйнує даний Деякі структурні і (або) фізичні характеристики гідрофобний кластер, виявляє тільки 33%-у ефекбазових пептидів формули (І) проілюстровані на тивність по активації LCAT, визначену в тому ж Фіг.3 і 4. На фігурі ЗВ показана схема у вигляді самому тесті. Ам фіпатичні пептиди, як очікується, решітки вибраного в якості прикладу пептиду 4 по зв'язують фосфоліпіди шляхом розміщення даному винаходу (PVLDLFRELLNELLEALKQKLK; гідрофобних поверхонь відносно алкільних ланSEQ ID NO 4): вона показує розподіл заряду цюгів в складі ліпідних складових. Отже, ввавздовж гідрофільної поверхні спіралі. На Фіг.3В жається, що даний високогідрофобний кластер циліндр зі спіраллю був розтінений вздовж цензабезпечує високий рівень афінності по тральної осі гідрофобної поверхні і сплощений. відношенню до ліпідів, характерний для базових Три гідрофобних залишки лейцину (L), які пептидів по даному винаходу. Оскільки скріплення замінюють залишки консенсусної 22-мірної ліпідів є умовою для активації LCAT, можна ввапослідовності Segrest (Фіг.3А), затінені. Як видно з жати, що даний гідрофобний кластер також є показаного на Фіг.3В, позитивно заряджені істотним з точки зору активації LCAT. амінокислотні залишки кластеризовані на останАроматичні амінокислоти часто розглядаютьньому С-кінцевому витку спіралі (С-кінцева частися, як важливі чинники прикріплення пептидів і на орієнтована догори малюнка). Не зв'язуючись з протеїнів до ліпідів (De Kruijff, 1990, Biosci. Rep., якою-небудь конкретною теорією, можна вважати, 10, 127-130; O'Neil & De Grado, 1990, Science, 250, що кластер основних залишків в С-кінцевій частині 645-551; Blondelle et al., 1993, Biochim. Biophys. стабілізує спіраль за рахунок електростатичних Acta, 1202, 331-336). Таким чином, далі можна взаємодій в зарядженому (NH3+) - спіральному вважати, що залишок фенілаланіна-6, який знаходиполі. Також передбачається, що стабілізація диться в центрі гідрофобного кластера, також момає місце за рахунок гідрофобних взаємодій між же грати ключову роль в прикріпленні базових бічними ланцюгами лізину і серцевинною частипептидів формули (І) до ліпідів. ною спіралі (див. Groebke et al., 1996, Proc. Natl. Взаємодії між базовими пептидами по даному Acad. Sci. USA, 93, 4025-4029; Esposito et al., 1997, винаходу і ліпідами приводять до утворення пепBiopolymers, 41, 27-35). тидно-ліпідних комплексів. Як показано на Фіг.11А, За винятком С-кінцевої частини, в якій кластехарактер такого комплексу (коміцели, дископодібні ризований позитивний заряд, по іншій частині структури, пухирцеподібні структури і багатошагідрофільної поверхні розподілені негативні зарярові утворення), залежить від молярного ди при наявності принаймні одного негативно завідношення в парі «ліпід/пептид», при тому, що рядженого (кислого) амінокислотного залишку з коміцели загалом утворюються при низькому морозрахунку на виток, внаслідок чого утвориться лярному відношенні в комплексі «ліпід/пептид», а безперервний тракт негативних зарядів по три інших типи комплексів утворюються при гідрофільній поверхні даної спіралі. Один позитивпослідовному зростанні молярних відносин у комний заряд пов'язаний з 7-м залишком, який, як плексів «ліпід/пептид». Цей параметр був охаракпередбачається, бере участь в забезпеченні теризований для амфіпатичних пептидів (Epand, стабільності спіралі за рахунок формування соль"The Amphipathic Helix", 1993) і для апопротеїна ового зв'язку («містка») з кислим залишком, розАроА-І (Jones, 1992, "Structure and Function of Apolташованим на сусідньому витку спіралі. ipoproteins", Ch. 8, pp.217-250). Молярне На Фіг.4В показана схема спіралі у вигляді відношення в комплексі «ліпід/пептид» також є решітки гідрофобної поверхні амфіпатичної чинником, що визначає розмір і склад таких комспіралі, що утворюється вибраним як приклад пепплексів (див. розділ 5.3.1 в даній заявці). тидом 4 (SEQ ID NO 4). На Фіг.4В циліндр Довга вісь α-спіралі, що утворюється базовими розтінений вздовж центральної осі гідрофільної пептидами формули (І), загалом має викривлену поверхні і сплощений. Гідрофобна поверхня базоформу. Для типових амфіпатичних спіралей було вого пептиду включає по два гідрофобних залишки встановлено, що довжина водневих зв'язків на на 1 виток спіралі, за винятком останнього Сгідрофільній і гідрофобній поверхнях варіюється кінцевого витка, в якому переважають основні затаким чином, що гідрофобна сторона спіралі є лишки. Аналіз методом ядерно-магнітного резоувігн утою (Barlow & Thornton, 1988, J. МоІ. Biol., нансу показує, що амінокислотні залишки 3, 6, 9 і 201, 601-619; Zhou et al., 1992, J.Amer. Chem. Soc, 10 даного базового пептиду формують 33, 11174-11183; Gasell et al., 1997, J.Biomol. NMR, 35 71553 36 9, 127-135). Поза зв'язком з якою-небудь конкретВзаємодії з утворенням водневого зв'язку будуть ною теорією, можна вважати, що загальне викриввідбуватися між амінокислотами Е8 і Q19 (Huyлення гідрофобної поверхні в спіралі може бути ghues-Despointes et al., 1995, Biochemistry, важливим чинником в процесі скріплення диско34,13267-13271). подібних комплексів - викривлена спіраль дозвоКрім того, при зіставленні в антипаралельній ляє пептиду краще «прилаштовуватися» до країв орієнтації спіралі щільно упаковуються: при цьому дископодібних часток, тим самим збільшуючи відсутні стеричні перешкоди тісному контакту між стабільність пептидно-дископодібного комплексу. спіралями. Зміни в амінокислотній послідовності у У принципі в складі структурної моделі, що базових пептидів, які порушують процес упаковки приймається АроА-І амфіпатичні α-спіралі упакоспіралей, негативно впливають на активність бавуються навколо країв дископодібних HDL (див. зових пептидів. Наприклад, звертаючись до Фіг.6В, Фіг.11У). В цій моделі спіралі, як вважається, укладимер пептидів, в складі яких глутамін-19 даються так, щоб їх гідрофобні поверхні розташозамінений на глутамінову кислоту (пептид 102: вувалися навпроти ацильних ланцюгів ліпідів PVLDLFRELLNLXLEALKEKLK, де X - це α(Brasseur et al., 1990, Biochim. Biophys. Acta, 1043, аміноізобутирова кислота (Aib); )( SEQ ID NO 102), 245-252). Спіралі організовані в антипаралельній тобто вони не можуть утворювати міжмолекулярні орієнтації, а кооперована дія таких спіралей розводневі зв'язки, не здатний активувати LCAT. глядається як важливий чинник стабільності дисІстотним є те, що при тому, що пептид 4 (SEQ ID коподібних комплексів HDL (Brasseur et al., цит. NO 4) виявляє 93%-ий рівень ефективності активище). Передбачається, що одним з чинників, що вації LCAT в тесті, описаному в даній заявці, пепзабезпечують стабільність дископодібних комтид 102 (SEQ ID NO 102) виявляє лише 2% такої плексів HDL, є існування іонних взаємодій між кисактивності в тому ж самому тесті. лими і основними залишками, внаслідок чого утвоТаким чином, не торкаючись якої-небудь конрюються міжмолекулярні сольові «містки» або кретної теорії, можна вважати, що здатність базоводневі зв'язки між залишками на сусідніх антипавих пептидів формули (І) упаковуватися щільним ралельних спіралях. У такій моделі пептиди розобразом і взаємодіяти по іонному типу з утворенглядаються не як окремі елементи, а як чинники ням внутрішньо- і (або) міжмолекулярних сольових взаємодії принаймні двох сусідніх пептидних мо«містків» і (або) водневих зв'язків в процесі лекул (фіг.11В). скріплення з ліпідами в антипаралельній орієнтації Також в принципі можна вважати прийнятим, є важливою властивістю базових пептидів по дащо утворення міжмолекулярного водневого зв'язку ному винаходу. або сольового «містка» між кислим і основним Здатність базових пептидів здійснювати перезалишками, відповідно, по положеннях важні міжмолекулярні пептид-пептидні взаємодії поліпептидної спіралі «і» та «і+3» стабілізує цю також розглядається як важливий чинник і у спіральну стр уктуру (Marqusee et al., 1985, Proc. відсутність ліпідів. Базові пептиди по даному винаNatl. Acad. Sci. USA, 84, 8898-8902). ходу можуть самоасоціюватися, зокрема, через Таким чином, додаткові ключові параметри високі значення їх , і гідрофобного кута базових пептидів формули (І) пов'язані з їх (див. табл.1). Явище самоасоціації залежить від здатністю утворювати міжмолекулярні водневі таких параметрів як рН, концентрація пептиду та зв'язки один з одним, коли вони іонна сила розчину і може виявлятися в утворенні взаєморозташовуються в антипаралельній декількох типів самоасоціацій - від наявності моорієнтації, при тому, що їх гідрофобні поверхні номерних до появи різних мультимерних форм орієнтуються в одному і тому ж напрямі, тобто так, (Фіг.11А). Гідрофобна серцевина пептидної агреяк це відбувається при скріпленні пептидів з гації сприяє гідрофобним взаємодіям з ліпідами. ліпідами (тобто між кислими залишками в 4-му і 8Здатність пептидів агрегуватися навіть при дуже му положеннях і основними залишками в 18-му, низьких їх концентраціях може сприяти їх 20-му і 22-му положеннях), а також з їх здатністю скріпленню з ліпідами. Передбачається, що в серутворювати міжмолекулярні водневі зв'язки або цевинній частині агрегації пептидів міжпептидні сольові «містки» поруч з N- і С-кінцевими взаємодії також мають місце і можуть конкурува ти ділянками конкретної спіралі (тобто між кислими і з пептидно-ліпідними взаємодіями. основними залишками в положеннях 4 і 7 і також У доповнення до описаних вище властиво15 і 18). стей, інші параметри, як можна передбачити, важЗдатність базових пептидів формули (І) утволиві з точки зору активності, включаючи такі вларювати міжмолекулярні водневі зв'язки стивості як загальне число гідрофобних залишків, проілюстровані на Фіг.6А. На Фіг.6А дві загальне число заряджених залишків і кумулятивідеалізовані α-спіралі, утворені вибраним як приний заряд пептидів. клад пептидом 4 (SEQ ID NO 4), зіставлені в антиУзагальнення переважних фізичних і струкпаралельній орієнтації так, щоб їх відповідні турних властивостей базових пептидів формули (І) гідрофобні поверхні розташовувалися в однаковій представлені нижче в таблиці 1. орієнтації (у напрямі від площини сторінки). 