Спосіб одержання модифікованого штаму дріжджів, здатного стабілізувати 3b-гідроксистероїди, спосіб одержання трансформованого штаму дріжджів та спосіб окислення субстрату







Формула / Реферат
1. Спосіб одержання модифікованого штаму дріжджів, здатного стабілізувати 3b-гідроксистероїди, в якому вихідний штам позбавляють ацетил-СоА-прегненолон-ацетилтрансферазної активності (АРАТ) за рахунок зміни гена ATF2 S. cerevisiae або його гомолога, що має гомологію 60 % з геном ATF2 S. cerevisiae.
2. Спосіб за п. 1, в якому зміненим геном є ген ATF2 S. cerevisiae.
3. Спосіб за п. 2, в якому ген ATF2 змінений шляхом вставки послідовності ДНК, що включає принаймні один нуклеотид.
4. Спосіб за п. 3, в якому ген ATF2 змінений шляхом вставки селекційного гена URA3 або блока експресії TEF1prom/PGKterm.
5. Спосіб за п. 4, в якому ген ATF2 змінений шляхом вставки селекційного гена URA3.
6. Спосіб за п. 5, в якому одержують модифіковані штами S. cerevisiae, які називаються TGY156 і TGY158.
7. Спосіб за п. 4, в якому ген ATF2 змінений шляхом вставки блока експресії TEF1prom/PGKterm.
8. Спосіб одержання трансформованого штаму дріжджів, в якому штам, одержаний у спосіб за п.1, трансформують таким чином, що він здатний експресувати принаймні один з ферментів шляху біосинтезу гідрокортизону з холестерину, який вибирається з-поміж таких ферментів:
фермент розщеплення бокового ланцюга холестерину (P450SCC);
3b-гідрокси-дельта5-стероїд-дегідрогеназа/дельта5-дельта4-стероїд-ізомераза (3b-HSD); і
17a-стероїд-гідроксилаза (Р45017a).
9. Спосіб за п. 8, в якому зміненим геном є ген ATF2 S. cerevisiae або його гомолог.
10. Спосіб за п. 9, в якому зміненим геном є ген ATF2 S. cerevisiae.
11. Спосіб за п. 10, в якому ген ATF2 змінений шляхом вставки послідовності ДНК, що включає принаймні один нуклеотид.
12. Спосіб за п. 10, в якому ген ATF2 змінений шляхом вставки селекційного гена URA3.
13. Спосіб за п. 12, який відрізняється тим, що штам експресує 3b-HSD.
14. Спосіб за п. 13, в якому одержують трансформований штам S. cerevisiae, що позначається TGY158/pTGY10862, депонований у CNCM за номером I-1978.
15. Спосіб за п. 10, в якому ген ATF2 змінений шляхом вставки блока експресії TEF1prom/PGKterm.
16. Спосіб за п. 15, який відрізняється тим, що штам експресує Р45017a.
17. Спосіб за п. 16, в якому одержують трансформований штам S. cerevisiae, що позначається TGY186/pTG10435, депонований у CNCM за номером I-2119.
18. Спосіб за п. 15, який відрізняється тим, що штам коекспресує 3b-HSD і Р45017a.
19. Спосіб за п. 18, в якому одержують трансформований штам S. cerevisiae, що позначається TGY186/pTG10417, депонований у CNCM за номером I-2118.
20. Спосіб окислення in vivo субстрату, що вибирається з-поміж ендогенного стерину, екзогенного стерину або екзогенного стероїду, згідно з яким використовують штам дріжджів, трансформований у спосіб за будь-яким з пп. 8-19, який або культивують окремо, коли штам генерує ендогенний стерин, або його інкубують зі стероліном чи екзогенним стероїдом, і, у разі необхідності, виділяють одержану окислену сполуку.
21. Спосіб окислення in vivo за п. 20, згідно з яким субстратом є 3b-гідроксистероїд і в якому використовують штам дріжджів, трансформований у спосіб за п. 13, і, у разі необхідності, виділяють одержаний 3-оксо-дельта4-стероїд.
22. Спосіб окислення in vivo за п. 21, згідно з яким 3b-гідроксистероїд вибирають з-поміж прегненолону або 17a-гідрокси-прегненолону.
23. Спосіб окислення in vivo за п. 20, згідно з яким субстратом є стероїд і в якому використовують штам дріжджів, трансформований у спосіб за п. 16, і, у разі необхідності, виділяють одержаний 17a-гідроксилований стероїд.
24. Спосіб окислення in vivo за п. 23, згідно з яким стероїдним субстратом є прегненолон або прогестерон.
25. Спосіб окислення in vivo за п. 20, згідно з яким субстратом є 3b-гідроксистероїд і в якому використовують штам дріжджів, трансформований у спосіб за п. 18, і, у разі необхідності, виділяють одержаний 17a-гідроксилований 3-оксо-дельта4-стероїд.
26. Спосіб окислення in vivo за п. 27, згідно з яким стероїдним субстратом є прегненолон.
Текст
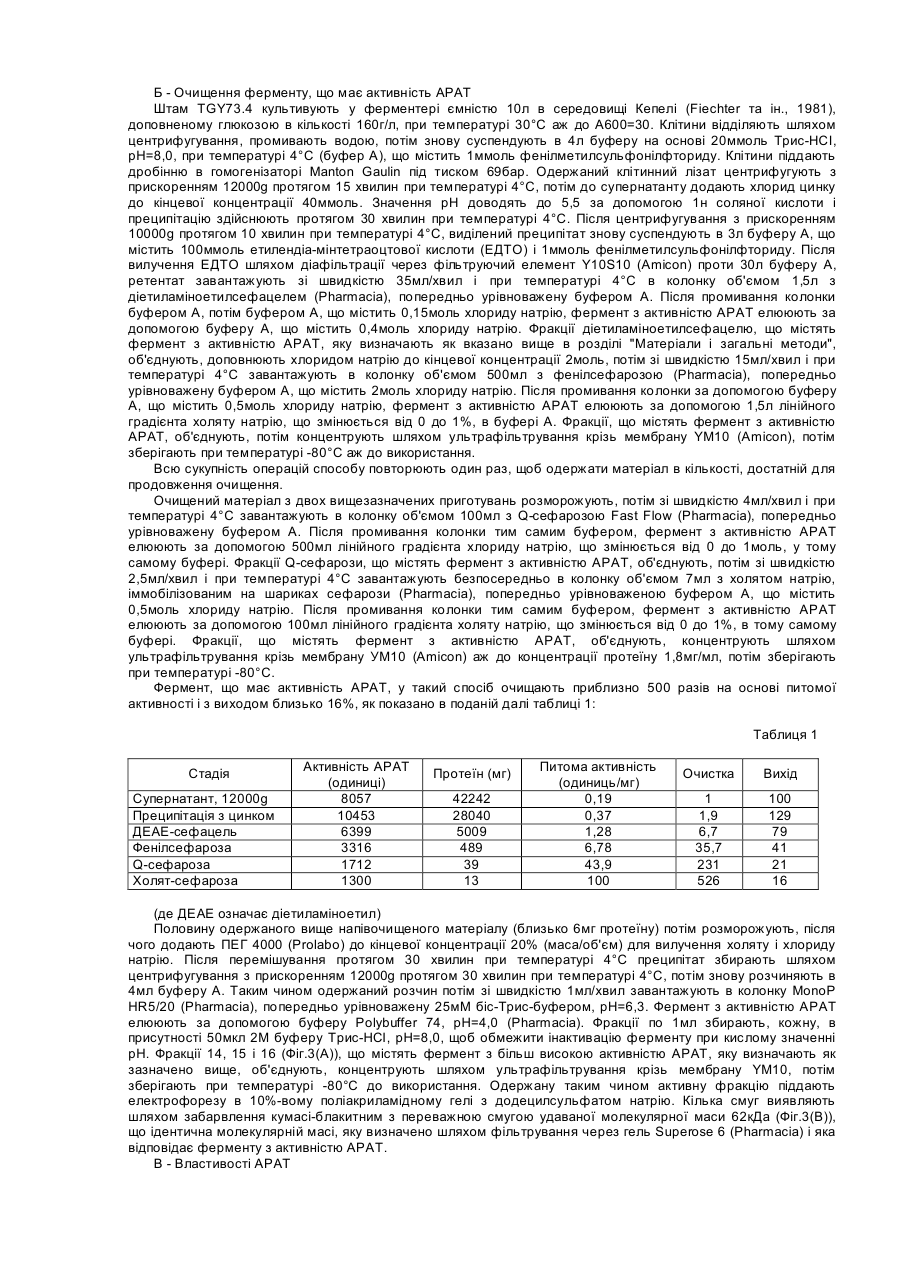
Утворення 3-оксо-дельта4-стероїдів з 3β-гідрокси-дельта5-попередників в біосинтезі всіх класів стероїдних гормонів у ссавців каталізується ферментативною системою 3β-гідрокси-дельта5-стероїд-дегідрогеназа (EC 1.1.1.145) і дельта5-дельта4-стероїд-ізомераза (EC 5.3.3,1), що називається 3β-ΗSD. 3β-HSD каталізує, наприклад, трансформацію прегненолону в прогестерон, 17α-гідроксипрегненолону в 17αгідроксипрогестерон, дегідроепіандростерону в дельта4-андростендіон або 5-андростен-3β-17β-діолу в тестостерон (Simard та ін., 1996). Отже, 3β-ΗSD являє собою один з ключових ферментів шляху біосинтезу гідрокортизону з холестерину в корі надниркових залоз ссавців (Фіг.1). Використання рекомбінантних мікроорганізмів, зокрема, трансформованих дріжджів, що дозволять здійснювати гетерологічну експресію одного чи кількох ферментів ссавців цього шляху біосинтезу для продукування гідрокортизону або проміжних продуктів цього біосинтезу, описується, наприклад, [в заявці на європейський патент 340878, в патенті США 5137822, Dumas та ін., 1994, і Cauet та ін., 1994]. У випадку експресії функціональної системи 3β-HSD в дріжджах, трансформовані клітини дріжджів не перетворюють повністю 3β-гідроксистероїди на відповідні 3-оксостероїди, наприклад, прегненолон на прогестерон, але акумулюють сполуку, що виявляється також у випадку клітин нетрансформованих дріжджів. Ідентифікація акумульованої сполуки з складним 3β-оцтовим ефіром вихідного стероїду і характеристика ферменту, що має ацилтрансферазну активність і відповідального за цю етерифікацію (далі позначається АРАТ: "ацетил-кофермент-А-прегненолон-ацетилтрансфераза") описуються в цій заявці. З іншого боку, акумуляція прегненолонацетату трансформованим штамом дріжджів, що продукує прегненолон, описується в заявці на європейський патент 727489, Ці дані дозволяють вважати, що здійснювана дріжджами етерифікація 3β-гідроксистероїдів є небажаною, оскільки вона відповідальна за вторинні реакції і побічні продукти, які призводять до зниження виходу 3β-гідроксистероїдів, що акумулюються, наприклад, прегненолону, або до зниження виходу від біоконверсії 3β-гідрокси-дельта5-стероїдів в 3-оксо-дельта4-стероїди, особливо при продукуванні прогестерону або 17α-гідроксипрогестерону, що призводить до зменшення подальшої продукції гідрокортизону вже описаним шляхом біосинтезу. У зв'язку з отриманими вищезазначеними результатами, цей винахід стосується конструювання штамів дріжджів, позбавлених небажаної активності АРАТ, шляхом зміни гена, який кодує цю активність, що веде до стабілізації 3β-гідроксистероїдів в їх присутності. Ці штами, отже, використовуються як вихідні штами для конструювання рекомбінантних штамів, здатних трансформувати 3β-гідроксистероїди в подальші продукти з покращеним виходом. Винахід також стосується конструювання штамів дріжджів, позбавлених активності АРАТ, шляхом зміни гена, який кодує цю активність, і або експресуючих 3β-HSD чи цитохром P45017α, або коекспресуючих 3β-HSD і цитохром Р45017α шляху біосинтезу гідрокортизону з холестерину. Штами, які експресують, наприклад, 3βHSD, дозволяють покращувати вихід від біоконверсії 3β-гідрокси-дельта5-стероїдів в 3-оксо-дельта4-стероїди і, отже, використовуються у вдосконалених способах одержання гідрокортизону або проміжних продуктів в дріжджах. Таким чином, об'єктом цього винаходу є модифікований штам дріжджів, позбавлений ацетил-СоАпрегненолон-ацетилтрансферазної активності (АРАТ) за рахунок зміни гена, який кодує цю активність, що приводить до стабілізації 3β-гідроксистероїдів. Зміна кодуючого активність АРАТ гена можна здійснювати, наприклад, шляхом вставки, делеції або заміни послідовності ДНК в функціональних елементах гена, наприклад, промоторі або послідовності, що кодує протеїн з активністю АРАТ. Потім може здійснюватись інтеграція зміненої таким чином послідовності ДНК в дріжджовий штам-хазяїн, наприклад, методом гомологічної рекомбінації, що веде до генерації хромосомних мутантів дріжджів, які відповідають модифікованим штамам відповідно до винаходу, в яких зникнення активності АРАТ і стабілізація 3β-гідроксистероїдів виявляються, наприклад, шляхом культивування клітин в присутності прегненолону і визначення вмісту прегненолону залежно від часу згідно з робочими умовами, описаними далі в експериментальній частині. В якості дріжджових штамів-хазяїв, що використовуються відповідно до винаходу, можна, зокрема, назвати штами Saccharomyces, такі, як S.cerevisiae, штами Candida, такі, як С.maltosa, штами Kluyveromyces, такі, як, K.lactis, або штами Pichia, такі, як P.pastoris. Об'єктом винаходу зокрема є вищезазначений модифікований штам дріжджів, в якому зміненим геном є ген ATF2 S.cerevisiae або його гомолог. Геном ATF2 називають ген S.cerevisiae, ідентифікований у геномі дріжджів під локусом ATF2 або YGR177c "бази даних геномів Saccharomyces" (SGD); (Cherry та ін., http://genome-www, Stanford.edu/Saccharomyces/), відкрита рамка зчитування якого, що позначається YGRl77c, виражається у вигляді амінокислотної послідовності в банку даних Mips, що