Домени, які зв’язуються з деімунізованою сироваткою, і їхнє застосування для збільшення часу напівжиття в сироватці
Номер патенту: 111612
Опубліковано: 25.05.2016
Автори: Джонсон Леслі С., Бонвіні Ецио, Барат Бхасваті, Хуан Лін

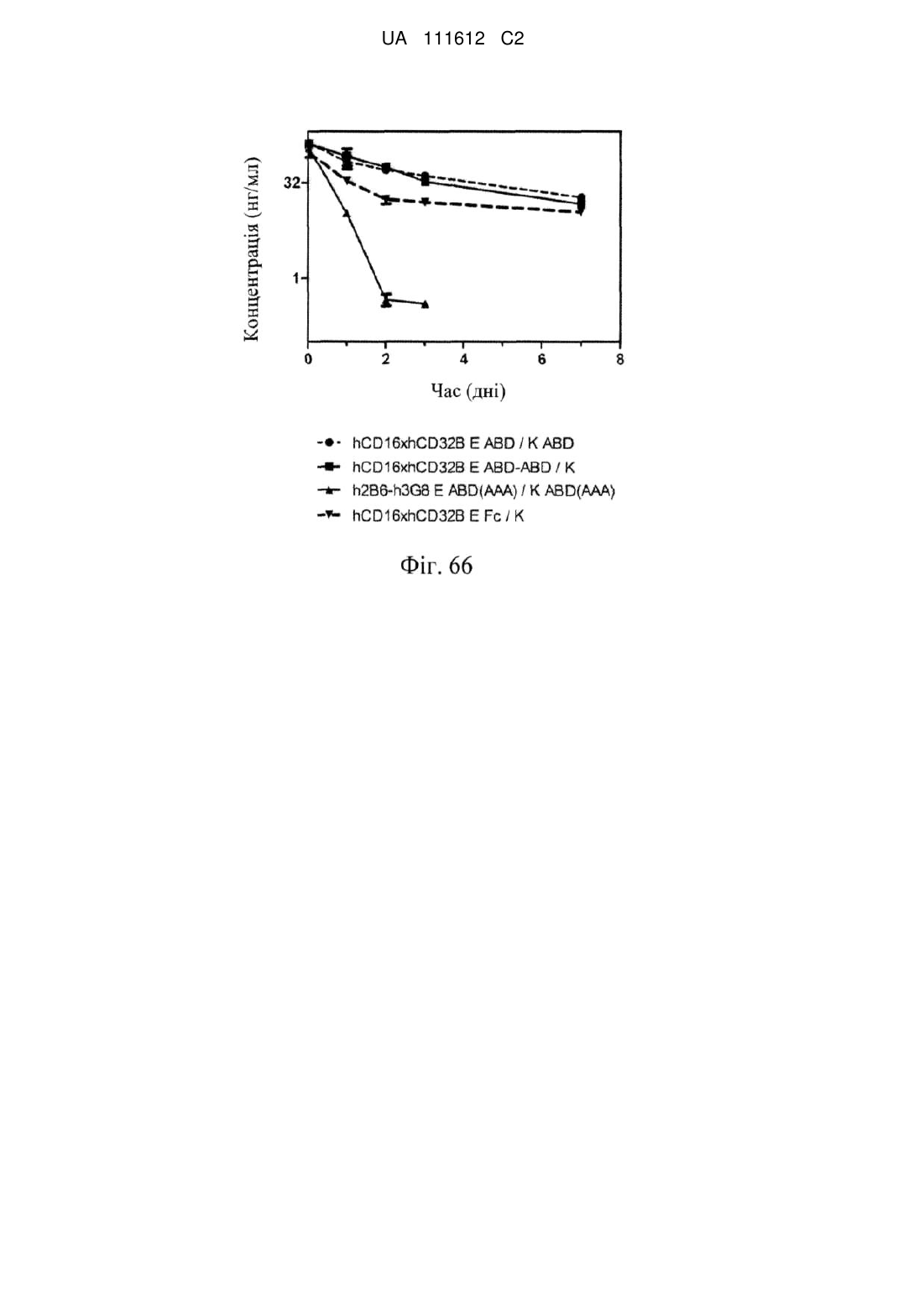



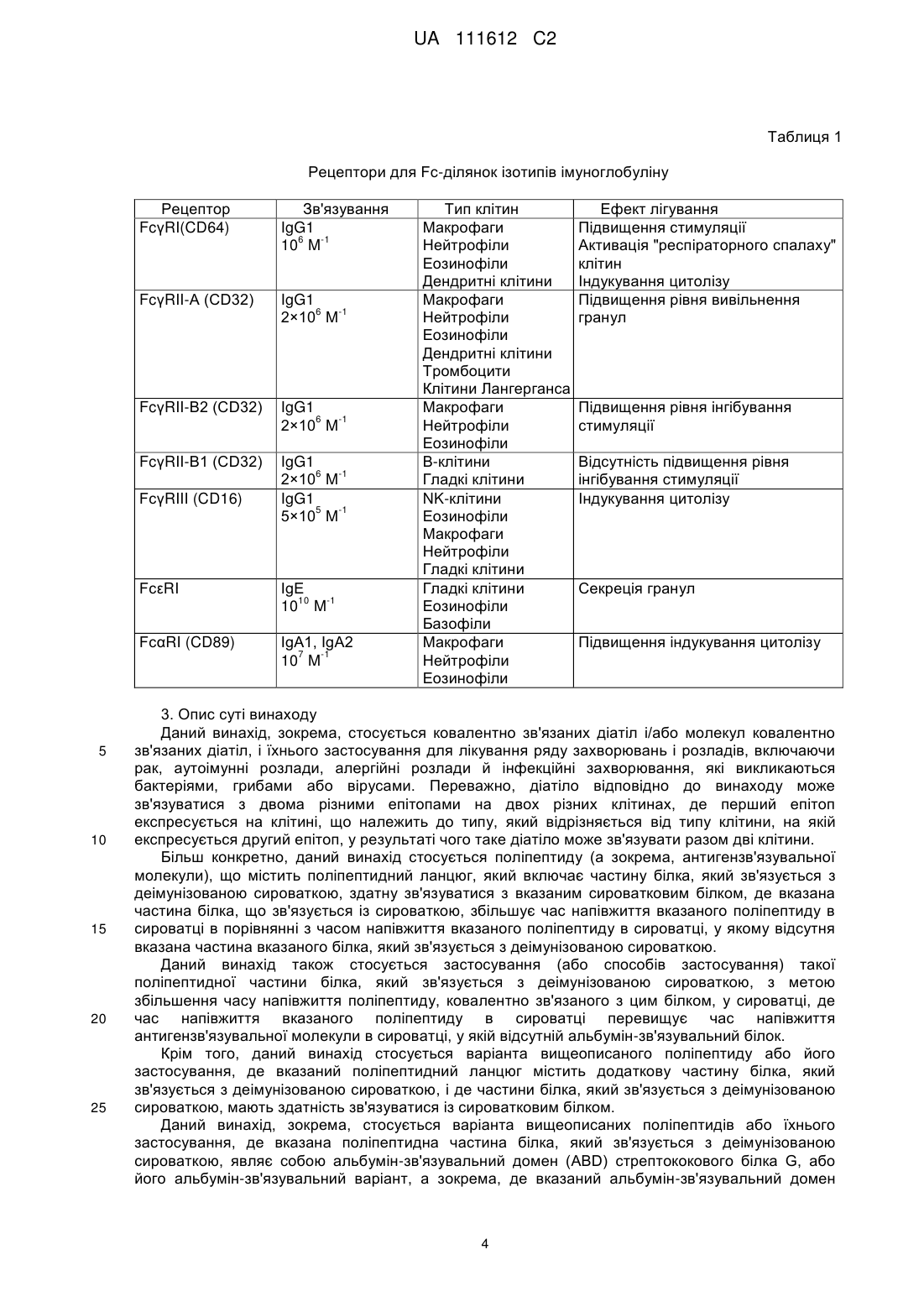


Формула / Реферат
1. Поліпептид, що містить частину білка, який зв'язується з деімунізованою сироваткою, здатну зв'язуватися з вказаним сироватковим білком, де вказана частина білка, що зв'язується із деімунізованою сироваткою, являє собою варіант деімунізованого альбумінзв'язувального домену (ABD) дикого типу стрептококового G-білка, де вказаний ABD дикого типу має амінокислотну послідовність SEQ ID NO: 304; де вказаний варіант ABD має амінокислотну послідовність, яка відрізняється від SEQ ID NO: 304 вмістом:
(A) заміни валіну на аланін в положенні 71 ABD, заміни валіну на аланін в положенні 74 ABD або заміни аспартату на аланін в положенні 79 ABD; де вказані положення замін в ABD відповідають положенням 31, 34 і 39 SEQ ID NO: 304;
(B) заміни лейцину на аланін в положенні 64 ABD, заміни ізолейцину на аланін в положенні 65 ABD і заміни валіну на аланін в положенні 71 ABD; де вказані положення замін в ABD відповідають положенням 24, 25 і 31 SEQ ID NO: 304; або
(C) заміни аспарагіну на аспарагінову кислоту в положенні 66 ABD, заміни треоніну на серин в положенні 70 ABD і заміни валіну на аланін в положенні 71 ABD; де вказані положення замін в ABD відповідають положенням 26, 30 і 31 SEQ ID NO: 304;
де вказана деімунізована частина білка, що зв'язується з сироваткою, збільшує час напівжиття вказаного поліпептиду в сироватці в порівнянні з часом напівжиття вказаного поліпептиду в сироватці, у якому відсутня вказана частина вказаного білка, який зв'язується з деімунізованою сироваткою.
2. Поліпептид за п. 1, де вказаний поліпептид містить додаткову частину білка, який зв'язується з деімунізованою сироваткою, і де вказані частини вказаного білка, який зв'язується з деімунізованою сироваткою, мають здатність зв'язуватися з вказаним сироватковим білком.
3. Поліпептид за будь-яким з пп. 1-2, де вказаний варіант ABD містить вказану заміну валіну на аланін в положенні 71 ABD, вказану заміну валіну на аланін в положенні 74 ABD або вказану заміну аспартату на аланін в положенні 79 ABD; де вказані положення замін в ABD відповідають положенням 31, 34 і 39 SEQ ID NO: 304.
4. Поліпептид за будь-яким з пп. 1-2, де вказаний варіант ABD містить вказану заміну лейцину на аланін в положенні 64 ABD, вказану заміну ізолейцину на аланін в положенні 65 ABD і вказану заміну валіну на аланін в положенні 71 ABD; де вказані положення замін в ABD відповідають положенням 24, 25 і 31 SEQ ID NO: 304.
5. Поліпептид за будь-яким з пп. 1-2, де вказаний варіант ABD містить вказану заміну аспарагіну на аспарагінову кислоту в положенні 66 ABD, вказану заміну треоніну на серин в положенні 70 ABD і вказану заміну валіну на аланін в положенні 71 ABD; де вказані положення замін в ABD відповідають положенням 26, 30 і 31 SEQ ID NO: 304.
6. Поліпептид за п. 1 або 2, де вказаний варіант ABD має послідовність SEQ ID NO: 323, SEQ ID NO: 324, SEQ ID NO: 325, SEQ ID NO: 326, SEQ ID NO: 327, SEQ ID NO: 328 або SEQ ID NO: 329.
7. Поліпептид за будь-яким з пп. 1-6, де вказаний поліпептид містить антигензв'язувальну молекулу.
8. Поліпептид за п. 7, де вказана антигензв'язувальна молекула являє собою діатіло, що складається щонайменше з першого і другого поліпептидних ланцюгів, що взаємодіють один з одним з утворенням двох антигензв'язувальних сайтів, де щонайменше один із вказаних поліпептидних ланцюгів містить вказану другу поліпептидну частину, що включає вказану частину білка, який зв'язується з деімунізованою сироваткою, здатну зв'язуватися з вказаним сироватковим білком.
9. Поліпептид за п. 8, де обидва вказані перший і другий поліпептидні ланцюги включають другу поліпептидну частину, яка містить вказану частину вказаного білка, який зв'язується з деімунізованою сироваткою, здатну зв'язуватися із сироватковим білком.
10. Поліпептид за будь-яким з пп. 8-9, де вказані перший і другий поліпептидні ланцюги ковалентно зв'язані один з одним.
11. Поліпептид за будь-яким з пп. 8-10, де вказане діатіло зв'язується:
(А) з рецептором групи 2D (NKG2D) природних кілерів або з Т-клітинним рецептором (TCR); і
(В) з пухлиноасоційованим антигеном.
12. Поліпептид за п. 7, де вказаним антигеном є антиген раку молочної залози, антиген раку яєчника, антиген раку передміхурової залози, антиген раку шийки матки, антиген карциноми підшлункової залози, антиген раку легень, антиген раку сечового міхура, антиген раку товстої кишки, антиген раку яєчок, антиген гліобластоми, антиген, асоційований зі злоякісною пухлиною В-клітин, антиген, асоційований із множинною мієломою, антиген, асоційований з неходжкінською лімфомою, або антиген, асоційований із хронічним лімфоцитарним лейкозом.
13. Поліпептид за будь-яким з пп. 8-11, де вказане діатіло має антигензв'язувальний сайт, який зв'язується з антигеном раку молочної залози, антигеном раку яєчника, антигеном раку передміхурової залози, антигеном раку шийки матки, антигеном карциноми підшлункової залози, антигеном раку легень, антигеном раку сечового міхура, антигеном раку товстої кишки, антигеном раку яєчок, антигеном гліобластоми, антигеном, асоційованим із злоякісною пухлиною В-клітин, антигеном, асоційованим з множинною мієломою, антигеном, асоційованим з неходжкінською лімфомою, або антигеном, асоційованим з хронічним лімфоцитарним лейкозом.
14. Застосування частини білка, який зв'язується з деімунізованною сироваткою, здатного зв'язуватися з вказаною сироваткою білка, з метою збільшення часу напівжиття ковалентно зв'язаного поліпептиду в сироватці, де вказана частина білка, яка зв'язується з деімунізованою сироваткою, являє собою варіант деімунізованого альбумінзв'язувального домену (ABD) дикого типу стрептококового G-білка, де вказаний ABD дикого типу має амінокислотну послідовність SEQ ID NO: 304; де вказаний варіант ABD має амінокислотну послідовність, яка відрізняється від SEQ ID NO: 304 вмістом:
(A) заміни валіну на аланін в положенні 71 ABD, заміни валіну на аланін в положенні 74 ABD або заміни аспартату на аланін в положенні 79 ABD; де вказані положення замін в ABD відповідають положенням 31, 34 і 39 SEQ ID NO: 304;
(B) заміни лейцину на аланін в положенні 64 ABD, заміни ізолейцину на аланін в положенні 65 ABD і заміни валіну на аланін в положенні 71 ABD; де вказані положення замін в ABD відповідають положенням 24, 25 і 31 SEQ ID NO: 304; або
(С) заміни аспарагіну на аспарагінову кислоту в положенні 66 ABD, заміни треоніну на серин в положенні 70 ABD і заміни валіну на аланін в положенні 71 ABD; де вказані положення замін в ABD відповідають положенням 26, 30 і 31 SEQ ID NO: 304;
де відбувається вказане збільшення часу напівжиття в сироватці в порівнянні з часом напівжиття в сироватці вказаної антигензв'язувальної молекули, в якій відсутня вказана частина білка, який зв'язується з деімунізованою сироваткою.
15. Застосування за п. 14, де вказаний поліпептид містить додаткову частину білка, який зв'язується з деімунізованою сироваткою, і де вказані частини вказаного білка, який зв'язується з деімунізованою сироваткою, мають здатність зв'язуватися з вказаним сироватковим білком.
16. Застосування за будь-яким з пп. 14-15, де вказаний варіант ABD містить вказану заміну валіну на аланін в положенні 71 ABD, вказану заміну валіну на аланін в положенні 74 ABD або вказану заміну аспартату на аланін в положенні 79 ABD; де вказані положення замін в ABD відповідають положенням 31, 34 і 39 SEQ ID NO: 304.
17. Застосування за будь-яким з пп. 14-15, де вказаний варіант ABD містить вказану заміну лейцину на аланін в положенні 64 ABD, вказану заміну ізолейцину на аланін в положенні 65 ABD і вказану заміну валіну на аланін в положенні 71 ABD; де вказані положення замін в ABD відповідають положенням 24, 25 і 31 SEQ ID NO: 304.
18. Застосування за будь-яким з пп. 14-15, де вказаний варіант ABD містить вказану заміну аспарагіну на аспарагінову кислоту в положенні 66 ABD, вказану заміну треоніну на серин в положенні 70 ABD і вказану заміну валіну на аланін в положенні 71 ABD; де вказані положення замін в ABD відповідають положенням 26, 30 і 31 SEQ ID NO: 304.
19. Застосування за п. 14, де вказаний варіант ABD має послідовність SEQ ID NO: 304, SEQ ID NO: 323, SEQ ID NO: 324, SEQ ID NO: 325, SEQ ID NO: 326, SEQ ID NO: 327, SEQ ID NO: 328 або SEQ ID NO: 329.
20. Застосування за будь-яким з пп. 14-19, де вказаний поліпептид містить антигензв'язувальну молекулу.
21. Застосування за п. 20, де вказана антигензв'язувальна молекула являє собою діатіло, що складається щонайменше з першого і другого поліпептидних ланцюгів, які взаємодіють один з одним з утворенням двох антигензв'язувальних сайтів, де щонайменше один з поліпептидних ланцюгів містить вказану другу поліпептидну частину, що включає вказану частину білка, який зв'язується з деімунізованою сироваткою, здатну зв'язуватися з вказаним сироватковим білком.
22. Застосування за п. 21, де обидва вказані перший і другий поліпептидні ланцюги включають другі поліпептидні частини, що містять вказану частину вказаного білка, який зв'язується з деімунізованою сироваткою, здатну зв'язуватися із сироватковим білком.
23. Застосування за будь-яким з пп. 21-22, де вказані перший і другий поліпептидні ланцюги ковалентно зв'язані один з одним.
24. Застосування за будь-яким з пп. 21-23, де вказане діатіло зв'язується:
(A) з рецептором групи 2D (NKG2D) природних кілерів або з Т-клітинним рецептором (TCR); і
(B) з пухлиноасоційованим антигеном;
для одержання лікарського засобу для лікування раку, експресуючого вказаний пухлиноасоційований антиген.
25. Застосування за п. 20, де вказаним антигеном є антиген раку молочної залози, антиген раку яєчника, антиген раку передміхурової залози, антиген раку шийки матки, антиген карциноми підшлункової залози, антиген раку легень, антиген раку сечового міхура, антиген раку товстої кишки, антиген раку яєчок, антиген гліобластоми, антиген, асоційований зі злоякісною пухлиною В-клітин, антиген, асоційований із множинною мієломою, антиген, асоційований з неходжкінською лімфомою, або антиген, асоційований із хронічним лімфоцитарним лейкозом.
26. Застосування за п. 21-24, де вказане діатіло має антигензв'язувальний сайт, який зв'язується з антигеном раку молочної залози, антигеном раку яєчника, антигеном раку передміхурової залози, антигеном раку шийки матки, антигеном карциноми підшлункової залози, антигеном раку легень, антигеном раку сечового міхура, антигеном раку товстої кишки, антигеном раку яєчок, антигеном гліобластоми, антигеном, асоційованим із злоякісною пухлиною В-клітин, антигеном, асоційованим з множинною мієломою, антигеном, асоційованим з неходжкінською лімфомою, або антигеном, асоційованим з хронічним лімфоцитарним лейкозом.
27. Композиція, яка містить поліпептид за будь-яким з пп. 1-13 і фармацевтично прийнятний носій.