37 71553 38 Таблиця 1 Фізичні властивості переважних агоністів АроА-І формули (І) Властивість Частка гідрофобних амінокислот, %% Кут «pho» Частка позитивно заряджених амінокислот Частка негативно заряджених амінокислот Сумарний заряд Гідрофобний кластер Кислий кластер Основний кластер Діапазон Переважний діапазон 40-70 50-60 від -0,050 до -0,070 від-0,030 до-0,055 0,90-1,2 0,94-1,1 0,45-0,65 0,50-0,60 160°-220° 180°-200° 3-5 4 3-5 4 від -1 до +1 0 в положеннях 3, 6, 9 і 10 знаходяться гідрофобні амінокислоти принаймні 1 кисла амінокислота на 1 виток за винятком останніх 5 С-кінцевих амінокислот принаймні 3 основні амінокислоти з 5 останніх С-кінцевих амінокислот Властивості амфіпатичних α-спіралей, що утворюються базовими пептидами по даному винаходу, в істотній мірі відрізняються від властивостей амфіпатичних α-спіралей Α-класу, зокрема, αспіралі Α-класу, характерної для 22-мірної консенсусної послідовності Segrest. Ці відмінності проілюстровані за допомогою вибраного як прикладу пептиду 4 (SEQ ID NO 4) на Фіг.3-5. Звертаючись до Фіг.4А і 4В, можна побачити, що гідрофобна поверхня пептиду 4 характеризується більш могутніми гідрофобними властивостями в порівнянні з гідрофобною поверхнею 22мірного консенсусного пептиду Segrest. Зокрема, залишки 5, 9 і 13 (затінені на Фіг.4В) є в послідовності пептиду 4 гідрофобними залишками лейцину (L) (SEQ ID NO 4) в порівнянні із зарядженими залишками, що є в консенсусному 22мірному пептиді (SEQ ID NO 75). Заміна цих трьох заряджених залишків в консенсусній 22-мірній послідовності Segrest на гідрофобні залишки лейцину (L) приводить до вияву істотних відмінностей по параметрах амфіпатичності, гідрофобності, величині кута «pho» і іншим характеристикам даної спіралі. Порівняння фізичних і структурних властивостей двох вибраних в якості прикладу базових пептидів формули (І) пептиду 4 (SEQ ID NO 4) і пептиду 8 (SEQ ID NO 8) і консенсусної 22-мірної послідовності Segrest (SEQ ID NO 75) представлено нижче в таблиці 2. Таблиця 2 Порівняння властивостей вибраних в якості прикладу пептидів з консенсусним 22-мірним поліпептидом Segrest Властивість Число амінокислот Число гідрофільних амінокислот Число гідрофобних амінокислот Частка гідрофобних амінокислот кут «pho» Число позитивно заряджених Число негативно заряджених Сумарний заряд Важливо зазначити, що базові пептиди формули (І) включають більше число гідрофобних залишків, характеризуються істотно більш високими значеннями і і характеризуються вдвічі більшим кутом «pho» (див. Фіг.5А і 5В). Ці відмінності у властивостя х зумовлюють істотну диференційовку їх активності. При тому, що консенсусний 22-мірний поліпептид Segrest (SEQ ID NO 75) виявляє тільки 10%-у активацію LCAT в порівнянні з нативним АроА-І в тесті, описаному в 22- мір 22 13 9 41% -0,293 0,960 0,425 100° 5 6 -1 пептид 4 22 10 12 55% -0,013 0,990 0,547 200° 4 4 0 пептид 8 22 10 12 55% -0,040 0,940 0,521 200° 4 4 0 даній заявці, пептиди 4 (SEQ ID NO 4) і 8 (SEQ ID NO 8) виявляють 93%-ий і 83%-ий рівні активації LCAT, відповідно, в порівнянні з нативним АроА-І, що оцінювався в тому ж самому тесті. Пептид 1 (PVLDLFRELLNELLEZLK-QKLK; SEQ ID NO 1) і пептид 2 (GVLDLFRELLNELLEALKQKLKK; SEQ ID NO 2) виявляють 120%-у і 105%-у активацію LCAT, відповідно, в порівнянні з нативним АроА-І, що оцінюється в тому ж самому тесті. 39 71553 40 Деякі амінокислотні залишки в складі базових ближче до одного з цих атомів. Полярними пептидів формули (І) можуть бути замінені іншими амінокислотами, що генетично кодуються є аспаамінокислотними залишками без істотного негарагін (Ν), глутамін (Q), серин (S) і треонін (Т). тивного впливу, а в багатьох випадках і з посиленТермін «гідрофобна амінокислота» означає ням активності цих пептидів. Отже, даним винахоамінокислоту, що виявляє властивість дом також представляються змінені або мутовані гідрофобності вище за 0 у відповідності зі станформи базових пептидів формули (І), при тому, що дартною шкалою гідрофобності за Ейзенбергом принаймні один певний амінокислотний залишок в (Eisenberg et al., 1984, J.МоІ. Biol., 179, 125-142). структурі замінюється іншим амінокислотним заГідрофільні амінокислоти, що генетично кодуються лишком. Оскільки одна з критичних властивостей, включають пролін (Р), ізолейцин (І), фенілаланін що впливають на активність базових пептидів по (F), валін (V), лейцин (L), триптофан (W), метіонін даному винаходу, розглядається як здатність (М), аланін (А), гліцин (G) і тирозин (Y). утворювати в присутності ліпідів α-спіралі, які виТермін «ароматична амінокислота» означає являють амфіпатичні і інші властивості, описані гідрофобну амінокислоту, в бічному ланцюгу якої є вище, то повинно бути зрозуміло, що в переважпринаймні одне ароматичне або гетероциклічне них варіантах даного винаходу амінокислотні кільце. Ароматичне або гетероциклічне кільце мозаміни є консервативними, тобто замінюючи же включати один або більше число заступників, амінокислотні залишки характеризуються таких як -ОН, - SH, -CN, -F, -СІ, -Br, -I, -NO 2, -NO, фізичними і хімічними властивостями, схожими з NH2, -NHR, -NRR, -C(O)R, -C(O)OH, -C(O)OR, такими у амінокислотних залишків, що C(O)NH2, -C(O)NHR, -C(O)NRR і подібне, де кожзамінюються. ний з R незалежно представлений Сі.6-алкілом, Для цілей визначення консервативних заміщеним С1-6-алкілом, С1-6-алкенілом, амінокислотних замін амінокислоти можуть бути заміщеним С1-6-алкенілом, С1-6-алкинілом, стандартним чином класифіковані на дві базові заміщеним С1-6-алкинілом, С5-20-арилом, категорії - гідрофільні і гідрофобні: насамперед, заміщеним С5-20-арилом, С6-26-алкарилом, розділення залежить від фізико-хімічних характезаміщеним С6-26-алкарилом, 5-20-атомним гетеристик бічних ланцюгів амінокислот. Ці дві основні роарилом, заміщеним 5-20-атомним гетероарикатегорії можуть бути далі класифіковані на дві лом, 6-26-атомним алкгетероарилом і заміщеним підгрупи, які більш детально характеризують вла6-26-атомним алкгетероарилом. Генетичними стивості бічних ланцюгів амінокислот. Наприклад, ароматичними амінокислотами, що кодуються є категорія гідрофільних амінокислот може бути далі фенілаланін (F), тирозин (Y) і триптофан (W). поділена на кислі, основні і полярні амінокислоти. Термін «неполярна амінокислота» означає Категорія гідрофобних амінокислот може бути далі гідрофобну амінокислоту, що характеризується поділена на неполярні і ароматичні амінокислоти. незарядженим бічним ланцюгом при фізіологічних Визначення різних категорій амінокислот, які визначеннях рН і що має зв'язки, в яких пара елекзначають склад формули (І), такі. тронів, що є спільними для двох атомів, знахоТермін «гідрофільна амінокислота» означає дяться на рівній відстані від кожного з цих атомів амінокислоту, що виявляє властивість (тобто бічний ланцюг не є полярним). Неполярнигідрофобності нижче за 0 у відповідності зі станми амінокислотами, що генетично кодуються є дартною шкалою гідрофобності за Ейзенбергом лейцин (L), валін (V), ізолейцин (І), метіонін (М), (Eisenberg et al., 1984, J.МоІ, Biol., 179, 125-142). гліцин (G) і аланін (А). Гідрофільні амінокислоти, що генетично кодуються Термін «аліфатична амінокислота» означає включають треонин (Т), серии (S), гістидин (Н), гідрофобну амінокислоту, що характеризується глутамінову кислоту (Е), аспарагін (Ν), глутамін наявністю аліфатичного вуглеводневого бічного (Q), аспарагін (D), лізин (К) і аргінін (R). ланцюга. Алі фатичними амінокислотами, що генеТермін «кисла амінокислота» означає тично кодуються є аланін (А), валін (V), лейцин (L) гідрофільну амінокислоту, в якої бічний ланцюг і ізолейцин (І). характеризується значенням рК, меншим 7. Кислі Амінокислотний залишок цистеїну (С) незвиамінокислоти звичайно мають негативно зарядчайний в тому, що він може утворювати дижені бічні ланцюги при фізіологічних значеннях рН сульфідні зв'язки з іншими залишками (С) або через втрату іона водню. Кислими іншими амінокислотами, що містять сульфонільну амінокислотами, що генетично кодуються є глюгруп у. Зда тність залишків цистеїну (С) (як і інших тамінова кислота (Е) і аспарагінова кислота (D). амінокислот, що мають групу -SH як бічний ланТермін «основна амінокислота» означає цюг) існува ти як у відновленій формі -SH, так і в гідрофільну амінокислоту, в якої бічний ланцюг окисленому стані у вигляді дисульфідного «містка» характеризується значенням рК, більшим за 7. впливає на внесок цистеїну (С) у величину Кислі амінокислоти звичайно мають позитивно гідрофобності/гідрофільності пептиду. Якщо цисзаряджені бічні ланцюги при фізіологічних значентеїн (С) характеризується величиною нях рН через втрату іона гідроксонія. Кислими гідрофобності 0,29 згідно з нормалізованою шкаамінокислотами, що генетично кодуються є лою за Ейзенбергом (Eisenberg, 1984, цит. вище), гістидин (Н), аргінін (R) і лізин (К). то повинно бути зрозуміло, що, ви ходячи з цілей Термін «полярна амінокислота» означає даного винаходу, цей залишок (С) класифікується гідрофільну амінокислоту з незарядженим бічним як полярна гідрофільна амінокислота, незалежно ланцюгом при фізіологічних значеннях рН, але в від загальних установок для описаної вище клаякої є принаймні один зв'язок, в якому пара елексифікації. тронів, спільна для двох атомів, знаходиться 41 71553 42 Як повинно бути зрозуміло фахівцеві в даній доповнення до амінокислот, що зустрічаються в області техніки, визначені вище категорії не є живій природі і тим, що кодуються генетично, взаємовиключними. Отже, амінокислоти, що хаамінокислоти базових пептидів формули (І) морактеризуються бічними ланцюгами, що виявляжуть бути заміщені з використанням амінокислот ють два і більше типів фізико-хімічних властивощо зустрічаються в природі, але й амінокислот, що стей можуть бути віднесені до «множинних генетично не кодуються, а також синтетичних категорій». Наприклад, бічні ланцюги амінокислот, амінокислот. що включають ароматичні складові, які крім того Деякі амінокислоти, що звичайно розглядазаміщені полярними заступниками, такі як тирозин ються, які представляють перспективний об'єкт (Y), можуть виявляти і ароматичні гідрофобні владля замін до складу базових пептидів формули (І), стивості, і полярні або гідрофільні властивості, включають, тим самим не обмежуючись, β-аланін тобто, відповідно, можуть бути включені в категорії (β-Ala) і інші ω-амінокислоти, такі як 3і ароматичних, і полярних амінокислот. Конкретна амінопропіонова кислота, 2,3-диамінопропіонова класифікація будь-якої амінокислоти буде очевидкислота (Dpr), 4-амінобутирова кислота і т.