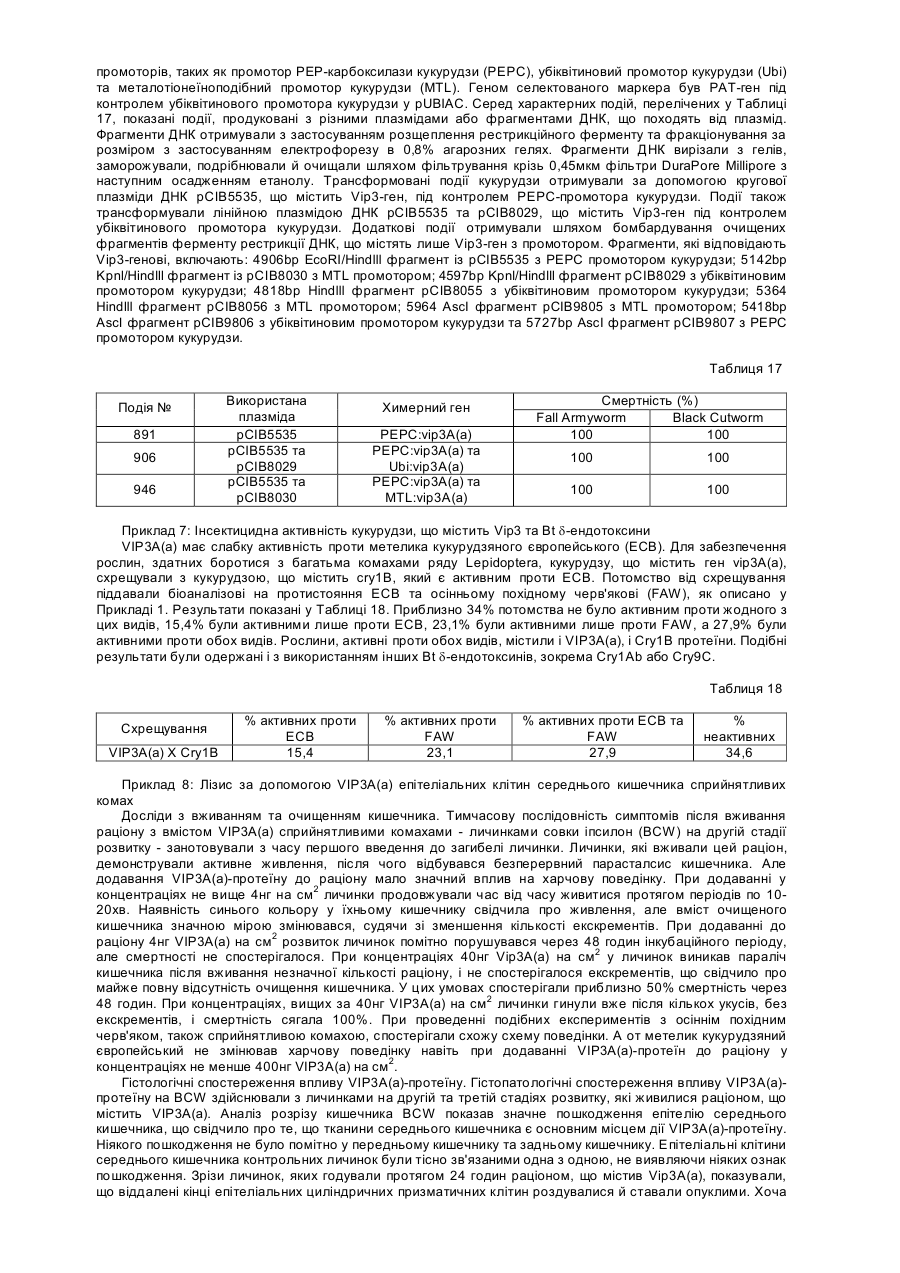
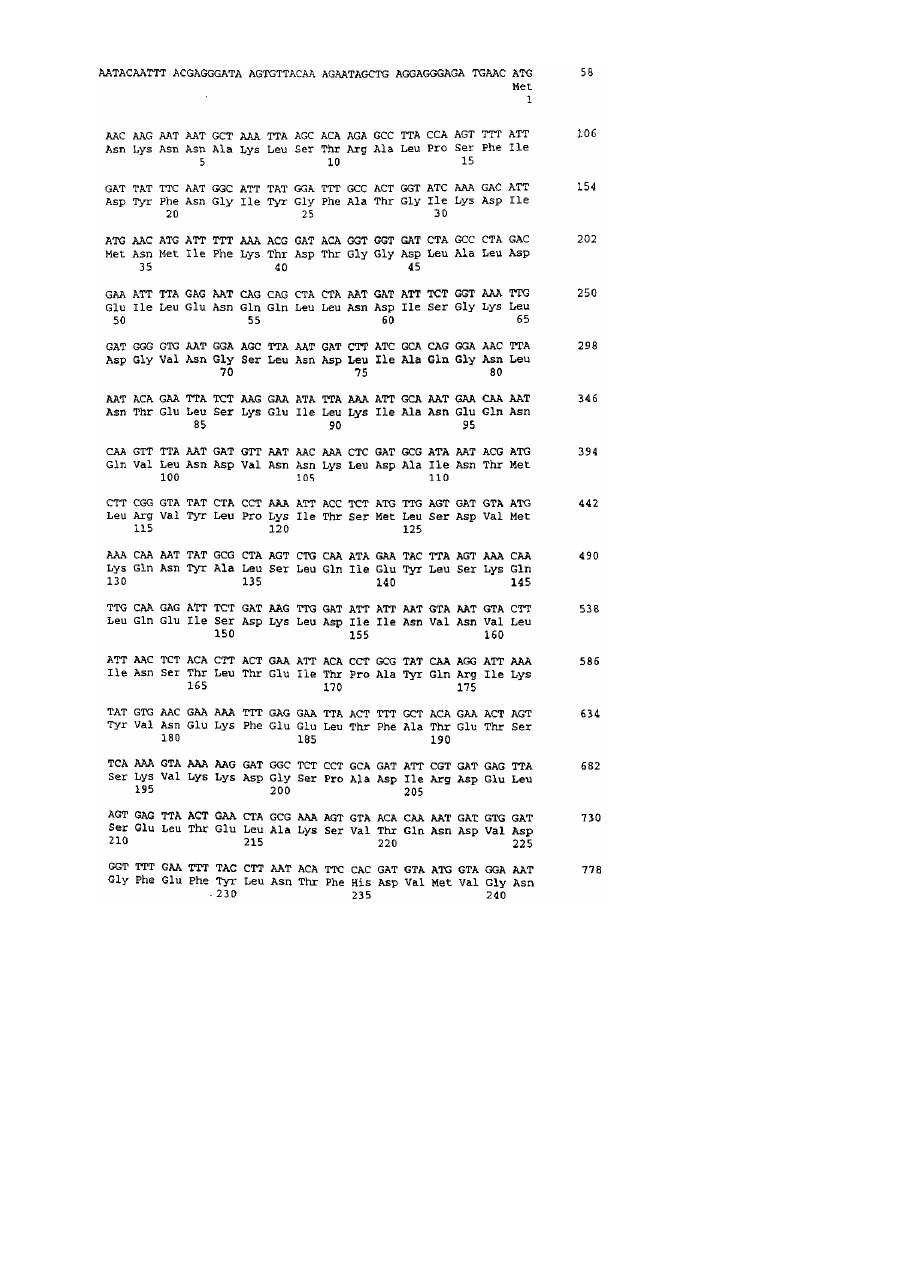
Рецептор до вегетативного інсектицидного протеїну vip3-класу, послідовність днк, антитіло до рецептора, спосіб ідентифікації гомологів рецептора, спосіб ідентифікації сполуки, спосіб боротьби з комахами
Номер патенту: 68345
Опубліковано: 16.08.2004
Автори: Десай Наліні Маной, Ю Као-Гуо, Козіел Майкл Джін, Уоррен Грегорі Уейн, Еструч Хуан Хосе, Най Гордон Джеймс


















































Формула / Реферат
1. Рецептор до вегетативного інсектицидного протеїну VIP3-класу, який кодується послідовністю ДНК, представленою в SEQ ID NO: 8 або послідовністю ДНК, яка гібридизується з нею в жорстких умовах.
2. Рецептор за пунктом 1, який включає домен смерті та повторюваний мотив епідермального фактора росту (EGF).
3. Рецептор за пунктом 1, який має послідовність, представлену в SEQ ID NO: 9.
4. Послідовність ДНК, представлена в SEQ ID NO: 8, або послідовність ДНК, яка гібридизується з нею в жорстких умовах, яка кодує рецептор до протеїну VIP3-класу.
5. Антитіло до рецептора за пунктом 1.
6. Спосіб ідентифікації гомологів рецептора до протеїну VIP3-класу через послідовність ДНК, яка кодує рецептор до протеїну VIP3-класу, який включає:
(а) гібридизацію послідовності ДНК за пунктом 4 із ДНК, отриманою із досліджуваного організму, виявлення гібридизації з ДНК з вказаного організму, та виділення гена, який гібридизується, з вказаного організму; або
(б) одержання зразка ДНК з організму з використанням праймерів до послідовності ДНК за пунктом 4, одержання продукту реакції і потім виділення послідовності ДНК, яка кодує рецептор до протеїну VIP3-класу, з вказаного організму.
7. Спосіб ідентифікації сполуки як хімічного ліганду VIP3-рецептора, що має пестицидну активність, який включає піддання VIP3-рецептора за пунктом 1 дії тестової сполуки та аналіз взаємодії між рецептором та тестовою сполукою.
8. Спосіб за пунктом 7, в якому VIP3-рецептор експресують у клітинах і аналізована взаємодія є запрограмованим відмиранням клітин.
9. Спосіб за пунктом 7, в якому аналізована взаємодія є специфічним зв'язуванням між VIP3-рецептором та тестовою сполукою.
10. Спосіб боротьби з комахами шляхом контактування комах з інсектицидною кількістю хімічного ліганду, отриманого способом за пунктом 7, або антитілом за пунктом 5.
Текст
Даний винахід стосується нового класу протеїнів для боротьби зі шкідниками рослин. Шкідники рослин є основним чинником втрат комерційно важливих сільськогосподарських культур у світі, призводячи як до економічних труднощів для фермерів, так і до недоїдання місцевого населення у багатьох частинах світу. Для боротьби зі шкідниками та їх знищення у сільському господарстві широко використовують найрізноманітніші хімічні пестициди. Однак існує й значний інтерес у розробці ефективних альтернативних пестицидів. Боротьба з різними шкідниками шляхом використання біологічних молекул можлива лише в обмеженій кількості випадків. Найкращим з відомих прикладів біологічних молекул пестицидного використання є dендотоксини з Bacillus thuringiensis (Bt), який є грам-позитивним мікроорганізмом, що утворює спори. Відомі різновиди Bt, які продукують понад 25 різних, але споріднених d-ендотоксинів. Штами Bt під час споруляції продукують d-ендотоксини, використання яких обмежується, бо вони є активними лише проти дуже обмеженої кількості з багатьох комах-шкідників. Обмежена специфічність Bt-ендотоксинів залежить, принаймні частково, як від активізації токсину в організмі комахи (Haider, М. Z. et al., 1986, Eur. J. Biochem. 156:531-540), так і від його здатності зв'язуватися зі специфічними рецепторами, присутніми в епітеліальних клітинах середнього кишечнику комах (Hofmann, С. P. et al., 1988, PNAS 85:7844-7848). Таким чином, можливість боротися з конкретними комахамишкідниками через використання d-ендотоксинів у даний час залежить від можливості знайти відповідний dендотоксин з потрібною сферою активності. У багатьох випадках подібний d-ендотоксин невідомий, і навіть не можна з певністю сказати, чи він взагалі існує. Рослини також зазвичай заражаються грибками та бактеріями, і багато видів мікроорганізмів розвинули здатність до користування різними нішами, які забезпечуються зростаючими рослинами. Крім зараження грибками та бактеріями, багато хвороб рослин викликається нематодами, які живуть у грунті і уражають коріння, як правило, завдаючи серйозної шкоди, коли один вид культури культивується кілька років поспіль на одній ділянці грунту. Серйозність викликаного захворюванням руйнівного процесу залежить від агресивності фітопатогену та хазяїна, і одна з цілей більшості програм вирощування рослин полягає у підвищенні опірності рослинихазяїна до хвороби. Нові джерела та комбінації генів, розроблені для забезпечення опірності до хвороб, як правило, мають лише обмежений період успішного застосування у багатьох патогенних для культур системах через швидкий розвиток фітопатогенів, які долають гени опірності. Таким чином, стає зрозумілим, що вчені повинні постійно перебувати у пошуку нових способів захисту культур від шкідників рослин. У даному винаході було виявлено новий клас протеїнів, які можуть бути використані для боротьби зі шкідниками рослин. Запрограмоване відмирання клітин є процесом, у якому еволюційні стимули або навколишнє середовище активізують генетичну програму, завершенням якої є смерть клітини (Jacobson, M.D. et аI., 1997, Cell 88: 347-354). Цей генетичний потенціал існує у більшості, якщо не у всіх багатоклітинних організмів. У разі безхребетних запрограмоване відмирання клітин відіграє подвійну роль, будучи невід'ємною частиною як процесу розвитку рослин, так і механізму відповіді на інфекції, зокрема, такі, що мають вірусне походження (Clem, R.J. et al., 1991, Science 254: 1388-1390). Запрограмоване відмирання клітин відбувається кількома різними шляхами, призводячи до апоптозу, атрофії або диференціації. Апоптоз є одним з найкраще охарактеризованих типів запрограмованого відмирання клітин, яке охоплює цитологічні зміни, включаючи зв'язані з мембраною апоптотичні тільця та цитоплазматичні пухирці, а також молекулярні зміни, такі як ендонуклеоліз, для якого характерним є утворення олігосомальної довжини фрагментів (Vaux, D.L and Strasser, А., 1996, PNAS 93:2239-2244). І хоча загальна апоптотична феноменологія є майже немінливою у різних організмів, цікаво зазначити, що для багатьох клітин комах скоріше цитоплазматична вакуолізація та бубнявіння, аніж конденсація, є цитологічними особливостями, пов'язаними з апоптотичними процесами (Bowen, I.D., et al., 1996, Micros. Res. Techniq. 34:202-217). Виявлено, що новий клас протеїнів, описаний у даному винаході, викликає запрограмоване відмирання клітин і має пестицидний вплив. Даний винахід стосується VІР3А(с)-протеїнів, включаючи їхні гомологи. Також передбачені винаходом домени протеїнів VІР3-класу, включаючи токсичний домен та стабілізуючий домен. Оптимальним варіантом згідно з винаходом є токсичний домен VІР3А(а)-протеїну та їхні гомологи. Іншим оптимальним варіантом є антитіла до протеїнів VІР3-класу, краще - до VIР3А(с)-протеїну. Винахід також забезпечує гібридні токсини, що включають токсичний домен протеїну VІР3-класу. В оптимальному варіанті гібридний токсин є химерними протеїнами, що мають токсичний стержневий домен, функціонально з'єднаний з гетерологічним стабілізуючим доменом. В іншому оптимальному варіанті гібридний токсин включає антитіло або його імунологічно активний фрагмент, який імунологічно визнає VІР3-рецептор, функціонально з'єднаний з токсичним доменом інших протеїнів, причому домен токсину одержують з багатьох цитотоксичних протеїнів, включаючи, крім інших, токсини Bacillus, включаючи ендотоксини та вегетативні інсектицидні протеїни. Винахід також охоплює рослини, що включають послідовність ДНК, яка кодує протеїн VІР3-класу, краще - VІР3А(с)-протеїн. В оптимальному варіанті це рослини, вибрані з групи, яка складається з кукурудзи, сорго, пшениці, соняшника, томатів, різних видів капусти, бавовни, рису, сої, цукрового буряка, цукрової тростини, тютюну, ячменю, та олійного рапсу. У найкращому варіанті рослиною є кукурудза. Винахід також забезпечує мікроорганізми, що включають гетерологічну послідовність ДНК, яка кодує протеїн VІР3-класу, краще - VІР3А(с)-протеїн. В оптимальному варіанті мікроорганізм вибирають з групи, яка складається з бактерій, бакуловірусів, водоростей та грибків. В іншому оптимальному варіанті мікроорганізм вибирають з групи, яка складається з Bacillus, Pseudomonas, Clavibacter та Rhizobium. Винахід також охоплює ентомоцидні композиції, що включають мікроорганізми, які включають гетерологічну послідовність ДНК, яка кодує протеїн VІР3-класу, краще - VІР3А(с)-протеїн. Винахід також стосується рослин та мікроорганізмів, які також включають другу послідовність ДНК, яка кодує другий інсектицидний протеїн. Особливу перевагу віддають другим послідовностям ДНК, які кодують б-ендотоксин, які кодують інший протеїн VІР3-класу або які кодують протеїн VІР1 або VIP2 класів. У ще кращому варіанті d-ендотоксин є активним проти комахи, вибраної з групи, яка складається з Lepidoptera та Coleoptera. У найкращому варіанті d-ендотоксин є активним проти Ostrinia або Diabrotica. В іншому оптимальному варіанті друга послідовність ДНК кодує d-ендотоксинний протеїн, вибраний з групи, яка складається з Сrу1, Сrу3, Сrу5 та Сrу9. У ще кращому варіанті d-ендотоксин вибирають з групи, яка складається з Cry1Aa, Cry1Ab, Cry1Aс, Cry1B, Cry1C, Cry1D, Cry1Ea, Cry1Fa, Сrу3А. Сrу9А, Сrу9С та Сrу9В. Найкращими вважають d-ендотоксини, вибрані з групи, яка складається з Cry1Ab, Cry1Ba та Сrу9С протеїнів. Винахід також забезпечує спосіб боротьби з комахами шляхом контактування комах з інсектицидною кількістю протеїну VІР3-класу, краще - VІР3А(с)-протеїну, або інсектицидною кількістю хімічного ліганду з рецептором протеїнів VІР3-класу. В одному оптимальному варіанті комахи контактують з трансгенною рослиною, що включає послідовність ДНК, яка експресує протеїн VІР3-класу, краще - VІР3А(с)-протеїн В іншому оптимальному варіанті комахи контактують з ентомоцидною композицією, що включає протеїн VІР3класу, краще - VІР3А(с)-протеїн, або що включає послідовність ДНК, яка експресує протеїн VІР3-класу, краще - VІР3А(с)-протеїн. В іще одному оптимальному варіанті трансгенна рослина включає послідовність ДНК, яка експресує VІР3А(а)-протеїн. В іншому оптимальному варіанті комаху вибирають з групи, яка складається з Coleoptera, Diptera, Hymenoptera, Lepidoptera, Mallophaga, Homoptera, Hemiptera, Orthroptera, Thysanoptera, Dermaptera, Isoptera, Anoplura, Siphonaptera, Trichoptera та Асагі. У найкращому варіанті комаха належить до Coleoptera або Lepidoptera. В іншому найкращому варіанті комаху вибирають з групи, яка складається з совки іпсилон (Agrotis ipsilon), осіннього похідного черв'яка (Spodoptera frugiperda), наземної совки (S. ехіguа), похідного черв'яка жовто-смугастого (S. omithogalli), метелика кукурудзяного південно-західного (Diatraea grandiosella), вогнівки цукрової тростини (D. saccharalis), совки (Helicoverpa zea), метелика кукурудзяного середземноморського (Sesamia nonagroides), совки ні (Thchoplusia nі), совки (Anticarsia gemmatalis), молі капустяної (Plutella xylostella) та совки тютюнової (Heliothis virescens). Винаходом також передбачається спосіб боротьби з комахами, згідно з яким трансгенна рослина або мікроорганізм також включає другу послідовність ДНК, яка кодує другий інсектицидний протеїн на зразок вищезгаданих. Винахід також забезпечує рекомбінантні послідовності ДНК, які кодують VІР3А(с)-протеїн, включаючи їхні гомологи. В іншому оптимальному варіанті послідовність ДНК є синтетичною послідовністю, яка була змінена для оптимальної експресії у рослині, зокрема, у випадках, коли послідовність ДНК була оптимізована для експресії у кукурудзі. Перевагу віддають також послідовностям ДНК, які включають як синтетичну частину, так і природну частину. У найкращому варіанті послідовність ДНК, що кодує VІР3А(с)протеїн, була оптимізована для експресії у кукурудзі. Іншим оптимальним варіантом є послідовності ДНК, які є гомологічними послідовностями ДНК, що кодує VІР3А(с)-протеїн. Зокрема, перевагу віддають послідовностям ДНК, які за помірковано суворих умов гібридизуються з wp3A(c) кодуючою послідовністю. Ще одним варіантом втілення винаходу є рекомбінантна послідовність ДНК, яка експресує протеїн VІР3класу, краще - VІР3А(с)-протеїн, під контролем гетерологічного промотора, або коли кодуюча ділянка включена у геном організму, у якому вона у природі не експресується, або експресується на більш високому рівні, ніж природний. Винахід також стосується способу ідентифікації та виділення гомологів VІР3А(с)-протеїну, або послідовності ДНК, яка кодує вищезгаданий протеїн. Винаходом також передбачені експресійні касети, що включають промотор, функціонально з'єднаний з послідовністю ДНК, яка кодує протеїн VІР3 класу, краще - VІР3А(с)-протеїн. В одному з оптимальних варіантів промотор вибирають з групи, яка складається з основних оптимальних для певних тканин та тканиноспецифічних промоторів для експресії у рослинах. У найкращому варіанті промотор вибирають з групи, яка складається з убіквітину, РЕР-карбоксилази (фосфоенолпіруваткарбоксилази), LPT та MTL промоторів. В іншому оптимальному варіанті промотор функціонує у мікроорганізмі. Винахід також забезпечує рецептор для протеїну VІР3-класу та послідовності ДНК. В одному варіанті винаходу рецептор включає домен смерті та повторюваний мотив EGF. У ще кращому варіанті винаходу це рецептор для VІР3A(a). Найкращий варіант - це послідовність рецепторного протеїну, показана у SEQ ID NO:9, та її гомологи. Винахід також охоплює послідовності ДНК, які кодують ці рецепторні протеїни, наприклад, послідовність ДНК, показану у SEQ ID NO:8 та її гомологи. КДНК для VІР3-рецептора міститься у плазміді рСІВ7113, яку було депоновано за Будапештською угодою у Зібранні патентних культур Служби сільськогосподарських досліджень (NRRL [Agricultural Research Service, Patent Culture Collection (NRRL), Northern Regional Research Center, 1815 North University Street, Peoria, Illinois 61604, USA]) 29 березня 1997p. під інвентарним номером В-21676. Винахід також охоплює антитіла до рецептора протеїнів VІР3класу. Винаходом також передбачений спосіб ідентифікації сполуки як VІР3-рецепторного хімічного ліганду, який має пестицидну активність, що включає піддавання клітини, краще - клітини комахи, дії тестової сполуки та аналіз вищезгаданої клітини на апоптотичну активність. В іншому варіанті винаходу спосіб включає вимірювання специфічного зв'язування між VІР3-рецептором та тестовою сполукою. Оптимальним варіантом є VІР3-рецепторні ліганди, ідентифіковані цим способом. Визначення «Шкідник рослин» означає будь-який організм, пов'язаний з рослинами, який в результаті цього зв'язку викликає шкідливий вплив на здоров'я та життєздатність рослин. Шкідники рослин включають, крім інших, грибки, бактерії, комах та нематоди. Вжитий нами термін «рослина» охоплює цілі рослини та частини рослин, такі як коріння, стебла, листя та насіння, а також клітини та тканини у рослинах або частинах рослин. «VІР3-клас протеїнів» включає VІР3A(a), VІР3A(b) VІР3A(c) та їхні гомологи. Термін «гомолог» в усьому тексті означає, що зазначений протеїн або поліпептид має певне визначене відношення до інших представників VІР3-класу протеїнів. Це визначене відношення включає, крім інших, 1) протеїни, які принаймні на 70%, краще - на 80%, найкраще - на 90% є ідентичними на рівні послідовності іншому представникові VІР3-класу протеїнів, водночас зберігаючи також пестицидну активність, 2) протеїни, які вступають у перехресну реакцію з антитілами, які імунологічно визнають іншого представника VІР3-класу протеїнів, 3) протеїни, які вступають у перехресну реакцію з рецептором іншого представника VІР3-класу протеїнів і зберігають здатність викликати запрограмоване відмирання клітин та 4) протеїни, які принаймні на 70%, краще - на 80%, найкраще - на 90% є ідентичними на рівні послідовності токсичній центральній ділянці іншого представника VІР3-класу протеїнів, водночас зберігаючи також пестицидну активність. Термін «гібридний токсин» використано для позначення генетичного злиття, що має домени, функціонально з'єднані таким чином, щоб при трансляції утворювався функціональний химерний протеїн, який в цілому має властивості окремих доменів. Термін «домен» використано для позначення ділянки або частини протеїну, що забезпечує функцію, яка може бути розпізнана, або структури, яка бере участь у загальній функціональності протеїну. Визнано, що це визначення охоплює також послідовність ДНК, яка кодує домен протеїну. Термін «гетерологічний» використано для позначення того, що протеїнова, поліпептидна