Композиція анти-гліпікан 3-антитіл, що мають модифікований цукровий ланцюг
Номер патенту: 93488
Опубліковано: 25.02.2011
Автори: Суго Ізумі, Танака Мегумі, Накано Кійотака, Іідзіма Сігеюкі, Ісігуро Такахіро, Сугімото Масаміті






Формула / Реферат
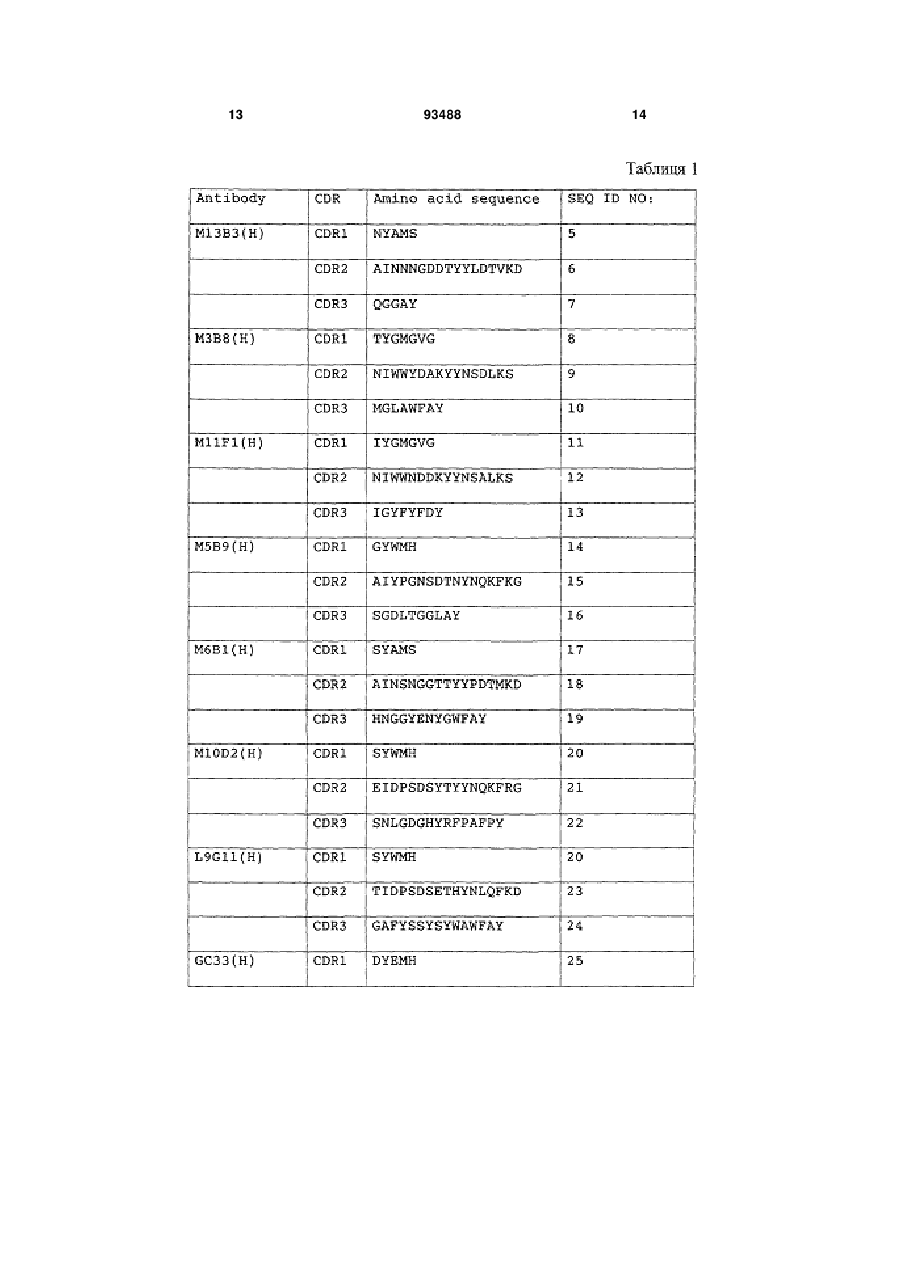
1. Композиція анти-гліпікан 3-антитіл, яка містить антитіло, що має варіабельну ділянку важкого ланцюга, яка містить CDR1, що має амінокислотну послідовність, представлену в SEQ ID NО: 25, CDR2, що має амінокислотну послідовність, представлену в SEQ ID NО: 26, і CDR3, що має амінокислотну послідовність, представлену в SEQ ID NО: 27, де компонент, який представляє цукровий ланцюг антитіла, був модифікований так, щоб мати підвищену частку антитіл, позбавлених фукози, або підвищену частку антитіл, що мають приєднаний дворозсікаючий N-ацетилглюкозамід (GlcNAc).
2. Композиція анти-гліпікан 3-антитіл, яка містить антитіло, що має варіабельну ділянку легкого ланцюга, яка містить CDR1, що має амінокислотну послідовність, представлену в SEQ ID NО: 45, CDR2, що має амінокислотну послідовність, представлену в SEQ ID NО: 46, і CDR3, що має амінокислотну послідовність, представлену в SEQ ID NО: 60, де компонент, який представляє цукровий ланцюг антитіла, був модифікований так, щоб матипідвищену частку антитіл, позбавлених фукози або підвищену частку антитіл, що мають приєднаний дворозсікаючий N-ацетилглюкозамід (GlcNAc).
3. Композиція анти-гліпікан 3-антитіл, в якій антитіло, визначене у п. 1, додатково має варіабельну ділянку легкого ланцюга, яка містить CDR1, що має амінокислотну послідовність, представлену в SEQ ID NО: 45, CDR2, що має амінокислотну послідовність, представлену в SEQ ID NО: 46, і CDR3, що має амінокислотну послідовність, представлену в SEQ ID NО: 60.
4. Композиція анти-гліпікан 3-антитіл, яка містить антитіло, що має одну з перерахованих (1) - (7) варіабельних ділянок важкого ланцюга:
(1) варіабельну ділянку важкого ланцюга, яка містить амінокислотну послідовність, представлену в SEQ ID NО: 81;
(2) варіабельну ділянку важкого ланцюга, яка містить амінокислотну послідовність, представлену в SEQ ID NО: 82;
(3) варіабельну ділянку важкого ланцюга, яка містить амінокислотну послідовність, представлену в SEQ ID NО: 83;
(4) варіабельну ділянку важкого ланцюга, яка містить амінокислотну послідовність, представлену в SEQ ID NО: 84;
(5) варіабельну ділянку важкого ланцюга, яка містить амінокислотну послідовність, представлену в SEQ ID NО: 85;
(6) варіабельну ділянку важкого ланцюга, яка містить амінокислотну послідовність, представлену в SEQ ID NО: 86; або
(7) варіабельну ділянку важкого ланцюга, яка містить амінокислотну послідовність, представлену в SEQ ID NО: 87,
де компонент, який представляє цукровий ланцюг антитіла, був модифікований так, щоб мати підвищену частку антитіл, позбавлених фукози або підвищену частку антитіл, що мають приєднаний дворозсікаючий N-ацетилглюкозамід (GlcNAc).
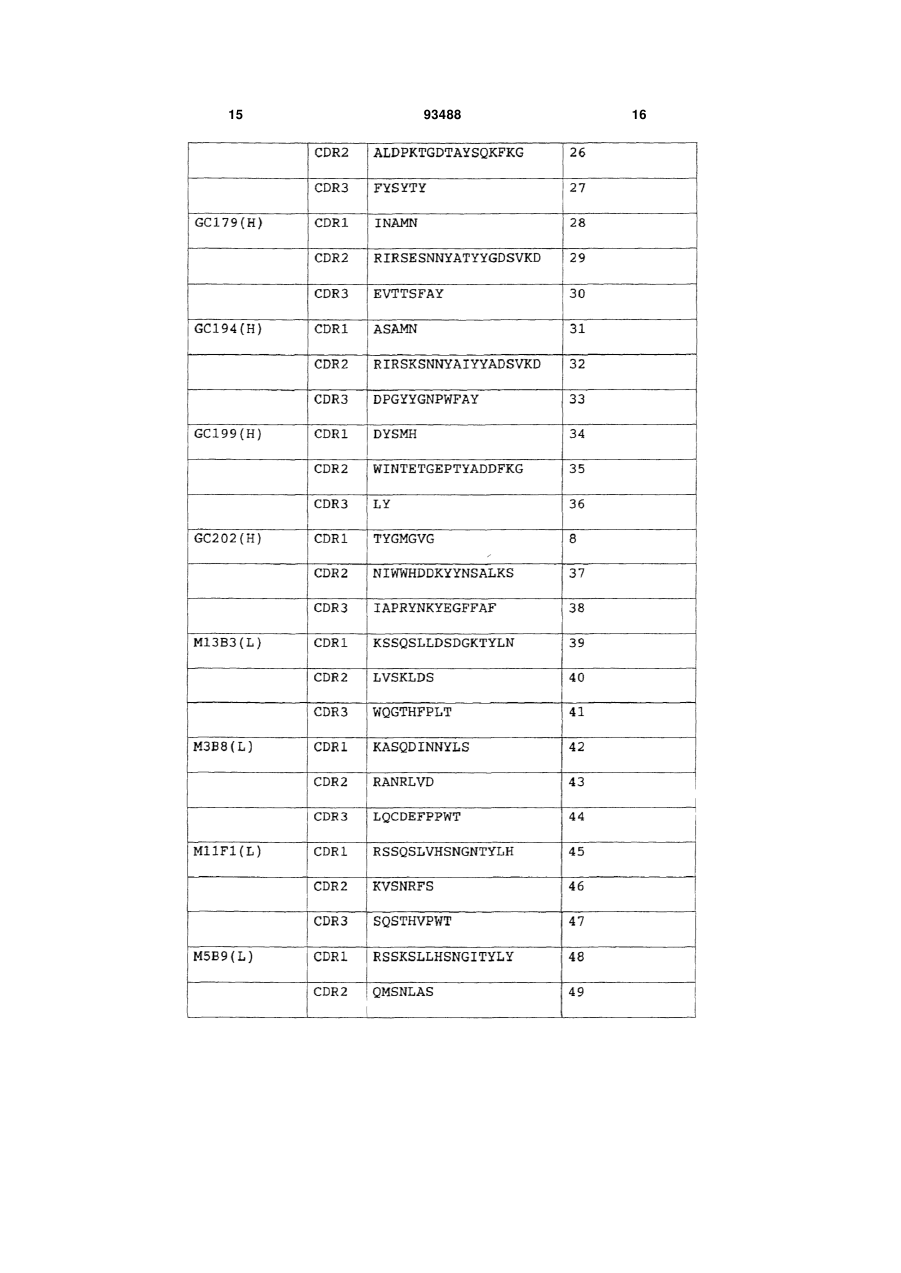
5. Композиція анти-гліпікан 3-антитіл, яка містить антитіло, що має варіабельну ділянку легкого ланцюга, яка містить амінокислотну послідовність, представлену в SEQ ID NО: 109, де компонент, який представляє цукровий ланцюг антитіла, був модифікований так, щоб мати підвищену частку антитіл, позбавлених фукози або підвищену частку антитіл, що мають приєднаний дворозсікаючий N-ацетилглюкозамід (GlcNAc).
6. Композиція анти-гліпікан 3-антитіл за п. 4, яка додатково містить антитіло, що має варіабельну ділянку легкого ланцюга, яка містить амінокислотну послідовність, представлену в SEQ ID NО: 109.
7. Композиція анти-гліпікан 3-антитіл, яка має активність, еквівалентну активності композиції антитіл за п. 3, де один або декілька амінокислотних залишків заміщені, делетовані або додані і/або вставлені з амінокислотних послідовностей, представлених в будь-якому з пп. 1-6.
8. Композиція анти-гліпікан 3-антитіл за п. 7, яка містить антитіло, що має варіабельну ділянку легкого ланцюга, яка містить одне з перерахованих (1) - (15) CDR1, 2 і 3:
(1) CDR 1, що містить амінокислотну послідовність, представлену в SEQ ID NО: 90, CDR 2, що містить амінокислотну послідовність, представлену в SEQ ID NО: 46, і CDR 3, що містить амінокислотну послідовність, представлену в SEQ ID NО: 60;
(2) CDR 1, що містить амінокислотну послідовність, представлену в SEQ ID NО: 91, CDR 2, що містить амінокислотну послідовність, представлену в SEQ ID NО: 46, і CDR 3, що містить амінокислотну послідовність, представлену в SEQ ID NО: 60;
(3) CDR 1, що містить амінокислотну послідовність, представлену в SEQ ID NО: 92, CDR 2, що містить амінокислотну послідовність, представлену в SEQ ID NО: 46, і CDR 3, що містить амінокислотну послідовність, представлену в SEQ ID NО: 60;
(4) CDR 1, що містить амінокислотну послідовність, представлену в SEQ ID NО: 93, CDR 2, що містить амінокислотну послідовність, представлену в SEQ ID NО: 46, і CDR 3, що містить амінокислотну послідовність, представлену в SEQ ID NО: 60;
(5) CDR 1, що містить амінокислотну послідовність, представлену в SEQ ID NО: 94, CDR 2, що містить амінокислотну послідовність, представлену в SEQ ID NО: 46, і CDR 3, що містить амінокислотну послідовність, представлену в SEQ ID NО: 60;
(6) CDR 1, що містить амінокислотну послідовність, представлену в SEQ ID NО: 95, CDR 2, що містить амінокислотну послідовність, представлену в SEQ ID NО: 46, і CDR 3, що містить амінокислотну послідовність, представлену в SEQ ID NО: 60;
(7) CDR 1, що містить амінокислотну послідовність, представлену в SEQ ID NО: 96, CDR 2, що містить амінокислотну послідовність, представлену в SEQ ID NО: 46, і CDR 3, що містить амінокислотну послідовність, представлену в SEQ ID NО: 60;
(8) CDR 1, що містить амінокислотну послідовність, представлену в SEQ ID NО: 97, CDR 2, що містить амінокислотну послідовність, представлену в SEQ ID NО: 46, і CDR 3, що містить амінокислотну послідовність, представлену в SEQ ID NО: 60;
(9) CDR 1, що містить амінокислотну послідовність, представлену в SEQ ID NО: 98, CDR 2, що містить амінокислотну послідовність, представлену в SEQ ID NО: 46, і CDR 3, що містить амінокислотну послідовність, представлену в SEQ ID NО: 60;
(10) CDR 1, що містить амінокислотну послідовність, представлену в SEQ ID NО: 99, CDR 2, що містить амінокислотну послідовність, представлену в SEQ ID NО: 46, і CDR 3, що містить амінокислотну послідовність, представлену в SEQ ID NО: 60;
(11) CDR 1, що містить амінокислотну послідовність, представлену в SEQ ID NО: 100, CDR 2, що містить амінокислотну послідовність, представлену в SEQ ID NО: 46, і CDR 3, що містить амінокислотну послідовність, представлену в SEQ ID NО: 60;
(12) CDR 1, що містить амінокислотну послідовність, представлену в SEQ ID NО: 101, CDR 2, що містить амінокислотну послідовність, представлену в SEQ ID NО: 46, і CDR 3, що містить амінокислотну послідовність, представлену в SEQ ID NО: 60;
(13) CDR 1, що містить амінокислотну послідовність, представлену в SEQ ID NО: 102, CDR 2, що містить амінокислотну послідовність, представлену в SEQ ID NО: 46, і CDR 3, що містить амінокислотну послідовність, представлену в SEQ ID NО: 60;
(14) CDR 1, що містить амінокислотну послідовність, представлену в SEQ ID NО: 103, CDR 2, що містить амінокислотну послідовність, представлену в SEQ ID NО: 47, і CDR 3, що містить амінокислотну послідовність, представлену в SEQ ID NО: 60;
(15) CDR 1, що містить амінокислотну послідовність, представлену в SEQ ID NО: 104, CDR 2, що містить амінокислотну послідовність, представлену в SEQ ID NО: 47, і CDR 3, що містить амінокислотну послідовність, представлену в SEQ ID NО: 60.