д.; αною для фахівця в даній області техніки, зокрема, аміноізобутирову кислоту (Aib), ε-аміногексаноєву в світлі докладних описів, включених в даний кислоту (Aha), δ-аміновалеріанову кислоту (Ava), текст. N-метилгліцин або саркозин (MeGly), орнитин Деякі амінокислотні залишки, що позначають(Оrn), цитрулин (Cit), трет-бутилаланін (t-BuA), ся як «розриваючи спіраль амінокислоти», мають трет-бутилгліцин (t-BuG), N-метилізолейцин властивість розривати структуру α-спіралей тоді, (Меllе), фенілгліцин (Phg), циклогексилаланін коли вони знаходяться у внутрішніх положеннях (Cha), норлейцин (Nle), нафтилаланін (Nal), 4такої спіралі. Амінокислотні залишки, що виявляхлорфенілаланін (Phe-4-Cl), 2-фторфенілаланін ють такі властивості розривання спіралей, добре (Phe-2-F), 3-фторфенілаланін (Phe-3-F), 4-фторвідомі в даній області техніки (див., наприклад, фенілаланін (Phe-4-F), пеніциламін (Pen), 1,2,3,4Chou & Fasman, Ann. Rev. Biochem., 47, 251-276) і тетрагідроізохінолін-3-карбонову кислоту (Tic), β-2включають пролін (Р), гліцин (G) і потенційно всі Dтієнілаланін (Thi), метіонінсульфоксид (MSO), гоамінокислоти (тоді, коли вони «попадають» до моаргінін (hArg), N-ацетилізин (AcLys), 2,4складу L-амінокислотного пептиду; і навпаки, Lдиамінобутировая кислота (Dbu), 2,3амінокислоти розривають спіральні структури, що диамінобутировая кислота (Dab), pутворюються D-пептидом). Хоч ці «розриваючи амінофенілаланін (Phe-pNH2), N-метилвалін (Meспіраль» амінокислоти і попадають в позначені Val), гомоцистеїн (hCys), гомофеніл-аланін (hPhe) і вище категорії, за винятком гліцину (G) (що буде гомосерин (hSer), гідроксипролін (Hyp), гомопролін обговорюватися в даному тексті), ці залишки не (hPro), N-метильовані амінокислоти і пептоїди (Nповинні використовуватися для заміни заміщені гліцини). амінокислотних залишків у внутрішні х положеннях Класифікації генетично кодованих і звичайних в межах спіралі - тобто вони можуть бути викоринекодованих амінокислот відповідно до категорій, стані тільки для заміни 1-3 амінокислотних завизначених вище, підсумовані нижче в табл.3. Полишків в складі N-кінцевої частини і (або) Свинно бути зрозуміло, що таблиця 3 наведена кінцевої частини конкретного пептиду. тільки як ілюстративний матеріал і не покликана При тому, що в якості прикладів в описаних служити вичерпним списком амінокислот, які б вище категоріях були приведені амінокислоти, могли бути використані для замін до складу базогенетично детерміновані (що кодуються), проте вих пептидів, що представляються даним винахонемає необхідності, а в деяких конкретних дом. Інші, конкретно не обумовлені в даному варіантах і переважно немає необхідності в тексті, амінокислоти легко можуть бути клазамінах, що обмежуються амінокислотами, що сифіковані на основі їх фізичних і хімічних властигенетично кодуються. Дійсно, багато які з перевостей в світлі характеристик, описаних в даному важних пептидів формули (І) включають тексті. амінокислоти, що не кодуються генетично. Отже, в Таблиця 3 Класифікація амінокислот, що звичайно використовуються Класифікація Гідрофобні -""- ароматичні -""- неполярні -””- аліфатичні Гідрофільні -""- кислі -""- основні -""- полярні «розривачі спіралі» Генетично кодовані Генетично некодовані F, Y, W L, V, І, М, G, А, Р А, V, L, І Phg, Nal, Thi, Tic, Phe-4-Cl, Phe-2-F, Phe-3-F, Phe-4-F, hPhe t-BuA, t-BuG, Melle, Nle, MeVal, Cha, MeGl y, Aib β-Ala, Dpr, Aib, Aha, MeGl y, t-BuA, t-BuG, Melle, Cha, Nle, Me Val D,E Η, ДО, R С, Q, Ν, S, T P, G Dpr, Orn, hArg, Phe-pNH2, Dbu, Dab Cit, AcL ys, MSO, β-Ala, hSer D-Pro і інші D-амінокислоти в складі L-пептидів 43 71553 При тому, що в більшості прикладів амінокислоти базових пептидів формули (І) заміняють з використанням L-амінокислот, заміни не обмежуються L-амінокислотами. Отже, також в поняття «змінені» або «мутовані» форми входять ті форми, які утворені внаслідок заміни Lамінокислоти на ідентичну D-амінокислоту (наприклад, L-аргінін →D-аргінін) або на Dамінокислоту тієї ж категорії або підгрупи (наприклад, L-аргінін →D-лізин), і навпаки. Дійсно, в деяких переважних варіантах, які є такими в зв'язку з пероральним введенням пацієнтам-ссавцям, доцільним є входження до складу пептиду, що вводиться принаймні однієї D-амінокислоти. Пептиди, що включають такі D-амінокислоти, як вважається, більш стійкі до руйнування факторами ротової порожнини, кишечнику або сироватки крові в порівнянні з пептидами, що складаються виключно з L-амінокислот. Як відзначалося вище, D-амінокислоти характеризуються тенденцією до руйнування α-спіралей у разі заняття ними внутрішніх положень в αспіралі, що утворюється L-пептидом. Більш того було встановлено, що деякі мутовані форми базових пептидів формули (І), які складаються виключно з D-амінокислот, виявляють істотно меншу здібність до активації ферменту LCAT, що визначається в описаному в даній заявці тесті, в порівнянні з ідентичними по послідовності пептидами, що складаються виключно з L-амінокислот. Отже, D-амінокислоти не повинні використовуватися для заміни L-амінокислот, що знаходяться у внутрішніх положеннях: заміни на D-амінокислоти повинні обмежуватися 1-3 амінокислотними залишками в N-кінцевій і (або) С-кінцевій частинах конкретного пептиду. Як обговорювалося вище, амінокислота гліцин (G) загалом є «руйнуючим спіраль» залишком в тих випадках, коли вона знаходиться у внутрішніх положеннях пептиду. Вельми дивним заявниками представляється встановлення того, що, хоч спіральна структура базових пептидів по даному винаходу руйн ується у відсутність ліпідів тоді, коли внутрішні по положенню залишки замінюються на гліцин (G), в присутності ліпідів такі включаючи гліцин (G) пептиди виявляють істотні позитивні властивості з утворення спіралі, так і по вияву активності. Наприклад, при тому, що пептид 8 (SEQ ID NO 8) виявляє тільки 20% утворення спіралі в буфері, 61-93%-а інтенсивність утворення спіралі спостерігалася в присутності ліпідів і 93%-а спіралізація була виявлена в присутності трифторетанола (TFE). Спіральна структура даного пептиду в присутності TFE була підтверджена із застосуванням ЯМР (див. тут розділ 7.3.5). Цікаво, що цей же пептид виявляв 83%-у ефективність по активації LCAT. Ін ші базові пептиди, що включають у внутрішніх положеннях залишки гліцину, також виявляють активацію LCAT на рівні >38% (див., наприклад, табл.10 в розділі 8.3.). Таким чином, хоч гліцин (G) загалом розглядається як «руйнуючий спіраль» залишок, проте гліцин (G) може бути використаний для заміни амінокислот у внутрішніх положеннях базових пептидів формули (І). Переважно, щоб тільки внутрішні залишки, роз 44 ташовані на відстані не більше за один виток в кожний з боків від центральної осі пептиду (зокрема, у випадку пептидів, що складаються з парного числа амінокислот), замінювалися на гліцин (G). Крім того, переважно, щоб тільки один внутрішній амінокислотний залишок пептиду був заміщений на залишок гліцину (G). Переважні варіанти агоністів АроА-І по даному винаходу, які включають у внутрішніх положеннях залишки гліцину, описані тут в розділі 5.1.2. З використанням класифікації амінокислотних залишків, представленої вище, в поєднанні з круговою схемою по Шифферу-Едмундсону і схемою у вигляді решітки, що застосовуються для базових пептидів формули (І), а також докладного опису бажаних властивостей, можуть бути легко отримані змінені або мутовані форми базових пептидів формули (І), які в істотній мірі зберігають амфіпатичні і інші властивості спіралі і які, отже, попадають в обсяг даного винаходу. У переважному варіанті по даному винаходу змінені і мутовані форми базових пептидів формули (І) отримують шляхом збереження незмінними (фіксації) гідрофільних або гідрофобних залишків відповідно до формули (І) і заміни принаймні одного нефіксованого залишку на іншу амінокислоту, переважно на іншу амінокислоту з тієї ж категорії або підгрупи. Залишки, що складають основний і (або) гідрофобний кластери також можуть бути фіксовані, а принаймні один нефіксований залишок - замінений. В іншому переважному варіанті змінені або мутовані форми базових пептидів формули (І) отримують шляхом фіксації гідрофільних амінокислотних залишків, розташованих на гідрофільній поверхні спіралі відповідно до формули (І) і заміни принаймні одного з нефіксованих амінокислотних залишків на іншу амінокислоту, переважно на іншу амінокислоту з складу тієї ж самої категорії або підгрупи. Звертаючись до Фіг.2 А, можна бачити, що залишки 1, 4, 7, 8, 11, 12, 15, 18, 19 і 22 розташовані на гідрофільній поверхні амфіпатичної спіралі, що утворюється базовими пептидами формули (І). Серед цих залишків всі є гідрофільними, за винятком 1-го залишку, який може бути або гідрофільним, або гідрофобним. Отже, в одному з переважних варіантів залишки 4, 1, 8, 11, 12, 15, 18, 19 і 22 є фіксованими відповідно до формули (І) і принаймні один із залишків 2, 3, 5, 6, 9, 10, 13, 14, 16, 17, 20 і 21 замінений на іншу амінокислоту тієї ж самої категорії, переважно на іншу амінокислоту тієї ж самої підгрупи. З іншого боку, 1-й залишок також є фіксованим відповідно до формули (І) і принаймні один із залишків 2, 3, 5, 6, 9, 10, 13, 14, 16, 17, 20 і 21 замінений відповідно до описаного. У конкретному переважному варіанті Скінцевий основний кластер (залишки 18, 19, 20 і 22) також є фіксованим відповідно до формули (І) і тільки залишки 2, 3, 5, 6, 9, 10, 13, 14, 16, 17 і (або) 21 можуть бути замінені. В іншому конкретному переважному варіанті гідрофобний кластер також фіксується і тільки залишки 2, 5, 13, 14, 16,17, 20 і (або) 21 можуть бути замінені. 