Розчинний мутантний ctla4 та його застосування
Номер патенту: 87432
Опубліковано: 27.07.2009
Автори: Піч Роберт Дж., Лінслі Петер С., Наємура Джозеф Р., Бейорет Юрген






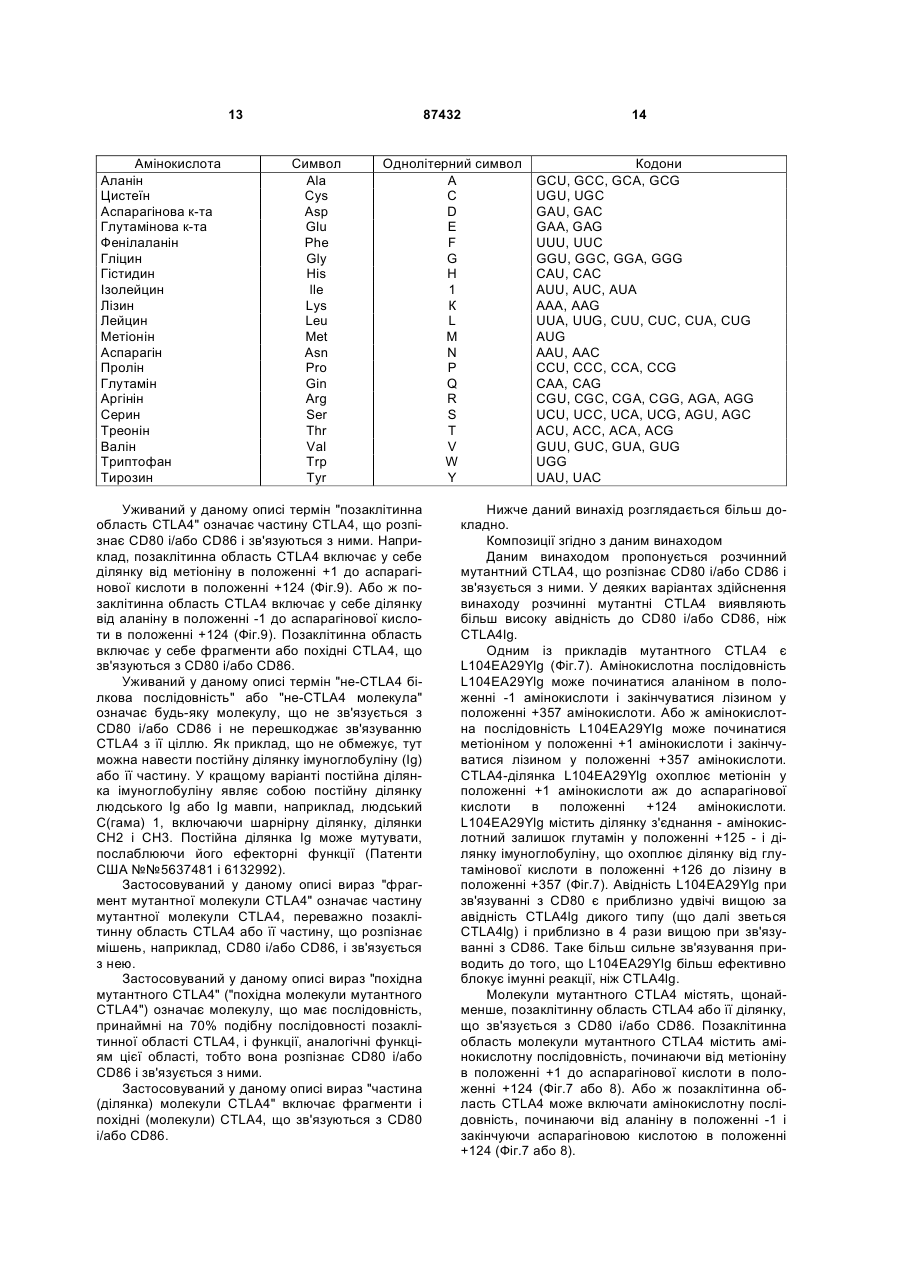

Формула / Реферат
1. Молекула мутантного CTLA4, яка зв'язується з CD80 і/або з CD86 і містить позаклітинну область CTLA4, як показано на Фіг.9, починається з аланіну в позиції -1 або метіоніну в позиції +1 та закінчується аспарагіновою кислотою в позиції +124, в позаклітинній області, (а) аланін у положенні +29 замінений на амінокислоту, яку вибирають із групи, що складається з тирозину, лейцину, триптофану і треоніну, та (б) лейцин у положенні +104 замінений на глутамінову кислоту.
2. Молекула мутантного CTLA4 за п. 1, яка відрізняється тим, що додатково містить амінокислотну послідовність, що змінює розчинність, афінність або валентність молекули мутантного CTLA4.
3. Молекула мутантного CTLA4 за п. 2, яка відрізняється тим, що амінокислотна послідовність містить постійну ділянку людського імуноглобуліну, або його частину, де постійна ділянка імуноглобуліну може включати одну або більше мутацій для зниження ефекторної функції.
4. Молекула мутантного CTLA4 за п. 2, яка відрізняється тим, що додатково містить амінокислотну послідовність, котра дозволяє секрецію молекули мутантного CTLA4.
5. Молекула мутантного CTLA4 за п. 4, яка відрізняється тим, що амінокислотна послідовність, що дозволяє мутантній молекулі CTLA4 секретувати, містить сигнальний пептид онкостатину М.
6. Молекула мутантного CTLA4 за п. 1, яка відрізняється тим, що містить амінокислотну послідовність, що є зв'язаною з метіоніном у положенні +1 і закінчується аспарагіновою кислотою в положенні +124, як показано на Фіг. 7.
7. Молекула мутантного CTLA4 за п. 1, яка відрізняється тим, що містить амінокислотну послідовність, що є зв'язаною з аланіном у положенні –1 і закінчується аспарагіновою кислотою в положенні +124, як показано на Фіг. 7.
8. Молекула мутантного CTLA4 за п. 3, яка відрізняється тим, що постійна ділянка людського імуноглобуліну мутує таким чином, щоб включити заміну цистеїну в положенні +130 на серин, заміну цистеїну в положенні +136 на серин, заміну цистеїну в положенні +139 на серин і заміну проліну в положенні +148 на серин, як показано на Фіг. 7.
9. Молекула мутантного CTLA4, яка із більш високою авідністю, ніж CTLA4, зв'язується з CD80 і/або CD86, містить позаклітинну область CTLA4, яка відрізняється тим, що в позаклітинній області аланін у положенні +29 замінений на тирозин, а лейцин у положенні +104 замінений на глутамінову кислоту, як показано на Фіг. 7.
10. Молекула мутантного CTLA4 за п. 9, яка відрізняється тим, що додатково містить амінокислотну послідовність, котра змінює розчинність, афінність або валентність молекули мутантного CTLA4.
11. Молекула мутантного CTLA4 за п. 10, яка відрізняється тим, що амінокислотна послідовність містить постійну ділянку людського імуноглобуліну, або його частину, де постійна ділянка імуноглобуліну може включати одну або більше мутацій для зниження ефекторної функції.
12. Молекула мутантного CTLA4 за п. 10, яка відрізняється тим, що додатково містить амінокислотну послідовність, котра дозволяє секрецію молекули мутантного CTLA4.
13. Молекула мутантного CTLA4 за п. 12, яка відрізняється тим, що амінокислотна послідовність, що дозволяє мутантній молекулі CTLA4 секретувати, містить сигнальний пептид онкостатину М.
14. Молекула мутантного CTLA4, яка містить амінокислотну послідовність, що є зв'язаною з метіоніном у положенні +1 і закінчується аспарагіновою кислотою в положенні +124, як показано на Фіг. 7.
15. Молекула мутантного CTLA4, яка містить амінокислотну послідовність, що є зв'язаною з аланіном у положенні -1 і закінчується аспарагіновою кислотою в положенні +124, як показано на Фіг. 7.
16. Молекула мутантного CTLA4 за п. 11, яка відрізняється тим, що постійна ділянка людського імуноглобуліну мутує таким чином, щоб включити заміну цистеїну в положенні +130 на серин, заміну цистеїну в положенні +136 на серин, заміну цистеїну в положенні +139 на серин і заміну проліну в положенні +148 на серин, як показано на Фіг. 7.
17. Молекула мутантного CTLA4, яка із більш високою авідністю, ніж CTLA4, зв'язується з CD80 і/або CD86, містить позаклітинну область CTLA4, яка відрізняється тим, що в позаклітинній області лейцин у положенні +104 замінений на глутамінову кислоту, як показано на Фіг. 8.
18. Молекула нуклеїнової кислоти, яка містить нуклеотидну послідовність, котра кодує амінокислотну послідовність, що відповідає молекулі мутантного CTLA4 за п. 1.
19. Молекула нуклеїнової кислоти, яка містить нуклеотидну послідовність, котра кодує амінокислотну послідовність, що відповідає молекулі мутантного CTLA4 за п. 9 або п. 17.
20. Молекула нуклеїнової кислоти, яка має нуклеотидну послідовність, котра починається з аденіну в нуклеотидному положенні +1 і закінчується аденіном у положенні +1071, як показано на Фіг. 8, або містить нуклеотидну послідовність, що починається з гуанідину в нуклеотидному положенні -3 та закінчується аденіном у нуклеотидному положенні +1071, як показано на Фіг. 8.
21. Молекула нуклеїнової кислоти, яка має нуклеотидну послідовність, котра починається з аденіну в нуклеотидному положенні +1 і закінчується аденіном у положенні +1071, як показано на Фіг. 7, або містить нуклеотидну послідовність, що починається з гуанідину в нуклеотидному положенні -3 та закінчується аденіном у нуклеотидному положенні +1071, як показано на Фіг. 7.
22. Молекула нуклеїнової кислоти, яка має нуклеотидну послідовність, котра починається з аденіну в положенні +1 і закінчується тиміном у положенні +372, як показано на Фіг. 8, або містить нуклеотидну послідовність, що починається з гуанідину в нуклеотидному положенні -3 та закінчується аденіном у нуклеотидному положенні +1071, як показано на Фіг. 8.
23. Молекула нуклеїнової кислоти, яка має нуклеотидну послідовність, котра починається з аденіну в положенні +1 і закінчується тиміном у положенні +372, як показано на Фіг. 7, або містить нуклеотидну послідовність, що починається з гуанідину в нуклеотидному положенні -3 та закінчується аденіном у нуклеотидному положенні +1071, як показано на Фіг. 7.
24. Вектор, що містить нуклеотидну послідовність за будь-яким з пп. 18-23.
25. Вектор, що кодує CTLA4 мутантну молекулу, де вектор позначений як pD16 L104EA29YIg і є депонований в АТСС як АТСС № РТА-2104.
26. Система вектор-хазяїн, що являє собою вектор за п. 24 або за п. 25 у відповідній клітині-хазяїні.
27. Система вектор-хазяїн за п. 26, яка відрізняється тим, що відповідною клітиною-хазяїном є бактеріальна клітина або еукаріотична клітина.
28. Клітина-хазяїн, що містить вектор за п. 24 або за п. 25.
29. Клітина-хазяїн за п.28, яка відрізняється тим, що є еукаріотичною клітиною.
30. Клітина-хазяїн за п. 29, яка відрізняється тим, що еукаріотичною клітиною є COS-клітина.
31. Клітина-хазяїн за п. 29, яка відрізняється тим, що еукаріотичною клітиною є клітина яєчника китайського хом'ячка (СНО).
32. Клітина-хазяїн за п. 31, яка відрізняється тим, що клітину СНО вибирають із групи, що складається із DG44, СНО-К1, СНО-К1 Tet-On клітинної лінії, СНО, позначеної як ЕСАСС 85050302, СНО клон 13, СНО клон В, CHO-K1/SF і RR-CHOK1.
33. Спосіб одержання мутантного білка, яким є CTLA4 мутантна молекула, за яким систему вектор-хазяїн за п. 26 вирощують таким чином, щоб продукувати у клітині-хазяїні мутантну молекулу CTLA4, і відділяють білок, котрий при цьому продукується.
34. Спосіб одержання мутантного білка, яким є CTLA4 мутантна молекула, за яким клітину-хазяїна за п. 28 вирощують таким чином, щоб продукувати у клітині-хазяїні CTLA4 мутантну молекулу, і відділяють білок, що при цьому продукується.
35. Мутантна молекула CTLA4, яку одержують за способом за п. 33.
36. Мутантна молекула CTLA4, яку одержують за способом за п. 34.
37. Спосіб регулювання взаємодії Т-клітини з CD80- і/або CD86-позитивною клітиною, при якому CD80- і/або CD86-позитивну клітину приводять у контакт з молекулою розчинного мутантного CTLA4 за п. 1 з утворенням комплексу: молекула мутантного CTLA4/CD80, або комплексу: молекула мутантного CTLA4/CD86, причому цей комплекс перешкоджає взаємодії між Т-клітиною і CD80- і/або CD86-позитивною клітиною.
38. Спосіб регулювання взаємодії Т-клітини з CD80- і/або СD86-позитивною клітиною, при якому приводять у контакт CD80- і/або CD86-позитивну клітину з молекулою розчинного мутантного CTLA4 за п. 9 з утворенням комплексу: молекула мутантного CTLA4/CD80, або комплексу: молекула мутантного CTLA4/CD86, причому цей комплекс перешкоджає взаємодії між Т-клітиною і CD80- і/або CD86-позитивною клітиною.
39. Спосіб регулювання взаємодії Т-клітини з CD80- і/або СD86-позитивною клітиною, при якому приводять у контакт CD80- і/або CD86-позитивну клітину з молекулою розчинного мутантного CTLA4 за п. 17 з утворенням комплексу: молекула мутантного CTLA4/CD80, або комплексу: молекула мутантного CTLA4/CD86, причому цей комплекс перешкоджає взаємодії між Т-клітиною і CD80- і/або CD86-позитивною клітиною.
40. Спосіб за п. 37, який відрізняється тим, що CD80- і/або СD86-позитивна клітина контактує з фрагментом або похідною молекули мутантного CTLA4.
41. Спосіб за п. 38, який відрізняється тим, що CD80- і/або CD86-позитивна клітина контактує з фрагментом або похідною молекули мутантного CTLA4.
42. Спосіб за п. 39, який відрізняється тим, що CD80- і/або CD86-позитивна клітина контактує з фрагментом або похідною молекули мутантного CTLA4 .
43. Спосіб за пп. 37, 38 або 39, який відрізняється тим, що CD80- і/або CD86-позитивною клітиною є клітина, що презентує антигени.
44. Спосіб за п. 37 або 39, який відрізняється тим, що взаємодія CTLA4-позитивних Т-клітин із CD80- і/або CD86-позитивними клітинами інгібується.
45. Спосіб за п. 38, який відрізняється тим, що взаємодія CTLA4-позитивних Т-клітин із CD80- і/або CD86-позитивними клітинами інгібується.
46. Спосіб лікування захворювань імунної системи, опосередкованих взаємодією Т-клітин з CD80- і/або CD86-позитивними клітинами, при якому пацієнту вводять молекули розчинного мутантного CTLA4 за п. 1 для регулювання Т-клітинної взаємодії з CD80- і/або CD86-позитивними клітинами.
47. Спосіб лікування захворювань імунної системи, опосередкованих взаємодією Т-клітин з CD80- і/або CD86-позитивними клітинами, при якому пацієнту вводять молекулу мутантного CTLA4 за п. 9 для регулювання Т-клітинної взаємодії з CD80- і/або CD86-позитивними клітинами.
