Мутанти fad-2 і високоолеїнові рослини










































Формула / Реферат
1. Виділена молекула нуклеїнової кислоти, що кодує білок дельта-12-олеатдесатуразу (FAD2), причому вказаний білок FAD2 має амінокислотну заміну в положенні, відповідному положенню 108 в білку FAD2 дикого типу, представленому SEQ ID NO: 4 або SEQ ID NO: 8.
2. Виділена молекула нуклеїнової кислоти, що кодує білок FAD2, причому вказаний білок FAD2 має амінокислотну заміну в положенні, відповідному положенню 118 в білку FAD2 дикого типу, представленому SEQ ID NO: 4 або SEQ ID NO: 8.
3. Виділена молекула нуклеїнової кислоти, що кодує білок FAD2, причому вказаний білок FAD2 має амінокислотну заміну в положенні, відповідному положенню 108, і в положенні, відповідному положенню 118 в білку FAD2 дикого типу, представленому SEQ ID NO: 4 або SEQ ID NO: 8.
4. Виділена молекула нуклеїнової кислоти за будь-яким з пп. 1-3, де вказаний білок FAD2 є білком FAD2 Brassica, більш конкретно - білком FAD2 Brassica napus.
5. Виділена молекула нуклеїнової кислоти за будь-яким з пп. 1-4, де вказана амінокислота в положенні 108 замінена на аспарагінову кислоту.
6. Виділена молекула нуклеїнової кислоти за будь-яким з пп. 2-5, де вказана амінокислота в положенні 118 замінена на фенілаланін.
7. Виділена молекула нуклеїнової кислоти, що включає нуклеїнову кислоту SEQ ID NO: 1, 5, 11 або 12, її комплементарна форма або РНК-форма.
8. Виділена молекула нуклеїнової кислоти, яка включає нуклеотидну послідовність, що має принаймні 80 %, переважно принаймні 85%, більш переважно принаймні 90 % і ще більш переважно принаймні 95 %, 96 %, 97 %, 98 % або 99 % ідентичності з SEQ ID NO: 1 або 11, або з їх комплементарною формою або РНК-формою, що кодує білок FAD2, який має амінокислотну заміну в положенні, відповідному положенню 108 в білку FAD2 дикого типу, представленому SEQ ID NO: 4 або SEQ ID NO: 8.
9. Виділена молекула нуклеїнової кислоти, яка включає нуклеотидну послідовність, що має принаймні 80 %, переважно принаймні 85 %, більш переважно принаймні 90 % і ще більш переважно принаймні 95 %, 96 %, 97 %, 98 % або 99 % ідентичності з SEQ ID NO: 5 або 12, або з їх комплементарною формою або РНК-формою, що кодує білок FAD2, який має амінокислотну заміну в положенні, відповідному положенню 118 в білку FAD2 дикого типу, представленому SEQ ID NO: 4 або SEQ ID NO: 8.
10. Виділена молекула нуклеїнової кислоти за будь-яким з пп. 1-9, де вказаний білок FAD2 дикого типу включає амінокислотну послідовність SEQ ID NO: 4 або 8.
11. Фрагмент з принаймні 10 нуклеотидів виділеної молекули нуклеїнової кислоти, причому вказаний фрагмент включає мутований кодон, який відповідає амінокислотній заміні в положеннівідповідному положенню 108 послідовності SEQ ID NO: 4 або SEQ ID NO: 8, і/або мутований кодон, що відповідає амінокислотній заміні в положенні, відповідному положенню 118, послідовності SEQ ID NO: 4 або SEQ ID NO: 8.
12. Білок FAD2, який має амінокислотну заміну в положенні 108 або положенні, відповідному положенню 108, відносно білка FAD2 дикого типу, такого як білок FAD2 дикого типу, представлений амінокислотною послідовністю SEQ ID NO: 4 або 8.
13. Білок FAD2, який має амінокислотну заміну в положенні 118 або положенні, відповідному положенню 118, відносно білка FAD2 дикого типу, такого як білок FAD2 дикого типу, представлений амінокислотною послідовністю SEQ ID NO: 4 або 8.
14. Білок FAD2, який має амінокислотну заміну в положенні 108 або положенні, відповідному положенню 108, і амінокислотну заміну в положенні 118 або положенні, відповідному положенню 118, відносно білка FAD2 дикого типу, такого як представлений амінокислотною послідовністю SEQ ID NO: 4 або 8.
15. Білок FAD2, який включає амінокислотну послідовність SEQ ID NO: 2 або 6.
16. Вектор, який включає молекулу нуклеїнової кислоти за будь-яким з пп. 1-10.
17. Клітина-хазяїн, яка включає вектор за п. 16.
18. Рослина, стабільно трансформована вектором за п. 16.
19. Рослина за п. 18, де вказана рослина для трансформації вибрана з групи, яка складається з олійних культур.
20. Рослина за п. 19, де вказані олійні культури являють собою соняшники, соєві боби, бавовни, кукурудзи і/або насіння рапсу.
21. Рослина або частина рослини або насіння, які містять нуклеотидну послідовність, що кодує білок FAD-2, який має амінокислотну заміну в положенні, відповідному положенню 108 в білку FAD-2 дикого типу, представленому SEQ ID NO: 4 або SEQ ID NO: 8.
22. Рослина або частина рослини або насіння за п. 21, які містять білок FAD-2, що має амінокислотну заміну в положенні, відповідному положенню 108, в білку FAD-2 дикого типу, представленому SEQ ID NO: 4 або SEQ ID NO: 8.
23. Рослина або частина рослини або насіння за п. 21 або 22, де вказана заміщувальна амінокислота являє собою аспарагінову кислоту.
24. Рослина або частина рослини або насіння, які містять нуклеотидну послідовність, що кодує білок FAD-2, який має амінокислотну заміну в положенні, відповідному положенню 118 в білку FAD-2 дикого типу, представленому SEQ ID NO: 4 або SEQ ID NO: 8.
25. Рослина або частина рослини або насіння за п. 24, які містять білок FAD-2, що має амінокислотну заміну в положенні, відповідному положенню 118, в білку FAD-2 дикого типу, представленому SEQ ID NO: 4 або SEQ ID NO: 8.
26. Рослина або частина рослини або насіння за п. 24 або 25, де вказана заміщувальна амінокислота являє собою фенілаланін.
27. Рослина або частина рослини або насіння, які містять нуклеотидну послідовність, що кодує білок FAD-2, який має амінокислотну заміну в положенні, відповідному положенню 108, і амінокислотну заміну в положенні, відповідному положенню 118 в білку FAD-2 дикого типу, представленому SEQ ID NO: 4 або SEQ ID NO: 8.
28. Рослина або частина рослини або насіння за п. 27, які містять білок FAD-2, що має амінокислотну заміну в положенні, відповідному положенню 108, і амінокислотну заміну в положенні, відповідному положенню 118, в білку FAD-2 дикого типу, представленому SEQ ID NO: 4 або SEQ ID NO: 8.
29. Рослина або частина рослини або насіння, які містять першу нуклеотидну послідовність, що кодує білок FAD-2, який має амінокислотну заміну в положенні, відповідному положенню 108, і другу нуклеотидну послідовність, що кодує білок FAD-2, який має амінокислотну заміну в положенні, відповідному положенню 118, в білку FAD-2 дикого типу, представленому SEQ ID NO: 4 або SEQ ID NO: 8.
30. Рослина або частина рослини або насіння за п. 29, які містять білок FAD-2, що має амінокислотну заміну в положенні, відповідному положенню 108, і інший білок FAD, що має амінокислотну заміну в положенні, відповідному положенню 118, в білку FAD-2 дикого типу, представленому SEQ ID NO: 4 або SEQ ID NO: 8.
31. Рослина або частина рослини або насіння за будь-яким з пп. 28-30, де вказана заміщувальна кислота в положенні 108 або положенні, відповідному положенню 108, являє собою аспарагінову кислоту, і вказана заміщувальна амінокислота в положенні 118 або положенні, відповідному положенню 118, являє собою фенілаланін.
32. Насіння, одержане від рослини за п. 29 або 31.
33. Потомство, одержане від рослини або частини рослини або насіння за будь-яким з пп. 29, 31 або 32.
34. Рослинна олія, одержана з насіння за п. 32, яка включає більше ніж (приблизно) 72 %, 75 %, 80 % або 85 % олеїнової кислоти відносно загальної ваги жирних кислот, присутніх в цій олії, і додатково включає менше ніж (приблизно) 4 %, 3,5 %, 3 %, 2 %, 1 % або 0,5 % ліноленової кислоти.
35. Спосіб збільшення вмісту олеїнової кислоти в рослині, який включає трансформування рослини вектором за п. 16.
36. Застосування фрагмента з принаймні 10 нуклеотидів за п. 11 як праймерів, зондів і/або маркерів селекції.
37. Спосіб селекції рослин видів Brassica за допомогою маркера з використанням молекули нуклеїнової кислоти за п. 11.
38. Набір для аналізу, який включає перший контейнер, що містить молекулу нуклеїнової кислоти за будь-яким з пп. 1-11.
