Композиція пухлино-асоційованих пептидів та відповідна протиракова вакцина
Номер патенту: 103882
Опубліковано: 10.12.2013
Автори: Левандровскі Петер, Вальтер Штеффен, Зінгх Харпреет, Траутвайн Клаудія, Вайншенк Тоні, Хільф Норберт, Шор Олівер





































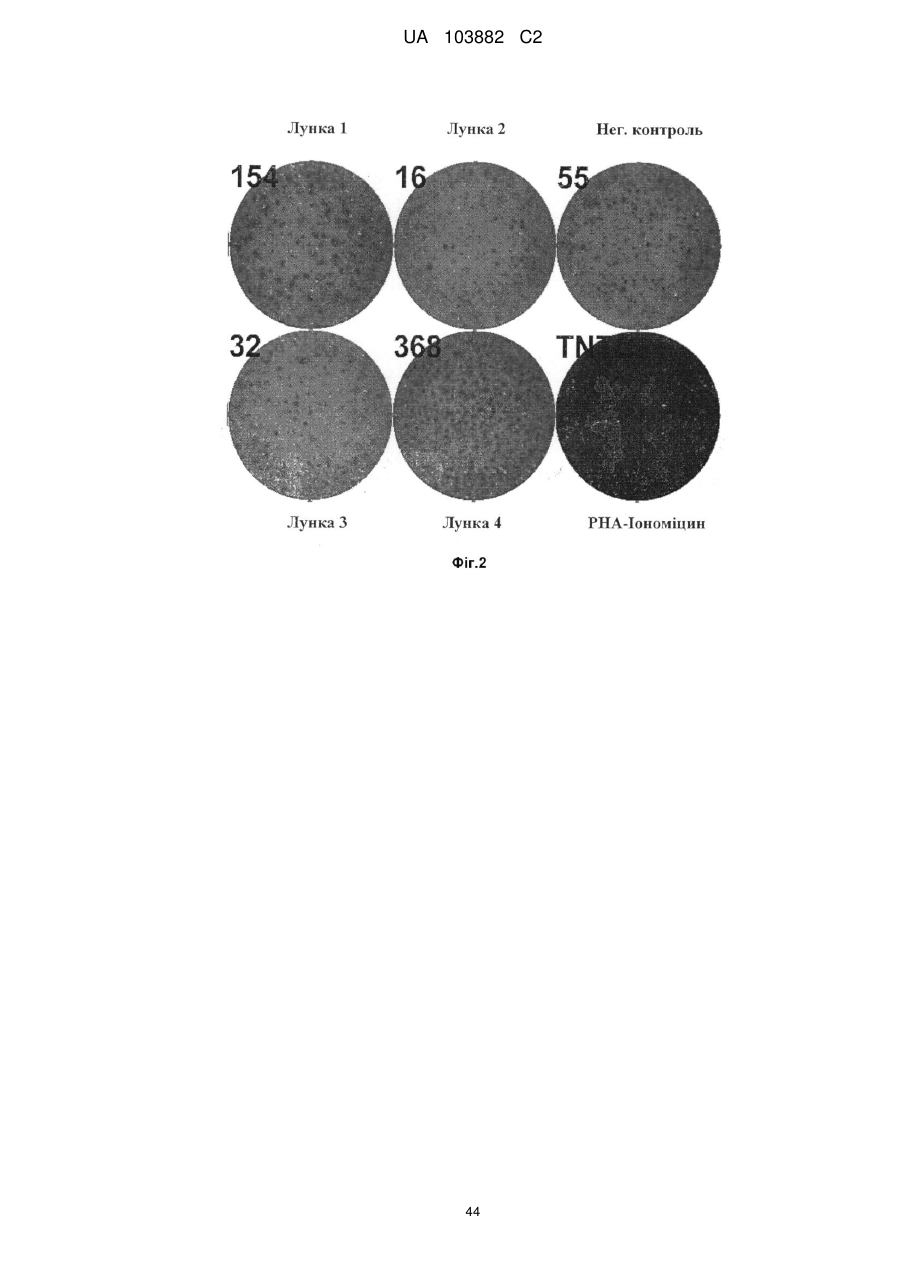
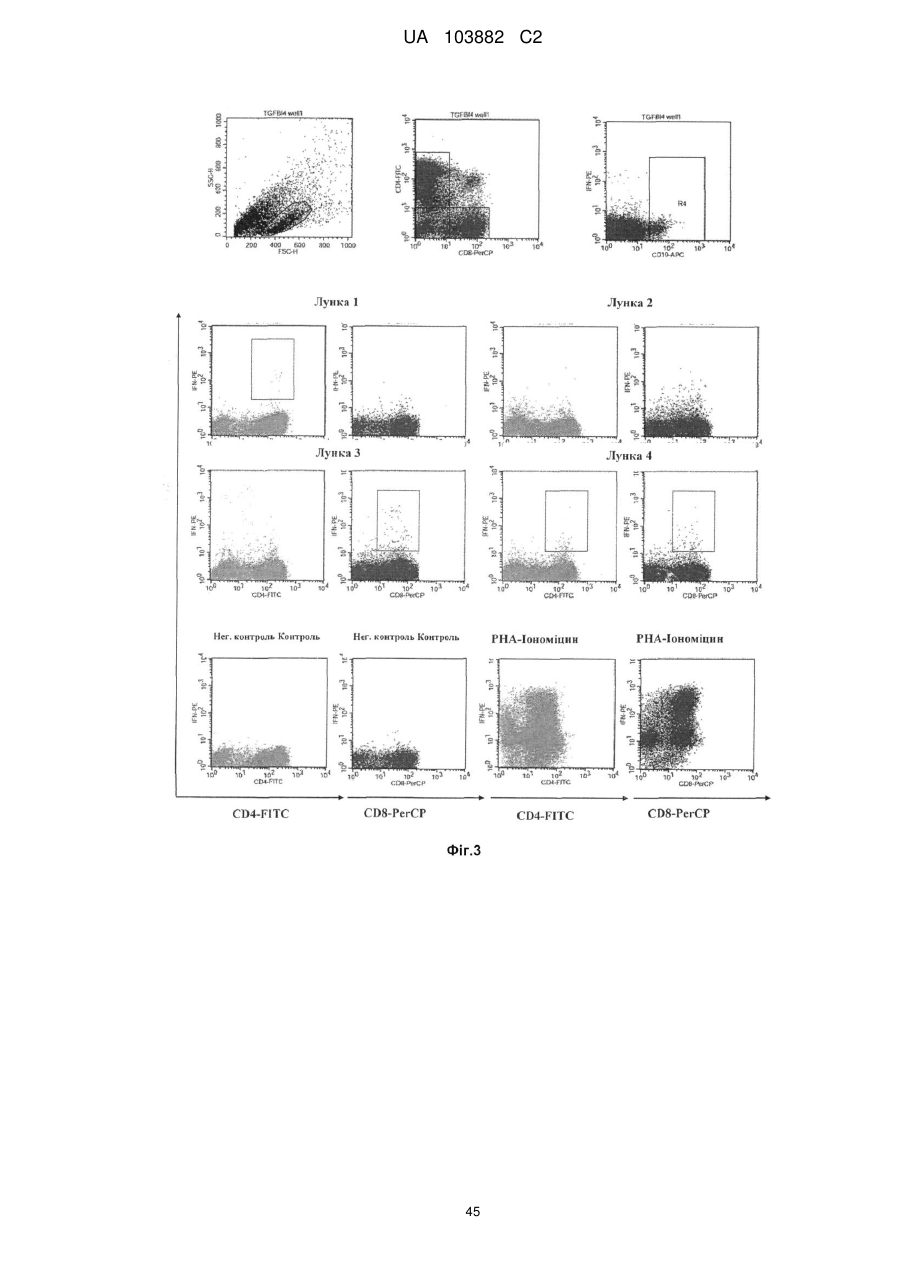
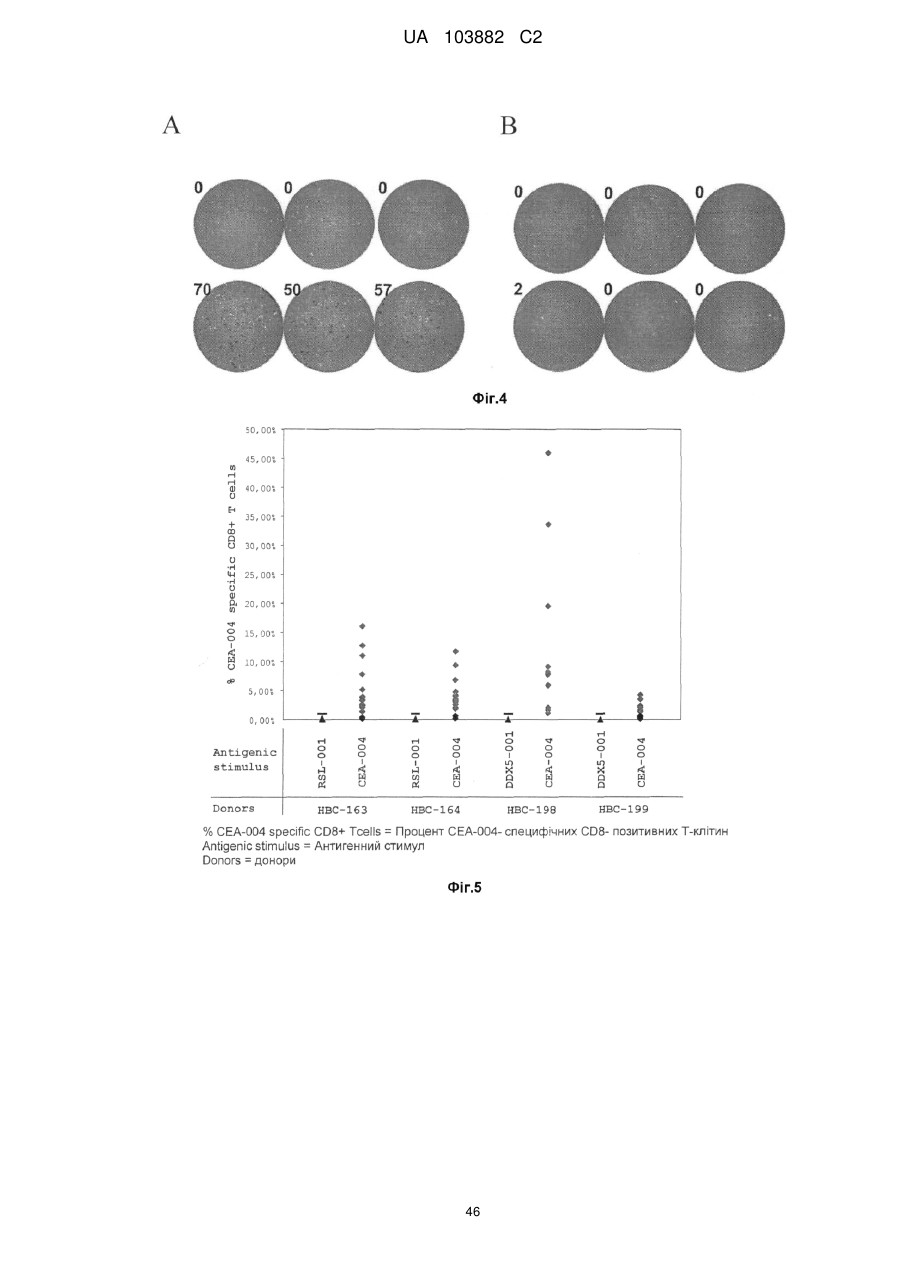
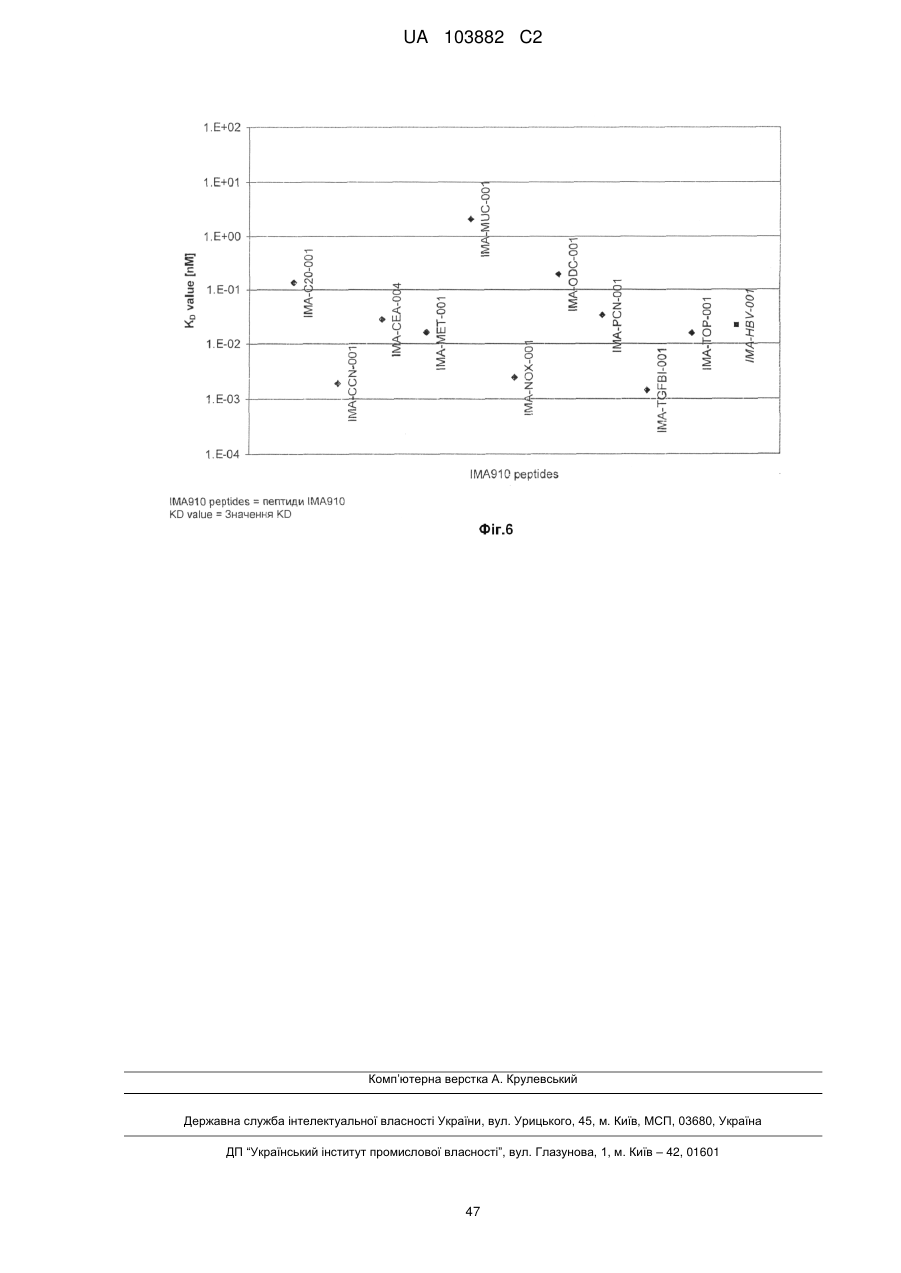
Формула / Реферат
1. Фармацевтична композиція, яка містить щонайменше два пептиди, що включають амінокислотну послідовність SEQ ID NO: 1 та SEQ ID NO: 2, та/або включають амінокислотну послідовність, яка щонайменше на 80 % ідентична SEQ ID NO: 1 та SEQ ID NO: 2, та фармацевтично прийнятний носій, яка відрізняється тим, що пептиди мають здатність зв'язуватися з молекулою головного комплексу гістосумісності людини (МНС) класу І або II класу, та зазначені пептиди мають загальну довжину від 8 до 100 амінокислот.
2. Фармацевтична композиція за п. 1, яка відрізняється тим, що додатково містить щонайменше один додатковий пептид, який включає амінокислотну послідовність, вибрану з групи, що складається з послідовностей SEQ ID NO: 8 − SEQ ID NO: 15, або включає амінокислотну послідовність, яка щонайменше на 80 % ідентична послідовності, вибраній з послідовностей SEQ ID NO: 8 − SEQ ID NO: 15.
3. Фармацевтична композиція за пп. 1 або 2, яка відрізняється тим, що пептиди мають загальну довжину від 8 до 30 або від 8 до 16 амінокислот.
4. Фармацевтична композиція за пп. 1 або 2, у якій щонайменше один пептид включає не-пептидні зв'язки.
5. Фармацевтична композиція за п. 1, яка містить щонайменше два пептиди, що складаються з амінокислотних послідовностей, відповідно до послідовностей SEQ ID NO: 1 − SEQ ID NO: 15.
6. Фармацевтична композиція за п. 1, у якій вибір, число та/або кількість пептидів, присутніх в композиції залежить/залежать від тканин, раку та/або пацієнтів.
7. Фармацевтична композиція за будь-яким з пп. 1-6, яка відрізняється тим, що додатково містить щонайменше один прийнятний ад'ювант, вибраний з групи, яка складається з таких, як: 1018 ISS, солі алюмінію, Amplivax, AS15, БЦЖ, СР-870,893, CpG7909, СуаА, dSLIM, GM-CSF, ІС30, ІС31, Іміквімод, ImuFact IMP321, IS Patch, ISCOMATRIX, JuvImmune, LipoVac, монофосфорильований ліпід А, Монтанід IMS 1312, Монтанід ISA 206, Монтанід ISA 50V, Монтанід ISA-51, OK-432, ОМ-174, OM-197-MP-EC, ONTAK, векторну систему PepTel, мікрочастинки PLG, резіквімод, SRL172, Віросоми та інші вірусоподібні частинки, YF-17D, стимулон Aquila's QS21, Ribi's Detox. Quil, Superfos, Фрейнда, холерний токсин, імунологічні ад'юванти, MF59 та цитокіни.
8. Фармацевтична композиція за п. 7, яка відрізняється тим, що ад'ювант являє собою гранулоцитарно-макрофагальний колонієстимулюючий фактор (GM-CSF).
9. Фармацевтична композиція за п. 1, яка відрізняється тим, що додатково містить щонайменше одну антиген-презентуючу клітину.
10. Фармацевтична композиція за п. 9, яка відрізняється тим, що антиген-презентуюча клітина являє собою дендритну клітину.
11. Фармацевтична композиція, за пп. 9 або 10, яка відрізняється тим, що щонайменше одна антиген-презентуюча клітина:
a) імпульсним шляхом наповнена або завантажена пептидом; або
б) містить компонент експресії, що кодує пептид.
12. Фармацевтична композиція за п. 1, яка призначена для внутрішньовенного введення, внутрішньоартеріального введення, внутрішньочеревного введення, внутрішньом'язового введення, підшкірного введення, введення в середину пухлини, перорального введення, дермального введення, назального, трансбукального введення, ректального введення, вагінального введення, введення шляхом інгаляції або місцевого застосування.
13. Застосування терапевтично ефективної кількості фармацевтичної композиції за п. 1 для отримання лікарського засобу для лікування або профілактики раку у пацієнта.
14. Застосування за пунктом 13, у якому фармацевтична композиція є протираковою вакциною.
15. Застосування за пунктом 14, у якому рак являє собою рак ротової порожнини і рак глотки, рак шлунково-кишкового тракту, рак товстої кишки, прямої кишки та анусу, рак дихальних шляхів, рак молочної залози, рак шийки матки, піхви та вульви, рак тіла матки та яєчників, рак статевих шляхів у чоловіків, рак сечовивідних шляхів, рак кістки та м'яких тканин, та саркому Капоші, меланому шкіри, меланому ока, рак ока немеланомного характеру, рак мозку та центральної нервової системи, рак щитовидної залози та інших ендокринних залоз, лімфому Ходжкіна, нехождкінську лімфому, мієлому, переважно рак нирки, колоректальний рак, рак легенів, рак підшлункової залози, рак передміхурової золози, рак шлунку, стромальні пухлини ШКТ або гліобластому.
16. Застосування, за пунктом 15, у якому рак являє собою колоректальний рак.
