Процес утворення імуногенного кон’югату, імуногенний кон’югат білка ab















































Формула / Реферат
1. Процес утворення імуногенного кон'югату, який включає у себе стадії:
(a) введення реактивної групи у амінокислотний залишок пептидного імуногена, де пептидний імуноген - це пептид Аb або його фрагмент чи аналог;
(b) дериватизації однієї чи більше функціональних груп білка-носія для створення активованої функціональної групи на білку-носії;
(c) проведення реакції пептидного імуногена за стадією (а) з білком-носієм за стадією (b) в таких умовах, щоб утворити кон'югат, де реактивна група пептидного імуногену ковалентно прикріплена до активованої функціональної групи на білку-носії; і
(d) проведення подальшої реакції кон'югата за стадією (с) з кепінг-реагентом для того, щоб інактивувати будь-яку з решти активованих функціональних груп на білку-носії, щоб створити імуногенний кон'югат.
2. Процес за п. 1, який відрізняється тим, що білок-носій вибирають із групи, що складається із альбуміну сироватки людини, гемоціаніну лімфи слимака (KLH), молекул імуноглобуліну, тироглобуліну, овальбуміну, гемаглютиніну грипу, PADRE поліпептиду, циркумспорозитного (CS) білка малярії, поверхневого антигена гепатиту В (HBSAg19-28), білка теплового шоку (HSP) 65, Mycobacterium tuberculosis, токсину холери, мутантів токсину холери зі зниженою токсичністю, токсину дифтерії, білка CRM197, що є перехресно-реактивним з токсином дифтерії, рекомбінантної стрептококової С5а пептидази, Streptococcus pyogenes ORF1224, Streptococcus pyogenes ORF1664, Streptococcus pyogenes ORF2452, пневмолізину Streptococcus pneumoniae, мутанти пневмолізину зі зниженою токсичністю, Chlamydia pneumoniae ORF T367, Chlamydia pneumoniae ORF T858, правцевого токсоїду, ВІЛ gp120 T1, компонентів, що розпізнають молекули адгезивної матриці мікробної поверхні (MSCRAMMS), факторів і гормонів росту, цитокінів і хемокінів.
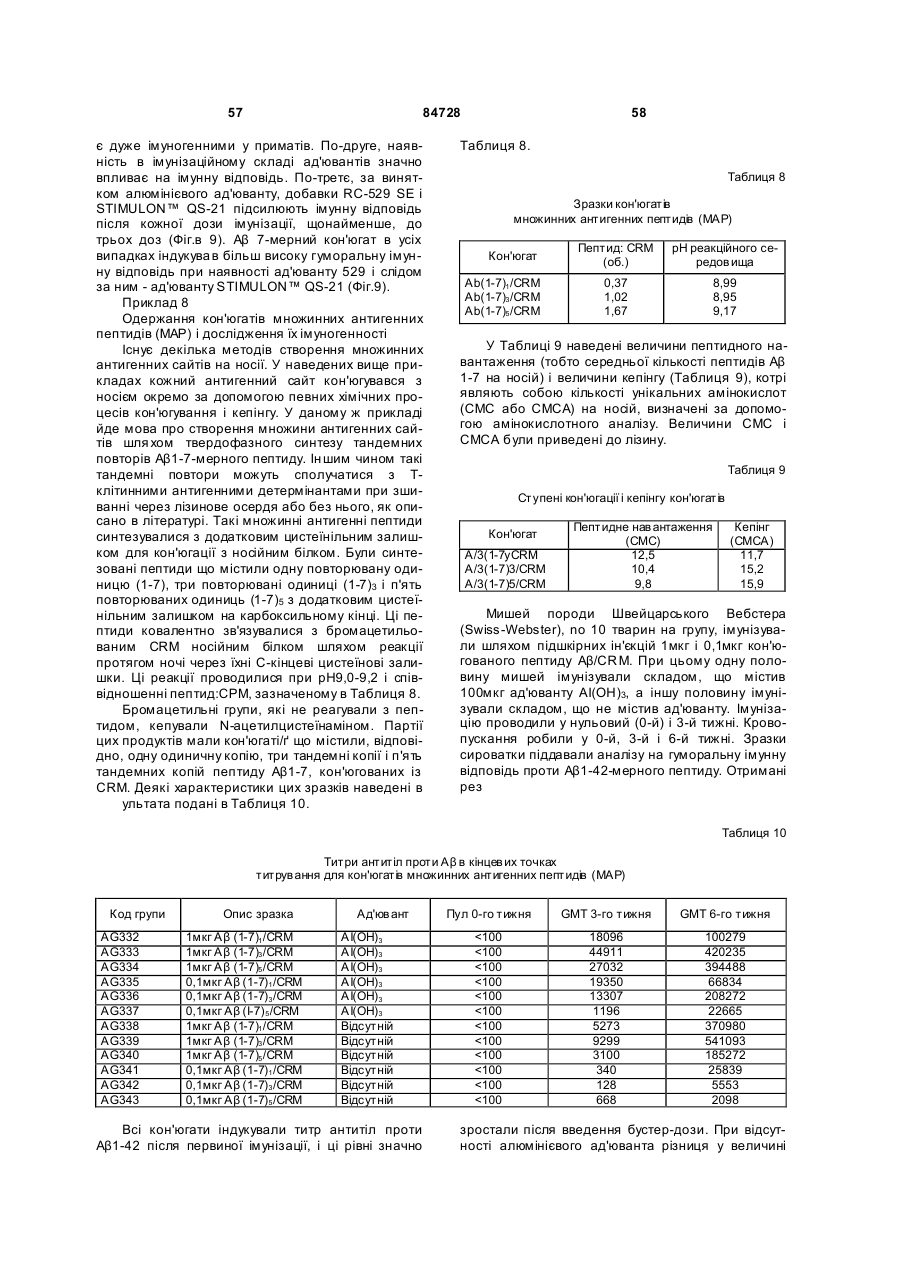
3. Процес за п. 2, який відрізняється тим, що білком-носієм є CRM197.
4. Процес за п. 1, який відрізняється тим, що пептидним імуногеном є фрагмент пептиду Аb.
5. Процес за п. 4, який відрізняється тим, що фрагмент пептиду Аb вибирають із групи, що складається із Аb1-3, 1-4, 1-5, 1-6, 1-7, 1-9, 1-10, 1-11, 1-12, 1-16, 1-28, 3-6, 3-7, 13-28, 15-24, 16-22, 16-23, 17-23, 17-24, 18-24, 18-25, 17-28, 25-35, 33-42, 35-40 і 35-42.
6. Процес за п. 1, який відрізняється тим, що функціональну групу білка-носія дериватизують за допомогою зшивального реагенту.
7. Процес за п. 6, який відрізняється тим, що функціональну групу дериватизують з галоацетилювальним агентом.
8. Процес за п. 1, який відрізняється тим, що кепінг-реагент, який використовують для дезактивації будь-якої активованої функціональної групи білка-носія, вибирають із групи реагентів, що складається із цистеїнаміну, N-ацетилцистеїнаміну, етаноламіну, гідроксиду натрію, карбонату натрію, бікарбонату амонію й аміаку.
9. Імуногенний кон'югат, що містить пептидний імуноген, ковалентно приєднаний до білка-носія, який відрізняється тим, що білок-носій має таку формулу:
![]() ,
,
де
С є білком-носієм, а X є перетворювана функціональна група амінокислотного залишку білка-носія, і де m є ціле число більше 0, але не більше 85, і де імуногенний кон'югат має таку формулу:
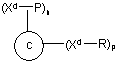 ,
,
де
С є білком-носієм, a Xd є дериватизована функціональна група амінокислотного залишку білка-носія, і де Р є пептидний імуноген, що містить пептид Аb або фрагменти пептиду Аb чи їх аналоги, ковалентно приєднані через реактивну групу амінокислотного залишку пептидного імуногену до дериватизованої функціональної групи амінокислотного залишку білка-носія, R є кепінг-молекула, ковалентно приєднана до дериватизованої функціональної групи амінокислотного залишку білка-носія, захищаючи цим функціональність білка-носія таким чином, що він зберігає свою здатність викликати бажані імунні відповіді проти пептидного імуногена, який містить пептид Аb або фрагменти пептиду Аb чи їх аналоги, що без носія було б неможливим, n є ціле число більше 0, але не більше 85, і р є ціле число більше 0, але менше ніж 85.
10. Імуногенний кон'югат за п. 9, який відрізняється тим, що білок-носій вибирають із групи, що складається із альбуміну сироватки, гемоціаніну лімфи слимака (KLH), молекул імуноглобуліну, тироглобуліну, овальбуміну, гемаглютиніну грипу, PADRE поліпептиду, циркумспорозитного (CS) білка малярії, поверхневого антигена гепатиту В (HBSAg19-28), білка теплового шоку (HSP) 65, Mycobacterium tuberculosis, токсину холери, мутантів токсину холери зі зниженою токсичністю, токсину дифтерії, білка CRM197, що є перехресно-реактивним з токсином дифтерії, рекомбінантної стрептококової С5а пептидази, Streptococcus pyogenes ORF1224, Streptococcus pyogenes ORF1664, Streptococcus pyogenes ORF2452, пневмолізину Streptococcus pneumoniae, мутантів токсину пневмолізину зі зниженою токсичністю, Chlamydia pneumoniae ORF T367, Chlamydia pneumoniae ORF T858, правцевого токсоїду, ВІЛ gp120 T1, компонентів, що розпізнають молекули адгезивної матриці мікробної поверхні (MSCRAMMS), факторів росту, гормонів, цитокінів і хемокінів.
11. Імуногенний кон'югат за п. 10, який відрізняється тим, що білком-носієм є CRM197.
12. Імуногенний кон'югат за п. 9, який відрізняється тим, що пептидним імуногеном є фрагмент пептиду Аb.
13. Імуногенний кон'югат за п. 12, який відрізняється тим, що фрагмент пептиду Аb вибирають із групи, що складається із Аb1-3,1-4, 1-5,1-6,1-7, 1-9, 1-10, 1-11, 1-12,1-16, 1-28, 3-6, 3-7, 13-28, 15-24, 16-22, 16-23, 17-23, 17-24, 18-24, 18-25, 17-28, 25-35, 33-42, 35-40 і 35-42.
14. Імуногенний кон'югат, який створений у відповідності з процесом за п. 1 і має формулу:
,
де
С є білком-носієм, a Xd є дериватизована функціональна група амінокислотного залишку білка-носія, і де Р є пептидний імуноген, що містить пептид Аb або фрагменти пептиду Аb чи їх аналоги, ковалентно приєднані через реактивну групу амінокислотного залишку пептидного імуногену до дериватизованої функціональної групи амінокислотного залишку білка-носія, R є кепінг-молекула, ковалентно приєднана до дериватизованої функціональної групи амінокислотного залишку білка-носія, захищаючи цим функціональність носія таким чином, що він зберігає свою здатність викликати бажані імунні відповіді проти пептидного імуногена, що без носія було б неможливим, n є ціле число більше 0, але не більше 85, і р є ціле число більше 0, але менше 85.
