Спосіб підвищення активності штамів маточних культур гливи звичайної
Номер патенту: 7869
Опубліковано: 15.07.2005
Автори: Фролов Олександр Кирилович, Кириченко Валентина Василівна, Копійка Віра Вікторівна, Кузьменко Олена Анатоліївна



Формула / Реферат
Спосіб підвищення активності штамів маточних культур гливи звичайної, що включає проведення 20-25 пасажів на сусло-агаровому середовищі, виготовлення на основі штамів маточних культур гливи звичайної посівного зернового міцелію, інокуляцію посівним зерновим міцелієм з штаму гливи звичайної карпофорного субстрату, вирощування плодових тіл з подальшим отриманням з карпофору вторинної чистої культури, який відрізняється тим, що як карпофорний субстрат використовують відпилки листяних порід дерев, а для отримання вторинної маточної культури використовують гіменіальний шар карпофору.
Текст
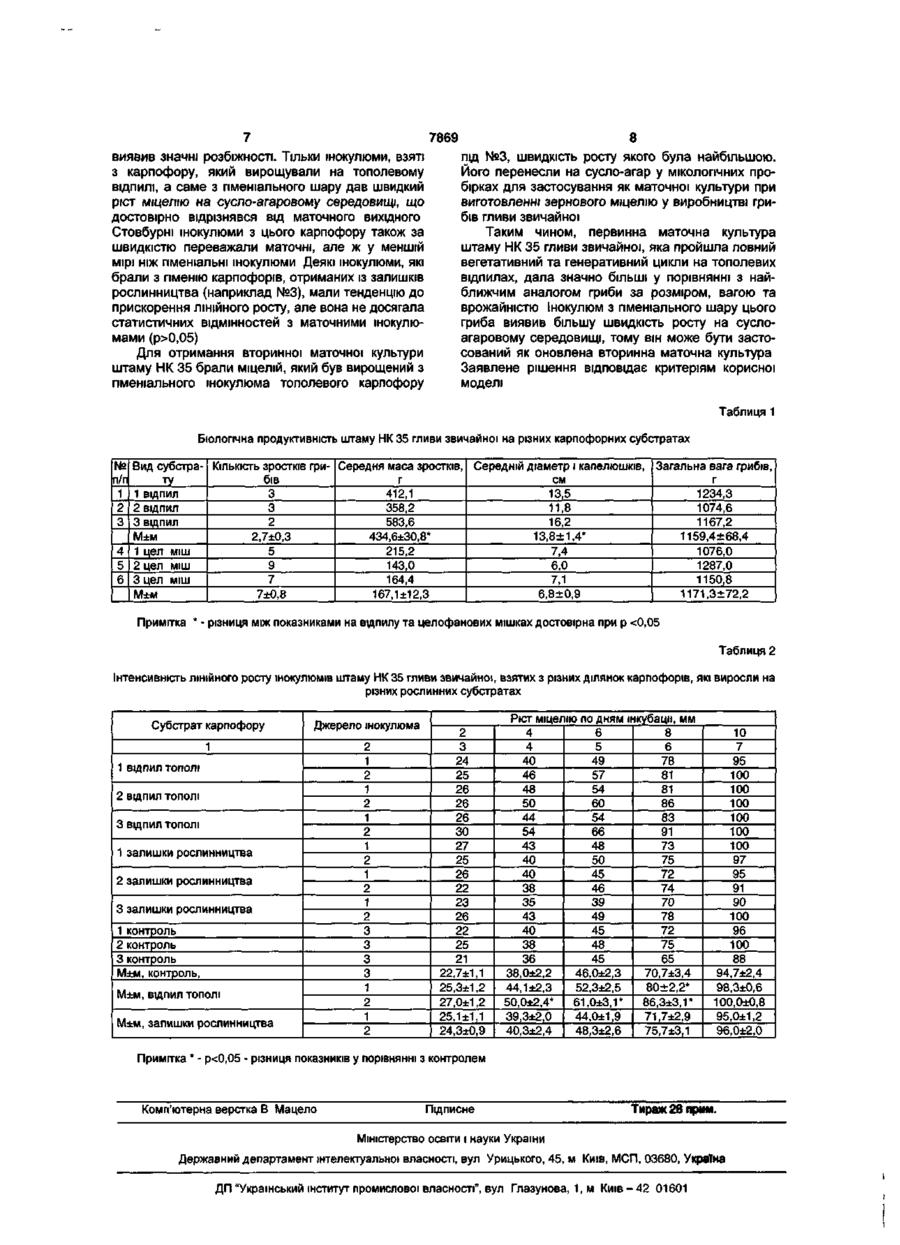
Спосіб підвищення активності штамів маточних культур гливи звичайної, що включає проведення 20-25 пасажів на сусло-агаровому середо CD 00 О) 7869 го зернового міцелію, інокуляція карпофорного субстрату посівним зерновим міцелієм з штаму гливи звичайної, вирощування плодових тіл з подальшим отриманням з карпофору вторинної чистої культури. Недоліком цього способу є недостатня стимуляція активності оновленої маточної культури гливи звичайної. Причинами, що перешкоджають досягненню результату, є: - низький вміст у даному карпофорному субстраті, який складається з залишків рослинництва (соломи злакових, лушпиння соняшника, качанів кукурудзи тощо), лігніну та целюлози; - неоднакова структура лігніну та целюлози в трав'яних рослинних залишках, що пролонговує дію метаболічних ферментів міцелію гриба і призводить до необхідності синтезу інших ферментів згідно з генотипом. В основу корисної моделі поставлено задачу розробити спосіб підвищення активності штамів маточних культур гливи звичайної, який шляхом 20-25 насувань на сусло-агаровому середовищі штамів маточних культур гливи звичайної, виготовлення на основі штамів маточних культур гливи звичайної посівного зернового міцелію, інокуляції карпофорного субстрату (відпил листяних порід дерев) посівним зерновим міцелієм, вирощування плодових тіл, отримання вторинної маточної культури з гіменіального шару карпофору дозволяє відновити активність колекційного штаму. Суттєвими ознаками запропонованого рішення є: - проведення 20-25 пасажів штамів маточних культур гливи звичайної на сусло-агаровому середовищі; - виготовлення на основі штамів маточних культур гливи звичайної посівного зернового міцелію; - інокуляція карпофорного субстрату з відпилу листяних порід дерев посівним зерновим міцелієм; - вирощування на відпилі плодових тіл (карпофорів); - отримання вторинної маточної культури з гемінеального шару карпофору. Відмінними від найближчого аналогу ознаками є: - інокуляція міцелієм карпофорного субстрату з відпилу листяних порід дерев; - отримання вторинної маточної культури з гіменіального шару карпофору. Спосіб здійснюють таким чином. Проводять 20-25 пасажів маточного штаму гливи звичайної на сусло-агаровому середовищі для отримання посівного міцелію для потреб грибівництва. Виготовляють на основі штамів маточних культур гливи звичайної посівний