знаходиться під присвоєним номером за каталогом S64491 (Hebling U., Hofmann В. і Delius Η. (травень 1996), та послідовність якого подано на Фіг.4. Цей ген кодує протеїн з активністю АРАТ, як це показано далі в експериментальній частині. Під геном, гомологічним гену ATF2, розуміють ген, що кодує протеїн, який має активність АРАТ і ідентичність послідовності приблизно на 60% і більше з послідовністю протеїну YGR177C. Особливо, об'єктом винаходу є вищезазначений модифікований штам дріжджів, в якому зміненим геном є ген ATF2 S.cerevisiae і який далі називається мутантним штамом atf2. Особливо об'єктом винаходу є вищезазначений модифікований штам дріжджів, в якому ген ATF2 змінений шляхом вставки послідовності ДНК, яка включає принаймні один нуклеотид. Послідовність ДНК, що вбудована в ген ATF2 так, щоб зникла активність АРАТ, може являти собою, наприклад, ауксотрофний селекційний ген, який доповнює живильну потребу штаму-хазяїна, такий, як ген URA3, ген LEU2, ген TRP1, ген HIS3 або ген ADE2, наприклад, домінантний селекційний ген, такий, як ген стійкості до антибіотика, як G418, флеоміцин або гігроміцин В, або, наприклад, репортерний ген, такий, як ген PGAL. Послідовність ДНК, яку вбудовують в ген ATF2, може являти собою також блок експресії дріжджів, утворений промотором і термінальною послідовністю транскрипції, наприклад, промотором дріжджів, таким, як PGK, TDH3, СYС1 або TEF1, наприклад, термінальною послідовністю дріжджів, такою, як CYC1, TDH3, TEF1 або PGK, Блок експресії може являти собою поєднання вищезазначених елементів, наприклад, як блок TEF1prom/PGKterm. Об'єктом винаходу більш конкретно є вищезазначений модифікований штам дріжджів, в якому ген ATF2 змінений шляхом вставки селекційного гена URA3 або блоку експресії TEF1prom/PGKterm. Об'єктом винаходу, зокрема, є вищезазначений модифікований штам дріжджів, в якому ген ATF2 змінений шляхом вставки селекційного гена URA3. Мутантні штами atf2 відповідно до винаходу, позбавлені активності АРАТ і в які вбудований ген URA3, що називатимуться далі atf2-Δ::URA3, також можуть бути селекційовані шляхом прототрофії з урацилом. Особливо, об'єктом винаходу є модифіковані штами S.cerevisiae, що називаються TGY156 і TGY158, конструкцію яких детально подано далі в експериментальній частині. Частковим об'єктом винаходу також є вищезазначений модифікований штам дріжджів, в якому ген ATF2 змінений шляхом вставки блоку експресії TEF1prom/PGKterm Мутантні штами atf2 відповідно до винаходу, позбавлені активності АРАТ і в які вбудований блок експресії TEF1prom/PGKterm, що називаються далі atf2Δ::TEF1prom/PGKterm, можуть бути сулукційовані у напрямку відсутності функціонального гена URA3, заміщеного блоком експресії, за їх стійкістю до 5-фтороротової кислоти. Особливо, об'єктом винаходу є модифікований штам S.cerevisiae, що називається TGY186, докладне конструювання якого подано далі в експериментальній частині. Об'єктом винаходу також є трансформований штам дріжджів, позбавлений ацетил-СоА-прегненолонацетилтрансферазної активності (ΑΡΆΤ) за рахунок зміни гена, який кодує цю активність, і експресує принаймні один з ферментів ссавців шляху біосинтезу гідрокортизону з холестерину, що вибирається серед таких ферментів: - фермент розщеплення бокового ланцюга холестерину (P450SCC); - 3β-гідрокси-дельта5-стероїд-дегідрогеназа /дельта5-дельта4-стероїд-ізомераза (3β-HSD); і - 17α-стероїд-гідроксилаза (Р45017α). Трансформовані штами дріжджів відповідно до винаходу можна одержати, наприклад, шляхом трансформації мутантних штамів atf2 відповідно до винаходу за допомогою відомих способів, наприклад, шляхом трансформації за допомогою вектора експресії P450SCC, а також ADX і ADR, вектора експресії 3β-HSD або вектора експресії Р45017α, Мутантні штами atf2 також можуть бути, у разі необхідності, котрансформовані, наприклад, за допомогою вектора експресії 3β-HSD і вектора експресії Р45017α, або можуть бути трансформовані за допомогою вектора коекспресії 3β-HSD і Р 45017α і можуть використовуватись, наприклад, у способі біоконверсії прегненолону в 17α-гідрокси-прогестерон. Сконструйовані вектори експресії P450SCC, а також ADX і ADR, 3β-HSD або Р45017α бичачого або людського походження в штамах дріжджів описуються, наприклад, Dumas та ін., 1994, в заявці на європейський патент 340878 або в патенті США 5137822. Винахід стосується, зокрема, вищезазначеного трансформованого штаму дріжджів, в якому зміненим геном є ген ATF2 S.cerevisiae або його гомолог. Особливо об'єктом винаходу є вищезазначений трансформований штам дріжджів, в якому змінений ген являє собою ген ATF2 S.cerevisiae і який відповідає трансформованому штаму atf2. Особливо, об'єктом винаходу є вищезазначений трансформований штам дріжджів, в якому ген ATF2 змінений за рахунок вставки послідовності ДНК, що включає принаймні один нуклеотид, і, конкретно, об'єктом є трансформований штам дріжджів, в якому ген ATF2 змінений за рахунок вставки селекційного гена URA3 і який відповідає трансформованому штаму atf2-Δ::URA3. Зміна гена з метою ліквідації активності АРАТ, ген ATF2 або його гомолог, а також штами-хазяї мають вищезазначені значення. Винахід більш прийнятно стосується вищезазначеного трансформованого штаму дріжджів atf2-Δ::URA3, що експресує 3β-HSD, і особливо трансформованого штаму S.cerevisiae, що називається TGY158/pTG108 62, докладне конструювання якого і описується далі в експериментальній частині. Об'єктом винаходу також особливо є вищезазначений трансформований штам дріжджів, в якому ген ATF2 змінений за рахунок вставки блоку експресії TEF1prom/PGKterm і який відповідає трансформованому штаму atf2Δ::TEF1prom/PGKterm. Винахід також більш прийнятно стосується вищезазначеного трансформованого штаму atf2Δ::TEF1prom/PGKterm, що експресує Р45017α, особливо трансформованого штаму S.cerevisiae, що називається TGY186/pTG10435. Винахід стосується зокрема вищезазначеного трансформованого штаму дріжджів atf2-Δ::TEF1prom/PGKterm, що коекспресує і 3β-ΗSD і Ρ 45017α, і більш прийнятно трансформованого штаму. S.cerevisiae, що називається TGYl86/pTG10417. Об'єктом винаходу також є спосіб окислення in vivo субстрату, що вибирається з-поміж ендогенного стерину, екзогенного стерину або екзогенного стероїду, згідно з яким використовують вищезазначений трансформований штам дріжджів, який або культивують індивідуально, коли штам генерує ендогенний стерин, або інкубують з екзогенним стерином або стероїдом, і, у разі необхідності, виділяють одержану окислену сполуку. Ендогенним стерином називають стерин, що акумулюється в штамі дріжджів і являє собою субстрат ферменту розщеплення бокового ланцюга (P450SCC), коли дріжджі, після трансформації, наприклад, за допомогою вектора експресії P450SCC, ADX і ADR, культивують за відсутності екзогенного стерину. Ендогенними стеринами, що використовуються для здійснення способу відповідно до винаходу, можуть бути, наприклад, ергоста-5-ен-3-ол, ергоста-5,24 (28)-дієн-3-ол або ергоста-5,22-дієн-3-ол. В заявці на європейський патент 727489 описується акумуляція таких стеролів у штамі дріжджів і розщеплення їх бокового ланцюга в культурі штаму після трансформації за допомогою вектора експресії P450SCC, ADX і ADR. Такій штам дріжджів, який також має активність АРАТ, може бути попередньо модифікований для одержання мутантного штаму atf2 відповідно до винаходу, потім його можна трансформувати за допомогою вектора експресії P450SCC,. ADX і ADR для одержання трансформованого мутантного штаму atf2 відповідно до винаходу. Екзогенним стерином називають стерин, що являє собою субстрат ферменту розщеплення P450SCC, одержуваний шляхом інкубації зі штамом дріжджів, трансформованим за допомогою вектора експресії P450SCC, ADX і ADR, наприклад, холестерин або ситостерин. Таким штамом може бути, наприклад, мутантний штам atf2, трансформований за допомогою вектора експресії P450SCC, ADX І ADR. 3β-гідроксистероїд, одержаний шляхом розщеплення бокового ланцюга ендогенного або екзогенного стерину, що використовується в якості субстрату, перебуває повністю в вільній формі, тобто не супроводжується відповідним складним 3β-оцтовим ефіром, в культурах трансформованих штамів atf2, що експресують P450SCC, ADX і ADR. Стероїдом називають стероїд, що являє собою субстрат ферменту 3β-HSD при інкубації зі штамом дріжджів, трансформованим, наприклад, за допомогою вектора експресії 3β-HSD, як прегненолон, 17αгідроксипрегненолон або дегідроепіандростерон, або стероїд, який являє собою субстрат ферменту P45017α при інкубації зі штамом дріжджів, трансформованим, наприклад, за допомогою вектора експресії Р45017α, як прогестерон або прегненолон. Таким штамом може бути, наприклад, мутантний штам atf2, трансформований за допомогою вектора експресії 3β-HSD або за допомогою вектора експресії P45017α відповідно до винаходу. Об'єктом винаходу особливо є вищезазначений спосіб окислення in vivo, згідно з яким субстратом є 3βгідроксистероїд і в якому використовують трансформований штам дріжджів atf2-Δ::UΚΑ3, який експресує 3βHSD, і у разі необхідності, виділяють одержаний 3-оксо-дельта4-стероїд, і особливо об'єктом винаходу є спосіб, згідно з яким 3β-гідроксистероїд вибирають з прегненолону або 17α-гідроксипрегненолону. Використовуваний