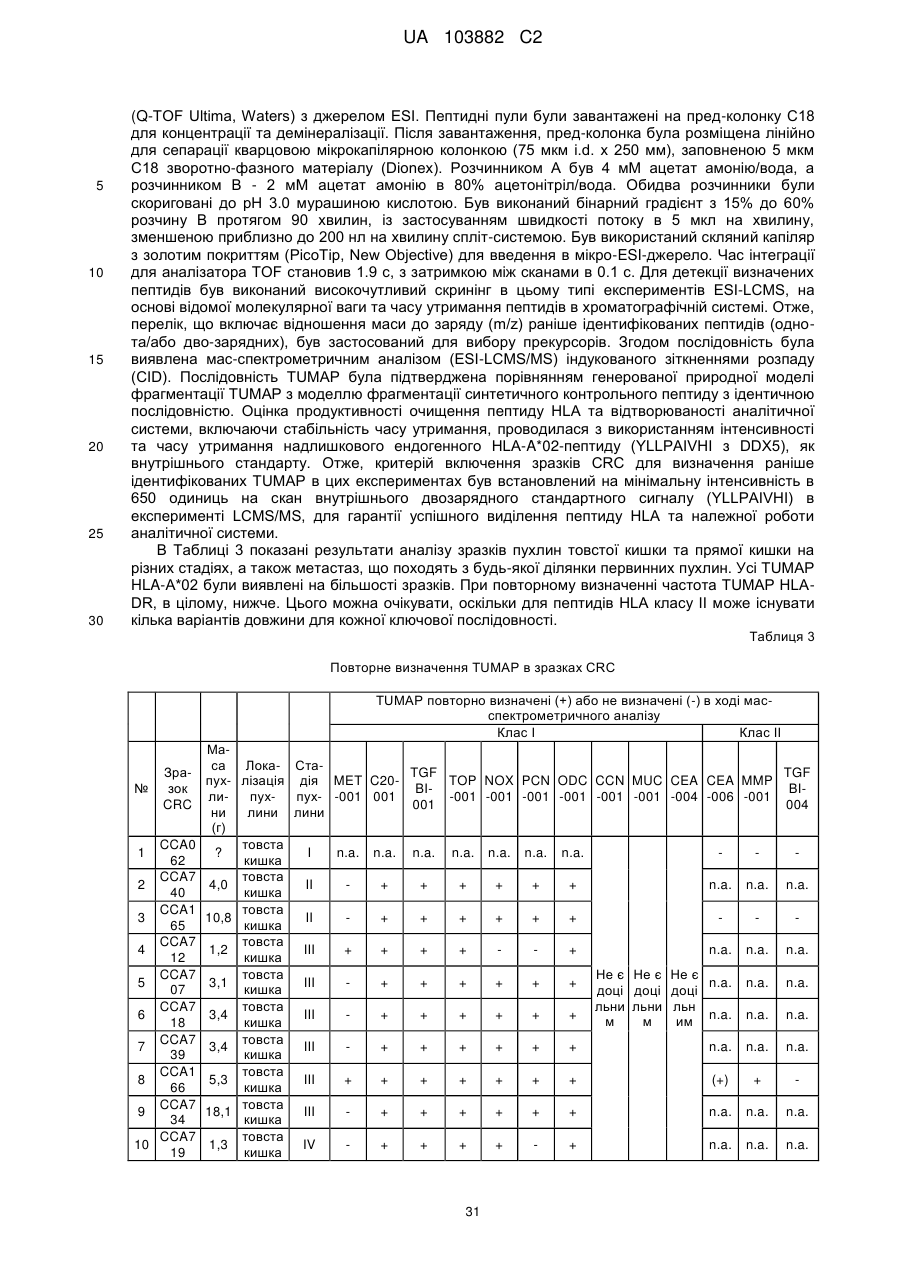
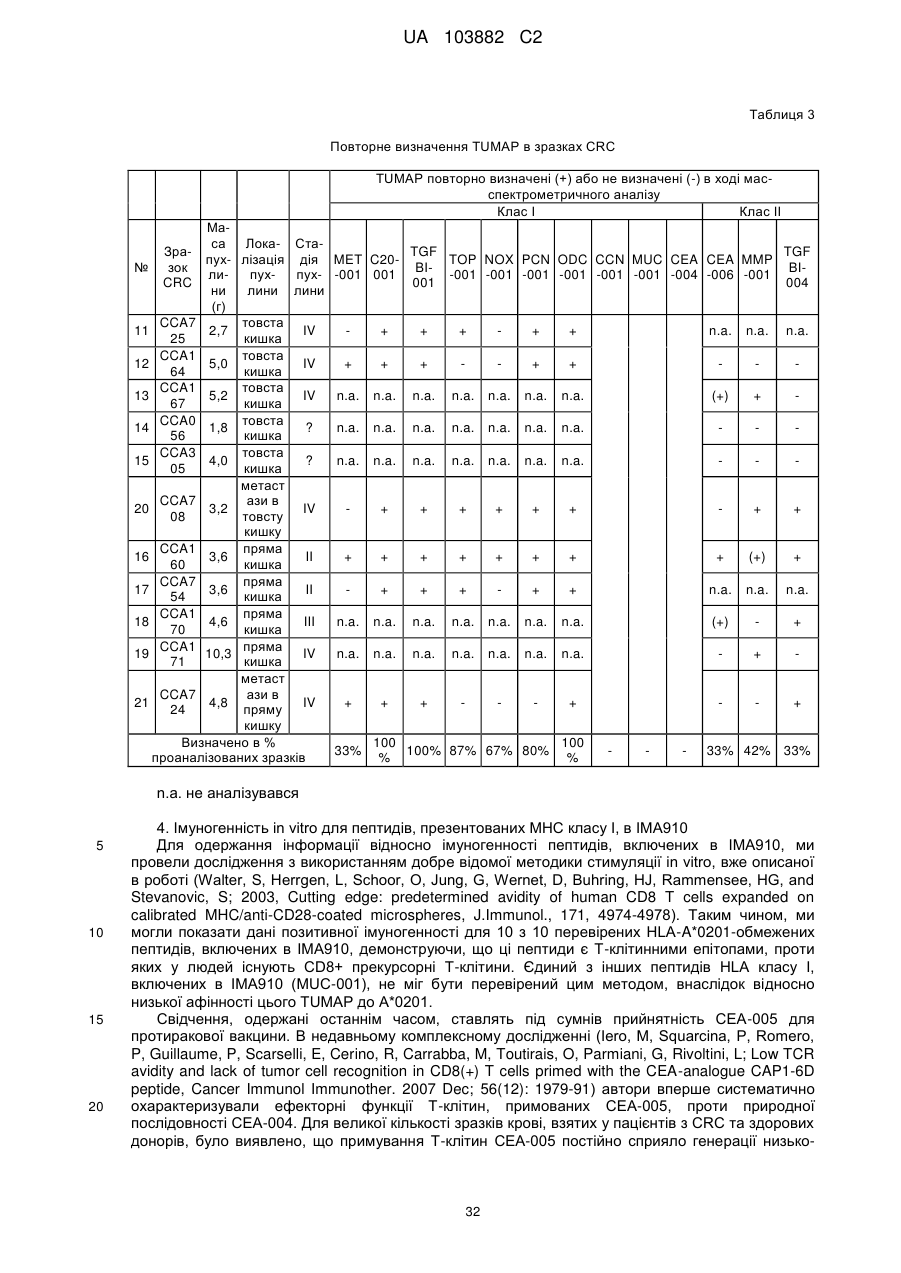
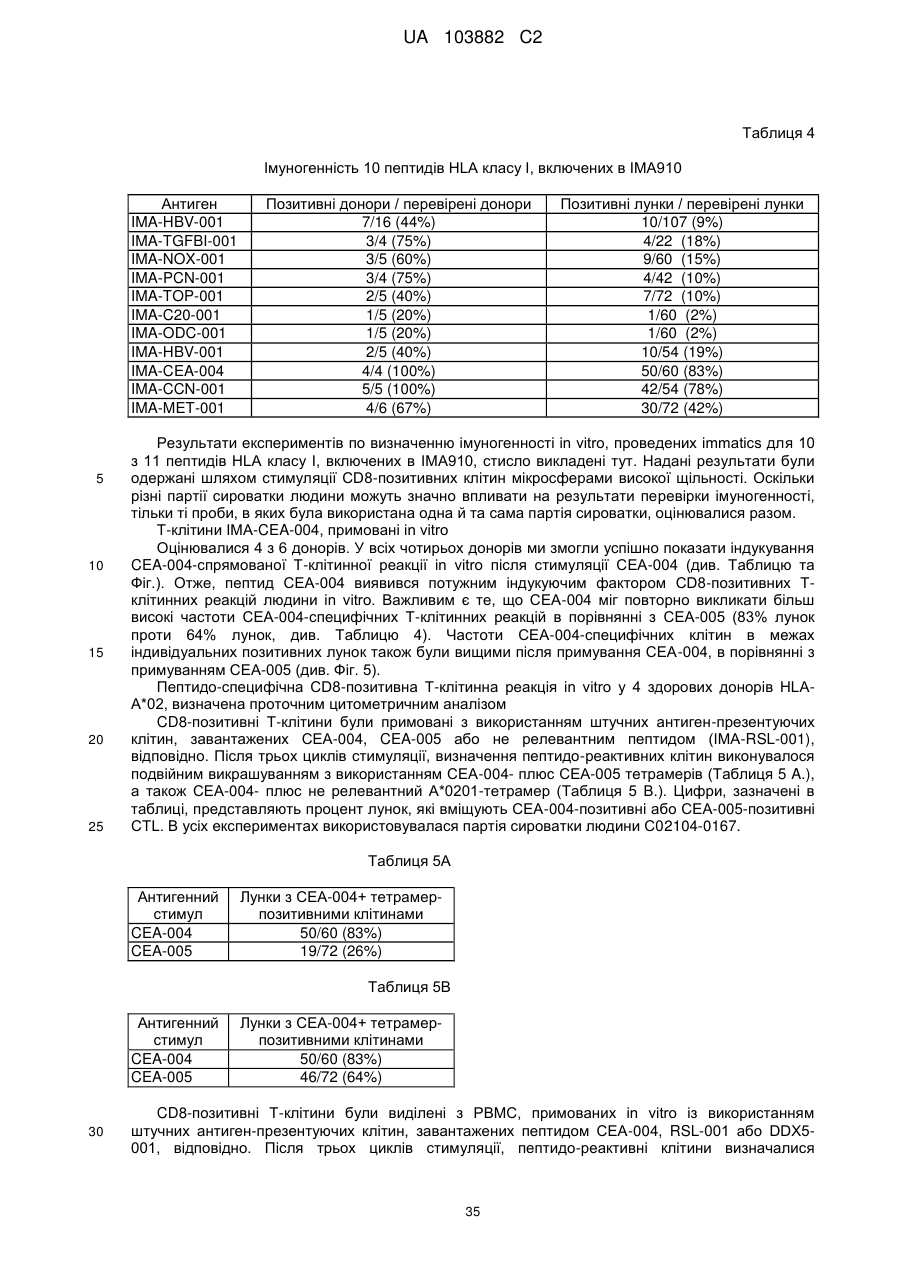
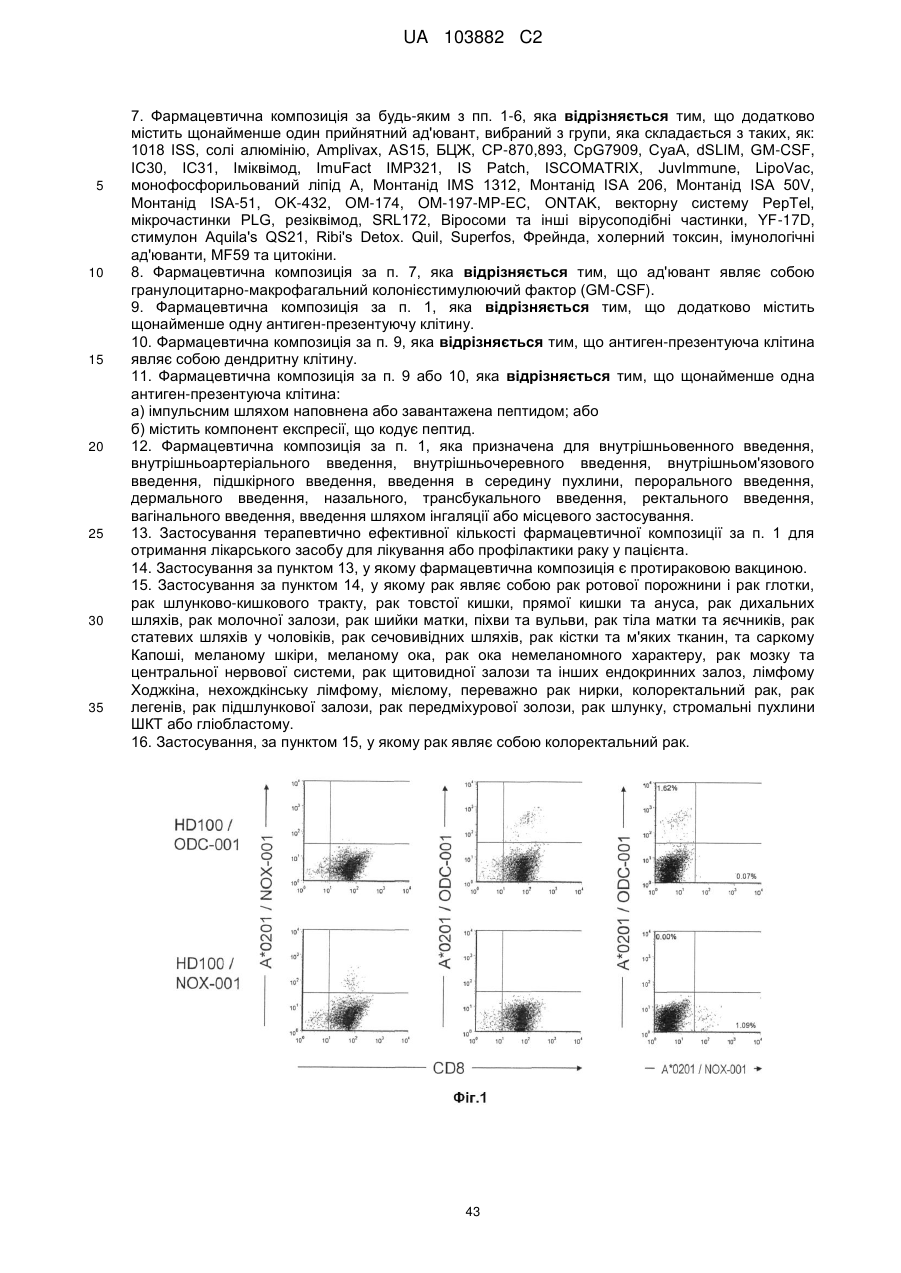
Текст
Реферат: Винахід належить до фармацевтичної композиції, яка містить щонайменше два пептиди, що включають амінокислотну послідовність SEQ ID NO: 1 і SEQ ID NO: 2 та/або включають амінокислотну послідовність, яка щонайменше на 80 % ідентична SEQ ID NO: 1 і SEQ ID NO: 2, та фармацевтично прийнятий носій, де пептиди мають здатність зв’язуватися з молекулою головного комплексу гістосумісності людини (МНС) класу І або ІІ класу, та зазначені пептиди мають загальну довжину від 8 до 100 амінокислот. UA 103882 C2 (12) UA 103882 C2 UA 103882 C2 5 10 15 20 25 30 35 40 45 50 55 60 Цей винахід відноситься до імунотерапевтичних пептидів та їхнього використання в імунотерапії, зокрема, в імунотерапії раку. В цьому винаході розкриваються епітопи пухлиноасоційованих пептидів T-клітин - хелперів, разом або в поєднанні з іншими пухлиноасоційованими пептидами, котрі служать активними фармацевтичними інгредієнтами композицій вакцин, стимулюючих протипухлинні імунні реакції. Зокрема, композиція пептидів цього винаходу може використовуватися в композиціях вакцин для викликання протипухлинних імунних реакцій проти колоректального раку. Для цілей даного винаходу, усі довідкові матеріали, котрі зазначені в ньому, включені шляхом посилань в усій своїй повноті. Рівень техніки Колоректальна карцинома Згідно із даними Американського Товариства по Боротьбі з Раком, колоректальний рак (CRC) є третім з найбільш розповсюджених видів раку в США, на який захворюють більш ніж 175 000 нових пацієнтів кожного року. В США, Японії, Франції, Німеччині, Італії, Іспанії та Великобританії він уражає більш ніж 480 000 людей. Колоректальний рак є однією з найчастіших причин смертності від раку в розвинених країнах. Дослідження показують ймовірність виникнення колоректального раку як результату взаємодії між успадкованими факторами та впливом зовнішнього середовища. В більшості випадків, попередниками колоректальних пухлин вважаються аденоматозні поліпи; однак, переродження може тривати багато років. Основним фактором ризику колоректального раку є вік, оскільки 90% випадків захворювання діагностуються у людей старше 50 років. Інші фактори ризику колоректального раку, за даними Американського Товариства по Боротьбі з Раком, включають вживання алкоголю, раціон харчування з великим вмістом жирів та/або червоного м’яса і недостатнім вживанням фруктів та овочів. Захворюваність продовжує зростати, зокрема, в таких регіонах, як Японія, де це може бути обумовлено переходом на західний стиль харчування, з надмірною кількістю жирів та м’яса в раціоні та зменшенням вживання харчових волокон. Однак, частота захворювань зростає не настільки швидко, як раніше, і це може бути обумовлено більш активним скринінгом та видаленням поліпів, що запобігає переродженню поліпів на рак. Як і у відношенні більшості солідних пухлин, терапією першого ряду є хірургічне втручання, однак, його переваги обмежуються ранніми стадіями захворювання, в той час як у значної частини пацієнтів діагностується запущена стадія. Стандартом лікування прогресуючого колоректального раку є режими хіміотерапії, які базуються на схемах з використанням фторурацила. Більшість таких режимів - це так звані схеми FOLFOX (інфузійне введення 5фторурацилу(5-FU)/лейковоріну плюс оксаліплатін) та FOLFIRI (ірінотекан, лейковорін, бюлюсне і тривале інфузійне введення 5-FU). Введення цитотоксичних препаратів третього покоління, таких як ірінотекан та оксаліплатін, збільшило надію на значне підвищення ефективності лікування, але прогноз все ще відносно несприятливий, та виживаність, в цілому, залишається на рівні приблизно 20 місяців при наявності метастаз, і, в результаті, незадоволені потреби стосовно лікування цієї хвороби поки що є високими. Останнім часом з’явилося нове покоління медикаментів, що діють цілеспрямовано на молекулярному рівні, таких як Avastin (бевакізумаб) та Erbitux (цетуксімаб), і близько 40 сполук знаходяться на завершальному етапі клінічної розробки для різних стадій колоректального раку. Комбінації декількох з цих сполук збільшують кількість потенційних варіантів терапії, які очікуються в майбутньому. Переважна більшість речовин є у фазі 2, при чому на EGFR (рецептори епідермального фактора росту) ці сполуки діють частіше, ніж будь-які інші ліки, розроблені для колоректального раку, і це обумовлено фактом, що приблизно у 80% хворих на колоректальний рак експресія EGFR підвищується. Зараз проводяться клінічні дослідження пацієнтів з II стадією захворювання, з поєднанням хіміотерапії та недавно ухвалених моноклональних антитіл (mAb) (цетуксімаб + ірінотекан або FOLFOX4; бевакізумаб як одиночний препарат або разом із FOLFOX4). Очікується, що період спостережень для одержання статистично значимих результатів цих досліджень становитиме від трьох до чотирьох років. Моноклональні антитіла (mAb), які на даний час використовуються в онкології, в цілому мають прекрасні шанси щодо їхнього використання без втручання в активну імунотерапію. Фактично, були одержані доклінічні дані, які свідчать про те, що зниження експресії VEGF (фактору росту ендотелія судин) (за допомогою бевакізумабу) позитивно впливає на активацію T-клітин, опосередковану DC (дендритними клітинами). Зараз проводяться приблизно 16 випробувань з метою визначення безпеки та 1 UA 103882 C2 5 10 15 20 25 30 35 40 45 50 55 60 потенціальних можливостей нових імунотерапевтичних підходів до лікування CRC. Імунотерапевтичні підходи до лікування Стимуляція імунної реакції залежить від наявності антигенів, які визнаються імунною системою хазяїна як чужорідні. Виявлення існування пухлино-асоційованих антигенів дало можливість використовувати імунну систему хазяїна для впливу на зріст пухлин. Зараз досліджуються різні механізми використання як гуморальних, так і клітинних важелів імунної системи для імунотерапії раку. Окремі елементи клітинної імунної реакції здатні специфічно розпізнавати та знищувати клітини пухлин. Виділення цитотоксичних T-лімфоцитів (CTL) з пухлино-інфільтруючих клітинних популяцій чи з периферичної крові наводить на думку, що такі клітини грають важливу роль в природному імунному захисті проти раку (Cheever et al., Annals N.Y. Acad. Sci. 1993 690:101-112; Zeh HJ, Perry-Lalley D, Dudley ME, Rosenberg SA, Yang JC; J Immunol. 1999, 162(2):989-94; High avidity CTLs for two self-antigens demonstrate superior in vitro and in vivo antitumor efficacy.). + Зокрема, CD8-позитивні T-клітини (TCD8 ), які розпізнають молекули Класу I головного комплексу гістосумісності (MHC), що презентують пептиди зазвичай з 8-10 залишків, одержані з білків чи дефектних продуктів рибосом (DRIPS) (Schubert U, Antón LC, Gibbs J, Norbury CC, Yewdell JW, Bennink JR.; Rapid degradation of a large fraction of newly synthesized proteins by proteasomes; Nature 2000; 404(6779):770-774), які розташовані у цитозолях, відіграють важливу роль в цій реакції. MHC-молекули людини також визначаються як лейкоцитарні антигени людини (HLA). Є два класи MHC-молекул: MHC-молекули класу I, котрі можна знайти на більшості клітин, що мають ядро; вони презентують пептиди, які є результатом протеолітичного розщеплення ендогенних протеїнів DRIPS та більшіх за розміром пептидів. MHC-молекули класу II, переважно, знаходяться на професійних антиген-презентуючих клітинах (APC); вони презентують пептиди екзогенних протеїнів, що поглинаються АPC в ході ендоцитозу, та проходять подальшу обробку. Комплекси пептиду та MHC-молекули класу I розпізнаються CD8позитивними цитотоксичними T-лімфоцитами, які презентують відповідні рецептори Т-клітин (TCR), а комплекси пептиду і MHC-молекули класу II розпізнаються CD4-позитивними Тклітинами- хелперами, що презентують відповідні TCR. CD4-позитивні T-клітини-хелпери відіграють важливу роль в регулюванні ефекторних функцій протипухлинних реакцій T-клітин, і з цієї причини ідентифікація епітопів CD4-позитивних T-клітин, які одержуються з пухлино-асоційованих антигенів (TAA) може мати велике значення для розробки фармацевтичних продуктів для викликання протипухлинних імунних реакцій (Kobayashi H., R. Omiya, M. Ruiz, E. Huarte, P. Sarobe, J. J. Lasarte, M. Herraiz, B. Sangro, J. Prieto, F. Borras-Cuesta, and E. Celis. Identification of an antigenic epitope for helper T lymphocytes from carcinoembryonic antigen. Clin. Cancer Res. 2002, 8:3219-3225., Gnjatic, S., D. Atanackovic, E. Jäger, M. Matsuo, A. Selvakumar, N.K. Altorki, R.G. Maki, B. Dupont, G. Ritter, Y.T. Chen, A. Knuth, and L.J. Old. Survey of naturally occurring CD4+ T-cell responses against NY-ESO-1 in cancer patients: Correlation with antibody responses. Proc. Natl. Acad. Sci. U.S.A. 2003, 100 (15): 8862-7). CD4-позитивні T-клітини можуть призвести до локального зростання рівнів гамма-інтерферону (Qin Z, Schwartzkopff J, Pradera F, Kammertoens T, Seliger B, Pircher H, Blankenstein T; A critical requirement of interferon gamma-mediated angiostasis for tumor rejection by CD8+ T cells; J Cancer Res; 2003, 63(14): 4095-4100). На моделях ссавців, наприклад, мишей, було продемонстровано, що навіть за відсутністю цитотоксичних Т-лімфоцитарних (CTL) ефекторних клітин (тобто, CD8-позитивних Tлімфоцитів), CD4-позитивні T-клітини є достатніми для візуалізації інгібування пухлин шляхом інгібування ангіогенезу завдяки секреції гамма-інтерферону (Qin, Z. and T. Blankenstein. CD4+ Tcell-mediated tumour rejection involves inhibition of angiogenesis that is dependent on IFN gamma receptor expression by nonhematopoietic cells. Immunity. 2000, 12:677-686). Крім того, було показано, що CD4-позитивні T-клітини, які розпізнають пептиди з пухлино-асоційованих антигенів, представлених молекулами HLA класу II, можуть протидіяти розвитку пухлин за допомогою індукування реакцій антитіл (Ab) (Kennedy, R.C., M.H. Shearer, A.M. Watts, and R.K. Bright. CD4+ T lymphocytes play a critical role in antibody production and tumour immunity against simian virus 40 large tumour antigen. Cancer Res. 2003, 63:1040-1045). На відміну від пухлиноасоційованих пептидів, які зв’язуються з молекулами HLA класу I, лише невелика кількість лігандів класу II TAA була описана на цей час (www.cancerimmunity.org, www.syfpeithi.de). Оскільки визначальна експресія молекул HLA класу II зазвичай обмежується клітинами імунної системи (Mach, B., V. Steimle, E. Martinez-Soria, and W. Reith. Regulation of MHC class II genes: lessons from a disease. Annu. Rev. Immunol. 1996, 14: 301-331), можливість виділення пептидів класу II безпосередньо з первинних пухлин не вважалася ймовірною. Однак, автори 2 UA 103882 C2 5 10 15 20 25 30 35 40 45 50 55 60 винаходів за останній час досягли успіху в ідентифікації ряду епітопів MHC класу II безпосередньо з пухлин (EP 1642905, EP 1760088; Dengjel J, Nastke MD, Gouttefangeas C, Gitsioudis G, Schoor O, Altenberend F, Müller M, Krämer B, Missiou A, Sauter M, Hennenlotter J, Wernet D, Stenzl A, Rammensee HG, Klingel K, Stevanović S.; Unexpected abundance of HLA class II presented peptides in primary renal cell carcinomas; Clin Cancer Res. 2006; 12:4163-4170). За відсутності запалення, експресія молекул MHC класу II, переважно, зводиться до клітин імунної системи, зокрема, до професійних антиген-презентуючих клітин (APC), наприклад, моноцитів, клітин, що походять з моноцитів, макрофагів, дендритних клітин. При дослідженні пацієнтів з пухлинами було несподівано виявлено, що клітини пухлини експресують MHCмолекули класу II (Dengjel J, Nastke MD, Gouttefangeas C, Gitsioudis G, Schoor O, Altenberend F, Müller M, Krämer B, Missiou A, Sauter M, Hennenlotter J, Wernet D, Stenzl A, Rammensee HG, Klingel K, Stevanović S.; Unexpected abundance of HLA class II presented peptides in primary renal cell carcinomas; Clin Cancer Res. 2006; 12:4163-4170). Для того, щоб пептид індукував (викликав) клітинну імунну реакцію, він повинен зв’язуватися з MHC-молекулою. Цей процес залежить від алелі MHC-молекули та специфічного поліморфізму амінокислотної послідовності пептиду. Пептиди, зв’язані з MHC класу I, як правило, мають довжину 8-10 амінокислотних залишків та зазвичай вміщують два консервативні залишки («якір») в своїй послідовності, що взаємодіє з відповідною порожниною зв’язування MHC-молекули. Таким чином, кожна MHC-алель має «мотив зв’язку», що визначає, які пептиди можуть специфічно зв’язуватися з порожниною зв’язування (Rammensee HG, Bachmann J, Stevanovic S. MHC ligands and peptide motifs, Landes Bioscience, USA, 1997). В імунній реакції, залежній від MHC класу I, пептиди повинні не тільки мати здатність зв’язуватися з певними молекулами MHC класу I, експресованими пухлинними клітинами, вони також мають розпізнаватися T-клітинами, презентуючими специфічні T-клітинні рецептори (TCR). Антигени, котрі розпізнаються пухлино-специфічними T-лімфоцитами, тобто, їхні епітопи, можуть бути молекулами, одержаними з усіх класів білків, таких як ферменти, рецептори, транскрипційні фактори та ін. Крім того, пухлино-асоційовані антигени, наприклад, також можуть бути присутні тільки в клітинах пухлин, зокрема, як продукти генів, що мутували. Іншим важливим класом пухлино-асоційованих антигенів є тканино-специфічні антигени, такі як CT ("cancer testis")-антигени, що експресуються в різних видах пухлин та в здоровій тканині сем’яників. Було ідентифіковано різні пухлино-асоційовані антигени. Крім того, проведена значна дослідницька робота по ідентифікації додаткових пухлино-асоційованих антигенів. Деякі групи пухлино-асоційованих антигенів, котрі також згадуються в літературі як пухлино-специфічні антигени, є тканино-специфічними. Приклади включають, без обмежень, тирозиназу для меланоми, простато-специфічні антигени (PSA) та простатичні специфічні мембранні антигени (PSMA) для раку простати і хромосомні кросовери, такі як bcr/abl у лімфомі. Однак, ряд ідентифікованих пухлино-асоційованих антигенів зустрічається в багатьох видах пухлин, а деякі з них, такі як онкогенні білки та/або гени-супресори пухлин (огляд генів-супресорів пухлин для раку нирки наведений, наприклад, в роботі Linehan WM, Walther MM, Zbar B. The genetic basis of cancer of the kidney. J Urol. 2003 Dec; 170 (6Pt1):2163-72), котрі фактично викликають трансформаційну дію, зустрічаються практично в усіх видах пухлин. Наприклад, звичайні білки клітин, що контролюють ріст та диференціацію клітин, такі як p53 (приклад гена-супресора пухлин), ras, c-met, myc, pRB, VHL та HER-2/neu, можуть накопичувати мутації, і це призводить до посилення експресії цих генних продуктів, таким чином роблячи їх онкогенними (McCartey et al. Cancer Research, 1998, 15:58 2601-5; Disis et al. Ciba Found. Symp. 1994, 187:198-211). Ці білки-мутанти також можуть бути мішенню пухлино-специфічної імунної реакції при багатьох видах ракових захворювань. Імунотерапія у пацієнтів, хворих на рак, спрямована на активацію клітин імунної системи специфічним чином, зокрема так званих цитотоксичних T-клітин (CTL, також відомих як «клітини-кілери» і як CD8-позитивні T-клітини), проти пухлинних клітин, а не проти здорових тканин. Клітини пухлинвідрізняються від здорових клітин експресією пухлино-асоційованих білків. Молекули HLA на клітинній поверхні презентують клітинний вміст назовні, таким чином дозволяючи цитотоксичній T-клітині проводити диференціацію між здоровою та пухлинною клітиною. Це здійснюється за допомогою розбивки усіх білків всередині клітини на короткі пептиди, котрі потім додаються до молекул HLA і презентуються на клітинній поверхні (Rammensee, HG, Falk, K, and Rotzschke, O; Peptides naturally presented by MHC class I molecules, Annu. Rev. Immunol., 1993, 11, 213-244). Пептиди, які презентуються на пухлинних клітинах, але не презентуються (або в незначній мірі презентуються) на здорових клітинах тіла, 3 UA 103882 C2 5 10 15 20 25 30 35 40 45 50 55 60 називаються пухлино-асоційованими пептидами (TUMAP). Для того, щоб білки розпізнавалися цитотоксичними T-лімфоцитами як пухлино-специфічні або пухлино-асоційовані антигени, та з метою використання в лікуванні, мають бути дотримані деякі попередні умови. Антиген повинен експресуватися, головним чином, клітинами пухлин, а не нормальними здоровими тканинами, чи бути присутнім в здорових клітинах в порівняно невеликих кількостях. Крім того, бажано, щоб відповідний антиген був не тільки присутнім в будь-якому виді пухлин, але також знаходився у високих концентраціях (наприклад, кількість копій відповідного пептиду на клітину). Пухлино-специфічні та пухлино-асоційовані антигени часто походять з білків, що беруть безпосередню участь у трансформації нормальної клітини в пухлинну клітину внаслідок їхньої функції, наприклад, в контролі клітинного циклу чи апоптозі. До того ж, наступні мішені цих білків, які є основною причиною трансформації, можуть мати підвищену експресію і, відповідно, бути непрямим чином асоційованими з пухлинами. Такі непрямо пухлино-асоційовані антигени також можуть бути мішенями у вакцинаційному підході. Істотно важливою в обох випадках є присутність епітопів в амінокислотній послідовності антигену, оскільки такий пептид («імуногенний пептид»), що одержується з пухлиноасоційованого антигену, повинен призводити до Т-клітинної реакції in vitro чи in vivo. По суті, будь-який пептид, здатний зв’язувати молекулу MHC, може функціонувати як Tклітинний епітоп. Передумовою для індукування Т-клітинної реакції in vitro чи in vivo є наявність T-клітини з відповідним рецептором TCR та відсутність толерантності до цього конкретного епітопу. T-клітини-хелпери відіграють важливу роль в регуляції ефекторної функції CTL в протипухлинному імунітеті. Епітопи Т-хелперів, які запускають реакцію цих клітин типу TH1, підтримують ефекторні функції CD8-позитивних T-клітин - кілерів, котрі включають цитотоксичні функції, спрямовані проти клітин пухлин, що експресують комплекси пухлино-асоційованого пептиду/MHC на своїх клітинних поверхнях. Таким чином, епітопи пухлино-асоційованих пептидів Т-хелперів, самостійно чи в комбінації з іншими пухлино-асоційованими пептидами, можуть служити активними фармацевтичними інгредієнтами композицій вакцин, які стимулюють протипухлинні імунні реакції. Оскільки обидва типи реакцій, залежні від CD8 та CD4, спільно та синергічно роблять свій внесок у протипухлинну дію, для розробки протипухлинних вакцин важливими є ідентифікація та характеристика пухлино-асоційованих антигенів, які розпізнаються CD8-позитивними CTL (молекули MHC класу I) чи CD4- позитивними CTL (молекули MHC класу II). Отже, задача цього винаходу полягає в знаходженні композицій пептидів, які вміщують пептиди, що зв’язуються з MHC-комплексами будь-якого класу. Перші клінічні дослідження з використанням пухлино-асоційованих пептидів розпочали в середині 90-х років минулого століття Бун (Boon) та його колеги, переважно, для індікації меланоми. Клінічні реакції в найбільш вдалих з цих досліджень варіювалися від 10% до 30%. Тяжкі побічні ефекти чи серйозна аутоімунність не повідомлялися в будь-яких клінічних дослідженнях із використанням монотерапії вакциною на основі пептидів. Є дані про вітіліго у слабкій формі, що спостерігалася у деяких пацієнтів, які проходили лікування меланомоасоційованими пептидами. Однак, примування одного виду CTL зазвичай є недостатнім для знищення усіх пухлинних клітин. Пухлини дуже мутагенні і тому здатні швидко реагувати на атаки CTL, змінюючи свою білкову модель для уникнення розпізнавання цитотоксичними Т-лімфоцитами. З метою протидії пухлинним механізмам уникнення, для вакцинації використовується широкий діапазон специфічних пептидів. Таким способом, проти клітини може бути розпочата широка одночасна атака відразу кількома CTL-клонами. Це може зменшити шанси того, що пухлина уникатиме імунної реакції. Така гіпотеза була недавно підтверджена в клінічному дослідженні лікування пацієнтів, хворих на меланому в останній стадії. Лише з кількома винятками, пацієнти, що мали якнайменш 3 визначені T-клітинні реакції, продемонстрували об'єктивні клінічні реакції чи стабілізацію захворювання (Banchereau, J, Palucka, AK, Dhodapkar, M, Burkeholder, S, Taquet, N, Rolland, A, Taquet, S, Coquery, S, Wittkowski, KM, Bhardwaj, N, Pineiro, L, Steinman, R, and Fay, J; Immune and clinical responses in patients with metastatic melanoma to CD34(+) progenitor-derived dendritic cell vaccine, Cancer Res., 2001, 61, 6451-6458), а також збільшену виживаність (інформація одержана при особистому спілкуванні з J. Banchereau), в той час як у широкої більшості пацієнтів з менш ніж трьома T-клітинними реакціями було діагностовано прогресуюче захворювання. Вивчення робіт заявників на патенти показало подібний ефект, коли пацієнти, хворі на гіпернефрому, проходили лікування вакциною, що складалася з 13 різних пептидів (H. SinghJasuja, S. Walter, T. Weinschenk, A. Mayer, P. Y. Dietrich, M. Staehler, A. Stenzl, S. Stevanovic, H. Rammensee, J. Frisch; Correlation of T-cell response, clinical activity and regulatory T-cell levels