Спосіб одержання поліненасичених жирних кислот в трансгенних організмах
Номер патенту: 90455
Опубліковано: 11.05.2010
Автори: Цанк Торстен, Шперлінг Петра, Хайнц Ернст, Домерг Фредерік, Квіу Ксяо, Цірпус Петра, Кірш Єлєна, Меєр Астрід, Бауер Йорг, Врінтен Патрісія, Аббаді Аміне








Формула / Реферат
1. Спосіб одержання сполук загальної формули І
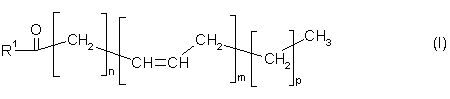
в трансгенних організмах із вмістом принаймні 1 мас.% даних сполук від загального вмісту ліпідів трансгенного організму, який відрізняється тим, що включає наступні етапи:
a) введення в організм принаймні однієї послідовності нуклеїнової кислоти, яка кодує Δ-9-елонгазну або Δ-6-десатуразну активність, та
b) введення в організм принаймні однієї послідовності нуклеїнової кислоти, яка кодує Δ-8-десатуразну або Δ-6-елонгазну активність, та
c) введення в організм принаймні однієї послідовності нуклеїнової кислоти, яка кодує Δ-5-десатуразну активність, та
d) введення в організм принаймні однієї послідовності нуклеїнової кислоти, яка кодує Δ-5-елонгазну активність, та
e) введення в організм принаймні однієї послідовності нуклеїнової кислоти, яка кодує Δ-4-десатуразну активність, та
де перемінні та замісники в формулі І мають наступні значення:
R1 = гідроксил-, коензимА-(тіоестер), лізофосфатидилхолін, лізофосфатидилетаноламін, лізофосфатидилгліцерол, лізодифосфатидилгліцерол, лізофосфатидилсерин, лізофосфатидилінозитол, сфінгооснова- або залишок загальної формули ІІ
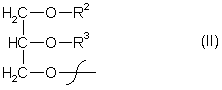
R2 = водень-, лізофосфатидилхолін, лізофосфатидилетаноламін, лізофосфатидилгліцерол, лізодифосфатидилгліцерол, лізофосфатидилсерин, лізофосфатидилінозитол або насичений або ненасичений C2-C24-алкілкарбоніл-,
R3 = водень, насичений або ненасичений C2-C24-алкілкарбоніл-, або R2 або R3 незалежно один від одного залишки загальної формули Іа:
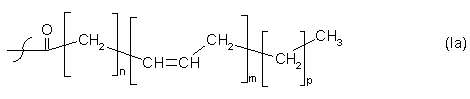
n = 2, 3, 4, 5, 6, 7 або 9, m = 2, 3, 4, 5 або 6 та p = 0 або 3.
2. Спосіб за пунктом формули 1, який відрізняється тим що, послідовності нуклеїнової кислоти, що кодують поліпептиди із Δ-9-елонгазною, Δ-6-десатуразною, Δ-8-десатуразною, Δ-6-елонгазною, Δ-5-десатуразною, Δ-5-елонгазною або Δ-4-десатуразною активністю, вибрані з групи, що включає:
a) послідовність нуклеїнової кислоти з наведених далі послідовностей SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5, SEQ ID NO:7, SEQ ID NO:9, SEQ ID NO:11, SEQ ID NO:13, SEQ ID NO:15, SEQ 1D NO:17, SEQ ID NO:19, SEQ ID NO:21, SEQ ID NO:23, SEQ ID NO:25, SEQ ID NO:27, SEQ ID NO:29, SEQ ID NO:31, SEQ ID NO:33, SEQ ID NO:35, SEQ ID NO:37, SEQ ID NO:39, SEQ ID NO:41, SEQ ID NO:43, SEQ ID NO:45, SEQ ID NO:47, SEQ ID NO:49, SEQ ID NO:51, SEQ ID NO:53, SEQ ID NO:59, SEQ ID NO:61, SEQ ID NO:63, SEQ ID NO:65, SEQ ID NO:67, SEQ ID NO:69, SEQ ID NO:71, SEQ ID NO:73, SEQ ID NO:75, SEQ ID NO:77, SEQ ID NO:79, SEQ ID NO:81, SEQ ID NO:83, SEQ ID NO:85, SEQ ID NO:89, SEQ ID NO:91, SEQ ID NO:93, SEQ ID NO:95, SEQ ID NO:97, SEQ ID NO:99, SEQ ID NO:101, SEQ ID NO:103, SEQ ID NO:111, SEQ ID NO:113, SEQ ID NO:117, SEQ ID NO:119, SEQ ID NO:131, SEQ ID NO:133, SEQ ID NO:135, SEQ ID NO:137 або SEQ ID NO:183 або
b) послідовності нуклеїнових кислот, які з урахуванням виродженості генетичного коду кодують наведені далі амінокислотні послідовності SEQ ID NO:2, SEQ ID NO:4, SEQ ID NO:6, SEQ ID NO:8, SEQ ID NO:10, SEQ ID NO:12, SEQ ID NO:14, SEQ ID NO:16, SEQ ID NO:18, SEQ ID NO:20, SEQ ID NO:22, SEQ ID NO: 24, SEQ ID NO:26, SEQ ID NO:28, SEQ ID NO:30, SEQ ID NO:32, SEQ ID NO:34, SEQ ID NO:36, SEQ ID NO:38, SEQ ID NO:40, SEQ ID NO:42, SEQ ID NO:44, SEQ ID NO:46, SEQ ID NO:48, SEQ ID NO:50, SEQ ID NO:52, SEQ ID NO:54, SEQ ID NO:60, SEQ ID NO:62, SEQ ID NO:64, SEQ ID NO:66, SEQ ID NO:68, SEQ ID NO:70, SEQ ID NO:72, SEQ ID NO:74, SEQ ID NO:76, SEQ ID NO:78, SEQ ID NO:80, SEQ ID NO:82, SEQ ID NO:84, SEQ ID NO:86, SEQ ID NO:88, SEQ ID NO:92, SEQ ID NO:94, SEQ ID NO:96, SEQ ID NO:98, SEQ ID NO:100, SEQ ID NO:102, SEQ ID NO:104, SEQ ID NO: 112, SEQ ID NO:114, SEQ ID NO:118, SEQ ID NO:120, SEQ ID NO:132, SEQ ID NO:134, SEQ ID NO:136, SEQ ID NO:138 або SEQ ID NO:184 або
c) похідні від наведених далі послідовностей нуклеїнових кислот SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5, SEQ ID NO:7, SEQ ID NO:9, SEQ ID NO:11, SEQ ID NO:13, SEQ ID NO:15, SEQ ID NO:17, SEQ ID NO:19, SEQ ID NO:21, SEQ ID NO:23, SEQ ID NO:25, SEQ ID NO:27, SEQ ID NO:29, SEQ ID NO:31, SEQ ID NO:33, SEQ ID NO:35, SEQ ID NO:37, SEQ ID NO:39, SEQ ID NO:41, SEQ ID NO:43, SEQ ID NO:45, SEQ ID NO:47, SEQ ID NO:49, SEQ ID NO:51, SEQ ID NO:53, SEQ ID NO:59, SEQ ID NO: 61, SEQ ID NO:63, SEQIDNO:65, SEQ ID NO:67, SEQ ID NO:69, SEQ ID NO:71, SEQ ID NO:73, SEQ ID NO:75, SEQ ID NO:77, SEQ ID NO:79, SEQ ID NO:81, SEQ ID NO:83, SEQ ID NO:85, SEQ ID NO:89, SEQ ID NO:91, SEQ ID NO:93, SEQ ID NO:95, SEQ ID NO:97, SEQ ID NO: 99, SEQ ID NO:101, SEQ ID NO:103, SEQ ID NO:111, SEQ ID NO:113, SEQ ID NO:117, SEQ ID NO:119, SEQ ID NO:131, SEQ ID NO:133, SEQ ID NO:135, SEQ ID NO:137 або SEQ ID NO:183, що кодують поліпептиди, які мають принаймні 60 % подібності до амінокислотних залишків послідовностей SEQ ID NO:2, SEQ ID NO:4, SEQ ID NO:6, SEQ ID NO:8, SEQ ID NO:10, SEQ ID NO:12, SEQ ID NO:14, SEQ ID NO:16, SEQ ID NO:18, SEQ ID NO:20, SEQIDNO:22, SEQ ID NO:24, SEQ ID NO:26, SEQ ID NO:28, SEQ ID NO:30, SEQ ID NO:32, SEQ ID NO:34, SEQ ID NO:36, SEQ ID NO:38, SEQ ID NO:40, SEQ ID NO:42, SEQ ID NO:44, SEQ ID NO:46, SEQ ID NO:48, SEQ ID NO:50, SEQ ID NO:52, SEQ ID NO:54, SEQ ID NO:60, SEQ ID NO:62, SEQ ID NO:64, SEQ ID NO:66, SEQ ID NO:68, SEQ ID NO:70, SEQ ID NO:72, SEQ ID NO:74, SEQ ID NO:76, SEQ ID NO:78, SEQ ID NO:80, SEQ ID NO:82, SEQ ID NO:84, SEQ ID NO:86, SEQ ID NO: 88, SEQ ID NO:92, SEQ ID NO:94, SEQ ID NO:96, SEQ ID NO:98, SEQ ID NO:100, SEQ ID NO:102, SEQ ID NO:104, SEQ ID NO:112, SEQ ID NO:114, SEQ ID NO:118, SEQ ID NO:120, SEQ ID NO:132, SEQ ID NO:134, SEQ ID NO:136, SEQ ID NO:138 або SEQ ID NO:184 та проявляють Δ-9-елонгазну, Δ-6-десатуразну, Δ-8-десатуразну, Δ-6-елонгазну, Δ-5-десатуразну, Δ-5-елонгазну або Δ-4-десатуразну активності.
3. Спосіб за пунктом формули 1 або 2, який відрізняється тим, що додатково в організм введено послідовність нуклеїнової кислоти, що кодує поліпептид з ω-3-десатуразною активністю, вибрану з групи, що складається з:
а) послідовності нуклеїнової кислоти з SEQ ID NO:87 або SEQ ID NO:105, представленої послідовності, або
b) послідовностей нуклеїнових кислот, які внаслідок виродженості генетичного коду являють собою наведені далі амінокислотні послідовності SEQ ID NO:88 або SEQ ID NO:106, або
c) похідних від наведених послідовностей нуклеїнової кислоти: SEQ ID NO:87 або SEQ ID NO:105, які кодують поліпептиди і мають, як мінімум, 60 % подібності до амінокислотних послідовностей, кодованих SEQ ID NO:88 або SEQ ID NO:106, та мають ω-3-десатуразну активність.
4. Спосіб за будь-яким з пунктів формули з 1 по 3, який відрізняється тим, що додатково в організм введено послідовність нуклеїнової кислоти, що кодує поліпептид із Δ-12-десатуразною активністю, вибрану з групи, що складається з:
a) послідовності нуклеїнової кислоти з SEQ ID NO:107 або SEQ ID NO:109, представленої послідовності, або
b) послідовностей нуклеїнової кислоти, які можуть бути виведені, як результат виродженості генетичного коду із наведених амінокислотних послідовностей: SEQ ID NO:108 або SEQ ID NO:110, або
c) похідних, представленої в SEQ ID NO:107 або SEQ ID NO:110 послідовності нуклеїнової кислоти, що кодують поліпептиди, які щонайменше на 60 % подібні амінокислотним ділянкам з SEQ ID NO:108 або SEQ ID NO:110 і проявляють Δ-12-десатуразну активність.
5. Спосіб за будь-яким з пунктів формули з 1 по 4, який відрізняється тим, що замісники R2 або R3 незалежно один від одного означають насичений або ненасичений С18-С22-алкілкарбоніл.
6. Спосіб за будь-яким з пунктів формули з 1 по 5, який відрізняється тим, що замісники R2 або R3 незалежно один від одного являють собою ненасичений С18-, С20- або С22-алкілкарбоніл із щонайменше двома подвійними зв'язками.
7. Спосіб за будь-яким з пунктів формули з 1 по 6, який відрізняється тим, що трансгенний організм є трансгенним мікроорганізмом або трансгенною рослиною.
8. Спосіб за будь-яким з пунктів формули з 1 по 7, який відрізняється тим, що трансгенний організм є олійною рослиною, овочевою рослиною або декоративною рослиною.
9. Спосіб за будь-яким з пунктів формули з 1 по 8, який відрізняється тим, що трансгенний організм є трансгенною рослиною, вибраною з групи сімейств рослин: Adetotheciaceae, Anacardiaceae, Asteraceae, Apiaceae, Betulaceae, Boraginaceae, Brassicaceae, Bromeliaceae, Caricaceae, Cannabaceae, Convolvulaceae, Chenopodiaceae, Crypthecodiniaceae, Cucurbitaceae, Ditrichaceae, Elaeagnaceae, Ericaceae, Euphorbiaceae, Fabaceae, Geraniaceae, Gramineae, Juglandaceae, Lauraceae, Leguminosae, Linaceae або Prasinophyceae.
10. Спосіб за будь-яким з пунктів формули з 1 по 9, який відрізняється тим, що сполуки загальної формули І є виділеними з організму у формі олій, ліпідів або вільних жирних кислот.
11. Спосіб за будь-яким з пунктів формули з 1 по 10, який відрізняється тим, що сполуки загальної формули І містяться в концентрації принаймні 5 ваг. % від загального вмісту ліпідів, виділених з трансгенного організму.
Текст
1. Спосіб одержання сполук загальної формули І CH3 CH2 (I) p в трансгенних організмах із вмістом принаймні 1 мас.% даних сполук від загального вмісту ліпідів трансгенного організму, який відрізняється тим, що включає наступні етапи: a) введення в організм принаймні однієї послідовності нуклеїнової кислоти, яка кодує Γ-9-елонгазну або Γ-6-десатуразну активність, та b) введення в організм принаймні однієї послідовності нуклеїнової кислоти, яка кодує Γ-8десатуразну або Γ-6-елонгазну активність, та c) введення в організм принаймні однієї послідовності нуклеїнової кислоти, яка кодує Γ-5десатуразну активність, та d) введення в організм принаймні однієї послідовності нуклеїнової кислоти, яка кодує Γ-5-елонгазну активність, та e) введення в організм принаймні однієї послідовності нуклеїнової кислоти, яка кодує Γ-4десатуразну активність, та де перемінні та замісники в формулі І мають наступні значення: 1 R = гідроксил-, коензимА-(тіоестер), лізофосфатидилхолін, лізофосфатидилетаноламін, лізофосфатидилгліцерол, лізодифосфатидилгліцерол, лізофосфатидилсерин, лізофосфатидилінозитол, сфінгооснова- або залишок загальної формули ІІ H2C O R HC O R H2C 2 O 2 3 (II) R =водень-, лізофосфатидилхолін, лізофосфатидилетаноламін, лізофосфатидилгліцерол, лізодифосфатидилгліцерол, лізофосфатидилсерин, лі C2 m (13) n CH2 CH CH 90455 CH2 (11) 1 UA O R (19) (21) a200602255 (22) 16.07.2004 (24) 11.05.2010 (86) PCT/EP2004/007957, 16.07.2004 (31) 103 35 992.3 (32) 01.08.2003 (33) DE (31) 103 44 557.9 (32) 24.09.2003 (33) DE (31) 103 47 869.8 (32) 10.10.2003 (33) DE (46) 11.05.2010, Бюл.№ 9, 2010 р. (72) ЦАНК ТОРСТЕН, DE, БАУЕР ЙОРГ, DE, ЦІРПУС ПЕТРА, DE, АББАДІ АМІНЕ, DE, ХАЙНЦ ЕРНСТ, DE, КВІУ КСЯО, CN/CA, ВРІНТЕН ПАТРІСІЯ, CA, ШПЕРЛІНГ ПЕТРА, DE, ДОМЕРГ ФРЕДЕРІК, FR/DE, МЕЄР АСТРІД, DE, КІРШ ЄЛЄНА, DE (73) БАСФ ПЛАНТ САЙЄНС ГМБХ, DE (56) DREXLER H ET AL: "Metabolic engineering of fatty acids for breeding of new oilseed crops: Strategies, problems and first results" JOURNAL OF PLANT PHYSIOLOGY, FISCHER, STUTTGART, DE, Bd. 160, Nr. 7, Juli 2003 (2003-07), Seiten 779-802, BEAUDOIN FREDERIC ET AL: "Heterologous reconstitution in yeast of the polyunsaturated fatty acid biosynthetic pathway" PROCEEDINGS OF THE NATIONAL ACADEMY OF SCIENCES OF USA, NATIONAL ACADEMY OF SCIENCE. WASHINGTON, US, Bd. 97, Nr. 12, 6. Juni 2000 (2000-06-06), Seiten 6421-6426, MEYER ASTRID ET AL: "Novel fatty acid elongases and their use for the reconstitution of docosahexaenoic acid biosynthesis." JOURNAL OF LIPID RESEARCH. OCT 2004, Bd. 45, Nr. 10, Oktober 2004 (2004-10), Seiten 1899-1909, DOMERGUE F ET AL: "Cloning and functional characterization of Phaeodactylum tricornutum frontend desaturases involved in eicosapentaenoic acid biosynthesis" EUROPEAN JOURNAL OF BIOCHEMISTRY, BERLIN, DE, Bd. 269, Nr. 16, August 2002 (2002-08), Seiten 4105-4113, ZANK T K ET AL: "Cloning and functional expression of the first plant fatty acid elongase specific for 2 3 90455 зофосфатидилінозитол або насичений або ненасичений C2-C24-алкілкарбоніл-, 3 R = водень, насичений або ненасичений C2-C242 3 алкілкарбоніл-, або R або R незалежно один від одного залишки загальної формули Іа: O CH3 CH2 CH2 n (Ia) CH2 CH CH m p n = 2, 3, 4, 5, 6, 7 або 9, m = 2, 3, 4, 5 або 6 та p = 0 або 3. 2. Спосіб за пунктом формули 1, який відрізняється тим, що послідовності нуклеїнової кислоти, що кодують поліпептиди із Γ-9-елонгазною, Γ-6десатуразною, Γ-8-десатуразною, Γ-6-елонгазною, Γ-5-десатуразною, Γ-5-елонгазною або Γ-4десатуразною активністю, вибрані з групи, що включає: a) послідовність нуклеїнової кислоти з наведених далі послідовностей SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5, SEQ ID NO:7, SEQ ID NO:9, SEQ ID NO:11, SEQ ID NO:13, SEQ ID NO:15, SEQ 1D NO:17, SEQ ID NO:19, SEQ ID NO:21, SEQ ID NO:23, SEQ ID NO:25, SEQ ID NO:27, SEQ ID NO:29, SEQ ID NO:31, SEQ ID NO:33, SEQ ID NO:35, SEQ ID NO:37, SEQ ID NO:39, SEQ ID NO:41, SEQ ID NO:43, SEQ ID NO:45, SEQ ID NO:47, SEQ ID NO:49, SEQ ID NO:51, SEQ ID NO:53, SEQ ID NO:59, SEQ ID NO:61, SEQ ID NO:63, SEQ ID NO:65, SEQ ID NO:67, SEQ ID NO:69, SEQ ID NO:71, SEQ ID NO:73, SEQ ID NO:75, SEQ ID NO:77, SEQ ID NO:79, SEQ ID NO:81, SEQ ID NO:83, SEQ ID NO:85, SEQ ID NO:89, SEQ ID NO:91, SEQ ID NO:93, SEQ ID NO:95, SEQ ID NO:97, SEQ ID NO:99, SEQ ID NO:101, SEQ ID NO:103, SEQ ID NO:111, SEQ ID NO:113, SEQ ID NO:117, SEQ ID NO:119, SEQ ID NO:131, SEQ ID NO:133, SEQ ID NO:135, SEQ ID NO:137 або SEQ ID NO:183 або b) послідовності нуклеїнових кислот, які з урахуванням виродженості генетичного коду кодують наведені далі амінокислотні послідовності SEQ ID NO:2, SEQ ID NO:4, SEQ ID NO:6, SEQ ID NO:8, SEQ ID NO:10, SEQ ID NO:12, SEQ ID NO:14, SEQ ID NO:16, SEQ ID NO:18, SEQ ID NO:20, SEQ ID NO:22, SEQ ID NO: 24, SEQ ID NO:26, SEQ ID NO:28, SEQ ID NO:30, SEQ ID NO:32, SEQ ID NO:34, SEQ ID NO:36, SEQ ID NO:38, SEQ ID NO:40, SEQ ID NO:42, SEQ ID NO:44, SEQ ID NO:46, SEQ ID NO:48, SEQ ID NO:50, SEQ ID NO:52, SEQ ID NO:54, SEQ ID NO:60, SEQ ID NO:62, SEQ ID NO:64, SEQ ID NO:66, SEQ ID NO:68, SEQ ID NO:70, SEQ ID NO:72, SEQ ID NO:74, SEQ ID NO:76, SEQ ID NO:78, SEQ ID NO:80, SEQ ID NO:82, SEQ ID NO:84, SEQ ID NO:86, SEQ ID NO:88, SEQ ID NO:92, SEQ ID NO:94, SEQ ID NO:96, SEQ ID NO:98, SEQ ID NO:100, SEQ ID NO:102, SEQ ID NO:104, SEQ ID NO: 112, SEQ ID NO:114, SEQ ID NO:118, SEQ ID NO:120, SEQ ID NO:132, SEQ ID NO:134, SEQ ID NO:136, SEQ ID NO:138 або SEQ ID NO:184 або c) похідні від наведених далі послідовностей нуклеїнових кислот SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5, SEQ ID NO:7, SEQ ID NO:9, SEQ ID NO:11, SEQ ID NO:13, SEQ ID NO:15, SEQ ID 4 NO:17, SEQ ID NO:19, SEQ ID NO:21, SEQ ID NO:23, SEQ ID NO:25, SEQ ID NO:27, SEQ ID NO:29, SEQ ID NO:31, SEQ ID NO:33, SEQ ID NO:35, SEQ ID NO:37, SEQ ID NO:39, SEQ ID NO:41, SEQ ID NO:43, SEQ ID NO:45, SEQ ID NO:47, SEQ ID NO:49, SEQ ID NO:51, SEQ ID NO:53, SEQ ID NO:59, SEQ ID NO: 61, SEQ ID NO:63, SEQIDNO:65, SEQ ID NO:67, SEQ ID NO:69, SEQ ID NO:71, SEQ ID NO:73, SEQ ID NO:75, SEQ ID NO:77, SEQ ID NO:79, SEQ ID NO:81, SEQ ID NO:83, SEQ ID NO:85, SEQ ID NO:89, SEQ ID NO:91, SEQ ID NO:93, SEQ ID NO:95, SEQ ID NO:97, SEQ ID NO: 99, SEQ ID NO:101, SEQ ID NO:103, SEQ ID NO:111, SEQ ID NO:113, SEQ ID NO:117, SEQ ID NO:119, SEQ ID NO:131, SEQ ID NO:133, SEQ ID NO:135, SEQ ID NO:137 або SEQ ID NO:183, що кодують поліпептиди, які мають принаймні 60 % подібності до амінокислотних залишків послідовностей SEQ ID NO:2, SEQ ID NO:4, SEQ ID NO:6, SEQ ID NO:8, SEQ ID NO:10, SEQ ID NO:12, SEQ ID NO:14, SEQ ID NO:16, SEQ ID NO:18, SEQ ID NO:20, SEQIDNO:22, SEQ ID NO:24, SEQ ID NO:26, SEQ ID NO:28, SEQ ID NO:30, SEQ ID NO:32, SEQ ID NO:34, SEQ ID NO:36, SEQ ID NO:38, SEQ ID NO:40, SEQ ID NO:42, SEQ ID NO:44, SEQ ID NO:46, SEQ ID NO:48, SEQ ID NO:50, SEQ ID NO:52, SEQ ID NO:54, SEQ ID NO:60, SEQ ID NO:62, SEQ ID NO:64, SEQ ID NO:66, SEQ ID NO:68, SEQ ID NO:70, SEQ ID NO:72, SEQ ID NO:74, SEQ ID NO:76, SEQ ID NO:78, SEQ ID NO:80, SEQ ID NO:82, SEQ ID NO:84, SEQ ID NO:86, SEQ ID NO: 88, SEQ ID NO:92, SEQ ID NO:94, SEQ ID NO:96, SEQ ID NO:98, SEQ ID NO:100, SEQ ID NO:102, SEQ ID NO:104, SEQ ID NO:112, SEQ ID NO:114, SEQ ID NO:118, SEQ ID NO:120, SEQ ID NO:132, SEQ ID NO:134, SEQ ID NO:136, SEQ ID NO:138 або SEQ ID NO:184 та проявляють Γ-9-елонгазну, Γ-6-десатуразну, Γ-8десатуразну, Γ-6-елонгазну, Γ-5-десатуразну, Γ-5елонгазну або Γ-4-десатуразну активності. 3. Спосіб за пунктом формули 1 або 2, який відрізняється тим, що додатково в організм введено послідовність нуклеїнової кислоти, що кодує поліпептид з ω-3-десатуразною активністю, вибрану з групи, що складається з: а) послідовності нуклеїнової кислоти з SEQ ID NO:87 або SEQ ID NO:105, представленої послідовності, або b) послідовностей нуклеїнових кислот, які внаслідок виродженості генетичного коду являють собою наведені далі амінокислотні послідовності SEQ ID NO:88 або SEQ ID NO:106, або c) похідних від наведених послідовностей нуклеїнової кислоти: SEQ ID NO:87 або SEQ ID NO:105, які кодують поліпептиди і мають, як мінімум, 60 % подібності до амінокислотних послідовностей, кодованих SEQ ID NO:88 або SEQ ID NO:106, та мають ω-3-десатуразну активність. 4. Спосіб за будь-яким з пунктів формули з 1 по 3, який відрізняється тим, що додатково в організм введено послідовність нуклеїнової кислоти, що кодує поліпептид із Γ-12-десатуразною активністю, вибрану з групи, що складається з: 5 90455 6 a) послідовності нуклеїнової кислоти з SEQ ID NO:107 або SEQ ID NO:109, представленої послідовності, або b) послідовностей нуклеїнової кислоти, які можуть бути виведені, як результат виродженості генетичного коду із наведених амінокислотних послідовностей: SEQ ID NO:108 або SEQ ID NO:110, або c) похідних, представленої в SEQ ID NO:107 або SEQ ID NO:110 послідовності нуклеїнової кислоти, що кодують поліпептиди, які щонайменше на 60 % подібні амінокислотним ділянкам з SEQ ID NO:108 або SEQ ID NO:110 і проявляють Γ-12десатуразну активність. 5. Спосіб за будь-яким з пунктів формули з 1 по 4, 2 3 який відрізняється тим, що замісники R або R незалежно один від одного означають насичений або ненасичений С18-С22-алкілкарбоніл. 6. Спосіб за будь-яким з пунктів формули з 1 по 5, 2 3 який відрізняється тим, що замісники R або R незалежно один від одного являють собою ненасичений С18-, С20- або С22-алкілкарбоніл із щонайменше двома подвійними зв'язками. 7. Спосіб за будь-яким з пунктів формули з 1 по 6, який відрізняється тим, що трансгенний організм є трансгенним мікроорганізмом або трансгенною рослиною. 8. Спосіб за будь-яким з пунктів формули з 1 по 7, який відрізняється тим, що трансгенний організм є олійною рослиною, овочевою рослиною або декоративною рослиною. 9. Спосіб за будь-яким з пунктів формули з 1 по 8, який відрізняється тим, що трансгенний організм є трансгенною рослиною, вибраною з групи сімейств рослин: Adetotheciaceae, Anacardiaceae, Asteraceae, Apiaceae, Betulaceae, Boraginaceae, Brassicaceae, Bromeliaceae, Caricaceae, Cannabaceae, Convolvulaceae, Chenopodiaceae, Crypthecodiniaceae, Cucurbitaceae, Ditrichaceae, Elaeagnaceae, Ericaceae, Euphorbiaceae, Fabaceae, Geraniaceae, Gramineae, Juglandaceae, Lauraceae, Leguminosae, Linaceae або Prasinophyceae. 10. Спосіб за будь-яким з пунктів формули з 1 по 9, який відрізняється тим, що сполуки загальної формули І є виділеними з організму у формі олій, ліпідів або вільних жирних кислот. 