39. Спосіб одержання ліній високоолійних рослин, який включає:
(a) індукцію мутагенезу в принаймні деяких клітинах з рослини, більш конкретно рослини Brassica, і переважно рослини Brassica napus, яка має вміст олеїнової кислоти в клітинах менший ніж 70 %;
(b) регенерацію рослин з принаймні однієї зі вказаних мутагенізованих клітин;
(с) вибір регенерованих рослин, які мають послідовність нуклеїнової кислоти за будь-яким з пп. 1-10 і/або які експресують білок FAD2 за будь-яким з пп. 12-15; і
(d) одержання подальших поколінь рослин зі вказаних регенерованих рослин.
Текст
Реферат: Винахід належить до трансгенних рослин, що містять виділену молекулу нуклеїнової кислоти, що кодує білок дельта-12-олеатдесатуразу (FAD2), причому вказаний білок FAD2 має одну або дві амінокислотні заміни в положеннях, відповідних положенням 108 та/або 118 відносно білка FAD2 дикого типу, що забезпечує вміст олеїнової кислоти понад 85% та вміст ліноленової кислоти менше 0,5% від загального вмісту жирних кислот в насінні зазначених рослин. Винахід також належить до олії, отриманої з насіння зазначених рослин та до способу одержання зазначених рослин. UA 99099 C2 (12) UA 99099 C2 UA 99099 C2 5 10 15 20 25 30 35 40 45 50 55 Даний винахід належить до рослин, насіння і продуктів, одержаних з них, особливо до рослин Brassica, насіннєвих продуктів, одержаних з них, що мають мутантні послідовності, які створюють високий профіль олеїнових кислот в олії насіння. Більш конкретно, винахід належить до мутантних послідовностей дельта-12-десатурази жирних кислот, також згадуваних тут як послідовності FAD2, в рослинах, які наділяють олію насіння високим профілем олеїнових кислот. Дельта-12-десатураза жирних кислот (також відома як олеїнова десатураза або олеатдесатураза) залучена до ензиматичного перетворення олеїнової кислоти в лінолеву кислоту. Різновиди з високим рівнем олеїнової кислоти (можливо в комбінації з низькими рівнями ліноленової кислоти) потрібні для множини різних застосувань (харчового використання, медичного використання, застосування в біодизелях і багатьох інших). Мутантне насіння, що дає олію, яка демонструє високий вміст олеїнової кислоти (олеїнова кислота вище ніж 70 вагових % відносно загальної ваги жирних кислот, присутніх в маслі), раніше описане в літературі, мало дуже мале агрономічне значення і/або погані характеристики коріння, і/або дуже низьку врожайність. Все ще існує потреба в матеріалі, що має стабільний, високий вміст олеїнової кислоти (можливо в поєднанні зі стабільно низьким вмістом ліноленової кислоти) в різних місцеположеннях протягом років, також з хорошою агрономічною продуктивністю і нормальною морфологією олійного рапсу. Особливо, рослини не повинні мати фасціації і повинні мати нормальний розвиток кореня. Даний винахід належить до молекули нуклеїнової кислоти, що включає (або складається з) послідовність нуклеїнової кислоти, яка кодує білок дельта-12-олеатдесатуразу (FAD2), причому вказаний білок FAD2 має заміну амінокислоти в положенні 108 відносно білка FAD2 дикого типу. Іншим об'єктом є молекула нуклеїнової кислоти, яка включає (або складається з) послідовність нуклеїнової кислоти, що кодує білок FAD2, причому вказаний білок FAD2 має заміну амінокислоти в положенні 118 відносно білка FAD2 дикого типу. Ще одним об'єктом є молекула нуклеїнової кислоти, яка включає (або складається з) послідовність нуклеїнової кислоти, що кодує білок FAD2, причому вказаний білок FAD2 має заміни амінокислот в положенні 108 і в положенні 118 відносно білка FAD2 дикого типу. Переважно, вказаний білок FAD2 являє собою білок FAD2 Brassica, більш конкретно - білок FAD2 Brassica napus. Переважно, вказана заміщена амінокислота в положенні 108 являє собою аспарагінову кислоту (що заміщує гліцин в білці FAD2 дикого типу). Переважно, вказана заміщена амінокислота в положенні 118 являє собою фенілаланінову кислоту (що заміщує лейцин в білці FAD2 дикого типу). Переважна молекула нуклеїнової кислоти за винаходом включає (або складається з) нуклеїнову кислоту з послідовністю SEQ ID NO: 1, 5, 11 або 12, комплементарною ним формою або їх РНК формою. Молекула нуклеїнової кислоти за винаходом може включати або складатися з нуклеотидної послідовності, що має принаймні 80%, переважно принаймні 85%, більш переважно принаймні 90% і ще більш переважно принаймні 95%, 96%, 97%, 98% або 99% ідентичності з SEQ ID NO: 1 або 11, або з їх комплементарною формою або їх РНК формою, що кодує білок FAD2, який має амінокислотну заміну в положенні 108 відносно білка FAD2 дикого типу. Молекула нуклеїнової кислоти за винаходом може включати або складатися з нуклеотидної послідовності, що має принаймні 80%, переважно принаймні 85%, більш переважно принаймні 90% і ще більш переважно принаймні 95%, 96%, 97%, 98% або 99% ідентичності з SEQ ID NO: 5 або 12, або з їх комплементарною формою або їх РНК формою, що кодує білок FAD2, який має амінокислотну заміну в положенні 118 відносно білка FAD2 дикого типу. Більш конкретно, вказаний білок FAD2 дикого типу включає (або складається з) амінокислотну послідовність з SEQ ID NO: 4 або 8. Також об'єкт даного винаходу являє собою фрагмент принаймні з 10, 15, 20, 25, 30, 40, 50, 100 або більше нуклеотидів молекули нуклеїнової кислотивідповідно до винаходу, вказаний фрагмент включає мутантний кодон, відповідний вказаній амінокислотній заміні в положенні 108 і/або мутантний кодон, відповідний вказаній амінокислотній заміні в положенні 118. Вказані фрагменти можуть бути використані як праймери, зонди і/або селективні маркери. Будь-яка з молекул нуклеїнових кислот за винаходом може бути використана в способі селекції рослин за допомогою маркера, переважно видів Brassica, більш переважно різновидів Brassica napus, що також є об'єктом даного винаходу. 1 UA 99099 C2 5 10 15 20 25 30 35 40 45 50 55 Інший об'єкт даного винаходу являє собою набір для аналізу, який може містити перший контейнер, що містить будь-яку з молекул нуклеїнових кислот за винаходом. Інший об'єкт даного винаходу являє собою білок FAD2, що має амінокислотну заміну в положенні 108 або відповідному положенню 108, відносно білка FAD2 дикого типу, представленого амінокислотною послідовністю SEQ ID NO: 4 або 8. Інший об'єкт даного винаходу являє собою білок FAD2, що має амінокислотну заміну в положенні 118 або відповідному положенню 118, відносно білка FAD2 дикого типу, представленого амінокислотною послідовністю SEQ ID NO: 4 або 8. Інший об'єкт даного винаходу являє собою білок FAD2, що має амінокислотну заміну в положенні 108 або відповідному положенню 108, і амінокислотну заміну в положенні 118 або відповідному положенню 118, відносно білка FAD2 дикого типу, представленого амінокислотною послідовністю SEQ ID NO: 4 або 8. Переважний білок FAD2 за винаходом включає (або складається з) амінокислотну послідовність SEQ ID NO: 2 або 6. Інший об'єкт даного винаходу являє собою вектор, що включає молекулу нуклеїнової кислоти, яка кодує мутантний білок FAD2, відповідно до винаходу. Інший об'єкт даного винаходу являє собою клітину-хазяїна, яка включає вектор за винаходом і/або послідовність нуклеїнової кислоти, що кодує мутантний білок FAD2 відповідно до винаходу. Інший об'єкт даного винаходу являє собою рослину, стабільно трансформовану вектором за винаходом. Рослина для трансформації може бути вибрана з групи, яка складається з олійних культур, більш конкретно з соняшників, соєвих бобів, бавовни, кукурудзи і/або рапсового насіння. Інший об'єкт даного винаходу являє собою рослину або частину рослини, або насіння, що містять нуклеотидну послідовність, яка кодує білок FAD-2, що має амінокислотну заміну в положенні 108 відносно білка FAD-2 дикого типу або положенні, відповідному йому. Більш конкретно, рослина або частина рослини, або насіння відповідно до винаходу містять (або експресують) білок FAD2, що має амінокислотну заміну, відповідну положенню 108 відносно білка FAD-2 дикого типу або положенню, відповідному йому. Інший об'єкт даного винаходу являє собою рослину або частину рослини, або насіння, що містять нуклеотидну послідовність, яка кодує білок FAD-2, що має амінокислотну заміну в положенні 118 відносно білка FAD-2 дикого типу або положенні, відповідному йому. Більш конкретно, рослина або частина рослини, або насіння відповідно до винаходу містять (або експресують) білок FAD2, що має амінокислотну заміну, відповідну положенню 118 відносно білка FAD-2 дикого типу або положенню, відповідному йому. Інший об'єкт даного винаходу являє собою рослину або частину рослини, або насіння, що містять нуклеотидну послідовність, яка кодує білок FAD-2, що має амінокислотну заміну