in 4 UA 103882 C2 5 10 15 20 25 30 35 40 45 50 55 60 renal cell carcinoma patients treated with IMA901, a novel multi-peptide vaccine; ASCO Meeting 2007 Poster # 3017; M. Staehler, A. Stenzl, P. Y. Dietrich, T. Eisen, A. Haferkamp, J. Beck, A. Mayer, S. Walter, H. Singh, J. Frisch, C. G. Stief; An open label study to evaluate the safety and immunogenicity of the peptide based cancer vaccine IMA901, ASCO meeting 2007; Poster # 3017). Основна задача розробки протипухлинної вакцини полягає, таким чином, не тільки в ідентифікації та характеристиці нових пухлино-асоційованих антигенів та епітопів імуногенних Тхелперів, які походять з них, але й в комбінуванні різних епітопів для збільшення ймовірності реакції більш, ніж на один епітоп для кожного пацієнта. Отже, задача цього винаходу полягає в знаходженні комбінацій амінокислотних послідовностей таких пептидів, що мають здатність зв’язуватися з молекулою головного комплексу гістосумісності людини (MHC) класу I (HLA класу I) чи II (HLA класу II). Додаткова задача цього винаходу полягає в наданні ефективної протиракової вакцини, яка базується на комбінації пептидів. Автори цього винаходу виділили і характеризували пептиди, які зв’язуються з молекулами HLA класу I або II, безпосередньо з пухлин, тобто, колоректальних карцином, ссавців. Цей винахід пропонує пептиди, що походять з антигенів, які асоціюються з пухлиногенезом, та мають здатність зв’язуватися в достатній мірі з молекулами MHC (HLA) класу II для запуску імунної реакції лейкоцитів людини, зокрема, лімфоцитів, в особливості, T-лімфоцитів, головним чином, CD4-позитивних T-лімфоцитів, а конкретніше, CD4-позитивних T-лімфоцитів, що виступають посередниками в імунних реакціях T H1-типу. Цей винахід також пропонує пептиди, що походять з антигенів, які асоціюються з пухлиногенезом, та мають здатність зв’язуватися в достатній мірі з молекулами MHC (HLA) класу I для запуску імунної реакції лейкоцитів людини, зокрема, лімфоцитів, в особливості, Tлімфоцитів, головним чином, CD8-позитивних цитотоксичних T-лімфоцитів, а також комбінацій з двох варіантів, що є особливо прийнятними для вакцинації пацієнтів, хворих на рак. Відповідно до цього винаходу, задача вирішується шляхом надання фармацевтичної композиції, котра складається принаймні з двох пептидів, що вміщують амінокислотну послідовність, обрану з групи, яка включає послідовності від SEQ ID NO 1 до SEQ ID NO 7, та/або вміщує варіант амінокислотної послідовності, що є принаймні на 80 % гомологічним послідовності SEQ ID NO 1 - SEQ ID NO 7, та/або полінуклеотид, який вміщує нуклеїнову кислоту, що кодує послідовності SEQ ID NO 1 - SEQ ID NO 7 або варіант амінокислотної послідовності, та фармацевтично прийнятний носій. Фармацевтичні композиції цього винаходу можуть також вміщувати якнайменш один додатковий пептид, який вміщує амінокислотну послідовність, обрану з групи, яка включає послідовності від SEQ ID NO: 8 до SEQ ID NO: 15, або включає варіант амінокислотної послідовності, що є принаймні на 80 % ідентичний послідовності SEQ ID NO: 8 - SEQ ID NO: 15, чи полінуклеотид, який вміщує нуклеїнову кислоту, що кодує послідовності SEQ ID NO: 8 - SEQ ID NO: 15 або варіант амінокислотної послідовності. Пептиди можуть мати загальну довжину від 8 до 100 амінокислот, переважно від 8 до 30 амінокислот, та найбільш переважно від 8 до 16 амінокислот. Пептиди також можуть мати непептидні зв’язки. Як описується нижче, всі пептиди, що є основою цього винаходу, були ідентифіковані як презентовані клітинами, що несуть в собі MHC класу I чи II. Отже, усі ці конкретні пептиди, а також інші пептиди, які включають послідовність (тобто, похідні пептиди) викликають специфічну T-клітинну реакцію, хоча ступінь, в якій ця реакція буде індукуватися, може варіюватися взалежності від конкретного пептиду та пацієнта. Різниця, наприклад, може бути викликана мутаціями в пептидах. Фахівцю, який має досвід в цій галузі, добре відомі методи, що можуть бути застосовані для визначення ступіню індукування реакції окремим пептидом, зокрема, із посиланням на приклади, наведені в цьому винаході, та відповідну довідкову літературу. Переважно, варіанти винаходу будуть індукувати T-клітини, що перехресно реагують з відповідним пептидом винаходу. Процент гомології між амінокислотною послідовністю пептиду чи послідовністю нуклеїнових кислот, що кодує пептид та варіант, може бути розрахований з використанням алгоритмів, добре відомих в цій галузі. Термін «гомологічний» в цьому винаході відноситься до ступеню ідентичності між послідовностями двох амінокислотних послідовностей, тобто, пептидними або поліпептидними послідовностями. Вищеназвана «гомологія» визначається порівнянням двох послідовностей, доведених в оптимальних умовах до порівнюваних послідовностей. Амінокислотні чи нуклеїновокислотні послідовності, які порівнюються в цьому винаході, можуть мати доповнення чи делеції (розриви і т.п.) в оптимальному ряду двох послідовностей. Гомологія таких послідовностей може бути розрахована шляхом створення порівняльного ряду із використанням, наприклад, алгоритму ClustalW (Nucleic Acid Res., 22(22): 4673 4680 (1994). 5 UA 103882 C2 5 10 15 20 Можуть також використовуватись наявні комп’ютерні програми для аналізу послідовностей, зокрема, Vector NTI, GENETYX чи засоби аналізу в загальнодоступних базах даних, таких як http://restools.sdsc.edu/biotools/biotools16.html. Фармацевтично прийнятні носії добре відомі; це, зазвичай, рідини, в яких формулюється активний терапевтичний препарат. Носій взагалі не додає формуляції ніякої фармакологічної функції, хоча він може забезпечити хімічну та/або біологічну стабільність, характеристики вивільнення, і таке інше. Зразки формуляцій можна знайти, наприклад, в роботі Alfonso R. Gennaro. Remington: The Science and Practice of Pharmacy, 20th Edition. Baltimore, MD: Lippincott Williams & Wilkins, 2000; вони включають розчини солей, воду, буферну воду, 0.3%-гліцин, гіалуронову кислоту, декстрозу і тому подібне, але не обмежуються ними. Останнім часом було виявлено, що певні жирові емульсії, які багато років використовувалися для парентерального харчування хворих людей, також можуть бути носіями для пептидів. Двома прикладами таких емульсій є наявні у продажу жирові емульсії, відомі під назвами Інтраліпід та Ліпофундін. «Інтраліпід» - це зареєстрована торгова марка жирової емульсії для парентерального харчування фірми Kabi Pharmacia, Швеція, яка описана в Патенті США № 3,169,094. «Ліпофундін» - це зареєстрована торгова марка B. Braun Melsungen, Німеччина. Обидві емульсії вміщують соєве масло, як жир (100 чи 200 г в 1000 мл дистильованої води: 10% чи 20%, відповідно). Фосфоліпіди яєчного жовтка використовуються як емульгатори в Інтраліпіді (12 г/л дистильованої води), а яєчний лецитин - в Ліпофундіні (12 г/л дистильованої води). Ізотонічність є результатом додання гліцерину (25 г/л), як в Інтраліпіді, так і в Ліпофундіні. Пептиди походять з пухлино-асоційованих антигенів, зокрема, пухлино-асоційованих антигенів з функціями, наприклад, в протеолізі, ангіогенезі, рості клітин, регуляції клітинного циклу, діленні клітин, регуляції транскрипції, регуляції трансляції, інвазії тканин і т. ін. В Таблиці 1 наведені пептиди та функція білку, з якого походять пептиди. 25 Таблиця 1 Пептиди цього винаходу і функція вихідного білку Ід. № послідовності 1 2 3 4 5 6 7 30 35 40 Ід. номер пептиду Послідовність Символ гена Функція Бере участь у приєднанні актиноC20orf42 вого цитоскелету до ECM (екстрацелюлярного матриксу) NOX-001 ILAPVILYI NOX1 НАДФ-оксидаза ODC-001 ILDQKINEV ODC1 Орнітин декарбоксилаза Дельта-ДНК-полімераза PCN-001 KLMDLDVEQL PCNA допоміжний білок Трансформуючий фактор росту, TGFBI-001 ALFVRLLALA TGFBI бета-індукований TOP2A/TO TOP-001 KIFDEILVNA Топоізомераза P2B TPPIDAHTRNLL Трансформуючий фактор росту, TGFBI-004 TGFBI RNH бета-індукований C20-001 ALSNLEVTL Зв’язується з MHC HLA-A*02 HLA-A*02 HLA-A*02 HLA-A*02 HLA-A*02 HLA-A*02 HLA-DR Відкрита рамка зчитування 42 20-ї хромосоми C20orf42 - це білок фокальної адгезії, який бере участь в приєднанні актинового цитоскелету до цитоплазматичної мембрани та в клітинних процесах, де посередником виступає інтегрин. Нестача C20orf42 в результаті мутацій з втратою функції викликає синдром Кіндлера, аутосомно-рецесивній генодерматоз, що характеризується утворенням пухирів на шкірі, прогресуючою атрофією шкіри, світлочутливістю та, в деяких випадках, канцерогенезом (Herz, C, Aumailley, M, Schulte, C, Schlotzer-Schrehardt, U, Bruckner-Tuderman, L, and Has, C; Kindlin-1 is a phosphoprotein involved in regulation of polarity, proliferation, and motility of epidermal keratinocytes, J Biol Chem., 2006, 281, 36082-36090). Нещодавно з’явилася інформація про серйозне ураження шлунково-кишкового тракту з геморагічним колітом у пацієнта, в якого спостерігалася мутація з втратою функції (Sadler, E, Klausegger, A, Muss, W, Deinsberger, U, Pohla-Gubo, G, Laimer, M, Lanschuetzer, C, Bauer, JW, and Hintner, H; Novel KIND1 gene mutation in Kindler syndrome with severe gastrointestinal tract involvement, Arch. Dermatol., 2006, 142, 16191624). 6 UA 103882 C2 5 10 15 20 25 30 35 40 45 50 55 60 Що стосується раку, то в дослідженнях по вивченню генної експресії C20orf42 був описаний серед послідовностей, безпосередньо пов’язаних з раком. Було виявлено, що він надмірно експресується в 70% колоректальних карцином та в 60% карцином легенів, які були досліджені (n = 10). Експресія в нормальних тканинах, виявлена за допомогою Норзерн-блотингу, зводилася до нейром’язових тканин (Weinstein, EJ, Bourner, M, Head, R, Zakeri, H, Bauer, C, and Mazzarella, R; URP1: a member of a novel family of PH and FERM domain-containing membraneassociated proteins is significantly over-expressed in lung and colon carcinomas, Biochim. Biophys. Acta, 2003, 1637, 207-216). Крім того, C20orf42 був ідентифікований як ген, що бере участь в TGF--опосередкованій міграції клітин та інвазії пухлин (Kloeker, S, Major, MB, Calderwood, DA, Ginsberg, MH, Jones, DA, and Beckerle, MC; The Kindler syndrome protein is regulated by transforming growth factor-beta and involved in integrin-mediated adhesion, J. Biol. Chem., 2004, 279, 6824-6833). Гомолог-1 НАДФ(нікотинамід-аденін-динуклеотидфосфат)-оксидази (NOX1) NOX1 являє собою фермент, чутливий до фактору росту, який каталізує формування активних форм кисню - супероксида (O2-) та пероксида водню (H2O2). Його експресія була первісно виявлена в товстій кишці, простаті, матці та в проліферуючих клітинах судин гладких м’язів (Suh, Y. A. et al. Cell transformation by the superoxide-generating oxidase Mox1. Nature 1999, 401, 79-82). Експресія цього ферменту пов’язана з рядом біологічних реакцій, включаючи клітинну проліферацію, ангіогенез та активацію клітинних шляхів передачі сигналів (Harper, R. W., Xu, C., Soucek, K., Setiadi, H. and Eiserich, J. P. A reappraisal of the genomic organization of human Nox1 and its splice variants. Arch. Biochem. Biophys. 2005, 435, 323-330). NOX1 в значній мірі експресується в товстій кишці, але його функція у фізіології або патології товстої кишки поки що недостатньо вивчена. В нормальних тканинах, експресія NOX1 виявилась низкою у клубовій кишці, середньою в висхідній ободовій кишці та високою у низхідній ободовій кишці. Не було продемонстровано статистичної розбіжності в експресії NOX1 між зразками, одержаними з аденом та високо- чи низько-диференційованих аденокарцином товстої кишки. NOX1 був експресований в значній мірі в тканинах епітелію товстої кишки, як у складках, так і на поверхні порожнини. На завершення треба відзначити, що NOX1 є ферментом, який конститутивно експресується в епітелії товстої кишки і прямо не асоціюється з пухлиногенезом (Szanto, I. et al. Expression of NOX1, a superoxide-generating NADPH oxidase, in colon cancer and inflammatory bowel disease. J Pathol. 2005, 207, 164-176). Дослідження в галузі імуногістохімії показали, що NOX1 конститутивно експресувався в поверхневих мукоїдних клітинах. Аденоми та високо-диференційовані аденокарциноми підвищують експресію NOX1. Ядерний фактор каппа-В (NF-kappaB) був переважно активований в клітинах аденоми та аденокарциноми, експресуючих NOX1 в надлишку, і це наводить на думку, що NOX1 може стимулювати NF-kappaB-залежні анти-апоптотичні шляхи в пухлинах товстої кишки (Fukuyama, M. et al. Overexpression of a novel superoxide-producing enzyme, NADPH oxidase 1, in adenoma and well differentiated adenocarcinoma of the human colon. Cancer Lett. 2005, 221, 97-104). Описано, що Wnt3a/бета-катеніновий шлях передачі сигналів індукує експресію NOX1 (Petropoulos, H. & Skerjanc, I. S. Beta-catenin is essential and sufficient for skeletal myogenesis in P19 cells. J Biol Chem. 2002, 277, 15393-15399). Нещодавно була висловлена думка, що активні форми кисню індукують ендотеліальний апоптоз, який згодом викликає експресію різних молекул адгезії для клітин пухлин. Це означає, що, вирішивши проблему із продукуванням ROS (активних форм кисню), можна реально запобігти рецидивам пухлин у віддалених ділянках (Ten, KM, van der Wal, JB, Sluiter, W, Hofland, LJ, Jeekel, J, Sonneveld, P, and van Eijck, CH; The role of superoxide anions in the development of distant tumour recurrence, 2006, Br.J Cancer). Орнітин декарбоксилаза 1 (ODC1) ODC1 - це лімітуючий швидкість фермент біосинтезу поліамінів, який каталізує перетворення орнітину в путресцин. Рівень активності ферменту змінюється у відповідь на стимули росту і демонструє високу інтенсивність обертів в порівнянні з іншими білками ссавців. Метаболізм поліамінів є невід’ємним компонентом механізму канцерогенезу в епітеліальних тканинах. Підвищення в ODC1 часто асоціюються з початком росту нормальних клітин і з підтриманням росту неопластичних клітин. Інгібітори ODC1 подавляють утворення пухлин в експериментальних моделях канцерогенезу сечового міхура, грудей, товстої кишки та шкіри. Надмірна експресія функції ODC1 є добре відомою характеристикою багатьох видів раку, і ODC1 вважається прото-онкогеном (Auvinen, M., Paasinen, A., Andersson, L. C. and Holtta, E. Ornithine decarboxylase activity is critical for cell transformation. Nature 1992, 360, 355-358). Генеративні мутації в гені аденоматозного поліпозу товстої кишки (APC) є одними з 7 UA 103882 C2 5 10 15 20 25 30 35 40 45 50 55 60 найбільш чітко визначених успадкованих схильностей до колоректального раку. Мутації APC викликають значне підвищення рівнів вільного -катеніну, який прямує до ядра, де формує комплекс з такими елементами, як лімфоїдний транскрипційний фактор (LEF)/T-клітинний фактор (Tcf) сімейства транскрипційних факторів, специфічних до послідовності. Онкоген c-myc є одним з генів-мішеней Tcf (He, T. C. et al. Identification of c-MYC as a target of the APC pathway (Science 281, 1509-1512 (1998). РНК c-Myc та білок надмірно експресуються і на ранніх, і на пізніх стадіях колоректального пухлиногенезу. ODC є геном-мішенню c-Myc. Втрата APC-функції викликає підвищену регуляцію ODC1 (Gerner, EW and Meyskens, FL, Jr., Polyamines and cancer: old molecules, new understanding, Nat. Rev. Cancer, 2004, 4, 781-792), та його надмірна експресія часто спостерігалася в колоректальній карциномі (Hu, H. Y. et al. Ornithine decarboxylase gene is overexpressed in colorectal carcinoma, World J. Gastroenterol. 2005, 11, 2244-2248; Kitahara, O. et al. Alterations of gene expression during colorectal carcinogenesis revealed by cDNA microarrays after laser-capture microdissection of tumor tissues and normal epithelia; Cancer Res. 2001, 61, 3544-3549; Nemoto, T., Kubota, S., Ishida, H., Murata, N. and Hashimoto, D. Ornithine decarboxylase, mitogen-activated protein kinase and matrix metalloproteinase-2 expressions in human colon tumors. World J. Gastroenterol. 2005, 11, 30653069). ODC1 має проангіогенні властивості, оскільки він діє як супресор ендостатіну (Nemoto, T., Hori, H., Yoshimoto, M., Seyama, Y. & Kubota, S. Overexpression of ornithine decarboxylase enhances endothelial proliferation by suppressing endostatin expression. Blood 2002, 99, 1478-1481). Інфекція CRC-клітинної лінії HT-29 аденовірусною антисенсною РНК до ODC1 та декарбоксилази S-аденозилметіоніну (ще один важливий фермент шляху біосинтезу поліаміну) призводить до зниженої регуляції CCND1 та затримання клітинного циклу. Крім того, ядерна транслокація бета-катеніну також інгібується (Gong, L, Jiang, C, Zhang, B, Hu, H, Wang, W, and Liu, X; Adenovirus-mediated Expression of Both Antisense Ornithine Decarboxylase and Sadenosylmethionine Decarboxylase Induces G(1) Arrest in HT-29 Cells, J Biochem. Mol. Biol, 2006, 39, 730-736). Більш того, аденовірус індукував регресію пухлин у випадках довготривалих пухлин у «голих» мишей (Zhang, B, Liu, XX, Zhang, Y, Jiang, CY, Hu, HY, Gong, L, Liu, M, and Teng, QS; Polyamine depletion by ODC-AdoMetDC antisense adenovirus impairs human colorectal cancer growth and invasion in vitro and in vivo, 2006, J Gene Med, 8, 980-989). Специфічним та незворотним інгібітором ODC1 є 2-дифторметил-орнітин (DMFO, Ефлорнітин (Sanofi-Aventis)). Він пропонується для лікування сонної хвороби (викликаної трипаносомами) та є активним інгредієнтом крему для епіляції Vaniqa. У відношенні до раку, DMFO широко використовувався у доклінічних моделях та показав багатообіцяючі протипухлинні ефекти завдяки зменшенню рівнів поліаміну (Gerner, EW and Meyskens, FL, Jr.; Polyamines and cancer: old molecules, new understanding, Nat. Rev. Cancer, 2004, 4, 781-792). Були проведені клінічні дослідження для кількох випадків раку, і зараз проводяться дослідження стосовно колоректального раку. Однак, вони є переважно комбінаційними підходами, що використовувались у превентивних призначеннях пацієнтам, особливо уразливим до CRC (при аденоматозних поліпах). Імуногенний ODC-пептид ODC-001 був ідентифікований раніше (M. Diehl, PhD Thesis, University of Tübingen, 1998) Ядерний антиген проліферуючих клітин (PCNA) PCNA знаходиться в ядрі та є ко-фактором дельта-ДНК-полімерази. Кодований білок діє як гомотример та сприяє збільшенню процесивності синтезу лідируючого ланцюгу в ході ДНКреплікації. Отже, він експресується в усіх проліферуючих клітинах, зокрема, в клітинах пухлин, та використовується як маркер для визначення проліферації. Проліферативні індекси в неопластичній та прилеглій нормальній слизовій оболонці, як визначено за допомогою імуногістохімічного аналізу PCNA, давно відомі як незалежні прогностичні фактори рецидивів та низької виживаності у пацієнтів з колоректальним раком (alSheneber, IF, Shibata, HR, Sampalis, J, and Jothy, S; Prognostic significance of proliferating cell nuclear antigen expression in colorectal cancer, Cancer, 1993, 71, 1954-1959; Mayer, A, Takimoto, M, Fritz, E, Schellander, G, Kofler, K, and Ludwig, H; The prognostic significance of proliferating cell nuclear antigen, epidermal growth factor receptor, and mdr gene expression in colorectal cancer, Cancer, 1993, 71, 2454-2460; Nakamura, T, Tabuchi, Y, Nakae, S, Ohno, M, and Saitoh, Y; Serum carcinoembryonic antigen levels and proliferating cell nuclear antigen labeling index for patients with colorectal carcinoma. Correlation with tumor progression and survival, Cancer, 1996, 77, 1741-1746) ДНК-топоізомераза II (TOP2) TOP2A і TOP2B кодують ізоформи ДНК-топоізомерази, ферменту, що контролює та змінює топологію ДНК в ході транскрипції. Цей ядерний фермент бере участь у таких процесах, як конденсація хромосом, розділення хроматид та зняття торсіонного напруження, що має місце в 8 UA 103882 C2 5 10 15 20 25 30 35 40 45 50 55 60 ході транскрипції та реплікації ДНК. Він каталізує тимчасове розділення та повторне з’єднання двох ланцюгів двоспіральної ДНК, що дозволяє ланцюгам проходити один крізь інший, таким чином, змінюючи топологію ДНК. Дві ізоформи цього ферменту є ймовірними продуктами події дуплікації генів. Ген, кодуючий альфа-форму, локалізується в хромосомі 17, а бета-ген локалізується в хромосомі 3. TOP2A є мішенню для декількох протиракових препаратів, і різноманітні мутації в цьому гені асоціюються з розвитком резистентності до ліків. Ген TOP2A знаходиться поблизу від онкогена HER-2, найбільш часто ампліфікованого онкогена у випадках раку молочної залози, на хромосомній ділянці 17q12-q21, та цей ген ампліфікується чи вищеплюється, з рівною частотою, майже в 90% первинних пухлин молочної залози, ампліфікованих HER-2 (Jarvinen, TA and Liu, ET; Topoisomerase IIalpha gene (TOP2A) amplification and deletion in cancer-more common than anticipated, Cytopathology, 14, 309-313). Крім того, про ампліфікації TOP2A повідомлялося для інших видів раку. Недавні експерименти, а також ряд комплексних великомасштабних досліджень наводять на думку, що ампліфікація (та/або делеція) гена TOP2A можуть бути причиною чутливості або резистентності до широко використовуваних цитотоксичних препаратів, тобто, інгібіторів топоізомерази II (антрацикліни та ін. Kellner, U, Sehested, M, Jeпnsen, PB, Gieseler, F, and Rudolph, P; Culprit and victim - DNA topoisomerase II, Lancet Oncol., 2002, 3, 