15. Імуногенний кон'югат за п. 14, який відрізняється тим, що білок-носій вибирають із групи, що складається із альбуміну сироватки людини, гемоціаніну лімфи слимака (KLH), молекул імуноглобуліну, тироглобуліну, овальбуміну, гемаглютиніну грипу, PADRE поліпептиду, циркумспорозитного (CS) білка малярії, поверхневого антигена гепатиту В (HBSAg19-28), білка теплового шоку (HSP) 65, Mycobacterium tuberculosis, токсину холери, мутантів токсину холери зі зниженою токсичністю, токсину дифтерії, білка CRM197, що є перехресно-реактивним з токсином дифтерії, рекомбінантної стрептококової С5а пептидази, Streptococcus pyogenes ORF1224, Streptococcus pyogenes ORF1664, Streptococcus pyogenes ORF2452, пневмолізину Streptococcus pneumoniae, мутантів пневмолізину зі зниженою токсичністю, Chlamydia pneumoniae ORF T367, Chlamydia pneumoniae ORF T858, правцевого токсоїду, ВІЛ gp120 T1, компонентів, що розпізнають молекули адгезивної матриці мікробної поверхні (MSCRAMMS), факторів і гормонів росту, цитокінів і хемокінів.
16. Імуногенний кон'югат за п. 15, який відрізняється тим, що білком-носієм є CRM197.
17. Імуногенний кон'югат за п. 14, який відрізняється тим, що пептидним імуногеном є фрагмент пептиду Аb.
18. Імуногенний кон'югат за п. 17, який відрізняється тим, що фрагмент пептиду Аb вибирають із групи, що складається із Аb1-3, 1-4, 1-5,1-6,1-7, 1-9, 1-10, 1-11, 1-12, 1-16, 1-28, 3-6, 3-7, 13-28, 15-24, 16-22, 16-23, 17-23, 17-24, 18-24, 18-25, 17-28, 25-35, 33-42, 35-40 і 35-42.
19. Імуногенна композиція, яка містить кон'югат пептидного імуногена з білком-носієм, створена у відповідності з процесом за п. 1, разом з одним чи більше фармацевтично прийнятними ексципієнтами, розчинниками і/або ад'ювантами.
20. Імуногенна композиція за п. 19, яка відрізняється тим, що білок-носій вибирають із групи, що складається із альбуміну сироватки людини, гемоціаніну лімфи слимака (KLH), молекул імуноглобуліну, тироглобуліну, овальбуміну, гемаглютиніну грипу, PADRE поліпептиду, циркумспорозитного (CS) білка малярії, поверхневого антигена гепатиту В (HBSAg19-28), білка теплового шоку (HSP) 65, Mycobacterium tuberculosis, токсину холери, мутантів токсину холери зі зниженою токсичністю, токсину дифтерії, білка CRM197, що є перехресно-реактивним з токсином дифтерії, рекомбінантної стрептококової С5а пептидази, Streptococcus pyogenes ORF1224, Streptococcus pyogenes ORF1664, Streptococcus pyogenes ORF2452, пневмолізину Streptococcus pneumoniae, мутантів пневмолізину зі зниженою токсичністю, Chlamydia pneumoniae ORF T367, Chlamydia pneumoniae ORF T858, правцевого токсоїду, ВІЛ gp120 T1, компонентів, що розпізнають молекули адгезивної матриці мікробної поверхні (MSCRAMMS), факторів і гормонів росту, цитокінів і хемокінів.
21. Імуногенна композиція за п. 20, яка відрізняється тим, що білком-носієм є CRM197.
22. Імуногенна композиція за п. 19, яка відрізняється тим, що пептидним імуногеном є фрагмент пептиду Аb.
23. Імуногенна композиція за п. 22, яка відрізняється тим, що фрагмент пептиду Аb вибирають із групи, що складається із Аb1-3, 1-4, 1-5,1-6, 1-7, 1-9, 1-10, 1-11, 1-12, 1-16, 1-28, 3-6, 3-7, 13-28, 15-24, 16-22, 16-23, 17-23, 17-24, 18-24, 18-25, 17-28, 25-35, 33-42, 35-40 і 35-42.
24. Імуногенна композиція за п. 19, яка відрізняється тим, що один або більше ад'ювантів вибирають із групи, що складається з GM-CSF, 529 SE, IL-12, фосфату алюмінію, гідроксиду алюмінію, Mycobacterium tuberculosis, Bordetella pertussis, бактеріальних ліпополісахаридів, аміноалкілглюкозамінфосфатних сполук, MPL™ (3-О-деацельованого монофосфорильного ліпіду А), поліпептиду, Quil A, STIMULON™ QS-21, токсину коклюшу (РТ), термолабільного токсину Е. соli (LT), IL-1a, ІL-1b, IL-2, IL-4, IL-5, IL-6, IL-7, IL-8, IL-10, IL-13, IL-14, IL-15, IL-16, IL-17, IL-18, інтерферону-a, інтерферону-b, інтерферону-g, G-CSF, TNF-a і TNF-b.
25. Процес за п. 1, який відрізняється тим, що стадія введення реактивної групи у пептидний імуноген містить додавання амінокислотного залишку, що має реактивну групу.
26. Процес за п. 25, який відрізняється тим, що амінокислотний залишок є цистеїновим залишком, в якому реактивна група містить -SH, або амінокислотний залишок є аргініновим залишком, і реактивна група містить гуанідильну групу, амінокислотний залишок є глутаматом або аспартатним залишком, і реактивна група містить -СООН, або амінокислотний залишок є лізиновим залишком, і реактивна група містить -NH2.
27. Процес за п. 5, який відрізняється тим, що реактивна група, яка вводиться в пептидний імуноген, містить -SH або цистеїновий залишок, і де цистеїновий залишок вводиться у пептидний імуноген Аb додаванням під час пептидного синтезу.
28. Процес за п. 27, який відрізняється тим, що цистеїновий залишок розташований на карбоксильному кінці пептидного імуногену Аb.
29. Процес за п. 1, який відрізняється тим, що стадія введення реактивної групи у пептидний імуноген містить бічну тіолову групу на амінокислотному залишку, що піддається такій модифікації через реагент тіолування.
30. Процес за п. 29, який відрізняється тим, що реагент тіолування містить тіолактон N-ацетилгомоцистеїну.
31. Процес за п. 25 або 29, який відрізняється тим, що амінокислотний залишок розташовується на амінному кінці пептидного імуногену Аb або на карбоксильному кінці пептидного імуногену Аb.
32. Процес за п. 1, який відрізняється тим, що білок-носій крім того містить один або більше поліпептидних лінкерів, ковалентно прикріплених до білка-носія, і одна або більше функціональних груп містять замісники одного або більше поліпептидних лінкерів.
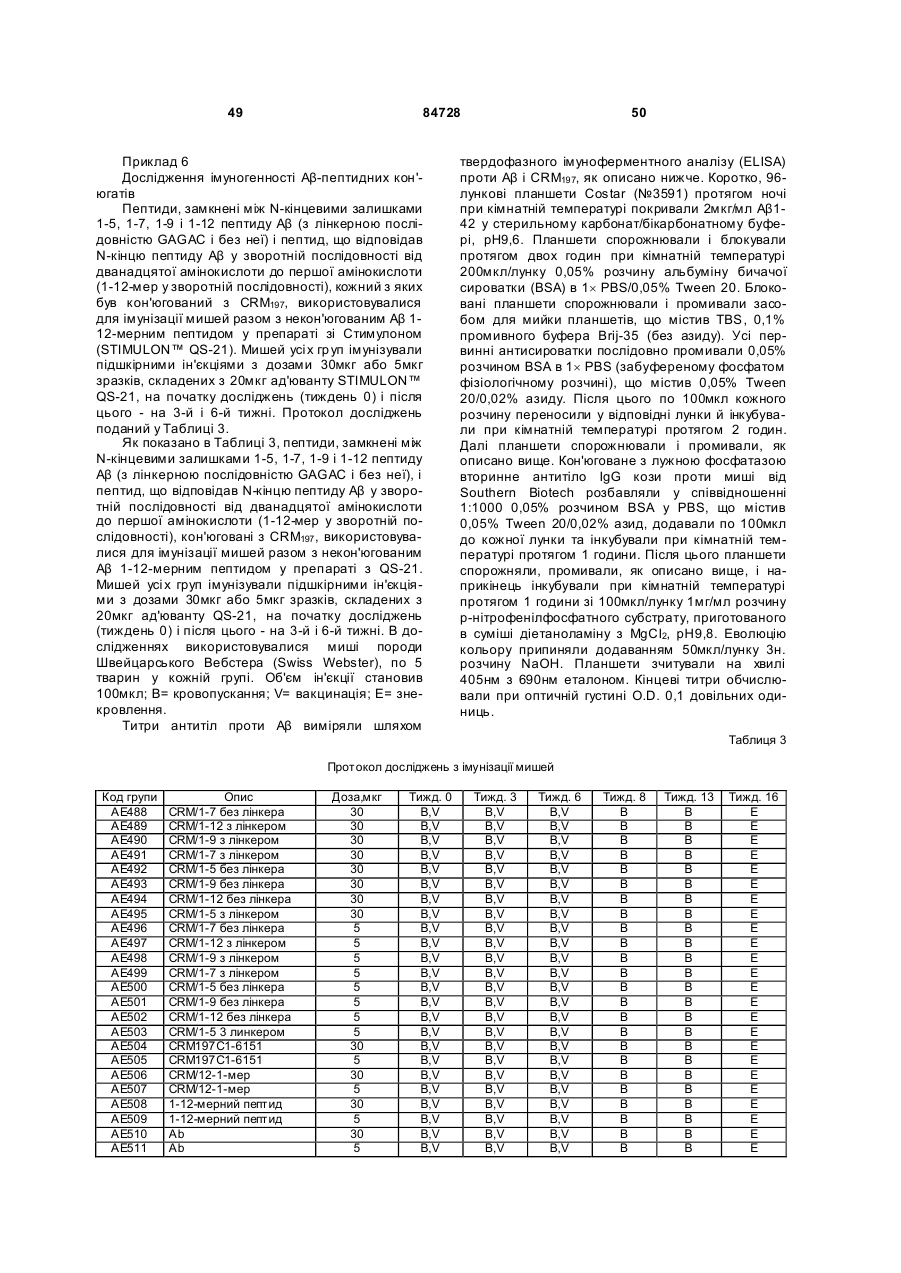
33. Імуногенний кон'югат за п. 9 або 14, який відрізняється тим, що реактивна група амінокислотного залишку пептидного імуногену містить вільну сульфгідрильну групу.
34. Імуногенний кон'югат за п. 33, який відрізняється тим, що вільна сульфгідрильна група є бічним ланцюгом цистеїнового залишку або бічним ланцюгом тіолування лізинового залишку.