9. Композиція анти-гліпікан 3-антитіл за п. 7, яка містить варіабельну ділянку легкого ланцюга, вибрану з (1) - (15):
(1) варіабельна ділянка легкого ланцюга, яка містить амінокислотну послідовність, представлену в SEQ ID NО: 107;
(2) варіабельна ділянка легкого ланцюга, яка містить амінокислотну послідовність, представлену в SEQ ID NО: 108;
(3) варіабельна ділянка легкого ланцюга, яка містить амінокислотну послідовність, представлену в SEQ ID NО: 109;
(4) варіабельна ділянка легкого ланцюга, яка містить амінокислотну послідовність, представлену в SEQ ID NО: 110;
(5) варіабельна ділянка легкого ланцюга, яка містить амінокислотну послідовність, представлену в SEQ ID NО: 111;
(6) варіабельна ділянка легкого ланцюга, яка містить амінокислотну послідовність, представлену в SEQ ID NО: 112;
(7) варіабельна ділянка легкого ланцюга, яка містить амінокислотну послідовність, представлену в SEQ ID NО: 113;
(8) варіабельна ділянка легкого ланцюга, яка містить амінокислотну послідовність, представлену в SEQ ID NО: 114;
(9) варіабельна ділянка легкого ланцюга, яка містить амінокислотну послідовність, представлену в SEQ ID NО: 115;
(10) варіабельна ділянка легкого ланцюга, яка містить амінокислотну послідовність, представлену в SEQ ID NО: 116;
(11) варіабельна ділянка легкого ланцюга, яка містить амінокислотну послідовність, представлену в SEQ ID NО: 117;
(12) варіабельна ділянка легкого ланцюга, яка містить амінокислотну послідовність, представлену в SEQ ID NО: 118;
(13) варіабельна ділянка легкого ланцюга, яка містить амінокислотну послідовність, представлену в SEQ ID NО: 119;
(14) варіабельна ділянка легкого ланцюга, яка містить амінокислотну послідовність, представлену в SEQ ID NО: 120; і
(15) варіабельна ділянка легкого ланцюга, яка містить амінокислотну послідовність, представлену в SEQ ID NО: 121.
10. Композиція анти-гліпікан 3-антитіл за будь-яким з пп. 1-9, де компонент, який представляє цукровий ланцюг, був модифікований так, щоб частка антитіл, позбавлених фукози, була більша ніж 20 %.
11. Композиція анти-гліпікан 3-антитіл за будь-яким з пп. 1-9, де компонент, який представляє цукровий ланцюг, був модифікований так, щоб частка антитіл, позбавлених фукози, була більша ніж 50 %.
12. Композиція анти-гліпікан 3-антитіл за будь-яким з пп. 1-9, де компонент, який представляє цукровий ланцюг, був модифікований так, щоб частка антитіл, позбавлених фукози, була більша ніж 90 %.
13. Спосіб одержання анти-гліпікан 3-антитіла з використанням клітини, де ген, який кодує анти-гліпікан 3-антитіло, визначене в будь-якому з пп. 1-9, був введений у вказану клітину, яка має зменшену здатність приєднання фукози до цукрових ланцюгів.
14. Спосіб отримання анти-гліпікан 3-антитіла п. 13, де клітина, яка має зменшену здатність приєднання фукози до цукрових ланцюгів, є клітиною, позбавленою транспортера фукози.
15. Спосіб отримання анти-гліпікан 3-антитіла, що включає стадії:
(а) введення гена, який кодує анти-гліпікан 3-антитіло, визначене в будь-якому з пп. 1-9, в клітину, яка має зменшену здатність приєднання фукози до цукрових ланцюгів; і
(b) культивування даної клітини.
16. Протираковий лікарський засіб, що має як активний інгредієнт композицію антитіл за будь-яким з пп. 1-12.
Текст
1. Композиція анти-гліпікан 3-антитіл, яка містить антитіло, що має варіабельну ділянку важкого ланцюга, яка містить CDR1, що має амінокислотну послідовність, представлену в SEQ ID NО: 25, CDR2, що має амінокислотну послідовність, представлену в SEQ ID NО: 26, і CDR3, що має амінокислотну послідовність, представлену в SEQ ID NО: 27, де компонент, який представляє цукровий ланцюг антитіла, був модифікований так, щоб мати підвищену частку антитіл, позбавлених фукози, або підвищену частку антитіл, що мають приєднаний дворозсікаючий N-ацетилглюкозамід (GlcNAc). 2. Композиція анти-гліпікан 3-антитіл, яка містить антитіло, що має варіабельну ділянку легкого ланцюга, яка містить CDR1, що має амінокислотну послідовність, представлену в SEQ ID NО: 45, CDR2, що має амінокислотну послідовність, представлену в SEQ ID NО: 46, і CDR3, що має амінокислотну послідовність, представлену в SEQ ID NО: 60, де компонент, який представляє цукровий ланцюг антитіла, був модифікований так, щоб мати підвищену частку антитіл, позбавлених фукози або підвищену частку антитіл, що мають приєднаний дворозсікаючий N-ацетилглюкозамід (GlcNAc). 3. Композиція анти-гліпікан 3-антитіл, в якій антитіло, визначене у п.1, додатково має варіабельну ділянку легкого ланцюга, яка містить CDR1, що має амінокислотну послідовність, представлену в SEQ ID NО: 45, CDR2, що має амінокислотну послідовність, представлену в SEQ ID NО: 46, і CDR3, що має амінокислотну послідовність, представлену в SEQ ID NО: 60. 4. Композиція анти-гліпікан 3-антитіл, яка містить антитіло, що має одну з перерахованих (1) - (7) варіабельних ділянок важкого ланцюга: UA (21) a200705812 (22) 26.10.2005 (24) 25.02.2011 (86) PCT/JP2005/020057, 26.10.2005 (31) 2004-311356 (32) 26.10.2004 (33) JP (46) 25.02.2011, Бюл.№ 4, 2011 р. (72) НАКАНО КІЙОТАКА, JP, СУГО ІЗУМІ, JP, СУГІМОТО МАСАМІТІ, JP, ІСІГУРО ТАКАХІРО, JP, ТАНАКА МЕГУМІ, JP, ІІДЗІМА СІГЕЮКІ, JP (73) ЧУГАІ СЕЙЯКУ КАБУСІКІ КАЙСЯ, JP (56) EP A1 1541680, 15.06.2005. EP A1 1411118, 21.04.2004. EP A1 1331266, 30.07.2003. EP A1 1176195, 30.01.2002. EP A1 1674111, 28.06.2006. EP A1 1411118, 21.04.2004. SHIELDS R L ET AL: "Lack of fucose on human IgG1 N-linked oligosaccharide improves binding to human FcgammaRIII and antibody-dependent cellular toxicity" JOURNAL OF BIOLOGICAL CHEMISTRY, AMERICAN SOCIETY OF BIOLOCHEMICAL BIOLOGISTS, BIRMINGHAM, US, vol. 277, no. 30, 26 July 2002 (2002-07-26), pages 26733-26740, XP002964542 ISSN: 0021-9258. SHINKAWA T ET AL: "The absence of fucose but not the presence of galactose or bisecting Nacetylglucosamine of human IgG1 complex-type oligosaccharides shows the critical role of enhancing antibody-dependent cellular cytotoxicity" JOURNAL OF BIOLOGICAL CHEMISTRY, AMERICAN SOCIETY OF BIOLOCHEMICAL BIOLOGISTS, BIRMINGHAM, US, vol. 278, no. 5, 31 January 2003 (2003-01-31), pages 3466-3473, XP002965857 ISSN: 0021-9258. NIWA RINPEI ET AL: "Defucosylated chimeric antiCC chemokine receptor 4 IgG1 with enhanced antibody-dependent cellular cytotoxicity shows potent therapeutic activity to T-cell leukemia and lymphoma" CANCER RESEARCH, AMERICAN ASSOCIATION FOR CANCER RESEARCH, BALTIMORE, MD., US, 2 (19) 1 3 (1) варіабельну ділянку важкого ланцюга, яка містить амінокислотну послідовність, представлену в SEQ ID NО: 81; (2) варіабельну ділянку важкого ланцюга, яка містить амінокислотну послідовність, представлену в SEQ ID NО: 82; (3) варіабельну ділянку важкого ланцюга, яка містить амінокислотну послідовність, представлену в SEQ ID NО: 83; (4) варіабельну ділянку важкого ланцюга, яка містить амінокислотну послідовність, представлену в SEQ ID NО: 84; (5) варіабельну ділянку важкого ланцюга, яка містить амінокислотну послідовність, представлену в SEQ ID NО: 85; (6) варіабельну ділянку важкого ланцюга, яка містить амінокислотну послідовність, представлену в SEQ ID NО: 86; або (7) варіабельну ділянку важкого ланцюга, яка містить амінокислотну послідовність, представлену в SEQ ID NО: 87, де компонент, який представляє цукровий ланцюг антитіла, був модифікований так, щоб мати підвищену частку антитіл, позбавлених фукози або підвищену частку антитіл, що мають приєднаний дворозсікаючий N-ацетилглюкозамід (GlcNAc). 5. Композиція анти-гліпікан 3-антитіл, яка містить антитіло, що має варіабельну ділянку легкого ланцюга, яка містить амінокислотну послідовність, представлену в SEQ ID NО: 109, де компонент, який представляє цукровий ланцюг антитіла, був модифікований так, щоб мати підвищену частку антитіл, позбавлених фукози або підвищену частку антитіл, що мають приєднаний дворозсікаючий Nацетилглюкозамід (GlcNAc). 6. Композиція анти-гліпікан 3-антитіл за п.4, яка додатково містить антитіло, що має варіабельну ділянку легкого ланцюга, яка містить амінокислотну послідовність, представлену в SEQ ID NО: 109. 7. Композиція анти-гліпікан 3-антитіл, яка має активність, еквівалентну активності композиції антитіл за п.3, де один або декілька амінокислотних залишків заміщені, делетовані або додані і/або вставлені з амінокислотних послідовностей, представлених в будь-якому з пп.1-6. 8. Композиція анти-гліпікан 3-антитіл за п.7, яка містить антитіло, що має варіабельну ділянку легкого ланцюга, яка містить одне з перерахованих (1) - (15) CDR1, 2 і 3: (1) CDR 1, що містить амінокислотну послідовність, представлену в SEQ ID NО: 90, CDR 2, що містить амінокислотну послідовність, представлену в SEQ ID NО: 46, і CDR 3, що містить амінокислотну послідовність, представлену в SEQ ID NО: 60; (2) CDR 1, що містить амінокислотну послідовність, представлену в SEQ ID NО: 91, CDR 2, що містить амінокислотну послідовність, представлену в SEQ ID NО: 46, і CDR 3, що містить амінокислотну послідовність, представлену в SEQ ID NО: 60; (3) CDR 1, що містить амінокислотну послідовність, представлену в SEQ ID NО: 92, CDR 2, що містить амінокислотну послідовність, представлену в SEQ ID NО: 46, і CDR 3, що містить амінокис 93488 4 лотну послідовність, представлену в SEQ ID NО: 60; (4) CDR 1, що містить амінокислотну послідовність, представлену в SEQ ID NО: 93, CDR 2, що містить амінокислотну послідовність, представлену в SEQ ID NО: 46, і CDR 3, що містить амінокислотну послідовність, представлену в SEQ ID NО: 60; (5) CDR 1, що містить амінокислотну послідовність, представлену в SEQ ID NО: 94, CDR 2, що містить амінокислотну послідовність, представлену в SEQ ID NО: 46, і CDR 3, що містить амінокислотну послідовність, представлену в SEQ ID NО: 60; (6) CDR 1, що містить амінокислотну послідовність, представлену в SEQ ID NО: 95, CDR 2, що містить амінокислотну послідовність, представлену в SEQ ID NО: 46, і CDR 3, що містить амінокислотну послідовність, представлену в SEQ ID NО: 60; (7) CDR 1, що містить амінокислотну послідовність, представлену в SEQ ID NО: 96, CDR 2, що містить амінокислотну послідовність, представлену в SEQ ID NО: 46, і CDR 3, що містить амінокислотну послідовність, представлену в SEQ ID NО: 60; (8) CDR 1, що містить амінокислотну послідовність, представлену в SEQ ID NО: 97, CDR 2, що містить амінокислотну послідовність, представлену в SEQ ID NО: 46, і CDR 3, що містить амінокислотну послідовність, представлену в SEQ ID NО: 60; (9) CDR 1, що містить амінокислотну послідовність, представлену в SEQ ID NО: 98, CDR 2, що містить амінокислотну послідовність, представлену в SEQ ID NО: 46, і CDR 3, що містить амінокислотну послідовність, представлену в SEQ ID NО: 60; (10) CDR 1, що містить амінокислотну послідовність, представлену в SEQ ID NО: 99, CDR 2, що містить амінокислотну послідовність, представлену в SEQ ID NО: 46, і CDR 3, що містить амінокислотну послідовність, представлену в SEQ ID NО: 60; (11) CDR 1, що містить амінокислотну послідовність, представлену в SEQ ID NО: 100, CDR 2, що містить амінокислотну послідовність, представлену в SEQ ID NО: 46, і CDR 3, що містить амінокислотну послідовність, представлену в SEQ ID NО: 60; (12) CDR 1, що містить амінокислотну послідовність, представлену в SEQ ID NО: 101, CDR 2, що містить амінокислотну послідовність, представлену в SEQ ID NО: 46, і CDR 3, що містить амінокислотну послідовність, представлену в SEQ ID NО: 60; (13) CDR 1, що містить амінокислотну послідовність, представлену в SEQ ID NО: 102, CDR 2, що містить амінокислотну послідовність, представлену в SEQ ID NО: 46, і CDR 3, що містить амінокислотну послідовність, представлену в SEQ ID NО: 60; (14) CDR 1, що містить амінокислотну послідовність, представлену в SEQ ID NО: 103, CDR 2, що містить амінокислотну послідовність, представле 5 93488 6 ну в SEQ ID NО: 47, і CDR 3, що містить амінокислотну послідовність, представлену в SEQ ID NО:60; (15) CDR 1, що містить амінокислотну послідовність, представлену в SEQ ID NО: 104, CDR 2, що містить амінокислотну послідовність, представлену в SEQ ID NО: 47, і CDR 3, що містить амінокислотну послідовність, представлену в SEQ ID NО: 60. 9. Композиція анти-гліпікан 3-антитіл за п.7, яка містить варіабельну ділянку легкого ланцюга, вибрану з (1) - (15): (1) варіабельна ділянка легкого ланцюга, яка містить амінокислотну послідовність, представлену в SEQ ID NО: 107; (2) варіабельна ділянка легкого ланцюга, яка містить амінокислотну послідовність, представлену в SEQ ID NО: 108; (3) варіабельна ділянка легкого ланцюга, яка містить амінокислотну послідовність, представлену в SEQ ID NО: 109; (4) варіабельна ділянка легкого ланцюга, яка містить амінокислотну послідовність, представлену в SEQ ID NО: 110; (5) варіабельна ділянка легкого ланцюга, яка містить амінокислотну послідовність, представлену в SEQ ID NО: 111; (6) варіабельна ділянка легкого ланцюга, яка містить амінокислотну послідовність, представлену в SEQ ID NО: 112; (7) варіабельна ділянка легкого ланцюга, яка містить амінокислотну послідовність, представлену в SEQ ID NО: 113; (8) варіабельна ділянка легкого ланцюга, яка містить амінокислотну послідовність, представлену в SEQ ID NО: 114; (9) варіабельна ділянка легкого ланцюга, яка містить амінокислотну послідовність, представлену в SEQ ID NО: 115; (10) варіабельна ділянка легкого ланцюга, яка містить амінокислотну послідовність, представлену в SEQ ID NО: 116; (11) варіабельна ділянка легкого ланцюга, яка містить амінокислотну послідовність, представлену в SEQ ID NО: 117; (12) варіабельна ділянка легкого ланцюга, яка міс тить амінокислотну послідовність, представлену в SEQ ID NО: 118; (13) варіабельна ділянка легкого ланцюга, яка містить амінокислотну послідовність, представлену в SEQ ID NО: 119; (14) варіабельна ділянка легкого ланцюга, яка містить амінокислотну послідовність, представлену в SEQ ID NО: 120; і (15) варіабельна ділянка легкого ланцюга, яка містить амінокислотну послідовність, представлену в SEQ ID NО: 121. 10. Композиція анти-гліпікан 3-антитіл за будьяким з пп.1-9, де компонент, який представляє цукровий ланцюг, був модифікований так, щоб частка антитіл, позбавлених фукози, була більша ніж 20 %. 11. Композиція анти-гліпікан 3-антитіл за будьяким з пп.1-9, де компонент, який представляє цукровий ланцюг, був модифікований так, щоб частка антитіл, позбавлених фукози, була більша ніж 50%. 12. Композиція анти-гліпікан 3-антитіл за будьяким з пп.1-9, де компонент, який представляє цукровий ланцюг, був модифікований так, щоб частка антитіл, позбавлених фукози, була більша ніж 90%. 13. Спосіб одержання анти-гліпікан 3-антитіла з використанням клітини, де ген, який кодує антигліпікан 3-антитіло, визначене в будь-якому з пп.19, був введений у вказану клітину, яка має зменшену здатність приєднання фукози до цукрових ланцюгів. 14. Спосіб отримання анти-гліпікан 3-антитіла п.13, де клітина, яка має зменшену здатність приєднання фукози до цукрових ланцюгів, є клітиною, позбавленою транспортера фукози. 