Композиція пухлиноасоційованих пептидів для лікування ракових захворювань
Номер патенту: 109891
Опубліковано: 26.10.2015
Автори: Вейншенк Тоні, Фрітше Йенс, Вальтер Штеффен, Зінгх Харпреет, Левандровскі Петер







Формула / Реферат
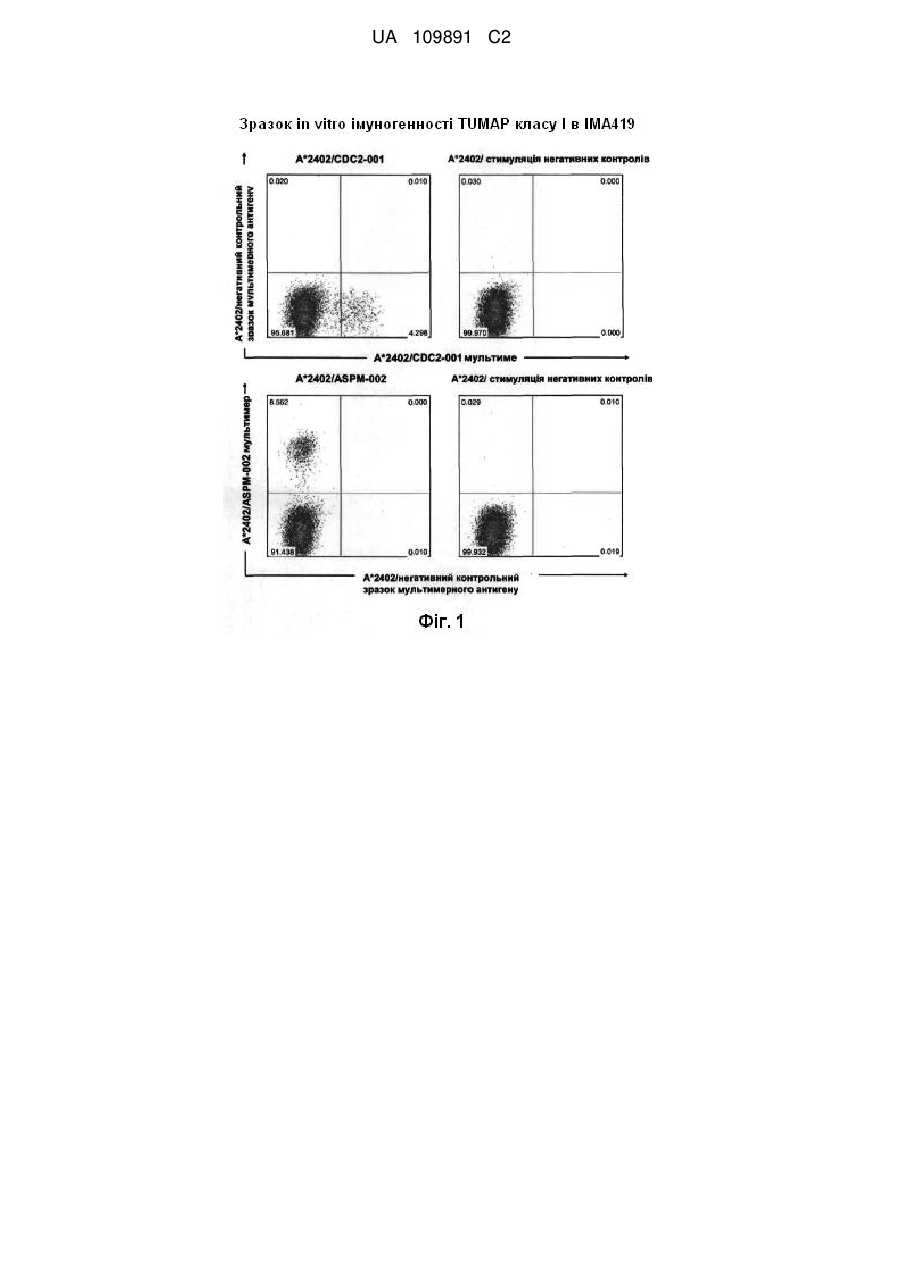
1. Фармацевтична композиція, яка включає принаймні два пептиди, де зазначені принаймні два пептиди являють собою HLA-A*024 ліганди, які складаються з амінокислотної послідовності відповідно до SEQ ID NO: 1 та SEQ ID NO: 2, та де зазначені пептиди представлені у формі фармацевтично прийнятної солі.
2. Фармацевтична композиція за п. 1, у якій принаймні один пептид включає непептидні зв'язки.
3. Фармацевтична композиція за п. 1, у якій вибір, число та/або кількість пептидів, присутніх у композиції, є специфічними для тканин, ракових захворювань та пацієнтів.
4. Фармацевтична композиція за п. 1, яка додатково включає принаймні один прийнятний ад'ювант.
5. Фармацевтична композиція за п. 4, у якій ад'ювант являє собою кoлонієстимулювальний фактор.
6. Фармацевтична композиція за п. 5, у якій кoлoнієстимулювальний фактор вибраний з групи, яка включає гранулоцитарно-макрофагальний колонієстимулювальний фактор (GM-CSF), іміквімод, та резиквімод.
7. Фармацевтична композиція за п. 1, яка додатково включає принаймні одну антигенпрезентуючу клітину.
8. Фармацевтична композиція за п. 7, у якій антигенпрезентуюча клітина являє собою дендритну клітину.
9. Фармацевтична композиція за п. 7, у якій принаймні одна антигенпрезентуюча клітина є
a) клітиною, в яку імпульсним методом введений чи в яку завантажений пептид, або
b) яка вміщує експресійну конструкцію, що кодує пептид.
10. Фармацевтична композиція за п. 1, де фармацевтична композиція призначена для введення методом внутрішньовенного, внутрішньоартеріального, внутрішньочеревного, внутрішньом'язового, внутрішньошкірного, внутрішньопухлинного, перорального, дермального, назального, букального, ректального, вагінального введення, введення шляхом інгаляцій або топічного застосування.
Текст
Реферат: Винахід стосується імунотерапевтичних пептидів та їхнього застосування в імунотерапії, зокрема в імунотерапії раку, зокрема, композиції пептидів для викликання протипухлинних імунних відповідей протиракових захворювань. UA 109891 C2 (12) UA 109891 C2 UA 109891 C2 5 10 15 20 25 30 35 40 45 50 55 60 ОПИС Цей винахід відноситься до імунотерапевтичних пептидів та їхнього використання в імунотерапії, зокрема, в імунотерапії раку. В цьому винаході розкриваються епітопи пухлиноасоційованих пептидів Т-клітин-хелперів, окремо або в поєднанні з іншими пухлиноасоційованими пептидами, котрі служать активними фармацевтичними інгредієнтами композицій вакцин, стимулюючих протипухлинні імунні реакції. Зокрема, композиція пептидів згідно цього винаходу може використовуватись в композиціях вакцин для викликання протипухлинних імунних відповідей проти раку шлунка та інших видів раку (РШ). Для цілей даного винаходу уся довідкова література:, наведена в ньому, включена шляхом посилань в усій повноті. Рівень техніки Рак шлунка є захворюванням, при якому в слизовій оболонці шлунка формуються злоякісні клітини. Пухлина шлунка або рак шлунка може розвиватися в будь-якій частині шлунка та поширюватися на весь шлунок і на інші органи; зокрема, стравохід, легені та печінку. Рак шлунка займає четверте місце серед найбільш розповсюджених видів раку у світі; у 2002 році було діагностовано 930 000 випадків даного виду раку. Це захворювання має високий коефіцієнт смертності (приблизно 800 000 смертей на рік), що виводить його на друге місце серед найбільш поширених причин смерті від раку в усьому світі, після раку легенів. Рак шлунка більш розповсюджений серед чоловіків та частіше зустрічається у країнах Азії та у країнах, що розвиваються. (Інформацію можна одержати у ВООЗ.) Щорічно він становить близько 2 % (25 500 випадків) усіх нових випадків раку у Сполучених Штатах, але є більш розповсюдженим в інших країнах. Це основний вид раку у Кореї (20,8 % злоякісних новоутворень). В Японії рак шлунка залишається найбільш розповсюдженим раковим захворюванням для чоловіків. Кожного року у Сполучених Штатах приблизно 13 000 чоловіків та 8000 жінок ставиться діагноз пухлина шлунка. Більшість з цих людей знаходяться у віці більше 70 років. Рак шлунка займає четверте місце серед найбільш розповсюджених видів раку у світі, після раку легенів, молочної залози, колоректального раку та раку прямої кишки. Крім того, пухлина шлунка залишається другою найбільш поширеною причиною смерті від раку. За даними Американського товариства по боротьбі з раком, у 2007 році оцінювана кількість нових випадків становила один мільйон, приблизно 70 % з них було виявлено у країнах, що розвиваються, та близько 800 000 смертельних випадків (див. публікації Американського товариства по боротьбі з раком). У розповсюдженості цього захворювання у світовому масштабі існує дуже значна географічна мінливість. Частота захворювань є найвищою в Азії та у деяких регіонах Південної Америки і найнижчою в Північній Америці. Найбільш висока смертність зафіксована у Чилі, Японії, Південній Америці та в колишньому Радянському Союзі. Рак шлунка часто діагностується на пізній стадії, оскільки в більшості країн світу, за винятком Японії (та в обмеженому режимі у Кореї), не ведеться скринінг, що часто дозволяє виявити хворобу на ранньому етапі. Таким чином, це й досі становить серйозну проблему для медичних працівників. Фактори ризику захворювання на рак шлунка - це інфікування Helicobacter pylori (H. pylori), паління, високий вміст солі у раціоні та інші фактори харчування. Дуже невеликий відсоток випадків раку шлунка (1 % - 3 %) асоціюється з синдромами спадкової схильності до раку шлунка. Мутації E-кадхеріну відбуваються приблизно у 25 % сімей з аутосомно-домінантним типом схильності до раку шлунка дифузного типу. Цей підвид раку шлунка був названий успадкованим дифузним раком шлунка. Може бути корисним надання генетичної консультації та розгляд можливості профілактичної гастректомії у молодих асимптоматичних носіїв усіченої гену зародкової лінії. Стінка шлунка складається з 3 шарів тканини: слизовий (внутрішній) шар, м'язовий (середній) шар та серозний (зовнішній) шар. Рак шлунка починається у клітинах, що вистилають слизовий шар, та розповсюджується на зовнішні шари, по мірі зростання пухлини. Використовується стандартне лікування чотирьох типів. Лікування раку шлунка може включати хірургічне втручання, хіміотерапію, променеву терапію або хіміо-променеве лікування. Хірургічна операція є основним методом лікування раку шлунка. Мета хірургічного втручання полягає у виконанні повної резекції з відсутністю пухлинних клітин на краї зрізу (R0-резекція). Однак, приблизно 50 % пацієнтів з місцево-розповсюдженим раком шлунка не можуть проходити R0-резекцію. R1 вказує на мікроскопічний резидуальний рак (край зрізу з наявністю пухлинних клітин); R2 означає значний (макроскопічний) резидуальний рак, але не метастатичну пухлину. Результат лікування пацієнта залежить від стадії раку на момент постановки діагнозу. (Клінічні рекомендації з онкології Національної онкологічної мережі США 1 UA 109891 C2 5 10 15 20 25 30 35 40 45 50 55 60 NCCN Clinical Practice Guidelines in Oncology™). 5-річна виживаність після радикального оперативного втручання становить 30-50 % для пацієнтів зі стадією II та 10-25 % для пацієнтів зі стадією III захворювання. У цих пацієнтів висока ймовірність місцевих і системних рецидивів. Метастази розвиваються у 80-90 % осіб з пухлиною шлунка, причому виживаність протягом шести місяців спостерігається у 65 % хворих, яким був поставлений діагноз на ранніх стадіях, та менш ніж 15 % у випадках, коли рак був діагностований на пізніх стадіях. Отже, залишається потреба у новому ефективному та безпечному варіанті лікування раку шлунка, раку передміхурової залози, раку ротової порожнини, плоскоклітинної карциноми ротової порожнини (OSCC), гострої мієлоїдної лейкемії (AML) (Qian і співавт., 2009), лімфоми MALT-типу, спричиненої мікроорганізмами H. Pylori (Banerjee і співавт., 2000), раку товстої кишки/колоректального раку, гліобластоми, немілкоклітинного раку легенів (NSCLC), карциноми шийки матки, раку молочної залози у людей, раку передміхурової залози, раку товстої кишки, раку підшлункової залози, протокової аденокарциноми підшлункової залози, раку яєчників, гепато-клітинної карциноми, раку печінки, пухлин головного мозку різних фенотипів, лейкемії, такої як гостра лімфобластна лейкемія (ALL), раку легенів, саркоми Юїнга, раку ендометрію, плоскоклітинної карциноми голови та шиї, епітеліального раку гортані, раку стравоходу, карциноми ротової порожнини, раку сечового міхура, раку яєчників, нирково-клітинної карциноми, атипової менінгіоми, папілярного раку щитовидної залози, пухлин головного мозку, карциноми протоків слинних залоз, раку шийки матки, екстранодальних T/NK-клітинних лімфом, неходжкінської лімфоми, злоякісних солідних пухлин легенів і молочної залози та інших пухлин. Також залишається необхідність у варіантах лікування, що покращують стан пацієнтів без використання хіміотерапевтичних речовин або інших препаратів, котрі можуть призвести до серйозних побічних ефектів. Колоректальна карцинома Згідно із даними Американського товариства по боротьбі з раком, колоректальний рак (CRC) є третім з найбільш розповсюджених видів раку в США, на який захворюють більш ніж 175 000 нових пацієнтів кожного року. В США, Японії, Франції, Німеччині, Італії, Іспанії та Великобританії він уражає більш ніж 480 000 людей. Колоректальний рак є однією з найчастіших причин смертності від раку в розвинених країнах. Дослідження показують ймовірність виникнення колоректального раку як результату взаємодії між успадкованими факторами та впливом зовнішнього середовища. В більшості випадків попередниками колоректальних пухлин вважаються аденоматозні поліпи; однак переродження може тривати багато років. Основним фактором ризику колоректального раку є вік, оскільки 90 % випадків захворювання діагностуються у людей старше 50 років. Інші фактори ризику колоректального раку, за даними Американського товариства по боротьбі з раком, включають вживання алкоголю, раціон харчування з великим вмістом жирів та/або червоного м'яса і недостатнім вживанням фруктів та овочів. Захворюваність продовжує зростати, зокрема, в таких регіонах, як Японія, де це може бути обумовлено переходом на західний стиль харчування, з надмірною кількістю жирів та м'яса в раціоні та зменшенням вживання харчових волокон. Однак частота захворювань зростає не настільки швидко, як раніше, і це може бути обумовлено більш активним скринінгом та видаленням поліпів, що запобігає переродженню поліпів на рак. Як і у відношенні більшості солідних пухлин, лікуванням першого ряду є хірургічне втручання, однак, його переваги обмежуються ранніми стадіями захворювання, в той час як у значної частини пацієнтів діагностуються пізні стадії. Стандартом лікування прогресуючого колоректального раку є режими хіміотерапії, які базуються на схемах з використанням фторурацила. Більшість таких режимів - це так звані схеми FOLFOX (інфузійне введення5FU/лейковорину плюс оксаліплатин) та FOLFIRI (іринотекан, лейковорин, болюсне і тривале інфузійне введення 5-FU). Введення цитотоксичних препаратів третього покоління, таких як іринотекан та оксаліплатин, збільшило надію на значне підвищення ефективності лікування, але прогноз все ще відносно несприятливий, і виживаність, в цілому, залишається на рівні приблизно 20 місяців при наявності метастазів, отже, незадоволені потреби стосовно лікування цієї хвороби поки що є високими. Останнім часом з'явилося нове покоління медикаментів, що діють цілеспрямовано на молекулярному рівні, таких як Авастин (Avastin) (бевацизумаб) та Ербітукс (Erbitux) (цетуксимаб), і близько 40 сполук знаходяться на завершальному етапі клінічної розробки для різних стадій колоректального раку. Комбінації декількох з цих сполук збільшують кількість потенційних варіантів терапії, які очікуються в майбутньому. Переважна більшість речовин є у фазі 2 досліджень, причому на EGFR (рецептори епідермального фактора росту) ці сполуки 2 UA 109891 C2 5 10 15 20 25 30 35 40 45 50 55 60 діють частіше, ніж будь-які інші ліки, що розроблюються для колоректального раку, і це обумовлено фактом, що приблизно у 80 % хворих на колоректальний рак експресія EGFR підвищується. Зараз проводяться клінічні дослідження пацієнтів з II стадією захворювання, з поєднанням хіміотерапії та недавно ухвалених моноклональних антитіл (mAb) (цетуксимаб + іринотекан або FOLFOX4; бевацизумаб як монотерапія або разом з FOLFOX4). Очікується, що період спостережень для одержання статистично значимих результатів цих досліджень становитиме від трьох до чотирьох років. Моноклональні антитіла (mAb), які на даний час використовуються в онкології, в цілому мають прекрасні шанси щодо їхнього використання без втручання в активну імунотерапію. Фактично були одержані доклінічні дані, які свідчать про те, що зниження експресії VEGF (фактору росту ендотелію судин) (за допомогою бевацизумабу) позитивно впливає на активацію T-клітин, опосередковану ДК (дендритними клітинами). Рак передміхурової залози та інші приклади пухлин Рак передміхурової залози є основною причиною смерті від раку у чоловіків: за наявними оцінками, в 2007 році було зареєстровано 27 050 смертельних випадків від цієї хвороби. Хоча з початку 1990-х років смертність серед білих чоловіків та афроамериканців знижувалася, показники смертності у афроамериканців залишаються більш ніж в два рази вищими, ніж у білих чоловіків. Рак передміхурової залози - це найчастіше діагностований вид раку у чоловіків. За невиясненими до цього часу причинами, захворюваність у афроамериканців значно більша, ніж у білих. Показники захворюваності на рак передміхурової залози значно змінилися за останні 20 років: із швидким збільшенням за період 1988-1992 років, різким спадом в 1992-1995 роках, та помірним ростом з 1995 року. Ці тенденції, значною мірою, відображають покращення скринінгу раку передміхурової залози з аналізом крові для визначення рівня простатоспецифічного антигену (PSA). Помірний зріст випадків захворювання за останнє десятиріччя, скоріше за все, пояснюється широко поширеним PSA-скринінгом серед чоловіків, молодших 65 років. Показники частоти захворювання на рак передміхурової залози вирівнялися у чоловіків віком 65 років і більше. Захворюваність досягла піку у білих чоловіків в 1992 році (237,6 на 100 000 осіб) та афроамериканців у 1993 році (342,8 на 100 000 осіб). Лікування раку передміхурової залози може включати пильне спостереження, хірургічне втручання, променеву терапію, терапію високоінтенсивним фокусованим ультразвуком (HIFU), хіміотерапію, кріохірургію, гормональну терапію або комбінацію деяких з цих методів. Вибраний варіант лікування залежить від стадії захворювання, показника за шкалою Глісона та рівня PSA. Інші важливі фактори включають вік чоловіка, загальний стан його здоров'я та відношення до потенційного лікування і можливих побічних ефектів. Оскільки будь-яке лікування може мати серйозні побічні ефекти, такі як еректильна дисфункція та недержання сечі, обговорення методу лікування часто зосереджується на досягненні балансу між задачами терапії та ризиками змін у стилі життя. Якщо рак поширюється за межі передміхурової залози, варіанти лікування значно змінюються, тому більшість лікарів, котрі займаються терапією раку передміхурової залози, використовують велику кількість томограм для прогнозування ймовірності поширення. Лікування пильним спостереженням, HIFU, променевою терапією, кріохірургією та хірургічним втручанням взагалі пропонуються чоловікам, у яких рак залишається в межах передміхурової залози. Гормональна терапія та хіміотерапія часто призначаються у випадку захворювання, котре поширюється за межі передміхурової залози. Однак є й винятки: променева терапія може використовуватись для деяких запущених пухлин, а гормональна терапія - для певних випадків пухлин на ранній стадії. Кріотерапія, гормональна терапія та хіміотерапія також можуть пропонуватись, якщо первісний варіант лікування не дає результату, і рак прогресує. У значної кількості пацієнтів, хворих на рак передміхурової залози, які перенесли радикальну простатектомію внаслідок клінічно очікуваного росту, обмеженого ураженим органом, заключний гістологічний аналіз хірургічної проби показує локальне екстенсивне поширення пухлини за межі органу. У цих пацієнтів відзначається високий ризик ранніх місцевих рецидивів, що, як правило, можуть бути виявлені як підвищений рівень PSA, тобто, біохімічний рецидив. Можливі методи лікування в цьому випадку включають дистанційну променеву терапію та гормон-деприваційну терапію; однак, цінність таких терапевтичних підходів, зокрема, для подовження тривалого виживання пацієнта, не може вважатися доказаним. Крім того, треба враховувати можливі ускладнення, пов'язані з лікуванням, такі як розвиток звуження уретри (променева терапія), втрата лібідо та імпотенція, ризик зменшення солей кальцію в кістках скелету, що проявляється як остеопороз, та помітно збільшений ризик патологічних переломів кісток (при гормон-деприваційній терапії). 