Молекули нуклеїнової кислоти, які кодують wrinkled1-подібні поліпептиди
Номер патенту: 89960
Опубліковано: 25.03.2010
Автори: Шенк Карін Дж., Міттендорф Фолькер, Хертель Хайко, Бхатт Гаріма























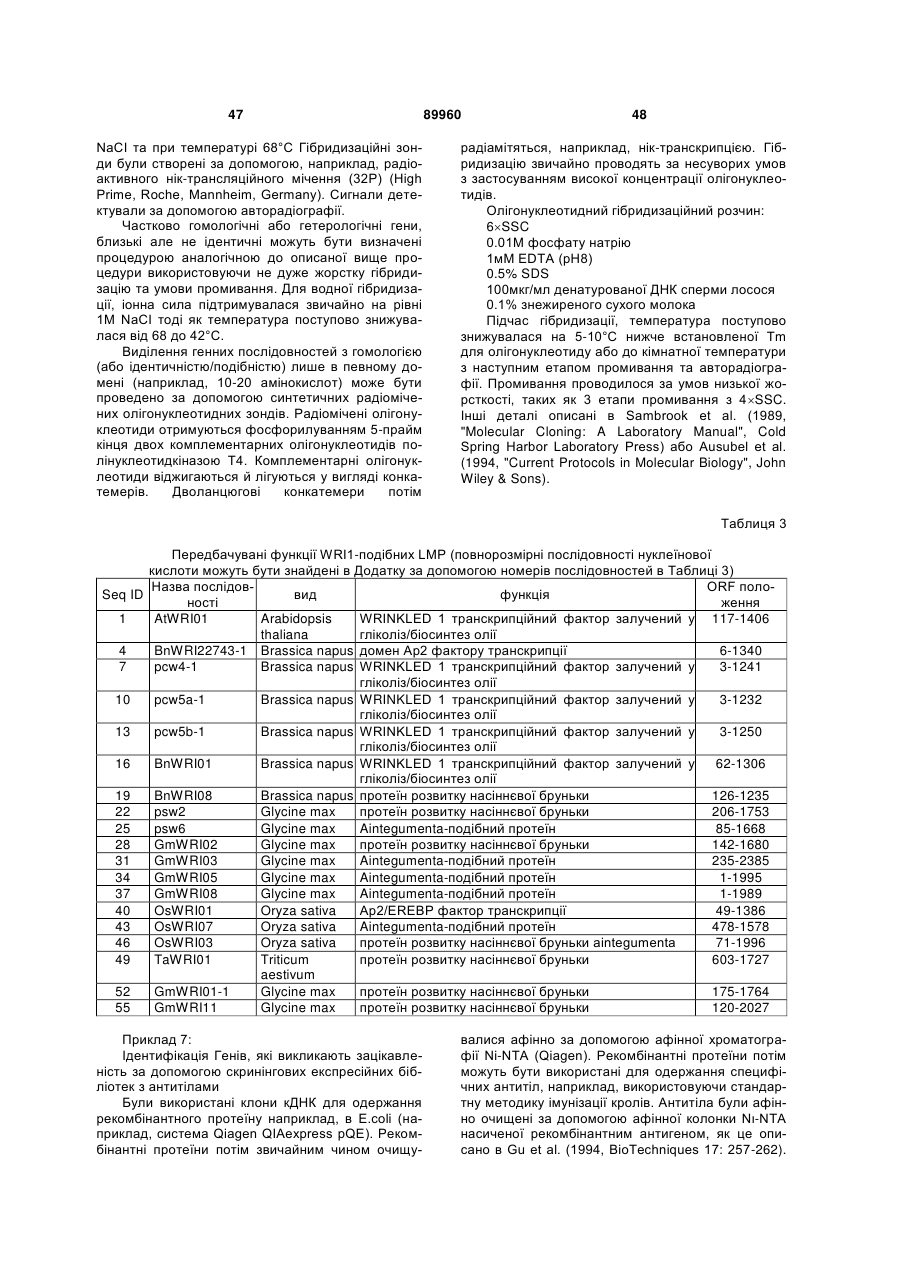



































































































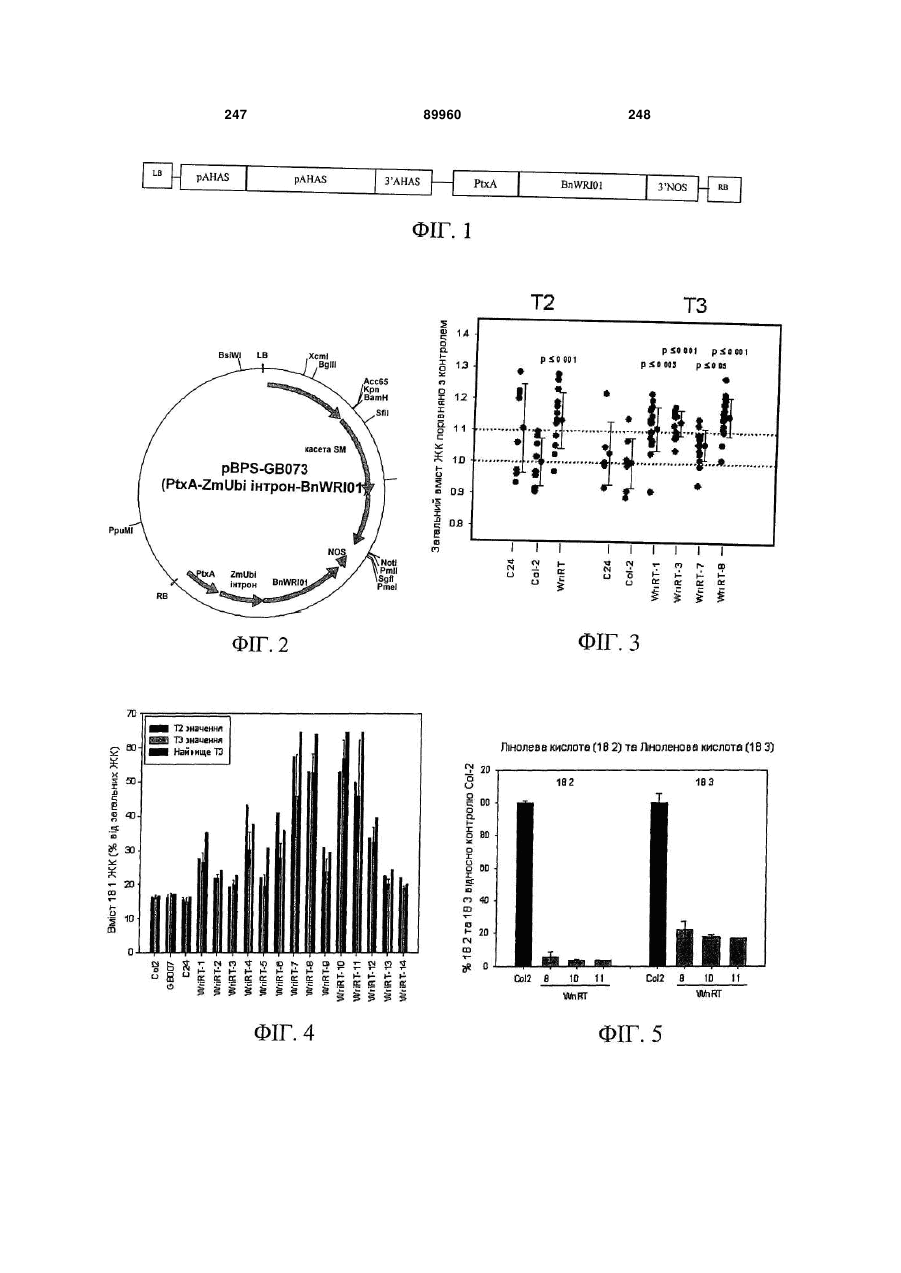
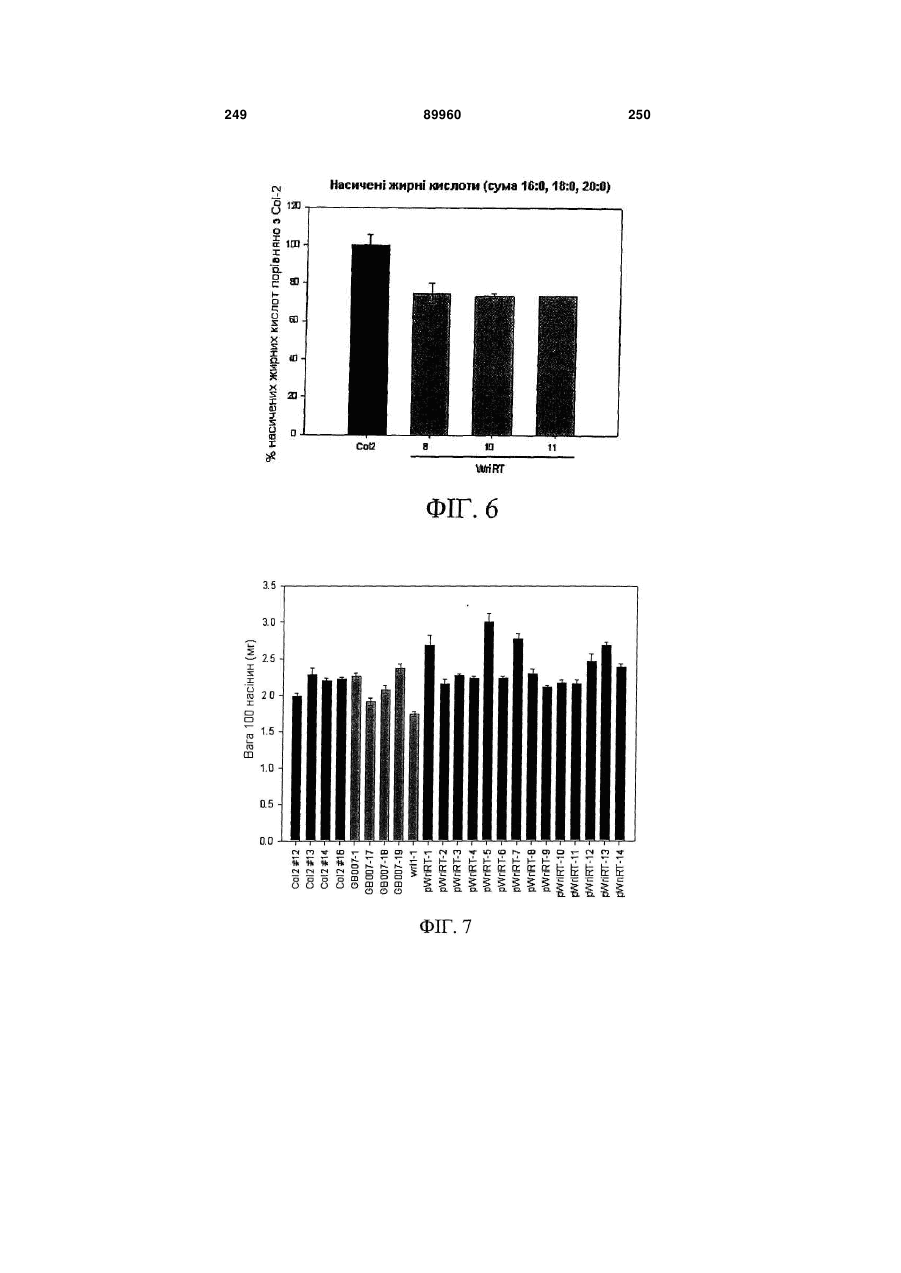

Формула / Реферат
1. Виділена нуклеїнова кислота, що кодує протеїн ліпідного метаболізму (LMP), яка містить полінуклеотидну послідовність, вибрану з групи, яка включає:
a) полінуклеотидну послідовність, як визначено в SEQ ID NО:4, SEQ ID NО:5, SEQ ID NО:7, SEQ ID NО:8, SEQ ID NО:10, SEQ ID NО:11, SEQ ID NО:13, SEQ ID NО:14, SEQ ID NО:16, SEQ ID NО:17, SEQ ID NО:19, SEQ ID NО:20, SEQ ID NО:22, SEQ ID NО:23, SEQ ID NО:25, SEQ ID NО:26, SEQ ID NО:28, SEQ ID NО:29, SEQ ID NО:31, SEQ ID NО:32, SEQ ID NО:34, SEQ ID NО:35, SEQ ID NО:37, SEQ ID NО:38, SEQ ID NО:40, SEQ ID NО:41, SEQ ID NО:43, SEQ ID NО:44, SEQ ID NО:46, SEQ ID NО:47, SEQ ID NО:49, SEQ ID NО:50, SEQ ID NО:52, SEQ ID NО:53, SEQ ID NО:55 або SEQ ID NО:56;
b) полінуклеотидну послідовність, яка кодує поліпептид, як визначено в SEQ ID NО:6, SEQ ID NО:9, SEQ ID NО:12, SEQ ID NО:15, SEQ ID NО:18, SEQ ID NО:21, SEQ ID NО:24, SEQ ID NО:27, SEQ ID NО:30, SEQ ID NО:33, SEQ ID NО:36, SEQ ID NО:39, SEQ ID NО:42, SEQ ID NО:45, SEQ ID NО:48, SEQ ID NО:51, SEQ ID NО:54 або SEQ ID NО:57;
c) полінуклеотидну послідовність, яка має принаймні 70 % ідентичності до повнорозмірної послідовності нуклеїнової кислоти LMP, наведеної вище в а) або b);
d) полінуклеотидну послідовність, яка є комплементарною до повнорозмірної послідовності нуклеїнової кислоти LMP, наведеної вище в а) або b); та
e) полінуклеотидну послідовність, яка гібридизується, за жорстких умов, до повнорозмірної послідовності нуклеїнової кислоти LMP, наведеної вище в а) або b).