48. Спосіб лікування захворювань імунної системи, опосередкованих взаємодією Т-клітин з CD80- і/або CD86-позитивними клітинами, при якому пацієнту вводять молекулу розчинного мутантного CTLA4 за п. 17 для регулювання Т-клітинної взаємодії з CD80- і/або CD86-позитивними клітинами.
49. Спосіб за п. 46 або 48, який відрізняється тим, що зазначена Т-клітинна взаємодія інгібується.
50. Спосіб за п. 47, який відрізняється тим, що зазначена Т-клітинна взаємодія інгібується.
51. Спосіб інгібування гомологічної хвороби у пацієнта, при якому пацієнту вводять молекулу мутантного CTLA4 за п. 1 і ліганду, реактивного у відношенні до IL-4.
52. Спосіб інгібування гомологічної хвороби у пацієнта, при якому пацієнту вводять молекулу мутантного CTLA4 за п. 9 і ліганду, реактивного у відношенні до IL-4.
53. Спосіб інгібування гомологічної хвороби у пацієнта, при якому пацієнту вводять молекулу мутантного CTLA4 за п. 17 і ліганду, реактивного у відношенні до IL-4.
54. Молекула мутантного CTLA4, що кодується молекулою нуклеїнової кислоти, депонованою як АТСС № РТА-2104.
55. Молекула ДНК, що кодує молекулу мутантного CTLA4, яка відрізняється тим, що молекула ДНК депонується як АТСС № РТА-2104.
56. Молекула мутантного CTLA4, що містить нуклеотидну послідовність, котра починається з метіоніну в положенні +1 і закінчується лізином у положенні +357, як показано на Фіг. 7, або яка починається з аланіну в положенні -1 та закінчується лізином у положенні +357, як показано на Фіг. 7.
57. Молекула нуклеїнової кислоти, що кодує молекулу мутантного CTLA4 за п.56.
58. Ділянка молекули мутантного CTLA4, котра (молекула) кодується нуклеотидною молекулою, депонованою як АТСС № РТА-2104, яка (ділянка) відрізняється тим, що вона містить повну позаклітинну область мутантного CTLA4, що зв'язується з CD80 і/або CD86.
59. Ділянка молекули мутантного CTLA4 за п. 58, яка відрізняється тим, що додатково містить Ig-половину.
60. Молекула нуклеїнової кислоти, яка містить ділянку молекули нуклеїнової кислоти, що кодує молекулу мутантного CTLA4 і має АТСС № РТА-2104, яка відрізняється тим, що вона кодує повну позаклітинну область молекули мутантного CTLA4, що зв'язується з CD80 і/або CD86.
61. Молекула нуклеїнової кислоти за п. 60, яка відрізняється тим, що додатково містить нуклеотидну молекулу, що кодує Ig-половину.
62. Фармацевтична композиція для лікування захворювання імунної системи, яка містить фармацевтично прийнятний носій і молекулу мутантного CTLA4 за п. 1 або 17.
63. Фармацевтична композиція для лікування захворювання імунної системи, яка містить фармацевтично прийнятний носій і молекулу мутантного CTLA4 за п. 9.
64. Молекула мутантного CTLA4 за п. 17, яка відрізняється тим, що амінокислотна послідовність, яка змінює розчинність, афінність або валентність, містить і ділянку імуноглобуліну, і тим, що ділянка імуноглобуліну може включати один або більше мутацій для зниження ефекторної функції.
65. Спосіб регулювання взаємодії Т-клітини з CD80- і/або СD86-позитивною клітиною, при якому приводять у контакт CD80- і/або CD86-позитивну клітину з молекулою розчинного мутантного CTLA4 за п. 56 таким чином, щоб утворювався комплекс: молекула мутантного CTLA4/CD80, або комплекс: молекула мутантного CTLA4/CD86, причому цей комплекс перешкоджає взаємодії між Т-клітиною і CD80- і/або CD86-позитивною клітиною.
66. Спосіб за п. 65, який відрізняється тим, що CD80- і/або СD86-позитивна клітина контактує з фрагментом або похідною молекули мутантного CTLA4.
67. Спосіб за п. 65, який відрізняється тим, що CD80- і/або CD86-позитивною клітиною є клітина, що презентує антигени.
68. Спосіб за п. 65, який відрізняється тим, що взаємодія CTLA4-позитивних Т-клітин із CD80- і/або CD86-позитивними клітинами інгібується.
69. Спосіб лікування захворювань імунної системи, опосередкованих взаємодією Т-клітин з CD80- і/або CD86-позитивними клітинами, при якому пацієнту вводять молекулу мутантного CTLA4 за п. 56 для регулювання Т-клітинної взаємодії з CD80- і/або CD86-позитивними клітинами.
70. Спосіб лікування захворювань імунної системи, опосередкованих взаємодією Т-клітин з CD80- і/або CD86-позитивними клітинами, при якому пацієнту вводять молекулу розчинного мутантного CTLA4 за п. 56 для регулювання Т-клітинної взаємодії з CD80- і/або CD86-позитивними клітинами.
71. Молекула мутантного CTLA4, яка зв'язується з CD80 і/або з CD86 і містить позаклітинну область CTLA4, як показано на Фіг.9, починається з аланіну в позиції -1 або метіоніну в позиції +1 та закінчується аспарагіновою кислотою в позиції +124, або її частиною, яка відрізняється тим, що в позаклітинній області або його частині, аланін у положенні 29 замінений на тирозин, та лейцин у положенні 104 замінений, та також містить амінокислотну послідовність, яка змінює розчинність, афінність або валентність молекули мутантного CTLA4.
72. Молекула мутантного CTLA4 за п. 71, яка відрізняється тим, що амінокислотна послідовність містить постійну ділянку людського імуноглобуліну, або його частину, де постійна ділянка імуноглобуліну може включати одну або більше мутацій для зниження ефекторної функції.
73. Молекула мутантного CTLA4, що містить амінокислотну послідовність, що є зв'язаною з метіоніном у положенні +1 і закінчується аспарагіновою кислотою в положенні +124, як показано на Фіг. 7, або її частину, яка зв'язується з CD80 і/або з CD86,
або містить амінокислотну послідовність, що є зв'язаною з аланіном у положенні -1 і закінчується аспарагіновою кислотою в положенні 124, як показано на Фіг. 7, або її частину, яка зв'язується з CD80 і/або з CD86.
74. Молекула мутантного CTLA4 за п. 73, яка відрізняється тим, що амінокислотна послідовність містить постійну ділянку людського імуноглобуліну, або його частину, де постійна ділянка імуноглобуліну може включати одну або більше мутацій для зниження ефекторної функції.
75. Молекула мутантного CTLA4 за пп. 1, 9, 14, 15, 35, 54, 56, 72 або 74, що має меншу швидкість дисоціації при зв'язуванні з CD86, ніж CTLA4 дикого типу.
76. Молекула мутантного CTLA4, що має менші швидкості асоціації і дисоціації при зв'язуванні з CD80 і/або CD86, ніж CTLA4 дикого типу за пп. 1, 9, 14, 15, 35, 54, 56, 72 або 74, що є розчинною.
Текст
1. Молекула мутантного CTLA4, яка зв'язується з CD80 і/або з CD86 і містить позаклітинну область CTLA4, як показано на Фіг.9, починається з аланіну в позиції -1 або метіоніну в позиції +1 та закінчується аспарагіновою кислотою в позиції +124, в позаклітинній області, (а) аланін у положенні +29 замінений на амінокислоту, яку вибирають із групи, що складається з тирозину, лейцину, триптофану і треоніну, та (б) лейцин у положенні +104 замінений на глутамінову кислоту. 2. Молекула мутантного CTLA4 за п.1, яка відрізняється тим, що додатково містить амінокислотну послідовність, що змінює розчинність, афінність або валентність молекули мутантного CTLA4. 3. Молекула мутантного CTLA4 за п.2, яка відрізняється тим, що амінокислотна послідовність містить постійну ділянку людського імуноглобуліну, або його частину, де постійна ділянка імуноглобу 2 (19) 1 3 13. Молекула мутантного CTLA4 за п.12, яка відрізняється тим, що амінокислотна послідовність, що дозволяє мутантній молекулі CTLA4 секретувати, містить сигнальний пептид онкостатину М. 14. Молекула мутантного CTLA4, яка містить амінокислотну послідовність, що є зв'язаною з метіоніном у положенні +1 і закінчується аспарагіновою кислотою в положенні +124, як показано на Фіг.7. 15. Молекула мутантного CTLA4, яка містить амінокислотну послідовність, що є зв'язаною з аланіном у положенні -1 і закінчується аспарагіновою кислотою в положенні +124, як показано на Фіг.7. 16. Молекула мутантного CTLA4 за п.11, яка відрізняється тим, що постійна ділянка людського імуноглобуліну мутує таким чином, щоб включити заміну цистеїну в положенні +130 на серин, заміну цистеїну в положенні +136 на серин, заміну цистеїну в положенні +139 на серин і заміну проліну в положенні +148 на серин, як показано на Фіг.7. 17. Молекула мутантного CTLA4, яка із більш високою авідністю, ніж CTLA4, зв'язується з CD80 і/або CD86, містить позаклітинну область CTLA4, яка відрізняється тим, що в позаклітинній області лейцин у положенні +104 замінений на глутамінову кислоту, як показано на Фіг.8. 18. Молекула нуклеїнової кислоти, яка містить нуклеотидну послідовність, котра кодує амінокислотну послідовність, що відповідає молекулі мутантного CTLA4 за п.1. 19. Молекула нуклеїнової кислоти, яка містить нуклеотидну послідовність, котра кодує амінокислотну послідовність, що відповідає молекулі мутантного CTLA4 за п.9 або п.17. 20. Молекула нуклеїнової кислоти, яка має нуклеотидну послідовність, котра починається з аденіну в нуклеотидному положенні +1 і закінчується аденіном у положенні +1071, як показано на Фіг.8, або містить нуклеотидну послідовність, що починається з гуанідину в нуклеотидному положенні -3 та закінчується аденіном у нуклеотидному положенні +1071, як показано на Фіг.8. 21. Молекула нуклеїнової кислоти, яка має нуклеотидну послідовність, котра починається з аденіну в нуклеотидному положенні +1 і закінчується аденіном у положенні +1071, як показано на Фіг.7, або містить нуклеотидну послідовність, що починається з гуанідину в нуклеотидному положенні -3 та закінчується аденіном у нуклеотидному положенні +1071, як показано на Фіг.7. 22. Молекула нуклеїнової кислоти, яка має нуклеотидну послідовність, котра починається з аденіну в положенні +1 і закінчується тиміном у положенні +372, як показано на Фіг.8, або містить нуклеотидну послідовність, що починається з гуанідину в нуклеотидному положенні -3 та закінчується аденіном у нуклеотидному положенні +1071, як показано на Фіг.8. 23. Молекула нуклеїнової кислоти, яка має нуклеотидну послідовність, котра починається з аденіну в положенні +1 і закінчується тиміном у положенні +372, як показано на Фіг.7, або містить нуклеотидну послідовність, що починається з гуанідину в нуклеотидному положенні -3 та закінчується аденіном у нуклеотидному положенні +1071, як показано на Фіг.7. 87432 4 24. Вектор, що містить нуклеотидну послідовність за будь-яким з пп.18-23. 25. Вектор, що кодує CTLA4 мутантну молекулу, де вектор позначений як pD16 L104EA29YIg і є депонований в АТСС як АТСС № РТА-2104. 26. Система вектор-хазяїн, що являє собою вектор за п.24 або за п.25 у відповідній клітині-хазяїні. 27. Система вектор-хазяїн за п.26, яка відрізняється тим, що відповідною клітиною-хазяїном є бактеріальна клітина або еукаріотична клітина. 28. Клітина-хазяїн, що містить вектор за п.24 або за п.25. 29. Клітина-хазяїн за п.28, яка відрізняється тим, що є еукаріотичною клітиною. 30. Клітина-хазяїн за п.29, яка відрізняється тим, що еукаріотичною клітиною є COS-клітина. 31. Клітина-хазяїн за п.29, яка відрізняється тим, що еукаріотичною клітиною є клітина яєчника китайського хом'ячка (СНО). 32. Клітина-хазяїн за п.31, яка відрізняється тим, що клітину СНО вибирають із групи, що складається із DG44, СНО-К1, СНО-К1 Tet-On клітинної лінії, СНО, позначеної як ЕСАСС 85050302, СНО клон 13, СНО клон В, CHO-K1/SF і RR-CHOK1. 33. Спосіб одержання мутантного білка, яким є CTLA4 мутантна молекула, за яким систему вектор-хазяїн за п.26 вирощують таким чином, щоб продукувати у клітині-хазяїні мутантну молекулу CTLA4, і відділяють білок, котрий при цьому продукується. 34. Спосіб одержання мутантного білка, яким є CTLA4 мутантна молекула, за яким клітину-хазяїна за п.28 вирощують таким чином, щоб продукувати у клітині-хазяїні CTLA4 мутантну молекулу, і відділяють білок, що при цьому продукується. 35. Мутантна молекула CTLA4, яку одержують за способом за п.33. 36. Мутантна молекула CTLA4, яку одержують за способом за п.34. 37. Спосіб регулювання взаємодії Т-клітини з CD80- і/або CD86-позитивною клітиною, при якому CD80- і/або CD86-позитивну клітину приводять у контакт з молекулою розчинного мутантного CTLA4 за п.1 з утворенням комплексу: молекула мутантного CTLA4/CD80, або комплексу: молекула мутантного CTLA4/CD86, причому цей комплекс перешкоджає взаємодії між Т-клітиною і CD80і/або CD86-позитивною клітиною. 38. Спосіб регулювання взаємодії Т-клітини з CD80- і/або СD86-позитивною клітиною, при якому приводять у контакт CD80- і/або CD86-позитивну клітину з молекулою розчинного мутантного CTLA4 за п.9 з утворенням комплексу: молекула мутантного CTLA4/CD80, або комплексу: молекула мутантного CTLA4/CD86, причому цей комплекс перешкоджає взаємодії між Т-клітиною і CD80і/або CD86-позитивною клітиною. 39. Спосіб регулювання взаємодії Т-клітини з CD80- і/або СD86-позитивною клітиною, при якому приводять у контакт CD80- і/або CD86-позитивну клітину з молекулою розчинного мутантного CTLA4 за п.17 з утворенням комплексу: молекула мутантного CTLA4/CD80, або комплексу: молекула мутантного CTLA4/CD86, причому цей комплекс 5 перешкоджає взаємодії між Т-клітиною і CD80і/або CD86-позитивною клітиною. 40. Спосіб за п.37, який відрізняється тим, що CD80- і/або СD86-позитивна клітина контактує з фрагментом або похідною молекули мутантного CTLA4. 41. Спосіб за п.38, який відрізняється тим, що CD80- і/або CD86-позитивна клітина контактує з фрагментом або похідною молекули мутантного CTLA4. 42. Спосіб за п.39, який відрізняється тим, що CD80- і/або CD86-позитивна клітина контактує з фрагментом або похідною молекули мутантного CTLA4. 43. Спосіб за пп.37, 38 або 39, який відрізняється тим, що CD80- і/або CD86-позитивною клітиною є клітина, що презентує антигени. 44. Спосіб за п.37 або 39, який відрізняється тим, що взаємодія CTLA4-позитивних Т-клітин із CD80і/або CD86-позитивними клітинами інгібується. 45. Спосіб за п.38, який відрізняється тим, що взаємодія CTLA4-позитивних Т-клітин із CD80- і/або CD86-позитивними клітинами інгібується. 