3 UA 109891 C2 5 10 15 20 25 30 35 Більш ніж 90 % усіх випадків раку передміхурової залози виявляються на локальній та регіональній стадії; відносний показник 5-річної виживаності для пацієнтів, у яких пухлини діагностувалися на цих стадіях, досягає 100 %. Протягом останніх 25 років 5-річна виживаність для усіх стадій в сукупності збільшилася з 69 % приблизно до 90 %. Відповідно до останніх даних, відносна 10-річна виживаність становить 93 %, а 15-річна виживаність - 77 %. Значне покращення в показниках виживаності, зокрема, на рівні 5 років, можна частково віднести на рахунок раннього діагностування та удосконалення лікування. Проте виживаність значно знижується після того, як пухлина поширюється на інші тканини та органи. Рак легенів За попередніми оцінками, в 2007 році в США очікується 210 000 нових випадків захворювання на рак легенів, що нараховує приблизно 15 % діагнозів раку. Захворюваність значно знижувалася у чоловіків, з 102 випадків на 100 000 осіб в 1984 році до 78,5 в 2003 році. У жінок зараз захворюваність залишається на одному й тому ж рівні після тривалого періоду зростання. Рак легенів клінічно класифікується як мілкоклітинний (13 %) або немілкоклітинний (87 %) рак, для цілей лікування. Рак легенів є причиною більшості смертей від ракових захворювань і у чоловіків, і у жінок. За прогнозами в 2007 році очікується 160 390 смертельних випадків від цього захворювання, що становить близько 29 % усіх смертей від раку. Щорічно, починаючи з 1987 року, від раку легенів вмирало більше жінок, ніж від раку молочної залози. Протягом періоду 1991-2003 років, показники смертності у чоловіків продовжували істотно знижуватися, приблизно на 1,9 % на рік. Зріст смертності від раку легенів у жінок припинився після постійного підвищення протягом кількох десятиліть. Ці тенденції в смертності від раку легенів відображають зменшення в кількості курців за останні 30 років. Методи лікування визначаються типом (мілкоклітинний чи немілкоклітинний) та стадією раку і включають хірургічне втручання, променеву терапію, хіміотерапію та спрямовану біологічну терапію, з використанням, наприклад, бевацизумабу (Avastin®) та ерлотинібу (Tarceva®). Для локалізованого раку, хірургічне втручання є зазвичай переважним методом лікування. Недавні дослідження вказують на те, що виживаність у випадку немілкоклітинного раку легенів на ранній стадії покращується завдяки хіміотерапії після проведення операції. Оскільки захворювання, як правило, вже є поширеним на момент його виявлення, часто використовуються променева терапія та хіміотерапія, іноді в поєднанні з хірургічною операцією. Хіміотерапія, окремо чи разом з променевою терапією, є взагалі переважним методом лікування мілкоклітинного раку легенів; при такій схемі лікування у великої кількості пацієнтів спостерігається ремісія, яка є в деяких випадках довготривалою. 1-річна відносна виживаність для раку легенів трохи підвищилася з 37 % в 1975-1979 роках до 42 % в 2002 році, значною мірою завдяки удосконаленням в хірургічній техніці та комбінованій терапії. Однак 5-річний рівень виживаності для усіх стадій разом - лише 16 %. Виживаність становить 49 % для випадків, визначених, коли захворювання ще локалізоване; однак, лише 16 % діагнозів раку легенів ставиться на такій ранній стадії. 40 Таблиця 1 Прогноз нових випадків захворювання на рак та смертельних випадків для обох статей для США в 2007 р. (Американське товариство по боротьбі з раком. Факти і цифри про рак - 2007 р. Атланта: American Cancer Society; 2007.) Локалізація Прогноз нових випадків захворювання Обидві Чоловіки Жінки статі 20 500 11 170 9 330 180 510 2 030 178 480 218 890 218 890 15 560 12 130 3 430 112 340 55 290 57 050 51 190 31 590 19 600 37 170 18 830 18 340 Прогноз смертельних випадків Обидві статі 12 740 40 910 27 050 13 940 52 180 12 890 33 370 Чоловіки Жінки Гліома та рак мозку 7 150 5 590 Рак молочної залози 450 40 460 Рак передміхурової залози 27 050 Рак стравоходу 10 900 3 040 Колоректальний рак 26 000 26 180 Рак нирки 8 080 4 810 Рак підшлункової залози 16 840 16 530 Плоскоклітинна карцинома; Не Не Не Не Не кератиноцитні 1 000 000 визначено визначено визначено визначено визначено новоутворення шкіри 4 UA 109891 C2 Таблиця 1 Прогноз нових випадків захворювання на рак та смертельних випадків для обох статей для США в 2007 р. (Американське товариство по боротьбі з раком. Факти і цифри про рак - 2007 р. Атланта: American Cancer Society; 2007.) Локалізація Лейкемія Рак легенів Неходжкінська лімфома Рак яєчників Меланома 5 10 15 20 25 30 35 40 45 Прогноз нових випадків захворювання Обидві Чоловіки Жінки статі 44,240 24 800 19 440 213 380 114 760 98 620 63 190 34 210 28 990 22 430 22 430 59 940 33 910 26 030 Прогноз смертельних випадків Обидві статі 21 790 160 390 18 660 15 280 8 110 Чоловіки Жінки 12 320 89 510 9 600 9 470 70 880 9 060 15 280 2 890 5 220 Отже, існує необхідність в новому ефективному та безпечному методі лікування гліобластоми, пухлини передміхурової залози, раку молочної залози, раку стравоходу, колоректального раку, світлоклітинного раку нирки, раку легенів, ЦНС, раку яєчників, меланоми, раку підшлункової залози, плоскоклітинної карциноми, лейкемії, медулобластоми та інших пухлин, в яких спостерігається надмірна експресія сурвівіну, для покращення стану пацієнтів без використання хіміотерапевтичних речовин або інших засобів, які можуть призвести до серйозних побічних ефектів. Детальний опис винаходу Усі терміни, що використовуються в цьому винаході, мають визначення, наведені нижче, якщо не вказано інше. Термін "пептид" використовується тут для визначення ряду амінокислотних залишків, з'єднаних один з одним, як правило, пептидними зв'язками між альфа-аміно- та карбонільною групами суміжних амінокислот. Переважно пептиди мають довжину 9 амінокислот, але можуть мати довжину лише 8 амінокислот чи досягати довжини 10, 11, 12, 13, 14, 15, 16, 17 або 18 амінокислот. Термін "олігопептид" використовується тут для визначення ряду амінокислотних залишків, з'єднаних один з одним, як правило, пептидними зв'язками між альфа-аміно- та карбонільною групами суміжних амінокислот. Довжина олігопептиду не є критичною для винаходу, якщо в ньому вміщені належний епітоп чи епітопи. Олігопептиди, як правило, за довжиною є меншими, ніж приблизно 30 амінокислот, та більшими, ніж приблизно 14 амінокислот. Термін "поліпептид" означає ряд амінокислотних залишків, з'єднаних один з одним, як правило, пептидними зв'язками між альфа-аміно- та карбонільною групами суміжних амінокислот. Довжина поліпептиду не є критичною для винаходу, якщо він вміщує належні епітопи. На відміну від термінів "пептид" чи "олігопептид", термін "поліпептид" вживається для визначення молекул, які вміщують більш ніж приблизно 30 амінокислотних залишків. Пептид, олігопептид, білок чи полінуклеотид, що кодує таку молекулу, є "імуногенним" (тобто, "імуногеном" в рамках цього винаходу), якщо він здатний індукувати імунну відповідь. У цьому винаході імуногенність більш специфічно визначена як здатність індукувати Т-клітинну відповідь. Отже, "імуноген" буде молекулою, здатною індукувати імунну відповідь, та у цьому винаході, - молекулою, здатною індукувати Т-клітинну відповідь. Для T-клітинного "епітопу" потрібний короткий пептид, що зв'язується з рецептором MHC класу I чи II, формуючи потрійний комплекс (альфа-ланцюг MHC класу I, бета-2-мікроглобулін та пептид), здатний розпізнаватися T-клітиною, яка має відповідний T-клітинний рецептор, що зв'язується з MHC/пептидним комплексом з належною афінністю. Пептиди, що зв'язуються з MHC-молекулами класу I, як правило, мають довжину 8-14 амінокислот, і найбільш типовою є довжина 9 амінокислот. Епітопи T-клітин, що зв'язуються з MHC-молекулами класу II, як правило, мають довжину 12-30 амінокислот. У випадку пептидів, котрі зв'язуються з молекулами MHC класу II, один і той самий пептид та відповідний T-клітинний епітоп можуть займати спільний коровий сегмент, але мати різну загальну довжину, внаслідок того, що бокові послідовності відрізняються за довжиною вище аміно-кінця корової послідовності та нижче її карбоксильного кінця, відповідно. Рецептори MHC класу II мають більш відкриту конформацію, пептиди, зв'язані з рецепторами MHC класу II, відповідно, не повністю заглиблені в структуру пептидно-зв'язувальної порожнини МНС-молекули класу II, як в пептидно-зв'язувальній порожнині MHC-молекули класу І. Дивно, але це не відноситься до пептиду послідовності SEQ ID NO: 1, оскільки невеликі варіації в довжині пептиду ведуть до значного зниження активності 5 UA 109891 C2 5 10 15 (див. нижче). У людей існують три різні генетичні локуси, які кодують МНС-молекули класу I (MHCмолекули людини також визначаються як лейкоцитарні антигени людини (HLA)): HLA-A, HLA-B та HLA-C. HLA-A*0l, HLA-A*02 та HLA-A*ll є прикладами різних алелей MHC класу І, що можуть експресуватися з цих локусів. В геномі людини є три різні локуси для генів MHC класу II: HLA-DR, HLA-DQ та HLA-DP. Рецептори MHC класу II є гетеродимерами, що складаються з альфа- та бета-ланцюгу, обидва з яких закріплюються в клітинній мембрані через трансмембранну ділянку. HLA-DRB1*04 та HLA-DRB1*07 - два приклади різних бета-алелей MHC класу II, котрі, як відомо, кодуються в цих локусах. Алелі класу II дуже поліморфні, наприклад, описано кількасот різних HLA-DRB1алелей. Для HLA-A*02 та серотипів HLA-DR, що зустрічаються найчастіше, частота експресії в різних популяціях показана в Таблиці 2. Таблиця 2: Надає частоту експресії F для HLA*A02 та серотипи HLA-DR, що зустрічаються найчастіше. Значення частоти експресії виводяться із частот гаплотипу G f в американській популяції, на основі даних роботи Mori і співавт. (Mori і співавт., 1997), де використовується формула Харді-Вейнберга F=1-(1-Gf)². Комбінації A*02 з визначеними алелями HLA-DR можуть бути збагаченими чи менш частими, ніж очікувалося від їхніх окремих частот, внаслідок нерівномірного розподілу зв'язків. За детальними даними звертайтесь до роботи Chanock і співавт. (Chanock і співавт., 2004). 20 Таблиця 2 Частота експресії HLA*02 та серотипів HLA-DR в межах Північно-Американських субпопуляцій HLA-алель A*02 DR1 DR2 DR3 DR4 DR5 DR6 DR7 DR8 DR9 25 Білі американці 49,1 % 19,4 % 28,2 % 20,6 % 30,7 % 23,3 % 26,7 % 24,8 % 5,7 % 2,1 % Афро-американці 34,1 % 13,2 % 29,8 % 24,8 % 11,1 % 31,1 % 33,7 % 19,2 % 12,1 % 5,8 % Монголоїди 43,2 % 6,8 % 33,8 % 9,2 % 28,6 % 30,0 % 25,1 % 13,4 % 12,7 % 18,6 % Латиноамериканці 48,3 % 15,3 % 21,2 % 15,2 % 36,8 % 20,0 % 31,1 % 20,2 % 18,6 % 2,1 % Таблиця 3: надає частоту експресії F для HLA*A024 та серотип HLA*A02402. Значення частоти експресії виводяться із частот гаплотипу G f в межах популяції, на основі даних роботи Mori і співавт. (Mori і співавт., 1997), де використовується формула Харді-Вейнберга F=1-(1-Gf)². Детальні дані див. у роботі Chanock і співавт. (Chanock і співавт., 2004). Таблиця 3 Частота експресії HLA*02 та серотипів A*2402 в усьому світі Алель A*24 A*24 A*2402 A*24 A*2402 A*24 A*24 A*24 A*24 A*2402 A*24 A*24 A*24 Популяція Філіппіни Росія, ненецький округ Японія Малайзія Філіппіни Індія Південна Корея Шрі-Ланка Китай Індія Австралія, Захід США Росія, Самара 6 Розрахований фенотип з частоти алеля 65 % 61 % 59 % 58 % 54 % 47 % 40 % 37 % 32 % 29 % 22 % 22 % 20 % UA 109891 C2 Таблиця 3 Частота експресії HLA*02 та серотипів A*2402 в усьому світі Алель A*24 A*24 5 10 15 20 25 30 35 40 45 50 Популяція Південна Америка Європа Розрахований фенотип з частоти алеля 20 % 18 % Отже, для терапевтичних та діагностичних цілей в значній мірі доцільним є пептид, що зв'язується з належною афінністю з кількома різними рецепторами HLA класу II. Пептид, який зв'язується з кількома різними молекулами HLA класу II, називається пептидом, що зв'язується нерозбірливо. Посилання на ДНК-послідовність в цьому винаході включає однониткові та двониткові ДНК. Отже, конкретна послідовність, якщо контекст не передбачає іншого, відноситься до однониткової ДНК такої послідовності, дуплексу такої послідовності з її комплементом (двониткова ДНК) та комплементу такої послідовності. Термін "кодуюча ділянка" відноситься до тієї частини гена, яка природним чи нормальним способом кодує продукт експресії того гена в його природному геномному середовищі, тобто, до ділянки, що кодує in vivo природний продукт експресії гена. Кодуюча ділянка може бути з нормального гена, або гена, що мутував чи був змінений, або навіть бути з послідовності ДНК чи гена, повністю синтезованого в лабораторії з використанням методів, добре відомих досвідченим фахівцям в галузі синтезу ДНК. Термін "нуклеотидна послідовність" відноситься до гетерополімеру дезоксирибонуклеотидів. Нуклеотидна послідовність, що кодує конкретний пептид, олігопептид чи поліпептид, може мати природне походження чи створюватися синтетичним шляхом. В цілому, сегменти ДНК, що кодують пептиди, поліпептиди та білки згідно цього винаходу, збираються з фрагментів кДНК та коротких олігонуклеотидних лінкерів або з серії олігонуклеотидів для одержання синтетичного гена, здатного експресуватися в рекомбінантній транскрипційній одиниці, яка вміщує регуляторні елементи, одержані з мікробного чи вірусного оперону. Термін "продукт експресії" означає поліпептид або білок, котрий є природним трансляційним продуктом гена та будь-якої нуклеїново-кислотної послідовності, що кодує еквіваленти, які є результатом деградації генетичного коду, і, таким чином, кодує ті ж амінокислоти. Термін "фрагмент", стосовно кодуючої послідовності, означає частину ДНК, яка вміщує менш ніж повну кодуючу ділянку, і продукт експресії якої зберігає по суті ту ж біологічну функцію чи активність, що й продукт експресії повної кодуючої ділянки. Термін "сегмент ДНК" відноситься до полімеру ДНК, у формі окремого фрагменту чи як компонент більшої структури ДНК, що одержаний з ДНК, виділеної принаймні один раз по суті в чистій формі, тобто, без забруднюючих ендогенних матеріалів та в кількості чи концентрації, яка дає змогу ідентифікувати, виконувати маніпуляції та відновлювати сегмент і його складові нуклеотидні послідовності за допомогою стандартних біохімічних методів, наприклад, з використанням вектору клонування. Такі сегменти надаються у формі відкритої рамки зчитування, без порушень внутрішніми нетрансльованими послідовностями чи інтронами, які зазвичай присутні в еукаріотичних генах. Послідовності нетрансльованих ДНК можуть бути присутні по низхідній від відкритої рамки зчитування, де вони не заважають маніпуляціям або експресії кодуючих ділянок. Термін "праймер" означає коротку нуклеїново-кислотну послідовність, котра може бути спарена з однією ниткою ДНК та надає вільний 3'OH-кінець, на якому ДНК-полімераза розпочинає синтез дезоксирибонуклеотидного ланцюга. Термін "промотор" означає ділянку ДНК, включену у зв'язування РНК-полімерази для ініціації транскрипції. Термін "виділений" означає, що матеріал видаляється зі свого первісного середовища (наприклад, природного середовища, якщо він має природне походження). Наприклад, природно існуючий полінуклеотид чи поліпептид, присутній у живих тварин, не є виділеним, але той самий полінуклеотид чи поліпептид, відокремлений від якихось чи всіх співіснуючих матеріалів в природній системі, є виділеним. Такі полінуклеотиди можуть бути часткою вектору, та/або такі полінуклеотиди чи поліпептиди можуть становити частину композиції і продовжувати бути виділеними, так що такий вектор чи композиція не є частиною свого природного середовища. Полінуклеотиди та рекомбінантні чи імуногенні поліпептиди, розкриті відповідно до цього винаходу, також можуть бути в "очищеній" формі. Термін "очищений" не вимагає абсолютної 7 UA 109891 C2 5 10 15 20 25 30 35 40 45 50 55 60 чистоти; він має на увазі відносне визначення та може включати препарати високого ступеню очищення або препарати, які тільки частково очищені, в тому сенсі, як спеціалісти в цій галузі розуміють такі терміни. Наприклад, окремі клони, виділені з бібліотеки кДНК, були у традиційний спосіб очищені до електрофоретичної однорідності. Очищення вихідного матеріалу чи природного матеріалу від домішок принаймні на порядок величини, переважно на два-три порядки, та більш переважно, на чотири-п'ять порядків величини, чітко передбачається в цьому винаході. Крім того, заявлений поліпептид, котрий має чистоту переважно в 99,999 %, або принаймні в 99,99 % чи 99,9 %; і навіть бажано 99 % за масою чи більше, також чітко пропонується. Нуклеїнові кислоти та продукти експресії поліпептидів, що розкриваються відповідно до цього винаходу, а також вектори експресії, які вміщують такі нуклеїнові кислоти та/або такі поліпептиди, можуть бути в "збагаченій формі". Термін "збагачений" в тому виді, в якому він використовується тут, означає, що концентрація матеріалу принаймні приблизно в 2, 5, 10, 100 або 1000 разів перевищує його природну концентрацію (наприклад), переважно 0,01 % за масою, принаймні переважно приблизно 0,1 % за масою. Також пропонуються збагачені препарати приблизно в 0,5 %, 1 %, 5 %, 10 % та 20 % за масою. Послідовності, конструкції, вектори, клони та інші матеріали, які складають цей винахід, можуть переважно бути у збагаченій чи виділений