Текст
Реферат: Винахід належить до поліпептиду, що містить частину білка, який зв’язується з деімунізованою сироваткою, здатну зв’язуватися з вказаним сироватковим білком, де вказана частина білка, що зв’язується з деімунізованою сироваткою, являє собою варіант деімунізованого альбумінзв’язувального домену (АВD) дикого типу стрептококового G-білка, способів одержання такого поліпептиду і його застосування. UA 111612 C2 (12) UA 111612 C2 UA 111612 C2 5 10 15 20 25 30 35 40 45 50 55 60 Перехресне посилання на споріднені заявки У даній заявці заявляється перевага заявки на патент США реєстр. № 61/488725 (поданої 21 травня 2011 р.; на розгляді), яка у всій своїй повноті вводиться в даний опис за допомогою посилання. Посилання на список послідовностей: Дана заявка включає один або декілька Списків послідовностей відповідно до статті 37 Зводу федеральних правил (C.F.R.) 1.821 і далі, які представлені у вигляді текстового документа і комп'ютерного файлу, і які у всій своїй повноті вводяться в даний опис за допомогою посилання. Передумови створення винаходу: 1. Галузь, до якої належить винахід Даний винахід стосується поліпептиду (наприклад, антигензв'язувальної молекули), що включає поліпептидну частину білка, який зв'язується з деімунізованою сироваткою, яка здатна зв'язуватися з вказаним сироватковим білком. Присутність білка, що зв'язується із сироваткою, сприяє збільшенню часу напівжиття поліпептиду в сироватці в порівнянні з часом напівжиття цього поліпептиду, у якому відсутня поліпептидна частина білка, що зв'язується з деімунізованою сироваткою. Даний винахід також стосується способів одержання таких молекул і їх застосування. Даний винахід, зокрема, стосується молекул діатіл, які іноді називаються "перенацілювальні реагенти з подвійною афінністю" ("DART"), і їхнього застосування для лікування ряду захворювань і розладів, включаючи імунологічні розлади і рак. Молекули діатіл відповідно до винаходу включають щонайменше два поліпептидні ланцюги, зв'язані так, що вони утворюють щонайменше два епітоп-зв'язувальні сайти, що можуть розпізнавати ті самі або різні епітопи. Крім того, ці епітопи можуть походити від тих самих або від різних молекул, або вони можуть знаходитися на тих самих або на різних клітинах. Окремі поліпептидні ланцюги молекул діатіл можуть бути ковалентно зв'язані непептидними ковалентними зв'язками, такими як, але не обмежуються ними, дисульфідні зв'язки з цистеїнових залишків, розташованих на кожному поліпептидному ланцюзі. У конкретних варіантах винаходу молекули діатіл відповідно до винаходу також містять Fc-ділянку, яка має антитіло-подібні функціональні властивості, що підходять для конструювання такої молекули. 2. Опис попереднього рівня техніки Конструювання ковалентно зв'язаних діатіл здійснюють на основі одноланцюжкової Fvконструкції (scFv) (Holliger et al. (1993) "Diabodies": Small Bivalent And Bispecific Antibody Fragments", Proc. Natl. Acad. Sci. USA 90:6444-6448; ця публікація у всій своїй повноті вводиться в даний опис за допомогою посилання). У інтактному немодифікованому IgG домени VL і VH розташовані на окремих поліпептидних ланцюгах, тобто на легкому ланцюзі і важкому ланцюзі, відповідно. Взаємодія легкого і важкого ланцюгів антитіла, зокрема, взаємодія доменів VL і VH, приводить до утворення одного з епітоп-зв'язувальних сайтів антитіла. На противагу цьому, scFv-конструкція включає VL- і VH-домени антитіла, що містяться в одному поліпептидному ланцюзі, де вказані домени розділені гнучким лінкером, який має довжину, достатню для самозбирання двох доменів в один функціональний епітоп-зв'язувальний сайт. У випадку, коли самозбирання неможливе через недостатню довжину лінкера (приблизно менше ніж 12 амінокислотних залишків), то дві scFv-конструкції піддавали взаємодії одна з одною з утворенням двовалентних молекул, тобто VL одиного ланцюга, зв'язаного з VH іншого ланцюга (див. огляд Marvin et al. (2005) "Recombinant Approaches To IgG-Like Bispecific Antibodies" Acta Pharmacol. Sin. 26:649-658). Крім того, ймовірно, що додавання цистеїнового залишку до С-кінця конструкції приводить до утворення дисульфідного зв'язку в поліпептидних ланцюгах, що стабілізує отриманий димер і не чинить негативного впливу на зв'язувальні властивості двовалентної молекули (див., наприклад, Olafsen et al. (2004) "Covalent Disulfide-Linked Anti-CEA Diabody Allows Site-Specific Conjugation And Radiolabeling For Tumor Targeting Applications", Prot. Engr. Des. Sel. 17:21-27). Крім того, при відборі VL- і VH-доменів з різною специфічністю може бути сконструйована не тільки двовалентна, але також і біспецифічна молекула. Двовалентні діатіла знаходять широке застосування, включаючи застосування в терапії й у імунодіагностиці. Двовалентність додає конструкції більшої гнучкості і дозволяє сконструювати діатіло, яке може мати різні застосування, де потрібна підвищена авідність відносно мультимерних антигенів, перехресне зв'язування з різними антигенами і пряма доставка в клітини конкретних типів, залежно від обох присутніх антигенів-мішеней. Фахівцям також відомо, що молекули діатіл, завдяки їхній підвищеній валентності, низькій швидкості дисоціації і швидкому кліренсу з кровотоку (у випадку діатіл невеликого розміру, а саме, ~50 кДа або менше), можуть бути, зокрема, використані для візуалізації ракової пухлини (Fitzgerald et al. 1 UA 111612 C2 5 10 15 20 25 30 35 40 45 50 55 60 (1997) "Improved Tumour Targeting By Disulphide Stabilized Diabodies Expressed In Pichia pastoris" Protein Eng. 10:1221). Особливо важливою властивістю є перехресне зв'язування різних клітин, наприклад, перехресне зв'язування цитотоксичних Т-клітин з пухлинними клітинами (Staerz et al. (1985) "Hybrid Antibodies Can Target Sites For Attack By T Cells", Nature 314:628-631, і Holliger et al. (1996) "Specific Killing Of Lymphoma Cells By Cytotoxic T-Cells Mediated By A Bispecific Diabody", Protein Eng. 9:299-305). Епітоп-зв'язувальні домени діатіл можуть бути також спрямовані проти поверхневої детермінанти будь-яких імунних ефекторних клітин, таких як CD3, CD16, CD32 або CD64, які експресуються на T-лімфоцитах, природних клітинах-кілерах (NK) або інших мононуклеарних клітинах. У багатьох дослідженнях було також виявлено, що зв'язування діатіла з детермінантами ефекторних клітин, наприклад, з Fсγ-рецепторами (FсγR), приводить до активації ефекторних клітин (Holliger et al. (1996) "Specific Killing Of Lymphoma Cells By Cytotoxic T-Cells Mediated By A Bispecific Diabody", Protein Eng. 9:299-305; Holliger et al. (1999) "Carcinoembryonic Antigen (CEA) -Specific T-cell Activation In Colon Carcinoma Induced By Anti-CD3×Anti-CEA Bispecific Diabodies And B7×Anti-CEA Bispecific Fusion Proteins", Cancer Res. 59:2909-2916). Звичайно, активація ефекторних клітин запускається зв'язуванням антигензв'язаного антитіла з ефекторними клітинами за допомогою взаємодії Fc-FсγR; і відповідно до цього молекули діатіл відповідно до винаходу можуть мати Ig-подібні функціональні властивості незалежно від того, чи містять вони Fc-домен чи ні (наприклад, як було визначено в аналізі на будь-яку ефекторну функцію, відомому фахівцям або описаному в даній заявці (наприклад, в аналізі на ADCC)). При перехресному зв'язуванні пухлинних і ефекторних клітин діатіло не тільки переносить ефекторні клітини на ділянку, розташовану поблизу від пухлинних клітин, але також ефективно здійснює знищення пухлини (див., наприклад, публікацію Cao et al. (2003) "Bispecific Antibody Conjugates In Therapeutics", Adv. Drug. Deliv. Rev. 55:171-197, що у всій своїй повноті вводиться в даний опис за допомогою посилання). 2.1 Рецептори ефекторних клітин і їхня роль в імунній системі При нормальному функціонуванні імунної системи взаємодія комплексів "антитіло-антиген" із клітинами імунної системи приводить до продукування відповідей широкого ряду, а саме, від ефекторних функцій, таких як антитіло-залежна цитотоксичність, дегрануляція тучних клітин і фагоцитоз, до виробляння імуномодулюючих сигналів, таких як сигнали регуляції проліферації лімфоцитів і секреції антитіл. Усі ці взаємодії ініціюються за допомогою зв'язування Fc-домену або антитіл імунних комплексів з поверхневими рецепторами спеціалізованих клітин, розташованими на гемопоетичних клітинах. Розмаїтість клітинних відповідей, що запускаються антитілами й імунними комплексами, обумовлено структурною гетерогенністю Fc-рецепторів. Fc-рецептори мають структурну подібність з антигензв'язувальними доменами, які переважно опосередковують передачу внутрішньоклітинного сигналу. Fсγ-рецептори, тобто члени суперсімейства білків, які кодуються генами імуноглобулінів, являють собою поверхневі глікопротеїни, які можуть зв'язуватися з Fсγ-частиною молекул імуноглобуліну. Кожен член цього сімейства розпізнає імуноглобуліни одного або більше ізотипів за допомогою домену розпізнавання, який присутній на альфа-ланцюзі Fсγ-рецептора. Fсγ-рецептори визначаються по їхній специфічності до підтипів імуноглобулінів. Fсγ-рецептори для IgG позначаються FсγR, для IgE-FcεR і для IgA-FcαR. Різні допоміжні клітини несуть Fсγрецептори для антитіл різних ізотипів, і по ізотипу антитіла визначають, які саме допоміжні клітини беруть участь у вироблянні даної відповіді (див. огляд Ravetch J.V. et al. (1991) "Fc Receptors" Annu. Rev. Immunol. 9:457-92; Gerber J.S. et al. (2001) "Stimulatory And Inhibitory Signals Originating From The Macrophage Fcgamma Receptors", Microbes and Infection, 3:131-139; Billadeau D.D. et al. (2002), "ITAMs Versus ITIMs: Striking A Balance During Cell Regulation", The Journal of Clinical Investigation, 2(109):161-1681; Ravetch J.V. et al. (2000) "Immune Inhibitory Receptors" Science, 290:84-89; Ravetch J.V. et al., (2001) "IgG Fc Receptors" Annu. Rev. Immunol. 19:275-90; Ravetch J.V. (1994) "Fc Receptors: Rubor Redux" Cell, 78(4):553-60). Різні Fсγрецептори, клітини, які експресують ці рецептори, і специфічність до їх ізотипів систематизовані в таблиці 1 (взятій з деякими змінами з публікації Immunobiology: The Immune System in Health th and Disease, 4 ed. 1999, Elsevier Science Ltd/Garland Publishing, New York). Fсγ-рецептори Кожен член цього сімейства являє собою інтегральний мембранний глікопротеїн, що має позаклітинні домени, споріднені C2-серії імуноглобулін-подібних доменів, один трансмембранний домен і один внутрішньоцитоплазматичний домен з довжиною, що варіюється. Фахівцям відомі три FсγR, які позначаються FсγRI(CD64), FсγRII(CD32) і FсγRIII(CD16). Ці три рецептори кодуються окремими генами; однак високий ступінь гомології між цими трьома членами такого сімейства дозволяє припустити, що вони, ймовірно, утворені від спільного попередника в результаті дуплікації генів. 2 UA 111612 C2 5 10 15 20 25 30 35 40 45 FсγRII(CD32) Білки FсγRII являють собою інтегральні мембранні глікопротеїни розміром 40 кДа, що зв'язуються тільки з IgG-комплексом, що зумовлено їх низькою афінністю відносно мономерного 6 -1 Ig (10 M ). Цей рецептор являє собою FcγR, який експресується у широкого кола хазяїнів і присутній на всіх гемопоетичних клітинах, включаючи моноцити, макрофаги, B-клітини, NKклітини, нейтрофіли, гладкі клітини і тромбоцити. FcγRII має тільки дві імуноглобулін-подібні ділянки у своєму імуноглобулін-зв'язувальному ланцюгу, а тому він має набагато меншу афінність відносно IgG, ніж рецептор FcγRI. Існують три людські гени FcγRII (FcγRII-A, FcγRII-B, FcγRII-C), кожний з який зчеплений з IgG у агрегатах або в імунних комплексах. Характерні розходження між цитоплазматичними доменами FcγRII-A і FcγRII-B продукують дві функціонально гетерогенні відповіді на зв'язування з рецептором. Фундаментальна відмінність полягає в тому, що ізоформа A ініціює внутрішньоклітинну передачу сигналу, що приводить до активації клітин, такої як фагоцитоз і "спалах" дихальної активності клітин, а ізоформа B ініціює передачу інгібуючих сигналів, наприклад, інгібування активації В-клітин. FcγRIII (CD16) Через гетерогенність усередині цього класу, розмір FсγRIII у мишей і людини становить від 40 до 80 кДа. Два людські гени кодують два транскрипти, FсγRIIIА, інтегральний мембранний глікопротеїн, і FсγRIIIB, тобто варіант, зв'язаний із глікозилфосфатидил-інозитом (GPI). Один мишачий ген кодує FсγRIII, гомологічний трансмембранному людському FсγRIIIA. FсγRIII має спільні структурні властивості з двома іншими FсγR. Подібно до FcγRII, FсγRIII зв'язується з IgG з низькою афінністю і містить два відповідні позаклітинні Ig-подібні домени. FсγRIIIA експресується в макрофагах і в тучних клітинах, і тільки один FсγR експресується в NK-клітинах. В даний час відомо, що GPI-зв'язаний FсγRIIIB експресується тільки в людських нейтрофілах. Передача сигналу за допомогою FсγR Сигнали активації і інгібування передаються за допомогою FсγR після їхнього зв'язування. Ці діаметрально протилежні функції обумовлені структурними розходженнями у рецепторів різних ізоформ. Два окремі домени в доменах, що передають цитоплазматичний сигнал рецептора, називаються імунорецепторними мотивами активації на основі тирозину (ITAM) або імунорецепторними мотивами інгібування на основі тирозину (ITIM) залежно від різних відповідей. Рекрутинг різних цитоплазматичних ферментів у ці структури залежить від типу FсγR-опосередкованих клітинних відповідей. ITAM-вмісні FсγR-комплекси включають FсγRI, FcγRIIA, FсγRIIIA, а ITIM-вмісні комплекси включають тільки FcγRIIB. Людські нейтрофіли експресують ген FсγRIIA. Кластеризація FсγRIIA за допомогою перехресного зв'язування з імунними комплексами або зі специфічним антитілом приводить до утворення агрегатів ITAM разом з кіназами, асоційованими з рецептором, що тим самим полегшує фосфорилування ITAM. Фосфорилування ITAM служить для заякорювання сайту кінази Syk, активація якої приводить до активації субстратів для наступного каскаду реакцій (наприклад, PI3K). Активація клітин приводить до вивільнення медіаторів прозапальної відповіді. Ген FсγRIIB експресується на B-лімфоцитах; і його позаклітинний домен на 96 % ідентичний FсγRIIA і зв'язується з IgG-комплексами незалежно від умов зв'язування. Присутність ITIM у цитоплазматичному домені FсγRIIB дозволяє визначити, до якого інгібуючого підкласу належить FсγR. Нещодавно був установлений механізм такого інгібування на молекулярному рівні. При зв'язуванні з активуючим FсγR ITIM в FсγRIIB стає фосфорилованим і контактує з SH2-доменом інозитполіфосфат-5’-фосфатази (SHIP), яка гідролізує фосфоінозитні месенджери, які вивільняються в результаті активації тирозинкінази, опосередковуваної ITAM-вмісним FсγR, що, ++ у свою чергу, запобігає припливу внутрішньоклітинного Ca . Таким чином, перехресне зв'язування FсγRIIB послаблює активуючу відповідь на зв'язування з FсγR і інгібує клітинну чутливість. У результаті цього припиняється активація В-клітин, проліферація B-клітин і секреція антитіл. 50 3 UA 111612 C2 Таблиця 1 Рецептори для Fc-ділянок ізотипів імуноглобуліну Рецептор FcγRI(CD64) Зв'язування IgG1 6 -1 10 M FcγRII-А (CD32) IgG1 6 -1 2×10 M FcγRII-В2 (CD32) IgG1 6 -1 2×10 M FcγRII-В1 (CD32) IgG1 6 -1 2×10 M IgG1 5 -1 5×10 M FсγRIII (CD16) FсεRI FсαRI (CD89) 5 10 15 20 25 IgЕ 10 -1 10 M IgА1, IgА2 7 -1 10 M Тип клітин Макрофаги Нейтрофіли Еозинофіли Дендритні клітини Макрофаги Нейтрофіли Еозинофіли Дендритні клітини Тромбоцити Клітини Лангерганса Макрофаги Нейтрофіли Еозинофіли В-клітини Гладкі клітини NK-клітини Еозинофіли Макрофаги Нейтрофіли Гладкі клітини Гладкі клітини Еозинофіли Базофіли Макрофаги Нейтрофіли Еозинофіли Ефект лігування Підвищення стимуляції Активація "респіраторного спалаху" клітин Індукування цитолізу Підвищення рівня вивільнення гранул Підвищення рівня інгібування стимуляції Відсутність підвищення рівня інгібування стимуляції Індукування цитолізу Секреція гранул Підвищення індукування цитолізу 3. Опис суті винаходу Даний винахід, зокрема, стосується ковалентно зв'язаних діатіл і/або молекул ковалентно зв'язаних діатіл, і їхнього застосування для лікування ряду захворювань і розладів, включаючи рак, аутоімунні розлади, алергійні розлади й інфекційні захворювання, які викликаються бактеріями, грибами або вірусами. Переважно, діатіло відповідно до винаходу може зв'язуватися з двома різними епітопами на двох різних клітинах, де перший епітоп експресується на клітині, що належить до типу, який відрізняється від типу клітини, на якій експресується другий епітоп, у результаті чого таке діатіло може зв'язувати разом дві клітини. Більш конкретно, даний винахід стосується поліпептиду (а зокрема, антигензв'язувальної молекули), що містить поліпептидний ланцюг, який включає частину білка, який зв'язується з деімунізованою сироваткою, здатну зв'язуватися з вказаним сироватковим білком, де вказана частина білка, що зв'язується із сироваткою, збільшує час напівжиття вказаного поліпептиду в сироватці в порівнянні з часом напівжиття вказаного поліпептиду в сироватці, у якому відсутня вказана частина вказаного білка, який зв'язується з деімунізованою сироваткою. Даний винахід також стосується застосування (або способів застосування) такої поліпептидної частини білка, який зв'язується з деімунізованою сироваткою, з метою збільшення часу напівжиття поліпептиду, ковалентно зв'язаного з цим білком, у сироватці, де час напівжиття вказаного поліпептиду в сироватці перевищує час напівжиття антигензв'язувальної молекули в сироватці, у якій відсутній альбумін-зв'язувальний білок. Крім того, даний винахід стосується варіанта вищеописаного поліпептиду або його застосування, де вказаний поліпептидний ланцюг містить додаткову частину білка, який зв'язується з деімунізованою сироваткою, і де частини білка, який зв'язується з деімунізованою сироваткою, мають здатність зв'язуватися із сироватковим білком. Даний винахід, зокрема, стосується варіанта вищеописаних поліпептидів або їхнього застосування, де вказана поліпептидна частина білка, який зв'язується з деімунізованою сироваткою, являє собою альбумін-зв'язувальний домен (ABD) стрептококового білка G, або його альбумін-зв'язувальний варіант, а зокрема, де вказаний альбумін-зв'язувальний домен 4 UA 111612 C2 5 10 15 20 25 30 35 40 45 50 55 60 (ABD) стрептококового білка G являє собою альбумін-зв'язувальний домен 3 (ABD3) білка G стрептококового штаму G148. Даний винахід, зокрема, стосується варіанта вищеописаних поліпептидів або їхнього застосування, де вказаний альбумін-зв'язувальний домен містить: (A) заміну валіну на аланін у положенні 71, заміну валіну на аланін у положенні 74 або заміну аспартату на аланін в положенні 79; (B) заміну лейцину на аланін у положенні 64, заміну ізолейцину на аланін у положенні 65 і заміну валіну на аланін у положенні 71; або (C) заміну аспарагіну на аспарагінову кислоту в положенні 66, заміну треоніну на серин у положенні 70 і заміну валіну на аланін у положенні 71. Даний винахід також стосується варіанта вищеописаних поліпептидів або їхнього застосування, де вказаний альбумін-зв'язувальний домен має послідовність SEQ ID NO: 304, SEQ ID NO: 323, SEQ ID NO: 324, SEQ ID NO: 325, SEQ ID NO: 326, SEQ ID NO: 327, SEQ ID NO: 328 або SEQ ID NO: 329. Даний винахід також стосується варіанта вищеописаних антигензв'язувальних молекул або їхнього застосування, де вказаною антигензв'язувальною молекулою є антигензв'язувальна молекула. Даний винахід також стосується варіанта, у якому антигензв'язувальною молекулою є діатіло, що складається щонайменше з першого і другого поліпептидних ланцюгів, які взаємодіють один з одним з утворенням двох антигензв'язувальних сайтів, де щонайменше один з поліпептидних ланцюгів містить другу поліпептидну частину, що включає частину білка, який зв'язується з деімунізованою сироваткою, здатну зв'язуватися із сироватковим білком. Даний винахід також стосується варіантів таких діатіл, у яких перший і другий поліпептидні ланцюги включають другі поліпептидні частини, що містять частину білка, який зв'язується з деімунізованою сироваткою, здатну зв'язуватися із сироватковим білком, і/або де перший і другий поліпептидні ланцюги ковалентно зв'язані один з одним. Даний винахід, зокрема, стосується варіанта вищеописаних діатіл або їхнього застосування, де вказане діатіло зв'язується: (A) з рецептором групи 2D (NKG2D) природних кілерів або з Т-клітинним рецептором (TCR); і (B) з пухлиноасоційованим антигеном. Даний винахід, зокрема, стосується варіантів усіх вищеописаних діатіл або їхнього