в положеннях 108 і 118 відносно білка FAD-2 дикого типу або положеннях, відповідних ним. Більш конкретно, рослина або частина рослини, або насіння відповідно до винаходу містять (або експресують) білок FAD2, що має амінокислотну заміну, відповідну положенням 108 і 118 відносно білка FAD-2 дикого типу або положенням, відповідним ним. Інший об'єкт даного винаходу являє собою рослину або частину рослини, або насіння, що містять першу нуклеотидну послідовність, яка кодує білок FAD2, що має амінокислотну заміну в положенні 108 або положенні, відповідному йому, і другу нуклеотидну послідовність, яка кодує білок FAD2, що має амінокислотну заміну в положенні 118 або положенні, відповідному йому, відносно білка FAD2 дикого типу. Більш конкретно, рослина або частина рослини, або насіння відповідно до винаходу містять (або експресують) два білки FAD2, один з яких має амінокислотну заміну, відповідну положенню 108 або положенню, відповідному йому, а інший має амінокислотну заміну, відповідну положенню 118 відносно білка FAD-2 дикого типу або положенню, відповідному йому. Переважно, вказана заміщена амінокислота в положенні 108 або положенні, відповідному йому, являє собою аспарагінову кислоту (що заміщує гліцин в білці FAD2 дикого типу). Переважно, вказана заміщена амінокислота в положенні 118 або в положенні, відповідному йому, являє собою фенілаланін (що заміщує лейцин в білці FAD2 дикого типу). Рослина або частина рослини, або насіння згідно з винаходом можуть бути одержані мутагенною обробкою, більш конкретно EMS обробкою. Потомство, що походить від вказаної рослини або частини рослини, або насіння, також є частиною винаходу. 2 UA 99099 C2 5 10 15 20 25 30 35 40 45 50 55 60 Інший об'єкт даного винаходу являє собою рослинну олія, одержану з насіння за винаходом, причому вказана олія містить більше ніж (приблизно) 72%, 75%, 80% або 85% олеїнової кислоти відносно загальної ваги жирних кислот, присутніх в рапсовій олії. Переважно, вказана олія також включає менше ніж (приблизно) 4%, 3,5% 3%, 2%, 1% або 0,5% ліноленової кислоти. Винахід також належить до харчових або кормових продуктів, які містять і/або приготовані з рослини, частини рослини, насіння і/або рослинної олії відповідно до винаходу. Спосіб поліпшення вмісту олеїнової кислоти в рослині може включати етап трансформації рослини вектором за винаходом. Альтернативно, спосіб одержання ліній рослини з високим вмістом олеїнової кислоти може включати: (а) схрещування першої рослини за винаходом з другою рослиною, (b) одержання насіння від схрещування на етапі (а), (с) вирощування фертильних рослин з такого насіння, (d) одержання насіння потомства від рослин етапу (с), і (е) ідентифікацію серед потомства того насіння, яке має високий вміст олеїнової кислоти. Альтернативно, спосіб одержання ліній рослини з високим вмістом олеїнової кислоти може включати: (a) індукцію мутагенезу в принаймні деяких клітинах з рослини, більш конкретно рослини Brassica, і переважно різновиду Brassica napus, яке має вміст олеїнової кислоти в клітинах менше ніж 70%; (b) регенерацію рослин з принаймні однієї з вказаних мутагенізованих клітин; (с) вибір регенерованих рослин, які мають будь-яку з послідовностей нуклеїнової кислоти за винаходом і/або які експресують білок FAD2 відповідно до винаходу; і (d) одержання подальших поколінь рослин з вказаних регенерованих рослин. Креслення відповідає списку послідовностей по представленому винаходу. Даний винахід належить до рослин, більш конкретно рослин Brassica, і переважно різновиду Brassica napus, які були одержані для одержання олії, що має вміст олеїнової кислоти вище ніж 70% відносно загальної ваги жирних кислот, присутніх в олії. Більш конкретно, рослина за винаходом мала принаймні один мутантний ген FAD2 за винаходом. Переважно, вказаний мутантний ген FAD2 приводить до високого вмісту олеїнової кислоти (тобто вміст олеїнової кислоти вище ніж 70% ваг. відносно загальної ваги жирних кислот, присутніх в олії) в насінні вказаних рослин і в олії, одержаній з вказаного насіння. Даний винахід належить також до будь-якої частини або будь-якого продукту вказаної рослини, що несе не менше одного вказаного мутантного гена FAD2. У контексті представленого винаходу, частина або продукт рослини розуміється таким, що включає лист, сім’ядолю, стебло, черешок, стеблинку, насіння або будь-яку іншу тканину або фрагмент тканини вказаної рослини. Даний винахід належить також до будь-якого потомства вказаної рослини, що несе принаймні один мутантний ген FAD2 за винаходом. У контексті представленого винаходу, термін «потомство» належить до прямих і непрямих спадкоємців, нащадків і похідних від рослини або рослин за винаходом і включає перше, друге, третє і/або подальші покоління, які можуть бути одержані самозапиленням, схрещуванням з рослинами з однаковим або іншими генотипами, і можуть бути модифіковані різними придатними методами генної інженерії. Даний винахід також належить до вказаних мутованих генів FAD2, які забезпечують високий вміст олеїнової кислоти в насінні при присутності в рослинах. Конкретно, винахід належить до нових ізольованих молекул нуклеїнової кислоти, які кодують нові варіанти форм білка FAD2, що має заміщену амінокислоту в положенні 108 (або відповідному положенню 108) і/або заміщену амінокислоту в положенні 118 (або відповідному положенню 118) відносно білка FAD2 дикого типу, такого як білок FAD2 дикого типу, представлений SEQ ID NO: 4 і/або SEQ ID NO: 8. Виділена молекула нуклеїнової кислоти за винаходом містить принаймні одну мутацію, яка приводить до заміщення, переважно до заміщення аспарагінової кислоти гліцином в положенні 108 (або відповідному йому), і/або приводить до заміни, переважно заміни фенілаланіну лейцином в положенні 118 (або відповідному йому) відносно білка FAD2 дикого типу, такого як білок FAD2 дикого типу, представлений SEQ ID NO: 4 і/або SEQ ID NO: 8. Вказана мутація(ї) змінює(ють) функціональність одержуваного продукту гена FAD2, причому рівень олеїнової кислоти змінюється, переважно збільшується, у рослини, що 3 UA 99099 C2 5 10 15 20 25 30 35 40 45 50 55 експресує мутантну послідовність(ті), в порівнянні з відповідним рівнем у рослин, що експресують послідовність(ті) дикого типу. У рамках даного винаходу, якщо не указано інакше, термін «в положенні 108» повинен розумітися як такий, що позначає амінокислоту в положенні 108 білка FAD2 дикого типу, представленого SEQ ID NO: 4 і/або SEQ ID NO: 8, але також такий, що належить до амінокислоти, відповідної вказаному положенню в білку FAD2 дикого типу, який може мати іншу амінокислотну послідовність в зв'язку з делеціями або додатковими амінокислотами в поліпептиді. Схожим чином, термін «в положенні 118» повинен розумітися як такий, що позначає амінокислоту в положенні 118 білка FAD2 дикого типу, представленого SEQ ID NO: 4 і/або SEQ ID NO: 8, але також такий, що належить до амінокислоти, відповідної вказаному положенню в білку FAD2 дикого типу, який може мати іншу амінокислотну послідовність в зв'язку з делеціями або додатковими амінокислотами в поліпептиді. Термін «відповідний положенню» як він використаний тут, означає, що положення не тільки визначається числом попередніх амінокислот. Положення вибраної амінокислоти відповідно до представленого винаходу може змінюватися через делеції або додаткові амінокислоти в поліпептиді. Таким чином, під «відповідним положенням» відповідно до представленого винаходу потрібно розуміти, що описувана амінокислота(и) може відрізнятися від вказаного номера, але все ще мати однакові сусідні амінокислоти в лінійній послідовності. У одному аспекті, молекула нуклеїнової кислоти за винаходом кодує білок FAD2, в якому заміна вказаної амінокислоти, відповідної положенню 108, являє собою глутамат, і переважно аспарагінову кислоту. Більш конкретно, молекула нуклеїнової кислоти за винаходом кодує білок FAD2, що має заміщену гліцином аспарагінову кислоту в положенні 108 відносно білка FAD2 дикого типу, представленого амінокислотною послідовністю SEQ ID NO: 4 або 8. Молекула нуклеїнової кислоти за винаходом може включати (або складатися з) послідовності нуклеїнової кислоти по SEQ ID NO: 3, 5, 7, 9, 10 або 12, причому кодон, що кодує амінокислоту в положенні 108, мав не менше однієї мутації (або був мутований) так, щоб він кодував амінокислоту, відмінну від гліцину, і переважно, щоб він кодував аспарагінову кислоту в положенні 108 відповідно до білка FAD2 за винаходом. Переважна молекула нуклеїнової кислоти за винаходом включає (або складається з) послідовності нуклеїнової кислоти по SEQ ID NO: 1 або 11. У іншому аспекті, молекула нуклеїнової