Спосіб одержання флавінмононуклеотиду (5′-фмн)
Номер патенту: 22568
Опубліковано: 25.04.2007
Автори: Яцишин Валентина Юріївна, Вороновський Андрій Ярославович, Сибірний Андрій Андрійович, Федорович Дарія Василівна, Дмитрук Костянтин Васильович, Іщук Олена Петрівна

Формула / Реферат
Спосіб одержання флавінмононуклеотиду (5'-ФМН), що полягає у введенні в геном мікроорганізмів генетичної інформації про синтез флавінмононуклеотиду, який відрізняється тим, що промотор гена FMN1 (кодує рибофлавінкіназу) дріжджів Debaryomyces hansenii вирізають, замінюють на сильний промотор фактора елонгації трансляції - TEF1 дріжджів Candida famata, конструюють вектори для надекспресії гена FMN1, за допомогою яких в геном Candida famata вводять фрагмент ДНК, що несе генетичну інформацію про синтез 5'-ФМН, отримують рекомбінантні штами з високою активністю рибофлавінкінази, в культуральній рідині яких нагромаджується 5'-ФМН.
Текст
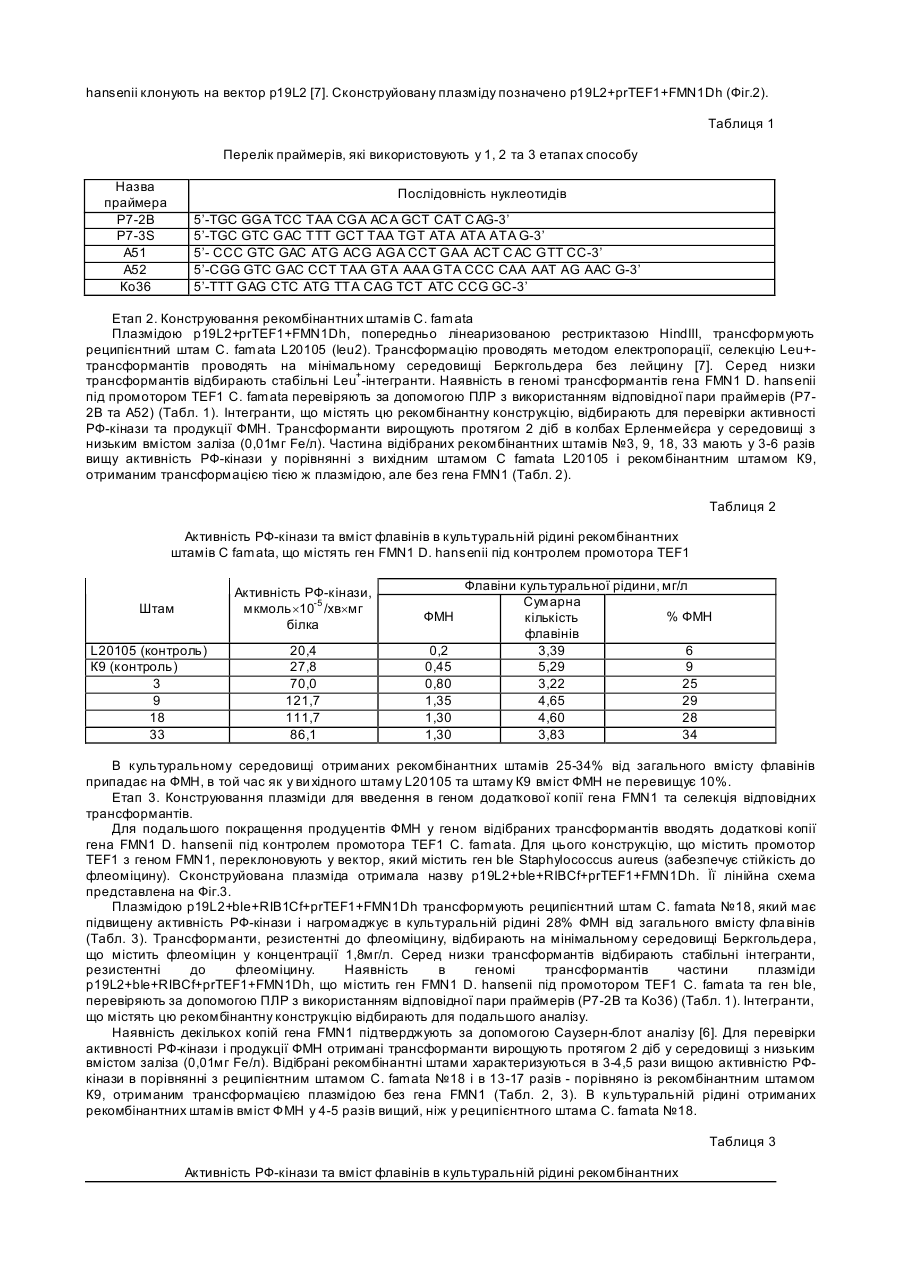
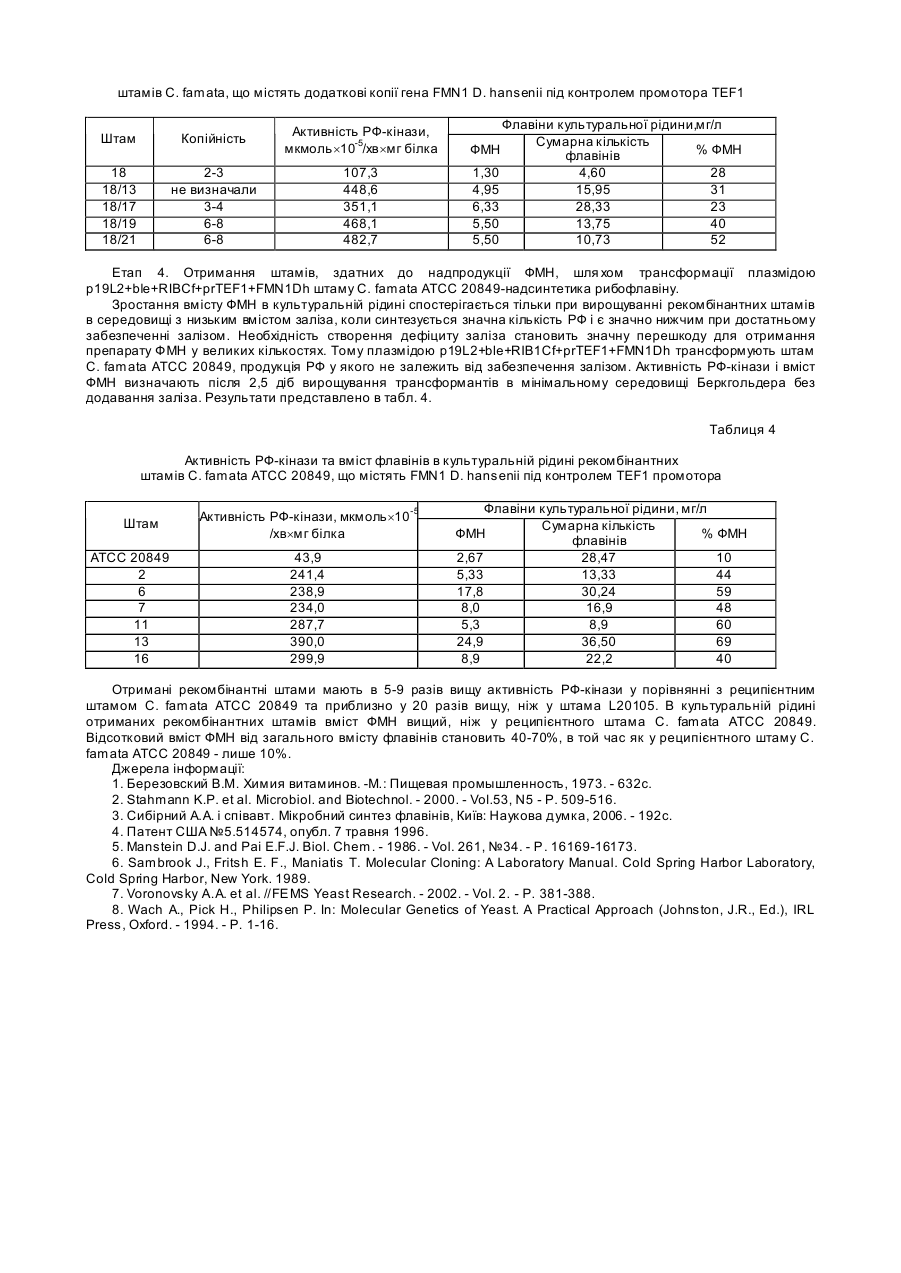
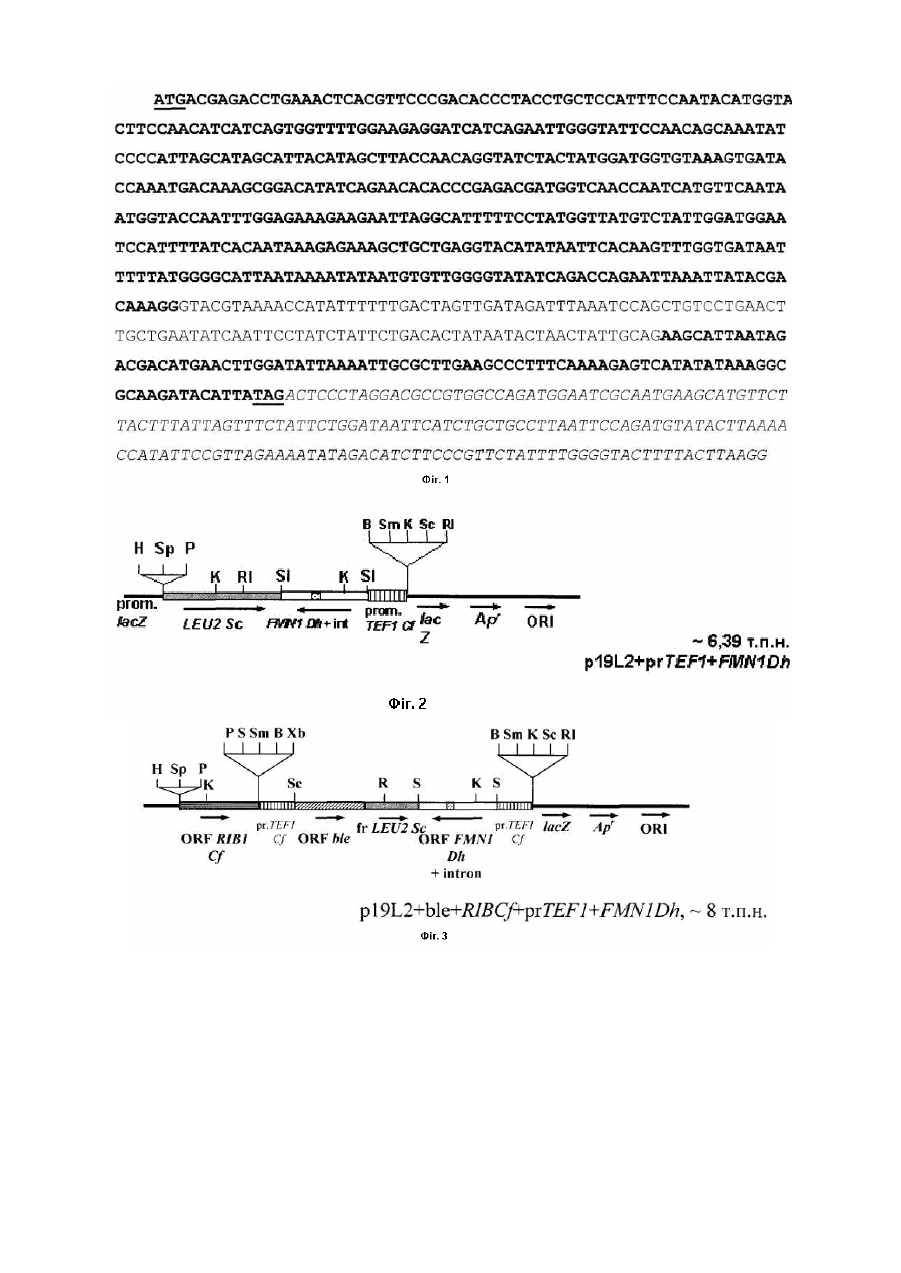
Корисна модель відноситься до галузі біотехнології і є способом отримання флавінмононуклеотиду (ФМН) за допомогою рекомбінантних штамів дріжджів Candida famata (C. famata). 5’-ФМН (рибофлавін-5’-фосфат) є сполукою, яка відіграє важливу роль як коензим у різноманітних ферментативних реакціях у живих організмах і є ефективним засобом профілактики і лікування захворювань, пов’язаних з порушенням біосинтезу цієї сполуки. Препарати ФМН, отримані ферментативним способом, можуть знайти застосування як: 1 - лікарські препарати, які не містять домішок з невстановленою біологічною активністю; 2 - реактиви високого ступеня чистоти; 3 - проміжні продукти для отримання флавінаденіндинуклеотиду (ФАД) та його похідних; 4 - компоненти системи біолюмінісцентного аналізу. Відомо, що 5’-ФМН у промислових масштабах отримують шляхом фосфорилювання рибофлавіну (РФ) хімічними агентами [1], що приводить до утворення препарату, який містить побічні продукти: ізомерні форми флавінмононуклеотиду, такі як 2’-ФМН і 4’-ФМН, рибофлавін динуклеотиди (5’,4’-ФДН і 5’,3’-ФДН), а також поліфосфати, які не мають вітамінної активності. Очищення 5’-ФМН від цих сполук є складним завданням. Комерційні препарати містять не більше 74% натрієвої солі 5’-ФМН, крім того, препарати ФМН, отримані таким способом, є дуже дорогими, що знижує