235-243), в залежності від специфічного генетичного дефекту в локусі TOP2A (Jarvinen, TA and Liu, ET; Simultaneous amplification of HER-2 (ERBB2) and topoisomerase IIalpha (TOP2A) genes-molecular basis for combination chemotherapy in cancer, Curr.Cancer Drug Targets., 2006, 6, 579-602). Без TOP2A, реплікація ДНК та клітинний поділ неможливі. Отже, він стає головною мішенню багатьох режимів протипухлинної терапії, навіть незважаючи на те, що точний механізм знищення клітин залишається неясним (Kellner, U, Sehested, M, Jensen, PB, Gieseler, F, and Rudolph, P; Culprit and victim - DNA topoisomerase II, Lancet Oncol., 2002, 3, 235-243). Успіх цього підходу обмежується розвитком природної резистентності, та індуковане ліками ушкодження ДНК може збільшити злоякісність. TOP2B, другий потенційний вихідний білок для TOP-001, не був у центрі досліджень в області раку, в значній мірі тому, що він знаходиться на хромосомній ділянці (3p24), і його часта ампліфікація в пухлинах не відома. Однак, TOP2B за своєю первинною структурою схожий на TOP2A і має майже ідентичні каталітичні властивості (Leontiou, C, Lightowlers, R, Lakey, JH, and Austin, CA; Kinetic analysis of human topoisomerase IIalpha and beta DNA binding by surface plasmon resonance, FEBS Lett., 2003, 554, 206-210). Ще одне дослідження також показало, що обидві ізоформи можуть замінювати одна одну (Sakaguchi, A and Kikuchi, A; Functional compatibility between isoform alpha and beta of type II DNA topoisomerase, J Cell Sci., 2004, 117, 1047-1054). Автори цього винаходу надають переконливі докази того, що пухлино-асоційовані пептиди, які в достатній мірі зв’язуються з HLA-молекулами класу I, здатні викликати імунні реакції, де посередниками виступають CD8-позитивні цитотоксичні T-лімфоцити, також демонструючи те, що заявлені пептиди є прийнятними для запуску реакцій імунної системи людини проти обраних пептидів в пептидомі пухлинної клітини. Подібним чином, було виявлено, що пухлино-асоційовані пептиди, які в достатній мірі зв’язуються з HLA-молекулами класу II, зокрема ті алелі HLA класу II, що генетично кодуються локусами HLA DR геному людини, здатні викликати імунні реакції, де посередниками виступають CD4-позитивні T-клітини людини. CD4-позитивні T-клітини були виділені з периферичної крові людини, і демонструється, що заявлені пептиди є прийнятними для запуску T-клітинних реакцій імунної системи людини проти обраних пептидів в пептидомі пухлинної клітини. Як показано нижче на прикладі пептиду TGFBI-004, цей зв’язуваний з HLA-DR пухлиноасоційований пептид розпізнається CD4-позитивними T-клітинами. Оскільки пептиди можуть синтезуватись хімічним способом та використовуватись як активні фармацевтичні інгредієнти фармацевтичних препаратів, пептиди, запропоновані в цьому винаході, можна використовувати для імунотерапії, переважно, імунотерапії раку. В іншому аспекті, фармацевтична композиція, крім того, має принаймні один додатковий пептид, котрий вміщує амінокислотну послідовність, обрану з групи, яка включає послідовності від SEQ ID NO 8 до SEQ ID NO 15, або вміщує варіант амінокислотної послідовності, принаймні на 80 % гомолочний послідовності від SEQ ID NO 8 до SEQ ID NO 15, чи полінуклеотид, який вміщує нуклеїнову кислоту, що кодує послідовності від SEQ ID NO 8 до SEQ ID NO 15 або варіант амінокислотної послідовності. Пептиди послідовності від SEQ ID NO 8 до SEQ ID NO 13 та 15 є імуногенними пептидами, ідентифікованими раніше, та зв’язуються з MHC-молекулами класу I і класу II (див. Таблицю 2). 9 UA 103882 C2 5 10 Ці пептиди, як було показано, викликають T-клітинні реакції in vivo у пацієнтів, хворих на гіпернефрому (RCC) (H. Singh-Jasuja, S. Walter, T. Weinschenk, A. Mayer, P. Y. Dietrich, M. Staehler, A. Stenzl, S. Stevanovic, H. Rammensee, J. Frisch; Correlation of T-cell response, clinical activity and regulatory T-cell levels in renal cell carcinoma patients treated with IMA901, a novel multipeptide vaccine; ASCO Meeting 2007 Poster # 3017; M. Staehler, A. Stenzl, P. Y. Dietrich, T. Eisen, A. Haferkamp, J. Beck, A. Mayer, S. Walter, H. Singh, J. Frisch, C. G. Stief; An open label study to evaluate the safety and immunogenicity of the peptide based cancer vaccine IMA901, ASCO meeting 2007; Poster # 3017). Оскільки вихідні білки надмірно експресуються не тільки в RCC, але й в CRC та інших видах раку, ці пептиди також є прийнятними у вакцинах для лікування інших типів пухлин, зокрема, у вакцинах проти колоректального раку. Таблиця 2 Додаткові імуногенні пептиди, прийнятні в композиції винаходу Ід. № Ід. номер послідовно пептиду сті 8 9 10 25 30 35 40 MMP-001 12 13 14 15 20 CCN-001 MUC-001 11 15 CEA-006 CEA-005 MET-001 (HBV-001) CEA-004 Послідовність SPQYSWRINGIPQQH T LLGATCMFV STAPPVHNV SQDDIKGIQKLYGKR S YLSGADLNL YVDPVITSI FLPSDFFPSV YLSGANLNL Символ гена Функція CCND1 MUC1 Карциноембріональний антиген Циклін D1 Муцин 1 MMP7 Металопротеїназа 7 CEACAM5 CEACAM5 Варіант пептиду CEA MET Протоонкоген met Контрольний пептид CEACAM5 Пептид CEA Зв’язується з MHC HLA-DR HLA-A*02 HLA-A*02 HLA-DR HLA-A*02 HLA-A*02 HLA-A*02 Молекула клітинної адзезії 5, що відноситься до карциноембріонального антигену Карциноембріональний антиген (CEA = CEACAM5) є високо глікозильованим мембранним білком з масою 180 kDa, що складається з трьох C2 Ig-подібних повторюваних одиниць, до яких прилягає N-термінальна Ig V-подібна ділянка та C-термінальна ділянка, що включає ділянку зв’язування з глікофосфатидилінозитолом (Hegde, P, Qi, R, Gaspard, R, Abernathy, K, Dharap, S, Earle-Hughes, J, Gay, C, Nwokekeh, NU, Chen, T, Saeed, AI, Sharov, V, Lee, NH, Yeatman, TJ, and Quackenbush, J; Identification of tumour markers in models of human colorectal cancer using a 19,200-element complementary DNA microarray, Cancer Res., 2001, 61, 7792-7797). Як онкофетальний антиген, CEA експресується на протязі розвитку плода, але також, на низьких рівнях, - в епітелії шлунково-кишкового тракту дорослих людей. Однак, CEA надмірно експресується у великій кількості (в процентному відношенні) пухлин людини, включаючи 90% клітин пухлин шлунково-кишкового тракту, товстої кишки та підшлункової залози, 70% ракових клітин у випадках немілкоклітинного раку легенів та в 50% клітин пухлин у випадках раку молочної залози (Thompson, JA, Grunert, F, and Zimmermann, W; Carcinoembryonic antigen gene family: molecular biology and clinical perspectives, J Clin Lab Anal., 5, 344-366 2005). Внаслідок його високої експресії клітинами пухлин та секреції у сироватку, CEA широко використовувався як маркер пухлин (Sikorska, H, Shuster, J, and Gold, P; Clinical applications of carcinoembryonic antigen, Cancer Detect.Prev., 12, 321-355 1988) та є стандартним маркером сироватки для моніторингу колоректального раку (Locker, GY, Hamilton, S, Harris, J, Jessup, JM, Kemeny, N, Macdonald, JS, Somerfield, MR, Hayes, DF, and Bast, RC, Jr.; ASCO 2006 update of recommendations for the use of tumour markers in gastrointestinal cancer, J Clin Oncol, 24, 53135327, 2006). Незважаючи на надмірну експресію CEA в клітинах пухлин, у пацієнтів з раковими захворюваннями, зазвичай, не виявляється імунної реакції проти цього антигену (Orefice, S, Fossati, G, Pietrojusti, E, and Bonfanti, G; Delayed cutaneous hypersensitivity reaction to carcinoembryonic antigen in cancer patients, Tumouri, 1982, 68, 473-475,). Імунна система, як правило, стає толерантною до CEA, оскільки він нормально експресується на низьких рівнях в тілі людини. Однак, в ряді клінічних досліджень вакцин була продемонстрована імуногенність CEA (Sarobe, P, Huarte, E, Lasarte, JJ, and Borras-Cuesta, F; Carcinoembryonic antigen as a target to induce anti-tumour immune responses, Curr. Cancer Drug Targets., 2004, 4, 443-454), особливо в 10 UA 103882 C2 5 10 15 20 25 30 35 40 45 50 55 60 колоректальній карциномі (CRC) (Mosolits, S, Ullenhag, G, and Mellstedt, H; Therapeutic vaccination in patients with gastrointestinal malignancies. A review of immunological and clinical results, Ann.Oncol., 2005, 16, 847-862), і CEA є пухлино-асоційованим антигеном (TAA) з найбільшою кількістю ділянок для створення вакцин, випробуваних в цьому типі пухлин (von Mehren, M; Colorectal cancer vaccines: what we know and what we don't yet know, Semin. Oncol., 2005, 32, 76-84). Щодо CEA, було описано декілька епітопів цитотоксичних Т-клітин і Т-клітин - хелперів (Crosti, M, Longhi, R, Consogno, G, Melloni, G, Zannini, P, and Protti, MP; Identification of novel subdominant epitopes on the carcinoembryonic antigen recognized by CD4+ T-cells of lung cancer patients, J Immunol., 2006, 176, 5093-5099; Novellino, L, Castelli, C, and Parmiani, G; A listing of human tumour antigens recognized by T-cells: March 2004 update, Cancer Immunol.Immunother., 2004, 54, 187-207; Ruiz, M, Kobayashi, H, Lasarte, JJ, Prieto, J, Borras-Cuesta, F, Celis, E, and Sarobe, P; Identification and characterization of a T-helper peptide from carcinoembryonic antigen, Clin Cancer Res., 2004, 10, 2860-2867), що дають можливість проведення різних досліджень по розробці вакцин на пептидній основі стосовно CRC (Babatz, J, Rollig, C, Lobel, B, Folprecht, G, Haack, M, Gunther, H, Kohne, CH, Ehninger, G, Schmitz, M, and Bornhauser, M; Induction of cellular immune responses against carcinoembryonic antigen in patients with metastatic tumours after vaccination with altered peptide ligand-loaded dendritic cells, Cancer Immunol. Immunother., 2006, 55, 268-276; Fong, L, Hou, Y, Rivas, A, Benike, C, Yuen, A, Fisher, GA, Davis, MM, and Engleman, EG; Altered peptide ligand vaccination with Flt3 ligand expanded dendritic cells for tumour immunotherapy, Proc. Natl. Acad. Sci. U.S.A, 2001, 98, 8809-8814; Liu, KJ, Wang, CC, Chen, LT, Cheng, AL, Lin, DT, Wu, YC, Yu, WL, Hung, YM, Yang, HY, Juang, SH, and Whang-Peng, J; Generation of carcinoembryonic antigen (CEA)-specific T-cell responses in HLA-A*0201 and HLAA*2402 late-stage colorectal cancer patients after vaccination with dendritic cells loaded with CEA peptides, Clin Cancer Res., 2004, 10, 2645-2651; Matsuda, K, Tsunoda, T, Tanaka, H, Umano, Y, Tanimura, H, Nukaya, I, Takesako, K, and Yamaue, H; Enhancement of cytotoxic T-lymphocyte responses in patients with gastrointestinal malignancies following vaccination with CEA peptidepulsed dendritic cells, Cancer Immunol. Immunother., 2004, 53, 609-616; Ueda, Y, Itoh, T, Nukaya, I, Kawashima, I, Okugawa, K, Yano, Y, Yamamoto, Y, Naitoh, K, Shimizu, K, Imura, K, Fuji, N, Fujiwara, H, Ochiai, T, Itoi, H, Sonoyama, T, Hagiwara, A, Takesako, K, and Yamagishi, H; Dendritic cell-based immunotherapy of cancer with carcinoembryonic antigen-derived, HLA-A24-restricted CTL epitope: Clinical outcomes of 18 patients with metastatic gastrointestinal or lung adenocarcinomas, Int. J Oncol., 2004, 24, 909-917; Weihrauch, MR, Ansen, S, Jurkiewicz, E, Geisen, C, Xia, Z, Anderson, KS, Gracien, E, Schmidt, M, Wittig, B, Diehl, V, Wolf, J, Bohlen, H, and Nadler, LM; Phase I/II combined chemoimmunotherapy with carcinoembryonic antigen-derived HLA-A2-restricted CAP-1 peptide and irinotecan, 5-fluorouracil, and leucovorin in patients with primary metastatic colorectal cancer, Clin Cancer Res., 2005, 11, 5993-6001). Ці та інші клінічні дослідження на даний момент продемонстрували безпеку вакцин на основі CEA та доказали викликання імунної реакції проти цього антигену (von Mehren, M; Colorectal cancer vaccines: what we know and what we don't yet know, Semin.Oncol., 2005, 32, 76-84). Дані про варіант CEA-006 були опубліковані раніше (Ruiz, M, Kobayashi, H, Lasarte, JJ, Prieto, J, Borras-Cuesta, F, Celis, E, and Sarobe, P; Identification and characterization of a T-helper peptide from carcinoembryonic antigen, Clin Cancer Res., 2004, 10, 2860-2867). CEA-005 є мутантом з однією зміненою амінокислотою, і відповідно до наявної інформації він переборює центральну імунну толерантність (Zaremba, S, Barzaga, E, Zhu, M, Soares, N, Tsang, KY, and Schlom, J; Identification of an enhancer agonist cytotoxic T lymphocyte peptide from human carcinoembryonic antigen, Cancer Res., 1997, 57, 4570-4577). Трансформуючий фактор росту, бета-індукований (TGFBI) TGFBI був вперше ідентифікований як TGF-бета-індуцибельний ген в клітинній лінії аденокарциноми легенів людини. Він кодує білок екстрацелюлярного матриксу, що секретується, котрий вважається впливаючим на адгезію клітин і на склад екстрацелюлярного матриксу. Було показано, що TGFBI є серед найбільш підвищених за кількістю генів у випадках колоректального раку; крім того, він характеризується високими рівнями експресії в аденомах. Результаті кількісного ПЛР-аналізу продемонстрували сильне підвищення як в неочищених, так і в очищених ракових епітеліальних клітинах. Відповідно, в експериментах по гібридізації in situ було виявлено експресію TGFBI в багатьох типах клітин, і в стромальному, і в епітеліальному компартментах (Buckhaults, P, Rago, C, St, CB, Romans, KE, Saha, S, Zhang, L, Vogelstein, B, and Kinzler, KW; Secreted and cell surface genes expressed in benign and malignant colorectal tumours, Cancer Res., 2001, 61, 6996-7001). 11 UA 103882 C2 5 10 15 20 25 30 35 40 45 50 55 60 В мета-аналізі досліджень по вивченню генної експресії в колоректальній карциномі, TGFBI був визначений як лише один з дев’яти генів, що описуються з постійною підвищеною експресією (4 дослідження щодо TGFBI) (Shih, W, Chetty, R, and Tsao, MS; Expression profiling by microarrays in colorectal cancer, Oncol.Rep., 2005, 13, 517-524). В тканинах пухлин підшлункової залози людини спостерігалося підвищення рівнів мРНК TGFBI в 32.4 рази, в порівнянні з нормальними контрольними тканинами. Гібридізація іn situ виявила експресію мРНК TGFBI, головним чином, в ракових клітинах в межах пухлинного утворення підшлункової залози (Schneider, D, Kleeff, J, Berberat, PO, Zhu, Z, Korc, M, Friess, H, and Buchler, MW; Induction and expression of betaig-h3 in pancreatic cancer cells, Biochim.Biophys.Acta, 2002, 1588, 1-6). TGFBI був ідентифікований як ген, що сприяє ангіогенезу в моделі in vitro. Крім того, в декількох пухлинах була визначена різко підвищена експресія TGFBI. Антисенсні олігонуклеотиди до TGFBI блокували генну експресію та утворення ендотеліальних трубок in vitro, і це наводить на думку, що TGFBI може грати вирішальну роль у взаємозв’язку між ендотеліальними клітинами та матриксом (Aitkenhead, M, Wang, SJ, Nakatsu, MN, Mestas, J, Heard, C, and Hughes, CC; Identification of endothelial cell genes expressed in an in vitro model of angiogenesis: induction of ESM-1, (beta)ig-h3, and NrCAM, Microvasc. Res., 2002, 63, 159-171). Муцин-1 (MUC1) Муцини являють собою епітеліальні глікопротеїни з великою молекулярною вагою, які мають високий вміст вбудованих олігосахаридів, з’єднаних за допомогою O-глікозидного зв’язку з пептидами тандемного повтору із значним вмістом треоніну, серину та проліну. Існують два структурно та функціонально відокремлюваних класи муцинів: трансмембранні муцини, до яких належить MUC1, та гелеутворюючі муцини, що секретуються. Муцини колоректального раку мають відмінності у вуглеводних структурах, що дослідним шляхом визначені як діагностичні та прогностичні маркери, а також як мішені для протиракових вакцин. Позаклітинний домен білку MUC1 складається з висококонсервативних повторів 20 амінокислот; фактична кількість варіюється від 25 до 100, в залежності від алелі. Кожний тандемний повтор вміщує п’ять потенційних сайтів глікозилювання, а між пар треоніну та серину знаходиться імунодомінантна ділянка, яка вміщує епітопи, що розпізнаються різними антиMUC1-антитілами (Taylor-Papadimitriou, J, Burchell, J, Miles, DW, and Dalziel, M; MUC1 and cancer, Biochim. Biophys. Acta, 1999, 1455, 301-313). В порівнянні з більшістю інших епітеліальних тканин, MUC1 товстої кишки є глікозильованим в більш значній мірі, що маскує білок MUC1 для імуногістохімічного викрашування MUC1специфічними антитілами. В колоректальних аденокарциномах MUC1 менше глікозильований, що дозволяє здійснювати імуно-детекцію. Аберрантно глікозильований MUC1 надає нові зв’язувальні властивості та може одночасно сприяти зв’язку з молекулами адгезії з певною молекулярною специфічністю та блокувати його, відіграючи подвійну роль в метастатичному поширенні клітин пухлин (McDermott, KM, Crocker, PR, Harris, A, Burdick, MD, Hinoda, Y, Hayashi, T, Imai, K, and Hollingsworth, MA; Overexpression of MUC1 reconfigures the binding properties of tumor cells, Int. J Cancer, 2001, 94, 783-791). MUC1, як це визначено імунологічним шляхом, має підвищену експресію в пухлинах товстої кишки, що корелює з більш несприятливим прогнозом (Byrd, JC and Bresalier, RS; Mucins and mucin binding proteins in colorectal cancer, Cancer Metastasis Rev., 2004, 23, 77-99), вказуючи на те, що підвищена регуляція MUC1 може грати активну роль в прогресуванні CRC. Пухлини товстої кишки з метастазами експресують MUC1 в більшій мірі, ніж пухлини без метастаз (Nakamori, S, Ota, DM, Cleary, KR, Shirotani, K, and Irimura, T; MUC1 mucin expression as a marker of progression and metastasis of human colorectal carcinoma, Gastroenterology, 1994, 106, 353361), та викрашування MUC1 було позитивним в усіх випадках колоректального раку з ураженням печінки, в одному з досліджень (Matsuda, K, Masaki, T, Watanabe, T, Kitayama, J, Nagawa, H, Muto, T, and Ajioka, Y; Clinical significance of MUC1 and MUC2 mucin and p53 protein expression in colorectal carcinoma, Jpn. J Clin Oncol., 2000, 30, 89-94). Недавнє дослідження 462 людей, хворих на колоректальний рак, виявило, що експресія MUC1 є незалежним прогностичним маркером несприятливого прогнозу (Duncan, TJ, Watson, NF, Al-Attar, AH, Scholefield, JH, and Durrant, LG; The role of MUC1 and MUC3 in the biology and prognosis of colorectal cancer, World J Surg. Oncol, 2007, 5, 31). Треба відзначити патофізіологічну значимість циркуляції анти-MUC1-антитіл в CRC: АнтиMUC1- антитіла були визначені у 5 з 31 (16.1%) здорових людей та у 27 з 56 (48.2%) пацієнтів з колоректальним раком (Nakamura, H, Hinoda, Y, Nakagawa, N, Makiguchi, Y, Itoh, F, Endo, T, and Imai, K; Detection of circulating anti-MUC1 mucin core protein antibodies in patients with colorectal cancer, J Gastroenterol., 1998, 33, 354-361). 12 UA 103882 C2 5 10 15 20 25 30 35 40 45 50 55 60 Крім виконання ролі мішені антитіл, MUC1 також являє собою визнану мішень для цитотоксичних T-клітин. Кілька наукових робіт продемонстрували, що цитотоксичні MHCнеобмежені T-клітини з пухлин яєчників, грудей, підшлункової залози та множинної мієломи можуть розпізнавати епітопи корового білку MUC1, локалізовані в тандемному повторі (Apostolopoulos, V and McKenzie, IF; Cellular mucins: targets for immunotherapy, Crit Rev. Immunol., 1994, 14, 293-309; Finn, OJ, Jerome, KR, Henderson, RA, Pecher, G, Domenech, N, MagarianBlander, J, and Barratt-Boyes, SM; MUC-1 epithelial tumor mucin-based immunity and cancer vaccines, Immunol. Rev., 1995, 145, 61-89; Barnd, DL, Lan, MS, Metzgar, RS, and Finn, OJ; Specific, major histocompatibility complex-unrestricted recognition of tumor-associated mucins by human cytotoxic T cells, Proc. Natl. Acad. Sci. U.S.A, 1989, 86, 7159-7163; Takahashi, T, Makiguchi, Y, Hinoda, Y, Kakiuchi, H, Nakagawa, N, Imai, K, and Yachi, A; Expression of MUC1 on myeloma cells and induction of HLA-unrestricted CTL against MUC1 from a multiple myeloma patient, J. Immunol., 1994, 153, 2102-2109; Noto, H, Takahashi, T, Makiguchi, Y, Hayashi, T, Hinoda, Y, and Imai, K; Cytotoxic T lymphocytes derived from bone marrow mononuclear cells of multiple myeloma patients recognize an underglycosylated form of MUC1 mucin, Int. Immunol., 1997, 9, 791-798). Однак, також були ідентифіковані HLA-A*02-обмежені епітопи T-клітин, одержані з білку MUC1 (Apostolopoulos, V, Karanikas, V, Haurum, JS, and McKenzie, IF; Induction of HLA-A2-restricted CTLs to the mucin 1 human breast cancer antigen, J Immunol., 1997, 159, 5211-5218; Brossart, P, Heinrich, KS, Stuhler, G, Behnke, L, Reichardt, VL, Stevanovic, S, Muhm, A, Rammensee, HG, Kanz, L, and Brugger, W; Identification of HLA-A2-restricted T-cell epitopes derived from the MUC1 tumor antigen for broadly applicable vaccine therapies, Blood, 1999, 93, 4309-4317). Одним з тих пептидів є MUC-001. Він одержується з ділянки тандемного повтору білку MUC1. Індукування реакцій цитотоксичних T-лімфоцитів in vivo після вакцинацій пацієнтів із запущеним раком грудей чи яєчників дендритними клітинами, в які імпульсним шляхом введені зазначені пептиди MUC1, було успішним (Brossart, P, Wirths, S, Stuhler, G, Reichardt, VL, Kanz, L, and Brugger, W; Induction of cytotoxic T-lymphocyte responses in vivo after vaccinations with peptide-pulsed dendritic cells, Blood, 2000, 96, 3102-3108; Wierecky, J, Mueller, M, and Brossart, P; Dendritic cell-based cancer immunotherapy targeting MUC-1, Cancer Immunol. Immunother., 2005 Apr. 28: 288-94). Крім того, зазначені вакцинації успішно індукували клінічні реакції у пацієнтів з гіпернефромою (Wierecky, J, Muller, MR, Wirths, S, Halder-Oehler, E, Dorfel, D, Schmidt, SM, Hantschel, M, Brugger, W, Schroder, S, Horger, MS, Kanz, L, and Brossart, P; Immunologic and clinical responses after vaccinations with peptide-pulsed dendritic cells in metastatic renal cancer patients, Cancer Res., 2006, 66, 5910-5918). Підвищена експресія імунореактивного MUC1 в колоректальному раку, переважно, не базується на надмірній експресії мРНК, а скоріше за все обумовлена його зменшеним глікозилюванням, що демаскує епітопи для розпізнавання антитіл, особливо в ділянці тандемного повтору MUC1. Разом з тим, таке деглікозилювання надає можливість утворення епітопів T-клітин за допомогою зміненого процесингу антигенів в клітинах пухлин, якому перешкоджає глікозилювання в нормальних клітинах. Цей механізм може пояснити надзвичайні властивості MUC-001 як пухлино-асоційованого епітопу Т-клітин, незважаючи на відсутність сильної надмірної експресії мРНК. Певні докази впливу змін у глікозилюванні на процесинг антигенів були одержані за даними недавнього дослідження, яке підтвердило, що змінене глікозилювання МUC1 у випадку колоректального раку може фактично визначатись антигенпрезентуючими клітинами за допомогою рецептору, що специфічно розпізнає глікоформи у випадках раку (Saeland, E, van Vliet, SJ, Backstrom, M, van, dB, V, Geijtenbeek, TB, Meijer, GA, and van, KY; 2006, The C-type lectin MGL expressed by dendritic cells detects glycan changes on MUC1 in colon carcinoma, Cancer Immunol.Immunother., 2007, 56(8): 1225-36). Специфічне поглинання і процесинг ракових глікоформ антиген-презентуючими клітинами також може пояснити той факт, що MUC-001-специфічні T-клітини спостерігалися природньо (без імунізації) у хворих на рак молочної залози (Rentzsch, C, Kayser, S, Stumm, S, Watermann, I, Walter, S, Stevanovic, S, Wallwiener, D, and Guckel, B; Evaluation of pre-existent immunity in patients with primary breast cancer: molecular and cellular assays to quantify antigen-specific T lymphocytes in peripheral blood mononuclear cells, Clin Cancer Res., 2003, 9, 4376-4386) та колоректальний рак (Dittmann, J, Keller-Matschke, K, Weinschenk, T, Kratt, T, Heck, T, Becker, HD, Stevanovic, S, Rammensee, HG, and Gouttefangeas, C; CD8+ T-cell response against MUC1-derived peptides in gastrointestinal cancer survivors, Cancer Immunol. Immunother., 2005, 54, 750-758). Не надається ніякої інформації про аутоімунні ефекти у цих пацієнтів. Це демонструє природну роль MUC-001 як пухлино-асоційованого пептиду, що індукує специфічні T-клітини, та дає можливість вважати безпечним введення MUC-001, хоча на рівні мРНК не можна визначити надмірну експресію антигену MUC1. 