формі. Термін "активний фрагмент" означає фрагмент, що генерує імунну відповідь, тобто має імуногенну функцію, при введенні (окремо чи, необов'язково, з прийнятним ад'ювантом) тварині, наприклад, ссавцю, такому як кролик чи миша, і також включаючи людину. Така імунна відповідь має вид стимуляції Т-клітинної відповіді у тварини-реципієнта, такої як людина. Альтернативно, "активний фрагмент" також може використовуватись для індукції Т-клітинної відповіді in vitro. В цьому винаході терміни "частка", "сегмент" і "фрагмент", якщо вони використовуються по відношенню до поліпептидів, означають безперервну послідовність залишків, таких як амінокислотні залишки, причому ця послідовність утворює підгрупу більшої послідовності. Наприклад, якщо поліпептид був підданий обробці будь-якою з типових ендопептидаз, таких як трипсин або хімотрипсин, олігопептиди, одержані в результаті такої обробки, будуть представляти частки, сегменти чи фрагменти початкового поліпептиду. Це означає, що будьякий такий фрагмент буде обов'язково вміщувати як частину своєї амінокислотної послідовності сегмент, фрагмент або частину, яка є по суті ідентичною, якщо не повністю ідентичною, послідовності від SEQ ID NO: 1 до 20, котра відповідає природним або "вихідним" білкам послідовностей від SEQ ID NO: 1 до 20. Якщо такі терміни використовуються стосовно полінуклеотидів, вони означають продукти, одержані після обробки зазначених полінуклеотидів будь-якою з типових ендонуклеаз. Відповідно до цього винаходу, термін "процентна ідентичність" або "ідентичний з процентною долею" відносно послідовності, означає, що послідовність порівнюється із заявленою або описаною послідовністю після вирівнювання послідовності, яка порівнюється ("Порівнювана послідовність"), з описаною або заявленою послідовністю ("Контрольна послідовність"). Процентна ідентичність визначається відповідно до наведеної нижче формули: Процентна ідентичність = 100 [I -(C/R)] де C - кількість відмінностей між Контрольною послідовністю та Порівнюваною послідовністю на довжині між Контрольною послідовністю та Порівнюваною послідовністю, де (i) кожна основа чи амінокислота в Контрольній послідовності, котра не має відповідної вирівняної основи чи амінокислоти у Порівнюваній послідовності, і (ii) кожній розрив у Контрольній Послідовності та (iii) кожна вирівняна основа чи амінокислота у Контрольній послідовності, яка відрізняється від вирівняної основи чи амінокислоти у Порівнюваній послідовності, являє собою різницю; та R - це кількість основ чи амінокислот в Контрольній послідовності на довжині вирівнювання з Порівнюваною послідовністю з будь-яким розривом, що утворився в Контрольній послідовності, також зарахованим як основа чи амінокислота. Якщо існує вирівнювання між Порівнюваною послідовністю та Контрольною послідовністю, для якої процентна ідентичність, розрахована вище, є приблизно рівною чи більшою, ніж зазначена мінімальна Процентна ідентичність, то Порівнювана послідовність має зазначену мінімальну процентну ідентичність до Контрольної послідовності, навіть якщо можуть існувати вирівнювання, в яких розрахована як описано вище Процентна ідентичність є меншою, ніж зазначена Процентна ідентичність. Первісні пептиди, що розкриваються в цьому винаході, можуть модифікуватися заміщенням одного чи кількох залишків на різних, можливо, селективних, ділянках пептидного ланцюгу, якщо не зазначається інше. Такі заміщення можуть бути консервативного характеру, наприклад, коли 8 UA 109891 C2 5 10 15 20 25 30 35 40 45 50 55 60 одна амінокислота замінюється амінокислотою подібної структури та характеристик, тобто, коли гідрофобна амінокислота замінюється іншою гідрофобною амінокислотою. Навіть більш консервативною буде заміна амінокислот такого ж чи подібного розміру та хімічного характеру, наприклад, коли лейцин замінюється ізолейцином. В дослідженнях варіацій послідовностей в сімействах природних гомологічних білків певні амінокислотні заміщення частіше допускаються, ніж інші, і вони часто демонструють кореляцію зі схожістю в розмірі, заряді, полярності та гідрофобності між первісною амінокислотою та її заміною, і це є основою для визначення "консервативних заміщень". Консервативні заміщення визначаються тут як заміни в межах однієї з наведених нижче п'яти груп: Група 1 - малі аліфатичні неполярні чи слабо полярні залишки (Ala, Ser, Thr, Pro, Gly); Група 2 - полярні негативно заряджені залишки та їхні аміди (Asp, Asn, Glu, Gln); Група 3 полярні позитивно заряджені залишки (His, Arg, Lys); Група 4 - великі аліфатичні неполярні залишки (Met, Leu, Ile, Val, Cys); та Група 5 - великі ароматичні залишки (Phe, Tyr, Trp). Менш консервативні заміщення можуть включати заміну однієї амінокислоти іншою, котра має подібні характеристики, але трохи відрізняється за розміром, наприклад, заміна аланіну ізолейциновим залишком. Високо-неконсервативні заміни можуть включати заміщення кислотної амінокислоти полярною, або навіть такою, що є основною за своїм характером. Такі "радикальні" заміщення не можуть, однак, відхилятись як потенційно неефективні, оскільки їхні хімічні наслідки не є повністю прогнозованими, та радикальні заміщення також можуть несподівано призвести до сприятливих ефектів, які неможливо передбачити за простими хімічними принципами. Безумовно, такі заміщення можуть включати структури, інші ніж звичайні L-амінокислоти. Отже, D-амінокислоти можуть заміщуватися L-амінокислотами, котрі зазвичай виявляються в антигенних пептидах згідно винаходу та охоплюються розкриттям в ньому. Крім того, амінокислоти, які мають нестандартні R-групи (тобто, R-групи, інші ніж можна знайти в типових 20 амінокислотах природних білків) також можуть використовуватись для заміщення з метою виробництва імуногенів та імуногенних поліпептидів відповідно до цього винаходу. Якщо виявляється, що заміщення в більш ніж одній позиції призводять до утворення пептиду по суті з еквівалентною чи більшою антигенною активністю, як визначено нижче, тоді комбінації цих заміщень будуть досліджуватись з метою визначення, чи мають комбіновані заміщення додатковий чи синергічний вплив на антигенність пептиду. Більшою частиною не більш ніж 4 позиції в пептиді замінюються одночасно. Термін "Т-клітинна відповідь" означає специфічну проліферацію та активацію ефекторних функцій, індукованих пептидом in vitro чи in vivo. Для ЦТЛ, рестриктованих за MHC класу I, ефекторними функціями можуть бути лізис клітин-мішеней із завантаженим пептидом, пептидним попередником чи клітин-мішеней, природно презентуючих пептид, секреція цитокінів, переважно гамма-інтерферону, TNF-альфа, або ІЛ-2, індукована пептидом, секреція ефекторних молекул, переважно, гранзимів чи перфоринів, індукована пептидом, або дегрануляція. Для Т-клітин- хелперів, рестриктованих за MHC класу II, ефекторними функціями ймовірно можуть бути індукована пептидом секреція цитокінів, переважно, IFN-гамма, TNFальфа, ІЛ-4, ІЛ-5, ІЛ-10 або ІЛ-2, чи індукована пептидом дегрануляція. Можливі ефекторні функції для ЦТЛ і Т-клітин-хелперів не обмежуються цим переліком. Імунотерапевтичні підходи до лікування Стимуляція імунної реакції залежить від наявності антигенів, які визнаються імунною системою хазяїна як чужорідні. Виявлення існування пухлино-асоційованих антигенів дало можливість використовувати імунну систему хазяїна для втручання у ріст пухлини. Зараз досліджуються різні механізми використання як гуморальної, так і клітинної ланки імунної системи для імунотерапії раку. Окремі елементи клітинної імунної відповіді здатні специфічно розпізнавати та знищувати клітини пухлин. Виділення цитотоксичних T-клітин (ЦТЛ) з пухлино-інфільтруючих клітинних популяцій чи з периферичної крові наводить на думку, що такі клітини грають важливу роль в природному імунному захисті проти раку. Зокрема, CD8-позитивні T-клітини, які розпізнають молекули класу I головного комплексу гістосумісності (MHC), що зв'язані з пептидами зазвичай з 8-10 залишків, що одержані з білків чи дефектних продуктів рибосом (DRIPS) (Schubert U, Antón LC, Gibbs J, Norbury CC, Yewdell JW, Bennink JR.; Rapid degradation of a large fraction of newly synthesized proteins by proteasomes; Nature 2000; 404(6779):770-774), котрі розташовані в цитозолях, відіграють значну роль в цій реакції. MHC-молекули людини також визначаються як лейкоцитарні антигени людини (HLA). Є два класи MHC-молекул: MHC-молекули класу I, котрі можна знайти на більшості клітин, що мають ядро; вони презентують пептиди, які є результатом протеолітичного розщеплення, 9 UA 109891 C2 5 10 15 20 25 30 35 40 45 50 55 60 переважно, ендогенних, цитозольних білків чи білків ядра, DRIPS та більших за розміром пептидів. Однак пептиди, одержані з ендосомних компартментів чи екзогенних джерел, також часто зустрічаються на молекулах MHC класу I. Цей некласичний спосіб презентації класом I визначається як крос-презентацією в науковій літературі. MHC-молекули класу II, переважно, знаходяться на професійних антигенпрезентуючих клітинах (АПК); вони презентують пептиди екзогенних білків, що поглинаються АPC в ході ендоцитозу та згодом процесуються. Стосовно класу І, описуються альтернативні способи процесингу антигенів, що дозволяють пептидам з ендогенних джерел презентуватися молекулами MHC класу II (наприклад, аутофагоцитоз). Комплекси пептиду та MHC-молекули класу I розпізнаються CD8-позитивними цитотоксичними T-лімфоцитами, які несуть відповідні Т-клітинні рецептори (ТКР), в той час як комплекси пептиду і MHC-молекули класу II розпізнаються CD4-позитивними хелперними Т-клітинами, що несуть відповідні ТКР. CD4-позитивні хелперні T-клітини відіграють важливу роль в регулюванні ефекторних функцій протипухлинних відповідей T-клітин, і з цієї причини ідентифікація епітопів CD4позитивних T-клітин, які одержуються з пухлино-асоційованих антигенів (TAA) може мати велике значення для розробки фармацевтичних продуктів для викликання протипухлинних імунних реакцій (Gnjatic, S., D. Atanackovic, E. Jäger, M. Matsuo, A. Selvakumar, N.K. Altorki, R.G. Maki, B. Dupont, G. Ritter, Y.T. Chen, A. Knuth, and L.J. Old. Survey of naturally occurring CD4+T-cell responses against NY-ESO-1 in cancer patients: Correlation with antibody responses. Proc. Natl. Acad. Sci. U.S.A. 2003, 100 (15): 8862-7). CD4-позитивні T-клітини можуть призвести до локального зростання рівнів гамма-інтерферону. На моделях ссавців, наприклад, мишей, було продемонстровано, що навіть за відсутністю ефекторних клітин ЦТЛ (тобто, CD8-позитивних T-лімфоцитів), CD4-позитивні T-клітини є достатніми для інгібування проявів пухлин шляхом інгібування ангіогенезу завдяки секреції гамма-інтерферону (IFNγ) (Qin, Z. and T. Blankenstein. CD4+T-cell--mediated tumor rejection involves inhibition of angiogenesis that is dependent on IFN gamma receptor expression by nonhematopoietic cells. Immunity. 2000, 12:677-686). Крім того, було показано, що CD4-позитивні T-клітини, які розпізнають пептиди з пухлино-асоційованих антигенів, представлених молекулами HLA класу II, можуть протидіяти розвитку пухлин за допомогою індукції реакцій антитіл (Ab) (Kennedy, R.C., M.H. Shearer, A.M. Watts, and R.K. Bright. CD4+T lymphocytes play a critical role in antibody production and tumor immunity against simian virus 40 large tumor antigen. Cancer Res. 2003, 63:1040-1045). На відміну від пухлино-асоційованих пептидів, які зв'язуються з молекулами HLA класу I, лише невелика кількість лігандів класу II TAA була описана на цей час (www.cancerimmunity.org, www.syfpeithi.de). Оскільки конститутивна експресія молекул HLA класу II зазвичай обмежується клітинами імунної системи, можливість виділення пептидів класу II безпосередньо з первинних пухлин не вважалася ймовірною. Однак автори винаходу за останній час досягли успіху в ідентифікації ряду епітопів MHC класу II безпосередньо з пухлин (EP 1642905, EP 1760088; Dengjel J, Nastke MD, Gouttefangeas C, Gitsioudis G, Schoor O, Altenberend F, Müller M, Krämer B, Missiou A, Sauter M, Hennenlotter J, Wernet D, Stenzl A, Rammensee HG, Klingel K, Stevanović S.; Unexpected abundance of HLA class II presented peptides in primary renal cell carcinomas; Clin Cancer Res. 2006; 12:4163-4170). За відсутності запалення, експресія молекул MHC класу II, переважно, зводиться до клітин імунної системи, особливо до АПК, наприклад, моноцитів, клітин, що походять з моноцитів, макрофагів, дендритних клітин. При дослідженні пацієнтів з пухлинами було несподівано виявлено, що клітини пухлини експресують MHC-молекули класу II (Dengjel J, Nastke MD, Gouttefangeas C, Gitsioudis G, Schoor O, Altenberend F, Müller M, Krämer B, Missiou A, Sauter M, Hennenlotter J, Wernet D, Stenzl A, Rammensee HG, Klingel K, Stevanović S.; Unexpected abundance of HLA class II presented peptides in primary renal cell carcinomas; Clin Cancer Res. 2006; 12:4163-4170). Для того, щоб пептид індукував (викликав) клітинну імунну відповідь, він повинен зв'язуватися з MHC-молекулою. Цей процес залежить від алеля MHC-молекули та специфічних поліморфізмів амінокислотної послідовності пептиду. Пептиди, зв'язані з MHC класу I, як правило, мають довжину 8-10 амінокислотних залишків та зазвичай вміщують два консервативні залишки ("якір") в своїй послідовності, що взаємодіє з відповідною зв'язувальною порожниною MHC-молекули. Таким чином, кожний MHC-алель має "зв'язувальний мотив", що визначає, які пептиди можуть специфічно зв'язуватися зі зв'язувальною порожниною (Rammensee HG, Bachmann J, Stevanovic S. MHC ligands and peptide motifs, Landes Bioscience, USA, 1997). В імунній реакції, залежній від MHC, пептиди повинні не тільки мати здатність зв'язуватися з 10 UA 109891 C2 5 10 15 20 25 30 35 40 45 50 55 60 певними молекулами MHC, експресованими пухлинними клітинами, вони також мають розпізнаватися T-клітинами, що несуть специфічні T-клітинні рецептори (ТКР). Антигени, котрі розпізнаються пухлино-специфічними цитотоксичними T-лімфоцитами, тобто, їхні епітопи, можуть бути молекулами, одержаними з усіх класів білків, таких як ферменти, рецептори, транскрипційні фактори та ін. Крім того, пухлино-асоційовані антигени, наприклад, також можуть бути присутніми тільки в клітинах пухлин, зокрема, як продукти генів, що мутували. Іншим важливим класом пухлино-асоційованих антигенів є тканино-специфічні антигени, такі як CT ("ракові тестикулярні")-антигени, що експресуються в різних видах пухлин та в здоровій тканині сім'яників. Було ідентифіковано різні пухлино-асоційовані антигени. Крім того, проведена значна дослідницька робота по ідентифікації додаткових пухлино-асоційованих антигенів. Деякі групи пухлино-асоційованих антигенів, котрі також згадуються в літературі як пухлино-специфічні антигени, є тканино-специфічними. Приклади включають, без обмежень, тирозиназу для меланоми, простато-специфічний антиген (PSA) та простато-специфічний мембранний антиген (PSMA) для раку передміхурової залози і хромосомні кросовери (транслокації), такі як bcr/abl у лімфомі. Однак ряд ідентифікованих пухлино-асоційованих антигенів зустрічається в багатьох видах пухлин, а деякі з них, такі як онкогенні білки та/або гени-супресори пухлин (огляд генівсупресорів пухлин для раку нирки наведений, наприклад, в роботі Linehan WM, Walther MM, Zbar B. The genetic basis of cancer of the kidney. J Urol. 2003 Dec; 170 (6Pt1):2163-72), котрі фактично викликають трансформаційну дію, зустрічаються практично в усіх видах пухлин. Наприклад, звичайні білки клітин, що контролюють ріст та диференціацію клітин, такі як p53 (приклад гена-супресора пухлин), ras, c-met, myc, pRB, VHL та HER-2/neu, можуть накопичувати мутації, і це призводить до посилення експресії цих генних продуктів, таким чином роблячи їх онкогенними (McCartey et al. Cancer Research, 1998, 15:58 2601-5; Disis et al. Ciba Found. Symp. 1994, 187:198-211). Ці білки-мутанти також можуть бути мішенню пухлино-специфічної імунної відповіді при багатьох видах ракових захворювань. Імунотерапія у пацієнтів, хворих на рак, чітко спрямована на активацію клітин імунної системи специфічним чином, зокрема так званих цитотоксичних T-клітин (ЦТЛ, також відомих як "клітини-кілери" і як CD8-позитивні T-клітини), проти пухлинних клітин, а не проти здорових тканин. Клітини пухлин відрізняються від здорових клітин експресією пухлино-асоційованих білків. Молекули HLA на клітинній поверхні презентують клітинний вміст назовні, таким чином дозволяючи цитотоксичній T-клітині проводити диференціацію між здоровою та пухлинною клітиною. Це здійснюється за допомогою розщеплення усіх білків всередині клітини на короткі пептиди, котрі потім приєднуються до молекул HLA і презентуються на клітинній поверхні (Rammensee, 1993). Пептиди, які презентуються на пухлинних клітинах, але не презентуються або в незначній мірі презентуються на здорових клітинах тіла, називаються пухлиноасоційованими пептидами (TUMAP). Для того, щоб білки розпізнавалися цитотоксичними T-лімфоцитами як пухлино-специфічні або пухлино-асоційовані антигени, та з метою застосування в лікуванні, мають бути дотримані деякі попередні умови. Антиген повинен експресуватися, головним чином, клітинами пухлин, а не нормальними здоровими тканинами, чи бути присутнім в здорових клітинах в порівняно