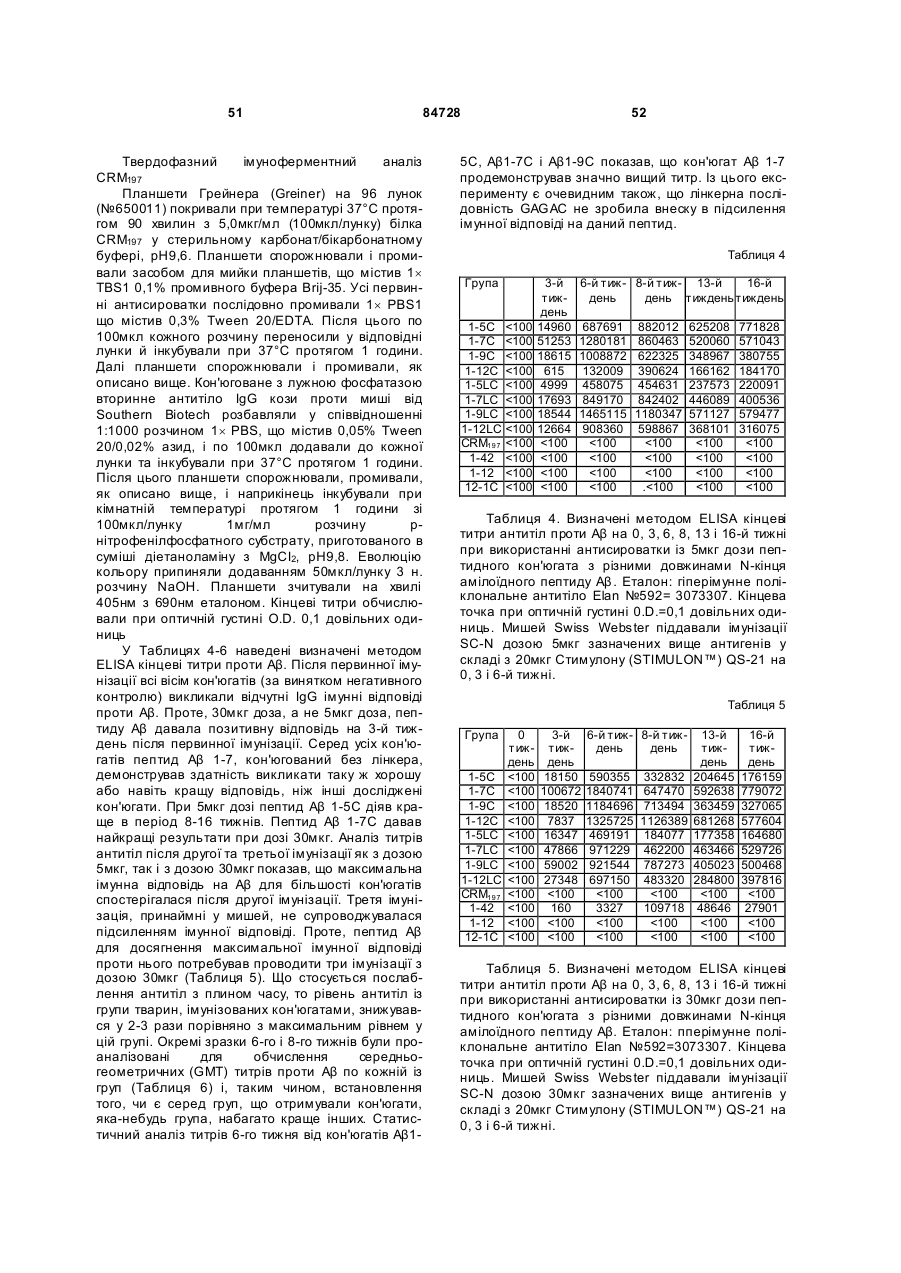
35. Імуногенний кон'югат за п. 9 або 14, який відрізняється тим, що реактивна група амінокислотного залишку пептидного імуногену містить гуанідильну групу, карбоксильну групу або e-амінну групу.
36. Імуногенний кон'югат за п. 13 або 18, який відрізняється тим, що реактивна група амінокислотного залишку пептидного імуногену містить вільну сульфгідрильну групу цистеїнового залишку.
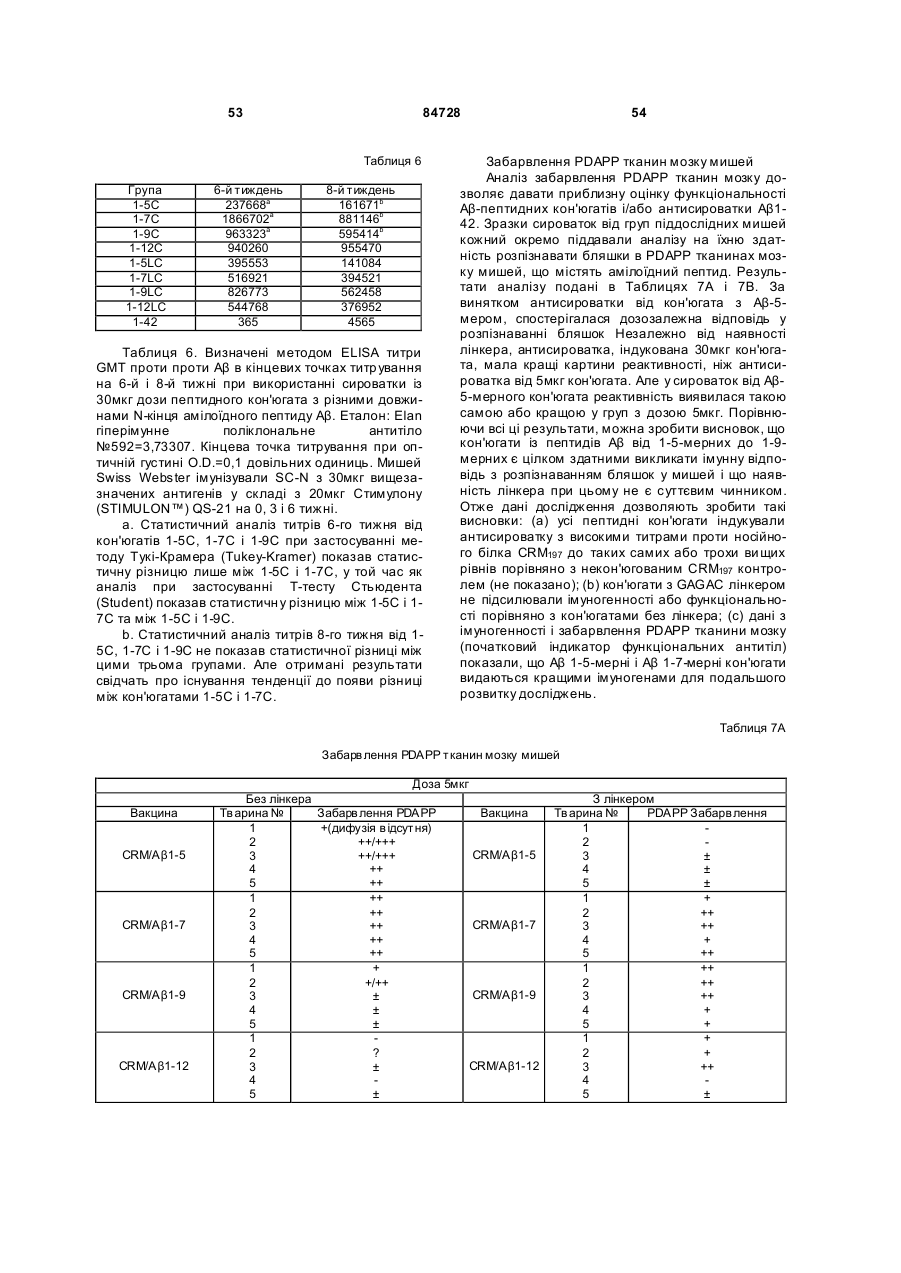
37. Імуногенний кон'югат за п. 36, який відрізняється тим, що цистеїновий залишок розташований на карбоксильному кінці пептидного імуногену.
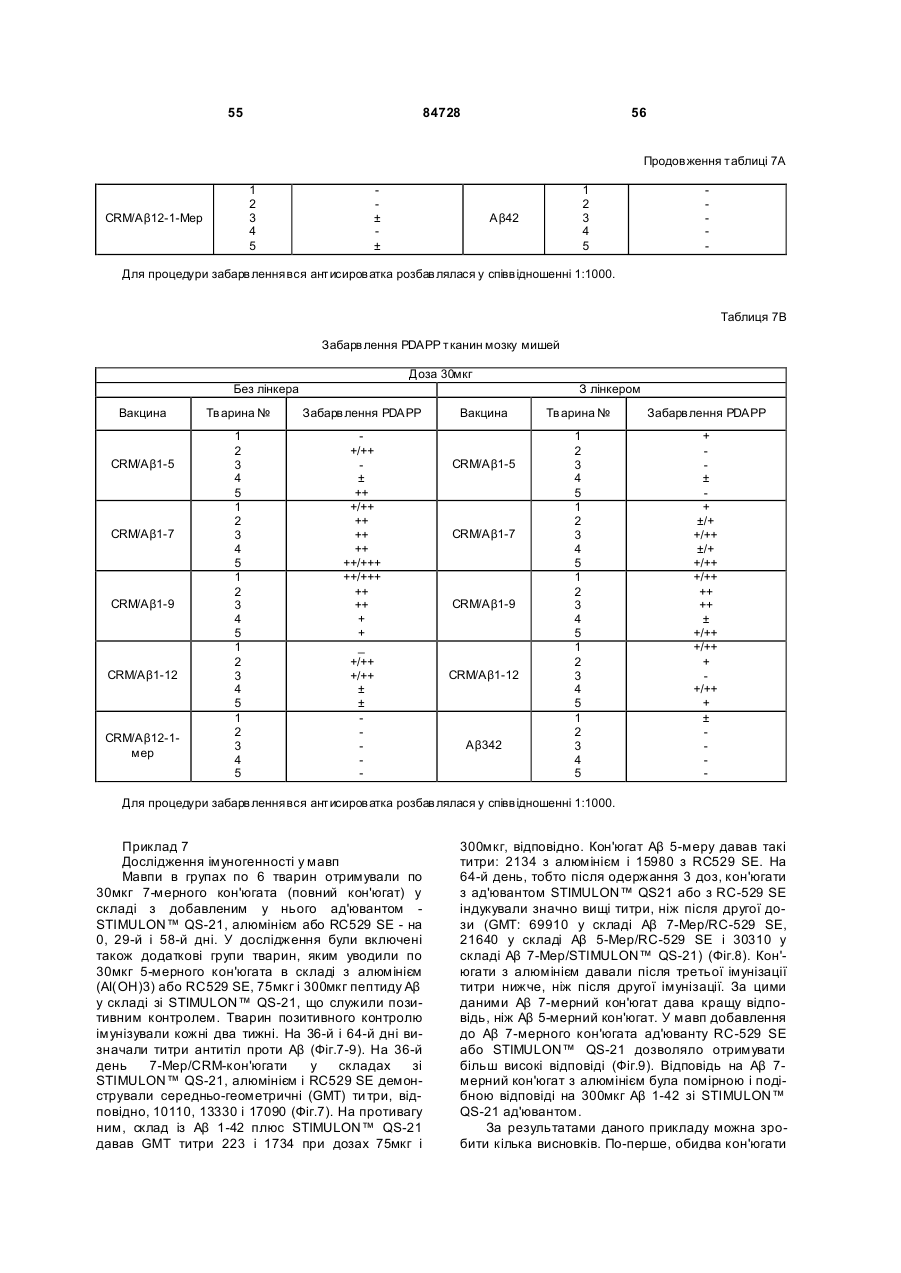
38. Імуногенний кон'югат за п. 9 або 14, який відрізняється тим, що білок-носій крім того містить пептидний лінкер, ковалентно прикріплений до білка-носія, і функціональна група містить замісник пептидного лінкера.