в якості субстрату 3β-гідроксистероїд є стабільним, коли його інкубують зі штамом atf2Δ::URA3 відповідно до винаходу, трансформованим за допомогою вектора експресії 3β-HSD. Таким чином, винахід стосується покращеного способу одержання 3-оксо-дельта4-стероїду в дріжджах, оскільки всю кількість 3β-гідрокси-субстрату може бути окислено до 3-оксо-дельта4-стероїду, як це показано далі в експериментальній частині. Також особливо об'єктом винаходу є вищезазначений спосіб окислення in vivo, згідно з яким субстратом є стероїд і в якому використовують трансформований штам дріжджів atf2-Δ::TEF1prom/PGKterm, який експресує Р45017α, і, у разі необхідності, виділяють одержаний 17α-гідроксильований стероїд, і особливо об'єктом винаходу є спосіб, згідно з яким стероїдним субстратом є прегненолон або прогестерон. Використовуваний в якості субстрату прегненолон є стабільним, коли його інкубують зі штамом atf2Δ::TEF1prom/PGKterm, трансформованим за допомогою вектора експресії Р45017α. Отже, винахід також стосується покращеного способу одержання 17α-гідроксистероїдів з 3βгідроксистероїдів, оскільки всю кількість 3β-гідрокси-субстрату може бути 17α-гідроксильовано. Також особливо об'єктом винаходу є вищезазначений спосіб окислення in vivo, згідно з яким субстратом є 3β-гідроксистероїд і в якому використовують трансформований штам дріжджів atf2-Δ::TEF1prom/PGKterm, що коекспресує 3β-HSD і Р45017α, і, у разі необхідності, виділяють одержаний 17α-гідроксильований 3-оксодельта4-стероїд. Особливо об'єктом винаходу є вищезазначений спосіб, згідно з яким субстратом є прегненолон. Трансформовані мутантні штами дріжджів atf2 і спосіб відповідно до винаходу дозволяють передбачати їх більш прийнятне використання для покращеного продукування гідрокортизону або його проміжних продуктів в дріжджах. Приклади конструювання штамів відповідно до винаходу і втілення способу, що пропонується у винаході, описуються далі в експериментальній частині. Матеріали і загальні методи 1. Штами і середовища Штами S.cerevisiae, що використовуються для здійснення винаходу, являють собою штам TGY73.4 (МАТа, URA3-Δ5, pra-1-1, prb-1-1, prc1-1, cps1-3, his), ізогенний похідний Leu+ с13ABYS86, описаний Achstetter та ін., 1992, і штам Fy1679 (МАТа, URA3-52, trp1-Δ63, leu2-Δ1, his3-Δ200, fen1, GAL), описаний Thierry та ін., 1990. Вирощування штамів здійснюють в повному середовищі YPD (Difco Laboratories), що містить 2% глюкози при температурі 28°С, згідно з умовами, описаними F.Sherman, 1991. Для трансформації S.cerevisiae, клітинам надають компетентності у спосіб з використанням ацетату літію (Ito та ін., 1983). Дріжджі як правило культивують в мінімальному синтетичному середовищі 3D, що містить 2% глюкози (F.Sherman, 1991) з доданням необхідних живильних речовин в концентрації 100мкг/мл. Штам Е.соli BJ5183 (D.Hanahan, 1983) використовують для рекомбінації in vivo та штам Е.coli C600, hsdR (Hubacek та ін., 1970) використовують в якості штаму-реципієнта для класичних реакцій лігування. 2. Маніпулювання з ДНК і рекомбінація in vivo в Е.соlі Використовувані загальні методи молекулярної біології описуються Sambrook та ін., 1989. Метод рекомбінації in vivo описується Е.Degryse, 1995, і Е.Degryse, 1996. 3. Тестування ферментної активності AРАT Ацетидтрансферазну активність АРАТ виявляють шляхом визначення включення [3Н]-ацетату в прегненолон з [3Н]-ацетил-CoA (New England Nuclear). Реакційне середовище (500мкл) містить [3Н]-ацетилСоА (20ммоль; 25Ки/моль) і прегненолон (30ммоль; Sigma). Прегненолон додають у вигляді розчину в 2мкл суміші тилоксаполу (Sigma) з етанолом у співвідношенні 1:1 у буфер на основі фосфату калію (20мкмоль) з рН=7,0. Після інкубації протягом 15 хвилин при температурі 30°С реакцію припиняють шляхом додання 2мл дихлорметану. Стероїди екстрагують дихлорметаном, потім поділяють шляхом високоефективної рідинної хроматографії з оберненою фазою (далі називається ВЕРХ з оберненою фазою) в ізократичних умовах елюювання за допомогою ацетонітрилу при використанні колонки Ultrasphere ODS (Beckman) при температурі 45°С на хроматографі HP 1090 (Hewlett Packard), що з'єднаний з paдіодетектором FLO-One 500 (Packard), який дозволяє визначати кількість [3Н]-ацетату прегненолону, що утворився. Одиницю АРАТ визначають як кількість ферменту, що продукує 1нмоль ацетату прегненолону на хвилину при температурі 30°С у вищезазначених умовах. 4. Визначення концентрації протеїну Концентрацію протеїну визначають шляхом використання набору "набір для кількісного аналізу протеїну" (Bio-Rad) з бичачим сироватковим альбуміном в якості стандарту. На Фіг., що додаються до цього опису, ілюструються деякі аспекти винаходу. На Фіг.1 подано шлях біосинтезу гідрокортизону з холестерину у ссавців. Фіг.2А і 2В ілюструють біоконверсію прегненолону в ацетат прегненолону в S.cerevisiae. Аналіз здійснюється шляхом високоефективної рідинної хроматографії з оберненою фазою при 205нм. На Фіг.2А подано кінетику утворення ацетату прегненолону і зникнення прегненолону в окремі інтервали часу протягом 12 годин інкубації. На Фіг.2В подано профіль стероїдів у час 0 (t=0) і 10 годин (t=10 годин) відносно до профілю стандартів прегненолону і ацетату прегненолону. Фіг.3 ілюструє очищення АРАТ шляхом хроматографії на MonoP HR 5/20: на (А) подано профіль активності АРАТ, наявної в фракціях 10-20; на (В) подано аналіз шляхом електрофорезу в поліакриламідному гелі з додецилсульфатом натрію очищених і концентрованих фракцій 14, 15 і 16 шляхом забарвлення за допомогою кумасі-блакитного (лінія 2) в присутності маркерів молекулярної маси (лінія 1). Стрілка вказує на смугу з удаваною молекулярною масою 62кДа. На Фіг.4 подано амінокислотну послідовність у вигляді рамки зчитування протеїну YGR177c. Секвеновані пептиди з очищеного і гідролізованого трипсином протеїну ΑΡΆΤ підкреслені. На Фіг.5 подано стратегію переривання гена ATF2 шляхом асоціації з геном URA3 відповідно до методу подвійного злиття полімеразної ланцюгової реакції (ПЛР). Порожні бруски і заповнені бруски означають, відповідно, послідовності ATF2 і URA3. Фіг.6 ілюструє вплив переривання гена ATF2 в S.cerevisiae на ацетилування прегненолону. Присутність лрегне-нолонацетату виявляється за допомогою високоефективної рідинної хроматографії з оберненою фазою при 205нм з шістнад-цятигодинних культур батьківського штаму TGY73.4 (А) або мутантного штаму TGY158 (В). На Фіг.7А, 7В і 7С подано схему конструювання плазмід експресії людського 3β-HSD в дріжджах pTG10832 і pTG10862. На Фіг.7А описується одержання фрагменту Mscl-Mlul, що містить послідовність, яка кодує людський 3βHSD, На Фіг.7В подано одержання фрагменту NotІ, що містить блок експресії CYC1p/3β-HSD/PGKt. На Фіг.7С описується одержання плазміди pTG10832 і плазміди pTG10862. На Фіг.8 подано рестрикційну карту плазміди pTG10832. На Фіг.9 подано рестрикційну карту плазміди pTG10862. На Фіг.10 подано схему конструювання плазміди експресії pTG10435. На Фіг.11 подано схему конструювання плазміди pTG10058. На Фіг.12 подано рестрикційну карту плазміди рTG10058. На Фіг.13 подано рестрикційну карту плазміди рTG10293. На Фіг.14 подано рестрикційну карту плазміди pTG10435. На Фіг.15А і 15В подано схему конструювання плазміди pTG10274: на Фіг.15А описується одержання плазміди pTG10214; на Фіг.15В описується одержання плазміди PTG10274. На Фіг.16 подано рестрикційну карту плазміди рTG10274. На Фіг.17 подано рестрикційну карту плазміди рTG10401. На Фіг.18 подано схему конструювання вектора експресії pTG10262. На Фіг.19 подано рестрикційну карту плазміди pTG10262. На Фіг.20 подано рестрикційну карту плазміди рTG10403. На Фіг.21 подано рестрикційну карту плазміди pTG10417. (Абревіатури рестрикційних ферментів: S=SalI; N=NotI; BII=BglII; M=MluI; C=ClaI; N°=без сайта Ncol; ХbаІ°=без сайта Xbal; E=EcoRI). Приклад 1 Ідентифікація активності АРАТ дріжджів А - Ацетилування in vivo прегненолону за допомогою дріжджів Штам TGY73.4 культивують при температурі 28°С в 10мл середовища YPD (Difco), інокульованого з розрахунку A600=0,1, виходячи з двадцятичотирьохгодинної вихідної культури, і в які додають 100мкл розчину прегненолону з концентрацією 10мг/мл в суміші тергитолу (Sigma) з етанолом у співвідношенні 1:1. Стероїди, що утворилися, ідентифікують при використанні аліквот об'ємом 250мкл бульйону, відібраних в різні інтервали часу протягом 10 годин. Після екстракції за допомогою 2мл дихлорметану органічні фази випарюють в атмосфері азоту, потім одержані залишки знову розчиняють в ацетонітрилі. Стероїди аналізують за допомогою ВЕРХ з оберненою фазою на колонці U1-trasphere ODS (Beckman), використовуючи послідовно в якості елююючого засобу: 60%-вий розчин ацетонітрилу у воді протягом 10 хвилин, потім 60-80%-вий розчин ацетонітрилу у воді протягом 5 хвилин, після цього 80%-вий розчин ацетонітрилу у воді протягом 5 хвилин, з витратою 1мл/хвил, при температурі 45°С і з детекціею при 205нм. Одержані хроматограми (Фіг.2В) показують, що прегненолон метаболізований в більш аполярний продукт, який має час утримування, ідентичний такому стандартного прегненолонацетату. На Фіг.2А показано, що прегненолон швидко перетворюється дріжджами на його метаболіт. Після лужної обробки (6%-вий розчин КОН в