45 71553 46 Ще в одному конкретному переважному Science, 240, 1648-1652; Harper et al., 1993, Bioваріанті і основний, і гідрофобний кластери chemistry, 32, 7605-7609; Dasgupta & Bell, 1993, фіксуються і тільки залишки 2, 5, 13, 14, 16, 17 і Intern. J.Peptide Protein Res, 41, 499-511; Seale et (або) 21 можуть бути замінені. al., 1994, Protein Sci, 3, 1741-1745; Doig et al., 1994, В іншому переважному варіанті даного винаBiochemistry, 33, 3396-3403; Zhou et al., 1994, Proходу змінені або мутовані форми базових пептидів teins, 18, 1-7; Doig & Baldwin, 1995, Protein Sci, 4, даного винаходу отримують шляхом фіксування 1325-1336; Odaert et al., 1995, Biochemistry, 34, гідрофобних амінокислотних залишків, розташо12820-12829; Petrukhov et al., 1996, Biochemistry, ваних на гідрофобній поверхні спіралі і заміни 35, 387-397; Doig et al., 1997, Protein Sci, 6, 147принаймні одного нефіксованого амінокислотного 155). 3 іншого боку, крайні 1-4 N-кінцеві і (або) Сзалишку на інший амінокислотний залишок, перекінцеві амінокислотні залишки формули (І) можуть важно на інший залишок з тієї ж категорії або бути замінені пептидоімітуючими складовими, які підгрупи амінокислот. імітують структуру і (або) властивості кепуючих Звертаючись до Фіг.2А, можна бачити, що зазалишків або сегментів. Відповідні міметики-кепи лишки 2, 3, 5, 6, 9, 10, 13, 14, 16, 17, 20 і 21 розтадобре відомі в даній області техніки і описані, нашовані в межах гідрофобної поверхні. З них всі приклад, у Richardson & Richardson, 1988, Science, амінокислоти є гідрофобними, за винятком 240, 1648-1652; Harper et al, 1993, Biochemistry, 32, гідрофільного 20-го залишку. Отже, в одному з 7605-7609; Dasgupta & Bell, 1993, Intern. J.Peptide переважних варіантів залишки 2, 3, 5, 6, 9, 10, 13, Protein Res, 41, 499-511; Seale et al, 1994, Protein 14, 16, 17 і 21 є фіксованими відповідно до форSci, 3, 1741-1745; Doig et al., 1994, Biochemistry, 33, мули (І) і принаймні один із залишків 1, 4, 7, 8, 11, 3396-3403; Zhou et al., 1994, Proteins, 18, 1-7; Doig 12, 15, 18, 19, 20 і 22 замінений на інший & Baldwin, 1995, Protein Sci., 4, 1325-1336; Odaert амінокислотний залишок, переважно на інший et al., 1995, Biochemistry, 34, 12820-12829; Petrukамінокислотний залишок з тієї ж самої категорії hov et al., 1996, Biochemistry, 35, 387-397; Doig et або підгрупи. al.. Protein Sci., 6,147-155. У конкретному переважному варіанті СПри тому, що формула (І) включає 22 кінцевий основний кластер також фіксують, і тільки амінокислотних залишки, повинно бути зрозуміло, залишки 1, 4, 7, 8,11, 12 і (або) 15 можуть бути що базові пептиди по даному винаходу можуть замінені. включати менше за 22 амінокислотних залишки. В іншому варіанті змінені або мутовані форми Дійсно, укорочені або делетовані по внутрішніх пептидів формули (І) отримують шляхом фіксації положеннях варіанти формули (І), що включають всіх амінокислотних залишків, що знаходяться в 18 і навіть 15 амінокислотних залишків, які в межах гідрофобної або гідрофільної поверхонь істотній мірі зберігають основні характеристики і даної спіралі, і заміни, переважно по консервавластивості амфіпатичної спіралі, що утворюється тивній схемі, принаймні одного амінокислотного базовими пептидами формули (І), розглядаються залишку, що знаходиться на протилежній пояк такі, що входять в обсяг даного винаходу. верхні, на інший амінокислотний залишок. ЗалишУкорочені форми пептидів формули (І) отрики, що формують гідрофобний кластер і (або) осмують шляхом делетування однієї або більшого новний кластер, також можуть бути необов'язково числа амінокислот з N- і (або) С-кінців формули (І). фіксованими відповідно до формули (І) відповідно Делетовані по внутрішніх ділянках варіанти фордо визначеного вище. мули (І) отримують шляхом делетування однієї В іншому варіанті даного винаходу змінені або або більшого числа амінокислот з внутрішніх помутовані форми формули (І) отримують шляхом ложень пептиду формули (І). Делетовані внутрішні заміни принаймні однієї амінокислоти на іншу амінокислотні залишки можуть розташовува тися, а амінокислоту по неконсервативній схемі. Для можуть і не розташовуватися підряд. фа хівця в даній області техніки буде зрозуміло, що Для фахівців в даній області техніки буде зроподібні заміни не будуть в істотній мірі змінювати зуміло, що делетування внутрішньої амінокислоти амфіпатичні і (або) структурні властивості спіралі, з базового пептиду формули (І) повинно обумовиякі розглядалися вище. Отже, у деяких випадках ти розворот площини гідрофільно-гідрофобного бажаною може бути заміна однієї або декількох розділу на 100° в точці делеції. пар амінокислот таким чином, щоб зберегти суОскільки таке обертання може привести до марні властивості всієї спіралі. Додаткові вказівки істотних змін амфіпатичних властивостей спіралі, по вибору відповідних амінокислотних замін предщо утворюється внаслідок, то в переважному ставлені послідовностями пептидів, приведеними варіанті даного винаходу амінокислотні залишки в табл.10 (див. розділ 8.3. даного тексту). делетують таким чином, щоб в істотній мірі збеЩе в одному варіанті даного винаходу крайні регти розташування площини гідрофільно1-4 амінокислотні залишки по N-і (або) С-кінцю гідрофобного зіткнення вздовж всієї довгої осі дабазових пептидів формули (І) замінюють на один ної спіралі. або декілька амінокислотних залишків або на один Цього можна досягти стандартним за рахунок або декілька пептидних сегментів, для яких відомі делетування достатнього числа розташованих або властивості по забезпеченню стабільності ділянок нерозташованих підряд амінокислот таким чином, вторинної структури, створюючи α-спіраль (залишщоб в результаті був видалений повний виток ки або сегменти т.зв. «кінцевих кепов», або сегспіралі. Ідеалізована α-спіраль містить 3,6 залишменти «кепування»). Такі «копіюючи» залишки і ки на виток. Отже, в переважному варіанті делесегменти добре відомі в даній області техніки тують групи з 3-4 послідовно розташованих або (див., наприклад, Richardson & Richardson, 1988, нерозташованих підряд амінокислот. Буде делету 47 71553 48 ватися 3 амінокислоти, або 4 амінокислоти, буде важно представлений лейцином (L); Х18 переважзалежати від положення в межах спіралі першого но представлений лізином (К); Х19 переважно із залишків, що делетуються. Визначення реальнопредставлений глутаміном (Q); Х20 переважно го числа розташованих або нерозташованих представлений лізином (К); Х21 переважно предпідряд амінокислотних залишків, які складають ставлений лейцином (L); і (або) Х22 переважно один повний виток спіралі, починаючи з будь-якої представлений лізином (К). конкретної початкової точки в межах амфіпатичної Також в обсяг даного винаходу входять «заспіралі, є досить простою задачею для фахівця в блоковані» форми агоніста АроА-І, тобто такі даній області техніки. варіанти агоністів АроА-І, в яких Ν- і (або) С-кінець Через передбачувану важливість основного блокуються складовими, спроможними реагувати з кластера, що знаходиться на С-кінці базових пепΝ- кінцевою групою -ΝΗ2 або з С-кінцевою групою тидів формули (І) в забезпеченні стабілізації СООН. Було встановлено, що видалення Ν- і (або) спіралі і важливості гідрофобного кластера в заС-кінцевого заряду у агоністів АроА-І по даному безпеченні ефективності скріплення ліпідів і активинаходу, що складаються з 18 або меншого чисвації LCAT, в переважних варіантах даного винала амінокислотних залишків (шляхом синтезу Nходу залишки, що складають основний і ацильованих пептидних гідрофобний кластери, не делетують. Отже, в пеамідів/ефірів/гідразидів/спиртів і їхніх заступників) реважних варіантах залишки 18, 19, 20 і 22 (осзумовлює утворення агоністів, які зберігають і в новний кластер) і залишки 3, 6, 9 і 10 (гідрофобний деяких варіантах навіть перевершують активність кластер) не делетують. незаблокованого варіанту конкретного агоніста. В Базові пептиди формули (І) також можуть бути деяких варіантах при наявності 22 або більшого подовжені по одному з своїх кінців або добудовані числа амінокислотних залишків N- або С-кінцеве по внутрішніх положеннях з використанням додатблокування зумовлює отримання агоністів АроА-І, кових амінокислот, які в істотній мірі не порушуваякі виявляють меншу активність в порівнянні з нели б, а в деяких варіантах навіть посилювали б заблокованими варіантами. Однак, блокування структурні і (або) функціональні параметри пепобох кінців (N і С) агоністів АроА-І, що складаютьтидів. Дійсно, подовження базових пептидів до 23, ся з 22 або більшого числа амінокислот, як 25, 26, 29 і навіть більшого числа амінокислотних очікується, приведе до відновлення активності. залишків потрапляє в обсяг даного винаходу. ПеОтже, в переважному варіанті по даному винаходу реважно, щоб такі подовжені пептиди в істотній або N- і (або) С-кінець (переважно обидва кінці) мірі зберігали загальні параметри амфіпатичності і базових пептидів, що складаються з 18 або менінші властивості пептидів формули (І). Звичайно, шого числа амінокислот, заблоковані, в той час як повинно бути зрозуміло, що додання амінокислот N- і С-кінці пептидів, що складаються з 22 або всередині пептиду приведе до розвороту площини більшого числа амінокислот, або обидва заблокогідрофобно-гідрофільного інтерфейсу в точки вбувані, або обидва не заблоковані. Звичайно Nдування по тому ж типу, як це відбувається у опикінцеві блокуючи групи включають групу RC(O)-, саних вище випадках внутрішнього делетування. де R представлений -Н, С1-6-алкілом, С1-6Отже, висновки, зроблені вище при описі алкенілом, С1-6-алкинілом, С5-20-арилом, С6-26внутрішніх делецій, в рівній мірі застосовні і для алкарилом, 5-20-атомними гетероарилом або 6внутрішніх вставок. 