Антагоністи тол-подібного рецептора 3
Номер патенту: 109397
Опубліковано: 25.08.2015
Автори: Раухенбергер Роберт, Рутц Марк, Тен Фан, Сан Матео Лані, Саріскі Роберт Т., Херінга Катаріна, Світ, Реймонд, Каннінгхем Марк, Тєпляков Алєксєй, Фен Іцин, Ло Цзиньцюань, Ву Шен-Дзюн








Формула / Реферат
1. Виділене антитіло або фрагмент, що взаємодіє з TLR3, яке містить як варіабельну ділянку важкого ланцюга, так і варіабельну ділянку легкого ланцюга, де антитіло містить:
а) амінокислотні послідовності CDR1, 2 і 3 важкого ланцюга, як показано в SEQ ID NO: 82, 196 і 84, де HCDR2 з послідовністю SEQ ID NO: 196 додатково визначена як показано у формулі (V):
Xaa24-I-D-P-S-D-S-Y-T-N-Y-Xaa25-P-S-F-Q-G, (V)
де
Хаа24 може бути Phe або Arg; і
Хаа25 може бути Ala або Ser; і
b) амінокислотні послідовності CDR1, 2 і 3 легкого ланцюга, як показано в SEQ ID NO: 79, 80 і 195, де LCDR3 з послідовністю SEQ ID NO: 195 додатково визначена як показано у формулі (VI):
Q-Q-Xaa18-Хаа19-Хаа20-Хаа21-Хаа22-Хаа23-Т, (VI)
де
Хаа18 може бути Туr, Gly або Ala;
Хаа19 може бути Gly, Glu або Asn;
Хаа20 може бути Ser або Thr;
Хаа21 може бути Val, Ilе або Leu;
Хаа22 може бути Ser або Leu; і
Хаа23 може бути Ilе, Ser, Pro або Туr.
2. Виділене антитіло або фрагмент, що взаємодіє з TLR3, яке містить як варіабельну ділянку важкого ланцюга, так і варіабельну ділянку легкого ланцюга, і де антитіло містить:
амінокислотні послідовності CDR1, 2 і 3, як показано в SEQ ID NО: 82, 86 і 84; і амінокислотні послідовності CDR1, 2 і 3, як показано в SEQ ID NО: 79, 80 і 87.
3. Виділене антитіло або його фрагмент, що взаємодіє з TLR3, яке містить як варіабельну ділянку важкого ланцюга, так і як варіабельну ділянку легкого ланцюга, і де антитіло містить:
а) варіабельну ділянку важкого ланцюга з послідовністю SEQ ID NO: 216;
або
b) варіабельну ділянку легкого ланцюга з послідовністю SEQ ID NO: 41; або
c) варіабельну ділянку важкого ланцюга з послідовністю SEQ ID NO: 216 і
варіабельну ділянку легкого ланцюга з послідовністю SEQ ID NO: 41; або
d) важкий ланцюг з послідовністю SEQ ID NO: 220; або
е) легкий ланцюг з послідовністю SEQ ID NO: 156; або
f) важкий ланцюг з послідовністю SEQ ID NO: 220 і легкий ланцюг з послідовністю SEQ ID NO: 156.
4. Виділене антитіло або фрагмент за п. 1, де антитіло є повністю людським або адаптованим для людини.
5. Виділене антитіло або фрагмент за п. 4, де антитіло кон'юговане з поліетиленгліколем.
6. Виділене антитіло або фрагмент за п. 4, що має ізотип ІgG4.
7. Виділене антитіло або фрагмент за п. 4, де домен Fc містить мутації S229P, Р235А або L236A.
8. Фармацевтична композиція, яка містить виділене антитіло або фрагмент за п. 4 і фармацевтично прийнятний носій.
9. Важкий ланцюг виділеного антитіла, що містить амінокислотну послідовність SEQ ID NO: 216.
10. Легкий ланцюг виділеного антитіла, що містить амінокислотну послідовність SEQ ID NО: 41.
11. Важкий ланцюг виділеного антитіла, що містить амінокислотну послідовність SEQ ID NO: 220.
12. Легкий ланцюг виділеного антитіла, що містить амінокислотну послідовність SEQ ID NO: 156.