невеликих кількостях. Крім того, бажано, щоб відповідний антиген був не тільки присутнім в будь-якому виді пухлини, але також знаходився у високих концентраціях (наприклад, кількість копій відповідного пептиду на клітину). Пухлино-специфічні та пухлино-асоційовані антигени часто походять з білків, що беруть безпосередню участь у трансформації нормальної клітини в пухлинну клітину внаслідок їхньої функції, наприклад, в контролі клітинного циклу чи апоптозі. До того ж низхідні мішені цих білків, які є основною причиною трансформації, можуть бути представлені у підвищеній кількості і, відповідно, можуть бути непрямим чином асоційованими з пухлинами. Такі непрямі пухлино-асоційовані антигени також можуть бути мішенями у вакцинаційному підході. В обох випадках істотно важливою є присутність епітопів в амінокислотній послідовності антигену, оскільки такий пептид ("імуногенний пептид"), що одержується з пухлино-асоційованого антигену, повинен призводити до Т-клітинної відповіді in vitro чи in vivo. По суті, будь-який пептид, здатний зв'язувати молекулу MHC, може функціонувати як Tклітинний епітоп. Передумовою для індукції Т-клітинної відповіді in vitro чи in vivo є наявність Tклітини з відповідним рецептором ТКР та відсутність толерантності до цього конкретного епітопу. Хелперні T-клітини відіграють важливу роль в регуляції ефекторної функції ЦТЛ в протипухлинному імунітеті. Епітопи Т-хелперів, які запускають реакцію цих клітин типу TH1, підтримують ефекторні функції CD8-позитивних T-клітин-кілерів, котрі включають цитотоксичні функції, спрямовані проти клітин пухлин, що виявляють комплекси пухлино-асоційованого 11 UA 109891 C2 5 10 15 20 25 30 35 40 45 50 55 60 пептиду/MHC на своїх клітинних поверхнях. Таким чином, епітопи пухлино-асоційованих пептидів Т-хелперів, самостійно чи в комбінації з іншими пухлино-асоційованими пептидами, можуть служити активними фармацевтичними інгредієнтами композицій вакцин, які стимулюють протипухлинні імунні відповіді. Оскільки обидва типи відповідей, залежні від CD8 та CD4, спільно та синергічно роблять свій внесок у протипухлинну дію, для розробки протипухлинних вакцин важливими є ідентифікація та характеристика пухлино-асоційованих антигенів, які розпізнаються CD8позитивними ЦТЛ (молекули MHC класу I) чи CD4- позитивними ЦТЛ (молекули MHC класу II). Отже, задача цього винаходу полягає в знаходженні композицій пептидів, які вміщують пептиди, що зв'язуються з MHC-комплексами будь-якого класу. Перші клінічні дослідження з використанням пухлино-асоційованих пептидів розпочали в середині 90-х років минулого століття Бун (Boon) та його колеги, переважно, стосовно меланоми. Клінічні відповіді в найбільш вдалих з цих досліджень варіювалися від 10 % до 30 %. Про тяжкі побічні ефекти чи серйозну аутоімунність не повідомлялося в будь-яких клінічних дослідженнях із використанням монотерапії вакциною на основі пептидів. Є дані про вітиліго у слабкій формі, що спостерігалася у деяких пацієнтів, які проходили лікування меланомоасоційованими пептидами. Однак праймінг одного виду ЦТЛ зазвичай є недостатнім для знищення усіх пухлинних клітин. Пухлини дуже мутагенні і тому здатні швидко реагувати на атаки ЦТЛ, змінюючи свою білкову модель для уникнення розпізнавання ЦТЛ. З метою протидії пухлинним механізмам уникнення для вакцинації використовується широкий діапазон специфічних пептидів. У такий спосіб проти клітини може бути розпочата широка одночасна атака відразу кількома ЦТЛклонами. Це може зменшити шанси того, що пухлина уникатиме імунної відповіді. Така гіпотеза була недавно підтверджена в клінічному дослідженні лікування пацієнтів, хворих на меланому на пізній стадії. Лише за кількома винятками, пацієнти, що мали якнайменш три визначені Tклітинні відповіді, продемонстрували об'єктивні клінічні відповіді чи стабілізацію захворювання (Banchereau і співавт., 2001), а також збільшену виживаність (інформація одержана при особистому спілкуванні з J. Banchereau), в той час як у широкої більшості пацієнтів з менш ніж трьома T-клітинними реакціями було діагностовано прогресуюче захворювання. Вивчення робіт заявниками на цей патент показало подібний ефект, коли пацієнти, хворі на нирково-клітинну карциному, проходили лікування вакциною, що складалася з 13 різних пептидів (H. Singh-Jasuja, S. Walter, T. Weinschenk, A. Mayer, P. Y. Dietrich, M. Staehler, A. Stenzl, S. Stevanovic, H. Rammensee, J. Frisch; Correlation of T-cell response, clinical activity and regulatory T-cell levels in renal cell carcinoma patients treated with IMA901, a novel multi-peptide vaccine; ASCO Meeting 2007 Poster # 3017; M. Staehler, A. Stenzl, P. Y. Dietrich, T. Eisen, A. Haferkamp, J. Beck, A. Mayer, S. Walter, H. Singh, J. Frisch, C. G. Stief; An open label study to evaluate the safety and immunogenicity of the peptide based cancer vaccine IMA901, ASCO meeting 2007; Poster # 3017). Основна задача розробки протипухлинної вакцини полягає, таким чином, не тільки в ідентифікації та характеристиці нових пухлино-асоційованих антигенів та епітопів імуногенних Тхелперів, які походять з них, але й в комбінуванні різних епітопів для збільшення ймовірності відповіді більш ніж на один епітоп для кожного пацієнта. Отже, задача цього винаходу полягає в знаходженні комбінацій амінокислотних послідовностей таких пептидів, що мають здатність зв'язуватися з молекулою головного комплексу гістосумісності людини (MHC) класу I (HLA класу I) чи II (HLA класу II). Додаткова задача цього винаходу полягає в наданні ефективної протиракової вакцини, яка базується на комбінації пептидів. Автори цього винаходу виділили і характеризували пептиди, які зв'язуються з молекулами HLA класу I або II, безпосередньо з пухлин ссавців, тобто, зразків первинних пухлин у пацієнтів, переважно, хворих на рак шлунка, але також з первинних зразків тканин гліобластоми, колоректальних карцином, нирково-клітинної карциноми, ракових пухлин легенів, підшлункової залози, злоякісної меланоми та ракової пухлини шлунка. Цей винахід пропонує пептиди, що походять з антигенів, які асоціюються з онкогенезом, та мають здатність зв'язуватися в достатній мірі з молекулами MHC (HLA) класу II для запуску імунної відповіді лейкоцитів людини, зокрема, лімфоцитів, в особливості, T-лімфоцитів, головним чином, CD4-позитивних T-лімфоцитів, а конкретніше, CD4-позитивних T-лімфоцитів, що виступають посередниками в імунних відповідях T H1-типу. Цей винахід також пропонує пептиди, що походять з антигенів, які асоціюються з онкогенезом, та мають здатність зв'язуватися в достатній мірі з молекулами MHC (HLA) класу I для запуску імунної відповіді лейкоцитів людини, зокрема, лімфоцитів, в особливості, Tлімфоцитів, головним чином, CD8-позитивних цитотоксичних T- лімфоцитів, а також комбінацій 12 UA 109891 C2 5 10 15 20 25 30 з двох варіантів, що є особливо прийнятними для вакцинації пацієнтів, хворих на рак. Відповідно до цього винаходу, задача вирішується шляхом надання фармацевтичної композиції, котра складається принаймні з двох пептидів, що вміщують амінокислотну послідовність, вибрану з групи, яка включає послідовності від SEQ ID NO 1 до SEQ ID NO 10, та/або вміщує варіант амінокислотної послідовності, що є принаймні на 85 % гомологічним послідовності від SEQ ID NO 1 до SEQ ID NO 10, та/або полінуклеотид, який вміщує нуклеїнову кислоту, що кодує послідовності від SEQ ID NO 1 до SEQ ID NO 10 або варіант амінокислотної послідовності, та фармацевтично прийнятний носій. Фармацевтичні композиції згідно цього винаходу можуть також вміщувати якнайменш один додатковий пептид, який вміщує амінокислотну послідовність, вибрану з групи, яка включає послідовності від SEQ ID NO: 11 до SEQ ID NO: 22, або включає варіант амінокислотної послідовності, що є принаймні на 85 % ідентичний послідовності від SEQ ID NO: 11 до SEQ ID NO: 22, чи полінуклеотид, який вміщує нуклеїнову кислоту, що кодує послідовності від SEQ ID NO: 11 до SEQ ID NO: 22 або варіант амінокислотної послідовності. Пептиди можуть мати загальну довжину від 8 до 100 амінокислот, переважно від 8 до 30 амінокислот, та найбільш переважно від 8 до 17 амінокислот. Пептиди також можуть мати непептидні зв'язки. Як описується нижче, пептиди, що є основою цього винаходу, були усі ідентифіковані як презентовані клітинами, що несуть в собі MHC класу I чи II. Отже, усі ці конкретні пептиди, а також інші пептиди, які включають цю послідовність (тобто, похідні пептиди) викликають специфічну T-клітинну відповідь, хоча ступінь, в якій ця відповідь буде індукуватися, може варіюватися в залежності від конкретного пептиду та пацієнта. Різниця, наприклад, може бути викликана мутаціями в пептидах. Фахівцю, який має досвід в цій галузі, добре відомі методи, що можуть бути застосовані для визначення ступеню індукції відповіді окремого пептиду, зокрема, із посиланням на приклади, наведені в цьому винаході, та відповідну літературу. Переважно, варіанти за винаходом будуть індукувати T-клітинну перехресну реакцію з відповідним пептидом згідно винаходу. Пептиди походять з пухлино-асоційованих антигенів, особливо, із пухлино-асоційованих антигенів з функціями, наприклад, в протеолізі, ангіогенезі, рості клітин, регуляції клітинного циклу, поділі клітин, регуляції транскрипції, регуляції трансляції, інвазії тканин і т. ін. В Таблиці 3 наведені пептиди та функція білка, з якого походять пептиди. Таблиця 4 Пептиди згідно цього винаходу і функція вихідного білка Ід. № послідовності 1 2 3 4 5 6 7 8 9 10 20 24 Ід. номер пептиду Послідовність Символ гена Зв'язується з MHC CDC2-001 ASPM-002 UCHL5-001 MET-006 PROM1-001 UQCRB-001 MST1R-001 PPAP2C-001 SMC4-001 MMP11-001 AVL9-001 ERBB3-001 LYQILQGIVF SYNPLWLRI NYLPFIMEL SYIDVLPEF SYIIDPLNL YYNAAGFNKL NYLLYVSNF AYLVYTDRL HYKPTPLYF VWSDVTPLTF FYISPVNKL VYIEKNDKL CDK1 ASPM UCHL5 MET PROM1 UQCRB MST1R PPAP2C SMC4 MMP11 AVL9 ERBB3 HLA-A*024 HLA-A*024 HLA-A*024 HLA-A*024 HLA-A*024 HLA-A*024 HLA-A*024 HLA-A*024 HLA-A*024 HLA-A*024 HLA-A*024 HLA-A*024 13 UA 109891 C2 Таблиця 5 Додаткові імуногенні пептиди, доцільні в композиції згідно винаходу Ід.№ послідовності 11 12 13 14 15 16 17 18 19 20 21 22 10 15 20 25 30 35 TLGEFLKLDRERAKN DDPSTIEKLAKNKQKP NKQKPITPETAEKLARD NGAYKAIPVAQDLNAPS TLGEFLKLDRERAKD FTELTLGEF LMLGEFLKL EPDLAQCFY VYGIRLEHF FYISPVNKL TYGNLLDYL RFLSGIINF 23 5 BIR-002 CDC42-001 CDC42-002 SPP1-001 BIR-002a BIR-002b BIR-002c BIR-002d NUF2-001 AVL9-001 ABL1-001 NUF2-002 (HBV-001) FLPSDFFPSV Ід.номер пептиду Послідовність Символ гена BIRC5 CDC42 CDC42 SPP1 сурвівін сурвівін сурвівін сурвівін NUF2 AVL9 ABL1 NUF2 контрольний пептид Зв'язується з MHC HLA-DR та HLA-A*02 HLA-DR HLA-DR HLA-DR HLA-DR та HLA-A*02 HLA-A1 HLA-A2 HLA-B35 HLA-A*024 HLA-A*024 HLA-A*024 HLA-A*024 Білок циклу поділу клітин 2 (CDC2) Білок CDC2, також відомий як p34cdc2 або Cdk1 (циклін-залежна кіназа 1), належить до Cdk, сімейства серин/треонінових протеїнкіназ, та відіграє ключову роль у контролі клітинного циклу. Він відомий як основний регулятор переходу від G2 до фази M. У кінці інтерфази він активується шляхом зв'язування з циклінами A-типу та сприяє початку мітозу. Після руйнування ядерної оболонки цикліни A-типу р та замінюються цикліном B. Комплекс між CDC2 та цикліном B утворює фактор стимуляції мітозу (MPF), котрий є суттєвим для процесу мітозу клітин. Активний CDC2 фосфорилює більш ніж 70 субстратів. Наприклад, фосфорилювання "лінкерного" гістону H1 призводить до послаблення хроматинової структури і транскрипції специфічних генів; фосфорилювання РНК-полімерази II посилює транскрипцію (Cisek і Corden, 1989). Фосфорилювання BRCA2 стимулює репарацію за участю гомологічної комбінації (Esashi і співавт., 2005), а фосфорилювання FOXO1 інгібує його транскрипційну активність, що сприяє проліферації та виживанню клітин. Фосфорилювання сепарази інгібує передчасне розходження сестринських хроматид (Stemmann і співавт., 2001). Активність CDC2 знов суттєво падає в ході анафази, оскільки комплекс стимуляції анафази/циклосома (APC/C) убіквітинізує циклін B, що веде до його деградації. Функція CDC2 у мітозі не є надлишковою та не може компенсуватися активністю інших Cdk, таких як Cdk2, 4 і 6. Навпаки, є інформація про участь CDC2 і в інших фазах клітинного циклу, наприклад, G1-S-переході, та його здатність замінювати "інтерфазні Cdk". Отже, CDC2 є, ймовірно, єдиною суттєвою Cdk клітинного циклу. Крім експресії та функцій CDC2 у клітинному циклі, є дані про його експресію в деяких випадках в умовах апоптозу, і про те, що посилена активність може призвести до мітотичної катастрофи. Надмірна експресія CDC2 була виявлена у декількох видах ракових пухлин, хоча експресія інших білків клітинного циклу, наприклад, циклінів, регулюється з порушеннями навіть частіше. До видів раку з надмірною експресією СDC2 відносяться карциноми передміхурової залози, карциноми ротової порожнини, плоскоклітинна карцинома ротової порожнини (OSCC), гостра мієлоїдна лейкемія (AML) (Qian і співавт., 2009), лімфома MALT-типу, спричинена H.pylori (Banerjee і співавт., 2000), та карцинома товстої кишки (Yasui і співавт., 1993). У кількох випадках надмірна експресія корелювала з несприятливим прогнозом. Щодо раку шлунка (РШ), повідомлялося про надмірну експресію та/або збільшену активність (в 14 з 23 випадків), і є припущення, що надмірна експресія CDC2 може бути причиною цього явища. Крім того, була підтверджена наявність CDC2 у наборі генів, активних під час мітозу, котрі при надмірній експресії ведуть до хромосомної нестабільності пухлин. Інгібітори CDC2 та інших Cdk вважаються кандидатами у лікарський засіб для лікування раку (Shapiro, 2006). Білок, асоційований з аномальною веретеноподібною мікроцефалією (ASPM) Аномальний ген білка веретена поділу, що асоціюється з мікроцефалією (ASPM), - це ортолог аномального веретена Drosophila (asp) у людини, та він є зазвичай мутованим геном 14 UA 109891 C2 5 10 15 20 25 30 35 40 45 50 55 60 аутосомно-рецесивної первинної мікроцефалії. Дефектний нейрогенез, спричинений гомозиготною мутацією ASPM у людей, призводить до мікроцефалії та затримки психічного розвитку. Найбільш частою причиною первинної аутосомно-рецесивної мікроцефалії (MCPH), як вважають, є мутації в гені ASPM, який бере участь у регуляції нейрогенезу. ASPM локалізується на полюсах веретена поділу під час мітозу. Інгібування ASPM за допомогою siРНК-опосередкованого нокдауну уповільнює проліферацію пухлинних клітин і нейрональних стовбурових клітин, підтверджуючи роль ASPM як потенційної молекулярної мішені у гліобластомі. Повідомлялося про надмірну експресію ASPM у гліобластомі у порівнянні з мозком у нормальному стані. Експресія ASPM може бути використана як маркер злоякісності гліоми та являє собою потенційну терапевтичну мішень. Надмірна експресія ASPM є молекулярним маркером, що може спрогнозувати збільшений інвазивний/метастатичний потенціал гепато-клітинної карциноми ГКК, підвищений ризик раннього рецидиву пухлини (ETR) незалежно від статусу мутації p53 і стадії пухлини, і, таким чином, несприятливий прогноз. Також продемонстрована підвищена експресія ASPM в іморталізованих клітинах, ракових клітинах і в тканині пухлини у випадку немілкоклітинного раку легенів (NSCLC) (Jung і співавт., 2009). Убіквітин карбоксил-термінальна гідролаза L5 (UCHL5) Убіквітин карбоксил-термінальна гідролаза L5 (UCHL5), також відома як Убіквітин Cтермінальна гідролаза (UCH37) чи INO80R, є деубіквітиназою, що асоціюється з протеасомою. Вона роз'єднує прикріплені до білка поліубіквітинові ланцюги з дистального кінця, розщеплюючи ізопептидний зв'язок між C-термінальними Cys76 та Lys48 (Nishio і співавт., 2009). UCHL5 зв'язується з hRpn13 (також відомим як Adrm1), компонентом 19S (PA700) регуляторної частинки протеасоми, що ініціює її ізопептидазну активність. hRpn13 також функціонує як рецептор убіквітину, таким чином зв'язуючи розпізнавання субстрату з деубіквітинізацією. UCHL5 також може зв'язуватися з Rpn10/S5a, іншим компонентом частинки 19S, що знаходиться між її "кришкою" та "основою", але така взаємодія не може активувати UCHL5. UCH37 може захищати від протеолізу субстрати, що погано убіквітинізуються, або інші кон'югати убіквітину, котрі повільно розщеплюються. Як альтернатива, вона також може посилювати деградацію, сприяючи вивільненню поліубіквітинованих субстратів з початкового сайту зв'язування в регуляторному комплексі 19S для транслокації у протеолітичне ядро частинку 20S протеасоми 26S. У ядрі UCHL5 також асоціюється з хроматин-ремоделюючим комплексом Ino80. Після зв'язування протеасоми вона стає активованою і може сприяти регуляції транскрипції чи репарації ДНК, що вважається опосередкованою Ino80 і протеасомою. Функція UCHL5 могла б, принаймні частково, виконуватися іншими білками, оскільки RNAiопосередкований нокдаун UCHL5 не має помітного впливу на ріст клітин, протеасомну структуру чи протеолітичну здатність, хоча й прискорює деградацію клітинного білка. Убіквітин-специфічні протеази, такі як UCHL5, беруть участь у декількох процесах, наприклад, у контролі прогресії клітинного циклу, диференціації клітин, реплікації та репарації ДНК, транскрипції, контролі якості білків, імунній відповіді та апоптозі. Є, принаймні, деякі підтвердження того, що UCHL5 сприяє