2. Виділена нуклеїнова кислота LMP за пунктом формули 1, де полінуклеотидна послідовність кодує поліпептидну послідовність, як визначено в SEQ ID NО:6, SEQ ID NО:9, SEQ ID NО:12, SEQ ID NО:15, SEQ ID NО:18, SEQ ID NО:21, SEQ ID NО:24, SEQ ID NО:27, SEQ ID NО:30, SEQ ID NO:33, SEQ ID NО:36, SEQ ID NO:39, SEQ ID NО:42, SEQ ID NО:45, SEQ ID NО:48, SEQ ID NО:51, SEQ ID NО:54 або SEQ ID NО:57.
3. Виділена нуклеїнова кислота LMP за пунктом формули 1, де полінуклеотидна послідовність має принаймні 90 % ідентичності до повнорозмірної послідовності нуклеїнової кислоти LMP, наведеної в а) або b) пункту формули 1, та де виділена нуклеїнова кислота LMP кодує поліпептид, який функціонує як модулятор накопичувальної речовини насіння рослин.
4. Виділена нуклеїнова кислота LMP за пунктом формули 1, розташована в векторі експресії.
5. Виділена нуклеїнова кислота LMP за пунктом формули 4, функціонально зв'язана із гетерологічним промотором, вибраним з групи, яка включає насіннєво-специфічний промотор, коренево-специфічний промотор та тканинно-неспецифічний промотор.