застосування, де вказаним антигеном є антиген раку молочної залози, антиген раку яєчника, антиген раку передміхурової залози, антиген раку шийки матки, антиген карциноми підшлункової залози, антиген раку легень, антиген раку сечового міхура, антиген раку товстої кишки, антиген раку яєчок, антиген гліобластоми, антиген, асоційований зі злоякісною пухлиною В-клітин; антиген, асоційований із множинною мієломою; антиген, асоційований з неходжкінською лімфомою, або антиген, асоційований із хронічним лімфоцитарним лейкозом. В одному зі своїх варіантів даний винахід стосується ковалентно зв'язаного біспецифічного діатіла, що містить перший і другий поліпептидні ланцюги, де вказаний перший поліпептидний ланцюг включає (і) перший домен, який містить зв'язувальну ділянку варіабельного домену легкого ланцюга першого імуноглобуліну (VL1), специфічного до першого епітопу, (ii) другий домен, який містить зв'язувальну ділянку варіабельного домену важкого ланцюга другого імуноглобуліну (VН2), специфічного до другого епітопу, і необов'язково (iii) третій домен, який містить щонайменше один цистеїновий залишок, де перший і другий домени ковалентно зв'язані один з одним так, що вказані перший і другий домени не утворюють епітопзв'язувальний сайт; а вказаний другий поліпептидний ланцюг включає (i) четвертий домен, який містить зв'язувальну ділянку варіабельного домену легкого ланцюга другого імуноглобуліну (VL2), (ii) п'ятий домен, який містить зв'язувальну ділянку варіабельного домену важкого ланцюга першого імуноглобуліну (VН1), і необов'язково (iii) шостий домен, який містить щонайменше один цистеїновий залишок, де вказані четвертий і п'ятий домени ковалентно зв'язані один з одним так, що вказані четвертий і п'ятий домени не утворюють епітопзв'язувальний сайт; і де вказані перший поліпептидний ланцюг і другий поліпептидний ланцюг ковалентно зв'язані один з одним за умови, що ковалентний зв'язок не є пептидним зв'язком; де перший домен і п'ятий домен утворюють перший зв'язувальний сайт (VL1)(VH1), який зв'язується з першим епітопом, а другий домен і четвертий домен утворюють другий зв'язувальний сайт (VL2)(VH2), що зв'язується з другим епітопом. В іншому своєму варіанті даний винахід стосується ковалентно зв'язаного біспецифічного діатіла, що містить перший і другий поліпептидні ланцюги, де вказаний перший поліпептидний ланцюг включає (i) перший домен, який містить зв'язувальну ділянку варіабельного домену легкого ланцюга першого імуноглобуліну (VL1), специфічного до першого епітопу, (ii) другий домен, який містить зв'язувальну ділянку варіабельного домену важкого ланцюга другого 5 UA 111612 C2 5 10 15 20 25 30 35 40 45 50 55 60 імуноглобуліну (VН2), специфічного до другого епітопу, і (iii) третій домен, який містить Fcдомен або його частину, де вказані перший і другий домени ковалентно зв'язані один з одним так, що вказані перший і другий домени не утворюють епітоп-зв'язувальний сайт; а вказаний другий поліпептидний ланцюг включає (i) четвертий домен, який містить зв'язувальну ділянку варіабельного домену легкого ланцюга другого імуноглобуліну (VL2), (ii) п'ятий домен, який містить зв'язувальну ділянку варіабельного домену важкого ланцюга першого імуноглобуліну (VН1), де вказані четвертий і п'ятий домени ковалентно зв'язані один з одним так, що вказані третій і четвертий домени не утворюють епітоп-зв'язувальний сайт; і де вказані перший поліпептидний ланцюг і другий поліпептидний ланцюг ковалентно зв'язані один з одним за умови, що ковалентний зв'язок не є пептидним зв'язком, де перший домен і п'ятий домен утворюють перший зв'язувальний сайт (VL1)(VH1), що зв'язується з першим епітопом, а другий домен і четвертий домен утворюють другий зв'язувальний сайт (VL2)(VH2), що зв'язується з другим епітопом. У деяких своїх аспектах даний винахід стосується молекули діатіла, що містить перший і другий поліпептидні ланцюги, де вказаний перший поліпептидний ланцюг включає (i) перший домен, який містить зв'язувальну ділянку варіабельного домену легкого ланцюга першого імуноглобуліну (VL1), специфічного до першого епітопу, (ii) другий домен, який містить зв'язувальну ділянку варіабельного домену важкого ланцюга другого імуноглобуліну (VН2), специфічного до другого епітопу, і (iii) третій домен, який містить Fc-домен або його частину, де вказані перший і другий домени ковалентно зв'язані один з одним так, що вказані перший і другий домени не утворюють епітоп-зв'язувальний сайт; а вказаний другий поліпептидний ланцюг включає (i) четвертий домен, який містить зв'язувальну ділянку варіабельного домену легкого ланцюга другого імуноглобуліну (VL2), (ii) п'ятий домен, який містить зв'язувальну ділянку варіабельного домену важкого ланцюга першого імуноглобуліну (VН1), і (iii) шостий домен, який містить щонайменше один цистеїновий залишок, де вказані четвертий і п'ятий домени ковалентно зв'язані один з одним так, що вказані четвертий і п'ятий домени не утворюють епітоп-зв'язувальний сайт; і де вказані перший поліпептидний ланцюг і другий поліпептидний ланцюг ковалентно зв'язані один з одним за умови, що ковалентний зв'язок не є пептидним зв'язком; де вказані перший домен і п'ятий домен утворюють перший зв'язувальний сайт (VL1)(VH1), що зв'язується з першим епітопом, а другий домен і четвертий домен утворюють другий зв'язувальний сайт (VL2)(VH2), що зв'язується з другим епітопом. У деяких своїх варіантах даний винахід стосується ковалентно зв'язаного біспецифічного діатіла, що являє собою димер молекул діатіла, де кожна молекула діатіла містить перший і другий поліпептидні ланцюги, де вказаний перший поліпептидний ланцюг включає (i) перший домен, який містить зв'язувальну ділянку варіабельного домену легкого ланцюга першого імуноглобуліну (VL1), специфічного до першого епітопу, (ii) другий домен, який містить зв'язувальну ділянку варіабельного домену важкого ланцюга другого імуноглобуліну (VН2), специфічного до другого епітопу, і (iii) третій домен, який містить Fc-домен або його частину, де вказані перший і другий домени ковалентно зв'язані один з одним так, що вони не утворюють епітоп-зв'язувальний сайт; а вказаний другий поліпептидний ланцюг включає (i) четвертий домен, який містить зв'язувальну ділянку варіабельного домену легкого ланцюга другого імуноглобуліну (VL2), (ii) п'ятий домен, який містить зв'язувальну ділянку варіабельного домену важкого ланцюга першого імуноглобуліну (VН1), і (iii) шостий домен, який містить щонайменше один цистеїновий залишок, де вказані четвертий і п'ятий домени ковалентно зв'язані один з одним так, що вказані четвертий і п'ятий домени не утворюють епітоп-зв'язувальний сайт; і де вказані перший поліпептидний ланцюг і другий поліпептидний ланцюг кожної молекули діатіла ковалентно зв'язані один з одним за умови, що ковалентний зв'язок не є пептидним зв'язком; де вказані перший домен і п'ятий домен кожної молекули діатіла утворюють перший зв'язувальний сайт (VL1)(VH1), що зв'язується з першим епітопом, а другий домен і четвертий домен кожної молекули діатіла утворюють другий зв'язувальний сайт (VL2)(VH2), що зв'язується з другим епітопом. В інших своїх варіантах даний винахід стосується ковалентно зв'язаного тетраспецифічного діатіла, який являє собою димер молекул діатіла, де перша молекула діатіла містить перший і другий поліпептидні ланцюги, де вказаний перший поліпептидний ланцюг включає (i) перший домен, який містить зв'язувальну ділянку варіабельного домену легкого ланцюга першого імуноглобуліну (VL1), специфічного до першого епітопу, (ii) другий домен, який містить зв'язувальну ділянку варіабельного домену важкого ланцюга другого імуноглобуліну (VН2), специфічного до другого епітопу, і (iii) третій домен, який містить Fc-домен або його частину, де вказані перший і другий домени ковалентно зв'язані один з одним так, що вони не утворюють епітоп-зв'язувальний сайт; а вказаний другий поліпептидний ланцюг включає (i) четвертий 6 UA 111612 C2 5 10 15 20 25 30 35 40 45 50 55 60 домен, який містить зв'язувальну ділянку варіабельного домену легкого ланцюга другого імуноглобуліну (VL2), (ii) п'ятий домен, який містить зв'язувальну ділянку варіабельного домену важкого ланцюга першого імуноглобуліну (VН1), і (iii) шостий домен, який містить щонайменше один цистеїновий залишок, де вказані четвертий і п'ятий домени ковалентно зв'язані один з одним так, що вказані четвертий і п'ятий домени не утворюють епітоп-зв'язувальний сайт; і де вказані перший поліпептидний ланцюг і другий поліпептидний ланцюг діатіла ковалентно зв'язані один з одним за умови, що ковалентний зв'язок не є пептидним зв'язком; де вказані перший домен і п'ятий домен утворюють перший зв'язувальний сайт (VL1)(VH1), що зв'язується з першим епітопом, а другий домен і четвертий домен утворюють другий зв'язувальний сайт (VL2)(VH2), що зв'язується з другим епітопом; а друга молекула діатіла містить перший і другий поліпептидні ланцюги, де вказаний перший поліпептидний ланцюг включає (i) перший домен, який містить зв'язувальну ділянку варіабельного домену легкого ланцюга третього імуноглобуліну (VL3), специфічного до третього епітопу, (ii) другий домен, який містить зв'язувальну ділянку варіабельного домену важкого ланцюга четвертого імуноглобуліну (VН4), специфічного до четвертого епітопу, і (iii) третій домен, який містить Fc-домен або його частину, де вказані перший і другий домени ковалентно зв'язані один з одним так, що вони не утворюють епітоп-зв'язувальний сайт; а вказаний другий поліпептидний ланцюг включає (i) четвертий домен, який містить зв'язувальну ділянку варіабельного домену легкого ланцюга четвертого імуноглобуліну (VL4), (ii) п'ятий домен, який містить зв'язувальну ділянку варіабельного домену важкого ланцюга третього імуноглобуліну (VН3), і (iii) шостий домен, який містить щонайменше один цистеїновий залишок, де вказані четвертий і п'ятий домени ковалентно зв'язані один з одним так, що вказані четвертий і п'ятий домени не утворюють епітоп-зв'язувальний сайт; і де вказані перший поліпептидний ланцюг і другий поліпептидний ланцюг діатіла ковалентно зв'язано один з одним за умови, що ковалентний зв'язок не є пептидним зв'язком; де вказані перший домен і п'ятий домен утворюють перший зв'язувальний сайт (VL3)(VH3), що зв'язується з третім епітопом, а другий домен і четвертий домен утворюють другий зв'язувальний сайт (VL4)(VH4), що зв'язується з четвертим епітопом. У деяких аспектах винаходу перший епітоп, другий епітоп, а також, якщо вони присутні, третій епітоп і четвертий епітоп, можуть бути однаковими. В інших аспектах винаходу перший епітоп, другий епітоп, а також, якщо вони присутні, третій епітоп і четвертий епітоп, можуть відрізнятися один від одного. У деяких аспектах винаходу, які включають домен, який зв'язується з третім епітопом, перший епітоп і третій епітоп можуть бути однаковими. У деяких аспектах винаходу, які включають домен, який зв'язується з четвертим епітопом, перший епітоп і четвертий епітоп можуть бути однаковими. У деяких аспектах винаходу, які включають домен, що зв'язується з третім епітопом, другий епітоп і третій епітоп можуть бути однаковими. У деяких аспектах винаходу, які включають домен, що зв'язується з четвертим епітопом, другий епітоп і четвертий епітоп можуть бути однаковими. У переважних аспектах винаходу перший епітоп і другий епітоп є різними. В інших аспектах винаходу, які включають домен, що зв'язується з третім епітопом, і домен, що зв'язується з четвертим епітопом, третій епітоп і четвертий епітоп можуть бути різними. Слід зазначити, що кожна з вищевказаних комбінацій входить в обсяг даного винаходу. У конкретних аспектах винаходу перший домен і п'ятий домен діатіла або молекули діатіла можуть походити від того самого імуноглобуліну. В іншому аспекті винаходу другий домен і четвертий домен діатіла або молекули діатіла можуть походити від того самого імуноглобуліну. В іншому аспекті винаходу перший домен і п'ятий домен діатіла або молекули діатіла можуть походити від іншого імуноглобуліну. В іншому аспекті винаходу другий домен і четвертий домен діатіла або молекули діатіла можуть походити від іншого імуноглобуліну. Слід зазначити, що кожна з вищевказаних комбінацій входить в обсяг даного винаходу. У деяких аспектах винаходу ковалентний зв'язок між першим поліпептидним ланцюгом і другим поліпептидним ланцюгом діатіла або молекули діатіла може бути утворений за допомогою дисульфідного зв'язку між щонайменше одним цистеїновим залишком на першому поліпептидному ланцюзі і щонайменше одним цистеїновим залишком на другому поліпептидному ланцюзі. Цистеїнові залишки на першому або другому поліпептидних ланцюгах, відповідальні за утворення дисульфідного зв'язку, можуть бути присутнім на будь-якій ділянці поліпептидного ланцюга, включаючи перший, другий, третій, четвертий, п'ятий і шостий домени. У конкретному варіанті винаходу цистеїновий залишок на першому поліпептидному ланцюзі присутній в першому домені, а цистеїновий залишок на другому поліпептидному ланцюзі присутній в п'ятому домені. Перший, другий, четвертий і п'ятий домени, що відповідають варіабельним ділянкам, відповідальні за зв'язування. У переважних варіантах винаходу цистеїнові залишки, відповідальні за утворення дисульфідних зв'язків між першим і другим 7 UA 111612 C2 5 10 15 20 25 30 35 40 45 50 55 60 поліпептидними ланцюгами, розташовані в третьому і шостому доменах, відповідно. У конкретному аспекті цього варіанта третій домен першого поліпептидного ланцюга містить 6 Cкінцевих амінокислот людського легкого ланцюга каппа, FNRGEC (SEQ ID NO: 23), що можуть входити в амінокислотну послідовність (SEQ ID NO: 17). В іншому аспекті цього варіанта шостий домен другого поліпептидного ланцюга містить 6 C-кінцевих амінокислот людського легкого ланцюга каппа, FNRGEC (SEQ ID NO: 23), що можуть входити в амінокислотну послідовність (SEQ ID NO: 17). У ще одному аспекті цього варіанта третій домен першого поліпептидного ланцюга містить амінокислотну послідовність VEPKSC (SEQ ID NO: 77), яка походить від шарнірного домену людського IgG і може кодуватися нуклеотидною послідовністю (SEQ ID NO: 78). В іншому аспекті цього варіанта шостий домен другого поліпептидного ланцюга містить амінокислотну послідовність VEPKSC (SEQ ID NO: 77), яка походить від шарнірного домену людського IgG і може кодуватися нуклеотидною послідовністю (SEQ ID NO: 78). У деяких аспектах цього варіанта третій домен першого поліпептидного ланцюга містить 6 C-кінцевих амінокислот людського легкого ланцюга каппа, FNRGEC (SEQ ID NO: 23), а шостий домен другого поліпептидного ланцюга містить амінокислотну послідовність VEPKSC (SEQ ID NO: 77). В інших аспектах цього варіанта шостий домен другого поліпептидного ланцюга містить 6 Cкінцевих амінокислот людського легкого ланцюга каппа, FNRGEC (SEQ ID NO: 23), а третій домен першого поліпептидного ланцюга містить амінокислотну послідовність VEPKSC (SEQ ID NO: 77). В інших аспектах цього варіанта третій домен першого поліпептидного ланцюга містить 6 C-кінцевих амінокислот людського легкого ланцюга каппа, FNRGEC (SEQ ID NO: 23), а шостий домен другого поліпептидного ланцюга містить шарнірний домен. В інших аспектах цього варіанта шостий домен другого поліпептидного ланцюга містить 6 C-кінцевих амінокислот людського легкого ланцюга каппа, FNRGEC (SEQ ID NO: 23), а третій домен першого поліпептидного ланцюга містить шарнірний домен. В інших аспектах цього варіанта третій домен першого поліпептидного ланцюга містить 6 C-кінцевих амінокислот людського легкого ланцюга каппа, FNRGEC (SEQ ID NO: 23), а шостий домен першого поліпептидного ланцюга містить Fc-домен або його частину. В інших аспектах цього варіанта шостий домен другого поліпептидного ланцюга містить 6 C-кінцевих амінокислот людського легкого ланцюга каппа, FNRGEC (SEQ ID NO: 23), а третій домен першого поліпептидного ланцюга містить Fc-домен або його частину. В інших варіантах винаходу цистеїнові залишки на першому або другому поліпептиді, відповідальні за утворення дисульфідного зв'язку, можуть бути розташовані за межами першого, другого або третього доменів на першому поліпептидному ланцюзі і за межами четвертого, п'ятого і шостого доменів на другому поліпептидному ланцюзі. Зокрема, цистеїновий залишок на першому поліпептидному ланцюзі може бути N-кінцевим відносно першого домену або С-кінцевим відносно першого домену. Цистеїновий залишок на першому поліпептидному ланцюзі може бути N-кінцевим відносно другого домену або С-кінцевим відносно другого домену. Цистеїновий залишок на першому поліпептидному ланцюзі може бути N-кінцевим відносно третього домену або С-кінцевим відносно третього домену. Крім того, цистеїновий залишок на другому поліпептидному ланцюзі може бути N-кінцевим відносно четвертого домену або С-кінцевим відносно четвертого домену. Цистеїновий залишок на другому поліпептидному ланцюзі може бути N-кінцевим відносно п'ятого домену або С-кінцевим відносно п'ятого домену. Відповідно до цього цистеїновий залишок на другому поліпептидному ланцюзі може бути С-кінцевим відносно шостого домену або N-кінцевим відносно шостого домену. У конкретному аспекті винаходу дисульфідний зв'язок може бути розташований щонайменше між двома цистеїновими залишками на першому поліпептидному ланцюзі і щонайменше між двома цистеїновими залишками на другому поліпептидному ланцюзі. У конкретному аспекті винаходу, де третій домен і шостий домен не містять Fc-домен або його частини, цистеїновий залишок може знаходитися в C-кінця першого поліпептидного ланцюга й біля С-кінця другого поліпептидного ланцюга. Слід зазначити, що кожна з вищевказаних комбінацій входить в обсяг даного винаходу. У конкретних варіантах даного винаходу, описаних вище, ковалентно зв'язане діатіло відповідно до винаходу включає димери молекул діатіл, де кожна молекула діатіла містить перший і другий поліпептидні ланцюги. У деяких аспектах цього варіанта молекули діатіла можуть бути ковалентно зв'язані один з одним з утворенням димера, за умови, що ковалентний зв'язок не є пептидним зв'язком. У переважних аспектах цього варіанта ковалентним зв'язком є дисульфідний зв'язок, розташований щонайменше між одним цистеїновим залишком на першому поліпептидному ланцюзі і щонайменше одним цистеїновим залишком кожного димера молекул діатіла. У ще більш переважних аспектах винаходу ковалентним зв'язком є дисульфідний зв'язок, розташований щонайменше між одним цистеїновим залишком на 8 UA 111612 C2 5 10 15 20 25 30 35 40 45 50 55 60 першому поліпептидному ланцюзі і щонайменше одним цистеїновим залишком кожної молекули діатіла, що утворює димер, де вказаний щонайменше один цистеїновий залишок присутній у третьому домені кожного першого поліпептидного ланцюга. У деяких аспектах винаходу перший домен на першому поліпептидному ланцюзі може бути N-кінцевим відносно другого домену або С-кінцевим відносно другого домену. Перший домен на першому поліпептидному ланцюзі може бути N-кінцевим відносно третього домену або Скінцевим відносно третього домену. Другий домен на першому поліпептидному ланцюзі може бути N-кінцевим відносно першого домену або С-кінцевим відносно першого домену. Крім того, другий домен на першому поліпептидному ланцюзі може бути N-кінцевим відносно третього домену або С-кінцевим відносно третього домену. Відповідно до цього третій домен на першому поліпептидному ланцюзі може бути N-кінцевим відносно першого домену або С-кінцевим відносно першого домену. Третій домен на першому поліпептидному ланцюзі може бути Nкінцевим відносно другого домену або С-кінцевим відносно другого домену. Що стосується другого поліпептидного ланцюга, то четвертий домен може бути N-кінцевим відносно п'ятого домену або С-кінцевим відносно п'ятого домену. Четвертий домен може бути N-кінцевим відносно шостого домену або С-кінцевим відносно шостого домену. П'ятий домен на другому поліпептидному ланцюзі може бути N-кінцевим відносно четвертого домену або С-кінцевим відносно четвертого домену. П'ятий домен на другому поліпептидному ланцюзі може бути Nкінцевим відносно шостого домену або С-кінцевим відносно шостого домену. Відповідно до цього шостий домен на другому поліпептидному ланцюзі може бути N-кінцевим відносно четвертого домену або С-кінцевим відносно четвертого домену. Шостий домен на другому поліпептидному ланцюзі може бути N-кінцевим відносно п'ятого домену або С-кінцевим відносно п'ятого домену. Слід