злоякісній трансформації. Було продемонстровано, що її активність збільшується у тканині карциноми шийки матки у жінок, у порівнянні із прилеглими нормальними тканинами. Вона здатна деубіквітинувати і, як наслідок, стабілізувати TGF-бетарецептор та його медіатори по низхідній, Smad, таким чином стимулюючи сигнальні каскади, які активує TGF-бета. Активація сигнального шляху за участю TGF-бета, може бути промотором пухлин на пізніх стадіях розвитку раку, хоча він має подвійну функцію і також може бути супресором пухлин на ранніх стадіях та перед ініціацією (Wicks і співавт., 2005; Wicks і співавт., 2006; Horton і співавт., 2007; Bierie і Moses, 2006). c-Met Див. приклади у EP 08008292.8 та EP1507795B1. Крім того, конститутивна активація c-Met шляхом фосфорилювання також виявилася важливим механізмом онкогенезу у випадку світлоклітинної карциноми нирки у людей (Nakaigawa і співавт., 2006). Надмірна експресія MET, що часто індукується гіпоксією пухлини, призводить до конститутивної активації рецептору і корелює з несприятливим прогнозом. Відключення ендогенного гена MET, надмірно експресованого у пухлинних клітинах, призводить до порушення виконання повної програми інвазивного росту in vitro, відсутності росту пухлини та зменшеному утворенню метастазів в експерименті in vivo. Заслуговує на увагу, що відключення MET у вже наявних метастазах веде до їхньої майже повної регресії (Corso і співавт., 2008). Білковий рецептор стимуляції макрофагів (MST1R) Рецептор MST1R (також відомий під назвою RON) є членом сімейства Met рецепторних 15 UA 109891 C2 5 10 15 20 25 30 35 40 45 50 55 60 тирозинкіназ поверхні клітин. Переважно, він експресується на епітеліальних клітинах та макрофагах. Подібно c-MET, RON експресується різними пухлинами, що походять з епітелію, та лініями ракових клітин і вважається таким, що відіграє функціональну роль в онкогенезі. Клінічні дослідження показали, що надмірна експресія MST1R асоціюється з гіршими результатами для пацієнта, а також з розвитком метастазів. Експресія MST1R є значною у тканині карциноми шлунка та відповідній паранеопластичній тканині, але він не експресується у нормальній слизовій оболонці шлунка (Zhou і співавт., 2008). Рецептор MST1R може індукувати міграцію, інвазію, проліферацію та виживання клітин під впливом відповідного ліганду. Крім того, MST1R має онкогенну активність in vitro, у тваринних моделях in vivo та часто дерегульований у ракових пухлинах у людей (Dussault і Bellon, 2009). Дані підтверджують, що "нокдаун" MST1R у клітинах пухлини передміхурової залози призводить до значно нижчого хемотаксису ендотеліальних клітин, у порівнянні з MST1R-експресуючими клітинами in vitro, а також меншого росту пухлини і зниженій щільності мікросудин після ортотопічної трансплантації у передміхурову залозу in vivo. Продемонстровано, що нокдаун MST1R-кінази, опосередкований РНК-інтерференцією, у високо-онкогенній клітинній лінії раку товстої кишки є причиною зниження проліферації у порівнянні з контрольними клітинами. Білки, що підтримують структуру хромосом (SMC4) Білки, що підтримують структуру хромосом (SMC), є хромосомними ATP-азами, висококонсервативними від бактерій до людини, які грають основні ролі у багатьох аспектах підтримки структури вищого порядку в хромосомах та їхньої динаміки. Білок SMC4 є ключовим компонентом конденсинового комплексу, який відіграє роль у конденсації хроматину, а також асоціюється із сегрегацією ядерець, ДНК-репарацією та утриманням хроматинового каркасу. Еукаріоти мають принаймні шість SMC-білків у індивідуальних організмах і формують три окремі гетеродимери зі спеціалізованими функціями: SMC2 та SMC4 функціонують як ядро конденсинових комплексів, що є суттєвим для збирання хромосом та їхньої сегрегації. Аналіз рівнів мРНК у 25 різних нормальних тканинах за допомогою ЗТ-ПЛР (зворотної транскрипції - полімеразної ланцюгової реакції) показує, що цей ген має високу експресію у нормальній передміхуровій залозі та слинній залозі, дуже слабку експресію у товстій кишці, підшлунковій залозі та кишковому тракті та взагалі не експресується в інших тканинах. Дослідження методом ЗТ-ПЛР, проведене на зразках ракових пухлин у людини, показують, що РНК в значній мірі експресується в багатьох лініях ракових клітин і зразках тканини ракових пухлин, у тому числі, 26 з 33 зразків ракових пухлин молочної залози у людини, 3 з 3 зразків ракових пухлин передміхурової залози, 3 з 3 зразків ракових пухлин товстої кишки і 3 з 3 зразків ракових пухлин підшлункової залози (Egland і співавт., 2006). AVL9 Дивним є той факт, що цей білок виявився вихідним білком, оскільки дуже мало даних є в наявності про білок AVL9 та функціонування відповідного гена. Білок кінетохора Nuf2 Ген NUF2 (CDCA-1) кодує білок, який є в значній мірі подібним до Nuf2 дріжджів, компоненту консервативного білкового комплексу, асоційованого з центромерою. Nuf2 дріжджів зникає з центромери в ході профази мейозу, коли центромери втрачають свій зв'язок із полярними тільцями веретена поділу, та відіграє регуляторну роль у сегрегації хромосом. Було продемонстровано, що csiРНК сурвівіну та hNuf2 здійснюють тимчасовий нокдаун їхніх мРНК, що призводить до утворення багатоядерних клітин та смерті клітин внаслідок мітотичного блоку, відповідно (Nguyen і співавт., 2006). Nuf2 і Hec1 потребуються для організації стабільних сайтів зв'язування плюсового кінця мікротрубочок на зовнішній пластині, котрі необхідні для постійної дії сил у напрямку полюсів, для біоорієнтації на кінетохорах (DeLuca і співавт., 2005). Імуногістохімічний аналіз із використанням мікроматриці тканини раку легенів підтвердив високі рівні білків CDCA1 та KNTC2 у значній більшості пухлин легенів різних гістологічних типів. Підвищена експресія цих білків асоціювалася із гіршим прогнозом для пацієнтів, хворих на немілкоклітинний рак легенів (NSCLC). Інгібування їхнього зв'язування клітинно-проникним пептидом, що несе отриманий із CDCA1 пептид із 19 амінокислот (11R-CDCA1(398-416)), який відповідає домену зв'язування з KNTC2, ефективно пригнічував ріст клітин NSCLC (Hayama і співавт., 2006). Опосередкований siРНК "нокдаун" CDCA1 або KNTC2, як виявилося, інгібує проліферацію клітин та індукція апоптозу у випадках NSCLC, раку яєчників, раку шийки матки, раку шлунка, колоректального раку та гліоми (Kaneko і співавт., 2009). Ген CDCA 1 диференційно експресується в клітинах раку шийки матки (експресія мРНК підтверджена ПЛР в реальному часі та білком - в ході імуногістохімічного аналізу) (Martin і співавт., 2009). Дані ЗТПЛР для тканин пухлин шлунка після хірургічної резекції (дифузний тип, 6; кишковий тип, 4) 16 UA 109891 C2 5 10 15 20 25 30 35 40 45 50 55 60 підтвердили, що два варіанти CDCA1 мали підвищену експресію у тканинах пухлин. В цьому дослідженні були визначені варіанти альтернативного сплайсингу, особливо у CDCA1, які можуть бути потенційно корисними як діагностичні маркери та/або нові мішені для протипухлинної терапії (Ohnuma і співавт., 2009). Ліпідна фосфат-фосфогідролаза 2 (PPAP2C) Білок, кодований цим геном, є членом сімейства фосфатаз фосфатидної кислоти (PAP). PAP перетворюють фосфатидну кислоту у діацилгліцерин та приймають участь у синтезі гліцероліпідів de novo, а також в активованій рецепторами передачі сигналів, опосередкованих фосфоліпазою D. Повідомлялося про три транскрипційних варіанти альтернативного сплайсингу, що кодують різні ізоформи. PPAP2C є потенційно новою мішенню, що присутня у підвищеній кількості у трансформованих первинних мезенхімальних стовбурових клітинах (MSC) у дорослих людей. "Нокдаун" PPAP2C знижує проліферацію клітин шляхом затримки їхнього входження у S-фазу клітинного циклу і транскрипційно регулюється за допомогою p53. Деякі дані свідчать про те, що надмірна експресія PPAP2C, яка спостерігається у різних видах раку у людини, може бути необхідною для підвищеної проліферації клітин (Flanagan і співавт., 2009). Дослідження демонструє, що PPAP2C є регулятором прогресії клітинного циклу у фібробластах. Надмірна експресія PPAP2C, на відміну від каталітично неактивного мутанту, викликала раннє входження до S-фази, що супроводжується передчасним накопиченням цикліну A. Це вказує на значні зміни у швидкості входження у S-фазу, що може впливати на такі процеси, як мітогенез, міграція, загоєння ран, зростання та онкогенез. Убіхінон-зв'язувальний білок комплексу убіхінол-цитохром-c-оксиредуктаза (UQCRB) Ген UQCRB кодує білок, що є частиною комплексу убіхінол-цитохром c-оксидоредуктази, який вміщує десять субодиниць, кодованих ядерною, та одну субодиницю - кодовану мітохондріальною ДНК. Кодований білок зв'язує убіхінон та бере участь у переносі електронів, коли убіхінон є зв'язаним. Мутації в цьому гені асоціюються з недостатністю мітохондріального комплексу III. Був описаний псевдоген на X-хромосомі. Ген UQCRB може бути потенційним онкогеном чи геном-супресором пухлин у протоковій аденокарциномі підшлункової залози (Harada і співавт., 2009). Ген UQCRB надмірно експресується у гепато-клітинній карциномі (Jia і співавт., 2007). Промінін 1 (Prom1) Промінін-1, також відомий під назвою CD133, був спочатку ідентифікований як молекула, специфічна до CD34-позитивних кровотворних клітин-попередників. Як було продемонстровано пізніше, він є маркером нормальних стовбурових клітин і ракових стовбурових клітин (CSC) різних тканин (Mizrak і співавт., 2008). Однак мало що відомо про його функцію. Оскільки він переважно локалізується у виступах плазматичної мембрани, таких як мікроворсинки епітеліальних клітин, промініну-1 відводиться функціональна роль "організатора" топології плазматичної мембрани. Оскільки була підтверджена його взаємодія з холестерином, він може бути важливим у підтриманні належного ліпідного складу в межах плазматичної мембрани. Промінін-1 використовується як CSC-маркер у багатьох видах пухлин у людей. Лише невеликий відсоток пухлинних клітин є зазвичай промінін-1-позитивними, як це очікується для CSC-маркера. В залежності від виду пухлини, кількість позитивних клітин на масу пухлини досягає від 1 до 15 % та в більшості випадків становить близько 2 %. Пухлини, в яких промінін1-експресуючі клітини є клітинами CSC, як показано функціональними випробуваннями (наприклад, утворення сфери, висока здатність ініціювати зростання пухлини у імунодефіцитних мишей та асиметричний поділ /самовідновлення/ плюрипотентність), наведені нижче: - рак товстої кишки (2 - 2.5 % маси пухлини) (Todaro і співавт., 2007; Ricci-Vitiani і співавт., 2007), - рак печінки (Ma і співавт., 2007; Suetsugu і співавт., 2006; Yin і співавт., 2007), - рак підшлункової залози (Hermann 2007; Wang 2009), - рак передміхурової залози (1 % маси пухлини) (Richardson і співавт., 2004), - пухлини мозку різних фенотипів (Singh і співавт., 2003; Singh і співавт., 2004), - лейкемія, наприклад, гостра лімфобластна лейкемія, ALL (Cox і співавт., 2009), - меланома (Monzani і співавт., 2007; Rappa і співавт., 2008), - рак легенів (Chen і O'Shea, 2008; Eramo і співавт., 2008; Tirino і співавт., 2009), - саркома Юїнга (Suva і співавт., 2009), - рак ендометрію (Rutella і співавт., 2009), - плоскоклітинна карцинома ротової порожнини (Zhang і співавт., 2009) і - плоскоклітинна карцинома голови та шиї (Harper і співавт., 2007). Крім того, декілька досліджень показали підвищену експресію промініну-1 у раковій тканині у 17 UA 109891 C2 5 10 15 20 25 30 35 40 45 50 55 60 порівнянні із здоровою тканиною, і в більшості з них знайдено кореляцію експресії промініну-1 з клінічними параметрами, такими як загальна виживаність, стадія пухлини або метастазування. Прикладами є немілкоклітинний рак легенів, злоякісна меланома, ретинобластома, нейробластома та синовіальна карцинома. Експресія промініну-1 також корелює з несприятливим прогнозом у випадках гліоми, рак підшлункової залози (до 15 % PROM1позитивних клітин), колоректального, ректального раку, раку товстої кишки і протокової карциноми молочної залози. Цікавим є той факт, що мРНК PROM1 має підвищену експресію в PBMC (мононуклеарах периферичної крові) у хворих на рак з метастазуючими пухлинами, особливо у пацієнтів з метастазами у кістках, та експресія PROM1 в PBMC є прогностичним фактором щодо загальної виживаності. Ніякої кореляції з прогнозом не було визначено у випадку раку яєчників. У випадку дифузного раку шлунка, було висловлене припущення про експресію PROM1 на основі аналізу in silico (Katoh і Katoh, 2007). Про його надмірну експресію у раковій пухлині шлунка, в порівнянні з нормальною тканиною шлунка на рівні білка говорилося у роботі (Smith і співавт., 2008). Однак автори (Boegl і Prinz, 2009) повідомили про те, що експресія промініну-1 була зниженою у випадку раку шлунка, зокрема, на пізніших стадіях, і заявили, що експресія промініну-1 скоріше корелює з ангіогенезом - також зниженим на більш пізніх стадіях - ніж з ростом пухлини. У дослідженні з використанням клітинних ліній раку шлунка (Takaishi і співавт., 2009) говорять про те, що CD44, а не промінін-1, є маркером CSC у випадку раку шлунка. Підтвердження про участь клітин, що експресують промінін-1, в утворенні пухлин, було надане у роботі (Zhu і співавт., 2009). Автори роботи повідомили, що у моделі раку кишечника у мишей усі неопластичні клітини походили з Prom1-позитивних клітин, але тільки 7 % зберегли Prom1-позитивний фенотип. В інших аспектах, було показано, що промінін-1-позитивні клітини сприяють ангіогенезу пухлин. Як і очікувалося для CSC, визначено, що промінін-1-позитивні клітини є хіміорезистентними завдяки активації антиапоптозного Akt-каскаду (Ma 2008). Інші автори (Bertolini і співавт., 2009) повідомляють, що ці клітини не реагують на лікування цисплатином. Вони стійкі до TRAIL- і Fas-індукованого апоптозу внаслідок підвищеної експресії FLIP. Вони захищають себе від апоптозу за допомогою секреції ІЛ-4. Однак вони можуть бути доступними для імунної системи, оскільки можливе їхнє знищення NK-клітинами (Castriconi і співавт., 2007; Pietra і співавт., 2009) і цитотоксичними T-клітинами (Brown і співавт., 2009). Матрична металопротеїназа 11 (MMP11) Вважається, що матрична металопротеїназа 11 (MMP11) відіграє роль у кількох фізіологічних процесах, що потребують ремоделювання тканин, наприклад, розвиток, інволюція молочної залози після лактації, загоєння ран і формування рубців, а також протягом менструального циклу. Є припущення щодо того, що вона негативно регулює жировий гомеостаз, знижуючи диференціацію адипоцитів. На відміну від інших MMP, вона не здатна розщеплювати типові молекули позаклітинного матриксу - крім колагену VI. Однак були ідентифіковані інші субстрати, такі як альфа-2-макроглобулін, окремі інгібітори серинпротеази (серпіни), у тому числі, альфа-1-антитрипсин, білок-1, що зв'язує інсуліноподібний фактор росту, та рецептор ламініну. MMP11 була виявлена як продукт, що надмірно специфічно експресується у стромальних клітинах, які оточують інвазивну карциному молочної залози. Подальші дослідження підтвердили її експресію у стромі, що оточує карциному молочної залози та інші види ракових пухлин, наприклад, у випадках раку шкіри, немілкоклітинного та мілкоклітинного раку легенів, плоскоклітинної карциноми голови та шиї, карциноми товстої кишки і колоректальної карциноми, епітеліального раку гортані, раку стравоходу, раку ротової порожнини, раку підшлункової залози, раку сечового міхура, карцином яєчників, нирково-клітинної карциноми, атипової менінгіоми, папілярного раку щитовидної залози, пухлин мозку (MMP11 експресувалася в астроцитомах, але тільки в незначній мірі в олігодендрогліомах та у злоякісних гліомах), раку слинних протоків, раку шийки матки, екстранодальних T/NK-клітинних лімфом, неходжкінської лімфоми і раку передміхурової залози. Було зазначено, що MMP11 надмірно експресується у стромі найбільш інвазивних карцином у людей, але рідко у саркомах та інших неепітеліальних пухлинах. Переважно MMP11 експресується у клітинах строми, безпосередньо прилеглих до пухлини, в той час як самі пухлинні клітини, нормальні тканини та клітини строми, віддалені від пухлини, є негативними. Однак це не можна узагальнювати, тому що в деяких випадках MMP11 також була виявлена у нераковій тканині, такій як тканина товстої кишки, або у пухлинних клітинах, наприклад, у пухлинах підшлункової залози, молочної залози, павутинної оболонки мозку та шлунка. Підвищені рівні MMP11 корелюють із злоякісним фенотипом / підвищеною інвазивністю та несприятливим прогнозом. Однак, у випадках папілярної карциноми щитовидної залози експресія MMP11 має зворотний зв'язок із характеристиками агресивності. 