13 UA 103882 C2 5 10 15 20 25 30 35 40 45 50 55 60 Протоонкоген Met (рецептор фактору росту гепатоцитів) (c-Met) Білковий продукт MET-протоонкогену являє собою рецептор фактору росту гепатоцитів. Він вміщує тирозин-кіназний домен, який активує шляхи передачі сигналів, що беруть участь в проліферації, рухомості, адгезії та інвазії клітин (Trusolino, L and Comoglio, PM; Scatter-factor and semaphorin receptors: cell signalling for invasive growth, Nat. Rev. Cancer, 2002, 2, 289-300). Дослідження різних типів пухлин продемонструвало декілька механізмів активації c-Met, включаючи аутокринну петлю HGF/c-Met, активуючу точкові мутації, гібридний білок TPR-Met, а також нездатність розділити c-Met на - та -ланцюги (Di Renzo, MF, Olivero, M, Martone, T, Maffe, A, Maggiora, P, Stefani, AD, Valente, G, Giordano, S, Cortesina, G, and Comoglio, PM; Somatic mutations of the MET oncogene are selected during metastatic spread of human HNSC carcinomas, Oncogene, 2000, 19, 1547-1555; Ebert, M, Yokoyama, M, Friess, H, Buchler, MW, and Korc, M; Coexpression of the c-met proto-oncogene and hepatocyte growth factor in human pancreatic cancer, Cancer Res., 1994, 54, 5775-5778; Mondino, A, Giordano, S, and Comoglio, PM; Defective posttranslational processing activates the tyrosine kinase encoded by the MET protooncogene (hepatocyte growth factor receptor), Mol. Cell Biol., 1991, 11, 6084-6092; Olivero, M, Valente, G, Bardelli, A, Longati, P, Ferrero, N, Cracco, C, Terrone, C, Rocca-Rossetti, S, Comoglio, PM, and Di Renzo, MF; Novel mutation in the ATP-binding site of the MET oncogene tyrosine kinase in a HPRCC family, Int. J Cancer, 1999, 82, 640-643; Park, M, Dean, M, Cooper, CS, Schmidt, M, O'Brien, SJ, Blair, DG, and Vande Woude, GF; Mechanism of met oncogene activation, Cell, 1986, 45, 895-904; Park, WS, Dong, SM, Kim, SY, Na, EY, Shin, MS, Pi, JH, Kim, BJ, Bae, JH, Hong, YK, Lee, KS, Lee, SH, Yoo, NJ, Jang, JJ, Pack, S, Zhuang, Z, Schmidt, L, Zbar, B, and Lee, JY; 1999, Somatic mutations in the kinase domain of the Met/hepatocyte growth factor receptor gene in childhood hepatocellular carcinomas, Cancer Res., 59, 307-310; Rahimi, N, Tremblay, E, McAdam, L, Park, M, Schwall, R, and Elliott, B; 1996, Identification of a hepatocyte growth factor autocrine loop in a murine mammary carcinoma, Cell Growth Differ., 7, 263-270;Schmidt, L, Duh, FM, Chen, F, Kishida, T, Glenn, G, Choyke, P, Scherer, SW, Zhuang, Z, Lubensky, I, Dean, M, Allikmets, R, Chidambaram, A, Bergerheim, UR, Feltis, JT, Casadevall, C, Zamarron, A, Bernues, M, Richard, S, Lips, CJ, Walther, MM, Tsui, LC, Geil, L, Orcutt, ML, Stackhouse, T, Lipan, J, Slife, L, Brauch, H, Decker, J, Niehans, G, Hughson, MD, Moch, H, Storkel, S, Lerman, MI, Linehan, WM, and Zbar, B; 1997, Germline and somatic mutations in the tyrosine kinase domain of the MET proto-oncogene in papillary renal carcinomas, Nat.Genet., 16, 68-73; Schmidt, L, Junker, K, Weirich, G, Glenn, G, Choyke, P, Lubensky, I, Zhuang, Z, Jeffers, M, Vande, WG, Neumann, H, Walther, M, Linehan, WM, and Zbar, B; 1998, Two North American families with hereditary papillary renal carcinoma and identical novel mutations in the MET proto-oncogene, Cancer Res., 58, 1719-1722). Що стосується механізму, надмірна експресія c-Met, разом із онкогенною мутацією Ki-ras, посилює туморогенність клітин пухлин товстої кишки in vivo (Long, IS, Han, K, Li, M, Shirasawa, S, Sasazuki, T, Johnston, M, and Tsao, MS; Met receptor overexpression and oncogenic Ki-ras mutation cooperate to enhance tumorigenicity of colon cancer cells in vivo, Mol.Cancer Res., 2003, 1, 393-401). Цікавим є факт існування певних доказів взаємодії передачі сигналів MET з Wnt/бетакатеніновим шляхом, що часто посилений у випадках раку товстої кишки. MET може активуватися Простагландином E2 (PGE2), і PGE2-активований c-Met асоціюється з -катеніном та підвищує фосфорилювання тирозину, таким чином індукуючи інвазивність клітин колоректального раку (Pai, R, Nakamura, T, Moon, WS, and Tarnawski, AS; Prostaglandins promote colon cancer cell invasion; signaling by cross-talk between two distinct growth factor receptors, FASEB J, 2003, 17, 1640-1647). Останнім часом, була описана взаємна активація MET і бетакатеніну, що призводить до формування позитивного зворотнього зв’язку між цими двома ключовими учасниками колоректального пухлиногенезу (Rasola, A, Fassetta, M, De, BF, D'Alessandro, L, Gramaglia, D, Di Renzo, MF, and Comoglio, PM; A positive feedback loop between hepatocyte growth factor receptor and beta-catenin sustains colorectal cancer cell invasive growth, Oncogene, 2007, 26, 1078-1087). Рівень експресії мРНК c-Met в первинних пухлинах CRC (n = 36) є важливим прогностичним маркером для інвазії на ранніх стадіях та регіонального метастазування, встановлюючи пряме співвідношення зі стадією колоректального раку (Takeuchi, H, Bilchik, A, Saha, S, Turner, R, Wiese, D, Tanaka, M, Kuo, C, Wang, HJ, and Hoon, DS; c-MET expression level in primary colon cancer: a predictor of tumor invasion and lymph node metastases, Clin Cancer Res., 2003, 9, 14801488). Інший аналіз експресії c-Met в 130 зразках CRC показав надмірну експресію (T/N > 2.0) cMet в 69% первинних CRC та значно вищі рівні c-Met в CRC з інвазією в кровоносні судини (P = 0.04) та на запущеній стадії (P = 0.04), що підтверджує роль c-Met в прогресуванні CRC та поширенні метастаз у людей (Zeng, Z, Weiser, MR, D'Alessio, M, Grace, A, Shia, J, and Paty, PB; Immunoblot analysis of c-Met expression in human colorectal cancer: overexpression is associated 14 UA 103882 C2 5 10 15 20 25 30 35 40 45 50 55 60 with advanced stage cancer, Clin Exp. Metastasis, 2004, 21, 409-417). Ще в одному дослідженні 69% та 48% з 60 аденокарцином товстої кишки показали, відповідно, більші ніж в 2 рази та в 10 раз рівні мРНК c-Met, в порівнянні з суміжною нормальною слизовою оболонкою (Kammula, US, Kuntz, EJ, Francone, TD, Zeng, Z, Shia, J, Landmann, RG, Paty, PB, and Weiser, MR; Molecular coexpression of the c-Met oncogene and hepatocyte growth factor in primary colon cancer predicts tumor stage and clinical outcome, Cancer Lett., 2007, 248, 219-228). Отже, підвищення сигналу cMet є типовим на ранніх стадіях CRC, з навіть більшим вираженням в запущених випадках хвороби та при метастазуванні. Циклін D1 (CCND1) CCND1 належить до сімейства високо-консервативних циклінів, члени якого характеризуються різким підвищенням концентрації білку на протязі клітинного циклу. Циклини функціонують як регулятори цикліно-залежних кіназ (CDK). Різні цикліни демонструють визначені моделі експресії та розпаду, що робить свій внесок у тимчасову координацію кожної мітотичної події. Такий циклін формує комплекс з CDK4 або CDK6 і функціонує як регуляторна підгрупа CDK4 або CDK6, активність яких вимагається для переходу G1/S клітинного циклу. Мутації, ампліфікація та надмірна експресія цього гену, що змінює розвиток клітинного циклу, часто спостерігаються в різноманітних пухлинах та можуть сприяти пухлиногенезу (Fu, M, Wang, C, LI, Z, Sakamaki, T, and Pestell, RG; Minireview: Cyclin D1: normal and abnormal functions, Endocrinology, 2004, 145, 5439-5447). Типовий однонуклеотидний поліморфізм A/G (A870G) дає в результаті дві окремі ізоформи мРНК - a та b. Альтернативно сплайсована ізоформа b кодує укорочений білок, який зв’язують з підвищеною частотою виникнення пухлин, включаючи рак легенів, рак товстої кишки та інші види раку (Fu, M, Wang, C, LI, Z, Sakamaki, T, and Pestell, RG; Minireview: Cyclin D1: normal and abnormal functions, Endocrinology, 2004, 145, 5439-5447). Відносно колоректального раку, часто описується надмірна експресія CCND1 на рівнях мРНК і білків (Sutter, T, Doi, S, Carnevale, KA, Arber, N, and Weinstein, IB; Expression of cyclins D1 and E in human colon adenocarcinomas, J Med, 1997, 28, 285-309; Mermelshtein, A, Gerson, A, Walfisch, S, Delgado, B, Shechter-Maor, G, Delgado, J, Fich, A, and Gheber, L; Expression of D-type cyclins in colon cancer and in cell lines from colon carcinomas, Br. J Cancer, 2005, 93, 338-345; Balcerczak, E, Pasz-Walczak, G, Kumor, P, Panczyk, M, Kordek, R, Wierzbicki, R, and Mirowski, M; Cyclin D1 protein and CCND1 gene expression in colorectal cancer, Eur. J Surg.Oncol., 2005, 31, 721-726; Bondi, J, Husdal, A, Bukholm, G, Nesland, JM, Bakka, A, and Bukholm, IR; Expression and gene amplification of primary (A, B1, D1, D3, and E) and secondary (C and H) cyclins in colon adenocarcinomas and correlation with patient outcome, J Clin Pathol., 2005, 58, 509-514; Perez, R, Wu, N, Klipfel, AA, and Beart, RW, Jr.; A better cell cycle target for gene therapy of colorectal cancer: cyclin G, J Gastrointest. Surg., 2003, 7, 884-889; Wong, NA, Morris, RG, McCondochie, A, Bader, S, Jodrell, DI, and Harrison, DJ; Cyclin D1 overexpression in colorectal carcinoma in vivo is dependent on beta-catenin protein dysregulation, but not k-ras mutation, J Pathol., 2002, 197, 128-135; McKay, JA, Douglas, JJ, Ross, VG, Curran, S, Murray, GI, Cassidy, J, and McLeod, HL; Cyclin D1 protein expression and gene polymorphism in colorectal cancer. Aberdeen Colorectal Initiative, Int.J Cancer, 2000, 88, 77-81; Bartkova, J, Lukas, J, Strauss, M, and Bartek, J; The PRAD-1/cyclin D1 oncogene product accumulates aberrantly in a subset of colorectal carcinomas, Int. J Cancer, 1994, 58, 568573). Це може пояснюватися визнаним фактом, що CCND1 є геном-мішенню шляху активації катенін-TCF/LEF, який часто має підвищену експресію в колоректальній карциномі (Shtutman, M, Zhurinsky, J, Simcha, I, Albanese, C, D'Amico, M, Pestell, R, and Ben-Ze'ev, A; The cyclin D1 gene is a target of the beta-catenin/LEF-1 pathway, Proc. Natl. Acad. Sci. U.S.A, 1999, 96, 5522-5527; Tetsu, O and McCormick, F; Beta-catenin regulates expression of cyclin D1 in colon carcinoma cells, Nature, 1999, 398, 422-426). Збільшену експресію CCND1 зв’язували з більш високим ступенем злоякісності пухлин, метастазуванням та зниженою виживаністю (Balcerczak, E, Pasz-Walczak, G, Kumor, P, Panczyk, M, Kordek, R, Wierzbicki, R, and Mirowski, M; Cyclin D1 protein and CCND1 gene expression in colorectal cancer, Eur. J Surg. Oncol., 2005, 31, 721-726; Bahnassy, AA, Zekri, AR, El-Houssini, S, El-Shehaby, AM, Mahmoud, MR, Abdallah, S, and El-Serafi, M; Cyclin A and cyclin D1 as significant prognostic markers in colorectal cancer patients, BMC. Gastroenterol., 2004, 4, 22; McKay, JA, Douglas, JJ, Ross, VG, Curran, S, Murray, GI, Cassidy, J, and McLeod, HL; Cyclin D1 protein expression and gene polymorphism in colorectal cancer. Aberdeen Colorectal Initiative, Int. J Cancer, 2000, 88, 77-81; Maeda, K, Chung, Y, Kang, S, Ogawa, M, Onoda, N, Nishiguchi, Y, Ikehara, T, Nakata, B, Okuno, M, and Sowa, M; Cyclin D1 overexpression and prognosis in colorectal adenocarcinoma, Oncology, 1998, 55, 145-151). 15 UA 103882 C2 5 10 15 20 25 30 35 40 45 50 55 60 Матрична металопептідаза 7 (матрілизін, маточний) (MMP7) Матричні металопротеїнази (MMP) є великим сімейством структурно зв’язаних цинкозалежних протеїназ, що, як правило, описуються як здатні руйнувати компоненти екстрацелюлярного матриксу. Були ідентифіковані окремі MMP, котрі демонструють збільшену експресію в пухлинах, і в більшості пухлин спостерігається підвищена активність MMP (Curran, S and Murray, GI; 1999, Matrix metalloproteinases in tumour invasion and metastasis, J Pathol., 189, 300-308; Curran, S and Murray, GI; 2000, Matrix metalloproteinases: molecular aspects of their roles in tumour invasion and metastasis, Eur.J Cancer, 36, 1621-1630). Базальна мембрана та екстрацелюлярний матрикс представляють два фізичні бар’єри злоякісної інвазії, та їхнє руйнування MMP відіграє ключову роль в прогресуванні пухлин та поширенні метастаз (Johnsen, M, Lund, LR, Romer, J, Almholt, K, and Dano, K; 1998, Cancer invasion and tissue remodeling: common themes in proteolytic matrix degradation, Curr.Opin.Cell Biol., 10, 667-671; Nelson, AR, Fingleton, B, Rothenberg, ML, and Matrisian, LM; 2000, Matrix metalloproteinases: biologic activity and clinical implications, J Clin Oncol., 18, 1135-1149; Wang, FQ, So, J, Reierstad, S, and Fishman, DA; 2005, Matrilysin (MMP-7) promotes invasion of ovarian cancer cells by activation of progelatinase, Int.J Cancer, 114, 19-31). Крім цієї функції ММР, тепер обговорюється їхня участь в розвитку та прогресуванні пухлин, включаючи ролі в апоптозі, проліферації клітин, а також диференціації клітин. Ці функції зв’язуються з MMPопосередкованим протеолізом нематричних білків та діяльністю, відмінною від їхньої ферментної активності (Egeblad, M and Werb, Z; 2002, New functions for the matrix metalloproteinases in cancer progression, Nat.Rev.Cancer, 2, 161-174; Leeman, MF, Curran, S, and Murray, GI; 2003, New insights into the roles of matrix metalloproteinases in colorectal cancer development and progression, J.Pathol., 201, 528-534). Недавні дослідження показали, що декілька матричних металопротеїназ, в особливості, матрілізін (MMP7), взаємодіють зі специфічними молекулярними генетичними напрямками і шляхами передачі сигналів, що задіяні в розвитку колоректального раку. Зокрема, матрілізін активується на ранній стадії колоректального пухлиногенезу бета-катеніновим шляхом передачі сигналів (Brabletz, T, Jung, A, Dag, S, Hlubek, F, and Kirchner, T; 1999, beta-catenin regulates the expression of the matrix metalloproteinase-7 in human colorectal cancer, Am.J Pathol., 155, 10331038; Leeman, MF, Curran, S, and Murray, GI; 2003, New insights into the roles of matrix metalloproteinases in colorectal cancer development and progression, J.Pathol., 201, 528-534; Zucker, S and Vacirca, J; 2004, Role of matrix metalloproteinases (MMPs) in colorectal cancer, Cancer Metastasis Rev., 23, 101-117). MMP7 надмірно експресується як в доброякісних, так і в злоякісних колоректальних пухлинах (Ishikawa, T, Ichikawa, Y, Mitsuhashi, M, Momiyama, N, Chishima, T, Tanaka, K, Yamaoka, H, Miyazakic, K, Nagashima, Y, Akitaya, T, and Shimada, H; 1996, Matrilysin is associated with progression of colorectal tumor, Cancer Lett., 107, 5-10; McDonnell, S, Navre, M, Coffey, RJ, Jr., and Matrisian, LM; 1991, Expression and localization of the matrix metalloproteinase pump-1 (MMP-7) in human gastric and colon carcinomas, Mol.Carcinog., 4, 527-533; Miyazaki, K, Hattori, Y, Umenishi, F, Yasumitsu, H, and Umeda, M; 1990, Purification and characterization of extracellular matrix-degrading metalloproteinase, matrin (pump-1), secreted from human rectal carcinoma cell line, Cancer Res., 50, 7758-7764; Nagashima, Y, Hasegawa, S, Koshikawa, N, Taki, A, Ichikawa, Y, Kitamura, H, Misugi, K, Kihira, Y, Matuo, Y, Yasumitsu, H, and Miyazaki, K; 1997, Expression of matrilysin in vascular endothelial cells adjacent to matrilysin-producing tumors, Int.J Cancer, 72, 441-445; Newell, KJ, Witty, JP, Rodgers, WH, and Matrisian, LM; 1994, Expression and localization of matrix-degrading metalloproteinases during colorectal tumorigenesis, Mol.Carcinog., 10, 199-206; Yoshimoto, M, Itoh, F, Yamamoto, H, Hinoda, Y, Imai, K, and Yachi, A; 1993, Expression of MMP-7(PUMP-1) mRNA in human colorectal cancers, Int.J Cancer, 54, 614-618). MMP7 є однією з небагатьох матричних металопротеїназ, котрі фактично секретуються клітинами пухлин (Overall, CM and Kleifeld, O; 2006, Tumour microenvironment - opinion: validating matrix metalloproteinases as drug targets and anti-targets for cancer therapy, Nat.Rev.Cancer, 6, 227-239). Крім того, встановлено співвідношення рівнів експресії мРНК MMP7 зі стадією прогресування CRC (Ishikawa, T, Ichikawa, Y, Mitsuhashi, M, Momiyama, N, Chishima, T, Tanaka, K, Yamaoka, H, Miyazakic, K, Nagashima, Y, Akitaya, T, and Shimada, H; 1996, Matrilysin is associated with progression of colorectal tumor, Cancer Lett., 107, 5-10; Mori, M, Barnard, GF, Mimori, K, Ueo, H, Akiyoshi, T, and Sugimachi, K; 1995, Overexpression of matrix metalloproteinase-7 mRNA in human colon carcinomas, Cancer, 75, 1516-1519). MMP7 також грає вирішальну роль в метастазуванні CRC (Adachi, Y, Yamamoto, H, Itoh, F, Hinoda, Y, Okada, Y, and Imai, K; 1999, Contribution of matrilysin (MMP-7) to the metastatic pathway of human colorectal cancers, Gut, 45, 252-258; Mori, M, Barnard, GF, Mimori, K, Ueo, H, Akiyoshi, T, and Sugimachi, K; 1995, Overexpression of matrix 16 UA 103882 C2 5 10 15 20 25 30 35 40 45 50 55 60 metalloproteinase-7 mRNA in human colon carcinomas, Cancer, 75, 1516-1519). Підвищені рівні MMP7 у сироватці асоціюються з несприятливим прогнозом у пацієнтів з запущеним колоректальним раком (Maurel, J, Nadal, C, Garcia-Albeniz, X, Gallego, R, Carcereny, E, Almendro, V, Marmol, M, Gallardo, E, Maria, AJ, Longaron, R, Martinez-Fernandez, A, Molina, R, Castells, A, and Gascon, P; 2007, Serum matrix metalloproteinase 7 levels identifies poor prognosis advanced colorectal cancer patients, Int.J Cancer, Published Online: 8 May 2007), та було висловлено думку, що надмірна експресія MMP7 у пацієнтів з CRC, що асоціюється із знижною виживаністю, також призводить до уникнення імунного контролю внаслідок розщеплення Fas на пухлинах клітин (Wang, WS, Chen, PM, Wang, HS, Liang, WY, and Su, Y; 2006, Matrix metalloproteinase-7 increases resistance to Fas-mediated apoptosis and is a poor prognostic factor of patients with colorectal carcinoma, Carcinogenesis, 27, 1113-1120). Білки винаходу можуть бути мішенню пухлино-специфічної імунної реакції при багатьох видах ракових захворювань. Пептид HBV-001 внутрішнього антигену вірусу Гепатиту B не має походження з ендогенного пухлино-асоційованого антигену людини, а одержується з внутрішнього антигену вірусу Гепатиту B. По-перше, він дозволяє кількісно порівняти величину T-клітинних реакцій, індукованих TUMAP, і таким чином, зробити важливі висновки щодо здатності викликати протипухлинні реакції. По-друге, він функціонує як важливий позитивний контроль за відсутністю будь-яких T-клітинних реакцій у пацієнта. По-третє, він також дозволяє визначити статус імунокомпетентності пацієнта. Інфекція, викликана вірусом Гепатиту B (HBV), є одною з основних причин захворювання печінки, від якого страждають приблизно 350 мільйонів людей в усьому світі (Rehermann, B and Nascimbeni, M; Immunology of hepatitis B virus and hepatitis C virus infection, Nat.Rev.Immunol., 2005, 5, 215-229). Внаслідок легкості передачі горизонтальним