11. Спосіб за будь-яким з пунктів формули з 1 по 10, який відрізняється тим, що сполуки загальної формули І містяться в концентрації принаймні 5 ваг. % від загального вмісту ліпідів, виділених з трансгенного організму. Даний винахід стосується способу одержання поліненасичених жирних кислот в організмі, в який були введені нуклеїнові кислоти, які кодують поліпептиди із -5 елонгазною активністю. Переважно, ці послідовності нуклеїнових кислот, за необхідності, з іншими послідовностями нуклеїнових кислот, що кодують поліпептиди біосинтезу жирних кислот або метаболізму ліпідів, можуть експресуватися в організмі. Особливо переважними є послідовності нуклеїнових кислот, що кодують -6-десатуразну. -5 десатуразну, -4-десатуразну, -12десатуразну та/або -6-елонгазну активність. Переважно ці десатурази та елонгази походять із Thalassiosira, Euglena або Ostreococcus. Також винахід стосується способу одержання олій та/або тригліцеридів із підвищеним вмістом довголанцюгових поліненасичених жирних кислот. Даний винахід відноситься, крім того, в переважній формі втілення винаходу до способу одержання ненасичених -3-жирних кислот, а також способу одержання тригліцеридів із підвищеним вмістом ненасичених жирних кислот, особливо, 3-жирних кислот із більш ніж трьома подвійними зв'язками. Винахід також стосується одержання трансгенного організму, переважно, трансгенної рослини або трансгенного мікроорганізму з підвищеним вмістом ненасичених -3-жирних кислот, олій або ліпідів із -3-подвійними зв'язками як наслідок експресії в способі використаних елонгаз та десатураз згідно даного винаходу, переважно, в комбінації із -3-десатуразами, наприклад, -3десатуразами із грибів сімейства Pythiaceae такими як рід Phytophtora, наприклад рід та вид Phytophtora infestans, або -3-десатуразами із водоростей, таких як сімейство Prasmophyceae наприклад, рід Ostreococcus та, зокрема, рід та вид Ostreococcus tauri або Diatomaceaе, такі як вид Thalassiosira та, зокрема, рід та вид Thalassiosira pseudonana. Винахід також стосується послідовностей нуклеїнових кислот, конструктів нуклеїнових кислот, векторів та організмів, що містять нуклеїнові кислоти та/або конструкти нуклеїнових кислот, а також трансгенних організмів, які містять вище згадані послідовності нуклеїнових кислот, конструкти нуклеїнових кислот та/або вектори. Наступна частина винаходу стосується олій, ліпідів та/або жирних кислот, одержаних способом згідно даного винаходу, та їх використання. Також винахід стосується ненасичених жирних кислот, а також тригліцеридів із підвищеним вмістом ненасичених жирних кислот та їхнього застосування. Жирні кислоти та триацилгліцериди мають широке застосування в харчовій промисловості, кормах для тварин, косметиці та фармацевтиці. Оскільки йдеться про вільні насичені або ненасичені жирні кислоти або триацилгліцериди, які мають підвищений вміст насичених або ненасичених жирних кислот, то вони можуть мати різноманітне застосування. Поліненасичені жирні кислоти, такі як лінолева та ліноленова кислота, є незамінними для ссавців, оскільки вони не можуть їх самі продукувати. От чому поліненасичені -3-жирні кислоти та -6-жирні кислоти є важливою складовою харчування тварин та людей. Поліненасичені довголанцюгові -3-жирні кислоти, такі як ейкозапентаєнова кислота (=ЕРА, 7 С20:5 ) або докозагексаєнова кислота 4,7,10,13,16,19 (=DHA, С22.6 ), є важливими компонентами харчування людини, оскільки відіграють важливу роль в таких аспектах здоров'я, як розвиток дитячого мозку, функціонування очей, синтез гормонів та інших сигнальних речовин, також схильність до хвороби Herz-Kreislauf, Krebs та діабету (Poulos, A Lipids 30:1-14,1995; Horrocks, LA and Yeo YK Pharmacol Res 40:211-225, 1999). На підставі цього виникає питання про виробництво поліненасичених довголанцюгових жирних кислот. Основою сучасного звичайного складу харчування людини є добавки поліненасичених -3жирних кислот, які є важливими для харчування та переважно зустрічаються у риб'ячому жирі. Таким чином, прикладами поліненасичених жирних кислот можуть бути докозагексаєнова кислота (=DHA, 4,7,10,13,16,19 С22.6 ) або ейкозапентаєнова кислота 5,8,11,14,17 (=ЕРА, С20:5 ), які мають надзвичайну харчову цінність в дитячому харчуванні. Ненасиченій жирній кислоті DHA приписують позитивний вплив на розвиток та забезпечення функціонування головного мозку. В подальшому, поліненасичені жирні кислоти, такі як PUFA, PUFAs, LCPUFA або LCPUFAs, означатимуть (polyunsaturated fatty acids, PUFA, поліненасичені жирні кислоти; long chain poly unsaturated fatty acids, LCPUFA, довголанцюгові поліненасичені жирні кислоти). Переважно, різноманітні жирні кислоти та тригліцериди одержують з мікроорганізмів, таких як Mortierella або Schizochytnm або з олійних рослин, таких як соя, рапс, водоростей, таких як Crypthecodinium або Phaeodactylum та інших, причому вони, як правило, накопичуються у формі триацилгліцеридів (=Тригліцерид =Тригліцерол). Також вони можуть бути одержані із тварин, наприклад, риб. Вільні жирні кислоти можуть бути одержані омиленням. Поліненасичені жирні кислоти з дуже довгими ланцюгами, такі як DHA, ЕРА, 5,8,11,14 арахідонова кислота (=ARA, С20:4 ), дигомо8,11,14 -ліноленова кислота (С20:3 ) або докозапен7,10,13,16,19 таєнова кислота (DPA, С22:5 ) не синтезуються в олійних культурах, таких як рапс, соя, соняшник, сафлор красильний. Звичайними природними джерелами таких жирних кислот є риби, такі як оселедець, лосось, сардина, йорж, вугор, короп, форель, палтус, макрель, судак або тунець або водорості. Залежно від мети застосування переважними є олії із насиченими або ненасиченими жирними кислотами. Тобто, наприклад, в харчуванні людини, переважними є ліпіди із ненасиченими жирними кислотами, особливо поліненасиченими жирними кислотами. Поліненасиченим -3-жирним кислотам приписують при цьому позитивний вплив на рівень холестерину в крові й, таким чином, на можливість запобігання серцевим захворюванням. Додаванням цих -3-жирних кислот до харчування суттєво знижується ризик серцевих захворювань, апоплексії або високого кров'яного тиску. Також 3-жирні кислоти справляють позитивний вплив на запальні, особливо, хронічні запальні процеси при імунологічних захворюваннях, таких як ревматоїд5,8,11,14,17 90455 8 ний артрит. От чому -6-жирні кислоти, такі як арахідонова кислота, які знаходять своє застосування в харчових продуктах, особливо дієтичних харчових продуктах або медикаментах, мають на фоні звичайного складу харчування скоріше негативний вплив на перебіг таких хвороб як ревматичні хвороби. -3- та -6-жирні кислоти є попередниками тканинних гормонів, так званих ейкозаноідів, таких як простагландини, які походять від дигомо-ліноленової кислоти, арахідонової кислоти та ейкозапентаєнової кислоти, тромоксагену та лейкотриєну, які походять від арахідонової кислоти та ейкозапентаєнової кислоти. Ейкозаноіди (т.зв. РG2-сімейство), які утворюються із -6-жирних кислот, як правило, сприяють запальним реакціям, тоді як ейкозаноіди (т.зв. РG3-сімейство), які утворюються із -3-жирних кислот мають незначний або зовсім не мають впливу на запалення. Зважуючи на їхні позитивні характеристики, в минулому було достатньо спроб одержати гени, які були б задіяні у синтезі жирних кислот або тригліцеридів для одержання олій в різних організмах із зміненим вмістом ненасичених жирних кислот. Так в WO91/13972 та його US-еквіваленті була описана -9-десатураза В WO93/11245 була заявлена -15-десатураза та в WO94/11516 заявлено -12-десатуразу. Також десатурази було описано в ЕР-А-0-550-162, WO94/18337, WO97/30582, WO97/21340, WO95/18222, ЕР-А-0-794-250, Stukey та ін., J.Biol.Chem., 265, 1990:20144-20149, Wada та ін., Nature 347, 1990:200-203 або Huang та ін., Lipids 34, 1999-649-659. Біохімічні характеристики різних десатураз ще до тепер недостатньо досліджені, ці вбудовані в мембрану ферменти є досить складними для виділення та дослідження (МсKеоn та ін., Methods in EnzyMol. 71, 1981:12141-12147, Wang та ін., Plant Physio.Biochem, 26, 1988:777792). Тут, як правило, описуються вбудовані в мембрану десатурази введенням у відповідний організм, та вивчення ферментної активності за допомогою дослідження едукту та продукту. -6десатурази були описані в WO93/06712, US5,614,393, US5,614,393, WO96/21022, WO00/21557 та WO99/27111. Також, у WO98/46763 WO98/46764, WO98/46765 було описано їхнє застосування та одержання в трансгенних організмах. Також, було описано та обговорено в WO99/64616 або WO98/46776 експресію різних десатураз та утворення поліненасичених жирних кислот, а також було відмічено ефективність експресії десатураз та їхній вплив на утворення поліненасичених жирних кислот, а саме те, що за допомогою експресії окремих десатураз, як було описано раніше, було досягнуто лише незначного вмісту поліненасичених жирних кислот/ліпідів, таких як -ліноленова кислота та стеаридонова кислота. Також, як правило, в одній суміші містилися -3- та -6-жирні кислоти. Особливо придатними мікроорганізмами для одержання PUFAs були такі мікроорганізми, як мікроводорості, такі як Phaeodactylum tricornutum, роди Porphindium роди Thraustochytrium, роди Schizochytrium або роди Crypthecodinium, Ciliates 9 такі як Stylonychia або Colpidium, гриби, такі як Mortierella, Entomophthora або Mucor та/або Moosen, такі як Physcomitrella, Ceratodon та Marchantia (R.Vazhappilly & F.Chen (1998) Botanic Marina 41:553-558, K.Totani & K.Oba (1987) Lipids 22:1060-1062; Μ.Akimoto та ін. (1998) Appl.Biochemistry and Biotechnology 73:269-278). Шляхом селекції штамів було одержано певну кількість мутантних штамів даного мікроорганізму, які виробляють ряд бажаних сполук, включаючи PUFAs. Мутація та селекція штамів із поліпшеним виробленням певних молекул, таких як поліненасичені жирні кислоти є однак тривалим та складним процесом. От чому завжди, коли можливо, переважно застосовується вище описаний генетично-технологічний спосіб. За допомогою вище описаних мікроорганізмів може вироблятися лише певний перелік бажаних сумішей поліненасичених жирних кислот, таких як DPA, ЕРА або ARA. При чому, як правило, може використовуватися такий мікроорганізм, який накопичує суміші жирних кислот, наприклад, ЕРА, DPA та ARA. Для синтезу арахідонової, ейкозапентаєнової (ЕРА) та докозагексаєнової (DHA) кислоти розглядалися різні шляхи синтезу (Фігура 1). Отже, вироблення ЕРА або DHA в морських бактеріях, таких як Vibrio sp. або Shewanella sp. відбувається полікетидним шляхом (Yu, R. та ін. Lipids 35:10611064, 2000; Takeyama, Ζ. та ін. Microbiology - 143: 2725-2731, 1997). Альтернативна стратегія полягає в змінній дії десатураз та елонгаз (Zank, Τ К та ін. Plant Journal 31:255-268, 2002; Sakuradani, E. та ін. Gene 238 445-453, 1999). Модифікація у ссавців описаних шляхів -6-десатурази, -6-елонгази, -5десатурази -5-елонгази, -4-десатурази є шляхом синтезу Sprecher (Sprecher 2000, Biochim Biophys. Acta 1486:219-231). Замість -4десатураци відбувається додатковий етап елонгації С24, -6-десатурація та, на завершення, окислення до довжини ланцюга С22. Зрозуміло що для одержання в рослинах та мікроорганізмах, так званий шлях синтезу Sprecher (дивись Фігуру 1) є непридатним, оскільки є невідомими механізми його регуляції. Поліненасичені жирні кислоти можуть бути поділені відповідно до їх нього типу десатурованості у два великих класи, -б- або -3-жирні кислоти, що мають різні метаболічні та функціональні властивості (Фіг.1). Вихідним продуктом для -6метаболічного шляху є жирна кислота -лінолева 9,12 кислота (18:2 ), тоді як -3-шлях завершується 9,12,15 ліноленовою кислотою (18:3 ). Ліноленова кислота при цьому утворюється завдяки дії -3десатурази (Tocher та ін. 1998, Prog.Lipid Res.37, 73-117, Domergue та ін. 2002, Eur.J.Biochem 269, 4105-4113). Ссавці, в тому числі й людина, не мають відповідної десатуразної активності (-12 та -3десатурази) та повинні поповнювати з їжею ці жирні кислоти (незамінні жирні кислоти). При послідовних реакціях десатурази та елонгази синтезуються із вихідних продуктів фізіологічно важливі поліненасичені жирні кислоти - арахідонова кисло 90455 10 та (=АRА 20:4 ), яка є -6-жирною кислотою та дві -3-жирні кислоти: ейкозапентаєнова (=ЕРА, 5,8,11,14,17 С20:5 ) та докозапентаєнова кислота 4,7,10,13,17,19 (DHA, С22.6 ). Показано використання 3-жирних кислот, яке, як описано вище, полягає в терапевтичному ефекті при лікуванні серцевосудинних захворювань (Shimikawa 2001, World Rev.Nutr.Diet.88, 100-108), запалень (Calder 2002, Proc.Nutr.Soc.61, 345-358) та артриту (Cleland and James 2000, J.Rheumatol.27, 2305-2307). Тому з точки зору фізіологічних норм харчування є важливим добитися при синтезі поліненасичених жирних кислот зміщення між -6метаболічного шляху синтезу та -3метаболічного шляху синтезу (дивись Фігуру 1), у бік збільшення продукування -3-жирних кислот. В літературі було описано ферментативні активності різних -3-десатураз, які десатурують С18:2- С22:4або С22:5-жирні кислоти (дивись Фігуру 1). Однак жодна з біохімічно описаних десатураз не конвертує широкий спектр субстратів шляхів -6-синтезу на шляхи -3-синтезу відповідних жирних кислот. Тому i надалі існує велика потреба у -3десатуразі, яка є придатною для одержання -3поліненасичених жирних кислот Всі відомі рослинні та ціанобактеріальні -3-десатурази десатурують С18-жирні кислоти із лінолевою кислотою у якості субстрату однак не здатні до десатурації С20- або С22-жирних кислотю Відомою є -3-десатураза гриба Saprolegnia dicilina [Pereira та ін 2004б Biochem.J 378 (Pt 2):665-71], яка здатна десатурувати С20поліненасичені жирні кислоти. Однак недоліком є те, що ці -3-десатурази не можуть десатурувати C18- або C22-PUFAs, а саме, такі важливі жирні кислоти як С18:2-, С2:24- або С22:5-жирні кислоти ω-6шляху синтезу Наступним недоліком цього ферменту є його нездатність десатурувати жирні кислоти сполучені із фосфоліпідами. Можна переробляти лише складні естери СоА- та жирної кислоти. Подовження жирних кислот елонгазами на 2 або 4 атоми вуглецю є важливим для одержання С20- або C22-PUFAs. Цей процес відбувається у 4 стадії. Перший етап полягає в конденсації малоніл-СоА до жирнокислотного-ацил-СоА за допомогою кетоацил-СоА-синтази (KCS, в подальшому тексті буде позначатися як елонгаза). За цим слідує етап скорочення (кетоацил-СоА-редуктаза, KCR), дегідратація (дептратаза) та заключний етап скорочення (еноіл-СоА-редуктаза). Було встановлено що активність елонгази впливає на специфічність та швидкість цілого процесу (Millar and Kunst, 1997 Plant Journal 12:121-131). В минулому робилися численні спроби одержання генів елонгази. В роботі Millar та Kunst, 1997 (Plant Journal 12:121-131) та в Millar та ін. 1999, (Plant Cell 11:825-838) описано характеристики рослинних елонгаз синтезу мононенасичених довголанцюгових жирних кислот (С22:1) або для синтезу дуже довголанцюгових жирних кислот при утворенні восків у рослинах (С28-С32). Опис синтезу арахідонової кислоти та ЕРА знаходиться, наприклад в WO01/59128, WO00/12720, WO02/077213 та WO02/08401. Синтез поліненаси5,8,11,14 11 чених С24 жирних кислот описано, наприклад в Tvrdik et al 2000, JCB 149:707-717 або WO02/44320. Досі не було описано жодної специфічної елонгази, для одержання DHA (С22:6 н-3) в організмах, які природно її не виробляють. Досі були описані лише елонгази для одержування С20- або С24жирних кислот. Досі не була описана -5елонгазна активність. Вищі рослини містять поліненасичені жирні кислоти, такі як лінолева кислота (С18:2) та ліноленова кислота (С18:3). ARA, ЕРА та DHA майже не зустрічаються в насіннєвій олії вищих рослин або присутні лише в слідових кількостях (E.Ucciani: Nouveau Dictionnaire des Huiles Vegetales. Technique & Documentation-Lavoisier, 1995. ISBN:27430-0009-0). Було б вигідно, виробляти, переважно у вищих рослинах, особливо в олійних рослинах, таких як рапс, льон, соняшник та соя LCPUFAs й таким чином одержувати великі кількості високоякісних LCPUFAs для харчової промисловості, тваринного харчування та фармацевтичних потреб. До того ж, методами генетичної інженері) мають бути створені та досліджені придатні гени, що кодують ферменти біосинтезу LCPUFAs в олійних культурах. Це є гени, які кодують, наприклад, -6-десатурази, -6-елонгази, -5десатурази або -4-десатурази. Ці гени, переважно, можуть бути виділені з мікроорганізмів або нижчих рослин, які виробляють LCPUFAs та вводять їх у мембрани або тригліцериди. Таким чином вже можуть виділятися гени -6-десатурази з моху Physcomitrella patens та гени -6-елонгази із P. patens та з нематоди С. elegans. Перші трансгенні рослини, в які було введено та експресовано ген ферменту біосинтезу LCPUFA та одержано LCPUFAs, вперше були описані, наприклад, в DE 10219203 (Verfahren zur Herstellung mehrfach ungesättigter Fettsäuren in Pflanzen - Спосіб одержання поліненасичених жирних кислот в рослинах). Однак, для того щоб ці рослини масово продукували LCPUFAs, вони ще мають бути оптимізовані для подальшого постпроцесингу в олійновмісних рослинах. Потрібно було створити простий, мало витратний метод одержання цих поліненасичених жирних кислот в еукарютичних системах, що зробило б можливим збагачення харчових продуктів та кормів цими поліненасиченими жирними кислотами. Тому постала задача надати гени, та відповідно ферменти, придатні для синтезу LCPUFAs, для одержання поліненасичених жирних кислот, тобто спеціальні гени, які проявляють -5-елонгазну, 5-десатуразну, -4-десатуразну, -12-десатуразну або -6-десатуразну активність. Наступним завданням даного винаходу було одержання генів й відповідно ферментів, які роблять можливим перехід від -6-жирних кислот до -3-жирних кислот. Наступним завданням було розробити спосіб одержання поліненасичених жирних кислот в організмі, зокрема в еукарютичному організмі, переважно в рослині або мікроорганізмі. Це завдання, згідно даного винаходу, було розв'язане способом одержання сполуки загальної формули І 90455 12 в трансгенних організмах із вмістом принаймні 1мас.% даної сполуки від загального вмісту ліпідів трансгенного організму, який відрізняється тим, що включає наступні етапи: a) введення в організм, принаймні однієї послідовності нуклеїнової кислоти, яка кодує -9елонгазну та/або -6-десатуразну активність, та b) введення в організм, принаймні однієї послідовності нуклеїнової кислоти яка кодує -8десатуразну або -6-елонгазну активність, та c) введення в організм, принаймні однієї послідовності нуклеїнової кислоти, яка кодує -5десатуразну активність, та d) введення в організм, принаймні однієї послідовності нуклеїнової кислоти, яка кодує -5елонгазну активність, та e) введення в організм, принаймні однієї послідовності нуклеїнової кислоти, яка кодує -4десатуразну активність, та де перемінні та замісники в формулі І мають наступні значення: 1 R =гідроксил-, коензимА-(тіоестер)-, лізофосфатидилхолін-, лізофосфатидилетаноламін-, лізофосфатидилгліцерол-, лізодифосфатидилгліцерол-, лізофосфатидилсерин-, лізофосфатидилінозитол-, сфінгооснова- або залишок загальної формули II 2 R =водень-, лізофосфатидилхолін-, лізофосфатидилетаноламін-, лізофосфатидилгліцерол-, лізодифосфатидилгліцерол-, лізофосфатидилсерин-, лізофосфатидилінозитол- або насичені або ненасичені С2-С24-алкiлкарбонiл-, 3 R =водень-, насичені або ненасичені С2-С242 3 алкілкарбоніл-, або R або R незалежно один від одного залишки загальної формули 1а: n=, 3, 4, 5, 6, 7 або 9, m=2, 3, 4, 5 або 6 та p=0 або 3. 1 R =в загальній формулі І означає гідроксил-, коензимА-тіоестер-, лізофосфатидилхолін-, лізофосфатидилетаноламін-, лізофосфатидилгліцерол-, лізодифосфатидилгліцерол-, лізофосфатидилсерин-, лізофосфатидилінозитол-, сфінгооснова-, або залишок загальної формули II. 13 90455 1 Вище названі залишки R завжди приєднані до сполуки загальної формули І у формі тіоестеру. 2 R =в загальній формулі II означає, водень, лізофосфатидилхолін-, лізофосфатидилетаноламін, лізофосфатидилгліцерол-, лізодифосфатидилгліцерол-, лізофосфатидилсерин-, лізофосфатидилінозитол- або насичений або ненасичений С2-С24алкілкарбоніл-, Алкільними залишками можуть бути заміщені або незаміщені насичені або ненасичені С2-С24алкілкарбонільні ланцюги, такі як етилкарбоніл-, нпропілкарбоніл-, н-бутилкарбоніл-, нпентилкарбоніл-, н-гексилкарбоніл-, нгептилкарбоніл-, н-октилкарбоніл-, ннонілкарбоніл-, н-децилкарбоніл-, нундецилкарбоніл-, н-додецилкарбоніл-, нтридецилкарбоніл-, н-тетрадецилкарбоніл-, нпентадецилкарбоніл-, н-гексадецилкарбоніл-, нгептадецилкарбоніл-, н-октадецилкарбоніл-, ннонадецилкарбоніл-, н-ейкозилкарбоніл-, ндокозанілкарбоніл- або н-тетракозанілкарбоніл-, які мають один або більше подвійних зв'язків. Особливо переважними є насичені або ненасичені С10-С22-алкілкарбонільні залишки, такі як ндецилкарбоніл-, н-ундецилкарбоніл-, ндодецилкарбоніл-, н-тридецилкарбоніл-, нтетрадецилкарбоніл-, н-пентадецилкарбоніл-, нгексадецилкарбоніл-, н-гептадецилкарбоніл-, ноктадецилкарбоніл-, н-нонадецилкарбоніл-, нейкозилкарбоніл- н-докозанілкарбоніл- або нтетракозанілкарбоніл-, які мають один або більше подвійних зв'язків. Особливо переважними є насичені або ненасичені С10-С22-алкілкарбонільні залишки, такі як С10-алюлкарбоніл-, С11алкілкарбоніл-, C12алкілкарбоніл-, С13алкілкарбоніл-, С14-алкілкарбоніл-, С16алкілкарбоніл-, С18-алкілкарбоніл-, С20алкілкарбоніл- або С22-алкілкарбонільний залишок, які мають один або більше подвійних зв'язків. Найбільш переважними є насичені або ненасичені С16-С22-алкілкарбонільні залишки, такі як С16алкілкарбоніл-, С18-алкілкарбоніл-, С20алкілкарбоніл- або С22-алкілкарбонільний залишок, які мають один або більше подвійних зв'язків. Ці переважні залишки можуть містити два, три, чотири, п'ять або шість подвійних зв'язків. Особливо переважні залишки із 20 або 22 атомами вуглецю у жирнокислотному ланцюзі містять до шести подвійних зв'язків, переважно три, чотири, п'ять або шість подвійних зв'язків, більш переважно п'ять або шість подвійних зв'язків. Усі зазначені залишки походять від відповідних жирних кислот. 3 R в загальній формулі II означає водень, насичений або ненасичении С2-С24-алкілкарбоніл. Алкільними залишками можуть бути заміщені або незаміщені насичені або ненасичені С2-С24алкілкарбонільні ланцюги, такі як етилкарбоніл нпропілкарбоніл-, н-бутилкарбоніл-, нпентилкарбоніл-, н-гексилкарбоніл-, н 14 гептилкарбоніл-, н-октилкарбоніл-, ннонілкарбоніл-, н-децилкарбоніл-, нундецилкарбоніл-, н-додецилкарбоніл-, нтридецилкарбоніл-, н-тетрадецилкарбоніл-, нпентадецилкарбоніл-, н-гексадецилкарбоніл-, нгептадецилкарбоніл-, н-октадецилкарбоніл-, ннонадецилкарбоніл-, н-ейкозилкарбоніл-, ндокозанілкарбоніл- або н-тетракозанілкарбоніл-, які мають один або більше подвійних зв'язків. Переважними є насичені або ненасичені С10-С22алкілкарбонільні залишки, такі як н-децилкарбоніл-, н-ундецилкарбоніл-, н-додецилкарбоніл-, нтридецилкарбоніл-, н-тетрадецилкарбоніл-, нпентадецилкарбоніл-, н-гексадецилкарбоніл-, нгептадецилкарбоніл-, н-октадецилкарбоніл-, ннонадецилкарбоніл-, н-ейкозилкарбоніл-, ндокозанілкарбоніл- або н-тетракозанілкарбоніл-, які мають один або більше подвійних зв'язків. Найбільш переважними є насичені або ненасичені С10-С22-алкілкарбонільні залишки, таю як С10алкілкарбоніл-, С11-алкілкарбоніл-, С12алкілкарбоніл-, С13-алкілкарбоніл-, С14алкілкарбоніл-, С16-алкілкарбоніл-, С18алкілкарбоніл-, С20-алкілкарбонілабо С22алкілкарбонільний залишок, які мають один або більше подвійних зв'язків. Ще найбільш переважними є насичені або ненасичені С16-С22алкілкарбонільні залишки, такі як С16алкілкарбоніл-, С18-алкілкарбоніл-, С20алкілкарбоніл- або С22-алкілкарбонільний залишок, які мають один або більше подвійних зв'язків. Ці переважні залишки можуть містити два, три, чотири, п'ять або шість подвійних зв'язків. Особливо переважні залишки із 20 або 22 атомами вуглецю у жирнокислотному ланцюгу містять до шести подвійних зв'язків, переважно три, чотири, п'ять або шість подвійних зв'язків, більш переважно п’ять або шість подвійних зв'язків. Усі зазначені залишки походять від відповідних жирних кислот. 