можливості їх широкого використання у практиці. Відомо досить багато мікроорганізмів, здатних до надсинтезу РФ, зокрема, рекомбінантні штами бактерій Bacillus subtilis, цвільовий гриб Ashbya gossypii та флавіногенні дріжджі С. famata [2, 3]. Однак не описано жодного прикладу прямого мікробного синтезу значних кількостей ФМН. Найбільш близьким до запропонованого є спосіб отримання ФМН, в якому використані бактерії родів Corynebacterium, Brevibacterium, Escherichia, Enterobacter i Pseudomonas, що містять рекомбінантну ДНК з фрагментом, який несе генетичну інформацію про синтез ФМН [4]. Однак у цих мікроорганізмів перетворення РФ до ФМН і наступну реакцію, фофорилювання ФМН до ФАД, здійснює той самий білок [5], тому в к ультура х нагромаджуються обидва нуклеотиди - ФМН і ФАД. В основу корисної моделі поставлено завдання за допомогою генноінженерних підходів сконструювати рекомбінантні штами дріжджів з високою активністю рибофлавінкінази (РФ-кіназа), здатні до надсинтезу ФМН. Поставлене завдання досягається тим, що у способі отримання флавінмононуклеотиду (5’-ФМН), який полягає у введенні в геном мікроорганізмів генетичної інформації про синтез ФМН, згідно з корисною моделлю промотор гена FMN1 (кодує РФ-кіназу) дріжджів Debaryomyces hansenii (D. hansenii) вирізають, замінюють на сильний промотор фактора елонгації трансляції - TEF1 дріжджів С famata, конструюють вектори для надекспресії гена FMN1, за допомогою яких в геном С. famata вводять фрагмент ДНК, що несе генетичну інформацію про синтез 5’-ФМН, отримують рекомбінантні штами з високою активністю РФ-кінази, в культуральній рідині яких нагромаджується 5’-ФМН. Суть корисної моделі полягає у введенні в геном дріжджів фрагменту ДНК, який несе генетичну інформацію про синтез ФМН під контролем сильного конститутивного промотора, що забезпечує високий рівень експресії гена FMN1 і нагромадження цільового продукту - ФМН, а не суміші флавінових н уклеотидів, ФМН і ФАД, як це має місце при використанні бактерійної моделі [5]. Біосинтез ФМН і ФАД у дріжджів С famata здійснюється за допомогою послідовної каталітичної дії 2 різних ферментів - РФ-кінази (КФ 2.7.1.26) та ФАД-синтетази (КФ 2.7.27.2), які забезпечують утворення ФМН і ФАД у незначній кількості, необхідній для життєдіяльності клітини. [3]. У запропонованому способі використовують такі методи: полімеразна ланцюгова реакція (ПЛР), конструювання рекомбінантних плазмід, виділення плазмід з Escherichia coli, рестрикційний аналіз, електрофорез в агарозному гелі, трансформація Е. coli методом електропорації, описані в [6]. Трансформацію С. famata проводять, як описано в [7]. Виділення сумарної ДНК з транс формантів С. famata проводиться як для S. cerevisiae [8]. Спосіб отримання ФМН ілюструється графічними матеріалами. На Фіг.1 зображено нуклеотидну послідовність відкритої рамки зчитування та термінатора гена РФ-кінази FMN1 дріжджів D. hansenii, де інтрон вказаний нежирним шрифтом, старт- та стоп-кодони підкреслені, термінатор позначено курсивом нежирним шрифтом. Послідовність взято з банку даних [„Genolevures: Debaryomyces Hansen”, http://cbi.labri.fr/Genolevures/elt/DEHA]. На Фіг.2. зображено лінійну схему плазміди p19L2+prTEF1+FMN1Dh, де фрагмент, що містить ген LEU2 S. cerevisiae, позначено сірою смугою; фрагмент, що містить промотор TEF1 С. famata, позначено вертикальними штри хами; фрагмент, що містить відкриту рамку зчитування FMN1 D. hansenii, позначено білою смугою; інтрон FMN1 D. hansenii, позначено крапками; тонкою чорною лінією позначено бактерійну частину вектора. Сайти рестрикції позначено: Н - HindІІІ; Sp - SphІ; P - PstІ; К - KpnІ; Rl -ЕсоІІІ; SI - SalІ; Xb - XbaІ; В - ВаmІІІ; Sm - SmaІ; Sc - SacI. На Фіг.3 зображено лінійну схему плазміни p19L2+ble+RIBCf+prTEF1+FMN1Dh, де фрагмент, що містить ген RIВ1 С. famata, позначено горизонтальними смугами; фрагмент, що містить ген blе S. aureus позначено діагональним штрихом; фрагмент гена LEU2 S. cerevisiae, позначено сірою смугою; фрагмент, що містить промотор TEF1 С. famata, позначено вертикальними штрихами; фрагмент, що містить відкриту рамку зчитування FMN1 D. hansenii, позначено білою смугою; інтрон FMN1 D. hansenii, позначено крапками; тонкою чорною лінією позначено бактерійну частину вектора. Сайти рестрикції позначено: Н - HindIII; Sp - SphI; P - PstI; К - КрnI, S - SalI; Sm - SmaI; Xb - XbaI; Sc - SacI; RI - EcoRI; В - BamHI. Спосіб здійснюють кількома етапами. Етап 1. Конструювання плазміди, що містить ген FMN1 Debaryomyces hansenii під промотором TEF1. Відкриту рамку зчитування (ВРЗ), що містить інтрон, а також термінатор гена FMN1 D. hansenii (Фіг.1) ампліфікують за допомогою ПЛР з використанням геномної ДНК D. hansenii як матриці та праймерів А51 і А52 (Табл. 1). Фрагмент очікуваної величини (824 пари нуклеотидів, (п.