метанолі) виявлений метаболіт вивільнює продукт з часом утримування, ідентичним такому прегненолону. Ідентифікацію метаболіту з прегненолонацетатом потім підтверджують методом мас-спектрометрії. Б - Очищення ферменту, що має активність АΡΑΤ Штам TGY73.4 культивують у ферментері ємністю 10л в середовищі Кепелі (Fiechter та ін., 1981), доповненому глюкозою в кількості 160г/л, при температурі 30°С аж до А600=30. Клітини відділяють шляхом центрифугування, промивають водою, потім знову суспендують в 4л буферу на основі 20ммоль Трис-НСІ, рН=8,0, при температурі 4°С (буфер А), що містить 1ммоль фенілметилсульфонілфториду. Клітини піддають дробінню в гомогенізаторі Manton Gaulin під тиском 69бар. Одержаний клітинний лізат центрифугують з прискоренням 12000g протягом 15 хвилин при температурі 4°С, потім до супернатанту додають хлорид цинку до кінцевої концентрації 40ммоль. Значення рН доводять до 5,5 за допомогою 1н соляної кислоти і преципітацію здійснюють протягом 30 хвилин при температурі 4°С. Після центрифугування з прискоренням 10000g протягом 10 хвилин при температурі 4°С, виділений преципітат знову суспендують в 3л буферу А, що містить 100ммоль етилендіа-мінтетраоцтової кислоти (ЕДТО) і 1ммоль фенілметилсульфонілфториду. Після вилучення ЕДТО шляхом діафільтрації через фільтруючий елемент Y10S10 (Amicon) проти 30л буферу А, ретентат завантажують зі швидкістю 35мл/хвил і при температурі 4°С в колонку об'ємом 1,5л з діетиламіноетилсефацелем (Pharmacia), попередньо урівноважену буфером А. Після промивання колонки буфером А, потім буфером А, що містить 0,15моль хлориду натрію, фермент з активністю АРАТ елююють за допомогою буферу А, що містить 0,4моль хлориду натрію. Фракції діетиламіноетилсефацелю, що містять фермент з активністю АРАТ, яку визначають як вказано вище в розділі "Матеріали і загальні методи", об'єднують, доповнюють хлоридом натрію до кінцевої концентрації 2моль, потім зі швидкістю 15мл/хвил і при температурі 4°С завантажують в колонку об'ємом 500мл з фенілсефарозою (Pharmacia), попередньо урівноважену буфером А, що містить 2моль хлориду натрію. Після промивання колонки за допомогою буферу А, що містить 0,5моль хлориду натрію, фермент з активністю АРАТ елююють за допомогою 1,5л лінійного градієнта холяту натрію, що змінюється від 0 до 1%, в буфері А. Фракції, що містять фермент з активністю АРАТ, об'єднують, потім концентрують шляхом ультрафільтрування крізь мембрану YM10 (Amicon), потім зберігають при температурі -80°С аж до використання. Всю сукупність операцій способу повторюють один раз, щоб одержати матеріал в кількості, достатній для продовження очищення. Очищений матеріал з двох вищезазначених приготувань розморожують, потім зі швидкістю 4мл/хвил і при температурі 4°С завантажують в колонку об'ємом 100мл з Q-сефарозою Fast Flow (Pharmacia), попередньо урівноважену буфером А. Після промивання колонки тим самим буфером, фермент з активністю АРАТ елююють за допомогою 500мл лінійного градієнта хлориду натрію, що змінюється від 0 до 1моль, у тому самому буфері. Фракції Q-сефарози, що містять фермент з активністю АРАТ, об'єднують, потім зі швидкістю 2,5мл/хвил і при температурі 4°С завантажують безпосередньо в колонку об'ємом 7мл з холятом натрію, іммобілізованим на шариках сефарози (Pharmacia), попередньо урівноваженою буфером А, що містить 0,5моль хлориду натрію. Після промивання колонки тим самим буфером, фермент з активністю АРАТ елююють за допомогою 100мл лінійного градієнта холяту натрію, що змінюється від 0 до 1%, в тому самому буфері. Фракції, що містять фермент з активністю АРАТ, об'єднують, концентрують шляхом ультрафільтрування крізь мембрану УМ10 (Amicon) аж до концентрації протеїну 1,8мг/мл, потім зберігають при температурі -80°С. Фермент, що має активність АРАТ, у такий спосіб очищають приблизно 500 разів на основі питомої активності і з виходом близько 16%, як показано в поданій далі таблиці 1: Таблиця 1 Стадія Супернатант, 12000g Преципітація з цинком ДЕАЕ-сефацель Фенілсефароза Q-сефароза Холят-сефароза Активність АРАТ (одиниці) 8057 10453 6399 3316 1712 1300 Протеїн (мг) 42242 28040 5009 489 39 13 Питома активність (одиниць/мг) 0,19 0,37 1,28 6,78 43,9 100 Очистка Вихід 1 1,9 6,7 35,7 231 526 100 129 79 41 21 16 (де ДЕАЕ означає діетиламіноетил) Половину одержаного вище напівочищеного матеріалу (близько 6мг протеїну) потім розморожують, після чого додають ПЕГ 4000 (Prolabo) до кінцевої концентрації 20% (маса/об'єм) для вилучення холяту і хлориду натрію. Після перемішування протягом 30 хвилин при температурі 4°С преципітат збирають шляхом центрифугування з прискоренням 12000g протягом 30 хвилин при температурі 4°С, потім знову розчиняють в 4мл буферу А. Таким чином одержаний розчин потім зі швидкістю 1мл/хвил завантажують в колонку MonoP HR5/20 (Pharmacia), попередньо урівноважену 25мМ біс-Трис-буфером, рН=6,3. Фермент з активністю АРАТ елююють за допомогою буферу Polybuffer 74, рН=4,0 (Pharmacia). Фракції по 1мл збирають, кожну, в присутності 50мкл 2М буферу Трис-НСІ, рН=8,0, щоб обмежити інактивацію ферменту при кислому значенні рН. Фракції 14, 15 і 16 (Фіг.3(А)), що містять фермент з більш високою активністю АРАТ, яку визначають як зазначено вище, об'єднують, концентрують шляхом ультрафільтрування крізь мембрану YM10, потім зберігають при температурі -80°С до використання. Одержану таким чином активну фракцію піддають електрофорезу в 10%-вому поліакриламідному гелі з додецилсульфатом натрію. Кілька смуг виявляють шляхом забарвлення кумасі-блакитним з переважною смугою удаваної молекулярної маси 62кДа (Фіг.3(В)), що ідентична молекулярній масі, яку визначено шляхом фільтрування через гель Superose 6 (Pharmacia) і яка відповідає ферменту з активністю АРАТ. В - Властивості АΡАΤ а) Специфічність субстрату Згідно з вищезазначеним методом для ацетил-СоА і прегненолону, використовуючи різні ацильні донори або різні стероїдні субстрати, напівочищена АРАТ переносить ацетат в 3β-ол, дельта4- або дельта5-стероїди з порівнянною ефективністю, тоді як перенос є незначним у випадку естрогенів і недетектованим у випадку стеролів і з помітною перевагою для ацетил-СоА в якості ацильного донора. В наступній таблиці 2, в якій у випадку а) досліди здійснюють з 30мкмоль кожного випробовуваного стероїду і 100мкмоль [3Н]-ацетил-СоА, а для б) досліди здійснюють з 100мкмоль кожного ацильного донора і 30мкмоль [3Н]-прегненолону, подані одержані результати: Таблиця 2 Субстрат а) прегненолон 17α-ОН-прегненолон DHEA 4-прегнен-3βол-20-он 5β-прегнан-3βол-20-он 17β-естрадіол естрон холестерин ергостерин Відн. активність (%) Субстрат б) 100 ацетил-СоА 89 пропіоніл-СоА 104 бутирил-СоА 70 гексаноїл-СоА 1,8 олеоїл-СоА 5 2,5 0,6 0,4 Відн. активність (%) 100 33 16 18 не детектується б) Інгібування Активність АРАТ сильно інгібується реагентами сульфгідрильних груп, такими, як N-етилмалеімід і 5,5'дитіобіс(2-нітробензойна кислота). Інгібування є повним в присутності хлориду цинку (1ммоль). Γ - Часткова амінокислотна послідовність Часткову амінокислотну послідовність визначають після гідролізу трипсином на зрізах гелю згідно з методом, описаним Rosenfeld та ін. (1992). З двох третин концентрату, одержаного вище, потім підданого поділу шляхом електрофорезу в поліакриламідному гелі з додецилсульфатом натрію, вирізають смугу з молекулярною масою 62кДа, потім інкубують з трипсином (Promega), Одержані пептиди після цього поділяють шляхом ВЕРХ з оберненою фазою на колонці Vydac 218TP (1,5´125мм) зі швидкістю 100мкл/хвил, використовуючи лінійний градієнт ацетонітрилу, що змінюється від 0 до 60% протягом 80 хвилин, потім від 60 до 100% протягом 20 хвилин, в 0,1%-вому розчині трифтороцтової кислоти (ТФОК) у воді, потім проводять секвенування амінокислот. N-Кінцеву послідовність визначають в секвенаторі протеїнів, модель 477А, зв'язаному з ВЕРХаналізатором РТН-амінокислот (Applied Biosystems), З секвенованих зразків, два піки х і у представляють однозначну послідовність, утворену, відповідно, такими 10 і 16 амінокислотами; для піку х: ISEQFKKDDF; для піку у: LIELISPVIIPLGNPK, Обидві таким чином одержані пептидні послідовності використовують для "просіювання" бази даних геному S.cerevisiae. Протеїн з молекулярною масою 62кДа, послідовність якого містить точні послідовності двох вищезазначених пептидів, може бути ідентифікований (Mips, номер за каталогом S64491, Hebling та ін,, травень 1996). Цей протеїн, послідовність якого подано на Фіг.4 і функцію якого не описано, кодує ген в локусі YGR177c. На основі ідентичності приблизно на 37% амінокислотної послідовності між протеїном відповідно до винаходу, що має активність ацилтрансферази, і генним продуктом ATF1 у випадку S.cerevisiae, описаним Fujii та ін. (1994) і для якого виявлена спиртацетилтрансферазна активність, автори використовують позначку ATF2 для гена, що кодує протеїн, відповідальний за активність АРАТ у S.cerevisiae. Приклад 2 Конструювання штамів дріжджів, що містять ген ATF2, перерваний геном URA3, і позбавлених активності АРАТ (atf2-Δ::URA3). А) Ген-мішень ATF2 Ген URA3 S.cerevisiae вводять шляхом заміни вибраної частини гена ATF2 S.cerevisiae, що дозволяє здійснювати подальшу селекцію мутантних штамів шляхом протографії з урацилом. Селекційний