1 2 3 Вище названі залишки R , R та R можуть бути заміщеними гідроксил та/або епоксигрупами, та/або можуть мати потрійні зв'язки. Переважні, одержані згідно даного винаходу жирні кислоти можуть містити мінімум два, переважно, три, чотири, п'ять або шість подвійних зв'язків. Особливо переважні жирні кислоти містять чотири, п'ять або шість подвійних зв'язків. Одержані в способі жирні кислоти переважно мають 18-, 20- або 22-С-атомів в жирнокислотному ланцюзі, переважно, жирні кислоти мають 20- або 22-Сатомів в жирнокислотному ланцюзі. Переважно насичені жирні кислоти перетворюються використовуваними в способі нуклеїновими кислотами в незначній мірі або зовсім не перетворюються. Під "в незначній мірі" слід розуміти що порівняно із поліненасиченими жирними кислотами насичені кислоти перетворюються з активністю менше 5%, переважно менше 3%, особливо переважно менше 2%, найбільш особливо переважно менше 1; 0,5; 0,25 або 0,125%. Ці зазначені жирні кислоти можуть бути одержані способом як єдиний продукт або як суміш жирних кислот. При використанні у способі згідно даного винаходу послідовності нуклеїнових кислот мається на увазі використання послідовності нуклеїнових 15 кислот, що кодують поліпептиди із -9-елогазною, -6-десатуразною, -8-десатуразною, -6елогазною, -5-десатуразною, -5-елогазною та/або -4-десатуразною активністю. Переважними, у способі згідно даного винаходу, будуть послідовності нуклеїнових кислот, які кодують поліпептиди із -9-елогазною, -6десатуразною, -8-десатуразною, -6-елогазною, -5-десатуразною, -5-елогазною та/або -4десатуразною активністю, вибрані із групи, яка складається із: а) послідовності нуклеїнової кислоти з наведених далі послідовностей: SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5, SEQ ID NO:7, SEQ ID NO:9, SEQ ID ΝO:11, SEQ ID NO:13, SEQ ID NO:15, SEQ ID NO:17, SEQ ID NO:19, SEQ ID NO:21, SEQ ID NO:23, SEQ ID NO:25, SEQ ID NO:27, SEQ ID NO:29, SEQ ID NO:31, SEQ ID NO:33, SEQ ID NO:35, SEQ ID NO:37, SEQ ID NO:39, SEQ ID NO:41 SEQ ID NO:43, SEQ ID NO:45, SEQ ID NO:47, SEQ ID NO:49, SEQ ID NO:51, SEQ ID NO:53, SEQ ID NO:59, SEQ ID NO:61, SEQ ID NO:63, SEQ ID NO:65, SEQ ID NO:67, SEQ ID NO:69, SEQ ID NO:71, SEQ ID NO:73, SEQ ID NO:75, SEQ ID NO:77, SEQ ID NO:79, SEQ ID NO:81, SEQ ID NO:83, SEQ ID NO:85, SEQ ID NO:89, SEQ ID NO:91, SEQ ID NO:93, SEQ ID NO:95, SEQ ID NO:97, SEQ ID NO:99, SEQ ID NO:101, SEQ ID NO:103, SEQ ID NO:111, SEQ ID NO:113, SEQ ID NO:117, SEQ ID NO:119, SEQ ID NO:131, SEQ ID NO:133, SEQ ID NO:135, SEQ ID NO:137 або SEQ ID NO:183 або b) послідовності нуклеїнових кислот, які внаслідок виродженості генетичного коду являють собою наведені далі амінокислотні послідовності. SEQ ID NO:2, SEQ ID NO:4, SEQ ID NO:6, SEQ ID NO:8, SEQ ID NO:10, SEQ ID NO:12, SEQ ID NO:14, SEQ ID NO:16, SEQ ID NO:18, SEQ ID NO:20, SEQ ID NO:22, SEQ ID NO:24, SEQ ID NO:26, SEQ ID NO:28, SEQ ID NO:30, SEQ ID NO:32, SEQ ID NO:34, SEQ ID NO:36, SEQ ID NO:38, SEQ ID NO:40, SEQ ID NO:42, SEQ ID NO:44, SEQ ID NO:46, SEQ ID NO:48, SEQ ID NO:50, SEQ ID NO:52, SEQ ID NO:54, SEQ ID NO:60, SEQ ID NO:62, SEQ ID NO:64, SEQ ID NO:66, SEQ ID NO:68, SEQ ID NO:70, SEQ ID NO:72, SEQ ID NO:74, SEQ ID NO:76, SEQ ID NO:78, SEQ ID NO:80, SEQ ID NO:82, SEQ ID NO:84, SEQ ID NO:86, SEQ ID NO:88, SEQ ID NO:92, SEQ ID NO:94, SEQ ID NO:96, SEQ ID NO:98, SEQ ID NO:100, SEQ ID NO:102, SEQ ID NO:104, SEQ ID NO:112, SEQ ID NO:114, SEQ ID NO:118, SEQ ID NO:120, SEQ ID NO:132, SEQ ID NO:134, SEQ ID NO:136, SEQ ID NO:138 або SEQ ID NO:184 або с) похідних від наведених далі послідовностей нуклеїнових кислот SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5, SEQ ID NO:7, SEQ ID NO:9, SEQ ID NO:11, SEQ ID NO:13, SEQ ID NO:15, SEQ ID NO:17, SEQ ID NO:19, SEQ ID NO:21, SEQ ID NO:23, SEQ ID NO:25, SEQ ID NO:27, SEQ ID NO:29, SEQ ID NO:31, SEQ ID NO:33, SEQ ID NO:35, SEQ ID NO:37, SEQ ID NO:39, SEQ ID NO:41, SEQ ID NO:43, SEQ ID NO:45, SEQ ID NO:47, SEQ ID NO:49, SEQ ID NO:51, SEQ ID 90455 16 NO:53, SEQ ID NO:59, SEQ ID NO:61, SEQ ID NO:63, SEQ ID NO:65, SEQ ID NO:67, SEQ ID NO:69, SEQ ID NO:71, SEQ ID NO:73, SEQ ID NO:75, SEQ ID NO:77, SEQ ID NO:79, SEQ ID NO:81, SEQ ID NO:83, SEQ ID NO:85, SEQ ID NO:89, SEQ ID NO:91, SEQ ID NO:93, SEQ ID NO:95, SEQ ID NO:97, SEQ ID NO:99, SEQ ID NO:101, SEQ ID NO:103, SEQ ID NO:111, SEQ ID NO:113, SEQ ID NO:117, SEQ ID NO:119, SEQ ID NO:131, SEQ ID NO:133, SEQ ID NO:135, SEQ ID NO:137 або SEQ ID NO:183 поліпептидів, які мають принаймні 40% подібності до амінокислотних залишків послідовностей, кодованих SEQ ID NO:2, SEQ ID NO:4, SEQ ID NO:6, SEQ ID NO:8, SEQ ID NO:10, SEQ ID NO:12, SEQ ID NO:14, SEQ ID NO:16, SEQ ID NO:18, SEQ ID NO:20, SEQ ID NO:22, SEQ ID NO:24, SEQ ID NO:26, SEQ ID NO:28, SEQ ID NO:30, SEQ ID NO:32, SEQ ID NO:34, SEQ ID NO:36, SEQ ID NO:38, SEQ ID NO:40, SEQ ID NO:42, SEQ ID NO:44, SEQ ID NO:46, SEQ ID NO:48, SEQ ID NO:50, SEQ ID NO:52, SEQ ID NO:54, SEQ ID NO:60, SEQ ID NO:62, SEQ ID NO:64, SEQ ID NO:66, SEQ ID NO:68, SEQ ID NO:70, SEQ ID NO:72, SEQ ID NO:74, SEQ ID NO:76, SEQ ID NO:78, SEQ ID NO:80, SEQ ID NO:82, SEQ ID NO:84, SEQ ID NO:86, SEQ ID NO:88, SEQ ID NO:92, SEQ ID NO:94, SEQ ID NO:96, SEQ ID NO:98, SEQ ID NO:100, SEQ ID NO:102, SEQ ID NO:104, SEQ ID NO:112, SEQ ID NO:114, SEQ ID NO:118, SEQ ID NO:120, SEQ ID NO:132, SEQ ID NO:134, SEQ ID NO:136, SEQ ID NO:138 або SEQ ID NO:184 та проявляють -9-елонгазну, -6-десатуразну, -8десатуразну, -6-елонгазну, -5-десатуразну, -5елонгазну або -4-десатуразну активності. 2 3 Переважно, замісники R або R в загальних формулах І та II означають незалежно один від одного насичені або ненасичені С18-С22алкілкарбоніл-, особливо переважно вони означають незалежно один від одного ненасичені С18-, С20- або С22-алкілкарбоніл- із, принаймні двома подвійними зв'язками. Ще одне переважне виконання способу, відрізняється тим. що в організм введено послідовність нуклеїнової кислоти, що кодує поліпептид з -3десатуразною активністю, вибрану з групи: a) послідовність нуклеїнової кислоти з SEQ ID NO:87 або SEQ ID NO:105, представленої послідовності, або b) послідовності нуклеїнових кислот, які внаслідок виродженості генетичного коду, являють собою наведені далі амінокислотні послідовності SEQ ID NO:88 або SEQ ID NO:106, або c) похідні від наведених послідовностей нуклеїнової кислоти SEQ ID NO:87 або SEQ ID NO:105, які кодують поліпептиди i мають, як мінімум 60% подібності до амінокислотних послідовностей кодованих SEQ ID NO:88 або SEQ ID NO:106 та мають -3-десатуразну активність. Ще одне наступне здійснення способу відрізняється тим, що в організм введено послідовність нуклеїнової кислоти, що кодує поліпептид з -12десатуразною активністю, вибрану з групи: 17 a) послідовності нуклеїнової кислоти з SEQ ID NO:107 або SEQ ID NO:109, представленої послідовності, або b) послідовності нуклеїнової кислоти, які можуть бути виведені як результат виродженості генетичного коду із наведених амінокислотних послідовностей SEQ ID NO:108 або SEQ ID NO:110, або c) похідні представленої в SEQ ID NO:107 або SEQ ID NO:109 послідовності нуклеїнової кислоти, що кодують поліпептиди які щонайменше на 60% подібні амінокислотним ділянкам з SEQ ID NO:108 90455 18 або SEQ ID NO:110 i проявляють -12-десатуразну активність. Ці вище згадані послідовності -12-десатурази можуть використовуватися окремо або в комбінації із послідовностями -3-десатурази використаних у способі послідовностей нуклеїнової кислоти, що кодують -9-елонгазу, -6-десатуразу, -8десатуразу, -6-елонгазу, -5-десатуразу, -5елонгазу та/або -4-десатуразу. В таблиці 1 наведено послідовності нуклеїнових кислот, організм походження та номер послідовності: 19 Поліненасичені жирні кислоти, одержані у способі, переважно зв'язані з мембранними ліпідами та/або триацилгліцеридами, можуть зустрічатися в організмі як вільні жирні кислоти або бути зв'язаними у формі естерів жирної кислотию До того ж, вони можуть існувати у вигляді "чистих продуктів" або переважно в суміші різних жирних кислот або в суміші різних гліцеридів. Зв'язаними у триацилгліцеридах стають різні жирні кислоти, при цьому вони є коротколанцюговими жирними кислотами з 4-6 С-атомами, середньоланцюговими жирними кислотами з 8-12 С-атомами або довголанцюговими жирними кислотами з 14-24 С-атомами, переважно довголанцюговими жирними кислотами, особливо переважно довголанцюговими жирними кислотами LCPUFAs з С18-, С20- та/або С22жирними кислотами. У способі згідно даного винаходу одержуються, переважно, естери жирних кислот із поліненасиченими С18-, С20- та/або С22-молекулами жирних кислот із, принаймні, двома подвійними зв'язками в естері жирної кислоти, переважно із, принаймні, трьома, чотирма п'ятьма, або шістьма подвійними зв'язками в естері жирної кислоти, найбільш переважно із, принаймні, п'ятьма, або шістьма подвійними зв'язками в естері жирної кислоти та прово9,12 дять синтез лінолевої кислоти (=LA, С18:2 ), 6,9,12 ліноленовоі кислоти (=GLA, С18:3 ), стеаридо6,9,12,15 нової кислоти (=SDA, C18:4 ), дигомо-8,11,14 ліноленової кислоти (=DGLA, 20:3 ), -35,8,11,14 ейкозатетраєновоі кислоти (=ЕТА, С20:4 ), 5,8,11,14 арахідонової кислоти (ARA, С20:4 ), ейкоза5,8,11,14,17 пентаєнової кислоти (ЕРА, С20:5 ), -64,7,10,13,16 докозапентаєновоі кислоти (С22:5 ), -6,7,10,13,16 докозатетраєновоі кислоти (С22:4 ), -3докозапентаєновоі кислоти (=DPA, 7,10,13,16,19 С22:5 ), докозагексаєнової кислоти 4,7,10,13,16,19 (=DHA, С22.6 ) або їхніх сумішей, переважно ARA, ЕРА та/або DHA Найбільш переважно одержуються -3-жирні кислоти, такі як ЕРА та/або DHA. Естери жирних кислот із поліненасиченими молекулами С18-, С20- та/або С22-жирних кислот можуть виділятися із організмів для одержання жирних кислот у формі олій або ліпідів, наприклад, у формі таких сполук як сфінголіпід, фосфогліце 90455 20 рид, ліпід, гліколіпід, такий як глікофосфоліпід, фосфоліпід, такий як фосфатидилетаноламін, фосфатидилхолін, фосфатидилсерин, фосфатидилгліцерол, фосфатидилінозитол або дифосфатидилгліцерол, моноацилгліцерид, діацилгліцерид, триацилгліцерид або наступний естер жирної кислоти, такий як ацетил-СоА-естер, поліненасичені жирні кислоти, що містять, принаймні, два, три, чотири, п'ять або шість, переважно містять, принаймні, п'ять, або шість подвійних зв'язків, виділені, переважно, у формі своїх діацилгліцеридів, триацилгліцеридів та/або у формі фосфатидилхоліну, зокрема переважно у формі триацилгліцериду. Поряд із цими естерами поліненасичені жирні кислоти також містяться в організмах, переважно рослинах, у формі вільних жирних кислот або зв'язаних в інших сполуках. Як правило, різні зазначені вище сполуки (естери жирних кислот або вільні жирні кислоти) містяться в організмах в наступному приблизному співвідношенні від 80 до 90ваг.% тригліцериди, від 2 до 5ваг.% дигліцериди, від 5 до 10ваг.% моногліцериди, від 1 до 5ваг.% вільні жирні кислоти, від 2 до 8ваг.% фосфоліпіди, при чому загальний вміст різних сполук становить 100ваг.%. У способі згідно даного винаходу одержані LCPUFAs становлять від загального вмісту, щонайменше, 3ваг.%, бажано, щонайменше, 5ваг.%, переважно, щонайменше, 8 ваг.%, особливо переважно, щонайменше, 10 ваг.%, найбільш особливо переважно, щонайменше, 15ваг.% від усіх жирних кислот, що містяться в трансгенних організмах, переважно, в трансгенних рослинах. При цьому переважно С18- та/або С20-жирні кислоти, які присутні в організмах хазяїнах принаймні на 10%, переважно, принаймні на 20%, особливо переважно, принаймні на 30%, найбільш особливо переважно, принаймні на 40%, містяться у вказаних продуктах, таких як DPA або DHA. Переважно, жирні кислоти знаходяться у зв'язаній формі. За допомогою використаних у способі згідно даного винаходу нуклеїнових кислот можна конвертувати вказані ненасичені жирні кислоти у sn1-,sn2- та/або sn3положення переважного одержаного три гліцериду. Також, в способі згідно даного винаходу з вихідних сполук лінолевої кислоти (С18:2) або ліноленової кислоти (С18:3) у декілька етапів 21 утворюються кінцеві продукти способу, такі як, наприклад, арахідонова кислота (ARA), ейкозапентаєнова кислота (ЕРА), -6-докозапентаєнова кислота або DHA, не у вигляді абсолютно чистого продукту, а із вмістом у кінцевому продукті домішок з попередніх етапів. Оскільки у вихідному організмі або у вихідній рослині знаходяться як лінолева кислота, так i ліноленова кислота, то i в кінцевому знаходиться суміш ARA, ЕРА або DHA. Вміст сполук з попередніх етапів бажано становить не більше ніж 20ваг.%, переважно не більше ніж 15ваг.%, особливо переважно не більше ніж 10ваг.%, найбільш особливо переважно не більше ніж 5ваг% від маси відповідного кінцевого продукту. Переважно в способі згідно даного винаходу в трансгенній рослині як кінцеві продукти виробляються лише ARA, ЕРА або лише DHA, зв'язані або як вільні кислоти Якщо одночасно виробляються сполуки ARA, ЕРА та DHA то вони виробляються в наступній пропорції, як мінімум 1:1:2 (EPA:ARA:DHA), бажано як мінімум 1:1:3, переважно 1:1:4, особливо переважно 1:1:5. Способом, запропонованим згідно з винаходом, одержують складні ефіри жирних кислот або суміш жирних кислот, що містять переважно від 6 до 15% пальмітинової кислоти, від 1 до 6% стеаринової кислоти; 7-85% олеїнової кислоти, від 0,5 до 8% вакценової кислоти, від 0,1 до 1% арахінової кислоти, від 7 до 25% насичених жирних кислот, від 8 до 85% мононенасичених жирних кислот та від 60 до 85% поліненасичених жирних кислот, відповідно, відносно 100% та загального вмісту жирних кислот організмів. Бажано, щоб частка поліненасичених жирних кислот в складних естерах жирних кислот або сумішах жирних кислот становила, переважно, принаймні 0,1; 0,2; 0,3; 0,4; 0,5; 0,6; 0,7; 0,8; 0,9 або 1% відносно загального вмісту жирних кислот, що приходиться на арахідонову кислоту. Також, складні естери жирних кислот або суміші жирних кислот, одержані способом згідно даного винаходу, переважно, жирні кислоти вибрані з групи жирних кислот: ерукова кислота (13-докозаєнова кислота), стеркулінова кислота (9,10-метилен октадек-9-енова кислота), малвалінова кислота (8,9-метилен-гептадек-8-енова кислота), хаульмугрова кислота, (циклопентдодеканова кислота), фуран-жирна кислота (9,12епоксиоктадека-9,11-дієнова кислота), вернова кислота (9,10-епоксиоктадек-12-енова кислота), таринова кислота (6-октадецинова кислота), 6нонадецинова кислота, санталбінова кислота (t11октадецен-9-інова кислота), 6,9-октадеценінова кислота, пірулінова кислота (t10-гептадецен-8інова кислота), крепенінова кислота (9-октадецен12-інова кислота), 13,14-дигідроорофенінова кислота октадецен-13-ен-9,11-діїнова кислота, петроселенова кислота (цис-6-октадеценова кислота), 9с,12t-октадекадієнова кислота, календулова (8t10t12c-октадекатрієнова кислота), катальпінова кислота (9t11t3с-октадекатрієнова кислота), елеостеринова кислота (9c11t13t-октадекатрієнова кислота), джакаринова кислота (8с10t12соктадекатрієнова кислота), пуніцинова кислота (9с11t13с-октадекатрієнова кислота) парінарінова кислота (9с11t13t15с-октадекатетраєнова кисло 90455 22 та), піноленова кислота (всі-цис-5,9,12октадекатрієнова кислота), лабаленова кислота (5,6-октадекадієналенова кислота), рицинолова кислота (12-гідроксимасляна кислота) та/або коріолінова кислота (13-гідрокси-9с,11t-октадієнонова кислота). Вище згадані жирні кислоти знаходяться у складних естерах жирних кислот або сумішах жирних кислот одержаних способом, згідно даного винаходу, як правило лише в слідових кількостях, це означає, що їхній вміст становить, по відношенню до загального вмісту жирних кислот, менше ніж 30%, переважно менше ніж 25%, 24%, 23%, 22% або 21%, особливо переважно менше ніж 20%, 15%, 10%, 9%, 8%, 7%, 6% або 5%, в найвищій мірі переважно менш ніж 4%, 3%, 2% або 1%. В наступній переважній формі здійснення винаходу вміст вище згаданих жирних кислот по відношенню до загального вмісту жирних кислот становить менше ніж 0,9%; 0,8%; 0,7%; 0,6%; або 0,5%, особливо переважно менше ніж 0,4%; 0,3%; 0,2%; 0, 1%. Переважно естери жирних кислот або суміші жирних кислот, одержані за способом згідно даного винаходу, містять менше ніж 0,1%, по відношенню до загального вмісту жирних кислот, та/або не містять масляної кислоти, холестерину, клупанодонової кислоти (=докозапентаєнова кислота, 4,8,12,15,21 С22:5 ), а також нізинової кислоти (тетра3,8,12,15,18,21 козагексаєнова кислота, С23:6 ). Послідовності нуклеїнової кислоти згідно даного винаходу або послідовності нуклеїнової кислоти, одержані способом згідно даного винаходу, можуть сприяти підвищенню виходу поліненасичених жирних кислот як мінімум на 50%, переважно як мінімум на 80%, особливо переважно як мінімум на 100%, в найвищій мірі переважно як мінімум на 150% порівняно із нетрансгенним вихідним організмом, наприклад, дріжджами, водоростями, грибами або рослинами, такими як Arabidopsis або льон при порівнянні GC-аналізом, дивись приклади. Також, хімічно чисті поліненасичені жирні кислоти або композиції жирних кислот можуть отримуватися описаним вище способом. Для цього жирні кислоти або композиції жирних кислот можуть виділятися з організму, такого як мікроорганізми або рослини, або з культурального середовища, в якому або на якому організми ростуть, або з організмів та культурального середовища відомими способами, такими як, екстракцією, дистиляцією кристалізацією, хроматографією або комбінаціями цих методів. Ці хімічно чисті жирні кислоти або композиції жирних кислот є переважно придатними для харчової промисловості, косметичної промисловості та, особливо, фармацевтичної промисловості. Як організм для одержання способом згідно даного винаходу принципово беруться до уваги будь-які організми, такі як мікроорганізми, тварини, за виключенням людей, або рослини. Як рослини, в принципі, беруться до уваги будь-які рослини, які можуть синтезувати жирні кислоти, як-то усі дводольні або однодольні рослини, водорості або мохи. Переважними є рослини, вибрані з групи сімейств рослин Adelotheciaceae, Anacardiaceae, Asteraceae, 23 Apiaceae, Betulaceae, Boraginaceae, Brassicaceae, Bromeliaceae, Caricaceae, Cannabaceae, Convolvulaceae, Chenopodiaceae Crypthecodiniaceae, Cucurbitaceae, Ditnchaceae, Elaeagnaceae, Ericaceae, Euphorbiaceae, Fabaceae, Geraniaceae, Gramineae, Juglandaceae, Lauraceae, Leguminosae, Linaceae, Euglenaceae, Prasinophyceae, або декоративні рослини, такі як чорнобривці. Для прикладу можуть бути наведені рослини вибрані з наступної групи: Adelotheciaceae, такі як роди Physcomitrella, наприклад, рід та вид Physcomitrella patens, Anacardiaceae, такі як роди Pistacia, Mangifera, Anacardium, наприклад, роди та види Pistacia vera [фісташка], Mangiferindica [манго] або Anacardium occidentale [кешью], Asteraceae такі як роди Calendula, Carthamus, Centaurea, Cichorium, Cynara, Helianthus, Lactuca, Locusta, Tagetes, Valeriana, наприклад, рід та вид Calendula officinalis [садова календула], Carthamus tinctorius [сафлор красильний], Centaurea cyanus [волошки], Cichorium intybus [цикорій], Cynara scolymus [артишок], Helianthus annuus [соняшник], Lactuca sativa, Lactuca crispa, Lactuca esculenta, Lactuca scanola L. ssp. sativa, Lactuca scanola L. var. integrata, Lactuca scariola L. var. integnfolia, Lactuca sativa subsp. romana, Locusta communis, Valenana locusta [салат], Tagetes lucida, Tagetes erecta або Tagetes tenuifolia [чорнобривці], Аріасеае такі як рід Daucus, наприклад, рід та вид Daucus carota [морква], Betulaceae, такі як рід Corylus, наприклад, роди та види Corylus avellana або Corylus colurna [ліщина], Boraginaceae, такі як рід Воrаgо, наприклад, рід та вид Borago officinalis [огуречник лікарський], Brassicaceae як роди Brassica, Camelina, Melanosinapis, Sinapis, Arabadopsis, наприклад, роди та види Brassica napus, Brassica rapa ssp. [рапс], Smapis arvensis, Brassica juncea, Brassica juncea var. juncea, Brassica juncea var. crispifolia, Brassica juncea var. foliosa, Brassica nigra, Brassica sinapioides, Camelina sativa, Melanosinapis communis [гірчиця], Brassica oleracea [кормовий буряк] або Arabidopsis thaliana, Bromeliaceae як рід Anana, Bromelia (ананас) наприклад роди та види Anana comosus, Ananas ananas або Bromelia comosa [ананас], Cancaceae як рід Carica та як рід та вид Саriса papaya [папайя], Cannabaceae як рід Cannabis та як рід та вид Cannabis sative [конопля], Convolvulaceae як роди Іроmеа, Convolvulus, наприклад, роди та види Іроmеа batatus, Ipomoea pandurata, Convolvulus batatas, Convolvulus tiliaceus, Ipomoea fastigiata, Ipomoea tiliacea, Ipomoea trioba або Convolvulus panduratus [солодка картопля, батат], Chenopodiaceae як рід Beta, наприклад, роди та види Beta vulgaris, Beta vulgaris var. altissima, Beta vulgäris var. Vulgaris, Beta maritima, Beta vulgaris var. perennis, Beta vulgaris var. conditiva або Beta vulgaris var. esculenta [цукровий буряк], Crypthecodiniaceae як рід Crypthecodinium, наприклад, рід та вид Crypthecodinium cohnii, Cucurbitaceae як рід Cucurbita, наприклад, роди та види Cucurbita maxima, Cucurbita mixta, Cucurbita pepo або Cucurbita moschata [гарбуз], Cymbellaceae як роди Amphora, Cymbella, 90455 24 Okedenia, Phaeodáctylum, Reimeria наприклад рід та вид Phaeodactylum tricornutum, Ditrichaceae як роди Ditrichaceae, Astomiopsis, Ceratodon, Chrysoblastella, Ditrichum, Distichium, Eccremidium, Lophidion, Philibertiella, Pleuridium, Saelania, Trichodon, Skottsbergia, наприклад, роди та види Ceratodon antarcticus, Ceratodon columbiae, Ceratodon heterophyllus, Ceratodon purpurascens, Ceratodon purpureus, Ceratodon purpureus ssp. convolutus, Ceratodon purpureus ssp. stenocarpus, Ceratodon purpureus var. rotundifolius, Ceratodon ratodon, Ceratodon stenocarpus, Chrysoblastella chilensis, Ditrichum ambiguum, Ditrichum brevisetum, Ditrichum crispatissimum, Ditrichum difficile, Ditrichum