6. Виділена нуклеїнова кислота LMP за пунктом формули 4, функціонально зв'язана з промотором ptxA.
Текст
1. Виділена нуклеїнова кислота, що кодує протеїн ліпідного метаболізму (LMP), яка містить полінуклеотидну послідовність, вибрану з групи, яка включає: a) полінуклеотидну послідовність, як визначено в SEQ ID NО:4, SEQ ID NО:5, SEQ ID NО:7, SEQ ID NО:8, SEQ ID NО:10, SEQ ID NО:11, SEQ ID NО:13, SEQ ID NО:14, SEQ ID NО:16, SEQ ID NО:17, SEQ ID NО:19, SEQ ID NО:20, SEQ ID NО:22, SEQ ID NО:23, SEQ ID NО:25, SEQ ID NО:26, SEQ ID NО:28, SEQ ID NО:29, SEQ ID NО:31, SEQ ID NО:32, SEQ ID NО:34, SEQ ID NО:35, SEQ ID NО:37, SEQ ID NО:38, SEQ ID NО:40, SEQ ID NО:41, SEQ ID NО:43, SEQ ID NО:44, SEQ ID NО:46, SEQ ID NО:47, SEQ ID NО:49, SEQ ID NО:50, SEQ ID NО:52, SEQ ID NО:53, SEQ ID NО:55 або SEQ ID NО:56; b) полінуклеотидну послідовність, яка кодує поліпептид, як визначено в SEQ ID NО:6, SEQ ID NО:9, SEQ ID NО:12, SEQ ID NО:15, SEQ ID NО:18, SEQ ID NО:21, SEQ ID NО:24, SEQ ID NО:27, SEQ ID NО:30, SEQ ID NО:33, SEQ ID NО:36, SEQ ID NО:39, SEQ ID NО:42, SEQ ID NО:45, SEQ ID C2 2 (19) 1 3 ники. Цей винахід, в загалі, стосується послідовностей нуклеїнових кислот, які кодують протеїни, які відносяться до накопичувальних речовин насіння в рослинах. Точніше, даний винахід стосується WRINKLED 1-подібних (WRI1-подібних) послідовностей нуклеїнових кислот, які кодують протеїни, регулятори метаболізму цукрів та ліпідів й використання таких послідовностей в трансгенних рослинах. Зокрема, винахід спрямовано на способи маніпулювання цукро-подібними речовинами, для підвищення рівнів олії, та для змінювання складу жирних кислот в рослинах та насінні. Також винахід стосується методів застосування цих нових рослинних поліпептидів для стимулювання росту рослин та/або для підвищення врожаю та/або складу запасних речовин насіння. Дослідження генетичних маніпулювань має довгу історію, яка починається ще до видатних досліджень Грегора Менделя (Gregor Mendel). Розвиваючи цю науку, вчені вдосконалили модифікування окремих ознак в ряді рослин від картоплі, бульби якої мають підвищений вміст крохмалю, до олійних рослин, таких як канола та соняшник із підвищеним або зміненим вмістом жирних кислот. Зі збільшенням споживання та використання рослинних олій, модифікування вмісту олій в насінні та рівнів олій в насінні стає широко розповсюдженим (наприклад, Topfer et al., 1995, Science 268: 681-686). Маніпулювання біосинтечними шляхами в трансгенних рослинах створює велику кількість можливостей для молекулярного біолога та біохіміка рослин впливати на рослинний метаболізм, що дає збільшення продукування специфічних цінних продуктів. Продукування насіннєвої олії або її склад, було змінено в багатьох традиційних олійних рослинах, таких як соя (U.S. Patent No.5,955,650), канола (U.S. Patent No.5,955,650), соняшник (U.S. Patent No.6,084,164), та рапс (Topfer et al., 1995, Science 268: 681-686), та нетрадиційних олійних рослинах, таких як тютюн (Cahoon et al., 1992, Proc. Natl. Acad. Sci. USA 89: 11184-11188). Рослинні олії містять як нейтральні, так і полярні ліпіди (Дивись Таблицю 1). Нейтральні ліпіди головним чином містять триацилгліцерол, який є основним засобом накопичення ліпіду, який накопичується в олійних тільцях в насінні. Полярні ліпіди знаходяться головним чином в різноманітних мембранах насіннєвих клітин, наприклад, ендоплазматичному ретикулумі, мікросомальних мембранах та клітинних мембранах. Нейтральні й полярні ліпіди мають декілька спільних жирних кислот (Дивись Таблицю 2) та велику кількість менш розповсюджених жирних кислот. Ліпіди позначені зірочкою в Таблиці 2 зазвичай не зустрічаються в рослинних оліях, але їхнє вироблення в оліях трансгенних організмів є важливим завданням біотехнології рослин. Жирнокислотний склад мембранних ліпідів є чітко визначеним, та лише певна кількість жирних кислот знаходиться в мембранних ліпідах. З іншого боку, велика кількість незвичайних жирних кислот може бути включена в нейтральні накопичувальні ліпіди в насіння багатьох видів рослин (Van de Loo et al., 1993, Unusual Fatty Acids in Lipid Metabolism in Plants pp.91-126, 89960 4 editor TS Moore Jr. CRC Press; Millar et al., 2000, Trends Plant Sci. 5: 95-101). Таблиця 1 Класи Ліпідів Рослин Нейтральні Триацилгліцерол (TAG) ліпіди Диацилгліцерол (DAG) Моноацилгліцерол (MAG) Полярні Моногалактозилацилгліцерол ліпіди (MGDG) Дигалактозилацилгліцерол (DGDG) Фосфатидилгліцерол (PG) Фосфатидилхолін (PC) Фосфатидилетаноламін (РЕ) Фосфатидилінозитол (РІ) Фосфатидилсерин (PS) Сульфохіновозилдіацилгліцерол Таблиця 2 Розповсюджені Рослинні Жирні кислоти 16:0 16:1 16:3 18:0 18:1 18:2 18:3 -18:3 20:0 20:1 22:6 20:2 20:4 20:5 22:1 Пальмітинова кислота Пальмітолеїнова кислота Пальмітоленова кислота Стеаринова кислота Олеїнова кислота Лінолеїнова кислота Ліноленова кислота Гамма-Ліноленова кислота* Арахінова кислота Ейкозова кислота Докозагексаенова кислота (DHA)* Ейкозодіенова кислота Арахідонова кислота (АА)* Ейкозапентаенова кислота (ЕРА)* Ерукова кислота Ліпіди синтезуються з жирних кислот, та їх синтез може бути поділений на дві частини: прокаріотичний шлях та еукаріотичний шлях (Browse et al., 1986, Biochemical J. 235: 25-31; Ohlrogge & Browse, 1995, Plant Cell 7: 957-970). Прокаріотичний шлях проходить в пластидах, які є первинним місцем біосинтезу жирних кислот. Синтез жирних кислот починається з перетворення ацетил-СоА у малонил-СоА за допомогою ацетил-СоА карбоксилази (АССази). Малонил-СоА перетворюється на білок-переносник малонил-ацил (АСР) за допомогою малонил-СоА:АСР трансацилази. Фермент бета-кето-ацил-АСР-синтетаза III (KAS III) каталізує реакцію конденсації, в якій ацильна група з ацетил-СоА переноситься на малонил-АСР для утворення 3-кетобутирил-АСР. В наступних послідовних реакціях конденсації, відновлення, та дегідратації, ланцюг створюваної жирної кислоти на кофакторі АСР подовжується поступовим додаванням (конденсацією) двох атомів вуглецю, які віддає малонил-АСР, до утворення 16- або 18вуглецевих насичених жирнокислотних ланцюгів. Пластидна дельта-9 ацил-АСР-десатураза вво 5 дить перший ненасичений подвійний зв'язок в жирну кислоту. Тіоестераза відщеплює жирну кислоту від кофактору АСР, й вільні жирні кислоти експортуються в цитоплазму, де вони беруть участь еукаріотичному шляху синтезу в складі ацил-СоА естерів. В цьому шляху біосинтезу, жирні кислоти естерифікуються гліцерол-3фосфатацилтрансферазою та лізофосфатидильною кислою ацилтрансферазою в положення sn-1 та sn-2 гліцерол-3-фосфату, відповідно, для отримання фосфатидної кислоти (РА). ФК (РА) є попередником інших полярних та неполярних нейтральних ліпідів, останні утворюються в циклі Kennedy (Voelker, 1996, Genetic Engineering ed.:Setlow 18: 111-113; Shanklin & Cahoon, 1998, Annu.Rev.Plant Physiol.Plant Mol.Biol. 49: 611-641; Frentzen, 1998, Lipids 100: 161-166; Millar et al., 2000, Trends Plant Sci. 5: 95-101). Запасні ліпіди в насінні синтезуються з попередників, які мають вуглеводневе походження. У рослин увесь гліколітичний цикл знаходиться в цитозолі (Plaxton, 1996, Annu.Rev.Plant Physiol.Plant Mol.Biol. 47: 185-214), й було показано, що у рапсу весь цикл також знаходиться в пластидах (Kang & Rawsthome, 1994, Plant J. 6: 795805). Сахароза є головним джерелом вуглецю та енергії, які транспортуються від листя до насіння, що розвивається. Підчас фази зберігання, сахароза перетворюється в цитозолі для забезпечення метаболічними попередниками, глюкоза-6фосфатом та піруватом. Вони транспортуються в пластиди й перетворюються на ацетил-СоА який слугує основним попередником для синтезу жирних кислот. Ацетил-СоА в пластидах є центральним попередником для біосинтезу ліпідів. АцетилСоА може утворюватися в пластидах у різних реакціях й остаточна роль кожної реакції все ще дискутується (Ohlrogge & Browse, 1995, Plant Cell 7: 957-970). Однак, загально прийнято, що велика частина Ацетил-СоА одержується із глюкоза-6фосфату й пірувату, які постачаються з цитоплазми в пластиди. Сахароза виробляється в продукуючих органах (листі, або будь-де де відбувається фотосинтез) й транспортується в насіння, яке також називають поглинаючими органами. В насінні, що розвивається, сахароза є попередником усіх накопичувальних речовин, таких як, крохмаль, ліпіди та частково насіннєві накопичувальні протеїни. Таким чином, стає зрозумілим метаболізм вуглеводів, в якому сахароза відіграє центральну роль, є дуже важливим для акумулювання накопичувальних речовин насіння. Накопичувальні речовини, такі як триацилгліцероли (насіннєві олії), слугують резервом вуглецю й енергії, які використовуються при дозріванні та рості молодих проростків. Насіннєва (рослинна) олія є важливою складовою харчування людини та важливим джерелом сировини для хімічної промисловості. Мутантний Arabidopsis у якого уражено метаболізм насіннєвих накопичувальних речовини має назву wrinkled1 (wri1) (Focks and Benning, 1998). Мутант характеризується 80%-им зниженням вмісту олії в насінні. Також, схоже було на те, що зазнала впливу експресія генів пов'язаних з метаболізмом цукру. 