та вертикальним шляхом та потенціалу переходу у хронічну форму, що може призвести до цирозу печінки та гепатоклітинної карциноми, HBV завдає серйозного впливу системі охорони здоров’я в багатьох країнах світу. Геном HBV (Previsani, N and Lavanchy, D; 2002, Hepatitis B, (Epidemic and Pandemic Alert and Response, World Health Organization, Geneva, 2002)) складається з частково двониткової кільцевої ДНК. У віріонах HBV він компонується разом із капсидним білком HBc та іншими білками для формування нуклео-капсиду, що знаходиться в зовнішній оболонці, яка вміщує ліпіди та HBs сімейства поверхневих білків (також називається «оболонковий білок»). Антигенні детермінанти, що асоціюються з HBc та HBs, відомі як HBcAg та HBsAg, відповідно. Ці антигени зв’язані з серологічними реакціями, тобто, реакціями антитіл, що визначаються у крові пацієнтів та є одними з найбільш клінічно прийнятних систем антигенів-антитіл для діагностування HBV-інфекції. HBc буде представляти новий чужорідний антиген для усіх осіб, які не мають попередньої історії HBV-інфекції. Оскільки імуногенні пептиди добре відомі для цього антигену (Bertoletti, A, Chisari, FV, Penna, A, Guilhot, S, Galati, L, Missale, G, Fowler, P, Schlicht, HJ, Vitiello, A, Chesnut, RC, and .; 1993, Definition of a minimal optimal cytotoxic T-cell epitope within the hepatitis B virus nucleocapsid protein, J.Virol., 67, 2376-2380; Livingston, BD, Crimi, C, Grey, H, Ishioka, G, Chisari, FV, Fikes, J, Grey, H, Chesnut, RW, and Sette, A; 1997, The hepatitis B virus-specific CTL responses induced in humans by lipopeptide vaccination are comparable to those elicited by acute viral infection, J.Immunol., 159, 1383-1392), один пептид з десяти амінокислот з HBcAg був обраний як позитивний контроль антигену в IMA. Індукування HBc-пептидо-специфічних CTL буде згодом використовуватись як маркер імунокомпетентності пацієнта та успішної вакцинації. Фармацевтична композиція може, крім того, вміщувати додаткові пептиди та/або наповнювачі для збільшення ефективності, як пояснюється нижче. Під терміном «варіант амінокислотної послідовності» даної амінокислотної послідовності автори винаходу мають на увазі, що бокові ланцюжки, наприклад, одного чи двох амінокислотних залишків змінюються (наприклад, шляхом заміни їх боковим ланцюжком іншого природно існуючого амінокислотного залишку чи якимось іншим боковим ланцюжком) таким чином, що пептид все ще здатний зв’язуватися з молекулою HLA, по суті, в такий же спосіб, як і пептид, що складається з даної амінокислотної послідовності. Наприклад, пептид може бути модифікований так, що він буде якнайменше зберігати, чи навіть поліпшувати, здатність взаємодіяти та зв’язуватися з прийнятною молекулою MHC, такою як HLA-A або -DR, та якнайменше зберігати, чи навіть поліпшувати, здатність утворювати активований CTL, що може розпізнавати та та знищувати клітини, котрі експресують поліпептид, вміщуючий амінокислотну послідовність, як визначено в аспектах винаходу. Як можна дізнатися з бази даних, окремі позиції HLA-A-зв’язуючих пептидів є, зазвичай, доменними залишками, що формують ключову послідовність, яка відповідає за зв’язок у порожнини зв’язування HLA. 17 UA 103882 C2 5 10 15 20 25 30 35 40 45 50 55 60 Ті амінокислотні залишки, що не суттєвими для взаємодії з T-клітинним рецептором, можуть бути модифіковані шляхом заміни іншою амінокислотою, введення якої не має значного впливу на реактивність Т-клітин та не виключає зв’язування із відповідним МНC. Отже, за винятком зазначеної умови, пептид винаходу може бути будь-яким пептидом (до цього терміну автори винаходу відносять олігопептид чи поліпептид), котрий включає амінокислотні послідовності або їхню частину чи варіант, що надається. Стосовно МНС-презентуючих пептидів (класу II) відомо, що ці пептиди складаються з «ключової послідовності», яка має певний HLA-специфічний амінокислотний фрагмент та, додатково, N- та/або C-термінальні подовження, які не втручаються у функцію ключової послідовності (тобто, вважаються не релевантними до взаємодії пептиду і T-клітини). N- та/або C-термінальні подовження можуть мати довжину, наприклад, від 1 до 10 амінокислот, відповідно. Ці пептиди можуть використовуватись безпосередньо для завантаження MHCмолекул класу II, або послідовність може клонуватись у вектори, згідно із наведеним нижче описом. Оскільки ці пептиди формують кінцевий продукт обробки більш крупних пептидів в межах клітини, також можуть використовуватися довші пептиди. Пептиди винаходу можуть мати будь-який розмір, але, як правило, вони можуть бути меншими, ніж 100 000, за молекулярною вагою, переважно, меншими, ніж 50 000, ще більш переважно меншими, ніж 10 000 та, зазвичай, приблизно дорівнюючими 5 000. Стосовно кількості амінокислотних залишків, пептиди винаходу можуть мати менше 1000 залишків, переважно менше, ніж 500 залишків, та ще більш переважно - менше 100 залишків. Відповідно, цей винахід також надає композиції пептидів та їхні варіанти, в яких пептид чи варіант має загальну довжину від 8 до 100, переважно, від 8 до 30, та найбільш переважно - від 8 до 16, а саме, 8, 9, 10, 11, 12, 13, 14, 15 чи 16 амінокислот. Відповідно, варіанти, що індукують перехресне реагування T-клітин з пептидом винаходу, часто є варіантами з різною довжиною. Якщо пептид, більший ніж приблизно 12 амінокислотних залишків, використовується безпосередньо для зв’язування з MHC-молекулою класу ІІ, бажано, щоб залишки, розташовані по краях основної ділянки зв’язування HLA, не мали значного впливу на здатність пептиду специфічно зв’язуватися з порожниною зв’язування MHC-молекули класу ІІ чи презентувати пептид в CTL. Однак, як вже зазначалося вище, перевага надається використанню більш крупних пептидів, особливо, коли вони кодуються полінуклеотидом, оскільки ці більш крупні пептиди можуть розділятися на частини відповідними антиген-презентуючими клітинами. Також можливо, щоб епітопи MHC класу I, хоча вони мають, зазвичай, довжину 8-10 амінокислот, утворювались шляхом процесингу пептидів з більш довгих пептидів чи білків, які включають фактичний епітоп. Подібно епітопам MHC класу II, бажано, щоб залишки, розташовані по краях ділянки зв’язування, не мали значного впливу на здатність пептиду специфічно зв’язуватися з порожниною зв’язування MHC-молекули класу I чи презентувати пептид в CTL, і не маскували ділянки для протеолітичного розпаду, необхідні для експозиції фактичного епітопу в ході процесингу. Відповідно, цей винахід також пропонує пептиди та варіанти епітопів MHC класу I, які мають загальну довжину від 8 до 100, переважно від 8 до 30, та найбільш переважно від 8 до 16, а саме, 8, 9, 10, 11, 12, 13, 14, 15 або 16 амінокислот. Звичайно, пептид чи варіант, відповідно до цього винаходу, матимуть здатність зв’язуватися з молекулою головного комплексу гістосумісності людини (MHC) класу I чи II. Зв’язування пептиду чи варіанту з MHC-комплексом може бути перевірене за допомогою методів, добре відомих в цій галузі, наприклад, описаних в наведених нижче прикладах цього винаходу чи в довідковій літературі для різних алелів MHC класу II (наприклад, в роботах Vogt AB, Kropshofer H, Kalbacher H, Kalbus M, Rammensee HG, Coligan JE, Martin R; Ligand motifs of HLA-DRB5*0101 and DRB1*1501 molecules delineated from self-peptides; J Immunol. 1994; 153(4):1665-1673; Malcherek G, Gnau V, Stevanovic S, Rammensee HG, Jung G, Melms A; Analysis of allele-specific contact sites of natural HLA-DR17 ligands; J Immunol. 1994; 153(3):1141-1149; Manici S, Sturniolo T, Imro MA, Hammer J, Sinigaglia F, Noppen C, Spagnoli G, Mazzi B, Bellone M, Dellabona P, Protti MP; Melanoma cells present a MAGE-3 epitope to CD4(+) cytotoxic T cells in association with histocompatibility leukocyte antigen DR11; J Exp Med. 1999; 189(5): 871-876; Hammer J, Gallazzi F, Bono E, Karr RW, Guenot J, Valsasnini P, Nagy ZA, Sinigaglia F; Peptide binding specificity of HLADR4 molecules: correlation with rheumatoid arthritis association; .J Exp Med. 1995 181(5):1847-1855; Tompkins SM, Rota PA, Moore JC, Jensen PE; A europium fluoroimmunoassay for measuring binding of antigen to class II MHC glycoproteins; J Immunol Methods. 1993;163(2): 209-216; Boyton RJ, Lohmann T, Londei M, Kalbacher H, Halder T, Frater AJ, Douek DC, Leslie DG, Flavell RA, Altmann DM; Glutamic acid decarboxylase T lymphocyte responses associated with susceptibility or resistance to type I diabetes: analysis in disease discordant human twins, non-obese diabetic mice and HLA-DQ 18 UA 103882 C2 5 10 15 20 25 30 35 40 45 50 55 60 transgenic mice; Int Immunol. 1998 (12):1765-1776). Додаткові N- та/або C-термінальні подовження амінокислот, які не обов’язково формують частину пептиду, що функціонує як епітоп для MHC-молекул, можуть, незважаючи на це, бути важливими для забезпечення ефективного введення пептиду, відповідно до винаходу, в клітини. В одному з втілень цього винаходу, пептид цього винаходу є гібридним білком, який вміщує, наприклад, 80 N-термінальних амінокислот HLA-DR-антиген-асоційованого інваріантного ланцюгу (p33, надалі - ”Ii”), одержаного з NCBI, інвентарний номер в генному банку (GenBank) X00497 (Strubin, M., Mach, B. and Long, E.O. The complete sequence of the mRNA for the HLA-DR-associated invariant chain reveals a polypeptide with an unusual transmembrane polarity EMBO J. 3 (4), 869-872 (1984)). Переважною є фармацевтична композиція, в якій пептиди мають загальну довжину від 8 до 100, переважно від 8 до 30, та найбільш переважно від 8 до 16 амінокислот. Крім того, пептид чи варіант може бути додатково модифікований для поліпшення його стабільності та/або зв’язку із молекулами MHC, щоб викликати сильнішу імунну реакцію. Методи для такої оптимізації пептидної послідовності добре відомі в цій галузі та включають, наприклад, введення реверсованих пептидних зв’язків чи непептидних зв’язків. Отже, відповідно до іншого аспекту, винахід пропонує фармацевтичну композицію, в якій принаймні один пептид чи варіант включає непептидні зв’язки. В реверсованому пептидному зв’язку амінокислотні залишки не з’єднуються пептидними зв’язками (-CO-NH-), а пептидний зв’язок реверсується. Такі ретро-інверсивні пептидні міметики можуть вироблятися за методами, відомими в даній галузі, наприклад, описаними в роботі Meziere et al (1997) J. Immunol. 159, 3230-3237, яка включена в даний документ шляхом посилання. Такий підхід включає формування псевдо-пептидів, які вміщують зміни із залученням основи, а не орієнтації бокових ланцюгів. Автори Meziere et al (1997) показують, що для реакцій клітин MHC і T-клітин - хелперів зазначені псевдо-пептиди є прийнятними. Ретроінверсивні пептиди, які вміщують зв’язки NH-CO замість пептидних зв’язків CO-NH, набагато більш стійкі до протеолізу. Непептидний зв’язок - це, наприклад, -CH2-NH, -CH2S-, -CH2CH2-, -CH=CH-, -COCH2-, CH(OH)CH2- та -CH2SO-. Патент США 4,897,445 пропонує метод твердофазного синтезу непептидних зв’язків (-CH2-NH) в поліпептидних ланцюгах, що включає поліпептиди, синтезовані за допомогою стандартних процедур, та непептидний зв'язок, синтезований шляхом реакції аміноальдегіду та амінокислоти в присутності NaCNBH3. Пептиди, що включають послідовності винаходу, як описано вище, можуть бути синтезовані з додатковими хімічними групами, присутніми на їхніх аміно- та/або карбоксильних кінцях, зокрема, для посилення стабільності, біологічної доступності та/або афінності пептидів. Наприклад, гідрофобні групи, такі як карбобензоксильні, данзильні чи t-бутилоксикарбонільні групи можуть додаватися до аміно-кінців пептидів. Подібним чином, на аміно-кінцях пептидів може розміщуватись ацетильна група чи 9-фторенілметокси-карбонільна група. До карбоксильних кінців пептидів може бути додана також, наприклад, гідрофобна група, tбутилоксикарбонільна чи амідо-група. Крім того, всі пептиди винаходу можуть бути синтезовані для зміни їхньої стеричної конфігурації. Наприклад, може використовуватися D-ізомер одного чи кількох амінокислотних залишків пептиду, а не звичайний L-ізомер. До того ж, принаймні один з амінокислотних залишків пептидів винаходу може замінюватись одним з добре відомих амінокислотних залишків не природного походження. Такі заміни можуть служити для підвищення стабільності, біологічної доступності та/або функції зв’язування пептидів винаходу. Подібним чином, пептид чи варіант винаходу може модифікуватися хімічно, шляхом реакції окремих амінокислот до чи після синтезу пептиду. Приклади таких модифікацій добре відомі в цій галузі та узагальнені, зокрема, в роботі R. Lundblad, Chemical Reagents for Protein Modification, 3rd ed. CRC Press, 2005, яка включена в цей документ шляхом посилання. Хімічна модифікація амінокислот включає, без обмежень, модифікацію шляхом ацилювання, амідинування, піридоксилювання лізину, відновлювального алкілювання, тринітробензилювання аміногруп 2, 4, 6-тринітробензол-сульфоновою кислотою (TNBS), амідну модифікацію карбоксильних груп та сульфгідрильну модифікацію шляхом окиснення цистеїну надмурашиною кислотою в цистеїнову кислоту, формування похідних ртуті, утворення змішаних дисульфідів з іншими тіольними сполуками, реакцію з імідом малеїнової кислоти, карбоксиметилювання йодооцтовою кислотою чи йодоацетамідом та карбамоїлування цианатом при лужному pH, хоча способи модифікації не обмежуються наведеними вище. В цьому відношенні, досвідчений спеціаліст повинен звернутися до Глави 15 роботи Current Protocols In Protein Science, Eds. Coligan et al. (John Wiley & Sons NY 1995-2000), де викладена детальна методика відносно 19 UA 103882 C2 5 10 15 20 25 30 35 40 45 50 55 60 хімічної модифікації білків. Стисло кажучи, модифікація, наприклад, аргінілових залишків в білках часто базується на реакції віцинальних дикарбонілових сполук, таких як фенілгліоксаль, 2,3-бутанедіон та 1,2циклогексанедіон, для утворення адукту. Ще один приклад - це реакція метилгліоксалю з аргініновими залишками. Цистеїн може модифікуватися без супутньої модифікації інших нуклеофільних ділянок, таких як лізин і гістидин. В результаті, доступна велика кількість реактивів для модифікації цистеїну. Веб-сайти Pierce Chemical Company, Sigma-Aldrich та інших надають інформацію про окремі реактиви. Типовим також є селективне відновлення дисульфідних зв’язків в білках. Дисульфідні зв’язки можуть формуватися та окиснюватися при тепловій обробці біофармацевтичних препаратів. К-реагент Вудварда може використовуватися для модифікації окремих залишків глутамінової кислоти. N-(3-(діметиламіно)пропіл)-N’-етилкарбодиімід може бути застосований для утворення внутрішньомолекулярних крос-лінків між лізиновим залишком та залишком глутамінової кислоти. Діетилпірокарбонат, наприклад, є реактивом для модифікації гістидилових залишків в білках. Гістидин також може бути модифікований із використанням 4-гідрокси-2-ноненалу. Реакція лізинових залишків та інших -аміно-груп є, наприклад, прийнятною у зв’язуванні пептидів з поверхнями або перехресному зшиванні білків/пептидів. Лізин - це місце приєднання полі(етилен)гліколю та основна ділянка модифікації при глікуванні білків. Метіонінові залишки в білках можуть бути модифіковані, наприклад, за допомогою йодоацетаміду, бромоетиленаміну, хлораміну T. Тетранітрометан та N-ацетилімідазол можуть використовуватися для модифікації тирозильних залишків. Перехресне зшивання шляхом формування дитирозину може здійснюватися за допомогою пероксиду водню/іонів міді. В недавніх дослідженнях щодо модифікації триптофану використовувався Nбромосуккінімід, 2-гідрокси-5-нітробензил-бромід чи 3-бромо-3-метил-2-(2-нітрофенілмеркапто)3H-індол (BPNS-скатол). Успішна модифікація PEG (поліетиленгліколем) терапевтичних білків та пептидів, що часто асоціюється з подовженням полу-періоду циркуляції при перехресному зшиванні білків з глютаральдегідом, поліетиленгліколь-діакрилатом та формальдегідом, використовується для приготування гідрогелів. Хімічна модифікація алергенів для імунотерапії часто досягається шляхом карбамоїлування з ціанатом калію. В цілому, пептиди та варіанти (принаймні, ті, що вміщують пептидні зв’язки між амінокислотними залишками) можуть бути синтезовані, наприклад, із використанням Fmocполіамідного способу синтезу пептидів твердої фази, як розкривається в роботі Lu et al (1981) J. Org. Chem. 46, 3433 та посиланнях на літературні джерела, наведених в ній. Очищення може виконуватися за допомогою будь-якого одного чи кількох методів, таких як рекристалізація, гель-хроматографія, іонообмінна хроматографія, хроматографія гідрофобної взаємодії та (зазвичай) зворотно-фазна високоефективна рідинна хроматографія із градієнтним розділенням, наприклад, ацетонітріл/ вода. Аналіз пептидів може виконуватися за допомогою тонкошарової хроматографії, електрофорезу, зокрема, капілярного електрофорезу, твердофазної екстракції (CSPE), зворотно-фазної високоефективної рідинної хроматографії, амінокислотного аналізу після гідролізу кислот та мас-спектрометричного аналізу аналізу із бомбардуванням прискореними атомами (FAB), а також мас-спектрометричного аналізу MALDI та ESI-Q-TOF. Ще один аспект винаходу пропонує нуклеїнову кислоту (наприклад, полінуклеотид), що кодує пептид чи варіант винаходу. Полінуклеотид, наприклад, може бути ДНК, кДНК, ПНК, СНК, РНК, одно- та/або двонитковою, природними чи стабілізованими формами полінуклеотидів, такими як, наприклад, полінуклеотиди з фосфоротіатним скелетом, чи їхніми комбінаціями; він може вміщувати або не вміщувати інтрони, за умови, що полінуклеотид кодує пептид. Звичайно, тільки ті пептиди, що вміщують природно існуючи амінокислотні залишки, з’єднані природними пептидними зв’язками, кодуються полінуклеотидом. Інший аспект винаходу пропонує вектор експресії, здатний експресувати поліпептид, відповідно до винаходу. Вектори експресії для різних типів клітин добре відомі в цій галузі та можуть обиратися без проведення зайвих експериментів. В цілому, ДНК вводиться у вектор експресії, такий як плазміда, в належній орієнтації та з відповідною рамкою зчитування для експресії. При необхідності, ДНК може зв’язуватися з належними нуклеотидними послідовностями транскрипційного і трансляційного регуляторного контролю, що розпізнаються бажаним хазяїном, хоча такі контрольні елементи взагалі є 20 UA 103882 C2 5 10 15 20 25 30 35 40 45 50 55 60 доступними у векторі експресії. Вектор потім вводиться хазяїну із використанням стандартних методик. Рекомендації можна знайти, наприклад, в роботі Sambrook et al (1989) Molecular Cloning, A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor, NY. Однак, в особливо переважному втіленні винаходу фармацевтична композиція вміщує принаймні два пептиди, які складаються з амінокислотних послідовностей, згідно із послідовностями від SEQ ID NO 1 до SEQ ID NO 15. Оптимальна кількість кожного пептиду, що включається у вакцину, та оптимальний режим дозування може визначатися досвідченим спеціалістом, без проведення зайвих експериментів. Наприклад, пептид чи його варіант може бути підготовлений для внутрішньовенного (i.v.) введення, підшкірного (s.c.) введення, інтрадермального (i.d.) введення, внутрішньочеревного (i.p.) введення, внутрішньом’язового (i.m.) введення. Переважні способи введення пептиду - це s.c., i.d., i.p., i.m. та i.v. Переважними способами введення ДНК є i.d., i.m., s.c., i.p. та i.v. Можуть вводитись, наприклад, дози від 1 до 500 мг, від 50 мкг до 1.5 мг, переважно, від 125 мкг до 500 мкг, пептиду чи ДНК, залежно від відповідного пептиду чи ДНК. Дози в цьому діапазоні успішно використовувались в попередніх дослідженнях (Brunsvig PF, Aamdal S, Gjertsen MK, Kvalheim G, Markowski-Grimsrud CJ, Sve I, Dyrhaug M, Trachsel S, Møller M, Eriksen JA, Gaudernack G;. Telomerase peptide vaccination: a phase I/II study in patients with non-small cell lung cancer; Cancer Immunol Immunother. 2006; 55(12):1553-1564; M. Staehler, A. Stenzl, P. Y. Dietrich, T. Eisen, A. Haferkamp, J. Beck, A. Mayer, S. Walter, H. Singh, J. Frisch, C. G. Stief; An open label study to evaluate the safety and immunogenicity of the peptide based cancer vaccine IMA901, ASCO meeting 2007; Abstract No 3017) Фармацевтична композиція винаходу може бути складена таким чином, щоб вибір, число та/або кількість пептидів, присутніх у композиції, були специфічними для тканин, ракових захворювань та/або окремих пацієнтів. Наприклад, точний вибір пептидів може керуватися профілями експресії вихідних білків в даній тканини, щоб уникнути побічних ефектів. Вибір може залежати від специфічного виду раку, на який хворіє пацієнт, що проходить лікування, а також стану захворювання, попередніх режимів лікування, імунного статусу пацієнта і, звичайно, HLAгаплотипу пацієнта. Крім того, вакцина, згідно із винаходом, може вміщувати індивідуалізовані компоненти, відповідно до особистих потреб конкретного пацієнта. Прикладами є різні кількості пептидів, згідно із експресією відповідних TAA у конкретного пацієнта, небажаними побічними ефектами внаслідок особистої алергії чи інших режимів лікування, а також корегування для другого курсу після проходження першого циклу чи курсу лікування. Для того, щоб, наприклад, композиція використовувалася як вакцина проти CRC, пептиди, вихідні білки яких експресуються у великих кількостях в нормальних тканинах, уникаються в композиції винаходу чи присутні в невеликих кількостях. З іншої сторони, коли відомо, що пухлина пацієнта експресує великі кількості окремного білку, відповідна фармацевтична композиція для лікування цього випадку раку може презентуватися у великих кількостях та/або може включатися більш ніж один пептид, специфічний для цього конкретного білку чи його шляху передачі сигналів. Досвідчений фахівець зможе обрати переважні комбінації імуногенних пептидів шляхом тестування, наприклад, генерації T-клітин in vitro, а також їхньої ефективності та загальної присутності, проліферації, афінності та розмноження окремих T-клітин для певних пептидів, і функціональності T-клітин, наприклад, аналізуючи утворення IFN- (див.