зерновий міцелій. Проводять інокуляцію карпофорного субстрату з відпилу листяних порід дерев посівним зерновим міцелієм, для чого готують неошкірений відпил з нещодавно зрубаної деревини листяних порід діаметром не менш 30см і довжиною 4045см. Якщо деревина, з якої готують відпил, суха (менш 60% вологості), то останню замочують на 23 доби у водопровідній відстояній воді. Інокулюм вносять у просвердлені отвори діаметром 8-10мм відпилу листяних порід дерев, отвори свердловин замощують вологою тирсою. Кількість свердловин становить 2-3 на 10см2. Вирощують на відпилі плодові тіла. Перші розвинені зростки плодових тіл зрізують, отримують вторинну маточну культуру з гемінеального шару капелюшка карпофору. Одним із головних прийомів підвищення активності штамів гливи звичайної є проведення її життєвого циклу (вегетативної та генеративної стадії) через філогенетично адаптований субстрат - відпил дерев листяних порід. На мертвій деревині у процесі філогенезу формується генотип метаболічного складу ферментів гливи звичайної. За багаторічний період виведення гливи звичайної отримано багато десятків її штамів, що відрізняються відповідними ознаками, які сформували селекціонери. Кожен із штамів підтримують у біотехнологічному процесі у вигляді колекційних маточних культур (колекційні штами) на штучному суслоагаровому середовищі. Ці колекційні штами використовують для отримання зернового міцелію (на зернівках пшениці, ячменю), які також є штучним середовищем. Зерновий міцелій далі використовують для інокуляції карпофорних субстратів (залишків рослинництва) при інтенсивній біотехнології гливи звичайної. Тривале ласування штамів на штучних середовищах: сусло-агар, зерно злакових культур - призводить до зниження фізіологічної активності міцелію, одною з контрастних проявів якої є зниження інтенсивності лінійного росту при засвоєнні штучного субстрату. Ймовірною причиною такого зниження є репресія генів, які відповідають за синтез ферментів, що розщеплюють лігнін та целюлозу в структурній агрегації деревини листяних порід дерев - основних біохімічних речовин живлення гливи звичайної як виду вищих базидіоміцетів. У сусло-агаровому та овочевих середовищах, на яких культивують маточні культури штамів гливи звичайної, містяться лише розчинні вуглеводи і немає потреби в експресії генів, що контролюють ферменти, які розчиняють лігноцелюлозу, що з часом призводить до їх репресування за типом зворотного зв'язку. Отримання вторинної маточної культури штаму гливи зі шматочка ніжки плодових тіл, які вирощували на карпофорному субстраті із залишків рослинництва, також не може в повній мірі вирішити проблему підвищення активності маточних штамів гливи звичайної, які пролонговано ласувались на штучних середовищах. Співвідношення лігніну та целюлози в рослинних залишках (солома злакових, лушпиння соняшника, качани кукурудзи) дуже відрізняється. Крім того, структурна організація лігніну та целюлози в цих субстратах відрізняється від такого комплексу в деревині листяних порід. Тому не весь набір метаболічних ферментів за видовим та штамовим генотипами використовують у життєвому циклі гриба при його культивуванні на штучних карпофорних субстратах з рослинних залишків. Враховуючи універсальну