89960 6 Хоча вміст та склад ліпідів та/або жирних кислот в насіннєвій олії може бути змінений традиційними методами селекції рослин, поява технології рекомбінантної ДНК дозволяє легше маніпулювати вмістом насіннєвих олій рослин, та в деяких випадках, дозволяє змінювати насіннєві олії таким чином, як це було б неможливо виконати традиційними методами селекції (Дивись, наприклад, Topfer et al., 1995, Science 268: 681-686). Наприклад, введення послідовності нуклеїнової кислоти а12-гідролази в трансгенний тютюн, призвело до введення нової жирної кислоти, рицинолеїнової кислоти, в насіннєву олію тютюну (Van de Loo et al., 1995, Proc. Natl. Acad. Sci USA 92: 6743-6747). Рослини тютюну, також створювалися для вироблення петроселінової кислоти шляхом уведення та експресування ацил-АСР-десатурази коріандру (Cahoon et al., 1992, Proc. Natl. Acad. Sci USA 89: 11184-11188). Модифікування вмісту насіннєвої олії в рослинах має великі наслідки для медицини, харчування та економіки. Стосовно медичного застосування, довголанцюгові жирні кислоти (С18 та довші) знайдені в багатьох насіннєвих оліях були пов'язані зі зниженням гіперхолестеринемії та іншими клінічними розладами пов'язаними з коронарною хворобою серця (Brenner, 1976, Adv. Exp. Med. Biol. 83: 85-101). Отже, вживання рослин, які мають підвищений рівень жирних кислот цього типу може знизити ризик серцевих захворювань. Збільшення рівня вмісту насіннєвої олії також збільшує вироблення насіннєвих олій, й таким чином знижує вартість цих олій. Для того, щоб збільшити або змінити рівень таких речовин як, наприклад, насіннєвих олій, в рослинах, потрібно ідентифікувати послідовності нуклеїнових кислот і білків, які регулюють ліпідний і жирнокислотний метаболізм. Оскільки раніше згадані, декілька десатураз нуклеїнових кислот 6 таких як, наприклад, нуклеїнова кислота □ 12 десатурази, нуклеїнова кислота □ -десатурази, і нуклеїнові кислоти ацил-АСР-десатурази були клоновані й показано, що вони здатні кодувати ферменти, потрібні для жирно-кислотного синтезу в різних видах рослин. Послідовності нуклеїнової кислоти олеозину із таких різних видів, як канола, соя, морква, сосна, і Arabidopsis thaliana також були клоновані й визначено, що вони кодують білки, пов'язані з мембранним фосфоліпідним моношаром олійних тіл в цих рослинах. Також було визначено, що два фітогормони, гіберелінова кислота (GA) і абсцизова кислота (ABA), залучаються до загальних процесів у розвитку насіння (e.g. Ritchie & Gilroy, 1998, Plant Physiol. 116: 765-776; Arenas-Huertero et al., 2000, Genes Dev. 14: 2085-2096). Обидва шляхи GA та ABA знаходяться під впливом окадайової кислоти, інгібітора протеїнової фосфатази (Kuo et al., 1996, Plant Cell. 8: 259-269). Регулювання фосфорилування білка кіназами і фосфатазами визнано універсальним механізмом клітинного контролю (Cohen, 1992, Trends Biochem. Sci. 17: 408-413). Також, рослинний гормон етилен (Дивись, наприклад, Zhou et al., 1998, Proc. Natl. Acad. Sci. USA 95: 10294-10299; Beaudoin et al., 2000, Plant Cell 7 2000: 1103-1115) та ауксин (e.g. Colon-Carmona et al., 2000, Plant Physiol. 124: 1728-1738) також залучені до контролю розвитку рослин. Хоча відомо, що декілька сполук, загалом впливають на рослину і розвиток насіння, є нагальна необхідність специфічно ідентифікувати фактори, які ε більш специфічними для регулювання розвитком і накопиченням запасних речовин та ідентифікувати гени, які мають здатність змінювати або збільшувати вихід олій до його рослини походження або інших видів рослин. Винаходом розкриваються послідовності нуклеїнової кислоти Arabidopsis thaliana, Brassica napus, Glycine max, Oryza sativa, або Triticum aestivum. Ці послідовності нуклеїнової кислоти можуть використовуватися, для змінювання або збільшення рівня накопичувальної речовини насіння такої як наприклад білки, цукор, й олії в рослинах, зокрема трансгенних рослинах, як наприклад канола, льон, соя, соняшник, кукурудза, овес, жито, ячмінь, пшениця, рис, перець, нагідки, бавовна, олійна пальма, кокосова пальма, сільниця і арахіс, які є олійними культурами з високим вмістом ліпідних сполук. Цим