кислоти за винаходом кодує білок FAD2, в якому заміна вказаної амінокислоти, відповідної положенню 118, являє собою фенілаланін. Більш конкретно, молекула нуклеїнової кислоти за винаходом кодує білок FAD2, що має заміщений фенілаланіном лейцин в положенні 118 відносно білка FAD2 дикого типу, представленого амінокислотною послідовністю SEQ ID NO: 4 або 8. Молекула нуклеїнової кислоти за винаходом може включати (або складатися з) послідовність нуклеїнової кислоти по SEQ ID NO: 1, 3, 7, 9, 10 або 11, причому кодон, що кодує амінокислоту в положенні 118, мав не менше однієї мутації (або був мутований) так, щоб він кодував амінокислоту, відмінну від лейцину, і переважно, щоб він кодував фенілаланін в положенні 118 відповідно до білка FAD2 за винаходом. Переважна молекула нуклеїнової кислоти за винаходом включає (або складається з) послідовність нуклеїнової кислоти SEQ ID NO: 5 або 12. У іншому аспекті, молекула нуклеїнової кислоти за винаходом може кодувати білок FAD2, що має делецію в положенні 118 відносно білка FAD2 дикого типу, представленого амінокислотною послідовністю SEQ ID NO: 4 або 8. Більш конкретно, молекула нуклеїнової кислоти за винаходом кодує білок FAD2, що має делецію лейцину в положенні 118 відносно білка FAD2 дикого типу, представленого амінокислотною послідовністю SEQ ID NO: 4 або 8. Молекула нуклеїнової кислоти за винаходом може включати (або складатися з) послідовність нуклеїнової кислоти по SEQ ID NO: 1, 3, 5, 7, 9, 10 або 11, причому кодон, що кодує амінокислоту в положенні 118, був видалений. Фахівцем буде визнано, що послідовності нуклеїнової кислоти SEQ ID NO: від 1 до 12 (тобто SEQ ID NO: 1, 3, 5, 7, 9, 10, 11 і 12) не єдині послідовності, які можуть бути використані для одержання білка FAD2 за винаходом. Також передбачається, що існують інші молекули нуклеїнової кислоти, які мають інші послідовності, але які, внаслідок виродженості генетичного коду, кодують білок FAD2, що включає заміну амінокислоти в положенні 108 (або відповідної положенню 108) і/або заміну амінокислоти в положенні 118 (або відповідної положенню 118) 4 UA 99099 C2 5 10 15 20 25 30 35 40 45 50 55 відносно амінокислотної послідовності дикого типу, такої як послідовність білка FAD2 дикого типу, представлена SEQ ID NO: 4 або 8. Зокрема, молекула нуклеїнової кислоти за винаходом може включати (або складатися з) нуклеотидну послідовність, що має принаймні 80%, переважно принаймні 85%, більш переважно принаймні 90%, і ще більш переважно принаймні 95%, 96%, 97%, 98% або 99% ідентичності з будь-яким з SEQ ID NO: від 1 до 12 (тобто SEQ ID NO: 1, 3, 5, 7, 9, 10, 11 і 12), або з їх комплементарною формою або їх РНК формою, що кодує білок FAD2, який має заміну амінокислоти в положенні 108 і/або 118 відносно білка FAD2 дикого типу, такого як білок FAD2 дикого типу, представлений SEQ ID NO: 4 або 8. Більш конкретно, молекула нуклеїнової кислоти за винаходом демонструє нуклеотидну послідовність, що має принаймні 80%, переважно принаймні 85%, більш переважно принаймні 90%, і ще більш переважно принаймні 95%, 96%, 97%, 98% або 99% ідентичності з будь-яким з SEQ ID NO: 1, 3, 5, 7, 9, 10, 11 і 12, або з їх комплементарною формою або їх РНК формою, і кодує білок FAD2, який має заміну гліцину аспарагіновою кислотою в положенні 108 (або відповідному положенню 108) і/або заміну фенілаланіну лейцином в положенні 118 (або відповідному положенню 118) відносно білка FAD2 дикого типу, представленого амінокислотною послідовністю SEQ ID NO: 4 або 8. Молекула нуклеїнової кислоти за винаходом може бути одержана з різновидів Brassica napus, таких як MSP05, MSP06, MSP07, MSP11 і/або 28DHS.059. Більш конкретно, молекула нуклеїнової кислоти за винаходом має мутацію в положенні 1540 (також згадувану як SNP1540) з послідовністю нуклеїнової кислоти SEQ ID NO: 11, яка викликає зміну генетичного кодону GGC на GAC, приводячи до заміни амінокислоти в положенні 108 (або відповідному положенню 108) відносно білка FAD2 дикого типу, представленого амінокислотною послідовністю SEQ ID NO: 4 або 8. Виділена молекула нуклеїнової кислоти за винаходом, що містить вказану мутацію SNP1540, яка приводить до заміни аспарагінової кислоти гліцином в положенні 108, змінює функціональність одержуваного продукту гена FAD2, причому рівень олеїнової кислоти збільшується в рослині, що експресує мутантну послідовність, в порівнянні з відповідним рівнем в рослині, що експресує послідовність дикого типу. У рамках винаходу, термін «SNP1540» належить до поліморфізму одиничного нуклеотиду, відповідного вказаній мутації в положенні 1540 нуклеїнової кислоти SEQ ID NO: 11, і може вказувати також на відповідну мутацію в будь-якій молекулі нуклеїнової кислоти, що кодує білок FAD2 за винаходом, який має заміщену амінокислоту в положенні 108 (або відповідному положенню 108) відносно білка FAD2 дикого типу, представленого SEQ ID NO: 4 або 8. Передбачається будь-який фрагмент молекули нуклеїнової кислоти за винаходом з принаймні 10, 15, 20, 25, 50, 100 або більше нуклеотидів, що включає вказану SNP1540. У іншому аспекті, нова молекула нуклеїнової кислоти за винаходом одержана з різновидів Brassica napus, таких як MSP05, MSP11 і/або 28DHS.059, що мають мутацію (SNP1590), яка приводить до заміни амінокислоти в положенні 118 послідовності FAD2 дикого типу, такої як представлена SEQ ID NO: 8. Більш конкретно, молекула нуклеїнової кислоти за винаходом має мутацію в положенні 1590 (також згадувану як SNP1590) з послідовністю нуклеїнової кислоти SEQ ID NO: 12, яка викликає зміну генетичного кодону CTT на TTT, приводячи до заміни амінокислоти в положенні 118 (або відповідному положенню 118) відносно амінокислотної послідовності дикого типу, такої як послідовність білка FAD2 дикого типу, представленого амінокислотною послідовністю SEQ ID NO: 4 або 8. Виділена молекула нуклеїнової кислоти за винаходом, що містить вказану мутацію SNP1590, яка приводить до заміни фенілаланіну лейцином в положенні 118, змінює функціональність одержуваного продукту гена FAD2, причому рівень олеїнової кислоти збільшується в рослині, що експресує мутантну послідовність, в порівнянні з відповідним рівнем в рослині, що експресує послідовність дикого типу. У рамках винаходу, термін «SNP1590» належить до поліморфізму одиничного нуклеотиду, відповідного вказаній мутації в положенні 1590 нуклеїнової кислоти SEQ ID NO: 12, і може вказувати також на відповідну мутацію в будь-якій молекулі нуклеїнової кислоти, що кодує білок FAD2 за винаходом, який має заміщену амінокислоту в положенні 118 (або відповідному положенню 118) відносно білка FAD2 дикого типу, представленого SEQ ID NO: 4 або 8. Передбачається будь-який фрагмент молекули нуклеїнової кислоти за винаходом з принаймні 10, 15, 20, 25, 50, 100 або більше нуклеотидів, що включає вказану SNP1590. 5 UA 99099 C2 5 10 15 20 25 30 35 40 45 50 55 Також передбачається будь-який фрагмент молекули нуклеїнової кислоти за винаходом з принаймні 10, 15, 20, 25, 50, 100 або більше нуклеотидів, що включає вказану SNP1540 і вказану SNP1590. Передбачається будь-який фрагмент молекули нуклеїнової кислоти за винаходом з принаймні 10, 15, 20, 25, 50, 100 або більше нуклеотидів, що включає принаймні одну мутацію, яка приводить до білка FAD2 за винаходом. Іншими словами, також передбачається будь-який фрагмент молекули нуклеїнової кислоти за винаходом з принаймні 10, 15, 20, 25, 30, 40, 50, 100, 500 або більше нуклеотидів, що включає принаймні одну мутацію в кодоні, який кодує вказану амінокислоту в положенні 108 (або відповідному положенню 108) і/або в кодоні, який кодує вказану амінокислоту в положенні 118 (або відповідному положенню 118) відносно білка FAD2 дикого типу, представленого SEQ ID NO: 4 або 8. Подібні фрагменти можуть бути використані як праймери, як зонди і/або як маркери. Фрагменти нуклеїнової кислоти за винаходом можуть бути використані як маркери при генетичному картуванні рослини і в програмах виведення рослини. Подібні маркери можуть включати, наприклад, дослідження поліморфізму довжини рестрикційних фрагментів (RFLP), детектування ампліфікацією з випадковими праймерами (RAPD), полімеразну ланцюгову реакцію (ПЛР) або самопідтримувану реплікацію послідовності (3SR). Методи розмноження із застосуванням маркера можуть бути використані для ідентифікації і відстеження рослини відповідно до винаходу або її потомства, також об'єкта винаходу, в процесі розмноження. Методи розмноження із застосуванням маркера можуть бути використані на доповнення до або як альтернатива іншим видам способів ідентифікації. Прикладом розмноження із застосуванням маркера є застосування праймерів ПЛР, які