18 UA 109891 C2 5 10 15 20 25 30 35 40 45 50 55 60 Її участь в ангіогенезі є малоймовірною, тому що експресія MMP11 не корелює зі щільністю мікросудин. Скоріш за все, вона посилює виживання ракових клітин і пригнічує апоптоз. Було зроблене припущення, що MMP11 з фібробластів веде до стимуляції IGF-1R сигнального шляху у клітинах карциноми, таким чином збільшуючи їхню проліферативну здатність. Здатність MMP11 призводити до диференціації адипоцитів стимулює рак завдяки акумуляції перитуморальних фібробласто-подібних клітин, що сприяють виживанню ракових клітин і прогресії пухлин (Motrescu і Rio, 2008). MMP11 була виявлена у пухлинній тканині, а також сироватці пацієнтів, хворих на рак шлунка; її експресія корелювала із розвитком метастазів (Yang і співавт., 2008). Крім того, автори (Deng і співавт., 2005) показали високу експресію MMP11 у лініях пухлинних клітин та первинній пухлині раку шлунка - на відміну від інших видів раку, не тільки у стромі, - і що вона, очевидно, посилює проліферацію клітин. ABL1 Протоонкоген ABL1 кодує цитоплазматичну та ядерну протеїнтирозинкіназу сімейства Src, що є важливими у процесах диференціації, поділу, адгезії клітин та стресовій реакції (Yoshida, 2007). C-Abl курсує меж ядерним та цитоплазматичним компартментами. Ядерний c-Abl бере участь в інгібуванні росту клітин та сприянні апоптозу. На відміну від цього, роль цитоплазматичного c-Abl описана менш детально. Існують припущення про його роль у морфогенезі та динаміці F-актину, а також роль у передачі сигналів, індукованих позаклітинними стимулами, такими як фактори росту і ліганди інтегринів. Є інформація про те, що цитоплазматичний c-Abl сприяє мітогенезу. Мітогенні субстрати C-Abl ще не були ідентифіковані, але вони, ймовірно, включатимуть регулятори малих ГТФаз сімейства Rho, зокрема, члени Vav та Sos. ДНК-зв'язувальна активність повсюдно експресованої ABL1-тирозинкінази регулюється опосередкованим CDC2 фосфорилюванням, що може означати участь ABL1 у клітинному циклі. Активність білка c-Abl негативно регулюється його доменом SH3, а делеція домену SH3 перетворює ABL1 на онкоген. Нерецепторна тирозинкіназа c-Abl регулює реакції актину у некровотворних клітинах. В деяких дослідженнях c-Abl ідентифікується як ключовий учасник сигнального каскаду, що приводить до реорганізації актину в ході активації T-клітин (Huang і співавт., 2008). Мутації у гені ABL1 асоціюються з хронічною мієлоїдною лейкемією (ХМЛ). При ХМЛ ген активується шляхом транслокації в межах гена BCR (ділянки локалізації сайту ініціації реаранжування) на хромосомі 22. Цей новий злитий ген BCR-ABL кодує нерегульовану тирозинкіназу, націлену на цитоплазму, що сприяє проліферації клітин без регуляції цитокінами. Це, в свою чергу, дозволяє клітинам ставати раковими (Zhao і співавт., 2009). Активована c-Abl тирозинкіназа, не як злитий білок, грає важливу роль у злоякісних солідних пухлинах легенів та молочної залози (Lin і Arlinghaus, 2008). Недавні дослідження вказують на те, що c-Abl також дерегулюється у солідних пухлинах. Висока активність цитоплазматичної кінази була визначена у карциномах молочної залози та NSCLC. Надмірна експресія, однак, не є достатньою, і для конститутивної кіназної активності є необхідним фосфорилювання білка. У клітинах ракової пухлини молочної залози фосфорилювання c-Abl індукується тирозинкіназами плазматичної мембрани, у тому числі, членами сімейств SFK, EGFR та рецептором IGF-1. Злиті білки ABL не були виявлені у солідних пухлинах. ABL1 та рак шлунка - В імуногістохімічному дослідженні експресії ABL1 був вивчений широкий діапазон нормальних ембріональних тканин і тканин дорослих людей, а також різні види пухлин. Більшість пухлин продемонструвала фокальну чи слабку імунореактивність ABL. Найбільш інтенсивне забарвлення спостерігалося у хондросаркомі, ліпосаркомі та дифузній (перснеподібній) аденокарциномі шлунка. У двох останніх випадках ABL також експресувався на мікросудинах пухлин, що вказує на його можливу роль в ангіогенезі. Останні дослідження виявили, що інфікування cagA-позитивними Helicobacter pylori відіграє суттєву роль у розвитку карциноми шлунка. H. pylori блокує ендоцитоз EGFR та деградацію рецепторів після тривалого інфікування епітеліальних клітин шлунка. Більше того, це інгібування відбувається шляхом залежної від CagA, але не залежної від фосфорилювання CagA, активації нерецепторної кінази c-Abl, яка, в свою чергу, фосфорилює сайт-мішень EGFRpY1173 (Bauer і співавт., 2009). Селективне інгібування c-Abl-кіназної активності за допомогою STI571 чи sh-РНК перериває стабільне фосфорилювання цитотоксин-асоційованого гена A (CagA) та міграцію епітеліальних клітин. Це вказує на ключову роль c-Abl в інфікуванні H. pylori та патогенності (Poppe і співавт., 2007). Прикладом блокаторів кіназного шляху є іматиніб (іматинібу мезилат, Gleevec, STI571), інгібітор онкобілка Bcr/Abl, який став медикаментом першої лінії при лікуванні хронічної 19 UA 109891 C2 5 10 15 20 25 30 35 40 45 50 55 60 мієлоїдної лейкемії (ХМЛ) (Pytel і співавт., 2009). Іматиніб був ухвалений для лікування пацієнтів із стромальними пухлинами шлунково-кишкового тракту (GIST) на пізніх стадіях, в яких аномально експресується KIT, рецептор тирозинкінази (Croom і Perry, 2003). Ще один інгібітор кінази, який останнім часом використовується при лікуванні раку, - це дазатиніб (BMS-354825), специфічний до нерецепторного цитоплазматичного ABL (Pytel і співавт., 2009). Нілотиніб - це тирозинкіназний інгібітор bcr-abl другого покоління для перорального застосування, показаний для лікування Ph-позитивної ХМЛ у хронічній фазі чи фазі акселерації у дорослих людей, коли спостерігається резистентність до іматинібу чи його непереносимість (Deremer і співавт., 2008). SEQ ID NO 13, SEQ ID NO 14 та SEQ ID NO 15 розкриті в патенті WO 2007/028574, CDC42 (цикл клітинного поділу 42) - це білок, який бере участь в регуляції клітинного циклу. Білок є малою ГТФазою підсімейства Rho, що регулює сигнальні шляхи, які контролюють різні клітинні функції, включаючи морфологію, міграцію, ендоцитоз клітин та хід клітинного циклу. Був визначений дуже високий рівень надмірної експресії CDC42 в гліобластомі. В патенті WO 2004/067023 описуються пептиди, рестриктовані за MHC класу I, що були одержані з пухлино-асоційованого антигену сурвівіну, пептиди якого мають здатність зв'язуватися з молекулами HLA класу І з високою афінністю. Секретований фосфопротеїн 1 (SPP1), також відомий як кістковий сіалопротеїн I (BSP-1), ранній активатор Т-лімфоцитів (ETA-1) та найчастіше відомий під назвою "остеопонтин" (OPN), є генним продуктом на основі генів людини, котрий також зберігає свої функції у інших видів. Остеопонтин вважається важливим фактором в ремоделюванні кісток. Зокрема, дослідження підказують, що він відіграє роль в закріпленні остеокластів на мінеральному матриксі кістної тканини. Органічна частина кістки становить близько 20 % сухої ваги та включає, крім остеопонтину, колаген типу I, остеокальцин, остеонектин, кістковий сіалопротеїн та лужну фосфатазу. Колаген типу I становить 90 % маси білка. OPN зв'язується з кількома інтегриновими рецепторами, включаючи α4β1, α9β1 та α9β4, що експресуються лейкоцитами. Ці рецептори добре відомі відносно їхніх функцій в адгезії, міграції клітин та виживання в цих клітинах. Отже, програми останніх досліджень зосереджуються на ролі OPN як посередника в цих відповідях. Остеопонтин експресується в ряді імунних клітин, включаючи макрофаги, нейтрофіли, дендритні клітини, T- та B-клітини, з варіабельною кінетикою. Є інформація про те, що OPN різними способами діє як імунний модулятор. По-перше, він має хемотактичні властивості, які сприяють рекрутингу клітин до запальних ділянок. Він також функціонує як адгезивний білок, який бере участь у прикріпленні клітин та загоєнні ран. До того ж, OPN опосередковує активацію клітин та продукування цитокінів, а також сприяє виживанню клітин шляхом регуляції апоптозу. Активовані Т-клітини диференціюють у Th1-тип під впливом стимуляції з боку ІЛ-12 та продукують цитокіни, включаючи ІЛ-12 та IFNγ. OPN інгібує продукування Th2 цитокіну ІЛ-10, що призводить до посиленої відповіді Th1. OPN впливає на клітинно-опосередковану імунність та має Th1- цитокінні функції. Він збільшує продукування В-клітинами імуноглобулінів та їх проліферацію. Недавні дослідження, проведені в 2008, наводять на думку, що OPN також індукує дегрануляцію мастоцитів. [Nagasaka A, Matsue H, Matsushima H, et al. (February 2008). "Osteopontin is produced by mast cells and affects IgE-mediated degranulation and migration of mast cells". Eur. J. Immunol. 38 (2): 489-99]. Вчені виявили, що IgE-опосередкована анафілаксія була значно знижена у OPN-нокаутованих мишей в порівнянні з мишами дикого типу. Роль OPN в активації макрофагів також була визначена в дослідженні раку, коли вчені виявили, що OPNпродукуючі пухлини здатні індукувати активацію макрофагів в порівнянні з пухлинами з нестачею OPN. OPN є важливим антиапоптотичним фактором в багатьох обставинах. OPN блокує індуковану активацією загибель макрофагів і Т-клітин, а також фібробластів та ендотеліальних клітин, що піддаються впливу шкідливих факторів. OPN перешкоджає незапрограмованій загибелі клітин при запальному коліті. Той факт, що OPN взаємодіє з великою кількістю рецепторів на поверхні клітин, і ці рецептори експресовані повсюдно, робить його активним учасником багатьох фізіологічних і патологічних процесів, включаючи загоєння ран, ремоделювання кісток, онкогенез, запалення, ішемію та імунні відповіді. Отже, маніпулювання рівнями OPN в плазмі може бути корисним в лікуванні аутоімунних захворювань, метастазів раку, остеопорозу та деяких форм стресу. Було показано, що OPN стимулює продукування ІЛ-17; OPN надмірно експресується в різних видах ракових захворювань, включаючи рак легенів, рак молочної залози, колоректальний рак, рак шлунка, рак яєчників, меланому та мезотеліому; OPN сприяє як гломерулонефриту, так і тубулоінтерстиціальному нефриту; крім того, OPN виявлений в атеросклеротичних бляшках в артеріях. Таким чином, маніпулювання рівнями OPN в плазмі може бути корисним в лікуванні 20 UA 109891 C2 5 10 15 20 25 30 35 40 45 50 55 60 аутоімунних захворювань, метастазів раку, остеопорозу та деяких форм стресу. Рецептор 3 епідермального фактору росту у людини (ERBB3) ERBB3 кодує одного з членів сімейства рецепторів епідермального фактору росту (EGFR) рецепторних тирозинкіназ. Він активується нейрегулінами, іншими ERBB та рецепторами, що відрізняються від ERBB, а також іншими кіназами і за допомогою нових механізмів. По низхідній він у значній мірі взаємодіє з фосфоінозитол-3-кіназою/АKT-сигнальним каскадом, що визначає виживання клітин/мітогенним каскадом, але також із GRB, SHC, SRC, ABL, rasGAP, SYK і регулятором транскрипції EBP1 (Sithanandam і Anderson 413-48). Дослідження експресії ERBB3 у первинних ракових пухлинах та механізмів його участі в культурах клітин підтвердили його причетність, з різними ступенями визначеності, до спричинення або підтримання ракових захворювань молочної залози, яєчників, передміхурової залози, окремих клітин мозку, сітківки, меланоцитів, товстої кишки, підшлункової залози, шлунка, ротової порожнини і легенів (Sithanandam і Anderson 413-48). Білок ERBB3 був виявлений за допомогою імуногістохімічного аналізу в епітеліальних клітинах в усьому шлунково-кишковому тракті, включаючи плаский епітелій ротової частини глотки та стравоходу, парієтальні клітини шлунка та поверхневі ентероцити тонкої і товстої кишки. ERBB3 продемонстрував підвищену експресію у ракових пухлинах шлунка (Poller і співавт. 275-80; Sanidas і співавт. 935-40). Усі клітинні лінії ракових пухлин шлунка експресували ERBB3 та усічений, секретований продукт. Переконливі докази ключової ролі ERBB3 у злоякісності пухлин шлунка були одержані в дослідженні слабко диференційованих персневидно-клітинних карциномах шлунка (Kobayashi і співавт. 1294-301). Zhang і співавт. вивчали експресію ERBB3 при ракових захворюваннях шлунка двох типів патології (кишковий тип і дифузний тип) з використанням імуногістохімічного аналізу (IHC). Дифузний тип РШ демонстрував значно більшій ступінь надмірної експресії ERBB3 в порівнянні з кишковим типом (26,2 % проти 5,0 %, p < 0,01). Селективна надмірна експресія ERBB3 у двох гістологічних типах раку шлунка була тісно пов'язана з несприятливим прогнозом (Zhang і співавт. 2112-18). Експресія ERBB3 в значній мірі асоціювалася з параметрами, пов'язаними з прогресією пухлини, включаючи глибину інвазії пухлини, ураження лімфовузлів, віддалені метастази, стадію пухлини і рецидиви (Hayashi і співавт. 7843-49). Була вивчена експресія та сумісна експресія EGFR, c-erbB-2 та cerbB-3 у 21 випадку раку шлунка та 20 випадках хронічного гастриту з використанням імуногістохімічного аналізу на свіжезаморожених тканинах, із врахуванням клініко-патологічних показників. В цілому, у хворих на рак шлунка частіше спостерігалась надмірна експресія EGFR, c-erbB-2 та d-erbB-3, ніж у групі з хронічним гастритом (81 % та 43 %; 38 % та 45 %; 35 % та 20 %, відповідно), однак статистично значима різниця була визначена тільки стосовно експресії EGFR (p=0,01) (Slesak і співавт. 2727-32). Були експериментально опробувані декілька терапевтичних підходів з націлюванням на ERBB3. РНК-аптамери до позаклітинного домену ERBB3 інгібували NRG-індуковану гетеродимерізацію ERBB3/ERBB2, фосфорилювання ERBB2 та ріст клітин ракової пухлини молочної залози MCF7 (Chen і співавт. 9226-31). Синтетичний транскрипційний фактор із доменом типу "цинкові пальці", що інгібує експресію гена ERBB3 в клітинах плоскоклітинної карциноми A431, призвів до зниженої проліферації та міграції; пригнічування експресії ERBB3 мало більший вплив, ніж зміна ERBB2 (Lund і співавт. 9082-91). Ізомер вітаміну E γ-токотриєнол уповільнив проліферацію клітин молочної залози за допомогою специфічного блокування активації ERBB3 та стимуляції сигнального шляху PI3K/AKT по низхідній (Samant і Sylvester 56374). Мікро-РНК 125a зменшила рівні РНК ERBB3 та білка, активацію AKT і ріст клітин, а також інвазивність клітин карциноми молочної залози SKBR3 (Scott і співавт. 1479-86). Зниження рівня ERBB3 за допомогою siРНК у клітинах раку молочної залози елімінувало їхню вторинну резистентність до тирозинкіназних інгібіторів та надало можливість здійснення індукції апоптозу (Sergina і співавт. 437-41). Мала РНК (siРНК), інгібіторна по відношенню до ERBB3 чи AKT, вважається багатообіцяючою як терапевтичний підхід до лікування аденокарциноми легенів (Sithanandam і співавт. 1847-59). Сурвівін (BIRC5) Експресія BIRC5 (сурвівіну), члена сімейства білків-інгібіторів апоптозу (IAP), підвищена в ембріональних тканинах та в різних видах ракових захворювань людини. У патенті WO 2004/067023 описуються пептиди, рестриктовані за MHC класу I, які походять із пухлиноасоційованого антигену сурвівіну, котрі здатні зв'язуватися з молекулами HLA класу І з високою афінністю. Передбачається, що сурвівін здатний регулювати клітинну проліферацію та загибель апоптотичних клітин. Дуже високі рівні експресії сурвівіну визначаються, зокрема, в гліобластомі (Angileri і співавт., 2008). Існує думка, що надмірна експресія сурвівіну в гліомах головного мозку може відігравати важливу роль в злоякісній проліферації, анти-апоптозі та ангіогенезі (Zhen і 21 UA 109891 C2 5 10 співавт., 2005; Liu і співавт., 2006). Особливо для гліобластоми, але також і для інших пухлинних об'єктів експресія сурвівіну була в значній мірі асоційована зі ступенем злоякісності (з найбільшою експресією сурвівіну в гліобластомі) та більш короткою загальною виживаністю в порівнянні з пацієнтами з сурвівін-негативними пухлинами (Kajiwara і співавт., 2003; Saito і співавт., 2007; Uematsu і співавт., 2005; Mellai і співавт., 2008; Grunda і співавт., 2006; Xie і співавт., 2006; Sasaki і співавт., 2002; Chakravarti і співавт., 2002). Коровий антиген вірусу Гепатиту B Імуногенні пептиди для корового білка HBc вірусу Гепатиту B (HBV) є добре відомими (Bertoletti і співавт., 1993; Livingston і співавт., 1997). Пептид із HBc, що складається із десяти амінокислот, може бути вибраний як позитивний контроль у протираковій вакцини на основі цього винаходу для визначення імунокомпетентності пацієнта та успішної вакцинації. Таблиця 6 Функції вихідних білків, пов'язані з раковими захворюваннями Категорія "-" < "(+)" < "+"; "?" означає, що ситуація на даний час неясна Пептиди TUMAP HLA класу І Функції вихідних білків, пов'язані з ASPM- CDC2- MET- MST1R- PROM1- UCHL5- MMP11- SMC4- PPAP2C- AVL9- UQCRBNUF2раковими ABL1-001 002 001 006 001 001 001 001 001 001 001 001 001 захворюваннями /активність TUMAP Онкофетальний білок (OF) / Раково? CT тестикулярний антиген (CT) Онкогенез (прогресія клітинного + + + + + ? + + ? + + циклу, проліферація) Інвазія пухлини, міграція, + + + + ? + ? + метастазування Шляхи EGFR, передачі PI3K, По Клітин- Ras / TGFсигналів, STAT3,+1, низхід- ний MAPK, Подібно бета IGF1RКлітинподіл пов'язані з Akt мітоз ? Rac/JNK, ній цикл PI3K, до MET (поси- шлях ний цикл клітин раковими Erk5, ЕGFR (G2/M) PLC лений) захворюбетаваннями катенін Антиапопто(+) (+) + + + ? + тичні ефекти Ангіогенез + (+) (+) ? + Ракові клітини, подібні + (+) (+) ? стовбуровим Надмірна експресія ? + + + (+) ? + ? ? ? ? + + в РШ 22 Пептиди TUMAP HLA класу II BIRC5002 OF + Інгібування апоптозу + + + UA 109891 C2 Таблиця 6 Функції вихідних білків, пов'язані з раковими захворюваннями Категорія "-" < "(+)" < "+"; "?" означає, що ситуація на даний час неясна Пептиди TUMAP HLA класу II Пептиди TUMAP HLA класу І Надмірна експресія в інших видах раку Несприятливий прогноз Пізні стадії 5 10 15 20 25 30 35 40 45 + + + + + (+) + + + ? + + + + + + + + + ? + ? ? + + + ? + ? + ? + Як видно з Таблиці 6, досвідчений спеціаліст зможе легко пристосувати композицію