або нуклеотидна послідовність має різне природне походження по відношенню до даного хазяїна. Наприклад, якщо ген wp3A(a) з Bacillus thuringiensis є генетично трансформованим у рослинну клітину, то ген описують як гетерологічний по відношенню до його нинішнього хазяїна, яким є рослинна клітина. Крім того, якщо ген wp3A(a) з Bacillus thuringiensis є генетично трансформованим у бактерію Pseudomonas, то ген також описують як гетерологічний по відношенню до Pseudomonas. Термін «гетерологічний» також використовують для позначення того, що один або кілька доменів, присутніх у химерній протеїновій, поліпептидній або нуклеотидній послідовності, відрізняються за природним походженням по відношенню до інших присутніх доменів. Наприклад, якщо токсичний домен з \/ІРЗ VІР3А(а)-протеїну зливається зі зв'язувальним доменом з VІР1А(а)-протеїну для створення функціонального інсектицидного протеїну, то химерне злиття матиме домени, гетерологічні один до одного. До того ж, гетерологічний химерний протеїн або поліпептид, що включає злиття токсичного домену з VІР3А(а)-протеїну зі зв'язувальним доменом з VІР1А(а)-протеїну, який, будучи експресованим у рослині, також має розглядатися як гетерологічний по відношенню до рослини-хазяїна. Термін «химерний» використано для позначення того, що протеїнова, поліпептидна або нуклеотидна послідовність складається з доменів, принаймні один з яких має походження, гетерологічне по відношенню до інших присутніх доменів. Ці химерні протеїни або поліпептиди кодуються химерними нуклеотидними послідовностями, які були злиті або ліговані, даючи в результаті кодуючу послідовність, яка у природі не трапляється. Подібні химерні конструкції можуть також називатися «рекомбінантними». Використаний авторами термін «касета експресії» означає послідовність ДНК, здатну на спрямування експресії гену у рослинних клітини, включаючи промотор, функціонально з'єднаний з амінокислотною кодуючою ділянкою, функціонально з'єднаною з кінцевою ділянкою. Ген може бути химерним, означаючи, що принаймні один компонент гена є гетерологічним по відношенню до принаймні одного іншого компонента гена. Ген також може траплятися у природі, але він має бути отриманим у рекомбінантній формі, яку використовують для генетичної трансформації рослини або мікроорганізму. Фігура 1: Амінокислотну послідовність рецептора для VІР3A(a) транслюють з кДНК. Показані деякі особливості протеїну: пунктирна лінія - сигнальний пептид (амінокислоти з 13 по 35); курсив - домен, що заповнює гіпотетичний домен смерті (амінокислоти 81-205); підкреслення подвійною лінією - послідовності з виразною гомологією з послідовностями, які містять узагальнюючі домени смерті; жирний шрифт - СКСмотив, який повторюється шість разів, заповнюючи EGF-мотиви; підкреслення однією лінією послідовності, що повторюються у EGF-мотивах. Членистоногі шкідники З точки зору даного винаходу, шкідники включають комах та павукоподібних, вибраних з-поміж рядів Coleoptera, Diptera, Hymenoptera, Lepidoptera, Mallophaga, Homoptera, Hemiptera, Orthroptera, Thysanoptera, Dermaptera, Isoptera, Anoplura, Siphonaptera, Trichoptera та Acah, зокрема, Coleoptera та Lepidoptera. Перелік шкідників основних культур подано, наприклад, у Таблицях 1-10 на сторінках з 13 по 20 WO 96/10083, на який нами робиться посилання. Інші шкідники представлені у нижчеподаних Таблицях 1-8. Такі шкідники включені до обсягу даного винаходу. З точки зору даного винаходу до шкідників також належать грибкові фітопатогени рослини. Перелік грибкових шкідників основних культур подано у Таблиці 9. Такі шкідники включені до обсягу даного винаходу. Протеїни VІР3-класу виділяються у середовище видами Bacillus на вегетативних стадіях розвитку. VIP3A(a) є представником нового відкритого класу протеїнів, що виявляє інсектицидну активність проти багатьох комах Lepidoptera, включаючи совку іпсилон (Agrofis ipsilon), осіннього похідного черв'яка (Spodoptera frugiperda), наземну совку (S. ехіguа), похідного черв'яка жовто-смугастого (S. omithogalli), метелика кукурудзяного південно-західного (Diatraea grandiosella), вогнівку цукрової тростини (D. saccharalis), совку бавовняну (Helicoverpa zea), метелика кукурудзяного середземноморського (Sesamia nonagroides), совку ні (Trichoplusia nі), совку (Anticarsia gemmatalis), міль капустяну (Plutella xylostelia) та совку тютюнову (Heliothis virescens). Деякі з цих комах Lepidoptera виявилися дуже стійкими до інших інсектицидних протеїнів, таких, як d-ендотоксин. Наприклад, LC50 для Сrу1А(с), одного з найбільш ефективних d-ендотоксинів проти совки іпсилон, потрібно понад 6000 нг/см2 (Macintosh et al, J. Invertebr. Pathol. 56:258-266 (1990)). На відміну від нього, VІР3А(а)-протеїну потрібно у 260 разів менше для знищення 50% личинок совки іпсилон. Таким чином VІР3А(а)-протеїн виявляє унікальну інсектицидну дію. Даний винахід пропонує нового представника VІР3-класу протеїнів, VІР3А(с)-протеїн, виділений зі штаму АВ51 (рСІВ7112, депонований 28 березня 1997 року під інвентарним номером NRRL В-21675; усі культури депонують згідно з Будапештською угодою у Зібранні патентних культур Служби сільськогосподарських досліджень (NRRL), Northern Regional Research Center, 1815 North University Street, Peoria, Illinois 61604, USA), як описано у SEQ ID NO: 5-6. VІР3А(с)-протеїн, що кодує послідовність ДНК, був ідентифікований і виділений з застосуванням технології полімеразної ланцюгової реакції (PCR). Зокрема, можуть бути створені послідовності праймерів, які розпізнають немінливі або мінливі ділянки кодуючої послідовності, а потім їх використовують для скринінгу зразків ДНК, одержаних з відомих або невідомих штамів. Було визнано, що існує багато підходів до ідентифікації та виділення гомологів у VІР3-класі протеїнів та послідовностей ДНК, які їх кодують, і ці підходи відомі спеціалістам. Послідовності ДНК та протеїнів для VIP3A(a)- та VІР3А(с)-протеїнів подано у Таблиці 12. Поліпептидні домени VIP3-wiacy протеїнів Було показано, що VІР3А(а)-протеїн зазнає протеолітичної обробки при змішуванні з рідинами кишечника личинок комах. Коли рідини кишечника, виділені з совки іпсилон, змішуються з очищеним VIP3A(a), можна ідентифікувати чотири основні похідні від VIP3A(a) протеолітичні продукти, що мають молекулярну масу приблизно 66, 45, 33 та 22кДа. Диск 22кДа включає N-термінальну частину VІР3А(а)протеїну від амінокислоти 1 до амінокислоти 198 SEQ ID NO:2. Диск 66кДа включає решту VІР3А(а)протеїну від амінокислоти 200 до амінокислоти 789 SEQ ID NO:2. Диски 45 та 33кДа одержують шляхом протеолізу з диску 66кДа і включають амінокислоти від 412 до амінокислоти 789 і від амінокислоти 200 до амінокислоти 455 SEQ ID NO:2 відповідно. Диск 33кДа є основним компонентом VІР3А(а)-протеїну, який залишається після інкубаційного періоду понад дві години. Цей 33кДа домен «токсичного кору» (амінокислоти від 200 до 455 SEQ ID NО:2) VІР3А(а)-протеїну повністю зберігає інсектицидні властивості проти багатьох комах Lepidoptera. Подібні результати одержують при інкубації VIP3A(a) з рідиною кишечника, виділеною з осіннього похідного черв'яка, іншої комахи, чутливої до VIP3A(a). Крім домену токсичного кору, VІР3А(а)-протеїн також має стабілізуючий домен на С-терміналі. Роль стабілізуючого домену вивчали з використанням мутантів VІР3А(а)-протеїну та VІР3А(с)-протеїну, жоден з яких не виявляє інсектицидних властивостей при вживанні комахами, чутливими до VIP3A(a). При проведенні подібних досліджень у рідині кишечника совки іпсилон з VІР3А(а)-мутантами, зокрема, з мутантом VІР3А(а)-протеїну, що містить три точкові мутації, розміщені у карбокси-термінальному домені (амінокислота 742 (E®D); амінокислота 770 (S®P) та амінокислота 784 (Y®H)), було виявлено, що протеїн був повністю гідролізований. Подібні результати були одержані для VIP3A(c) (SEQ ID NО:6) протеїну, виділеного з АВ51, який в цілому на 96% є ідентичним VІР3А(а)-протеїну, але не має карбокситермінального домену VIP3A(a). Однак і мутант, і VІР3А(с)-протеїн є активними проти лінії клітин комах Sf-9. Ці результати показують, що функція карбокси-термінального домену протеїнів VІР3-класу полягає у забезпеченні стабільності для протеїну у кишечнику сприйнятливої комахи. Гібридні токсини, що включають VІР3-ділянку та гетерологічну ділянку Токсини, ферменти, фактори транскрипції, антитіла, зв'язувальні складові клітин або інші домени протеїнів можуть бути функціонально з'єднаними з новими протеїнами даного винаходу шляхом утворення генетичного злиття каркасів, які, будучи трансльовані рибосомами, продукують протеїн злиття з комбінованими властивостями VIP та іншого компоненту, який використовують для злиття. Крім того, якщо домен протеїну, злитий з VIP, має спорідненість з іншим протеїном, нуклеїновою кислотою, вуглеводом, ліпідом або іншим хімічним реагентом або фактором, то може бути утворений трикомпонентний комплекс. Цей комплекс матиме властивості усіх його компонентів. Подібне пояснення може бути використане для утворення комплексів з чотирьох або більшої кількості компонентів. Ці комплекси використовують як інсектицидні токсини, фармацевтичні, лабораторні реагенти, діагностичні реагенти і т. ін. Прикладами використання подібних комплексів є токсини злиття для лікування потенційних ракових захворювань, реагенти для ферментних імуносорбентних тестів (ELISA) та імуноблотного аналізу. Гібридні токсини винаходу включають химерні протеїни, що мають домен токсичного кору, який є гетерологічним стабілізуючому доменові. Гібридні токсини також утворюють шляхом комбінування антитіла або його імунологічно активного фрагмента, який імунологічно відрізняє VІР3-рецептор з токсичним доменом від інших протеїнів. Домен токсину одержують від багатьох цитотоксичних протеїнів. Вони включають, крім інших, токсини Bacillus, включаючи ендотоксини та вегетативні інсектицидні протеїни. Див., наприклад, Заявку на патент США №08/037,057, подану 25 березня 1993р. та Заявку на патент США №07/951,715, подану 25 вересня 1992р., на які авторами робиться посилання. Інші токсини включають каталітичні рибосомні інактиватори, такі, як гелонін, екзотоксин A Pseudomonas або фітолакцин, (структура екзотоксину Pseudomonas була добре охарактеризована у роботі Chaudhary et al., J. Вiol. Chem. 265:1630316310 (1990)); переривачі метаболізму клітин, такі як рибонуклеази, (див., наприклад, Mariani et al. Nature 347:737-741 (1990)); токсин Barnase (або РЕ-Ваr), химерний токсин, що походить від екзотоксину A Pseudomonas та рибонуклеази, (див. Prior et al. Cell 64:1017-1023 (1991)); гідрофільні пептиди, які утворюють пори у мембранах (див. Frohlich and Wells, Int. J. Peptide Protein Res. 37:2-6 (1991)). Принцип дії VIP3A(a) VІР3А(а)-протеїн виявив свою активність проти багатьох шкідників рослин. Наприклад, гістопатологічні спостереження показують, що вживання VIP3A(a) сприйнятливими до нього комахами, такими як совка іпсилон (Agrotis ipsilon) та осінній похідний черв'як (Spodoptera frugiperda), викликає параліч кишечника при концентраціях раціону 4нг/см2, з повним лізисом епітеліальних клітин кишечника, що веде до смерті личинки при концентраціях понад 40нг/см2. У менш сприйнятливих комах, таких як метелик кукурудзяний європейський (Ostrinia nubilalis) не розвивається ніякої патології після вживання VIP3A(a). Тоді як протеолітична обробка VІР3А(а)-протеїну рідиною середнього кишечника, одержаною від сприйнятливих та несприйнятливих комах, піддається порівнянню, in vivo дослідження імунолокалізації показують, що VIP3A(a)-зв'язування обмежується клітинами кишечника сприйнятливих комах. Таким чином, список комаххазяїв для VIP3A(a) визначається його здатністю зв'язуватися з клітинами кишечника. Гістопатологічні спостереження показують, що епітеліальні клітини середнього кишечника сприйнятливих комах є основною мішенню для інсектицидного протеїну VIP3A(a), а їхній наступний лізис є основним механізмом смертності. Запрограмоване відмирання клітин є активним процесом саморуйнування, який є важливим для розвитку та підтримання багатоклітинних організмів (Clem, R.J. et al. Science 254: 1388-1390 (1991)). Клітини, що зазнають апоптозу, який є формою запрограмованого відмирання клітин, утворюють зв'язані з мембраною апоптотичні організми й активізують ендогенні нуклеази, яка розщеплює хроматин на окремі фрагменти. SF-9-клітини комахи S. frugiperda, піддані дії VІР3А(а)-протеїну, зазнають цитологічних та молекулярних змін, включаючи протрузію мембрани, значну вакуолізацію та ендонуклеоліз, які свідчать про запрограмоване відмирання клітин апоптотичного типу. Гістологічні дослідження показали, що VІР3А(а)протеїн є спрямованим на епітеліальні клітини середнього кишечника сприйнятливих комах, викликаючи цитологічні зміни, які включають значну вакуолізацію та бубнявіння перед лізисом клітин та смертю личинок. Ці клітини середнього кишечника також зазнають ендонуклеолізу, будучи підданими дії VІР3А(а)протеїну, про що свідчить in situ виявлення фрагментації ДНК. Ці результати показують, що VIP3A(a) виявляє свої інсектицидні властивості на клітинах сприйнятливих комах шляхом ініціювання запрограмованого відмирання клітин апоптотичного типу. Рецептор для VIP3A(a) виділено Представлені вище імуногістохімічні результати показують, що VIP3A(a) має здатність зв'язуватися з апікальними мембранами епітеліальних клітин середнього кишечника і що це зв'язування ініціює процес, який зрештою закінчується лізисом клітин. Це означає, що існує один або кілька протеїнів, розташованих в апікальній мембрані, які розпізнають VIP3A(a) і зв'язуються з ним, діючи як рецептор. Цей рецептор сигналізує про взаємодію з VIP3A(a) і ініціює процес апоптозу. Таким чином, через рецептор клітина комахи реагує на VIP3A(a). Для виділення цього рецептора шляхом скринінгу відбирають бібліотеку кДНК з мРНК виділеної з тканини середнього кишечника совки іпсилон. Мета цього скринінгу полягає у тому, щоб ідінтифікувати й виділити послідовності ДНК, які кодують протеїни, що взаємодіють з VIP3A(a) у двогібридній системі (див. Fields. S. and Song, O.-K. Nature 340:245-246 (1989)). Цей підхід дозволяв в результаті ідентифікувати й виділити кДНК, кодований протеїн якої активно взаємодіяв з VІР3А(а)-протеїном. Ця кДНК довжиною в 1,75 т.п.н. (SEQ ID NO:8) кодує протеїн приблизно у 48кДа (396 амінокислот; див. SEQ ID NO:9). Клонована кДНК має майже такий самий розмір, що й мРНК, яка кодує кДНК, про що свідчить нозерн-блотинг. Частина послідовності ДНК, яка кодує перші, від 5 до 20, амінокислоти, може бути відсутня. У кодованому кДНК протеїні можуть бути розпізнані такі особливості (див. Фігуру 1): 1) він містить сигнальний пептид; 2) він містить домен, гомологічний так званому доменові смерті (Feinstein, Е. Et at. Trends in Biochem. 20:342-344 (1995)); і 3) він містить EGF-подібні мотиви або повтори (Fantl, W. J. et al., Annu. Rev. Biochem. 62:453-481 (1993)). Пошук баз даних протеїнів з застосуванням рецептора VIP3A(a) виявив гомологію з родиною позаклітинних глікопротеїнів, відомих під назвами Tenascins (Pearson, C.A. et al. EMBO J. 7:2677-2681 (1988)) або Hexabrachion (Nies. D.E. et al. J. Biol. Chem. 266:2818-2823 (1991)). Ця родина протеїнів містить EGF-подібні повтори, взаємодіє зі складними лігандами й виконує певну роль в адгезії клітин та/або передачі сигналу. Комбінація домену смерті та повторюваних EGF-мотивів, що спостерігається в VІР3рецепторі, є унікальною серед рецепторів запрограмованого відмирання клітин. До того ж, частина VІР3А(а)-рецептора має гомологію з так званим «доменом смерті». Домен смерті є мотивом довжиною у 60-70 амінокислот, який бере участь у взаємодії між протеїнами і є спільним для протеїнів з різними функціями клітин (Feinstein, E. et aІ. Trends in Biochem. 20:342-344 (1995)). Деякі протеїни, що містять мотиви домену смерті, включають рецептори, пов'язані з апоптотичними процесами. Вони, наприклад, включають Fas-рецептор (Brakebush, С. etal. EMBO J. 11:943-950 (1992)) та фактор некрозу пухлин (TNF) (Tartaglia, LA. etal. Cell 74:845-853 (1993)). Гомологи VІР3А(а)-рецептора можуть бути ідентифіковані й виділені різними способами, наприклад, шляхом гібридизації нуклеїнових кислот. Саузерн-блотинг може бути здійснений на зразках ДНК, взятих з клітин комах або грибків, підданих ферментній рестрикції, хроматографії в агарозі та блотингу на нітроцелюлозних та/або нейлонових фільтрах. Саузерн-блотинг може бути зондований повною або частковою нуклеїновою кислотою, що кодує рецептор VІР3А(а)-протеїну в умовах поміркованої гібридизації та промивання. Гени можуть бути легко клоновані й секвеновані з кДНК або геномної бібліотеки. Відібрана за розміром геномна бібліотека також може бути одержана для забезпечення клонування потрібного(их) гена(ів). Вищезазначені технічні протоколи для здійснення експериментів є легкодоступними (див., наприклад, Molecular Cloning, A Laboratory Manual, Second Edition, Vols. 1-3, Sambrook et aІ. (eds.) Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY (1989) та посилання). Антитіла до VIP3A(a) та його рецептор Пропонуються поліклональні та моноклональні антитіла до VІР3-протеїну або його рецептора, включаючи їхні фрагменти, які імунологічно розпізнають частини кожного з протеїнів. Антитіло та моноклональні антитіла даного винаходу можуть бути приготовлені з використанням VІР3-протеїну або його рецептора як антигена. Антитіла винаходу включають поліклональні та моноклональні антитіла, а також їхні фрагменти, які зберігають здатність до зв'язування VІР3-протеїну або його рецептора. Зазначено, що антитіло, моноклональне антитіло або його фрагмент здатні зв'язувати молекулу, якщо вони здатні специфічно реагувати з молекулою для зв'язування таким чином молекули з антитілом, моноклональним антитілом або його фрагментом. Термін «антитіло» (Аb) або «моноклональне антитіло» (Mab) включає цілу молекулу, а також її фрагменти або зв'язувальні ділянки чи домени (такі як, наприклад, Fab та F(ab)2-фрагменти), здатні зв'язувати гаптен. Такі фрагменти, як правило, утворюються шляхом протеолітичного розщеплення, такі як папаїн або пепсин. Або ж гаптен-зв'язуючі фрагменти можуть бути утворені застосуванням технології рекомбінантних ДНК або синтетичної хімії. Способи приготування антитіл даного винаходу спеціалістам, як правило, відомі. Наприклад, див. Antibodies, A Laboratory Manual, Ed Hahow and David Lane (eds.) Cold Spring Harbor Laboratory, NY (1988), а також посилання авторів. Типові роботи, у яких викладено загальні принципи імунології, включають: Klein, J. Immunology: The Science of Cell-Noncell Discrimination, John Wiley & Sons. NY (1982); Dennett, R., et al. Monoclonal Antibodies, Mybridoma: A New Dimension in Biological Analyses, Plenum Press, NY (1980); and Campbell, A. «Monoclonal Antibody Technology», In Laboratory Techniques in Biochemistry and Molecular Biology, Vol. 13, Burdon et al. (eds.), Elsevier, Amsterdam (1984). Див. також Патенти США №№: 4,609,893; 4,713,325; 4,714,681; 4,716,111; 4,716,117 та 4,720,459. Було визнано, що описаними авторами способами можна одержати антитіла, специфічні до конкретного VІР3-протеїну або його рецептора. Підгрупа Mab-ліній, яка має описану зв'язувальну специфічність, може бути використана як джерело РНК-посередника для клонування кДНК для конкретного моноклонального антитіла. Клонована ДНК після цього може бути секвенована відомими спеціалістам способами. Див., наприклад, Sambrook et al., Molecular Cloning: A Laboratory Manual, 2nd. Edition, Cold Spring Harbor Laboratory Press, NY (1989) vol. 1-3, та посилання авторів. З послідовності нуклеїнової кислоти може бути виключена протеїнова послідовність зв'язувальної ділянки з вибраної МАb. Один спосіб використання антитіл та моноклональних антитіл винаходу включає, крім інших, утворення молекул гібридних токсинів. Тобто, будучи з'єднаним, моноклональне антитіло або фрагмент антитіла зберігає свої зв'язувальні властивості, а частина токсину - свої цитотоксичні властивості. Відомі різні способи одержання генів антитіл. Одним зі способів є клонування випадково вибраної бібліотеки генів антитіл у фазі та скринінг бібліотеки на здатність до зв'язування VІР3-протеїну або його рецептора. Інший можливий підхід полягає у виробленні моноклональних антитіл, які зв'язуються з VІР3протеїном або його рецептором, а потім клонують гени антитіл з таких ліній. У даному разі застосовують другий спосіб. Гени антитіл можуть бути клоновані з клітин гібридоми з використанням праймерів до збережених послідовностей ДНК у постійних ділянках та каркасних ділянках мінливих ділянок і ампліфіковані для клонування з застосуванням полімеразної ланцюгової реакції (PCR). Див. Mullis et al., Meth. Enzymol., 155:335-350 (1987); Erlich, (ed.), PCR Technology, Stockton Press (New York 1989). База даних для важколанцюгових та легколанцюгових послідовностей мишей, зібрана у роботі Kabat et al., US Dept Health and Human Services. US Government Printing Offices (1991), була успішно використана для вироблення ізотипічно специфічних та дегенеративних праймерів для клонування антитіл генів. (Jones et al. Bio/technology 9:88-89 (1991)). Крім того, відомі способи клонування менших фрагментів антитіл (Fab), які мають зв'язувальні властивості первісного антитіла. Повні антитіла є великими молекулами (150кДа), але було виявлено, що значно менші Fab та Fv антиген-зв'язувальні фрагменти (12кДа-50кДа) зберігають повну зв'язувальну спорідненість. Одноланцюгові Fv-фрагменти (scFv), у яких Vh та VI домени з'єднані гідрофільним і гнучким пептидом, були успішно використані для спрямування ферментів та токсинів на конкретні клітини (Bird, Science 423:423-426 (1988); Huston, PNAS 85:5879-5883 (1988)). Єдині Vh-домени (Dabs) та єдині комплементарні детермінуючі ділянки довжиною у 20 амінокислот, які називають мінімальними розпізнавальними одиницями (m.r.u.), також використовували для зв'язування антигенів (Ward, Nature 341:544-546 (1989); Taub, J. Biol. Chem 264:259-265 (1989); Williams, PNAS 86:5537-5541 (1989)). Таким чином, можна максимально зменшити зв'язувальний домен, специфічний для VIP3 або його рецептора. Технологію полімеразної ланцюгової реакції та специфічні олігонуклеотидні праймери застосовують для клонування генів імуноглобуліну або ділянок генів імуноглобуліну. PCR-праймери, специфічні і для важких, і для легких ланцюгів ІgМ, та три IgG ізотипи вибирали з описаної вище бази даних Kabat. Праймери для ділянки, що кодує NН2-термінальний кінець сформованої мінливої ділянки призначали для стимулювання у першій каркасній ділянці й конструювали з певною виродженістю, що дозволяло використовувати їх як «універсальні праймери». 3'-праймери, використовувані для специфічної PCRампліфікації мінливих ділянок, конструювали зі збережених послідовностей першого постійного домену (СН1) і легких, і важких ланцюгів. Інший 3'-праймер використовують для ізотипів імуноглобуліну lgG1, lgG3 та IgM. Ізотипи lgG2A та lgG2B можуть бути ампліфіковані тими самими праймерами, які використовували для IgG1 Мінливі ділянки антитіл клонують у вектор експресії легкого та важкого ланцюга, що містить сигнальний пептид ендоплазматичної сітки та постійні ділянки lgG1 легких та важких ланцюгів відповідно. З послідовностями праймерів, використовуваними для PCR-клонування легких та важких мінливих ділянок імуноглобуліну мишей, можна ознайомитися в опублікованій літературі (Coloma et al. Bio/Techniques 11:152-156 (1991); Jones et al. Biotechnology 9:88-89 (1991)). Олігонуклеотиди створювали на синтезаторі ДНК 380В від Applied Biosystems (Applied Biosystems, Foster City, СА) з застосуванням описаних нижче стандартних умов. PCR-праймери включають сайти рестрикції і після ампліфікації та розщеплення можуть бути клоновані у вектор експресії рослин під контролем промотора, здатного експресуватися в рослинах. Вибирали сайти рестрикції, яких немає у секвенованих генах антитіл. Інший спосіб використання поліклональних та/або моноклональних антитіл винаходу включає стимулювання апоптозу шляхом спрямування рецептора на Vip3A з антитілами. Взаємодія антитіл, вироблених проти протеїнів, розташованих на поверхні клітин, які беруть участь у контролюванні росту клітин, в результаті веде до початку апоптозу шляхом перешкоджання зв'язуванню вищезгаданого рецептора з його природним(и) лігандом(ами). Наприклад, анти-АРО-1 антитіло повністю блокує проліферацію лейкоцитів, що несуть АРО-1-протеїн, і ініціює апоптоз у цих клітинах (Trauth, B.C. et al. Science 245:301-305 (1989)). Крім того, активність, що виникає в результаті взаємодії між даним рецептором та лігандом, імітується шляхом заміщення ліганду для антитіл, вироблених проти рецептора. Наприклад, додавання певної кількості анти-Fas антитіл до клітин, що несуть Fas-рецептор у поверхнях їхніх клітин, опосередковує апоптоз так само, як це було б при додаванні ліганду Fas-рецептора (Itoh, N. etal. Cell 66:233-243 (1991)). Рецептор для Vip3A(a), виділений з совки іпсилон, має гомологію з родиною позаклітинних глікопротеїнів, відомих як Tenascins, зокрема, з Tenascin-X (Bristow, J. et al. J. Cell Biol. 122:265-278 (1993)). Відомо, що Tenascin-X беруть участь в адгезії клітина-клітина та передачі сигналу. Брак функціональності Tenascin-X, чи то через мутацію, чи через видалення гена, веде до смерті. Таким чином, антитіла, вироблені проти різних доменів рецептора для Vip3A(a) або ефективно блокують зв'язування рецептора з його лігандом(ами), або імітують взаємодію VІР3А(а)-протеїну, що ініціює апоптоз. Цей підхід поширюється на різні рецептори з подібними біологічними функціями. У цьому аспекті антитіла, вироблені проти рецепторів клітин комах, задіяних в основних процесах росту, та взаємодії клітин викликають апоптоз і використовуються як стратегічний засіб боротьби з комахами. Скринінг нових інсектицидних речовин, дією яких є апоптоз Матеріали, описані у даному винаході, використовують для скринінгу хімічних лігандів, що мають пестицидні властивості, які викликають апоптотичну реакцію. Хімічні ліганди включають невеликі органічні молекули, пептиди та протеїни. В одному варіанті винаходу лінії клітин комах використовують як модель організмів комах для скринінгу сполук, які є інсектицидними внаслідок їхньої здатності ініціювати апоптозу. Ці лінії клітин піддають скринінгові з високою пропускною здатністю, вирощуючи клітини у багатолункових планшетах і піддаючи дії різних сполук. Як модель організму використовують також дріжджі. З застосуванням описаних авторами або відомих спеціалістам процедур визначають, чи є сполука пестицидною внаслідок викликання апоптозу. Одним зі способів ідентифікації сполук, що ініціюють апоптотичну реакцію через взаємодію з відомим рецептором, є застосування ідентифікованих рецепторів, які беруть участь у трансдукції сигналу, ініційованій в апоптотичних лініях клітин комах. Ці рецептори трансформують у гетерологічні лінії клітин, що створюють ізогенні лінії, одна з яких містить ген експресії специфічного рецептора, а інша не має й не експресує такого гена. Ці лінії клітин піддають скринінгові з високою пропускною здатністю, причому трансформовані лінії клітин, що експресують рецептор, виявляють різну реакцію на сполуки, які ініціюють апоптоз через їхню специфічну взаємодію з вищезгаданим рецептором. Даний винахід також охоплює визначення біохімічних та/або молекулярних маркерів, які специфічно ідентифікують лінії клітин комахи, що зазнають апоптозу. Наприклад, можна виділити конкретні кДНК, що виникли під час апоптотичного процесу у конкретних лініях клітин комах. Хоча шлях кору смерті здається філогенетично незмінним (Nagata, S. Cell 88:355-365 (1997)), шлях трансдукції сигналу від рецептора до кору смерті може бути різним у різних організмах. РНК-посередники, які диференціально експресуються у клітинах комах, що зазнають апоптозу, ідентифікують багатьма доступними способами, наприклад, за допомогою диференціального відображення (Bauer, D. et al. Nucleic Acid Res. 21:4272-4280 (1993)) або субтрактивних бібліотек (Sommer, H. et al. EMBO J. 9:605-613 (1990)). Кодовані диференціально експресованою кДНК протеїни використовують як маркери для апоптозу у специфічних лініях клітин комах. Трансгенні рослини, що включають послідовність ДНК, яка кодує протеїн VІР3-класу Рослина-хазяїн, що експресує принаймні одну з послідовностей винаходу, виявляє більшу опірність до нападів шкідників рослин і, таким чином, має більше можливостей протистояти втратам, пов'язаним з такими нападами. Під рослиною розуміють будь-який вид рослин, який може бути генетично трансформований відомими спеціалістам способами. Ці способи трансформації рослин обговорюються нижче. Рослини-хазяї включають, крім інших, перелічені вище культури, які бувають уражені шкідниками. ЕКСПРЕСІЙНІ КАСЕТИ РОСЛИН Методологія побудови касет експресії у рослинах, а також введення чужорідної ДНК у рослини описані у спеціальній літературі. Такі експресійні касети можуть включати промотори, термінатори, енхансери, лідерні послідовності, інтрони та інші регулюючі послідовності, функціонально з'єднані з кодуючою послідовністю пестицидного протеїну. Визнано також, що промотори або термінатори VІР3-генів можуть бути використані в експресійних касетах. Гени токсинів, що походять від мікроорганізмів, можуть також відрізнятися від генів рослин. Гени рослин відрізняються від генів мікроорганізмів, у яких їхня транскрибована РНК не має визначеної послідовності зв'язування рибосом, що прилягає до ініціюючого метіоніну. Отже, гени мікроорганізмів можуть бути посилені шляхом включення еукаріотного консенсусного ініціатора трансляції в ATG (Kozak, Cell 44:283-292 (1986)). У каталозі Clontech (1993/1994, стор. 210) пропонується послідовність GTCGACCATGGTC (SEQ ID NO:—) як спільний ініціатор трансляції для експресії гена uidA E. соIі у рослинах. Крім того, у роботі Joshi (Nucleic Acid Res. 15: 6643-6653 (1987)) міститься порівняння багатьох послідовностей рослин, прилеглих до ATG, й пропонується консенсусна TAAACAATGGCT (SEQ ID NO:—). У разі труднощів з експресією відкритих рамок зчитування (ORF) мікробів у рослинах включення однієї з цих послідовностей в ініціюючу ATG може поліпшити трансляцію. У таких випадках останні три нуклеотиди консенсусу можуть бути непридатними для включення у модифіковану послідовність через їх модифікацію другого АА-залишку. Оптимальні послідовності, прилеглі до ініціюючого метіоніну, можуть бути різними у різних видів рослин. Розглянувши послідовність генів кукурудзи, занесених до бази даних GenBank/EMBL, можна визначити, які нуклеотиди, прилеглі до ATG, мають бути модифікованими для посилення трансляції гена токсину, введеного у кукурудзу. До того ж, було показано, що видалення невідповідних сплайсових сайтів може посилити експресію та стабільність введених генів. Гени, клоновані з нерослинних джерел і не оптимізовані для експресії у рослинах, можуть містити мотиви, які можуть бути розпізнані у рослинах як 5’ або 3’ сплайсові сайти. Отже, процес транскрипції може бути передчасно припинений з утворенням зрізаної або делегованої мРНК. Гени токсинів можуть бути піддані інженерії для видалення цих невідповідних сплайсових сайтів з застосуванням багатьох різних технологій. Наприклад, застосовують кілька доступних способів для ідентифікації потенційних сплайсових сайтів у послідовності ДНК. По-перше, потенційні сплайсові сайти можуть бути ідентифіковані шляхом комп'ютерного аналізу послідовності ДНК. Консенсусні послідовності, які ідентифікують сплайсові сайти, спеціалістам відомі. Див., наприклад, Goodall, G.J. and Filipowicz. W., EMBO J. 10, 2635-2644 (1991) and Brown, J.W.S., Nucleic Acid Research 14, 9549-9559 (1986). Або ж можна ідентифікувати сплайсові сайти, фактично перероблені рослиною шляхом порівняння PCR-аналізу кДНК, що походить від гена, з дійсними генними продуктами. Продукти, що виявляють здатність до сплайсингу, є коротшими, ніж передбачалося. Ці менші продукти потім клонують, секвенують і визначають точне місце сплайсу. Визнано також, що можна застосовувати поєднання комп'ютерного дослідження та PCR-аналізу. Нові гени токсинів даного винаходу, чи то як природна послідовність, чи як оптимізовані синтетичні послідовності, як було описано вище, можуть бути функціонально злиті з різними промоторами для експресії у рослинах, включаючи конститутивні, індуцибельні, тимчасово регульовані, еволюційно регульовані, хімічно регульовані, тканиноселективні і тканиноспецифічні промотори для приготування молекул рекомбінантної ДНК, тобто химерних генів. Оптимальні конститутивні промотори включають CaMV 35S та 19S промотори (Патент США №5,352,605, виданий Фрейлі (Fraley et al.) 4 жовтня 1994p.). Додатковий оптимальний промотор походить від будь-якого одного або кількох генів актину, які експресуються у більшості типів клітин. Промоторні експресійні касети, описані у роботі McElroyet al. (МоI. Gen. Genet. 