маркер URA3 об'єднують з геном ATF2 шляхом подвійного злиття за рахунок полімеразної ланцюгової реакції (ПЛР) згідно з способом, описаним Amberg та ін., 1995. Така стратегія, яку подано на Фіг.5, включає в цілому 4 полімеразні ланцюгові реакції. Дві перші реакції (що їх позначено ПЛР1) дозволяють, відповідно, ампліфікувати ділянки 5' і 3', що фланкують інсерційний сайт маркера URA3 в перерваному генімішені ATF2, і третя реакція (що її позначено ПЛР2) дозволяє ампліфікувати ген-маркер URA3. В результаті подвійного злиття (яке позначено ПЛР3), нарешті, ділянки 5' і 3' гена-мішені ATF2 з'єднуються з геноммаркером URA3 (позначається як 5'ATF2-URA3-3'ATF2). Спочатку зразок цілих клітин штаму Fyl679, що використовується в якості джерела ДНК гена-мішені ATF2, ампліфікують в ПЛР-буфері, що містить 2ммоль dNTP (Pharmacia), за таких умов: 25 циклів; 93°С - 30 секунд; 54°С - 2 хвилини; 68°С - 3 хвилини; з подальшим подовженням на 5 хвилин при 72°С; полімераза Ampli Taq (Perkin Elmer). З одного боку, ділянку 5' гена ATF2 ампліфікують шляхом ПЛР, використовуючи в якості прямого і непрямого праймерів олігонуклеотиди, що мають такі послідовності: OTG10841 AAAAGTCGACAAAATGGAAGATATAGAAGGATACGAACeACATAICACTC (послідовність №1) OTG10844 ATCAATCTCCAATTAGGCCTCTTCGGATTACCC (послідовність №2) які містять ділянку, гомологічну ділянці 5' послідовності гена ATF2 (SGD:YGR177c), і з доданням сайта рестрикції Sall для OTG10841. З іншого боку, область 3' гена ATF2 ампліфікують шляхом ПЛР, використовуючи в якості прямого і непрямого праймерів олігонуклеотиди, що мають такі послідовності: OTG10846 CATTCGACATTCCCGAAGGTGACAATGACAA G (послідовність №3) OTG10842 AAAAACGCGTAACTATTAAAGCGACGCAAATTCGCCGATGGTTTGG (послідовність №4) які містять ділянки, гомологічні ділянці 3' послідовності гена ATF2 (SGD:YGR177c), і з доданням сайта рестрикції Mlul для OTG10842. Потім, ген URA3 S.cerevisiae ампліфікують шляхом ПЛР, використовуючи в якості прямого праймера олігонуклеотид, що має послідовність: GGGTAATCCGAAGAGGCCTAATTGGAGATTGATAAGCTTTTCAATTCAATTCATC OTG10843 (послідовність№5) ATTTTTTTTTTATTCTTTTTTTTG який містить послідовність, гомологічну ділянці 5' опублікованої послідовності гена URA3 [Rose та ін., 1984; банк генів; YSCODCD; номер за каталогом: КО2207; SGD: YEL021w], об'єднану з послідовністю, яка гомологічна ділянці 5' гена ATF2 (комплементарна OTG10844), і в якості непрямого праймера - олігонуклеотид, що має послідовність: CTTGTCATTGTCACCTTCGGGAATGTCGAATGGGGTAATAACTGATAT OTG10845 (послідовність №6) AATTAAATTGAACTC який містить послідовність, гомологічну ділянці 3' гена URA3, об'єднану з послідовністю, що гомологічна ділянці 3' гена ATF2 (комплементарна OTG10846). Зразок масою 20нг ДНК гена URA3, виділений з шаттлвектора Е.соlі - дріжджі pTG10021 (Degryse та ін., 1995) шляхом гідролізу за допомогою рестрикційного ферменту Hindlll, ампліфікують у вищезазначених умовах. Одержані, відповідно, продукти ПЛР очищають, використовуючи набір під назвою "Набір для очистки генів" (Віо 101 Inc., La Jolla, США), потім піддають реакції подвійного злиття, використовуючи в якості праймерів вищезазначені олігонуклеотиди OTG10841 і OTG10842, в умовах ампліфікації, що використовуються для попередніх полімеразних ланцюгових реакцій з програмою 20 циклів. Після очищення кінцевого продукту злиття, що містить фланкуючі ділянки гена ATF2, злиті з функціональним геном URA3, наявність гена URA3 підтверджують шляхом гідролізу за допомогою рестрикційного ферменту EcoRV, що показує присутність цього сайта в ампліфікованому матеріалі. Б) Генерація штамів дріжджів atf2-Δ::URA3 Одержаний вище продукт злиття трансформують безпосередньо в компетентних клітинах штаму Fy1679 або штаму IGY73.4 і трансформанти селекціонують шляхом вирощування в середовищі 3D (F.Sherman, 1991) в присутності необхідних живильних речовин штаму і за відсутності урацилу, У виділених клонах виявляють нову сполуку між ділянкою 5' гена ATF2 і гена URAS (5'ATF2-URA3-3'ATF2) шляхом ампліфікації за допомогою ПЛР в цілих клітинах, використовуючи вищезазначені праймери OTG10841 і OTG10845. Відсутність активності АРАТ потім визначають ін вітро згідно з описаним тестом, виходячи з клітинного гомогенату, в порівнянні з спорідненим штамом, що має чітко виражену активність АРАТ. Штами, що відповідають цим критеріям, таким чином були охарактеризовані як мутантні штами atf2, які позначаються atf2-Δ::URA3. Мутантний штам, одержаний таким чином з спорідненого штаму Fy167 9, позначають TGY156, а мутантний штам, одержаний з спорідненого штаму TGY73.4, позначають TGY158. Зразок штаму TGY156 депонований в Національній колекції культур мікроорганізмів. Інститут Пастера, 25, вулиця Docteur Roux, 75724, PARIS CEDEX 15, Франція, 2 лютого 1998p. під номером 1-1977. Зразок штаму TGY158 депонований в Національній колекції культур мікроорганізмів. Інститут Пастера, 25, вулиця Docteur Roux, 75724, PARIS CEDEX 15, Франція, 2 лютого 1998р., під номером l-1976. В) - Стабілізація прегненолону in vivo в культурах штамів дріжджів atf2-Δ::URA3 Клітини одержаного штаму TGY158 вносять, з розрахунку A600=0,1, в середовище YPD (Difco), що містить 100мкг/мл прегненолону. Після інкубації протягом 16 годин при температурі 28°С стероїди екстрагують дихлорметаном і аналізують за допомогою ВЕРХ з оберненою фазою, як вказано вище. З фігури б(В) видно, що мутант TGY158 позбавлений здатності етерифікувати прегненолон, тоді як в тих самих умовах культивування споріднений штам TGY73.4 перетворює прегненолон на прегненолонацетат (Фіг.6(А)). Ці результати дозволяють зробити висновок про те, що генний продукт ATF2 є відповідальним за етерифікацію прегненолону дріжджами, тоді як переривання гена ATF2 не приводить до помітних змін зростання клітин за нормальних умов. Приклад 3 Конструювання штаму дріжджів, що містись ген ATF2, перерваний геном URA3, і екепресує 3β-ΗSD Плазміди 2μ, які містять послідовність кДНК, що кодує людський 3β-HSD під контролем промотору CYC1 S.cerevisiae або промотору TEF1 S.cerevisiae і включають ген стійкості до G418, конструюють згідно з наведеною на Фіг.7А - 7С схемою, потім трансформують у мутантних штамах дріжджів atf2-Δ::URA3. Спочатку вектор переносу pTG10095, що містить послідовність ДНК, яка кодує людський 3β-HSD типу II, описану Е.Rheaume та ін., 1991, фланковану сайтами Sail і Mlul та розташовану нижче промотору дріжджів GAL10/CYC1, одержують так: Кодуючу 3β-HSD послідовність субклонують згідно з Е.Rheaume та ін., 1991, як рестрикційний фрагмент Sall-NotI в тих самих сайтах вектора Bluescript II (Stratagene). Одержаний вектор, описаний Е.Rheaume та ін., 1991, містить сайт NotІ, локалізований у кінця 3' послідовності, що кодує 3β-HSD. Цей вектор потім гідролізують за допомогою рестрикційного ферменту NotІ, після чого обробляють фрагментом Кленова в присутності dNTP, щоб заповнити "липкі" кінці, потім знову піддають лігатурі в присутності олігонуклеотиду, який має таку послідовність: OTG4461: CACACGCGTGTG (послідовність №7), попередньо фосфорилованого та гібридизованого з самим собою, щоб ввести сайт Mlul. Одержаний таким чином вектор pTGl0082 (Фіг.7А) містить послідовність, яка кодує 3β-HSD, що містить сайт Bglll і "облямовану" сайтами Sall і Mlul, тоді як сайт NotІ втрачений. Цей вектор містить також природну неко-дуючу ділянку 5' , що ідентифікується за наявністю сайта Bglll. Для того, щоб наблизити сайт Sall до ATG (антитимоцитарний глобулін) - ініціатору, вище якого хочуть ввести промотор GAL10/CYC1, вектор pTG10082 гідролізують за допомогою рестрикційного ферменту MscI, сайт якого знаходиться в некодуючій області 5' і безпосередньо перед ATG-ініціатором, і за допомогою рестрикційного ферменту Mlul. Фрагмент MscI-Mlul завдовжки 1,8т.н., що містить послідовність, яка кодує 3βΗSD (Фіг.7А) , виділяють, потім лігують у плазміді pTG10033 (E. Degryse та ін., 1995), що містить промотор GAL10/CYC1, попередньо гідролізований за допомогою рестрикційного ферменту Sall, після чого обробляють фрагментом Кленова в присутності dNTP і потім гідролізують за допомогою рестрикційного ферменту Mlul. Таким чином одержують вектор PTG10095. Потім конструюють рекомбінаційний вектор pTG10268, що містить плазміду 2μ дріжджів, реплікон Е.