н.)) обробляють рестриктазою SalI (сайти цього фермента фланкують одержаний продукт ПЛР). Фрагмент, що містить промотор TEF1 С famata, ампліфікують за допомогою ПЛР з використанням праймерів Р7-2В і P7-3S (Табл. 1) та геномної ДНК С. famata як матриці. Продукт очікуваної величини (658п.н.) обробляють рестриктазами ВатНІ та SalI, сайти яких фланкують цей продукт. BаmНІ/SаlI-фрагмент з промотором TEF1 С. famata та SalI-фрагмент з ВРЗ і термінатором FMN1 D. hansenii клонують на вектор p19L2 [7]. Сконструйовану плазміду позначено p19L2+prTEF1+FMN1Dh (Фіг.2). Таблиця 1 Перелік праймерів, які використовують у 1, 2 та 3 етапах способу Назва праймера Р7-2В P7-3S А51 А52 Ко36 Послідовність нуклеотидів 5’-TGC GGA ТСС ТАА CGA АС А GCT CAT CAG-3’ 5’-TGC GTC GAC TTT GCT ТАА TGT ATA ATA ATA G-3’ 5’- CCC GTC GAC ATG ACG AGA CCT GAA ACT C AC GTT CC-3’ 5’-CGG GTC GAC CCT TAA GTA AAA GTA CCC CAA AAT AG AAC G-3’ 5’-TTT GAG CTC ATG TTA CAG TCT АТС CCG GC-3’ Етап 2. Конструювання рекомбінантних штамів С. famata Плазмідою p19L2+prTEF1+FMN1Dh, попередньо лінеаризованою рестриктазою HindIII, трансформують реципієнтний штам С. famata L20105 (leu2). Трансформацію проводять методом електропорації, селекцію Leu+трансформантів проводять на мінімальному середовищі Беркгольдера без лейцину [7]. Серед низки трансформантів відбирають стабільні Leu+-інтегранти. Наявність в геномі трансформантів гена FMN1 D. hansenii під промотором TEF1 С. famata перевіряють за допомогою ПЛР з використанням відповідної пари праймерів (Р72В та А52) (Табл. 1). Інтегранти, що містять цю рекомбінантну конструкцію, відбирають для перевірки активності РФ-кінази та продукції ФМН. Трансформанти вирощують протягом 2 діб в колбах Ерленмейєра у середовищі з низьким вмістом заліза (0,01мг Fe/л). Частина відібраних рекомбінантних штамів №3, 9, 18, 33 мають у 3-6 разів вищу активність РФ-кінази у порівнянні з вихідним штамом С famata L20105 і рекомбінантним штамом К9, отриманим трансформацією тією ж плазмідою, але без гена FMN1 (Табл. 2). Таблиця 2 Активність РФ-кінази та вміст флавінів в культуральній рідині рекомбінантних штамів С famata, що містять ген FMN1 D. hansenii під контролем промотора TEF1 Активність РФ-кінази, мкмоль´10-5 /хв´мг білка Штам L20105 (контроль) К9 (контроль) 3 9 18 33 20,4 27,8 70,0 121,7 111,7 86,1 ФМН 0,2 0,45 