26-атомними алкгетероарилом. Переважними NВ одному з варіантів базові пептиди подовжукінцевими блокуючими групами є ацетил, форміл і ють по N- і (або) С-кінцю принаймні на один виток дансил. Звичайні С-кінцеві блокуючи групи вклюспіралі. Переважно, щоб такі добудови зумовлючають групи -C(O)NRR і -C(O)OR, де R незалежно вали стабілізацію повторної спіральної структури в визначаються відповідно до вказаного вище. Пеприсутності ліпідів в тій же мірі, що і описані вище реважними С-кінцевими блокуючими групами є ті, кеповані амінокислотами і сегментами пептиди. в яких гр упа R незалежно представлена метилом. У конкретному переважному варіанті базові Не торкаючись якої-небудь конкретної теорії, можпептиди формули (І) добудовують по С-кінцю одна вважати, що такі кінцеві блокуючи групи ним амінокислотним залишком, переважно стабілізують α-спіраль в присутності ліпідів (див., лізином (К). Після добудови X1 переважно преднаприклад, Venkatachelapathi et al., 1993, Proteins: ставлений D-проліном (р) або гліцином (G); Х2 пеSctruct, Funct. & Genet., 15, 349-359). реважно представлений валіном (V); Х3 переважно Нативна структура Аро А-І включає вісім представлений лейцином (L); Х4 переважно предспіральних одиниць, які, як вважається, діють ставлений аспарагіновою кислотою (D); Х5 перевідповідно до процесу скріплення ліпідів (Nakaважно представлений лейцином (L); Х6 переважно gawa et al., 1985, J.Amer. Chem. Soc, 107, 7087представлений фенілаланіном (F); Х7 переважно 7092; Anantharamaiah et al., 1985, J.Biol. Chem., представлений аргініном (R); Х8 переважно пред260, 10248-10262; Vanloo et al., 1991, J.Lipid Res., ставлений глутаміновою кислотою (Е); Х9 пере32, 1253-1264; Mendez et al., 1994, J.Clin. Invest., важно представлений лейцином (L); Х10 переваж94, 1698-1705; Palgunari et al., 1996, Arterioscleroно представлений лейцином (L); Х11 переважно sis, Thrombosis & Vasuclar Biol., 16, 328-338; Deпредставлений аспарагіном (N); Х12 переважно moor et al., 1996, Eur. J.Biochem., 239, 74-84). Отпредставлений глутаміновою кислотою (Е); Х13 же, також в обсяг даного винаходу входять переважно представлений лейцином (L); Χ14 переагоністи АроА-І, що є димерами, тримерами, тетважно представлений лейцином (L); Х15 переважрамерами і навіть полімерами більш високого поно представлений глутаміновою кислотою (Е); Х16 рядку· («мультимери») базових пептидів, описаних переважно представлений аланіном (А); Хі7 перев даній заявці. Такі мультимери можуть бути 49 71553 50 влаштовані у вигляді тандемних повторів, розгаються одиночними залишками проліна. У цьому лужених конструкцій або поєднувати їх. Базові значенні в таких варіантах, де базові пептиди пептиди можуть бути напряму приєднані один до включають на N- або С-кінці залишок проліна, таодного або розділені одним або декількома ких як наявність на місці Х1 в формулі (І) проліна лінкерами. (Р) або D-проліна (р), то m в формулі (II) переважБазовими пептидами, які утворюють мультино дорівнює 0. В тих варіантах, в яких на N- або Смери, можуть бути пептиди формули (І), аналоги кінці пептиду немає залишку проліна, лінкером формули (І), муто вані варіанти формули (І), укоропереважно є пролін (Р) або D-пролін (р), a m перечені або делетовані по внутрішніх положеннях важно дорівнює 1. варіанти формули (І), добудовані варіанти формуУ деяких варіантах даного винаходу бажаним ли (І) і (або) їх поєднання. Базові пептиди можуть може бути використаний лінкерів, що розщеплюбути сполучені «голова-до-хвоста» (тобто N-кінець ються, за допомогою яких можна відділяти один до С-кінця), «голова-до-голови» (тобто N-кінець до або декілька спіральних сегментів (НН) за конN-кінця), «хвіст-до-хвоста» (тобто С-кінець до Скретних умов. Відповідними лінкерами, що розщекінця) або шляхом поєднання цих орієнтацій. плюються є пептиди, які характеризуються В одному з варіантів даного винаходу мультиамінокислотними послідовностями, які мери організовані у вигляді тандемних повторів, розпізнаються протеазами, олігонуклеотиди, які що складаються з двох, трьох, чотирьох і аж до розщеплюються ендонуклеа-зами, і органічні сподесяти базових пептидів. Переважними є мультилуки, які можуть бути розщеплені хімічним шлямери, складені тандемними повторами від двох до хом, наприклад, в умовах кислої або лужної серевосьми базових пептидів. Отже, в одному з ди, або в інших умовах. Переважно, умови варіантів агоністи АроА-І по даному винаходу розщеплення повинні бути досить м'якими, щоб не включають мультимери, що характеризуються зумовлювати денатурацію або які-небудь інші понаступною структурною формулою: рушення спіральних сегментів і (або) непризначеНН[LLm-HH]nLLm-HH (ІІ) них до розщеплення лінкерів в складі мультимерде: них агоністів АроА-І. кожний показник m незалежно один від іншого Пептидні і олігонуклеотидні лінкери, які можуть є цілим числом від 0 до 1, переважно рівний 1; бути вибірним чином розщеплені, а також способи n є цілим числом від 0 до 10, переважно від 0 розщеплення лінкерів добре відомі і легко доступні до 8; фа хівцям в даній області техніки. Відповідні кожний «НН» незалежно представляє базовий лінкери у вигляді органічних сполук, які можуть пептид або пептидний аналог (І) або її мутовану, бути вибірним чином розщеплені, будуть очевидні укорочену, делетовану по внутрішньому положендля фа хівців в даній області техніки і включають ню або добудовану форму відповідно до описаноте, що, наприклад, було описано в міжнародній го в даній заявці; заявці WO 94/08051 і в матеріалах, що цитуються кожний «LL» незалежно представляє лінкер; і в ній. кожний знак «-» незалежно означає ковалентний У переважному варіанті лінкерами, що викоризв'язок. стовуються є пептиди, які служать субстратом для У формулі (II) лінкер LL може бути представендогенних циркулюючих (позаклітинних) ферлений будь-якою біфункціональною молекулою, ментів, що тим самим забезпечує мультимерним здатною забезпечувати ковалентний зв'язок одноагонистам АроА-І здатність виборчо розщеплюваго пептиду з іншим. Отже, відповідні лінкери - це тися in vivo. Ендогенним ферментом, придатним біфункціональні молекули, в яких функціональні для розщеплення таких лінкерів, є, наприклад, групи здатні утворювати ковалентний зв'язок з N-i пропептидаза проаполіпопротеїна А-І. Відповідні (або) С-кінцем пептида. Функціональні групи, приферменти, а також пептидні сегменти, що служать датні для приєднання до N- або С-кінцю пептидів, субстратом для таких ферментів, добре відомі в добре відомі в даній області техніки, рівно як і даній області техніки (див., наприклад, Edelstein et відомі відповідні хімічні прийоми для забезпечення al., 1983, J.Biol. Chem., 258, 11430-11433; Zanis, ефективного утворення таких зв'язків. 1983, Proc. Natl. Acad. Sci. USA, 80, 2574-2578). Лінкер може мати гнучку, жорстку або Як обговорювалося вище, ключовою вланапівжорстку структур у, що залежить від бажаних стивістю базових пептидів по даному винаходу є їх властивостей конкретного мультимера. Відповідні здатність утворювати міжмолекулярні водневі лінкери включають, наприклад, амінокислотні зазв'язки або сольові «містки» при вибудуванні в лишки, такі як пролін або гліцин, або пептидні сегантипаралельній орієнтації. Отже, в переважному менти, що включають від приблизно 2 до приблизваріанті даного винаходу лінкери достатньої довно 5, 10, 15 або 20, або навіть більшого числа жини і гнучкості використовуються таким чином, амінокислот, біфункціональні органічні сполуки, щоб забезпечувати спіральним сегментам (НН) такі як H2N(CH2)nCOOH, де n є цілим числом від 1 формули (II) можливість шикуватися в антипарадо 12, і подібне. Приклади таких лінкерів, рівно як і лельній орієнтації і в присутності ліпідів утворюваспособи отримання таких лінкерів і пептидів, що ти міжмолекулярні водневі зв'язки або сольові включають такі лінкери, добре відомі в даній об«містки». ласті техніки (див., наприклад, Hunig et al., 1974, Лінкери достатньої довжини і гнучкості вклюChem. Ber., 100, 3039-3044; Basak et al., 1994, Bioчають, тим самим не обмежуючись, пролін (Р), conjug. Chem., 5, 301-305). гліцин (G), Cys-Cys, H 2N-(CH2) n-COOH, де n рівне У переважному варіанті даного винаходу танвід 1 до 12, переважне від 4 до 6, H2N-арил-COOH демні повтори у внутрішніх своїх дільницях уриваі вуглеводи. 51 71553 52 З іншого боку, оскільки нативні зв'язок і присутній залишок проліна у аполіпопротеїни забезпечують скріплення між розвнутрішньому положенні. Для ознайомлення з огташованими антипаралельно спіральними сегменлядом, що описує різні типи і послідовності βтами, пептидні лінкери, які відповідають по своїй вигинів в білках і пептидах, читач може ознайомиамінокислотній послідовності пептидним сегментися з працею Wilmot et al., 1988, J.МоІ. Biol., 203, там, що примикають до спіралей нативних 221-232. аполіпопротеїнів, включаючи, наприклад, АроА-І, Конформації і послідовності багатьох пептидАроА-ІІ, Apo A-IV, АроС-І, АроС-ІІ, АроС-ІІІ, ApoD, них γ-вигинів також добре відомі в даній області ApoE і ApoJ, можуть бути стандартним чином витехніки (див., наприклад. Rose et al., 1985, Adv. користані для скріплення базових пептидів. Такі Protein Chem., 37, 1-109; Wilmer-White et al., 1987, послідовності добре відомі в даній області техніки Trends Biochem. Sci., 12, 189-192; Wilmot et al., (див., наприклад, Rosseneu et al., 1992, "Analysis of 1988, J.МоІ., Biol./ 203, 221-232; Si-banda et al./ the primary and of the secondary structure of the 1989, J.