231: 150-160 (1991)), можуть бути легко модифіковані для експресії нового гена токсину і є особливо придатними для використання в однодольних рослинах-хазяях. Ще один оптимальний конститутивний промотор походить від убіквітину, який є ще одним генним продуктом, що накопичується у багатьох типах клітин. Убіквітиновий промотор було клоновано з кількох видів для використання у трансгенних рослинах (наприклад, соняшнику - Binet et al. Science 79: 87-94 (1991), кукурудзи - Christensen et al. Plant Molec. ВЫ. 12: 619-632 (1989)). Убіквітиновий промотор кукурудзи розвивався у трансгенних однодольних системах, і його послідовність та вектори, побудовані для трансформації однодольних, описані у публікації патенту ЕР 0 342 926. Убіквітиновий промотор є придатним для експресії нових генів токсинів у трансгенних рослинах, особливо в однодольних. Інші промотори, які використовують для експресії нового гена токсину у рослинах, зокрема, у кукурудзі, є, наприклад, тканиноспецифічними або тканиноселективними промоторами, такими як описані у WO 93/07278; хімічно індуцибельні промотори, описані в ЕР-А 0 332 104, на який авторами робиться посилання у його повному обсязі. Крім промоторів, доступними є також транскрипційні термінатори для використання у послідовностях химерних генів з використанням нових генів токсинів даного винаходу. Транскрипційні термінатори відповідають за термінування транскрипції поза трансгеном та його правильне поліаденілювання. Відповідні транскрипційні термінатори та термінатори, які функціонують у рослинах, включають CaMV 35Sтермінатор, tml-термінатор, термінатор нопалінсинтази, rbcS Е9-термінатор гороху та інші відомі спеціалістам термінатори. Вони можуть бути використані як в однодольних, так і у дводольних рослинах. Відомо, що багато нетрансльованих лідерних послідовностей, що походять від вірусів, такі як описані, наприклад, у WO 96/10083, також посилюють експресію, і вони є особливо ефективними у клітинах дводольних. Зокрема, лідерні послідовності з вірусу тютюнової мозаїки (TMV, the «W-послідовність»), вірусу хлоротичної мозаїки кукурудзи (MCMV) та вірусу мозаїки люцерни (AMV) виявили свою ефективність у посиленні експресії (див., наприклад, Gallie etal. Nucl. Acid Res. 15: 8693-8711 (1987); Skuzeski et al. Plant Molec. Biol. 15; 65-79(1990)). Різні інтронні послідовності виявили посилення експресії при додаванні до 5’ регуляторних ділянок, особливо у клітинах однодольних. Наприклад, було виявлено, що інтрони Adh1-гена кукурудзи значною мірою посилюють експресію природного гена з його спорідненим по жіночій лінії промотором при введенні у клітини кукурудзи (Callis et al., Genes Develop. 1:1183-1200(1987)). ОПТИМІЗАЦІЯ vip3-ГЕНІВ ДЛЯ ЕКСПРЕСІЇ У РОСЛИНАХ Пестицидні гени винаходу можуть бути оптимізовані для посиленої експресії у рослинах. Див., наприклад, ЕРА 0359472; ЕРА 0385962; WO 91/16432 та Periak et al., Proc. Natl. Acad. Sci. 88:3324-3328 (1991). Таким чином можуть бути синтезовані кодуючі послідовності, оптимізовані для експресії у рослинах. В одному варіанті винаходу vip3A(a) одержують згідно зі способом, описаним у Заявці на Патент США за сер. №07/951,715, на який авторами робиться посилання. За цією процедурою використовують оптимальні кодони кукурудзи, тобто єдиний кодон, який найчастіше кодує цю амінокислоту у кукурудзі. Оптимальний кодон кукурудзи для конкретної амінокислоти може бути одержаний, наприклад, із відомої генної послідовності кукурудзи. Використання кодону кукурудзи для 28 генів кукурудзи згадується у роботі Murray et al., Nucleic Acid Research 17: 477-498 (1989), на яку авторами робиться посилання. Приклади синтетичних послідовностей, створених за допомогою оптимізованих кодонів кукурудзи, подані у SEQ ID NO:7 (VIP3A(a)), SEQ ID NO:19 (VIP3A(b)) та SEQ ID NO:20 (VIP3A(c)). Таким чином, нуклеотидні послідовності можуть бути оптимізовані для експресі у будь-якій рослині. Було визнано, що оптимізовані або синтезовані можуть бути усі частини генної послідовності або будь-яка частина. Тобто можуть також використовуватися синтетичні або частково оптимізовані послідовності. ТРАНСФОРМАЦІЯ РОСЛИН Молекули рекомбінантної ДНК можуть бути введені у клітину рослини багатьма способами, визнаними спеціалістами. Спеціалісти знають, що вибір способу може залежати від типу призначеної для трансформації рослини, тобто від того, чи є вона однодольною або дводольною. Відповідні способи трансформації клітин рослин описано, наприклад, у WO 96/10083 та WO 97/46105, відповідно, включаючи мікроін'єкцію, електропорацію, опосередковану Agrobacterium трансформацію, пряме перенесення генів та прискорення балістичних частинок з застосуванням пристроїв від Agracetus, Inc., Медісон, Віконсін, та Dupont, Inc., Вілмінгтон, Делавер. Оптимальним варіантом є спосіб трансформації протопластів для кукурудзи, описаний у Заявці ЕР 0 292 435, а також у Патенті США №5,350,689, на які авторами робиться посилання у їх повному обсязі. Один з кращих варіантів введення експресійних касет даного винаходу у пшеницю шляхом бомбардування мікрочастинками можна знайти у Патенті США №5,610,042, на який авторами робиться посилання у його повному обсязі. Трансформацію рослин здійснюють за допомогою однієї молекули ДНК або багатьох молекул ДНК (тобто, котрансформація), і обидві ці технології є придатними для застосування з експресійними касетами даного винаходу. Існує багато векторів для трансформації рослин, і експресійні касети даного винаходу можуть бути використані разом з будь-якими подібними векторами. Вибір вектора залежить від оптимального способу трансформації та виду, призначеного для трансформації. Існує багато векторів для трансформації з використанням Agrvbacterium tumefaciens. Вони, як правило, несуть принаймні одну крайню Т-ДНК-послідовність і включають такі вектори, як pBIN19 (Bevan, Nucl. Acids Res. (1984)). В одному з оптимальних варіантів новий ген токсину даного винаходу може бути введений у будь-який з бінарних векторів рСІВ200 та рСІВ2001 для використання з Agrobactenum, побудову якого описано, наприклад, у WO 95/33818 (приклад 35) (див. також ЕР 0 332 104, приклад 19). Додатковим вектором, який використовують для опосередкованої Agrobacterium трансформації, є бінарний вектор Рсів10, що містить ген, який кодує резистентність до канаміцину для відбору у рослинах, праву та ліву крайні Т-ДНК-послідовності і включає послідовності з плазміди багатьох хазяїв pRK252, дозволяючи їй відтворюватися в Е. соlі та Agrobactenum. її будову описано у роботі Rothstein et al. (Gene 53:153-161 (1987)). Були побудовані різні похідні рСІВ10, які включають ген для гігроміцину В фосфотрансферази, описаний у роботі Gritz et al. (Gene 25: 179-188 (1983)). Ці похідні дозволяють здійснювати відбір клітин трансгенних рослин лише на гігроміцині (рСІВ743) або гігроміцині та канаміцині (рСІВ715, рСІВ717). Способи з застосуванням прямого перенесення генів або опосередкованого Agrobacterium перенесення зазвичай, але не обов'язково, здійснюють з селектованим маркером, який може забезпечувати резистентність до антибіотика (наприклад, канаміцину, гігроміцину або метотрексату) або гербіциду (наприклад, фосфінонітрицину). Вибір селектованого маркера для трансформації рослин, однак, не є ключовим для винаходу, якщо експресія цієї резистентності та його біохімічна активність не впливає на вибір способу перетворення протоксину на токсин для використання при забезпеченні умовної фертильності. Для деяких видів рослин можуть бути вибрані різні антибіотики або маркери відбору гербіциду. Маркери відбору, які зазвичай використовують при трансформації, включають nptll-геи, який забезпечує резистентність до канаміцину та подібних антибіотиків (Messing & Vierra, Gene 19: 259-268 (1982); Bevan et al., Nature 304:184-187 (1983)), bar-ген, який забезпечує резистентність до гербіциду фосфінонітрицину (White et al., Nucl Acids Res 18:1062 (1990), Spencer et al., TheorAppI Genet 79:625-631 (1990)), hph-геи, який забезпечує резистентність до антибіотика гігроміцину (Blochinger & Diggelmann, Моl Cell Biol 4: 2929-2931), dhfr-геи, який забезпечує резистентність до метотрексату (Bourouis et al., EMBO J. 2: 1099-1104 (1983)), ген манозофосфатізомерази, який дозволяє відбір манози як джерела вуглецю (ЕР 530 129, WO 94/20627). Одним таким вектором, який використовують для прямого перенесення генів у поєднанні з відбором за допомогою гербіциду Basta (або фосфінонітрицину), є рСІВ3064. Основою цього вектора є плазміда рСІВ246, яка включає CaMV 35S промотор у функціональному злитті з GUS геном Е. соlі, а транскрипційний термінатор CaMV 35S описаний в опублікованій згідно з РСТ заявці WO 93/07278, на яку авторами робиться посилання. Інший використовуваний селектований маркер одержують шляхом функціонального з'єднання убіквітинового промотора, синтетичного РАТ-гена та nos-термінатора. Одним з прикладів вектора, що включає цей маркер, є плазміда рСІВ9804. Додатковим вектором трансформації є pSOG35, який використовує ген дигідрофолатредуктази (DHFR) Е. соlі як селектований маркер, що забезпечує резистентність до метотрексату і побудову якого описано, наприклад, у WO 95/33818 (приклад 35). Ще одним вектором трансформації є вектор pGL2 (Shimamoto et al. Nature 338, 274-276 (1989)), який містить ген гігроміцинфосфотрансферази (hpt) Streptomyces, функціонально з'єднаний з послідовностями 35S промотора та 35S термінатора. Трансгенні рослини також можуть бути ідентифіковані використанням маркера, який піддається кількісному аналізові. Прикладами таких маркерів, які використовують у винаході, є b-глюкуронідаза, зелений флуоресцентний протеїн та С1 та В-peru регуляторні гени шляху антоціаніну кукурудзи. Крім того, трансгенні рослини, що експресують VІР3-протеїн, можуть бути ідентифіковані шляхом їх скринінгу на інсектицидну активність без потреби у маркерах, що піддаються кількісному аналізу, або селектованих маркерах. Трансформації кукурудзи з послідовністю ДНК, що кодує протеїн VIP3-класу, краще - VІР3А(с)-протеїн згідно з будь-яким з вищезгаданих способів, легко досягають шляхом бомбардування мікрочастинками або недорозвинених зиготних ембріонів, або ембріогенного калюсу І типу, який піддається серійному розведенню. Для трансформації з використанням недорозвинених зиготних ембріонів, колосся самозапилюють, і приблизно через 10 днів одержують недорозвинені зиготні ембріони. Приблизно вісімсот недорозвинених зиготних ембріонів розподіляють по різних планшетах-мішенях, що містять середовище, здатне викликати й підтримувати утворення ембріогенного калюсу. Недорозвинені зиготні ембріони одразу переносять до такого самого середовища, яке додатково містить 12% цукрозу. Через 5 годин недорозвинені зиготні ембріони бомбардують плазмідою або плазмідами з використанням пристрою PDS-1000/Не від BioRad. Плазміда або плазміди включають селектований маркер, такий як ген, що забезпечує резистентність до фосфінонітрицину, або маркер, що піддається кількісному аналізові, такий як зелений флуоресцентний протеїн, та ген, що кодує протеїн VІР3-класу, підготовлений для введення та експресії у кукурудзі згідно з поданим вище описом. Плазміда або плазміди осаджують на 1мкм частинках золота практично за опублікованою процедурою, запропонованою BioRad. Частинки вводять з застосуванням тиску розриву 1550фунт-сил/квадратний дюйм гелію. У кожен планшет-мішень двічі вносять композицію плазміди та частинок золота. Оскільки в одному варіанті винаходу плазміда або плазміди включають химерний ген, що кодує резистентність до фосфінонітрицину, ця речовина може бути використана для відбору трансформованих клітини in vitro. При використанні агент відбору застосовують у концентрації 10мг/л у день введення гена й збільшують її до 40мг/л приблизно через один місяць. Ембріогенний калюс, отриманий таким чином, може бути регенерований у присутності фосфінонітрицину при використанні селектованого маркера. Рослини одержують з відібраних ліній ембріогенного калюсу. Регенеровані рослини аналізують на резистентність до сприйнятливих комах. Усі рослини, які є резистентними до комах, також експресують введений химерний ген, що кодує протеїн або протеїни VІР3-класу, про що свідчить виявлення VІР3-протеїну у рослинах з застосуванням ферментного імуносорбентного тесту ELISA. Трансформують рослини, які є резистентними до комах і експресують VІР3-протеїн. Для трансформації кукурудзи з використанням ембріогенного калюсу І типу калюс одержують з недорозвинених зиготних ембріонів з використанням стандартних способів культивування. Для введення генів приготовляють приблизно 300мг калюсу І типу шляхом подрібнення за допомогою скальпеля або пересівання протягом 3-5 днів перед введенням гена. Перед введенням гена підготовлений калюс поміщають на напівтверде культуральне середовище, яке також містить 12% цукрозу. Через приблизно 4 години тканину бомбардують, використовуючи біолістичний пристрій PDS-1000/He від BioRad. Плазміда або плазміди включають селектований маркер, такий як ген, що забезпечує резистентність до фосфінонітрицину, або маркер, що піддається кількісному аналізові, такий як зелений флуоресцентний протеїн, та ген, що кодує протеїн VІР3-класу, підготовлений для введення та експресії у кукурудзі згідно з представленим вище описом. Плазміди осаджують на 1мкм частинках золота, застосовуючи практично стандартний протокол, запропонований BioRad. Приблизно через 16 годин після введення гена калюс переносять у стандартне культуральне середовище, що містить 2% цукрозу і, у разі використання селектованого маркера, до 1мг/л фосфінонітрицину. Калюс пересівають для відбору на 8 тижнів, після чого калюс, який продовжує розвиватися, переносять у стандартне регенеративне середовище для утворення рослин. Регенеровані рослини аналізують на резистентність до сприйнятливих комах. Усі рослини, які є резистентними до комах, також експресують введений химерний ген, що кодує протеїн VІР3-класу, про що свідчить виявлення VІР3-протеїну у рослині через ELISA-тест. Трансформують рослини, які є резистентними до комах і експресують протеїн VІР3-класу. ДОДАТКОВІ ЕЛЕМЕНТИ БОРОТЬБИ З КОМАХАМИ Пестицидні протеїни винаходу можуть використовуватися у поєднанні з Bt 5-ендотоксинами або іншими інсектицидними протеїнами для підвищення уражуваності комах. Крім того, використання VIP-протеїнів даного винаходу, краще - VІР3А(с)-протеїну, у поєднанні з Bt 5-ендотоксинами або іншими конкретно визначеними інсектицидними елементами має конкретне застосування для запобігання резистентності комах та/або боротьби з нею. Були класифіковані різні інсектицидні кристалічні протеїни Bacillus thuhngiensis на основі їхнього спектру активності та подібності послідовностей. Класифікація, запропонована Hofte and Whiteley, MicrobioL Rev. 53: 242-255 (1989), розбила відомі на той час інсектицидні кристалічні протеїни на чотири основні класи. Взагалі, основні класи визначають за спектром активності, коли Cry 1-протеїни є активними проти Lepidoptera, Сrу2-протеїни активні проти Lepidoptera та Diptera, Сrу3-протеїни активні проти Coleoptera, а Сгу4-протеїни активні проти Diptera. У кодному з основних класів 6-ендотоксини згруповані згідно з подібністю послідовностей. Сrу1протеїни, як правило, продукуються як 130-140кДа протоксинові протеїни, які протеолітично розщеплюються для вироблення активних токсинних протеїнів приблизно по 60-70кДа. Активна частина dендотоксину міститься у NH-термінальній частині молекули повної довжини. Вищезгадані Hofte and Whiteley розбили відомі Сrу1- протеїни на шість груп, 1Аа, 1Аb, 1Ас, 1В, 1С та 1D. З того часу були також охарактеризовані протеїни, класифіковані як Cry1Ea, Cry1Fa, Сrу9А, Сrу9С та Сrу9В. Спектр інсектицидної активності окремого d-ендотоксину з Bacillus thuringiensis значно звужується, і даний d-ендотоксин є активним лише проти кількох комах. Специфічність є результатом ефективності різних етапів вироблення активного токсинного протеїну і, як наслідок, його здатності до взаємодії з епітеліальними клітинами травного тракту комах. В одному оптимальному варіанті експресія VIP-протеїнів у трансгенній рослині супроводжується експресією одного або кількох Bt d-ендотоксинів. Особливу перевагу віддають Bt d-ендотоксинам, описаним у заявці на Патент США за серійним номером 07/951,715, на яку авторами робиться посилання. Добре відомо, що багато d-ендотоксинних протеїнів з Bacillus thuringiensis дійсно експресуються як протоксини. Ці протоксини розчиняють у лужному середовищі кишечника комах і протеолітично перетворюють за допомогою протеаз на токсичний фрагмент кору (Hofte and Whiteley, Microbiol Rev. 53: 242-255 (1989)). Для d-ендотоксинних протеїнів Сrу1-класу токсичний фрагмент кору знаходять в Nтермінальній половині протоксину. Обсяг даного винаходу передбачає, що гени, які кодують будь-якої форми протоксин повної довжини або зрізаний токсичний фрагмент кору нових токсинних протеїнів, можуть бути використані при трансформації рослинних векторів для надання інсектицидних властивостей рослинам-хазяям. Інші інсектицидні елементи включають інгібітори протеази (серинового та цистеїнового типів), лектини, a-амілазу, пероксидазу та холестеролоксидазу. Інші VIP-гени, такі як vip1A(a) та vip2A(a), як описано у заявці на Патент США за серійним номером 08/463,483, на яку авторами робиться посилання, також використовують у даному винаході. Цієї коекспресії більш ніж одного інсектицидного елемента в одній трансгенній рослині досягають шляхом генної інженерії рослини, яка б містила й експресувала усі необхідні гени. Або ж Вихідна рослина 1 може бути піддана генній інженерії для експресії VIP-протеїнів. Друга