0,80 1,35 1,30 1,30 Флавіни культуральної рідини, мг/л Сумарна % ФМН кількість флавінів 3,39 6 5,29 9 3,22 25 4,65 29 4,60 28 3,83 34 В культуральному середовищі отриманих рекомбінантних штамів 25-34% від загального вмісту флавінів припадає на ФМН, в той час як у ви хідного штаму L20105 та штаму К9 вміст ФМН не перевищує 10%. Етап 3. Конструювання плазміди для введення в геном додаткової копії гена FMN1 та селекція відповідних трансформантів. Для подальшого покращення продуцентів ФМН у геном відібраних трансформантів вводять додаткові копії гена FMN1 D. hansenii під контролем промотора TEF1 С. famata. Для цього конструкцію, що містить промотор TEF1 з геном FMN1, переклоновують у вектор, який містить ген ble Staphylococcus aureus (забезпечує стійкість до флеоміцину). Сконструйована плазміда отримала назву p19L2+ble+RIBCf+prTEF1+FMN1Dh. Її лінійна схема представлена на Фіг.3. Плазмідою p19L2+ble+RIB1Cf+prTEF1+FMN1Dh трансформують реципієнтний штам С. famata №18, який має підвищену активність РФ-кінази і нагромаджує в культуральній рідині 28% ФМН від загального вмісту фла вінів (Табл. 3). Трансформанти, резистентні до флеоміцину, відбирають на мінімальному середовищі Беркгольдера, що містить флеоміцин у концентрації 1,8мг/л. Серед низки трансформантів відбирають стабільні інтегранти, резистентні до флеоміцину. Наявність в геномі трансформантів частини плазміди p19L2+ble+RIBCf+prTEF1+FMN1Dh, що містить ген FMN1 D. hansenii під промотором TEF1 С. famata та ген ble, перевіряють за допомогою ПЛР з використанням відповідної пари праймерів (Р7-2В та Ко36) (Табл. 1). Інтегранти, що містять цю рекомбінантну конструкцію відбирають для подальшого аналізу. Наявність декількох копій гена FMN1 підтверджують за допомогою Саузерн-блот аналізу [6]. Для перевірки активності РФ-кінази і продукції ФМН отримані трансформанти вирощують протягом 2 діб у середовищі з низьким вмістом заліза (0,01мг Fe/л). Відібрані рекомбінантні штами характеризуються в 3-4,5 рази вищою активністю РФкінази в порівнянні з реципієнтним штамом С. famata №18 і в 13-17 разів - порівняно із рекомбінантним штамом К9, отриманим трансформацією плазмідою без гена FMN1 (Табл. 2, 3). В к ультуральній рідині отриманих рекомбінантних штамів вміст ФМН у 4-5 разів вищий, ніж у реципієнтного штама С. famata №18. Таблиця 3 Активність РФ-кінази та вміст флавінів в культуральній