falcifolium, Ditrichum flexicaule, Ditrichum giganteum, Ditrichum heteromallum, Ditrichum lineare, Ditrichum lineare, Ditrichum montanum, Ditrichum montanum, Ditrichum pallidum, Ditrichum punctulatum, Ditrichum pusillum, Ditrichum pusillum var. tortile, Ditrichum rhynchostegium, Ditrichum schimperi, Ditrichum tortile, Distichium capillaceum, Distichium hagenii, Distichium inclinatum, Distichium macounii, Eccremidium floridanum, Eccremidium whiteleggei, Lophidion strictus, Pleuridium acuminatum, Pleuridium altemifolium, Pleuridium holdridgei, Pleuridium mexicanum, Pleuridium ravenelii, Pleuridium subulatum, Saelania glaucescens, Trichodon borealis, Trichodon cylindricus або Trichodon cylindricus var. oblongus, Elaeagnaceae як рід Elaeagnus, наприклад рід та вид ОІеа еurораеа [оливка], Ericaceae як рід Kаlmіа, наприклад, роди та види Kalmia latifolia, Kalmia angustifolia, Каlmіа microphylla, Kаlmіа polifolia, Kаlmіа occidentalis, Cistus chamaerhodendros або Kаlmіа lucida [гірський лавр], Еuglеnасеае як роди Ascoglena, Astasia, Colacium, Cyclidiopsis, Euglena, Euglenopsis, Hyalaphacus, Khawkinea, Lepocinclis, Phacus, Strombomonas, Trachelomonas, наприклад, рід та вид Euglena gracilis; Euphorbiaceae як роди Manihot, Janipha, Jatropha, Ricinus, наприклад, роди та види Manihot utilissima, Janipha manihot., Jatropha manihot., Manihot aipil, Manihot dulcis, Manihot manihot, Manihot melanobasis, Manihot esculenta [маніока] або Ricinus communis [кліщовина], Fabaceae як роди Pisum, Albizia, Cathormion, Feuillea, Inga, Pithecolobium, Acacia, Mimosa, Medicajo, Glycine, Dolichos, Phaseolus, Soja, наприклад, роди та види Pisum sativum, Pisum arvense, Pisum humile [горох], Albizia berteriana, Albizia julibrissin, Albizia lebbeck, Acacia berteriana, Acacia littoralis, Albizia berteriana, Albizzia berteriana, Cathormion berteriana, Feuillea berteriana, Inga fragrans, Pithecellobium berierianum, Pithecellobium fragrans, Pithecolobium berterianum, Pseudalbizzia berteriana, Acacia julibriissin, Acacia nemu, Albizia nemu, Feuilleea julibrissm, Mimosa julibrissin, Mimosa speciosa, Sericanrda julibrissin, Acacia lebbeck, Acacia macrophylla, Albizia lebbek, Feuilleea lebbeck, Mimosa lebbeck, Mimosa speciosa [шовковиця], Medicago sativa, Medicago falcata, Medicago varia [люцерна] Glycine max Dolichos soja, Glycine gracilis, Glycine hispida, Phaseolus max, Soja hispida або Soja max [соя], Funariaceae як роди Aphanorrhegma, Entosthodon, Funaria, 25 Physcomitrella, Physcomitrium, наприклад, роди та види Aphanorrhegma erratum, Entosthodon attenuatus, Entosthodon bolanderi, Entosthodon bonplandii, Entosthodon californicus, Entosthodon drummondii, Entosthodonjamesonii, Entosthodon leibergii, Entosthodon neoscoticus, Entosthodon rubrisetus, Entosthodon spathulifolius, Entosthodon tucsoni, Funaria americana, Funaria bolanderi, Funaria calcarea, Funaria californica, Funaria calvescens, Funaria convoluta, Funaria flavicans, Funaria groutiana, Funaria hygrometrica, Funaria hygrometrica var. arctica, Funaria hygrometrica var. calvescens, Funaria hygrometrica var. convoluta, Funaria hygrometrica var. muralis, Funaria hygrometrica var. utahensis, Funaria microstoma, Funaria microstoma var. obtusifolia, Funaria muhlenbergii, Funaria orcuttii, Funaria plano-convexa, Funaria polaris, Funaria ravenelii, Funaria rubriseta, Funaria serrata, Funaria sonorae, Funaria sublimbatus, Funaria tucsoni, Physcomitrella californica, Physcomitrella patens, Physcomitrella readeri, Physcomitrium australe, Physcomitrium californicum, Physcomitrium collenchymatum, Physcomitrium coloradense, Physcomitrium cupuliferum, Physcomitrium drummondii, Physcomitrium eurystomum, Physcomitrium flexifolium, Physcomitrium hookeri, Physcomitrium hookeri var. serratum, Physcomitrium immersum, Physcomitrium kellermanii, Physcomitrium megalocarpum, Physcomitrium pyriforme, Physcomitrium pyriforme var. serratum, Physcomitrium rufipes, Physcomitrium sandbergii, Physcomitrium subsphaericum, Physcomitrium washingtoniense, Geraniaceae як роди Pelargonium, Cocos, Oleum, наприклад, роди та види Cocos nucifera, Pelargonium grossulanoides або Oleum cocois [кокосовий горіх], Grammeae як рід Saccharum, наприклад, рід та вид Saccharum officmarum, Juglandaceae як роди Juglans, Wallia, наприклад, роди та види Juglans regia, Juglans atlanthifolia, Juglans sieboldiana, Juglans cineria, Wallia cineria, Juglans bixbyi, Juglans californica, Juglans hindsii, Juglans intermedia, Juglans jamaicensis, Juglans major, Juglans microcarpa, Juglans nigra або Wallia nigra [грецький горіх], Lauraceae як роди Persea, Laurus, наприклад, роди та види Laurus nobilis [лавр], Persea americana, Persea gratissima або Persea persea [авокадо], Leguminosae як рід Arachis, наприклад, рід та вид Arachis hypogaea [арахіс], Linaceae як роди Linum, Adenolinum, наприклад, роди та види Linum usitatissimum, Linum humile, Linum austriacum, Linum bienne, Linum angustifolium, Linum cathaticum, Linum flavum, Linum grandiflorum, Adenolinum grandiflorum, Linum lewisii, Linum narbonense, Linum perenne, Linum perenne var. lewisii, Linum pratense або Linum trigynum [льон], Lythrarieae як рід Punica, наприклад рід та вид Рunіса granatum [гранат], Malvaceae як рід Gossypium, наприклад, роди та види Gossypium hirsutum, Gossypium arboreum, Gossypium barbadense, Gossypium herbaceum або Gossypium thurben [бавовна], Marchantiaceae як рід Marchantia, наприклад, роди та види Marchantia berteroana, Marchantia foliacea, Marchantia 90455 26 macropora, Musaceae як рід Musa, наприклад, роди та види Musa nana, Musa acummata, Musa paradisiaca, Musa spp. [банан], Оnаgrасеае як роди Camissonia, Oenothera, наприклад, роди та види Oenothera biennis або Camissonia brevipes [ослинник], Palmae як рід Elacis, наприклад рід та вид Elacis guineensis [маслична пальма], Papaveraceae як рід Papaver, наприклад, роди та види Papaver orientale, Papaver rhoeas, Papaver dubium [мак], Pedaliaceae як рід Sesamum, наприклад, рід та вид Sesamum indicum [сезам], Piperaceae як роди Piper, Artanthe, Peperomia, Steffensia, наприклад, роди та види Piper aduncum, Piper amalago, Piper angustifolium, Piper auritum, Piper betel, Piper cubeba, Piper longum, Piper nigrum, Piper retrofractum, Artanthe adunca, Artanthe elongata, Peperomia elongata, Piper elongatum, Steffensia elongata. [перець чілі], Роасеае як роди Hordeum, Secale, Avena, Sorghum, Andropogon, Holcus, Panicum, Oryza, Zea (кукурудза), Triticum, наприклад, роди та види Hordeum vulgare, Hordeum jubatum, Hordeum murinum, Hordeum secalinum, Hordeum distichon Hordeum aegiceras, Hordeum hexastichon, Hordeum hexastichum, Hordeum irregulare, Hordeum sativum, Hordeum secalinum [ячмінь], Secale cereale [жито], Avena sativa, Avena fatua, Avena byzantina, Avena fatua var. sativa, Avena hybrida [овес], Sorghum bicolor, Sorghum halepense, Sorghum saccharatum, Sorghum vu/gare, Andropogon drummondii, Holcus bicolor, Holcus sorghum, Sorghum aethiopicum, Sorghum arundinaceum, Sorghum caffrorum, Sorghum cernuum, Sorghum dochna, Sorghum drummondii, Sorghum durra, Sorghum guineense, Sorghum lanceolatum, Sorghum nervosum, Sorghum saccharatum, Sorghum subglabrescens, Sorghum verticilliflorum, Sorghum vulgare, Holcus halepensis, Sorghum miliaceum, Panicum militaceum [просо], Oryza sativa, Oryza latifolia [рис], Zea mays [кукурудза] Triticum aestivum, Triticum durum, Triticum turgidum, Triticum hybemum, Triticum macha, Triticum sativum або Triticum vulgare [пшениця], Porphyridiaceae як роди Chroothece, Flintiella, Petrovanella, Porphyridium, Rhodella, Rhodosorus, Vanhoeffenia, наприклад, рід та вид Porphyridium cruentum, Proteaceae як рід Macadamia, наприклад, рід та вид Macadamia intergnfolia [макадамія], Prasinophyceae як роди Nephroselmis, Prasinococcus, Scherffelia, Tetraselmis, Mantoniella, Ostreococcus, наприклад, роди та види Nephroselmis olivacea, Prasinococcus capsulatus, Scherffelia dubia, Tetraselmis chui, Tetraselmis suecica, Mantoniella squamata, Ostreococcus tauri, Rubiaceae як рід Coffea, наприклад, роди та види Cofea spp., Coffea arabica, Coffea canephora або Coffea liberica [кава], Scrophulariaceae як рід Verbascum, наприклад, роди та види Verbascum blattaria, Verbascum chaixii, Verbascum densiflorum, Verbascum lagurus, Verbascum longifolium, Verbascum lychnitis, Verbascum nigrum, Verbascum olympicum, Verbascum phlomoides, Verbascum phoenicum, Verbascum pulverulentum або Verbascum thapsus [коров'як], Solanaceae як роди Capsicum, Nicotiana, Solanum, Lycopersicon, наприклад, роди та види Capsicum annuum, Capsicum 27 annuum var. glabriusculum, Capsicum frutescens [перець], Capsicum annuum [паприка, червоний солодкий перець], Nicotiana tabacum, Nicotiana alata, Nicotiana attenuate, Nicotiana glauca, Nicotiana langsdorffii, Nicotiana obtusifolia, Nicotiana quadrivalvis, Nicotiana repanda, Nicotiana rustic, Nicotiana sylvestris [тютюн], Solanum tuberosum [картопля], Solanum melongena [баклажан] Lycopersicon esculentum, Lycopersicon lycopersicum., Lycopersicon pyriforme, Solanum integnfolium або Solanum lycopersicum [томат, помідор], Sterculiaceae як рід Theobroma наприклад, рід та вид Theobroma cacao [какао] або Theaceae як рід Camellia наприклад, рід та вид Camellia sinensis [чай]. Переважними мікроорганізмами є, наприклад, гриби, вибрані з групи сімейств: Chaetomiaceae, Choanephoraceae, Cryptococcaceae, Cunninghamellaceae, Demetiaceae, Moniliaceae, Mortiereilaceae, Mucoraceae, Pythiaceae, Sacharomycetaceae, Saprolegniaceae, Schizosacharomycetaceae, Sodariaceae або Tuberculariaceae. Для прикладу можуть бути наведені мікроорганізми вибрані з наступної групи: Choanephoraceae, такі як роди Blakeslea, Choanephora, наприклад, роди та види Blakeslea trispora, Choanephora cucurbitarum, Choanephora infundibulifera var. cucurbitarum, Mortiereilaceae, такі як рід Mortierella, наприклад, роди та види Mortierella isabellina, Mortierella polycephala, Mortierella ramanniana, Mortierella vinacea, Mortierella zonata, Pythiaceae, такі як роди Phytium, Phytophthora, наприклад, роди та види Pythium debaryanum, Pythium intermedium, Pythium irregulare, Pythium megalacanthum, Pythium paroecandrum, Pythium sylvaticum, Pythium ultimum, Phytophthora cactorum, Phytophthora cinnamomi, Phytophthora citricola, Phytophthora citrophthora, Phytophthora cryptogea, Phytophthora drechsleri, Phytophthora erythroseptica, Phytophthora lateralis, Phytophthora megasperma, Phytophthora nicotianae, Phytophthora nicotiana var. parasitica, Phytophthora palmivora, Phytophthora parasitica, Phytophthora syringe, Saccharomycetaceae, такі як роди Hansenula, Pichia, Saccharomyces, Saccharomycodes, Yarrowia, наприклад, роди та види Hansenula anomala, Hansenula californica, Hansenula canadensis, Hansenula capsulata, Hansenula ciferrii, Hansenula glucozyma, Hansenula henricii, Hansenula holstii, Hansenula minuta, Hansenula nonfermentans, Hansenula philodendri, Hansenula polymorpha, Hansenula saturnus, Hansenula subpelliculosa, Hansenula wickerhamii, Hansenula wingei, Pichia alcoholophila, Pichia angusta, Pichia anomala, Pichia bispora, Pichia burtonn, Pichia canadensis, Pichia capsulata, Pichia carsonii, Pichia cellobiosa, Pichia ciferrii, Pichia fannosa, Pichia fermentans, Pichia finlandica, Pichia glucozyma, Pichia guilliermondii, Pichia haplophila, Pichia henricii, Pichia holstii, Pichia jadinii, Pichia lindnerii, Pichia membranaefaciens, Pichia methanolica, Pichia minuta var. minuta, Pichia minuta var. nonfermentans, Pichia norvegensis, Pichia ohmeri, Pichia pastors, Pichia philodendri, Pichia ріnі, 90455 28 Pichia polymorpha, Pichia quercuum, Pichia rhodanensis, Pichia sargentensis, Pichia stipitis, Pichia strasburgensis, Pichia subpelliculosa, Pichia toletana, Pichia trehalophila, Pichia vini, Pichia xylosa, Saccharomyces aceti, Saccharomyces bailii, Saccharomyces bayanus, Saccharomyces bisporus, Saccharomyces capensis, Saccharomyces carlsbergensis, Saccharomyces cerevisiae, Saccharomyces cerevisiae var. ellipsoideus, Saccharomyces chevalieri, Saccharomyces delbrueckii, Saccharomyces diastaticus, Saccharomyces drosophilarum, Saccharomyces elegans, Saccharomyces ellipsoideus, Saccharomyces fermentati, Saccharomyces florentinus, Saccharomyces fragilis, Saccharomyces heterogenicus, Saccharomyces hienipiensis, Saccharomyces musitatus, Saccharomyces italicus, Saccharomyces kluyven, Saccharomyces krusei, Saccharomyces lactis, Saccharomyces marxianus, Saccharomyces microellipsoides, Saccharomyces montanus, Saccharomyces norbensis, Saccharomyces oleaceus, Saccharomyces paradoxus, Saccharomyces pastorianus, Saccharomyces pretoriensis Saccharomyces rosei, Saccharomyces rouxii, Saccharomyces uvarum, Saccharomycodes ludwigii, Yarrowia lipolytica, Schizosacharomycetaceae, такі як роди Schizosaccharomyces, наприклад, види Schizosaccharomyces japonicus var. japonicus, Schizosaccharomyces japomcus var. versatilis, Schizosaccharomyces malidevorans, Schizosaccharomyces octosporus, Schizosaccharomyces pombe var. malidevorans, Schizosaccharomyces pombe var. pombe, Thraustochytriaceae, такі як роди Althornia, Aplanochytrium Japonochytrium, Schizochytrium, Thraustochytrium такі як види Schizochytrium aggregatum, Schizochytrium limacmum, Schizochytrium mangrovei, Schizochytrium minutum, Schizochytrium octosporum, Thraustochytrium aggregatum, Thraustochytrium amoeboideum, Thraustochytrium antacticum, Thraustochytrium arudimentale, Thraustochytrium aureum, Thraustochytrium benthicola, Thraustochytrium globosum, Thraustochytrium indicum, Thraustochytrium kerguelense, Thraustochytrium kinnei, Thraustochytrium motivum, Thraustochytrium multirudimentale, Thraustochytrium pachydermum, Thraustochytrium proliferum, Thraustochytrium roseum, Thraustochytrium rossii, Thraustochytrium stratum або Thraustochytrium visurgense. Наступні переважні мікроорганізми є, наприклад, бактеріями, вибраними з групи сімейств: ВасіІІасеае, Enterobacteriacae або Rhizobiaceae. Як приклад можуть бути наведені мікроорганізми вибрані з наступної групи: ВасіІІасеае, такі як рід Bacillus, наприклад, як роди та види Bacillus acidocaldarius, Bacillus acidoterrestris, Bacillus alcalophilus, Bacillus amyloliquefaciens, Bacillus amylolyticus, Bacillus brevis, Bacillus cereus, Bacillus circulans, Bacillus coagulans, Bacillus sphaericus subsp. fusiformis, Bacillus galactophilus, Bacillus globisporus, Bacillus globisporus subsp. marinus, Bacillus halophilus, Bacillus lentimorbus, Bacillus lentus, Bacillus licheniformis, Bacillus megaterium, 29 Bacillus polymyxa, Bacillus psychrosaccharolyticus, Bacillus pumilus, Bacillus sphaericus, Bacillus subtilis subsp. spizizenu, Bacillus subtilis subsp. subtilis або Bacillus thuringiensis, Enterobacteriacae, такі як роди Citrobacter, Edwardsiella, Enterobacter, Erwinia, Escherichia, Klebsiella, Salmonella або Serratia, наприклад, як роди та види Citrobacter amalonaticus, Citrobacter diversus, Citrobacter freundii, Citrobacter genospecies, Citrobacter gillenii, Citrobacter intermedium, Citrobacter koseri, Citrobacter murliniae, Citrobacter sp., Edwardsiella hoshinae, Edwardsiella ictaluri, Edwardsiella tarda, Erwinia alni, Erwinia amylovora, Erwinia ananatis, Erwinia aphidicola, Erwinia billingiae, Erwinia cacticida, Erwinia cancerogena, Erwinia carnegieana, Erwinia carotovora subsp. atroseptica, Erwinia carotovora subsp. betavasculorum, Erwima carotovora subsp. odorifera, Erwinia carotovora subsp. wasabiae, Erwinia chrysanthemi, Erwinia cypripedii, Erwinia dissolvens, Erwima herbicola, Erwinia mallotivora, Erwinia milletiae, Erwinia nigrifluens, Erwinia nimipressuralis, Erwinia persicina, Erwinia psidii, Erwinia pyrifoliae, Erwinia quercina, Erwinia rhapontici, Erwinia rubrifaciens, Erwinia salicis, Erwinia stewartii, Erwinia tracheiphila, Erwinia uredovora, Escherichia adecarboxylata, Escherichia anindolica, Escherichia aurescens, Escherichia blattae, Escherichia coli, Escherichia coli var. communior, Escherichia coli-mutabile, Escherichia fergusonii, Escherichia hermannii, Escherichia sp, Escherichia vulneris, Klebsiella aerogenes, Klebsiella edwardsii subsp. atlantae, Klebsiella ornithinolytica, Klebsiella oxytoca, Klebsiella planticola, Klebsiella pneumoniae, Klebsiella pneumoniae subsp. pneumoniae, Klebsiella sp., Klebsiella terrigena, Klebsiella trevisanii, Salmonella abony, Salmonella anzona, Salmonella bongori, Salmonella choleraesuis subsp. arizonae, Salmonella choleraesuis subsp. bongori, Salmonella choleraesuis subsp. choleraesuis, Salmonella choleraesuis subsp. diarizonae, Salmonella choleraesuis subsp. houtenae, Salmonella choleraesuis subsp. indica, Salmonella choleraesuis subsp. salamae, Salmonella daressalaam, Salmonella enterica subsp. houtenae, Salmonella enteric subsp. salamae, Salmonella enteritidis, Salmonella gallinarum, Salmonella heidelberg, Salmonella panama, Salmonella senftenberg, Salmonella typhimurium, Serratia entomophila, Serratia ficaria, Serratia fonticola, Serratia grimesii, Serratia liquefaciens, Serratia marcescens, Serratia marcescens subsp. marcescens, Serratia marinorubra, Serratia odonfera, Serratia plymouthensis, Serratia plymuthica, Serratia proteamaculans, Serratia proteamaculans subsp. quinovora, Serratia quinivorans або Serrätia rubidaea, Rhizobiaceae такі як роди Agrobacterium, Carbophilus, Chelatobacter, Ensifer, Rhizobium, Sinorhizobium, наприклад як роди та види Agrobacterium atlanticum, Agrobacterium ferrugineum, Agrobacterium gelatinvorum, Agrobacterium larrymoorei, Agrobacterium meteori, Agrobacterium radiobacter, Agrobacterium rhizogenes, Agrobacterium rubi, Agrobacterium stellulatum, Agrobacterium tumefaciens, Agrobacterium vitis, Carbophilus carboxidus, 90455 30 Chelatobacter heintzii, Ensifer adhaerens, Ensifer arboris, Ensifer fredii, Ensifer kostiensis, Ensifer kummerowiae, Ensifer medicae, Ensifer meliloti, Ensifer saheli, Ensifer terangae, Ensifer xinjiangensis, Rhizobium ciceri Rhizobium etli, Rhizobium fredii, Rhizobium galegae, Rhizobium gallium, Rhizobium giardinii, Rhizobium hainanense, Rhizobium huakuii, Rhizobium huautlense, Rhizobium indigoferae, Rhizobium japonicum, Rhizobium leguminosarum, Rhizobium loessense, Rhizobium loti, Rhizobium lupini, Rhizobium mediterraneum, Rhizobium meliloti, Rhizobium mongolense, Rhizobium phaseoli, Rhizobium radiobacter, Rhizobium rhizogenes, Rhizobium rubi, Rhizobium sullae, Rhizobium tianshanense, Rhizobium trifolii, Rhizobium tropici, Rhizobium undicola, Rhizobium vitis, Smorhizobium adhaerens, Sinorhizobium arboris, Smorhizobium fredii, Smorhizobium kostiense, Smorhizobium kummerowiae, Sinorhizobium medicae, Sinorhizobium meliloti, Sinorhizobium morelense, Smorhizobium saheli або Sinorhizobium xinjiangense. Наступними переважними мікроорганізмами способу, згідно даного винаходу є, наприклад, протисти або діатомові водорості, вибрані з групи сімейств: Dinophyceae, Turaniellidae або Oxytrichidae, наприклад, роди та види: Crypthecodinium cohnii, Phaeodactylum tricornutum, Stylonychia mytilus, Stylonychia pustulata, Stylonychia putrina, Stylonychia notophora, Stylonychia sp., Colpidium campylum або Colpidium sp. Переважними трансгенними організмами у способі, згідно даного винаходу є такі гриби, як Mortierella або Thraustochytrium, дріжджі, такі як Saccharomyces або Schizosaccharomyces, мохи, такі як Physcomitrella або Ceratodon, тварини крім людини, такі як Caenorhabditis, водорості, такі як Nephroselmis, Pseudoscourfielda, Prasinococcus, Scherffelia, Tetraselmis, Mantoniella, Ostreococcus, Crypthecodinium або Phaeodactylum або рослини, такі як однодольні або дводольні. Особливо переважними організмами, які застосовуються у способі, згідно даного винаходу є такі, що належать до олійних організмів, тобто таких, які використовуються для одержання олив, такі гриби, як Mortierella або Thraustochytrium, водорості, також як Nephroselmis, Pseudoscourfielda, Prasinococcus, Scherffelia, Tetraselmis, Mantoniella, Ostreococcus, Crypthecodinium, Phaeodactylum або рослини, зокрема рослини переважно олійні рослини, які містять великі кількості ліпідних сполук, такі як арахіс, рапс, канола, соняшник, сафлор (Carthamus tinctoria), мак, гірчиця, конопля, кліщовина, оливки, сезам, календула, гранат, ослинник, коров'як, осот, шипшина, ліщина, мигдаль, макадамія, авокадо, лавр, гарбуз, льон, соя, фісташка, огуречник, дерева (маслична пальма, кокосовий горіх або грецький горіх) або польові культури, такі як кукурудза, пшениця, жито, овес, трітікале, рис, ячмінь, бавовна, манюка, перець, чорнобривці, пасльонові рослини, такі як картопля, тютюн, баклажан та помідор, бобові культури, горох, люцерна або кущові рослини (кава, какао, чай), івові рослини, також багаторічні трави та кормові польові культури. 