рослина, Вихідна рослина 2, може бути піддана генній інженерії для експресії додаткового елемента боротьби з комахами. Через схрещування Вихідної рослини 1 з Вихідною рослиною 2 одержують потомство, яке експресує усі гени, введені у Вихідні рослини 1 та 2. Рекомбінантні мікроорганізми, що включають гени та протеїни VIP3-класу Було визнано, що виділені гени даного винаходу, які кодують протеїн VIP3-класу, краще - VІР3А(с)протеїн, можуть бути перенесені до будь-якого мікроорганізму-хазяїна і надати цьому хазяїнові інсектицидних властивостей. Різних хазяїв для нових генів даного винаходу вибирають як таких, що є придатними для клонування, для характеризації форми та функцій гена або кодованого протеїну, для використання як хазяїв для ферментації з метою збільшення продукування токсинного протеїну, з метою більш ефективного доставляння принаймні одного токсинного протеїну до організму комахи-шкідника або введення нового гена токсину до організму патогенних комах, таких як бакуловірус (вірус ядерного поліедрозу, наприклад, Autographies califomica) для підвищення їх ефективності. За задумом, вищезгадані хазяї мають бути застосовані у середовищі або до рослини або тварин для боротьби з комахами. Відбирають мікроорганізми-хазяї, які займають своє місце у «фітосфері» (філоплані, філосфері, ризосфері та/або ризоплані) однієї або кількох культур. Ці мікроорганізми вибирають за здатністю успішно конкурувати у конкретному середовищі з дикими мікроорганізмами, вони забезпечують підтримання та експресію гена, що експресує поліпептидний пестицид, і бажано, щоб забезпечували поліпшений захист пестициду від деградації та інактивації у середовищі. Такі мікроорганізми включають бактерії, водорості та грибки. Особливо цікавими є мікроорганізми, такі як бактерії, наприклад, Bacillus, Caulobacter, Agmenellum, Pseudomonas, Erwinia, Serratia, Klebsiella, Xanthomonas, Streptomyces, Rhizobium, Rhodopseudomonas, Methyfius, Agmbacterium, Acetobacter, Lactobadllus, Arthrobacter, Azotobacter, Leuconostoc та Alcaligenes; грибки, зокрема, дріжджі, наприклад, Saccharomyces, Cryptococcus, Kluyveromyces, Sporobolomyces, Rhodotomla та Aureobasidium. Особливий інтерес викликають такі види фітосферних бактерій, як Bacillus, Pseudomonas syringae, Pseudomonas fiuorescens, Serratia marcescens, Acetobacter xylinum, Agrobacteria, Rhodopseudomonas spheroides, Xanthomonas campestris, Rhizobium melioti, Alcaligenes entrophus, Clavibacter xyli та Azotobacter vinlandii; та фітосферні види дріжджів, такі як Rhodotomla rubra, R. glutinis, R. marina, R. aurantiaca, Cryptococcus albidus, C. diffluens, С laurentii, Saccharomyces rosei, S. pretoriensis, S. cerevisiae, Sporobolomyces rosues, S. odorus, Kluyveromyces veronae та Aureobasidium pollulans. Особливий інтерес викликають пігментовані мікроорганізми. Придатні клітини-хазяї для обробки клітин, що містять пестициди, з метою продовження активності токсину у клітині при застосуванні обробленої клітини до середовища конкретних шкідників, можуть включати або прокаріоти, або еукаріоти, які зазвичай обмежуються клітинами, що не продукують речовин, токсичних для вищих організмів, таких як ссавці. Однак можуть використовуватися й організми, які продукують речовини, токсичні для вищих організмів, якщо токсин є нестійким або рівень застосування є достатньо низьким для того, щоб уникнути будь-якої можливості токсичності для ссавця-хазіїна. Як хазяї особливий інтерес викликають прокаріоти та нижчі еукаріоти, такі як грибки. Прикладами прокаріотів, як грам-негативних, так і грам-позитивних, є Enterobactenaceae, такі як Eschenchia, Erwinia, Shigella, Salmonella та Proteus, Bacillaceae; Rhizobiceae, такі як Rhizobium; Spinllaceae, такі як photobacterium, Zymomonas, Serratia, Aeromonas, Vibrio, Desulfovibrio, Spirillum; Lactobacillaceae; Pseudomonadaceae, такі як Pseudomonas та Acetobacter, Azotobacteraceae та Nitrobacteraceae. Еукаріоти включають грибки, такі як Phycomycetes та Ascomycetes, який включає дріжджі, такі як Saccharomyces та Schizosaccharromyces; а також дріжджі Basidiomycetes, такі як Rhodotorula, Aureobasidium, Sporobolomyces, і т. ін. Особливо цікавими характеристиками у відборі клітини-хазяїна для продукування є легкість введення протеїнового гена до клітини-хазяїна, доступність експресійних систем, ефективність експресії, стабільність протеїну у хазяїні та наявність додаткових генетичних можливостей. Характеристики, пов'язані з використанням як пестициду мікрокапсули, включають захисні якості для пестициду, такі як товсті стінки клітин, пігментація та внутрішньоклітинна упаковка або утворення включень; листова спорідненість; нетоксичність для ссавців; привабливість для вживання шкідниками; легкість знищення та засвоювання без шкоди для токсинів і т. ін. Серед інших характеристик - легкість формулювання й застосування, економічність, стабільність при зберіганні тощо. Особливо цікавими організмами-хазяями є такі види дріжджів, як Rhodotorula, Aureobasidium, Saccharomyces, та Sporobolomyces; філопланні організми, такі, як Pseudomonas, Erwinia та Flavobactehum; або інші подібні організми, такі як Eschehchia, Lactobacillus, Bacillus та інші. Серед конкретних організмів Pseudomonas aeurginosa, Pseudomonas fluorescens, Saccharomyces cerevisiae, Bacillus thuhngiensis, Escherichia coli, Bacillus subtilis і т. ін. Існує багато доступних способів введення гена, що експресує пестицидний протеїн, до мікроорганізмухазяїна за умов, які дозволяють стабільно підтримувати й експресувати ген. Наприклад, можуть бути побудовані експресійні касети, які включають потрібні послідовності ДНК, функціонально з'єднані з транскрипційними та трансляційними регуляторними сигналами для експресії послідовностей ДНК, і послідовність ДНК, гомологічна послідовності організму-хазяїна, у якому відбувається інтеграція, та/або система реплікації, яка функціонує у хазяїні, коли відбувається інтеграція або стабільне підтримання. Транскрипційні та трансляційні регуляторні сигнали включають, крім інших, промотор, сайт ініціації транскрипції, оператори, активатори, енхансери, інші регуляторні елементи, рибосомні зв'язувальні сайти, ініціаторний кодон, сигнали термінації і т. ін. Див., наприклад, Патент США №5,039,523; Патент США №4,853,331; ЕРО 0480762А2; Sambrook et al. supra; Molecular Cloning, A Laboratory Manual. Maniatis et al. (eds) Cold Spring Harbor Laboratory, Cold Spring Harbor, NY (1982); Advanced Bacterial Genetics, Davis et al. (eds.) Cold Spring Harbor Laboratory, Cold Spring Harbor, NY (1980) та посилання на них. Нові гени або їхні рекомбінантні форми можуть бути трансформовані у таких хазяїв з використанням багатьох загальновизнаних способів. Одним з таких оптимальних способів є електропорація клітини мікроорганізму, описана наприклад, як спосіб Дауера (Dower) (Патент США №.5,186,800). Іншим оптимальним способом є спосіб Шуртера (Schurter et al.) (МоІ. Gen. Genet. 218: 177-181 (1989)), також описаний у заявці на Патент США за серійним номером 07/353,565, на яку авторами робиться посилання у повному обсязі. Гени, що кодують VІР3-клас протеїнів, можуть бути введені у мікроорганізми, які розмножуються на рослинах (епіфіти) або у рослинах (ендофіти) для доставляння протеїнів VІР3-класу до шкідників, призначених для знищення. Багато видів бактерій здатні жити у судинних тканинах рослин. Більшість із цих ендофітів та епіфітів виявляють слабкий фізіологічний вплив на ріст та продуктивність рослин. Бактерії, які утворюють колонії у корінні, наприклад, можуть бути виділені з потрібних рослин відомими спеціалістам способами. Так штам Bacillus cereus, який утворює колонії у корінні, може бути виділений з коріння рослини (наприклад, див. J. Handelsman, S. Raffel, Е. Mester, L. Wunderlich and С. Grau, Appl. Environ. Microbiol. 56:713-718, (1990)). VІР3-гени також можуть бути введені у Bacillus cereus, що утворює колонії у корінні, з застосуванням стандартних способів, відомих спеціалістам. Так ген, що кодує протеїн VІР3-класу, який походить від штаму АВ88, може бути введений у Bacillus cereus, що утворює колонії у корінні, за допомогою кон'югації з застосуванням стандартних способів (J. Gonzalez, В. Brown and В. Cariton, Proc. Natl. Acad. Sci. 79:6951-6955, (1982)). Нові гени винаходу також можуть бути введені у Bacillus, що утворює колонії у корінні, шляхом електротрансформації. Наприклад, vip3A(a) може бути клонований у шатл-вектор, наприклад, рНТ3101 (D. Lereclus et al., FEMS Microbiol. Letts., 60:211-218 (1989)). Шатл-вектор рНТ3101, який містить кодуючи послідовність, може бути трансформований у Bacillus, що утворює колонії у корінні, шляхом електропорації (D. Lereclus et al. 1989, FEMS Microbiol. Letts. 60:211-218). Можливе також використання Bacillus megatehum, що утворює колонії у бавовні. Іншим прикладом є ендофіт Clavibacter xyli з роду/виду, який містить фітопатогеннні бактерії, що уповільнюють ріст рослин. Ця бактерія може виростати до дуже високого рівня у судинній системі рослин. dендотоксин вводили у цей ендофіт, який після цього інокулювали у рослину, що забезпечувало ефективну боротьбу з метеликом кукурудзяним. Відомі також інші ендофіти. Експресійні системи можуть бути побудовані таким чином, щоб VIP3-протеїни секретувалися за межами цитоплазми грам-негативних бактерій, наприклад, Е. соІі. Переваги секретування VІР3-протеїнів полягають у тому, що (1) це може підвищити рівень експресованого VІР3-протеїну і (2) може сприяти ефективному очищенню VІР3-протеїну. Секреція VІР3-протеїнів Е. соІі може забезпечуватися, наприклад, шляхом злиття відповідного сигнального пептиду Е. соІі з аміно-термінальним кінцем \/ІРЗ-сигнального пептиду або заміщення VІР3сигнального пептиду сигнальним пептидом Е. соІі. Сигнальні пептиди, розпізнані Е. соІі, знаходять у протеїнах, які секретуються в Е. соїі, наприклад, OmpA-протеїні (J. Ghrayeb. H. Kimura, M. Takahara, Y. Masui and М. Inouye, EMBO J., 3:2437-2442 (1984)). ОmрА є основним протеїном зовнішньої мембрани Е. соlі і, таким чином, його сигнальний пептид має бути ефективним у процесі транслокації. Також немає необхідності у модифікації OmpA-сигнального пептиду перед обробкою, як у випадках з іншими сигнальними пептидами, наприклад, сигнальним пептидом ліпопротеїну (G. Duffaud, P. March and M. Inouye, Methods in Enzymology 153:492 (1987)). Так, унікальні BamHI сайти рестрикції можуть бути введені на аміно-термінальному та карбокситермінальному кінцях VIP кодуючих послідовностей з застосуванням стандартних способів, відомих спеціалістам. BamHI фрагменти можуть бути клоновані у каркасі у вектор plN-lll-ompA1, A2 або A3 (J. Ghrayeb, Н. Kіmurа, М. Takahara, Н. Hsiung, Y. Masui and М. Inouye, EMBO J., 3:2437-2442 (1984)), утворюючи таким чином ompA: VIP злитий ген, який секретується у периплазматичний простір. Інші сайти рестрикції у полілінкері pIN-lll-ompA можуть бути видалені відомими спеціалістам стандартними способами, щоб VIP3 аміно-термінальна амінокислотна кодуюча послідовність розміщувалася безпосередньо за сайтом розщеплення ompA-сигнального пептиду. Таким чином, секретована VІР3-послідовність в Е. соlі має бути ідентичною природній VІР3-послідовності. Якщо VIP3 природний сигнальний пептид не потрібен для належного складання сформованого протеїну, такі сигнальні послідовності можуть бути видалені й заміщені ompA-сигнальною послідовністю. Унікальні BamHI сайти рестрикції можуть бути введені на аміно-кінцях пропротеїнової кодуючої послідовності безпосередньо після кодуючої послідовності сигнального пептиду VIP3 і на карбокси-кінці VІР3-кодуючої послідовності. Ці BamHI фрагменти після цього можуть бути клоновані у pIN-lll-ompA вектори, як описано вище. Спеціалістам відомі загальні способи застосування штамів винаходу для пестицидного контролю або інженерії інших організмів як пестицидних агентів. Див., наприклад Патент США №5,039,523 та ЕР 0480762А2. VIP3 можуть бути ферментовані у бактеріях-хазяях, а одержані в результаті бактерії обробляють і використовують для розпилювання мікробів так само, як штами Bacillus thuhngiensis використовували в інсектицидних розпилювачах. У разі з VIP3, який секретується з Bacillus, сигнал секреції видаляють або мутують із застосуванням відомих спеціалістам процедур. Такі мутації та/або видалення перешкоджають секреції VІР3-протеїну(ів) у середовище росту у процесі ферментації. VІР3-протеїни утримуються у клітині, а клітини після цього обробляють для отримання інкапсульованого VІР3-протеїну. З цією метою використовують будь-який придатний мікроорганізм. Psuedomonas використовували для експресії ендотоксинів Bacillus thuringiensis як інкапсульованих протеїнів, а одержані в результаті клітини обробляли й розпилювали як інсектицид (Н. Gaertner et al. 1993, In Advanced Engineered Pesticides, L Kirn ed.). Таким чином використовують різні штами Bacillus thuringiensis. Такі штами Bt продукують ендотоксинний(і) протеїн(и), а також VIP3. Або ж, такі штами можуть продукувати лише VIP3. Штам Bacillus subtilis з недостатньою споруляцією продукує високий рівень Сrу3А ендотоксину з Bacillus thuringiensis (Agaisse, H. and Lereclus, D., «Expression in Bacillus subtilis of the Bacillus thuringiensis CryifIA toxin gene is not dependent on a sporulation-specific sigma sigma factor and is increased in a spoOA mutant», J. Bacteriol., 176:4734-4741 (1994)). Подібний spoOA мутант може бути приготовлений у Bacillus thuringiensis і використаний для продукування інкапсульованого VIP3, який не секретується у середовище, а зберігається у клітині. Культури, які мають бути піддані захистові у межах даного винаходу, включають, наприклад, такі види рослин: злаки (пшениця, ячмінь, жито, овес, рис, сорго та інші подібні культури), буряки (цукровий буряк та кормовий буряк), фуражні трави (грястиця, вівсяниця і т. ін.), кісточкові, насіннячкові та ягоди (яблуня, груша, слива, персик, мигдаль, вишня, суниця, малина та ожина), бобові рослини (боби, сочевиця, горох, соя), олійні рослини (рапс, гірчиця, мак, оливки, соняшник, кокос, рицина, какао-боби, арахіс), огіркові рослини (огірок, кабачок, диня) волокнисті рослини (бавовна, льон, коноплі, джут), цитрусові (апельсин, лимон, грейпфрут, мандарин), овочі (шпинат, салат, спаржа, капуста та інші Brassicae, цибуля, томати, картопля, паприка), Lauraceae (авокадо, морква, коричне дерево, камфор), листяні та хвойні дерева (наприклад, липа, тис, дуб, вільха, тополя, береза, ялина, модрина, сосна) або такі рослини як кукурудза, тютюн, горіхи, кофейне дерево, цукрова тростина, чай, виноград, хміль, банан та каучукові рослини, а також декоративні рослини (включаючи складноцвітні). Мікроорганізми які були генетично змінені для того, щоб містити пестицидний ген та протеїн, можуть бути використані для захисту сільськогосподарських культур та продуктів від шкідників. В одному аспекті винаходу цілі, тобто нелізовані клітини організму, що продукує токсин (пестицид) обробляють реагентами, які подовжують активність токсину, виробленого у клітині, коли клітину застосовують у середовищі шкідника(ів). Або ж пестициди продукують шляхом гетерологічного гена у клітинах-хазяях. Експресія гетерологічного гена в результаті, безпосередньо або опосередковано, веде до продукування й збереження пестициду всередині клітин. Ці клітини потім обробляють в умовах, які подовжують активність токсину, виробленого у клітині, коли клітину застосовують у середовищі шкідника(ів). Виникаючий в результаті продукт зберігає токсичність токсину. Ці природно інкапсульовані пестициди після цього можуть бути формульовані згідно з традиційними технологіями для застосування у середовищі - хазяїні шкідника, наприклад, у грунті, воді або листі рослини. Див., наприклад ЕРА 0192319 та посилання на нього. Активні інгредієнти даного винаходу, як правило, застосовують у формі композицій на площах, зайнятих під культури, або наносять на рослини, які підлягають обробці, одночасно або переміжно з іншими сполуками. Ці сполуки можуть бути добривами або джерелом мікронутрієнтів, або іншими композиціями, які сприяють розвитку рослин. Вони також можуть бути гербіцидами, інсектицидами, фунгіцидами, бактерицидами, нематицидами, молюскоцидами або, у разі потреби, сумішами кількох цих композицій разом з іншими прийнятними у сільському господарстві носіями, поверхнево-активними речовинами або ад'ювантами, які традиційно застосовують при розробці. Придатні носії та ад'юванти можуть бути твердими або рідкими і відповідають речовинам, традиційно застосовуваним при розробці рецептури, наприклад, природним або регенерованим мінеральним речовинам, розчинникам, диспергаторам, зволожувачам, речовинам для підвищення клейкості, в'яжучим речовинам або добривами. Оптимальними способами застосування активного інгредієнта даного винаходу або агрохімічної композиції даного винаходу, що містить принаймні один з пестицидних