цієї заявки до конкретного пацієнту та/або пухлини, та відповідно вибрати пухлино-асоційовані пептиди. У переважному втіленні винаходу фармацевтична композиція вміщує принаймні два пептиди, один з яких включає амінокислотну послідовність відповідно до SEQ ID No 1 та додатково вміщує пептид, який включає амінокислотну послідовність відповідно до SEQ ID No 11. У переважному втіленні винаходу, фармацевтична композиція вміщує принаймні два пептиди, один з яких включає амінокислотну послідовність відповідно до SEQ ID No 1 та амінокислотну послідовність відповідно до SEQ ID No 2 та/або SEQ ID No 11. У переважному втіленні винаходу, фармацевтична композиція вміщує принаймні два пептиди, які включають амінокислотну послідовність відповідно до SEQ ID No 3 та амінокислотну послідовність відповідно до SEQ ID No 2 та/або SEQ ID No 11. У переважному втіленні винаходу, фармацевтична композиція вміщує принаймні два пептиди, що включають амінокислотну послідовність відповідно до SEQ ID No 1 та амінокислотну послідовність відповідно до SEQ ID No 7 і необов'язково SEQ ID No 11. У переважному втіленні винаходу, фармацевтична композиція вміщує принаймні два пептиди, що включають амінокислотну послідовність відповідно до SEQ ID No 2 та амінокислотну послідовність відповідно до SEQ ID No 7 і необов'язково SEQ ID No 11. У переважному втіленні винаходу, фармацевтична композиція вміщує принаймні два пептиди, що включають амінокислотну послідовність відповідно до SEQ ID No 3 та амінокислотну послідовність відповідно до SEQ ID No 7 і необов'язково SEQ ID No 11. В ще більш переважному втіленні фармацевтична композиція вміщує принаймні ще один з пептидів, який включає амінокислотну послідовність, вибрану з групи, що складається з послідовностей від SEQ ID NO 2 до SEQ ID NO 10 та від SEQ ID No 11 до SEQ ID No 22 та SEQ ID No 24 та/або амінокислотну послідовність, котра є принаймні на 85 % ідентичною послідовності від SEQ ID No 2 до SEQ ID No 10 та від SEQ ID No 11 до SEQ ID No 22 та SEQ ID No 24, та/або полінуклеотид, що вміщує нуклеїнову кислоту, яка кодує послідовності від SEQ ID Nо 2 до SEQ ID Nо 10 та від SEQ ID No 11 до SEQ ID No 22 та SEQ ID No 24 чи варіант амінокислотної послідовності, та фармацевтично прийнятний носій. Інші переважні втілення винаходу вміщують принаймні 3, 4, 5, 6, 7, 8, 9, 10, 11 12, 13, 14, 15, 16, 17 або 18 пептидів, які включають амінокислотну послідовність, вибрану з групи, що складається з послідовностей від SEQ ID NO 1 до SEQ ID NO 10 та від SEQ ID No 11 до SEQ ID No 22 та SEQ ID No 24 та/або амінокислотну послідовність, яка є принаймні на 85 % ідентичною послідовності від SEQ ID NO 1 до SEQ ID NO 10 та/або полінуклеотид, що вміщує нуклеїнову кислоту, яка кодує послідовності від SEQ ID NO 1 до SEQ ID NO 10 та від SEQ ID No 11 до SEQ ID No 22 та SEQ ID No 24 чи варіант амінокислотної послідовності, та фармацевтично прийнятний носій. Фармацевтична композиція може додатково вміщувати додаткові пептиди та/або допоміжні речовини для збільшення ефективності, як пояснюється нижче. Під терміном "варіант амінокислотної послідовності" автори винаходу мають на увазі, що бокові ланцюжки, наприклад, одного чи двох амінокислотних залишків змінюються (зокрема, шляхом заміни їх боковим ланцюжком іншого природно існуючого амінокислотного залишку чи якимось іншим боковим ланцюжком) таким чином, що пептид все ще здатний зв'язуватися з 23 UA 109891 C2 5 10 15 20 25 30 35 40 45 50 55 60 молекулою HLA по суті в такий же спосіб, як і пептид, що складається з даної амінокислотної послідовності. Наприклад, пептид може бути модифікований так, що він буде якнайменше зберігати, чи навіть поліпшувати, здатність взаємодіяти та зв'язуватися з прийнятною молекулою MHC, такою як HLA-A або -DR, та якнайменше зберігати, чи навіть поліпшувати, здатність генерувати активовані ЦТЛ, що можуть розпізнавати та знищувати клітини, котрі експресують поліпептид, що вміщує амінокислотну послідовність, як визначено в аспектах винаходу. Як можна дізнатися з бази даних, окремі позиції пептидів, що зв'язуються з HLA-A, є типово якірними залишками, формуючими корову послідовність, яка відповідає зв'язувальному мотиву зв'язувальної порожнини для HLA. Ті амінокислотні залишки, що не суттєвими для взаємодії з T-клітинним рецептором, можуть бути модифіковані шляхом заміни іншою амінокислотою, введення якої не має значного впливу на реактивність Т-клітин та не виключає зв'язування із відповідним МНC. Отже, за винятком зазначеної умови, пептид згідно винаходу може бути будь-яким пептидом (до цього терміну автори винаходу відносять олігопептид чи поліпептид), котрий включає амінокислотні послідовності або їхню частину чи варіант, як наведено у винаході. Крім того, стосовно пептидів, презентованих МНС класу II, відомо, що ці пептиди складаються з "корової послідовності", яка має певний HLA-специфічний амінокислотний мотив та, необов'язково, N- та/або C-термінальні подовження, які не втручаються у функцію корової послідовності (тобто, вважаються нерелевантними до взаємодії пептиду і T-клітини). N- та/або C-термінальні подовження можуть мати довжину, наприклад, від 1 до 10 амінокислот, відповідно. Ці пептиди можуть використовуватись безпосередньо для завантаження MHCмолекул класу II, або послідовність може клонуватись у вектори згідно із наведеним нижче описом. Оскільки ці пептиди формують кінцевий продукт процесингу більш крупних пептидів в межах клітини, то також можуть використовуватися довші пептиди. Пептиди згідно винаходу можуть мати будь-який розмір, але, як правило, вони можуть бути меншими ніж 100 000 за молекулярною масою, переважно, меншими ніж 50 000, ще більш переважно меншими ніж 10 000, більш переважно меншими ніж 5000, більш переважно меншими ніж 2500 та, зазвичай, дорівнюючими приблизно від 1000 до 2000. Стосовно кількості амінокислотних залишків, пептиди згідно винаходу можуть мати менше 1000 залишків, переважно менше ніж 500 залишків, і ще більш переважно - менше 100 залишків. Відповідно, цей винахід також надає композиції пептидів та їхні варіанти, в яких пептид чи варіант має загальну довжину від 8 до 100, переважно від 8 до 30, та найбільш переважно від 8 та 17, а саме, 8, 9, 10, 11, 12, 13, 14, 15 або 16 амінокислот. Переважними є пептиди з коровою послідовністю, вибраною з групи, яка складається з послідовностей від SEQ ID NO 11 до SEQ ID NO 22 і SEQ ID NO 24 з подовженнями від 1 до 10 амінокислот на C-кінці та/або N-кінці, більш переважною є загальна довжина цих бокових амінокислот від 1 до 12, більш переважною - від 1 до 10, більш переважною - від 1 до 8, і більш переважною від 1 до 6, де бокові амінокислоти можуть розподілятися в будь-якому співвідношенні на C-кінці та N-кінці (наприклад, усі бокові амінокислоти можуть додаватися до одного кінця, чи амінокислоти можуть додаватися в рівних кількостях до обох кінців, чи в будьякому іншому співвідношенні), за умови, що пептид зберігає здатність зв'язуватися з молекулою HLA в такий же спосіб, як і зазначений пептид, відповідно до будь-якої з послідовностей від SEQ ID NO 11 до SEQ ID NO 22 та SEQ ID No 24. Відповідно, варіанти, що індукують перехресне реагування T-клітин з пептидом згідно винаходу, часто є варіантами з різною довжиною. Якщо пептид більший, ніж приблизно 12 амінокислотних залишків, він використовується безпосередньо для зв'язування з молекулою MHC класу II. Переважно, щоб залишки, розташовані по краях основної ділянки зв'язування HLA, не мали значного впливу на здатність пептиду специфічно зв'язуватися зі зв'язувальною порожниною MHC-молекули класу ІІ чи презентувати пептид ЦТЛ. Однак, як вже зазначалося вище, перевага надається використанню більш крупних пептидів, особливо коли вони кодуються полінуклеотидом, оскільки ці більш крупні пептиди можуть розділятися на частини відповідними антигенпрезентуючими клітинами. Крім того, фланкуючі амінокислоти також можуть зменшувати швидкість пептидної деградації іn vivo, отже, кількість фактичного пептиду, доступного для ЦТЛ, є вищою в порівнянні з пептидом без фланкуючих амінокислот. Також можливо, щоб епітопи MHC класу I, хоча вони мають, зазвичай, довжину 8-10 амінокислот, утворювались шляхом процесингу пептидів з більш довгих пептидів чи білків, які включають фактичний епітоп. Подібно епітопам MHC класу II, бажано, щоб бокові залишки подовжених пептидів-попередників за та/або до N- та C-кінця фактичного епітопу не завдавали значного впливу на презентування пептиду ЦТЛ та не маскували сайти протеолітичного 24 UA 109891 C2 5 10 15 20 25 30 35 40 45 50 55 60 розпаду, необхідного для утворення фактичного епітопу, опосередкованого процесингом подовженого пептиду. Переважними є пептиди з коровою послідовністю, що складається з послідовностей від SEQ ID NO 1 до SEQ ID NO 10 та SEQ ID 11 з подовженнями від 1 до 10 амінокислот на C-кінці та/або N-кінці, більш переважною є загальна довжина цих фланкуючих амінокислот від 1 до 12, більш переважною - від 1 до 10, більш переважною - від 1 до 8, і ще більш переважною від 1 до 6, де фланкуючі амінокислоти можуть розподілятися в будь-якому співвідношенні на C-кінці та N-кінці (наприклад, усі бокові амінокислоти можуть додаватися до одного кінця, чи амінокислоти можуть додаватися в рівних кількостях до обох кінців, чи в будь-якому іншому співвідношенні), за умови, що пептид зберігає здатність зв'язуватися з молекулою HLA в такий же спосіб, як і зазначений пептид, відповідно до будь-якої з послідовностей від SEQ ID NO 1 до SEQ ID NO 10 та SEQ ID No 11. Відповідно, цей винахід також пропонує пептиди та варіанти епітопів MHC класу I, які мають загальну довжину від 8 до 100, переважно від 8 до 30, та найбільш переважно від 8 до 18, а саме, 8, 9, 10, 11, 12, 13, 14, 15, 16 або 17 амінокислот. Звичайно, пептид чи варіант відповідно до цього винаходу матиме здатність зв'язуватися з молекулою MHC класу I чи II людини. Зв'язування пептиду чи варіанту з MHC-комплексом може бути перевірене за допомогою методів, добре відомих в цій галузі, наприклад, описаних в прикладах цього винаходу нижче або в довідковій літературі для різних алелей MHC класу II (наприклад, в роботах Vogt AB, Kropshofer H, Kalbacher H, Kalbus M, Rammensee HG, Coligan JE, Martin R; Ligand motifs of HLA-DRB5*0101 and DRB1*1501 molecules delineated from selfpeptides; J Immunol. 1994; 153(4):1665-1673; Malcherek G, Gnau V, Stevanovic S, Rammensee HG, Jung G, Melms A; Analysis of allele-specific contact sites of natural HLA-DR17 ligands; J Immunol. 1994; 153(3):1141-1149; Manici S, Sturniolo T, Imro MA, Hammer J, Sinigaglia F, Noppen C, Spagnoli G, Mazzi B, Bellone M, Dellabona P, Protti MP; Melanoma cells present a MAGE-3 epitope to CD4(+) cytotoxic T cells in association with histocompatibility leukocyte antigen DR11; J Exp Med. 1999; 189(5): 871-876; Hammer J, Gallazzi F, Bono E, Karr RW, Guenot J, Valsasnini P, Nagy ZA, Sinigaglia F; Peptide binding specificity of HLA-DR4 molecules: correlation with rheumatoid arthritis association;.J Exp Med. 1995 181(5):1847-1855; Tompkins SM, Rota PA, Moore JC, Jensen PE; A europium fluoroimmunoassay for measuring binding of antigen to class II MHC glycoproteins; J Immunol Methods. 1993;163(2): 209-216; Boyton RJ, Lohmann T, Londei M, Kalbacher H, Halder T, Frater AJ, Douek DC, Leslie DG, Flavell RA, Altmann DM; Glutamic acid decarboxylase T lymphocyte responses associated with susceptibility or resistance to type I diabetes: analysis in disease discordant human twins, non-obese diabetic mice and HLA-DQ transgenic mice; Int Immunol. 1998 (12):1765-1776). Пептиди згідно цього винаходу можуть мати додаткові N- та/або C-термінальні ділянки амінокислот, які не обов'язково формують частину пептиду, що функціонує як фактичний епітоп для молекул MHC, та все ж таки можуть бути важливими для забезпечення ефективного введення пептиду відповідно до цього винаходу в клітини (див. вище). В одному з втілень цього винаходу пептид згідно цього винаходу є злитим білком, який вміщує, наприклад, 80 Nтермінальних амінокислот HLA-DR-антиген-асоційованого інваріантного ланцюгу (p33, надалі "Ii"), одержаного з банка даних NCBI, інвентарний номер (GenBank Accession-number) X00497 (Strubin, M., Mach, B. and Long, E.O. The complete sequence of the mRNA for the HLA-DRassociated invariant chain reveals a polypeptide with an unusual transmembrane polarity EMBO J. 3 (4), 869-872 (1984)). Цей винахід також пропонує фармацевтичну композицію, що вміщує принаймні один з пептидів згідно цього винаходу, в яких пептиди мають загальну довжину від 8 до 100, переважно від 8 до 30 та більш переважно від 8 до 17 або 9, 10, 11, 12, 13, 14, 15 чи 16 амінокислот. Крім того, пептид чи варіант може бути додатково модифікований для поліпшення його стабільності та/або зв'язку із молекулами MHC, щоб викликати сильнішу імунну відповідь. Методи для такої оптимізації пептидної послідовності добре відомі в цій галузі та включають, наприклад, введення реверсованих пептидних зв'язків чи непептидних зв'язків. Таким чином, відповідно до іншого аспекту винахід надає фармацевтичну композицію, в якій принаймні один пептид чи варіант включає непептидні зв'язки. В реверсованому пептидному зв'язку амінокислотні залишки не з'єднуються пептидними зв'язками (-CO-NH-), а пептидний зв'язок реверсується. Такі ретро-інвертовані пептидні міметики можуть бути отримані за методами, відомими в даній галузі, наприклад, описаними в роботі Meziere і співавт (1997) J. Immunol. 159, 3230-3237, яка включена в даний документ шляхом посилання. Такий підхід включає формування псевдопептидів, які вміщують зміни із 25 UA 109891 C2 5 10 15 20 25 30 35 40 45 50 55 60 залученням остова, а не орієнтації бокових ланцюгів. Meziere і співавт (1997) показують, що для відповідей клітин MHC і хелперних T-клітин зазначені псевдопептиди є прийнятними. Ретроінвертовані пептиди, які вміщують зв'язки NH-CO замість пептидних зв'язків CO-NH, набагато більш стійкі до протеолізу. Непептидний зв'язок - це, наприклад, -CH2-NH, -CH2S-, -CH2CH2-, -CH=CH-, -COCH2-, CH(OH)CH2-, та -CH2SO-. Патент США 4 897 445 пропонує метод твердофазного синтезу непептидних зв'язків (-CH2-NH) в поліпептидних ланцюгах, що включають поліпептиди, синтезовані за допомогою стандартних процедур, та непептидний зв'язок, синтезований шляхом реакції аміноальдегіду та амінокислоти в присутності NaCNBH 3. Пептиди, що включають послідовності за винаходом, описані вище, можуть бути синтезовані з додатковими хімічними групами, присутніми на їхніх аміно- та/або карбоксильних кінцях, зокрема, для посилення стабільності, біологічної доступності та/або афінності пептидів. Наприклад, гідрофобні групи, такі як карбобензоксильні, данзильні чи трет-бутилоксикарбонільні групи можуть додаватися до аміно-кінців пептидів. Подібним чином, на аміно-кінцях пептидів може розміщуватись ацетильна група чи 9-фторенілметоксикарбонільна група. До карбоксильних кінців пептидів може бути додана також, наприклад, гідрофобна група, третбутилоксикарбонільна чи амідна група. Крім того, усі пептиди згідно винаходу можуть бути синтезовані для зміни їхньої просторової конфігурації. Наприклад, може використовуватися D-ізомер одного чи кількох амінокислотних залишків пептиду, а не звичайний L-ізомер. До того ж, принаймні один з амінокислотних залишків пептидів згідно винаходу може замінюватись одним з добре відомих амінокислотних залишків не природного походження. Такі заміни можуть служити для підвищення стабільності, біологічної доступності та/або зв'язувальних властивостей пептидів згідно винаходу. Подібним чином, пептид чи варіант за винаходом може модифікуватися хімічно, шляхом реакції окремих амінокислот до чи після синтезу пептиду. Приклади таких модифікацій добре відомі в цій галузі та узагальнені, зокрема, в роботі R. Lundblad, Chemical Reagents for Protein Modification, 3rd ed. CRC Press, 2005, яка включена в цей документ шляхом посилання. Хімічна модифікація амінокислот включає, без обмежень, модифікацію шляхом ацилювання, амідинування, піридоксилювання лізину, відновлювального алкілування, тринітробензилювання аміногруп 2,4,6-тринітробензолсульфоновою кислотою (TNBS), амідну модифікацію карбоксильних груп та сульфгідрильну модифікацію шляхом окиснення цистеїну надмурашиною кислотою в цистеїнову кислоту, формування похідних ртуті, утворення змішаних дисульфідів з іншими тіольними сполуками, реакцію з малеімідом, карбоксиметилювання йодооцтовою кислотою чи йодоацетамідом та карбамоїлування ціанатом при лужному pH, хоча способи модифікації не обмежуються наведеними тут. В цьому відношенні досвідчений спеціаліст повинен звернутися до Глави 15 роботи Current Protocols In Protein Science, Eds. Coligan et al. (John Wiley & Sons NY 1995-2000), де викладена детальна методика відносно хімічної модифікації білків. Успішна модифікація PEG (поліетиленгліколем) терапевтичних білків та пептидів, що часто асоціюється з подовженням напівперіоду циркуляції, при перехресному зшиванні білків з глутаральдегідом, поліетиленглікольдіакрилатом та формальдегідом використовується для приготування гідрогелів. Хімічна модифікація алергенів для імунотерапії часто досягається шляхом карбамоїлування ціанатом калію. В цілому, пептиди та варіанти (принаймні, ті, що вміщують пептидні зв'язки між амінокислотними залишками) можуть бути синтезовані, наприклад, із використанням Fmocполіамідного способу твердофазного синтезу пептидів, як розкривається в роботі Lu і співавт (1981) J. Org. Chem. 46, 3433, та посиланнях на літературні джерела, наведених в ній. Очищення може виконуватися за допомогою будь-якого одного чи кількох методів, таких як рекристалізація, ексклюзійна хроматографія, іонообмінна хроматографія, хроматографія гідрофобної взаємодії та (зазвичай) зворотно-фазова високоефективна рідинна хроматографія з використанням, наприклад, градієнтного розділення у системі ацетонітрил/ вода. Аналіз пептидів може виконуватися за допомогою тонкошарової хроматографії, електрофорезу, зокрема, капілярного електрофорезу, твердофазної екстракції (ТФЕ), зворотнофазової високоефективної рідинної хроматографії, амінокислотного аналізу після кислотного гідролізу та мас-спектрометричного аналізу із бомбардуванням прискореними атомами (FAB), а також мас-спектрометричного аналізу MALDI та ESI-Q-TOF. Ще один аспект винаходу пропонує нуклеїнову кислоту (наприклад, полінуклеотид), що кодує пептид чи варіант за винаходом. Полінуклеотид, наприклад, може бути ДНК, кДНК, ПНК, ЦНК, РНК, чи їхньою комбінацією, одно- та/або двонитковою, природними чи стабілізованими формами полінуклеотидів, такими як, наприклад, полінуклеотиди з фосфоротіоатним остовом 26 UA 109891 C2 5 10 15 20 25 30 35 40 45 50 55 60 або їхніми комбінаціями; він може вміщувати або не вміщувати інтрони, за умови, що полінуклеотид кодує пептид. Звичайно, тільки ті пептиди, що вміщують природно існуючи амінокислотні залишки, з'єднані природними пептидними зв'язками, кодуються полінуклеотидом. Інший аспект винаходу пропонує вектор експресії, здатний експресувати поліпептид відповідно до винаходу. Вектори експресії для різних типів клітин добре відомі в цій галузі та можуть бути вибрані без неналежного експериментування. Зазвичай ДНК вводиться у вектор експресії, такий як плазміда, в належній орієнтації та з відповідною рамкою зчитування експресії. При необхідності ДНК може зв'язуватися з належними нуклеотидними послідовностями, які забезпечують координацію транскрипції і трансляції, що розпізнаються бажаним хазяїном, хоча такі контрольні елементи взагалі є доступними у векторі експресії. Вектор згодом вводиться хазяїну із використанням стандартних методик. За рекомендаціями можна звернутись, наприклад, до роботи Sambrook et al. (1989) Molecular Cloning, A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor, NY. Разом із тим, фармацевтична композиція в особливо переважному втіленні винаходу вміщує принаймні два пептиди, які включають амінокислотні послідовності відповідно з SEQ ID NO 1 до SEQ ID NO 12. Оптимальну кількість кожного пептиду, що включається у вакцину, та оптимальний режим дозування може визначити досвідчений спеціаліст без неналежного експериментування. Наприклад, пептид чи його варіант може бути підготовлений для внутрішньовенного (i.v.) введення, підшкірного (s.c.) введення, внутрішньошкірного (i.d.) введення, внутрішньочеревного (i.p.) введення, внутрішньом'язового (i.m.) введення. Переважні способи введення пептиду - це s.c., i.d., i.p., i.m. та i.v. Переважними способами введення ДНК є i.d., i.m., s.c., i.p. та i.v. Можуть вводитись, наприклад, дози від 1 до 500 мг, та 50 мкг і 1,5 мг, переважно від 125 мкг до 500 мкг, пептиду чи ДНК, залежно від відповідного пептиду чи ДНК. Дози в цьому діапазоні успішно використовувались в попередніх дослідженнях (Brunsvig PF, Aamdal S, Gjertsen MK, Kvalheim G, Markowski-Grimsrud CJ, Sve I, Dyrhaug M, Trachsel S, Møller M, Eriksen JA, Gaudernack G;. Telomerase peptide vaccination: a phase I/II study in patients with non-small cell lung cancer; Cancer Immunol Immunother. 2006; 55(12):1553-1564; M. Staehler, A. Stenzl, P. Y. Dietrich, T. Eisen, A. Haferkamp, J. Beck, A. Mayer, S. Walter, H. Singh, J. Frisch, C. G. Stief; An open label study to evaluate the safety and immunogenicity of the peptide based cancer vaccine IMA901, ASCO meeting 2007; Abstract No 3017). Фармацевтичні композиції згідно цього винаходу можуть бути складені таким чином, щоб вибір, число та/або кількість пептидів, присутніх у композиції, були специфічними для тканин, ракових захворювань та/або окремих пацієнтів. Наприклад, точний вибір пептидів може визначатися профілями експресії вихідних білків в даній тканини, щоб уникнути побічних ефектів. Вибір може залежати від конкретного виду раку, на який хворіє пацієнт, що проходить лікування, а також статусу захворювання, попередніх режимів лікування, імунного статусу пацієнта і, звичайно, HLA-гаплотипу пацієнта. Крім того, вакцина згідно із винаходом може вміщувати індивідуалізовані компоненти, відповідно до особистих потреб конкретного пацієнта. Прикладами є різні кількості пептидів згідно із експресією відповідних TAA у конкретного пацієнта, небажаними побічними ефектами внаслідок алергії чи інших видів лікування, а також корегування для вторинного лікування після проходження першого циклу чи схеми лікування. Для того, щоб, наприклад, композиція використовувалася як вакцина проти гліобластоми, пептидів, вихідні білки яких експресуються у великих кількостях в нормальних тканинах, уникають в композиції згідно винаходу чи допускають їхню присутність у невеликих кількостях. З другого боку, коли відомо, що пухлина пацієнта експресує великі кількості конкретного білка, відповідна фармацевтична композиція для лікування цього випадку раку може бути присутньою у великих кількостях, та/або може включати більш ніж один пептид, специфічний для цього конкретного білка чи його сигнального шляху. Досвідчений фахівець зможе вибрати переважні комбінації імуногенних пептидів шляхом тестування, наприклад, популяції T-клітин in vitro, а також їхньої ефективності та загальної присутності, проліферації, афінності та розмноження окремих T-клітин для певних пептидів, і функціональності T-клітин, наприклад, аналізуючи утворення IFN-γ (див. також приклади нижче). Зазвичай найбільш ефективні пептиди згодом комбінуються як вакцина в цілях, описаних вище. Прийнятна вакцина буде переважно вміщувати від 1 до 20 пептидів, більш переважно 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 чи 20 різних пептидів, ще більш переважно 6, 7, 8, 9, 10 11, 12, 13 або 14 різних пептидів та найбільш переважно 10, 11, 12, 13 або 14 різних пептидів. Довжина пептиду для використання в протираковій вакцині може бути будь-якою прийнятною довжиною, як розкривається в цьому винаході. Зокрема, це може бути прийнятний пептид із 9 амінокислотних залишків або прийнятний пептид із 8 чи 9 чи 10 чи 11 27 UA 109891 C2 5 10 15 20 25 30 35 40 45 50 55 60 амінокислотних залишків, або пептид із 12, 13, 14 чи 15 амінокислотних залишків. Пептиди більшої довжини також можуть використовуватись; пептиди із 9 чи 10 амінокислотних залишків, як описано в доданих Таблицях 1 і 2, є переважними для пептидів MHC класу I, в той час як 1215 амінокислотних залишків переважні для пептидів MHC класу II. Пептид(и) становить(становлять) протипухлинну чи протиракову вакцину. Вона може вводитись безпосередньо пацієнту в уражений орган чи системно, або застосовуватись ex vivo до клітин, виділених з пацієнта або клітинної лінії людини, що згодом вводяться пацієнту, чи використовуватись in vitro для селекції субпопуляції з імунних клітин, які отримані від пацієнта і потім знов вводяться йому. Пептид може бути, по суті, чистим, або поєднаним з імуностимулюючим ад'ювантом (див. нижче) чи може використовуватись в комбінації з імуностимулюючими цитокінами, або вводитись з належною системою доставки, наприклад, ліпосомами. Пептид також може бути кон'югованим із належним носієм, таким як гемоціанін фісурели (KLH) або манан (див. заявку WO 95/18145 та роботу Longenecker et al (1993) Ann. NY Acad. Sci. 690,276-291). Пептид також може бути міченим, і бути злитим білком чи гібридною молекулою. Очікується, що пептиди, послідовність яких надається в цьому винаході, стимулюють CD4 T-клітини або CD8 ЦТЛ. Однак стимуляція більш ефективна у присутності підтримки, яка надається T-клітинами, позитивними до протилежного CD. Отже, для епітопів MHC класу II, котрі стимулюють CD4 T-клітини, партнер у злитті або частини гібридної молекули належним чином надають епітопи, що стимулюють CD8-позитивні T-клітини. З другого боку, для епітопів MHC класу I, які стимулюють CD8 ЦТЛ, партнер у злитті або частини гібридної молекули належним чином надають епітопи, котрі стимулюють CD4-позитивні T-клітини. CD4- та CD8-стимулюючі епітопи добре відомі в цій галузі та включають епітопи, ідентифіковані в цьому винаході. Фармацевтично прийнятні носії добре відомі; це, зазвичай, рідини, в яких формулюється активна терапевтична речовина. Носій взагалі не додає композиції ніякої фармакологічної активності, хоча він може забезпечити хімічну та/або біологічну стабільність, характеристики вивільнення і таке інше. Зразки композицій можна знайти, наприклад, в роботі Alfonso R. Gennaro. Remington: The Science and Practice of Pharmacy, 20th Edition. Baltimore, MD: Lippincott Williams & Wilkins, 2000, і вони включають фізіологічний розчин, воду, воду з буфером, 0,3 % гліцин, гіалуронову кислоту, глюкозу і тому подібне, але не обмежуються ними. Останнім часом було виявлено, що певні жирові емульсії, які багато років використовувалися для внутрішньовенного харчування людей, також можуть бути носіями для пептидів. Двома прикладами таких емульсій є наявні у продажу жирові емульсії, відомі під назвами Інтраліпід та Ліпофундин. "Інтраліпід" - це зареєстрована торгова марка жирової емульсії для внутрішньовенного харчування фірми Kabi Pharmacia, Швеція, яка описана в патенті США 3,169,094. "Ліпофундин" - це зареєстрована торгова марка фірми B. Braun Melsungen, Німеччина. Обидві емульсії вміщують соєве масло як жир (100 або 200 г в 1000 мл дистильованої води: 10 % чи 20 %, відповідно). Фосфоліпіди яєчного жовтка використовуються як емульгатори в Інтраліпіді (12 г/л дистильованої води), а лецитин яєчного жовтка - в Ліпофундині (12 г/л дистильованої води). Ізотонічність є результатом додавання гліцерину (25 г/л), як в Інтраліпіді, так і в Ліпофундині. Для викликання імунної відповіді зазвичай необхідно включення ад'ювантів, що надають композиції більшої імуногенності. Тобто, в переважному втіленні винаходу фармацевтична композиція також вміщує принаймні один прийнятний ад'ювант. Прийнятні ад'юванти включають, без обмежень, 1018 ISS, солі алюмінію, Amplivax, AS15, BCG, CP-870,893, CpG7909, CyaA, dSLIM, флагелін чи ліганди TLR5, які походять з флагеліну, ліганд FLT3, GM-CSF, IC30, IC31, іміквімод, (ALDARA®), резиквімод, ImuFact IMP321, интерлейкіни, такі як ІЛ-2, ІЛ-13, ІЛ-21, інтерферон-альфа чи -бета, або їхні пегільовані похідні, IS Patch, ISS, імуностимулюючі комплекси ISCOM, JuvImmune, LipoVac, MALP2 MF59, монофосфорил ліпід A, Монтанід IMS 1312, Монтанід ISA 206, Монтанід ISA 50V, Монтанід ISA51, емульсії "вода у маслі" та "масло у воді", OK-432, OM-174, OM-197-MP-EC, ONTAK, OspA, векторну систему PepTel®, мікрочастинки на основі полі(лактид когліколіду) [PLG] та декстрану, талактоферин SRL172, віросоми та інші вірусоподібні частинки, YF-17D, VEGF trap, R848, бетаглюкан, Pam3Cys, стимулон QS21 Aquila, який виділяється з сапоніну, мікобактеріальні екстракти та синтетичні імітатори стінок бактеріальних клітин, а також інші патентовані ад'юванти, наприклад, Detox компанії Ribi, Quil чи Superfos. Переважними є такі ад'юванти, як ад'ювант Фрейнда або GM-CSF. Кілька імунологічних ад'ювантів (наприклад, MF59), специфічних для дендритних клітин, та їхнє приготування були описані раніше (Aucouturier і співавт., 2001; Allison і Krummel, 1995). Також можуть застосовуватися цитокіни. Окремі цитокіни були прямо співвіднесені з впливом на міграцію дендритних клітин до лімфоїдних тканин 28
ДивитисяДодаткова інформація
Назва патенту англійськоюComposition of tumor-associated peptides treatment of cancerous diseases
Автори англійськоюFritsche, Jens, Weinschenk, Toni, Walter, Steffen, Lewandrowski, Peter, Singh, Harpreet
Назва патенту російськоюКомпозиция опухолеассоциированных пептидов для лечения раковых заболеваний
Автори російськоюФритше Йенс, Вейншенк Тони, Вальтер Штеффен, Левандровски Петер, Зингх Харпреет
МПК / Мітки
МПК: A61K 38/17, A61P 35/00, C07K 7/06
Мітки: композиція, пухлиноасоційованих, захворювань, ракових, лікування, пептидів
Код посилання
<a href="https://ua.patents.su/59-109891-kompoziciya-pukhlinoasocijjovanikh-peptidiv-dlya-likuvannya-rakovikh-zakhvoryuvan.html" target="_blank" rel="follow" title="База патентів України">Композиція пухлиноасоційованих пептидів для лікування ракових захворювань</a>
Фармацевтична композиція пухлиноасоційованих пептидів та її застосування для лікування ракових захворювань

Номер патенту: 103202
Опубліковано: 25.09.2013
Автори: Вальтер Штеффен, Хільф Норберт, Вейншенк Тоні, Сінгх Харпреет, Шор Олівер, Траутвейн Клаудія
МПК: A61K 38/08, A61K 38/16, A61K 39/00, A61K 35/00, A61P 35/00
Мітки: пептидів, застосування, ракових, композиція, лікування, фармацевтична, захворювань, пухлиноасоційованих
Формула / Реферат:
1. Фармацевтична композиція, яка включає (і) принаймні два імуногенних пептиди, де один з зазначених пептидів складається з амінокислотної послідовності, ідентичної до SEQ ID NO: 3, та (іі) фармецевтично прийнятний носій.2. Фармацевтична композиція за п. 1, яка додатково включає принаймні один додатковий імуногенний пептид, що складається з амінокислотної послідовності, вибраної з групи, що включає SEQ ID NO: 1, SEQ ID NO: 2 та...
Фармацевтична композиція для лікування ракових захворювань

Номер патенту: 84948
Опубліковано: 11.11.2013
Автор: Тарасов Міхаіл Вячєславовіч
МПК: A61K 31/7016, A61K 33/16, A61K 9/28
Мітки: фармацевтична, захворювань, ракових, композиція, лікування
Формула / Реферат:
Фармацевтична композиція для лікування ракових захворювань, яка виконана у формі таблетки, що складається з основи та плівкового покриття, причому основа таблетки містить капецитабін, лактозу, мікрокристалічну целюлозу, стеарат магнію, яка відрізняється тим, що основа таблетки додатково містить кармелозу натрію, сухий кукурудзяний крохмаль, повідон К-30, колоїдний безводний діоксид кремнію та тальк, при наступному співвідношенні компонентів...
Нові композиції пухлиноасоційованих пептидів, що зв’язуються з молекулами і або іі класу лейкоцитарного антигену людини (hla) для розробки вакцин

Номер патенту: 104572
Опубліковано: 25.02.2014
Автори: Левандровскій Петер, Флор Крістіан
МПК: A61K 38/10
Мітки: людини, класу, hla, нові, антигену, зв'язуються, розробки, пептидів, пухлиноасоційованих, лейкоцитарного, молекулами, вакцин, композиції
Формула / Реферат:
1. Фармацевтична композиція, яка містить:від 2 до 18, переважно від 2 до 15, більш переважно від 2 до 13, ще більш переважно від 2 до 12 пептидів та ще більш переважно від 3 до 12 пухлиноасоційованих пептидів;де кожний пептид має довжину від 8 до 22 амінокислот, переважно від 9 до 16 амінокислот;де зазначені пептиди проявляють розчинність в 90 % оцтовій кислоті, що складає щонайменше 2,7 мг/мл;манітол і...
Стабільний розчин мітоксантрону, спосіб його виготовлення, інгредієнт для виготовлення лікарського засобу для лікування ракових захворювань та лікарський засіб

Номер патенту: 72461
Опубліковано: 15.03.2005
Автори: Вольф-Хойсс Елізабет, Вайнгарт Маріо, Мошнер Катрін, Пірот Міхаель, Морік Вольфганг
МПК: A61K 47/18, A61P 35/00, A61K 31/135
Мітки: розчин, інгредієнт, стабільний, захворювань, засобу, спосіб, лікарський, ракових, мітоксантрону, засіб, лікування, виготовлення, лікарського
Формула / Реферат:
1. Стабільний розчин мітоксантрону, який містить від 1 мг/мл до 5 мг/мл розчину мітоксантрон-гідрохлориду, від 0,001 до 0,15% (мас.) едетату натрію і хлорид натрію, ацетат натрію і оцтову кислоту як додаткові компоненти і який є водним і вільним від сульфітних сполук.2. Стабільний розчин мітоксантрону для ін'єкцій за п. 1, який відрізняється тим, що вміст едетату натрію у варіанті, якому віддається перевага, складає 0,04 %...
Спосіб гістохімічної діагностики ракових захворювань

Номер патенту: 43904
Опубліковано: 10.09.2009
Автори: Хатковий Сергій Андрійович, Підгаєвський Ігор Юрійович, Підгаєвська Тетяна Петрівна, Трунова Олена Костянтинівна, Суслов Євгеній Іванович
МПК: G01N 33/49
Мітки: діагностики, гістохімічної, спосіб, ракових, захворювань
Формула / Реферат:
Спосіб гістохімічної діагностики ракових захворювань шляхом дослідження продуктів гістохімічної реакції на мазку крові в зоні дії діагностичних реагентів, який відрізняється тим, що на мазок крові пацієнта послідовно, з інтервалом 10 хв наносять по 1 краплині розчину комплексоноутворювача - оксіетилендифосфонової кислоти, в концентрації 6 ваг.%-10 ваг.%, стандартного біологічно активного донатора лужноземельного металу кальцію - кальцієвого...
Попередній патент: Композиція, що включає пестицид і алкоксилат 2-пропілгептиламіну
Наступний патент: Спосіб термомеханічної обробки титанового сплаву (варіанти)
Випадковий патент: Фармацевтична композиція у формі іонізованого настою для лікування атеросклерозу