Текст
Реферат: Винахід стосується виділеного антитіла або його фрагмента, що зв'язується з тол-подібним рецептором 3 (TLR3). UA 109397 C2 (12) UA 109397 C2 UA 109397 C2 5 10 15 20 25 30 35 40 45 50 55 60 АНТАГОНІСТИ ТОЛ-ПОДІБНОГО РЕЦЕПТОРА 3 Дана заявка вимагає пріоритет заявок США: сер. № 61/109974 від 31 жовтня 2008 р. і сер. № 61/161860 від 20 березня 2009 р., а також сер. № 61/165100 від 31 березня 2009 р. і сер. № 61/173686 від 29 квітня 2009 р., повний зміст яких включено в дану заявку шляхом посилання. Галузь застосування винаходу Даний винахід стосується антитіл, які є антитілами-антагоністами тол-подібного рецептора 3 (TLR3), полінуклеотидів, кодуючих антитіла-антагоністи TLR3 або їх фрагменти, а також способів одержання й використання названих продуктів. Передумови створення винаходу Тол-подібний рецептор (TLR) регулює активацію вродженої імунної відповіді, а також розвиток адаптивного (специфічного) імунітету, активуючи каскади внутрішньоклітинної передачі сигналу у відповідь на появу бактеріальних, вірусних, паразитичних, а в деяких випадках і ендогенних лігандів (Lancaster et al., J. Physiol. 563:945-955, 2005). Розташовані на плазматичній мембрані TLR, TLR1, TLR2, TLR4 і LR6 розпізнають ліганди, включаючи білкові або ліпідні компоненти бактерій і грибків. Переважно внутрішньоклітинні тол-подібні рецептори, TLR3, TLR7 і TLR9 активуються дволанцюжковими РНК, одноланцюжковими РНК і неметильованими CpG ДНК, відповідно. Вважається, що дисрегуляція сигнального шляху TLR є причиною множини проблем, і терапевтичні стратегії розвиваються в цьому напрямку (Hoffman et al., Nat. Rev. Drug Discov. 4:879-880, 2005; Rezaei, Int. Immunopharmacol. 6:863-869, 2006; Wickelgren, Science 312:184-187, 2006). Наприклад, антагоністи TLR4, а також TLR7 і TLR9 знаходяться у стадії клінічних випробувань у лікуванні важкого сепсису й системного червоного волчака, відповідно (Kanzler et al., Nat. Med. 13:552-559, 2007). Сигнальний шлях TLR3 активується дволанцюжковою РНК, матричною РНК або РНК, вивільнюваною некротичними (загиблими) клітинами в процесі запалення або вірусної інфекції. Активація TLR3 індукує секрецію інтерферонів і прозапальних цитокінів, а також приводить до активації й збільшення числа імунних клітин, захищаючи, таким чином, від деяких мікробних інфекцій. Наприклад, домінантно негативний алель TLR3 був асоційований з підвищеною сприйнятливістю до вірусу герпесу форми Herpes Simplex encephalitis після первинного інфікування вірусом HSV-1 у дитячому віці (Zheng et al., Science 317:1522-1527 2007). У мишей нестача TLR3 пов'язана зі зниженою виживаністю при перевірочному зараженні вірусом Коксакі (Richer et al., PLoS One 4:e4127, 2009). Було показано, проте, що порушена регуляція сигнального шляху TLR3 має відношення до захворюваності й смертності при певних вірусних інфекціях, включаючи вірус західнонільської лихоманки, флебовірус, вірус ваксінія й вірус грипу типу A (Wang et al., Nat. Med. 10:1366-1373, 2004; Gowen et al., J. Immunol. 