Текст
1. Процес утворення імуногенного кон'югату, який включає у себе стадії: (a) введення реактивної групи у амінокислотний залишок пептидного імуногена, де пептидний імуноген - це пептид Аb або його фрагмент чи (b) дериватизації однієї чи більше функціональних аналог; груп білка-носія для створення активованої функціональної групи на білку-носії; (c) проведення реакції пептидного імуногена за стадією (а) з білком-носієм за стадією (b) в таких умовах, щоб утворити кон'югат, де реактивна група пептидного імуногену ковалентно прикріплена до активованої функціональної групи на білкуносії; і (d) проведення подальшої реакції кон'югата за стадією (с) з кепінг-реагентом для того, щоб інактивувати будь-яку з решти активованих функціональних груп на білку-носії, щоб створити імуногенний кон'югат. 2. Процес за п.1, який відрізняється тим, що білок-носій вибирають із групи, що складається із альбуміну сироватки людини, гемоціаніну лімфи слимака (KLH), молекул імуноглобуліну, тироглобуліну, овальбуміну, гемаглютиніну грипу, PADRE 2 (19) 1 3 84728 9. Імуногенний кон'югат, що містить пептидний імуноген, ковалентно приєднаний до білка-носія, який відрізняється тим, що білок-носій має таку формулу: (X)m C , де С є білком-носієм, а X є перетворювана функціональна група амінокислотного залишку білка-носія, і де m є ціле число більше 0, але не більше 85, і де імуногенний кон'югат має таку формулу: (Xd P)n C (X d R)P , де С є білком-носієм, a Xd є дериватизована функціональна група амінокислотного залишку білка-носія, і де Р є пептидний імуноген, що містить пептид Аb або фрагменти пептиду Аb чи їх аналоги, ковалентно приєднані через реактивну груп у амінокислотного залишку пептидного імуногену до дериватизованої функціональної групи амінокислотного залишку білка-носія, R є кепінг-молекула, ковалентно приєднана до дериватизованої функціональної групи амінокислотного залишку білка-носія, захищаючи цим функціональність білка-носія таким чином, що він зберігає свою здатність викликати бажані імунні відповіді проти пептидного імуногена, який містить пептид Аb або фрагменти пептиду Аb чи їх аналоги, що без носія було б неможливим, n є ціле число більше 0, але не більше 85, і р є ціле число більше 0, але менше ніж 85. 10. Імуногенний кон'югат за п.9, який відрізняється тим, що білок-носій вибирають із групи, що складається із альбуміну сироватки, гемоціаніну лімфи слимака (KLH), молекул імуноглобуліну, тироглобуліну, овальбуміну, гемаглютиніну грипу, PADRE поліпептиду, циркумспорозитного (CS) білка малярії, поверхневого антигена гепатиту В (HBSAg19-28), білка теплового шоку (HSP) 65, Mycobacterium tuberculosis, токсину холери, мутантів токсину холери зі зниженою токсичністю, токсину дифтерії, білка CRM197 , що є перехреснореактивним з токсином дифтерії, рекомбінантної стрептококової С5а пептидази, Streptococcus pyogenes ORF1224, Streptococcus pyogenes ORF1664, Streptococcus pyogenes ORF2452, пневмолізину Streptococcus pneumoniae, мутантів токсину пневмолізину зі зниженою токсичністю, Chlamydia pneumoniae ORF T367, Chlamydia pneumoniae ORF T858, правцевого токсоїду, ВІЛ gp120 T1, компонентів, що розпізнають молекули адгезивної матриці мікробної поверхні (MSCRAMMS), факторів росту, гормонів, цитокінів і хемокінів. 11. Імуногенний кон'югат за п.10, який відрізняється тим, що білком-носієм є CRM197. 12. Імуногенний кон'югат за п.9, який відрізняється тим, що пептидним імуногеном є фрагмент пептиду Аb. 4 13. Імуногенний кон'югат за п.12, який відрізняється тим, що фрагмент пептиду Аb вибирають із групи, що складається із Аb1-3,1-4, 1-5,1-6,1-7, 1-9, 1-10, 1-11, 1-12,1-16, 1-28, 3-6, 3-7, 13-28, 15-24, 16-22, 16-23, 17-23, 17-24, 18-24, 18-25, 17-28, 2535, 33-42, 35-40 і 35-42. 14. Імуногенний кон'югат, який створений у відповідності з процесом за п.1 і має формулу: (Xd P) n C (Xd R)P , де С є білком-носієм, a Xd є дериватизована функціональна група амінокислотного залишку білка-носія, і де Р є пептидний імуноген, що містить пептид Аb або фрагменти пептиду Аb чи їх аналоги, ковалентно приєднані через реактивну груп у амінокислотного залишку пептидного імуногену до дериватизованої функціональної групи амінокислотного залишку білка-носія, R є кепінг-молекула, ковалентно приєднана до дериватизованої функціональної групи амінокислотного залишку білка-носія, захищаючи цим функціональність носія таким чином, що він зберігає свою здатність викликати бажані імунні відповіді проти пептидного імуногена, що без носія було б неможливим, n є ціле число більше 0, але не більше 85, і р є ціле число більше 0, але менше 85. 15. Імуногенний кон'югат за п.14, який відрізняється тим, що білок-носій вибирають із групи, що складається із альбуміну сироватки людини, гемоціаніну лімфи слимака (KLH), молекул імуноглобуліну, тироглобуліну, овальбуміну, гемаглютиніну грипу, PADRE поліпептиду, циркумспорозитного (CS) білка малярії, поверхневого антигена гепатиту В (HBSAg19-28), білка теплового шоку (HSP) 65, Mycobacterium tuberculosis, токсину холери, мутантів токсину холери зі зниженою токсичністю, токсину дифтерії, білка CRM197 , що є перехреснореактивним з токсином дифтерії, рекомбінантної стрептококової С5а пептидази, Streptococcus pyogenes ORF1224, Streptococcus pyogenes ORF1664, Streptococcus pyogenes ORF2452, пневмолізину Streptococcus pneumoniae, мутантів пневмолізину зі зниженою токсичністю, Chlamydia pneumoniae ORF T367, Chlamydia pneumoniae ORF T858, правцевого токсоїду, ВІЛ gp120 T1, компонентів, що розпізнають молекули адгезивної матриці мікробної поверхні (MSCRAMMS), факторів і гормонів росту, цитокінів і хемокінів. 16. Імуногенний кон'югат за п.15, який відрізняється тим, що білком-носієм є CRM197. 17. Імуногенний кон'югат за п.14, який відрізняється тим, що пептидним імуногеном є фрагмент пептиду Аb. 18. Імуногенний кон'югат за п.17, який відрізняється тим, що фрагмент пептиду Аb вибирають із групи, що складається із Аb1-3, 1-4, 1-5,1-6,1-7, 19, 1-10, 1-11, 1-12, 1-16, 1-28, 3-6, 3-7, 13-28, 15-24, 16-22, 16-23, 17-23, 17-24, 18-24, 18-25, 17-28, 2535, 33-42, 35-40 і 35-42. 19. Імуногенна композиція, яка містить кон'югат пептидного імуногена з білком-носієм, створена у відповідності з процесом за п.1, разом з одним чи 5 84728 6 більше фармацевтично прийнятними ексципієнтами, розчинниками і/або ад'ювантами. 20. Імуногенна композиція за п.19, яка відрізняється тим, що білок-носій вибирають із групи, що складається із альбуміну сироватки людини, гемоціаніну лімфи слимака (KLH), молекул імуноглобуліну, тироглобуліну, овальбуміну, гемаглютиніну грипу, PADRE поліпептиду, циркумспорозитного (CS) білка малярії, поверхневого антигена гепатиту В (HBSAg19-28), білка теплового шоку (HSP) 65, Mycobacterium tuberculosis, токсину холери, мутантів токсину холери зі зниженою токсичністю, токсину дифтерії, білка CRM197 , що є перехреснореактивним з токсином дифтерії, рекомбінантної стрептококової С5а пептидази, Streptococcus pyogenes ORF1224, Streptococcus pyogenes ORF1664, Streptococcus pyogenes ORF2452, пневмолізину Streptococcus pneumoniae, мутантів пневмолізину зі зниженою токсичністю, Chlamydia pneumoniae ORF T367, Chlamydia pneumoniae ORF T858, правцевого токсоїду, ВІЛ gp120 T1, компонентів, що розпізнають молекули адгезивної матриці мікробної поверхні (MSCRAMMS), факторів і гормонів росту, цитокінів і хемокінів. 21. Імуногенна композиція за п.20, яка відрізняється тим, що білком-носієм є CRM197. 22. Імуногенна композиція за п.19, яка відрізняється тим, що пептидним імуногеном є фрагмент пептиду Аb. 23. Імуногенна композиція за п.22, яка відрізняється тим, що фрагмент пептиду Аb вибирають із групи, що складається із Аb1-3, 1-4, 1-5,1-6, 1-7, 19, 1-10, 1-11, 1-12, 1-16, 1-28, 3-6, 3-7, 13-28, 15-24, 16-22, 16-23, 17-23, 17-24, 18-24, 18-25, 17-28, 2535, 33-42, 35-40 і 35-42. 24. Імуногенна композиція за п.19, яка відрізняється тим, що один або більше ад'ювантів вибирають із групи, що складається з GM-CSF, 529 SE, IL-12, фосфа ту алюмінію, гідроксиду алюмінію, Mycobacterium tuberculosis, Bordetella pertussis, бактеріальних ліпополісахаридів, аміноалкілглюкозамінфосфатних сполук, MPL™ (3-Одеацельованого монофосфорильного ліпіду А), поліпептиду, Quil A, STIMULON™ QS-21, токсину коклюшу (РТ), термолабільного токсину Е. соli (LT), IL-1a, ІL-1b, IL-2, IL-4, IL-5, IL-6, IL-7, IL-8, IL10, IL-13, IL-14, IL-15, IL-16, IL-17, IL-18, інтерферону-a, інтерферону-b, інтерферону-g, G-CSF, TNF-a і TNF-b. 25. Процес за п.1, який відрізняється тим, що стадія введення реактивної групи у пептидний імуноген містить додавання амінокислотного залишку, що має реактивну гр уп у. 26. Процес за п.25, який відрізняється тим, що амінокислотний залишок є цистеїновим залишком, в якому реактивна група містить -SH, або амінокислотний залишок є аргініновим залишком, і реак тивна група містить гуанідильну груп у, амінокислотний залишок є глутаматом або аспартатним залишком, і реактивна група містить -СООН, або амінокислотний залишок є лізиновим залишком, і реактивна група містить -NH2. 27. Процес за п.5, який відрізняється тим, що реактивна група, яка вводиться в пептидний імуноген, містить -SH або цистеїновий залишок, і де цистеїновий залишок вводиться у пептидний імуноген Аb додаванням під час пептидного синтезу. 28. Процес за п.27, який відрізняється тим, що цистеїновий залишок розташований на карбоксильному кінці пептидного імуногену Аb. 29. Процес за п.1, який відрізняється тим, що стадія введення реактивної групи у пептидний імуноген містить бічну тіолову гр уп у на амінокислотному залишку, що піддається такій модифікації через реагент тіолування. 30. Процес за п.29, який відрізняється тим, що реагент тіолування містить тіолактон Nацетилгомоцистеїну. 31. Процес за п.25 або 29, який відрізняється тим, що амінокислотний залишок розташовується на амінному кінці пептидного імуногену Аb або на карбоксильному кінці пептидного імуногену Аb. 32. Процес за п.1, який відрізняється тим, що білок-носій крім того містить один або більше поліпептидних лінкерів, ковалентно прикріплених до білка-носія, і одна або більше функціональних гр уп містять замісники одного або більше поліпептидних лінкерів. 