46. Спосіб лікування захворювань імунної системи, опосередкованих взаємодією Т-клітин з CD80і/або CD86-позитивними клітинами, при якому пацієнту вводять молекули розчинного мутантного CTLA4 за п.1 для регулювання Т-клітинної взаємодії з CD80- і/або CD86-позитивними клітинами. 47. Спосіб лікування захворювань імунної системи, опосередкованих взаємодією Т-клітин з CD80і/або CD86-позитивними клітинами, при якому пацієнту вводять молекулу мутантного CTLA4 за п.9 для регулювання Т-клітинної взаємодії з CD80і/або CD86-позитивними клітинами. 48. Спосіб лікування захворювань імунної системи, опосередкованих взаємодією Т-клітин з CD80і/або CD86-позитивними клітинами, при якому пацієнту вводять молекулу розчинного мутантного CTLA4 за п.17 для регулювання Т-клітинної взаємодії з CD80- і/або CD86-позитивними клітинами. 49. Спосіб за п.46 або 48, який відрізняється тим, що зазначена Т-клітинна взаємодія інгібується. 50. Спосіб за п.47, який відрізняється тим, що зазначена Т-клітинна взаємодія інгібується. 51. Спосіб інгібування гомологічної хвороби у пацієнта, при якому пацієнту вводять молекулу мутантного CTLA4 за п.1 і ліганду, реактивного у відношенні до IL-4. 52. Спосіб інгібування гомологічної хвороби у пацієнта, при якому пацієнту вводять молекулу мутантного CTLA4 за п.9 і ліганду, реактивного у відношенні до IL-4. 53. Спосіб інгібування гомологічної хвороби у пацієнта, при якому пацієнту вводять молекулу мутантного CTLA4 за п.17 і ліганду, реактивного у відношенні до IL-4. 54. Молекула мутантного CTLA4, що кодується молекулою нуклеїнової кислоти, депонованою як АТСС № РТА-2104. 55. Молекула ДНК, що кодує молекулу мутантного CTLA4, яка відрізняється тим, що молекула ДНК депонується як АТСС № РТА-2104. 56. Молекула мутантного CTLA4, що містить нуклеотидну послідовність, котра починається з меті 87432 6 оніну в положенні +1 і закінчується лізином у положенні +357, як показано на Фіг.7, або яка починається з аланіну в положенні -1 та закінчується лізином у положенні +357, як показано на Фіг.7. 57. Молекула нуклеїнової кислоти, що кодує молекулу мутантного CTLA4 за п.56. 58. Ділянка молекули мутантного CTLA4, котра (молекула) кодується нуклеотидною молекулою, депонованою як АТСС № РТА-2104, яка (ділянка) відрізняється тим, що вона містить повну позаклітинну область мутантного CTLA4, що зв'язується з CD80 і/або CD86. 59. Ділянка молекули мутантного CTLA4 за п.58, яка відрізняється тим, що додатково містить Igполовину. 60. Молекула нуклеїнової кислоти, яка містить ділянку молекули нуклеїнової кислоти, що кодує молекулу мутантного CTLA4 і має АТСС № РТА2104, яка відрізняється тим, що вона кодує повну позаклітинну область молекули мутантного CTLA4, що зв'язується з CD80 і/або CD86. 61. Молекула нуклеїнової кислоти за п.60, яка відрізняється тим, що додатково містить нуклеотидну молекулу, що кодує Ig-половину. 62. Фармацевтична композиція для лікування захворювання імунної системи, яка містить фармацевтично прийнятний носій і молекулу мутантного CTLA4 за п.1 або 17. 63. Фармацевтична композиція для лікування захворювання імунної системи, яка містить фармацевтично прийнятний носій і молекулу мутантного CTLA4 за п.9. 64. Молекула мутантного CTLA4 за п.17, яка відрізняється тим, що амінокислотна послідовність, яка змінює розчинність, афінність або валентність, містить і ділянку імуноглобуліну, і тим, що ділянка імуноглобуліну може включати один або більше мутацій для зниження ефекторної функції. 65. Спосіб регулювання взаємодії Т-клітини з CD80- і/або СD86-позитивною клітиною, при якому приводять у контакт CD80- і/або CD86-позитивну клітину з молекулою розчинного мутантного CTLA4 за п.56 таким чином, щоб утворювався комплекс: молекула мутантного CTLA4/CD80, або комплекс: молекула мутантного CTLA4/CD86, причому цей комплекс перешкоджає взаємодії між Тклітиною і CD80- і/або CD86-позитивною клітиною. 66. Спосіб за п.65, який відрізняється тим, що CD80- і/або СD86-позитивна клітина контактує з фрагментом або похідною молекули мутантного CTLA4. 67. Спосіб за п.65, який відрізняється тим, що CD80- і/або CD86-позитивною клітиною є клітина, що презентує антигени. 68. Спосіб за п.65, який відрізняється тим, що взаємодія CTLA4-позитивних Т-клітин із CD80- і/або CD86-позитивними клітинами інгібується. 69. Спосіб лікування захворювань імунної системи, опосередкованих взаємодією Т-клітин з CD80і/або CD86-позитивними клітинами, при якому пацієнту вводять молекулу мутантного CTLA4 за п.56 для регулювання Т-клітинної взаємодії з CD80і/або CD86-позитивними клітинами. 7 87432 8 70. Спосіб лікування захворювань імунної системи, опосередкованих взаємодією Т-клітин з CD80і/або CD86-позитивними клітинами, при якому пацієнту вводять молекулу розчинного мутантного CTLA4 за п.56 для регулювання Т-клітинної взаємодії з CD80- і/або CD86-позитивними клітинами. 71. Молекула мутантного CTLA4, яка зв'язується з CD80 і/або з CD86 і містить позаклітинну область CTLA4, як показано на Фіг.9, починається з аланіну в позиції -1 або метіоніну в позиції +1 та закінчується аспарагіновою кислотою в позиції +124, або її частиною, яка відрізняється тим, що в позаклітинній області або його частині, аланін у положенні 29 замінений на тирозин, та лейцин у положенні 104 замінений, та також містить амінокислотну послідовність, яка змінює розчинність, афінність або валентність молекули мутантного CTLA4. 72. Молекула мутантного CTLA4 за п.71, яка відрізняється тим, що амінокислотна послідовність містить постійну ділянку людського імуноглобуліну, або його частину, де постійна ділянка імуноглобуліну може включати одну або більше мутацій для зниження ефекторної функції. 73. Молекула мутантного CTLA4, що містить амінокислотну послідовність, що є зв'язаною з метіоніном у положенні +1 і закінчується аспарагіновою кислотою в положенні +124, як показано на Фіг.7, або її частину, яка зв'язується з CD80 і/або з CD86, або містить амінокислотну послідовність, що є зв'язаною з аланіном у положенні -1 і закінчується аспарагіновою кислотою в положенні 124, як показано на Фіг.7, або її частину, яка зв'язується з CD80 і/або з CD86. 74. Молекула мутантного CTLA4 за п.73, яка відрізняється тим, що амінокислотна послідовність містить постійну ділянку людського імуноглобуліну, або його частину, де постійна ділянка імуноглобуліну може включати одну або більше мутацій для зниження ефекторної функції. 75. Молекула мутантного CTLA4 за пп.1, 9, 14, 15, 35, 54, 56, 72 або 74, що має меншу швидкість дисоціації при зв'язуванні з CD86, ніж CTLA4 дикого типу. 76. Молекула мутантного CTLA4, що має менші швидкості асоціації і дисоціації при зв'язуванні з CD80 і/або CD86, ніж CTLA4 дикого типу за пп.1, 9, 14, 15, 35, 54, 56, 72 або 74, що є розчинною. Даний винахідстосується розчинного CTLA4, який є мутантним варіантом CTLA4 дикого типу, що зберігає спроможність зв'язувати CD80 і/або CD86. Антиген-неспецифічні міжклітинні взаємодії між Т-лімфоцитами і клітинами, що презентують антигени (АРС), викликають Т-клітинні сигнали, що співстимулюють, які індукують Т-клітинну відповідь на антиген [Jenkins and Johnson (1993) Curr. Opin. Immunol. 5: 361-367]. Сигнали, що співстимулюють, визначають величину Т-клітинної відповіді на антиген і те, активує чи інактивує ця відповідь наступні реакції на антиген [Mueller et al. (1989) Annu. Rev. Immunol. 7: 445-480]. Активація Т-клітин у відсутність співстимуляції приводить до припинення Т-клітинної відповіді або до анергічної Т-клітинної відповіді [Schwartz, R. Η. (1992) Cell 71: 1065-1068]. Один ключовий сигнал, що співстимулює, утворюється при взаємодії рецептора Т-клітинної поверхні CD28 із родинними В7 молекулами на клітинах, що презентують антигени, (відомими також, наприклад, як В7-1 і В7-2 або CD80 і CD86, відповідно) [Р. Linsley and J. Ledbetter (1993) Annu. Rev. Immunol. 11: 191-212]. Молекула, відома сьогодні як CD80 (В7-1), була спочатку описана як активаційний антиген, пов'язаний із людськими В-клітинами [Yokochi, Т. et al. (1381) J. Immunol 128.823-827; Freeman, G. J. et al. (1989) J. Immunol. 143: 2714-2722], і який, відповідно, ідентифікується як протирецептор у відношенні до родинних Т-клітинних молекул CD28 і CTLA4 [Linsley, P., et al. (1990) Proc. Natl. Acad. Sci. USA 87: 5031-5035; Linsley, P.S. et al., (1991a) J. Exp. Med. 173:721-730; Linsley, P.S. et al. (1991b) J. Exp. Med. 174: 561-570]. Пізніше на клітинах, що презентують антигени, був ідентифікований інший протирецептор для CTLA4 [Azuma, N. et al. (1993) Nature 366: 76-79; Freeman (1993a) Science 262: 909-911; Freeman. G. J. et al. (1993b) J. Exp. Med. 178: 2185-2192: Hathcock, K.L.S., et al. (1994) J. Exp. Med. 180: 631640; Lenschow, D. J. et al., (1993) Proc. Natl. Acad. Sci. USA 90: 11054-11058: Ravi-Wolf, Z. et al. (1993) Proc. Natl. Acad. Sci. USA 90: 11182-11186; Wu, Y., et al., (1993) J. Exp. Med. 178: 1789-1793]. Ця молекула, відома сьогодні як CD86 [Caux, С., et al. (1994) J. Exp. Med. 180: 1841-1848], але що зветься також В7-0 [Azuma et al. (1993, див. вище] або В7-2 [Freeman et al. (1993a), див. вище], має послідовність, приблизно, на 25% ідентичну послідовності CD80 на її позаклітинній ділянці [Azuma et al. (1993), див. вище; Freeman et al. (1993a), див. вище, (1993b), див. вище]. Клітини, що трансфікуються молекулами CD86, ініціюють опосередковані CD28 Т-клітинні реакції [Azuma et al. (1993), див. вище; Freeman et al. (1993a), (1993b), див. вище]. Порівняння експресії CD80 і CD86 було предметом декількох досліджень [Azuma et al. (1993), див. вище; Hathcock, et al. (1994), див. вище; Larsen, СР., et al. (1994) J, Immunol. 152: 52085219: Stack, R. M., et al., (1994) J. Immunol. 152: 5723-5733]. Сучасні дані показують, що експресія CD80 і CD86 регулюється по-різному, і наводять на думку, що експресія CD86 має тенденцію у процесі імунної відповіді передувати експресії CD80. Розчинні форми CD28 і CTLA4 були створені злиттям варіабельних (v)-подібних позаклітинних областей CD28 і CTLA4 із постійними областями імуноглобуліну (Іg), що дає в результаті CD28lg і CTLA4lg. CTLA4lg зв'язує як СD80-позитивні, так і 9 СD86-позитивні клітини сильніше, ніж CD28lg [Linsley, P., et al. (1994) Immunity 1: 793-80]. Багато серед залежних від Т-клітин імунних реакцій блокуються CTLA4lg як in vitro, так і in vivo [Linsley, et al., (1991b), див. вище; Linsley, P.S. et al. (1992a) Science 257: 792-795; Linsley, Ρ. S. et al. (1992b) J Exp.Med. 176: 1595-1604; Lenschow, D. J. et al., (1992), Science 257: 789-792; Tan, P. et al., (1992) J. Exp. Med. 177: 165-173; Turka, L Α., (1992) Proc. Natt. Acad. Sci. USA 89: 11102-11105]. Пітч зі співробітниками [Peach, et al, J. Exp. Med. (1994) 180: 2049-2058] ідентифікували ділянки позаклітинної області CTLA4, що є важливими для міцного зв'язування з CD80. Конкретно, гексапептидний мотив (MYPPPY) на ділянці, подібній гіперваріабелльній ділянці 3 (CDR3), був ідентифікований як цілком консервативна область в усіх членів сімейства CD28 і CTLA4. Мутагенез із скануванням аланіну за допомогою мотиву MYPPPY у CTLA4 і в обраних залишках CD28lg послабляє або припиняє зв'язування з CD80. Були також створені химерні молекули, що взаємодіють із гомологічними ділянками CTLA4 і CD28. Молекули HS4, HS4-A і HS4-B були сконструйовані "пересаджуванням" СDР3-подібних ділянок CTLA4, що містили також частину карбоксильного кінця, продовжену так, щоб включати певні неконсервативні амінокислотні залишки на CD28lg. Ці гомологічні мутанти проявили більш високу авідність зв'язування з CD80, ніж CD28lg. В іншій групі химерних гомологічних мутантів CDRI-подібну ділянку CTLA4, що не є консервативною в CD28 і, як прогнозувалося, просторово прилягає до CDR3-подібної ділянки, "пересаджували" до HS4 і HS4-A. Ці химерні гомологічні мутантні молекули (позначені як HS7 і HS8) продемонстрували навіть більш високу авідність зв'язування з CD80, ніж CD28lg. Химерні гомологічні мутантні молекули були також отримані шляхом пересаджування до HS7 і HS8 CDR2-nofli6HOi ділянки CTLA4, але ця комбінація не дає додаткового поліпшення авідності зв'язування з CD80. Отже, мотив MYPPPY CTLA4 і CD28 був визначений як критичний для зв'язування з CD80, але деякі неконсервативні амінокислотні залишки CDR1- і CDR3-подібних ділянок були також відповідальними за підвищену авідність зв'язування CTLA4 із CD80. Було показано, що CTLA4lg ефективно блокує обумовлену CD80 Т-клітинну додаткову стимуляцію (співстимуляцію), але не є ефективним щодо блокади CD86-обумовлених імунних реакцій. Були створені молекули розчинного мутантного CTLA4, зокрема такого, що має більш високу авідність до CD86, ніж CTLA4 дикого типу, які, можливо, більш спроможні блокувати примування антигенспецифічних активованих клітин, ніж CTLA4lg. Залишається необхідність у більш досконалих молекулах CTLA4 для створення поліпшених фармацевтичних композицій для супресії імунної відповіді і для терапії раку, ніж відомі до тих пір розчинні форми CTLA4. Відповідно до вищевикладеного винаходом пропонується розчинний мутантний CTLA4, молекули якого зв'язуються з CD80 і/або CD86. До чис 87432 10 ла мутантних молекул згідно з даним винаходом належать такі молекули, що можуть розпізнавати CD80 і CD86 як поодинці, так і обидва, і зв'язуватися або з CD80 або CD86, або з ними обома. У деяких варіантах здійснення даного винаходу запропоновані мутантні молекули зв'язують CD80 і/або CD86 із більш високою авідністю, ніж CTLA4. Одним із прикладів мутантної молекули CTLA4 є описана тут L104EA29Ylg (Фіг.7). Іншим прикладом мутантної молекули CTLA4 є L104Elg (Фіг.8), також подана в даному описі. Молекули L104EA29Ylg і L104Elg зв'язуються з CD80 і CD86 із більш високою авідністю, ніж CTLA4lg. На Фіг.1 наведені результати аналізу рівноважного зв'язування L104EA29Ylg, L104Elg і CTLA4lg дикого типу з CD86lg. На Фіг.2А і 2В наведені результати FACSаналізу (FACS: клітинний сортер зі збудженням флуоресценції), що ілюструє зв'язування L104EA29Ylg, L104Elg і CTLA4lg із людськими CD80- і СD86-трансфікованими СНО клітинами, як описано в Прикладі 2, див. нижче. На Фіг.3А і 3В показане інгібування проліферації СD80-позитивних і CD86-позитивних СНОклітин, як описано нижче, в Прикладі 2. На Фіг.4А і 4В продемонстровано, що L104EA29Ylg більш ефективно інгібує проліферацію первинних і вторинних алостимульованих Тклітин, ніж CTLA4lg, як описано нижче, в Прикладі 2. На Фіг.5А-С показано, що L104EA29Ylg більш ефективно, ніж CTLA4lg, інгібує продукування цитокінів IL-2 (Фіг.5А), IL-4 (Фіг.5В) і g-інтерферону (Фіг.5С) алостимульованих людських Т-клітин, як описано нижче, у Прикладі 2. На Фіг.6 показано, що L104EA29Ylg більш ефективно, ніж CTLA4lg, інгібує проліферацію фітогематоглютинін(РНА)-стимульованих Т-клітин мавп, як описано нижче, у Прикладі 2. На Фіг.7 зображені нуклеотидна й амінокислотна послідовності мутантної молекули CTLA4 (L104EA29Ylg), що містять сигнальний пептид; мутантна позаклітинна область CTLA4, починаючи від метіоніну в положенні +1 і закінчуючи аспарагіновою кислотою в положенні +124, або починаючи від аланіну в положенні -1 і закінчуючи аспарагіновою кислотою в положенні +124; і ділянка Іg, як описано нижче, у Прикладі 1. На Фіг.8 зображені нуклеотидна й амінокислотна послідовності мутантної молекули CTLA4 (L104Elg), що містять сигнальний пептид; мутантна позаклітинна область CTLA4, починаючи від метіоніну в положенні +1 і закінчуючи аспарагіновою кислотою в положенні +124, або починаючи від аланіну в положенні -1 і закінчуючи аспарагіновою кислотою в положенні +124; і ділянка Іg, як описано нижче, у Прикладі 1. На Фіг.9 зображені нуклеотидна й амінокислотна послідовності CTLA4lg, що містять сигнальний пептид; амінокислотна послідовність позаклітинної області CTLA4 дикого типу, починаючи від метіоніну в положенні +1 і закінчуючи аспарагіновою кислотою в положенні +124, або починаючи від аланіну в положенні -1 і закінчуючи аспарагіновою кислотою в положенні +124; і ділянка Іg. 11 На Фіг.10А-С подані результати PAGE-аналізу в SDS-гелі (Фіг.10А) для CTLA4lg (доріжка 1), L104Elg (доріжка 2) і L104EA29Ylg (доріжка 3А) і результати ексклюзивної за розміром хроматографії для CTLA4lg (Фіг.10В) і L104EA29Ylg (Фіг.10С). На Фіг.11А і 11В зображена смугова діаграма V-подібної складки позаклітинного Іg CTLA4, отримана шляхом визначення структури за допомогою ЯМР-спектроскопії. На Фіг.11В поданий розгорнутий вигляд ділянки S25-R33 і ділянки MYPPPY, що демонструє положення та орієнтацію бічного ланцюга мутацій L104 і А29, що підвищують авідність. На Фіг.12 зображена схематична діаграма вектора piLN-LEF29Y із інсерцією L104EA29Ylg. Термінологія Нижче дані визначення застосованої у даному описі спеціальної термінології. Термін "CTLA4 дикого типу" стосується CTLA4, який має амінокислотну послідовність природного повномірного CTLA4 (Патенти США №№5434131,5844095, 5851795), або його позаклітинної області, що зв'язує CD80 і/або CD86, і/або перешкоджає CD80 і/або CD86 зв'язуватися з їхніми лігандами. У конкретних варіантах здійснення винаходу позаклітинна область CTLA4 дикого типу починається з метіоніну в положенні+1 і закінчується аспарагіновою кислотою в положенні+124, або позаклітинна область CTLA4 дикого типу починається з аланіну в положенні -1 і закінчується аспарагіновою кислотою в положенні +124. CTLA4 дикого типу являє собою білок клітинної поверхні, що містить N-кінцеву позаклітинну область, трансмембранну область і С-кінцеву цитоплазматичну область. Позаклітинна область зв'язується з цільовими антигенами (мішенями), такими як CD80 і CD86. У природній клітині білок CTLA4 дикого типу транслюється у вигляді незрілого поліпептиду, що включає у себе сигнальний пептид на N-кінці. Незрілий поліпептид зазнає посттрансляційного процесінгу, який включає у себе відщеплення і видалення сигнального пептиду з утворенням продукту розщеплення CTLA4, котрий має новостворений N-кінець, що відрізняється від N-кінця незрілої форми. Для фахівця у даній галузі зрозуміло, що може відбуватися додатковий посттрансляційний процесінг, при якому із новоствореного N-кінця продукту розщеплення CTLA4 видаляється одна чи більше амінокислот. Молекула зрілої форми CTLA4 включає позаклітинну область CTLA4 або будь-яку її частину, що зв'язується з CD80 і/або CD86. "CTLA4lg" являє собою розчинний злитий білок, що містить позаклітинну область CTLA4 дикого типу або її частину, котра зв'язує CD80 і/або CD86, з'єднаної з кінцем Іg. Кращий варіант даного винаходу включає у себе позаклітинну область CTLA4 дикого типу, починаючи від метіоніну в положенні +1 і закінчуючи аспарагіновою кислотою в 87432 12 положенні +124; або починаючи від аланіну в положенні -1 і закінчуючи аспарагіновою кислотою в положенні +124; ділянку з'єднання - амінокислотний залишок глутамін у положенні +125; і імуноглобулінову ділянку, що охоплює ділянку від глутамінової кислоти в положенні +126 до лізину в положенні +357 (Фіг.9). Застосовуване в даному описі поняття "злитий білок" визначається як одна або більше амінокислотних послідовностей, з'єднаних між собою за допомогою добре відомих способів і способів, описаних у патентах США №5434131 або №5637481. При цьому з'єднані амінокислотні послідовності утворюють один злитий білок. Застосовуваний у· даному описі вираз "молекула мутантного CTLA4" ("мутантний CTLA4") може позначати молекулу повномірного CTLA4 (повномірний CTLA4) або її (його) частини (похідні або фрагменти), що містять мутацію або множинні мутації в CTLA4 (переважно у позаклітинній області CTLA4), так, що вона (він) стає аналогічною (аналогічним), але не ідентичною (ідентичним) молекулі CTLA4 дикого типу (CTLA4 дикого типу). Молекули мутантного CTLA4 зв'язуються або з CD80, або з CD86 або з ними обома. Молекули мутантного CTLA4 можуть містити в собі біологічно або хімічно активну не- CTLA4 молекулу, або ця молекула може бути приєднана до молекули мутантного CTLA4. Молекули мутантного з'єднання можуть бути розчинними (тобто такими, що циркулюють) або можуть бути пов'язані з поверхнею. Молекули мутантного CTLA4 можуть включати у себе всю позаклітинну область CTLA4 або її частини, наприклад, фрагменти або похідні. Мутантний CTLA4 можна одержувати шляхом синтезу або за методами рекомбінантних ДНК. Застосовуваний у даному описі термін "мутація" означає зміну в нуклеотидній або амінокислотній послідовності поліпептиду дикого типу. У даному випадку він означає зміну в позаклітинній області CTLA4 дикого типу. Такою зміною може бути зміна амінокислоти, що включає у себе заміни, делеції, додавання або усікання (скорочення). Мутантна молекула може містити одну або декілька мутацій. Мутації в нуклеотидній послідовності можуть викликати або не викликати мутації в амінокислотній послідовності, що добре зрозуміло з рівня техніки. Так, деякі нуклеотидні кодони кодують одну й ту саму амінокислоту. Прикладами тут можуть служити кодони CGU, CGG, CGC і CGA, що кодують амінокислоту, аргінін (R); або кодони GAU і GAC, що кодують амінокислоту, аспарагінову кислоту (D). Отже, білок може кодуватися однією і більше нуклеїновими кислотами, що відрізняються специфічними нуклеотидними послідовностями, проте кодують білки з ідентичними послідовностями. Нижче перелічені послідовності, що кодують амінокислоти. 13 Амінокислота Аланін Цистеїн Аспарагінова к-та Глутамінова к-та Фенілаланін Гліцин Гістидин Ізолейцин Лізин Лейцин Метіонін Аспарагін Пролін Глутамін Аргінін Серин Треонін Валін Триптофан Тирозин 87432 Символ Ala Cys Asp Glu Phe Gly His lle Lys Leu Met Asn Pro Gin Arg Ser Thr Val Trp Туr Однолітерний символ A С D Ε F G Η 1 К L Μ Ν Ρ Q R S Τ V W Υ Уживаний у даному описі термін "позаклітинна область CTLA4" означає частину CTLA4, що розпізнає CD80 і/або CD86 і зв'язуються з ними. Наприклад, позаклітинна область CTLA4 включає у себе ділянку від метіоніну вположенні +1 до аспарагінової кислоти в положенні +124 (Фіг.9). Або ж позаклітинна область CTLA4 включає у себе ділянку від аланіну в положенні -1 до аспарагінової кислоти в положенні +124 (Фіг.9). Позаклітинна область включає у себе фрагменти або похідні CTLA4, що зв'язуються з CD80 і/або CD86. Уживаний у даному описі термін "не-СТLА4 білкова послідовність" або "не-СТLА4 молекула" означає будь-яку молекулу, що не зв'язується з CD80 і/або CD86 і не перешкоджає зв'язуванню CTLA4 з її ціллю. Як приклад, що не обмежує, тут можна навести постійну ділянку імуноглобуліну (Іg) або її частину. У кращому варіанті постійна ділянка імуноглобуліну являє собою постійну ділянку людського Іg або Іg мавпи, наприклад, людський С(гама) 1, включаючи шарнірну ділянку, ділянки СН2 і СН3. Постійна ділянка Іg може мутувати, послаблюючи його ефекторні функції (Патенти США №№5637481 і 6132992). Застосовуваний у даному описі вираз "фрагмент мутантної молекули CTLA4" означає частину мутантної молекули CTLA4, переважно позаклітинну область CTLA4 або її частину, що розпізнає мішень, наприклад, CD80 і/або CD86, і зв'язується з нею. Застосовуваний у даному описі вираз "похідна мутантного CTLA4" ("похідна молекули мутантного CTLA4") означає молекулу, що має послідовність, принаймні на 70% подібну послідовності позаклітинної області CTLA4, і функції, аналогічні функціям цієї області, тобто вона розпізнає CD80 і/або CD86 і зв'язується з ними. Застосовуваний у даному описі вираз "частина (ділянка) молекули CTLA4" включає фрагменти і похідні (молекули) CTLA4, що зв'язуються з CD80 і/або CD86. 14 Кодони GCU, GCC, GCA, GCG UGU, UGC GAU, GAC GAA, GAG UUU, UUC GGU, GGC, GGA, GGG CAU, CAC AUU, AUC, AUA AAA, AAG UUA, UUG, CUU, CUC, CUA, CUG AUG AAU, AAC CCU, CCC, CCA, CCG CAA, CAG CGU, CGC, CGA, CGG, AGA, AGG UCU, UCC, UCA, UCG, AGU, AGC ACU, ACC, ACA, ACG GUU, GUC, GUA, GUG UGG UAU, UAC Нижче даний винахід розглядається більш докладно. Композиції згідно з даним винаходом Даним винаходом пропонується розчинний мутантний CTLA4, що розпізнає CD80 і/або CD86 і зв'язується з ними. У деяких варіантах здійснення винаходу розчинні мутантні CTLA4 виявляють більш високу авідність до CD80 і/або CD86, ніж CTLA4lg. Одним із прикладів мутантного CTLA4 є L104EA29Ylg (Фіг.7). Амінокислотна послідовність L104EA29Ylg може починатися аланіном в положенні -1 амінокислоти і закінчуватися лізином у положенні +357 амінокислоти. Або ж амінокислотна послідовність L104EA29YIg може починатися метіоніном у положенні +1 амінокислоти і закінчуватися лізином у положенні +357 амінокислоти. СТLА4-ділянка L104EA29Ylg охоплює метіонін у положенні +1 амінокислоти аж до аспарагінової кислоти в положенні +124 амінокислоти. L104EA29Ylg містить ділянку з'єднання - амінокислотний залишок глутамін у положенні +125 - і ділянку імуноглобуліну, що охоплює ділянку від глутамінової кислоти в положенні +126 до лізину в положенні +357 (Фіг.7). Авідність L104EA29Ylg при зв'язуванні з CD80 є приблизно удвічі вищою за авідність CTLA4lg дикого типу (що далі зветься CTLA4lg) і приблизно в 4 рази вищою при зв'язуванні з CD86. Таке більш сильне зв'язування приводить до того, що L104EA29Ylg більш ефективно блокує імунні реакції, ніж CTLA4lg. Молекули мутантного CTLA4 містять, щонайменше, позаклітинну область CTLA4 або її ділянку, що зв'язується з CD80 і/або CD86. Позаклітинна область молекули мутантного CTLA4 містить амінокислотну послідовність, починаючи від метіоніну в положенні +1 до аспарагінової кислоти в положенні +124 (Фіг.7 або 8). Або ж позаклітинна область CTLA4 може включати амінокислотну послідовність, починаючи від аланіну в положенні -1 і закінчуючи аспарагіновою кислотою в положенні +124 (Фіг.7 або 8). 15 В однім з варіантів здійснення винаходу розчинний мутантний CTLA4 являє собою злитий білок, що містить позаклітинну область CTLA4, котра має одну чи більше мутацій на ділянці амінокислотної послідовності, починаючи серином в положенні +25 і закінчуючи аргініном у положенні +33 (S25-R33). Наприклад, аланін у положенні +29 CTLA4 дикого типу можна замінити на тирозин (кодони: UAU, UAC). Або ж аланін можна замінити на лейцин (кодони: UUA, UUG, CUU, CUC, CUA, CUG), фенілаланін (кодони: UUU, UUC), триптофан (кодон: UGG) або треонін (кодони: ACU, АСС, АСА, ACG). Як легко зрозуміють фахівці у даній галузі, урацил (U), нуклеотид послідовності РНК, відповідає тиміну (Т), нуклеотиду послідовності ДНК. В іншому варіанті здійснення винаходу розчинний мутантний CTLA4 являє собою злитий білок, що містить позаклітинну область CTLA4, котра має одну й більше мутацій на ділянці або біля ділянки амінокислотної послідовності, що починається метіоніном в положенні +97 і закінчується гліцином у положенні +107 (M97-G107). Наприклад, лейцин у положенні +104 CTLA4 дикого типу можна замінити на глутамінову кислоту (кодони GAA, GAG) Мутантний CTLA4, що містить таку заміну, називається в даному описі L104Elg (Фіг.8) Ще в одному варіанті здійснення даного винаходу розчинний мутантний CTLA4 являє собою злитий білок, що має позаклітинну область CTLA4, котра містить одну і більше мутацій на ділянках S25-R33 і M97-G107. Наприклад, згідно з одним із варіантів винаходу мутантний CTLA4 містить тирозин у положенні +29 замість аланіну; і глутамінову кислоту в положенні +104 замість лейцину. Мутантний CTLA4, що містить таку заміну, фігурує в даному описі під назвою L104EA29Ylg (Фіг.7). Нуклеїнова кислота, що кодує L104EA29Ylg, міститься в pD16 L104EA29Ylg і депонована 19 червня 2000 року Американською Колекцією Культур Клітин (АТСС), 10801 University Blvd., Manasas, VA 201102209 (АТСС No. PTA-2104). Вектор pD16 L104EA29Ylg є похідним від вектора pcDNA3 (INVITROGEN). Винаходом охоплюється також розчинний мутантний CTLA4, що містить позаклітинну область CTLA4, як показано на Фіг.7 або 8, або його частину (частини), і фрагмент, що змінює розчинність, афінність і/або валентність мутантного CTLA4. Практика застосування даного винаходу показує, що зазначений фрагмент може являти собою постійну ділянку імуноглобуліну або її частину. Для застосування in vivo краще, якщо постійна ділянка імуноглобуліну не викликає шкідливої імунної відповіді у пацієнта. Наприклад, бажано, щоб у клінічних протоколах молекули мутанта включали постійні ділянки людського або мавпячого імуноглобуліну. Одним із прикладів підхожої ділянки імуноглобуліну є людський С(гама)1, що містить шарнірну ділянку, СН2 і СН3 ділянки. Можливі також інші ізотипи. Крім того, можливі й інші постійні ділянки (бажано, щоб це були слабкоімуногенні або неімуногенні постійні ділянки імуноглобуліну). Фрагментами можуть бути також поліпептидні мітки. Підхожими мітками можуть бути, наприклад, 87432 16 молекули р97, env gp120, E7 і ova [Dash, В., et al. (1994) J. Gen. Virol. 