МоІ. Biol., 206, 759-777; Tramontano et al., apolipopro-teins", "In Structure & Function of Lipopro1989, Proteins: Struct., Funct. & Genet., 6, 382-394). teins", Ch.6, pp.159-183, CRC Press Inc.). Всі дані типи β-вигинів і γ-вигинів і відповідні ним Іншими лінкерами, що забезпечують утворенамінокислотні послідовності, рівно як і пізніше опиня міжмолекулярних водневих зв'язків або сольосані структури і послідовності пептидних β-вигинів і вих «містків» між тандемними повторами антипаγ-вигинів конкретним чином представляються даралельно орієнтованих спіральних сегментів, є ним винаходом. зворотні пептидні витки, такі як β-вигини і γ-вигини, З іншого боку, лінкер (LL) може включати оррівно як і органічні молекули, які імітують структуганічну молекулу або складову, яка імітує стр уктури пептидних β-вигинів і (або) γ-вигинів. Загалом, ру пептидного β-вигину або γ-вигину. Такі складові, зворотні витки - це сегменти пептидів, які розверщо імітують β-вигини і (або) γ-вигини, рівно як і тають напрям поліпептидного ланцюжка таким способи синтезу пептидів, що включають такі чином, щоб забезпечити одинарному складові, добре відомі в даній області техніки і поліпептидному ланцюжку можливість пристосувавключають, крім іншого, те, що описано в- працях ти такі свої дільниці, як антипаралельна βGiannis & Kolter, 1993, Angew. Chem. Intl. Ed. Eng., площина або α-спіраль, β-вигин загалом складе32, 1244-1267; Kahn et al., 1988, J.МоІ. Recognition, ний чотирма амінокислотними залишками, а γ1, 75-79; Kahn et al., 1987, Tetrahedron Lett., 28, вигин звичайно складений трьома 1623-1626. амінокислотними залишками. Ще в одному варіанті даного винаходу мульКонформації і послідовності багатьох пептидтимери мають форму розгалуженої структури них β-вигинів добре відомі в даній області техніки і (див., наприклад, Фіг.7). Такі структури стандартвключають, тим самим не обмежуючись, названі в ним чином отримують з використанням багатоякості прикладів тип-І, тип-І', тип-ІІ, тип-ІІ', тип-ІІІ, функціональних зв'язуючих складових, які забезтип-ІІІ', тип-IV, тип-V, тип-V', тип-VІa, тип-VIb, типпечують приєднання до простої зв'язуючої VII і тип-VIII (див. Richardson, 1981, Adv. Protein складової більш ніж двох спіральних одиниць. ОтChem, 34, 167-339; Rose et al., 1985, Adv. Protein же, розгалужені структури засновані на молекулах, Chem., 37, 1-109; Wilmot et al., 1988, J.МоІ. Biol., що мають три, чотири або навіть більше число 203, 221-232; Sibanda et al., 1989, J.МоІ. Biol., 206, функціональних груп, які здатні ковалентно 759-777; Tramontane et al., 1989, Proteins: Struct., приєднуватися до N-і (або) С-кінцю пептиду. Funct. & Genet, 6, 382-394). Відповідні зв'язуючі складові, наприклад, включаВигнутість конкретних конформацій коротких ють амінокислотні залишки, що характеризуються пептидів з утворенням β-вигину насамперед залебічними ланцюгами, що несуть гідроксильні, сульжить від розташування деяких амінокислотних фанільні, аміно, карбоксильні, амідні і (або) ефірні залишків у вигині (звичайно Gly, Asn або Pro). Звифункціональні групи, такі як, наприклад, Ser (S), чайно β-вигин типу-І сумісний з будь-яким Thr (T), Cys (С), Туr (Υ), Asn (N), Gin (Q), Lys (K), амінокислотним залишком в положенні від 1-го до Arg (R), Orn, Asp (D) і Glu (E) або інші органічні 4-го, за винятком того, що пролін не повинен займолекули, що включають такі функціональні групи. мати 3-е положення. У складі типу-І і типу-ІІ в поСпіральні сегменти, приєднані до однієї ложенні 4 переважає залишок гліцина, а в полозв'язуючої складової, не обов'язково повинні бути женні 2 переважає пролін. У положенні 1 часто приєднані однаковими кінцями. Дійсно, в деяких знаходяться залишки Asp, Asn, Ser і Cys, при тому, варіантах спіральні сегменти приєднані до однієї що їх бічні ланцюги часто утворюють водневі зв'язуючої складової таким чином, щоб забезпечизв'язки з NH в складі 3-го залишку. ти антипаралельну орієнтацію, і при цьому деякі У вигинах типу-ІІ залишки гліцина і аспарагіна спіралі приєднані по своїх N-кінцях, а інші - по найчастіше знаходяться в 3-му положенні, тому своїх С-кінцях. що вони найбільш легко забезпечують пристосуСпіральні сегменти можуть бути приєднані навання необхідних кутів у вуглецевому ланцюгу. В пряму до зв'язуючої складової або можуть ідеальній ситуації вигини типу-І' мають залишки відділені від зв'язуючої складової за допомогою гліцина в положеннях 2 і 3, а у вигині типу-ІІ' гліцин одного або декількох біфункціональних лінкерів знаходиться в положенні 2. У вигині типу-ІІІ можуть (LL) відповідно до описаного вище. знаходитися більшість амінокислотних залишків, в Звертаючись до Фіг.7А і 7В, можна бачити, що той час як в типі-ІІІ', як правило, у 2-му і 3-му порозгалужена структура може бути описана в ложеннях потрібна присутність гліцина. У вигинах термінах числа «вузлів гілкування», що складають типу-VІa і типа-VIb звичайно є циспептидний таку структур у, де вузол являє собою багато 53 71553 54 функціональну зв'язуючу складову. На Фіг.7А і 7В варіантом відповідно до описаного в даному спіральні сегменти (наприклад, базові пептиди по тексті; даному винаходу) зображені у вигляді циліндрів, а кожний LL незалежно представлений багатофункціональні зв'язуючі складові (або «вузбіфункціональним лінкером; ли») позначені кільцями (•), де число ліній, що викожний m незалежно є цілим числом від 0 ходять з кільця, означає «порядок» (суть число до 1; функціональних груп) для багато функціональної кожний n незалежно є цілим числом від 0 до 8; зв'язуючої складової. Nya і N yb незалежно один від іншого є багатоЧисло «вузлів гілкування» в даній структурі зафункціональною зв'язуючою складовою, де уа і уb галом залежить від загальної бажаної кількості представляють число функціональних гр уп, спіральних сегментів і звичайно складає від 1 до 2. відповідно, в Nya і N yb; Звісно, повинно бути зрозуміло, що для даного кожний уа або уb незалежно є цілим числом від числа бажаних спіральних сегментів структури, що 3 до 8; базуються на зв'язуючих складових більш високоp є цілим числом від 0 до 7; і го порядку, зможуть включати менше число кожний знак «-» незалежно означає ковалент«вузлів». Наприклад, звертаючись до Фіг.7А і 7В, ний зв'язок. структура 3-го порядку (тобто стр уктура, що вклюУ переважному варіанті розгалужена структучає 3-функціональні зв'язуючі складові) для семи ра включає «лізинове дерево», тобто є структуспіральних одиниць має три «вузли» (Фіг.7А), в той рою, в якій багатофункціональна зв'язуюча склачас як структура 4-го порядку (тобто структура, що дова є одиночним або серією залишків лізина (К) включає 4-функціональні зв'язуючі складові) для (див., наприклад, Фіг.7D). семи спіральних одиниць має тільки два «вузли» В одному ілюстративному варіанті «лізинове (Фіг.7В). дерево» в розгалуженій структурі по даному винаСтруктури можуть характеризуватися одноходу описується такою формулою: типними порядками: тобто в таких структура х всі «вузли» є, наприклад, 3-функціональними або 4функціональними зв'язуючими складовими, або ж можуть мати змішані по порядку «вузли гілкування», наприклад, при одночасній наявності 3-функціональньїх і 4-функціональних зв'язуючих складових. Звичайно, повинно бути зрозуміло, що навіть в структурах з гілкуваннями одного порядку зв'язуючі складові не повинні бути ідентичними. У структурі з гілкуваннями третього порядку можуть використовува тися, наприклад, дві, три, чотири або навіть більше різних трифункціональних де зв'язуючих складових. кожний X незалежно представлений НН-[LLmЯк і лінійні мультимери, спіральні сегменти, HH]n-LLm-HH; що входять до складу розгалужених структур, мокожний НН незалежно представлений базовим жуть, а можуть і не бути ідентичними. пептидом формули (І) або його аналогом, або муПриклад такої структури зі змішаними порядтованим, укороченим, делетованим по ками гілкування показаний на Фіг.1С. На Фіг.1С внутрішньому положенню або добудованим спіральні сегменти (тобто базові пептиди по дановаріантом відповідно до описаного в даному му винаходу) показані у вигляді циліндрів, а багатексті; тофункціональні зв'язуючі складові показані у викожний LL незалежно представлений гляді кільців (•), де число ліній, що ви ходять з біфункціональним лінкером, кільця, відповідає «порядку» (суть числу кожний n незалежно є цілим числом від 0 до 8; функціональних груп) багатофункціональної кожний m незалежно є цілим числом від 0 зв'язуючої складової. Лінії, зв'язуючі спіральні сегдо 1; менти, представляють біфункціональні лінкери LL R1 є -OR або -NRR; і у відповідності до раніше описаного. Спіральні кожний R незалежно представлений -Н, С1-6сегменти, які складають розгалужені структури, алкілом, С1-6-алкенілом, С1-6-алкинілом, С5-20можуть бути тандемними повторами базових пепарилом, С6-26-алкарилом, 5-20-атомним гетероатидів у відповідності до раніше описаного. рилом або 6-26-атомним алкгетероарилом. В одному з ілюстративних варіантів розгалу5.1.1. Аналіз структури і функцій жені структури по даному винаходу описуються Структура і функції базових пептидів або пепнаступною формулою: тидних аналогів по даному винаходу, рівно як і X-N ya-X( ya-1)- (N yb-X( yb-1) )ρ/ (III) агоністи АроА-І, складені такими базовими пептиде дами, включаючи мультимерні форми, описані кожний X незалежно представлений НН-[LLmвище, можуть бути протестовані з метою вибору HH]n-LLm-HH; активних агоністів або міметиків АроА-І. Наприккожний НН незалежно представлений базовим лад, базові пептиди або пептидні аналоги можуть пептидом формули (І) або його аналогом, або мубути протестовані по їх здатності утворювати αтованим, укороченим, делетованим по спіралі в присутності ліпідів, зв'язуватися з внутрішньому положенню або добудованим ліпідами, утворювати комплекси з ліпідами, акти 55 71553 56 вувати LCAT, сприяти виходу холестерину з клітин флуоресцентної спектроскопії, тест на основі якої і т.