специфічно ампліфікують молекулу нуклеїнової кислоти за винаходом. Винахід надає методи сегрегації і відбору аналізів генетичних схрещувань визначених рослин, які мають послідовності нуклеїнової кислоти згідно з винаходом. Спосіб за винаходом може, наприклад, залучати визначення присутності в геномі конкретних алелів FAD2, що містять не менше однієї мутації, яка приводить до заміщення (переважно заміщення аспарагінової кислоти гліцином) в положенні 108 (або відповідному) і/або приводить до заміщення (переважно заміщення фенілаланіну лейцином) в положенні 118 (або відповідному) відносно білка FAD2 дикого типу, представленого SEQ ID NO: 4 або 8. Таке визначення може бути, наприклад, досягнуте набором способів, таких як ампліфікація ПЛР, методом «відбитків пальців» ДНК, методом «відбитків пальців» РНК, гель-блотингом і RFLP аналізами, аналізом захисту від нуклеаз, секвенуванням відповідного фрагмента нуклеїнової кислоти, одержанням антитіл (моноклональних і поліклональних) або альтернативними методами, адаптованими для розрізнення білків, продукованих схожими алелями з інших варіантів форм цього білка або від дикого типу. Більш конкретно, такі фрагменти можуть бути використані в способі селекції за допомогою маркера ознак високої олійності в рослинах, переважно видах Brassica, більш переважно в різновидах Brassica napus. Інший аспект представленого винаходу пов'язаний з рекомбінантною нуклеотидною послідовністю, що включає одну або більше прилеглих регуляторних нуклеотидних послідовностей, оперативно зв'язаних з нуклеотидною послідовністю відповідно до винаходу. Вказана прилегла регуляторна послідовність(ті) переважно походять з гомологічного організму. Однак, вказані прилеглі регуляторні послідовності можуть також походити з гетерологічних організмів. Вказані прилеглі регуляторні послідовності являють собою специфічні послідовності, такі як промотори, енхансери, секреторні сигнальні послідовності і/або термінатори. Інший аспект винаходу належить до вектора, який містить молекулу нуклеїнової кислоти за винаходом, можливо оперативно зв'язану з однією або більше прилеглими регуляторними послідовностями, що походять з гомологічних або гетерологічних організмів. У даному контексті слово «вектор» визначене як будь-яка біохімічна конструкція, яка може бути використана для введення нуклеотидної послідовності (шляхом трансдукції, трансфекції, трансформації, інфекції, кон’югації і т. д.) в клітину. Переважно, вектор відповідно до винаходу вибирається з групи, яка складається з плазмід (включаючи реплікативні і інтегративні плазміди), вірусів, фагмід, хромосом, транспозонів, ліпосом, катіонних везикул або їх суміші. Вказаний вектор може вже містити одну або більше 6 UA 99099 C2 5 10 15 20 25 30 35 40 45 50 55 60 прилеглих регуляторних послідовностей, дозволяючи експресію вказаної молекули нуклеїнової кислоти і її транскрипцію в поліпептид за винаходом. Винахід також належить до поліпептиду FAD2, який має амінокислотну заміну в положенні 108 (або відповідному йому) відносно білка FAD2 дикого типу, представленого SEQ ID NO: 4 або 8. Більш конкретно, поліпептид FAD2 за винаходом включає (або складається з) амінокислотну послідовність SEQ ID NО: 6, яка крім того включає заміну аспарагінової кислоти гліцином в положенні 108. Переважний поліпептид FAD2 за винаходом включає (або складається з) амінокислотну послідовність SEQ ID NО: 2. Винахід також належить до поліпептиду FAD2, який має амінокислотну заміну в положенні 118 (або відповідному йому) відносно білка FAD2 дикого типу, представленого SEQ ID NO: 4 або 8. Більш конкретно, поліпептид FAD2 за винаходом включає (або складається з) амінокислотну послідовність SEQ ID NO: 2, яка крім того включає заміну фенілаланіну лейцином в положенні 118. Переважний поліпептид FAD2 за винаходом включає (або складається з) амінокислотну послідовність SEQ ID NО: 6. Даний винахід також включає будь-які фрагменти білка FAD2 за винаходом, які мають дельта-12-олеатдесатуразну активність і включають вказану(і) заміну(и) в положенні 108 і/або 118. Молекули нуклеїнової кислоти, рекомбінантні молекули нуклеїнової кислоти і/або вектори за представленим винаходом придатні для трансформування рослин-мішеней, і таким чином надання продукту зміненого гена FAD2, причому рівень олеїнової кислоти змінюється, переважно збільшується, в рослині, що експресує мутантний FDA2 згідно з винаходом, в порівнянні з відповідним рівнем в рослині, що експресує послідовність дикого типу. Даний винахід також належить до трансформованої клітини-хазяїна або рекомбінантної клітини-хазяїна, що містить (або має вбудовану) одну або більше нуклеотидних послідовностей і/або векторів відповідно до винаходу. У представленому контексті, «трансформована клітина-хазяїн» або «рекомбінантна клітина», також згадувана як «трансформант», являє собою клітину, яка має включеними одну або більше нуклеотидних послідовностей і/або векторів відповідно до винаходу. Трансформована клітина-хазяїн може бути клітиною з вказаним(и) вектором(ами) і/або вказаною(ими) нуклеотидною(ими) послідовністю(ями), включеною(ими) засобами генетичної трансформації, переважно засобами гомологічної рекомбінації або іншими добре відомими способами, використовуваними для одержання рекомбінантного організму. Будь-який спосіб, за допомогою якого нова послідовність може бути включена в геном хазяїна, передбачається представленим винаходом. Більш переважно, будь-який спосіб, за допомогою якого нова послідовність може бути включена в геном хазяїна і стабільно успадковує його потомство, передбачається представленим винаходом. У цей час існує широкий спектр відомих способів досягнення прямої або непрямої трансформації вищих рослин екзогенною ДНК. Трансформація рослинних клітин може бути опосередкована використанням векторів. Звичайний спосіб досягнення трансформації являє собою використання Agrobacterium tumefaciens для включення чужорідного гена в цільову клітину рослини. Віруси рослини також пропонують можливі засоби для передачі екзогенної ДНК. Також може бути застосоване пряме введення в рослинні клітини. Звичайно, протопласти цільової рослини поміщуються в культуру в присутності молекул нуклеїнової кислоти, що переносяться, і фактора, який збільшує поглинання вказаних молекул нуклеїнової кислоти протопластом. Корисними факторами в цьому відношенні є поліетиленгліколь і фосфат кальцію. Альтернативно, поглинання молекул нуклеїнової кислоти може бути стимульоване електропорацією. У цьому способі, електричний імпульс використовується для відкривання тимчасових пор в клітинній мембрані протопласта, і вказані молекули нуклеїнової кислоти з оточуючого розчину проникають в клітину через пори. Схожим чином, для доставки молекул вказаної нуклеїнової кислоти безпосередньо в клітину і переважно безпосередньо в ядро клітини може застосовуватися мікроін’єкція. У цих способах, трансформація відбувається в рослинній клітині в культурі. Після події трансформації рослинні клітини можуть бути регенеровані в цілі рослини. Способи для регенерації дорослих рослин з калюсу або протопластів добре відомі. 7 UA 99099 C2 5 10 15 20 25 30 35 40 45 50 55 60 Також доступні альтернативні способи, які не вимагають застосування ізольованих клітин і, таким чином, способів регенерації рослин, для досягнення трансформації. Вони звичайно згадуються як «балістичні» або способи з «прискоренням частинок», в яких молекули нуклеїнової кислоти, що покривають металеві частинки, заштовхуються в клітини рослини пороховим зарядом або електричним розрядом. Цим способом рослинні клітини в культурі або репродуктивні органи або клітини рослин, наприклад, пилок, можуть бути стабільно трансформовані цікавлячими молекулами нуклеїнової кислоти. Даний винахід може бути застосований до трансформації практично будь-якого типу рослин, однодольних або дводольних. Придатні рослини для трансформації являють собою переважно олійні культури, такі як соняшник, соєвий біб, бавовна, кукурудза і т. п., переважно види Brassica, більш переважно різновиди Brassica napus. У одному аспекті винаходу, рослини включають не менше однієї послідовності, яка кодує FAD2 за винаходом. Рослина за винаходом може включати послідовність нуклеїнової кислоти SEQ ID NО: 5 або послідовність нуклеїнової кислоти SEQ ID NО: 12. Переважно, рослина за винаходом містить послідовність нуклеїнової кислоти SEQ ID NO: 11 або послідовність нуклеїнової кислоти SEQ ID NO: 1, таку як MSP06 або MSP07. У іншому аспекті винаходу, рослина містить дві послідовності, які кодують FAD2 за винаходом. Зокрема, рослина за винаходом містить послідовність нуклеїнової кислоти