15. Спосіб отримання анти-гліпікан 3-антитіла, що включає стадії: (а) введення гена, який кодує анти-гліпікан 3антитіло, визначене в будь-якому з пп.1-9, в клітину, яка має зменшену здатність приєднання фукози до цукрових ланцюгів; і (b) культивування даної клітини. 16. Протираковий лікарський засіб, що має як активний інгредієнт композицію антитіл за будь-яким з пп.1-12. Даний винахід належить до антитіла до антигену гліпікану 3 (тобто анти-гліпікан 3-антитіла), в якому цитотоксична активність, зокрема антитілозалежна клітинноопосередкована цитотоксичність (ADCC), є посиленою, а також до способу одержання цього антитіла. Гліпікан 3 (GPC3) є членом сімейства гепарансульфат-протеогліканів, присутніх на поверхні клітин. Було передбачено, що GPC3 може брати участь у клітинному поділі при розвитку і рості ракових клітин. Було виявлено, що тип антитіла, який зв'язується з GPC3, інгібує ріст клітин внаслідок активності ADCC (антитілозалежної клітинноопосередкованої цитотоксичності) і активності CDC (комплементзалежної цитотоксичності) (WO 2003/00883). Крім того, оскільки GPC3 розщеплюється в тілі і секретується в кров у вигляді розчинної форми GPC3, було зроблене припущення, що рак може діагностуватися з використанням антитіла, яке може детектувати розчинну форму GPC3 (WO 2004/022739, WO 2003/100429, WO 2004/018667). 7 При розвитку протиракового агента, який використовує цитотоксичну активність антитіла, використовуване антитіло переважно буде мати високий рівень активності ADCC. Таким чином, було потрібне анти-GPC3-антитіло з високим рівнем цитотоксичної активності. Відомо, що модифікація цукрових ланцюгів антитіла посилює його ADCC-активність. Наприклад, в WO 99/54342 описано, що ADCC-активність посилюється модифікацією глікозилування антитіла. Крім того, в WO 00/61739 описано, що ADCCактивність регулюється контролюванням присутності або відсутності фукози в цукрових ланцюгах антитіла. У WO 02/31140 описане одержання антитіла, що має цукрові ланцюги, які не містять 1,6-фукози основної структури (кора) цукрового ланцюга, продукуванням цього антитіла в клітинах YB2/0. В WO 02/79255 описане антитіло з цукровими ланцюгами, що мають дворозсічений GlcNAc. Однак, анти-GРС3-антитіло з підвищеною активністю ADCC внаслідок модифікації цукрового ланцюга досі не було описане. Даний винахід забезпечує композицію антиGРС3-антитіла з підвищеною ADCC-активністю, зумовленою зміною його компонента, що представляє цукровий ланцюг, а також спосіб одержання такого антитіла. Після численних досліджень автори даного винаходу виявили, що антитіла проти GPC3 з цукровими ланцюгами, позбавленими -1,6-фукози основної структури (кора), мають високий рівень цитотоксичної активності. Таким чином, даний винахід забезпечує композицію анти-GРС3-антитіл, в яких компонент антитіла, що представляє цукровий ланцюг, був змінений, і, більш конкретно, композицію антитіл з більшою фракцією фукозанедостатніх анти-GРС3-антитіл. Композиція анти-GРС3антитіл з модифікованим цукровим ланцюгом даного винаходу має високий рівень цитотоксичної активності і, отже, застосовна як інгібітор росту клітин, такий як протираковий агент. Даний винахід забезпечує також спосіб одержання композиції анти-GРС3-антитіл, в яких цукровий ланцюгантитіла є модифікованим, який передбачає стадії: введення нуклеїнової кислоти, яка кодує анти-GРС3-антитіло, в клітину-хазяїна із зменшеною здатністю приєднання фукози, таку як клітини YB2/0, і культивування цієї клітини-хазяїна для одержання цього антитіла. Переважно, клітиною із зменшеною здатністю приєднання фукози до цукрових ланцюгів є клітина, позбавлена транспортера фукози. Фіг.1 показує основну структуру Nглікозидзв'язаних цукрових ланцюгів; Фіг.2 показує активність ADCC химерного антитіла, коли мішенями є клітини HepG2, з використанням моноцитів периферійної крові людини (РВМС); Фіг.3 показує активність ADCC химерного антитіла, коли мішенню є клітини HuH-7, з використанням РВМС людини; Фіг.4 показує активність ADCC антитіл, коли мішенями є клітини HuH-7, з використанням РВМС людини; 93488 8 Фіг.5 показує хроматограми ВРХ з нормальною фазою цукрових ланцюгів, модифікованих агалактозил-2-АВ, одержаних з антитіл (а, b, с), що продукуються клітинами FT-KO і клітинами СНО; Фіг.6 показує передбачені структури для піків G(0)- і G(0)-Fuc, показаних на Фіг.5; і Фіг.7 показує графік вимірювань диференціальної сканувальної калориметрії (DSC) для антитіл, що продукуються клітинами FT-KO (а) і клітинами СНО (b). Докладний опис винаходу Даний винахід описує композицію анти-GРС3антитіл, в яких компонент, що представляє цукровий ланцюг, цього антитіла був модифікований. Відомо, що структура цукрового ланцюга, зв'язаного з антитілом, суттєво впливає на виявлення цитотоксичної активності антитіла. Цукрові ланцюги, які зв'язані з антитілом, включають в себе Nглікозидзв'язані цукрові ланцюги, які приєднані до атома азоту на бічному ланцюгу залишку аспарагіну, і О-глікозидзв'язані цукрові ланцюги, які приєднані до гідроксильної групи на бічному ланцюгу залишку серину або треоніну на молекулі антитіла. Даний винахід сфокусований на присутності або відсутності фукози в N-глікозидзв'язаному цукровому ланцюгу. Фіг.1 показує основну структуру Nглікозидзв'язаних цукрових ланцюгів, приєднаних до антитіла. Як показано в основних цукрових ланцюгах IgG (1) і (3) Фіг.1, N-глікозидзв'язані цукрові ланцюги мають основну структуру (кор), в якій один залишок манози (Man) і два залишки Nацетилглюкозаміну (GlcNAc) зв'язані Р-1,4зв'язками [-Man 1-4GlcNAc -1-4GlcNAc-]. «GlcNAc» на правому боці цієї структури називають відновним кінцем, a "Man" на лівому боці цієї структури називають невідновним кінцем. Коли фукоза зв'язана з відновним кінцем, вона звичайно приймає форму α-зв'язку між положенням 6 Nацетилглюкозаміну на відновному кінці і положенням 1 фукози. З іншого боку, в цукровому ланцюгу, показаному в основному цукровому ланцюгу IgG (2) Фіг.1, нарівні з вищезазначеними двома цукровими ланцюгами, N-ацетилглюкозамін (GlcNAc) зв'язаний з невідновним кінцем основної структури (кора) через -1,4-зв'язок. Цей тип Nацетилглюкозаміну (GlcNAc) називають «дворозсікальним N-ацетилглкжозаміном». Цукровим ланцюгом, що має дворозсікальний Nацетилглюкозамін, може бути О-глікозидзв'язаний цукровий ланцюг або N-глікозидзв'язаний цукровий ланцюг, і він утворюється перенесенням Nацетилглюкозаміну до цукрового ланцюга Nацетилглюкозамінтрансферазою III (GnTIII). Ген, що кодує цей фермент, вже був клонований, і повідомлялися як амінокислотна послідовність, так і нуклеотидна послідовність ДНК, яка кодує цей фермент (База даних NCBI (ACCESSION D13789)). У даному винаході композицією антитіл з модифікованим або зміненим компонентом, який представляє цукровий ланцюг (композицією антитіл з модифікованим цукровим ланцюгом), називають композицію антитіл, які мають компонент, що представляє цукровий ланцюг, який відрізня 9 ється від композиції антитіл, що продукуються клітиною-хазяїном, яка є референсним стандартом. У даному винаході можна визначити, був або не був компонент, що представляє цукровий ланцюг, модифікований, з використанням як референсного стандарту композиції антитіл, що продукуються клітиною-хазяїном, яка є референсним стандартом. Якщо композиція антитіл має компонент, що представляє цукровий ланцюг, відмінний від композиції антитіл з референсного стандарту, то цю композицію антитіл вважають композицією антитіл з модифікованим компонентом, який представляє цукровий ланцюг. Клітиною-хазяїном, яка служить посилальним стандартом, в даному винаході є клітина СНО DG44. Клітка СНО DG44 може бути одержана, наприклад, з Invitrogen Corporation. Приклади композиції антитіл з модифікованим компонентом, що представляє цукровий ланцюг, включають в себе, наприклад, композицію антитіл із збільшеною часткою фукозанедостатніх (наприклад, недостатніх відносно -1,6-фукози кора (основної структури)) антитіл в цій композиції антитіл, і композицію антитіл із збільшеною часткою антитіл, що мають приєднаний дворозсікальний Nацетилглюкозамін (GlcNAc) в цій композиції антитіл. У переважному варіанті здійснення даного винаходу, ця композиція антитіл має більш високу частку фукозанедостатніх антитіл, ніж композиція антитіл, яка використовується як референсний стандарт. Оскільки деякі антитіла мають множину Nглікозидних цукрових ланцюгів, фукозанедостатнє антитіло даного винаходу включає в себе не тільки антитіла, в яких фукоза не приєднана, але також антитіла, в яких кількість фукозних залишків, приєднаних до антитіла, є зменшеною (антитіла, що мають щонайменше один або більше цукрових ланцюгів, в яких не присутня фукоза). При одержанні антитіла з модифікованим цукровим ланцюгом з використанням клітин-хазяїнів часто важко одержати композицію, що містить однорідні антитіла, в якій всі антитіла мають ідентичні цукрові ланцюги. Таким чином, якщо композиція антитіл з модифікованим компонентом, який представляє цукровий ланцюг, є, наприклад, композицією антитіл із збільшеною часткою фукозанедостатніх антитіл, то ця композиція антитіл з модифікованим компонентом, який представляє цукровий ланцюг, даного винаходу може містити як антитіла, недостатні у відношенні фукози, так і антитіла, що не є недостатніми у відношенні фукози, але загальна частка антитіл, недостатніх у відношенні фукози, буде більш високою, ніж в композиції антитіл, що продукуються клітинамихазяїнами, що служать як референсний стандарт. Даний винахід особливо не обмежується щодо конкретної частки фукозанедостатніх антитіл в композиції антитіл з високою часткою фукозанедостатніх антитіл даного винаходу, але переважно ця частка дорівнює не менше ніж 20%, більш переважно не менше ніж 50% і найбільш переважно не менше ніж 90%. 93488 10 Даний винахід особливо не обмежується щодо конкретної частки антитіл з доданим дворозсікальним N-ацетилглюкозаміном в композиції антитіл, що мають високу частку антитіл з доданим дворозсікальним N-ацетилглюкозаміном, але переважно ця частка дорівнює не менше ніж 20%, більш переважно не менше ніж 50% і найбільш переважно не менше ніж 90%. Композиція анти-GРС3-антитіл з модифікованим компонентом, який представляє цукровий ланцюг, даного винаходу може бути одержана способами, відомими кваліфікованому фахівцеві в даній галузі. Наприклад, фукозанедостатнє антитіло може бути одержане експресією анти-GРС3-антитіла в клітинах-хазяїнах, які або позбавлені здатності, або мають низьку здатність приєднувати -1,6фукозу кора (основної структури). Даний винахід особливо не обмежується щодо клітин-хазяїнів, які позбавлені здатності або мають низьку здатність приєднувати фукозу, але клітинахазяїн без активності фукозотрансферази або із зменшеною активністю фукозотрансферази, клітина-хазяїн з більш низькою концентрацією фукози в тільцях Гольджі, і т. п.можуть бути використані в даному винаході. Більш конкретно, приклади клітин-хазяїнів включають в себе клітини мієломи щура YB2/3HL.P2.G11.16Ag.2O (скорочено YB2/0) (що зберігаються як АТСС CRL 1662), клітини СНО з нокаутом FTVIII (WO 02/31140), клітини Lec13 (WO 03/035835) і недостатні відносно транспортера фукози клітини (WO 2005/017155). У даному контексті, термін «недостатні відносно транспортера фукози клітини» належить до клітини, в якій кількість транспортера фукози в клітині менше, ніж в нормальних клітинах, або функція транспортера фукози є ослабленою внаслідок відхилення від норми в структурі транспортера фукози. Приклади клітин, недостатніх відносно транспортера фукози, включають в себе, наприклад, клітини, в яких зруйнований ген транспортера фукози (тобто клітини з нокаутом цього гена) (далі звані клітинами FT-KO), клітини, в яких або відсутня, або є мутованою частина гена транспортера фукози, клітини, недостатні у відношенні системи експресії гена транспортера фукози, і т. п.Нуклеотидна послідовність гена, який кодує транспортер фукози Китайського хом'ячка, і його амінокислотна послідовність показані в SEQ ID NO:126 і 127, відповідно. Крім того, можна одержати недостатню відносно транспортера фукози клітину даного винаходу з використанням інтерференції РНК (RNAi) із застосуванням нуклеотидної послідовності, представленої SEQ ID NO:126. RNAi являє собою наступний феномен: при введенні дволанцюгової РНК (dsRNA) в клітину внутрішньоклітинна мРНК, відповідна цій послідовності РНК, специфічно руйнується і не може експресуватися у вигляді білка. Звичайно dsRNA використовується з RNAi, але даний винахід не обмежується цим, і, наприклад, може бути використана також дволанцюгова РНК, утворена самокомплементарними одноланцюговими молекулами РНК. Що стосується районів, які утворюють дволанцюгову молекулу, ця молекула 11 може бути дволанцюговою у всіх районах або може бути одноланцюговою в деяких районах (наприклад, в одному або обох кінцях). Даний винахід не обмежується конкретною довжиною оліго-РНК, використовуваного в RNAi. Довжина оліго-РНК в даному винаході може бути рівною, наприклад, 51000 основам (або 5-1000 пар основ в дволанцюговій молекулі), переважно, 10-100 основам (або 10-100 парам основ в дволанцюговій молекулі) і, найбільш переважно, 15-25 основам (15-25 парам основ в дволанцюговій молекулі); однак, особливо переважною є довжина 19-23 основи (19-23 пари основ в дволанцюговій молекулі). Вищезазначений процес RNAi використовує феномен, в якому dsRNA, що складається як із смислової РНК, так і антисмислової РНК, гомологічних конкретному гену, буде руйнувати гомологічну частину транскрипту (мРНК) цього гена. Може бути використана dsRNA, відповідна всій послідовності гена транспортера фукози, або більш коротка dsRNA (наприклад, 21-23 п.