75: 1389-97: Ikeda, Т., et al. (1994) Gene 138:193-6; Falk, K., et al. (1993) Cell. Immunol. 150: 447-52: Fujisaka, K., et al. (1994) Virology 204: 789-93]. Як мітки можуть бути застосовані також інші молекули [Gerard, С, et al. (1994) Neuroscience 62: 721-739; Byrn, R., et al., (1989) J. Virol 63: 4370-4375; Smith, D. et al. (1987) Science 238:1704-1707; Lasky, L, (1996) Scieuse 233: 209212]. Крім того, винаходом охоплюються розчинні мутантні злиті білки CTLA4lg, переважно, більш реактогенні щодо антигена CD80 і/або CD86 порівняно з CTLA4 дикого типу. Одним із прикладів таких білків є L104EA29Ylg, показаний на Фіг.7. В іншому варіанті здійснення даного винаходу молекула розчинного мутантного білка CTLA4 включає у себе з'єднувальний амінокислотний залишок, розташований між ділянкою CTLA4 і ділянкою імуноглобуліну. Ділянка з'єднання амінокислот може бути будь-якою амінокислотою, що містить глутамін. З'єднувальна амінокислота (ділянка з'єднання) може вводитися за методами молекулярної генетики або за допомогою відомих методів хімічного синтезу. Ще в одному варіанті здійснення винаходу молекула розчинного мутантного білка CTLA4 включає ділянку імуноглобуліну (наприклад, шарнірну ділянку, СН2 і СН3 ділянки), де будь-який цистеїновий залишок або всі цистеїнові залишки в шарнірній області імуноглобулінової ділянки замінені на серин, як, наприклад, цистеїнові залишки в положеннях +130, +136 або +139 (Фіг.7 або 8). Мутантні молекули можуть також включати у себе пролін у положенні +148, замінений на серин, як показано на Фіг.7 або 8. Молекула розчинного мутантного білка CTLA4 може містити послідовність сигнального пептиду, пов'язану з N-кінцем позаклітинної області СТLА4частини мутантної молекули. Сигнальний пептид може являти собою будь-яку послідовність, що дозволяє мутантній молекулі секретувати; ним може бути, наприклад, сигнальний пептид онкостатину Μ [Malik, et al., (1989) Molec. Cell. Biol. 9: 2847-2853] або CD5 [Jones, Ν. Η. et al., (1986) Nature 323: 346-349] або сигнальний пептид будьякого позаклітинного білка. Молекула мутантної сполуки може включати у себе сигнальний пептид онкостатину М, пов'язаний із N-кінцем позаклітинної області CTLA4, і молекулу людського імуноглобуліну (наприклад, шарнірну ділянку, СН2 і СН3 ділянки), пов'язану із Скінцем позаклітинної області CTLA4. Ця молекула містить сигнальний пептид онкостатину М, що охоплює амінокислотну послідовність, починаючи від метіоніну в положенні -26 і закінчуючи аланіном в положенні -1, ділянку CTLA4, що охоплює амінокислотну послідовність від метіоніну в положенні +1 до аспарагінової кислоти в положенні +124, ділянку з'єднання - амінокислотний залишок глутамін у положенні +125 і ділянку імуноглобуліну, що охоплює амінокислотну послідовність від глутамінової кислоти в положенні +126 до лізину в положенні +357. 17 Розчинний мутантний білок CTLA4 можна одержувати за молекулярними методами рекомбінантних ДНК або за хімічними методами синтезу. Молекулярні методи включають такі стадії: інтродукцію підхожої клітини-хазяїна з молекулою нуклеїнової кислоти, що експресує і кодує молекулу розчинного мутантного CTLA4; культивування інтродукованої у такий спосіб клітини-хазяїна в умовах, що дозволяють клітині-хазяїну експресувати мутантні молекули; і виділення мутантного білка, що експресує. Ділянка сигнального пептиду мутантної молекули робить можливою експресію білка на поверхні клітини і секрецію його клітиноюхазяїном. Мутантні молекули, що транслюються, можуть піддаватися посттрансляційній модифікації, котра включає у себе відщеплення сигнального пептиду з утворенням зрілого білка, який містить ділянки CTLA4 й імуноглобуліну. Відщеплення може здійснюватися після аланіну в положенні -1, що дає молекулу зрілого мутантного білка, котра містить метіонін у положенні +1 які першу амінокислоту (Фіг.7 або 8). Або ж відщеплення може відбуватися після метіоніну в положенні -2, що дає молекулу зрілого мутантного білка, котра як першу амінокислоту містить аланін у положенні -1. Кращим варіантом здійснення винаходу є молекула розчинного мутантного білка CTLA4, що містить позаклітинну область людського CTLA4, пов'язану з цілою молекулою імуноглобуліну або її частиною (наприклад, шарнірною ділянкою, СН2 і СН3 ділянками). Молекула такого білка містить СТЦ\4-ділянку молекули розчинного білка, що охоплює амінокислотну послідовність від метіоніну в положенні +1 до аспарагінової кислоти в положенні +124, ділянку з'єднання - амінокислотний залишок глутамін у положенні +125 і ділянку імуноглобуліну, що охоплює амінокислотну послідовність від глутамінової кислоти в положенні +126 до лізину в положенні +357. Частина, що містить позаклітинну область CTLA4, мутує так, що аланін у положенні +29 заміняється на тирозин, а лейцин у положенні +104 заміняється на глутамінову кислоту. Імуноглобулінова частина мутантної молекули може мутувати так, що цистеїнові залишки в положеннях +130, +136 і +139 заміняються на серин, а пролін у положенні +148 також заміняється на серин. Цей мутантний білок (мутантна молекула) позначена в даному описі як L104EA29Ylg (Фіг.7). Іншим варіантом мутантної молекули L104EA29Ylg згідно з даним винаходом є мутантна молекула, що має амінокислотну послідовність від аланіну в положенні -1 до аспарагінової кислоти в положенні +124, ділянку з'єднання амінокислот глутаміновий залишок - у положенні +125 і ділянку імуноглобуліну, що охоплює глутамінову кислоту в положенні +126 (наприклад, від +126 до лізину в положенні +357). Ділянка, що містить позаклітинну область CTLA4, мутує так, що аланін у положенні +29 заміняється на тирозин, а лейцин у положенні +104 заміняється на глутамінову кислоту. Імуноглобулінова частина мутантної молекули мутує так, що цистеїнові залишки в положеннях +130, +136 і +139 заміняються на серин, а пролін у положенні +148 також заміняється на серин. Цей мутантний білок (мутантна молекула) позначена в 87432 18 даному описі як L104EA29Ylg (Фіг.7). Після відщеплення сигнальної послідовності L104EA29Ylg може починатися з метіоніну в положенні +1 або з аланіну в положенні -1. Іншою мутантною молекулою за даним винаходом є молекула розчинного мутантного білка CTLA4, що має позаклітинну область людського CTLA4, пов'язану з молекулою людського імуноглобуліну (наприклад, шарнірною ділянкою, СН2 і СН3). Ця молекула включає у себе частину амінокислотної послідовності, що кодує CTLA4, починаючи від метіоніну в положенні +1 і закінчуючи аспарагіновою кислотою в положенні +124, ділянку з'єднання амінокислот - глутаміновий залишок - у положенні +125 і ділянку імуноглобуліну, що охоплює амінокислотну послідовність від глутамінової кислоти в положенні +126 до лізину в положенні +357. Ділянка, що містить позаклітинну область CTLA4, мутує так, що лейцин у положенні +104 заміняється на глутамінову кислоту. Шарнірна ділянка молекули мутує так, що цистеїнові залишки в положеннях +130, +136 і +139 заміняються на серин, а пролін у положенні +148 також заміняється на серин. Цей мутантний білок (мутантна молекула) позначений у даному описі як L104Elg (Фіг.8). Альтернативним варіантом L104Elg є молекула розчинного мутантного CTLA4, що містить позаклітинну область CTLA4 пов'язану з молекулою людського імуноглобуліну (наприклад, шарнірною ділянкою, СН2 і СН3). Ця молекула включає у себе частину CTLA4, що охоплює амінокислотну послідовність від аланіну в положенні -1 до аспарагінової кислоти в положенні +124, ділянку з'єднання амінокислотний залишок глутамін у положенні +125 і ділянку імуноглобуліну, що охоплює область від глутамінової кислоти в положенні +126 до лізину в положенні +357. Ділянка, що містить позаклітинну область CTLA4, мутує так, що лейцин у положенні +104 заміняється на глутамінову кислоту. Шарнірна ділянка мутантної молекули мутує так, що цистеїнові залишки в положеннях +130, +136 і +139 заміняються на серин, а пролін у положенні+148 також заміняється на серин. Цей мутантний білок (мутантна молекула) позначений у даному описі як L104Elg (Фіг.8). Крім того, даним винаходом охоплюється молекула розчинного мутантного CTLA4, що має: (а) першу амінокислотну послідовність мембранного глікопротеїну, наприклад, CD28, CD86, CD80, CD40 і gр39, що блокує проліферацію Т-клітин, злиту з другою амінокислотною послідовністю; (Ь) другу амінокислотну послідовність, що є фрагментом позаклітинної області мутантного CTLA4, що блокує проліферацію Т-клітин, таку як, наприклад, молекула амінокислоти, що включає у себе ділянку від метіоніну в положенні+1 до аспарагінової кислоти в положенні +124 (Фіг.7 або 8); і (с) третю амінокислотну послідовність, що діє як ідентифікаційна мітка або підвищує розчинність молекули. Наприклад, третя амінокислотна послідовність може складатися виключно із амінокислотних залишків шарнірної ділянки, СН2 і СН3-ділянок молекули неімуногенного імуноглобуліну. Підхожими молекулами імуноглобуліну можуть бути, наприклад, людський або мавпячий імуноглобулін, на 19 приклад, С(гама)1. Можливими є також інші ізотипи. Крім того, винаходом охоплюються нуклеотидні молекули, котрі мають нуклеотидні послідовності, що кодують амінокислотні послідовності, які відповідають молекулам запропонованого розчинного мутантного CTLA4. В одному з варіантів здійснення винаходу нуклеотидна молекула являє собою ДНК (наприклад, кДНК) або її гібрид. Нуклеотидними молекулами можуть бути також РНК або їхні гібриди. Крім того, даний винахід включає у себе вектор, що містить запропоновані нуклеотидні послідовності. Винаходом охоплюється також система вектора-хазяїна. Система вектора-хазяїна включає у себе вектор за даним винаходом у підхожій клітині-хазяїні. Підхожими клітинами-хазяїнами можуть бути, наприклад, клітини прокаріотів і еукаріотів. Винаходом охоплюються фармацевтичні композиції для лікування захворювань імунної системи, що містять фармацевтично ефективні кількості розчинного мутантного CTLA4. У деяких варіантах винаходу захворювання імунної системи опосередковуються взаємодією CD28- і/або СТLA4позитивних клітин із CD80- і/або СD86позитивними клітинами. Молекули розчинного мутантного CTLA4, переважно, являють собою молекули CTLA4, що містять одну чи більше мутацій у позаклітинній області CTLA4. Фармацевтична композиція може містити розчинний мутантний білок CTLA4 і/або нуклеїнову кислоту і/або вектори, що їх кодують. У кращих варіантах здійснення винаходу розчинний мутантний CTLA4 має амінокислотну послідовність позаклітинної області CTLA4, як показано на Фіг.7 або на Фіг.8 (L104EA29Y або L104E, відповідно). У ще кращому варіанті розчинним мутантним CTLA4 є L104EA29Ylg, поданий у даному описі. Композиції можуть містити також інші терапевтичні агенти, включаючи без обмеження, лікарські токсини, ферменти, антитіла або кон'югати. Фармацевтичні композиції також можуть включати у себе підхожі носії або ад'юванти, що містять будь-який матеріал, який при з'єднанні з молекулою за даним винаходом (наприклад, розчинним мутантним CTLA4, таким як L104EA29Y або L104E) зберігає активність молекули і не є реактивним по відношенню до імунної системи пацієнта. Підхожими носіями і ад'ювантами можуть бути, але без обмеження лише ними: людський сироватковий альбумін; іонообмінні речовини; оксид алюмінію; лецитин; буфери, такі як фосфати; гліцин; сорбінова кислота; сорбат калію; і солі або електроліти, такі як протамінсульфат. У числі інших прикладів можна назвати будь-які стандартні фармацевтичні носії, такі як фізіологічний розчин із фосфатним буфером; вода; емульсії, наприклад, емульсія олії у воді; і різноманітні зволожуючі засоби. Носіями можуть бути також стерильні розчини; таблетки, включаючи таблетки, покриті оболонкою, і капсули. Зазвичай такі носії містять ексципієнти, такі як крохмаль, молоко, цукор, деякі види глин, желатинів, стеаринову кислоту або її солі, стеарат магнію або кальцію, тальк, рослинні 87432 20 жири або олії, смоли, гліколі або інші відомі ексципієнти. Такі носії можуть містити також смакові добавки і барвники або інші інгредієнти. Композиції з такими носіями .