також приклади нижче). Зазвичай, найбільш ефективні пептиди згодом комбінуються як вакцина в цілях, описаних вище. Прийнятна вакцина буде переважно вміщувати від 1 до 20 пептидів, більш переважно, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 чи 20 різних пептидів, ще більш переважно 6, 7, 8, 9, 10 11, 12, 13 або 14 різних пептидів та найбільш переважно 14 різних пептидів. Довжина пептиду для використання в протираковій вакцині може бути будь-яким прийнятним пептидом. Зокрема, це може бути прийнятний 9-мерний пептид чи прийнятний 7-мерний або 8-, 10-, 11-, 12-, 13-мерний, 14-мерний чи 15-мерний пептид. Пептиди більшої довжини також можуть використовуватись, але 9-мерні чи 10-мерні пептиди, як описано в доданих таблицях 1 і 2, є переважними для пептидів MHC класу I, в той час як пептиди з 12- до 15-мерних переважні для пептидів MHC класу II. Пептид(и) становить(становлять) протипухлинну чи протиракову вакцину. Вона може вводитись безпосередньо пацієнту в уражений орган чи системно, або застосовуватись ex vivo до клітин, виділених з пацієнта або клітинної лінії людини, що згодом вводяться пацієнту, чи використовуватись in vitro для обирання субпопуляції з імунних клітин, які виділяються з пацієнта і потім знов вводяться йому. 21 UA 103882 C2 5 10 15 20 25 30 35 40 45 50 55 60 Пептид може бути, по суті, чистим, або поєднаним з імуно-стимулюючим ад’ювантом (див. нижче) чи використовуватись в комбінації з імунно-стимулюючими цитокінами, або вводитись з належною системою доставки, наприклад, ліпосомами. Пептид також може поєднуватись з належним носієм, таким як гемоцианін лімфи равлика (KLH) або маннан (див. WO 95/18145 та Longenecker et al (1993) Ann. NY Acad. Sci. 690,276-291). Пептид також може бути міченим, і бути гібридним білком чи гібридною молекулою. Очікується, що пептиди, послідовність яких надається в цьому винаході, стимулюють CD4 або CD8 CTL. Однак, стимуляція більш ефективна у присутності підтримки, яка надається T-клітинами, позитивними до протилежного CD. Отже, для епітопів MHC Класу II, які стимулюють CD4 CTL, гібридний партнер або частини гібридної молекули належним чином надають епітопи, що стимулюють CD8-позитивні T-клітини. З іншої сторони, для епітопів MHC Класу I, які стимулюють CD8 CTL, гібридний партнер або частини гібридної молекули належним чином надають епітопи, котрі стимулюють CD4-позитивні T-клітини. CD4- та CD8-стимулюючі епітопи добре відомі в цій галузі та включають епітопи, ідентифіковані в цьому винаході. Для викликання імунної реакції, зазвичай необхідно включення наповнювачів, що надають композиції більшої імуногенності. Тобто, в переважному втіленні винаходу фармацевтична композиція також вміщує принаймні один прийнятний ад’ювант. Ад’ювантами є речовини, котрі неспецифічно збільшують чи посилюють імунну реакцію (наприклад, імунні реакції, де посередниками виступають CTL і Т-клітини-хелпери (TH) на антиген, отже, вони вважаються доцільними для медикаменту цього винаходу. Прийнятні ад’юванти включають, без обмежень, 1018 ISS, солі алюмінію, Amplivax, AS15, БЦЖ, CP870,893, CpG7909, CyaA, dSLIM, GM-CSF, IC30, IC31, Іміквімод, ImuFact IMP321, IS Patch, ISS (імуно-стимулюючі послідовності), ISCOMATRIX, JuvImmune, LipoVac, MF59, монофосфориловий ліпід A, Монтанід IMS 1312, Монтанід ISA 206, Монтанід ISA 50V, Монтанід ISA-51, OK-432, OM-174, OM-197-MP-EC, ONTAK, векторну систему PepTel®, мікрочастинки PLG, резіквімод, SRL172, Віросоми та інші вірусоподібні частинки, YF-17D, VEGF trap, R848, бета-глюкан, Pam3Cys, стимулон QS21 Aquila (Aquila Biotech, Ворчестер, Масачусетс, США), який виділяється з сапоніну, мікобактеріальні екстракти та синтетичні імітатори стінок бактеріальних клітин, а також інші патентовані ад’юванти, наприклад, Ribi's Detox, Quil або Superfos. Переважними є такі ад’юванти, як неповний ад’ювант Фрейнда або GM-CSF. Кілька імунологічних ад’ювантів (наприклад, MF59), специфічних для дендритних клітин, та їхнє приготування, були описані раніше (Dupuis M, Murphy TJ, Higgins D, Ugozzoli M, van Nest G, Ott G, McDonald DM; Dendritic cells internalize vaccine adjuvant after intramuscular injection; Cell Immunol. 1998; 186(1):18-27; Allison AC; The mode of action of immunological adjuvants; Dev Biol Stand. 1998; 92:3-11). Також можуть застосовуватися цитокіни. Окремі цитокіни були прямо співвіднесені з впливом на міграцію дендритних клітин до лімфоїдних тканин (наприклад, TNF), прискорюючи перетворення дендритних клітин в ефективні антиген-презентуючі клітини для T-лімфоцитів (наприклад, GM-CSF, IL-1 та IL-4) (Патент США №5,849,589, окремо включений в цей документ шляхом посилання в усій повноті) та діючи як імуно-ад’юванти (наприклад, IL-12) (Gabrilovich DI, Cunningham HT, Carbone DP; IL-12 and mutant P53 peptide-pulsed dendritic cells for the specific immunotherapy of cancer; J Immunother Emphasis Tumor Immunol. 1996 (6):414418). Також доповідалося про те, що імуно-стимулюючі CpG-олігонуклеотиди посилюють ефект ад’ювантів у вакцинному складі. Теоретично, не будучи зв’язаними, CpG олігонуклеотиди діють шляхом активації природної (не здобутої) імунної системи за допомогою Toll-подібних рецепторів (TLR), переважно TLR9. Активація TLR9, ініційована CpG, посилює антигенспецифічні гуморальні та клітинні реакції на широкий спектр антигенів, включаючи пептидні чи білкові антигени, живі або знищені віруси, вакцини на основі дендритних клітин, аутологічні клітинні вакцини та полісахаридні кон’югати в профілактичних і терапевтичних вакцинах. Більш важливо те, що така активація посилює визрівання та диференціацію дендритних клітин, що в результаті збільшує активацію T H1-клітин та генерацію цитотоксичних T-лімфоцитів (CTL), навіть за відсутності допомоги CD4 T-клітин. Активація TH1, індукована стимуляцією TLR9, зберігається навіть у присутності вакцинних ад’ювантів, таких як солі алюмінію чи неповний ад’ювант Фрейнда (IFA), котрі зазвичай сприяють активації TH2. CpG-олігонуклеотиди демонструють навіть більшу ад’ювантну активність, коли формулюються або вводяться разом з іншими ад’ювантами чи в таких формуляціях, як мікрочастинки, нано-частинки, ліпідні емульсії або подібні формуляції, особливо необхідні для індукування сильної реакції, коли антиген відносно слабкий. Вони також прискорюють імунну реакцію та дозволяють зменшити дози антигену приблизно на два порядки, в порівнянні з реакціями антитіл на вакцину в повній дозі без CpG в деяких експериментах (Arthur M. Krieg, Therapeutic potential of Toll-like receptor 9 22 UA 103882 C2 5 10 15 20 25 30 35 40 45 50 55 60 activation, Nature Reviews, Drug Discovery, 2006, 5, 471-484). В Патенті США 6,406,705 B1 описується комбіноване використання CpG-олігонуклеотидів, ад’ювантів, що не вміщують нуклеїнових кислот, та антигену для індукування антиген-специфічної імунної реакції. Наявним у продажу CpG TLR9-антагоністом є dSLIM (імуномодулятор з подвійно-нитчастим контуром) фірми Mologen (Берлін, Німеччина), котрий є переважним компонентом фармацевтичної композиції цього винаходу. Також можуть використовуватись інші TLR-зв’язувальні молекули, наприклад, TLR 7, TLR 8 та/або TLR 9, зв’язані з РНК. Інші приклади прийнятних ад’ювантів включають, без обмежень, хімічно модифіковані CpGs (наприклад, CpR, Idera), Poly(I:C) (наприклад, polyI:C12U), бактеріальні ДНК або РНК, відмінні від CpG, а також невеликі імуноактивні молекули та антитіла, такі як імідазокіноліни, циклофосфамід, сунітініб (sunitinib), бевацизумаб, целебрекс (celebrex), NCX-4016, сілденафіл (sildenafil), тадалафіл (tadalafil), варденафіл (vardenafil), сорафеніб (sorafinib), XL-999, CP547632, пазопаніб (pazopanib), ZD2171, AZD2171, іпілімумаб (ipilimumab), тремелімумаб (tremelimumab) та SC58175, котрі можуть діяти терапевтично та/або як ад’ювант. Кількості та концентрації ад’ювантів та добавок, прийнятих в контексті цього винаходу, можуть бути легко визначені досвідченим спеціалістом без неналежного експериментування. Переважними ад’ювантами є dSLIM, БЦЖ, OK432, іміквімод, PeviTer та JuvImmune. В переважному втіленні фармацевтичної композиції, відповідно до винаходу, ад’ювант обирається з групи, що складається з факторів стимулювання колоній, таких як Фактор Стимулювання Колоній Гранулоцитів-макрофагів (GM-CSF, сарграмостим). В переважному втіленні фармацевтичної композиції, відповідно до винаходу, ад’ювантом є іміквімод. Ця композиція використовується для парентерального введення, наприклад, підшкірного, інтрадермального, внутрішньом’язового чи перорального введення. Для цього пептиди і додатково інші молекули розчиняються або суспендуються у фармацевтично прийнятному, переважно водному, носії. До того ж, композиція може вміщувати наповнювачі, такі як буферні елементи, зв’язувальні речовини, очисники, розріджувачі, ароматизатори, мастильні речовини та ін. Пептиди також можуть вводитись разом із імуностимулюючими речовинами, наприклад, цитокінами. Вичерпний перелік наповнювачів, які можуть використовуватись у такій композиції, наведений, наприклад, в книзі A. Kibbe, Handbook of Pharmaceutical Excipients, 3. Ed. 2000, American Pharmaceutical Association and pharmaceutical press. Композиція може використовуватися для попередження, профілактики та/або терапії аденомних чи ракових захворювань, переважно CRC. Цитотоксичні T-клітини (CTL) розпізнають антиген у формі пептиду, зв’язаного з MHCмолекулою, а не неушкоджений інорідний антиген. Сама молекула MHC знаходиться на клітинній поверхні антиген-презентуючої клітини. Отже, активація CTL можлива тільки в тому випадку, якщо присутній тримерний комплекс пептидного антигену, MHC-молекули та АPC. Відповідно, імунна реакція може посилюватись, якщо для активації CTL використовується не тільки пептид, але також APC з відповідною молекулою MHC. Отже, у переважному втіленні фармацевтична композиція, відповідно до цього винаходу, додатково вміщує принаймні одну антиген-презентуючу клітину. Антиген-презентуюча клітина (чи клітина-стимулятор), як правило, має на своїй поверхні MHC-молекулу класу I або II та в одному з втілень винаходу вона переважно не здатна, сама по собі, завантажувати зазначену MHC-молекулу класу I або II обраним антигеном. Як описується більш детально нижче, MHC-молекула класу I або II може легко завантажуватись обраним антигеном in vitro. Переважно, клітина ссавців не має пептидного транспортера ТАР, чи має його знижений рівень або зменшену функцію. Відповідні клітини, в яких немає пептидного транспортера ТАР, включають T2, RMA-S та клітини дрозофіли. TAР - це Транспортер, асоційований з процесінгом антигенів. Клітинна лінія Т2, яка не здатна завантажувати пептиди людини, є доступною для придбання в American Type Culture Collection, 12301 Parklawn Drive, Rockville, Меріленд 20852, США, номер за каталогом CRL 1992; лінія клітин дрозофіли, лінія Schneider 2, доступна для придбання в ATCC, номер за каталогом CRL 19863; клітинна лінія миші RMA-S описана авторами Karre та Ljunggren (1985) J. Exp. Med. 162,1745. Ці клітинні лінії можуть використовуватись як APC, та внаслідок відсутності TAP приблизно усі пептиди, презентовані MHC класу I, будуть пептидами, що вивчаються з метою зовнішнього завантаження порожніх MHC-молекул класу I цих клітинних ліній, оскільки усі ефекти будуть чітко приписуватись використовуваним пептидам. Переважно, антиген-презентуючі клітини є дендритними клітинами. Відповідно, дендритні 23 UA 103882 C2 5 10 15 20 25 30 35 40 45 50 55 60 клітини є аутологічними дендритними клітинами, які імпульсним методом завантажуються антигенним пептидом. Антигенний пептид може бути належним антигенним пептидом, який викликає відповідну T-клітинну реакцію. Т-клітинна терапія із використанням аутологічних дендритних клітин, в які імпульсним методом завантажуються пептиди з пухлино-асоційованого антигену, розкривається в роботі Murphy et al (1996) The Prostate 29, 371-380 and Tjua et al (1997) The Prostate 32, 272-278. Отже, в переважному втіленні цього винаходу фармацевтична композиція, яка вміщує принаймні одну антиген-презентуючу клітину, імпульсним шляхом наповнюється чи зався пептидом, наприклад, за методом прикладу 4. Як альтернатива, антиген-презентуюча клітина вміщує компонент експресії, що кодує пептид. Полінуклеотид може бути будь-яким відповідним полінуклеотидом, та переважно, щоб він був здатний трансдукувати дендритні клітини, таким чином забезпечуючи презентування пептиду та стимуляцію імунітету. Зручним є те, що нуклеїнова кислота винаходу може бути включена у вірусний полінуклеотид чи вірус. Наприклад, показано, що дендритні клітини, трансдуковані аденовірусом, стимулюють антиген-специфічний протипухлинний імунітет відносно MUC1 (див. Gong et al (1997) Gene Ther. 4, 1023-1028). Подібним чином, можуть використовуватись системи на основі аденовірусів (див., наприклад, роботу Wan et al (1997) Hum. Gene Ther. 8, 1355-1363); і ретровірусні системи (Specht et al (1997) J. Exp. Med. 186, 1213-1221 and Szabolcs et al (1997). Крім того, можна використовувати перенос в дендритні клітини за допомогою кров’яних часток (Tuting et al (1997) Eur. J. Immunol. 27, 2702-2707); також можна використовувати РНК (Ashley et al (1997) J. Exp. Med. 186, 1177 1182). В цілому, фармацевтична композиція винаходу, яка вміщує (a) нуклеїнову кислоту(кислоти) винаходу, може вводитись способом, подібним до введення композиції з вмістом пептидів винаходу, наприклад, внутрішньовенного, внутрішньоартеріального, внутрішньочеревного, внутрішньом’язового, інтрадермального, внутрішньопухлинного, перорального, дермального, назального, букального, ректального, вагінального введення, введення шляхом вдихання або місцевого застосування. Внаслідок механізмів уникнення, в пухлині часто розвивається резистентність до препаратів, за допомогою яких вона лікується. Резистентність до ліків може наступити під час лікування; це проявляється в метастазах та в рецидивуючих пухлинах. Щоб уникнути такої резистентності, пухлину, зазвичай, лікують комбінацією препаратів, і наявність метастаз і рецидивів пухлини після періоду, вільного від хвороби, часто вимагає використання іншої комбінації. Отже, в одному з аспектів винаходу фармацевтична композиція вводиться в поєднанні з другим протираковим препаратом. Другий препарат може вводитись до, після чи одночасно з фармацевтичною композицією винаходу. Одночасне введення може, наприклад, досягатись змішуванням фармацевтичної композиції винаходу з другим протираковим препаратом, якщо їхні хімічні властивості є сумісними. Ще один спосіб одночасного введення - це введення композиції та протиракового препарату в той же день, незалежно від способу введення, таким чином, щоб фармацевтична композиція винаходу, наприклад, могла вводитися як ін’єкція, а другий протираковий препарат - перорально. Фармацевтична композиція та другий протираковий препарат також можуть вводитись протягом одного курсу лікування, але в різні дні та/або в ході роздільних курсів лікування. Ще один аспект цього винаходу пропонує метод для лікування чи попередження раку у пацієнта, котрий полягає у введенні пацієнту терапевтично ефективної кількості будь-якої з фармацевтичних композицій винаходу. Терапевтично ефективна кількість - це кількість, достатня для індукування імунної реакції, зокрема, активації субпопуляції CTL. Досвідчений фахівець може достатньо легко визначити, чи є кількість препарату ефективною, використовуючи стандартні імунологічні методи, наприклад, такі, що надані в прикладах цих специфікацій. Ще один спосіб моніторингу ефекту певної кількості фармацевтичної композиції - це спостерігання за ростом пухлини, що лікується, та/або її рецидивами. В особливо переважному втіленні цього винаходу фармацевтична композиція використовується як протиракова вакцина. Композиція, яка вміщує пептиди або нуклеїнові кислоти, що кодують пептид, також може вміщувати протипухлинну чи протиракову вакцину. Вона може вводитись безпосередньо пацієнту, в уражений орган, або системно, чи застосовуватися ex vivo до клітин, виділених у пацієнта чи клітинної лінії людини, котрі згодом вводяться пацієнту, або використовуватись in vitro для обирання субпопуляції з імунних клітин, які виділяються у пацієнта і потім знов вводяться йому. 24 UA 103882 C2 5 10 15 20 25 30 35 40 45 50 55 60 Композиція винаходу може використовуватись в методі лікування раку або як вакцина проти раку. Це може бути рак ротової порожнини та рак глотки, рак шлунково-кишкового тракту, рак товстої кишки, прямої кишки та анусу, рак дихальних шляхів, рак молочної залози, рак шийки матки, вагіни та вульви, рак матки та яєчника, рак статевих шляхів у чоловіків, рак сечовивідних шляхів, рак кістки та м’яких тканин, саркома Капоші, меланома шкіри, меланома ока, рак ока немеланомного характеру, рак мозку та центральної нервової системи, рак щитовидної залози та інших ендокринних залоз, лімфома Ходжкіна, не-хождкінська лімфома, мієлома, переважно, рак нирки, колоректальний рак, рак легенів, рак грудей, рак підшлункової залози, рак простати, рак шлунку, рак мозку, стромальні пухлини ШКТ або гліобластома. В найбільш переважному втіленні методу лікування або вакцини, відповідно до винаходу, вакцина є протипухлинною вакциною з кількох пептидів для лікування колоректальної карциноми. Переважно, вакцина вміщує ряд пухлино-асоційованих пептидів, обраних з послідовностей від SEQ ID No. 1 до SEQ ID No. 15, котрі розташовані та ідентифіковані на клітинах первинних колоректальних пухлин. Цей ряд включає пептиди HLA класу I і класу II. Ряд пептидів може також вміщувати принаймні один пептид, такий як з внутрішнього антигену HBV, використовуваний як пептид позитивного контролю, що служить як імунний маркер для тестування ефективності інтрадермального введення. В одному з конкретних втілень, вакцина складається з 14 індивідуальних пептидів (згідно з послідовностями від SEQ ID No. 1 до SEQ ID No. 15) від приблизно 1500 мкг до приблизно 75 мкг, переважно від 1000 мкг до близько 750 мкг, та більш переважно від приблизно 500 мкг до приблизно 600 мкг, і і найбільш переважно приблизно 578 мкг кожного пептиду, всі з яких можуть очищуватися за допомогою HPLC та іонообмінної хроматографії та мають форму білого чи жовтуватого порошку. Ліофілізат переважно розчиняється в гідрокарбонаті натрію та використовується для інтрадермальних ін’єкцій протягом 30 хвилин після відновлення при кімнатній температурі. Відповідно до цього винаходу, переважні кількості пептидів можуть варіюватися від приблизно 0.1 та 100 мг, переважно, від приблизно 0.1 до 1 мг, та найбільш переважно - від приблизно 300 мкг до 800 мкг на 500 мкл розчину. Термін «приблизно» тут означає +/- 10 процентів від даного значення, якщо не зазначено інше. Спеціаліст у цій галузі здатний відкорегувати фактичну кількість пептиду, який використовується, на основі кількох факторів, наприклад, імунного статусу індивідуального пацієнта та/або кількості TUMAP, що є присутньою в конкретному виді раку. Пептиди цього винаходу можуть бути надані в інших прийнятних формах (стерильні розчині та ін.) замість ліофілізату. Фармацевтичний препарат цього винаходу, який вміщує пептиди та/або нуклеїнову кислоту (кислоти), згідно із винаходом, вводиться пацієнту, який страждає на аденоматозне чи ракове захворювання, асоційоване з відповідним пептидом чи антигеном. Таким чином, може бути запущена імунна реакція з посередництвом Т-клітин. Переважною є фармацевтична композиція, відповідно до винаходу, в якій кількість пептидів чи нуклеїнових кислот (зокрема, пухлино-асоційованих), відповідно до винаходу, чи векторів експресії, відповідно до винаходу, присутня у композиції, є специфічною для тканин, ракових захворювань та/або окремих пацієнтів. В іншому втіленні винаходу вакцина є нуклеїново-кислотною вакциною. Відомо, що введення нуклеїново-кислотної вакцини, такої як ДНК-вакцина, що кодує поліпептид, веде до Тклітинної реакції. Вона може вводитись безпосередньо пацієнту, в уражений орган, або системно, чи застосовуватися ex vivo до клітин, виділених у пацієнта чи клітинної лінії людини, котрі згодом вводяться пацієнту, або використовуватись in vitro для обирання субпопуляції з імунних клітин, які виділяються у пацієнта і потім знов вводяться йому. Якщо нуклеїнова кислота вводиться у клітини in vitro, вона може бути корисною для клітин, що трансфекуються, щоб спільно експресувати імунно-стимулюючі цитокіни, наприклад, інтерлейкін-2 чи GM-CSF. Нуклеїнова кислота (кислоти) можуть бути, по суті, чистими, або поєднаними з імуностимулюючим ад’ювантом, чи використовуватись в комбінації з імуно-стимулюючими цитокінами, або вводитись з належною системою доставки, наприклад, ліпосомами. Нуклеїново-кислотна вакцина також може вводитися з ад’ювантом, таким як описані для пептидних вакцин вище. Переважно, якщо нуклеїново-кислотна вакцина вводиться без ад’юванту. Полінуклеотид може бути, по суті, чистим, або включеним в належний вектор чи в систему доставки. Відповідні вектори та системи доставки включають вірусні системи, основані на аденовірусі, вірусі коров’ячої віспи, ретровірусах, вірусі герпесу, адено-асоційованому вірусі чи гібридах, які включають елементи більш, ніж одного вірусу. Невірусні системи доставки включають катіонні ліпіди та катіонні полімери, добре відомі як засоби доставки ДНК. Також може використовуватись фізична доставка, наприклад, за допомогою «генної пушки». Пептид, 25 UA 103882 C2 5 10 15 20 25 30 35 40 45 50 55 60 або пептид, кодований нуклеїновою кислотою, може бути гібридним білком, наприклад, з епітопом з правцевого анатоксину, що стимулює CD4-позитивні T-клітини. Відповідно, будь-яка нуклеїнова кислота, яка вводиться пацієнту, є стерильною і такою, що не вміщує пірогену. Чиста ДНК може вводитися внутрішньом’язовим, інтрадермальним чи підшкірним шляхом. Зручним є те, що вакцина з нуклеїновою кислотою може включати будьякий прийнятний засіб доставки нуклеїнової кислоти. Нуклеїнова кислота, переважно ДНК, може також доставлятись з ліпосомами чи як частина вірусної векторної системи доставки. Переважно, якщо нуклеїново-кислотна вакцина, така як ДНК-вакцина, вводиться в м’язи, в той час як пептидні вакцини переважно вводяться s.c. або i.d. Також переважно, якщо вакцина вводиться в шкіру. Вважається, що поглинання нуклеїнової кислоти та експресія поліпептиду, кодованого професійними антиген-презентуючими клітинами, такими як дендритні клітини, може бути механізмом ініціації імунної реакції; однак, дендритні клітини не можуть бути трансфековані, але є все ж таки важливими, оскільки вони можуть забирати експресований пептид з трансфекованих клітин в тканині («крос-примування», наприклад, в роботі Thomas AM, Santarsiero LM, Lutz ER, Armstrong TD, Chen YC, Huang LQ, Laheru DA, Goggins M, Hruban RH, Jaffee EM. Mesothelin-specific CD8(+) T cell responses provide evidence of in vivo cross-priming by antigen-presenting cells in vaccinated pancreatic cancer patients. J Exp Med. 2004 Aug 2;200(3):297306). Імунізаційна терапія раку, в якій посередниками виступають полінуклеотиди, описана в роботах Conry et al (1996) Seminars in Oncology 23, 135-147; Condon et al (1996) Nature Medicine 2, 1122-1127; Gong et al (1997) Nature Medicine 3, 558-561; Zhai et al (1996) J. Immunol. 