протеїнів, продукованих штамами бактерій даного винаходу, є нанесення на листя, вкривання насіння та нанесення на грунт. Кількість прийомів та норми застосування залежать від інтенсивності нападів тих чи інших шкідників. Ентомоцидні композиції, що включають рекомбінантний штам Bacillus thuringiensis Даний винахід також забезпечує ентомоцидну композицію, що включає рекомбінантний штам Bacillus thuhngiensis, який містить принаймні один з нових генів токсинів у рекомбінантній формі або його похідні або мутанти разом з таким сільськогосподарським ад'ювантом, як носій, розріджувач, поверхнево-активна речовина або ад'ювант, що полегшує нанесення. Композиція також може містити іншу біологічно активну сполуку, вибрану з-поміж добрив, джерел мікронутрієнтів, композицій для росту рослин, гербіцидів, інсектицидів, фунгіцидів, бактерицидів, нематицидів та молюскоцидів, а також їх сумішей. Композиції можуть включати від 0,1 до 99% за масою рекомбінантного штаму Bacillus thuringiensis, що містить принаймні один з нових генів у рекомбінантній формі, або його похідні або мутанти, від 1 до 99,9% за масою твердого або рідкого ад'юванту та від 0 до 25% за масою поверхнево-активної речовини. Рекомбінантний штам Bacillus thuringiensis, що містить принаймні один з нових генів у рекомбінантній формі, або композиція, що його містить, може бути введений у рослину або культуру, яка потребує захисту, разом з деякими іншими інсектицидами або хімікатами (1993 Crop Protection Chemicals Reference, Chemical and Pharmaceutical Press, Canada) без втрати ефективності. Він є рівноцінним іншим загальновживаним матеріалам, які застосовують у сільському господарстві для розпилення, але його неможна використовувати у розчинах для розпилювання з сильною лужною реакцією. Його можна застосовувати як порошок, суспензію, порошок для змочування або у будь-якій іншій формі, придатній для застосування у сільському господарстві. Рекомбінантний штам Bacillus thuringiensis, що містить принаймні один з нових генів у рекомбінантній формі, зазвичай застосовують у формі ентомоцидних композицій і можуть застосовувати на ділянках або до рослин, які підлягають обробці, одночасно або переміжно з іншими біологічно активними сполуками. Ці сполуки можуть бути добривами або джерелом мікронутрієнту, або іншими композиціями, які сприяють розвиткові рослин. Вони також можуть бути гербіцидами, інсектицидами, фунгіцидами, бактерицидами, нематицидами, молюскоцидами або сумішами кількох цих композицій, якщо потрібно - разом з іншими традиційно застосовуваними спеціалістами носіями, поверхнево-активними речовинами або ад'ювантами, що полегшують нанесення. Придатні носії та ад'юванти можуть бути твердими або рідкими й відповідають речовинам, які зазвичай застосовують при розробці, тобто природним або регенерованим мінеральним речовинам, розчинникам, диспергаторам, зволожувачам, речовинам для підвищення клейкості, в'яжучим речовинам або добривам. Ентомоцидні композиції, композиції або суміші, що містять рекомбінантний штам Bacillus thuringiensis, який містить новий ген у рекомбінантній формі як активний інгредієнт, або його комбінацію з іншими активними інгредієнтами і, якщо потрібно, з твердим або рідким ад'ювантом, приготовляють традиційним способом, наприклад, шляхом змішування до однорідного стану та/або розтирання активних інгредієнтів з розріджувачами, наприклад, розчинникам, твердими носіями та, у деяких випадках, поверхнево-активними сполуками (поверхнево-активними речовинами). Розчинники, носії, поверхнево-активні речовини, поверхнево-активні сполуки і т. ін., які традиційно застосовують при розробці композицій і які можуть бути використані у межах даного винаходу, описані, наприклад, у WO 96/10083. Іншою оптимальною характеристикою ентомоцидної композиції даного винаходу є стійкість активного інгредієнта при застосуванні до рослин та грунту. Можливими причинами втрати активності є інактивація ультрафіолетовим світлом, висока температура, ексудати листя та рН. Наприклад, при високому рівні рН, зокрема, у присутності відновника, кристали d-ендотоксину розчиняються і стають більш придатними для протеолітичної інактивації. Високий рівень рН у листі також може бути важливим, особливо якщо на поверхні листа він становить рН 8-10. Розробка ентомоцидної композиції даного винаходу може бути застосована для розв'язання цих проблем або через включення добавок, які допомагають запобігти втраті активного інгредієнта, або інкапсуляції матеріалу таким чином, щоб активний інгредієнт був захищений від інактивації. Інкапсуляцію здійснюють хімічним (McGuire and Shasha, 1992) або біологічним шляхом (Barnes and Cummings, 1986). Хімічна інкапсуляція включає процес, згідно з яким активний інгредієнт вкривають полімером, тоді як біологічна інкапсуляція включає експресію d-ендогенних токсинів у мікроорганізмі. Для біологічної інкапсуляції використовують цілий мікроорганізм, що містить d-ендотоксинний протеїн як активний інгредієнт у композиції. Додавання засобів захисту від ультрафіолетових променів може ефективно знизити шкоду від опромінення. З інактивацією через високу температуру також можна боротися шляхом включення відповідної добавки. Ентомоцидні композиції, які зазвичай містять від 0,1 до 99%, краще - від 0,1 до 95% рекомбінантного штаму Bacillus thuringiensis, що містить принаймні один з нових генів у рекомбінантній формі, або його комбінацію з іншими активними інгредієнтами, від 1 до 99,9% твердого або рідкого ад'юванту та від 0 до 25%, краще - від 0,1 до 20%, поверхнево-активної речовини. Оскільки комерційні продукти в оптимальному варіанті приготовляють як концентрати, споживач, як правило, застосовує розведені композиції значно нижчої концентрації. Ентомоцидні композиції також можуть містити інші інгредієнти, такі як стабілізатори, протиспінювачі, регулятори в'язкості, в'яжучі речовини, речовини для підвищення клейкості, а також добрива або інші активні інгредієнти для отримання потрібного ефекту. Способи боротьби з комахами З огляду на попередній опис винаходу стає зрозумілим, що існує кілька способів, згідно з якими з комахами можна боротися, використовуючи протеїни VІР3-класу як інсектицидний елемент, або один, або у поєднанні з допоміжними елементами боротьби з комахами, такими як d-ендотоксини. Будь-який спосіб введення VІР3-протеїну для вживання сприйнятливою комахою веде до загибелі цієї комахи. В одному варіанті винаходу рослини трансформують геном, що кодує протеїн VІР3-класу. Експресія протеїну може відбуватися у будь-який момент під час росту та розвитку рослини, залежно від особливостей комахи, на яку спрямовано боротьбу. Наприклад, протеїн VІР3-класу згідно з винаходом може бути експресованим у корінні, стеблах, листі, насінні, пилку і т. ін. Він забезпечує перевагу експресії протеїну лише у клітинах або тканинах, де живиться комаха, на яку спрямовано боротьбу. Вживання сприйнятливою комахою клітин або тканин рослини, що експресує VIP3 протеїн, веде до загибелі цієї комахи. В одному варіанті винаходу VІР3-протеїн експресується у стеблах або черешках рослини для боротьби з совкою іпсилон. Рослини вирощують або у польових, або у тепличних умовах. Насіння, що містить VІР3-протеїн, також може бути захищене від комах при зберіганні. ПРИКЛАДИ У прикладах з 16 по 18 на сторінках з 73 по 82 WO 96/10083 описано виділення та біологічна специфікація штамів Bacillus thuringiensis AB88 та АВ424, очищення та характеризація VІР3А(а)-протеїну, клонування vip3A(a) та vip3A(b) генів та ідентифікація нових іф-генів шляхом гібридизації. Нами робиться посилання на вищезгадані приклади у їх повному обсязі. У представлених нижче прикладах також описуються матеріали та способи, застосовані при втіленні винаходу, та результати цього втілення. Вони пропонуються як ілюстрація, і їх перерахування не повинно розглядатися як таке, що обмежує заявлений винахід. Приклад 1: Присутність vір3-подібних генів та VІР3-подібних протеїнів в ізолятах Bacillus Інші ізоляти Bacillus, крім АВ88, виявили інсектицидну активність проти личинок Lepidoptera при випробуванні супернатантів використаних культур. Деякі ізоляти, які були активними проти совки іпсилон, аналізували на присутність VІР3-подібних генів та на продукування VІР3-подібних протеїнів. Стандартний аналіз шляхом полімеразної ланцюгової реакції (PCR) застосовували для того, щоб визначити, чи містять активні проти совки іпсилон ізоляти Bacillus vір3-подібний ген. Використовуючи пару праймерів PCR GW110 (5’-CGA ТТА ATG TTG GCC ТС-3'; SEQ ID NO:17) та GW111 (S'-CAT TAG CAT СТС CGG АСА CAG-3'; SEQ ID NO:18), виявили, що усі активні проти совки іпсилон ізоляти продукують 728 п. н. продукт vip3-гена, який за розміром дорівнював тому, який продукується типовим штамом, АВ88. Один ізолят Bacillus, AB51, який не був активним проти совки іпсилон, продукував vip3-продукт того самого розміру. Жоден з інших неактивних проти совки іпсилон ізолятів Bacillus не продукував vip3-продукту PCR. Аналіз продукування VІР3-протеїну здійснювали з застосуванням стандартної процедури вестернблотингу. Антитіла, вироблені проти VIP3A(a)-протеїну, описані у попередньому прикладі, використовували для виявлення імунореактивних протеїнів. Аліквотні проби супернатантів вільних від клітин спорульованих культур піддавали електрофорезові з додецилсульфатом натрію в поліакриламідному гелі (SDS-PAGE) з застосуванням стандартних способів. Після цього здійснювали стандартний вестерн-блотинг для визначення присутності VІР3-подібних протеїнів. Усі ізоляти Bacillus, які мали 728 п. н. продукту PCR і були активні проти совки іпсилон, продукували 80кДа протеїн, який був імунореактивним до VІР3А(а)-антитіла. Ізолят АВ51, який мав належного розміру vip3-продукт PCR, але не був активним проти совки іпсилон, продукував імунореактивний протеїн, який був зрізаним, що свідчило про те, що це може бути причиною відсутності біологічної активності проти совки іпсилон. Приклад 2: Характеризація штаму Bacillus thuringiensis AB51, що містить vір3-подібний ген Штам Bacillus thuringiensis, позначений як АВ51, виявив вміст протеїнів VІР3-класу при вестернблотингу з використанням поліклональних антитіл кроля проти Vip3A(a). \/ір3-подібний ген клонували у pKS, що утворює рСІВ7112. Цей ген позначали як vip3A(c). Послідовність ДНК для vip3A(c) описано у SEQ ID NO:5, а послідовність кодованого протеїну описано у SEQ ID NO:6. VIP3A(c)-протеїн має довжину 746 амінокислот, на 43 амінокислоти коротше його гомологи VIP3A(a) та VIP3A(b). Приклад 3: Вироблення антитіл до Vip3А(а)-протеїну Антисироватку проти очищеного Vip3A(a) інсектицидного протеїну продукували у кролях та козах. Для кролів зв'язаний нітроцелюлозою протеїн (50мкг) розчиняли у DMSO, емульгували повним ад'ювантом Фройнда (Difco) і вводили підшкірно двічі на місяць протягом трьох місяців. Для кіз активний розчинний чистий \/ір3А-протеїн (300мкг) вводили внутрішньом'язово двічі на місяць протягом трьох місяців. У них брали кров через 10 днів після другої та третьої ін'єкцій і зі зразків крові відновлювали сироватку (Hahow, E. and Lane, D. Antibodies: A Manual Laboratory, Cold Spring Harbor Lab. Press, NY, 1988). Антисироватки після цього фракціонували шляхом афінної хроматографії, використовуючи протеїн А стафілококу, і одержану в результаті lgG-фракцію далі очищали шляхом фільтрації через колонку, яка містила іммобілізований лізат Е. coli (Yu, C.G. etal. Appl. Environ. Microbiol. 63:532-536 (1997)). Специфікацію антисироваток кролів та кіз складали, аналізуючи VIP3A(a)-протеїн шляхом вестернблотингу. Протеїни відокремлювали за допомогою SDS/PAGE і переносили на нітроцелюлозу. Плями нітроцелюлози блокували у суміші 20мМ Tris-HCI, рН 7,5 / 0,15М NaCI / 0,02% NaN3 / 5% нежирного сухого молока. Потім їх проявляли з використанням або кролячих, або козячих антитіл проти Vip3A(a) у концентрації 200нг/мл або 100нг/мл відповідно. Кон'юговані лужною фосфатазою антитіло кози IgG проти кроля або антисироватку кроля проти кози використовували як вторинні антитіла у концентрації 1мкг/мл (Kirkegaard & Perry Laboratories, Inc.). Бромхлоріндоїлфосфат та нітротетразол використовували як субстрат для реакції лужної фосфатази. Обидва анти-Vip3A(a) антитіла, і від кроля, і від кози, є поліклональними. Анти-Vip3А(а) антитіла, отримані від кози, мають вищий титр, ніж отримані від кроля. При експериментальному підході анти-Vір3А(а) антитіла кроля мають використовувати як розведені у пропорції 1/500 з первісної сироватки (200нг/мл). Якщо порівняти, анти-Vір3А(а) антитіла, отримані від кози, можуть бути розведені до 1/2000 (100нг/мл) з первісної сироватки. Якщо отримані від кроля антитіла лише розпізнають N-термінальну частину VІР3А(а)-протеїну, то антитіла, отримані від кіз, реагують з епітопами, присутніми по всій довжині VІР3А(а)-протеїну. Приклад 4: Побудова касет експресії у рослинах Касети експресії у рослинах складаються з промоторів, які можуть стимулювати експресію кодуючої послідовності або конститутивно, або тканиноспецифічно, кодуючих послідовностей, які мають бути експресовані, та термінуючих послідовностей, які дозволяють здійснювати поліаденілювання мРНК та її належну трансляцію. Промотори, вибрані у ДНК послідовностях даного винаходу, включають конститутивні промотори, такі як промотор з гена убіквітину кукурудзи (Christensen et al. Plant Мої. Biol. 12:619-632, 1989) (рСІВ8029, Фігура 4; рСІВ8055, Фігура 5; рСІВ9806, Фігура 6), та тканиноспецифічні промотори, такі як промотори з металотіонеїноподібного гена кукурудзи (de Framond, A. FEBS 290:103-106, 1991) (рСІВ8030. Фігура 7; рСІВ8056, рСІВ9805), який забезпечує оптимальну для коріння експресію, з гена РЕРС кукурудзи (Hudspeth. R.L. and Grula, J.W. Plant МоI. Biol. 12:579-589, 1989) (pCIB5535, Фігура 8; pCIB9807), який забезпечує специфічну для зеленої тканини експресію, та з протеїну неспецифічного перенесення ліпідів ячменю LTP4 (рСІВ9819, Фігура 9) (Molina, A. and Garcia-Olmedo, F. Plant J. 4:983-991, 1993), який забезпечує оптимальну для стебел експресію. Усі послідовності, використані у даному винаході, містять послідовність термінатора, яка походить від 35S CaMV, та інтрон 9, що походить від гена РЕРС кукурудзи, для посилення експресії генів. Плазміди рСІВ8029, рСІВ8055 та рСІВ9806 містять інтрон №1 гена убіквітину кукурудзи, розміщений між убіквітиновим промотором кукурудзи та viр3А(а)-геном. Послідовність, що включає кодуючу послідовність vір3А(а)-гена, інтрон №9 та 35S термінуючу послідовність, вводили шляхом інженерії у реципієнтну плазміду, що несе різні промотори як подвійні гідролізати BamHI-EcoRI. Касети експресії у рослинах використовували як такі в експериментах з трансформації рослин, або ж їх лінеаризували через використання рестрикційних ферментів, які приєднують до AmpR гена каркасної плазміди. У деяких експериментах фрагменти, що включають промотор, потрібний ген, інтрон та термінатор, виділяли з решти каркасу плазміди шляхом рестрикційного розщеплення та очищення фрагмента. У цих випадках очищення фрагмента здійснювали так: 500мкг ДНК розщеплюють відповідним ферментом і відокремлюють на 0,8% агарозному гелі. Потрібний фрагмент ідентифікують, вирізають з гелю й очищають з використанням фільтру Durapore Millipore (0,45мкм). Фільтрат, що містить фрагмент, осаджують ацетатом натрію та етанолом. Фрагмент ресуспендують у ТЕ і використовують в експериментах з трансформації. Приклад 5: Інсектицидна активність кукурудзи, що експресує VІР3А(а) Кукурудзу, що експресує VІР3А(а)-протеїн, перевіряли на інсектицидну дію на види комах, перелічені нижче у Таблиці, з застосуванням такої процедури. Від листя трансгенної та контрольної кукурудзи відрізали від одного до чотирьох фрагментів по 4см. Кожен фрагмент листя поміщали на зволожений фільтрувальний диск у 50X9мм чашці Петрі. П'ять новонароджених зразків випробуваних видів поміщали на кожен фрагмент листя, з загальною кількістю по 5-20 личинок, які випробували для кожної рослини. Чашки Петрі інкубували при 30°С у темряві. Смертність оцінювали через 48-72 години. Результати показані у Таблиці 16. Таблиця 16 Випробувані види комах Шкідники Кукурудзи Совка іпсилон (Agrotis ipsilon) Осінній похідний черв'як (Spodoptera frugiperda) Вогнівка цукрової тростини (Diatrea saccharalis) Метелик кукурудзяний південно-західний (Diatraea grandiosella) Совка бавовняна (Helicoverpa zea) Метелик кукурудзяний середземноморський(Sesamia nonagroides) Інші шкідники Lepidoptera Наземна совка (S. exigua) Похідний черв'як жовто-смугастий (S. omithogalli) П'ядун капустяний (Thchoplusia nі) Відсоток смертності VIP3A(a) Контроль 100 100 100 100 100 100 0 0 0 0 10 15 100 100 100 0 0 20 Приклад 6: Експресія vір3А(а) у кукурудзі Трансформації інбредних ліній елітної кукурудзи Ciba CG00526 та 2154 за допомогою Vір3-гена досягали з застосуванням бомбардування