готують у добре відомі способи. Такі композиції можна також поміщати в різноманітні ліпідні середовища, такі як ліпосоми, або у різні полімерні середовища, наприклад, полімерні мікрсфери. Фармацевтичні композиції за даним винаходом можна вводити пацієнтам у звичайні способи, включаючи без обмеження внутрішньовенний (вв), внутрішньочеревинний (вч), внутрішньом'язовий (вм), підшкірний, пероральний шляхи, супозиторний спосіб або місцевий контакт, або ж імплантацію пацієнту медикаментозно-постачального пристрою пролонгованої дії, такого як мініосмотичний насос. Фармацевтичні композиції згідно з даним винаходом можуть мати вигляд різноманітних лікарських форм, які містять без обмеження рідкі розчини або суспензії, таблетки, пілюлі, порошки, супозиторії, полімерні мікрокапсули або мікровезикули, ліпосоми і розчини для ін'єкцій або вливань. Краща форма залежить від способу введення і терапевтичного застосування. Найбільш ефективний спосіб застосування і схема приймання композицій за даним винаходом залежать від тяжкості і перебігу хвороби, стану здоров'я хворого і реакції на лікування, а також від рішення лікаря-куратора. Відповідно до цього, дозування композицій варто визначати для кожного пацієнта індивідуально. Молекули розчинного мутантного CTLA4 можна вводити пацієнту в кількості і в часі (наприклад, період і/або кількість прийомів), достатніми для того, щоб блокувати зв'язування ендогенних В7 (наприклад, CD80 і/або CD86) із Їхніми відповідними лігандами в організмі пацієнта. Тим самим блокада зв'язування ендогенного В7/ліганда інгібує взаємодію В7-позитивних клітин (наприклад, CD80- і/або СD86-позитивних клітин) із CD28-і/або СTLA4-позитивними клітинами. Дозування терапевтичного агента залежить від багатьох чинників, включаючи без обмеження характер тканини, в яку вводять агент, вид аутоімунного (аутоалергічного) захворювання, що підлягає лікуванню, тяжкість захворювання, стан здоров'я пацієнта і реакція пацієнта на лікування за допомогою агентів. Відповідно до цього, дозування агентів може змінюватися в залежності від пацієнта і способу введення. Молекули розчинного мутантного CTLA4 можна вводити в кількості 0,1-20,0мг/кг маси хворого на день, краще, якщо 0,5-10,0мг/кг/день. Вводити фармацевтичні композиції можна з різноманітними проміжками часу. В одному з варіантів фармацевтичну композицію за даним винаходом можна вводити протягом однієї години і більше. Крім того, введення композиції можна повторювати відповідно до тяжкості захворювання, а також інших чинників, добре відомих фахівцям. Винаходом охоплюються також способи одержання білка, що полягають у вирощуванні системи хазяїн-вектор згідно з винаходом таким чином, щоб продукувати білок у хазяїні, і у відділенні утвореного при цьому білка. 21 Крім того, винаходом охоплюються способи регулювання функціональної взаємодії CTLA4- і СD28-позитивних Т-клітин із CD80- і/або СD86позитивними клітинами. До цих способів належить приведення у контакт CD80- і/або СD86позитивних клітин із молекулою розчинного мутантного CTLA4 за винаходом таким чином, щоб утворилися комплекси мутантний CTLA4/CD80 і/або мутантний CTLA4/CD86, причому ці комплекси перешкоджають реакції антигена ендогенного CTLA4 із CD80 і/або CD86, і/або реакції антигена ендогенного CD28 із CD80 і/або CD86. В одному з варіантів здійснення винаходу молекула розчинного мутантного CTLA4 являє собою злитий білок, який містить принаймні частину позаклітинної області мутантного CTLA4. В іншому варіанті молекула розчинного мутантного CTLA4 містить: першу амінокислотну послідовність, що включає позаклітинну область CTLA4 від метіоніну в положенні +1 до аспарагінової кислоти в положенні +124, що містить принаймні одну мутацію; і другу амінокислотну послідовність, що включає шарнірну ділянку, СН2 і СН3-ділянки молекули людського імуноглобуліну гама 1 (Фіг.7 або 8). Відповідно до практики застосування даного винаходу CD80- і/або СD86-позитивні клітини приводять у контакт із фрагментами або похідними молекули запропонованого розчинного мутантного CTLA4. Або ж розчинний мутантний CTLA4 за даним винаходом використовується у формі злитого білка CD28lg/CTLA4lg, що має першу амінокислотну послідовність, яка відповідає частині позаклітинної області рецептора CD28 і злита з другою амінокислотною послідовністю, що відповідає частині позаклітинної області мутантного рецептора CTLA4, і третю амінокислотну послідовність, що відповідає шарнірній ділянці, СН2 і СН3-ділянкам молекули людського імуноглобуліну С-гамма-1. Передбачається, що розчинний мутантний CTLA4 виявляє інгібувальні властивості in vivo. В умовах, коли взаємодія Т-клітина/АРС-клітина, наприклад, взаємодія Т-клітина/В-клітина, відбувається в результаті контакту Т-клітин і АРС-клітин, зв'язування введених із метою реакції з CD80і/або СD86-позитивними клітинами, наприклад, із В-клітинами, молекул мутантного CTLA4 може перешкоджати (тобто інгібувати) взаємодії Тклітина/АРС-клітина, що викликає регуляцію імунних реакцій. Винахід охоплює способи негативної регуляції (інгібування) імунних реакцій. Негативна регуляція (модуляція) імунної відповіді молекулами розчинного мутантного CTLA4 може відбуватися шляхом інгібування або блокади імунної відповіді вже в процесі розвитку або може запобігати індукції імунної відповіді. Молекули розчинного CTLA4 за винаходом можуть інгібувати функції активованих Тклітин, такі як проліферація Т-лімфоцитів і секреція цитокінів, пригнічуючи Т-клітинні реакції або викликаючи специфічну толерантність у Тклітинах, або у той та інший спосіб одночасно. Крім того, даним винаходом охоплюються способи лікування захворювань імунної системи й індукції толерантності. В особливо прийнятних варіантах здійснення винаходу захворювання іму 87432 22 нної системи опосередковуються взаємодією CD28і/абр CTLA4-позитивних клітин із СD80/СD86-позитивними клітинами. В іншому варіанті винаходу Т-клітинна взаємодія інгібується. До числа вищезгаданих захворювань імунної системи входять, але без обмеження, аутоімунні захворювання, імунопроліферативні захворювання і захворювання, пов'язані з трансплантацією. Способи згідно з винаходом включають у себе уведення пацієнту молекул запропонованого розчинного мутантного CTLA4 з метою регуляції Тклітинної взаємодії з CD80- і/або СD86позитивними клітинами. Або ж можна вводити гібрид мутантного CTLA4, що містить мембранний глікопротеїн, з'єднаний із молекулою мутантного CTLA4. Серед захворювань, пов'язаних із трансплантацією, можна назвати гомологічну хворобу (GVHD) (наприклад, таку, що може явитися результатом пересадження кісткового мозку або при індукції толерантності), розлади імунної системи, обумовлені відторгненням трансплантата, хронічним відторгненням, а також ало- і ксенотрансплантатами тканин або клітин, включаючи щільні тканини (органи), острівці, м'язи, гепатоцити, нейрони. У числі прикладів імунопроліферативних захворювань можна назвати, без обмеження, псоріаз; Т-клітинну лімфому; Т-клітинний гострий лімфобластний лейкоз; доброякісний лімфоцитарний ангіїт; і аутоімунні захворювання, такі як вовчак (наприклад, системний червоний вовчак, lupus nephritis), тиреоїдит Хасімото, первинна мікседема, хвороба Грейвса, злоякісна анемія, аутоімунний атрофічний гастрит, хвороба Аддісона, діабет (наприклад, інсулінозалежний цукровий діабет типу І), синдром Гудпасчера, тяжка псевдопаралітична міастенія, пузирчатка, хвороба Крона, симпатична офтальмія, симпатичний увеїт, розсіяний склероз, аутоімунна гемолітична анемія, ідіопатична тромбоцитопенія, первинний біліарний цироз, хронічний активний гепатит, неспецифічний виразковий коліт, синдром Сегрена, ревматичні захворювання (наприклад, ревматоїдний артрит), поліміозит, склеродерма і змішана сполучнотканинна хвороба. У даний винахід входить також спосіб інгібування відторгнення пацієнтом трансплантатів однорідних органів i/або щільних тканин, де пацієнт є реципієнтом пересадженої тканини. Як правило, при трансплантації тканини відторгнення трансплантата ініціюється розпізнаванням його Тклітинами як чужорідного з наступною імунною відповіддю, що руйнує трансплантат. Молекули розчинного мутантного CTLA4 за даним винаходом, інгібуючи проліферацію Т-лімфоцитів і/або секрецію цитокінів, можуть привести до зменшення деструкції тканини, а індукція антигенспецифічної імунологічної толерантності Т-клітин може дати тривале приживлення трансплантата при відсутності необхідності в загальній імуносупресії. Крім того, молекули розчинного мутантного CTLA4 згідно з даним винаходом можна застосовувати разом з іншими фармацевтичними препаратами, без обмеження, такими як кортикостероїди, циклоспорин, рапаміцин, 23 мікофенолятмофетил, азатіоприн, такролізмус, базиліксимаб і/або інші біологічні сполуки. Даним винаходом також охоплюється спосіб інгібування гомологічної хвороби у пацієнта. Цей засіб полягає у введенні пацієнту (молекул) розчинного мутантного CTLA4 за винаходом, самого або разом із додатковими лігандами, реактогенними з IL-2, IL-4 або γ-інтерфероном. Наприклад, розчинний мутантний CTLA4 згідно з даним винаходом можна вводити реципієнту трансплантата кісткового мозку з метою інгібувати алореактивність донорних Т-клітин. Або ж донорні Т-клітини трансплантата кісткового мозку можна позбавляти імуногенності щодо алоантигенів реципієнта ex vivo перед трансплантацією. Інгібування Т-клітинної імунної відповіді розчинним мутантним CTLA4 може також бути корисним при лікуванні аутоімунних порушень. Багато аутоімунних порушень є наслідком непідходящої активації Т-клітин, що є реактивними до аутоантигенів і активують продукування цитокінів і аутоантитіл, "залучених" до патології захворювання. Введення (молекул) розчинного мутантного CTLA4 пацієнту, що страждає на аутоімунне порушення, може попередити активацію аутореактивних Тклітин і зменшити або зняти симптоми захворювання. Цей спосіб може також полягати у введенні пацієнту (молекул) розчинного мутантного CTLA4 за даним винаходом, самого або разом із додатковими лігандуми, реактогенними з IL-2, IL-4 або gінтерфероном. Даний винахід також включає у себе застосування (молекул) розчинного мутантного CTLA4 разом з іншими імуносупресорами, наприклад, із циклоспорином, див. [Mathiesen, "Prolonged Survival and Vascularization of Xenografted Human Glioblastoma Cells in the Central Nervous System of Cyclosporin Α-treated Rats" (1989) Cancer Lett. 44: 151-156], преднізоном, азатіоприном і метотрексатом [R. Handschumacher "Chapter 53: Drugs Used for Immunosuppression" pages 1264-1276]. Можливим є застосування інших імуносупресорів. Наприклад, при лікуванні ревматоїдного артриту (молекули) розчинного мутантного CTLA4 можна вводити з фармацевтичними препаратами, включаючи, але без обмеження, кортикостероїди, нестероїдні протизапальні лікарські препарати/інгібітори Сох-2, метотрексат, гідроксихлорохін, сульфасалазопірин, солі золота, етанерцепт, інфліксімаб, анакінру, азатіоприн і/або інші біологічні препарати, такі як проти-ТГФ-препарати. При лікуванні системного червоного вовчака молекули розчинного мутантного CTLA4 можна вводити з фармацевтичними препаратами, включаючи, але без обмеження, кортикостероїди, цитоксан, азатіоприн, гідроксихлорохін, мікофенолятмофетил і/або інші біологічні препарати. Крім того, при лікуванні розсіяного склерозу молекули розчинного мутантного CTLA4 можна вводити з фармацевтичними препаратами, включаючи, але без обмеження, кортикостероїди, інтерферон бета-1а, інтерферон бета-1b, глатирамеру ацетат, метоксантрону гідрохлорид і/або інші біологічні препарати. Молекули розчинного мутантного CTLA4 (переважно, L104EA29Ylg) можна також застосовува 87432 24 ти в комбінації з одним і більше із таких агентів для регуляції імунної відповіді: розчинний др39 (також відомий як ліганд CD40 (CD40L), CD-154, ТВАМ, TRAP), розчинний CD29, розчинний CD40, розчинний CD80, розчинний CD86, розчинний CD28, розчинний CD56, розчинний Thy-1, розчинний CD3, розчинний TCR, розчинний VLA-4, розчинний VCAM-1, розчинний LECAM-1, розчинний ELAM-1, розчинний CD44, антитіла, реактивні у відношенні до др39; антитіла, реактивні у відношенні до CD40; антитіла, реактивні у відношенні до В7; антитіла, реактивні у відношенні до CD28; антитіла, реактивні у відношенні до LFA-1; антитіла, реактивні у відношенні до LFA-2; антитіла, реактивні у відношенні до IL-2, антитіла реактивні у відношенні до IL-12, антитіла реактивні у відношенні до IFN-гамма; антитіла, реактивні у відношенні до CD2; антитіла, реактивні у відношенні до CD48; антитіла, реактивні у відношенні до будьяких ІСАМ (наприклад, ІСАМ-2); антитіла, реактивні у відношенні до CTLA4; антитіла, реактивні у відношенні до Thy-1; антитіла, реактивні у відношенні до CD56; антитіла, реактивні у відношенні до CD3; антитіла, реактивні у відношенні до CD29; антитіла, реактивні у відношенні до TCR; антитіла, реактивні у відношенні до VLA-4; антитіла, реактивні у відношенні до VCAM-1; антитіла, реактивні у відношенні до LECAM-1; антитіла, реактивні у відношенні до ELAM-1; антитіла, реактивні у відношенні до CD44. В окремих варіантах здійснення винаходу кращими є моноклональні антитіла (mAbs). В інших варіантах винаходу кращими є фрагменти антитіл. Для фахівця у даній галузі добре зрозуміло, що комбінація такого типу може включати у себе розчинний мутантний CTLA4 за винаходом й інший імуносупресор, розчинний мутантний CTLA4 із двома іншими імуносупресорами, розчинний мутантний CTLA4 із трьома іншими імуносупресорами і т.