зазначити, що кожна з вищевказаних комбінацій входить в обсяг даного винаходу. У деяких варіантах винаходу перший домен і другий домен можуть знаходитися з боку Скінця відносно третього домену на першому поліпептидному ланцюзі, або перший домен і другий домен можуть знаходитися з боку N-кінця відносно третього домену на першому поліпептидному ланцюзі. Що стосується другого поліпептидного ланцюга, то четвертий домен і п'ятий домен можуть знаходитися з боку С-кінця відносно шостого домену, або четвертий домен і п'ятий домен можуть знаходитися з боку N-кінця відносно шостого домену. У деяких своїх аспектах цього варіанта, даний винахід стосується ковалентно зв'язаного біспецифічного діатіла, що являє собою димер молекул діатіла, кожна з яких містить перший і другий поліпептидні ланцюги, де вказаний перший поліпептидний ланцюг включає (i) перший домен, який містить зв'язувальну ділянку варіабельного домену легкого ланцюга першого імуноглобуліну (VL1), специфічного до першого епітопу, (ii) другий домен, який містить зв'язувальну ділянку варіабельного домену важкого ланцюга другого імуноглобуліну (VН2), специфічного до другого епітопу, і (iii) третій домен, який містить Fc-домен або його частину, де вказані перший і другий домени ковалентно зв'язані один з одним так, що вказані перший і другий домени не утворюють епітоп-зв'язувальний сайт, і де третій домен знаходиться з боку Nкінця відносно першого домену і другого домену, а вказаний другий поліпептидний ланцюг включає (i) четвертий домен, який містить зв'язувальну ділянку варіабельного домену легкого ланцюга другого імуноглобуліну (VL2), (ii) п'ятий домен, який містить зв'язувальну ділянку варіабельного домену важкого ланцюга першого імуноглобуліну (VН1), і (iii) шостий домен, який містить щонайменше один цистеїновий залишок, де вказані четвертий і п'ятий домени ковалентно зв'язані один з одним так, що вказані четвертий і п'ятий домени не утворюють епітоп-зв'язувальний сайт; і де вказані перший поліпептидний ланцюг і другий поліпептидний ланцюг кожної молекули діатіла ковалентно зв'язані один з одним за умови, що ковалентний зв'язок не є пептидним зв'язком; де вказані перший домен і п'ятий домен кожної молекули діатіла утворюють перший зв'язувальний сайт (VL1)(VH1), що зв'язується з першим епітопом, а другий домен і четвертий домен кожної молекули діатіла утворюють другий зв'язувальний сайт (VL2)(VH2), що зв'язується з другим епітопом. В іншому своєму варіанті даний винахід стосується ковалентно зв'язаного тетраспецифічного діатіла, що являє собою димер молекул діатіла, де перша молекула діатіла містить перший і другий поліпептидні ланцюги, де вказаний перший поліпептидний ланцюг включає (i) перший домен, який містить зв'язувальну ділянку варіабельного домену легкого ланцюга першого імуноглобуліну (VL1), специфічного до першого епітопу, (ii) другий домен, який містить зв'язувальну ділянку варіабельного домену важкого ланцюга другого імуноглобуліну (VН2), специфічного до другого епітопу, і (iii) третій домен, який містить Fc-домен або його частину, де вказані перший і другий домени ковалентно зв'язані один з одним так, що вони не утворюють епітоп-зв'язувальний сайт; і де третій домен знаходиться з боку N-кінця відносно 9 UA 111612 C2 5 10 15 20 25 30 35 40 45 50 55 60 першого домену і другого домену, а вказаний другий поліпептидний ланцюг включає (i) четвертий домен, який містить зв'язувальну ділянку варіабельного домену легкого ланцюга другого імуноглобуліну (VL2), (ii) п'ятий домен, який містить зв'язувальну ділянку варіабельного домену важкого ланцюга першого імуноглобуліну (VН1), і (iii) шостий домен, який містить щонайменше один цистеїновий залишок, де вказані четвертий і п'ятий домени ковалентно зв'язані один з одним так, що вказані четвертий і п'ятий домени не утворюють епітопзв'язувальний сайт; і де вказані перший поліпептидний ланцюг і другий поліпептидний ланцюг діатіла ковалентно зв'язані один з одним за умови, що вказаний ковалентний зв'язок не є пептидним зв'язком; де вказані перший домен і п'ятий домен утворюють перший зв'язувальний сайт (VL1)(VH1), що зв'язується з першим епітопом, а другий домен і четвертий домен утворюють другий зв'язувальний сайт (VL2)(VH2), що зв'язується з другим епітопом; де друга молекула діатіла містить перший і другий поліпептидний ланцюг, і де вказаний перший поліпептидний ланцюг включає (i) перший домен, який містить зв'язувальну ділянку варіабельного домену легкого ланцюга третього імуноглобуліну (VL3), специфічного до третього епітопу, (ii) другий домен, який містить зв'язувальну ділянку варіабельного домену важкого ланцюга четвертого імуноглобуліну (VН4), специфічного до четвертого епітопу, і (iii) третій домен, який містить Fc-домен або його частину, де вказані перший і другий домени ковалентно зв'язані один з одним так, що вказані перший і другий домени не утворюють епітопзв'язувальний сайт; і де третій домен знаходиться з боку N-кінця відносно першого домену і другого домену, а вказаний другий поліпептидний ланцюг включає (i) четвертий домен, який містить зв'язувальну ділянку варіабельного домену легкого ланцюга четвертого імуноглобуліну (VL4), (ii) п'ятий домен, який містить зв'язувальну ділянку варіабельного домену важкого ланцюга третього імуноглобуліну (VН3), і (iii) шостий домен, який містить щонайменше один цистеїновий залишок, де вказані четвертий і п'ятий домени ковалентно зв'язані один з одним так, що вказані четвертий і п'ятий домени не утворюють епітоп-зв'язувальний сайт; і де вказані перший поліпептидний ланцюг і другий поліпептидний ланцюг діатіла ковалентно зв'язані один з одним за умови, що ковалентний зв'язок не є пептидним зв'язком; де вказані перший домен і п'ятий домен утворюють перший зв'язувальний сайт (VL3)(VH3), що зв'язується з третім епітопом, а другий домен і четвертий домен утворюють другий зв'язувальний сайт (VL4)(VH4), що зв'язується з четвертим епітопом. Як обговорювалося вище, домени на окремих поліпептидних ланцюгах є ковалентно зв'язаними. У конкретних аспектах винаходу ковалентний зв'язок між першим і другим доменом, першим і третім доменом, другим і третім доменом, четвертим і п'ятим доменом, четвертим і шостим доменом і/або п'ятим і шостим доменом може являти собою пептидний зв'язок. Зокрема, перший і другий домени, і четвертий і п'ятий домени можуть бути розділені третіми доменом і шостими доменом, відповідно, або додатковими амінокислотними залишками, за умови, що перший і другий домени, і четвертий і п'ятий домени не утворюють зв'язувальний сайт. Число амінокислотних залишків може становити 0, 1, 2, 3, 4, 5, 6, 7, 8 або 9 амінокислотних залишків. В одному з переважних аспектів винаходу число амінокислотних залишків між доменами дорівнює 8. У деяких аспектах винаходу домени першого і другого поліпептидних ланцюгів, що містять Fc-домен, тобто, необов'язково третій і шостий домени, відповідно, можуть також включати шарнірний домен, який містить шарнірну ділянку-Fc-ділянку. В альтернативних варіантах винаходу перший поліпептидний ланцюг або другий поліпептидний ланцюг можуть містити шарнірний домен, що також не включає Fc-домен. Важкі ланцюги, легкі ланцюги, шарнірні ділянки, Fc-домени і/або шарнірні ділянки-Fc-домени, використовувані в даному винаході, можуть походити від імуноглобуліну будь-якого типу, включаючи IgA, IgD, IgE, IgG або IgM. У переважному аспекті винаходу імуноглобулін належить до типу IgG, або до будь-якого його підтипу, тобто IgG1, IgG2, IgG3 або IgG4. В інших аспектах винаходу імуноглобулін, від якого походять легкі і важкі ланцюги, є гуманізованим або химерним. Крім того, перший і другий епітопи, і, якщо вони присутні, то третій і четвертий епітопи, з якими зв'язуються діатіло або молекула діатіла, можуть являти собою різні епітопи того самого антигену або різні епітопи різних антигенів. Антигени можуть являти собою будь-яку молекулу, проти якої може вироблятися антитіло. Так, наприклад, такими молекулами є білки, нуклеїнові кислоти, бактеріальні токсини, маркери клітинної поверхні, аутоімунні маркери, вірусні білки, лікарські засоби і т. п. У конкретних аспектах винаходу щонайменше один епітоп-зв'язувальний сайт діатіла є специфічним до антигену на конкретній клітині, такій як B-клітина, T-клітина, фагоцит, природна клітина-кілер (NK) або дендритна клітина. У деяких аспектах варіанта даного винаходу, щонайменше один епітоп-зв'язувальний сайт діатіла або молекули діатіла є специфічним до Fc-рецептора, де вказаний Fc-рецептор може 10 UA 111612 C2 5 10 15 20 25 30 35 40 45 50 55 60 являти собою активуючий Fc-рецептор або інгібуючий Fc-рецептор. У конкретних аспектах винаходу Fc-рецептором є Fсγ-рецептор, а Fсγ-рецептором є FсγRI-, FсγRII- або FсγRIIIрецептор. У більш переважних аспектах винаходу FсγRIII-рецептором є рецептор FсγRIIIA (CD16A) або рецептор FсγRIIIB (CD16B), а більш переважно FсγRIII-рецептором є рецептор FсγRIIIA (CD16A). В іншому переважному аспекті винаходу FсγRII-рецептор являє собою рецептор FсγRIIA (CD32A) або рецептор FсγRIIB (CD32B), а більш переважно рецептор FсγRIIB (CD32B). В особливо переважному аспекті винаходу один зв'язувальний сайт діатіла є специфічним до CD32B, а інший зв'язувальний сайт є специфічним до CD16A. У конкретному варіанті винаходу щонайменше один епітоп-зв'язувальний сайт діатіла або молекули діатіла є специфічним до активуючого Fc-рецептора, а щонайменше один інший сайт є специфічним до інгібуючого Fc-рецептора. У деяких аспектах цього варіанта активуючим Fc-рецептором є CD32A, а інгібуючим Fc-рецептором є CD32B. В інших аспектах цього варіанта активуючим Fcрецептором є BCR, а інгібуючим Fс-рецептором є CD32B. В інших аспектах цього варіанта активуючим Fc-рецептором є IgERI, а інгібуючим Fс-рецептором є CD32B. У тих випадках, коли один епітоп-зв'язувальний сайт є специфічним до CD16A, домени VL і VH можуть бути ідентичними або аналогічними доменам VL і VH мишачого антитіла 3G8, послідовність якого була клонована і представлена в даному описі. В інших випадках, коли один епітоп-зв'язувальний сайт є специфічним до CD32A, домени VL і VH можуть бути ідентичними або аналогічними доменам VL і VH мишачого антитіла IV.3. В інших випадках, коли один епітопзв'язувальний сайт є специфічним до CD32В, домени VL і VH можуть бути ідентичними або аналогічними доменам VL і VH мишачого антитіла 2B6, послідовність якого була клонована і представлена в даному описі. Слід зазначити, що будь-які домени VL або VH антитіл 3G8, 2B6 і IV.3 можуть бути використані в будь-якій комбінації. Даний винахід також стосується біспецифічного діатіла або молекули біспецифічного діатіла, де перший епітоп є специфічним до CD32В, а другий епітоп є специфічним до CD16A. В інших аспектах винаходу епітоп-зв'язувальні сайт може бути специфічним до патогенного антигену. Використовуваний тут патогенний антиген являє собою антиген, що викликає конкретне захворювання, яке викликається патогенами, включаючи рак, інфекцію й аутоімунне захворювання. Таким чином, патогенними антигенами можуть бути пухлинний антиген, бактеріальний антиген, вірусний антиген або аутоімунний антиген. Репрезентативними патогенними антигенами є, але не обмежуються ними, ліпополісахарид, вірусні антигени, вибрані з групи, яка складається з вірусних антигенів, які походять від вірусу імунодефіциту людини, аденовірусу, респіраторно-синцитіального вірусу, вірусу Західного Нілу (наприклад, антигени E16 і/або E53) і вірусу гепатиту; нуклеїнові кислоти (ДНК і РНК) і колаген. Переважним патогенним антигеном є нейтралізуючий антиген. У переважному аспекті винаходу, один епітопзв'язувальний сайт є специфічним до CD16A або CD32A, а інший епітоп-зв'язувальний сайт є специфічним до патогенного антигену, за винятком аутоімунних антигенів. В іншому переважному аспекті винаходу один епітоп-зв'язувальний сайт є специфічним до CD32В, а інший епітоп-зв'язувальний сайт є специфічним до будь-якого патогенного антигену. У конкретних варіантах винаходу молекула діатіла відповідно до винаходу зв'язується з двома різними антигенами на одній і тій же клітині, наприклад, один антигензв'язувальний сайт є специфічним до активуючого Fс-рецептору, а інший є специфічним до інгібуючого Fс-рецептору. В інших варіантах винаходу молекула діатіла зв'язується з двома різними віруснейтралізуючими епітопами, наприклад, такими як, але не обмежуються ними, E16 і E53 вірусу Західного Нілу. В іншому варіанті даного винаходу діатіла відповідно до винаходу можуть бути використані для лікування ряду захворювань і розладів. Відповідно до цього, даний винахід стосується способу лікування захворювання або розладу, що включає введення пацієнту, який потребує цього, ефективного кількості ковалентно зв'язаного діатіла або молекули діатіла відповідно до винаходу, у яких щонайменше один зв'язувальний сайт є специфічним до патогенного антигену, такого як антиген, який експресується на поверхні ракових клітин або на поверхні бактерій або віріона, а щонайменше один інший зв'язувальний сайт є специфічним до Fс-рецептору, наприклад, до CD16A. В іншому своєму варіанті даний винахід стосується способу лікування захворювання або розладу, що включає введення пацієнту, який потребує цього, ефективного кількості діатіла або молекули діатіла відповідно до винаходу, у яких щонайменше один зв'язувальний сайт є специфічним до CD32B, а щонайменше один інший зв'язувальний сайт є специфічним до CD16A. В іншому своєму варіанті даний винахід стосується способу індукування імунологічної толерантності до патогенного антигену, де вказаний спосіб включає введення пацієнту, який 11 UA 111612 C2 5 10 15 20 25 30 35 40 45 50 55 60 потребує цього, ефективної кількості ковалентно зв'язаного діатіла або двовалентної молекули діатіла відповідно до винаходу, у яких щонайменше один зв'язувальний сайт є специфічним до CD32B, а щонайменше один інший зв'язувальний сайт є специфічним до вказаного патогенного антигену. В аспектах цього варіанта винаходу патогенним антигеном може бути алерген або інша молекула, до яких було б бажано виробляння імунологічної толерантності, таких як білок, який експресується на трансплантованій тканині. У ще одному своєму варіанті даний винахід стосується способу детоксифікації, де вказаний спосіб включає введення пацієнту, який потребує цього, ефективної кількості ковалентно зв'язаного діатіла або молекули діатіла відповідно до винаходу, у яких щонайменше один зв'язувальний сайт є специфічним до маркера клітинної поверхні, а щонайменше один інший зв'язувальний сайт є специфічним до токсину. У конкретних аспектах винаходу діатілом, що вводиться, відповідно до винаходу є діатіло, у якому один зв'язувальний є специфічним до маркера клітинної поверхні, такого як Fc, а інший зв'язувальний сайт є специфічним до бактеріального токсину або до лікарського засобу. В одному з аспектів винаходу маркер клітинної поверхні не присутній на еритроцитах. Даний винахід також стосується способу збільшення часу напівжиття молекули, що містить антигензв'язувальний домен антитіла, де вказаний спосіб включає зв'язування (наприклад, пряме зв'язування або зв'язування за допомогою кон'югування, приєднання і т. п.) двох або більше альбумін-зв'язувальних доменів з молекулою. Даний винахід, зокрема, стосується варіанта такого способу, у якому зв'язування двох або більше альбумін-зв'язувальних доменів включає приєднання 2, 3, 4, 5, 6, 7, 8, 9, 10 або більше альбумін-зв'язувальних доменів до молекули, а більш конкретно, до варіанта такого способу, у якому зв'язування двох або більше альбумін-зв'язувальних доменів включає приєднання 2 альбумін-зв'язувальних доменів до молекули. Такі два або більше альбумін-зв'язувальних доменів можуть бути однаковими або різними, і переважно вони можуть бути незалежно вибрані з групи, яка складається з послідовностей SEQ ID NO: 304, SEQ ID NO: 323, SEQ ID NO: 324, SEQ ID NO: 325, SEQ ID NO: 326, SEQ ID NO: 327, SEQ ID NO: 328 або SEQ ID NO: 329. Даний винахід, зокрема, стосується варіанта таких способів, у яких два або більше альбумін-зв'язувальних доменів є однаковими, або в яких 2 альбумін-зв'язувальні домени є однаковими. Даний винахід, зокрема, стосується варіанта такого способу, у якому молекулою, що містить антигензв'язувальний домен антитіла, є діатіло. 3.1 Визначення Якщо це не обговорено особливо, то усі використовувані тут спеціальні терміни, примітки й інші наукові терміни або термінологія мають загальноприйняті значення, зрозумілі фахівцю в галузі, до якої належить винахід. У деяких випадках терміни, що мають загальноприйняті значення, приводяться тут для зрозумілості і/або кращого розуміння винаходу, і такі приведені тут визначення термінів не повинні бути витлумачені як визначення, які істотно відрізняються від тих визначень, що, по суті, зрозумілі фахівцям. Для практичного здійснення даного винаходу можуть бути застосовані, якщо це не обговорено особливо, стандартні методи, застосовувані в молекулярній біології (включаючи методи рекомбінантних ДНК), мікробіології, біології клітини, біохімії, хімії нуклеїнових кислот і імунології, і відомі фахівцям. Такі методи докладно описані в літературі, наприклад, у публікаціях Current Protocols in Immunology (J. E. Coligan et al, eds., 1999, включаючи доповнення, що вийшли до 2001 р. включно); Current Protocols in Molecular Biology (F. M. Ausubel et al, eds., 1987, включаючи доповнення, що вийшли до 2001 р. включно); Molecular Cloning: A Laboratory Manual, третє видання (Sambrook and Russel, 2001); PCR: The Polymerase Chain Reaction, (Mullis et al., eds., 1994); The Immunoassay Handbook (D. Wild, ed., Stockton Press NY, 1994); Bioconjugate Techniques (Greg T. Hermanson, ed., Academic Press, 1996); Methods of Immunological Analysis (R. Masseyeff, W. H. Albert, and N. A. Staines, eds., Weinheim: VCH Verlags gesellschaft mbH, 1993), Harlow and Lane Using Antibodies: A Laboratory Manual Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., 1999; and Beaucage et al. eds., Current Protocols in Nucleic Acid Chemistry John Wiley & Sons, Inc., New York, 2000). Використовувані тут терміни "антитіло" і "антитіла" означають моноклональні антитіла, мультиспецифічні антитіла, людські антитіла, гуманізовані антитіла, синтетичні антитіла, химерні антитіла, поліклональні антитіла, верблюжі антитіла, одноланцюжкові Fv (scFv), одноланцюжкові антитіла, Fab-фрагменти, F(ab')-фрагменти, біспецифічні Fv (sdFv), зв'язані дисульфідним зв'язком, інтраантитіла й антиідіотипові (анти-Id) антитіла (включаючи, наприклад, анти-Id і анти-анти-іd антитіла проти антитіл відповідно до винаходу) і епітопзв'язувальні фрагменти будь-яких перерахованих вище антитіл. Зокрема, антитілами є молекули імуноглобуліну і імунологічно активні фрагменти молекул імуноглобуліну, тобто молекули, що містять антигензв'язувальний сайт. Молекули імуноглобуліну можуть належати до 12 UA 111612 C2 5 10 15 20 25 30 35 40 45 50 55 60 будь-якого типу (наприклад, IgG, IgE, IgM, IgD, IgA і IgY), класу (наприклад, IgG1, IgG2, IgG3, IgG4, IgA1 і IgA2) або підкласу. Використовувані тут терміни "імуноспецифічно зв'язується", "імуноспецифічно розпізнає", "специфічно зв'язується", "специфічно розпізнає" і аналогічні терміни стосуються молекул, що специфічно зв'язуються з антигеном (наприклад, з епітопом або з імунним комплексом), але не зв'язуються специфічно з іншою молекулою. Молекула, що специфічно зв'язується з антигеном, може зв'язуватися з іншими пептидами або поліпептидами з більш низькою афінністю, як було визначено, наприклад, за допомогою імуноаналізів, BIAcore або інших аналізів, відомих фахівцям. Переважно, щоб молекули, що специфічно зв'язуються з антигеном, не мали здатність перехресно реагувати з іншими білками. Молекули, що специфічно зв'язуються з антигеном, можуть бути ідентифіковані, наприклад, за допомогою імуноаналізів, BIAcore або іншими методами, відомими фахівцям. Використовуваний тут термін "імунний комплекс" означає структуру, що утворюється в тому випадку, коли щонайменше одна молекула-мішень і щонайменше один гетерологічний поліпептид, що містить Fсγ-ділянку, зв'язуються один з одним з утворенням комплексу з більш високою молекулярною масою. Прикладами імунних комплексів є комплекси "антиген-антитіло", що можуть бути розчинними або складатися з частинок (наприклад, комплекс антиген/антитіло на клітинній поверхні). Використовувані тут терміни "важкий ланцюг", "легкий