соlі, полігенний експресувальний кластер СУС1prom - PGKterm і селекційний маркер LEU2. Цей вектор ідентичний вектору pTG10159, описаному Е.Degryse та ін. (1995), за винятком сайта Xbal, що міститься в області 2μ, який замінено маркером Xbal°, одержуваним шляхом заповнення природного сайта Xbal в присутності фрагменту Кленова, потім шляхом повторної лігатури. Потім одержують плазміду експресії pTG10268 шляхом гомологічної рекомбінації за рахунок введення блоку експресії, що містить промотор GAL10/CYC1 p, що одержаний з сконструйованої вище плазміди pTG10095, потім гідролізованої за допомогою рестрикційного ферменту NotІ, в плазміду Ptg10260, попередньо гідролізовану за допомогою рестрикційних ферментів Sall і Mlul, Плазміда pTG10268 (Фіг.7В) містить послідовність, що кодує людський 3β-ΗSD типу II під контролем промотору СYС1. Потім плазміду експресії pTG10862, яка містить послідовність, що кодує 3β-HSD під контролем промотору TEF1, конструюють так (Фіг.7С): Спочатку конструюють плазміду pTG108 32 (Фіг.8) шляхом гомологічної рекомбінації між фрагментом NotI, одержаним з сконструйованої вище плазміди pTG10268, потім гідролізованої за допомогою рестрикційного ферменту NotI, і рекомбінаційною плазмідою pTG10164 (Ε.Degryse та ін., 1995), попередньо гідролізованою за допомогою рестрикційних ферментів Sall і Mlul. Потім одержують плазміду експресії pTG10862 шляхом введення промотору TEF1, що міститься в фрагменті Clal-Sall, виділеному з плазміди pTG10085 (Ε.Degryse та ін., 1995), замість промотору CYC1, усуненого шляхом гідролізу сконструйованої вище плазміди pTG10832 за допомогою рестрикційних ферментів СlаІ та Sall. Таким чином одержану плазміду pTGlO862 (Фіг.9) , що містить послідовність кДНК, яка кодує людський 3βHSD типу II, трансформують, відповідно, в спорідненому штамі TGY73/4 або в його мутанті atf2-Δ::URA3, який відповідає одержаному в прикладі 2 штаму TGY158, також, як в спорідненому штамі FY1679 або в його мутанті atf2-Δ::URA3, що відповідає одержаному в прикладі 2 штаму TGY156. Трансформанти виділяють як вказано вищев середовищі YPD (Difco), що містить 250мкг/мл G418. Далі одержані таким чином колонії-"кандидати" попередньо культивують у середовищі 3D, що містить необхідні для кожного штаму живильні елементи (гістидин і урацил для штаму TGY73.4; гістидин для TGY158; триптофан, гістидин, лейцин і урацил для штаму FY1679; триптофан, гістидин і лейцин для штаму TGY156; в концентрації по 100мкг/мл кожного), потім вносять в середовище, що містить 100мкг/мл прегненолону, Після культивування і біоконверсії протягом 24-х годин при температурі 28°С, стероїди екстрагують і аналізують за допомогою ВЕРХ з оберненою фазою, як вказано вище. Одержані результати, визначені у випадку трьох клонів кожного штаму, подані в наведеній далі таблиці 3: Таблиця 3 Прегненолон Прогестерон (мкг/мл) (мкг/мл) TGY73.4 PTG10862 1 0 TGY158 “ 94 15,2 FY1679 “ 1 1,3 TGY156 “ 100 13 Штам Плазміда Ці результати показують, що прегненолоновий субстрат практично повністю рекуперують у випадку мутантних штамів TGY156 або TGY158, в яких спостерігають початок біоконверсії прегненолону в прогестерон, тоді як зникнення прегненолону є повним в споріднених штамах TGY73.4 або FY1679, які не продукують або дуже мало продукують прогестерон, але акумулюють ацетат прегненолону, як це показано в прикладі 2. Зразок трансформованого штаму TGY158/pTG10862 депонований в Національній колекції культур мікроорганізмів. Інститут Пастера, 25, вулиця Docteur Roux 75724, PARIS CEDEX 15, Франція, 2 лютого 1998p., під номером l-1978. Приклад 4 Конструювання штаму дріжджів, що містить ген ATF2, перерваний за допомогою TEF1prom/PGKterm (atf2Δ::TEF1/PGK) Штам TGY186, що являє собою штам, який походить від штаму TGY156 (atf2-Δ::URA3) , описаного в прикладі 2, в якому ген URA3 в локусі ATF2 замінений блоком експресії TEF1prom/PGKterm, конструюють у такий спосіб: Спочатку блок експресії TEF1prom/PGKterm об'єднують з геном ATF2 шляхом подвійного злиття за допомогою ПЛР згідно з умовами, описаними в прикладі 2, але використовуючи блок експресії TEF1prom/PGKterm S.cerevisiae, описаний Ε.Degryse та ін. (1995), замість селекційного маркера URA3. Дві перші полімеразні ланцюгові реакції (ПЛР1), які дають змогу, відповідно, ампліфікувати кодуючі ділянки 5' і 3' гена ATF2, що фланкує інсерційний сайт блоку TEF1prom/PGKterm. У перерваному гені-мішені ATF2, здійснюють при використанні, відповідно, в якості прямого і непрямого праймерів для ділянки 5', олігонуклеотиди, що мають такі послідовності; OTG11049 CTCTCTGTCGACAAAATGGAAGATATAGAAGGATACGAACCACATАТСАСТС (послідовність №8) OTG10844 ATCAATCTCGAATTAGGCCTCTTCGGATTACCC (послідовність №9) а для ділянки 3' - олігонуклеотиди, які мають такі послідовності: OTG10846 CATTCGACATTCCCGAAGGTGACAATGACAAG (послідовність №10) OTG11050 AACAACACGCGTAACTATTAAAGCGACGCAAATTCGCCGATGCTTTGG (послідовність №11) Праймери OTG1104 9 і OTG11050 призначені для введення, відповідно, сайтів рестрикції Sall і Mlul. Третю полімеразну ланцюгову реакцію (ПЛР2), що дозволяє ампліфікувати блок TEF1prom/PGKterm, здійснюють при використанні, відповідно, в якості прямого і непрямого праймерів, олігонуклеотиди, які мають такі послідовності: GGGTAATCCGAAGAGGCCTAATTGGAGATTGATATCGATCACACACCATAGCTTC OTG11052 (послідовність№12) AAAATGTTTCTAC CTTGTCATTGTCACCTTCGGGAATGTCGAAGCTTCGAAACGCAGAATTTTCGAGTTАТТ OTG11053 (послідовність№13) АААСТТАА які вводять сайти рестрикції СlаІ та Hindlll. Нарешті, об'єднання здійснюють шляхом подвійного злиття одержаних вище продуктів ПЛР, використовуючи в якості праймерів олігонуклеотиди, що мають послідовності OTG11049 (послідовність №8) і OTG11050 (послідовність №11), які вводять сайти рестрикції Sall і Mlul в сайти зв'язування ATF2. Потім, після очищення, кінцевий продукт злиття рекомбінують з геном ATF2, що міститься в плазміді pTG10885, сконструйованій як вказано вище і попередньо гідролізованій за допомогою рестрикційних ферментів BstI і StuI. Таким чином одержують плазміду pTG10888, що містить сигнал TEF1prom/PGKterm в сайтах Clal та Hindlll, "облямований" фланкуючими ділянками гена ATF2. Одержання плазміди pTG10885 включає ампліфікацію гена ATF2 з штаму FY167 9 згідно з описаними в прикладі 2 умовами, але при використанні, відповідно, в якості прямого і непрямого праймерів вищезазначені олігонуклеотиди, які мають послідовності OTG11049 (послідовність №8) і OTG11050 (послідовність №11), які вводять сайти рестрикції Sall і Mlul. В одержаному продукті ПЛР ці сайти потім вилучають шляхом гідролізу за допомогою рестрикційних ферментів Sall і Mlul, потім шляхом обробки за допомогою фрагменту Кленова полімерази І Е.соlі, щоб заповнити "липкі" кінці. Одержаний фрагмент потім лігують в експресійному векторі pTG10031, описаному Е.Degryse та ін. (1995), попередньо гідролізованому за допомогою ферментів Clal та Hindlll, потім обробленому за допомогою фрагменту Кленова. Шляхом трансформації в Е.соli таким чином одержують плазміду pTG10885 в результаті лігування і сайта Sall продукту ПЛР, доповненого за рахунок використання фрагменту Кленова, щоб одержати послідовність GTCGA, з сайтом Hindlll вектора, який доповнений за рахунок використання фрагменту Кленова, щоб одержати послідовність AGCTT, так, щоб реконструювати сайт Hindlll (GTCGAAGCTT) (послідовність №14) та виключити сайт Clal. Сайт Clal вектора, доповнений за рахунок використання фрагменту Кленова, щоб одержати послідовність ATCG, зникає після лігування з продуктом ПЛР. Сигнал TEF1prom/PGKterm потім вилучають з плазміди pTG10888 у вигляді фрагменту NotI завдовжки 1,8т.н., потім заміняють маркером URA3 в штамі TGY156 (atf2-Δ::URA3). Одержаний в прикладі 2 і використовуваний в якості штаму-хазяїна штам TGY156 котрансформують з ДНК, вилученою з плазміди pTG10888, і з вектором дріжджів, що містять джерело ARS (СНІД-асоційований комплекс), що називається YRp7, описаний Struhl та ін. (1979), що дозволяє виконати потребу у триптофані штаму TGY156 і здійснювати селективну детекцію колоній за їх стійкістю до 5-фтороротової кислоти. 2-5мкг ДНК, вилученої з плазміди pTG10888 шляхом гідролізу за допомогою рестрикційного ферменту NotI, і плазміди YRp7 вводять в штам TGY156 за методом з ацетатом літію (Ito та ін., 1983), Потім здійснюють селекцію по виконанню потреби штаму в триптофані після утворення моношарів в чашках з агаром в середовищі YNGB (Difco), доповненому гістидином і лейцином (по 100мкг/мл кожного). Колонії-"кандидати", відібрані за допомогою зубочистки, потім приміщують в середовище, що містить 5-фтороротовую кислоту, яке одержують згідно з Boeke та ін, (1984), потім стійкість до 5-фтороротової кислоти підтверджують при використанні того самого середовища, причому можлива втрата вектора YKp7 в стійких до 5-фтороротової кислоти колоніях вказує на потребу в триптофані. Серед відібраних таким чином клонів об'єднання гена ATF2 з TEF1prom/PGKterm контролюється за допомогою ПЛР. Таким чином одержують штам TGY186. Приклад 5 Конструювання штаму дріжджів, що містить ген ATF2, перерваний за допомогою TEF1prom/PGKterm і експресує Р45017α Плазміду (pTG10435), що містить ділянку ініціації реплікації дріжджів ARSH4/CEN6, селекційний маркер URA3 і включає послідовність кДНК, що кодує бичачий цитохром P45017α під контролем промотору TEF1 S.cerevisiae, конструюють відповідно до наведеної на фігурі 10 схеми, потім трансформують в мутантному штамі дріжджів atf2-Δ::TEF1/PGK (TGY186). Спочатку плазміду pTG10058, що містить послідовність кДНК, яка кодує бичачий цитохром Р 45017α, описаний Zuber та ін, (1986), фланковану сайтами Sall та Mlul і розташовану нижче промотору дріжджів CYC1, одержують згідно з наведеною на Фіг.11 схемою: Плазміду pGB17α-5, яку описано в міжнародній заявці на патент 89/10963 і яка містить послідовність, що кодує бичачий цитохром Ρ45017α, розкривають шляхом гідролізу за допомогою рестрикційного ферменту Xhol, потім обробляють лужною фосфатазою. Після фосфорилування і гібридизації, олігонуклеотиди, які мають такі послідовності: OTG4511 TCGACGGACGCGTGG (послідовність №15) OTG4512 TCGACCACGCGTCCG (послідовність №16) вводять в сайт Xhol з одержанням плазміди pTG10104. Плазміду pTG10104 потім обробляють рестрикційними ферментами Sail і Міні, потім вводять в плазміду pTG10031, описану Е. Degryse та ін, (1995), що містить промотор дріжджів CYC1, попередньо гідролізовану за допомогою рестрикційних ферментів Sall і Mlul і оброблену лужною фосфатазою. Таким чином одержують вектор pTG10058, що містить кДНК, яка кодує бичачий цитохром Р45017α (Фіг.12). Плазміду pTG10058 потім гідролізують за допомогою рестрикційних ферментів Sall та Mlul і обробляють лужною фосфатазою. Фрагмент Sall-Mlul зовдовжки 1,7т.