31 Переважними рослинами згідно даного винаходу є олійні культури, такі як арахіс, рапс, канола, соняшник сафлор, мак, гірчиця, конопля, кліщовина, оливки, календула, гранат, ослинник, гарбуз, льон, соя, огуречник, дерева (маслична пальма, кокосовий горіх). Особливо переважними є рослини багаті на С18:2- та/або С18:3-жирні кислоти, такі як соняшник, сафлор красильний, тютюн, коров'як, сезам, бавовна, гарбуз, мак, ослинник, волоський горіх, льон, коноплі, будяк або сафлор красильний. Найбільш переважними є рослини, такі як сафлор красильний, соняшник, мак, ослинник, волоський горіх, льон або коноплі. Для описаного способу згідно даного винаходу є переважним, що в організм додатково на етапах опису способу (а) до (d) до введених нуклеїнових кислот, а також, за потреби вводяться послідовності нуклеїнової кислоти, які кодують -3десатурази, додатково вводяться інші нуклеїнові кислоти, які кодують ферменти обміну жирної кислоти або ліпідів. В принципі, усі гени, що кодують обмін жирної кислоти або ліпідів, можуть використовуватися в способі одержання поліненасичених жирних кислот, переважно гени обміну жирних кислот або ліпідів, вибраних з групи: ацил-СоАдегідрогеназа(и), ацил-АСР[=асуІ carrier рrotein]десатураза(и), ацил-АСР-тіоестераза(и), жирнокислотна-ацил-Ттрансфераза(и), ацил-СоА лізофосфоліпід-ацилтрансфераза жирнокислотнасинтаза(и), жирнокислотна-гідроксилаза(и), ацетил-коензим А-карбоксилаза(и), ацил-коензимАоксидаза(и), жирнокислотна-десатураза(и), жирнокислотна-ацетиленази, ліпооксигенази, триацилгліцерол-ліпази алленоксид-синтази, гідропероксид-ліази або жирнокислотна-елонгаза(и), переважно в комбінації із винайденими -5елонгазою(ами), -6-елонгазою(ами) та/або -3десатуразою(ами) [в сенсі даної заявки однина може означати множину та навпаки]. Особливо переважними є гени вибрані з групи -4десатурази, -5-десатурази, -6-десатурази, -8десатурази, -9-десатурази, -12-десатурази, -6елонгази або -9-елонгази в комбінації із вищезгаданими генами -5-елонгази, -6-елонгази та/або -3-десатурази, причому може використовуватися один ген або більше генів в комбінації. -5-елонгази, згідно даного винаходу, мають порівняно із елонгазами людини або елонгазами тварини, крім людини, такої як Oncorhynchus, Xenopus або Сionа таку властивість, як елонгування С22-жирних кислот, але не елонгування згадуваних С24-жирних кислот. В подальшому вони переважно не конвертують ніякі жирні кислоти з одним подвійним зв'язком у -6-позици, як це відбувається з елонгазами людини або елонгазами тварин, відмінних від людини. Особливо бажано, що -5-елонгази перетворюють переважно лише ненасичені С20-жирні кислоти. Ці переважні -5елонгази мають деякі передбачувані трансмембранні спіралі (5-7). Переважно конвертуються лише С20-жирні кислоти з подвійним зв'язком в -5позиції, причому -3-С20 жирні кислоти мають перевагу (ЕРА). Далі, в переважному варіанті здійс 90455 32 нення винаходу, вони мають здатність поряд із 5-елонгазною активністю не проявляти переважно ніякої або тільки відносно незначну -6-елонгазну активність. На противагу цьому, елонгази людини або елонгази тварин, що є відмінними від людей, проявляють приблизно схожу активність по відношенню до жирних кислот з -6- або -5-подвійним зв'язком. Ці переважні елонгази позначаються, як так звані, монофункцюнальні елонгази. Елонгази людини або елонгази тварин, що є відмінними від людей, позначаються, на противагу, як мультифункцюнальні елонгази, які поряд з вищезгаданими субстратами також конвертували мононенасичені С16- та С18-жирні кислоти, наприклад, з -9- або 11-подвійним зв'язком. Переважно, монофункцюнальні елонгази конвертувалися згідно опису живлення дріжджів, в якому як субстрат дріжджам додавався ЕРА, щонайменше 15мас.% доданого EPAs до докозапентаєнової кислоти (DPA, 7,10,13,16,19 С22:5 ), більш бажано щонайменше 20мас.%, особливо бажано щонайменше 25мас.%. Якщо як субстрат було додано у-ліноленову кис6,9,12 лоту (=GLA, С18:3 ), то елонгація переважно не відбувалася. Також не елонгуються i 5,9,12 С18:3 . В іншому бажаному випадку здійснення менше ніж 60мас.% доданої GLA до дигомоілі8,11,14 ноленової кислоти (=С20:3 ), бажано менше ніж 55мас,%, переважно менше ніж 50мас.%, особливо переважно менше ніж 45мас.%, надзвичайно особливо переважно менше ніж 40мас.%. В одній наступній, найбільш переважній формі здійснення даного винаходу -5-елонгазна активність GLA не конвертується. На Фігурах 27 та 28 приведено виміряну специфічність до субстрату різноманітних елонгаз. На Фігурі 27 приведено специфічності багатофункціональних елонгаз з Xenopus laevis (Фіг.27А), Ciona intestinalis (Фіг.27В) та Oncorhynchus mykiss (Фіг.27С). Усі ці елонгази переробляють широкий спектр субстратів. Це може призвести у способі згідно даного винаходу до появи побічних продуктів, які мають бути перероблені дією інших ферментів. Тому ці ферменти є менш бажаними в способі згідно даного винаходу. Переважні монофункцюнальні ферменти та їхні специфічності до субстрату приведено на Фігурі 28. На Фігурі 28А показано специфічність -5-елонгази Ostreococcus tauri. Вони конвертують жирні кислоти з подвійними зв'язками в -5-положенні. Переважно можуть конвертуватися лише С20-жирні кислоти. Також високу специфічність до субстрату проявляє -5-елонгаза із Thalassiosira pseudonana (Фіг.28С). -6-Елонгаза із Ostreococcus tauri (Фіг.28В), так само як i з Thalassiosira pseudonana (Фіг.28D), конвертує жирні кислоти із подвійним зв'язком в -6-положенні. Переважно можуть конвертуватися лише С18-жирні кислоти. Також, -5елонгаза із Arabidopsis thaliana та Euglena gracilis відрізняються своїми специфічностями. Пераважні -6-елонгази згідно даного винаходу відрізняються високою специфічністю, це означає що вони переважно елонгують цис-жирні кислоти. Вони переважно конвертують жирні кислоти із подвійним зв язком в -6-положенні. Особливо 33 переважні -6-елонгази переробляють переважно С18-жирні кислоти із трьома або чотирма подвійними зв'язками в молекулі, причому вони повинні містити подвійний зв'язок в -6-положенні. Далі, в переважному варіанті здійснення винаходу вони мають здатність поряд з -6-елонгазною активністю не проявляти переважно ніяку або тільки відносно незначну -5-елонгазну активність. На противагу цьому елонгази людини або елонгази тварин, що є відмінними від людей, проявляють приблизно схожу активність по відношенню до жирних кислот з -6 або -5-подвійним зв'язком. Ці переважні елонгази позначаються як так звані монофункцюнальні елонгази. Елонгази людини або елонгази тварин, що є відмінними від людей, позначаються, на противагу, як мультифункцюнальні елонгази, які поряд з вищезгаданими субстратами також конвертують мононенасичені С16-та С18-жирні кислоти, наприклад, з -9- або -11-подвійним зв'язком. Переважно, монофункцюнальні елонгази в описі живлення дріжджів, в якому як субстрат дріжджам додавався ЕРА, щонайменше 10ваг.% до9,12,15 даної -ліноленовоі кислоти (=АLА, С18:3 ), та відповідно, щонайменше 40ваг.% доданої 6,9,12 ліноленовоі кислоти (=GLA, С18:3 ), переважно щонайменше 20ваг.% та відповідно 50ваг.% особливо переважно 25ваг.% та відповідно 60ваг.%. Особливо переважно, також елонгується 6,9,12,15 С18:4 (стеаридонова кислота). При цьому SDA буде перероблюватися як мінімум на 40ваг.%, переважно як мінімум на 50ваг.%, особливо переважно як мінімум на 60ваг.%, в найвищій мірі переважно як мінімум на 70ваг.%. Особливо переважні -6-елонгази не проявляють або проявляють дуже незначну активність (менше ніж 0,1мас.% перетворення) по відношенню до насту6 9 11 пних субстратів С18:1 , С18:1 , С18:1 , 11,14 11,14,17 8,11,14 5,8,11,14 С20:2 , С20:3 , С20:3 , С20:4 , 5,8,11,14,17 7,10,18,16 С20:5 або С22:4 . На Фігурах 29 i 30, а також в таблиці 18 показано визначені субстрат-специфічності різних елонгаз. -3-Десатураза згідно даного винаходу по відношенню до відомої -3-десатурази має переважну властивість, що вона здатна десатурувати більш широкий спектр -6-жирних кислот, переважно десатуруються С20- та С22-жирні кислоти, такі як С20:2-, С20:3-, С20:4-, С22:4- або С20:5-жирні кислоти. Але, також можуть переважно десатуруватися й більш короткі С18-жирні кислоти, такі як С18:2- або С18:3-жирні кислоти. За допомогою цих властивостей -3-десатурази переважно можна конвертувати спектр жирних кислот від -6-жирних кислот до -3-жирних кислот, організму переважно можна рослини або гриба. Переважно -3-десатураза, згідно даного винаходу десатурує С20-жирні кислоти. В організмі ці жирні кислоти із пулу жирних кислот як мінімум на 10%, 15%, 20%, 25% або 30% конвертуються у вказані -3-жирні кислоти. По відношенню до С18-жирних кислот -3-десатураза проявляють більш незначну активність до фактора 10, це означає, що лише приблизно від 1,5 до 3% жирних кислот в жирнокислотному пулі конвертуються до -3-жирних кислот. Переважним суб 90455 34 стратом -3-десатурази згідно даного винаходу є фосфоліпіди зв'язані із -6-жирними кислотами. Фігура 19 чітко показує на прикладі десатураци 8,11,14 дигомо--ліноленової кислоти [20:4 ], що -3десатураза при десатурації переважно не розрізняє зв'язані жирні кислоти в позиції sn1- або sn2. Будуть десатуровані жирні кислоти зв'язані із фосфоліпідом в позиції або sn1- або sn2. Також є бажаним, що -3-десатураза може конвертувати широку палітру фосфоліпідів, таких як фосфатидилхолін (=РС), фосфатидилінозитол (=PIS) або фосфатидилетаноламін (=РЕ). Також продукти десатураци можуть знаходитися в нейтральних ліпідах (=NL), тобто в тригліцеридах. -4-десатурази, -5-десатурази та -6десатурази, згідно даного винаходу мають, на відміну від відомих -4-десатураз, -5-десатураз та -6-десатураз, ту перевагу що вони можуть конвертувати жирні кислоти, зв'язані із фосфоліпідами або СоА-естером жирної кислоти, переважно із СоА-естером жирної кислоти. Переважно в способі згідно даного винаходу використовували -12-десатурази для перетворення масляної кислоти (С18:19) в лінолеву кис9,12 6,9 6,9,12 лоту (С18:2 ) або С18:2 в С18:3 (=GLA). Переважно використовували -12-десатурази для перетворення жирних кислот, зв'язаних із фосфоліпідами або СоА-естером жирної кислоти, переважно із СоА-естером жирної кислоти. За допомогою ферментативної активності, способу, згідно даного винаходу, яка кодується нуклеїновими кислотами, що кодують поліпептиди із -5елонгазною, -6-елонгазною та/або -3десатуразною активністю, переважно в комбінації з послідовністю нуклеїнової кислоти, яка кодує поліпептид жирнокислотного або ліпідного обміну, такі як поліпептиди з -4-, -5-, -6-, -8-, -12десатуразною або -5--6- або -9-елонгазною активністю, можуть бути одержані різноманітні поліненасичені жирні кислоти, способом, згідно даного винаходу. В залежності від вибору використовуваних в способі згідно даного винаходу організмів, переважно рослин, можна отримувати суміші різноманітних поліненасичених жирних кислот або окремі поліненасичені жирні кислоти, такі як ЕРА або ARA у вільній або зв'язаній формі. Залежно від того, який склад жирних кислот переважає у вихідній рослині (С18:2- або С18:3-жирні кислоти), утворюються такі жирні кислоти, які походять від С18:2-жирних кислот як то GLA, DGLA або ARA, або які походять від С18:3-жирних кислот, як то SDA, ЕТА або ЕРА. Якщо рослина, що використовується у способі, в якості ненасиченої жирної кислоти містить тільки лінолеву кислоту (=LA, 9,12 С18:2 ), то у якості продуктів процесу можуть утворюватися лише GLA, DGLA та ARA, які можуть бути вільними жирними кислотами або зв'язаними. Якщо ненасиченою жирною кислотою рослини, що використовується у способі, є тільки альфа9,12,15 ліноленова кислота (=АLА, С18:3 ), наприклад як у льону то у якості продуктів процесу можуть утворюватися лише SDA, ЕТА, ЕРА та/або DHA які як зазначено вище, можуть бути вільними жирними кислотами або зв’язаними. Шляхом модифікації 35 активності ферменту, що бере участь у синтезі 5-елонгази переважно у комбінації з -4-, -5-, -6, -12-десатуразою та/або -6-елонгазою або -4-, -5-, -8-, -12-десатуразою та/або -9-елонгазою, можна цілеспрямовано отримувати у згаданих вище організмах, переважно у згаданих вище рослинах, лише певні продукти. Залежно від вихідної рослини та ненасиченої жирної кислоти дією -6десатурази та -6-елонгаз утворюються, наприклад, GLA та DGLA і, відповідно, SDA та ЕТА. Переважно, утворюються DGLA і, відповідно, ЕТА або їх суміші. Якщо додатково в організм, переважно в рослину, введено -5-десатураза, -5елонгаза та -4-десатураза, то додатково утворюються ARA, ЕРА та/або DHA. Це стосується також організмів в яких раніше були введені -8десатураза та -9-елонгаза. Переважно синтезуються тільки ARA, ЕРА або DHA або їх суміші, в залежності від організму або рослини в котрих певні жирні кислоти служать вихідними сполуками для синтезу. Коли йдеться про ланцюг біосинтезу, то відповідні кінцеві продукти в організмі містяться не у вигляді чистих речовин. Також, завжди в кінцевому продукті міститься незначна кількість сполук-попередників. Ця незначна кількість складає менше, ніж 20мас.%, переважно, менше, ніж 15мас.%, особливо бажано, менше, ніж 10мас.%, найбільш бажано, менше, ніж 5, 4, 3, 2 або 1мас.% відносно кінцевого продукту DGLA, ETA або їх сумішей і, відповідно, ARA, EPA, DHA або їх сумішей, переважно, ЕРА або DHA або їх сумішей. Згідно з винаходом, протеїн, кодований нуклеїновою кислотою, показує високу специфічність по відношенню до обох проміжних сполук, 6,9,12,15 5,8,11,14,17 С18:4 та С20:5 -жирних кислот синтезу DHA (проміжні сполуки та синтез DHA показано на Фігурі 1). Протеїн, кодований SEQ NO:53, має разом з тим специфічність до -6 та -5жирних кислот з додатковим -3-подвійним зв'язком (Фігура 2). -5-Елонгаза має кето-ацил-СоАсинтазну активність, яка подовжує переважно жирнокислотнии залишок від ацил-СоА-естерів на 2 вуглецевих атома. За допомогою гену -5-елонгази, -5десатурази з Phaeodactylum а також -4десатурази з Euglena було доведено синтез DHA в дріжджах (Saccharomyces cerevisiae) (Фігура 3). Поряд із продукуванням вихідних жирних кислот для -5-елонгази, -6-елонгази та/або -3десатурази безпосередньо в організмі жирні кислоти можуть поглинатися ззовні Зважаючи на вартість, переважним є їх утворення в організмі. Переважно субстратом -3-десатурази є лінолева 9,12 кислота (С18:2 ), -ліноленова кислота 11,14 6,9,12 (С18:3 ), ейкозадієнова кислота (С20:2 ), 8,11,14 дигомо--ліноленова кислота (С20:3 ), арахі5,8,11,14 донова кислота (С20:4 ), докозатетраєнова 7,10,13,16 кислота (С22:4 ) та докозапентаєнова кис4,7,10,13,15 лота (С22:5 ). Для підвищення в описаному способі виходу одержуваних олій та/або тригліцеридів, бажано із підвищеним вмістом поліненасичених жирних кислот, бажано збільшувати масу вихідного продукту для синтезу жирної кислоти, що може досягатися, 90455 36 наприклад, введенням в організм нуклеїнової кислоти, яка кодує поліпептид -12-десатурази. Особливо переважними серед олійно-продукуючих організмів є ті, що мають підвищений вміст жирних кислот, таких як сімейство Brassicaceae, рід Brassica, наприклад, рапс; сімейство ЕІаеаgnасеае рід Elaeagnus, наприклад, рід та вид ОІеа еurораеа або сімейство Fabaceae рід Glycine, наприклад, рід та вид Glycine max. Оскільки ці організми мають незначний вміст лінолевої кислоти (Mikokläjczak та ін., Journal of the American Oil Chemical Society, 38, 1961, 678-681) переважним є застосування згаданої -12-десатурази для одержання вихідного продукту ліноленової кислоти. Нуклеїнові кислоти застосовані у способі згідно даного винаходу походять переважно з рослин таких як водорості, наприклад, водоростей сімейства Prasinophyceae, родів Heteromastix, Mammella, Mantoniella, Micromonas, Nephroselmis, Ostreococcus, Prasinocladus, Prasinococcus, Pseudoscourfielda, Pycnococcus, Pyramimonas, Scherffelia або Tetraselmis, родів та видів Heteromastix longifillis, Mamiella gilva, Mantoniella squamata, Micromonas pusilla, Nephroselmis olivacea, Nephroselmis pyriformis, Nephroselmis rotunda, Ostreococcus tauri, Ostreococcus sp. Prasinocladus ascus, Prasinocladus lubricus, Pycnococcus provasolii, Pyramimonas amylifera, Pyramimonas disomata, Pyramimonas obovata, Pyramimonas orientalis, Pyramimonas parkeae, Pyramimonas spinifera, Pyramimonas sp., Tetraselmis apiculata, Tetraselmis carteriaformis, Tetraselmis chui, Tetraselmis convolutae, Tetraselmis desikacharyi, Tetraselmis gracilis, Tetraselmis hazeni, Tetraselmis impellucida, Tetraselmis inconspicua, Tetraselmis levis, Tetraselmis maculata, Tetraselmis marina, Tetraselmis striata, Tetraselmis subcordiformis, Tetraselmis suecica, Tetraselmis tetrabrachia, Tetraselmis tetrathele, Tetraselmis verrucosa, Tetraselmis verrucosa fo. rubens або Tetraselmis sp. або з водоростей сімейства Euglenaceae родів Ascoglena, Astasia, Colacium, Cyclidiopsis, Euglena, Euglenopsis, Hyalophacus, Khawkinea, Lepocinclis, Phacus, Strombomonas або Trachelomonas родів та видів Euglena acus, Euglena geniculata, Euglena gracilis, Euglena mixocylindracea, Euglena rostrifera, Euglena viridis, Colacium stentorium, Trachelomonas cylindrica або Trachelomonas volvocina. Переважно застосовані нуклеїнові кислоти походять із водоростей Algen родів Euglena, Mantoniella або Ostreococcus. Наступними переважними рослинами є водорості, такі як Isochrysis або Crypthecodinium, діатомові водорості, такі як Thalassiosira або Phaeodactylum, мохи, такі як Physcomitrella або Ceratodon або вищі рослини такі як Primulaceae такі як Aleuritia, Calendula stellata, Osteospermum spinescens або Osteospermum hyoseroides, мікроорганізми, такі як гриби як Aspergillus, Thraustochytrium, Phytophthora, Entomophthora, Mucor або Mortierella, бактерії, такі як Shewanella, дріжджі або тварини, такі як нематоди такі як Caenorhabditis, комахи, жаби, морські кубишки або риби. Виділені послідовності нуклеїнової кислоти, згідно даного винаходу, переважно походять від 37 тварин з порядку хребетних. Послідовності нуклеїнової кислоти, переважно походять з класу Vertebrata; Euteleostomi, Actinopterygii; Neopterygii; Teleostei; Euteleostei, Protacanthopterygii, Salmoniformes, Salmonidae або Oncorhynchus або Vertebrata, Amphibia, Anura, Pipidae, Xenopus або Evertebrata, такі як Protochordata, Tunicata, Holothuroidea, Cionidae як то Amaroucium constellatum, Botryllus schlosseri, Ciona intestinalis, Molgula citrina, Molgula manhattensis, Perophora vindis або Styela partita. Нуклеїнові кислоти, особливо переважно походять з грибів, тварин або з рослин таких як водорості або мохи, переважно з порядку Salmoniformes як наприклад сімейство Salmonidae таких як рід Salmo наприклад, рід та вид Oncorhynchus mykiss, Trutta trutta або Salmo trutta fario, з водоростей таких як Mantoniella або Ostreococcus або з діатомових водоростей таких як рід Thalassiosira або Phaeodactylum водоростей таких як Crypthecodinium. Переважно у способі згідно даного винаходу поліпептид, кодований згаданими послідовностями нуклеїнової кислоти або їхніми похідними або гомологами також буде мати ферментативну активність протеїну, кодованого послідовностями нуклеїнової кислоти. Ці послідовності можуть використовуватися окремо або в комбінації із послідовностями, що кодують -12-десатуразу, 4-десатуразу, -5-десатуразу, -6-десатуразу, -5елонгазу, -6-елонгазу та/або -3-десатуразу, бути клонованими в експресійному конструкті й використаними для введення й експресії в організмі. Ці експресійні конструкти роблять можливим, за допомогою своєї конструкції, переважний оптимальний синтез поліненасичених жирних кислот способом згідно даного винаходу. У переважній формі здійснення спосіб включає етап отримання клітини або цілого організму, який містить послідовності нуклеїнової кислоти використані у способі, причому клітина та/або організм є трансформованими послідовністю нуклеїнової кислоти, згідно даного винаходу, яка кодує -12десатуразу, -4-десатуразу, -5-десатуразу, -6десатуразу, -5-елонгазу, -6-елонгазу та/або -3десатуразу генетичний конструкт або вектор, як описано далі, окремо або в комбінації із наступної послідовністю нуклеїнової кислоти, яка кодує протеїни жирнокислотного або ліпідного обміну. В наступному переважному варіанті здійснення спосіб включає подальший етап виділення масел, ліпідів або вільних жирних кислот із організму або з культури. Під культурою мається на увазі, наприклад, ферментаційна культура наприклад у випадку культивування мікроорганізмів, таких, наприклад, як Mortierella, Thalassiosira, Mantoniella, Ostreococcus, Saccharomyces або Thraustochytrium, або мається на увазі теплична чи польова культура рослини. Такою одержаною клітиною або таким одержаним організмом є переважно клітина організму, що продукує масла, така як рослина олійної культури, як то, наприклад, арахіс, рапс(ріпак), канола, льон, коноплі, арахіс, соя, сафлор, коноплі, соняшник або огуречник. Під вирощуванням необхідно розуміти, наприклад, культивування на або в поживному середо 90455 38 вищі у випадку рослинних клітин, тканин або органів, або цілої рослини на або в субстраті, наприклад, у гідрокультурі, ґрунті квіткового горщика або на ріллі. "Трансген" та, відповідно, "Рекомбінант" згідно з винаходом означає, наприклад, послідовність нуклеїнової кислоти, експресійну касету (=генний конструкт) або вектор, що містить послідовність нуклеїнової кислоти згідно з винаходом, або організм, трансформований послідовностями нуклеїнових кислот, згідно з винаходом, експресійною касетою або вектором конструкції, одержані генноінженерними методами в яких або, a) послідовність нуклеїнової кислоти, згідно з винаходом, або b) функціонально зв'язана з послідовністю нуклеїнової кислоти згідно з винаходом, генетична регуляторна послідовність, наприклад промотор, або c) (а) та (b) не містяться в їхньому природному генетичному середовищі або модифіковані генноінженерними методами, причому модифікацією може бути, як приклад, заміщення, доповнення, делеція, інверсія або вставка одного або декількох нуклеотидних залишків. Під природнім генетичним середовищем мається на увазі природній геномний та, відповідно, хромосомний локус у вихідному організмі або знаходження у геномній бібліотеці. У випадку геномної бібліотеки природне, генетичне середовище бажано щонайменше ще частково містить послідовність нуклеїнової кислоти. Оточення фланкує послідовності нуклеїнової кислоти щонайменше з одного боку та має довжину послідовності від щонайменше 50bр (пар основ), переважно, щонайменше 500bр, особливо бажано, щонайменше 1000bр, найбільш бажано, щонайменше 5000bр. Експресійна касета, що зустрічається в природі, наприклад, природна комбінація промоторів послідовності нуклеїнової кислоти, згідно даного винаходу що зустрічаються в природі згадуваних генів -12-десатурази-, -4-десатурази-, -5 десатурази-, -6-десатурази-, -8-десатурази-, -3-десатурази-, -9-елонгази-, -6-елонгазита/або -5-елонгази- знаходяться в одній експресійній касеті, де вони змінюються неприродним, синтетичним ("штучним") шляхом, як наприклад, мутуванням. Відповідні способи описані, наприклад, в US5,565,350 або WO00/15815. Під трансгенним організмом та, відповідно, трансгенною рослиною згідно з винаходом як вказано вище, розуміють, що нуклеїнові кислоти, що застосовуються у способі, знаходяться не на їх природному місці у геномі організму, при цьому можуть експресуватися гомологічні або гетеролопчні нуклеїнові кислоти. Трансген означає, як було зазначено, що послідовності нуклеїнової кислоти, згідно даного винаходу, знаходяться в геномі організму на своєму природному місці, однак при цьому послідовність порівняно із природною послідовністю була змінена та/або змінені регуляторні послідовності природних послідовностей. Переважно, під трансгеном розуміється експресія нуклеїнової кислоти, згідно даного винаходу в неприродному місці в геномі, що означає гомологічну або 39 переважно гетеролопчну експресію нуклеїнової кислоти. Переважними трансгенними організмами є гриби, такі як Mortierella або Phytophtora, мохи, такі як Physcomitrella, водорості, такі як Mantoniella, Euglena, Crypthecodinium або Ostreococcus, діатомові водорості, такі як Thalassiosira або Phaeodactylum або рослини, такі як олійні культури. Як організми або організми-хазяїни для застосованої в способі згідно даного винаходу нуклеїнової кислоти, експресійної касети або вектору переважно принципово придатні всі організми, які можуть спеціально синтезувати жирні кислоти, особливо ненасичені жирні кислоти або експресувати рекомбінантні гени. Прикладом можуть бути рослини, такі як Arabidopsis, Asteraceae, такі як Calendula або культурні рослини, такі як соя, арахіс, рицина, соняшник, кукурудза, хлопок, лен, рапс, кокосовий горіх, оливкова пальма, сафлор (Carthamus tinctorius) або какао боби, мікроорганізми, такі як гриби, наприклад родів Mortierella, Thraustochytrium, Saprolegnia, Phytophtora або Pythium, бактерії, такі як роди Escherichia або Shewanella, дріжджі, такі як роди Saccharomyces, Cyanobacteria, Ciliates, Alga, такі як Mantoniella, Euglena, Thalassiosira або Ostreococcus або найпростіші такі динофлагеляти як Crypthecodinium. Переважно, організмами які природно можуть синтезувати олії у великих кількостях є гриби, такі як Mortierella alpina, Pythium insidiosum, Phytophtora infestans або рослини, такі як соя, рапс, кокосовий горіх, оливкова пальма, сафлор, льон, конопля, рицина, календула, арахіс, какао боби, або соняшник або дріжджі, такі як Saccharomyces cerevisiae, особливо переважними є соя, льон, рапс, сафлор, соняшник, календула, Mortierella або Saccharomyces cerevisiae. Принципово, як господарські організми, поряд із зазначеними трансгенними організмами також можуть застосовуватися трансгенні тварини, переважно тварини, за виключенням людей, наприклад, C.elegans, Ciona intestinalis або Xenopus laevis. Придатні клітини-хазяїни, приведені в: Goeddel, Gene Expression Technology:Methods in Enzymology 185, Academic Press, San Diego, CA (1990). Придатні до застосування експресійні лінії, наприклад, такі, що проявляють більш значну протеазну активність описані в: Gottesman, S., Gene Expression Technology:Methods in Enzymology 185, Academic Press, San Diego, California (1990) 119128. До них належать згадані рослинні клітини та певні рослинні тканини, органи та частини в усіх їх формах, таких як пильники, волокна, кореневі волоски, стебла, ембріони, калюси, сім'ядолі, черешки, матеріал врожаю, рослинні тканини, репродуктивні тканини та культури тканин, які можуть бути одержані з даної трансгенної рослини та/або використані з цією метою. Трансгенні рослини, які містять синтезовані способом згідно даного винаходу поліненасичені жирні кислоти, переважно можуть безпосередньо продаватися без виділення з них синтезованих олій, ліпідів або жирних кислот. Під рослинами у 90455 40 способі згідно даного винаходу розуміють рослини взагалі, такі як частини рослин, органи рослин або такі частини рослин як лист, стебло, сім'я, корінь, бульби, пильники, волокна, кореневі волоски, стебла, ембріони, калюси, сім'ядолі, черешки, матеріал врожаю, рослинні тканини, репродуктивні тканини, клітинні культури, які можуть використовуватися для виведення трансгенної рослини або створення трансгенної рослини. При цьому насіння охоплює всі частини насіння, такі як насіннєва оболонка, епідерміс та насіннєва клітина, ендосперм або ембріональна тканина. Сполуки, одержані способом згідно даного винаходу, також можуть бути виділені з організмів, переважно рослин, у формі олій, жирів, ліпідів та/або вільних жирних кислот. За допомогою цього способу поліненасичені жирні кислоти можна одержувати або збиранням організмів з культури, в якій вони продукуються, або з поля. Це може відбуватися чавленням або екстракцією частин рослин, переважно насіння рослин. При цьому олії, жири, ліпіди та/або вільні жирні кислоти можуть одержуватися так званим холодним розбиттям та холодним пресуванням без подавання тепла пресами. Щоб частини рослин, особливо насіння легше розкривалися, вони подрібнюються, розм'якшуються або жаряться. Потім насіння яке пройшло таку попередню обробку, може віджиматися або екстрагуватися розчинником таким як теплий гексан. Потім розчинник видаляється. У випадку із мікроорганізмами вони обробляються після збирання, наприклад, екстрагуються, безпосередньо без наступних технологічних етапів або екстрагуються методами відомими спеціалісту. Таким чином можуть виділятися більше 96% сполук одержаних даним способом. Потім ці одержані продукти далі перероблюються, тобто рафінуються. При цьому видаляються, наприклад, слизові речовини та замутнюючі матеріали. Під час цієї, так званої, рафінації, може відбуватися ферментне або наприклад фізичне/хімічне приєднання кислоти, наприклад, фосфорної кислоти. Потім вільні жирні кислоти видаляються шляхом обробки основою, наприклад, розчином їдкого натру. Одержаний продукт інтенсивно промивається водою для видалення залишків лугу та висушується. Для видалення забарвлюючих речовин, які ще містяться в продукті, проводять обробку відбілювачами, такими як відбілююча глина або активоване вугілля. На останок продукт додатково дезодорується, наприклад водяною парою. Переважно одержані цим способом PUFAs або LCPUFAs представляють собою С18-, С20- або С22молекули жирних кислот, переважно С20- або С22молекули жирних кислот із як мінімум двома подвійними зв'язками в молекулі жирної кислоти, переважно трьома, чотирма, п'ятьма або шістьма подвійними зв'язками. Ці С18-, С20- або С22молекули жирної кислоти можуть бути виділені із організму у вигляді олій, ліпідів або вільних жирних кислот. Придатними є, наприклад, вище перераховані організми. Переважними організмами є трансгенні рослини. Тому реалізацією винаходу є олії, ліпіди або жирні кислоти або їхні фракції, які одержані описа 41 ним вище способом, зокрема олія, ліпід або композиція жирних кислот, які містять PUFAs та одержані із трансгенних рослин. Ці олії, ліпіди або жирні кислоти містять, як описано вище, переважно від 6 до 15% пальмітинової кислоти, від 1 до 6% стеаринової кислоти; від 7-85% масляної кислоти; від 0,5 до 8% вакценової кислоти, від 0,1 до 1% арахінової кислоти, від 7 до 25% насичених жирних кислот, від 8 до 85% мононенасичених жирних кислот та від 60 до 85% поліненасичених жирних кислот відповідно до 100% та повний вміст жирних кислоти в організмі. Бажано, щоб частка поліненасичених жирних кислот в складних естерах жирних кислот або сумішах жирних кислот становила, переважно, принаймні 0,1; 0,2; 0,3; 0,4; 0,5; 0,6; 0,7; 0,8; 0,9 або 1% у перерахунку на загальний вміст арахідонової кислоти. Далі, складні естери жирних кислот або суміші жирних кислот, одержані способом згідно даного винаходу, бажано жирні кислоти вибрані з групи жирних кислот: ерукова кислота (13докозаєнова кислота), стеркулінова кислота (9,10метилен октадек-9-енова кислота), малвалінова кислота (8,9-метилен гептадек-8-енова кислота), хаульмугрова кислота, (циклопентдодеканова кислота), фуран-жирна кислота (9,12-епоксиоктадека-9,11-дієнова кислота), фернова кислота (9,10-епоксиоктадек-12-енова кислота), таринова кислота (6-октадецинова кислота), 6нонадецинова кислота, санталбінова кислота (t11октадецен-9-інова кислота), 6,9-октадеценінова кислота, пірулінова кислота (t10-гептадецен-8інова кислота), крепенінова кислота (9-октадецен12-інова кислота), 13,14-дигідроорофеїнова кислота, октадецен-13-ен-9,11-діінова кислота, петроселенова кислота (цис-6-октадеценова кислота), 9с,12t-октадекадієнова кислота, календулова кислота (8t10t12c-октадекатриєнова кислота), катальтнова кислота (9t11t13с-октадекатриєнова кислота), елеостеринова кислота (9c11t13tоктадекатриенова кислота), якаринова кислота (8с10t12с-октадекатриєнова кислота), пініцинова кислота (9c11t13c-октадекатетраенова кислота) парінарінова кислота (9с11t13t15соктадекатетраєнова кислота), піноленова кислота (аІІ-цис-5,9,12-октадекатриєнова кислота), лабаленова кислота (5,6-октадекадієналенова кислота), ріцинолова кислота (12-гідроксимасляна кислота) та/або коріолінова кислота (13-гідрокси9c,11t-октадієнова кислота). Вище згадані жирні кислоти знаходяться у складних естерах жирних кислот або сумішах жирних кислот, одержаних за способом згідно даного винаходу, як правило лише в слідових кількостях, це означає, що їхній вміст становить, по відношенню до загального вмісту жирних кислот менше ніж 30%, переважно менше ніж 25%, 24%, 23%, 22% або 21%, особливо переважно менше ніж 20%, 15%, 10%, 9%, 8%, 7%, 6% або 5%, в найвищій мірі переважно менш ніж 4%, 3%, 2% або 1%. В наступній переважній формі здійснення винаходу вміст згаданих вище цих жирних кислот, по відношенню до загального вмісту жирних кислот, становить менше ніж 0,9%; 0,8%, 0,7%; 0,6%; або 0,5%, особливо переважно менше ніж 0,4%, 0,3%, 0,2%; 0,1%. Переважно 90455 42 естери жирних кислот або суміші жирних кислот, одержані за способом згідно даного винаходу, містять менше ніж 0,1%, по відношенню до загального вмісту жирних кислот, та/або не містять масляної кислоти, холестерину, клупанодонової кислоти 4,8,12,15,21 (=докозапентаєнова кислота, С22:5 ), а також нізинової кислоти (тетракозагексаєнова кис3,5,12,15,18,21 лота, С23:6 ). Переважно олії, ліпіди або жирні кислоти, згідно даного винаходу містять як мінімум 0,5%, 1%, 2%, 3%, 4% або 5%, переважно як мінімум 6%, 7%, 8%, 9% або 10%, особливо переважно як мінімум 11%, 12%, 13%, 14% або 15% ARA або як мінімум 0,5%, 1%, 2%, 3%, 4% або 5%, переважно як мінімум 6%, або 7%, особливо переважно як мінімум 8%, 9% або 10% ЕРА та/або DHA по відношенню до загального вмісту жирних кислот продукуючого організму, переважно рослини, особливо переважно олійної рослини, такої як соя, рапс, кокосовий горіх, оливкова пальма, сафлор красильний льон, конопля, рицини, календула, арахіс, какао боби, соняшник або вищеназвані одно- або дводольні олійні культури. Наступною реалізацією згідно даного винаходу є одержання олій, ліпідів, жирних кислот та/або композицій жирних кислот у кормах, продуктах харчування, косметиці або фармацевтиці. Олії, ліпіди, жирні кислоти або суміші жирних кислот, згідно даного винаходу, можуть бути змішані способом, відомим професіоналу, із оліями, ліпідами, жирними кислотами та/або сумішами жирних кислот тваринного походження, такими як, наприклад, риб'ячий жир. Також ці олії, ліпіди, жирні кислоти або суміші жирних кислот, які складаються із рослинних або тваринних складових, можуть використовуватися для одержання кормів, продуктів харчування, косметики або фармацевтики. Під поняттям "олія", "ліпід" або "жир" розуміється суміш жирних кислот, яка містить ненасичену, насичену, переважно естерифіковану(і) жирну(і) кислоту(і). Переважно, олія, ліпід або жир мають підвищений вміст поліненасичених вільних або бажано естерифікованих жирних кислот, зокрема, лінолеву кислоту, -ліноленову кислоту, дигомо--ліноленову кислоту, арахідонову кислоту, ліноленову кислоту, стеаридонову кислоту, ейкозатетраєнову кислоту, ейкозапентаєнову кислоту, докозапентаєнову кислоту або докозагексаєнову кислоту. Переважно, вміст ненасичених естерифікованих жирних кислот становить приблизно 30%, більш переважно частка становить від 50%, ще більш переважно частка становить від 60%, 70%, 80% або більше. Визначення, наприклад, частки жирної кислоти після перетворення її на метиловий естер переестерифікацією, може бути проведено газохроматографічно. Олія, ліпід або жир можуть містити різні інші насичені або ненасичені жирні кислоти, наприклад, календулову кислоту, пальмітинову, стеаринову, масляну та ін. Зокрема, залежно від вихідного організму, може коливатися частка різних жирних кислот в олії або жирі. У способі одержання поліненасичених жирних кислот, які містять як мінімум два подвійні зв'язки, йдеться, як описано вище про, наприклад, сфінголіпіди, фосфатидилгліцериди, ліпіди, гліколіпіди, 43 фосфоліпіди, моноацилгліцерин, диацилгліцерин, триацилгліцерин або інший естер жирної кислоти. Із одержаних способом згідно даного винаходу поліненасичених жирних кислот із принаймні п'ятьма або шістьма подвійними зв'язками, поліненасичені жирні кислоти можна вивільняти, наприклад, за допомогою лужної обробки, наприклад, водними KОН або NaOH, або кислотним гідролізом бажано в присутності спиртів, таких як метанол або етанол, або ферментативним вивільненням, та виділяти, наприклад, розділенням фаз й наступним підкисленням за допомогою, наприклад, H2SO4. Вивільнення жирних кислот може відбуватися без вище описаної дообробки. Використані у способі нуклеїнові кислоти можуть бути інтегровані введенням в організм, наприклад, рослинну клітину або рослину або введенням в плазміду або переважно в геном клітинихазяїна. Під час інтеграції в геном може відбуватися інтеграція випадковим чином або подібним чином, так що, нативний ген замінюється введеною копією, внаслідок чого клітиною модулюється виробництво бажаної сполуки, або таким використанням гену, при якому ген виявляється зв'язаним із функціональною одиницею експресії, яка містить, як мінімум одну експресію гену, яка забезпечує послідовність та як мінімум одне поліаденілування функціонально транскрибованого гену, який забезпечує послідовність. Переважно нуклеїнові кислоти переносяться в організми мультиекспресійними касетами або конструктами із мультипаралельною експресією, переважно мультипаралельною насіннєвоспецифічною експресією генів в рослині. Мохи та водорості є єдиними відомими рослинними системами, які виробляють значні кількості поліненасичених жирних кислот, таких як арахідонова кислота (ARA) та/або ейкозапентаєнова кислота (ЕРА) та/або докозапентаєнова кислота (DHA). Мохи містять PUFAs в мембранних ліпідах, тоді як водорості, організми подібні до водоростей, та деякі гриби також акумулюють значні маси PUFAs в триацилгліцерольній фракції. Тому молекули нуклеїнової кислоти виділяються із таких родів, які акумулюють PUFAs також в триацилгліцерольній фракції, особливо бажано для способу згідно даного винаходу та поряд із цим для модифікації систем продукування ліпідів та PUFA в хазяїні, зокрема в рослинах, таких як олійні рослини, наприклад рапс, канола, льон, конопля, соя, соняшник, огірочник. Тому вони є переважно застосовуваними у способі згідно даного винаходу. Як субстрат для нуклеїнових кислот, використаних у способі згідно даного винаходу, які кодують поліпептиди із -12-десатуразною, -5десатуразною, -4-десатуразною, -6десатуразною, -8-десатуразною, -9-елонгазною, -5-елонгазною, -6-елонгазною та/або -3десатуразною активністю, та/або подальшого використання таких нуклеїнових кислот як нуклеїнові кислоти, що кодують поліпептиди обміну жирних кислот або ліпідів, вибраних з групи ацил-СоАдегідрогеназ(и), ацил-АСР [=acyl carrier protein]десатураз(и), ацил-АСР-тіоестераз(и), жирнокислотної(их)-ацил-трансфераз(и), ацил-СоА: лізофо 90455 44 сфоліпід-ацилтрансфераз(и), жирнокислотної(их)синтаз(и), жирнокислотної(их)-гідроксилаз(и), ацетил-коензим А-карбоксилаз(и), ацил-коензимАоксидаз(и), жирнокислотної-десатураз(и), жирнокислотної-ацетиленаз(и), ліпооксигенази, триацилгліцерол-ліпази, алленоксид-синтази, гідропероксид-ліази або жирнокислотної(их)-елонгаз(и), переважно С16-, С18- або С20-жирні кислоти. Переважно, у способі як субстрат застосовуються жирні кислоти у формі їхніх Ацил-СоА-естерів та/або їхніх фосфоліпідних естерів. Для одержання довголанцюгових PUFAs згідно даного винаходу, поліненасичені С18-жирні кислоти повинні спочатку бути десатуровані за допомогою ферментативної десатуразної дії, а потім за допомогою елонгази подовжені, як мінімум на два атоми вуглецю. Після циклу елонгації ця елонгазна дія приводить до одержання С20-жирних кислот, а наступні два цикли елонгації до С22-жирних кислот. Дія десатураз та елонгаз у способі згідно даного винаходу приводить до переважно С18-, С20- та/або С22-жирних кислот, переважно, як мінімум, із двома подвійними зв'язками в молекулі жирної кислоти, переважно із трьома, чотирма, п'ятьма, або шістьма подвійними зв'язками, особливо переважно, С20- та/або С22-жирних кислот, як мінімум, із двома подвійними зв'язками в молекулі жирної кислоти, переважно із трьома, чотирма, п'ятьма, або шістьма подвійними зв'язками, у найвищій мірі переважно п'ятьма, або шістьма подвійними зв'язками у молекулі. Після того, як відбулися перші десатурація та подовження, можуть продовжуватися подальші етапи десатурації та елонгації, такі як, наприклад, десатурації в -5- та -4положеннях. Особливо переважно продуктом способу згідно винаходу ε дигомо--лінолева кислота, арахідонова кислота, ейкозапентаєнова кислота, докозапентаєнова кислота та/або докозагексаєнова кислота. С20-Жирні кислоти, як мінімум, із двома подвійними зв'язками в жирній кислоті можуть бути подовжені ферментативною дією, згідно даного винаходу у формі вільних жирних кислот або у формі естерів, таких як фосфоліпід, гліколіпід, сфінголіпід, фосфогліцерид, моноацилгліцерин, диацилгліцерин або триацилгліцерин. Бажаним місцем біосинтезу жирних кислот, олій, ліпідів або жирів у переважно використовуваних рослинах є, наприклад, в основному насіння або клітинні шари насіння, так що раціональною є насіннєспецифічна експресія нуклеїнових кислот що використовуються у способі. Однак є зрозумілим, що біосинтез жирних кислот олій або ліпідів не повинен бути обмежений тканинами насіння, але він також може відбуватися тканиноспецифічно в усіх частинах рослини, наприклад, в клітинах епідермісу або в бульбах. В запропонованому згідно з винаходом способі як організми застосовуються такі мікроорганізми, як дріжджі, такі як Saccharomyces або Schizosaccharomyces, грибки, такі як Mortierella, Aspergillus, Phytophtora, Entomophthora, Mucor або Thraustochytrium, водорості, такі як Isochrysis, Mantoniella, Euglena, Ostreococcus, Phaeodactylum або Crypthecodinium тому що ці організми є ферментативно активними 45 Застосовуючи нуклеїнові кислоти, згідно даного винаходу, які кодують -5-елогазу, можливо в способі підвищити одержані поліненасичені жирні кислоти до як мінімум 5%, бажано як мінімум 10%, особливо бажано як мінімум 20%, найбільш бажано як мінімум 50% порівняно із організмами дикого типу, які не містять рекомбінантних нуклеїнових кислот. Способом, згідно даного винаходу, можна збільшити одержані поліненасичені жирні кислоти у організмах, що використовуються у способі у двох видах. Можливо що бажаним є збільшити пул вільних поліненасичених жирних кислот та/або збільшити частку естерифікованих поліненасичених жирних кислот, одержаних цим способом. Переважно за допомогою способу, згідно даного винаходу збільшується пул естерифікованих поліненасичених жирних кислот в трансгенних організмах. Якщо в способі згідно даного винаходу як організми використовуються мікроорганізми, то вони залежно від організму хазяїна вирощуються способом, відомим професіоналам. Мікроорганізми, як правило, вирощуються в рідкому середовищі, яке містить джерело вуглецю, переважно, у формі цукру, джерело азоту, переважно, у формі органічного джерела азоту, таке як дріжджовий екстракт або солі, такі як сульфат амонію, мікроелементи, такі як солі заліза, марганцю, магнію та, за потреби, вітаміни, при температурі від 0°С до 100°С, переважно між 10°С та 60°С та при подачі кисню. При цьому значення рН поживного середовища може підтримуватися на певному рівні, тобто під час розведення може або регулюватися, або ні. Розведення може бути завантажувального типу, напівзавантажувального типу або безперервного. Поживні речовини можуть вводитися на початку ферментації або додаватися напівнеперервно або безперервно. Одержані поліненасичені жирні кислоти можуть бути виділені з організмів способом відомим досвідченому професіоналу, як описано вище. Наприклад, екстракцією дистиляцією, кристалізацією, за потреби, сольовим осадженням та/або хроматографією. Бажано, щоб організми для цього були попередньо подрібнені. При роботі із мікроорганізмами, спосіб, згідно даного винаходу здійснюється при температурі від 0°С до 95°, бажано від 10°С до 85°С, особливо переважно бажано від 15°С до 75°С, найбільш переважно бажано від 15°С до 45°С. Значення рН при цьому бажано становить від рН 4 до 12, переважно бажано від рН 6 до 9, особливо переважно бажано від рН 7 до 8. Спосіб згідно даного винаходу може бути порційним, напівпорційним або безперервним. Опис відомих способів культивування може бути знайдений у посібнику Chmiel (Bioprozeßtechnik 1.Emführung in die Bioverfahrenstechnik (Gustav Fischer Verlag, Stuttgart, 1991)) або в посібнику Storhas (Bioreaktoren und penphere Einnchtungen (Vieweg Verlag, Braunschweig/Wiesbaden, 1994)). Використовуване культуральне середовище має задовольняти вимогам означеним у способі відповідних штамів. Описи культуральних середовищ різних мікроорганізмів знаходяться в підруч 90455 46 нику "Manual of Methods for General Bacteriology" Американського