д. Визначення оптимальної комбінації і дозувань можна визначити й оптимізувати за добре відомими фахівцям методами. Серед деяких конкретних комбінацій можна назвати такі: L104EA29Ylg і CD80 mAbs; L104EA29Ylg і CD86 mAbs; L104EA29Ylg, CD80 mAbs i CD86 mAbs; L104EA29Ylg і gp39 mAbs; L104EA29Ylg і CD40 mAbs; L104EA29Ylg і CD28 mAbs; L104EA29Ylg, CD80 і CD86 mAbs і gp39 mAbs; L104EA29Ylg, CD80 і CD86 mAbs і CD40 mAbs; і L104EA29Ylg, анти-LFAI mAb і анти-gр39 mAb. Конкретним прикладом gp39 mAb є MR1. Існує також безліч інших комбінацій, які зможуть легко скласти і гідно оцінити фахівці у даній галузі. Розчинний мутантний CTLA4 (молекули), наприклад, L104EA29Y, можна вводити як єдиний (самостійний) активний інгредієнт або разом з іншими лікарськими речовинами для імуномодуляції або для лікування чи відвертання гострого або хронічного відторгнення ало- або ксенотрансплантата, або запальних чи аутоімунних захворювань, або ж для індукції толерантності. Наприклад, його можна використовувати в комбінації з інгібітором кальциневрину, наприклад, циклоспорином А або FK506; із імуносупресорним макролідом, наприклад, рапаміцином або його похідною, наприклад, 40-О-(2-гідроксі)етилрапаміцином; із лімфоцитар 25 ним агентом хомінгу, наприклад, FTY720 або його аналогом; із кортикостероїдами; із циклофосфамідом; з азатіоприном; із метотрексатом; лефлюномідом або його аналогом; із мізорибіном; із мікофенольною кислотою; із мікофенолятом мофетилу; із 15-дезоксипергуаліном або його аналогом; із імуносупресорними моноклональними антитілами, наприклад, моноклональними антитілами проти рецепторів лейкоцитів, утому числі МНС, CD2, CD3, CD4, CD11a/CD18, CD7, CD25, CD27, В7, CD40, CD45, CD58, CD137, ICOS, CD150 (SLAM), OX40, 4-ІВВ або інші ліганди; або з іншими імуномодуляторними сполуками, наприклад, CTLA4/CD28-lg, або молекулами інших інгібіторів адгезії, наприклад, mAbs чи з інгібіторами з низькомолекулярною вагою, включаючи антагоністи LFA-1, антагоністи селектину й антагоністи VLA4. Дана сполука є особливо ефективною у застосуванні в комбінації зі сполукою, що заважає CD40 і його ліганду, наприклад, антитілам проти CD40 і антитілам проти CD40-L досягти вищеописаних показань, наприклад, індукції толерантності. Коли молекули розчинного мутантного CTLA4 згідно з даним винаходом вводять у сполученні з іншими імуносупресорними/імуномодуляторними або протизапальними терапевтичними засобами, наприклад, такими, як наведені вище, дозування спільно вживаного імуносупресора, імуномодулятора або протизапального засобу, цілком зрозуміло, залежить від типу спільно вживаної лікарської речовини, наприклад, від того, є вона стероїдом чи циклоспорином, від конкретної застосовуваної лікарської речовини, від стану хворого і т.п. Відповідно до вищеописаного даний винахід пропонує в додатковому аспекті зазначені вище способи, що полягають у спільному, наприклад, одночасному (супутньому) або послідовному введенні терапевтично ефективної кількості (молекул) розчинного мутантного CTLA4 за даним винаходом, L104EA29Ylg, у вільному вигляді або у вигляді фармацевтично прийнятної солі, і другої лікарської речовини, при цьому зазначена друга лікарська речовина є імуносупресором, імуномодулятором або протизапальною лікарською речовиною, наприклад, такою, як зазначено вище. Крім того, винаходом передбачаються терапевтичні комбінації, наприклад, такі, як набір, що містить L104EA29Ylg, у вільному вигляді або у вигляді фармацевтично прийнятної солі, призначений для застосування у будь-який із вищезазначених способів шляхом введення одночасно або послідовно принаймні з однією фармацевтичною композицією, що містить імуносупресор, імуномодулятор або протизапальний лікарський засіб. Набір може містити інструкції для їх застосування. Способи одержання молекул згідно з даним винаходом Експресію молекул мутантного CTLA4 можна здійснювати в клітинах прокаріотів. Найчастіше прокаріоти мають форму різноманітних штамів бактерій. Бактерії можуть бути грампозитивними або грамнегативними. Як правило, кращими є грамнегативні бактерії, такі як Е. соlі. Можна використовувати також інші штами мікробів. 87432 26 Послідовності, що кодують молекули мутантного CTLA4, можна вбудовувати у вектор, створений для експресування чужорідних послідовностей у клітинах прокаріотів, таких як Е. соlі. До числа цих векторів можуть входити звичайно використовувані прокаріотні регулярні послідовності, котрі, згідно з даним описом, включають промотори ініціації транскрипції, у разі необхідності, з оператором, поряд із сайтами зв'язування рибосом, такі звичайно використовувані промотори, як промоторні системи бета-лактамази (пеніцилінази) і лактози (lac) [Chang, et al., (1977) Nature 198: 1056)], промоторна система триптофану (trp) [Goeddel, et al., (1980) Nucleic Acids Res. 8: 4057)], промотор PL і сайт зв'язування рибосом N-гена лямбда-фага [Shimatake, et al., (1981) Nature 292: 128]. Такі вектори експресії також включають ориджини реплікації і селективні маркери, такі як ген бета-лактамази або неоміцин-фосфотрансферази, що надають антибіотикам стійкості, завдяки чому вектори можуть реплікувати у бактеріях, і можна відбирати клітини, що несуть плазміди, вирощені при наявності антибіотиків, таких як ампіцилін або канаміцин. Плазміду експресії можна вводити в клітини прокаріотів за допомогою різноманітних стандартних методів, включаючи, без обмеження лише ними, СаСІ2-шок [Cohen, (1972) Proc. Natl. Acad. Sci. USA 69: 2110; Sambrook et al., (eds.), "Molecular Cloning: A Laboratory Manual", 2nd Edition, Cold Spring Harbor Press, (1989)] і електропорацію. Згідно з практикою застосування даного винаходу клітини еукаріотів також підхожі для використання як клітини-хазяїни. Прикладами клітин еукаріотів можуть бути будь-які тваринні клітини, як первинні, так і іморталізовані, дріжджові клітини (наприклад, Saccharomyces cerevisiae, Schizosaccharomyces pombe i Pichia pastoris) j рослинні клітини. Як приклади тваринних клітин, котрі можна використовувати як клітини-хазяїни, можна назвати клітини мієломи, COS і СНО. Підходящими СНО-клітинами можуть бути, зокрема, але без обмеження, DG44 [Chasin, et al., (1986) Som. Cell. Molec. Genet. 12: 555-556; Kolkekar (1997) Biochemistry 36: 10901-10909], CHO-K1 [ATCC No.CCL-61], CHO-K1 Tet-On клітинна лінія [Clontech], СНО, позначені як ЕСАСС 85050302 [CAMR, Salisbury, Wiltshire, UK], СНО-клон 13 [GEIMG, Geneva, IT], СНО-клон В [GEIMG, Genova, IT], CHO-K1/SF, позначений як ЕСАСС 93061607 [CAMR, Salisbury, Wiltshire, UK] і RR-CHOK1, позначений як ЕСАСС 92052129 [CAMR, Salisbury, Wiltshire, UK]. До числа підходящих рослинних клітин належать, наприклад, клітини тютюну (ціла рослина, культури клітин або калюс), пшениці, сої культурної і рису. Застосовувати можна також насіння пшениці, сої культурної і рису. Нуклеотидні послідовності, що кодують молекули мутантного CTLA4, можна також вбудовувати у вектор, призначений для експресування чужорідних послідовностей у клітинах еукаріотів. Регуляторні елементи вектора можуть варіюватися відповідно до конкретної еукаріотичної клітинихазяїна. 27 Використовувані звичайно еукаріотичні регулярні послідовності для застосування у векторах експресії включають до свого числа промотори і регулярні послідовності, сумісні з клітинами ссавців, такі, наприклад, як промотор CMV (вектор CDM8) і вірус саркоми птахів (ASV) (вектор pπLΝ). Серед інших застосовуваних звичайно промоторів можна назвати, наприклад, ранні і пізні промотори вірусу зеленої мартишки (SV40) [Fiers, et al., Nature 273: 113], або інші вірусні промотори, такі як вірусні промотори поліоми, аденовірусу 2 і вірусу бичачої папіломи. Можна також використовувати індуцибельний промотор, такий як hMTII [Karin, et al., (1982) Nature 299: 797-802]. Вектори експресії (молекул) мутантного CTLA4 в клітинах еукаріотів можуть також нести послідовності, називані енхансерами (енхансерна ділянка). Вони є важливими для оптимізації експресії гена і знаходяться в ряду послідовності вище або нижче промотора. Як приклади векторів експресії еукаріотичних клітин-хазяїнів можна назвати, без обмеження лише ними, вектори клітин-хазяїнів ссавців (наприклад, BPV-1, pHyg, pRSV, hSV2, pTK2 (Maniatis); pIRES (Clontech); pRc/CMV2, pRc/RSV, pSFV1 (Life Technologies); вектори pVPakc, вектори pCMV, вектори pSG5 (Stratagene)), ретровірусні вектори (наприклад, вектори pFB (Stratagene)), pCDNA-3 (Invitrogen) або їхні модифіковані форми, аденовірусні вектори; вектори аденозв'язаних вірусів, вектори бакуловірусів, дріжджові вектори (наприклад, вектори pESC (Stratagene)). Нуклеотидні послідовності, що кодують молекули мутантного CTLA4, можна інтегрувати в геном еукаріотичної клітини-хазяїна і реплікувати у вигляді реплік геному клітини-хазяїна. Або ж вектор, що несе молекули мутантного CTLA4, може містити ориджини реплікації, що роблять можливою позахромосомну реплікацію. Для експресії нуклеотидних послідовностей у Saccharomyces cerevisiae можна використовувати ориджин реплікації ендогенної дріжджової плазміди, виток (кільце) 2μ [Broach, (1983) Meth. Enz. 101: 307]. Або ж можна використовувати послідовності дріжджового геному, спроможні активувати автономну реплікацію, див., наприклад, [StinchMomb et al., (1979) Nature 282: 39); Tschemper et al., (1980) Gene 10: 157; Clarke et al., (1983) Meth. Enz. 101: 300]. Транскрипційні регулярні послідовності дріжджових векторів включають промотори синтезу гліколітичних ферментів [Hess et al., (1968) J Adv. Enzvme Reg. 7: 149; Holland et al., (1978) Biochemistry 17: 4900]. Додаткові промотори, відомі в техніці, містять CMV-промотор, створений у векторі CDM8 [Toyama and Okayama, (1990) FEBS 268: 217-221]; промотор 3-фосфогліцераткінази [Hitzeman et al., (1980) J. Biol. Chem. 255: 2073] і промотори інших гліколітичних ферментів. Інші промотори є індуцибельними, оскільки вони можуть регулюватися подразниками із навколишнього середовища або із живильного середовища для вирощування клітин. До таких індуцибельних промоторів належать промотори генів білків теплового шоку, алкогольдегідрогенази 2, 87432 28 ізоцитохрому С, кислої фосфатази, ферментів, пов'язаних із катаболізмом азоту, і ферментів, відповідальних за утилізацію мальтози і галактози. Регуляторні послідовності можна поміщати на 3'-кінець кодувальних послідовностей. Ці послідовності можуть стабілізувати матричну РНК. Такі термінатори виявлені на 3'- ділянці, що не транслюється і розташовується слідом за кодувальними послідовностями декількох генів дріжджів і ссавців. Серед прикладів векторів рослин і рослинних клітин можна назвати, без обмеження, плазміди Ті агробактерій, вірус мозаїки цвітної капусти (CaMV) і вірус золотої мозаїки томатів (TGMV). Загальні аспекти трансформації системихазяїна клітини ссавця описані Акселом [Axel, Патент США №4399216, виданий 16 серпня 1983 року]. Клітини ссавців можна трансформувати за допомогою різноманітних методів і, у тому числі, шляхом трансфекції у присутності фосфату кальцію, мікроін'єкції, електропорації або трансдукції за участі вірусних векторів. До числа способів уведення послідовності чужорідної ДНК у геноми рослин або дріжджів належать: (1) механічні способи, такі як мікроін'єкція ДНК в одиночні клітини або протопласти, здійснювана при інтенсивному перемішуванні клітин зі скляними кульками у присутності ДНК, або при "вистрілюванні" вольфрамовими чи золотими кульками в клітини або протопласти; (2) уведення ДНК шляхом перетворення клітинних мембран на такі, що є проникними для макромолекул, обробкою їх поліетиленгліколем або за допомогою електроімпульсів високої напруги (електропорація); або (3) застосування ліпосом (що містять кДНК), котрі зливаються з клітинними мембранами. Експресію молекул мутантного CTLA4 можна виявити за допомогою добре відомих методів. Наприклад, мутантні молекули можна виявити по забарвленню геля SDS-PAGE Кумасі блакитним або за допомогою імуноблотінгу із застосуванням антитіл, що зв'язуються з CTLA4. Виділення білка можна здійснювати за допомогою стандартних способів очищення білків, наприклад, шляхом афінної хроматографії або іонообмінної хроматографії, одержуючи таким чином практично чистий продукт [R. Scopes, "Protein Purification. Principles and Practice", Third Edition, Springer-Verlag (1994)]. Даним винаходом також охоплюються молекули розчинного мутантного CTLA4 (розчинний мутантний CTLA4), що продукуються відповідно до описаного вище. Мутагенез CTLA4lg за допомогою кодонів В одному із варіантів здійснення даного винаходу застосовуються сайт-скерований мутагенез і нова методика скринінгу для ідентифікації декількох мутацій у позаклітинній області CTLA4, що підвищують авідність зв'язування з CD86. У цьому варіанті здійснюються мутації в залишках на ділянках позаклітинної області CTLA4 від серину 25 до аргініну 33, С'-ланцюга (аланін 49 і треонін 51), Fланцюга (лізин 93, глутамінова кислота 95 і лейцин 96) і на ділянці від метіоніну до тирозину 102, від тирозину 103 до гліцину 107 і в G-ланцюзі в положеннях глутамін 111, тирозин 114 і ізолейцин 115. Ці сайти обрані на основі досліджень злитих білків 29 химерного CD28/CTLA4 [Peach et al., J. Exp. Med. (1994) 180: 2049-2058] і на основі моделі, що дозволяє спрогнозувати те, бічні ланцюги яких амінокислотних залишків піддаються дії розчинника, і відсутність ідентичності або гомології амінокислотних залишків у деяких положеннях між CD28 і CTLA4. Далі, будь-який залишок, який розташований у безпосередній близькості (5-20 ангстремів) до залишків, що ідентифікуються, розглядається як частина даного винаходу. З метою синтезу і скринінгу молекул розчинного мутантного CTLA4 зі зміненою афінністю до CD80 і/або CD86 обрана стратегія, що складається з двох стадій. На базі результатів експериментів спочатку була створена бібліотека мутацій у специфічному кодоні позаклітинної ділянки CTLA4, а потім проведено її скринінг шляхом аналізу за методом Віасоrе з метою ідентифікації мутантів зі зміненою реактивністю у відношенні до CD80 або CD86. В аналітичному приладі Віасоге (Pharmacia, Piscataway, NJ.) використовується детекторна система резонансу поверхневого плазмону, яка включає у себе ковалентне зв'язування або CD80lg або CD86lg із покритим декстраном сенсорним чипом, розташованим в детекторі. Після цього речовину (молекули випробуваної речовини), що аналізується, упорскують у камеру, що містить сенсорний чип, і оцінюють кількість комплементарного білка, що зв'язується, виходячи зі зміни молекулярної маси, що фізично зв'язується з покритим декстраном боком сенсорного чипа; зміну молекулярної маси можна вимірювати за допомогою детекторної системи. Переваги даного винаходу Оскільки зв'язування CTLA4 із CD80 і CD86 характеризується високими швидкостями протікання ("on", "до" ) і високими швидкостями дисоціації ("off", "від") і оскільки комплекси CTLA4lg-CD86 дисоціюють приблизно у 5-8 разів швидше, ніж комплекси CTLA4lg-CD80, резонно припустити, що уповільнення швидкості дисоціації CTLA4lg у його комплексі з CD80 і/або CD86 приводить до утворення сполук (молекул) із більш сильними імуносупресорними властивостями. Отже, можна очікувати, що молекули