SEQ ID NO: 11 і послідовність нуклеїнової кислоти SEQ ID NO: 12, таку як MSP05, MSP11 або 28DHS.059. Переважно, рослина за винаходом містить послідовність нуклеїнової кислоти SEQ ID NO: 1 і послідовність нуклеїнової кислоти SEQ ID NO: 5, таку як MSP05, MSP11 або 28DHS.059. Різновид MSP06 підтримується як патентний депонент за Будапештською Угодою в NCIMB під номером доступу NCIMB 41367, поміщено 22 грудня 2005 р. Різновид MSP07 підтримується як патентний депонент за Будапештською Угодою в NCIMB під номером доступу NCIMB 41368, поміщено 22 грудня 2005 р. Різновид 28DHS.059 підтримується як патентний депонент за Будапештською Угодою в NCIMB під номером доступу NCIMB 41364, поміщено 22 грудня 2005 р. Різновид MSP05 підтримується як патентний депонент за Будапештською Угодою в NCIMB під номером доступу NCIMB 41233, поміщено 9 липня 2004 р. Різновид MSP11 підтримується як патентний депонент за Будапештською Угодою в NCIMB під номером доступу NCIMB 41234, поміщено 9 липня 2004 р. Інший об'єкт винаходу являє собою спосіб виробництва високоолеїнових ліній рослини, який включає: (a) схрещування першої рослини з другою рослиною, що має принаймні один ген мутантного FAD2 відповідно до винаходу, (b) одержання насіння від схрещування за етапом (a); (с) вирощування фертильних рослин з подібного насіння; (d) одержання насіння потомства від рослин етапу (с), і (е) ідентифікацію серед потомства насіння, яке має високий вміст олеїнової кислоти. У іншому аспекті, винахід пропонує спосіб збільшення вмісту олеїнової кислоти в рослинах, більш конкретно в рослинах Brassica, і переважно в рослинах Brassica napus, який включає етапи: (a) індукції мутагенезу в принаймні деяких клітинах рослини, більш конкретно рослини Brassica, і переважно рослини Brassica napus, яка має вміст олеїнової кислоти менше ніж 70%; (b) регенерації рослин з принаймні однієї мутагенізованої клітини; (с) відбору регенерованих рослин, які мають послідовність нуклеїнової кислоти за винаходом і/або які експресують білок FAD2 за винаходом; і (d) одержання подальших поколінь рослин з вказаних регенерованих рослин. Переважно, насіння, одержане від вказаних рослин, дає олію, що має вміст олеїнової кислоти більше ніж 70% ваг., більш переважно більше ніж 75% ваг., відносно загальної ваги жирних кислот, присутніх в олії. Інший об'єкт винаходу являє собою рослинну олію, одержану з принаймні однієї рослини відповідно до винаходу, яка містить більше ніж (приблизно) 70%, 72%, 75%, 80% або 85% олеїнової кислоти. Більш конкретно, рослинна олія за винаходом, одержана переважно з принаймні одного виду Brassica за винаходом, більш переважно з одного з різновидів Brassica napus відповідно до винаходу, містить більше ніж (приблизно) 70%, 72%, 75%, 80% або 85% олеїнової кислоти. Вказана олія може, крім того, містити менше ніж (приблизно) 4%, 3,5%, 3%, 2%, 1% або 0,5% ліноленової кислоти відносно загальної ваги жирних кислот, присутніх в олії. 8 UA 99099 C2 5 10 15 20 25 30 35 40 45 50 55 60 Переважно, вказана олія містить більше ніж (приблизно) 70%, 72%, 75%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89% або 90%, переважно між (приблизно) 70% і (приблизно) 90%, більш переважно між (приблизно) 72% і (приблизно) 89%, олеїнової кислоти. Вказана олія може також включати менше ніж (приблизно) 4%, 3,5%, 3%, 2%, 1% або 0,5%, переважно між (приблизно) 4% і (приблизно) 0,4% ліноленової кислоти відносно загальної ваги жирних кислот, присутніх в олії. Відповідно до переважного варіанта здійснення, два подвійні низькі озимі різновиди рапсу (ENVOL і LIBERATOR) були відправлені на обробку етилметансульфонатом (EMS) в 1992 р. Обробка EMS проводилася при 2,5% і 5% протягом 4 год. або 8 год. Покоління М1 було вирощене в оранжереї після 8 тижнів яровизації в ростовій камері і потім зібране в липні 93 р. Насіння М1 було висаджене в полі у вересні 93 р., закрите мішками на початку цвітіння, і насіння М2 було зібране в липні 94 р. Насіння М2 було висаджене у вересні 94 р., закрите мішками на початку цвітіння, і насіння М3 було зібране в липні 95 р. Потім склад жирних кислот потомства був проаналізований із застосуванням аналітичного методу, основаного на газовій хроматографії, який добре відомий в цій галузі технології. Всі потомки, що показують вміст олеїнової кислоти вище 68%, були збережені. Вибране потомство було знов висаджене в полі у вересні 1995 р., закрито мішками в квітні і зібране в липні 1996 р. На цій стадії потомки були досліджені на хороші агрономічні і морфологічні характеристики, такі як хороша здатність до проростання, хороша сила восени, хороша стійкість взимку, хороша коренева система, хороша стійкість до чорної ніжки і світлої плямистості (light leaf spot), також як і чудова стійкість до вилягання. Матеріал, який був дуже високим або дуже пізнім, був видалений, також як і матеріал, що демонструє високу фасціацію. Був знов проведений аналіз потомства, що залишилося, за допомогою газової хроматографії для відбору індивідуумів з рівнями олеїнової кислоти вище ніж 68%. Всі ці індивідууми були висаджені в полі у вересні 1996-1997 рр. Потомство, назване MUT152-96, виглядало особливо цікаво відносно агрономічних і морфологічних характеристик, також як і відносно вмісту олеїнової кислоти в ньому. Воно було культивоване в ізоляції під час посівного сезону у вересні 1996-1997 рр. Найбільш цікаве потомство відносно агрономічних і морфологічних характеристик було вибране для ізоляції мішками і схрещування. Схрещування проводилося з різновидами подвійного нижнього озимого олійного рапсу, що мають звичайний профіль жирної кислоти (тобто олеїнову кислоту нижче 70%) або з низьким вмістом ліноленової кислоти (тобто менше ніж приблизно 3,5%) для одержання ліній з високим вмістом олеїнової кислоти, пов'язаним з низьким вмістом ліноленової кислоти (HOLL). Матеріал був проведений через лінійне розведення, самозапилення проводилося принаймні до покоління F7. На всіх поколіннях застосовувався сильний селективний тиск проти фасціації і на користь нормального розвитку рослини і нормальної кореневої системи. Склад жирної кислоти відстежувався в кожному поколінні і зберігався тільки матеріал із вмістом олеїнової кислоти вище ніж 75% і ліноленової кислоти нижче ніж 3,5%. В результаті цього процесу були одержані наступні різновиди HOLL: MSP05, MSP06, MSP07, MSP11, 28DHS059. Подвійними низькими різновидами із звичайними профілями жирної кислоти, використаними в даній роботі, були BRISTOL, CAPITOL, CAPVERT, VIVOL і CAIMAN і розмноження або підтримання їх проводилося за такою ж схемою догляду, що і описана тут вище для ліній HOLL. Основне насіння було використане для визначення вмісту жирної кислоти у випробуваннях 2 невеликих дослідницьких випробуваннях (від 6 до 12 м ) або випробуваннях при розробці (500 2 м ) і для робіт з секвенування. ПРИКЛАДИ Приклад 1 Насіння було подрібнене в першому розчині, який складається з метанолу (800 мл), триметилпентану (200 мл) і 5 г NaOH. Приблизно 3 мл розчину було використано для приблизно 10 г насіння (іншими словами, приблизно від 10 до 50 насінин на 1 мл розчину). Екстракція проводилася протягом 20 хвилин, після чого був доданий другий розчин, який складається з триметиламіну (900 мл) і пропанолу-2 (100 мл) в тому ж об'ємі, що і перший розчин. 9 UA 99099 C2 5 Одержаний розчин був збовтаний і залишений у спокої до формування верхньої фази. Верхня фаза була відібрана і перенесена в посудини. Один мікролітр її був впорснутий в газовий хроматограф (Fisons thermo-electron з колонкою DB3 - 30 метрів з діаметром 0,25 мм і товщиною 25 мкм). Час пробігу був приблизно 4 хв. Вміст олеїнової кислоти був узагальнений в таблиці 1. Таблиця 1 Різновиди MSP05 MSP06 MSP07 MSP11 28DHS059 BRISTOL VIVOL CAPVERT CAIMAN CAPITOL 10 15 20 25 30 35 40 45 Вміст олеїнової кислоти 78,1-81,9 75,6-78,5 76,7-79,4 80,2-83,9 83,8-84,9 61,4-65,7 60,8-63,2 58,9-65,9 61,9-64,0 59,7-64,6 Оцінка Дуже висока Висока Висока Дуже висока Дуже висока Нормальна Нормальна Нормальна Нормальна Нормальна Вміст олеїнової кислоти відносно загальної ваги жирної кислоти у виділеній олії. Приклад 2 Рослинні матеріали, використані для секвенування: - мутантні лінії з підвищеним вмістом олеїнової кислоти MSP05, MSP06, MSP07, MSP11, 28DHS.059; і - різновиди дикого типу з нормальним вмістом олеїнової кислоти: Bristol, Capitol, Vivol, Capvert і Caiman. Всі ці лінії були вирощені в ростовій камері, і у 7-денних рослин були зібрані стебла. Рослинні тканини були ліофілізовані і використані для виділення ДНК. ДНК була виділена з використанням наборів для виділення рослинної ДНК Qiagen (Qiagen INC. - USA, Valencia