рідині рекомбінантних штамів С. famata, що містять додаткові копії гена FMN1 D. hansenii під контролем промотора TEF1 Штам Копійність Активність РФ-кінази, мкмоль´10-5/хв´мг білка 18 18/13 18/17 18/19 18/21 2-3 не визначали 3-4 6-8 6-8 107,3 448,6 351,1 468,1 482,7 Флавіни культуральної рідини,мг/л Сумарна кількість ФМН % ФМН флавінів 1,30 4,60 28 4,95 15,95 31 6,33 28,33 23 5,50 13,75 40 5,50 10,73 52 Етап 4. Отримання штамів, здатних до надпродукції ФМН, шля хом трансформації плазмідою p19L2+ble+RIBCf+prTEF1+FMN1Dh штаму С. famata ATCC 20849-надсинтетика рибофлавіну. Зростання вмісту ФМН в культуральній рідині спостерігається тільки при вирощуванні рекомбінантних штамів в середовищі з низьким вмістом заліза, коли синтезується значна кількість РФ і є значно нижчим при достатньому забезпеченні залізом. Необхідність створення дефіциту заліза становить значну перешкоду для отримання препарату ФМН у великих кількостях. Тому плазмідою p19L2+ble+RIB1Cf+prTEF1+FMN1Dh трансформують штам С. famata ATCC 20849, продукція РФ у якого не залежить від забезпечення залізом. Активність РФ-кінази і вміст ФМН визначають після 2,5 діб вирощування трансформантів в мінімальному середовищі Беркгольдера без додавання заліза. Результати представлено в табл. 4. Таблиця 4 Активність РФ-кінази та вміст флавінів в культуральній рідині рекомбінантних штамів С. famata АТСС 20849, що містять FMN1 D. hansenii під контролем TEF1 промотора Штам АТСС 20849 2 6 7 11 13 16 Активність РФ-кінази, мкмоль´10-5 /хв´мг білка 43,9 241,4 238,9 234,0 287,7 390,0 299,9 Флавіни культуральної рідини, мг/л Сумарна кількість ФМН % ФМН флавінів 2,67 28,47 10 5,33 13,33 44 17,8 30,24 59 8,0 16,9 48 5,3 8,9 60 24,9 36,50 69 8,9 22,2 40 Отримані рекомбінантні штами мають в 5-9 разів вищу активність РФ-кінази у порівнянні з реципієнтним штамом С. famata АТСС 20849 та приблизно у 20 разів вищу, ніж у штама L20105. В культуральній рідині отриманих рекомбінантних штамів вміст ФМН вищий, ніж у реципієнтного штама С. famata АТСС 20849. Відсотковий вміст ФМН від загального вмісту флавінів становить 40-70%, в той час як у реципієнтного штаму С. famata АТСС 20849 - лише 10%. Джерела інформації: 1. Березовский В.М. Химия витаминов. -М.: Пищевая промышленность, 1973. - 632с. 2. Stahmann K.P. et al. Microbiol. and Biotechnol. - 2000. - Vol.53, N5 - P. 509-516. 3. Сибірний А.