177:6301-6307, 2006; Hutchens et al., J. Immunol. 180:483-491, 2008; Le Goffic et al., PLoS Pathog. 2:E53, 2006). Також було продемонстровано, що TLR3 визначає механізм патогенезу при цілому ряді запальних, імуноопосередкованих і аутоімунних захворювань, які включають, наприклад, септичний шок (Cavassani et al., J. Exp. Med. 205:2609-2621, 2008), гостре ушкодження легенів (Murray et al., Am. J. Respir. Crit. Care Med. 178:1227-1237, 2008), ревматоїдний артрит (Kim et al., Immunol. Lett. 124:9-17, 2009; Brentano et al., Arth. Rheum. 52:2656-2665, 2005), астму (Sugiura et al., Am. J. Resp. Cell Mol. Biol. 40:654-662, 2009; Morishima et al., Int. Arch. Allergy Immunol. 145:163-174, 2008; Stowell et al., Respir. Res. 10:43, 2009), запальні захворювання кишечнику, такі як хвороба Крона й виразковий коліт (Zhou et al., J. Immunol. 178:4548-4556, 2007; Zhou et al., Proc. Natl. Acad. Sci. (USA) 104:7512-7515, 2007), аутоімунні захворювання печінки (Lang et al., J. Clin. Invest. 116:2456-2463, 2006) і діабет І типу (Dogusan et al., Diabetes 57:1236-1245, 2008; Lien and Zipris, Curr. Mol. Med. 9:52-68, 2009). Більше того, було показано, що органфспецифічне збільшення експресії TLR3 корелює з багатьма патологіями, які виникають внаслідок порушення регуляції запальної відповіді, такими як біліарний (холангіолітичний) цироз печінки (Takii et al., Lab Invest. 85:908-920, 2005), ревматоїдний артрит суглобів (Ospelt et al., Arthritis Rheum. 58:3684-3692, 2008), і зі станом слизової оболонки носа у пацієнтів з алергічним ринітом (Fransson et al., Respir. Res. 6:100, 2005). При некрозі звільнення клітинного вмісту, що включає ендогенні матричні РНК, індукує секрецію цитокінів, хемокінів і інших факторів, які ініціюють процес місцевого запалення, полегшують знищення клітинних залишків і відновний процес. Некроз часто приводить до запального процесу, що не припиняється, збільшуючи можливість розвитку хронічного або гострого запалення (Bergsbaken et al., Nature Reviews 7:99-109, 2009). Активація TLR3 у місці некрозу може сприяти розвитку ненормального запального процесу й привести до виникнення прозапального позитивного замкненого зворотного зв'язку за рахунок вивільнених лігандів TLR3. Таким чином, антагонізм до TLR3 може відігравати позитивну роль у багатьох захворюваннях, які супроводжуються хронічним або гострим запаленням і/або некрозом. 1 UA 109397 C2 5 10 15 20 25 30 35 40 45 50 55 Зменшення активації TLR3 може розглядатися як нова стратегія в лікуванні онкологічних захворювань, таких як нирковоклітинний рак і плоскоклітинний (епідермальний) рак шиї (Morikawa et al., Clin. Cancer Res. 13:5703-5709, 2007; Pries et al., Int. J. Mol. Med. 21: 209-215, L423F 2008). Крім того, алель TLR3 , що приводить до виробництва білка зі зменшеною активністю, був асоційований із захисною функцією у випадку "сухої", викликуваної