н.), відповідна частині цієї послідовності (мала інтерферуюча РНК; siRNA). dsRNA може бути безпосередньо перенесена в клітину, або може бути одержаний вектор, який продукує dsRNA, і перенесений в клітинухазяїна, і потім ця dsRNA може бути продукована в цій клітині. Наприклад, вся ДНК або частина ДНК фукози, що кодує транспортер, може бути вбудована у вектор таким чином, що вона утворює послідовність оберненого повтору, і цей вектор може бути потім перенесений в клітину-хазяїна. Процедура RNAi може проводитися відповідно до описів в наступних посиланнях: Fire A. et al., Nature (1998), 391, 806-811; Montgomery Μ. Κ. et al., Proc. Natl. Acad. Sci. USA (1998), 95, 15502-15507; Timmons L. et al., Nature (1998), 395, 854; Sanchez A. et al., Proc. Natl. Acad. Sci. USA (1999), 96, 50495054; Misquitta L. et al., Proc. Natl. Acad. Sci. USA (1999), 96, 1451-1456; Kennerdell J. R. et al. Cell (1998), 95, 1017-1026; Waterhouse P. M. et al., Proc. Natl. Acad. Sci. USA (1998), 95 13959-13964; and Wianny F. et al. Nature Cell Biol. (2000), 2, 70-75. Недостатні відносно транспортера фукози клітини, одержані процедурою RNAi, можуть бути піддані скринінгу, як указано, за допомогою активності транспортера фукози. Скринінг може також проводитися на основі транскрипції і експресії гена транспортера фукози, з використанням Вестернблотингу або Нозерн-блотингу. Антитіло з дворозсікальним Nацетилглюкозаміном (GlcNAc), доданим до цукрового ланцюга, може бути одержане експресією анти-GРС3-антитіла в клітині-хазяїні, здатній утворювати структуру дворозсікального Nацетилглюкозаміну (GlcNAc) на цукровому ланцюгу. Спосіб одержання антитіла з цукровим ланцюгом, який має доданий N-ацетилглюкозамін, вже 93488 12 відомий (WO 02/79255). Клітина-хазяїн, здатна утворювати структуру дворозсікального Nацетилглюкозаміну (GlcNAc) на цукровому ланцюгу, особливо не обмежується в даному винаході, але може включати в себе, наприклад, клітинухазяїна, яка має експресуючий вектор, що містить ДНК, яка кодує GnTIII. Таким чином, анти-GРС3антитіло, що має цукровий ланцюг з доданим дворозсікальним N-ацетилглюкозаміном, може бути одержане з використанням клітини-хазяїна, яка містить як експресуючий вектор з ДНК, що кодує Gnlll, так і експресуючий вектор, що кодує антиGРС3-антитіло. Як ДНК, що кодує Gnlll, так і ген, що кодує анти-GРС3-антитіло, можуть бути присутніми на одному і тому ж векторі або можуть бути присутніми на різних векторах. Іншим способом збільшення частки композиції фукозанедостатніх антитіл або антитіл з приєднаним дворозсікальним N-ацетилглюкозаміном в цій композиції антитіл є збільшення частки цих антитіл в композиції очищенням фукозанедостатніх антитіл або антитіл з приєднаним дворозсікальним Nацетилглюкозаміном. Аналіз цукрових ланцюгів може проводитися будь-якими способами, відомими кваліфікованим в даній галузі фахівцям. Наприклад, цукровий ланцюг може бути вивільнений з антитіла реакцією цього антитіла з N-глікозидазою F (Roche) і т. п.Потім ці цукрові ланцюги можуть бути знесолені твердофазною екстракцією з використанням целюлозного картриджа (Shimizu Y. et al., Carbohydrate Research 332 (2001), 381-388), сконцентровані і висушені, і помічені флуоресцентно 2амінопіридином (Kondo A. et al., Agricultural and Biological Chemistry 54:8 (1990), 2169-2170). Реагент видаляють з піридиламіносахарних ланцюгів (ΡΑ-цукрових ланцюгів) твердофазною екстракцією з використанням целюлозного картриджа, потім цукрові ланцюги концентрують центрифугуванням з одержанням очищених ΡΑ-цукрових ланцюгів. Ці цукрові ланцюги можуть аналізуватися ВРХ із оберненою фазою з використанням колонки з октадецилсиланом (ODS). Одержані таким чином ΡΑцукровіланцюги можуть аналізуватися двомірним картуванням з використанням комбінування ВРХаналізу із оберненою фазою з ODS-колонкою і ВРХ-аналізу з нормальною фазою з аміноколонкою. Анти-GРС3-антитіло з модифікованим цукровим ланцюгом даного винаходу че обмежується якими-небудь конкретними антитілами, при умові, що воно зв'язується з GPC3. Переважно, зв'язування з GPC3 може бути специфічним. Переважні анти-GРС3-антитіла даного винаходу включають в себе антитіла, які мають послідовність району (CDR), що визначає компліментарність, показану в таблиці 1 нижче. 13 93488 14 15 93488 16 17 93488 1) Антитіло, 2) Амінокислотна послідовність 18 19 Антитіла з послідовністю CDR зі списку, наведеного в таблиці вище, мають високий рівень цитотоксичної активності. Антитіла з послідовністю CDR зі списку, наведеного в таблиці вище, упізнають епітопи амінокислот 524-563 на GPC3. Оскільки антитіла, які упізнають епітопи амінокислот 524563, мають високий рівень цитотоксичної активності, вони є переважними як анти-GРС3-антитіла даного винаходу. У одному переважному варіанті здійснення даного винаходу, композиція антитіл, які мають модифікований компонент, що представляє цукровий ланцюг, даного винаходу, характеризується виявом підвищеної активності ADCC. У даному винаході присутність або відсутність підвищеної активності ADCC можна визначити порівнянням активності ADCC композиції антитіл даного винаходу з активністю ADCC композиції антитіл референсного стандарту. Якщо композиція антитіл даного винаходу виявляє більш високу активність ADCC, ніж референсний стандарт, то вважають, що активність ADCC є підвищеною. Активність ADCC може бути виміряна способом, відомим кваліфікованим в даній галузі фахівцям, наприклад, змішуванням анти-GPC3-антитіла з ефекторними клітинами і клітинами-мішенями і потім визначенням рівня ADCC. Більш конкретно, клітини селезінки миші, моноцити людини, виділені з периферійної крові (РВМС) і кісткового мозку і т. п., можуть бути використані як ефекторні клітини, а клітини людини, експресуючі GPC3, такі як лінія клітин НuН-7 печінково-клітинного раку людини, можуть бути використані як клітини-мішені. Спочатку ці клітини-мішені мітять 51Cr, додають антиGPC3-антитіло, клітини інкубують і потім додають ефекторні клітини у придатному співвідношенні з клітинами-мішенями і їх інкубують разом. Після інкубації супернатант збирають і активність ADCC вимірюють рахуванням радіоактивності в цьому супернатанті. Анти-GPC3-антитіло Анти-GPC3-антитіло може бути одержане способом, відомим кваліфікованим в даній галузі фахівцям. Наприклад, це антитіло може бути одержане з використанням GPC3 як сенсибілізуючого антигену для імунізації відповідно до звичайного способу імунізації, злиттям цих імунних клітин з відомими вихідними клітинами за допомогою звичайної процедури злиття клітин і скринінгом на продукуюче моноклональне антитіло клітин за допомогою звичайного способу скринінгу. Більш конкретно, моноклональні антитіла можуть бути одержані таким чином. Спочатку GPC3 для використання як сенсибілізуючого антигену одержують експресією GPC3 (MXR7) на основі послідовності гена/амінокислотної послідовності, описаних в Lage, Η. et al., Gene 188 (1997), 151-156. Іншими словами, послідовність гена, яка кодує GPC3, вбудовують у відомий експресуючий вектор. Після трансформації придатних клітинхазяїнів цим вектором білок-мішень гліпікан 3 людини очищають з клітин-хазяїнів або супернатанту культурального середовища відомим способом. Потім очищений білок GPC3 використовують як 93488 20 сенсибілізуючий антиген. Альтернативно, як сенсибілізуючий антиген може бути використаний частковий пептид GPC3. У такому способі цей частковий пептид може бути одержаний хімічним синтезом відповідно до амінокислотної послідовності GPC3 людини. Епітопи на молекулі GPC3, упізнавані анти-GPC3-антитілом даного винаходу, не обмежуються, і анти-GPC3-антитіло даного винаходу може упізнавати будь-який епітоп, присутній на молекулі GPC3. Це пояснюється тим, що анти-GPC3-антитіло виявляє інгібуючу ріст клітин активність через його ADCC-активність, CDCактивність або інгібування активності фактора росту, і тим, що ріст клітин також інгібується дією цитотоксичної речовини, такої як радіоактивний ізотоп, хіміотерапевтичний лікарський засіб, бактеріальний токсин, приєднаний до анти-GPC3антитіла. Таким чином, антигеном для одержання анти-GPC3-антитіла даного винаходу може бути будь-який фрагмент GPC3, при умові, що він містить епітоп, присутній на молекулі GPC3. У особливо переважному варіанті здійснення, як сенсибілізуючий антиген для генерування антитіла, яке упізнає епітоп з амінокислот 524-563 GPC3, може бути використаний пептид, який містить амінокислоти 524-563. Ссавець, який використовується для імунізації сенсибілізуючим антигеном, особливо не обмежується в даному винаході, але переважно він повинен бути вибраний з урахуванням сумісності з вихідними клітинами, які повинні бути використані в злитті клітин, і може включати в себе гризуна, наприклад, мишу, щура або хом'яка, або кролика, мавпу і т. п.Ця тварина може бути імунізована сенсибілізуючим антигеном з використанням відомого способу. Звичайно, наприклад, ссавець може бути ін'єктований внутрішньочеревинно або підшкірно сенсибілізуючим антигеном. Більш конкретно, сенсибілізуючий антиген може бути розбавлений і суспендований у придатній кількості забуференого фосфатом сольового розчину (ЗФР) або фізіологічного сольового розчину, змішаний з придатною кількістю загальноприйнятого ад'юванту, такого як повний ад'ювант Фрейнда, якщо бажано, емульгований і введений ссавцеві багато разів кожні 4 дні - 21 день. Крім того, при імунізації сенсибілізуючим антигеном може бути використаний придатний носій. Після імунізації ссавця описаним вище чином і детектування рівня бажаного антитіла в сироватці імунні клітини збирають з цього ссавця і піддають клітинному злиттю. Клітини селезінки є особливо переважними імунними клітинами для злиття клітин. Клітини мієломи ссавців використовують як вихідні клітини-партнери для злиття з імунними клітинами. Відомі клітинні лінії, придатні для використання як клітини мієломи, включають в себе Р3 (P3x63Ag8.653) (J. Immnol. (1979) 123, 1548-1550), P3x63Ag8U.1 (Current Topics in Microbiology and Immunology (1978) 81, 1-7), NS-1 (Kohler. G. and Milstein, С Eur. J. Immunol. (1976) 6, 511-519), MPC11 (Margulies. D.H. et al., Cell (1976) 8, 405-415), SP2/0 (Shulman, M. et al., Nature (1978) 276, 269270), FO (de St. Groth, S. F. et al., J. Immunol. 21 Methods (1980) 35, 1-21), S194 (Trowbridge, I. S. J. Exp. Med. (1978) 148, 313-323), R210 (Galfre, G. et al., Nature (1979) 277, 131-133). Злиття імунних клітин і клітин мієломи в основному може проводитися відповідно до відомого способу, наприклад, способом, описаним Kohler and Milstein (Kohler, G. And Milstein, З., Methods Enzymol. (1981) 73, 3-46). Більш конкретно, злиття клітин може проводитися, наприклад, в загальноприйнятому рідкому культуральному середовищі, яке містить стимулятор злиття клітин. Приклади стимулюючих злиття клітин хімічних речовин включають в себе поліетиленгліколь (PEG) і вірус Сендай (HVJ) і т. п.Якщо бажано, для збільшення ефективності злиття може бути доданий додатковий агент, такий як диметилсульфоксид і т. п.Відношення імунних клітин до клітин мієломи може бути встановлено довільно. Наприклад, переважним є встановлення відношення імунних клітин до клітин мієломи 1:1 10:1. Загальноприйняте рідке культуральне середовище, що використовується для культивування цих типів клітин, таке як рідке середовище RPMI-1640, рідке середовище MEM або інше рідке середовище, придатне для росту лінії клітин мієломи, може бути використане як рідке середовище в процедурі злиття клітин. Разом з середовищем може бути також використана сироваткова добавка, така як фетальна теляча сироватка. У процедурі злиття клітин вказані кількості імунних клітин і мієломних клітин ретельно змішують в рідкому культуральному середовищі і потім розчин PEG (наприклад, з середньою молекулярною масою приблизно 1000-6000), який був нагрітий до 37°С, додають звичайно при концентрації 30-60% (маса/об'єм) і перемішують для створення можливості утворення злитих клітин (гібридом). Потім додають відповідне рідке середовище і центрифугують для видалення супернатанту. За допомогою повторення цієї процедури видаляють будь-які хімікалії, використані для клітинного злиття, несприятливі для росту гібридом. Одержані таким чином гібридоми відбирають культивуванням їх в загальноприйнятому рідкому селективному середовищі, такому як середовище HAT (культуральне середовище, що містить гіпоксантин, аміноптерин і тимідин). Культивування в середовищі HAT продовжують протягом достатнього періоду часу (звичайно протягом декількох днів - декількох тижнів), поки клітини, інші, ніж гібридоми-мішені (незлиті клітини), не загинуть. Потім, проводять загальноприйняту процедуру лімітуючого розведення з подальшим скринінгом і моноклонуванням гібридом, які продукують антитіло-мішень. Нарівні з імунізацією тварини, що не є людиною, антигеном для одержання гібридом, як описано вище, бажані антитіла людини, що мають GPC3-зв'язуючу активність, можуть бути одержані сенсибілізацією лімфоцитів людини GPC3 in vitro і потім злиттям цих сенсибілізованих лімфоцитів з імморталізованими клітинами мієломи людини (див. Публікацію патенту Японії з номером Н159878). Крім того, можна ввести GPC3 як антиген трансгенній тварині, що має повний репертуар генів антитіл людини, для генерування клітин, які продукують анти-GPC3-антитіла, і зібрати антитіла 93488 22 до GPC3 людини з імморталізованих клітин (див. Міжнародні заявки на патент з номерами WO 94/25585, WO 93/12227, WO 92/03918 і WO 94/02602). Гібридоми, що продукують моноклональні антитіла, одержані вищезгаданим чином, можуть бути субкультивовані в загальноприйнятому рідкому культуральному середовищі і можуть зберігатися протягом тривалого періоду часу в рідкому азоті. Рекомбінантні антитіла Моноклональне антитіло, використане в даному винаході, є рекомбінантним моноклональним антитілом, яке може бути одержане клонуванням гена цього антитіла з гібридоми, вбудовуванням цього гена у придатний вектор і інтеграцією цього вектора в клітину-хазяїна (наприклад, див. Vandamme, A.M. et al., Eur. J. Biochem. (1990) 192: Ί61-115, 1990). Більш конкретно, мРНК, яка кодує варіабельну (V) область анти-GPC3-антитіла, виділяють з гібридоми, що продукує анти-GPC3антитіло. мРНК може бути виділена з використанням відомого способу, такого як спосіб центрифугування з гуанідином (Chirgwin, J.M. et al., Biochemistry (1979) 18, 5294-5299) і AGPC-спосіб (Chomczynski, P., et al., Anal. Biochem. (1987) 162, 156-159), для одержання тотальної РНК і потім одержання мРНК з використанням набору для очищення мРНК (Pharmacia) і т. п.мРНК може бути також одержана безпосередньо з використанням набору для одержання мРНК QuickPrep (Pharmacia). Потім синтезують кДНК варіабельної (V) області цього антитіла з одержаної таким чином мРНК з використанням зворотної транскриптази. кДНК може бути синтезована з використанням набору для синтезу кДНК першого ланцюга зворотною транскриптазою AMV (Seikagaku Corporation), і т. п.Може бути також використаний 5'-RACE-cnociб з використанням набору 5!AmpliFINDER RACE Kit (Clontech) і ПЛР для синтезу і ампліфікації кДНК (Frohman, M.A. et al., Proc. Natl. Acad. Sci. USA (1988) 85, 8998-9002, Belyavsky, A. et al., Nucleic Acids Res. (1989) 17, 2919-2932). ДНК-фрагмент-мішень очищають з продукту ПЛР і лігують з ДНК-вектором. Бажаний рекомбінантний вектор готують вбудовуванням цих векторів. Його вводять в Е. соlі і бажану колонію відбирають для приготування бажаного рекомбінантного вектора. Нуклеотидну послідовність ДНК-мішені підтверджують відомим способом, таким як спосіб термінації ланцюга з використанням дидезоксинуклеотиду. Після одержання ДНК, яка кодує варіабельну (V) область анти-GPC3антитела-мішені, її вбудовують в експресуючий вектор, що містить ДНК, яка кодує константну область бажаного антитіла (С-область). Для одержання анти-GPC3-антитіла, що використовується в даному винаході, ген цього антитіла вбудовують в цей експресуючий вектор таким чином, що він буде експресуватися під контролем району регуляції експресії, такого як енхансер, промотор і т. п.