властивість генів - їх плейотропну дію на декілька ознак, зменшення активності та репресія трофічних генів негативно відбивається і на інших фізіологічних реакціях гриба: швидкості поділу клітин, який проявляється у відставанні швидкості 7869 лінійного росту Міцелію та засвоєнні ним субстрату. У заявленому способі підвищення активності гриба штам гливи звичайної проходить увесь біологічний цикл від вихідної маточної культури до плодового тіла на філогенетично адаптованому субстраті - мертвої деревині листяних порід, який містить оптимальний склад та структуру лігноцелюлозного комплексу. У процесі живлення і росту в клітинах максимально діє видообумовлена генетична програма штаму. її активний стан зберігається у вторинній маточній культурі даного штаму гливи, який отримали з плодового тіла, вирощеного на відпилі дерев листвяних порід. Другою головною умовою підвищення фізіологічної активності вихідних штамів гливи звичайної, згідно із запропонованим способом, є виділення інокулюму з гіменіального шару капелюшка карпофору, який виріс на відпилі дерев листяних порід. Біопсія інокулюму з ніжки того ж карпофору не дає адекватного результату по підвищенню швидкості росту міцелію та засвоєнню субстрату. Біологічне значення плодового тіла в життєвому циклі капелюшних грибів є в розмноженні гаплоїдними спорами, при створенні яких має місце клітинний поділ за типом мейозу. Завдяки мейозу відбуваються усі форми статевого розмноження, воно є вершиною клітинної еволюції. Тому, ймовірно, повнота розгортання генетичної програми в гіфах гіменіального шару йде повніше, ніж у гіфах ніжки та інших частинах грибниці. Підвищення швидкості росту оновленого штаму гливи звичайної згідно із запропонованим способом прискорить засвоєння субстрату, що особливо важливо на першому тижні при інтенсивній біотехнології цього гриба. Це підвищує його конкурентну здатність порівняно з іншими контамінантами в трав'яних субстратах. Останні, особливо термофільні мікроорганізми, залишаються в трав'яних субстратах після їх пастеризації при інтенсивному способі вирощування грибів. Тому при зниженні активності росту міцелію контамінантна мікрофлора його буде ще більше пригнічувати, що призводить до зниження врожайності культури гриба. Прискорений ріст міцелію також скоротить вегетативну та генеративну стадії гриба, що підвищить кількість біологічних циклів, які можна провести при грибництві гливи звичайної і отримати більший економічний ефект. Приклад конкретного виконання. Проводили 20-25 пасажів маточного штаму гливи звичайної на сусло-агаровому середовищі для отримання посівного міцелію для потреб грибівництва. Виготовляли на основі штамів маточних культур гливи звичайної посівний зерновий міцелій. Заготовляли 3 тополеві неошкурені відпили діаметром 40-45см. Відпили замочували на 3 доби у проточній воді, потім просвердлили отвори діаметром 1см, довжиною до 10см. Кількість свердловин складала 2-3 на см 2 відпилу. В отвори інокулювали посівний зерновий міцелій гливи звичайної (штам НК-35), фізіологічну активність якого ми хотіли підвищити. Для порівняння інокулювали цим же міцелієм 3 целофанові мішки вагою 10кг з пастеризованою сумішшю: 70% соломи пшениці та 30% лушпиння соняшника, згідно зі способом