н., що містить послідовність, яка кодує бичачий цитохром P45017α, виділяють, потім лігують в експресійному векторі pTG10085, який описано Е.Degryse та ін. (1995) і який містить промотор дріжджів TEF1, попередньо гідролізованому за допомогою ферментів Sall і Mlul. Таким чином одержують плазміду pTG10293 (Фіг.13), в якій кодуюча цитохром Р45017α послідовність перебуває під контролем промотору TEF1. Потім плазміду експресії pTG10435 одержують шляхом гомологічної рекомбінації між рекомбінаційною ллазмідою PTG10434, описаною Е.Degryse та ін. (1995) і тією, яка містить послідовність ARSH4/CEN6, попереденьо гідролізованою за допомогою ферментів Sall і Mlul, і фрагментом NotІ завдовжки 2,8т.н., одержаним з сконструйованої вище плазміди PTG10293. Одержану таким чином плазміду pTG10435 (Фіг.14), що містить послідовність, яка кодує бичачий цитохром Р45017α під контролем промотору TEF1, потім трансформують, відповідно, в спорідненому штамі FY1679 або в штамі TGY186 (atf2-Δ::TEF1/PGK) , одержаному в прикладі 4. Трансформанти виділяють як вказано вище в середовищі YNBG (Difco), доповненому триптофаном, гістидином і лейцином (по 100мкг/мл кожного). Потім одержані таким чином колонії попередньо культивують протягом 16 годин в середовищі YNB (Difco), що містить 2% глюкози і 0,5% cas-амінокислот, після чого розбавляють свіжим середовищем до А600=0,2. Після вирощування протягом 6 годин додають 100мкг/мл прегненолону або прогестерону. Після інкубації і біоконверсії протягом 48 годин при температурі 28°С стероїди екстрагують і аналізують за допомогою ВЕРХ з оберненою фазою, як вказано в прикладі 1, використовуючи, відповідно, стандарти прегненолону і 17α-гідроксипрегненолону або прогестерону і 17αгідроксипрогестерону. Одержані результати, виражені в мкг/мл, подано в таблиці 4: Таблиця 4 Штам Субстрат: прегненолон прегненолон 17α-гідроксипрегненолон Субстрат: прогестерон прогестерон 17α-гідроксипрогестерон FYl679/ TGY186/ pTG10435 pTG10435 0 0 67,1 42,0 46,5 29,3 41,4 33,1 Ці результати показують, що здатність до біоконверсії цитохрому Р45017α, експресія якого здійснюється вектором pTG10435, майже така сама в штамі дикого типу (FY) або його мутанті atf2 (TGY186) при використанні прогестерону в якості субстрату. З іншого боку, при використанні прегненолону в якості субстрату досягають такої ж конверсії порівняно з прогестероном, але тільки з мутантом atf2 (TGY186). В штамі дикого типу FY водночас субстрат і продукт ацетиловані, і вільний гідроксипрогестерон не виявляється. Зразок трансформованого штаму TGY186/pTG10435 депонований в Національній колекції культур мікроорганізмів, Інститут Пастера, 25, вулиця Docteur Roux 75724, PARIS CEDEX 15, Франція, 20 січня 1999p. під номером l-2119. Приклад 6 Конструювання штаму дріжджів, що містить ген ATF2, перерваний за допомогою TEF1prom/PGKterm, і коекспресуе 3β-HSD і Ρ45017α Плазміду pTG10417 (Фіг.21), що містить реплікон дріжджів 2μ і два блоки експресії, з яких один кодує людський 3β-HSD, а іншій кодує бичачий цитохром Р45017α, причому обидва перебувають під контролем промотору СУСl S.cerevisiae і містять селекційний маркер URA3, конструюють послідовно згідно з описаними нижче стадіями 1-3 (Фіг.15А і В), потім описаними нижче стадіями 4-6, після чого трансформують в мутантному штамі дріжджів atf2-Δ::TEF1prom/PGKterm (TGY186). Стадія 1: конструювання плазміди pTG10210 Експресійний вектор pTG10033, описаний Е.Degryse та ін. (1995) і який містить гібридний промотор дріжджів GAL10/CYC1 , попередньо гідролізований за допомогою рестрикційного ферменту PvuII, обробляють лужною фосфатазою, потім знову лігують в присутності олігонуклеотиду, що має таку послідовність: OTG1050: CCCGAATTCGGG (послідовність№17) попередньо фосфорилованого і гібридизованого з самим собою, щоб ввести сайти EcoRI у вигляді "облямівки" блоку експресії, що містить промотор GAL10/CYC1. Таким чином одержують вектор pTG10210. Стадія 2: конструювання плазміди pTG10214 Блок експресії, що містить промотор GAL10/CYC1, наявний у векторі pTG10210, потім вводять в шатлвектор Е.соlі - дріжджі PTG10013, описаний Е.Degryse та ін. (1995) і який містить селекційний маркер URA3-d. Вектор pTG10013, після гідролізу за допомогою рестрик-ційного ферменту EcoRI і обробки лужною фосфатазою, лігують з одержаним в стадії 1 вектором pTG10210, попередньо гідролізованим за допомогою ферменту EcoRI. Одержаний таким чином вектор pTG10214 містить блок експресії, що включає промотор GAL10/CYC1, спрямований до реплікону 2μ. Стадія 3: конструювання плазміди pTG10274 В плазміді pTG10214 промотор GAL10/CYC1 замінюють промотором CYC1 шляхом гомологічної рекомбінації після вилучення плазміди за рахунок обробки рестрикційними ферментами СlаІ і Sall. Експресійний вектор pTG10031, описаний Е.Degryse та ін. (1995) і який містить промотор CYC1, гідролізують за допомогою рестрикційних ферментів HindlІІ і Fspl, потім рекомбінують з плазмідою pTG10214, одержаною в стадії 2 і попередньо гідролізованою за допомогою рестрикційних ферментів СlаІ і Sall, одержуючи таким чином плазміду pTG10274 (Фіг.16). Стадія 4: конструювання плазнідн pTGWAOl З плазміди pTG10274, що містить промотор CYC1, і плазміди pTG10293, що містить послідовність, яка кодує цитохром Р45017α під контролем промотору TEF1, шляхом гомологічної рекомбінації одержують нову плазміду pTG10401, що містить послідовність, яка кодує цитохром P45017α під контролем промотору TEF1. Промотор CYC1 і частину реплікону Е.соlі вилучають з плазміди pTG10274, одержаної в стадії 3, шляхом гідролізу за допомогою рестрикційних ферментів Mlul і Dral. Одержану в прикладі 5 плазміду pTG10293 гідролізують за допомогою рестрикційних ферментів Hindlll і PvuII, потім рекомбінують з плазмідою pTG10274, гідролізованою за допомогою рестрикційних ферментів Mlul і Dral, одержуючи плазміду pTG10401 (Фіг.17). Стадія 5: конструювання плазміди pTG10403 кДНК, що кодує людський 3β-HSD типу II, вводять в плазміду pTG10401 під контролем промотору CYC1. Спочатку вектор експресії pTG10262, що містить кДНК, яка кодує людську 3β-HSD типу II, конструюють згідно з наведеною на Фіг.18 схемою, з вектора-переносника pTG10095, одержаного в прикладі 3, і рекомбінаційного вектора pTG10257, що містить реплікон дріжджів 2μ, реплікон Е.соlі, полігенний експресувальний кластер дріжджів CYC1prom-PGKterm і селекційний маркер URA3-d. Цей вектор pTG10257 ідентичний рекомбінаційному вектору pTG10042, описаному Е.Degryse та ін. (1995), за винятком сайта Xbal, що міститься в області 2μ, який замінений маркером Xbal°, одержаним шляхом доповнення природного сайта Xbal фрагментом Кленова, потім повторної лігатури. Блок експресії людського 3β-ΗSO типу II вилучають з век-тора-переносника pTG10095 шляхом гідролізу за допомогою рестрикційного ферменту NotІ, потім вводять в рекомбінаційний вектор Ptg10257, попередньо гідролізований за допомогою рес-трикційних ферментів Sall і Mlul, одержуючи вектор експресії pTG10262 (Фіг.19). Слід зазначити, що внаслідок рекомбінації блоку GAL10/CYC1-кДНК, що походить з плазміди pTG10095, в рекомбінаційному векторі pTG10257, що містить промотор CYC1, одержують вектор експресії, який містить блок CYC1-кДНК. Потім сконструйований вище вектор експресії pTG10262 гідролізують за допомогою рестрикційного ферменту XmnI. Одержаний фрагмент, що містить кДНК, яка кодує 3β-HSD, рекомбінують з фрагментом плазміди pTG10401, одержаної в стадії 4, що містить кДНК, яка кодує цитохром Р45017α, в свою чергу одержаний шляхом гідролізу за допомогою рестрикційного ферменту Scall. Одержана таким чином плазміда pTG10403 (Фіг.20) містить два блоки експресії, з яких один кодує 3β-HSD під контролем промотору CYC1, а інший кодує цитохром Р45017α під контролем промотору TEF1. Стадія 6: конструювання ллазміди pTG10417 Нарешті, у вищезазначеній плазміді pTG10403 промотор TEF1 заміняють промотором CYC1. З одного боку, описану в прикладі 5 плазміду pTG10058 гідролізують за допомогою рестрикційного ферменту PvuII, що дає змогу вивільнювати частину послідовності, що кодує цитохром P45017α, асоційованої з промотором CYC1, також, як переважну частину реплікону Е.соlі. З іншого боку, частину реплікону Е.