віковими змінами, макулярної дегенерації (Yang et al., N. Engl. J. Med. 359:1456-1463, 2008), вказуючи на те, що антагоністи TLR3 можуть відігравати позитивну роль при цьому захворюванні. Захворювання, пов'язані із запаленням, також як і захворювання, що викликаються інфекціями, мають значний вплив на здоров'я й, отже, велике економічне значення. Проте, незважаючи на прогрес у багатьох сферах медицини, існує відносно мало можливостей для лікування більшості названих захворювань. Таким чином, існує необхідність у супресії активності TLR3 для лікування пов'язаних з функцією цього рецептора захворювань. Короткий опис креслень На фіг. 1 показаний ефект моноклональних антитіл (mАb) до TLR3 людини (huTLR3) в експерименті з використанням гена-репортера NF-kB. На фіг. 2A і 2В показаний ефект (%) інгібування або ефект mАb до huTLR3 в експерименті з використанням гена BEAS-2B. На фіг. 3A і 3В показаний ефект mАb до huTLR3 в експерименті з використанням гена NHBE. На фіг. 4 показаний ефект mАb до huTLR3 в експерименті з використанням гена PBMC. На фіг. 5A і 5В показаний ефект mАb до huTLR3 в експерименті з використанням гена HASM. На фіг. 6A, 6В і 6С показане зв'язування mАb до huTLR3 з мутантами TLR3. На фіг. 7A показані епітопи, розпізнавані mАb 15EVQ (чорним кольором) і C1068 (сірим кольором) (верхня частина малюнка), і епітопи, розпізнавані mАb 12QVQ/QSV (чорним кольором, нижня частина малюнка), накладені на зображення структури TLR3 ECD людини. На Фіг. 7В показана пертурбаційна карта вибіркового заміщення водню дейтерієм у випадку, коли TLR3 ECD утворює комплекс із mАb 15EVQ. На фіг. 8A і 8В показаний ефект сурогатних щурячих/мишачих mАb (mАb до мишачого TLR3, mАb 5429) в експериментах з використанням (A) гена-репортера NF-kB і (В) гена-репортера ISRE. На фіг. 9 показаний ефект сурогатних mАb (mАb 5429, mАb c1811) в експерименті з MEF CXCL10/IP-10. На фіг. 10 показана специфічність зв'язування сурогатних mАb з TLR3. Верхня частина малюнка: ізотипний контроль; нижня частина малюнка: mАb c1811. На фіг. 11 показаний вплив сурогатних mАb на рівень penH у моделі AHR. На фіг. 12 показаний ефект сурогатних mАb на загальне число нейтрофілів у рідині бронхоальвеолярного лаважу (BAL) при гіперчутливості дихальних шляхів (AHR). На фіг. 13 показаний ефект впливу сурогатних mАb на рівень продукції CXCL10/IP-10 у рідині BAL при AHR. На фіг. 14 показаний ефект сурогатних mАb на гістопатологічні показники при використанні декстрану сульфату натрію (DSS). На фіг. 15 показаний ефект сурогатних mАb на (A) гістопатологічні показники й (В) приплив нейтрофілів при перенесенні Т-клітин. На фіг. 16 показаний ефект сурогатних mАb на клінічні показники при колаген-індукованому артриті (CIA). На фіг. 17 показаний ефект сурогатних mАb на площу під кривою, що описує динаміку клінічних показників, при CIA. На фіг. 18 показаний ефект сурогатних mАb на виживаність мишей C57BL/6 після інтраназального введення