най ближчого аналога. Інокульовані відпили ставили в піддон з вологою тирсою листяних дерев. Відпили та мішки розміщували у окремих мікотронах та інкубували при температурі 26±0,5°С і вологості 85-95%. Вологість підтримували розпилювачем та зволоженням тирси. При появі примордіїв, що свідчить про закінчення вегетативної стадії росту міцелію і початок генеративної стадії життєвого циклу гливи звичайної в мікотронах знизили температуру до 12-13° С в перші 4 дні, потім її підтримували на рівні 15-17°С до початку плодоношення. Вологість доводили до 85-95%, освітлювали камери лампами денного освітлення інтенсивністю 400±50 люкс протягом 9-10 годин на добу. Зібрали розвинені зростки грибів першої хвилі плодоношення, характеристики яких представлені в таблиці 1. Аналіз плодових тіл, зібраних при першій хвилі плодоношення на двох видах субстратів: відпилах тополі та залишках рослинництва, свідчить, що зростки на деревині крупніші і окремі карпофори розвинені більше, ніж ті, які виросли на трав'янистих залишках. Як правило, в тополевих зростках грибів домінували 1-3 карпофори, у яких діаметр шляпки був більше 15см. У зростках на трав'янистих залишках домінувало 4-5 карпофорів, діаметр капелюшок яких у середньому складав 7см і рідко досягав 10см. Привертає увагу те, що загальна вага зібраних грибів з кожного зразка культури мало залежала від виду субстрату. Однак для трав'яного субстрату врожайність першої хвилі плодоношення складала 65-70% від загальної врожайності, яку збирали додатково на 2 та 3 хвилях плодоношення. На відпилах листяних порід дерев перша хвиля плодоношення складала 20-25% від загальної врожайності, а основна біологічна продуктивність припадала на 2-3 хвилю плодоношення. Вказані розбіжності обумовлені щільністю деревини та повільним її засвоєнням міцелієм, у порівнянні з пухким трав'яним субстратом. З конкретного виду субстрату (відпили тополі і відходи рослинництва) взяли по одному найбільш розвиненому карпофору, з яких отримували вторинну маточну культуру. Для цього в стерильному боксі карпофори обробляли 96°-ним етиловим спиртом, потім 3% перекисом водню, з різних ділянок ніжки та капелюшка брали по 3 зразки: з ніжки зрізали верхній пошкоджений стерилізацією шар гіфів, з цією метою в капелюшках знімали пластинки, а інокулюм брали з оновленої поверхні зрізу. Інокулюм розміщували в центрі суслоагарового середовища в чашках Петрі, які мають загальний діаметр 10см. Інокульовані чашки Петрі інкубували в термостаті при t=26±0,5°C. Інтенсивність росту міцелію оцінювали через 2 доби. Результати замірювань лінійного росту міцелію представлені в таблиці 2. За 1 приймали інокулюм, який брали з ніжки карпофору; за 2 - з гіменіального шару; за 3 - маточну культуру штаму, який пройшов 25 пасажів на сусло-агаровому середовищі. Аналіз лінійного росту інокулюмів, які отримували з різних ділянок плодових тіл, які, у свою чергу, отримували згідно з найближчим аналогом (із залишків рослинництва) та заявленим способом, 7869 виявив значні розбіжності. Тільки інокулюми, взяті з карпофору, який вирощували на тополевому ВІДПИЛІ, а саме з пменіального шару дав швидкий ріст міцелію на суспо-агаровому середовищі, що достовірно відрізнявся від маточного вихідного Стовбурні