винаходом пропонуються нові виділені послідовності нуклеїнових кислот та амінокислот, які пов'язані з метаболізмом накопичувальних речовин насіння в рослинах, зокрема з послідовностями, які є WRI1 -подібними. Цим винаходом пропонуються також виділені послідовності нуклеїнових кислот з Arabidopsis thaliana, Brassica napus, Glycine max, Oryza sativa, та Triticum aestivum які кодують протеїн ліпідного метаболізму (LMP), або його частину. Ці послідовності, можуть використовуватися, для змінювання або збільшення ліпідів і жирних кислот, кофакторів і ферментів в мікроорганізмах і рослинах. Відомо, що рослини Arabidopsis виробляють значну кількість жирних кислот, таких як лінолева та ліноленова кислоти (Див., наприклад, Таблицю 2) і мають значну подібність за багатьма показниками (гомологія генів та ін.) до такої олійної культури як Brassica. Отже, молекули нуклеїнової кислоти, які походять з такої рослини як Arabidopsis thaliana, Brassica napus, Glycine max, Oryza sativa, або Triticum aestivum, або близьких до них, є особливо придатними об'єктами модифікування метаболізму ліпідів та жирних кислот, особливо в мікроорганізмах та рослинах. Також, нуклеїнові кислоти з Arabidopsis thaliana, Brassica napus, Glycine max, Oryza sativa, або Triticum aestivum, або близьких до них організмів можуть використовуватися для ідентифікування таких послідовностей ДНК та ферментів у інших видів, придатних для модифікування біосинтезу молекул попередників жирних кислот у відповідних організмах. Цим винаходом також пропонується виділена послідовність нуклеїнової кислоти з рослини (Arabidopsis thaliana, Brassica napus,Glycine max, Oryza sativa, або Triticum aestivum) яка кодує протеїн ліпідного метаболізму (LMP), або його частину. Також даним винаходом пропонуються поліпептиди, кодовані нуклеїновими кислотами, гетерологічні поліпептиди, які включають поліпептиди 89960 8 кодовані нуклеїновими кислотами, та антитіла до цих поліпептидів. Також, даний винахід стосується та пропонує використання нуклеїнових кислот LMP при отримування трансгенних рослин, які мають змінений рівень або склад накопичувальної речовини насіння. Стосовно зміненого складу, даний винахід може бути застосований, наприклад, для підвищення відсотку олеїнової кислоти по відношенню до інших рослинних олій. Спосіб одержання трансгенної рослини, з модифікованим рівнем або складом накопичувальної речовини насіння, який включає етапи трансформації рослинної клітини за допомогою вектора експресії, який містить нуклеїнову кислоту LMP та отримання рослини з модифікованим рівнем або складом накопичувальної речовини насіння з рослинної клітини. В одній з реалізацій, рослина є видом з високим продукуванням олії як описано в Kinney et al. (1994, Current Opin.
ДивитисяДодаткова інформація
Назва патенту англійськоюNucleic acid molecules encoding wrinkled1-like polypeptides
Автори англійськоюHaertel Heiko, Bhatt Garima, Mittendorf Volker, Shank Karin J.
Назва патенту російськоюМолекулы нуклеиновой кислоты, которые кодируют wrinkled1-подобные полипептиды
Автори російськоюХертель Хайко, Бхатт Гарима, Миттендорф Фолькер, Шенк Карин Дж.
МПК / Мітки
МПК: A01H 1/06, C07H 21/04, C12N 9/02
Мітки: кодують, поліпептиди, молекули, wrinkled1-подібні, кислоти, нуклеїнової
Код посилання
<a href="https://ua.patents.su/126-89960-molekuli-nuklenovo-kisloti-yaki-koduyut-wrinkled1-podibni-polipeptidi.html" target="_blank" rel="follow" title="База патентів України">Молекули нуклеїнової кислоти, які кодують wrinkled1-подібні поліпептиди</a>
Спосіб визначення послідовності молекули нуклеїнової кислоти (варіанти) та набір для використання при визначенні послідовності нуклеїнової кислоти

Номер патенту: 48119
Опубліковано: 15.08.2002
Автор: Дрманач Радоє
МПК: G01N 37/00, C12Q 1/68, C12N 15/09, G01N 33/53
Мітки: визначенні, нуклеїнової, молекули, визначення, спосіб, набір, кислоти, варіанти, послідовності, використання
Формула / Реферат:
1. Способ определения последовательности молекулы нуклеиновой кислоты, отличающийся тем, что проводят гибридизацию молекулы с комплементарными последовательностями олигонуклеотидов из двух наборов малых олигонуклеотидных зондов известной последовательности, причем первый набор зондов прикреплен к твердой подложке, а второй набор зондов представляет собой меченые зонды, находящиеся в растворе, создают ковалентные связи между гибридизированным...