п. описаний тут в розділі 7. Здатність пептидів і (або) Методи і тести, призначені для аналізу струкпептидних аналогів активувати фермент LCAT тури і (або) функцій пептидів добре відомі в даній може бути легко визначена з використанням тесту області техніки. Переважні методи охарактеризона активацію LCAT, описаного тут в розділі 8. Тесвані в прикладах, описаних в даному тексті. Нати in vitro та in vivo, описані тут в розділах 9, 10 і приклад, тести на основі кругового ди хроїзма (КД) і 11, можуть бути використані для оцінки часу ядерного магнітного резонансу (ЯМР), описані тут півжиття, розподілу, інтенсивності виходу холестев розділі 7, можуть бути використані для аналізу рину і впливу на ОТХ. структури пептидів або пептидних аналогів Загалом, базові пептиди і (або) пептидні аназокрема, для визначення «ступеня спіральності» в логи по даному винаходу, які виявляють властиприсутності ліпідів. Здатність зв'язуватися з вості, підсумовуванні в табл.4, розглядаються як ліпідами може бути визначена з використанням активні. Таблиця 4 Властивості активних пептидів Діапазон «Частка спіральності» в присутності ліпідів (Ri=30) (незаблокований 22³60% амінокислотний пептид) «Частка спіральності» в присутності ліпідів (Ri=30) (незаблокований 18³40% амінокислотний пептид) «Частка спіральності» в присутності ліпідів (Ri=30) (заблокований 18³60% амінокислотний пептид і більш короткі пептиди) Зв'язування ліпідів (в присутності SUV) 0,5-10мкМ пептида (Ri=1-50) Активація ферменту LCAT >38% Ri - молярне відношення «ліпід/пептид». Як ілюструється в робочих прикладах даної заявки, базові пептиди, які виявляють високу міру активації LCAT (³38%), звичайно виявляють наявність істотної міри α-спіральності в присутності невеликих одношарових жирових пухирців (SUV) (³60% спіральності у випадку з незаблокованими пептидами, що складаються з 22 або більшого числа амінокислотних залишків і заблокованих пептидів, що складаються з 18 або меншого числа амінокислотних залишків; ³40% спіральності у випадку незаблокованих пептидів, що складаються з 18 або меншого числа амінокислот), в той час як пептиди, що виявляють низьку активність в активації LCAT або позбавлені її зовсім, характеризуються слабою α-спіральністю. Однак, в деяких випадках пептиди, що характеризуються високою інтенсивністю утворення спіралі в присутності ліпідів, не були ефективні в активації LCAT. Схожим чином, при тому, що пептиди, які забезпечують істотн у активацію LCAT, звичайно зв'язують ліпіди, в деяких варіантах пептиди, що проявляють скріплення ліпідів, не зумовлюють скільки-небудь істотної активації LCAT. Як наслідок, для фахівців в даній області техніки повинно бути зрозуміло, що, при тому, що здатність базових пептидів, описаних в даному тексті, утворювати а-спіралі (в присутності ліпідів) і зв'язувати ліпіди є критичною для їх активності, в багатьох варіантах ці властивості можуть не бути вираженими. Отже, в переважному варіанті базові пептиди по даному винаходу є об'єктами серії тестів, націлених на вибір базових пептидів, що виявляють виразну фармакологічну активність. Переважний діапазон ³80% ³60% ³80% ³80% На першому етапі базовий пептид тестують по його здатності утворювати α-спіраль в присутності ліпідів з використанням КД-тесту, описаного тут в розділі 7. Ті пептиди, які характеризуються принаймні 40%-ою «спіральністю» (для незаблокованих пептидів, що складаються з 18 або меншого числа амінокислот) або 60%-ою спіральністю (для заблокованих пептидів, що складаються з 18 або меншого числа амінокислот, а також для незаблокованих пептидів, що складаються з 22 або більшого числа амінокислот) в присутності ліпідів (в концентрації приблизно 5мкМ і молярному відношенні «ліпід/пептид» приблизно 30), потім тестують по їх здатності зв'язувати ліпіди з використанням тесту на флуоресценцію, описаного тут в розділі 7. Зрозуміло, що тільки ті пептиди, які включають флуоресцуючі залишки триптофана (W) або нафтилаланіна (Nal), тестують на скріплення ліпідів в тесті на флуоресценцію. Однак, у випадку пептидів, які не включають флуоресцуючих амінокислотних залишків, скріплення ліпідів оцінюється по збільшенню спіральності в присутності ліпідів. Базові пептиди, які виявляють скріплення ліпідів в присутності SUV (0,5-10мкМ пептиди; молярне відношення «ліпід/пептид» в діапазоні від 1 до 50), потім піддають скринінгу на фармакологічну активність. Зрозуміло, що фармакологічна активність, на яку проводиться скринінг, буде залежати від передбачуваного використання конкретних агоністів АроА-І. У переважному варіанті базові пептиди тестують по їх здатності активува ти фермент LCAT, тому що пептиди, здатні активувати LCAT, є конкретно застосовними в способах, що описуються даним текстом. Базові пептиди, які виявляють принаймні 38%-у активацію LCAT в 57 71553 58 порівнянні з нативним АроА-l людини (відповідно Х22 представлений лізином (К) або орнитином, до оцінки в тесті на активацію LCAT, описаним тут а Х3, Х5, Х6 , Х9, Х10 і Х14 відповідають попереднім в розділі 8), є переважними, при тому, що визначенням для формули (І). найбільш переважними є базові пептиди, що виЩе більш переважним варіантом відповідно до являють активацію на рівні 50%, 60%, 70%, 80% даного аспекту даного винаходу є ті пептиди, в або навіть 90%. яких: 5.1.2. Переважні варіанти Х2 представлений валіном (V); Агоністи Аро А-І по даному винаходу можуть Х3 представлений лейцином (L); бути далі визначені шляхом обумовлювання переX5 представлений лейцином (L); важних варіантів. Х6 представлений фенілаланіном (F); В одному з переважних варіантів агоністи Х7 представлений аргініном (R) або лізином АроА-І складаються з 22 амінокислотних залишків (К); відповідно до формули (І) або є їх ацильованими X8 представлений глутаміновою кислотою (Е); по Текінцю і (або) амідованими по С-кінцю, або Х9 представлений лейцином (L); етерифікованими формами. Х10 представлений лейцином (L); В іншому переважному варіанті агоністи АроАХ11 представлений аспарагіном (N) або глуІ є 22-амінокислотними пептидами відповідно до таміновою кислотою (Q); формули (І) або їх N-ацетильованими і (або) СХ12 представлений глутаміновою кислотою (Ε); амідованими, або етерифікованими формами, в і (або) яких Х7 є основною амінокислотою, аспарагіном Х15 представлений глутаміновою кислотою (Е); (N) або глутаміновою кислотою (Е); Х8 є кислою і Х1 , X4, Х13 , Х14, Х16 , Х17, Х18, Х19 , Х20, Х21 і X22 амінокислотою або аргініном (R); Х12 є кислою відповідають попереднім визначенням, даним для амінокислотою або аспарагіном (N); і (або) X15 є формули (І), або відповідають визначенням попекислою амінокислотою, глутаміном (Q) або реднього абзацу. лізином (К); і Х1 Х2, Х3, Х4 , X5, Х6, Х9, Х10 , Х11 , Х13, Ще в одному переважному варіанті агонистаХ14, Х16 , Х17, Х18 , Х19, Х20 і Х21 відповідають попеми АроА-І є 22-амінокислотні пептиди, відповідні реднім визначенням, даним для формули (І). формулі (І), або їх ацильовані по N-кінцю і (або) В іншому переважному варіанті агоністи АроАамідовані по С-кінцю, або етерифікоовані форми, в І є 22-амінокислотними пептидами, відповідними яких тільки один із залишків Х18 і Х19 є основною формулі (І), або їх N-ацильованими і (або) Самінокислотою, а інший мономер з Х18 і X19 є глуамідованими, або етерифікованими формами, в таміном (Q) або аспарагіном (N). яких: Ще в одному варіанті агоністи АроА-І є пептиХ1 представлений проліном (Р), гліцином (G), дами формули (І) або їх ацильованими по N-кінцю аланіном (А), глутаміном (Q), аспарагіном (N), асі (або) амідованими по С-кінцю, або етепарагіновою кислотою (D) або D-проліном (р); рифікованими формами, в яких один з X18 або Xiq Х2 представлений аланіном (А), валіном (V) представлений лізином (К) або орнитином, а інший або лейцином (L); мономер з Х18 і Х19 є глутаміном (Q) або аспаХ4 представлений аспарагіновою кислотою (D) рагіном (N). або глутаміновою кислотою (Е); Ще в одному переважному варіанті агонистаХ7 представлений лізином (К), аргініном (R), ми АроА-І є 22-амінокислотні пептиди, відповідні орнитином, аспарагіном (N) або глутаміновою киформулі (І), або їх ацильовані по N-кінцю і (або) слотою (Е); амідовані по С-кінцю, або етерифіковані форми, в Х11 представлений аспарагіном (N), глуяких один з Х9, Х10 , Х13, Х14, Х16 або Х17 представтаміном (Q), глутаміновою кислотою (Е) або лений гліцином (G), а інші амінокислоти з цього аргініном (R); переліку не є гліцином (G). Х12 представлений аспарагіновою кислотою Ще в одному переважному варіанті агониста(D), глутаміновою кислотою (Е) або аспарагіном ми АроА-І є 22-амінокислотні пептиди, відповідні (N); формулі (І), або їх ацильовані по N-кінцю і (або) Х13 представлений лейцином (L), гліцином (G) амідовані по С-кінцю, або етерифіковані форми, в або α-аміноізобутировою кислотою (Aib); яких Х13 є гліцином (G), а кожний з Х9, Х10, Х14 , Х16 і Х15 представлений аспарагіновою кислотою Х17 не є гліцином (G). (D), глутаміновою кислотою (Е), глутаміном (Q) Ще в одному переважному варіанті агонистаабо лізином (К); ми АроА-І є 22-амінокислотні пептиди, відповідні Х16 представлений аланіном (А), триптофаном формулі (І), або їх ацильовані по N-кінцю і (або) (W), гліцином (G), лейцином (L), фенілаланіном (F) амідовані по С-кінцю, або етерифіковані форми, в або нафтилаланіном (Nal); яких: Х17 представлений лейцином (L), гліцином (G) Х1 представлений проліном (Р), гліцином (G) або нафтилаланіном (Nal); або D-проліном (Р); Χ18 представлений лізином (К), орнитином, Х2 представлений валіном (V); Х3 представлеглутаміном (Q) або аспарагіном (Ν); ний лейцином (L); Х19 представлений лізином (К), орнитином, Х4 представлений аспарагіновою кислотою (D) глутаміном (Q) або аспарагіном (Ν); або глутаміновою кислотою (Е); Х20 представлений лізином (К) або орнитином; Х6 представлений фенілаланіном (Г); Х21 представлений лейцином (L); і (або) Х7 представлений аргініном (R); X8 представлений глутаміновою кислотою (Е); Х9 представлений лейцином (L); 59 71553 60 Х10 представлений лейцином (L) або триптоХ17 представлений лейцином (L) або нафтифаном (W); лаланіном (Nal); Х11 представлений аспарагіном (N); Х18 представлений лізином (К) або орнитином; Х12 представлений глутаміновою кислотою (Е); Х19 представлений глутаміном (Q); Х13 представлений гліцином (G); Х20 представлений лізином (К) або орнитином; Х14 представлений лейцином (L); Х21 представлений лейцином (L); і Х15 представлений глутаміновою кислотою (Е); Х22 представлений лізином (К) або орнитином. Х16 представлений аланіном (А) або триптоКонкретні переважні агоністи АроА-І відповідно фаном (W); до даного аспекту даного винаходу вибирають з групи, що включає наступне: пептид 3 пептид 13 пептид 19 пептид 137 пептид 138 пептид 139 пептид 140 пептид 141 пептид 142 PVLDLFRELLNEGLEALKQKLK GVLDLFRELLNEGLEALKQKLK PVLDLFRELLNEGLEALKQKLK