Потім це антитіло експресують трансформацією клітинихазяїна цим експресуючим вектором. Ген цього антитіла може бути експресований в клітинаххазяїнах вбудовуванням ДНК, яка кодує важкий ланцюг (Η-ланцюг) і легкий ланцюг (L-ланцюг) ан 23 титіла, в окремі експресуючі вектори і одночасною трансформацією клітин-хазяїнів або вбудовуванням ДНК, яка кодує як Η-ланцюг, так і L-ланцюг, в єдиний експресуючий вектор і трансформацією клітин-хазяїнів (див. WO 94/11523). Крім того, рекомбінантне антитіло може бути одержане не тільки з використанням вищезазначених клітинхазяїнів, але також з використанням трансгенної тварини. Наприклад, ген антитіла може бути вбудований в середину гена, що кодує білок, який продукується специфічно в молоці (такий як казеїн кози), з одержанням злитого гена. Потім ДНК-фрагмент, що містить ген цього антитіла ін'єктують в зародок кози і цей зародок імплантують в козу. Бажане антитіло може бути одержане з молока, що продукується трансгенною козою, народженою козою, імплантованою цим зародком, і її потомством. Крім того, можуть бути використані придатні гормони в трансгенній козі для збільшення кількості молока, що містить бажане антитіло, яке продукується цією трансгенною козою (Ebert, Κ.Μ. et al., Bio/Technology (1994) 12, 699-702). Змінені антитіла Крім описаних вище антитіл, в даному винаході може бути використано штучно змінене рекомбінантне антитіло, наприклад, химерне антитіло, гуманізоване антитіло і т. п., з метою зменшення ксеноантигенності відносно людей. Такі модифіковані антитіла можуть бути одержані відповідно до відомих способів. Химерне антитіло може бути одержане лігуванням ДНК, яка кодує V-область антитіла, одержаної, як описано вище, з ДНК, яка кодує С-область антитіла людини, і потім вбудовуванням цих ДНК в експресуючий вектор. Вектор, в якому вбудовані ці ДНК, інтегрують в клітинихазяїни для одержання цього антитіла. Химерне антитіло, застосовне в даному винаході, може бути одержане з використанням такого загальноприйнятого способу. Гуманізоване антитіло, також зване реконструйованим антитілом людини, містить CDR антитіла з ссавця не людини, такого як миша, трансплантований на CDR антитіла людини. Загальні способи генної інженерії для одержання гуманізованих антитіл відомі в даній області (див. ЕР 125023 і WO 96/02576). Більш конкретно, ДНК-послідовність, призначену для зв'язування CDR мишачого антитіла і каркасної області (FR) антитіла людини, синтезують за допомогою ПЛР з використанням як праймерів множини олігонуклеотидів, приготованих таким чином, що вони перекривають кінцеві райони CDR і FR (цей спосіб описаний в WO 98/13388). Каркасну область антитіла, яка повинна бути зв'язана через CDR, вибирають таким чином, що цей CDR буде утворювати придатний сайт зв'язування антитіла. Якщо необхідно, амінокислоти цієї каркасної області у варіабельному районі антитіла можуть бути замінені таким чином, що цей реконструйований CDR антитіла людини буде утворювати придатний сайт зв'язування антитіла (Sato, K. et ah, Cancer res. (1993) 53, 851-856). Для С-області химерного антитіла і гуманізованого антитіла використовують Собласть антитіла людини. Наприклад, Су1, Су2, Су3 і Су4 можуть бути використані в Η-ланцюзі, а С і С можуть бути використані в L-ланцюгу. Крім 93488 24 того, С-область антитіла людини може бути модифікована для поліпшення стабільності або продуктивності цього антитіла. Химерне антитіло містить варіабельну область антитіла з ссавця не людини і константну область з антитіла людини. З іншого боку, гуманізоване антитіло містить CDR антитіла з ссавця не людини і каркасну область і С-область з антитіла людини. Оскільки гуманізоване антитіло має більш низьку антигенність в тілі людини, воно є більш застосовним як активний інгредієнт в терапевтичному агенті даного винаходу. Модифіковані антитіла Антитіло, що використовується в даному винаході, не обмежується цілою молекулою антитіла і може бути фрагментом антитіла або модифікованою формою антитіла, при умові, що воно зв'язується з GPC3 і інгібує активність GPC3. Даний винахід включає в себе також бівалентні антитіла, а також моновалентні антитіла. Приклади фрагмента антитіла включають в себе Fab, F(ab')2, Fv, Fab/c, що має один Fab і повний Fc, або одноланцюговий Fv (scFv), де Fv Η-ланцюга або L-ланцюга зв'язаний придатним лігандом. Більш конкретно, для одержання фрагмента антитіла це антитіло може бути оброблене ферментом, таким як папаїн або пепсин, або ген, що кодує такий фрагмент антитіла, може бути сконструйований, вбудований в експресуючий вектор і експресований у придатній клітині-хазяїні (див., наприклад, Co, M.S. et al., J. Immunol. (1994) 152, 2968-2976, Better, M. & Horwitz, A. H. Methods in Enzymology (1989) 178, 476-496, Academic Press, Inc., Plueckthun, A. & Skerra, A. Methods in Enzymology (1989) 178, 476496, Academic Press, Inc., Lamoyi, E., Methods in Enzymology (1989) 121, 652-663, Rousseaux, J. et al., Methods in Enzymology (1989) 121, 663-669, Bird, R. E. et al., TIBTECH (1991) 9, 132-137). scFv може бути одержаний з'єднуванням V-області Ηланцюга і V-області L-ланцюга антитіла. У scFv Vобласть Η-ланцюга і V-область L-ланцюги з'єднують за допомогою лінкера, переважно пептидного лінкера (Huston, J.S. et al., Proc. Natl. Acad. Sci. USA (1988) 85, 5879-5883). V-область Н-ланцюга i V-область L-ланцюга в scFv можуть походити з будь-яких описаних тут антитіл. Будь-який одноланцюговий пептид, що містить 12-19 амінокислотних залишків, може бути використаний як пептидний лінкер, що з'єднує ці V-області. ДНК, яка кодує scFv, може бути одержана ампліфікацією фрагмента за допомогою ПЛР з використанням як матриці частини ДНК, яка кодує всю або бажану амінокислотну послідовність, з послідовностей ДНК, яка кодує Η-ланцюг або V-область Н-ланцюга вищезазначеного антитіла, і ДНК, яка кодує L-ланцюг або V-область L-ланцюга вищезазначеного антитіла, з парою праймерів, яка визначає обидва його кінці. Потім цей фрагмент ампліфікують з комбінацією ДНК, яка кодує пептидну лінкерну частину, і парою праймерів, яка визначає обидва кінці, які повинні бути ліговані з Η-ланцюгом і L-ланцюгом. Після одержання ДНК, яка кодує scFv, можуть бути одержані відповідно до стандартних способів експресуючий вектор, що містить цю ДНК, і клітинахазяїн, трансформована цим експресуючим векто 25 ром. scFv може бути одержаний з такого хазяїна відповідно до стандартних способів. Ці фрагменти антитіл можуть бути продуковані в хазяїні за допомогою одержання їх гена таким же чином, як описано вище, і його експресією. У даному винаході термін «антитіло» включає в себе також фрагмент цього антитіла. Анти-GPC3-антитіло, приєднане до різних молекул, таких як PEG і т. п., може бути використане як модифіковане антитіло. У даному винаході термін «антитіло» включає в себе також таке модифіковане антитіло. Модифіковане антитіло може бути одержане хімічною модифікацією антитіла, одержаного, як описано вище. Способи модифікації антитіл вже створені в даній галузі. Крім того, антитіло, що використовується в даному винаході, може бути біспецифічним антитілом. Біспецифічне антитіло може бути антитілом, що має антигензв'язувальний сайт, який упізнає інший епітоп на молекулі GPC3, або воно може бути антитілом, в якому один антигензв'язувальний сайт упізнає GPC3, а інший антигензв'язувальний сайт зв'язування упізнає цитотоксичну речовину, таку як хіміотерапевтичний лікарський засіб і токсин, що походить з клітини. У такому випадку ця цитотоксична речовина буде діяти безпосередньо на клітини, експресуючі GPC3, і, конкретно, на пухлинні клітини пошкодження, і пригнічувати ріст пухлинних клітин. Біспецифічне антитіло може бути одержане зв'язуванням двох типів HL-пap антитіл. Воно може бути також одержане одержанням злитої клітини, яка продукує біспецифічне антитіло, за допомогою злиття гібридом, які продукують різні моноклональні антитіла. Біспецифічне антитіло може бути також одержане способами генної інженерії. Експресія і одержання рекомбінантних антитіл або модифікованих антитіл Ген антитіла, сконструйований, як відмічалося вище, може бути експресований або одержаний відомими способами. У випадку клітин ссавців, звичайний застосовний промотор, ген, який повинен бути експресований, і полі-А-сигнальна послідовність праворуч від 3'-кінця можуть бути функціонально зв'язані разом і експресовані. Наприклад, як промотор/енхансер можуть бути використані негайно ранні промотор/енхансер цитомегаловірусу людини. Крім того, інші промотори/енхансери, які можуть бути використані для експресії антитіла даного винаходу, включають в себе вірусні промотори/енхансери ретровірусу, поліомавірусу, аденовірусу і вірусу 40 мавпи (SV40) або промотори/енхансери з клітин ссавців, таких як фактор елонгації 1а людини (HEF-1a). Антитіла можуть бути легко експресовані за способом Mulligan et al., (Nature (1979) 277, 108), коли використовується промотор/енхансер SV40, і за способом Mizuschima et al., (Nucleic Acids Res. (1990) 18, 5322), коли використовується промотор/енхансер HTF1. Антитіло даного винаходу може бути одержане з використанням еукаріотичної системи експресії, здатної приєднувати цукровий ланцюг до експресованого антитіла. Еукаріотичні клітини включають в себе, наприклад, встановлені клітинні 93488 26 лінії ссавців і клітинні лінії комах, клітини тварин, клітини грибів і клітини дріжджів. Переважно, антитіло даного винаходу експресують в клітинах ссавців, наприклад, в клітинах СНО, COS, клітинах мієломи, ВНК, Vero або HeLa. Бажане антитіло одержують культивуванням трансформованих клітин-хазяїнів in vitro або in vivo. Клітини-хазяїни можуть культивуватися з використанням відомих способів. Наприклад, як культуральне середовище можуть бути використані DMEM, MEM, RPMI-1640 або IMDM, і може бути доданий сироватковий комплемент, наприклад, фетальна теляча сироватка (ФТС). Виділення і очищення антитіла Антитіло, експресоване і одержане описаним вище чином, може бути виділене з клітин-хазяїнів або тварин-хазяїнів і очищене до гомогенності. Антитіло даного винаходу може бути виділене і очищене з використанням афінної колонки, наприклад, Білок Α-колонки, такий як Hyper D, POROS і Sepharose F.F. (Pharmacia). Крім того, в даному винаході можуть бути використані будь-які загальноприйняті способи. Наприклад, антитіло може бути виділене і очищене відповідним чином за допомогою вибору і комбінування афінних колонок, таких як Білок Α-колонка, з хроматографічними колонками, процедурами фільтрації, ультрафільтрації, висолювання і діалізу (Antibodies: A Laboratory Manual, Ed. Harlow, David Lane, Cold Spring Harbor Laboratory, 1988). Антитіло, яке має бажаний цукровий ланцюг, може бути виділене з використанням колонки з лектином способом, відомим в даній галузі, і цей спосіб описаний в WO 02/30954. Визначення активності антитіл Антигензв'язувальна активність (Antibodies: A Laboratory Manual, Ed. Harlow, David Lane, Cold Spring Harbor Laboratory, 1988) і інгібування зв'язування ліганд-рецептор (Harada, A. et al., International Immunology (1993) 5, 681-690) антигену, що використовується в даному винаході, можуть бути виміряні відомими способами. ELISA (твердофазний імуноферментний аналіз), ЕІА (ферментний імуноаналіз), RIA (радіоімуноаналіз) або спосіб флуресцентного антитіла можуть бути використані для вимірювання антигензв'язувальної активності анти-GPC3-антитіла даного винаходу. Наприклад, ЕІА проводять таким чином. Пробу, що містить анти-GPC3-антитіло, таку як культуральний супернатант клітин, що продукують антиGPC3-антитіло, або очищене антитіло, додають в чашку, покриту GPC3. Додають вторинне антитіло, мічене ферментом, таким як лужна фосфатаза, цю чашку інкубують і промивають і потім додають субстрат ферменту, такий як п-нітрофенілфосфат, і визначають оптичну густину для оцінки антигензв'язу вальної активності. Фармацевтична композиція Даний винахід забезпечує фармацевтичну композицію, що містить анти-GPC3-антитіло з модифікованим компонентом, який представляє цукровий ланцюг, даного винаходу. Фармацевтична композиція, що містить композицію антитіл даного винаходу, застосовна для попередження і/або лікування захворювань, асоці 27 йованих із ростом клітин, таких як рак, і особливо застосовна для попередження і/або лікування раку печінки. Фармацевтична композиція, що містить антитіло даного винаходу, може бути приготована способами, відомими кваліфікованим в даній галузі фахівцям. Ця фармацевтична композиція може вводитися парентерально в формі ін'єкційного препарату, який містить стерильний розчин або суспензію у воді або іншій фармацевтично прийнятній рідині. Наприклад, ця фармацевтична композиція може бути приготована відповідним чином за допомогою об'єднання цього антитіла з фармацевтично прийнятними носіями і середовищами, такими як стерильна вода і фізіологічний розчин, рослинна олія, емульгатор, суспендуючий агент, поверхнево-активна речовина, стабілізатор, ароматизуючий ексципієнт, розріджувач, носій, консервант, зв'язувальний агент, з подальшим змішуванням в уніфікованій стандартній формі, необхідній для звичайно прийнятної фармацевтичної практики. Кількість активного інгредієнта, включеного в ці фармацевтичні препарати, є такою, що забезпечується відповідна доза в межах визначеного діапазону. Ця стерильна композиція для ін'єкції може бути приготована відповідно до загальноприйнятої фармацевтичної практики з використанням дистильованої води для ін'єкції як носія. Наприклад, фізіологічний розчин або ізотонічний розчин, що містить глюкозу або інші добавки, такі як D-сорбіт, D-маноза, D-маніт і хлорид натрію, можуть бути використані як водний розчин для ін'єкції, необов'язково в комбінації з придатним солюбілізуючим агентом, наприклад, спиртом, таким як етанол і поліспирт, такий як пропіленгліколь або поліетиленгліколь, і неіоногенною поверхнево-активною речовиною, такою як полісорбат 80, НСО-50 і т. п. Приклади маслянистої рідини включають в себе кунжутну олію і соєву олію, і вона може бути об'єднана з бензилбензоатом або бензиловим спиртом як солюбілізуючим агентом. Іншими компонентами, які можуть бути включені, є фосфатний буфер або натрій-ацетатний буфер, болезаспокійливий засіб, такий як гідрохлорид прокаїну, стабілізатор, такий як бензиловий спирт або фенол, і антиоксидант. Приготований ін'єкційний розчин буде звичайно упакований у відповідну ампулу. Спосіб введення є звичайно парентеральним, наприклад, введення за допомогою ін'єкції, трансназальне введення, черезлегеневе введення, черезшкірне введення. Введення може бути системним або локальним, за допомогою внутрішньовенного введення, внутрішньом'язової ін'єкції, внутрішньочеревинної ін'єкції, підшкірної ін'єкції. Придатні способи введення можуть бути вибрані відповідно до віку і стану пацієнта. Однократна доза фармацевтичної композиції, що містить антитіло або полінуклеотид, який кодує це антитіло, може бути вибрана з діапазону 0,001 1000 мг/кг маси тіла. З іншого боку, можна також вибирати дозу в діапазоні 0,001 100000 мг/тіло, але даний винахід ні в якій мірі не обмежується цими чисель 93488 28 ними діапазонами. Доза і спосіб введення будуть залежати від маси, віку, стану і т. п.