33. Імуногенний кон'югат за п.9 або 14, який відрізняється тим, що реактивна група амінокислотного залишку пептидного імуногену містить вільну сульфгідрильну гр упу. 34. Імуногенний кон'югат за п.33, який відрізняється тим, що вільна сульфгідрильна група є бічним ланцюгом цистеїнового залишку або бічним ланцюгом тіолування лізинового залишку. 35. Імуногенний кон'югат за п.9 або 14, який відрізняється тим, що реактивна група амінокислотного залишку пептидного імуногену містить гуанідильну груп у, карбоксильну гр упу або e-амінну гр упу. 36. Імуногенний кон'югат за п.13 або 18, який відрізняється тим, що реактивна група амінокислотного залишку пептидного імуногену містить вільну сульфгідрильну гр упу цистеїнового залишку. 37. Імуногенний кон'югат за п.36, який відрізняється тим, що цистеїновий залишок розташований на карбоксильному кінці пептидного імуногену. 38. Імуногенний кон'югат за п.9 або 14, який відрізняється тим, що білок-носій крім того містить пептидний лінкер, ковалентно прикріплений до білка-носія, і функціональна група містить замісник пептидного лінкера. Дана заявка згідно зі Зводом законів США [35 U.S.C. §119(е) U.S.] претендує на корисний ефект попередньої заявки на [патент США сер. №60/530,481, поданої 17 грудня, 2003p.], яка включена тут шля хом посилання в усій її повноті і для всіх цілей. Набутий імунітет є не що інше, як здатність організму регувати на наявність в ньому чужорідних субстанцій і виробляти компоненти (антитіла і клі 7 84728 тини), спроможні специфічним чином взаємодіяти з ними і захищати хазяїна від їх інвазії. "Антиген" або "імуноген" являє собою субстанцію, здатну викликати імунну відповідь цього типу, а також взаємодіяти із сенсибілізованими клітинами й антитілами, виробленими проти нього. Антигени або імуногени, зазвичай, є макромолекулами, що містять чіткі антигенні сайти або детермінанти, які розпізнаються і з якими взаємодіють різноманітні компоненти імунної системи. Вони можуть існувати у формі індивідуальних молекул, що складаються із синтетичних органічних хімічних речовин, білків, ліпопротеїдів, глікопротеїдів, РНК, ДНК, полісахаридів, або ж можуть бути частинами клітинних стр уктур (бактерій або грибків) чи вірусів [Harlow і Lane 1988 а, b, с; Male et al., 1987]. Малі молекули, подібно коротким пептидам, хоча вони зазвичай є здатними взаємодіяти з продуктами імунної відповіді, часто не можуть викликати відповідь самостійно. Такі пептидні імуногени або, як їх часто звуть, "гаптени" ("haptens") фактично є неповними антигенами і, хоча самостійно вони не є здатними викликати імуногенність або добиватися вироблення антитіл, вони можуть бути зроблені імуногенними шляхом сполучання їх з відповідним носієм. Носіями, зазвичай, є білкові антигени більшої молекулярної маси, що є здатними викликати імунологічну відповідь при їх уведенні в організм-хазяїн in vivo. При імунній відповіді антитіла виробляються і виділяються В-лімфоцитами у взаємодії з Тклітинами-хелперами (Тн). У більшості систем типу гаптен-носій B-клітини виробляють антитіла, що є специфічними як до гаптена, так і до носія. У цих випадках T-лімфоцити мають на носії домени специфічного зв'язування, але не розпізнають самі лише гаптени (тобто без носія). В- і Т-клітини, об'єднуючи свої дії в синергічному ефекті, викликають гаптен-специфічну гуморальну імунну відповідь. Після такої імунної відповіді, якщо хазяїн слідом за нею стимулюється самим лише гаптеном, то звичайно він відповідає виробленням гаптен-специфічних антитіл із клітин пам'яті, утворених після початкової імунізації. Синтетичні гаптени, що імітують деякі критичні антиген-детермінантні структури на більш великих макромолекулах, часто кон'югують з носіями, створюючи імунну відповідь на більш велику "батьківську" молекулу. Наприклад, короткі пептидні сегменти можуть бути синтезовані із відомої послідовності білка і сполучені з носієм для індукування імуногенності до нативного білка. Цей тип синтетичного підходу до вироблення імуногена став основою для багатьох сучасних досліджень зі створення вакцин. Проте, в багатьох випадках одне лише генерування гуморальної імунної відповіді шляхом використання синтетичних кон'югатів пептид-носій, хоча і добре сконструйованих, не завжди гарантує повний захисний імунітет проти інтактного антигена. Імунна відповідь, генерована короткою пептидною детермінантою із більш великої вірусної частки або бактеріальної клітини, може бути достатньою лише для генерування пам'яті на рівні B-клітин. Стосовно цих випадків існує сучасна 8 загальноприйнята думка, що цитотоксична Tклітинна реакція є більш важливим індикатором захисного імунітету. Сьогодні одним із найбільш цікавих об'єктів досліджень у галузі імунології є проектування пептидних імуногенів з належними імуногенними детермінатними сайтами зв'язування для розпізнавання як В-клітинами, так і Тклітинами. Підхід до збільшення імуногенності малих або слабоімуногенних молекул шляхом кон'югування цих молекул з великими молекулами "носіїв" з успіхом використовувався протягом десятиріч [Goebel et al., (1939) J. Exp. Med. 69: 53]. Наприклад, було описано багато імуногенних композицій, в яких очищені капсульовані полімери кон'югувалися з носійними білками, утворюючи більш ефективні імуногенні склади, що використовували цей "ефект носія" [Schneerson et al., (1984) Infect, lmmun. 45: 582-591]. Було також показано, що кон'югація шунтує слабку гуморальну імунну відповідь, що звичайно спостерігається у дітей при їх імунізації вільним полісахаридом [Anderson et al., J. Pediatr. 107: 346 (1985); lnsel etal., J. Exp. Med. 158: 294 (1986)]. Кон'югати гаптен-носій з успіхом генерувалися при використанні різноманітних зшивальних та зв'язувальних реагентів, включаючи гомобіфункціональні, гетеробіфункціональні та лінкери нульової довжини. Багато з цих методів сьогодні використовується для сполучання сахаридів, білків і пептидів з пептидними носіями. Більшість з таких методів дозволяє створювати амінні, амідні, уретанові, ізотіосечовинні та дисульфідні зв'язки, а в деяких випадках - тіоетери. Недоліком використання зв'язувальних реагентів, які вводять хімічно активні сайти в бічні ланцюги хімічно активних амінокислотних молекул в молекулах носія і/або гаптена, полягає в тому, що активні сайти, якщо вони не нейтралізовані, можуть вільно реагувати з будь-якою небажаною молекулою чи то in vitro (несприятливо цим впливаючи на функціональність або стабільність кон'югата (чи кон'югатів), чи in vivo (створюючи цим потенційний ризик виникнення несприятливих подій у людей або тварин, імунізованих даними препаратами). На такі надмірно активні сайти можна впливати шляхом проведення з ними хімічних реакцій або шляхом їх "кепування" таким чином, щоб їх дезактивувати, використовуючи різноманітні відомі хімічні реакції, але з іншого боку ці реакції можуть бути р уйнівними для функціональності зазначених кон'югатів. Це може ставати особливо проблематичним при спробах створити кон'югат шляхом уведення хімічно активних сайтів в молекулу носія, оскільки більш великі розміри та більш складна структура останньої (порівняно з гаптеном) може зробити її більш уразливою для руйнівних ефектів хімічної обробки. Підтвердженням цьому є також відсутність прикладів застосування методів, за допомогою яких кон'югат одержують, застосовуючи спочатку активацію носія, потім реакцію кон'югування його з гаптеном і, наприкінець, "кепінгування" (або "кепування") решти хімічно активних сайтів, зберігаючи при цьому здатність отримуваного кон'югата функціонувати як імуногенний склад, що має ба 9 84728 жаний "носійний ефект". Метою даного винаходу є створення процесів для одержання імуногенного кон'югата пептидного імуногена, що містить пептид Аβ або фрагменти пептиду Аβ чи їх аналоги, з білковим або поліпептидним носієм, де пептид Аβ або фрагмент пептиду Аβ чи його аналог кон'югується з носієм через дериватизовані функціональні групи амінокислотних залишків носія, таких як лізини, і де будь-які некон'юговані, дериватизовані функціональні групи амінокислотних залишків дезактивуються шляхом кепінгу для запобігання реакції їх з іншими молекулами, включаючи білки та поліпептиди, і тим самим зберігається функціональність носія, який в результаті залишається спроможним добиватися бажаних імунних відповідей проти даного пептидного імуногена, генерування котрих без носія було б неможливим. Крім того, винахід стосується кон'югатів, що одержуються за допомогою вищезазначених процесів, та імуногенних композицій, що включають у себе такі кон'югати. В одному з варіантів його здійснення даний винахід спрямований на перший процес для кон'югації пептидного імуногена через хімічно активну груп у амінокислотного залишку пептидного імуногена з білковим або поліпептидним носієм, що має одну або більше функціональних гр уп, де даний процес включає у себе стадії: (а) дериватизації однієї чи більше функціональних гр уп білкового або поліпептидного носія для створення дериватизованих молекул з хімічно активними сайтами; (b) проведення реакції дериватизованого білкового або поліпептидного носія стадії (а) з хімічно активною групою амінокислотного залишку пептидного імуногена в таких умовах, що даний пептидний імуноген кон'югується з дериватизованим білковим або поліпептидним носієм через функціональні групи; і (с) проведення подальшої реакції кон'югата з кепінг-реагентом для дезактивації вільних, хімічно активних функціональних груп активованого білкового або поліпептидного носія, захищаючи цим функціональність носія від пептидного імуногена, що без носія було б неможливим. В одному з варіантів здійснення винаходу білковий або поліпептидний носій вибирають із групи, що складається із альбуміну сироватки людини, гемоціаніну лімфи слимака, молекул імуноглобуліну, тироглобулін у, овальбуміну, гемаглютиніну грипу, PAN-DR зв'язувального пептиду (PADRE поліпептиду), циркумспорозитного (CS) білка малярії, поверхневого антигена гепатиту В (HBSAg1928), білка теплового шоку (HSP) 65, Bacillus Calmette-Guerin (BCG), токсину холери, мутантів токсину холери зі зниженою токсичністю, токсину дифтерії, білка CRM197 , що є перехреснореактивним з токсином дифтерії, рекомбінантної стрептококової С5а пептидази, Streptococcus pyogenes ORF1224, Streptococcus