розчинного мутантного CTLA4, що характеризуються більш високою авідністю до CD80-або СР86-позитивних клітин порівняно з CTLA4 дикого типу або немутантними формами CTLA4lg, будуть блокувати примування антигенспецифічних активованих клітин із більш високою ефективністю, ніж CTLA4 дикого типу або немутантні форми CTLA4lg. Крім того, витрати на одержання CTLA4lg є дуже високими. Мутантний CTLA4lg (молекули), що володіє більш сильними імуносупресорними властивостями, може застосовуватися в клініці в значно менших дозах, ніж немутантний CTLA4lg, даючи аналогічні рівні імуносупресії. Отже, застосування розчинного мутантного CTLA4, наприклад, L104EA29Ylg, може бути дуже ефективним із погляду вартості. Нижче наведені приклади, метою яких є ілюстрація даного винаходу і надання допомоги фахівцям щодо його здійснення і застосування. Наведе 87432 30 ні приклади жодним чином не обмежують об'єму винаходу. Приклади Приклад 1 У даному прикладі описані способи одержання нуклеотидних послідовностей, що кодують молекули розчинного мутантного CTLA4 за даним винаходом. Отриманий "односайтовий" мутант (мутант по одному сайту) L104Elg, і вивчена кінетика його зв'язування з CD80 і/або з CD86. Нуклеотидна послідовність L104Elg використана як матриця для створення послідовності "двосайтового" мутантного (мутантного по двох сайтах) CTLA4, L104EA29Ylg, і вивчена кінетика його зв'язування з CD80 і/або з CD86. Мутагенез CTLA4 із використанням кодонів Для визначення мутантних молекул CTLA4lg із меншими швидкостями дисоціації («від»швидкості) у молекулах із CD80 і/або CD86 вироблена стратегія мутагенезу і скринінгу. "Односайтові" мутантні нуклеотидні послідовності одержують, використовуючи як матрицю CTLA4lg [патенти США №№5844095; 5851795; і 5885796; АТСС №68629]. Мутагенні олігонуклеотидні PCRпраймери створені для випадкового мутагенезу кодону специфічної кДНК, при цьому в положеннях 1 і 2 кодону припускається будь-яка основа, але в положенні 3 перебуває тільки гуанін або тимін (XXG/T; відомий також як NNG/T). Таким шляхом можна здійснювати довільну мутацію специфічного кодону, що кодує амінокислоту, щоб кодувати кожну з 20 амінокислот. При цьому мутагенез XXG/T дає 32 потенційних кодони, що кодують кожну з 20 амінокислот. PCR-продукти, що кодують мутації в безпосередній близькості до -M97G107 CTLA4lg (див. Фіг.7 або 8), відщеплюють за допомогою Sacl/Xbal і субклонують в аналогічним чином розрізаний вектор експресії CTLA4lg pLΝ. Цей спосіб застосовують, щоб одержати "односайтову" СТLА4-мутантну молекулу L104Elg (Фіг.8). З метою мутагенезу в просторовій близькості до S25-R33 CTLA4lg сайт рестрикції Nhel, що мовчить, спочатку вводять 5і до цієї петлі за допомогою PCR праймер-скерованого мутагенезу. PCRпродукти розщеплюють під дією Nhel/Xbal і субклонують у розрізані аналогічним чином вектори експресії CTLA4lg або L104Elg. Цей метод використовують для одержання молекули "двосайтового" мутантного CTLA4, L104EA29Ylg (Фіг.7). Зокрема, нуклеотидна молекула, що кодує "односайтову" мутантну молекулу CTLA4, L104Elg, використана як матриця для одержання молекули "двосайтового" мутантного CTLA4, L104EA29Ylg. Вектор piLN, що містить L104EA29Ylg, показаний на Фіг.12. Приклад 2 Нижче поданий опис способів скринінгу, застосовуваних для ідентифікації "одно-" і "двосайтових" мутантних СТLА4-поліпептидів, які експресують за допомогою описаних у Прикладі 1 конструкцій, що виявляють більш високу авідність зв'язування з антигенами CD80 і CD86 порівняно з молекулами немутантного CTLA4lg. Сучасні дослідження in vitro і in vivo показують, що CTLA4lg сам по собі не спроможний цілком блокувати примування антиген-специфічних акти 31 вованих Т-клітин. Дослідження in vitro з визначення інгібування проліферації Т-клітин, проведені з CTLA4lg і будь-яким моноклональним антитілом, що зв'язує CD80 або CD86, показують, що моноклональне антитіло, специфічне до CD80, не збільшує інгібування CTLA4lg. Проте, моноклональне антитіло проти CD86 підвищує інгібування, засвідчуючи те, що CTLA4lg неефективно блокує взаємодію з CD86. Ці дані підтверджують виявлений раніше Лінслеєм той факт, що для інгібування опосередкованих CD80 клітинних реакцій потрібно, приблизно, у 100 разів нижча концентрація CTLA4lg, ніж для СD86-опосередкованих реакцій [Linsley et al. (Immunity. (1994), 1: 793-801]. Виходячи з цього, було зроблене припущення, що молекули розчинного мутантного CTLA4, які характеризуються більш високою авідністю у відношенні до CD86, ніж CTLA4 дикого типу, спроможні краще блокувати примування антиген-специфічних активованих клітин, ніж CTLA4lg. З цією метою молекули розчинного мутантного CTLA4, описаного у Прикладі 1, піддають скринінгу, застосовуючи нову методику, для того, щоб ідентифікувати деякі мутації в позаклітинній області CTLA4, що підвищують авідність зв'язування з CD80 і CD86. Така стратегія скринінгу дає ефективний метод безпосередньої ідентифікації мутантів із порівняно меншими "від"-шидкостями, що не потребує очищення білка або кількісного визначення, оскільки визначення "від"-швидкості не залежить від концентрації [O'Shannessy D. J., et al., (1993) Anal. Biochem. 212: 457-468]. Клітини COS трансфікують, використовуючи мініпрепарат очищеної плазмідної ДНК, і культивують протягом декількох днів. До біосенсорних чипів BIAcore (Pharmacia Biotech AB, Uppsala, Sweden), покритих розчинними CD80lg або CD86lg, добавляють триденні кондиціоновані культурні середовища. Специфічне зв'язування і дисоціацію мутантних білків визначають за допомогою резонансу поверхневого плазмону [O'Shannessy et al., (1993) Anal. Biochem. 212: 457-468]. Всі експерименти проводять на біосенсорах BIAcore™ і BIAcore™ 2000 при температурі 25°С. Ліганди імобілізують на сенсорних чипах NCM5 для пошуку сполук (Pharmacia), використовуючи стандартне зв'язування з N-етил-N'(диметиламiнопропіл)карбодіїмід-Nгідроксисукцинімідом [Johnsson, В., et al., (1991) Anal. Biochem. 198: 268-277; Khilko, S. N.. et al., (1993) J. Biol. Chem. 268: 15425-15434]. Методика скринінгу Клітини COS, вирощені в 24-ямкових планшетах для тканинних культур транзиторно трансфікують за допомогою ДНК, що кодує мутантний CTLA4lg. Через 3 дні збирають культурні середовища, що містять секретований розчинний мутантний CTLA4lg. Потік кондиціонованих COS-клітинних культурних середовищ пропускають через біосенсорні чипи BIAcore, дериватизовані при використанні CD80lg або CD86lg (як описано в [Greene et al., (1996) J. Biol. Chem. 271: 26762-26771), або мутантні молекули ідентифікують при "від"-швидкостях, менших за "від"-швидкості, що спостерігаються 87432 32 для CTLA4lg дикого типу. Далі секвенують кДНК, що відповідають зразкам обраних середовищ, і одержують ДНК для здійснення транзиторної трансфекції COS-клітин у великих масштабах, за допомогою якої одержують мутантний білок CTLA4lg після очищення від протеїну А культурного середовища. Умови аналізу BIAcore і методика аналізу даних рівноважного зв'язування є такими, як описано в [Greene et al., (1996) J. Biol. Chem. 271: 2676226771] і як подано в даному описі. Анзліз даних BIAcore Діаграму чутливості перед аналізом нормалізують до нульового рівня відгуку (RU). Для визначення фонового рівня відгуку (RU) на основі великої різниці значень коефіцієнтів рефракції розчинів зразки пропускають через макет-дериватизовані проточні кювети. Константи рівноважної дисоціації (Кд) розраховують із графіка залежності Req від С, де Req означає відгук (відповідь) у стаціонарному стані мінус відгук на макет-дериватизованому чипі, а С означає молярну концентрацію аналіту. Криві зв'язування аналізують, використовуючи програми для креслення нелінійних кривих (Prism, GraphPAD Software), що випускаються промисловістю. Експериментальні дані спочатку наближують до моделі зв'язування одиночного ліганду з одиночним рецептором (1-сайтова модель, тобто проста ленгмюрова система А+В«АВ), і константи рівноважної асоціації (Kd=[A]·[B]\[AB] обчислюють за допомогою рівняння R=RmaxC/(Kd+C). Потім наближують дані до найпростішої двосайтової моделі зв'язування ліганду (тобто до рецептора, що має два невзаємодіючих незалежних сайти зв'язування згідно з рівнянням R=Rmax1·C\(Kd1+C)+Rmax2·C\(Kd2+C). Відповідність (придатність) цих двох моделей аналізують візуально, порівнюючи з експериментальними даними, і статистично, за допомогою Fкритерію суми квадратів. Як найбільш відповідну вибирають найпростішу односайтову модель, якщо двосайтова модель не виявляє значно більшої відповідності (р
ДивитисяДодаткова інформація
Назва патенту англійськоюSoluble ctla4 mutant molecules and uses thereof
Автори англійськоюPeach Robert J., Naemura Joseph R., Linsley Peter S., Bajorath Jurgen
Назва патенту російськоюРастворимый мутантный ctla4 и его применение
Автори російськоюПич Роберт Дж., Наемура Джозеф Р., Линсли Петер С., Беёрет Юрген
МПК / Мітки
МПК: C07K 14/705
Мітки: розчинний, застосування, мутантний, ctla4
Код посилання
<a href="https://ua.patents.su/31-87432-rozchinnijj-mutantnijj-ctla4-ta-jjogo-zastosuvannya.html" target="_blank" rel="follow" title="База патентів України">Розчинний мутантний ctla4 та його застосування</a>
Мутантний штам вірусу гепатиту b, виділена молекула нуклеїнової кислоти, що кодує мутантний головний поверхневий антиген штаму вірусу гепатиту в, вектор та векторна система для одержання поліпептиду, спосіб оде

Номер патенту: 73476
Опубліковано: 15.08.2005
Автори: Чен Уей Нінг, Лім Гек Кіоу, Ун Чонг Джін, Жао Йі
МПК: C12N 7/00, C12P 21/08, A61P 1/16, A61K 45/00, G01N 33/576, G01N 33/68, C07K 16/08, A61P 31/20, C12N 15/51, G01N 33/50, C12N 15/09, C12P 21/02, G01N 33/15, C12Q 1/68, A61P 35/00, C07K 14/02, A61K 39/29
Мітки: система, кислоти, поверхневий, векторна, виділена, штаму, оде, вірусу, нуклеїнової, кодує, антиген, поліпептиду, молекула, гепатиту, вектор, спосіб, мутантний, штам, одержання, головний
Формула / Реферат:
1. Виділений штам вірусу гепатиту В, визначений як штам Оon вірусу гепатиту В людини, що несе поверхневий антиген-'S'-133 (метіонін на треонін), що зберігається під номером доступу № Р97121504, Р97121505 і Р97121506 в Європейській колекції культур клітин від 15 грудня 1997 року.2. Виділена молекула нуклеїнової кислоти, яка кодує поліпептид, що є мутантним головним поверхневим антигеном штаму вірусу гепатиту В, яка відрізняється тим, що...
Мутантний поліпептид інтерлейкіну-18, виділена днк, що кодує мутантний поліпептид інтерлейкіну-18, вектор та фармацевтична композиція

Номер патенту: 79235
Опубліковано: 11.06.2007
Автори: Кім Соо Хіун, Дінарелло Чарльз
МПК: C12N 15/24, C07K 14/54, A61P 31/12, A61K 38/20, A61P 35/00
Мітки: мутантний, інтерлейкіну-18, фармацевтична, композиція, вектор, поліпептид, днк, кодує, виділена
Формула / Реферат:
1. Мутантний поліпептид IL-18, що включає мутації в одному або декількох амінокислотних залишках, які беруть участь в його взаємодії з IL-18-зв'язуючим білком, де мутації присутні в залишку, вибраному з Glu-42 і Lys-89 попередника IL-18 людини, що кодується кДНК з послідовністю SEQ ID NO:1, і де мутації є неконсервативними замінами.2. Поліпептид за п.1, де мутація присутня в залишку Glu-42.3. Поліпептид за п. 2, де мутація...
Розчинний у воді полімер, спосіб його одержання, продукт на його основі та спосіб стимулювання росту рослин
Номер патенту: 80395
Опубліковано: 25.09.2007
Автори: Сандерс Джон Л., Мазо Джекоб, Мазо Грігорі
МПК: C08F 222/00, C05G 3/10, A01P 21/00, C05G 3/00, A01G 29/00, C08F 220/00
Мітки: спосіб, росту, води, стимулювання, рослин, продукт, полімер, основі, розчинний, одержання
Формула / Реферат:
1. В основному розчинний у воді полімер, який в основному складається з повторюваних полімерних ланок, кожна з яких утворена щонайменше з двох різних полімерних ланок, індивідуально вибраних з групи, яка складається з ділянок В і С, де ділянка В описується загальною формулою або
Мутантний інтерлейкін-6 людини, молекула днк, що його кодує, вектор, клітина-хазяїн, а також спосіб одержання та використання мутантних il-6 для лікування захворювань, при яких il-6 має патогенний вплив

Номер патенту: 72426
Опубліковано: 15.03.2005
Автори: Гротзінгер Йоахім, Елерс Марк, Розе-Йон Штефан
МПК: C12P 21/02, C12N 15/24, C12N 15/09, A61P 43/00, A61K 38/00, C07K 14/54, A61K 31/00, C12N 1/21
Мітки: яких, молекула, має, інтерлейкін-6, кодує, патогенний, спосіб, вектор, використання, одержання, клітина-хазяїн, мутантних, днк, захворювань, вплив, лікування, людини, мутантний, також
Формула / Реферат:
1. Мутантный интерлейкин-6 (IL-6) человека, имеющий аминокислотную последовательность SEQ ID NО:1, содержащую следующие точечные мутации по сравнению с аминокислотной последовательностью встречающегося в природе ІL-6 человека: Pro в положении 54, Glu в положении 159, Pro в положении 162, Leu в положении 170 и Аrg в положении 176, или его фрагмент, обладающий такой же IL-6-антагонистической активностью, как и указанный мутантный...
Розчинний сухий кавовий напій (варіанти) та спосіб його одержання

Номер патенту: 73093
Опубліковано: 15.06.2005
Автор: Фрітц Міхаель
МПК: A23F 5/24
Мітки: одержання, спосіб, розчинний, сухий, кавовий, напій, варіанти
Формула / Реферат:
1. Розчинний сухий кавовий напій, який після додавання води здатний утворювати кавовий напій з піною на поверхні, що включає розчинну газовмісну основу, що включає тверді частки розчинної кави і розчинний наповнювач, який є олігофруктозою, причому розчинний наповнювач складає не менше 35 мас.% речовини основи і ефективний для збільшення об'єму розчинного сухого кавового напою, що включає газ.2. Розчинний сухий кавовий напій за п.1, у...
Попередній патент: Специфічні для насіння льону промотори
Наступний патент: Гонадотропіни для стимулювання фолікулогенезу
Випадковий патент: Спосіб дистанційного біологічного тестування наявності гострої токсичності водного середовища