156, 700710; Graham et al (1996) Int J. Cancer 65, 664-670; та Burchell et al (1996) 309-313 In: Breast Cancer, Advances in biology and therapeutics, Calvo et al (eds), John Libbey Eurotext; всі вони включені в цей документ шляхом посилань в усій повноті. Також може бути доцільним спрямування вакцини на конкретні клітинні популяції, наприклад, антиген-презентуючі клітини, по місцю введення, з використанням цільових векторів і систем доставки або селективної очистки такої клітинної популяції у пацієнта та введення пептиду чи нуклеїнової кислоти ex vivo (наприклад, дендритні клітини можуть сортуватися, як описано в роботі Zhou et al (1995) Blood 86, 3295-3301; Roth et al (1996) Scand. J. Immunology 43, 646-651). Наприклад, цільові вектори можуть вміщувати тканино- чи пухлино-специфічний промотор, який спрямовує експресію антигену в належне місце. Нарешті, вакцина згідно із винаходом може залежати від специфічного виду раку, яким хворіє пацієнт, що проходить лікування, а також стану захворювання, попередньої схеми лікування, імунного статусу пацієнта та, звичайно, від HLA-гаплотипу пацієнта. Крім того, вакцина, згідно із винаходом, може вміщувати індивідуалізовані компоненти, відповідно до особистих потреб конкретного пацієнта. Прикладами є різні кількості пептидів, згідно із експресією відповідних ТАА у зазначених пацієнтів, небажані побічні ефекти внаслідок алергії чи інших режимів лікування, а також корегування для другого курсу після проходження першого циклу чи курсу лікування. На додаток до використання з метою лікування раку, пептиди цього винаходу також є прийнятними як діагностичні реактиви. Оскільки пептиди були генеровані з гліобластоми, і було визначено, що ці пептиди не присутні в нормальних тканинах, дані пептиди можуть використовуватися для діагностування наявності раку. Присутність пептидів цього винаходу на біоптатах тканин може допомогти патологоанатому в діагностуванні раку. Визначення окремих пептидів цього винаходу за допомогою антитіл, масспектрометрії чи інших методів, відомих у цій галузі, може підказати патологоанатому, що тканина є злоякісною чи запаленою або взагалі ураженою хворобою. Наявність груп пептидів цього винаходу може дозволити проведення класифікації чи суб-класифікації хворих тканин. Визначення пептидів цього винаходу на зразку хворої тканини може підказати рішення про необхідність терапії із включенням імунної системи, зокрема, якщо відомо або передбачається, що T-лімфоцити беруть участь в механізм дії. Втрата MHC-експресії є добре описаним механізмом уникнення імунного контролю інфікованими або злоякісними клітинами. Тобто, присутність пептидів цього винаходу показує, що цей механізм не використовується проаналізованими клітинами. Пептиди цього винаходу могли б використовуватися для аналізу реакцій лімфоцитів проти пептидів цього винаходу, таких як T-клітинні реакції або реакції антитіл проти пептидів цього винаходу, чи пептидів цього винаходу в комплексі з молекулами MHC. Такі лімфоцитарні реакції можуть використовуватися як прогностичні маркери для прийняття рішення щодо подальших етапів лікування. Ці реакції також можна застосовувати як сурогатні маркери в 26 UA 103882 C2 5 10 15 20 25 30 35 40 45 50 55 60 імунотерапевтичних підходах, спрямованих на індукування реакцій лімфоцитів різними засобами, наприклад, вакцинацією білком, нуклеїновими кислотами, аутологічними матеріалами, адоптивним переносом лімфоцитів. В генній терапії, реакції лімфоцитів проти пептидів цього винаходу можуть розглядатись в ході оцінки побічних ефектів. Моніторинг реакцій лімфоцитів також може стати корисним засобом для подальшого обстеження при трансплантаційній терапії, наприклад, для виявлення реакцій «трансплантат проти хазяїна» та «хазяїн проти трансплантата». Пептиди цього винаходу можуть використовуватися для створення та розробки специфічних антитіл проти комплексів MHC/пептид. Вони можуть використовуватись для терапії, із націленням токсинів чи радіоактивних речовин на тканину, уражену хворобою. Ще одним способом використання цих антитіл може бути спрямування радіонуклідів на тканину, уражену хворобою, в цілях томографії, наприклад, PET (позитронно-емісійної томографії). Це може допомогти у визначенні початку розвитку метастаз або визначити розмір і точну локалізацію хворих тканин. Крім того, пептиди можуть використовуватись для підтвердження діагнозу раку, поставленого патологоанатомом на основі біопсії. Цей винахід, ще в одному своєму аспекті, відноситься до комплекту, який вміщує (a) контейнер, який вміщує фармацевтичну композицію, як описано вище, в розчині або в ліофілізованій формі; (b) додатково, другий контейнер, який вміщує розріджувач або відновлюючий розчин для ліофілізованої формуляції; та (c) додатково, інструкції по (i) використанню розчину або (ii) відновленню та/або використанню ліофілізованої формуляції. Комплект може, крім того, включати один чи кілька (iii) буферів, (iv) розріджувачів, (v) фільтрів, (vi) голок або (v) шприців. Контейнер, переважно, є пляшкою, пробіркою, шприцом чи тестовою пробіркою; він може бути контейнером багаторазового використання. Фармацевтична композиція є переважно ліофілізованою. Комплекти цього винаходу, переважно, включають ліофілізовану формуляцію цього винаходу в належному контейнері та інструкції по її відновленню та/або використанню. Належні контейнери включають, наприклад, пляшки, пробірки (наприклад, пробірки з двох частин), шприці (такі як двокамерні шприці) і тестові пробірки. Контейнер може бути виготовлений з різних матеріалів, таких як скло чи пластмаса. Переважно, комплект та/або контейнер вміщує інструкції на контейнер або такі, що відносяться до нього, із вказівками щодо відновлення та/або використання. Наприклад, на ярлику може вказуватися, що ліофілізована формуляція повинна бути відновлена до концентрацій пептидів, як зазначено вище. Ярлик, крім того, може зазначати, що формуляція є прийнятною або призначена для підшкірного введення. Контейнер, вміщуючий формуляцію, може бути пробіркою багаторазового використання, котра дозволяє робити повторні введення (наприклад, від 2 до 6 введень) відновленої формуляції. Комплект, крім того, може включати другий контейнер з прийнятним розріджувачем (наприклад, розчином бікарбонату натрію). Після змішування розріджувача та ліофілізованої формуляції, остаточна концентрація пептиду у відновленій формуляції становить, переважно, принаймні 0.15 мг/мл/пептид (=75 мкг) та переважно не більше 3 мг/мг/пептид (=1500 мкг). Комплект, крім того, може включати інші матеріали, бажані з комерційної точки зору та із врахуванням використання, включаючи інші буфери, розріджувачі, фільтри, голки, шприці та вкладиші з інструкціями по використанню. Комплекти цього винаходу можуть включати один контейнер, який вміщує формуляцію фармацевтичних композицій, відповідно до цього винаходу, з іншими компонентами чи без них (наприклад, інші сполуки або фармацевтичні композиції інших сполук) чи включати окремий контейнер для кожного компоненту. Переважно, комплекти винаходу включають формуляцію винаходу, упаковану для використання, в комбінації з одночасним введенням другої сполуки, такої, як ад’юванти (наприклад, GM-CSF), хіміотерапевтична речовина, природний продукт, гормон чи антагоніст, анти-ангіогенезний агент чи інгібітор, апоптоз-індукуючий агент або хелатор, чи їхню фармацевтичну композицію. Компоненти комплекту можуть бути завчасно змішані, чи кожний компонент може бути в окремому контейнері, до введення пацієнту. Компоненти комплекту можуть бути надані в одному чи кількох рідких розчинах, переважно, водному розчині, більш переважно, стерильному водному розчині. Компоненти комплекту також можуть надаватись як речовини у твердому стані, які можуть перетворюватися на рідини шляхом додання прийнятних розчинників, котрі переважно вміщені в іншому окремому контейнері. Контейнер терапевтичного комплекту може являти собою пробірку, тестову пробірку, колбу, пляшку, шприц або будь-який інший засіб для зберігання твердого тіла чи рідини. Зазвичай, коли є більш ніж один компонент, комплект буде включати другу пробірку чи другий контейнер, який дозволяє окреме дозування. Комплект може також вміщувати інший контейнер для 27 UA 103882 C2 5 10 15 20 25 30 35 40 45 50 55 60 фармацевтично прийнятної рідини. Переважно, терапевтичний комплект буде включати апарат (наприклад, одну чи кілька голок, шприців, крапельниць, піпетку та ін.), який робить можливим введення речовин винаходу, що становлять компоненти цього комплекту. Фармацевтична формуляція цього винаходу є прийнятною для введення пептидів будь-яким зручним методом, наприклад, пероральним (ентеральним), назальним, очним, підшкірним, інтрадермальним, внутрішньом’язовим, внутрішньовенним або трансдермальним. Переважно, введення здійснюється s.c., та найбільш переважно, i.d.-методом. Введення може виконуватись інфузійним насосом. Слід взяти до уваги, що ознаки винаходу, розкриті та описані тут, можуть використовуватися не тільки у відповідній комбінації, як зазначається, але й окремо, не відходячи від передбачуваного обсягу цього винаходу. З метою цього винаходу, усі довідкові матеріали, згадані тут, включені шляхом посилань в усій повноті. Тепер винахід може бути описаний більш детально із посиланням на наведені нижче Малюнки, перелік Послідовностей та Приклади. Ці приклади надані тільки для ілюстративних цілей, без наміру обмежувати винахід. Стислий опис Малюнків Фіг. 1: Тетрамерний аналіз проліферації ODC-001 та NOX-001-специфічних CD8-позитивних 6 лімфоцитів з периферичної крові з використанням мікросфер. 110 збагачених CD8позитивними PBMC (мононуклеарів периферичної крові) на лунку здорового HLA-A*0201позитивного донора HD100 стимулювалися кожного тижня мікросферами в поєднанні з антиCD28 плюс пухлинний антиген високої щільності A*0201/ODC-001 (верхня панель) чи анти-CD28 плюс пухлинний антиген високої щільності A*0201/NOX-001 (нижня панель). Після трьох стимуляцій in vitro, усі клітини були викрашені антитілами CD8 FITC плюс тетрамери A*0201/NOX-001 PE та A*0201/ODC-001 APC. Клітини обмежуються популяцією лімфоцитів або CD8-позитивних лімфоцитів (права панель); цифри представляють процент тетрамерпозитивних в межах CD8-позитивних лімфоцитів. Фіг. 2: Імуногенність іn vitro TGFBI-004, визначена імуноферментним спот-аналізом ELISPOT IFNγ (гамма-інтерферонів) після п’яти циклів стимуляції. Клітини були примовані та повторно рестимульовані TGFBI-004, а потім інкубовані з релевантним пептидом TGFBI-004 (Лунка 1, 2, 3 та 4) і не релевантним пептидом (Нег. контроль), відповідно. Аналіз після IFN ELISPOT був проведений на ELISPOT-рідері (CTL, Клівленд, США). PHA-іономіцин служив позитивним контролем. Цифри вказують на кількість позитивних плям. Фіг. 3: Імуногенність іn vitro TGFBI-004, визначена аналізатором ICS після п’яти циклів стимуляції. Клітини були примовані аутологічними DC з введеним TGFBI-004 та повторно рестимульовані аутологічними PBMC плюс TGFBI-004. Для зчитування, клітини були інкубовані з релевантним пептидом TGFBI-004 (Лунка 1, 2, 3 та 4) і не релевантним пептидом (Нег. контроль), відповідно. Додатково до внутрішньоклітинного IFN-викрашування, клітини також були викрашені антитілами CD4-FITC та CD8-PerCP. Аналіз виконувався на чотирьохкольоровому цитометрі FACSCalibur (BD Biosciences, Німеччина). Фіг. 4: Аналіз ELISPOT виробництва IFN T-клітинними лініями після рестимуляції in vitro пептидом NOX-001. A. T-клітинна лінія 7+ від донора HBC-154 (сортована CD8+ NOX-001 тетрамер+); B. T-клітинна лінія 7 - від донора HBC-154 (сортована CD8+ NOX-001 тетрамер-). Сортовані CD8+ NOX-001 тетрамер-позитивні (A.) та CD8+ NOX-001 тетрамер-негативні (B.) клітини були проаналізовані за допомогою IFN ELISPOT після рестимуляції не релевантним пептидом (MLA-001) (верхні лунки) та релевантним пептидом (NOX-001) (нижні лунки) (10 мкг/мл). Цифри вказують на кількість позитивних плям. Фіг. 5: Частоти of CEA-004-специфічних CD8+ T-клітин в 4 HLA-A2 здорових донорів після стимуляції CEA-004 in vitro, як визначено проточним цитометричним аналізом. Фіг. 6: Афінність пептидів HLA класу I винаходу до MHC-молекули, кодованої алеллю HLAA*0201. Приклади 1. Синтез і структура Пептиди були синтезовані за допомогою стандартного і добре відомого твердофазного синтезу з використанням Fmoc-хімії. Після очищення препаративною HPLC (високоефективною рідинною хроматографією), була виконана іонообмінна процедура для включення фізіологічно сумісних протилежно заряджених іонів (у вигляді ацетатів чи хлоридів). Нарешті, після ліофілізації були одержані тверді речовини, які мали колір від білого до жовтуватого. Усі TUMAP вводяться як ацетати, крім IMA-CCN-001, що надавався як хлорид, по технічним причинам, в ході процедури виготовлення. Важливим є факт, що ідентичність та чистота пептидів можуть визначатися простим 28
ДивитисяДодаткова інформація
Назва патенту англійськоюComposition of tumour-associated peptides and related anti-cancer vaccine
Автори російськоюSingh, Harpreet, Schoor, Oliver, Trautwein, Claudia, Hilf, Norbert, Weinschenk, Toni, Walter, Steffen, Lewandrowski, Peter
МПК / Мітки
МПК: A61K 38/08, A61K 38/16, A61K 39/00, A61P 35/00
Мітки: протиракова, пухлино-асоційованих, відповідна, вакцина, пептидів, композиція
Код посилання
<a href="https://ua.patents.su/49-103882-kompoziciya-pukhlino-asocijjovanikh-peptidiv-ta-vidpovidna-protirakova-vakcina.html" target="_blank" rel="follow" title="База патентів України">Композиція пухлино-асоційованих пептидів та відповідна протиракова вакцина</a>
Протиракова вакцина, що містить пухлиноасоційований пептид, що зв’язується з молекулами лейкоцитарного антигену людини (hla) класу i чи ii, та її застосування у виробництві лікарського засобу

Номер патенту: 97095
Опубліковано: 10.01.2012
Автори: Вальтер Штеффен, Вайншенк Тоні, Зінгх Харпреет, Еммеріх Нільс
МПК: C12N 15/12, A61P 35/00, A61K 38/17, C07K 14/47
Мітки: протиракова, пептид, класу, застосування, засобу, вакцина, людини, антигену, пухлиноасоційований, лікарського, містить, hla, зв'язується, молекулами, виробництві, лейкоцитарного
Формула / Реферат:
1. Протиракова вакцина, що містить пухлиноасоційований пептид, який включає послідовність відповідно до SEQ ID NО: 1, що має загальну довжину від 16 до 30 амінокислот.2. Протиракова вакцина за п. 1, у якій пептид складається з амінокислотної послідовності відповідно до SEQ ID NО: 1.3. Протиракова вакцина за п. 1 або п. 2, у якій пептид включає непептидні зв'язки.4. Протиракова вакцина за будь-яким з пп. 1-3, яка...
Надчиста композиція коротких пептидів з мікроелементами

Номер патенту: 56208
Опубліковано: 10.01.2011
Автори: Косінов Микола Васильович, Каплуненко Володимир Георгійович
МПК: C07C 51/41, C07C 229/16, A23J 1/18, B82B 3/00, A23L 1/30, A61K 36/06
Мітки: пептидів, мікроелементами, коротких, композиція, надчиста
Формула / Реферат:
1. Надчиста композиція коротких пептидів з мікроелементами, що містить воду і щонайменше один короткий пептид і щонайменше один мікроелемент з групи, що включає мідь, цинк, залізо, молібден, кобальт, хром, селен, вісмут, марганець, срібло, золото, паладій, платину, іридій, олово, титан, кремній, германій, ванадій, магній, яка відрізняється тим, що містить мікроелементи у вигляді протеїнатів вказаних металів, отриманих взаємодією наночастинок...
Водна композиція коротких пептидів з мікроелементами

Номер патенту: 59070
Опубліковано: 10.05.2011
Автори: Косінов Микола Васильович, Каплуненко Володимир Георгійович
МПК: A23L 1/305, A23L 1/30
Мітки: коротких, мікроелементами, пептидів, водна, композиція
Формула / Реферат:
1. Водна композиція коротких пептидів з мікроелементами, що містить щонайменше одну сполуку, вибрану з числа цукрів, ферментів, вітамінів, нуклеотидів, коферментів, фосфоліпідів, ароматів, фарбників, консервантів, і щонайменше один короткий пептид і щонайменше один мікроелемент з групи, що включає мідь, цинк, залізо, молібден, кобальт, хром, селен, вісмут, марганець, срібло, золото, паладій, платину, іридій, олово, титан, кремній, германій,...
Протиракова композиція, яка містить стабільні наночастинки, та спосіб лікування раку

Номер патенту: 27746
Опубліковано: 16.10.2000
Автори: САРПОТДАР Прамонд, ЛАЙВЕРСІДЖ Ілейн, ЛАЙВЕРСІДЖ Гарі Дж.
Мітки: протиракова, яка, наночастинки, композиція, лікування, спосіб, стабільні, містить, раку
Текст:
...исходная смесь подвергалась такому предварительному измельчению дисперсии, когда для истирания частиц используют мельницу с циркуляцией измельчающего материала. Из механических средств, используемых для снижения размеров частиц противоопухолевого агента, удобно воспользоваться дисперсионной мельницей. Подходящими дисперсионными мельницами могут быть шаровая мельница, жерновая мельница, вибрационная мельница, планетарная мельница, измельчающим...
Фармацевтична композиція ранозагоюючої та регенеруючої дії на основі низькомолекулярних пептидів, що має специфічну активність

Номер патенту: 68224
Опубліковано: 26.03.2012
Автори: Борщевська Марина Іллінічна, Жебровська Філя Іванівна, Лісничук Наталія Євгенівна, Костюк Григорій Вікторович, Бігуняк Володимир Васильович
МПК: A61K 31/56, A61K 9/127
Мітки: ранозагоюючої, специфічну, активність, дії, низькомолекулярних, регенеруючої, фармацевтична, композиція, має, пептидів, основі
Формула / Реферат:
Фармацевтична композиція ранозагоюючої та регенеруючої дії на основі низькомолекулярних пептидів, що має специфічну активність, яка полягає у підвищенні регенеративної активності епідермісу, колагенізації дерми та помірній запальній інфільтрації, яка відрізняється тим, що висока антимікробна резистентність препарату, отриманого із замороженого дермального шару шкіри свиней, здатність його до ефективної адсорбції, інактивації токсинів,...
Попередній патент: Профілактика або терапевтичне лікування інфекції, викликаної вірусом імунодефіциту людини
Наступний патент: Термооборотно зшита бітумно-полімерна композиція
Випадковий патент: Спосіб контролю вуглеводного обміну у рисистих коней