pyogenes ORF1664, Streptococcus pyogenes ORF2452, пневмолізину Streptococcus Pneumoniae, мутантнів пневмолізину зі зниженою токсичністю, Chlamydiapneumoniae ORF Т367, Chlamydiapneumoniae ORF Т858, правцевого токсоїду, ВІЛ gp120 T1, компонентів мікробної поверхні, що розпізнають молекули адгезивної матриці 10 (MSCRAMMS), факторів і гормонів росту, цитокінів і хемокінів. В іншому варіанті здійснення винаходу білковий або поліпептидний носій містить Т-клітинну антигенну детермінанту. Передбачений також варіант здійснення винаходу, в якому білковим або поліпептидним носієм є бактеріальний токсоїд, такий як правцевий токсоїд, токсин холери або мутант токсину холери, як зазначено вище. У кращому варіанті білковим або поліпептидним носієм є CRM197. У ще одному варіанті здійснення винаходу білковим або поліпептидним носієм може бути гемаглютинін грипу, PADRE поліпептид, CS білок маляріїї, поверхневий антиген гепатиту В (HSBAg19- 28), білок теплового шоку 65 (HSP 65) або поліпептид із Mycobacterium tuberculosis (BCG). У кращому варіанті білковий або поліпептидний носій вибирають серед стрептококової rС5а пептидази, Streptococcus pyogenes ORF1224, Streptococcus pyogenes ORF1664 або Streptococcus pyogenes ORF2452, пневмолізину Streptococcus Pneumoniae, мутантнів пневмолізину зі зниженою токсичністю, Chlamydiapneumoniae ORF Т367 і Chlamydiapneumoniae ORF Т858. В одному з варіантів здійснення винаходу білковим або поліпептидним носієм є фактор або гормон росту, який стимулює або підсилює імунну відповідь і який вибирають з сукупності, що складається із IL-1, IL-2, у-інтерферону, IL-10, GM-CSF, MIP-1α, MIP-1β i R ANTES. В іншому варіанті здійснення даного винаходу пропонується пептидний імуноген, що містить пептид Аβ або фрагменти пептиду Аβ чи їх аналоги, що викликають імуногенну відповідь проти деяких антигенних детермінант усередині Аβ. До числа імуногенних пептидів за даним винаходом входять імуногенні гетерологічні пептиди. У деяких імуногенних пептидів фрагмент пептиду Аβ зшивають з носієм, утворюючи імуногенний гетерологічний пептид, а потім цей гетерологічний пептид зшивають з носієм у відповідності з процесом за даним винаходом, отримуючи кон'югат. Відповідно до іншого варіанту здійснення даного винаходу пептидним імуногеном є поліпептид що містить N-кінцевий сегмент, що складається принаймні із залишків 1-5 пептиду Аβ де першим залишком пептиду Аβ є N-кінцевий залишок даного поліпептиду, де цей поліпептид не має Скінцевого сегмента пептиду Аβ У відповідності з ще одним варіантом здійснення даного винаходу пептидним імуногеном є поліпептид що містить Nкінцевий сегмент пептиду Аβ де цей сегмент починається на залишках 1-3 пептиду Aβ, а закінчується на залишках 7-11 пептиду Аβ У деяких варіантах здійснення даного винаходу пептидним імуногеном є речовина, що викликає імуногенну відповідь проти N-кінцевого сегмента пептиду Аβ де цей сегмент починається на залишках 1-3 пептиду Аβ, а закінчується на залишках 7-11 пептиду Аβ не викликаючи імуногенної відповіді проти антигенної детермінанти в інтервалі залишків 12-43 пептиду Аβ43. Відповідно до ще одного варіанту здійснення даного винаходу пептидним імуногеном є гетерологічний поліпептид, що містить сег 11 84728 мент пептиду Аβ зшитий з гетерологічною амінокислотною послідовністю, що індук ує відповідь Тклітин-хелперів проти цієї гетерологічної амінокислотної послідовності, а отже і відповідь В-клітин проти даного N-кінцевого сегмента. У деяких пептидних імуногенах N-кінцевий сегмент пептиду Аβ зши тий принаймні на його С-кінці з гетерологічним поліпептидом. У деяких пептидних імуногенів N-кінцевий сегмент пептиду Аβ зшитий принаймні на його N-кінці з гетерологічним поліпептидом. У деяких пептидних імуногенів Nкінцевий сегмент пептиду Аβ зшитий принаймні на його N і C кінцях з першим і другим гетерологічними поліпептидами. У деяких пептидних імуногенів N-кінцевий сегмент пептиду Аβ зши тий принаймні на його N-кінці з гетерологічним поліпептидом, а на його С-кінці принаймні з однією додатковою копією N-кінцевого сегмента. У деяких пептидних імуногенів поліпептид містить від N-кінця до Скінця N-кінцевий сегмент пептиду Аβ множину додаткових копій N-кінцевого сегмента і гетерологічний амінокислотний сегмент. У деяких із вищезгаданих пептидних імуногенів поліпептид містить також щонайменше одну додаткову копію N-кінцевого сегмента. У деяких із вищезгаданих пептидних імуногенів зазначений фрагмент не має, щонайменше, 5 С-кінцевих амінокислот в Аβ43. У деяких варіантах ви щеописаних пептидних імуногенів зазначений фрагмент містить до 10 суміжних амінокислот із Аβ Відповідно до іншого аспекту даного винаходу пропонується пептидний імуноген, що містить пептид Аβ або фрагменти пептиду Аβ чи їх аналоги, що викликають імуногенну відповідь проти деяких антигенних детермінант усередині Aβ, і може мати конфігурацію, відому під назвою конфігурації множинного антигенного пептиду (МАР). У деяких вищеописаних варіантах здійснення даного винаходу пептидний імуноген є із Nкінцевої половини пептиду Аβ. У деяких вищеописаних варіантах здійснення даного винаходу пептидним імуногеном є фрагмент пептиду Аβ вибраний із сукупності, що складається із Аβ1-3, 1-4, 1-5, 1-6, 1-7, 1-Ю, 1-11, 1-12, 1-16, 3-6 і 3-7 У деяких вищеописаних варіантах здійснення даного винаходу пептидний імуноген є із внутрішньої ділянки пептиду Аβ. У деяких вищеописаних варіантах здійснення даного винаходу пептидним імуногеном є фрагмент пептиду Аβ вибраний із групи, що складається із Аβ13-28, 15-24, 17-28 і 25-35. Відповідно до деяких вищеописаних варіантів здійснення даного винаходу пептидний імуноген є із Скінцевої частини пептиду Аβ Відповідно до деяких варіантів здійснення даного винаходу пептидним імуногеном є фрагмент Аβ вибраний із сукупності, що складається із Аβ33-42, 35-40 і 35-42. Відповідно до деяких варіантів здійснення даного винаходу пептидним імуногеном є фрагмент пептиду Аβ вибраний із сукупності, що складається із Аβ1-3, 14, 1-5, 1-6, 1-7, 1-10, 1-11, 1-12, 1-16, 1-28, 3-6, 3-7, 13-28, 15-24, 17-28, 25-35, 33-42, 35-40 і 35-42. Відповідно до деяких варіантів здійснення даного винаходу пептидним імуногеном є фрагмент пептиду Аβ вибраний із сукупності, що складається із 12 Аβ1-5, Аβ1-7, Аβ1-9 і Аβ1-12. Відповідно до деяких варіантів здійснення даного винаходу пептидним імуногеном є фрагмент пептиду Аβ вибраний із сукупності, що складається із пептиду Аβ1-5-L, Аβ1-7-L, Аβ1-9-L і Аβ1-12-L, де L є лінкер. Відповідно до деяких варіантів здійснення даного винаходу пептидним імуногеном є фрагмент пептиду Аβ вибраний із групи, що складається із Аβ1-5-L-C, Аβ1-7-L-C, Аβ1-9-L-C і Аβ1-12-L-C, де C є цистеіновий амінокислотний залишок. Відповідно до деяких варіантів здійснення даного винаходу пептидним імуногеном є фрагмент пептиду Аβ вибраний із сукупності, що складається із Аβ16-22, Аβ16-23, Аβ17-23, Аβ17-24, Аβ18-24 і Аβ18-25. Відповідно до деяких варіантів здійснення даного винаходу пептидним імуногеном є фрагмент пептиду Аβ вибраний із групи, що складається із Аβ16-22-С, Аβ16-23-С, Аβ17-23-С, Аβ1724-С, Аβ18-24-С і Aβ18-25-C, де C є цистеїновий амінокислотний залишок. Відповідно до інших варіантів здійснення даного винаходу пептидним імуногеном є фрагмент пептиду Аβ вибраний із групи, що складається із С-Аβ16-22, С-Аβ16-23, САβ17-23, С-Аβ17-24, С-Аβ18-24 і С-Аβ18-25, де C є цистеїновий амінокислотний залишок. У деяких із вищезгаданих пептидних імуногенів гетеролопчний поліпептид вибирають із групи, що складається із пептидів, які мають Т-клітинну антигенну детермінанту, В-клітинну антигенну детермінанту та їх комбінації. В одному з варіантів здійснення винаходу функціональну груп у однієї чи більше амінокилотних молекул білкового або поліпептидного носія або приєднаного в разі потреби поліпептидного лінкера дериватизують за допомогою зшивального реагента. В іншому варіанті здійснення винаходу агентом дериватизації є зшивальний реагент нульової довжини. Можливим є варіант здійснення винаходу, в якому агентом дериватизації є гомобіфункціональний зшивальний реагент. Передбачений також варіант здійснення винаходу, в якому агентом дериватизації є а гетеробіфункціональний зшивальний реагент. У кращому варіанті здійснення винаходу гетеробіфункціональним реагентом є реагент, що реагує з первинною або є-амінною функціональною групою однієї чи більше амінокислотних молекул білкового або поліпептидного носія і з бічною тіоловою групою однієї чи більше амінокислотних молекул пептидного імуногена. В одному з варіантів здійснення винаходу ге теробіфункціональним реагентом є N-сукцинімідилбромацетат. В іншому варіанті здійснення винаходу первинною або є-амінною функціональною групою є лізин. Передбачений також варіант здійснення винаходу, в якому дериватизація первинної або eамінної функціональної групи лізину білкового або поліпептидного носія N-сукцинімідилбромацетатом має результатом бромацетилювання первинних або e-амінних залишків у лізинових молекул білкового або поліпептидного носія. У ще кращому варіанті бічною тіоловою групою є цистеїновий залишок пептидного імуногена, який може бути локалізований на амінному кінці пептидного імуногена, на карбоксильному кінці пептидного імуноге 13 84728 на або усередині пептидного імуногена. В іншому варіанті здійснення винаходу бічна тіолова група створюється реагентом тіолування, таким, як N-ацетилгомоцистеїнтіолактон, реагент Трота (Traut) (2-імінотілан), SATA (М-сукцинімідилБ-ацетилтіоацетат), SMPT (4-сукцинімідилоксикарбонілметил-2-піридилтіотолуол), сульфоLC SPDP (сульфосукцинімідилпіридилдитіопропіонамідгексаноат), SPDP (сукцинімідилпіридилдитіопропіонат). У кращому варіанті кепінг-реагент, який використовують для дезактивації вільної, хімічно активної функціональної групи активованого білкового або поліпептидного носія, вибирають із групи реагентів, що складається із цистеїнаміну, N-ацетилцистаміну й етаноламіну. В особливо кращому варіанті здійснення винаходу кепінг-реагент, який використовують для дезактивації вільної, хімічно активної функціональної групи активованого білкового або поліпептидного носія, вибирають із групи реагентів, що складається із гідроксиду натрію, карбонату натрію, бікарбонату амонію й