пацієнта і може бути відповідним чином вибрана кваліфікованими в даній галузі фахівцями. Зміст всіх патентів і посилальних документів, що спеціально цитуються в описі цієї заявки, включений тим самим як посилання в їх повному об'ємі. Крім того, зміст опису і креслення Заявки на патент Японії 2004-311356, яка є основою для заяви пріоритету цієї заявки, включені тим самим як посилання в їх повному вигляді. Приклади Даний винахід пояснюється детально за допомогою наступних прикладів, але ніяким чином не обмежується цими прикладами. Приклад 1 Одержання мишачого анти-GPC3-антитіла Розчинний білок GPC3, позбавлений гідрофобного району С-кінця (амінокислот 564-580) одержували як імунізуючий білок для одержання антиGPC3-антитіла для імунізації. Як тварину для імунізації використовували мишу MRL/MpJUmmCrjlpr/lpr (далі звану мишею MRL/lpr, що купується з Charles River Japan), яка є мишею з аутоімунним захворюванням. Імунізацію починали, коли вік миші був 7 або 8 тижнів, і препарат для початкової імунізації коректували до дози 100 мкг/миша розчинного GPC3. Емульсію готували з використанням повного ад'юванту Фрейнда (FCA, Bekton Dickinson) і ін'єктували підшкірно. Після серії з п'яти імунізацій дозу кінцевої імунізації розбавляли в ЗФР до 50 мкг/миша і ін'єктували внутрішньовенно через хвостову (каудальну) вену. У день 4 після кінцевої імунізації клітини селезінки витягували, змішували з клітинами мієломи миші P3-X63Ag8U1 (далі званими P3U1, що купуються з АТСС) у співвідношенні 2:1, і проводили злиття клітин поступовим додаванням PEG-1500 (Roche Diagnostics). Гібридоми піддавали скринінгу за допомогою ELISA з використанням імунопланшетів з іммобілізованим розчинним білком GPC3 основної структури (кора). Позитивні клони моноклонували процедурою граничного розведення. У результаті одержували 11 клонів антитіл з сильною активністю зв'язування GPC3 (М3С11, М13В3, М1Е7, М3В8, M11F1, L9G11, М19В11, М6В1, M18D4, М5В9 і M10D2). Серед одержаних анти-GPC3-антитіл особливо сильну CDC-активність виявляли M11F1 і МЗВ8. Таким чином, GST-злитий білок, що містить епітопи M11F1 і М3В8 (GC-3), який є злитим білком, що містить пептид від 524 Ala до 563 Lys GPC3 і GST, використовували як імуногени для імунізації 3 мишей Balb/c (Charles River Japan) і 3 мишей MRL/lpr. Для першої імунізації препарат GC-3 в концентрації 100 мкг/миша емульгували з ФТС і ін'єктували підшкірно. Через два тижні препарат 50 мкг/миша емульгували з неповним ад'ювантом Фрейнда (FIA) і ін'єктували підшкірно. Після п'яти імунізацій кінцеву імунізацію (50 мкг/миша) ін'єктували внутрішньовенно всім мишам через каудальну вену і піддавали злиттю клітин. Гібридоми піддавали скринінгу за допомогою ELISA з використанням імунопланшетів з іммобілізованим розчинним білком GPC3 кора, позбавленим гідро 29 фобного району С-кінця (амінокислот 564-580). Позитивні клони моноклонували з використанням процедури граничного розведення. У результаті одержували 5 клонів антитіл з сильною активністю зв'язування GPC3 (GC199, GC202, GC33, GC179 і GC194). Варіабельні області Н-ланцюга і L-ланцюга клонували і кожну послідовність визначали стандартним способом. Крім того, CDR-райони визначали порівнянням з базою даних амінокислотних послідовностей відомого антитіла і перевіряли на гомологію. Послідовності цих CDR-районів показані в таблицях 1 і 2. Приклад 2 Одержання химерного антитіла анти-GPC3антитіла миша-людина Послідовності варіабельної області Η-ланцюга і L-ланцюга анти-GPC3-антитіла GC33 лігували з послідовностями константної області lgG1 і ланцюга людини. ПЛР проводили з використанням синтетичного олігонуклеотиду, комплементарного 5'-кінцевій нуклеотидній послідовності варіабельної області Н-ланцюга антитіла, що має послідовність Козака, і синтетичного олігонуклеотиду, комплементарного 3'-кінцевій нуклеотидній послідовності, що має сайт Nhel. Одержаним таким чином продукт ПЛР клонували у вектор рВ-СН, в якому константна область lgG1 людини була вбудована у вектор pBluescript KS+ (Toyobo Co., Ltd.). Варіабельну область Η-ланцюга миші і константну область Н-ланцюга людини (1-ланцюг) лігували через сайт Nhel. Одержаний таким чином фрагмент гена Η-ланцюга клонували в експресуючий вектор pCXND3. Крім того, проводили ПЛР з використанням синтетичного олігонуклеотиду, комплементарного 5'-кінцевій нуклеотидній послідовності варіабельної області L-ланцюга антитіла, що має послідовність Козака, і синтетичного олігонуклеотиду, комплементарного 3'-кінцевій нуклеотидній послідовності, що має сайт BsiWI. Одержаний таким чином продукт ПЛР клонували у вектор pB-CL, в якому константна область -ланцюга людини була вбудована у вектор pBluescript KS+ (Toyobo Co., Ltd.). Варіабельну область і константну область L-ланцюга людини лігували через сайт BsiWI. Одержаний таким чином фрагмент гена Lланцюга клонували в експресуючий вектор pUCAG. Вектор pUCAG є вектором, одержаним розщепленням pCXN (Niva et al., Gene, 1991, 108, 193-200) рестриктазою BamHI з одержанням фрагмента 2,6 т.п.н., який лігували з сайтом рестриктази BamHI вектора pUC 19 (Toyobo Co., Ltd.). Для одержання експресуючого вектора химерного анти-GPC3-миша-людина-антитіла фрагмент гена одержували розщепленням вектора pUCAG, в якому був вставлений фрагмент гена L-ланцюга, рестриктазою Hindlll (Takara Shuzo Co., Ltd.). Цей фрагмент гена лігували з сайтом розщеплення рестриктази Hindlll pCXND3, що містить ген Ηланцюга, і потім клонували. Одержана таким чином плазміда експресувала ген стійкості до неоміцину, ген DHFR і ген химерного анти-GPC3-мишалюдина-антитіла в клітинах тварини. (Амінокислотна послідовність варіабельної області Η-ланцюга показана в SEQ ID NO:3, а амінокислотна послідо 93488 30 вність варіабельної області L-ланцюга показана в SEQ ID NO:4. Приклад 3 Одержання химерного анти-GPC3-антитіла з низьким вмістом фукози Спочатку клітини YB2/0 (АТСС, CRL-1662) культивували як клітини-хазяїни в середовищі RPMI1640, що містить 10% ФТС. Потім 25 мкг експресуючого химерне анти-GPC3-антитіло вектора, одержаного, як описано в прикладі 2, вводили в клітини YB2/0 (АТСС CRT-1662) електропорацією в концентрації 7,5х10б клітин/0,75 мл ЗФР(-) при 1,4 кВ і 25 мкФ. Після періоду відновлення клітин протягом 10 хв. при кімнатній температурі ці клітини, оброблені електропорацією, суспендували в 40 мл середовища RPMI-1640, що містить 10% ФТС. Готували 10-кратне розведення з використанням того ж самого середовища. Після культивування протягом 24 годин в СО2-термостаті (5% СО2) додавали генетицин (Invitrogen Corp.) в концентрації 0,5 мг/мл і ці клітини культивували протягом 2 тижнів. Клітинні лінії з високим рівнем експресії химерного антитіла піддавали скринінгу з використанням сендвіч-ELISA з антитілом проти IgG людини і встановлювали клітинні лінії, стабільно експресуючі це антитіло. Кожне химерне антиGPC3-миша-людина-антитіло очищали з використанням Ні Trap Protein G HP (Amersham). Приклад 4 Вимірювання ADCC-активності з використанням РВМС з периферійної крові людини Одержання розчину РВМС людини Периферійну кров з доданим гепарином, зібрану із здорової дорослої людини, розбавляли 2кратно ЗФР(-) і наносили на Ficoll-Paque PLUS (Amersham). Після центрифугування (500xg, 30 хвилин, 20°С) виділяли проміжний шар, який є фракцією мононуклеарних клітин. Після промивання цього шару 3 рази ці клітини суспендували в 10% ФТС/RPMI для одержання розчину РВМС людини. Одержання клітин-мішеней Клітини HepG2 (ATCC) і клітини HuH-7 (Health Science Research Resources Bank), що культивуються в середовищі 10% ФТС/RPMI-1640, відділяли від чашки з використанням буфера для дисоціації клітин (Invitrogen), наносили у вигляді аліквот в 96-ямковий U-донний планшет (Falcon) при концентрації 1x10 клітин на ямку і культивували протягом одного дня. Після культивування додавали 5,55 МБк 51Cr і клітини культивували 1 годину при 37°С в термостаті з 5% газоподібним СО2. Клітини промивали один раз культуральним середовищем і додавали 50мкл середовища 10% ФТС/RPMI1640 для одержання клітин-мішеней. Аналіз вивільнення хрому (активності ADCC) Об'єм 50мкл розчину антитіла, приготованого в різних концентраціях, додавали до клітинмішеней і давали реагувати на льоду протягом 15 хвилин. Потім додавали 100 мкл розчину РВМС людини (5×105 клітин на ямку) і ці клітини інкубували протягом 4 годин при 37°С в термостаті з 5% газоподібним СО2. Після культивування цей планшет центрифугували і радіоактивність в 100 мкл культурального супернатанту вимірювали з вико 31 93488 ристанням гамма-лічильника. Питому швидкість вивільнення хрому визначали за наступною фор 32 мулою. Питома швидкість вивільнення хрому (%)=(АС)×100/(В-С) У цій формулі А представляє середню величину радіоактивності (імп./хв.) в кожній лунці; В представляє середню величину радіоактивності (імп./хв.) в лунці, в якій до клітин-мішеней додавали 100мкл 2% водного розчину NP-40 (Нонідет Р40, Номер коду 252-23, Nacalai Tesque) і 50мкл середовища 10% ФТС/RPMI; і С представляє середню величину радіоактивності (імп./хв.) в лунці, в якій до клітин-мішеней додавали 150 мкл середовища 10% ФТС/RPMI. Аналіз проводили в трьох повторностях і розраховували середні величини і стандартні відхилення ADCC-активності (%). Фіг.2 і 3 показують ADCC-активність химерного анти-GPC3-антитіла, виміряну з використанням РВМС. На цих фігурах вертикальна вісь представляє цитотоксичну активність, а горизонтальна вісь представляє концентрацію (мкг/мл) доданого антитіла. Фіг.2 показує результати, коли як клітинимішені використовували клітини HepG2, а Фіг.3 показує результати для клітин HuH-7. Білі кружки показують активність химерного антитіла GC33, що продукується клітинами СНО, а чорні кружки показують активність химерного антитіла GC33, що продукується клітинами YB2/0. Химерне антитіло GC33 з низькою фукозою, що продукується клітинами YB2/0, виявляє більш сильну активність ADCC, ніж химерне антитіло GC33, що продукується клітинами СНО, ясно свідчачи про те, що ADCC-активність анти-GPC3-антитіла посилюється модифікацією цукрового ланцюга. Приклад 5 Створення антитілопродукуючих клітин Гігроміцин В додавали в середовище SFMII(+) в кінцевій концентрації 1 мг/мл і клітинну лінію, недостатню відносно транспортера фукози (клон 3F2), субкультивували в цьому середовищі. Готували суспензію клітин 3F2 у фосфатному буфері Дульбекко (8×106 клітин/0,8мл). До цієї суспензії клітин додавали 25мкг експресуючого антитіло вектора (Посилальні приклади 1 і 2) і цю суспензію клітин переносили в Gene Pulser Cuvette. Після залишення цієї кювети стояти на льоду протягом 10 хвилин цей вектор вводили в клітини електропорацією з використанням GENE-PULSER II при 1,5кВ і 25мкФ. Ці клітини суспендували в 40 мл середовища SFMII(+) і переносили в 96-ямковий плоскодонний планшет (Iwaki) при 100мкл на ямку. Після інкубування цього планшета в СO2термостаті протягом 24 годин при 37°С, додавали генетицин (Invitrogen Cat. No. 10131-027) в кінцевій концентрації 0,5мг/мл. Кількість антитіла, що продукується стійкими до лікарського засобу клітинами, вимірювали для встановлення клітинних ліній, які продукують гуманізоване анти-GPC3-антитіло. Приклад 6 Очищення антитіл Супернатант з експресуючої антитіло клітинної лінії збирали і наносили на колонку Hitrap rProtein A (Pharmacia Cat. No. 17-5080-01) з використанням насоса Р-1 (Pharmacia). Після проми вання цієї колонки буфером для зв'язування (20мМ фосфат натрію, (рН 7,0)) цей білок елюювали буфером для елюції (0,1 Μ гліцин-НСl (рН 2,7)). Елюат відразу ж нейтралізували нейтралізуючим буфером (1 Μ Тріс-HCl (рН 9,0)). Фракції елюції антитіла збирали з використанням DCаналізу білка (BIO-RAD Cat. No. 500-0111) і об'єднували і концентрували до приблизно 2мл з використанням Centriprep-YMIO (Millipore Cat. No. 4304). Потім ці антитіла розділяли гельфільтрацією з використанням колонки Superdex 200 26/60 (Pharmacia), врівноваженою 20 мМ ацетатним буфером з 150 мМ NaCl (рН 6,0). Піки фракції мономера збирали, концентрували з використанням Centriprep-YMIO і фільтрували через фільтрувальний елемент MILLEX-GW (Millipore Cat. No. SLGV 013SL) і потім зберігали при 4°С. Вимірювали поглинання при 280 нм і концентрацію очищеного антитіла розраховували з коефіцієнта молярного поглинання. Приклад 7 Активність ADCC in vitro гуманізованого антиGPC3-антитіла, що продукується клітинами FT-KO Фіг.4 показує активність ADCC in vitro антиGPC3-антитіла, що продукується клітинами FT-KO, при використанні РВМС людини. Цей спосіб є способом, описаним в прикладі 4. На цій фігурі вертикальна вісь представляє цитотоксичну активність (%), а горизонтальна вісь представляє концентрацію (мкг/мл) доданого антитіла. Як клітини-мішені використовували клітини HuH-7. Білі кружки показують активність анти-GPC3-антитіла, що продукується клітинами СНО дикого типу, а чорні кружки показують активність анти-GPC3-антитіла, що продукується клітинами FT-KO. Ahth-GPC3антитіло з низькою фукозою, яке продукується клітинами FT-KO, виявляє більш сильну активність ADCC, ніж анти-GPC3-антитіло, що продукується клітинами СНО дикого типу, що ясно вказує на те, що активність ADCC анти-GPC3-антитіла, що продукується клітинами FT-KO, є підвищеною. Приклад 8 Аналіз цукрових ланцюгів гуманізованого антиGPC3-антитіла, що продукується клітинами FT-KO 1. Одержання 2-амінобензамідмічених цукрових ланцюгів (2АВ-мічених цукрових ланцюгів) Антитіло, що продукується клітинами FT-KO даного винаходу, і антитіла, що продукуються клітинами СНО, як контрольну пробу обробляли Nглюкозидазою F (Roche Diagnostics) для вивільнення цих цукрових ланцюгів з білка (Weitzhandler Μ. et al., Journal of Pharmaceutical Sciences 83:12 (1994), 1670-1675). Після видалення білка етанолом (Schenk В. et al., The Journal of Clinical Investigation 108:11 (2001), 1687-1695) цукрові ланцюги концентрували і сушили і флуоресцентно мітили 2-амінопіридином (Bigge J.C. et al., Analitical Biochemistry 230:2 (1995), 229-238). Реагент видаляли з 2-АВ-мічених цукрових ланцюгів твердофазною екстракцією з використанням целюлозного 33 93488 картриджа і після концентрування центрифугуванням одержували 2-АВ-мічені цукрові ланцюги. Потім ці очищені 2-АВ-мічені цукрові ланцюги обробляли -галактозидазою (Seikaguku Corp.) з одержанням -галактозил-2-АВ-мічених цукрових ланцюгів. 2. Аналіз -галактозил-2-АВ-мічених цукрових ланцюгів з використанням ВРХ з нормальною фазою Антитіла, що продукуються клітинами FT-KO даного винаходу, і антитіла, що продукуються клітинами СНО, як контрольну пробу одержували у вигляді -галактозил-2-АВ-мічених цукрових ланцюгів відповідно до описаного вище способу і аналізували ВРХ з нормальною фазою з використанням амідної колонки (Tosoh Corp. TSKgel Amide80) і хроматограми порівнювали. У антитілах, що продукуються клітинами СНО, основним компонентом є G(0), a G(0)-Fuc відповідальний за приблизно 4% площі піка. З іншого боку, в антитілах, що продукуються клітинами FT-KO, основним компонентом є G(0)-Fuc, і він присутній щонайменше в 90% площі піка в кожній з клітинних ліній (Фіг.5 і таблиця 2). Фіг.6 показує передбачувані структури для піків G(0) і G(0)-Fuc. Таблиця 2 Відносна частка цукрових ланцюгів, визначена з аналізу ВРХ з нормальною фазою -галактозил2АВ-цукрових ланцюгів Цукровий ланцюг G(0)-Fuc G(0) СНО 4,0%96,0% FT-KO-a FT-KO-b FT-KO-c 92,4% 7,6% 92,5% 7,5% 93,2% 6,8% Приклад 9 Аналіз термостабільності гуманізованого антиGPC3-антитіла, що продукується клітинами FT-KO 1. Приготування розчину проби для DSCвимірювання Зовнішній діалізний розчин був 20 моль/л натрієм-ацетатним буфером (рН 6,0), що містить 200ммоль/л хлориду натрію. Діалізну мембрану, наповнену 700мкг-еквівалентами розчину антитіла, діалізували зануренням в зовнішній діалізний розчин протягом ночі для одержання розчину проби. 