ланцюг", "варіабельна ділянка", "каркасна ділянка", "константний домен" і т. п. мають своє загальноприйняте значення, відоме фахівцям в галузі імунології, і означають домени в природних імуноглобулінах і відповідні домени синтетичних (наприклад, рекомбінантних) білків, що зв'язуються (наприклад, гуманізованих антитіл, одноланцюжкових антитіл, химерних антитіл і т. п.). Основною структурною одиницею природних імуноглобулінів (наприклад, IgG) є тетрамер, що має два легкі ланцюги і два важкі ланцюги і звичайно експресується у вигляді глікопротеїну розміром приблизно 150000 Да. Аміно-кінцева ("N") частина кожного ланцюга включає варіабельну ділянку приблизно з 100-110 або більше амінокислот, що, головним чином, відповідальні за розпізнавання антигену. Карбокси-кінцева ("C") частина кожного ланцюга означає константну ділянку, де легкі ланцюги мають один константний домен, а важкі ланцюги звичайно мають три константні домени і шарнірну ділянку. Таким чином, структура легких ланцюгів молекули IgG являє собою n-VL-CL-с, а структура важких ланцюгів IgG являє собою n-VH-CН1-H-CH2-CH3-c (де H являє собою шарнірну ділянку). Варіабельні ділянки молекули IgG складаються з гіперваріабельних ділянок (CDR, ділянок, які визначають комплементарність), що містять залишки, які контактують з антигеном і не-CDR-сегментами, що є каркасними сегментами, які, по суті, зберігають структуру і визначають положення CDR-петель (хоча деякі каркасні залишки можуть також контактувати з антигеном). Таким чином, VL- і VH-домени мають структуру n-FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4-c. Що стосується зв'язувальних білків або антитіл (визначених тут загалом), то присвоєння амінокислот кожному домену здійснюють відповідно до визначень по Кебату, Sequences of Proteins of Immunological Interest (National Institutes of Health, Bethesda, Md., 1987 and 1991). Амінокислоти варіабельних ділянок зрілих важких і легень ланцюгів імуноглобулінів позначають по положенню амінокислоти в ланцюзі. Кебатом були описані різні амінокислотні послідовності для антитіл, ідентифікована амінокислотна консенсусна послідовність для кожної підгрупи, і кожній амінокислоті був присвоєний номер залишку. Схема нумерації по Кебату поширюється і на антитіла, не включені в його короткий посібник з вирівнювання розглянутого антитіла з однієї з консенсусних послідовностей, пронумерованих по консервативних амінокислотах відповідно до нумерації Кебата. Цей метод присвоєння номерів залишкам стає традиційним у даній галузі і дозволяє легко ідентифікувати амінокислоти в еквівалентних положеннях різних антитіл, включаючи химерні або гуманізовані варіанти. Так, наприклад, амінокислота в положенні 50 легкого ланцюга людського антитіла займає положення, еквівалентне положенню 50 амінокислоти в легкому ланцюзі мишачого антитіла. Використовуваний тут термін "важкий ланцюг" визначений як важкий ланцюг антитіла IgG. У інтактному нативному IgG важкий ланцюг містить VH-домен, CH1-домен, шарнірну ділянку, CH2-домен і CH3-домен імуноглобуліну. В описі даного винаходу залишки у важкому ланцюзі IgG пронумеровані відповідно до Європейської нумерації (EU-нумерації), як описано в публікації th Kabat et al., Sequences of Proteins of Immunological Interest, 5 Ed. Public Health Service, NH1, MD (1991), що точно вводиться в даний опис за допомогою посилання. "EU-нумерація по Кебату" означає EU-нумерацію людського антитіла IgG1. Приклади амінокислотних послідовностей, що містять шарнірну ділянку, CH2- і CH3-домени людського IgG1, представлені на фіг. 1A і 1B, як описано нижче. На фіг. 1A і 1B також представлені амінокислотні послідовності шарнірної 13 UA 111612 C2 5 10 15 20 25 30 35 40 45 50 55 ділянки, CH2- і CH3-доменів важких ланцюгів IgG2, IgG3 і IgG4. Амінокислотні послідовності ізотипів IgG2, IgG3 і IgG4 вирівнювали з послідовністю IgG1 шляхом уведення першого й останнього цистеїнових залишків відповідних шарнірних ділянок, які утворюють S-S зв'язки між важкими ланцюгами, у ті самі положення. У шарнірній ділянці IgG2 і IgG3 не всі залишки пронумеровані відповідно до EU-нумерації. "Шарнірна ділянка" або "шарнірний домен" загалом визначені як фрагмент, розташований від Glu216 до Pro230 людського IgG1. Приклад амінокислотної послідовності шарнірної ділянки людського IgG1 представлений на фіг. 1А (амінокислотні залишки на фіг. 1А пронумеровані відповідно до системи нумерації Кебата). Шарнірні ділянки IgG інших ізотипів можуть бути вирівняні з послідовністю IgG1 шляхом уведення першого й останнього цистеїнових залишків, що утворюють S-S зв'язки між важкими ланцюгами, у ті самі положення, як показано на фіг. 1А. Використовувані тут терміни "Fc-ділянка", "Fc-домен" або аналогічні терміни визначають як C-кінцеву ділянку важкого ланцюга IgG. Приклад амінокислотної послідовності, що містить людський IgG1, представлений на фіг. 1B. Хоча межі можуть варіюватися дуже незначно, як було визначено відповідно до системи нумерації Кебата, однак було встановлено, що Fc-домен розташований від амінокислоти 231 до амінокислоти 447 (амінокислотні залишки на фіг. 1B пронумеровані відповідно до системи нумерації Кебата). На фіг. 1B також представлені приклади амінокислотних послідовностей Fc-ділянок ізотипів IgG, а саме, IgG2, IgG3 і IgG4. Fc-ділянка IgG містить два константні домени, CH2 і CH3. CH2-домен Fc-ділянки людського IgG звичайно розташований від амінокислоти 231 до амінокислоти 341, як було визначено відповідно до системи нумерації Кебата (фіг. 1B). CH3-домен Fc-ділянки людського IgG звичайно розташований від амінокислоти 342 до амінокислоти 447, як було визначено відповідно до системи нумерації Кебата (фіг. 1B). Унікальність CH2-домену Fc-ділянки людського IgG (який також називається доменом "Сγ2") полягає в тому, що він не утворює тісно зв'язану пару з іншим доменом. Точніше, два N-зв'язаних розгалужених вуглеводних ланцюги убудовані між двома CH2-доменами інтактного нативного IgG. Використовувані тут терміни "FсγR-зв'язувальний білок", "FсγR-антитіло" і "анти-FсγR антитіло" є синонімами й означають різні імуноглобулін-подібні білки або білки, які походять від імуноглобуліну. "FсγR-зв'язувальні білки" зв'язуються з FсγR за допомогою взаємодії з VL- і/або VR-доменами (на відміну від Fсγ-опосередковуваного зв'язування). Прикладами FсγRзв'язувальних білків є повністю людські, поліклональні, химерні і гуманізовані антитіла (наприклад, що містять 2 важкі і 2 легкі ланцюги), їхні фрагменти (наприклад, Fab-, Fab'-, F(ab')2і Fv-фрагменти), біфункціональні або мультифункціональні антитіла (див., наприклад, Lanzavecchia et al. (1987) "The Use Of Hybrid Hybridomas To Target Human Cytotoxic T Lymphocytes" Eur. J. Immunol. 17:105-111), одноланцюжкові антитіла (див., наприклад, Bird et al. (1988) "Single-Chain Antigen-Binding Proteins" Science 242:423-26), злиті білки (наприклад, злиті білки фагового представлення), "мініантитіла" (див., наприклад, патент США № 5837821) і інші антигензв'язувальні білки, що містять VL- і/або VН-домен або його фрагмент. В одному з аспектів винаходу FсγRIIIA-зв'язувальним білком є "тетрамерне антитіло", тобто антитіло, що в основному має структуру природного IgG і містить варіабельні і константні домени, тобто два легкі ланцюги, що містять VL-домен і константний домен легкого ланцюга, і два важкі ланцюги, що містять VН-домен, шарнірну ділянку і константні домени важкого ланцюга. Використовуваний тут термін "антагоністи FсγR" і аналогічні терміни означають білкові і небілкові речовини, включаючи невеликі молекули, які інгібують щонайменше одну біологічну активність FсγR, наприклад, блокують передачу сигналу. Так, наприклад, молекули відповідно до винаходу блокують передачу сигналу за допомогою інгібування зв'язування IgG з FсγR. Використовуваний тут термін "похідне", що стосується поліпептидів або білків, означає поліпептид або білок, що містить амінокислотну послідовність, яка була модифікована шляхом уведення замін, делецій або додавань амінокислотних залишків. Використовуваний тут термін "похідне" також означає поліпептид або білок, який був модифікований, наприклад, за допомогою ковалентного зв'язування молекули будь-якого типу з поліпептидом або білком. Прикладами способів модифікації антитіла є, але не обмежуються ними, глікозилування, ацетилування, ПЕГилування, фосфорилування, амідування, дериватизація відомими захисними/блокуючими групами, протеолітичне розщеплення, зв'язування з клітинним антигеном або з іншим білком і т. п. Похідне поліпептиду або білка може бути отримане шляхом хімічних модифікацій із застосуванням методів, відомих фахівцям, включаючи, але не обмежуючись ними, специфічне хімічне розщеплення, ацетилування, формілування, метаболічний синтез тунікаміцину і т. п. Крім того, похідне поліпептиду або білка має функцію, аналогічну або ідентичну функції поліпептиду або білка, від якого воно походить. 14 UA 111612 C2 5 10 15 20 25 30 35 40 45 50 55 60 Використовуваний тут термін "похідне", що стосується небілкового похідного, означає вторинну органічну або неорганічну молекулу, що утворюється на основі структури першої органічної або неорганічної молекули. Похідне органічної молекули включає, але не обмежується ними, молекулу, модифіковану, наприклад, у результаті додавання або делеції гідроксильної, метильної, етильної, карбоксильної групи або аміногрупи. Органічна молекула може бути також етерифікованою, алкилованою і/або фосфорилованою. Використовуваний тут термін "молекула діатіла" означає комплекс двох або більш поліпептидних ланцюгів або білків, кожні з яких містять щонайменше один V L-домен і один VHдомен або їхні фрагменти, де обидва домени містяться в одному поліпептидному ланцюзі. У деяких варіантах винаходу, "молекула діатіла" включає молекули, що містять Fc або шарнірну ділянку-Fc-домен. Вказані поліпептидні ланцюги в комплексі можуть бути однаковими або різними, тобто, молекула діатіла може являти собою гомомультимер або гетеромультимер. У конкретних аспектах винаходу "молекула діатіла" включає димери або тетрамери, або вказані поліпептидні ланцюги, що містять VL- і VН-домен. Окремі поліпептидні ланцюги, що містять мультимерні білки, можуть бути ковалентно зв'язані щонайменше з одним іншим пептидом мультимера міжланцюговими дисульфідними зв'язками. Використовувані тут терміни "розлад" і "захворювання" є синонімами й означають патологічний стан в індивідуума. Зокрема, термін "аутоімунне захворювання" є синонімом терміна "аутоімунний розлад" і означає стан індивідуума, що характеризується ушкодженням клітин, тканин і/або органів, що викликається імунологічною реакцією індивідуума на свої власні клітини, тканини і/або органи. Термін "запальне захворювання" є синонімом терміна "запальний розлад" і означає стан індивідуума, що характеризується запаленням, а переважно хронічним запаленням. Аутоімунні розлади можуть асоціюватися, а можуть і не асоціюватися з запаленням. Крім того, запалення може викликатися, а може і не викликатися аутоімунним розладом. Таким чином, деякі розлади можуть бути охарактеризовані як аутоімунні і запальні розлади. Використовуваний тут термін "ідентичні поліпептидні ланцюги" також означає поліпептидні ланцюги, що мають майже ідентичні амінокислотні послідовності, наприклад, включаючи ланцюги, що мають розходження в одній або більше амінокислотах, а переважно консервативні амінокислотні заміни, що не чинять значного впливу на активність двох поліпептидних ланцюгів. Використовуваний тут термін "рак" означає новоутворення або пухлину, що утворюються в результаті аномального неконтрольованого росту клітин. Більш конкретно, використовуваний тут термін "рак" включає лейкоз і лімфоми. У деяких варіантах винаходу рак означає доброякісну пухлину, що зберігає свою локалізацію. В інших варіантах винаходу рак означає злоякісну пухлину, що проникає в прилеглі структури організму і руйнує їх, а також поширюється і на віддалені ділянки. У деяких варіантах винаходу рак асоціюється зі специфічним раковим антигеном. Використовуваний тут термін "імуномодулюючий агент" і його варіанти означає агент, що модулює імунну систему хазяїна. У деяких варіантах винаходу імуномодулюючим агентом є імуносупресорний агент. У деяких інших варіантах винаходу імуномодулюючим агентом є імуностимулюючий агент. Імуномодулюючими агентами є, але не обмежуються ними, невеликі молекули, пептиди, поліпептиди, злиті білки, антитіла, неорганічні молекули, агенти-міметики й органічні молекули. Використовуваний тут термін "епітоп" означає фрагмент поліпептиду або білка або небілкову молекулу, що мають антигенну або імуногенну активність у тварини, переважно в ссавця, а найбільш переважно в людини. Епітоп, який має імуногенну активність, являє собою фрагмент поліпептиду або білка, що виробляє гуморальну відповідь у тварини. Епітоп, що має антигенну активність, являє собою фрагмент поліпептиду або білка, з яким імуноспецифічно зв'язується антитіло, як може бути визначено будь-яким методом, добре відомим фахівцям, наприклад, за допомогою імуноаналізів. Антигенні епітопи необов'язково повинні бути імуногенними. Використовуваний тут термін "фрагмент" означає пептид або поліпептид, що містить амінокислотну послідовність, яка складається щонайменше з 5 суміжних амінокислотних залишків, щонайменше з 10 суміжних амінокислотних залишків, щонайменше з 15 суміжних амінокислотних залишків, щонайменше з 20 суміжних амінокислотних залишків, щонайменше з 25 суміжних амінокислотних залишків, щонайменше з 40 суміжних амінокислотних залишків, щонайменше з 50 суміжних амінокислотних залишків, по меншій мері з 60 суміжних амінокислотних залишків, щонайменше з 70 суміжних амінокислотних залишків, щонайменше з 80 суміжних амінокислотних залишків, щонайменше з 90 суміжних амінокислотних залишків, щонайменше з 100 суміжних амінокислотних залишків, щонайменше з 125 суміжних 15 UA 111612 C2 5 10 15 20 25 30 35 40 45 50 55 амінокислотних залишків, щонайменше з 150 суміжних амінокислотних залишків, щонайменше з 175 суміжних амінокислотних залишків, щонайменше з 200 суміжних амінокислотних залишків, або щонайменше з 250 суміжних амінокислотних залишків амінокислотної послідовності іншого поліпептиду. У конкретному варіанті винаходу фрагмент поліпептиду зберігає щонайменше одну функцію цього поліпептиду. Використовувані тут терміни "нуклеїнові кислоти" і "нуклеотидні послідовності" включають молекули ДНК (наприклад, кДНК або геномну ДНК), молекули РНК (наприклад, мРНК), комбінації молекул ДНК і РНК або гібридні молекули ДНК/РНК і аналоги молекул ДНК або РНК. Такі аналоги можуть бути отримані з використанням, наприклад, нуклеотидних аналогів, якими є, але не обмежуються ними, інозин або тритиловані основи. Такі аналоги можуть також включати молекули ДНК або РНК, що містять модифіковані кістяки, які надають молекулам цінні властивості, такі як, наприклад, резистентність до нуклеаз або підвищена здатність проходити через клітинні мембрани. Нуклеїнові кислоти або нуклеотидні послідовності можуть бути одноланцюжковими і дволанцюжковими, можуть містити одноланцюжкові і дволанцюжкові частини, і можуть містити триланцюжкові частини, але переважними є дволанцюжкові ДНК. Використовуваний тут термін "терапевтично ефективна кількість" означає кількість терапевтичного агента, достатню для терапії або лікування захворювання або розладу. Терапевтично ефективна кількість може означати кількість терапевтичного агента, достатню для уповільнення розвитку захворювання або мінімізації тяжкості захворювання, наприклад, для уповільнення прогресування раку або мінімізації поширення раку. Терапевтично ефективна кількість може також означати кількість терапевтичного агента, що буде давати терапевтичний ефект при терапії або лікуванні захворювання. Крім того, терапевтично ефективна кількість терапевтичного агента відповідно до винаходу означає кількість терапевтичного агента, узятого окремо або в комбінації з іншими терапевтичними агентами, що буде давати терапевтичний ефект при терапії або лікуванні захворювання. Використовувані тут терміни "профілактичний агент" і "профілактичні агенти" означають будь-який(і) агент(и), який(і) може (можуть) бути використаний(і) для попередження розладу або попередження рецидивів або поширення такого розладу. Профілактично ефективна кількість може означати кількість профілактичного агента, достатню для попередження рецидивів або поширення гіперпроліферативного захворювання, а зокрема, раку, зниження або випадків захворюваності на рак пацієнтів, включаючи, але не обмежуючись ними, пацієнтів зі схильністю до розвитку гіперпроліферативного захворювання, наприклад, пацієнтів з генетичною схильністю до раку, або пацієнтів, що раніше піддавалися впливу канцерогенів. Профілактично ефективна кількість може також означати кількість профілактичного агента, що буде давати профілактичний ефект при попередженні захворювання. Крім того, профілактично ефективна кількість профілактичного агента відповідно до винаходу означає кількість профілактичного агента, узятого окремо або в комбінації з іншими агентами, що буде давати профілактичний ефект при попередженні захворювання. Використовувані тут терміни "попереджати", "попереджуючий" і "попередження" стосуються попередження рецидивів або початку розвитку одного або більше симптомів розладу в індивідуума після введення профілактичного або терапевтичного агента. Використовуваний тут термін "у комбінації" означає використання більше ніж одного профілактичного і/або терапевтичного агентів. При вживанні терміна "у комбінації" не мається на увазі обмеження порядку введення профілактичного і/або терапевтичного агентів індивідууму, що страждає на розлад. Перший профілактичний або терапевтичний агент може бути введений до (наприклад, за 5 хвилин, 15 хвилин, 30 хвилин, 45 хвилин, 1 годину, 2 години, 4 години, 6 годин, 12 годин, 24 години, 48 годин, 72 години, 96 годин, 1 тиждень, 2 тижні, 3 тижні, 4 тижні, 5 тижнів, 6 тижнів, 8 тижнів або 12 тижнів) уведення другого профілактичних або терапевтичного агента; або одночасно з уведенням вказаного другого профілактичного або терапевтичного агента, або після (наприклад, через 5 хвилин, 15 хвилин, 30 хвилин, 45 хвилин, 1 годину, 2 години, 4 години, 6 годин, 12 годин, 24 години, 48 годин, 72 години, 96 годин, 1 тиждень, 2 тижні, 3 тижні, 4 тижні, 5 тижнів, 6 тижнів, 8 тижнів або 12 тижнів) уведення вказаного другого профілактичного або терапевтичного агента індивідууму, що страждає на розлад. Використовуваний тут термін "ефекторна функція" означає біохімічний процес, обумовлений взаємодією Fc-ділянки антитіла з Fc-рецептором або антигеном. Ефекторними функціями є, але не обмежуються ними, антитіло-залежна клітинно-опосередковувана цитотоксичність (ADCC), антитіло-залежний клітинно-опосередковуваний фагоцитоз (ADCP) і комплемент-залежна цитотоксичність (CDC). Ефекторними функціями є функції, що здійснюються після зв'язування з антигеном і незалежно від зв'язування з антигеном. 