частинками тканини калюсу І типу. Для трансформації з використанням ембріогенного калюсу І типу калюс отримували з зиготних ембріонів з застосуванням стандартних способів і інокулювали 1-2 дні перед бомбардуванням. Тканину калюсу підготовляли до бомбардування, розміщуючи ~20 фрагментів діаметром по 3-5мм у формі кільця на культуральному середовищі, що містило 12% цукрозу. Тканину калюсу поміщали на середовище на чотири години перед бомбардуванням. ДНК, використана для трансформації калюсу кукурудзи, являла собою кругову плазміду ДНК, лінійну плазміду ДНК або очищені фрагменти ДНК, що містять Vір3-ген, під контролем різних рослинних промоторів. В експериментах, де використовують селектабельний агент, ген забезпечував резистентність до фосфінонітрицину або ріст у присутності манози. Фрагменти плазмід або ДНК, виділені шляхом фільтрації, осаджували на 0,3мкм частинках золота згідно з опублікованою процедурою від BioRad Laboratories, Hercules, CA. Частинки золота подавали, застосовуючи тиск розриву 650фунт-сил/квадратний дюйм гелію. Кожен планшет-мішень двічі обстрілювали вкритими ДНК частинками. Через шість-двадцять годин після бомбардування CG00526 калюс переносили до стандартного культурального стабілізуючого середовища. Через сім днів після бомбардування тканину переносили до середовища, яке містило селекційний агент Basta у концентрації 100мг/л. Basta являє собою комерційну композицію глюфозинатамонію виробництва Hoechst. Калюс 2154 зберігали на 12% цукрозі протягом 1-7 днів після бомбардування й переносили до стандартного культурального середовища, що містило 20-30мг/л Basta на 7 день. 2154 та CG00526 калюс інокулювали у присутності 30 або 100мг/л Basta, відповідно, протягом восьми тижнів. Тканину, яка пройшла відбір, інокулювали на нижчих рівнях Basta (5-40мг/л) протягом періоду приблизно від п'яти до десяти тижнів, щоб можна було зібрати тканину, а потім переносили на стандартне регенераційне середовище без відбору на продукування рослин. Взагалі, 12% підданих бомбардуванню фрагментів калюсу давали трансформований калюс, який проходив Basta-відбір. Окремі трансформовані калюси, як правило, регенеруються, даючи 20-30 рослин. Події отримували в експериментах, у яких не застосовували відбору. У цих експериментах калюс вирощували протягом періоду 9-10 тижнів на стабілізуючому середовищі перед перенесенням до регенераційного середовища. Подія 1337 є прикладом трансформованої VІР3-події в результаті експерименту з трансформації без маркера, що піддається селекції або кількісному аналізові, шляхом скринінгу рослин на інсектицидну активність. Трансформовані калюси також отримували в експериментах, у яких застосовували відбір манози. У цих трансформаціях ген манозо-6-фосфат-ізомерази під контролем убіквітинового промотора кукурудзи рСІВ9818 бомбардували Vір3-геном. Манозу при 0,5-1,5% включали у стабілізуюче середовище протягом періоду у дванадцять тижнів і не включали у регенераційне середовище. Трансгенні рослини оцінювали на експресію VІР3А(а)-протеїну шляхом біоаналізу комах та твердофазного імуноферментного аналізу (ELISA). Фрагменти листя брали з рослин на стадії 2-4 листків для оцінки, використовуючи біоаналізи совки іпсилон та осіннього похідного черв'яка. Біоаналізи здійснювали з використанням десяти щойно інкубованих личинок, поміщених у посудини з фрагментами листя. Розраховували відсоток смертності за 72 години. Тканини трансгенних рослин також аналізували шляхом ELISA-аналізу з використанням стандартних протоколів для кількісного визначення рівня Vір3протеїну у різних тканинах рослин. Брали тканини рослин, і у Таблиці 17 показані характерні події та їхні відповідні результати біоаналізу комах. Трансгенну кукурудзу трансформували різними плазмідами, що містили Vір3-ген, під контролем різних промоторів, таких як промотор РЕР-карбоксилази кукурудзи (РЕРС), убіквітиновий промотор кукурудзи (Ubi) та металотіонеїноподібний промотор кукурудзи (MTL). Геном селектованого маркера був РАТ-ген під контролем убіквітинового промотора кукурудзи у pUBIAC. Серед характерних подій, перелічених у Таблиці 17, показані події, продуковані з різними плазмідами або фрагментами ДНК, що походять від плазмід. Фрагменти ДНК отримували з застосуванням розщеплення рестрикційного ферменту та фракціонування за розміром з застосуванням електрофорезу в 0,8% агарозних гелях. Фрагменти ДНК вирізали з гелів, заморожували, подрібнювали й очищали шляхом фільтрування крізь 0,45мкм фільтри DuraPore Millipore з наступним осадженням етанолу. Трансформовані події кукурудзи отримували за допомогою кругової плазміди ДНК рСІВ5535, що містить Vір3-ген, під контролем РЕРС-промотора кукурудзи. Події також трансформували лінійною плазмідою ДНК рСІВ5535 та рСІВ8029, що містить Vip3-гeн під контролем убіквітинового промотора кукурудзи. Додаткові події отримували шляхом бомбардування очищених фрагментів ферменту рестрикції ДНК, що містять лише Vip3-гeн з промотором. Фрагменти, які відповідають Vip3-генові, включають: 4906bp EcoRI/Hindlll фрагмент із рСІВ5535 з РЕРС промотором кукурудзи; 5142bp Kpnl/Hindlll фрагмент із рСІВ8030 з MTL промотором; 4597bp Kpnl/Hindlll фрагмент рСІВ8029 з убіквітиновим промотором кукурудзи; 4818bp Hindlll фрагмент рСІВ8055 з убіквітиновим промотором кукурудзи; 5364 Hindlll фрагмент рСІВ8056 з MTL промотором; 5964 AscI фрагмент рСІВ9805 з MTL промотором; 5418bp AscI фрагмент рСІВ9806 з убіквітиновим промотором кукурудзи та 5727bp AscI фрагмент рСІВ9807 з РЕРС промотором кукурудзи. Таблиця 17 Подія № 891 906 946 Використана плазміда рСІВ5535 рСІВ5535 та рСІВ8029 рСІВ5535 та рСІВ8030 Химерний ген РЕРС:vip3А(а) PEPC:vip3A(a) та Ubi:vip3A(a) PEPC:vip3A(a) та MTL:vip3A(a) Смертність (%) Fall Armyworm Black Cutworm 100 100 100 100 100 100 Приклад 7: Інсектицидна активність кукурудзи, що містить Vір3 та Bt d-ендотоксини VIP3A(a) має слабку активність проти метелика кукурудзяного європейського (ЕСВ). Для забезпечення рослин, здатних боротися з багатьма комахами ряду Lepidoptera, кукурудзу, що містить ген vip3A(a), схрещували з кукурудзою, що містить сrу1В, який є активним проти ЕСВ. Потомство від схрещування піддавали біоаналізові на протистояння ЕСВ та осінньому похідному черв'якові (FAW), як описано у Прикладі 1. Результати показані у Таблиці 18. Приблизно 34% потомства не було активним проти жодного з цих видів, 15,4% були активними лише проти ЕСВ, 23,1% були активними лише проти FAW, a 27,9% були активними проти обох видів. Рослини, активні проти обох видів, містили і VIP3A(a), і Сrу1В протеїни. Подібні результати були одержані і з використанням інших Вt d-ендотоксинів, зокрема Сrу1Аb або Сrу9С. Таблиця 18 Схрещування VIP3A(a) X Cry1B % активних проти ЕСВ 15,4 % активних проти FAW 23,1 % активних проти ЕСВ та FAW 27,9 % неактивних 34,6 Приклад 8: Лізис за допомогою VІР3А(а) епітеліальних клітин середнього кишечника сприйнятливих комах Досліди з вживанням та очищенням кишечника. Тимчасову послідовність симптомів після вживання раціону з вмістом VIP3A(a) сприйнятливими комахами - личинками совки іпсилон (BCW) на другій стадії розвитку - занотовували з часу першого введення до загибелі личинки. Личинки, які вживали цей раціон, демонстрували активне живлення, після чого відбувався безперервний парасталсис кишечника. Але додавання VIP3A(a)-протеїну до раціону мало значний вплив на харчову поведінку. При додаванні у концентраціях не вище 4нг на см2 личинки продовжували час від часу живитися протягом періодів по 1020хв. Наявність синього кольору у їхньому кишечнику свідчила про живлення, але вміст очищеного кишечника значною мірою змінювався, судячи зі зменшення кількості екскрементів. При додаванні до раціону 4нг VIP3A(a) на см2 розвиток личинок помітно порушувався через 48 годин інкубаційного періоду, але смертності не спостерігалося. При концентраціях 40нг Vip3A(a) на см2 у личинок виникав параліч кишечника після вживання незначної кількості раціону, і не спостерігалося екскрементів, що свідчило про майже повну відсутність очищення кишечника. У цих умовах спостерігали приблизно 50% смертність через 48 годин. При концентраціях, вищих за 40нг VIP3A(a) на см2 личинки гинули вже після кількох укусів, без екскрементів, і смертність сягала 100%. При проведенні подібних експериментів з осіннім похідним черв'яком, також сприйнятливою комахою, спостерігали схожу схему поведінки. А от метелик кукурудзяний європейський не змінював харчову поведінку навіть при додаванні VІР3А(а)-протеїн до раціону у концентраціях не менше 400нг VIP3A(a) на см2. Гістологічні спостереження впливу VІР3А(а)-протеїну. Гістопатологічні спостереження впливу VІР3А(а)протеїну на BCW здійснювали з личинками на другій та третій стадіях розвитку, які живилися раціоном, що містить VIP3A(a). Аналіз розрізу кишечника BCW показав значне пошкодження епітелію середнього кишечника, що свідчило про те, що тканини середнього кишечника є основним місцем дії VІР3А(а)-протеїну. Ніякого пошкодження не було помітно у передньому кишечнику та задньому кишечнику. Епітеліальні клітини середнього кишечника контрольних личинок були тісно зв'язаними одна з одною, не виявляючи ніяких ознак пошкодження. Зрізи личинок, яких годували протягом 24 годин раціоном, що містив Vip3A(a), показували, що віддалені кінці епітеліальних циліндричних призматичних клітин роздувалися й ставали опуклими. Хоча келихоподібні клітини виявляли певні морфологічні зміни, у них не спостерігалося ознак пошкодження на цій стадії. Дегенерація епітеліальних циліндричних призматичних клітин тривала, і через 48год. після вживання раціону, що містив Vip3A(a), просвіт був заповнений рештками зруйнованих клітин. Келихоподібні клітини також виявляли ознаки пошкодження через 48 годин, але обидва типи клітин були все ще прикріплені до основної мембрани. Личинки совки іпсилон гинули через 72 години, і лущення епітеліального шару завершувалося. Якщо в осіннього похідного черв'яка спостерігалася подібна гістопатологія, метелик кукурудзяний європейський не виявляв ніякого пошкодження тканин за тих самих умов експерименту. In vivo імунолокалізація VІР3А(а)-протеїну. Личинок совки іпсилон та метелика кукурудзяного європейського на третій стадії розвитку, які живилися штучним раціоном з додаванням 100-200нг VIP3A(a) на см2, використовували для імуноцитохімічної характеризації VIP3A(a), зв'язаного з фрагментами середнього кишечника. Зв'язаний VIP3A(a) візуалізували з використанням антитіла кроля проти VIP3A(a), попередньо очищеного через сефарозу протеїну А та іммобілізовані колонки Е. coli (Yu, C.G. et al. Appl. Environ. Microbiol. 63:532-536, 1997). Зв'язування VIP3A(a) виявляли в епітелії середнього кишечника совки іпсилон, не виявляючи ніякого зв'язування з середнім кишечником метелика кукурудзяного європейського. Фрагменти середнього кишечника личинок совки іпсилон, які живилися контрольним раціоном, не виявляли зв'язування VIP3A(a). Очевидно, зв'язування VIP3A(a) специфічно пов'язане з апікальними мікроворсинками і найбільш пов'язане з циліндричними призматичними клітинами, без жодних ознак сигналу у келихоподібних клітинах. Приклад 9: VІР3А(а) та VІР3А(b) викликають апоптоз у клітинах комах VIP3A(a) та VIP3A(b) виявилися викликаючими апоптоз протеїнами, що виникають в результаті характеризації інсектицидної дії, спрямованої на лінії клітин комах (Sf-9), що походять від Spodaptera frugiperda, комахи, сприйнятливої до VIP3A(a). VІР3А(а) виявляв інсектицидну активність проти лінії клітин комах, якщо його присутність забезпечували протягом усього експерименту. При тимчасовому піддаванні SF-9 клітин комах дії VIP3A(a) та VIP3A(b) життєздатність їхніх клітин значною мірою знижувалася, навіть при такому короткому періоді, як 5хв. Коли час інкубації перевищив 10 хвилин, вплив VIP3A(a) та VIP3A(b) на життєздатність клітин комах протягом періоду у 6 годин був максимальним, виявляючи 90% зниження життєздатності клітин. Цитологічні зміни, які відбуваються у SF-9 клітинах, тимчасово підданих дії VIP3A(a), відстежували шляхом мікроскопії. На поверхні оброблених клітин виникали невеличкі виступи у проміжку часу від 10 до 15 хвилин після того, як їх піддали дії VІР3А(а)-протеїну. На цій стадії мітохондрії клітин залишалися функціонально неушкодженими, як показало забарвлення родаміном 123, барвником, який накопичується у мітохондріях з активним мембранним потенціалом (Johnson, L.V. et al. Proc. Natl. Acad. Sci. USA 77:990-994, 1980). Ці виступи зрештою зникали, і клітини вступали у фазу активної вакуолізації, яка тривала ще від 30 до 60хв. На останніх стадіях клітин комах перед розпадом бубнявіють. Для окремої клітини весь процес вимагає від 1 до 2 годин. Усі ці клітинні події відповідають даним попередніх досліджень клітин, які зазнають апоптозу, особливо якщо брати до уваги, що запрограмоване відмирання клітин під час перетворення певних комах супроводжується клітинною вакуолізацією та бубнявінням (Schwartz, L.M. et al. Proc. Natl. Acad. Sci. USA 90:980-984 (1993)). Останні дослідження показали, що розподіл фосфоліпідів у плазматичній мембрані піддається впливу на перших стадіях тваринних клітин, які зазнають апоптозу (Martin, S. J., et al. J. Bxp. Med.:i82, 1545-1556, 1995), зокрема, втілення фосфатидилсерину (PS). Цей процес можуть бути візуалізований через використання Аnnехіn V, антикоагулюючого протеїну з високою спорідненістю до фосфатидилсерину (PS). Коли оброблені VIP3A(a) SF-9 клітини інкубували з Аnnехіn V, втілення PS виявляло себе у мембранах клітин комах вже через 5-10 хвилин після піддання дії VIP3A(a), що, можливо, позначало початок апоптозу. Одною з основних молекулярних подій, що є критерієм апоптозу, є ендонуклеоліз, який в результаті веде до розриву дволанцюгової ДНК, звільняючи олігнуклеосомного розміру фрагменти, що мають 200 пар нуклеотидів та у кілька разів більше. Ми досліджували випадки ендонуклеолізу в SF-9 клітинах, оброблених VIP3A(a), застосовуючи спосіб in situ виявлення та аналізуючи ДНК шляхом електрофорезу в агарозному гелі. На основі здатності ферменту Кленова включати модифіковані нуклеотиди з застосуванням кінців ДНК, отриманих шляхом фрагментації ДНК, SF-9 клітини комах виявляли ознаки ендонуклеолізу вже через 30 хвилин після підцання їх дії VІР3А(а)-протеїну. Цей етап збігається з появою зв'язаних з мембраною субклітинних апоптотичних організмів, візуалізованих при мікроскопічному спостереженні. Ці ранні ознаки ендонуклеотичної активності підтверджувалися виявленням фрагментів ДНК в агарозних гелях, які характеризуються хроматиновими сходами на дещо пізніших стадіях процесу. Ці результати підтверджували ознаки, виявлені з цитологічних спостережень, згідно якими SF-9 клітини викликають запрограмоване відмирання клітин апоптотичного типу після їх піддання дії VІР3А(а)-протеїну. VIP3A(a) та VIP3A(b) протеїни були виявлені на основі їхніх інсектицидних властивостей проти деяких комах роду Lepidoptera. Таким чином, нам потрібно було знати, чи викликає VІР3ЗА(а)-протеїн апоптотичний шлях у клітинах кишечника сприйнятливих комах після його вживання ними і чи виявляє він, таким чином, свої інсектицидні властивості при ініціюванні активного процесу відмирання клітин. Гістологічні та гістохімічні досліди показали, що VIP3A(a)-протеїн є специфічно спрямованим на циліндричні призматичні клітини епітелію середнього кишечника сприйнятливих комах, викликаючи зміни у клітинах, які характеризуються виступами на мембрані та активною вакуолізацією, що призводить до відмирання клітин. Ці цитологічні зміни, викликані VIP3A(a) у клітинах кишечника комах, схожі на описані вище для SF-9 клітин. Отже, ми намагалися перевірити, чи зазнають клітини епітелію середнього кишечника сприйнятливих комах ендонкуклеолізу після вживання раціону, що містить VIP3A(a), шляхом in situ виявлення (Cuvillier, О., et al. Nature 381:800-803 (1996)) фрагментації ДНК. Коли фрагменти тканини середнього кишечника личинок совки іпсилон підживлювали раціоном, що містить VIP3A(a), або контрольним раціоном, забарвлення ядер, яке свідчило про фрагментацію ДНК, виявляло себе лише у циліндричних призматичних клітинах епітелію середнього кишечника, підданого дії VІР3А(а)-протеїну. Цей результат свідчить про те, що VІР3А(а)-протеїн викликає процес ендонуклеолізу в епітеліальних клітинах середнього кишечника одночасно з цитологічними змінами, про які повідомлялося раніше. Ми дійшли висновку, що VІР3А(а)-протеїн так само виявляє свої інсектицидні властивості шляхом активізації апоптотичного типу запрограмованого відмирання клітин епітелію середнього кишечника сприйнятливих комах. Приклад 10: Виділення рецептора для VІР3А(а) совки іпсилон Совка іпсилон є чутливою до VIP3A(a) і, таким чином, цю комаху використовували для виділення VIP3A(a) рецептора. Середній кишечник личинок совки іпсилон на третій стадії розвитку вибирали шляхом розтину і відразу