2. Вимірювання температури термодеструкції з використанням DSC (диференціальної сканувальної калориметрії) Після ретельного дегазування розчину проби і посилального розчину (зовнішнього діалізного розчину) кожний з них помішували в калориметр і термічно врівноважували при 20°С. Потім проводили DSC-вимірювання від 20°С до 100°С при швидкості сканування приблизно 1К/хв. Цей результат представлений нахилом піка розкладання як функції температури (Фіг.7). Було виявлено, що температура термодеструкції антитіл, що продукуються клітинами СНО, і антитіл, що продукуються клітинами FT-KO, є еквівалентною. Посилальний приклад 1 Гуманізація GC33 34 Інформацію про послідовність антитіла одержували з публічно доступної бази даних Кабата (ftp://ftp.ebi.ac.uk.pub/databases/kabat/) і варіабельну область Н-ланцюга і варіабельну область Lланцюга по окремості піддавали пошуку гомології. Було виявлено, що варіабельна область Ηланцюга має високий рівень гомології з DN13 (Smithson et al., Mol. Immunol. 1999; 36: 113-124). Було також виявлено, що варіабельна область Lланцюга має високий рівень гомології з мРНК IgK Homo sapiens відносно VLJ-області легкого ланцюга каппа, часткових CDS імуноглобуліну, клону: К64 з Accession Number AB064105. Сигнальну послідовність Accession Number S40357, яка має високий рівень гомології з АВ064105, використовували як сигнальну послідовність L-ланцюга. Потім CDR трансплантували в FR цих антитіл для одержання гуманізованого антитіла. Більш конкретно, синтетичні оліго-ДНК з довжиною приблизно 50 основ конструювали таким чином, що приблизно 20 основ з них гібридизувалися одна з одною, і ці синтетичні оліго-ДНК збирали за допомогою ПЛР для одержання гена, що кодує кожну варіабельну область. їх розщеплювали в послідовності Hindlll, вбудованій на кінці 5'кінця синтетичного ДНК-оліго, і в послідовності ВатНІ, вбудованій на кінці 3'-кінця оліго-ДНК, і цей синтетичний ДНК-оліго клонували в експресуючий вектор HEFg1, коли клонували константну область lgG1 людини, або експресуючий вектор HEFg, коли клонували константну область ланцюга людини (Sato et al., Mol. Immunol., 1994; 371-381). Η-ланцюг і L-ланцюг цього гуманізованого GC33, сконструйовані, як описано вище, кожний, означали ver.a. Гуманізоване антитіло GC33 (ver.a/ver.a), в якому обидва ланцюги, Η-ланцюг і L-ланцюг, були ver.a, мали більш низьку активність зв'язування, ніж антитіло з варіабельними областями GC33 миші (миша/миша). Химерні антитіла одержували об'єднанням мишачих послідовностей GC33 і послідовностей ver.a для Н-ланцюгів і Lланцюгів (миша/ver.a, ver.a/миша) і оцінювали активність зв'язування. Більш низька активність зв'язування була виявлена з антитілом ver.a, що свідчить про те, що зменшення активності зв'язування зумовлене заміною амінокислоти, яка була зв'язана з Η-ланцюгом. Потім готували модифіковані Ηланцюги, названі ver.c, ver.f, ver.h, ver.i, ver.j і ver.k. Всі гуманізовані антитіла GC33 виявляли той же самий рівень активності зв'язування, що і химерне антитіло, яке має варіабельні області мишачого GC33. Нуклеотидні послідовності варіабельних областей Н-ланцюга гуманізованого GC33 ver.a, ver.c, ver.f, ver.h, ver.i, ver.j і ver.k показані в SEQ ID NO:74, 75, 76, 78, 79 і 80, а всі їх амінокислотні послідовності показані в SEQ ID NO:81, 82, 83, 84, 85, 86 і 87, відповідно. Нуклеотидна послідовність варіабельної області ver.a L-ланцюга гуманізованого GC33 показана в SEQ ID NO:88, а її амінокислотна послідовність показана в SEQ ID NO:89, відповідно. У варіабельних областях ver.i, ver.j і ver.k Η-ланцюга гуманізованого GC33 шоста глутамінова кислота замінена глутаміном. Ці антитіла виявляли явно збільшену термостабільність. Посилальний приклад 2 35 Зміна L-ланцюга гуманізованого GC33 Що стосується деамідування білка, було відомо, що константа швидкості реакції деамідування залежить від первинної послідовності. Відомо також, що послідовність Asn-Gly особливо чутлива до деамідування (Rocinson et al., Proc. Natl. Acad. Sci. USA 2001; 98: 944-949). Оскільки Asn 33 в CDR1 варіабельної області ver.a L-ланцюга гуманізованого GC33 SEQ ID NO:88 має первинну послідовність Asn-Gly, передбачається, що цей залишок є чутливим до деамідування. Для оцінки дії деамідування Asn 33 на активність зв'язування цього антитіла одержували модифіковане антитіло, в якому Asn 33 був замінений Asp. Набір Quick Change Site-Directed Mutagenesis Kit (Stratagene) використовували для введення точкової мутації. Більш конкретно, 50 мкл реакційного розчину, що містить 125нг смислового праймеру (СТТ GTA CAC AGT GAC GGA ААС АСС ТАТ: SEQ ID NO:124), 125нг антисмислового праймеру (ATA GGT GTT ТСС GTC ACT GTG ТАС AAG: SEQ ID NO:125), 5мкл 10х реакційного буфера, 1 мкл суміші dNTP, 10 нг вектора HEFg, в які був клонований L-ланцюг ver.a гуманізованого GC33, і 1мкл ДНК-полімерази Pfu Turbo, піддавали 12 циклам, які складаються з 30 сек. при 95°С, 1хв. при 55°С і 9 в. при 68°С. Продукт реакції розщеплювали рестриктазою Dpnl протягом 2 годин при 37°С і вводили в XL I-Blue компетентні клітини для одержання трансформантів. Варіабельну область вирізали з клонів, що містять коректну мутацію, і знову клонували в експресуючий вектор HEFg. Експресуючий вектор HEFg1, що містить Η-ланцюг ver.k гуманізованого GC33, вводили в клітини C0S7 з використанням Fugene 6 (Roche). Супернатант культурального середовища збирали з клітин, тимчасово експресуючих це модифіковане антитіло. Концентрацію антитіла кількісно визначали сендвіч-ELISA з використанням антитіла проти IgG людини і активність зв'язування цього модифікованого антитіла оцінювали за допомогою ELISA з використанням планшета, покритого розчинним білком-кором GPC3. Активність зв'язування втрачалася в модифікованому антитілі (N33D), в якому Asn 33 був замінений Asp, що дозволяє передбачити, що на активність зв'язування суттєво впливає деамідування в Asn 33. Повідомлялося, що деамідування Asn 33 пригнічується заміною Gly 34 іншим амінокислотним залишком (WO 03057881 А1). Відповідно до цього способу, одержували ряд модифікованих антитіл заміною G34 17 іншими амінокислотними залишками, за винятком Cys і Met, з використанням набору Quick Change Site-Directed Mutagenesis Kit з одержанням G34A, G34D, G34E, G34F, G34H, G34N, G34P, G34Q, G34I, G34K, G34L, G34V, G34W, G34Y, G34R, G34S і G34T. Активність зв'язування цих антитіл оцінювали з використанням культурального супернатанту клітин COS7, тимчасово експресуючих ці антитіла. Було виявлено, що активність зв'язування зберігалася, навіть якщо G34 замінюють іншим амінокислотним залишком, іншим, ніж Pro (G34P) і Val (G34V). Амінокислотні послідовності CDR1 L-ланцюга модифікованих антитіл представлені SEQ ID NO: 93488 36 90 (G34A), SEQ ID NO: 91 (G34D), SEQ ID NO: 92 (G34E), SEQ ID NO: 93 (G34F), SEQ ID NO: 94 (G34H), SEQ ID NO: 95 (G34N), SEQ ID NO: 96 (G34T), SEQ ID NO: 97 (G34Q), SEQ ID NO: 98 (G34I), SEQ ID NO: 99 (G34K), SEQ ID NO: 100 (G34L), SEQ ID NO: 101 (G34S), SEQ ID NO: 102 (G34W), SEQ ID NO: 103 (G34Y), SEQ ID NO: 104 (G34R), SEQ ID NO: 105 (G34V), і SEQ ID NO: 106 (G34P), відповідно. Амінокислотні послідовності варіабельних областей L-ланцюга модифікованих антитіл представлені SEQ ID NO: 107 (G34A), SEQ ID NO: 108 (G34D), SEQ ID NO: 109 (G34E), SEQ ID NO: 110 (G34F), SEQ ID NO:111 (G34H), SEQ ID NO: 112 (G34N), SEQ ID NO:113 (G34T), SEQ ID NO:114 (G34Q), SEQ ID NO: 115 (G34I), SEQ ID NO: 116 (G34K), SEQ ID NO: 117 (G34L), SEQ ID NO: 118 (G34S), SEQ ID NO: 119 (G34W), SEQ ID NO: 120 (G34Y), SEQ ID NO: 121 (G34R), SEQ ID NO: 122 (G34V), і SEQ ID NO: 123 (G34P), відповідно. Посилальний приклад 3 Деструкція гена транспортера фукози в клітинах СНО 1. Конструювання націлюючого вектора (1). Одержання вектора КО1 Ген стійкості до гігроміцину (Hygr) конструювали за допомогою ПЛР з праймерами Hyg5-BH і Hyg3-NT з плазміди pcDNA3.1/Hygro (Invitrogen), яка має послідовність, ідентичну 5'-частині стартового кодону гена транспортера фукози, приєднанням сайта BamHI і послідовності TGCGC до 5'частини стартового кодону і сайта NotI, доданого до 3'-частини, що містить цей район, до сигналу приєднання поліА SV40, і Hygr-фрагмент вирізали. Прямий праймер Hyg5-BH 5'-GGA ТСС TGC GCA TGA ААА AGC CTG ААС ТС А СС-3' (SEQ ID NO:128) Зворотний праймер Hyg3-NT 5'-GCG GCC GCC TAT TCC TTT GCC CTC GGA CG-3' (SEQ ID NO:129) Націлюючий на транспортер фукози вектор ver.1 (далі званий вектором КO1) конструювали вбудовуванням 5'-частини нуклеотидної послідовності транспортера фукози (від Smal при основі № 2780 до ВатНІ при основі № 4323 нуклеотидної послідовності, показаної в SEQ ID NO:126), 3'частини (від основи № 4284 до Sad при основі № 10934) і Hygr-фрагмента у вектор рМС1DТ-А (Yagi Τ, Proc. Natl. Acad. Sci. USA, Vol. 87, p. 9918-9922, 1990). Характеристикою цього вектора КО1 є те, що цей Hygr-фрагмент буде експресуватися від промотору транспортера фукози, коли має місце гомологічна рекомбінація, оскільки до цього Hygrфрагмента не приєднаний промотор. Однак, Hygr не завжди експресується до такої міри, що утворюється стійкість до гігроміцину В, якщо тільки одна копія цього вектора інсертується в клітину гомологічною рекомбінацією. Вектор КО1 розщеплювали NotI і вводили в клітину. Очікується, що транспортер фукози буде втрачати 41 пару основ екзону 1, в тому числі стартовий код он, внаслідок введення вектора КО1, що буде приводити до втрати його функції. (2) Одержання pBSK-pgk-1-Hygr 37 Вектор pBSK-pgk-1 одержували відрізанням промотору гена pgk-Ι миші від вектора pKJ2 (Роро Н, Biochemical Genetics, Vol. 28, p. 299-308, 1990) рестриктазами EcoRI-PstI і клонуванням його в сайт EcoRI-PstI pBluescript (Stratagene). За допомогою ПЛР з праймерами Hyg5-AV і Hyg3-BH з pcDNA3.1/Hygro сайт ЕсоТ22І і послідовність Козака приєднували до 5'-частини Hygr, а сайт BamHI додавали до 3'-частини, що містить район до сигналу приєднання полі A SV40, і потім вирізали Hygr-фрагмент. Прямий праймер Hyg5-AV ATG CAT GCC ACC ATG AAA AAG CCT GAA CTC ACC-3' (SEQ ID NO:130) Зворотний праймер Hyg3-BH 5'-GGA TCC CAG GCT TTA CAC TTT ATG CTT C-3' (SEQ ID NO:131) Вектор pBSK-pgk-1-Hygr одержували вбудовуванням Hygr (EcoT22I-ВатНІ)-фрагмента в сайт Pstl-BamHI pBSK-pgk-1. (3) Одержання вектора КО2 Націлюючий на транспортер фукози вектор ver.2 (далі званий вектором КО2) конструювали вбудовуванням 5'-частини нуклеотидної послідовності транспортера фукози (від Smal при основі № 2780 до ВатНІ при основі № 4323 нуклеотидної послідовності, показаної в SEQ ID NO:126), 3'частини (від основи № 4284 до Sad при основі № 10934) і pgk-1-Hygr-фрагмента у вектор pMC1DTА. На відміну від вектора КО1, вектор КО2 буде додавати стійкість до гігроміцину В, навіть якщо тільки одна копія цього вектора вбудована гомологічною рекомбінацією, оскільки промотор гена pgk1 приєднаний до Hygr. Вектор КО2 розщеплювали NotI і вводили в клітини. Очікується, що транспортер фукози буде втрачати 46 пар основ екзону 1, в тому числі стартовий кодон, внаслідок введення вектора КО2, що буде приводити до втрати його функції. (4) Одержання pBSK-pgk-1-Puror Вектор pBSK-pgk-1-Puror одержували розщепленням вектора pPUR (BD Biosciences) PstI і BamHI і вбудовуванням цього розщепленого фрагмента (Puror) в сайт Pstl-BamHI pBSK-pgk-1. (5) Одержання вектора КО3 Націлюючий на транспортер фукози вектор ver.3 (далі званий вектором КО3) конструювали вбудовуванням 5'-частини нуклеотидної послідовності транспортера фукози (від Smal при основі № 2780 до ВатНІ при основі № 4323 нуклеотидної послідовності, показаної в SEQ ID NO:126), 3'частини (від основи № 4284 до Sad при основі № 10934) і pgk-1-Puror-фрагмента у вектор pMC1DTА. Крім того, до 3'-кінця pgk-1-Puror приєднували послідовність для зв'язування з праймером для описаного нижче скринінгу. Вектор КО3 розщеплювали Notl і вбудовували в клітини. Очікується, що транспортер фукози буде втрачати 46 пар основ екзону 1, в тому числі стартовий кодон, внаслідок введення вектора КО3, що буде приводити до втрати його функції. Зворотний праймер RSGR-A 5'-GCT GTC TGG AGT ACT GTG CAT CTG C-3' (SEQ ID NO:132) 93488 38 Описані вище три види націлюючих векторів використовували для нокауту гена транспортера фукози. 2. Введення векторів в клітини СНО НТ-добавку (100х) (Invitrogen Cat. No. 11067030) і пеніцилін-стрептоміцин (Invitrogen Cat. No. 15140-122) додавали до CHO-S-FMII НТ (Invitrogen Cat. No. 12052-098), кожний в об'ємі 1/100 відносно об'єму CHO-S-FMII НТ. Клітини СНО DXB11 субкультивували в культуральному середовищі (далі званому SEMII(+)), і це середовище SEMII(+) використовували для культивування цих клітин після перенесення гена. Клітини СНО суспендували в фосфатному буфері Дульбекко (далі званому ЗФР, Invitrogen cat. No. 14190-144) при концентрації (8x106 клітин/0,8 мл). Потім до цієї суспензії клітин додавали 30 мкг націлюючого вектора і цю суспензію клітин переносили в Gene Pulser Cuvette (4 мм) (Віо-Rad, Cat. No. 1652088). Після залишення цієї кювети стояти на льоду протягом 10 хвилин цей вектор вводили в клітини електропорацією з використанням GENE-PULSER II (Bio-Rads, Code No. 340BR) при 1,5 кВ і 25 мкФ. Після введення цього вектора клітини суспендували в 200 мл середовища SFMII(+) і переносили в двадцять 96-ямкових плоскодонних планшетів (Iwaki, Cat. No. 1860-096) при 100 мкл на ямку. Після інкубування цих планшетів в С02-термостаті протягом 24 годин при 37°С, додавали реагент. 3. Стадія нокауту 1 Або вектор КО1, або вектор КО2 вводили в клітини СНО і після 24 годин ці клітини піддавали відбору з використанням гігроміцину В (Invitrogen, Cat. No. 10687-010). Гігроміцин В розчиняли в SFMII(+) до концентрації 0,3 мг/мл і додавали при 100 мкл на ямку. 4. Скринінг на гомологічні рекомбінанти за допомогою ПЛР (1) Приготування ПЛР-проби Скринінг на гомологічні рекомбінанти проводили за допомогою ПЛР. Клітини СНО, що використовуються в скринінгу, культивували в 96-ямкових планшетах. Після видалення супернатанту додавали 50 мкл на ямку буфера для цитолізу і ці клітини спочатку нагрівали при 55°С протягом 2 годин і потім при 95°С протягом 15 хвилин для інактивації протеази К для одержання ПЛРматриці. Буфер для цитолізу складався з 5 мкл 10xLA-буфер II (Takara Bio Inc., доповнений La Taq), 2,5 мкл 10% NP-40 (Roche, Cat. No. 1 332 473), 4 мкл протеїнази К (20 мг/мл, Takara Bio Inc. Cat. No. 9033) і 38,5 мкл дистильованої води (Nacalai Tesque Cat. No. 36421-35) на ямку. (2) Умови ПЛР Реакційна суміш ПЛР містила 1 мкл вищезгаданої проби ПЛР, 5мкл 10 x LA-буфера II, 5мкл MgCl2 (25мМ), 5мкл dNTP (2,5мМ), 2мкл праймеру (10 мкМ кожного), 0,5мкл LA Taq (5МЕ/мкл, Cat. No. RR002B) і 29,5мкл дистильованої води (загальний об'єм 50 мкл). Для скринінгу клітин, що містять вектор КО1, як ПЛР-праймери використовували TP-F4 і THygro-R1, а для скринінгу клітин, що містять вектор КО2, як праймери використовували TP-F4 і THygro-F1. 