вірусу грипу типу A/PR/8/34. На фіг. 19 показаний ефект сурогатних mАb на клінічні показники після зараження вірусом грипу типу A/PR/8/34. На фіг. 20 показаний ефект сурогатних mАb на вагу тіла протягом 14 днів після зараження вірусом грипу типу A/PR/8/34. На фіг. 21 показаний ефект сурогатних mАb на рівень глюкози в крові у тварин (A) WT DIO і (В) TLR3KO DIO після пробного введення глюкози. На фіг. 22 показаний ефект сурогатних mАb на рівень інсуліну у тварин WT DIO. На фіг. 23 показаний ефект mАb 15EVQ на (A) NTHi і (В) продукцію CXCL10/IP-10 і CCL5/RANTES у клітинах NHBE, індуковану риновірусом. 2 UA 109397 C2 5 10 15 20 25 30 35 40 45 50 55 На фіг. 24 показаний вплив mАb 15EVQ на (A) рівень sICAM-1 і (В) життєздатність клітин HUVEC. Короткий опис винаходу Перший аспект даного винаходу стосується опису виділених антитіл або антигензв’язувальних фрагментів, причому ці антитіла зв'язують щонайменше один амінокислотний залишок рецептора TLR3, вибраний із залишків K467, R488 або R489 в SEQ ID NO: 2. Наступний аспект даного винаходу стосується опису виділених антитіл або антигензв’язувальних фрагментів, причому антитіла зв'язують щонайменше один амінокислотний залишок рецептора TLR3, вибраний із групи залишків D116 або K145 в SEQ ID NO: 2. Наступний аспект даного винаходу стосується виділених антитіл, здатних взаємодіяти з TLR3, причому антитіла мають щонайменше одну з наступних властивостей: a) зменшують біологічну активність TLR3 в експериментах in vitro з використанням генарепортера NF-kB і поліінозинової:поліцитидилової кислоти (poly(I:C)), >50% при 60% продукцію IL-6 або CXCL10/IP-10 у клітинах BEAS-2B, стимульованих впливом
ДивитисяДодаткова інформація
Назва патенту англійськоюToll-like receptor 3 antagonists
Автори російськоюCunningham, Mark, San Mateo, Lani, Sarisky, Robert, T., Sweet, Raymond, Rauchenberger, Robert, Rutz, Mark, Feng, Yiqing, Heeringa, Katharine, Luo, Jinguan, Teng, Fang, Teplyakov, Alexey, Wu, Sheng-Jiun
МПК / Мітки
МПК: A61P 29/00, A61K 39/395
Мітки: антагоністи, тол-подібного, рецептора
Код посилання
<a href="https://ua.patents.su/180-109397-antagonisti-tol-podibnogo-receptora-3.html" target="_blank" rel="follow" title="База патентів України">Антагоністи тол-подібного рецептора 3</a>
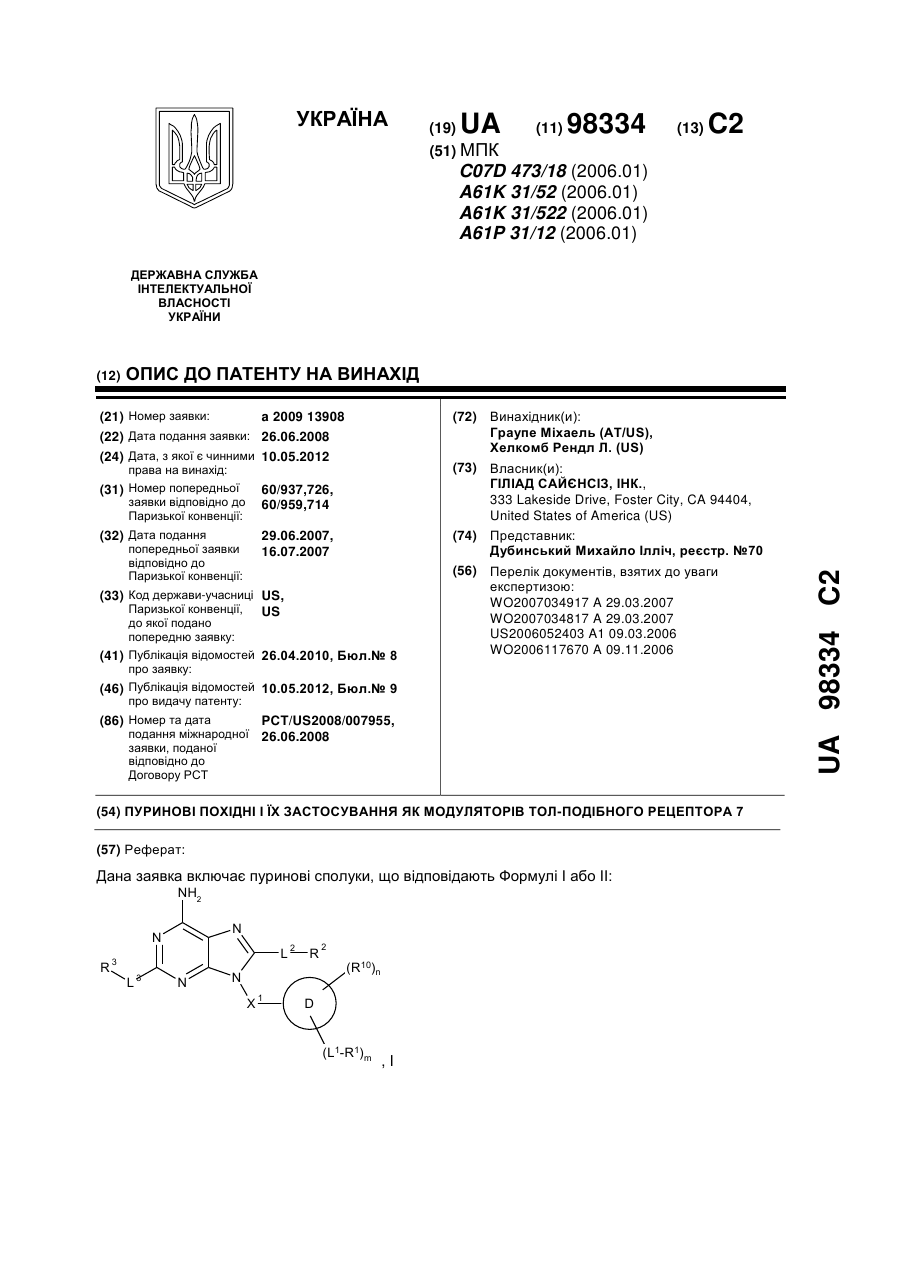
Пуринові похідні і їх застосування як модуляторів тол-подібного рецептора 7
Номер патенту: 98334
Опубліковано: 10.05.2012
Автори: Граупе Міхаель, Хелкомб Рендл Л.
МПК: A61K 31/52, A61K 31/522, A61P 31/12, C07D 473/18
Мітки: застосування, рецептора, пуринові, похідні, тол-подібного, модуляторів
Формула / Реферат:
1. Сполука Формули Іа: Iaабо фармацевтично прийнятна сіль зазначеної сполуки, у якій: R1 являє собою -NR4R5; R2 являє собою Н, алкіл, заміщений алкіл, карбоцикліл, заміщений карбоцикліл, карбоциклілалкіл, заміщений карбоциклілалкіл, арилалкіл, заміщений арилалкіл, гетероциклілалкіл, заміщений гетероциклілалкіл,...