аміаку. В одному з варіантів здійснення винаходу хімічно активною групою амінокислотного залишку пептидного імуногена є вільна сульфгідрильна група. В іншому варіанті здійснення винаходу одна або більше функціональних груп є на лінкері, який у разі потреби є приєднаним до білкового або поліпептидного носія. У кращому варіанті зазначеним лінкером є пептидний лінкер. У ще кращому варіанті пептидним лінкером є полілізин. В одному з варіантів його здійснення даний винахід спрямований на другий процес для кон'югування пептидного імуногена, що містить пептид Аβ або фрагмент пептиду Аβ чи їх аналоги, з білковим або поліпептидним носієм, який має таку структур у: де: C є білковий або поліпептидний носій, a X є перетворюваною функціональною групою амінокислотного залишку в білковому або поліпептидному носії або, в разі потреби, амінокислотного залишку пептидного лінкера, ковалентно приєднаного до білкового або поліпептидного носія, і де m є ціле число більше 0, але не більше 85, причому даний процес включає у себе стадії: (a) дериватизації однієї чи більше функціональних груп білкового або поліпептидного носія або приєднаної в разі потреби молекули лінкера для створення дериватизованих молекул з хімічно активними сайтами; (b) проведення реакції дериватизованого білкового або поліпептидного носія стадії (а) з хімічно активною групою амінокислотного залишку пептидного імуногена з утворенням ковалентно зв'язаного кон'югата пептидного імуногена з білковим або поліпептидним носієм; і (c) проведення реакції вищезазначеного кон'ю 14 гата з кепінг-реагентом для дезактивації вільної, хімічно активної функціональної групи в активованому білковому або поліпептидному носії таким чином, щоб кеповані групи не могли реагувати з іншими молекулами, включаючи білки та поліпептиди, і цим зберігалася функціональність носія та його здатність викликати бажані імунні відповіді проти даного пептидного імуногена, що без носія було б неможливим, у результаті чого утворюється кепований кон'югат пептидного імуногена з білковим або поліпептидним носієм, який описується такою формулою: де: C є білковий або поліпептидний носій, a Xd є дериватизована функціональна група амінокислотного залишку білкового або поліпептидного носія або, в разі потреби, амінокислотного залишку пептидного лінкера, ковалентно приєднаного до білкового або поліпептидного носія, і де P є пептидна імуногенна молекула, ковалентно приєднана до дериватизованої функціональної групи амінокислотного залишку білкового носія або, в разі потреби, амінокислотниого залишку пептидного лінкера, ковалентно приєднаного до білкового або поліпептидного носія; R є кепінгмолекула, ковалентно приєднана до дериватизованої функціональної групи амінокислотного залишку білкового або поліпептидного носія або, в разі потреби, амінокислотниого залишку пептидного лінкера, ковалентно приєднаного до білкового або поліпептидного носія; n є ціле число більше 0, але не більше 85, а р є ціле число більше 0, але менше 85. Подані вище детальні описи варіантів здійснення першого процесу є правомірними також для кон'югатів, що одержуються за допомогою другого процесу. В одному з варіантів його здійснення даний винахід спрямований на кон'югати пептидного імуногена з білковим або поліпептидним носієм, де білковий або поліпептидний носій має таку формулу: де: C є білковий або поліпептидний носій, a X є перетворювана функціональна група амінокислотного залишку білкового або поліпептидного носія або, в разі потреби, амінокислотного залишку пептидного лінкера, ковалентно приєднаного до білкового або поліпептидного носія, і де m є ціле число більше 0, але не більше 85, а також де кепований кон'югат пептидного імуногена з білковим або поліпептидним носієм має таку формулу: 15 84728 де C є білковий або поліпептидний носій, a Xd є дериватизована функціональна група амінокислотного залишку білкового або поліпептидного носія або, в разі потреби, амінокислотного залишку пептидного лінкера, ковалентно приєднаного до білкового або поліпептидного носія, і де P є пептидна імуногенна молекула, ковалентно приєднана до дериватизованої функціональної групи амінокислотного залишку білкового носія або, в разі потреби, амінокислотного залишку пептидного лінкера, ковалентно приєднаного до білкового або поліпептидного носія, R є кепінг-молекула, ковалентно приєднана до дериватизованої функціональної групи амінокислотного залишку білкового або поліпептидного носія або, в разі потреби, амінокислотного залишку пептидного лінкера, ковалентно приєднаного до білкового або поліпептидного носія, захищаючи цим функціональність носія таким чином, що він зберігає свою здатність викликати бажані імунні відповіді від пептидного імуногена, що без носія було б неможливим, n є ціле число більше 0, але не більше 85, а р є ціле число більше 0, але менше 85. Поданий вище детальний опис варіантів здійснення першого і другого процесів є правомірним також для описаних ви ще кон'югатів. В іншому варіанті його здійснення даний винахід спрямований на кон'югати пептидного імуногена, що містить пептид Аβ або фрагменти пептиду Аβ чи їх аналоги, з білковим або поліпептидним носієм, які створені за допомогою другого процесу згідно з даним винаходом і мають таку формулу: де: C є білковий або поліпептидний носій, a Xd є дериватизована функціональна група амінокислотного залишку білкового або поліпептидного носія або, в разі потреби, амінокислотного залишку пептидного лінкера, ковалентно приєднаного до білкового або поліпептидного носія, і де P є молекула пептидного імуногена, ковалентно приєднана до дериватизованої функціональної групи амінокислотного залишку білкового носія або, в разі потреби, амінокислотного залишку пептидного лінкера, ковалентно приєднаного до білкового або поліпептидного носія, R є кепінг-молекула, ковалентно приєднана до дериватизованої функціональної групи амінокислотного залишку білкового або поліпептидного носія або, в разі потреби, амінокислотного залишку пептидного лінкера, ковалентно приєднаного до білкового або поліпептидного носія, захищаючи цим функціональність носія таким чином, що він зберігає свою здатність викликати бажані імунні відповіді від пептидного імуногена, що без носія було б неможливим, n є ціле число більше 0, але не більше 85, а р є ціле число більше 0, але менше 85. Поданий вище детальний опис варіантів здійснення другого процесу є правомірним також для кон'югатів, створених за допомогою другого про 16 цесу, як описано вище. В іншому варіанті здійснення винаходу пропонуються імуногенні композиції, що містять кон'югат пептидного імуногена з білковим або поліпептидним носієм, створений за допомогою другого процесу за даним винаходом, разом з одним чи більше фармацевтично прийнятними ексципієнтами, розріджувачами та ад’ювантами. Поданий вище детальний опис варіантів здійснення другого процесу і створених за його допомогою кон'югатів є правомірним також для імуногенних композицій, що містять ці кон'югати згідно з описаним вище. В іншому варіанті здійснення винаходу пропонується процес для індукування імунної відповіді у ссавця, де зазначений процес включає у себе введення ссавцю ефективної кількості імуногенної композиції за даним винаходом. Подані вище детальні описи варіантів здійснення імуногенної композиції, що містить кон'югат за даним винаходом, є правомірними також до варіантів здійснення даного винаходу, що стосуються процесу для використання такої імуногенної композиції. Фіг.1. Блок-схема процесу кон'югації фрагментів пептиду Аβ з білковим або поліпептидним носієм CRM197 для утворення Аβ/CRM197-кон'югата. Фіг.2. Блок-схема процесу кислотного гідролізу, що використовується для визначення кількості S-карбоксиметилцистеїну і Sкарбоксиметилцистеїнаміну при оцінці ступеню злиття кон'югатів пептидного імуногена з білком або поліпептидом, таких, як кон'югат Аβ/CRM197. Фіг.3. Ілюстрація рН-залежності реакції кон'югації пептиду Аβ з білком CRM. Фіг.4. Ілюстрація залежності реакції кон'югації пептиду Аβ з білком CRM від співвідношення пептид:CRM. Фіг.5. Перевірка кепінг-процесу в кон'югації Аβ1-7/CRM. Величина рН реакційної суміші складала 9,15. Тривалість реакції з пептидом становила 16 годин. Кепінг N-ацетилцистеїнаміном тривав 8 годин. Фіг.6. Кон'югація і кепінг пептидом при різних співвідношеннях пептид:CRM. Величина рН реакційної суміші складала 9,0. Тривалість реакції з пептидом становила 16 годин. Кепінг Nацетилцистеїнаміном тривав 8 годин. Фіг.7. Титри сироваток приматів на 36 день після імунізації приматів кон'югатами пептиду Аβ з різними ад’ювантами. Фіг.8. Титри сироваток приматів на 64 день після імунізації приматів кон'югатами пептиду Аβ з різними ад’ювантами. Фіг.9. Титри приматів по днях і групах імунізації. Примати імунізувалися кон'югатами Аβ1-7 або Аβ1-5 з CRM197 з галуном або 529 як ад’ювантами, а титри антитіл проти Аβ виміряли на 29, 36, 57 і 54 дні. Фіг.10. Пептид-білкові кон'югати характеризувалися за допомогою аналізу методом SDS-PAGE (електрофорез у поліакриламідному гелі з додецилсульфатом натрію) вестерн-блотінгу з тристрициновим гелем промислового виготовлення. Смугами на фото є: маркер (смуга 1); L-28375 17 24/01 (смуга 2); L-28375 24/02 (смуга 3); L-28375 24/03 (смуга 4); L-28375 24/04 (смуга 5); L-28375 24/05 (смуга 6); L-28375 24/06 (смуга 7) L-28375 84728 18 24/07 (смуга 8); L-28375 24/08 (смуга 9); L-28375 24/09 (сурогат) (смуга 10); і BrAcCR M197 (см уга 11). Скорочений опис послідовностей 19 84728 Даний винахід стосується процесів для створення кон'югатів типу пептидний імуноген-носій, де непрореаговані активні функціональні групи носія, що створюються під час активації, дезактивуються за допомогою кепінг-реагентів і, зокрема, Nацетилцистеїнаміну для запобігання їх подальшому реагуванню. Винахід стосується також кепованих кон'югатів типу носій-пептидний імуноген, створених за допомогою запропонованих процесів, та імуногенних композицій, що містять зазначені кон'югати. Підхід до збільшення імуногенності малих або слабоімуногенних молекул таких, як сахариди, шляхом кон'югації з успіхом використовувався протягом десятиріч [Goebel et аl., (1939) J. Exp. Med. 69. 