Невірусний вектор для введення нуклеїнової кислоти та спосіб його одержання, конденсований комплекс поліпептид/нуклеїнова кислота та спосіб вбудовування нуклеїнової кислоти, що представляє інтерес, в еукаріотич

Номер патенту: 77393
Опубліковано: 15.12.2006
Автори: Руссель Віллі, Алтон Ерік, Тагава Тошіакі, Мюррей Карл, Пєрузель Ерік, Меттьюз Девід, Менвелл Мішель, Міллєр Ендрю Девід
МПК: C12N 15/88, A61K 48/00, A61P 43/00, C12N 15/09, A61K 9/127, A61P 25/00, A61K 47/48, A61P 25/28, C07K 14/075, A61P 35/02, A61P 35/00, A61K 38/00, C12N 15/34
Мітки: нуклеїнової, представляє, конденсований, інтерес, введення, одержання, невірусний, комплекс, вбудовування, вектор, кислоти, спосіб, кислота, еукаріотич
Формула / Реферат:
1. Невірусний вектор для введення нуклеїнової кислоти, який містить конденсований комплекс поліпептид/нуклеїнова кислота і катіонний ліпід, де комплекс включає(а) нуклеотидну послідовність (NOI), що представляє інтерес; і(б) один або декілька аденовірусних поліпептидів або їх похідних, які упаковують нуклеїнову кислоту, де поліпептиди або їх похідні (І) мають здатність зв'язуватися з NOI; і (II) мають здатність конденсувати NOI;...
Олігонуклеотид для ампліфікації послідовностей нуклеїнових кислот, спосіб ампліфікації цільової послідовності нуклеїнової кислоти, набір для здійснення реакції ампліфікації нуклеїнової кислоти

Номер патенту: 49843
Опубліковано: 15.10.2002
Автор: Уілл Стівен Гордон
МПК: C12Q 1/68, C07H 21/00, C12N 15/09
Мітки: олігонуклеотид, спосіб, здійснення, нуклеїнових, ампліфікації, кислоти, набір, послідовності, кислот, реакції, послідовностей, цільової, нуклеїнової
Формула / Реферат:
1. Олігонуклеотид для ампліфікації послідовностей нуклеїнових кислот, який має загальну структуру:або,де: S1 - перша послідовність нуклеотидів довжиною, приблизно, від 5 до 50 нуклеотидів;S2 - друга послідовність довжиною від одного до трьох нуклеотидів;Nu - нуклеотид, до складу якого входить пуринова або піримідинова основа, яка включає в себе екзоциклічний амін;R - модифікаторна...
Рекомбінантна термостабільна днк-полімераза, фрагмент нуклеїнової кислоти, композиція для застосування в реакції секвенування днк, спосіб секвенування нуклеїнової кислоти-мішені, набір для секвенування нуклеїно

Номер патенту: 47423
Опубліковано: 15.07.2002
Автори: Рейчерт Фред Лоуренс, Кальман Ліса Вівіан, Гельфанд Девід Харроу
МПК: C12R 1/19, C12Q 1/68, C12N 15/54, C12N 9/12, C12N 15/09
Мітки: композиція, днк, фрагмент, термостабільна, нуклеїнової, нуклеїно, набір, спосіб, застосування, секвенування, реакції, кислоти, кислоти-мішені, днк-полімераза, рекомбінантна
Формула / Реферат:
1. Рекомбінантна термостабільна ДНК-полімераза, яка являє собою мутовану форму нативної термостабільної ДНК-полімерази, що має амінокислотну послідовність, яка містить амінокислотний фрагмент Ser Gln Ile Glu Leu Arg Xaa, в якому Xaa в положенні 7 вказаного фрагменту є залишком валіну Val або ізолейцину Ilе, причому вказана мутована форма модифікована таким чином, що вона містить відмінну від Glu амінокислоту в положенні 4 послідовності...
Виділений поліпептид омр106 зовнішньої мембрани moraxella catarrhalis, його пептидний фрагмент та молекули днк, що їх кодують, виділене антитіло, вакцина (варіанти) та антигенна композиція (варіанти), а також с

Номер патенту: 68332
Опубліковано: 16.08.2004
Автори: Такер Кеннет, Плосіла Лаура
МПК: C12R 1/36, C12N 15/31, C12Q 1/68, C07K 14/21, G01N 33/53, A61P 31/04, C07K 16/12, A61K 39/095
Мітки: moraxella, вакцина, мембрани, catarrhalis, кодують, молекули, виділене, антигенна, днк, композиція, зовнішньої, виділений, пептидний, антитіло, варіанти, омр106, також, поліпептид, фрагмент
Формула / Реферат:
1. Выделенный полипептид ОМР106, который является полипептидом наружной мембраны Moraxella catarrhalis и имеет молекулярную массу приблизительно от 180 кДа до приблизительно 230 кДа, как определено электрофорезом в SDS-полиакриламидном геле с использованием миозина скелетных мышц кролика и -галактозидазы Е.соlі в качестве стандартов молекулярных масс 200 кДа и 116,25 кДа,...
Попередній патент: Коаксіально-колінеарна антена
Наступний патент: Спосіб обтиснення безперервнолитого блюма або заготовки у рідкотвердому стані
Випадковий патент: Спосіб одержання солі 3-етил-5-метил-2-(2-аміноетоксиметил)-4-(2-хлорфеніл)-6-метил-1,4-дигідро-3,5-піридиндикарбоксилату і бензолсульфокислоти (безилату амлодипіну)