заморожували у рідкому азоті. Один грам тканини середнього кишечника використовували для виділення мРНК згідно з протоколом, описаним для комплекту побудови двогібридної бібліотеки кДНК від фірми Clontech (1997). За початковий матеріал брали десять мікрограмів полі А + РНК. При синтезуванні першого ланцюга і випадкові, і зціплювальні оліго(dТ)25d(А/С/G)-праймери використовують в окремому синтезі з MML зворотною транскриптазою. Другий ланцюг кДНК отримували шляхом оптимального співвідношення активності ДНК-полімерази та активності РНКази Н у ферментному коктейлі другого ланцюга. Щойно синтезовані кДНК з подвійним ланцюгом після цього лігують з EcoRI-NotI-SalI адапторами. КДНК лігували у pGADIO (Vijaychander, S. et al. CLONTECHniques IX-3:8-10, 1994), що забезпечує домен активізації. Vip3А(а) ген вводили шляхом інженерії у сайт полілінкера плазміди pGBT9 у рамці з GAI-4-ДНК зв'язувальним доменом (Bartel, P.L. et al. Cellular Interactions in Development: Practical Approach, pp. 153-179, Oxford University Press, 1993). Рекомбінантний pGBT9-vip3A(a) трансформували у штам дріжджів GGY1::171 (Gill, G. and Ptashne, M. Cell51:121-126, 1987) шляхом електропорації (Estruch, J.J. et al. BioTechniques 16:610-612, 1994). Трансформовані дріжджі відбирали у мінімальному середовищі без триптофану (Bartel, P.L. et af. Cellular Interactions in Development: Practical Approach, pp. 153-179, Oxford University Press, 1993). Експресію VІР3А(а)-протеїну у рекомбінантних дріжджах підтверджували вестерн-блотингом. Штам дріжджів GGY1::171- VIP3A(a) трансформували за допомогою бібліотеки кДНК совки іпсилон, представленої у pGADIO. GGY1::171 має HIS3 маркер під контролем GAL4 сайтів розпізнавання. HISS ген дозволяє здійснювати відбір позитивного росту для клонів, трансформованих двома взаємодіючими гібридними послідовностями. Після посіву понад 200000 рекомбінантних клонів лише один виявився здатним до розвитку у мінімальному середовищі без гістидину. Плазмідну ДНК позитивної колонії дріжджів виділяли буферним способом лізису дріжджів (Kaiser, P. and Auer, B. BioTechniques 14:552 (1993)) і піддавали електропорації в Е. соlі. Вставку, що містить кДНК, субклонували у EcoRl сайт pBluescript (Stratagene) і секвенували способом дидеокситермінації (Sanger et al., Proc. Natl. Acad. Sci. USA, 74: 5463-5467 (1977)), застосовуючи комплекти для секвенування PRISM Ready Reaction Dye Deoxy Terminator Cycle Sequencing Kits та PRISM SequenaseÒ Terminator Double-Stranded DNA Sequencing Kit і аналізували на автоматичному секвенаторі АВІ 373. Альтернативний спосіб ідентифікації клонів, що кодують протеїн(и), які взаємодіють з Vip3A, складається з посіву дріжджів, трансформованих бібліотекою «ДНК совки іпсилон, присутньою у pGADIO. Після перенесення вищезгаданої популяції трансформованих дріжджів на нітроцелюлозні фільтри їх піддавали скринінгові з застосуванням міченого біотином Vір3А-протеїну, після чого здійснювали інкубацію з розчином, що містив стрептавідин разом з лужною фосфатазою (АР). Після активного промивання клон або клони, що експресують протеїн зі здатністю зв'язуватися з Vip3A, візуалізували з використанням субстрату - 5-бром-4-хлор-3-індоїлфосфату (ВСІР) у поєднанні з нітротетразолом синім (NBT). АР каталізує утворення нерозчинних осадів, які розвиваються у вигляді темних плям на нітроцелюлозних фільтрах. Експериментальні процедури детально описано у роботі Sambrook. J, et al. in Molecular Cloning; Laboratory Manual, pp. 12-21-12.24, Cold Spring Harbor Laboratory, 1989. Після повторного вирощування планшетів з дріжджами протягом доби проявлені місця порівнювали з колоніями дріжджів у первісних планшетах. Ці колонії вирощували, їхні плазміди виділяли, а їхні вставки характеризували, як описано вище. Приклад 11: Клітини комах, трансформовані геном для рецептора, виявляють апоптоз під дією VІР3А(а)-протеїну Рецептор клітин середнього кишечника совки іпсилон для VIP3A(a)-протеїну клонували у XhoI-BamHLcavn вектора косміди Smart 2 (Speek, M. et al. Gene 64: 173-177 (1988)) і рекомбінантну послідовність використовували для трансформації лінії клітин Schneider 2 (S2) виду Drosophila з застосуванням способу копреципітації фосфату кальцію (Clem, R.J. and Miller. L.K. МоI. Cel. Biol. 14: 5212-5222 (1994)). Smart 2 несе селектований маркер tet (тетрациклін) для бактеріальної трансформації та пео (неоміцин) для трансформації клітин виду Drosophila. Селектований маркером пео експресується під контролем hsp70 промотора виду Drosophila. Трансформовані S2 клітини відбирали в S2 середовищі виду Drosophila з 10% сероальбуміну плоду з G418 (1мг/мл) при 30°С (див. каталог GIBCO за 1997p.). Через 45 днів відбору у вищеописаному середовищі утворювалися кілька стабільно трансформованих S2 ліній клітин. Чутливість трансформованих S2 клітин до VIP3A(a) випробували шляхом додавання VІР3А(а)-протеїну (в остаточній концентрації 1,7мкг на мл) до середовища, що містить трансформовані S2 клітини і було попередньо піддане збовтуванню при 42°С протягом 30 хвилин. Викликання апоптозу у трансформованих S2 клітинах підтверджувалося і мікроскопічними спостереженнями, і через використання комплекту TAGS, а також комплекту для виявлення апоптозу in sftu Apoptosis Detection (детальний опис див. у каталозі Trevigen catalogue за 1996p.). Приклад 12: Виділення гомологів рецептора з інших комах Клітини епітелію середнього кишечника личинок совки іпсилон мають рецептор, який розпізнається VІР3А(а)-протеїном. Рецептори інших комах, сприйнятливих до VIP3A(a), виділяють шляхом ідентифікації послідовностей ДНК за допомогою саузерн-блотингу. ДНК підготовляють, фермент рестриктують, пропускають через агарозні гелі і піддають блотингові на нітроцелюлозних та/або нейлонових фільтрах. Ці фільтри зондують за допомогою кДНК, що кодує рецептор совки іпсилон, у не дуже суворих умовах гібридизації та промивання. Ідентифікували гени з нижчою за 50% подібністю до рецептора совки іпсилон до VIP3A(a). Саузерн-блотинг також міг бути зондований на часткових послідовностях кДНК, які кодують такі специфічні домени, як домен смерті або EGF-подібні мотиви, з метою виділення генів, що містять подібні домени, навіть якщо вони є функціонально відмінними від рецептора совки іпсилон до VIP3A(a). Виділення гомологів до рецептор совки іпсилон до VІР3A(a) здійснюють за допомогою двох гібридних систем, описаних у роботі Fields, S. and Song. O.-К. Nature 340:245-246 (1989). Виділену мРНК одержують з потрібних організмів, синтезують кДНК і клонують їх у pGADIO або рівнозначні плазміди. Бібліотеку кДНК котрансформують за допомогою pGB9, що несе vip3A(a)-гeн (або гомологи цього гена) і звільняють передбачувані рецептори у дріжджах шляхом активізації маркера на основі взаємодії протеїн-протеїн між VІР3А(а)-протеїном (або гомологами) та передбачуваним рецептором. Гомологи рецептора совки іпсилон до VIP3A(a) виділяють шляхом експресії бібліотек кДНК, виділених з потрібних організмів, клонують у відповідні вектори експресії й трансформують у клітини-хазяї, такі як дріжджі або клітини комах, які не здатні зв'язуватися та/або бути чутливими до VIP3A(a). Трансформовані клітини піддають скринінгові на основі їхньої набутої властивості зв'язувати VIP3A(a) або зазнавати апоптотичної реакції при інкубації з VIP3A(a). У цьому разі протеїн VIP3A(a) використовують як зонд, і його зв'язування відстежують або за допомогою антитіл проти VIP3A(a) або міток, таких як біотин, прикріплений до VIP3A(a). Приклад 13: Скринінг нових сполук, які викликають апоптоз у клітинах комах Модельні лінії клітин для різних рядів комах (деякі приклади включають Sf-9 клітини для ряду Lepidoptera, колорадського картопляного жука ряду Coleoptera, S2 виду Drosophila ряду Diptera) використовують для скринінгу нових сполук, дія яких полягає у викликанні апоптозу. Клітини вирощують у багатолункових планшетах, які використовують для скринінгу з високою пропускною здатністю для тисяч сполук (і великої, і малої молекулярної маси). Сполуку(и) додають як єдиний компонент або як суміші. Сполука(и), що викликають апоптоз, ідентифікують так: 1) у мембрані клітин спостерігаються мембранні виступи, 2) при використанні специфічних протеїнів з високою спорідненістю з фосфатидилсерином, таким як Annexin-V, зв'язаний з видимим маркером, виявляється реорганізація фосфатидилсерину, що містить ліпіди мембрани, 3) у цитоплазмі клітин спостерігаються згустки цитоплазми, 4) активні мітохондрії можуть бути візуалізовані шляхом використання вітальних барвників, таких як родамін 123, які накопичуються у мітохондріях, 5) фрагментація ДНК виявляється або шляхом аналізу ДНК в агарозних гелях, шляхом ELISAаналізу вивільнення нуклеосом або шляхом виявлення in vivo одноланцюгового розриву ДНК. Усі ці цитологічні та молекулярні особливості є ознаками апоптозу. Рецептор совки іпсилон до VIP3A(a) трансформують у лінії S2 клітин. Таким чином, ізогенні S2 лінії можна одержати з вищезгаданим рецептором і , без нього. Ці лінії клітин використовують для скринінгу сполук, які забезпечують диференційну реакцію через присутність вищезгаданого рецептора. Трансформовані S2 клітини, що зазнають апоптозу під дією певних сполук, ідентифікують так, як зазначено вище. Різниця реакції трансформованих та нетрансформованих клітин свідчить про те, що дія сполуки опосередковується клонованим рецептором. Подібні підходи застосовують до клітин комах, трансформованих рецепторами, гомологічними рецепторові совки іпсилон до VIP3A(a). Усі публікації та патентні заявки, згадані у цьому описі, вказують на рівень підготовки спеціалістів, яких стосується даний винахід. На усі публікації та патентні заявки авторами робиться посилання у тій же мірі, як це було б, якби кожна окрема публікація або патентна заявка була конкретно й окремо вказана як включена шляхом посилання. Хоча даний винахід був описаний з деякими деталями, що використовувалися як ілюстрація та приклад для кращого розуміння, у межах формули, яка додається, можуть передбачатися певні зміни та модифікації. ПЕРЕЛІК ПОСЛІДОВНОСТЕЙ (1) ЗАГАЛЬНА ІНФОРМАЦІЯ: (1) ЗАГАЛЬНА ІНФОРМАЦІЯ: (і) ПОДАВЕЦЬ: (A) НАЗВА: NOVARTIS AG (Б) ВУЛИЦЯ: Schwarzwaldallee 215 (B) МІСТО: Базель (Д) КРАЇНА: Швейцарія (Е) ПОШТОВИЙ ІНДЕКС (ZIP): 4058 (Е) ТЕЛЕФОН: +4161 324 11 11 (Ж) ТЕЛЕФАКС: +4161 322 75 32 ....(іі) НАЗВА ВИНАХОДУ: Новий клас протеїнів для боротьби зі шкідниками рослин (ііі) КІЛЬКІСТЬ ПОСЛІДОВНОСТЕЙ: 20 (iv) ФОРМА КОМП'ЮТЕРНОГО ЗЧИТУВАННЯ: (A) ТИП НОСІЯ: дискета (Б) КОМП'ЮТЕР: IBM PC (B) ОПЕРАЦІЙНА СИСТЕМА: PC-DOS/MS-DOS (Г) ПРОГРАМА: Patentin Release #1.0, Version #1.30 (2) ІНФОРМАЦІЯ ДЛЯ SEQ ID NО:1: (і) ХАРАКТЕРИСТИКИ ПОСЛІДОВНОСТІ: (A) ДОВЖИНА: 2378 пар нуклеотидів (Б) ТИП: нуклеїнова кислота (B) ЛАНЦЮГОВІСТЬ: одинична (Г) ТОПОЛОГІЯ: лінійна (іі) ТИП МОЛЕКУЛИ: ДНК (геномна) (ііі) ГІПОТЕТИЧНІ ВАРІАНТИ: НЕМАЄ (іх) ОСОБЛИВОСТІ: (А) НАЗВА/КОД: CDS (Б) РОЗТАШУВАННЯ: 9..2375 (Г) ІНША ІНФОРМАЦІЯ: /ознака= «Природна послідовність ДНК, що кодує VІР3А(а)-протеїн з АВ88, що міститься у рСІВ7104» (хі) ОПИС ПОСЛІДОВНОСТІ: SEQ ID NО:1: (2) ІНФОРМАЦІЯ ДЛЯ SEQ ID NО:2: (і) ХАРАКТЕРИСТИКИ ПОСЛІДОВНОСТІ: (А) ДОВЖИНА: 789 амінокислот (Б) ТИП: амінокислота (Г) ТОПОЛОГІЯ: лінійна (іі) ТИП МОЛЕКУЛИ: протеїн (хі) ОПИС ПОСЛІДОВНОСТІ: SEQ ID NО:2:
ДивитисяДодаткова інформація
Назва патенту англійськоюA method for pest control of the plants
Назва патенту російськоюСпособ борьбы с вредителями растений
МПК / Мітки
МПК: C12R 1/07, C07K 14/415, C12N 15/62, C12N 15/82, C07K 14/32, A01P 7/04, C12N 15/32, C12N 15/31, A01N 63/00, A01H 5/00, C07K 16/18, G01N 33/53, C07H 21/04, C07K 14/325, C07K 14/435
Мітки: гомологів, спосіб, ідентифікації, послідовність, комахами, рецептора, вегетативного, vip3-класу, днк, протеїну, рецептор, боротьби, сполуки, антитіло, інсектицидного
Код посилання
<a href="https://ua.patents.su/53-68345-receptor-do-vegetativnogo-insekticidnogo-protenu-vip3-klasu-poslidovnist-dnk-antitilo-do-receptora-sposib-identifikaci-gomologiv-receptora-sposib-identifikaci-spoluki-sposib-borotb.html" target="_blank" rel="follow" title="База патентів України">Рецептор до вегетативного інсектицидного протеїну vip3-класу, послідовність днк, антитіло до рецептора, спосіб ідентифікації гомологів рецептора, спосіб ідентифікації сполуки, спосіб боротьби з комахами</a>
Похідні арилпіролу, що мають інсектицидну, нематоцидну і акарицидну активність, спосіб їх одержання, проміжні сполуки для їх одержання і спосіб боротьби з комахами, нематодами і кліщами

Номер патенту: 41241
Опубліковано: 17.09.2001
Автори: Діл Роберт Юджін, РАЙТ Дональд Перрі, мол., Сідденс Джек Кеннет, БРАУН Дейл Гордон
МПК: A01P 7/04, A01N 43/36, A01P 7/02, C07D 207/34, A01P 5/00, C07D 207/42
Мітки: похідні, проміжні, сполуки, інсектицидну, акарицидну, спосіб, одержання, нематодами, кліщами, боротьби, арилпіролу, активність, нематоцидну, мають, комахами
Формула / Реферат:
1. Производные арилпиррола общей формулы (I)где Х - хлор, бром, йод или СF3;Y - хлор, бром, йод, СF3 или CN;W - CN или NO2;А - водород, С1-С4-алкил, незамещенный или замещенный галогеном, гидрокси-, С1-С4-алкокси-, С1-С4-алкилтио или фенильной группой, фенокси- или С1-С2-алкоксифенильной группой, С1-С4-карбалкоксиметил, С3-С4-алкенил, незамещенный или замещенный одним галогеном, цианогруппа, С3-С4-алкинил,...
Спосіб приваблювання та боротьби з комахами

Номер патенту: 48176
Опубліковано: 15.08.2002
Автори: Девідсон Томас Чарльз, Вернер Георгіна М.
МПК: A01N 43/56, A01N 47/02
Мітки: комахами, спосіб, боротьби, приваблювання
Формула / Реферат:
1. Спосіб приваблювання та боротьби з комахами, який включає пропонування згаданим комахам для ковтання ефективної кількості сполуки формули:в якій:R1 - є CN або метилом;R2 – є S(O)nR3;R3 - є алкілом або галогеналкілом;R4 - вибраний із групи, що включає атом водню, атом галогену та радикал, який може бути -NR5R6, -S(O)mR7,...
Інсектицидна композиція та спосіб боротьби з комахами

Номер патенту: 56140
Опубліковано: 15.05.2003
Автори: Гаутіер Мартін, Дероіс Жан
МПК: A01N 47/02, A01N 43/56
Мітки: інсектицидна, спосіб, боротьби, композиція, комахами
Формула / Реферат:
1. Інсектицидна композиція, що містить:а) від 0,001 до 5%, краще - від 0,05 до 0,5% сполуки формули (I)(I),у якій :- R1 означає атом галогену, групу CN або метил;- R2 означає S(O)nR3;- R3 означає алкіл або галогеналкіл;- R4 означає атом водню або галогену; або радикал NR5R6, S(O)mR7, C(O)R7 або C(O)O-R7, алкіл, галогеналкіл...
Спосіб інгібування тирозинкінази рецептора епідермального фактора росту (варіанти), біциклічні сполуки піримідину, фармацевтична композиція, яка має активність, що інгібує тирозинкіназу рецептора епідермального

Номер патенту: 62028
Опубліковано: 15.12.2003
Автори: Денні Уілліам Александер, Крейкер Алан, Бріджес Александер Джеймс, Фрай Девід, Томпсон Ендрю Марк, Рукастл Гордон Уілліам, Майєр Роберт
МПК: C07D 495/04, A61P 13/02, A61K 31/495, C07D 473/34, A61P 1/00, A61K 31/505, A61P 15/00, A61P 43/00, A61P 29/00, C07D 487/04, C07D 471/04, C07D 513/04, A61P 35/00, C07D 498/04, C07D 491/048, A61P 17/00, C07D 471/14
Мітки: фактора, яка, має, сполуки, інгібування, тирозинкінази, інгібує, тирозинкіназу, спосіб, фармацевтична, активність, епідермального, варіанти, росту, біциклічні, композиція, рецептора, піримідину
Формула / Реферат:
1. Способ ингибирования тирозинкиназы рецептора эпидермального фактора роста при лечении млекопитающего путем введения млекопитающему бициклического производного, отличающийся тем, что в качестве бициклического производного используют бициклическое производное пиримидина общей формулы (I), (I)где1 - 2 из радикалов А - Е означают азот, а остальные...
Похідні 1-арилпіролів, що мають інсектоакарицидонематоцидну активність, інсектоакарицидонематоцидна композиція та спосіб боротьби із комахами

Номер патенту: 41248
Опубліковано: 17.09.2001
Автори: Філіп Рейд Тіммонс, Жан-П'єр А. Вор, Пол А. Кейн, Рассел Джеймс Аутколт, Лі С. Холл, Девід Ніл Сінодіс, Патрісіа Л. Квятковські, Енібел Лоупс
МПК: A01N 47/02, C07D 295/145, C07D 207/36, A01N 43/36, C07C 255/30, C07D 207/34
Мітки: інсектоакарицидонематоцидну, похідні, боротьби, мають, композиція, інсектоакарицидонематоцидна, активність, спосіб, 1-арилпіролів, комахами
Формула / Реферат:
1. Производные 1 - арилпирролов общей формулы (I):где для соединений (Іа)Х - группа R2S(O)n, где n = 0,1 или 2, R2 - метил или (С1 С3)галогеналкил, линейный или разветвленный, в котором галоген означает атом фтора, хлора или брома или их сочетание,R1 - водород, алкил, алкилтио, цианоалкил с 1-3 атомами углерода,R3 - хлор, алкил или фторалкил с 1-3 атомами углерода,Y - трифторметил, при условии,...
Попередній патент: Порошковий інгалятор (варіанти)
Наступний патент: Тренажер для навчання і контролю оптимальної посадки водія
Випадковий патент: Спосіб борування алюмінію та його сплавів