товариства бактеріологи (Washington D. С., USA, 1981). Середовища згідно даного винаходу, як описано вище зазвичай повинні мати одне або більше джерел вуглецю, азоту, неорганічних солей, вітамінів та/або мікроелементів. Переважно джерелом вуглецю є цукор, такий як моно-, ди- або полісахариди. Дуже добрими джерелами вуглецю є наприклад, глюкоза, фруктоза, манноза, галактоза, рибоза, сорбоза, рибулоза, лактоза, мальтоза, сахароза, раффіноза, крохмаль або целюлоза. Можна вводити в середовище цукор в комплексних речовинах таких як меляси, або інші побічні продукти рафінування цукру. Також можливо додавати суміші різних джерел вуглецю. Іншими можливими джерелами вуглецю є олії та жири, такі як, наприклад, соєва олія, соняшникова олія, арахісова олія, та/або кокосове масло, жирні кислоти, такі як, наприклад, пальмітинова кислота, стеаринова кислота та/або лінолева кислота, алкоголі та/або поліалкоголі, такі як, наприклад, гліцерин, метанол та/або етанол, та/або органічні кислоти, такі як, наприклад, оцтова кислота та/або молочна кислота. Джерелами азоту зазвичай є органічні або неорганічні сполуки азоту або матеріали, які містять ці сполуки. Прикладом джерел азоту є, наприклад, аміак у рідкій або газоподібній формі або солі амонію, такі як сульфат амонію, хлорид амонію, фосфат амонію, карбонат амонію або нітрат амонію, нітрати, сечовина, амінокислоти, або комплексні джерела азоту, такі як кукурудзяний екстракт, соєве борошно, соєвий білок, дріжджовий екстракт, м'ясний екстракт та інші. Джерела азоту можуть використовуватися як окремо так i як суміш. Неорганічні сполуки, які можуть міститися в середовищах, включають, хлоридні, фосфорні або сульфатні солі кальцію, магнію, натрію, кобальту, молібдену, калію, марганцю, цинку та заліза. Як джерело сірки для одержання сірковмісних речовин, зокрема, метіоніну, можуть застосовуватися неорганічні сірковмісні сполуки, як наприклад, сульфати, сульфіти, дитіоніти, тетратіоніти, тіосульфати, сульфіди або також органічні сполуки сірки, такі як меркаптани та тіоли. Джерелом фосфору може бути фосфорна кислота, дигідрофосфат калію, гідрофосфат калію або перераховані натрійвмісні солі. В середовище можуть додаватися хелатоутворювачі для утримування іонів металу в розчині. Особливо придатний хелатоутворювач включає дигідроксифеноли, такі як катехол або протокатехат, або органічні кислоти, такі як лимонна кислота. Згідно даного винаходу ферментаційне середовище для культивування мікроорганізмів містить зазвичай різноманітні фактори росту, такі як вітаміни, стимулятори росту, до яких, наприклад, належать, біотин, рибофлавін, тіамін, фолієва кислота, нікотинова кислота, пантотенат та піридоксин. Фактори росту та солі часто знаходяться в комплексних компонентах середовища, таких як дріжджовий екстракт, меляса, кукурудзяний екстракт й подібні. До культурального середовища можуть 47 додаватися потрібні компоненти. Точний склад речовин поживного середовища в значній мірі залежить від поточного експерименту й для кожного випадку підбирається окремо. Інформація щодо оптимізації середовища є доступною в підручнику "Applied MicroBiol Physiology, A Practical Approach" (Hrsg.P.M.Rhodes, P.F.Stanbury, IRL Press (1997) S.53-73, ISBN 0199635773). Ростові середовища можуть одержуватися від комерційних постачальників, таких як Standard 1 (Merck) або ВНІ (Brain heart infusion, DIFCO) й подібні. Усі без виключення компоненти середовища були стерилізовані або нагріванням (20хв. при 1,5бар та 121°С) або стерилізацією при фільтрації. Компоненти можуть стерилізуватися усі разом або за потреби окремо. Усі компоненти можуть бути присутніми в середовищі від початку росту або можуть безперервно або періодично подаватися. Температура культури зазвичай знаходиться в межах від 15°С до 45°С, переважно від 25°С до 40°С, та під час експерименту може залишатися сталою або змінюватися. Значення рН середовища повинно бути в межах від 5 до 8,5, переважно знаходитись біля 7,0. Значення рН при розведенні можна контролювати введенням основних сполук, таких як гідроксид натрію, гідроксид калію, аміак або аміачна вода, або кислими сполуками, такими як фосфорна кислота або сірчана кислота. Для керування піноутворенням можуть бути введений антиспінювач, такий як, наприклад, поліглікоестер жирної кислоти. Для підтримування стабільності плазмід до середовища можуть додаватися певні речовини із селективною дією, такі як, наприклад, антибіотики. Для підтримування аеробних умов в культуру може вноситися кисень або кисневмісні суміші, як наприклад, зовнішнє повітря. Температура культури зазвичай знаходиться в межах від 20°С до 45°С й переважно від 25°С до 40°С. Культивування продовжується до того часу, поки утворюється максимум бажаного продукту. Зазвичай цього досягають протягом від 10 до 160 годин. Ці одержані ферментаційні бульйони, які містять зокрема поліненасичені кислоти, як правило мають від 7,5 до 25мас.% сухого залишку. Потім цей ферментаційний бульйон може перероблятися. Якщо є потреба, біомаса може повністю або частково бути видалена з ферментаційного бульйону за допомогою методів сепарації, таких як центрифугування, фільтрація, декантація або комбінацією цих методів. Переважно біомаса після відділення проходить дообробку. Ферментаційний бульйон також може бути зконцентрований відомими методами без впливу на клітини, наприклад, за допомогою ротаційного випаровувального апарату, плівкового випарову 90455 48 вального апарату, випаровувального апарату із падаючою плівкою, оберненим осмосом або нанофільтрацією. Концентрований ферментаційний бульйон потім може бути перероблений для одержання жирних кислот, що містяться в ньому. Одержані в способі жирні кислоти можуть бути вихідним матеріалом для хімічного синтезу подальших продуктів. В подальшому вони можуть використовуватися наприклад, в комбінації з іншими або окремо для виробництва фармацевтичних препаратів, продуктів харчування, кормів для тварин або косметики. Наступним об'єктом даного винаходу є виділені послідовності нуклеїнової кислоти, які кодують -5-елонгазу, причому кодовані послідовностями нуклеїнової кислоти -5-елонгази, перетворюють С20-жирні кислоти із як мінімум чотирма подвійними зв'язками в молекулі; переважно, в діацилгліцеридах та/або триацилгліцеридах. Переважними виділеними послідовностями нуклеїнової кислоти є послідовності нуклеїнової кислоти, які кодують поліпептиди із -5елонгазною активністю, та в яких амінокислотна послідовність містить послідовність, вибрану з групи амінокислотних представлених послідовностей: SEQ ID NO:115, SEQ ID NO:116, SEQ ID NO:139, SEQ ID NO:140, SEQ ID NO:141 або SEQ ID NO:142. Наступними переважними виділеними послідовностями нуклеїнової кислоти ε послідовності нуклеїнової кислоти, які кодують пептиди із -5елонгазною активністю та мають комбінацію амінокислотних послідовностей вибраних із групи: a) SEQ ID NO:115 та SEQ ID NO:139, SEQ ID NO:115 та SEQ ID NO:140 або SEQ ID NO:139 та SEQ ID NO:140; або b) SEQ ID NO:116 та SEQ ID NO:141, SEQ ID NO:116 та SEQ ID NO:142 або SEQ ID NO:141 та SEQ ID NO:142; або c) SEQ ID NO:115, SEQ ID NO:139 та SEQ ID NO:140 або SEQ ID NO:116, SEQ ID NO:141 та SEQ ID NO:142. В послідовностях: SEQ ID NO:115 (NXXXHXXMYXYYX), SEQ ID NO:116 (HHXXXXWAWW), SEQ ID NO:139 (LHXXHH), SEQ ID NO:140 (TXXQXXQF), SEQ ID NO:141 (DTXFMV) та SEQ ID NO:142 (TQAQXXQF) наведеним послідовностям вставляють консервативні ділянки різних елонгаз. В таблиці 2 наведено значення амінокислот, позначених X (колонка 3) в згаданих послідовностях нуклеїнової кислоти. Також, переважні амінокислоти можуть бути взяті з різних позицій таблиці (колонка 3). В колонці 1 наведені SEQ ID NO, в колонці 2 - позиція в послідовності. 49 Особливо переважними послідовностями нуклеїнової кислоти є послідовності вибрані з групи: а) послідовність нуклеїнової кислоти, вибрана з приведених послідовностей SEQ ID NO:43, SEQ ID NO:45, SEQ ID NO:47, SEQ ID NO:49, SEQ ID NO:59, SEQ ID NO:61, SEQ ID NO:63; SEQ ID NO:65, SEQ ID NO:67, SEQ ID NO:75, SEQ ID NO:77, SEQ ID NO:79, SEQ ID NO:83, SEQ ID NO:85, SEQ ID NO:113, SEQ ID NO:131 або SEQ ID NO:133, b) послідовності нуклеїнової кислоти, які походять, в результаті виродженості генетичного коду, від наступних амінокислотних послідовностей SEQ ID NO:44, SEQ ID NO:46, SEQ ID NO:48, SEQ ID NO:50, SEQ ID NO:60, SEQ ID NO:62, SEQ ID NO:64, SEQ ID NO:66, SEQ ID NO:68, SEQ ID NO:76, SEQ ID NO:78, SEQ ID NO:80, SEQ ID NO:84, SEQ ID NO:86, SEQ ID NO:114, SEQ ID NO:132 або SEQ ID NO:134, або c) похідні від наведених послідовностей нуклеїнової кислоти: SEQ ID NO:43, SEQ ID NO:45, SEQ 90455 50 ID NO:47, SEQ ID NO:49, SEQ ID NO:59, SEQ ID NO:61, SEQ ID NO:63; SEQ ID NO:65, SEQ ID NO:67, SEQ ID NO:75, SEQ ID NO:77, SEQ ID NO:79, SEQ ID NO:83, SEQ ID NO:85, SEQ ID NO:113, SEQ ID NO:131 або SEQ ID NO:133, які кодують поліпептиди із, як мінімум 40% гомології до амінокислотних послідовностей, кодованих: SEQ ID NO:44, SEQ ID NO:46, SEQ ID NO:48, SEQ ID NO:50, SEQ ID NO:60, SEQ ID NO:62, SEQ ID NO:64, SEQ ID NO:66, SEQ ID NO:68, SEQ ID NO:76, SEQ ID NO:78, SEQ ID NO:80, SEQ ID NO:84, SEQ ID NO:85, SEQ ID NO:113, SEQ ID NO:131 або SEQ ID NO:133 та проявляють -5елонгазну активність. Наступним предметом даного винаходу є виділені послідовності нуклеїнової кислоти, які кодують поліпептиди із -6-елонгазною активністю, вибрані з групи: a) послідовність нуклеїнової кислоти з представлених послідовностей SEQ ID NO:69, SEQ ID NO:81, SEQ ID NO:111 або SEQ ID NO:183, 51 b) послідовності нуклеїнової кислоти, які можуть виводитись як результат виродженості генетичного коду із приведених амінокислотних послідовностей: SEQ ID NO:70, SEQ ID NO:82, SEQ ID NO:112 або SEQ ID NO:184 або c) похідні від приведених послідовностей нуклеїнової кислоти: SEQ ID NO69, SEQ ID NO:81, SEQ ID NO:111 або SEQ ID NO:183 які кодують поліпептиди із, як мінімум 40% гомології до амінокислотних послідовностей, кодованих: SEQ ID NO:70, SEQ ID NO:82, SEQ ID NO:112 або SEQ ID NO:184 та проявляють -6-елонгазну активність. Наступним предметом даного винаходу є виділені послідовності нуклеїнової кислоти, які кодують поліпептиди із -3-десатуразною активністю, вибрані з групи: a) послідовність нуклеїнової кислоти вибрана з групи представлених послідовностей SEQ ID NO:87 або SEQ ID NO:105, b) послідовності нуклеїнових кислот, які внаслідок виродженості генетичного коду, являють собою наведені далі амінокислотні послідовності SEQ ID NO:88 або SEQ ID NO:106, або c) похідні від приведених послідовностей нуклеїнової кислоти: SEQ ID NO:87 або SEQ ID NO:105 які кодують поліпептиди і мають, як мінімум 60% подібності до амінокислотних послідовностей SEQ ID NO:88 або SEQ ID NO:106 та мають -3-десатуразну активність. Наступним предметом даного винаходу є виділені послідовності нуклеїнової кислоти, які кодують поліпептиди із -6-десатуразною активністю, вибрані з групи: a) послідовності нуклеїнової кислоти з представленої в SEQ ID NO:89 або SEQ ID NO:97 послідовностей, b) послідовності нуклеїнової кислоти, які можуть виводитись як результат виродженості генетичного коду із приведених амінокислотних послідовностей: SEQ ID NO:90 або SEQ ID NO:98, або c) похідні від наведених послідовностей нуклеїнової кислоти: SEQ ID NO:89 або SEQ ID NO:97, які кодують поліпептиди із, як мінімум 40% гомології до амінокислотних послідовностей, кодованих: SEQ ID NO:90 або SEQ ID NO:98 та проявляють 6-десатуразну активність. Наступним предметом даного винаходу є виділені послідовності нуклеїнової кислоти, які кодують поліпептиди із -5-десатуразною активністю, вибрані з групи: а) послідовність нуклеїнової кислоти з послідовностей, представлених в SEQ ID NO:91, SEQ ID NO:93, SEQ ID NO:99 або в SEQ ID NO:101, b) послідовності нуклеїнової кислоти, які можуть виводитись як результат виродженості генетичного коду із приведених амінокислотних послідовностей: SEQ ID NO:92, SEQ ID NO:94, SEQ ID NO:100 або SEQ ID NO:102 або c) похідні від наведених послідовностей нуклеїнової кислоти: SEQ ID NO:91, SEQ ID NO:93, SEQ ID NO:99 або SEQ ID NO:101 які кодують поліпептиди із, як мінімум 40% гомології до амінокислотних послідовностей, кодованих: SEQ ID NO:92, SEQ ID NO:94, SEQ ID NO:100 або SEQ ID NO:102 та проявляють -5-десатуразну активність. 90455 52 Наступним предметом даного винаходу є виділені послідовності нуклеїнової кислоти, які кодують поліпептиди із -4-десатуразною активністю, вибрані з групи: a) послідовності нуклеїнової кислоти з SEQ ID NO:95 або SEQ ID NO:103 представленої послідовності, b) послідовності нуклеїнових кислот, які внаслідок виродженості генетичного коду можуть походити від представлених в SEQ ID NO:96 або SEQ ID NO:104 амінокислотних послідовностей, або c) похідні від наведених послідовностей нуклеїнової кислоти: SEQ ID NO:95 або SEQ ID NO:103, які кодують поліпептиди із, як мінімум 40% гомології до амінокислотних послідовностей, кодованих: SEQ ID NO:96 або SEQ ID NO:104 та проявляють -4-десатуразну активність. Наступним предметом даного винаходу є виділені послідовності нуклеїнової кислоти, які кодують поліпептиди із -12-десатуразною активністю, вибрані з групи: a) послідовність нуклеїнової кислоти вибрана з групи представлених послідовностей: SEQ ID NO:107 або SEQ ID NO:109, b) послідовності нуклеїнової кислоти, які можуть бути виведені, як результат виродженості генетичного коду із наведених амінокислотних послідовностей: SEQ ID NO:108 або SEQ ID NO:110, або с) похідні від наведених послідовностей нуклеїнової кислоти: SEQ ID NO:107 або SEQ ID NO:109, які кодують поліпептиди із, як мінімум 50% гомології до амінокислотних послідовностей, кодованих: SEQ ID NO:108 або SEQ ID NO:110 та проявляють -12-десатуразну активність. Наступним предметом винаходу є генетичні конструкти, чиї послідовності нуклеїнової кислоти знаходяться в: SEQ ID NO:43, SEQ ID NO:45, SEQ ID NO:47, SEQ ID NO:49, SEQ ID NO:59, SEQ ID NO:61, SEQ ID NO:63; SEQ ID NO:65, SEQ ID NO:67, SEQ ID NO:69, SEQ ID NO:75, SEQ ID NO:77, SEQ ID NO:79, SEQ ID NO:81, SEQ ID NO:83, SEQ ID NO:85, SEQ ID NO:87, SEQ ID NO:89, SEQ ID NO:91, SEQ ID NO:93, SEQ ID NO:95, SEQ ID NO:97, SEQ ID NO:99, SEQ ID NO:101, SEQ ID NO:103, SEQ ID NO:105, SEQ ID NO:107, SEQ ID NO:109, SEQ ID NO:111, SEQ ID NO:113, SEQ ID NO:117, SEQ ID NO:119, SEQ ID NO:131, SEQ ID NO:133, SEQ ID NO:135, SEQ ID NO:137 або SEQ ID NO:183, причому нуклеїнова кислота знаходиться у функціональному зв'язку із одним або більше регуляторними сигналами. В генетичному конструкті можуть додатково міститися гени біосинтезу жирної кислоти або обміну ліпідів, вибрані з групи ацил-СоА-дегідрогеназ(и), ацил-АСР[=асуІ carrier рrоtеіn]-десатураз(и), ацилАСР-тіоестераз(и), жирнокислотної-ацилТтрансфераз(и), ацил-СоА: лізофосфоліпідацилтрансферази, жирнокислотних(ої)-синтаз(и), жирнокислотних(ої)-гідроксилаза(и), ацетилкоензимА-карбоксилаз(и), ацил-коензимАоксидаз(и), жирнокислотних(ої)-десатураз(и), жирнокислотних(ої)-ацетиленаз(и), ліпооксигенази, триацилгліцерол-ліпази, алленоксид-синтази, гід 53 ропероксид-ліази або жирнокислотних(ої)елонгаз(и). Бажано, додаткові гени біосинтезу жирної кислоти або обміну ліпідів вибрані з групи: 4-десатураза, -5-десатураза, -6-десатураза, -8десатураза, -9-десатураза, -12-десатураза, -6елонгаза, -9-елонгаза або -3-десатураза. Переважно, використані послідовності нуклеґнової кислоти у способі згідно даного винаходу походять із еукаріотичного організму, такого як рослина, мікроорганізму або тварини. Переважно, послідовності нуклеїнової кислоти походять із порядку Salmoniformes, Xenopus або Ciona, водоростей таких як Mantoniella, Crypthecodinium, Euglena або Ostreococcus, грибів таких як рід Phytophtora або з діатомових водоростей таких як роди Thalassiosira або Phaeodactylum. Використані в способі послідовності нуклеїнових кислот, які кодують протеїни із -3десатуразною, -4-десатуразною, -5десатуразною, -6-десатуразною, -8десатуразною, -9-десатуразною, -12десатуразною, -5-елонгазною, -6-елонгазною або -9-елонгазною активністю, переносяться в експресійній касеті (=нуклеїновокислотний конструкт) окремо або бажано в комбінації, яка робить можливою експресію нуклеїнової кислоти в організмі, переважно рослині або мікроорганізмі. Може бути в нуклеїновокислотному конструкті більш ніж одна послідовність нуклеїнової кислоти із ферментативною активністю, наприклад, із -12десатуразною, -4-десатуразною, -5десатуразною-, -6-десатуразною-, -5елонгазною-, -6-елонгазною та/або -3десатуразною. Для введення використовувані нуклеїнові кислоти бажано ампліфікуються та лігуються відомим способом, який переважно проводять згідно протоколу Pfu-ДНК-полімерази або Рfu/Taq-ДНК(полімеразні суміші). Праймер вибирається залежно від ампліфікованої послідовності. За доцільності, праймер повинен вибиратися так, щоб ампліфікат охоплював усю послідовність від старт-кодону до стоп-кодону. Після ампліфікації ампліфікат аналізується відповідним способом. Наприклад, кількісний та якісний аналіз може проводитися після гель-елекрофорезного розділення. Потім ампліфікат може бути очищений згідно стандартного протоколу (z.b. Qiagen). Одна аліквота очищеного ампліфікату залишається для подальшого клонування за потреби. Придатні вектори для клонування є загально відомими для спеціалістів. Сюди ж належать зокрема вектори, які можуть бути репліковані в мікробних системах, особливо усі вектори, які забезпечують клонування в дріжджах та грибах та роблять можливою стабільну трансформацію в рослинах. Треба назвати зокрема різні бінарні та ко-інтегровані векторні системи, придатні для Т-ДНК-опосередкованої трансформації. Такі векторні системи, як правило, відрізняються тим, що потребують vir-гени для трансформації опосередкованої Agrobakterium, а також охоплюють послідовності обмежені Т-ДНК (T-DNABorder). Переважно ці векторні системи охоплюють інші cis-регуляторні регіони, такі як промотори 90455 54 та термінатори та/або селективні маркери, завдяки яким згадані трансформовані організми можуть бути ідентифіковані. Якщо при інтегрованих векторних системах vir гени та Т-ДНК-послідовності розташовані на одному векторі, бінарні системи базуються як мінімум на двох векторах, один з яких несе vir-гени, але без Т-ДНК i другого який несе ТДНК, але без vir-гену. Внаслідок цього, останні гени є відносно маленькими и легкими для маніпулювання й можуть бути репліковані як в Δ.-coli так i в Agrobacterium. До цих бінарних векторів належать вектори із серій pBIB-HYG, pPZP, pBecks, pGreen. Згідно даного винаходу переважно використовуються Віn19, рВ1101, pBinAR, pGPTV та рСАМВІА. Огляд бінарних векторів та їхнього використання наведено в Hellens та ін., Trends in Plant Science (2000) 5, 446-451. Для підготування вектору спершу вектор може бути лінеаризований рестрикційною ендонуклеазою а потім придатним способом ферментативно модифікований. Потім вектор очищують й аліквоту вставляють для клонування. При клонуванні ферментативно порізаний та за потреби очищений ампліфікат із відповідно подготованими векторними фрагментами та з вставками клонується за допомогою лігази. При цьому певний нуклеїновокислотний конструкт, відповідно векторний або плазмідний конструкт, може мати один або декілька кодуючих генних відрізків. Переважно кодуючі генні відрізки в конструкті є функціонально зв'язаними із реуляторними послідовностями. До регуляторних послідовностей, зокрема, належать рослинні послідовності, такі як вище описані промотори та термінатори. Котнструкт переважно може стабільно розмножуватися за селективних умов в мікроорганізмах, зокрема Escherichia coli та Agrobacterium tumefaciens, та робить можливим трансфер гетеролопчної ДНК в рослини або мікроорганізми. При бажаному застосуванні клонування векторів, нуклеїнові кислоти, використані у способі, винайдені нуклеїнові кислоти та нуклеїновокислотні конструкти введені в організми, такі як мікроорганізми або переважно рослини, й таким чином здійснена рослинна трансформація, інформація про яку опублікована та процитована в Plant Molecular Biology and Biotechnology (CRC Press, Boca Raton, Florida), Kapitel 6/7, S.71-119 (1993); F.F.White, Vectors for Gene Transfer in Higher Plants; in:Transgenic Plants, Bd.1, Engineering and Utilization, Hrsgb.:Kung and R.Wu, Academic Press, 1993, 15-38; В.Jenes та ін., Techniques for Gene Transfer, in:Transgenic Plants, Bd.1, Engineering and Utilization, Hrsgb.: Kung and R.Wu, Academic Press (1993), 128-143, Potrykus, Annu.Rev.Plant Physio.Plant Molec.Biol.42 (1991), 205-225)). Використані у способі нуклеїнові кислоти, винайдені нуклеїнові кислоти та нуклеїновокислотні конструкти та/або вектори можуть використовуватися для генно-технологічного змінювання широкого спектру організмів, переважно рослин, таким чином, щоб вони стали кращими та/або більш ефективними виробниками PUFAs. Є ряд механізмів, за допомогою яких є можливим змінювання згідно винаходу протеїнів -12десатурази, -5-елонгази, -6-елонгази, -5 55 десатурази, -4-десатурази, -6-десатурази та/або -3-десатурази, а також різних способів використання протеїнів таких як протеїни -12-десатурази, -9-елонгази, -6-десатурази, -8-десатурази, -6елонгази, -5-десатурази або -4-десатурази, таким чином, що це впливає на вихід, продукування та/або ефективність продукування бажаних поліненасичених жирних кислот в рослинах, особливо, в олійних рослинах або мікроорганізмах, напряму через змінені протеїни. Кількість або активність протеїнів -12-десатурази, -3-десатурази, -9елонгази, -6-десатурази, -8-десатурази, -6елонгази, -5-десатурази, -5-елонгази або -4десатурази або генів може бути підвищена, таким чином, що буде одержано більша кількість генного продукту й в результаті більша кількість сполуки, загальної формули І. Також можливий de novo синтез в організмі, в якому до введення згаданого(их) гену/генів була відсутня дана активність або здатність до біосинтезу цих сполук. Відповідно, це є справедливим для комбінації інших десатураз або елонгаз або інших ферментів із жирно кислотного та ліпідного обміну. При цьому переважним може бути також використання різних дивергентних послідовностей, тобто на рівні ДНК послідовності, або використання промоторів генетичної експресії, що уможливлює іншу тимчасову генетичної експресії, наприклад, залежно від ступеня зрілості насіння або тканин, що накопичують олію. За допомогою введення генів -12-десатурази, -3-десатурази, -9-елонгази, -6-десатурази, -8десатурази, -6-елонгази, -5-десатурази, -5елонгази та/або -4-десатурази в організм окремо або в комбінації із іншим геном в клітині може не лише підвищитися рівень біосинтезу кінцевого продукту, але й підвищуватиметься загальний вміст триацилгліцеридів або створюватиметься de novo. Так само може підвищитися кількість або активність різних генів, які необхідні для імпорту поживних речовин біосинтезу однієї або більше жирних кислот, олій, полярних та/або нейтральних ліпідів таким чином, що концентрація цих попередників, кофакторів або проміжних сполук в клітині або в накопичувальних компартментах буде підвищена, внаслідок чого здатність клітин до продукування PUFAs, як далі буде описано, збільшиться. Оптимізацією активності або збільшенням кількості однієї або більше генів -12-десатурази, -3-десатурази, -9-елонгази, -6-десатурази, -8десатурази, -6-елонгази, -5-десатурази, -5елонгази та/або -4-десатурази, які беруть участь в біосинтезі цих сполук, або порушенням активності одного або більше генів, які беруть участь в розкладі цих сполук, може бути збільшений вихід, продукування та/або ефективність продукування жирнокислотних або ліпідних молекул з організмів, переважно рослин. У способі згідно винаходу використовуються молекули нуклеїнової кислоти які кодують протеїни або їхні частини, при чому протеїни або цілий протеїнабо його частина містить амінокислотну послідовність, її достатній гомолог, вибраний з наведених далі послідовностей: SEQ ID NO:2, SEQ ID NO:4, SEQ ID NO:6, SEQ ID NO:8, SEQ ID 90455 56 NO:10, SEQ ID NO:12, SEQ ID NO:14, SEQ ID NO:16, SEQ ID NO:18, SEQ ID NO:20, SEQ ID NO:22, SEQ ID NO:24, SET NID NO:26, SEQ ID NO:28, SEQ ID NO:30, SEQ ID NO:32, SEQ ID NO:34, SEQ ID NO:36, SEQ ID NO:38, SEQ ID NO:40, SEQ ID NO:42, SEQ ID NO:44, SEQ ID NO:46, SEQ ID NO:48, SEQ ID NO:50, SEQ ID NO:52, SEQ ID NO:54, SEQ ID NO:60, SEQ ID NO:62, SEQ ID NO:64, SEQ ID NO:66, SEQ ID NO:68, SEQ ID NO:70, SEQ ID NO:72, SEQ ID NO:74, SEQ ID NO:76, SEQ ID NO:78, SEQ ID NO:80, SEQ ID NO:82, SEQ ID NO:84, SEQ ID NO:86, SEQ ID NO:88, SEQ ID NO:90, SEQ ID NO:92, SEQ ID NO:94 SEQ ID NO:96 SEQ ID NO:98, SEQ ID NO:100, SEQ ID NO:102, SEQ ID NO:104, SEQ ID NO:106, SEQ ID NO:108, SEQ ID NO:110, SEQ ID NO:112, SEQ ID NO:114, SEQ ID NO:118, SEQ ID NO:120, SEQ ID NO:132, SEQ ID NO:134, SEQ ID NO:136, SEQ ID NO:138 або SEQ ID NO:184 таким чином, що протеїни проявляють активність -12-десатурази, -3-десатурази, -9елонгази, -6-десатурази, -8-десатурази, -6елонгази, -5-десатурази, -5-елонгази та/або -4десатурази. Переважно, протеїн або його частини, яка(ий.