CA). ПЛР проводився по протоколу TaqGold (AB Biosystem, Inc.). Реакційна суміш включає 2,5 мкл 10× буфера, 0,2 мкл TaqGold, 0,2 мкл dNTP (25 мМ), 2 мкл праймерів (5 мкМ) і 10 мкл ДНК-матриці (2 нг/мкл) і 10,1 мкл Н2О. Цикли ПЛР були наступними: 94°С 5 хв.; 8 циклів 94°С 40 сек., 62°С 40 сек., 72°С 1 хв., 94°С 40 сек., 60°С 40 сек., 72°С 1 хв., 94°С 40 сек., 58°С 40 сек., 72°С 1 хв., 94°С 40 сек., 56°С 40 сек., 72°С 1 хв., 3 цикли 94°С 40 сек., 55°С 40 сек., 72°С 1 хв.; тримати при 72°С 7 хв. Продукти ПЛР були проаналізовані в 1% агарозному гелі. Для секвенування 5 мкл продуктів ПЛР було відділено в нову пробірку і туди був доданий 1 мкл Екзонуклеази I (розведення 1:50) і 1 мкл лужної фосфатази з креветок (розведення 1:5). Суміш була інкубована при 37°С протягом 20 хв. і потім при 80°С протягом 15 хв. для інактивації ферментів. Було додано 40 мкл Н2О і 6 мкл було використано як матриця з 1 мкл праймера для секвенування. Секвенування проводили на ДНК-аналізаторі 3730 (Applied Biosystems). Послідовності були зібрані і порівняні з використанням програми SeqMan II від LAserGene (DNASTAR, INC., Madison. WI). Приклад 3 Чотири послідовності генів дельта-12-олеатдесатурази (FAD2) Brassica napus, 4684997, 46399190, 8705228 і 4092878 були завантажені з Генбанку (NCBI). Ці послідовності були використані як запити BLAST проти бази даних послідовностей Monsanto. З використанням програми «blastn» (NCBI), було одержано декілька висококомплементарних результатів. Всі ці послідовності були завантажені і зібрані наново програмою SeqmanII (DNASTAR, INC., Madison, Wisconsin, USA). Два різних транскрипти були ідентифіковані і позначені як Fad2-1 (SEQ ID NO: 9) і Fad2-2 (SEQ ID NO: 10). Fad2-1 і Fad2-2 мають високу гомологію послідовності, з 97% ідентичності послідовності. Для ідентифікації значущих мутацій, пов'язаних з високим вмістом олеїнової кислоти в мутантних лініях і їх потомстві, були створені гніздові локус-специфічні праймери для покривання цілих послідовностей. 10 UA 99099 C2 5 10 3' кінець праймера завжди знаходився на нуклеотиді, який відрізняв Fad2-1 від Fad2-2 за винятком тих, які розташовані на 5' і 3' кінцях консенсусних послідовностей, в яких немає розрізнюваних у двох генів нуклеотидів. Праймери були також створені таким чином, що один амплікон буде перекривати інший для забезпечення повного покривання всієї послідовності. Ці праймери були складені і використані для одержання локус-специфічних ампліконів мутантного і дикого типу. Результати секвенування показали, що всі локус-специфічні ПЛР праймери поводилися як очікувалося. Послідовності, що належать до одного гена, були зібрані разом з використанням програми SeqManII. Консенсусні геномні послідовності мутантних генів Fad2-1 і Fad2-2 представлені відповідно SEQ ID NO: 11 і 12. Таблиця 2 узагальнює властивості послідовностей обох генів Fad2-1 і Fad2-2. Таблиця 2 Елементи Ген 5'UTR Екзон Інтрон Екзон CDS 3'UTR 15 20 25 30 35 40 45 Положення в Fad2-1 1-2614 1-1217 1-108 109-1213 1214-2614 1218-2372 2373-2614 Положення в Fad2-2 1-2666 1-1238 1-111 112-1234 1235-2619 1239-2393 2394-2666 Елементи основані на консенсусних геномних послідовностях з різних прочитань різних генотипів. І ген Fad2-1 і ген Fad2-2 мають по одному інтрона кожний. Розміри інтронів злегка розрізнюються у двох генів. Для Fad2-1 розмір інтрона становить 1105 п.о., починаючи з положення 109 до положення 1213, в той час як для Fad2-2 інтрон складається з 1123 п.о., починаючи з положення 112 до положення 1234 на консенсусних послідовностях. Інтрон розташований на ділянці 5'UTR. Передбачувані кодони ініціації трансляції розташовані в положеннях 1218 і 1239, відповідно, для Fad2-1 і Fad2-2. Кодони термінації трансляції розташовані в положеннях 2370-2372 і 2391-2393, відповідно, для Fad2-1 і Fad2-2. Послідовності 3'UTR являють собою 247 пар основ у випадку гена Fad2-1 і 273 пари основ у випадку гена Fad2-2. Була виявлена мутація трансляції в положенні 1540 (звана SNP1540) гена FAD2-1 (як представлено SEQ ID NO: 11), яка викликала зміну генетичного кодону GGC на GAC, приводячи до зміни амінокислотного залишку з гліцину на аспарагінову кислоту. Оскільки гліцин і аспарагінова кислота мають дуже різні властивості відносно гідрофобності, зарядів і полярності і т. д., мутація викликає радикальні зміни в роботі ферменту в мутантних лініях. Також були описані висококонсервативні амінокислотні послідовності серед рослинних дельта-12-десатураз жирної кислоти і рослинних дельта-15-десатураз жирної кислоти (патент США US6872872B1). Серед інших, одна з вказаних консервативних ділянок амінокислотної послідовності являє собою AHECGH. Виявилося, що SNP1540 знаходиться в тій же ділянці. «G» цієї ділянки була мутована в «D». Оскільки консервативність ділянок звичайно означає їх функціональну або структурну значущість, мутація в цій консервативній ділянці викликала побічні ефекти в ферменті Fad2-1, приводячи до високого вмісту олеїнової кислоти в мутантних лініях MSP11, MSP05, MSP06, MSP07 і 28DHS.059. Точкова мутація в положенні 1590 (названа SNP1590) гена FAD2-2 (як представлено SEQ ID NO: 12) викликала зміну амінокислотного залишку з лейцину (CTT) на фенілаланін (TTT). І лейцин і фенілаланін мають гідрофобну природу і мають деякі схожі амінокислотні властивості, але фенілаланін містить велику жорстку ароматичну групу на бічному ланцюгу, яка викликає деякі зміни в роботі ферменту. Більше того, в комбінації з мутацією SNP1540, мутація викликає більш помітний вплив на фенотип. 11 UA 99099 C2 5 10 15 20 Поєднання різних алелів при таких мутаціях створювало градієнт на вміст олеїнової кислоти, як відмічено в різних мутантних лініях (див. таблицю 1). Три мутантні лінії, MSP11, MSP05 і 28DHS.059, несли подвійні мутації в SNP1540 і SNP1590. Оскільки обидві мутації були місенс-мутаціями, функції гена FAD2 були серйозно порушені, приводячи до найбільшого вмісту олеїнової кислоти серед мутантних ліній. Вміст олеїнової кислоти для MSP05 був нижчим, ніж для інших двох мутантів. Це відбулося внаслідок того, що вміст олеїнової кислоти був одержаний за даними тільки одного року, які можуть бути об'єктом варіацій у зв'язку з впливом навколишнього середовища. Дві мутантні лінії, MSP06 і MSP07, несли одиничну точкову мутацію в SNP1540. Оскільки вони менш серйозні, ніж у подвійних мутантів, вміст олеїнової кислоти для цих двох ліній був злегка нижчим, ніж для подвійних мутантів. Загалом, дані послідовності чітко показували, що ці мутації в Fad2-1 і Fad2-2 сильно пов'язані з вмістом олеїнової кислоти в різних мутантних лініях. Поєднання різних алелів пояснює всі фенотипічні варіації вмісту олеїнової кислоти в одержаних рослинних матеріалах. Ідентифікація кількісних варіацій послідовності критично важлива для створення діагностичних аналізів, специфічних для кожного мутантного алеля. Знання про зв'язок між варіаціями послідовності і фенотипом можуть дозволити створити маркерні аналізи для точного прогнозу вмісту олеїнової кислоти в рослинах без необхідності хімічного аналізу вмісту жирної кислоти. Перелік послідовностей 12 UA 99099 C2 13 UA 99099 C2 14 UA 99099 C2 15 UA 99099 C2 16 UA 99099 C2 17 UA 99099 C2 18 UA 99099 C2 19 UA 99099 C2 20 UA 99099 C2 21 UA 99099 C2 22 UA 99099 C2 23 UA 99099 C2 24 UA 99099 C2 25 UA 99099 C2 26 UA 99099 C2 27 UA 99099 C2 28
ДивитисяДодаткова інформація
Назва патенту англійськоюMutants fad-2 and high-oleinic plants
Автори англійськоюDespeghel, Jean-Pierre, Grainier Kristel
Назва патенту російськоюМутанты fad-2 и высокоолеиновые растения
Автори російськоюДеспегель Жан-Пьер, Граньер Кристель
МПК / Мітки
МПК: A01H 5/00, C12N 15/82, C12N 9/02
Мітки: fad-2, мутанти, високоолеїнові, рослини
Код посилання
<a href="https://ua.patents.su/42-99099-mutanti-fad-2-i-visokoolenovi-roslini.html" target="_blank" rel="follow" title="База патентів України">Мутанти fad-2 і високоолеїнові рослини</a>
Fad-2 мутанти і рослини з високим вмістом олеїнової кислоти

Номер патенту: 96442
Опубліковано: 10.11.2011
Автори: Гуген Неллі, Деспегель Жан-П'єр, Ву Куншенг
МПК: C12N 15/82, C12Q 1/68, C12N 9/02, A01H 5/02
Мітки: кислоти, високим, олеїнової, мутанти, вмістом, fad-2, рослини
Формула / Реферат:
1. Ізольована молекула нуклеїнової кислоти, що містить послідовність SEQ ID NO: 1 або 5 з додатково видаленим нуклеотидом в положенні 215 або (варіантну) послідовність нуклеїнової кислоти, що кодує білок FAD2, щонайменше на 80 % ідентичну SEQ ID NO: 1 або 5 з додатково видаленим нуклеотидом, що відповідає положенню 215.2. Ізольована молекула нуклеїнової кислоти за п. 1, що містить послідовність SEQ ID NO: 9 з додатково видаленим...