16 UA 111612 C2 5 10 15 20 25 30 35 40 45 50 55 Використовуваний тут термін "ефекторна клітина" означає клітину імунної системи, яка експресує один або більше Fc-рецепторів і опосередковує здійснення однієї або більше ефекторних функцій. Ефекторними клітинами є, але не обмежуються ними, моноцити, макрофаги, нейтрофіли, дендритні клітини, еозинофіли, гладкі клітини, тромбоцити, B-клітини, великі гранулярні лімфоцити, клітини Лангерганса, природні клітини-кілери (NK), і ці клітини можуть походити від будь-яких організмів, таких як, але не обмежуваних ними, людина, миша, щур, кролики і мавпи. Використовуваний тут термін "специфічно зв'язується з імунним комплексом" і аналогічні терміни стосуються молекул, що специфічно зв'язуються з імунним комплексом і не зв'язуються специфічно з іншою молекулою. Молекула, що специфічно зв'язується з імунним комплексом, може зв'язуватися з іншими пептидами або поліпептидами з більш низькою афінністю, і може бути визначена, наприклад, за допомогою імуноаналізів, BIAcore або інших аналізів, відомих фахівцям. При цьому переважно, щоб молекули, які специфічно зв'язуються з імунним комплексом, були нездатні перехресно реагувати з іншими білками. Молекули, які специфічно зв'язуються з імунним комплексом, можуть бути ідентифіковані, наприклад, за допомогою імуноаналізів, BIAcore, або іншими методами, відомими фахівцям. Використовуваний тут термін "стабільний злитий білок" означає злитий білок, який піддається мінімального ступеня розкладання або взагалі не піддається розкладанню в процесі його продукування і/або збереження, як може бути визначено за допомогою стандартних біологічних і функціональних аналізів, відомих фахівцям, і який може зберігатися протягом тривалого періоду часу, не втрачаючи при цьому свою біологічну активність, наприклад, активність зв'язування з FсγR. 4. Короткий опис графічного матеріалу Фіг. 1A-B. Амінокислотна послідовність CH1-ділянки, шарнірної ділянки і Fc-ділянки людського IgG На фігурах 1(A) і (B) представлені амінокислотні послідовності шарнірного домену (A) і Fcдомену (B) людських IgG1, IgG2, IgG3 і IgG4. (шарнірний домен IgG1 (SEQ ID NO: 1); шарнірний домен IgG2 (SEQ ID NO: 2); шарнірний домен IgG3 (SEQ ID NO: 3); шарнірний домен IgG4 (SEQ ID NO: 4); Fc-домен IgG1 (SEQ ID NO: 5); Fc-домен IgG2 (SEQ ID NO: 6); Fc-домен IgG3 (SEQ ID NO: 7); Fc-домен IgG1 (SEQ ID NO: 8)). Амінокислотні залишки, вказані на фігурах 1A і 1B, пронумеровані відповідно до системи нумерації EU Кебата. Послідовності ізотипів вирівнювали з послідовністю IgG1 шляхом уведення першого й останнього цистеїнових залишків відповідних шарнірних ділянок, які утворюють S-S зв'язки між важкими ланцюгами, у ті самі положення. На фігурі 1B залишки в CH2-домені позначені +, а залишки в CH3-домені позначені ~. Фіг. 2. Схематичне представлення поліпептидних ланцюгів ковалентно зв'язаних біфункціональних діатіл Поліпептиди ковалентно зв'язаних біфункціональних діатіл складаються з VL-домену антитіла і VH-домену антитіла, відділених один від одного коротким пептидним лінкером. Лінкер, який складається з 8 амінокислотних залишків, запобігає самозбиранню одного поліпептидного ланцюга в scFv-конструкції, і замість цього, в основному, відбувається взаємодія між VL- і VH-доменами різних поліпептидних ланцюгів. Були створені 4 конструкції (кожна конструкція зчитується з аміно-кінця ("n"), тобто з лівого краю конструкції, до карбокси-кінця ("c"), тобто на фігурі праворуч): конструкція (1) (SEQ ID NO: 9) містить n-VL-домен Hu2B6 лінкер (GGGSGGGG (SEQ ID NO: 10)) - VH-домен Hu3G8 - і C-кінцеву послідовність (LGGC)-c; конструкція (2) (SEQ ID NO: 11) містить n-VL-домен Hu3G8 - лінкер (GGGSGGGG (SEQ ID NO: 10)) - VH-домен Hu2B6 - і C-кінцеву послідовність (LGGC)-c; конструкція (3) (SEQ ID NO: 12) містить n-VL-домен Hu3G8 - лінкер (GGGSGGGG (SEQ ID NO: 10)) - VH-домен Hu3G8 - і Cкінцеву послідовність (LGGC)-c; конструкція (4) (SEQ ID NO: 13) містить n-VL-домен Hu2B6 лінкер (GGGSGGGG (SEQ ID NO: 10)) - VH-домен Hu2B6 - і C-кінцеву послідовність (LGGC)-c. Фіг. 3. Аналіз афінно очищених діатіл за допомогою електрофорезу в ДСН-ПААГ Афінно очищені діатіла піддавали ДСН-ПААГ-аналізу у відновних умовах (доріжки 1-3) або у невідновлювальних умовах (доріжки 4-6). Вказані приблизні молекулярні маси стандарту (між доріжками 3 і 4). Доріжки 1 і 4, h3G8 CMD; доріжки 2 і 5, h2B6 CMD; і доріжки 3 і 6, h2B6-h3G8 CBD. Фіг. 4 A-B. Аналіз афінно очищених діатіл за допомогою ексклюзійної хроматографії (ЕХ) Афінно очищені діатіла піддавали ЕХ-аналізу. (A) Профіль елюювання відомих стандартів: повнорозмірного IgG (~150 кДа), Fab-фрагмента IgG (~50 кДа) і scFv (~30 кДа); (B) Профіль елюювання h2B6 CMD, h3G8 CMD і h2B6-h3G8 CBD. Фіг. 5. Зв'язування h2B6-h3G8 CBD з sCD32B і sCD16A 17 UA 111612 C2 5 10 15 20 25 30 35 40 45 50 55 60 Зв'язування h2B6-h3G8 CBD з sCD32B і sCD16A аналізували за допомогою "сендвіч»-ELISA. sCD32B використовували як білок-мішень. Другим зондом був ПХ-кон'югований sCD16A. h3G8 CMD, яке зв'язується з CD16A, використовували як контроль. Фіг. 6 A-C. BIACORE-аналіз на зв'язування діатіла з sCD16A, SCD32B і sCD32А Зв'язування h2B6-h3G8 CBD, h2B6 CMD і h3G8 CMD з sCD16A, sCD32B і sCD32A (негативний контроль) аналізували за допомогою ППР-аналіза. h3G8 scFv також тестували як контроль. (A) Зв'язування з sCD16; (B) зв'язування з sCD32B і (C) зв'язування з sCD32A. Діатіла наносили шляхом інжекції в концентрації 100 нM, а scFv - у концентрації 200 нM, по всій поверхні рецептора зі швидкістю потоку 50 мл/хв протягом 60 секунд. Фіг. 7 A-C. BIACORE-аналіз на зв'язування діатіла з sCD16A і sCD32B Зв'язування h2B6-h3G8 CBD, h2B6 CMD і h3G8 CMD з sCD16A і sCD32B аналізували за допомогою ППР-аналіза. h3G8 scFv також тестували як контроль. (A) Зв'язування h3G8 CMD з sCD16A; (B) зв'язування h2B6-h3G8 CBD з sCD16A; (C) зв'язування h3G8 scFv з sCD16A; (D) зв'язування h2B6 CMD з sCD32B; і (E) зв'язування h2B6-h3G8 CBD з sCD32B. Діатіла наносили шляхом інжекції в концентраціях 6,25-200 нМ по всій поверхні рецептора зі швидкістю потоку 70 мл/хв протягом 180 секунд. Фіг. 8. Схематичне представлення взаємодії поліпептидних ланцюгів, які містять VL- і VHдомени, з утворенням ковалентно зв'язаної біспецифічної молекули діатіла NH2 і COOH являють собою аміно-кінець і карбокси-кінець кожного поліпептидного ланцюга, відповідно. S являє собою C-кінцевий цистеїновий залишок на кожному поліпептидному ланцюзі. VL і VH означають варіабельний домен легкого ланцюга і варіабельний домен важкого ланцюга, відповідно. Два поліпептидні ланцюги розділені штриховими і пунктирними лініями, що, зокрема, представляють лінкерні частини вказаних ланцюгів. h2B6 Fv і h3G8 Fv означають епітоп-зв'язувальний сайт, специфічний до CD32B і CD16, відповідно. Фіг. 9. Схематичне представлення поліпептидних ланцюгів, які містять Fc-домени ковалентно зв'язаних біспецифічних функціональних діатіл Представлені поліпептидні конструкції молекул діатіла відповідно до винаходу (кожна конструкція зчитується з аміно-кінця ("n"), тобто з лівої сторони конструкції, до карбокси-кінця ("c"), тобто на фігурі праворуч): конструкція (5) (SEQ ID NO: 14) містить n-VL-домен Hu2B6 перший лінкер (GGGSGGGG (SEQ ID NO: 10)) - VH-домен Hu3G8 - другий лінкер (LGGC)- і Cкінцевий Fc-домен людського IgG1-С; конструкція (6) (SEQ ID NO: 15) містить n-VL-домен Hu3G8 - лінкер (GGGSGGGG (SEQ ID NO: 10)) - VH-домен Hu2B6 - і другий лінкер (LGGC)- і Cкінцевий Fc-домен людського IgG1-с; конструкція (7) (SEQ ID NO: 16) містить n-VL-домен Hu3В6 - перший лінкер (GGGSGGGG (SEQ ID NO: 10)) - VH-домен Hu3G8 - і C-кінцеву послідовність (LGGCFNRGEC) (SEQ ID NO: 17)-c; конструкція (8) (SEQ ID NO: 18) містить n-VL-домен Hu3G8 лінкер (GGGSGGGG (SEQ ID NO: 10)) - VH-домен Hu2B6 - другий лінкер (LGGC)- і C-кінцевий шарнірний домен/Fc-домен людського IgG1 (з амінокислотною заміною A215V)-c. Фіг. 10. Зв'язування молекул діатіла, що містять Fc-домени, з sCD32B і sCD16A Зв'язування молекул діатіла, що містять Fc-домени, з sCD32B і sCD16A аналізували за допомогою "сендвіч»-ELISA. Аналізовані антитіла були отримані з використанням 3 систем рекомбінантної експресії: контрансфекції pMGX669 і pMGX674, експресуючих конструкції 1 і 6, відповідно; контрансфекції pMGX667 і pMGX676, експресуючих конструкції 2 і 5, відповідно; і контрансфекції pMGX674 і pMGX676, експресуючих конструкції 5 і 6, відповідно. sCD32B використовували як білок-мішень. Другим зондом був ПХ-кон'югований sCD16A. Фіг. 11. Схематичне представлення взаємодії двох поліпептидних ланцюгів, кожен з який містить Fc-домен, з утворенням двовалентного ковалентно зв'язаного діатіла NH2 і COOH являють собою аміно-кінець і карбокси-кінець кожного поліпептидного ланцюга, відповідно. S являє собою щонайменше один дисульфідний зв'язок між цистеїновими залишками в другій лінкерній послідовності кожного поліпептидного ланцюга. VL і VH означають варіабельний домен легкого ланцюга і варіабельний домен важкого ланцюга, відповідно. Два поліпептидні ланцюги розділені штриховими і пунктирними лініями, що, зокрема, представляють перші лінкерні частини вказаних ланцюгів. СН2 і СН3 означають константні СН2- і СН3-домени Fc-домену. h2B6 Fv і h3G8 Fv означають епітоп-зв'язувальний сайт, специфічний до CD32B і CD16, відповідно. Фіг. 12. Зв'язування молекул діатіла, що містять шарнірні домени/Fc-домени, з sCD32B і sCD16A Зв'язування молекул діатіла, що містять Fc-домени, з sCD32B і sCD16A аналізували за допомогою "сендвіч»-ELISA. Аналізовані антитіла були отримані з використанням 4 систем рекомбінантної експресії: котрансфекції pMGX669+pMGX674, експресуючих конструкції 1 і 6, відповідно; котрансфекції pMGX669+pMGX678, експресуючих конструкції 2 і 8, відповідно; 18 UA 111612 C2 5 10 15 20 25 30 35 40 45 50 55 60 котрансфекції pMGX677+pMGX674, експресуючих конструкції 7 і 6, відповідно; і котрансфекції pMGX677+pMGX678, експресуючих конструкції 7 і 8, відповідно. sCD32B використовували як білок-мішень. Другим зондом був ПХ-кон'югований sCD16A. Фіг. 13. Схематичне представлення взаємодії поліпептидних ланцюгів з утворенням тетрамерної молекули діатіла NH2 і COOH являють собою аміно-кінець і карбокси-кінець кожного поліпептидного ланцюга, відповідно. S являє собою щонайменше один дисульфідний зв'язок між цистеїновим залишком у другій лінкерній послідовності Fc-несучого "більш важкого" поліпептидного ланцюга і цистеїновим залишком у С-кінцевій послідовності, що не несе Fc, "більш легкого" поліпептидного ланцюга. VL і VH означають варіабельний домен легкого ланцюга і варіабельний домен важкого ланцюга, відповідно. Два поліпептидні ланцюги розділені штриховими і пунктирними лініями, що, зокрема, представляють перші лінкерні частини вказаних "більш важких" або ланцюгів лінкер вказаних "більш легких" ланцюгів. СН2 і СН3 означають константні СН2- і СН3-домени Fc-домену. h2B6 Fv і h3G8 Fv означають епітопзв'язувальний сайт, специфічний до CD32B і CD16, відповідно. Фіг. 14. Схематичне представлення поліпептидних ланцюгів, які містять Fc-домени, що утворюють ковалентно зв'язані біспецифічні діатіла Представлені поліпептидні конструкції, які утворюють молекули діатіла відповідно до винаходу (кожна конструкція зчитується з аміно-кінця ("n"), тобто з лівої сторони конструкції, до карбокси-кінця ("c"), тобто на фігурі праворуч): конструкція (9) (SEQ ID NO: 19) містить nшарнірний домен/Fc-домен людського IgG1-VL-домен Hu3G8 - лінкер (GGGSGGGG (SEQ ID NO: 10)) - VH-домен Hu2B6 - лінкер (GGGSGGGG) (SEQ ID NО 10)) і C-кінцеву послідовність LGGCз; конструкція (10) (SEQ ID NO: 20) містить n-Fc-домен людського IgG1-VL-домен Hu3G8 - лінкер (GGGSGGGG (SEQ ID NO: 10)) - VH-домен Hu2B6 - лінкер (GGGSGGGG (SEQ ID NO: 10)) - і Cкінцеву послідовність LGGC-c; конструкція (11) (SEQ ID NO: 21) містить n-VL-домен Hu2B6 (G105C) - лінкер (GGGSGGGG (SEQ ID NO: 10)) - VH-домен Hu3G8 - і C-кінцевий шарнірний домен/Fc-домен людського IgG1 з амінокислотною заміною A215V-c; конструкція (12) (SEQ ID NO: 22) містить n-VL-домен Hu3G8 - лінкер (GGGSGGGG (SEQ ID NO: 10)) - VH-домен Hu2B6 (G44C) - і C-кінцеву послідовність FNRGEC (SEQ ID NO: 23)-с. Фіг. 15 A-B. Електрофорез у ДСН-ПААГ і вестерн-блот-аналіз афінних тетрамерних діатіл Діатіла, продуковані з використанням систем рекомбінантної експресії, котрансфікованних векторами, експресуючими конструкції 10 і 1, конструкції 9 і 1 і конструкції 11 і 12, піддавали електрофорезу в ДСН-ПААГ у невідновлювальних умовах (A) і вестерн-блот-аналізу з використанням козячого антитіла проти людських IgG1 H+L як зонда (B). Білки в ДСН-ПААГ-гелі візуалізували з використанням синього барвника Simply Blue Safestain (Invitrogen). Для обох панелей A і B молекули діатіл, що містять конструкції 10 і 1, конструкції 9 і 1 і конструкції 11 і 12А, представлені на доріжках 1, 2 і 3, відповідно. Фіг. 16. Зв'язування молекул діатіл, що містять Fc-домени і сконструйовані міжланцюгові дисульфідні зв'язки, з SCD32B і sCD16A Зв'язування молекул діатіл, що містять Fc-домени і сконструйовані міжланцюгові дисульфідні зв'язку між "більш легким" і "більш важким" поліпептидними ланцюгами, з sCD32B і sCD16A, оцінювали за допомогою "сендвіч»-ELISA. Аналізовані діатіла були отримані з використанням 3 систем рекомбінантної експресії: систем, експресуючих конструкції 1 і 10; систем, експресуючих конструкції 1 і 9; і систем, експресуючих конструкції 11 і 12, відповідно. sCD32B використовували як білок-мішень. Другим зондом був ПХ-кон'югований sCD16A. Зв'язування h3G8 використовували як контроль. Фіг. 17. Схематичне представлення поліпротеїнового попередника молекули діатіла і схематичне представлення поліпептидних ланцюгів, що містять легкий ланцюг лямбда і/або шарнірні домени Представлені поліпептидні конструкції, що містять молекули діатіла відповідно до винаходу (кожна конструкція зчитується з аміно-кінця ("n"), тобто з лівої сторони конструкції, до карбоксикінця ("c"), тобто на фігурі праворуч): конструкція (13) (SEQ ID NO: 95) містить n-VL-домен 3G8 перший лінкер (GGGSGGGG (SEQ ID NO: 10)) - VH-домен 2.4G2VH - другий лінкер (LGGC) сайт розпізнавання фурину (RAKR (SEQ ID NO: 93)) - VL-домен 2.4G2 - третій лінкер (GGGSGGG (SEQ ID NO: 10)) - VH-домен 3G8 - і C-кінцевий домен LGGC; (нуклеотидна послідовність, яка кодує SEQ ID NO: 95, представлена в SEQ ID NO: 96). Конструкція (14) (SEQ ID NO: 97) містить n-VL домен 3G8 - перший лінкер (GGGSGGGG (SEQ ID NO: 10)) - VH-домен 2.4G2VH - другий лінкер (LGGC) - сайт розпізнавання фурину (RAKR (SEQ ID NO: 93)) - сайт FMD (протеаза С3 вірусу ящуру) - VL-домен 2.4G2 - третій лінкер (GGGSGGG (SEQ ID NO: 10)) VH домен 3G8 - і C-кінцевий домен LGGC; (нуклеотидна послідовність, яка кодує SEQ ID NO: 19 UA 111612 C2 5 10 15 20 25 30 35 40 45 50 55 97, представлена в SEQ ID NO: 98). Конструкція (15) (SEQ ID NO: 99) містить n-VL-домен Hu2B6 - лінкер (GGGSGGGG (SEQ ID NO: 10)) - VH-домен Hu3G8-C-кінцевий-домен FNRGEC (SEQ ID NO: 23); (нуклеотидна послідовність, яка кодує SEQ ID NO: 99, представлена в SEQ ID NO: 100). Конструкція (16) (SEQ ID NO: 101) містить n-VL-домен Hu3G8 - лінкер (GGGSGGGG (SEQ ID NO: 10)) - VH-домен Hu2B6 - і C-кінцевий-домен VEPKSC (SEQ ID NO: 77); (нуклеотидна послідовність, яка кодує SEQ ID NO: 101, представлена в SEQ ID NO: 102). Фіг. 18. Зв'язування молекул діатіл, які походять від поліпротеїнової молекули-попередника, з mCD32B і sCD16A Зв'язування молекул діатіл, які походять від конструкції 13 поліпротеїнової молекулипопередника (SEQ ID NO: 95), з мишачим CD32B (mCD32B) і розчинним CD16A (sCD16A), аналізували за допомогою "сендвіч»-ELISA. sCD32B використовували як білок-мішень. Другим зондом був кон'югований з біотином sCD16A. Фіг. 19. Зв'язування молекул діатіл, що містять ланцюг лямбда і/або шарнірні домени, з sCD32B і sCD16A Зв'язування молекул діатіл, що містять домени, які походять від C-кінця людського легкого ланцюга лямбда і/або шарнірного домену IgG, з sCD32B і sCD16A, аналізували, і отримані результати порівнювали з результатами для діатіл, що містить конструкції 1 і 2 (фіг. 5), за допомогою "сендвіч»-ELISA. Аналізовані діатіла були отримані з використанням системи рекомбінантної експресії, яка експресує конструкції 15 і 16 (SEQ ID NO: 99 і SEQ ID NO: 101, відповідно). sCD32B використовували як білок-мішень. Другим зондом був ПХ-кон'югований sCD16A. Стовпці з невеликими "боксами" представляють комбінацію конструкцій 15/16, а стовпці з великими "боксами" представляють комбінацію конструкцій 1/2. Фіг. 20. Схематичне представлення DART 2В6/4420, зв'язаних з CD32B, локалізованому на поверхні клітин, і з молекулою, кон'югованою із флуоресцеїном На діаграмі проілюстрована гнучкість "універсального адаптера" анти-флуоресцеїнової гілки DART, а також можливість заміни гілки 2B6 на гілку з іншою специфічністю. V-ділянки представлені у вигляді боксів, лінкери GGGSGGGG (SEQ ID NO: 10) показані лініями, а дисульфідний зв'язок представлений як зв'язок, що з'єднує два ланцюги. Компоненти одного ланцюга показані синім, а компоненти іншого ланцюга показані рожевим. N - аміно-кінець; C карбокси-кінець; FL - флуоресцеїн, VL - варіабельна ділянка легкого ланцюга; VH - варіабельна ділянка важкого ланцюга. Фіг. 21 (Панелі A і B). DART 2B6/4420 специфічно зв'язується з молекулами, кон'югованими з флуоресцеїном, і може одночасно зв'язуватися з CD32B (A) 2B6/4420 або 2B6/3G8 були зв'язані з ELISA-планшетами, покритими білком ФІТЦ-S. Зв'язування і функцію гілки 2B6 детектували шляхом контактування з розчинним CD32B, а потім з антитілом, специфічним до CD32B, і з "другим" детектучим антитілом, кон'югованим із ПХ. (B) 2B6/4420 або 2B6/3G8 були зв'язані з ELISA-планшетами, покритими HuIgG або ФІТЦ-HuIgG (кон'югованими з флуоресцеїном). Зв'язування детектували шляхом контактування з поліклональною сироваткою, специфічною до Fv 2B6, а потім із ПХ-кон'югованим "другим" антитілом. Фіг. 22 (Панелі A і B). Активація очищених В-клітин з використанням антитіл проти людського CD79B Очищені B-клітини активували з використанням зростаючих концентрацій ФІТЦ-кон'югованих антитіл проти людського CD79b, CB3.1-ФІТЦ (A) або CB3.2-ФІТЦ (B) і 50 мкг/мл F(ab')2фрагмента, специфічного до Fc IgG, GAM (вісь x). B-клітини активували в присутності PBS (білі стовпці) або 5 мкг/мл αFITCαCD32BDART (чорні стовпці) або αCD16αCD32BDART (сірі стовпці). Реакції здійснювали з трьома повторностями й обчислювали стандартні відхилення. Фіг. 23 (Панелі A і B). Активація очищених B-клітин Очищені B-клітини, узяті в другого здорового донора, активували, як описано на фіг. 22, панель B. Індекс проліферації вимірювали в клітинах, активованих у присутності ФІТЦкон'югованого анти-CD79b антитіла CB3.2 (A), і порівнювали з індексом проліферації клітин, активованих у присутності неміченого антитіла CB3.2 (B). Фіг. 24 (Верхня і нижня панелі). Виснаження мишачих В-клітин in vivo у HCD16A/Bтрансгенних мишей з використанням MGD261 Мишам mCD32-/hCD16A+C57B1/6, mCD32-/hCD32B+C57B1/6 і mCD32-/hCD16A+hCD32B+C57B1/6, виведеним шляхом схрещування клітинних колоній MacroGenics, i.v. ін'єктували MGD261 (10, 3, 1 або 0,3 мг/кг) або нерелевантне антитіло (hE16, 10 мг/кг) на дні 0, 3, 7, 10, 14 і 17. Кров брали на день -19 (попередній забір крові) і на дні 4, 11, 18, 25 і 32 для FACS-аналізу. Здоров'я й активність тварин реєстрували три рази на тиждень. Верхня панель: 20 UA 111612 C2 5 10 15 20 25 30 35 40 45 50 55 h2B6-3G8 і WNV mAb; нижня панель: миші, яким уводили h2B6-3G8-hCD16A або -hCD32B, і миші, яким уводили WNV mAb-hCD16A або - hCD32B. Фіг. 25. Виснаження мишачих В-клітин in vivo у HCD16A/B-трансгенних мишей з використанням 2.4G2-3G8 DB Мишам mCD16-/-, mCD16-/- hCD16A+C57B1/6, mCD16-/- hCD16B+ і mCD16-/hCD16A+hCD16B+, виведеним шляхом схрещування клітинних колоній MacroGenics, i.