Антагоністи b2 адренергічного рецептора

Номер патенту: 73965
Опубліковано: 17.10.2005
Автори: Чой Сеок-Кі, Моран Едмунд
МПК: A61K 39/00, A61K 31/167, A61P 11/08, A61K 39/44, A61P 1/00, A61P 15/06, A61P 3/04, A61K 31/137, G01N 33/53, A61P 43/00, A61K 31/135, A61K 31/138, C07C 233/43, A61K 31/165, A61P 11/06, C07C 213/00, A61K 39/395, C07C 215/00, G01N 33/566, A61P 25/00, A61K 38/00, A61K 51/00, A61P 11/00, A61P 3/10, G01N 33/543
Мітки: рецептора, адренергічного, антагоністи
Формула / Реферат:
1. Сполука формули:або її фармацевтично прийнятна сіль.2. Сполука за п. 1 для застосування як активного компонента лікувального засобу, особливо лікувального засобу для лікування захворювання, що опосередковане адренергічним рецептором у ссавців.3. Сполука за пунктом 2 для...
Антагоністи прогестеронового рецептора

Номер патенту: 98312
Опубліковано: 10.05.2012
Автори: Роттманн Антьє, Хассельманн Райнер, Шмідт Аня, Шульце-Мосгау Маркус, Фурманн Ульріке, Брудні-Кльоппель Маргарете, Прюс Штефан, Мьоллер Карстен, Гарке Гуннар, Клеве Арвед, Петроф Орлін
МПК: A61K 31/567, C07J 1/00, A61P 5/36, C07J 21/00, C07J 51/00
Мітки: рецептора, прогестеронового, антагоністи
Формула / Реферат:
1. Антагоніст прогестероновогорецептора загальної формули І:, формула І у якій R1 може бути атомом водню і R2 гідроксильною групою або R1 і R2 разом можуть бути оксогрупою. 2. Антагоніст прогестеронового рецептора загальної формули І за п. 1, що являє собою 11
Модифіковані антагоністи (мутеїни) рецептора il-4

Номер патенту: 94021
Опубліковано: 11.04.2011
Автори: Рожняк Стів, Лонгфре Малінда, Юнг Стефані Л., Пен Кларк, Грев Джері Майкл, Вонг Тереза Мо-фан, Томкінсон Адріан
МПК: A61P 11/00, C12N 15/24, A61K 38/20
Мітки: антагоністи, мутеїни, модифіковані, рецептора
Формула / Реферат:
1. Очищений полінуклеотид, який включає: (a) нуклеотидну послідовність, наведену в SEQ ID NO: 6; або(b) нуклеотидну послідовність, яка кодує поліпептид, що має амінокислотну послідовність, наведену в SEQ ID NO: 14.2. Експресуючий вектор, який включає полінуклеотид за п. 1.3. Клітина-хазяїн, яка включає експресуючий вектор за п. 2.4. Спосіб одержання модифікованого антагоніста (мутеїна) рецептора IL-4, який...
Антагоністи рецептора il-8

Номер патенту: 98456
Опубліковано: 25.05.2012
Автор: Буш-Петерсен Якоб
МПК: A61K 31/44, A01N 43/76, C07D 211/84, A01N 43/40, C07D 413/00, C07D 211/72, C07D 498/00, C07D 263/60, A61K 31/42, C07D 213/63, C07D 213/70, C07D 263/52
Мітки: рецептора, антагоністи
Формула / Реферат:
1. Сполука відповідно до Формули (І):, (I) в якій X вибраний з групи, що складається з галогену, С1-3алкілу, С1-3алкокси, ціано, CF3 і OCF3; R2 вибраний з групи, що складається з С3-6циклоалкілу, фенілу й гетероарилу, причому феніл або гетероарил за необхідності заміщені одним або двома замісниками,...
Попередній патент: Брикет для виплавки сталі та спосіб виробництва брикету
Наступний патент: Одноразовий контейнер, спосіб створення тиску у одноразовому контейнері та застосування контейнера
Випадковий патент: Спосіб лікування кавернозної гемангіоми нижньої кінцівки