PVLDLFRELLNEGLEAZKQKLK PVLDLFRELLNEGLEWLKQKLK PVLDLFRELWNEGLEALKQKLK PVLDLFRELLNEGLEALOQOLO PVLDFFRELLNEGLEALKQKLK PVLELFRELLNEGLEALKQKLK а також їх амідовані по С-кінцю або етерифіковані, або ацильовані по N- кінцю варіанти. Ще в одному переважному варіанті агонистами АроА-І є 22-амінокислотні пептиди, відповідні формулі (І), або їх ацильовані по N-кінцю і (або) амідовані по С-кінцю, або етерифіковані форми, в яких Х9 представлений гліцином (G), а кожний з Х10, Х13, Х14 , Х16 і Х17 гліцином (G) не є. Конкретним переважним агоністом АроА-І відповідно до даного аспекту даного винаходу є пептид 20: PVLDLFREGLNELLEALKQKLK (SEQ ID NO 20). Ще в одному переважному варіанті агоністами АроА-І є 22-амінокислотні пептиди, відповідні формулі (І), або їх ацильовані по N-кінцю і (або) амідовані по С-кінцю, або етерифіковані форми, в яких Х10 представлений гліцином (G), а кожний з Х9 , Х13, Х14, Х16 і Х17 гліцином (G) не є. Конкретним переважним агоністом АроА-І відповідно до даного аспекту даного винаходу є пептид 9: PVLDLFRELGNELLEALKQKLK (SEQ ID NO 9). Ще в одному переважному варіанті агоністами АроА-І є 22-амінокислотні пептиди, відповідні формулі (І), або їх ацильовані по N-кінцю і (або) амідовані по С-кінцю, або етерифіковані форми, в яких Х14 представлений гліцином (G), а кожний з Х9 , Х10, Х13, Х16 і Х17 гліцином (G) не є. Конкретним переважним агоністом АроА-І відповідно до даного аспекту даного винаходу є пептид 126: PVLDLFRELLNELGEALKQKLK (SEQ ID NO 126). Ще в одному переважному варіанті агоністами АроА-І є 22-амінокислотні пептиди, відповідні формулі (І), або їх ацильовані по N-кінцю і (або) амідовані по С-кінцю, або етерифіковані форми, в яких Х]6 представлений гліцином (G), а кожний з Х9 , Х10, Х13, Х14 і Х17 гліцином (G) не є. Конкретним переважним агоністом АроА-І відповідно до даного аспекту даного винаходу є пептид 22: PVLDLFRELLNELLEGLKQKLK (SEQ ID NO 22). Ще в одному переважному варіанті агоністами АроА-І є 22-амінокислотні пептиди, відповідні формулі (І), або їх ацильовані по N-кінцю і (або) амідовані по С-кінцю, або етерифіковані форми, в яких XJ 7 представлений гліцином (G), а кожний з Х9 , Х10, Х13, Х14 і Х16 гліцином (G) не є. Конкретним переважним агоністом АроА-І відповідно до даного (SEQ ID NO 3); (SEQ ID NO 13); (SEQ ID NO 19); (SEQ ID NO 137); (SEQ ID NO 138); (SEQ ID NO 139); (SEQ ID NO 140); (SEQ ID NO 141); (SEQ ID NO 142); аспекту даного винаходу є пептид 12: PVLDLFRELLNELLEAGKQKLK (SEQ ID NO 12). Варіанти, що включають внутрішні залишки гліцина, можуть бути легко синтезовані з високим виходом шляхом сегментної конденсації, тим самим забезпечуючи істотні переваги для крупномасштабного виробництва. Технологія «сегментної конденсації», тобто сполучення один з одним невеликих пептидних ланцюжків з утворенням більш великого ланцюжка, була застосована для отримання різноманітних біологічно активних пептидів, включаючи 44-амінокислотний міметик АроА-І (див., наприклад, Nakagawa et al., 1985, J.Amer. Chem. Soc, 107, 7087-7083; Nokihara et al., 1989, Peptides, 1988, 166-168; Kneib-Cordonnier et al., 1990, Intern. J.Peptide Protein Res., 35, 527538), і вважається найбільш економічним методом масового високопродуктивного синтезу базових пептидів по даному винаходу. Переваги синтезу за допомогою сегментної конденсації включають можливість поєднувати заздалегідь підготовлені сегменти в розчині і простоту очищення кінцевого продукту. Обмеженнями даного методу є низькі ефективність реакції поєднання і вихід на етапі конденсації, а також низька розчинність деяких поліпептидних послідовностей, що отримуються. Ефективність поєднання на етапі конденсації може бути в істотній мірі підвищена шляхом збільшення часу поєднання. Звичайно підвищення тривалості поєднання приводить до підвищення рацемізації кінцевого продукту (Sieber et al., 1970, Helv. Chem. Acta, 53/ 2135-2150). Однак, оскільки в молекулі гліцину центр обертання відсутній, то він і не зазнає рацемізації (пролін через наявність стеричних перешкод також при подовженому часі поєднання зазнає рацемізації в слабій мірі або не зазнає її зовсім). Отже, варіанти з наявністю у внутрішніх положеннях залишків гліцину можуть бути синтезовані в значній кількості з високим виходом за допомогою сегментної конденсації заздалегідь синтезованих сегментів з урахуванням переваги, що дається відсутністю рацемізації у гліцину. Отже, варіанти, що мають всередині залишки гліцину, забезпечують значні переваги з
ДивитисяДодаткова інформація
Назва патенту англійськоюAgonists of apolipoprotein a-i and their use for treatment of disorders associated with dyslipidemia
Назва патенту російськоюАгонисты аполипопротеина a-i и их применение для лечения заболеваний, связанных с дислипидемией
МПК / Мітки
МПК: C07K 7/08, C12N 15/09, C12N 5/10, A61K 9/19, C12P 21/02, A61K 47/48, A61P 9/02, A61P 9/10, C07K 14/775, A61K 38/00, C07K 14/725, A61P 3/06
Мітки: дисліпідемічних, а-і, агоніста, apoa-i, мультимірний, аполіпопротеїну, агоніст, комплекс, ліпіду, лікування, варіанти, порушень, спосіб
Код посилання
<a href="https://ua.patents.su/90-71553-agonist-apolipoprotenu-a-i-apoa-i-multimirnijj-apoa-i-varianti-kompleks-agonista-apoa-i-i-lipidu-ta-sposib-likuvannya-dislipidemichnikh-porushen.html" target="_blank" rel="follow" title="База патентів України">Агоніст аполіпопротеїну а-і (apoa-i), мультимірний apoa-i (варіанти), комплекс агоніста apoa-i і ліпіду та спосіб лікування дисліпідемічних порушень</a>
Азадициклічні ліганди рецептора 5ht1 та проміжні сполуки для їх отримання, фармацевтична композиція (варіанти) та спосіб лікування різних порушень або станів (варіанти)

Номер патенту: 65608
Опубліковано: 15.04.2004
Автор: Брайт Джін Майкл
МПК: A61P 25/22, C07D 471/04, A61P 7/00, A61P 25/04, A61P 15/00, A61P 35/00, A61P 25/00, A61P 1/00, A61P 25/24, A61K 31/4985, A61P 5/00, A61P 25/16, A61P 43/00, A61P 9/12, A61P 25/06, A61P 3/04, A61P 25/28, A61P 25/30, A61P 25/18, A61P 13/00, A61P 15/10
Мітки: ліганди, варіанти, фармацевтична, композиція, сполуки, спосіб, проміжні, різних, порушень, лікування, азадициклічні, станів, отримання, рецептора
Формула / Реферат:
Азадициклічні ліганди рецептора 5НТ1 формули (І) , (I)деR3, R4 та Z незалежно вибрано з гідрогену, галогену (наприклад, хлору, флуору, брому чи йоду), (С1-С4)алкілу, що необов'язково заміщено 1-3 атомами флуору, (С1-С4)алкоксилу, що необов'язково заміщено 1-3 атомами флуору, та (С1-С4)алкоксил(С1-С4)алкілу, що необов'язково заміщено 1-3 атомами флуору,W - -СН2-O-(С1-С6)алкіл, де алкіл може мати лінійний чи...
Комплекс елетриптану та похідної циклодекстрину, фармацевтична композиція, способи їх одержання та спосіб лікування мігрені (варіанти)

Номер патенту: 66932
Опубліковано: 15.06.2004
Автор: Біллотт Анн
МПК: A61P 25/06, A61K 31/715, A61K 31/404, A61K 47/48
Мітки: елетриптану, композиція, одержання, варіанти, комплекс, мігрені, циклодекстрину, спосіб, лікування, фармацевтична, способи, похідної
Формула / Реферат:
1. Комплекс елетриптану та похідної циклодекстрину формули (І):(І),де R1a-q, R2a-q та R3a-q - кожний, незалежно, являє собою -ОН або -O(СH2)4SО3Н;принаймні один з R1a-q являє собою -O(СH2)4SО3Н:або його фармацевтично прийнятної солі.2. Комплекс за п. 1, в якому середнє число груп -O(СH2)4SО3Н на молекулу похідної формули (І) становить 6,1-6,9.3. Комплекс за п. 1 або 2, в якому кожна група...
Піридазинохіноліни, спосіб їх одержання (варіанти), проміжні сполуки, фармацевтична композиція (варіанти) для лікування або профілактики нейродегенеративних порушень та способи лікування

Номер патенту: 60291
Опубліковано: 15.10.2003
Автори: Бер Томас Майкл, Емпфілд Джеймс Рой, Спаркс Річард Брюс, МакКінні Джеффрі Алан, Девенпорт Тімоті Вейн
МПК: C07D 471/14, A61P 25/00, A61P 43/00, A61P 3/08, A61P 25/28, A61P 25/08, A61P 3/10, C07D 495/14, A61K 31/50, C07D 215/56, C07D 471/04
Мітки: варіанти, профілактики, спосіб, проміжні, порушень, нейродегенеративних, композиція, сполуки, одержання, піридазинохіноліни, способи, фармацевтична, лікування
Формула / Реферат:
1. Пиридазинохинолины формулы II или их фармацевтически приемлемые соли и таутомеры:,где:Z выбран из О, S или NH, или, когда кольцо В N-таутомеризовано, группа Z может быть выбрана из Н, ОН, SH или NH2,кольцо А выбрано из ортоконденсированного ароматического или гетероароматического пяти- или шестичленного кольца, выбранного из фенила,...
Фармацевтична композиція, що містить твердий іонний комплекс аналога lhrh (варіанти), спосіб лікування та шприц

Номер патенту: 68343
Опубліковано: 16.08.2004
Автори: Гефтер Малколм Л., Молінекс Крістофер Дж., Баркер Ніколас, Муссо Гарі
МПК: A61P 15/00, A61K 38/09, A61K 47/48
Мітки: спосіб, містить, варіанти, лікування, аналога, іонний, фармацевтична, композиція, шприц, комплекс, твердий
Формула / Реферат:
1. Фармацевтична композиція, що включає активну сполуку і носій, яка відрізняється тим, що містить твердий іонний комплекс аналога LHRH і аніонного полімеру, причому вміст пептиду у вищезгаданому комплексі складає принаймні 57 % мас.2. Фармацевтична композиція за п.1, яка відрізняється тим, що вміст пептиду у вищезгаданому комплексі складає від 57 % до 79 % мас.3. Фармацевтична композиція за п.1, яка відрізняється тим, що в...
Фармацевтична композиція, що містить 5-гідроксііндол та модулятор агоніста нікотинового рецептора

Номер патенту: 65617
Опубліковано: 15.04.2004
Автори: Герлей Девід, Ленторн Томас
МПК: A61K 31/404, A61P 25/28
Мітки: агоніста, нікотинового, композиція, фармацевтична, модулятор, рецептора, містить, 5-гідроксііндол
Формула / Реферат:
1. Фармацевтична композиція, яка відрізняється тим, що включає позитивний модулятор агоніста нікотинового рецептора разом з фармацевтично прийнятним носієм, при цьому позитивний модулятор має здатність збільшувати ефективність вказаного агоніста нікотинового рецептора, де вказаний позитивний модулятор є 5-гідроксііндолом.2. Фармацевтична композиція за п. 1, яка відрізняється тим, що вона додатково включає агоніст нікотинового...