р. ін'єктували 2.4G2-3G8 DB (75 мкг/миша) або PBS на дні 0, 2, 4, 7, 9, 11, 14, 16 і 18. Кров брали на день -10 (попередній забір крові) і на дні 4, 11 і 18 для FACS-аналізу. Здоров'я й активність тварин реєстрували три рази на тиждень. Фіг. 26. Ілюстрація протипухлинної активності MGD261 на моделі, якій внутрішньовенно (i.v.) уводили людські пухлинні клітини лінії RAJI 12-20-тижневим мишам mCD16-/-, hCD16A+, RAG1-/- C57B1/6, виведеним шляхом 6 схрещування клітинних колоній MacroGenics, i.v. ін'єктували 5×10 клітин Raji на день 0. На дні 6, 9, 13, 16, 20, 23, 27 і 30 мишей внутрішньоочеревинно обробляли (i.p.) 250, 25 або 2,5 мкг MGD261 або PBS (негативний контроль). Потім мишей щодня спостерігали, і масу тіла реєстрували два рази на тиждень. Мишей, у яких розвивався параліч задніх кінцівок, умертвляли. Фіг. 27. Експресія DART у хазяїнів, які не є ссавцями Клітини BL21DE3 (Novagen) трансформували плазмідою pET25b(+) T7-lac+3G8/3G8, і колонії, резистентні до ампіциліну, використовували для посіву бульйонної культури. Коли культура досягала OD600=0,5 одиниць, для індукування експресії додавали 0,5 мM IPTG. Культуру вирощували при 30 °C протягом 2 годин і збирали безклітинне середовище. Фіг. 28. ELISA DART ELISA-аналіз на зв'язування з h3G8-h3G8 DART проводили на 96-ямкових планшетах Maxisorp. Після реакції планшет три рази промивали PBS-T і виявляли з використанням 80 мкл/ямку субстрату TMB. Після 5-хвилинного інкубування реакцію припиняли додаванням 40 мкл/ямку 1 % H2SO4. OD450 нм зчитували на 96-ямковому планшеті-рідері і реєстрували з використанням комп'ютерної програми SOFTmax. За даними зчитування будували криву за допомогою комп'ютерної програми GraphPadPrism 3.03. Фіг. 29. DART-індукована загибель людських B-клітин Людські МКПК інкубували протягом ночі з вказаними молекулами. Апоптоз оцінювали за + + + допомогою FACS-аналізу, і дані виражали як відсоток PI -анексин-V -популяцій В-клітин (CD20 клітин) від загального числа популяцій, що не пройшли FSC/SSC-сортинг. Фіг. 30. Конструкції 8B5-CB3.1 DART Для ілюстрації даного винаходу була отримана множина конструкцій 8B5-CB3.1 DART. Конструкції 5 і 6, або 6 і 7, або 8 і 9, або 9 і 10, які кодуються експресійними плазмідами, спільно переносили в клітини HEK-293 для експресії 8B5-CB3.1 DART у присутності або під час відсутності антитіла проти flag-мітки з використанням ліпофектаміну 2000 (Invitrogen). Кондиціоноване середовище збирали три рази в кожні три дні. Потім кондиціоноване середовище очищали на CD32B-афінній колонці. Фіг. 31. ELISA 8B5-CB3.1 DART ELISA-аналіз на конкурентне зв'язування з 8B5-c3.1 DART/ch8b5 проводили на 96-ямкових планшетах Maxisorp. Після реакції планшет три рази промивали PBS-T і виявляли з використанням 80 мкл/ямку субстрату TMB. Після 5-хвилинного інкубування реакцію припиняли додаванням 40 мкл/ямку 1 % H2SO4. OD450 нм зчитували на 96-ямковому планшеті-рідері і реєстрували з використанням комп'ютерної програми SOFTmax. За даними зчитування будували криву за допомогою комп'ютерної програми GraphPadPrism 3.03. Фіг. 32. Схематична ілюстрація чотиривалентної структури DART На цій фігурі проілюстрована загальна структура молекул DART, отриманих шляхом збирання чотирьох поліпептидних ланцюгів. Чотири антигензв'язувальні домени Ig-подібного DART показані у вигляді смуг і темно-сірих еліпсів. Фіг. 33. Ig-подібний чотиривалентний DART На цій фігурі схематично представлені епітоп-зв'язувальні сайти Ig-подібного чотиривалентного DART. Фіг. 34. ELISA-аналіз на зв'язування mCD32-hCD16A На цій фігурі представлені результати ELISA-аналізу, які показали, що молекули Igподібного чотиривалентного DART приклада 6.10 зв'язуються з антигеном з більшою афінністю, ніж контрольне антитіло (ch-mCD32 mAb) або інші молекули DART. Фіг. 35. Схематична ілюстрація Ig-подібних молекул DART 21 UA 111612 C2 5 10 15 20 25 30 35 40 45 50 55 60 На цій фігурі схематично представлені Ig-подібні молекули DART. Специфічність показана у вигляді затінених, візерункових або білих ділянок, константні ділянки зафарбовані чорними, а дисульфідні зв'язки показані чорними пунктирними лініями. На цій фігурі N-кінці всіх білкових ланцюгів знаходяться зверху, а C-кінці всіх білкових ланцюгів знаходяться внизу. Панелі A-E представляють біспецифічні антитіла, а панелі F-J представляють триспецифічні антитіла. Панелі A і E представляють тривалентні антитіла. Панелі B, C, F, I і J представляють шестивалентні антитіла. Панелі D, G і H представляють восьмивалентні антитіла. Докладний опис окремих доменів представлено на фігурах 1, 2, 9, 14 і 17 і в Розділі 3.1. Фіг. 36. Активність зв'язування біспецифічного антитіла HU2B6 4.5-HU3G8 5.1 На фігурі 36 проілюстрована здатність біспецифічного антитіла Hu2B6 4.5-Hu3G8 5.1 (квадрати) зв'язуватися з CD32b і CD16a у порівнянні з діатілами Hu2B6 4.5 або Hu3G8 5.1 (трикутники). Фіг. 37. Схематичне представлення E-спіральних і K-спіральних похідних DART На фігурі 37 проілюстрована загальна конформація E-спіральних і K-спіральних похідних DART. Фіг. 38. Спіральна структура переважних E-спіральних і K-спіральних розділювальних послідовностей На фігурі 38 показана спіральна структура переважної "E-спіральної" послідовності (EVAALEK)4 (SEQ ID NO: 299) і переважної "K-спіральної" послідовності (KVAALKE)4 (SEQ ID NO: 300). Фіг. 39. Fc-вмісні E-спіральні і K-спіральні похідні DART На фігурі 39 проілюстровані різні типи Fc-вмісних E-спіральних і K-спіральних похідних DART, які можуть бути утворені за допомогою перестановки ланцюгів. Фіг. 40. Ексклюзійна хроматографія на E-спіральних і/або K-спіральних похідних і на Fcвмісних E-спіральних і/або K-спіральних похідних h2B6YAhCB3 DART На фігурі 40 представлені результати ексклюзійної хроматографії на E-спіральних і/або Kспіральних похідних і Fc-вмісних E-спіральних і/або K-спіральних похідних h2B6YAhCB3 DART. Було проаналізовано чотири види таких молекул, кожна з яких мала Е-спіраль і К-спіраль: EK (без Fc-ділянки), 2,1 мг; EFc/K (Fc, зв'язаний з E-спіраллю), 2,7 мг; E/KFc (Fc, зв'язаний з Kспіраллю), 1,8 мг; EFc/KFc (Fc, зв'язаний з K-спіраллю і E-спіраллю того самого DART), 2,0 мг. Фіг. 41. Структура отриманих димерних молекул На фігурі 41 показана можлива структура отриманої димерної молекули, ідентифікованої на ексклюзійному хроматографі, представленому на фігурі 40. Фіг. 42. Аналіз E-спіральних і/або K-спіральних похідних і Fc-вмісних E-спіральних і/або Kспіральних похідних h2B6YAhCB3 DART, проведений за допомогою електрофорезу в поліакриламідному гелі з ДСН На фігурі 42 представлені результати аналізу фракцій E-спіральних і/або K-спіральних похідних і Fc-вмісних E-спіральних і/або K-спіральних похідних h2B6YAhCB3 DART, отриманих за допомогою ексклюзійної хроматографії (фігура 40), де вказаний аналіз проводили за допомогою електрофорезу в поліакриламідном гелі з ДСН. Крайні доріжки: контрольні молекулярні маркери; доріжка 1: EK (без Fc-ділянки); доріжка 2: EFc/K, агрегована фракція; доріжка 3: EFc/K, мономерна фракція; доріжка 4: E/KFc, агрегована фракція; доріжка 5: E/KFc, мономерна фракція; доріжка 6: EFc/KFc, агрегована фракція; доріжка 7: EFc/KFc, димерна фракція; доріжка 8: EFc/KFc, мономерна фракція. Фіг. 43. ELISA-аналіз на біспецифічне зв'язування На фігурі 43 представлені результати порівняльного ELISA-аналізу на біспецифічне зв'язування Fc-вмісних E-спіральних/K-спіральних похідних h2B6YAhCB3 DART (EFc/K або E/KFc), h2B6YAhCB3 DART, контролю і похідного EFc/KFc h2B6YAhCB3 DART. Фіг. 44. Здатність E-спіральних і/або K-спіральних похідних і Fc-вмісних E-спіральних і/або Kспіральних похідних h2B6YAhCB3 DART інгібувати проліферацію Т-клітин На фігурі 44 проілюстрована здатність E-спіральних і/або K-спіральних похідних і Fc-вмісних E-спіральних і/або K-спіральних похідних h2B6YAhCB3 DART інгібувати проліферацію Т-клітин. Фіг. 45. hCD16-hCD32B ABD-DART На фігурі 45 схематично представлена рекомбінантна молекула антитіла hCD16-hCD32B ABD-DART, яка складається з домену ABD3 стрептококового білка G, злитого з рекомбінантним біспецифічним DART, що вступає в імунну реакцію з антигенами hCD16 і hCD32B. Фіг. 46A-46J. Афінність зв'язування hCD16-hCD32B ABD-DART ELISA-планшети покривали антигеном CD16 (фігура 46A) або альбуміном людської сироватки (фігура 46B) у концентрації 2 мкг/мл. hCD16-hCD32B ABD-DART ( ) і контрольний hCD16-hCD32B DART (o) зв'язувалися при різних концентраціях, починаючи з 2 мкг/мл. У 22 UA 111612 C2 5 10 15 20 25 30 35 40 45 50 55 планшет додавали біотинільований sCD32B, а потім додавали ПХ-кон'югований стрептавідин для детектування. На фігурі 46C показано, що молекула ABD-DART має здатність одночасно зв'язуватися з CD32B, CD16A і HSA. На фігурах 46D-46I показано, що hCD16-hCD32B ABDDART і його ABD-гібридне похідне мають еквівалентну здатність зв'язуватися з розчинним CD32B (sCD32B) і з розчинним CD16A(158F) (sCD16A). На фігурі 46J показано, що ABD-гібрид DART зберігає здатність батьківського DART до біспецифічного зв'язування з sCD16A і CD32B незалежно від присутності альбуміну людської сироватки. Фіг. 47A/47B. Цитотоксичність білків DART На фігурі 47A проілюстрована МКПК-опосередковує цитотоксичність білків DART. Аналізи на ADCC здійснювали з використанням людських B-клітинних ліній Дауді як клітин-мішеней, інкубованих із клітинами МКПК, використовуваними як ефекторі клітини. Окремі аналізи проводили з трьома повторностями з використанням клітин у відношенні ефектор:мішень = 20:1 і з титруванням антитіл: hCD16A-hCD32B DART (-) і hCD16A-hCD32B ABD DART ( ). Клітинноопосередковувану цитотоксичність оцінювали в аналізі на вивільнення LDH. Нижня крива при 0 10 відповідає hCD16A-hCD32B ABD DART ( ). На фігурі 47B показано, що CD16xCD32B DART зв'язується з CD16B-експресуючими ефекторними клітинами, що приводить до рекрутингу вказаних клітин і знищення (за допомогою перенаціленого цитолізу) клітин, експресуючих CD32B. Фіг. 48. Поліпшені фармакокінетичні властивості hCD16-hCD32B ABD-DART у мишей C57B1/6 Мишам (n=3) вводили одну внутрішньовенну ін'єкцію (A) hCD16-hCD32B ABD-DART (-) і (B) hCD16-hCD32B DART (▲) при 5 мг/кг. Мишачу сироватку брали в різні періоди часу, і концентрацію білка в сироватці оцінювали за допомогою ELISA. Оцінку фармакокінетичних властивостей проводили за допомогою програми WinNonlin Professional 5.1. Фіг. 49A-49C. Біологічна активність ABD-DART in vivo На фігурах 49A-49C продемонстровано, що hCD16-hCD32B ABD-DART, після його введення мишам in vivo, зберігав свою біологічну активність. Було виявлено, що одна доза ABD-DART або батьківського DART, уведена Tg(mCD16-/-hCD16A+32B+)- мишам, приводить до селективного зниження концентрацій В-клітин (фігура 49A), але не чинить впливу на концентрації Т-клітин (фігура 49B) або мононуклеарних периферичних клітин (фігура 49C). Фіг. 50A-50E. Активність HER2×TCRb DART на панелі клітинних ліній, експресуючих низькі рівні HER2 Молекули DART, що мають домени, які зв'язуються з Her2 і з T-клітинним рецептором (TCR), тестували на їхню здатність опосередковувати цитотоксичність клітинних ліній при первинномножинному раку молочної залози, раку товстої кишки і раку сечового міхура, де вказані клітинні лінії були охарактеризовані як клітинні лінії, що мають низькі рівні експресії HER2 (а значить, були не сприйнятливими до лікування анти-Her2/neu антитілом, Herceptin®). Тестованими клітинними лініями раку молочної залози були ZR75-1 (HER2 2+) (фіг. 50A), MCF-7 (HER2 1+) (фіг. 50B) і MDA-MB468 (HER 2-ve) (фіг. 50C). Тестованими клітинними лініями, що не стосуються раку молочної залози, були HT-29 (клітинна лінія раку товстої кишки) (фіг. 50D) і SW780 (клітинна лінія раку сечового міхура) (фіг. 50E). Фіг. 51A-51B. Очищення поліпептидів KYK2VL-4420VH-GFNRGEC (SEQ ID NO: 313) - Eспіраль і 4420VL-KYK2VH-GVEPKSC (SEQ ID NO: 314) На фіг. 51A-51B проілюстроване очищення поліпептидів KYK2VL-4420VH-GFNRGEC (SEQ ID NO: 313) - E-спіраль і 4420VL-KYK2VH-GVEPKSC (SEQ ID NO: 314). Фіг. 52. Специфічність зв'язування KYK2-4420 VF DART На фіг. 52 проілюстрована специфічність зв'язування KYK2-4420 VF DART. Фіг. 53. Конструювання плазміди pPAL7 KCOIL-ABD На фіг. 53 схематично представлений опис конструкції плазміди pPAL7 Kcoil-ABD. Фіг. 54. Вестерн-блот-аналіз K-спірального білка ABD, очищеного на смолі для очищення PROFINITY EXACT™ На фігурі 54 проілюстрований вестерн-блот-аналіз K-спірального білка ABD, очищеного на смолі для очищення PROFINITY EXACT™ (у невеликому масштабі виходячи з 10 мл культури). Доріжки 1 і 5: неочищені лізати; доріжки 2 і 6: протоковий матеріал, доріжки 3, 4, 8 і 9: очищений білок; доріжка 7: маркер. "Перше" антитіло: кроляче поліклональне анти-EK антитіло; "друге" антитіло: ПХ-кон'юговане антикроляче антитіло. Фіг. 55. ДСН-ПААГ-аналіз K-спірального білка ABD, очищеного на смолі для очищення PROFINITY EXACT™ 23 UA 111612 C2 5 10 15 20 25 30 35 40 45 50 55 60 На фігурі 55 представлені результати ДСН-ПААГ-аналіза фракцій, елюйованих зі смоли для очищення PROFINITY EXACT™. L: неочищений лізат E. coli; FT: протоковий матеріал; W: промивання; M: маркер. Фіг. 56A-56C. ELISA-аналіз на зв'язування KYK2 4420 EK ABD DART з подвійною афінністю На фігурах 56A-56C представлені результати ELISA-аналізу, які продемонстрували, що комплекси KYK2-4420 VF DART з молекулою K-спіраль-ABD, що утворюють E-спіраль - Kспіраль-ABD DART, мають поліпшені властивості. Фігура 56A: DART захоплювалися NKG2D-Fc і були детектовані з використанням біотинільованого моноклонального анти-EK антитіла. Фігура 56B: DART захоплювалися моноклональним анти-EK антитілом і були детектовані з використанням ПХ-людського альбуміну. Фігура 56C: DART захоплювалися NKG2D-Fc і були детектовані з використанням біотинільованого анти-ФІТЦ антитіла. Результати ELISA-аналізу показали, що всі домени в цьому комплексі можуть зв'язуватися з їх відповідними лігандами. Фіг. 57. ELISA-аналіз деімунізованого ABD На фігурі 57 представлені результати ELISA, що продемонстрували зміну рівня зв'язування, обумовлена модифікаціями в послідовності альбумін-зв'язувальних доменів. Фіг. 58. Аналіз на зв'язування з альбуміном На фігурі 58 представлені результати аналізів на зв'язування з альбуміном для ABDваріантів Y61A/I65A, Y61A/N66D, L64A/I65A* і L64A/N66D і для ABD дикого типу. Фіг. 59. Аналіз на зв'язування з альбуміном На фігурі 59 представлені результати аналізів на зв'язування з альбуміном для ABDваріантів L64G/N66D, I65A/N66D, N66S/T70S* і Y61A/N66S/T70S і для ABD дикого типу. Фіг. 60. Аналіз на зв'язування з альбуміном На фігурі 60 представлені результати аналізів на зв'язування з альбуміном для ABDваріантів Y60A/Y61A, Y60T/Y61A, I65A/N66D/V71A, Y61A/I65A/V71A, L64A/N66D/V71A і для ABD дикого типу. Фіг. 61. Аналіз на зв'язування з альбуміном На фігурі 61 представлені результати аналізів на зв'язування з альбуміном для ABDваріантів Y60A/Y61A/V71A, Y60T/Y61A/V71A, L64A/I65A/V71A і для ABD дикого типу. Фіг. 62. Аналіз на зв'язування з альбуміном На фігурі 62 представлені результати аналізів на зв'язування з альбуміном для ABDваріантів L64G/I65A/V71A, L64G/N66D/V71A, Y61A/T70S, N66S, T70S, Y61A/N66S, L64A/I65A і для ABD дикого типу. Фіг. 63. Поліпшені фармакокінетичні властивості CD16xCD32B ABD-DART і варіантів ABDDART у мишей C57BL/6 На фігурі 63 проілюстровані поліпшені фармакокінетичні властивості CD16xCD32B ABDDART і варіантів ABD-DART у мишей C57BL/6. Мишам (n=3) вводили одну внутрішньовенну ін'єкцію білків ABD-DART при 5 мг/кг. Мишачу сироватку брали в різні періоди часу, і концентрацію білка в сироватці оцінювали за допомогою ELISA. Оцінку фармакокінетичних властивостей проводили за допомогою програми WinNonlin Professional 5.1. Фіг. 64A і 64B. Полівалентні білки ABD DART На фігурах 64A і 64B проілюстровані структури DART, що містять полівалентні ABD, сконструйовані шляхом приєднання ABD по кінцях обох ланцюгів (hCD16xhCD32B E ABD/K ABD DART) (фігура 64A) або шляхом приєднання тандемних ABD по кінці одного ланцюга (hCD16xhCD32B E ABD ABD/K DART) (фігура 64B). Фіг. 65(A-C). Біспецифічна активність hCD16xhCD32B ABD DART На фігурах 65A-65C проілюстрована біспецифічна активність hCD16xhCD32B ABD DART, оцінена за допомогою ELISA з подвійною специфічністю. ELISA-планшети покривали 2 мкг/мл антигени CD16. ABD DART і Fc DART у різних концентраціях зв'язували з планшетом. І нарешті, у планшет додавали біотинільований антиген sCD32B, а потім ПХ-кон'югований стрептавідин для детектування. Фіг. 66. Час напівжиття ABD DART і FC DART у сироватці На фігурі 66 представлені результати дослідження фармакокінетичних властивостей in vivo, у якому мишам C57B1/6 вводили одну i.v. ін'єкцію ABD DART або Fc DART (5 мг/кг) для оцінки часу їхнього напівжиття в сироватці. 5. Опис переважних варіантів винаходу Кожен поліпептидний ланцюг молекули діатіла містить VL- домен і VH-домен, які були ковалентно зв'язані так, щоб це запобігало самозбиранню. Взаємодія двох поліпептидних ланцюгів приводить до продукування пар VL-VH, що утворюють два епітоп-зв'язувальні сайти, тобто двовалентну молекулу. Жоден з VH- або VL-доменів не обмежений яким-небудь положенням у поліпептидному ланцюзі, тобто, він обмежений тільки аміно (N)- або карбокси (C) 24 UA 111612 C2 5 10 15 20 25 30 35 40 45 50 55 60 кінцем, і жоден з цих доменів не обмежений їхніми положеннями один відносно одного, тобто, VL-домен може бути N-кінцевим відносно VH-домену і навпаки. Одним лише обмеженням є те, що комплементарний поліпептидний ланцюг повинний бути доступним для утворення функціонального діатіла. Якщо VL- і VH-домени походять від того самого антитіла, то два комплементарні поліпептидні ланцюги можуть бути ідентичними. Так, наприклад, якщо зв'язувальні домени походять від антитіла, специфічного до епітопу А (тобто, якщо зв'язувальний домен утворюється в результаті взаємодії VLA-VHA), то кожен поліпептид буде містити VHA і VLA. Гомодимеризація двох поліпептидних ланцюгів антитіла буде приводити до утворення двох VLA-VHA-зв'язувальних сайтів і, тим самим, до утворення двовалентного моноспецифічного антитіла. Якщо VL- і VH-домени походять від антитіл, специфічних до різних антигенів, то для утворення функціонального біспецифічного діатіла необхідна взаємодія двох різних поліпептидних ланцюгів, тобто утворення гетеродимера. Так, наприклад, для біспецифічного діатіла один поліпептидний ланцюг буде містити VL A і VLB; а гомодимеризація вказаного ланцюга буде приводити до утворення двох VLA-VHB-зв'язувальних сайтів, які або не зв'язуються, або їхнє зв'язування є непередбаченим. На противагу цьому, якщо два відмінні один від одного поліпептидні ланцюги є вільними для взаємодії, наприклад, у рекомбінантній експресійній системі, тобто в одній системі, що містить VLA і VHB, і в іншій системі, що містить VLB і VHA, то два відмінні один від одного зв'язувальні сайти будуть утворювати VL A-VHA і VLBVHB. Для всіх пар поліпептидних ланцюгів діатіл існує імовірність помилкового вирівнювання або помилкового зв'язування на двох ланцюгах, тобто взаємодії доменів VL-VL або VH-VH, однак, очищення функціональних діатіл може бути легко здійснене виходячи з імуноспецифічності "правильно" димеризованого зв'язувального сайта з застосуванням будьякого афінного методу, відомого фахівцям або описаного в даній заявці, наприклад, методу афінної хроматографії. В інших варіантах винаходу один або більше поліпептидних ланцюгів діатіла містять Fcдомен. Fc-домени в поліпептидних ланцюгах молекул діатіла переважно димеризуються з утворенням молекули діатіла, що має імуноглобулін-подібні властивості, наприклад, здатність до Fc-FсγR-взаємодій. Fc-вмісні діатіла можуть бути димерами, наприклад, вони можуть складатися з двох поліпептидних ланцюгів, кожен з який містить VH-домен, VL-домен і Fcдомен. Димеризація вказаних поліпептидних ланцюгів приводить до утворення двовалентного діатіла, що містить Fc-домен, хоча структура цього антитіла відрізняється від структури немодифікованого двовалентного антитіла (фіг. 11). Такі молекули діатіла, на відміну від імуноглобуліну дикого типу, мають змінені фенотипи, наприклад, вони мають змінений час напівжиття в сироватці, модифіковані зв'язувальні властивості і т. п. В інших варіантах винаходу молекули діатіла, що містять Fc-домени, можуть бути тетрамерами. Такі тетрамери містять два "більш важкі" поліпептидні ланцюги, тобто поліпептидний ланцюг, що містить VL-, VH- і Fcдомен, і два "більш легкі" поліпептидні ланцюги, тобто поліпептидний ланцюг, що містить VL і VH. Вказані "більш легкі" і "більш важкі" ланцюги взаємодіють один з одним з утворенням мономера, а вказані мономери взаємодіють один з одним за допомогою їх неспарених Fcдоменів з утворенням Ig-подібної молекули. Таке Ig-подібне діатіло є чотиривалентним і може бути моноспецифічним, біспецифічним або тетраспецифічним. Щонайменше два зв'язувальні сайти молекули діатіла можуть розпізнавати той самий або різні епітопи. Різні епітопи можуть походити від того самого антигену або від різних антигенів. В одному з варіантів винаходу епітопи походять від різних клітин. В іншому варіанті винаходу епітопами є антигени клітинної поверхні, які присутні на одній і тій же клітині або на тому самому вірусі. Епітоп-зв'язувальні сайти можуть розпізнавати будь-який антиген, проти якого може продукуватися антитіло. Так, наприклад, такими молекулами є білки, нуклеїнові кислоти, бактеріальні токсини, маркери клітинної поверхні, аутоімунні маркери, вірусні білки, лікарські засоби і т. п. У конкретних аспектах винаходу щонайменше один епітоп-зв'язувальний сайт діатіла є специфічним до антигену на конкретній клітині, такій як B-клітина, T-клітина, фагоцит, природна клітина-кілер (NK) або дендритна клітина. Кожен домен поліпептидного ланцюга діатіла, тобто VL-, VH- і Fс-домени, можуть бути розділені пептидним лінкером. Пептидний лінкер може мати довжину в 0, 1, 2, 3, 4, 5, 6, 7, 8 або 9 амінокислот. У деяких варіантах винаходу амінокислотна лінкерна послідовність являє собою послідовність GGGSGGGG (SEQ ID NO: 10), яка кодується послідовністю нуклеїнової кислоти (SEQ ID NO: 74). У деяких варіантах винаходу кожен поліпептидний ланцюг молекули діатіла був сконструйований так, щоб він містив щонайменше один цистеїновий залишок, який буде взаємодіяти з аналогом у положенні щонайменше одного цистеїнового залишку на другому поліпептидному ланцюзі відповідно до винаходу з утворенням міжланцюгового дисульфідного 25 UA 111612 C2 5 10 15 20 25 30 35 40 45 50 55 60 