інокулюми з цього карпофору також за швидкістю переважали маточні, але ж у меншій мірі ніж пменіальні інокулюми Деякі інокулюми, які брали з гіменію карпофорів, отриманих із залишків рослинництва (наприклад №3), мали тенденцію до прискорення ЛІНІЙНОГО росту, але вона не досягала статистичних відмінностей з маточними інокулюмами (р>0,05) Для отримання вторинної маточної культури штаму НК 35 брали міцелій, який був вирощений з пменіального інокулюма тополевого карпофору 8 під №3, швидкість росту якого була найбільшою. Його перенесли на сусло-агар у МІКОЛОГІЧНИХ про бірках для застосування як маточної культури при виготовленні зернового міцелію у виробництві грибів гливи звичайної Таким чином, первинна маточна культура штаму НК 35 гливи звичайної, яка пройшла повний вегетативний та генеративний цикли на тополевих відпилах, дала значно більші у порівнянні з найближчим аналогом гриби за розміром, вагою та врожайністю Інокулюм з пменіального шару цього гриба виявив більшу швидкість росту на суслоагаровому середовищі, тому він може бути застосований як оновлена вторинна маточна культура Заявлене рішення відповідає критеріям корисної моделі Таблиця 1 Біологічна продуктивність штаму НК 35 гливи звичайної на різних карпофорних субстратах № п/п 1 2 3 Вид субстра- КІЛЬКІСТЬ зростків гри- Середня маса зростків, Середній діаметр і капелюшків, Загальна вага грибів, см г бів г ту 1 ВІДПИЛ 3 412,1 13,5 1234,3 3 358,2 11,8 2 відпил 1074,6 2 16,2 1167,2 3 ВІДПИЛ 583,6 1159,4±68,4 М±м 434,6±30,8* 2,7±0,3 13,8±1,4* 4 1 цел міш 5 215,2 7,4 1076,0 9 6,0 5 2 цел міш 143,0 1287,0 7 164,4 1150,8 6 3 цел міш 7,1 1171,3±72,2 М±м 167,1 ±12,3 6,8±0,9 7±0,8 Примітка * - різниця між показниками на відпилу та целофанових мішках достовірна при р
ДивитисяДодаткова інформація
Назва патенту англійськоюMethod of improving activity of strains of stock cultures of oyster fungus
Автори англійськоюFrolov Oleksandr Kyrylovych, Kyrychenko Valentyna Vasylivna, Kopiika Vira Viktorivna
Назва патенту російськоюСпособ повышения активности штаммов маточных культур вешенки обыкновенной
Автори російськоюФролов Александр Кириллович, Кириченко Валентина Васильевна, Копейка Вера Викторовна
МПК / Мітки
МПК: A01G 1/04
Мітки: звичайної, гливи, активності, підвищення, маточних, штамів, спосіб, культур
Код посилання
<a href="https://ua.patents.su/4-7869-sposib-pidvishhennya-aktivnosti-shtamiv-matochnikh-kultur-glivi-zvichajjno.html" target="_blank" rel="follow" title="База патентів України">Спосіб підвищення активності штамів маточних культур гливи звичайної</a>
Спосіб активації посівного міцелію їстівного гриба гливи звичайної (pleuroutus ostereatus)

Номер патенту: 53900
Опубліковано: 17.02.2003
Автори: Бухало Ася Сергіївна, Потьомкіна Жанна В'ячеславівна, Поєдинок Наталія Леонідівна
МПК: A01G 1/04
Мітки: спосіб, гливи, їстивного, посівного, активації, ostereatus, звичайної, міцелію, pleuroutus, гриба
Формула / Реферат:
Спосіб активації посівного міцелію їстівного гриба гливи звичайної (Pleuroutus ostereatus) впливом лазерного випромінювання на посівний міцелій, який відрізняється тим, що ведуть обробку на 14 добу інкубації на зерні пшениці дикаріотичного міцелію в червоній області спектра при дозі 230 мДж/см2.