39 Умови ПЛР для клітин, що містять вектор КО1, складалися з попереднього нагрівання при 95°С протягом 1 хвилини, проведення 40 циклів, що складаються з циклу ампліфікації 30 секунд при 95°С, 30 секунд при 60°С і 2 хвилин при 60°С, і потім повторного нагрівання при 72°С протягом 7 хвилин. Умови ПЛР для клітин, що містять вектор КО2, складалися з попереднього нагрівання при 95°С протягом 1 хвилини, проведення 40 циклів, що складаються з циклу ампліфікації 30 секунд при 95°С і 3 хвилин при 70°С, і потім повторного нагрівання при 70°С протягом 7 хвилин. Використовувані праймери наведені нижче. У пробах клітин, в яких відбувається гомологічна рекомбінація з вектором КО1 або вектором КО2, буде ампліфікуватися ДНК приблизно 1,6 т.п.н. або 2,0 т.п.н., відповідно. Праймер ТР-F4 був сконструйований для 5'-району геному транспортера фукози зовні від вектора, a THygro-F1 і THygro-R1 були сконструйовані для гена Hygr всередині цього вектора. Прямий праймер (КО1, ΚΟ2) TP-F4 5'-GGA ATG CAG CTT CCT САА GGG ACT CGC-3' (SEQ ID NO:133) Зворотний праймер (КО1) THygro-R1 5'-TGC АТС AGG TCG GAG ACG CTG TCG AAC-3' (SEQ ID NO:134) Зворотний праймер (КО2) THygro-Fl 5'-GCA CTC GTC CGA GGG CAA AGG AAT AGC-3' (SEQ ID NO:135) 5. Результати ПЛР-скринінгу Загалом аналізували 918 клітин, що містять вектор КО1, і одна клітина була, мабуть; гомологічним рекомбінантом (ступінь гомологічної рекомбінації: приблизно 0,1%). Загалом аналізували 537 клітин, що містять вектор КО2, і 17 клітин, мабуть, є гомологічними рекомбінантами (ступінь гомологічної рекомбінації: приблизно 3,2%). 6. Блот-аналіз за Саузерном Гомологічну рекомбінацію додатково підтверджували блотингом за Саузерном. У цілому 10 мкг геномної ДНК одержували з культивованих клітин згідно зі стандартним способом для аналізу в блотингу за Саузерном. ПЛР проводили з використанням двох праймерів, наведених нижче, для одержання зонда 387 п.н., відповідного району основ з номерами 2113-2500 нуклеотидної послідовності, показаної в SEQ ID NO:126, який використовували в блотингу за Саузерном. Цю геномну ДНК розщеплювали Bglll. Прямий праймер Bgl-F: 5'- TGT GCT GGG ААТ TGA ACC CAG GAC-3' (SEQ ID NO:136) Зворотний праймер Bgl-R: 5'- СТА СТТ GTC TGT GCT ТТС ТТС С3' (SEQ ID NO:137) У результаті розщеплення Bglll, цей блот буде показувати смугу приблизно 30 т.п.н. з хромосоми транспортера фукози, приблизно 4,6 т.п.н. з хромосоми, в якій відбулася гомологічна рекомбінація з вектором КО1, і приблизно 5,0 т.п.н. з хромосоми, в якій відбулася гомологічна рекомбінація з вектором КО2. Цей експеримент містив 1 клітину з гомологічної рекомбінації з вектором КО1 і 7 клітин з гомологічної рекомбінації з вектором КО2. Ця 93488 40 єдина клітина, одержана з гомологічної рекомбінації з вектором КО1, була спочатку названа 5С1, але більш пізній аналіз виявив, що ця клітина складалася з популяцій множинних клітин. Таким чином, ці клітини клонували лімітуючим розведенням перед використанням в цьому експерименті. Одна з клітин, одержана з вектором КО2, була названа 6Е2. 7. Стадія нокауту 2 Ці три вектори використовували для створення клітинних ліній, повністю дефектних по гену транспортера фукози, з клітин, в яких мала місце гомологічна рекомбінація з векторами КО1 і КО2. Комбінації векторів і клітин були наступними: Спосіб 1 об'єднував вектор КО2 і клітини 5С1 (КО1), Спосіб 2 об'єднував вектор КО2 і клітини 6Е2 (КО2) і Спосіб 3 об'єднував вектор КО3 і клітини 6Е2 (КО2). Кожний вектор вводили у відповідні клітини і після 24 годин починали відбір з використанням гігроміцину В і пуроміцину (Nacalai Tesque, Cat. No. 294-12). Кінцева концентрація гігроміцину В була встановлена на 1 мг/мл в Способі 1 і 7 мг/мл в Способі 2. У Способі 3 гігроміцин додавали при кінцевій концентрації 0,15 мг/мл, а пуроміцин в кінцевій концентрації 8 мкг/мл. 8. Скринінг на гомологічні рекомбінанти за допомогою ПЛР Для скринінгу клітин зі Способу 1 ПЛР проводили для детектування клітин, що мають гомологічну рекомбінацію як з вектором КО1, так і з вектором КО2. Для клітин зі Способу 2 були сконструйовані наступні праймери ПЛР: TPS-F1 був утворений в районі основ з номерами 39243950 SEQ ID NO:126, a SHyhroR1 був утворений в районі основ з номерами 4248-4274. Ці праймери будуть ампліфікувати 350 п.н. району гена транспортера фукози, що містить делецію внаслідок вектора КО2. Таким чином, в ПЛР-скринінгу в Способі 2 клітини, що не забезпечують продукту ампліфікації 350 п.н. вважаються повністю позбавленими гена транспортера фукози. Умови ПЛР складалися з попереднього нагрівання протягом 1 хвилини при 95°С, 35 циклів, що складаються з циклу ампліфікації 30 секунд при 95°С і 1 хвилини при 70°С, і повторного нагрівання 7 хвилин при 70°С. Прямий праймер TPS-F1: 5'- СТС GAC TCG ТСС СТА ТТА GGC AAC AGC-3' (SEQ ID NO:138) Зворотний праймер SHygroR1: 5'- ТСА GAG GCA GTG GAG CCT CCA GTC AGC-3' (SEQ ID NO:139) Для способу 3 TP-F4 використовували як прямий праймер і RSGR-A використовували як зворотний праймер. Умови ПЛР складалися з попереднього нагрівання протягом 1 хвилини при 95°С, 35 циклів: циклу ампліфікації 30 секунд при 95°С, 30 секунд при 60°С і 2 хвилини при 72°С, і повторного нагрівання 7 хвилин при 72°С. У пробі клітин, що мають гомологічну рекомбінацію з вектором КО3, буде ампліфікуватися ДНК приблизно 1,6 т.п.н. Ця процедура ПЛР буде детектувати клітини, що мають гомологічну рекомбінацію з вектором КО3, а також клітини, які все ще мають гомологічну рекомбінацію з вектором КО2. 41 9. Результати ПЛР-скринінгу У Способі 1 аналізували в цілому 616 клітин, і 18 клітин, мабуть, були гомологічними рекомбінантами (ступінь гомологічної рекомбінації: 2,9%). У способі 2 аналізували в цілому 524 клітини, і 2 клітини, мабуть, були гомологічними рекомбінантами (ступінь гомологічної рекомбінації: 0,4%). Крім того, в Способі 3 аналізували в цілому 382 клітини, і 7 клітин, мабуть, були гомологічними рекомбінантами (ступінь гомологічної рекомбінації: 1,8%). 10. Блот-аналіз за Саузерном Блотинг за Саузерном проводили відповідно до описаного вище способу. Серед аналізованих клітин була виявлена 1 клітина, повністю позбавлена гена транспортера фукози. У стадії нокауту 1 результати аналізу ПЛР і блотингу за Саузерном узгоджуються, але в стадії нокауту 2 вони не узгоджуються. Можливі причини є наступними: 1. У Способі 1 змішували разом клітини, що мають гомологічну рекомбінацію незалежно від КО1 або КО2; 2. Ген транспортера фукози був присутнім не у вигляді однієї пари (2 генів), а у вигляді множинних пар (або у вигляді більше ніж 3 генів); і 3. Під час культивування клітинних ліній, створених в стадії нокауту 1, кількість копій гена транспортера фукози, що залишається в субкультивованих клітинах, збільшувалася. 11. Аналіз експресії фукози Експресію фукози аналізували за допомогою ПЛР в 26 клітинах, які, як було виявлено, є гомологічними рекомбінантами. У цілому 1x10 клітин забарвлювали на льоду протягом 1 години 100 мкл 93488 42 ЗФР, що містить 5 мкг/мл аглютиніну Lens culinaris, кон'югата FITC (Vector Laboratories, Cat. No. FL1041), 2,5% ФТС і 0,02% азиду натрію (далі званого FACS-розчином). Потім ці клітини промивали 3 рази FACS-розчином і аналізували з використанням FACSCalibur (Becton Dickinson). Ці результати ясно показали, що експресія фукози зменшується тільки в клітинах, які, як було виявлено в блотинганалізі за Саузерном, позбавлені гена транспортера фукози. Наведені вище результати виявили наступні: На основі того факту, що тільки одна клітина з 616 підданих скринінгу клітин має повну делецію гена транспортера фукози, частота гомологічної рекомбінації була дуже низькою, приблизно 0,16%. Як відмічалося вище, є декілька можливих причин того, що результати ПЛР і блотинг-аналізу за Саузерном не узгоджувалися. Однак, клітинні лінії, одержані в Способі 3, можуть не містити суміші клітин, що мають гомологічну рекомбінацію незалежно від векторів КО2 і КО3, оскільки відбір виконували з використанням двох типів лікарських засобів. Крім того, навряд чи всі інші клітинні лінії, що мають гомологічну рекомбінацію з використанням ПЛР, містять численні популяції клітин. Як відмічалося вище, якщо присутні 3 або більше генів транспортера фукози, націлювання на цей ген в клітинах було б особливо важким. Гомологічні рекомбінанти могли б бути одержані тільки з використанням такого вектора, як вектор КО1, де ген Hygr навряд чи експресується, і скринінгом великої кількості клітин. 43 93488 44 45 93488 46 47 93488 48 49 93488 50 51 93488 52 53 93488 54 55 93488 56 57 93488 58 59 93488 60
ДивитисяДодаткова інформація
Назва патенту англійськоюAnti-glypican 3 antibody having modified sugar chain
Автори англійськоюNakano Kiyotaka, Sugo Izumi, Sugimoto Masamichi, Ishiguro Takahiro, Tanaka Megumi, Iijima Shigeyuki
Назва патенту російськоюКомпозиция анти-глипикан 3-антител, которые имеют модифицированную сахарную цепь
Автори російськоюНакано Кийотака, Суго Изуми, Сугимото Масамити, Исигуро Такахиро, Танака Мегуми, Иидзима Сигеюки
МПК / Мітки
МПК: A61K 39/395, C12N 15/09, C07K 16/18, A61P 35/00, C12P 21/08
Мітки: модифікований, анти-гліпікан, ланцюг, композиція, 3-антитіл, мають, цукровий
Код посилання
<a href="https://ua.patents.su/81-93488-kompoziciya-anti-glipikan-3-antitil-shho-mayut-modifikovanijj-cukrovijj-lancyug.html" target="_blank" rel="follow" title="База патентів України">Композиція анти-гліпікан 3-антитіл, що мають модифікований цукровий ланцюг</a>
Химерне антитіло проти cd40 людини, молекула нуклеїнової кислоти, вектор експресії, фармацевтична композиція для лікування захворювань, опосередкованих т-клітинами, яка містить химерне антитіло

Номер патенту: 71909
Опубліковано: 17.01.2005
Автори: Ву Херрен, Сайдек Ентоні В., Бейорет Юрген, Херріс Лінда Дж., Беррі Карен К., Холленбау Даєн, Аруффо Аледжандро А., Хьюз Уільям Д., Торн Барбара А., Уоткінс Джеффрі Д.
МПК: A61P 29/00, C07K 16/28, C12N 15/09, A61P 43/00, A61K 39/395, A61P 37/06, C07K 14/46, C12N 15/13, C12N 1/21, C07K 14/725, A61P 37/02, A61P 19/02
Мітки: опосередкованих, містить, фармацевтична, кислоти, композиція, людини, захворювань, експресії, антитіло, нуклеїнової, т-клітинами, лікування, химерне, вектор, молекула, яка
Формула / Реферат:
1. Варіабельна ділянка легкого ланцюга химерного антитіла, що зв’язується з CD40 людини, яка містить амінокислотну послідовність SEQ ID NО:1 (Фіг. 4а).2. Варіабельна ділянка важкого ланцюга химерного антитіла, що зв’язується з CD40 людини, яка містить амінокислотну послідовність SEQ ID NО:2 (Фіг. 4b).3. Химерне антитіло, що зв'язується з CD40 людини, яке містить легкий і важкий ланцюг, причому зазначений легкий ланцюг містить ...
Сполуки, що мають гіпотензивну, кардіопротекторну, анти-ішемічну та антиліполітичну властивості, фармацевтична композиція та способи лікування
Номер патенту: 51716
Опубліковано: 16.12.2002
Автори: Чой-Следескі Йонг-Мі, Майєрс Майкл Р., Спада Альфред П., Магір Мартін П., Евінг Уільям Р., Полс Генрі В.
МПК: A61P 9/10, A61K 31/7076, A61K 31/52, C07D 473/34, A61P 9/12
Мітки: композиція, кардіопротекторну, гіпотензивну, фармацевтична, лікування, мають, антиліполітичну, сполуки, способи, властивості, анти-ішемічну
Формула / Реферат:
1. Сполука формули,де: К - N, N→ О або СН;Q - СН2 або О;R6 - водень, алкіл, аліл, 2-метилаліл, 2-бутеніл або циклоалкіл; X являє собою або ,де азот в кільці X заміщений Y; Е - О або S; Y - водень, алкіл, аралкіл, заміщений аралкіл, арил,заміщений арил, гетероцикліл, заміщений гетероцикліл, гетероциклілалкіл або заміщений гетероциклілалкіл; n та р- незалежно один від...
Тест-система імуноферментна для одночасного виявлення антигену р24 вірусу імунодефіциту людини першого та другого типів (віл- 1/2) та анти-віл антитіл (dia-hiv1/2-ag/at)

Номер патенту: 68845
Опубліковано: 16.08.2004
Автори: Касьяненко Тетяна Вальтерівна, Семиноженко Володимир Петрович, Ліщенко Людмила Петрівна, Ніколаєнко Ігор Васильович, Горлов Юрій Іванович, Терещенко Михайло Іванович, Пилипенко Віталій Григорович, Гураль Анатолій Леонтійович, Троянський Василь Васильович, Іванська Наіля Валєєвна, Раєвська Галина Євгенівна, Шевчук Олександр Анатолійович, Костенко Оксана Георгіївна
МПК: A61K 39/21
Мітки: вірусу, першого, анти-віл, імунодефіциту, другого, віл, антигену, людини, імуноферментна, антитіл, виявлення, одночасного, тест-система, типів
Формула / Реферат:
Тест-система імуноферментна для одночасного виявлення антигену р24 вірусу імунодефіциту людини першого та другого типів (ВІЛ-1/2) та анти-ВІЛ антитіл (DIA-HIVl/2-Ag/AT), в якій на полістироловий планшет сорбовані антитіла проти антигену р24 ВІЛ та очищені ВІЛ-1 і ВІЛ-2 антигени, а як кон'югат використовують біотинільовані анти-ВІЛ-1 антитіла та стрептавідин, зв'язаний з пероксидазою, яка відрізняється тим, що імуносорбент включає...
Анти-ефрр одноланцюжковий fv,молекула днк,яка його кодує, анти-ефрр антитіло,спосіб одержання анти-ефрр одноланцюжкового fv,спосіб одержання повного анти-ефрр антитіла та фармацевтична композиція (варіанти)

Номер патенту: 41929
Опубліковано: 15.10.2001
Автори: Бласко Франческ, Адан Жом, Бендіг Мері М., Мітжанс Франческ, П'юлат Жом, Кеттлбаре Кетрін А., Гюссов Детлеф, Анселл Кіт Х., Роселл Елізабет
МПК: C12P 21/08, C07K 16/28, C12N 15/09, C12N 15/13, G01N 33/53, A61P 35/00, C07K 16/00, A61K 39/395, C12P 21/02, G01N 33/563
Мітки: анти-ефрр, повного, кодує, одноланцюжковий, fv,спосіб, антитіла, варіанти, фармацевтична, днк,яка, композиція, fv,молекула, одноланцюжкового, антитіло,спосіб, одержання
Формула / Реферат:
1. Анти-ЭФРР одноцепочечный FV, полученный из фаг-антитело библиотеки, сконструированный из клеток иммунизированного млекопитающего, отличающийся тем, что вариабельные области одноцепочечного FV содержат тяжелую цепь аминокислотной последовательности выбранную из одной из...
Застосування анти-a5b1 антитіл для пригнічення проліферації ракових клітин

Номер патенту: 88294
Опубліковано: 12.10.2009
Автори: Бхаскар Вінай, Ло Деббі, Рамакрішнан Ваніта, Хо Сунь, Муррей Річард
МПК: A61P 35/00, A61K 38/00, A61K 39/395, A61K 39/00, C07K 16/00
Мітки: ракових, застосування, проліферації, антитіл, анти-a5b1, пригнічення, клітин
Формула / Реферат:
1. Спосіб пригнічення проліферації ракової клітини, яка експресує α5β1 інтегрин на своїй поверхні, у пацієнта, який включає введення пацієнту терапевтично ефективної кількості фармацевтичної композиції, яка включає рідку лікарську форму, що містить:приблизно від 1,0 мг/мл до 15 мг/мл анти-α5β1 антитіла; приблизно від 22 мМ до 27 мМ цитрату;приблизно від 145 мМ до 165 мМ хлориду натрію; приблизно від...
Попередній патент: Спосіб об’єднання технічних даних і система для обслуговування і нагляду промислової установки
Наступний патент: Гетероциклічні сполуки як антагоністи ccr2b
Випадковий патент: Термостатично керований змішувальний кран