А. і співавт. Мікробний синтез флавінів, Київ: Наукова думка, 2006. - 192с. 4. Патент США №5.514574, опубл. 7 травня 1996. 5. Manstein D.J. and Pai E.F.J. Biol. Chem. - 1986. - Vol. 261, №34. - P. 16169-16173. 6. Sambrook J., Fritsh E. F., Maniatis T. Molecular Cloning: A Laboratory Manual. Cold Spring Harbor Laboratory, Cold Spring Harbor, New York. 1989. 7. Voronovsky A.A. et al. //FEMS Yeast Research. - 2002. - Vol. 2. - P. 381-388. 8. Wach A., Pick H., Philipsen P. In: Molecular Genetics of Yeast. A Practical Approach (Johnston, J.R., Ed.), IRL Press, Oxford. - 1994. - P. 1-16.
ДивитисяДодаткова інформація
Назва патенту англійськоюProcess for the preparation of flavin mononucleotide (5'-fmn)
Автори англійськоюVoronovskyi Andrii Yaroslavovych, Dmytruk Kostiantyn Vasyliovych, Ischuk Olena Petrivna, Sybirnyi Andrii Andriiovych, Fedorovych Dariia Vasylivna, Yatsyshyn Valentyna Yuriivna
Назва патенту російськоюСпособ получения флавинмононуклеотида (5'-фмн)
Автори російськоюВороновский Андрей Ярославович, Дмитрук Константин Васильевич, Ищук Елена Петровна, Сибирный Андрей Андреевич, Федорович Дария Васильевна, Яцишин Валентина Юрьевна
МПК / Мітки
МПК: C12P 25/00
Мітки: спосіб, одержання, флавінмононуклеотиду, 5'-фмн
Код посилання
<a href="https://ua.patents.su/4-22568-sposib-oderzhannya-flavinmononukleotidu-5-fmn.html" target="_blank" rel="follow" title="База патентів України">Спосіб одержання флавінмононуклеотиду (5′-фмн)</a>
Спосіб одержання модифікованого штаму дріжджів, здатного стабілізувати 3b-гідроксистероїди, спосіб одержання трансформованого штаму дріжджів та спосіб окислення субстрату

Номер патенту: 72438
Опубліковано: 15.03.2005
Автори: Дегріз Ерік, Ахштеттер Тільман, Кое Жілль
МПК: C12N 15/54, C12N 1/19, C12N 9/10, C12P 33/00, C12N 15/09
Мітки: модифікованого, спосіб, стабілізувати, одержання, штаму, субстрату, окислення, трансформованого, 3b-гідроксистероїди, здатного, дріжджів
Формула / Реферат:
1. Спосіб одержання модифікованого штаму дріжджів, здатного стабілізувати 3b-гідроксистероїди, в якому вихідний штам позбавляють ацетил-СоА-прегненолон-ацетилтрансферазної активності (АРАТ) за рахунок зміни гена ATF2 S. cerevisiae або його гомолога, що має гомологію 60 % з геном ATF2 S. cerevisiae.2. Спосіб за п. 1, в якому зміненим геном є ген ATF2 S. cerevisiae.3. Спосіб за п. 2, в якому ген ATF2 змінений шляхом вставки...