соlі вилучають з одержаної в стадії 5 плазміди pTG10403 шляхом гідролізу за допомогою рестрикційного ферменту Dral. Шляхом рекомбінації двох плазмід, попередньо гідролізованих таким чином, нарешті, одержують плазміду pTG10417. Плазміда pTG10417 містить реплікон дріжджів 2μ, селекційний маркер URA3-d і два блоки експресії, з яких один кодує людський 3β-HSD, а іншій кодує цитохром Р45017α бичачого походження, причому обидва перебувають під контролем промотору дріжджів CYC1 (Фіг.21). Плазміду pTG10417 потім трансформують, відповідно, в спорідненому штамі FY1679 або в штамі TGY186 (atf2-Δ::TEF1prom/PGKterm), одержаному в прикладі 4. Трансформанти виділяють в агаровому середовищі YNB (Difco), що містить 0,5% глюкози та яке доповнено триптофаном, гістидином і лейцином (по 100мкг/мл кожного). Одержані таким чином колонії потім попередньо культивують протягом 24-х годин при температурі 28°С в середовищі YNB (Difco), що містить 0,5% глюкози і 0,1% cas-амінокислот, потім розбавляють до А600=0,1 і додають 100мг/мл прегненолону, при використанні або того самого свіжого середовища (середовище 1), або середовища YNB (Difco), що містить 0,1% глюкози, 2% гліцерину і 0,2% cas-амінокислот (середовище 2). Після культивування і біоконверсії протягом 48 годин стероїди екстрагують і аналізують за допомогою ВЕРХ з оберненою фазою як вказано в прикладі 1, використовуючи стандарти прегненолону, 17αгідроксипрегненолону, прогестерону і 17α-гідроксипрогестерону. Одержані результати, виражені в мкг/мл, подано в наведених далі таблиці 5 (середовище 1) і таблиці 6 (середовище 2): Таблиця 5 Штам прегненолон прегненолонацетат 17α-гідроксипрегненолон 17α-гідроксипрегненолонацетат прогестерон FY1679/ TGY186/ рТG10417pTG10417 0 58,1 87,3 0 0 3,1 0 10,6 0 1,4 17α-гідроксипрогестерон 1,3 23,4 Таблиця 6 Штам прегненолон прегненолонацетат 17α-гідроксипрегненолон 17α-гідроксипрегненолонацетат прогестерон 17α-гідроксипрогестерон FY1679/ TGY186/ pTG10417pTG10417 0 54,0 52,1 0 0 3,4 12,8 0 8,6 0 1,4 31,4 Ці результати показують, що біоконверсія в штамі дикого типу (FY), трансформованому плазмідою pTG10417, приводить до акумуляції прегненолонацетату і 17α-гідроксипрегненолонаце-тату, які надалі не трансформуються ферментами 3β-HSD або P45017α, і баланс біоконверсії гірший від такого, що спостерігається при використанні тарнсформованого мутанта atf2 (TGY186). Зразок трансформованого штаму TGY186/pTG10417 депонований в Національній колекції культур мікроорганізмів, Інститут Пастера, 25, вулиця Docteur Roux 75724, PARIS CEDEX 15, 20 січня 1999p. під номером l-2118. Бібліографічні посилання - Achstetter Т., Nguyen-Juilleret M., Findeli Α., Merkamm Μ. and Lemoine Υ. (1992), Gene 110, 25-31. - Amberg D.C., Botstein D. and Beasley E.M., (1995), Yeast 11: 1275-1280. - Boeke Jef D., Lacroute F. and Fink G R., (1984) Моl Gen Genet 197: 345-346. - Cauet G., Dumas В., Degryse E., Spagnoli R. and Achstetter T. (1994) in Cytochrome P-450 biochemistry, biophysics and molecular biology (Lechner M.C., ed.) pp.583-586, John Libbey Eurotext. - Cherry J.M., Adier C., Ball C., Dwight S., Chervitz S., Jia Y., Juvik G., Roe Т., Weng S. and Botstein D., "Saccharomyces Genome Database" http://genome-www.stan-ford.edu/Saccharomyces/. - Degryse E., J.Biotech. 39 (1995) 181-187. - Degryse E., Dumas В., Dietrich M., Laruelle L. and Achstetter T. (1995). Yeast 11: 629-640. - Degryse E. (1996), Gene 170, 45-50. - Dumas В., Cauet G., Degryse E., Spagnoli R. & Achtetter T. (1994). Cytochrome P450. 8 th International Conference. Ed. M.C.Lechner. John Libbey Eurotext, Paris pp.527-530. - Fiechter Α., Fuhrmann G.F. and Kappeli O., (1981). Adv. Microb. Physiol. 22, 123-183. - Fujii Т., Nagasawa Ν., Iwamatsu Α., Bogaki Т., Tamai Y. and Hamachi M. (1994) Appl. Environ. Microbiol, 60, 2786-2792. - Hanahan D., J.Моl. Biol. 166 (1983) 557-580. - Hubacek J. and Glover S.W., (1970), J. Моl. Biol. 50: 111-127. - Ito H., Fukuda Y., Murata K. and Kimura Α., 1983, Journal of Bacteriology, 163-168. - Rheaume E., Lachance Y., Zhao H.-F., Breton N., Dumont M., de Launoit Y., Trudel C., Luu-The V., Simard J. & Labrie F. (1991), Моl. Endocrinol. 5, 1147-1157. - Rose M., Grisafi P. and Botstein D., Gene 29, 113-124 (1984). - Rosenfeld J., Capdevielle J., Guillemot J.C. and Ferrara P. (1992) Anal. Biochem. 203, 173-179. - Sambrook J., Fritsch E.F, and Maniatis Т., (1989). Molecular Cloning: A Laboratory Manual. Cold Spring Harbor University Press, 2nd edition, Cold Spring Harbor. Sherman F. (1991) Methods in Enzymology 194, 3-21. - Simard J., Durocher F., Mebarki P., Turgeon C, Sanchez R., Labrie Y., Couet J., Trudel C, Rheaume E., Morel Y., Luu-The V. And Labrie F. (1996) J. Endocrinol. 150, 5189-5207 - Struhl K,, Stinchomb DT., Scherer S. and Davis RW., 1979, Proc. Natl. Acad. Sci. USA Vol.76, №3, 1035-1039. - Thierry Α., Fairhead C. and Dujon В., Yeast; Vol.6: 521-534 (1990). - Zuber MX., Simpson ER. and Waterman MR., (1986a) Science 234, 1258-1261. Список послідовностей де: Хехст Маріон Руссель Штами дріжджів, що містять перерваний ген ATF2, та їх застосування 2482 РСТ Послідовності у Франції ДНК штучна послідовність ознака помилки комплемент олігонуклеотид
ДивитисяДодаткова інформація
Назва патенту англійськоюYeast strains containing broken atf-2 gene and use thereof
Назва патенту російськоюШтаммы дрожжей, которые содержат прерванный ген ат2, и их применение
МПК / Мітки
МПК: C12N 9/10, C12N 1/19, C12P 33/00, C12N 15/54, C12N 15/09
Мітки: 3b-гідроксистероїди, окислення, одержання, здатного, модифікованого, трансформованого, штаму, субстрату, стабілізувати, спосіб, дріжджів
Код посилання
<a href="https://ua.patents.su/26-72438-sposib-oderzhannya-modifikovanogo-shtamu-drizhdzhiv-zdatnogo-stabilizuvati-3b-gidroksisterodi-sposib-oderzhannya-transformovanogo-shtamu-drizhdzhiv-ta-sposib-okislennya-substratu.html" target="_blank" rel="follow" title="База патентів України">Спосіб одержання модифікованого штаму дріжджів, здатного стабілізувати 3b-гідроксистероїди, спосіб одержання трансформованого штаму дріжджів та спосіб окислення субстрату</a>
Штам дріжджів saccharomyces cerevisiae для одержання сушених хлібопекарських дріжджів

Номер патенту: 36624
Опубліковано: 16.06.2003
Автори: Рудніченко Людмила Вікторівна, Левандовський Леонід Вікторович, Ткаченко Алла Феодосіївна, Хоменко Анатолій Іванович, Коваль Катерина Олександрівна, Олійнічук Сергій Тимофійович, Шустікова Наталія Євгенівна
МПК: C12N 1/18, C12R 1/865
Мітки: хлібопекарських, сушених, одержання, cerevisiae, saccharomyces, дріжджів, штам
Формула / Реферат:
Штам дріжджів Saccharomyces cerevisiae ІMB У-5009 для одержання сушених хлібопекарських дріжджів.
Штам дріжджів saccharomyces cerevisiae хл-1 для одержання хлібопекарських дріжджів

Номер патенту: 30677
Опубліковано: 15.03.2002
Автори: Шустікова Наталія Євгенівна, Коваль Катерина Олександрівна, Олійнічук Сергій Тимофійович, Рудніченко Людмила Вікторівна, Левандовський Леонід Вікторович, Ткаченко Алла Феодосіївна
МПК: C12R 1/865, C12N 1/18
Мітки: cerevisiae, одержання, хл-1, штам, хлібопекарських, saccharomyces, дріжджів
Формула / Реферат:
Штам дріжджів Saccharomyces cerevisiae ХЛ-1 для одержання хлібопекарських дріжджів.
Штам дріжджів saccharomyces cerevisiae imb-у-5016 для одержання пресованих хлібопекарських дріжджів в двопродуктовому виробництві спирту і хлібопекарських дріжджів

Номер патенту: 47821
Опубліковано: 15.01.2004
Автори: Левандовський Леонід Вікторович, Коваль Катерина Олександрівна, Ткаченко Алла Феодосіївна, Хоменко Анатолій Іванович, Бейко Наталія Євгенівна, Рудніченко Людмила Вікторівна
МПК: C12R 1/865, C12N 1/18
Мітки: двопродуктовому, одержання, штам, imb-у-5016, хлібопекарських, saccharomyces, пресованих, спирту, cerevisiae, дріжджів, виробництві
Формула / Реферат:
Штам дріжджів Saccharomyces cerevisiae ІМВ-У-5016 для одержання хлібопекарських дріжджів в двопродуктовому виробництві спирту і хлібопекарських дріжджів.
Спосіб одержання правастатину з компактину за допомогою штаму micromonospora

Номер патенту: 67854
Опубліковано: 15.07.2004
Автори: Ілкой Єва, Мозес Юлія, Йєккел Антоніа, Борош Шандор, Коніа Аттіла, Нагі Жужанна, Амбрус Габор, Барта Іштван, Хорват Ільдіко, Сомодьі Дьордь, Салат Янош, Хорват Дьюла, Шабо Іштван Міхай
МПК: C12P 7/62, C12P 7/40, C12P 17/02
Мітки: компактину, допомогою, штаму, правастатину, одержання, micromonospora, спосіб
Формула / Реферат:
1. Мікробний спосіб одержання сполуки формули (І) зі сполуки загальної формули (II),де R може бути лужним металом або іоном амонію,за допомогою зануреної культури штаму Micromonospora, котрий здатний
Спосіб виробництва пресованих хлібопекарських дріжджів з меляси

Номер патенту: 38449
Опубліковано: 15.09.2004
Автори: Хоменко Анатолій Іванович, Левандовський Леонід Вікторович, Ткаченко Алла Феодосіївна, Коваль Катерина Олександрівна, Олійнічук Сергій Тимофійович, Рудніченко Людмила Вікторівна, Янчевський Віктор Казимирович, Ткаченко Любов Володимирівна
МПК: C12R 1/865, C12N 1/18
Мітки: пресованих, спосіб, меляси, хлібопекарських, виробництва, дріжджів
Формула / Реферат:
Спосіб виробництва пресованих хлібопекарських дріжджів з меляси, який включає приготування розчину меляси з додаванням поживних речовин, вирощування на вказаному розчині чистої культури і товарних дріжджів в декілька стадій з використанням штаму Saccharomyces cerevisiae, виділення дозрілих дріжджів з культуральної рідини та їх пресування і формування, який відрізняється тим, що для одержання пресованих хлібопекарських дріжджів використовують...
Попередній патент: Пристрій для перевантаження сільськогосподарських матеріалів із збиральної машини на транспортний контейнер
Наступний патент: Фармацевтична композиція, що має активність пригнічувати апетит (варіанти)
Випадковий патент: Спосіб відновлення деталей машин