і) кодується молекулою(ами) нуклеїнової кислоти проявляють основну ферментативну активність та здатність брати участь в обміні речовин спголук, необхідних для побудови клітинних мембран або ліпдних тілець в організмі, переважно в рослинах, або в транспортуванні молекул через ці мембрани. Переважно, молекула нуклеїнової кислоти, що кодує протеїни як мінімум приблизно на 50%, переважно як мінімум приблизно на 60% та більш переважно як мінімум приблизно на 70%, 80% або 90% та найбільш переважно як мінімум приблизно на 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% або більше, подібна до наведених амінокислотних послідовностей: SEQ ID NO:2, SEQ ID NO:4, SEQ ID NO:6, SEQ ID NO:8, SEQ ID NO:10, SEQ ID NO:12, SEQ ID NO:14, SEQ ID NO:16, SEQ ID NO:18, SEQ ID NO:20, SEQ ID NO:22, SEQ ID NO:24, SEQ ID NO:26, SEQ ID NO:28, SEQ ID NO:30, SEQ ID NO:32, SEQ ID NO:34, SEQ ID NO:36, SEQ ID NO:38, SEQ ID NO:40, SEQ ID NO:42, SEQ ID NO:44, SEQ ID NO:46, SEQ ID NO:48, SEQ ID NO:50, SEQ ID NO:52, SEQ ID NO:54, SEQ ID NO:60, SEQ ID NO:62, SEQ ID NO:64, SEQ ID NO:66, SEQ ID NO:68, SEQ ID NO:70, SEQ ID NO:72, SEQ ID NO:74, SEQ ID NO:76, SEQ ID NO:78, SEQ ID NO:80, SEQ ID NO:82, SEQ ID NO:84, SEQ ID NO:86, SEQ ID NO:88, SEQ ID NO:90, SEQ ID NO:92, SEQ ID NO:94, SEQ ID NO:96, SEQ ID NO:98, SEQ ID NO:100, SEQ ID NO:102, SEQ ID NO:104, SEQ ID NO:106, SEQ ID NO:108, SEQ ID NO:110, SEQ ID NO:112, SEQ ID NO:114, SEQ ID NO:118, SEQ ID NO:120, SEQ ID NO:132, SEQ ID NO:134, SEQ ID NO:136, SEQ ID NO:138 або SEQ ID NO:184. У контексті даного винаходу під гомологією або гомологічний розуміють ідентичність або ідентичний. Гомологія розраховується по загальній амінокислотній або виведеній нуклеїновокислотній послідовності. Для порівнювання різних послідовностей спеціалісти можуть використовувати ряд 57 програм, які базуються на різних алгоритмах. При цьому, особливо достовірні результати дають алгоритми Needleman та Wunsch або Smith та Waterman. Для порівняння послідовностей використовувалася програма PileUp (J.Mol. Evolution., 25, 351-360, 1987, Higgins та ін., CABIOS, 5 1989:151153) або програми Gap та BestFit [Needleman and Wunsch (J.Mol. Biol. 48; 443-453 (1970) та Smith and Waterman (Adv.Appl.Math.2; 482-489 (1981)], які містилися в пакеті програм GCG [Genetics Computer Group, 575 Science Drive, Madison, Wisconsin, USA 53711 (1991)]. Вищезазначені проценти гомології послідовностей були одержані в програмі GAP при загальній послідовності та наступних установлених значеннях: Gap Weight:50, Length Weight:3, Average Match:10.000 та Average Mismatch:0.000. Ці установки використовувалися замість стандартних параметрів для порівняння послідовностей, крім випадків коли інше зазначалося. Під суттєвою ферментативною активністю, використаних у способі згідно винаходу -12десатураз, -3-десатураз, -9-елонгаз, -6десатураз, -8-десатураз, -6-елонгаз, -5десатураз, -5-елонгаз та/або -4-десатураз, розуміють, що вони порівняно із послідовностями: SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5, SEQ ID NO:7, SEQ ID NO:9, SEQ ID NO:11, SEQ ID NO:13, SEQ ID NO:15, SEQ ID NO:17, SEQ ID NO:19, SEQ ID NO:21 SEQ ID NO:23, SEQ ID NO:25, SEQ ID NO:27, SEQ ID NO:29, SEQ ID NO:31, SEQ ID NO:33, SEQ ID NO:35, SEQ ID NO:37, SEQ ID NO:39, SEQ ID NO:41, SEQ ID NO:43, SEQ ID NO:45, SEQ ID NO:47, SEQ ID NO:49, SEQ ID NO:51, SEQ ID NO:53, SEQ ID NO:59, SEQ ID NO:61, SEQ ID NO:63, SEQ ID NO:65, SEQ ID NO:67, SEQ ID NO:69, SEQ ID NO:71, SEQ ID NO:73, SEQ ID NO:75, SEQ ID NO:77, SEQ ID NO:79, SEQ ID NO:81, SEQ ID NO:83, SEQ ID NO:85, SEQ ID NO:87, SEQ ID NO:89, SEQ ID NO:91, SEQ ID NO:93, SEQ ID NO:95, SEQ ID NO:97, SEQ ID NO:99, SEQ ID NO:101, SEQ ID NO:103, SEQ ID NO:105, SEQ ID NO:107, SEQ ID NO:109, SEQ ID NO:111, SEQ ID NO:113, SEQ ID NO:117, SEQ ID NO:119, SEQ ID NO:131, SEQ ID NO:133, SEQ ID NO:135, SEQ ID NO:137 або SEQ ID NO:183 та їхніми похідними, що кодують протеїни/ферменти мають ферментативну активність щонайменше 10%, переважно 20%, особливо переважно 30% та в особливо переважно 40% i таким чином можуть брати участь в обміні речовин сполук, необхідних для побудови жирних кислот, естерів жирних кислот та диацилгліцеридів та/або триацилгліцеридів в організмі, переважно рослині або рослинній клітині, або в транспортуванні молекул через мембрани, причому маються на увазі С18-, С20- або С22-вуглецеві ланцюги у молекулі жирної кислоти із подвійними зв'язками, як мінімум в двох, трьох, чотирьох, п'ятьох або шістьох положеннях. Переважно придатні нуклеїнові кислоти походять із бактерій, грибів, діатомових водоростей, тварин, таких як Caenorhabditis або Oncorhynchus, або рослин, таких як водорості або мохи, такі як роди Shewanella, Physcomitrella, Thraustochytrium, 90455 58 Fusarium, Phytophthora, Ceratodon, Mantoniella, Ostreococcus, Isochrysis, Aleunta, Muscarioides, Mortierella, Borago, Phaeodactylum, Crypthecodinium, особливо із видів та родів Oncorhynchus mykiss, Xenopus laevis, Ciona intestinalis, Thalassiosira pseudonona, Mantoniella squamata, Ostreococcus sp., Ostreococcus tauri, Euglena gracilis, Physcomitrella patens, Phytophtora infestans, Fusarium graminaeum, Cryptocodinium cohnii, Ceratodon purpureus, Isochrysis galbana, Aleunta fannosa, Thraustochytrium sp., Muscarioides vialii, Mortierella alpina, Borago officinahs, Phaeodactylum tricornutum, Caenorhabditis elegans або особливо переважно із Oncorhynchus mykiss, Euglena gracilis, Thalassiosira pseudonana або Crypthecodinium cohnii. Також у способі згідно даного винаходу можуть бути використані послідовності нуклеїнової кислоти, які кодують -12-десатуразу, -3десатуразу, -9-елонгазу, -6-десатуразу, -8десатуразу, -6-елонгазу, -5-десатуразу, -5елонгазу та/або -4-десатуразу, які наведені в послідовностях: SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5, SEQ ID NO:7, SEQ ID NO:9, SEQ ID NO:11, SEQ ID NO:13, SEQ ID NO:15, SEQ ID NO:17 SEQ ID NO:19, SEQ ID NO:21, SEQ ID NO:23, SEQ-ID NO:25, SEQ ID NO:27, SEQ ID NO:29, SEQ ID NO:31, SEQ ID NO:33, SEQ ID NO:35, SEQ ID NO:37, SEQ ID NO:39, SEQ ID NO:41, SEQ ID NO:43, SEQ ID NO:45, SEQ ID NO:47, SEQ ID NO:49, SEQ ID NO:51, SEQ ID NO:53, SEQ ID NO:59, SEQ ID NO:61, SEQ ID NO:63, SEQ ID NO:65, SEQ ID NO:67, SEQ ID NO:69, SEQ ID NO:71, SEQ ID NO:73, SEQ ID NO:75, SEQ ID NO:77, SEQ ID NO:79, SEQ ID NO:81, SEQ ID NO:83, SEQ ID NO:85, SEQ ID NO:87, SEQ ID NO:89, SEQ ID NO:91, SEQ ID NO:93, SEQ ID NO:95, SEQ ID NO:97, SEQ ID NO:99, SEQ ID NO:101, SEQ ID NO:103, SEQ ID NO:105, SEQ ID NO:107, SEQ ID NO:109, SEQ ID NO:111, SEQ ID NO:113, SEQ ID NO:117, SEQ ID NO:119, SEQ ID NO:131, SEQ ID NO:133, SEQ ID NO:135, SEQ ID NO:137 або SEQ ID NO:183, та за певних умов гібридизовані в нуклеотидну послідовність. Використані в способі послідовності нуклеїнових кислот, переважно в експресійній касеті, яка робить можливою експресію нуклеїнові кислоти в організмах таких як мікроорганізми або рослини. Таким чином, послідовності нуклеїнових кислот, які кодують -12-десатуразу, -3-десатуразу, -9-елонгазу, -6-десатуразу, -8-десатуразу, -6елонгазу, -5-десатуразу, -5-елонгазу та/або -4десатуразу, функціонально пов'язані з одним або кількома регуляторними сигналами, переважно для посилення генної експресії. Ці регуляторні послідовності роблять можливою спрямовану експресію генів та протеїнову експресію. Це може означати, що наприклад, залежно від організму хазяїна, лише після індукції ген стане експресуватися та/або доекспресуватися, або одразу ж стане експресуватися та/або доекспресуватися. Наприклад, йдеться про ці регуляторні послідовності, тобто послідовності, які зв'язані із індукторами або репресорами, й таким чином регулюють експресію нуклеїнової кислоти. Додатково до цих нових регу 59 ляторних послідовностей або замість цих послідовностей може відбуватися природне регулювання цих послідовностей, які знаходяться перед природними структурними генами та за потреби можуть бути генетично змінені таким чином, щоб відключалася природна регуляція й підвищувалася експресія генів. Експресійна касета (=експресійний конструкт=генний конструкт) може бути сформована більш просто, що означає, що не були вставлені додаткові регуляторні сигнали перед послідовністю нуклеїнової кислоти або її похідним, та природний промотор із своєю регуляцією не видалявся. Замість цього природна регуляторна послідовність настільки змінювалася, що не відбувалася регуляція та/або збільшувалася генна експресія. Ці змінені промотори можуть бути у формі часткової послідовності (=промотор із частками нуклеїнової послідовності згідно винаходу) введені також перед природним геном для підвищення активності. Генний конструкт, крім того, переважно може містити один або більше так званих "енхансерних послідовностей", функціонально зв'язаних із промотором, що робить можливим підвищення експресії послідовності нуклеїнової кислоти. Також до 3'-кінця послідовності ДНК можуть додатково бути приєднані послідовності, такі як різноманітні регуляторні елементи або термінатори. Гени -12-десатурази, -3-десатурази, -9елонгази, -6-десатурази, -8-десатурази, -6елонгази, -5-десатурази, -5-елонгази та/або -4десатурази можуть бути вставлені в одній експресійній касеті (=генний конструкт) в одній або більше копіях. Переважно в експресійній касеті знаходиться лише одна копія генів. Цей генний конструкт або генні конструкти можуть бути сукупно експресовані в організмі хазяїна. При цьому генний конструкт або генні конструкти можуть бути вставлені в один або декілька векторів й вільно існувати в клітині, або бути вставленими в геном. Це є бажаним для вставляння інших генів в геном хазяїна, якщо гени, що експресуються, лежать в одному генетичному конструкті. Регуляторні послідовності або фактори, можуть при цьому, як описано вище, переважно позитивно впливати на експресію введених генів і таким чином м підвищувати. Таким чином, може відбуватися підсилююча дія регуляторних елементів на транскрипційну матрицю, шляхом використання сильних транскрипційних сигналів, таких як промотори та/або енхансери. Однак, поряд із цим можливе підсилення трансляції тим, що наприклад, покращується стабільність мРНК. Наступна форма здійснення винаходу полягає в одному або більше генних конструктах, в яких міститься одна або більше послідовностей з наведених далі: SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5, SEQ ID NO:7, SEQ ID NO:9, SEQ ID NO:11, SEQ ID NO:13, SEQ ID NO:15, SEQ ID NO:17, SEQ ID NO:19, SEQ ID NO:21, SEQ ID NO:23, SEQ ID NO:25, SEQ ID NO:27, SEQ ID NO:29, SEQ ID NO:31, SEQ ID NO:33, SEQ ID NO:35, SEQ ID NO:37, SEQ ID NO:39, SEQ ID NO:41, SEQ ID NO:43, SEQ ID NO:45, SEQ ID NO:47, SEQ ID NO:49, SEQ ID NO:51, SEQ ID NO:53, SEQ ID NO:59, SEQ ID NO:61, SEQ ID NO:63, SEQ ID 90455 60 NO:65, SEQ ID NO:67, SEQ ID NO:69, SEQ ID NO:71, SEQ ID NO:73, SEQ ID NO:75, SEQ ID NO:77, SEQ ID NO:79, SEQ ID NO:81, SEQ ID NO:83, SEQ ID NO:85, SEQ ID NO:87, SEQ ID NO:89, SEQ ID NO:91, SEQ ID NO:93, SEQ ID NO:95, SEQ ID NO:97, SEQ ID NO:99, SEQ ID NO:101, SEQ ID NO:103, SEQ ID NO:105, SEQ ID NO:107, SEQ ID NO:109, SEQ ID NO:111, SEQ ID NO:113, SEQ ID NO:117, SEQ ID NO:119, SEQ ID NO:131, SEQ ID NO:133, SEQ ID NO:135, SEQ ID NO:137 або SEQ ID NO:183 або їхні похідні, які є визначеними як поліпептиди, що кодуються згідно: SEQ ID NO:2, SEQ ID NO:4, SEQ ID NO:6, SEQ ID NO:8, SEQ ID NO:10, SEQ ID NO:12, SEQ ID NO:14, SEQ ID NO:16, SEQ ID NO:18, SEQ ID NO:20, SEQ ID NO:22, SEQ ID NO:24, SEQ ID NO:26, SEQ ID NO:28, SEQ ID NO:30, SEQ ID NO:32, SEQ ID NO:34, SEQ ID NO:36, SEQ ID NO:38, SEQ ID NO:40, SEQ ID NO:42, SEQ ID NO:44, SEQ ID NO:46, SEQ ID NO:48, SEQ ID NO:50, SEQ ID NO:52, SEQ ID NO:54, SEQ ID NO:60, SEQ ID NO:62, SEQ ID NO:64, SEQ ID NO:66, SEQ ID NO:68, SEQ ID NO:70, SEQ ID NO:72, SEQ ID NO:74, SEQ ID NO:76, SEQ ID NO:78, SEQ ID NO:80, SEQ ID NO:82, SEQ ID NO:84, SEQ ID NO:86, SEQ ID NO:88, SEQ ID NO:90, SEQ ID NO:92, SEQ ID NO:94, SEQ ID NO:96, SEQ ID NO:98, SEQ ID NO:100, SEQ ID NO:102, SEQ ID NO:104, SEQ ID NO:106, SEQ ID NO:108, SEQ ID NO:110, SEQ ID NO:112, SEQ ID NO:114, SEQ ID NO:118, SEQ ID NO:120, SEQ ID NO:132, SEQ ID NO:134, SEQ ID NO:136, SEQ ID NO:138 або SEQ ID NO:184. Згадані протеїни -12десатурази, -3-десатурази, -9-елонгази, -6деструктази, -8-десатурази, -6-елонгази, -5десатурази, -5-елонгази та/або -4-десатурази, ведуть переважно десатурацію або елонгацію жирних кислот, причому субстратом виступає молекула жирної кислоти із однією, двома, трьома, чотирма, п'ятьмя або шістьма подвійними зв'язками та переважно із 18, 20 або 22 атомами карбону. Це саме стосується їхніх гомологів, похідних та аналогів, які функціонально зв'язані з одним або більше регуляторними сигналами, переважно для підвищення генної експресії. Переважними регуляторними послідовностями для нового способу є наприклад, промотори, такі як: cos-, tac-, trp-, tet-, trp-tet-, Ipp-, lac-, Ipp-lac-, laclq-, T7-, T5-, T3-, gal-, trc-, ara-, SP6-, -PR- або -PL-промотор, що переважно застосовуються в Грам-негативних бактеріях. Наступними переважними регуляторними послідовностями є, наприклад, Грам-позитивні промотори аmу та SP02, та дріжджові або грибкові промотори ADC1, MFa, АС, Р-60, CYC1, GAPDH, TEF, rp28, ADH або рослинні промотори CaMV/35S [Franck та ін., Cell 21 (1980) 285294], PRP1 [Ward та ін., Plant.Mol. ВioІ 22 (1993)], SSU, OCS, Iib4, usp, STLS1, В33, nos або Ubiquitin або Phaseolin промотор. В цьому відношенні також є бажаними індуковані промотори, такі як промотори, що описані в ЕР-А-0388186 (індукований бензилсульфонамідом), Plant J.2,1992 397-404 (Gatz та ін., індукований тетрацикліном), ЕР-А-0335528 (індукований амбіциновою кислотою) або WO93/21334 (індукований етанолом або
ДивитисяДодаткова інформація
Назва патенту англійськоюProcess for preparation of polyunsaturated fatty acids in transgenic organisms
Автори англійськоюZANK THORSTEN, BAUER JOERG, CIRPUS PETRA, ABBADI AMINE, HEINZ ERNST, QIU XIAO, VRINTEN PATRICIA, SPERLING PETRA, DOMERGUE FREDERIC, MEYER ASTRID, KIRSCH JELENA
Назва патенту російськоюСпособ получения полиненасыщенных жирных кислот в трансгенных организмах
Автори російськоюЦанк Торстен, Бауер Йорг, Цирпус Петра, Аббади Аминэ, Хайнц Эрнст, Квиу Ксяо, Вринтен Патрисия, Шперлинг Петра, Домерг Фредерик, Мейер Астрид, Кирш Елена
МПК / Мітки
МПК: C12N 9/02, C12N 9/10, C12N 15/82
Мітки: трансгенних, жирних, одержання, кислот, поліненасичених, спосіб, організмах
Код посилання
<a href="https://ua.patents.su/309-90455-sposib-oderzhannya-polinenasichenikh-zhirnikh-kislot-v-transgennikh-organizmakh.html" target="_blank" rel="follow" title="База патентів України">Спосіб одержання поліненасичених жирних кислот в трансгенних організмах</a>
Застосування довголанцюжкових поліненасичених жирних кислот для одержання рідкої поживної суміші для годування дітей

Номер патенту: 89207
Опубліковано: 11.01.2010
Автори: Берманн Крістофер, Бем Гюнтер, Шталь Бернд, Гарссен Йохан
МПК: A61K 31/202, A61P 43/00, A23L 1/29
Мітки: дітей, поліненасичених, довголанцюжкових, жирних, кислот, суміші, одержання, поживної, годування, застосування, рідкої
Формула / Реферат:
1. Застосування довголанцюжкових поліненасичених жирних кислот для одержання рідкої поживної суміші для годування дітей, матері яких страждали під час вагітності на діабет І типу, діабет II типу, ожиріння або надлишкову вагу, причому вказана суміш містить n-3 фракцію поліненасичених жирних кислот, що містить щонайменше 0,1 ваг. % докозагексаєнової кислоти (DHA) від повної ваги ліпіду, щонайменше 0,01 ваг. % n-3 докозапентаєнової кислоти...
Спосіб оцінки порушень метаболізму поліненасичених жирних кислот ліпідів ліквору
Номер патенту: 22884
Опубліковано: 05.05.1998
Автори: Цимбалюк Віталій Іванович, Пічкур Ліонид Дмитрович, Яценко Валентин Порфір'євич, Брюзгіна Тетяна Семенівна
МПК: G01N 33/53, C07C 57/00, G01N 30/68
Мітки: поліненасичених, ліпідів, порушень, оцінки, спосіб, метаболізму, ліквору, кислот, жирних
Формула / Реферат:
Способ оценки нарушений метаболизма полиненасыщенных жирных кислот липидов ликвора путем газохроматографического анализа, отличающийся тем, что определяют содержание арахидоновой и линолевой кислот липидов ликвора и по коэффициенту соотношения между ними устанавливают нарушение метаболизма полиненасыщенных жирных кислот.
Спосіб отримання гідропероксидів поліненасичених жирних кислот

Номер патенту: 13716
Опубліковано: 25.04.1997
Автори: Кухар Валерій Павлович, Бутович Ігор Анатолійович, Бридня Віта Петрівна, Цись Олена Валентинівна
МПК: C12P 7/64, C12N 11/00, G01N 1/28
Мітки: поліненасичених, гідропероксидів, спосіб, кислот, жирних, отримання
Формула / Реферат:
Способ получения гидропероксидов полиненасыщенных жирных кислот, включающий окисление полиненасыщенных жирных кислот в присутствии лйпоксигеназы в качестве катализатора и последующее выделение целевого продукта, отличающийся тем, что, с целью увеличения выхода целевого продукта и упрощения способа, окисление проводят в непрерывном режиме путем пропускания водного раствора полинепасыщсн-ных жирных кислот с концентрацией 0,01-0,25 мМ/л...
Спосіб одержання жирних кислот з відходів масложирової промисловості

Номер патенту: 30047
Опубліковано: 11.02.2008
Автор: Булдов Юрій Юрійович
МПК: C11B 13/00
Мітки: жирних, відходів, кислот, масложирової, одержання, спосіб, промисловості
Формула / Реферат:
Спосіб одержання жирних кислот з відходів масложирової промисловості, що включає омилення лугом масложирових компонентів відходів, нейтралізацію отриманої суміші кислотою з наступним виділенням жирних кислот із суміші, який відрізняється тим, що як відходи масложирової промисловості використовують відпрацьовані фільтрувальні порошки на основі перлітів або кізельгурів.
Спосіб одержання моноетаноламідів синтетичних жирних кислот

Номер патенту: 21191
Опубліковано: 04.11.1997
Автори: Рудак Валерій Борисович, Мулява Мар'ян Петрович, Степанов Михайло Васильович, Степанов Василь Михайлович
МПК: C07C 231/00
Мітки: спосіб, жирних, синтетичних, моноетаноламідів, одержання, кислот
Формула / Реферат:
Спосіб одержання моноетаноламідів синтетичних жирних кислот амідуванням їх моноетаноламіном, який відрізняється тим, що моноетаноламін беруть з надлишком від 20 до 50% і процес проводять в присутності ксилолу при температурі 160-180°С і атмосферному або пониженому тиску з одночасною азеотропною відгонкою утворюваної води в складі гетероазеотропу з ксилолом і наступним видаленням спочатку непрореагованого етаноламіну азеотропною відгонкою з...
Попередній патент: Захищена реєстрація для широкомовної багатоадресної мультимедійної системи (mbms)
Наступний патент: Розширений канал підтвердження і керування швидкістю
Випадковий патент: Спосіб прогнозування зрощення перелому