Поживне середовище для селекції та гібридизації штамів гливи звичайної

Номер патенту: 28397
Опубліковано: 16.10.2000
Автори: Каліберда Генадій Володимирович, Негруцький Сергій Федорович, Сичьов Петро Антонович, Ткаченко Наталя Петрівна, Тімофеєв Олексій Анатолієвич
МПК: A01G 1/04, A01H 15/00, A01H 1/04
Мітки: звичайної, поживне, штамів, селекції, гібридизації, середовище, гливи
Формула / Реферат:
Питательная среда для селекции и гибридизации штаммов вешенки обыкновенной, включающая воду, агар-агар, отличающаяся тем, что дополнительно содержит плоды лоха узколистного при следующем соотношении компонентов, г/л: плоды лоха узколистного зрелые в воздушно-сухом состоянии 99,85-100,15 агар-агар 19,85-20,15 вода остальное.
Спосіб підвищення активності біологічної фіксації азоту і урожайності сільськогосподарських культур

Номер патенту: 47305
Опубліковано: 17.06.2002
Автори: Волкогон Віталій Васильович, Лохова Валентина Іванівна, Штанько Надія Петрівна
МПК: C05D 7/00
Мітки: урожайності, біологічно, активності, спосіб, підвищення, фіксації, азоту, сільськогосподарських, культур
Формула / Реферат:
Спосіб підвищення активності біологічної фіксації азоту і урожайності сільськогосподарських культур шляхом локального внесення в грунт фізіологічно активних речовин при висіві та посадці сільськогосподарських культур, який відрізняється тим, що як джерело фізіологічно активних сполук використовується попередньо гранульований біогумус.
Спосіб закладення інтенсивних маточних насаджень плодових та ягідних культур і садів лугового типу

Номер патенту: 24251
Опубліковано: 07.07.1998
Автори: Книга Микола Мусійович, Гордановська Галина Петрівна
МПК: A01G 1/00
Мітки: типу, садів, насаджень, лугового, ягідних, закладення, плодових, спосіб, інтенсивних, культур, маточних
Формула / Реферат:
1. Спосіб закладення інтенсивних маточних насаджень плодових та ягідних культур і садів лугового типу, що включає висаджування відсадків чи саджанців під гострим кутом до поверхні грунту, який відрізняється тим, що при садінні використовують спеціально сформований садівний матеріал, пагони подовження у якого відходять під кутом до коренегенної зони, і його висаджуванням забезпечують горизонтальну орієнтацію пагоноутворюючої частини відсадків...
Субстракт для вирощування істівного гриба гливи звичайної

Номер патенту: 8596
Опубліковано: 30.09.1996
Автори: Негруцький Сергій Федорович, Ерьоменко Микола Павлович, Зубченко Лілія Івановна, Сичьов Петро Антонович, Кіпень Ана Миколайовна, Кисаров Генадій Миколайович
МПК: A01G 1/04
Мітки: вирощування, гливи, гриба, істівного, субстракт, звичайної
Формула / Реферат:
Использование отхода хлопчатобумажного производства - орешка и пуха второго пропуска в качестве субстрата для выращивания съедобного гриба вешенки обыкновенной.
Попередній патент: Пісочний кондитерський виріб
Наступний патент: Кондитерський виріб
Випадковий патент: Радіозахисний біопрепарат із молюсків "біполан"