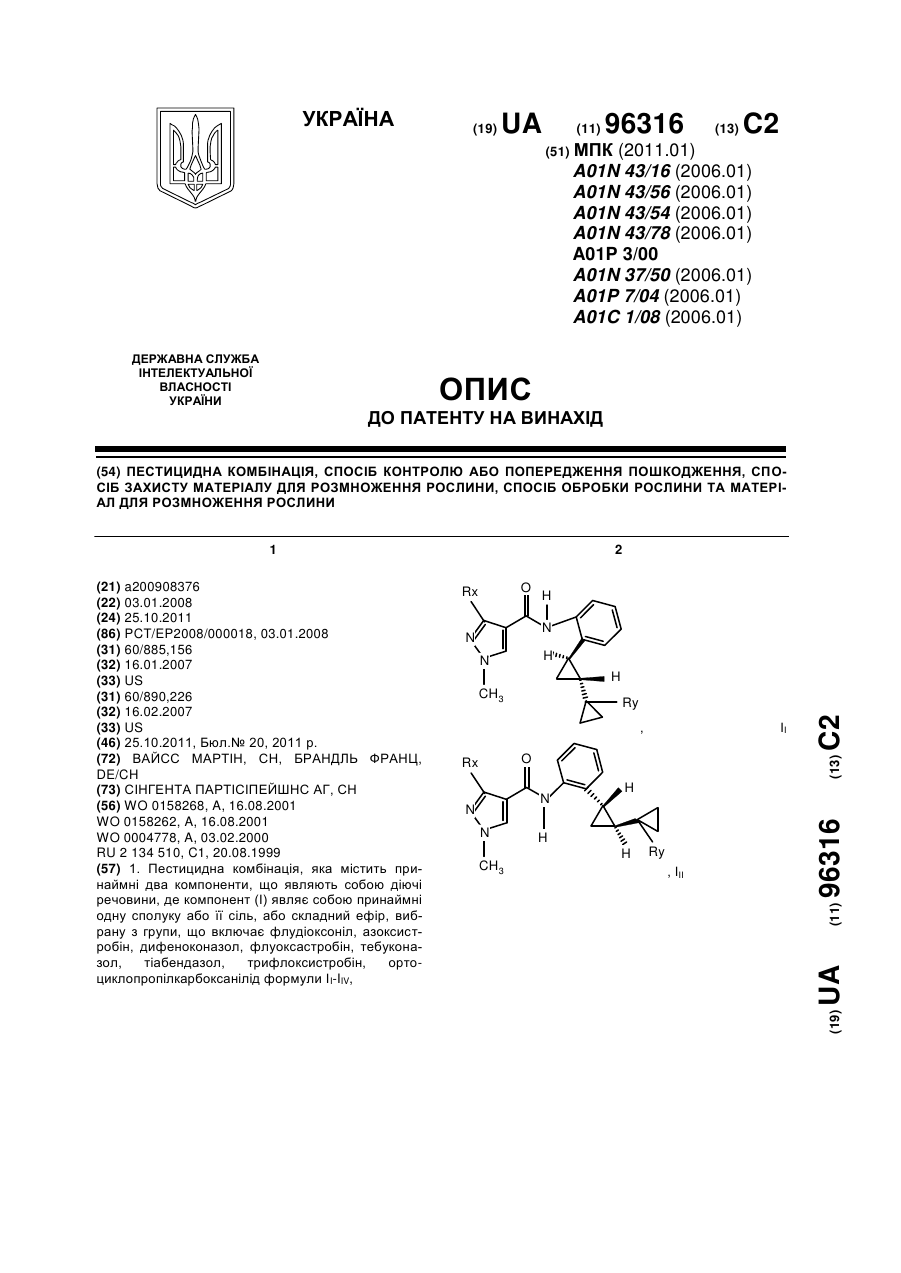
Пестицидна комбінація, спосіб контролю або попередження пошкодження, спосіб захисту матеріалу для розмноження рослини, спосіб обробки рослини та матеріал для розмноження рослини
Номер патенту: 96316
Опубліковано: 25.10.2011
Автори: Брандл Франц, Вайсс Мартін
МПК: A01P 7/04, A01N 43/56, A01C 1/08, A01N 43/54, A01P 3/00, A01N 37/50, A01N 43/16, A01N 43/78
Мітки: матеріалу, пошкодження, матеріал, контролю, захисту, спосіб, пестицидна, комбінація, обробки, попередження, рослини, розмноження
Формула / Реферат:
1. Пестицидна комбінація, яка містить принаймні два компоненти, що являють собою діючі речовини, де компонент (І) являє собою принаймні одну сполуку або її сіль, або складний ефір, вибрану з групи, що включає флудіоксоніл, азоксистробін, дифеноконазол, флуоксастробін, тебуконазол, тіабендазол, трифлоксистробін, opтo-циклопропілкарбоксанілід формули II-IIV,, ІІ
Плазміда pslc1-1/prd400, штам gv3101 agrobacterium tumefaciens, спосіб одержання трансгенної рослини з масловмісним насінням, спосіб виділення їстівної або неїстівної олії з масловмісного насіння рослини

Номер патенту: 49833
Опубліковано: 15.10.2002
Автори: Тейлор Девід С., Маккензі Самюел Л., Зоу Джітао, Келлер Вілфред А., Катавік Весна
МПК: C12N 15/09, A01H 5/00, C11B 1/00, C12N 1/21, C12N 15/00, C12N 5/10, C12N 9/10, C11B 1/10, C12N 15/82
Мітки: gv3101, неїстівної, штам, насіння, виділення, tumefaciens, спосіб, масловмісним, плазміда, олії, їстівної, рослини, насінням, agrobacterium, одержання, трансгенної, масловмісного
Формула / Реферат:
1. Плазміда pSLC1-1/pRD400 (ATCC 97545).2. Штам GV3101 Agrobacterium tumefaciens, який відрізняється тим, що вищезгаданий штам модифікований введенням дріжджового гена SLC1-1, що має нуклеотидну послідовність SEQ ID NO: 1.3. Спосіб одержання трансгенної рослини з масловмісним насінням, який відрізняється тим, що в геном вищезгаданої рослини вводять дріжджовий ген SLC1-1, що здатен до експресії і який має нуклеотидну...
Теплостабільні мутанти ферментів біосинтезу крохмалю

Номер патенту: 84669
Опубліковано: 25.11.2008
Автори: Берджер Браян, Ханнан Кертіс Л., Грін Томас У.
МПК: C12N 9/14, C12N 15/82, C12N 15/29, A01H 5/00, C12N 15/55
Мітки: мутанти, ферментів, біосинтезу, крохмалю, теплостабільні
Формула / Реферат:
1. Полінуклеотид, що кодує мутантну велику субодиницю поліпептиду рослинної ендоспермної АДФ-глюкозопірофосфорилази або біологічно активний фрагмент згаданого мутантного поліпептиду, причому згаданий мутантний поліпептид включає амінокислотні мутації у двох або більше сайтах амінокислотної послідовності згаданого поліпептиду, та такий, що, коли згаданий мутантний поліпептид експресується з малою субодиницею АДФ-глюкозопірофосфорилази з...
Застосування рослини або екстракту з рослини для приготування композиції, призначеної для попередження, полегшення та/або лікування захворювань кісток або для підтримання здоров’я кісток у людини або хатніх тварин

Номер патенту: 84123
Опубліковано: 25.09.2008
Автори: Федерічі Ерманно, Оффорд-Кейвін Елізабет, Лемор Бернар, Куртуа Дідьє
МПК: A61P 19/10, A61P 3/14, A61K 127/00, A61K 36/185, A61K 36/48, A61K 36/23, A61P 19/08, A61P 19/00, A61K 125/00, A23K 1/16, A61K 135/00, A61K 36/73, A61K 36/53, A61K 131/00, A61K 133/00, A61K 36/28, A23L 1/30
Мітки: полегшення, тварин, застосування, призначеної, композиції, людини, здоров'я, захворювань, хатніх, приготування, екстракту, лікування, кісток, підтримання, попередження, рослини
Формула / Реферат:
1. Застосування рослини або екстракту однієї або більше рослин, що містить фітохімічні сполуки, здатні стимулювати кістковий морфогенний білок та/або інгібувати резорбцію кісток, де рослину вибрано з групи, яка включає: надземні частини Lindera benzoin, надземні частини Artemisia vulgaris, кореневища (різома) Acorus calamus, насіння або квіти Carthamus tinctorius, плоди Amelanchier ovalis, плоди Amelanchier alnifolia, листя або коріння...
Попередній патент: Спосіб затвердіння однокомпонентних пінополіуретанів, спосіб виготовлення виробів з поліуретану у формі та пристрій для їх здійснення
Наступний патент: Спосіб діагностики пошкодження теплозахисного покриття тонкостінної системи
Випадковий патент: Частина, що охоплюється, і частина, що охоплює, для вузла зношуваних елементів ковша машини для земляних робіт