Спосіб одержання ендоспермальних рекомбінантів кукурудзи

Номер патенту: 20433
Опубліковано: 15.01.2007
Автори: Ніколенко Ірина Анатоліївна, Тимчук Сергій Михайлович
МПК: A01H 1/04
Мітки: рекомбінантів, одержання, ендоспермальних, кукурудзи, спосіб
Формула / Реферат:
Спосіб одержання ендоспермальних рекомбінантів кукурудзи, що включає гібридизацію між собою джерел неалельних ендоспермальних мутацій, отримання та самозапилення гібридних рослин першого покоління і другого покоління та виділення в третьому поколінні рекомбінантних форм за фенотипом насіння, який відрізняється тим, що для отримання всіх можливих рекомбінацій між генами структури ендосперму серед насіння другого покоління виділяється таке, що...
Рекомбінантна молекула днк, яка може бути використана для одержання рослини з чоловічою стерильністю, спосіб одержання рослин із чоловічою стерильністю, спосіб одержання самозаплідненого насіння рослин з чолові

Номер патенту: 39169
Опубліковано: 15.06.2001
Автори: Мол Йозефус Ніколас Марія, Ван Тунен Адріанус Йоханес, Ван Дер Мер Інгрід Марія
МПК: C12N 9/10, C12N 15/09, C12N 9/02, C12N 9/00, A01H 1/02, A01H 5/00, C12N 15/11, C12N 15/29, C12N 15/82
Мітки: стерильністю, насіння, використана, спосіб, одержання, днк, чолові, рослин, молекула, рослини, яка, самозаплідненого, рекомбінантна, чоловічою
Формула / Реферат:
1. Рекомбинантная молекула ДНК, которая может быть использована для получения растения с мужской стерильностью и которая содержит ингибиторный ген, способный ингибировать экспрессию гена-мишени, присутствующего в указанном растении и кодирующего фермент пути биосинтеза халкона, и промотор, который является активным в клетках тапетума пыльников указанного растения и который оперативно сшит с указанным ингибиторным геном в целях осуществления...
Спосіб одержання рекомбінантної плазмідной днк рrнw11 або prhw12, кодуючий омега-інтерферон

Номер патенту: 8029
Опубліковано: 26.12.1995
Автори: Ева Растль-Дворкін, Рудольф Гауптманн, Петер Светли, Гюнтер Адольф, Петер Мейндль, Норберт Гауель, Крістіан Пілер
Мітки: рrнw11, кодуючий, рекомбінантної, спосіб, prhw12, одержання, днк, плазмідной, омега-інтерферон
Формула / Реферат:
Формула изобретенияСпособ получения рекомбинантной плазмидной ДНК pRHW11 или pRHW12, кодирующей омега-интерферон, включающий обработку плазмиды Р9А2 или Е79Е9 эндонуклеазой AvaII, выделение вставки кДНК, обработку полученного фрагмента эндонуклеазой Sau3A с последующим выделением фрагмента размером 189 п.о., обработку плазмиды энзимами EcoRI и PvuII, обработку полученного фрагмента размером 389 п.о., содержащего trp-промотор, место...
Спосіб одержання імунної сироватки проти набрякової хвороби (коліентеротоксемії) поросят

Номер патенту: 59914
Опубліковано: 15.09.2003
Автори: Головко Анатолій Миколайович, Ушкалов Валерій Олександрович, Горбенко Олександр Віталійович
МПК: A61K 39/085
Мітки: набрякової, коліентеротоксемії, спосіб, одержання, сироватки, імунної, поросят, хвороби
Формула / Реферат:
Спосіб одержання імунної сироватки проти набрякової хвороби (коліентеротоксемії) поросят, призначеної для пасивної імунізації і імунотерапії набрякової хвороби у поросят, шляхом введення до складу антигенів для імунізації донорів сироватки крові полівалентного комплексу протективних антигенів збудників (гемолізинів, екзотоксинів, адгезивних антигенів), який відрізняється тим, що адгезивні антигени відокремлюють із суспензії бакмаси виробничих...
Попередній патент: Спосіб лікування хворих на вторинну неоваскулярну глаукому
Наступний патент: Спосіб діагностики папілярних карцином щитовидної залози
Випадковий патент: Спосіб лікування гострого некротичного панкреатиту