зв'язку. Вказані міжланцюгові дисульфідні зв'язки служать для стабілізації молекули діатіла, підвищують рівень експресії і вихід продукту в рекомбінантних системах, дозволяють одержати стабільний препарат з однорідним складом, а також підвищують стабільність виділеного і/або очищеного продукту in vivo. Вказаний щонайменше один цистеїновий залишок може бути введений як одна амінокислота або як частина більш великої амінокислотної послідовності, наприклад, шарнірного домену, у будь-яку частину поліпептидного ланцюга. У конкретному варіанті винаходу вказаний щонайменше один цистеїновий залишок уводять так, щоб він був присутній біля С-кінця поліпептидного ланцюга. У деяких варіантах винаходу вказаний щонайменше один цистеїновий залишок вводять у поліпептидний ланцюг амінокислотної послідовності LGGC. У конкретному варіанті винаходу C-кінець поліпептидного ланцюга, що входить до складу молекули діатіла відповідно до винаходу, містить амінокислотну послідовність LGGC. В іншому варіанті винаходу вказаний щонайменше один цистеїновий залишок вводять у поліпептид, який присутній в амінокислотній послідовності, що містить шарнірний домен, наприклад, SEQ ID NO: 1 або SEQ ID NO: 4. У конкретному варіанті винаходу C-кінець поліпептидного ланцюга молекули діатіла відповідно до винаходу містить амінокислотну послідовність шарнірного домену IgG, наприклад, SEQ ID NO: 1. В іншому варіанті винаходу C-кінець поліпептидного ланцюга молекули діатіла відповідно до винаходу містить амінокислотну послідовність VEPKSC (SEQ ID NO: 77), що може кодуватися нуклеотидною послідовністю (SEQ ID NO: 78). В інших варіантах винаходу вказаний щонайменше один цистеїновий залишок вводять у поліпептидний ланцюг в амінокислотній послідовності LGGCFNRGEC (SEQ ID NO: 17), що може кодуватися нуклеотидною послідовністю (SEQ ID NO: 76). У конкретному варіанті винаходу C-кінець поліпептидного ланцюга, що входить до складу молекули діатіла відповідно до винаходу, містить амінокислотну послідовність LGGCFNRGEC (SEQ ID NO: 17), що може кодуватися нуклеотидною послідовністю (SEQ ID NO: 76). В інших варіантах винаходу вказаний щонайменше один цистеїновий залишок вводять у поліпептидний ланцюг в амінокислотній послідовності FNRGEC (SEQ ID NO: 23), що може кодуватися нуклеотидною послідовністю (SEQ ID NO: 75). У конкретному варіанті винаходу C-кінець поліпептидного ланцюга, що входить до складу молекули діатіла відповідно до винаходу, містить амінокислотну послідовність FNRGEC (SEQ ID NO: 23), що може кодуватися нуклеотидною послідовністю (SEQ ID NO: 75). У деяких варіантах винаходу молекула діатіла містить щонайменше два поліпептидні ланцюги, кожен з яких включає амінокислотну послідовність LGGC і ковалентно зв'язаний дисульфідного зв'язком, розташованим між цистеїновими залишками в вказаних послідовностях LGGC. В іншому конкретному варіанті винаходу молекули діатіла містять щонайменше два поліпептидні ланцюги, один з яких включає послідовність FNRGEC (SEQ ID NO: 23), а інший включає шарнірний домен (який містить щонайменше один цистеїновий залишок), де вказані щонайменше два поліпептидні ланцюги ковалентно зв'язані дисульфідним зв'язком, розташованим між цистеїновим залишком у FNRGEC (SEQ ID NO: 23) і цистеїновим залишком у шарнірному домені. У конкретних аспектах винаходу цистеїновий залишок, відповідальний за утворення дисульфідного зв'язку в шарнірному домені, являє собою Cys-128 (пронумерований відповідно до нумерації EU Кебата і розташований у шарнірному домені важкого ланцюга немодифікованого інтактного IgG), а цистеїновий залишок аналога в SEQ ID NO: 23 являє собою Cys-214 (пронумерований відповідно до нумерації EU Кебата і розташований у шарнірному домені легкого ланцюга немодифікованого інтактного IgG) (Elkabetz et al. (2005) "Cysteines In CHI Underlie Retention Of Unassembled Ig Heavy Chains" J. Biol. Chem. 280:1440214412; ця публікація у всій своїй повноті вводиться в даний опис за допомогою посилання). В інших варіантах винаходу щонайменше один цистеїновий залишок сконструйований так, щоб він був присутній біля N-кінця амінокислотного ланцюга. В інших варіантах винаходу щонайменше один цистеїновий залишок сконструйований так, щоб він був присутній в лінкерній частині поліпептидного ланцюга молекули діатіла. В інших варіантах винаходу VH- або VLдомен сконструйований так, щоб він містив щонайменше одну амінокислотну модифікацію в порівнянні з батьківським VH- або VL-доменом, де вказана амінокислотна модифікація включає заміну батьківської амінокислоти цистеїном. Даний винахід охоплює молекули діатіл, які містять Fc-домен або його частину (наприклад, CH2-домен або CH3-домен). Fc-домен або його частину можуть походити від імуноглобуліну будь-якого ізотипу або алотипу, включаючи, але не обмежуючись ними, IgA, IgD, IgG, IgE і IgM. У переважних варіантах винаходу Fc-домен (або його частина) походить від IgG. У конкретних варіанті винаходу ізотипами IgG є IgG1, IgG2, IgG3 або IgG4 або їх алотипи. В одному з варіантів винаходу молекула діатіла містить Fc-домен, де вказаний Fc-домен включає CH2домен і CH3-домен, незалежно вибрані з імуноглобулінів будь-яких ізотипів (тобто, Fc-домен, 26 UA 111612 C2 5 10 15 20 25 30 35 40 45 50 55 60 який містить CH2-домен, походить від IgG, а CH3-домен походить від IgG, або CH2-домен походить від IgG1, а CH3-домен походить від IgG2, і т. п.). Вказаний Fc-домен може бути введений у поліпептидний ланцюг, який складає молекулу діатіла відповідно до винаходу, у будь-якому положенні відносно інших доменів або частин вказаного поліпептидного ланцюга (наприклад, Fc-домен або його частина можуть бути С-кінцевими відносно VL- і VH-доменам поліпептиду вказаного ланцюга; N-кінцевими відносно VL- і VH-доменам; або N-кінцевими відносно одного домену і С-кінцевими відносно іншого домену (тобто, вони можуть бути розташовані між двома доменами поліпептидного ланцюга)). Даний винахід також охоплює молекули, що містять шарнірний домен. Шарнірний домен походить від імуноглобуліну будь-якого ізотипу або алотипу, включаючи IgA, IgD, IgG, IgE і IgM. У переважних варіантах винаходу шарнірний домен походить від IgG, де IgG має ізотип IgG1, IgG2, IgG3 або IgG4 або їх алотип. Вказаний шарнірний домен може бути введений у поліпептидний ланцюг, який складає молекулу діатіла, разом з Fc-доменом, так, щоб молекула діатіла містила шарнірний домен-Fc-домен. У деяких варіантах винаходу шарнірний домен і Fcдомен незалежно вибрані з імуноглобуліну будь-якого ізотипу, відомого фахівцям або описаного в даній заявці. В інших варіантах винаходу шарнірний домен і Fc-домен розділені щонайменше одним іншим доменом поліпептидного ланцюга, наприклад, VL-доменом. Шарнірний домен або необов'язково шарнірний домен-Fc-домен можуть бути введені в поліпептид відповідно до винаходу в будь-якому положенні відносно інших доменів або частин вказаного поліпептидного ланцюга. У деяких варіантах винаходу поліпептидний ланцюг відповідно до винаходу містить шарнірний домен, де вказаний шарнірний домен знаходиться біля С-кінця поліпептидного ланцюга, де вказаний поліпептидний ланцюг не містить Fc-домену. В інших варіантах винаходу поліпептидний ланцюг відповідно до винаходу містить шарнірний домен-Fc-домен, де вказаний шарнірний домен-Fc-домен знаходиться біля С-кінця поліпептидного ланцюга. В інших варіантах винаходу поліпептидний ланцюг відповідно до винаходу містить шарнірний домен-Fcдомен, що знаходиться біля N-кінця поліпептидного ланцюга. Як обговорювалося вище, даний винахід охоплює мультимери поліпептидних ланцюгів, кожен з яких містить VH- і VL-домени. У деяких аспектах винаходу поліпептидні ланцюги в вказаних мультимерах також містять Fc-домен. Димеризація Fc-доменів приводить до утворення молекули діатіла, що має імуноглобулін-подібні властивості, тобто Fcопосередковувані функціональні властивості (такі як, наприклад, взаємодія Fc-FcγR, зв'язування з комплементом і т. п.). У деяких варіантах винаходу VL- і VH-домени, що містять кожен поліпептидний ланцюг, мають однакову специфічність, а вказана молекула діатіла є двовалентною і моноспецифічною. В інших варіантах винаходу VL- і VH-домени, що містять кожен поліпептидний ланцюг, мають різні специфічності, а вказане діатіло є двовалентним і біспецифічним. В інших варіантах винаходу молекули діатіла відповідно до винаходу охоплюють тетрамери поліпептидних ланцюгів, кожен з яких містить VH- і VL-домени. У деяких варіантах винаходу два поліпептидні ланцюги тетрамера також включають Fc-домен. Отже, тетрамер складається з двох "більш важких" поліпептидних ланцюгів, кожен з яких містить VL-, VH- і Fc-домени, і двох "більш легких" поліпептидних ланцюгів, що містять VL- і VH-домени. Взаємодія "більш важкого" і "більш легкого" ланцюгів приводить до утворення двовалентного мономера, що, після димеризації за допомогою Fc-доменів "більш важких" ланцюгів, утворює чотиривалентну імуноглобулін-подібну молекулу (описану в прикладі 6.2 і в прикладі 6.3). У деяких аспектах винаходу вказані мономери є однаковими, а чотиривалентна молекула діатіла є моноспецифічною або біспецифічною. В інших аспектах винаходу вказані мономери відрізняються один від одного, а чотиривалентна молекула діатіла є біспецифічною або тетраспецифічною. Для утворення тетраспецифічної молекули діатіла, описаної вище, необхідна взаємодія чотирьох різних поліпептидних ланцюгів. Такі взаємодії представляють певні труднощі для їхнього ефективного здійснення в одноклітинній системі рекомбінантного продукування, що обумовлено множиною варіантів потенційно помилкових спарювань ланцюга. Однією з можливостей підвищення імовірності помилкових спарювань є введення мутацій типу "вузли в дірках" у потрібні пари поліпептидних ланцюгів. Такі мутації відбуваються переважно при гетеродимеризації, а не при гомодимеризації. Так, наприклад, що стосується Fc-Fc-взаємодій, то амінокислотна заміна (переважно, заміна амінокислотою, що містить об'ємну бічну групу, яка утворює "вузол", наприклад, триптофаном) може бути введена в CH2- або CH3-домени, так, щоб стеричне утруднення запобігало взаємодії з аналогічним чином мутованим доменом і змушувало мутований домен спарюватися з доменом, у який була введена комплементарна або адаптуюча мутація, тобто "діра" (наприклад, заміна гліцином). Такі серії мутацій можуть 27 UA 111612 C2 5 10 15 20 25 30 35 40 45 50 55 60 бути введені в будь-яку пару поліпептидів, що складають молекулу діатіла, і, крім того, у будьяку частину поліпептидних ланцюгів вказаної пари. Методи конструювання білків для досягнення переважно гетеродимеризації, а не гомодимеризації, добре відомі фахівцям, зокрема, фахівцям з конструювання імуноглобулін-подібних молекул, і ці методи описані в літературі (див., наприклад, публікації: Ridgway et al. (1996) "Knobs-Into-Holes" Engineering Of Antibody CH3 Domains For Heavy Chain Heterodimerization, "Protein Engr. 9:617-621, Atwell et al. (1997) "Stable Heterodimers From Remodeling The Domain Interface Of A Homodimer Using A Phage Display Library" J. Mol. Biol. 270:26-35, і Xie et al. (2005) "A New Format Of Bispecific Antibody: Highly Efficient Heterodimerization, Expression И Tumor Cell Lysis" J. Immunol. Methods 296:95-101; кожна з який у всій своїй повноті вводиться в даний опис за допомогою посилання). Даний винахід також охоплює молекули діатіла, які містять варіант Fc-домену або варіант шарнірного домену-Fc-домену (або їхньої частини), де вказаний варіант Fc-домену має щонайменше одну амінокислотну модифікацію (наприклад, заміну, інсерцію, делецію) у порівнянні з Fc-доменом дикого типу або шарнірним доменом-Fc-доменом дикого типу (або їх частинами). Молекули, що містять варіанти Fc-доменів або варіанти шарнірних доменів-Fcдоменів (або їхніх частин) (наприклад, антитіл), звичайно мають змінені фенотипи в порівнянні з молекулами, що містять Fc-домени або шарнірні домени-Fc-домени або їхні частини дикого типу. Змінений фенотип може бути охарактеризований як змінений час напівжиття в сироватці, змінена стабільність, змінена сприйнятливість до клітинних ферментів або змінена ефекторна функція, які були визначені в аналізі на NK-залежний цитоліз або макрофаг-залежний цитоліз. Варіанти Fc-домену, ідентифіковані як варіанти, які мають змінені ефекторні функції, описані в Міжнародній заявці WO04/063351, у публікаціях заявок на патент США 2005/0037000 і 2005/0064514, у попередній заявці на патент США 60/626510, поданій 10 листопада 2004 р., у попередній заявці на патент США 60/636663, поданій 15 грудня 2004 р., і в попередній заявці на патент США 60/781564, поданій 10 березня 2006 р., а також у заявці на патент США 11/271140, поданій 10 листопада 2005 р., і в заявці на патент США 11/305787, поданій 15 грудня 2005 р., і в одночасно поданих заявках авторів винаходу, які у всій своїй повноті вводяться в даний опис за допомогою посилання. Біспецифічні діатіла відповідно до винаходу можуть одночасно зв'язуватися з двома окремими і різними епітопами. У деяких варіантах винаходу епітопи походять від того самого антигену. В інших варіантах винаходу епітопи походять від різних антигенів. У переважних варіантах винаходу щонайменше один епітоп-зв'язувальний сайт є специфічним до детермінанти, яка експресується на імунних ефекторних клітинах (наприклад, CD3, CD16, CD32, CD64 і т. п.) і на T-лімфоцитах, природних клітинах-кілерах (NK) або на інших імунонуклеарних клітинах. В одному з варіантів винаходу молекула діатіла зв'язується з детермінантою ефекторної клітини, а також активує вказану ефекторну клітину. Відповідно до цього молекули діатіла відповідно до винаходу можуть мати Ig-подібні функціональні властивості незалежно від того, чи містять вони Fc-домен чи ні (як може бути проаналізовано в будь-якому аналізі на ефекторну функцію, відомому фахівцям або описаному в даній заявці (наприклад, в аналізі на ADCC)). У деяких варіантах винаходу біспецифічне діатіло відповідно до винаходу зв'язується з раковим антигеном на пухлинній клітині і з детермінантою ефекторної клітини й активує вказану клітину. В альтернативних варіантах винаходу біспецифічне діатіло або молекула діатіла відповідно до винаходу можуть інгібувати активацію мішені, наприклад, ефекторної клітини за допомогою одночасного зв'язування і тим самим об'єднання активуючого і інгібуючого рецепторів на одній і тій же клітині (наприклад, зв'язування з CD32A і CD32B, BCR і CD32B або IgERI і CD32B), як описано вище (див. розділ "Попередній рівень техніки"). В іншому аспекті цього варіанта біспецифічне діатіло може мати противірусні властивості завдяки одночасному зв'язуванню двох нейтралізуючих епітопів на вірусі (наприклад, епітопів RSV; епітопів WNV, таких як E16 і E53). У деяких варіантах винаходу молекули біспецифічних діатіл відповідно до винаходу мають унікальну здатність націлюватися на клітини конкретних типів. Так, наприклад, біспецифічне діатіло або молекула біспецифічного діатіла можуть бути сконструйовані так, щоб вони містили комбінацію епітоп-зв'язувальних сайтів, що розпізнають серію антигенів, унікальних для клітиниабо тканини-мішені конкретного типу. Крім того, якщо будь-який з окремих антигенів або обидва ці антигени є досить ізольованими, як це звичайно має місце, якщо вони знаходяться в тканині і/або в клітині інших типів, то для конструювання діатіла або молекули діатіла можуть бути використані домени з низькою афінністю зв'язування. Такі домени з низькою афінністю зв'язування будуть нездатні зв'язуватися з окремим епітопом або антигеном з авідністю, достатньою для терапевтичного застосування. Однак, якщо епітопи або антигени присутні на одній клітині- або тканині-мішені, то авідність діатіла або молекули діатіла для цієї або клітини 28
ДивитисяДодаткова інформація
Назва патенту англійськоюDeimmunized serum-binding domains and their use for extending serum half-life
Автори англійськоюBonvini, Ezio, Barat, Bhaswati, Huang, Ling, Johnson, Leslie, S.
Автори російськоюБонвини Эцио, Барат Бхасвати, Хуан Лин, Джонсон Лесли С.
МПК / Мітки
МПК: A61K 39/395, C07K 16/06, C07K 1/107
Мітки: їхнє, збільшення, домени, деімунізованою, застосування, зв'язуються, сироватці, сироваткою, часу, напівжиття
Код посилання
<a href="https://ua.patents.su/370-111612-domeni-yaki-zvyazuyutsya-z-deimunizovanoyu-sirovatkoyu-i-khneh-zastosuvannya-dlya-zbilshennya-chasu-napivzhittya-v-sirovatci.html" target="_blank" rel="follow" title="База патентів України">Домени, які зв’язуються з деімунізованою сироваткою, і їхнє застосування для збільшення часу напівжиття в сироватці</a>
Антитіла до людського tweak та їхнє застосування

Номер патенту: 105520
Опубліковано: 26.05.2014
Автори: Бенер Моніка, Кньотген Хендрік, Нівьонер Йєнс
МПК: C07K 16/28, A61P 35/00, A61K 39/395
Мітки: застосування, людського, антитіла, їхнє, tweak
Формула / Реферат:
1. Антитіло, що зв'язується з людським TWEAK, яке відрізняється тим, що міститьa) CDR1H, послідовність якого представлена у SEQ ID NO:22, CDR2H, послідовність якого представлена у SEQ ID NО:23, CDR3H, послідовність якого представлена у SEQ ID NО:24, і CDR1L, послідовність якого представлена у SEQ ID NО:18, CDR2L, послідовність якого представлена у SEQ ID NO: 19, CDR3L, послідовність якого представлена у SEQ ID NО:20; абоb)...
Химерні віруси, які представляють неприродні поверхневі білки, і їхнє застосування

Номер патенту: 96140
Опубліковано: 10.10.2011
Автори: Гастрія-Састре Адольфо, Пейліз Пітер
МПК: A61K 39/17, A61K 39/15, A61K 35/76
Мітки: білки, застосування, неприродні, поверхневі, вірусі, химерні, їхнє, представляють
Формула / Реферат:
1. Химерний вірус пташиного грипу, який міститьa) упакований сегмент NA вірусу грипу, що кодує злитий з нейрамінідазою (NA) білок, у якому відкрита рамка зчитування NA модифікована так, що кодуючі ектодомен NA нуклеотиди замінені нуклеотидами, що кодують ектодомен антигену нейрамінідази інфекційного агента, відмінного від вірусу грипу, який прикріплюється N-кінцем, так що злитий з нейрамінідазою білок експресується і вбудовується в...
Спосіб збільшення часу розвитку захворювання або виживаності у ракових пацієнтів

Номер патенту: 95902
Опубліковано: 26.09.2011
Автори: Дерінк Міка К., Келсі Стівен М.
МПК: A61P 35/00, A61K 39/395
Мітки: спосіб, збільшення, виживаності, захворювання, розвитку, пацієнтів, ракових, часу
Формула / Реферат:
1. Спосіб збільшення часу розвитку захворювання (ТТР) або виживаності у пацієнта, хворого на рак, що включає введення пацієнтові інгібітора димеризації HER, що являє собою антитіло, яке зв'язує Домен II зовнішньоклітинного домену HER2 та інгібує димеризацію HER2 або гетеродимеризацію HER, в кількості, яка збільшує ТТР або виживаність пацієнта, де рак пацієнта вибирають з групи, що складається з раку яєчників, очеревинного раку, раку...
Кон’югати, їхнє одержання і застосування в терапії

Номер патенту: 108473
Опубліковано: 12.05.2015
Автори: Бушар Ерве, Брен Марі-Прісцилл, Чжан Цзидун, Коммерсон Ален
МПК: A61P 35/00, C07D 273/00, A61K 31/395
Мітки: їхнє, кон'югати, застосування, одержання, терапії
Формула / Реферат:
1. Похідне криптофіцину формули (II):, (II) в якій: R1 означає атом галогену і R2 означає групу -ОН або (С1-С4)алканоїлоксигрупу; або R1 і R2 разом утворюють епоксидну ланку; R3 означає (С1-С6)алкільну групу; R4 і R5 обидва означають Н або разом утворюють подвійний...
1,5-бензотіазепіни і їхнє застосування як протиліпемічних засобів

Номер патенту: 79085
Опубліковано: 25.05.2007
Автори: Бломберг Давід, Дальстром Мікаель, Старке Інгемар
МПК: A61P 3/06, A61P 9/06, A61P 29/00, C07F 9/6536, A61K 45/06, A61K 31/785, A61K 38/00, A61K 31/505, A61P 9/10, A61K 31/192, A61P 31/00, A61P 9/08, A61K 31/255, A61P 9/04, A61P 17/02, C07D 281/00, C07D 417/12, C07K 5/065, A61K 31/675, A61P 9/00, A61P 43/00, A61K 31/40, A61P 7/02, A61K 45/00, A61K 31/397, A61K 31/554, A61K 31/366, A61P 17/00
Мітки: 1,5-бензотіазепіни, застосування, протиліпемічних, їхнє, засобів
Формула / Реферат:
1. Сполука формули (І):(І),деRv і Rw незалежно вибрані з водню або С1-6алкілу;R1 і R2 незалежно вибрані з С1-6алкілу;Rx і Ry незалежно вибрані з водню або С1-6алкілу, або один з Rx і Ry являє собою водень або С1-6алкіл, а інший являє собою гідроксил або С1-6алкокси;Rz вибраний з галогену, нітро, ціано, гідроксилу, аміно, карбокси,...
Попередній патент: Кульовий млин з перемішувачем, який містить систему запобігання зношенню
Наступний патент: Парасолька
Випадковий патент: В'яжуче