53]. У результаті було описано багато імуногенних композицій, в яких очищені капсульовані полімери кон'югувалися з носійними білками, утворюючи більш ефективні імуногенні композиції, що використовували цей "ефект носія". Наприклад, Шнірсон зі співробітниками [Schneerson et al., J. Exp. Med. 152: 361-376, 1980] описали кон'югати "Haemophilus influenzae b полісахаридний білок", які надають імунітет до інвазійних хвороб, викликаних цим мікроорганізмом. Кон'югати PRP (полірибосилрибітолфосфату, капсульованого полімеру мікроорганізму H. influenzae b) виявили свою більш високу ефективність, ніж імуногенні композиції на основі лише полісахариду [Chu et аl., (1983) Infect. Іmmun. 40: 245; Schneerson et al. (1984), Infect. Іmmun. 45: 582-591]. Було також показано, що кон'югація шунтує слабку гуморальну імунну відповідь, що звичайно спостерігається у дітей при їх імунізації вільним полісахаридом 20 [Anderson et аl., J. Pediatr. 107: 346 (1985); lnsel et аl., J. Exp. Med. 158: 294 (1986)]. Ще однією перевагою використання бактеріального токсину або токсоїду як білкового носія порівняно з імунізацією людей (наприклад, від правця або дифтерії) у стандартній практиці є те, що бажаний імунітет до токсину або токсоїду індукується разом з імунітетом проти патогенів, пов'язаних з даним капсульованим полімером. Для вироблення високоспецифічних моноклональних антитіл, здатних розпізнавати дискретні хімічні антигенні детермінанти на зв'язаному гаптені, використовуються також кон'югати типу "антигенна детермінанта/гаптен-носій". Одержані за їх допомогою моноклональні антитіла часто використовуються в дослідженнях антиген-детермінантної структури і взаємодій між нативними білками. У багатьох випадках такі сполучення антигенних детермінант з гаптенами, що використовуються для створення цих моноклональних антитіл, є малими пептидними сегментами, які являють собою ключові антигенні сайти на поверхні більш великих білків. Критеріями для добору носія, підходящого для застосування у створенні кон'югата "антигенна детермінанта/гаптен-носій", є його потенційна імуногенність, наявність у ньому підходящої функціональної групи для кон'югації із субстанцією "антигенна детермінанта/гаптен", прийнятна розчинність навіть після дериватизації і відсутність токсичності in vivo. Цим критеріям задовольняють кон'югати, створені за допомогою процесів згідно з даним винаходом. Підходящими є будь-які стабільні кон'югати "пептидний імуноген-носій", створені за 21 84728 допомогою описаного тут процесу кон'югації. Ці кон'югати одержують, застосовуючи процес згідно з даним винаходом, де білковий або поліпептидний носій, який має структуру ковалентно зв'язується з білковим або поліпептидним носієм, де: C є білковий або поліпептидний носій, a X є перетворювана функціональна група на амінокислотному залишку білкового або поліпептидного носія або, в разі потреби, на амінокислотному залишку пептидного лінкера, ковалентно приєднаного до білкового або поліпептидного носія, і де m є ціле число більше 0, але не більше 85, ковалентно зв'язується з пептидним імуногеном і де кон'югат пептидного імуногена з білковим або поліпептидним носієм має таку формулу: де: C є білковий або поліпептидний носій, a Xd є дериватизована функціональна група на амінокислотному залишку білкового або поліпептидного носія або, в разі потреби, на амінокислотному залишку пептидного лінкера, ковалентно приєднаного до білкового або поліпептидного носія, P є пептидний імуноген, ковалентно зв'язаний з дериватизованою функціональною групою амінокислотного залишку білкового або поліпептидного носія або, в разі потреби, на амінокислотному залишку пептидного лінкера, ковалентно приєднаного до білкового або поліпептидного носія, R є кепінг-молекула, ковалентно приєднана до дериватизованої функціональної групи на амінокислотному залишку білкового або поліпептидного носія або, в разі потреби, амінокислотниого залишку пептидного лінкера, ковалентно приєднаного Гемаглютинін грипу: Н А307-319 PAN-DR Пептид (PADRE пептид) CS білок малярії: ТЗ антигенна детермінанта Поверхневий антиген гепатиту В: HBs Ag19-28 Білок 65 теплового шоку: hsp65153-171 Bacillus Calmette-Guerin (BCG) Правцевий токсоїд: ТТ830-844 Правцевий токсоїд: ТТ947-967 ВІЛ gр120Т1: CRM197 Фрагмент альбуміну 22 до білкового або поліпептидного носія, захищаючи цим функціональність носія таким чином, що він зберігає свою здатність викликати бажані імунні відповіді від пептидного імуногена, що без носія було б неможливим, n є ціле число більше 0, але не більше 85, а р є ціле число більше 0, але менше 85. Добір носіїв Деякі пептидні імуногени містять відповідну антигенну детермінанту для індукування імунної відповіді, але є занадто малими для того, щоб бути імуногенними. У зв'язку з цим, пептидні імуногени об'єднують з підходящим носієм для сприяння індукції імунної відповіді. У поданій вище схематичній уяві кон'югата "пептидний імуноген-носій", створеного за допомогою процесу згідно з даним винаходом, C є білковий або поліпептидний носій, з яким пептидні імуногени кон'юговані безпосередньо через дериватизовані функціональні групи на амінокислотних залишках самого носія або непрямо - через дериватизовані функціональні групи на пептидних лінкерах, ковалентно зв'язаних з цими носіями. Підходящими білковими або поліпептидними носіями можуть бути, наприклад, альбумін (включаючи альбумін людської сироватки), гемоціанін лімфи слимака, молекули імуноглобуліну, тироглобулін, овальбумін, правцевий токсоїд або токсоїд від інших патогенних бактерій, що мають знижену токсичність, включаючи таких мутантів, як дифтерія, E. соlі, холера, H. pylori, або пригнічена похідна токсину. Одним з таких носіїв є, наприклад, білок CRM197 (SEQ ID NO.: 40), що володіє перехресною реактивністю з токсином дифтерії. Підходящими носіями можуть бути також Тклітинні антигенні детермінанти, що зв'язуються з множинними алелями головного комплексу гістосумісності (MHC), наприклад, щонайменше 75% всіх алелей MHC людини. Такі носії іноді називають "універсальними Т-клітинними антигенними детермінантами". Нижче наведені типові носії з універсальними Т-клітинними антигенними детермінантами. PKYVKQNTLKL AT (SEQ. ID NO. 11) AKXVAAWTLKAAA (SEQ. ID NO. 12) EKKIAKMEKASSVFNV (SEQ. ID NO. 13) FELLTRILTI (SEQ. ID NO. 14) QSIGDLIAEAMDKVGNEG (SEQ. ID NO. 15) QVHFQPLPPAWKL (SEQ. ID NO. 16) QYIKANSKFIGITEL (SEQ. ID NO. 17) NNFTVSFWLRVPKVSASHLE (SEQ. ID NO. 18) KQIIN MWQEVGKAMY (SEQ. ID NO. 19) Див. Скорочений опис послідовностей (SEQ ID NO.: 40) ISQAVHAAH AEINEAGR (SEQ ID NO: 41) До носіїв, які є підходящими для стимулювання або підсилення імунної відповіді і з якими може бути кон'югований пептидний імуноген або гаптен, належать також цитокіни, наприклад, IL-1, IL-1 α i β пептиди, IL-2, γlNF, IL-10, GM-CSF, і хемокіни, наприклад, МІР 1α i β, і RANTES. Імуногенні пептиди можна також зв'язувати з білковими або пептидними носіями, що підсилюють транспортування через тканини, як описано в заявках [O'Mahony, WO 97/17163, WO 97/17614], включених тут в усій їхній повноті і для всіх цілей шля хом посилання. Серед підходящих носіїв можна назвати також рекомбінантну стрептококову С5а пептидазу, Streptococcus pyogenes ORF 1224, 1664 і 2452, Chlamydia pnewrioniae ORF Т367 і Т858, фактори і гормони росту. В одному з кращих варіантів здійснення даного винаходу носійним білком є CRM197, нетоксич 23 84728 ний мутант токсину дифтерії із заміною однієї амінокислоти в його первинній послідовністі. Гліцин у положенні амінокислоти 52 молекули заміщується на глутамінову кислоту внаслідок заміни одного нуклеїнокислотного кодону. У результаті цієї заміни білок втрачає ADP-рибозилтрансферазну активність і стає нетоксичним. Він має молекулярну масу 58408 Да. Білок CRM197 виробляється у великих кількостях шляхом рекомбінантної експресії у відповідності з [U.S. Patent 5,614,382], включеним тут шля хом посилання. Кон'югування сахаридів, а також пептидів з білком CRM197 здійснюють шляхом зв'язування через є-аміногрупи лізинових залишків. На декількох промислових продуктах було добре встановлено, що білок CRM197 є чудовим і безпечним носієм для В-клітинних антигенних детермінант. Імуногенні пептиди Використовуваний тут термін "пептидний імуноген" або "гаптен" означає будь-яку білкову або субблочну стр уктур у, її фрагмент або виведений з неї її аналог, що може викликати, полегшувати або індукуватися до вироблення імунної відповіді при введенні її (або його) ссавцю. Цей термін, зокрема, використовується для позначення поліпептидної антигенної детермінанти із будь-якого джерела (бактерії, вірусу або еукаріота), яка може бути зв'язана з носієм за допомогою описаного тут процесу. Такі поліпептидні імуноген/антигенні детермінанти можуть походити із вірусних, бактеріальних або еукаріотних клітин. Для того, щоб пептидні імуногени використовувати як імунотерапевтичні засоби у відвертанні, лікуванні, профілактиці або поліпшенні симптомів різноманітних захворювань людей їх можна кон'югувати з носієм. Джерелами для виведення таких пептидних імуногенів можуть бути: пептид Аβ, що складається із 39-43 амінокислот, краще - із 42 амінокислот, і є головним компонентом характеристичних бляшок хвороби Альцгеймера (AD) [US 4,666,829; Glenner & Wong (l9S4)Biochem.Biophys.Res.Commun.l2Q: 1131, Hardy (1984) TINS 20: 1131; Hardy (1977) TINS 20: 154]; амілоїдні пептиди аміліну; поліпептидний матеріал, що виробляється клітинами панкреатичних острівців, які були залучені до діабету Il типу; генні продукти ліпопротеїнів низької густини, які були залучені до атеросклерозу, та антигенні пептиди, виведені із запальних цитокінів і факторів росту таких, як інтерлейкін 6 (DL-6), фактор α росту пухлин (TNF-α) і GDF-8. Такі еукаріотні пептидні імуногени можуть включати у себе Т-клітинну (CTL) або В-клітинну антигенну детермінанту, відому також як β-амілоїдний пептид або пептид А4. Пептид Аβ відомий також як Β-амілоїдний пептид або пептид А4, див. [US 4,666,829; Glenner & Wong, Biochem. Biophys. Res. Com/mm., 120,1131 (1984)], складається із 39-43 амінокислот і є головним компонентом характеристичних бляшок хвороби Альцгеймера. Пептид Аβ створюють шляхом процесінгу білка APP більшої величини двома ферментами, що звуться β і γ-секретазами [Hardy, TESTS 20,154 (1997)] Відомі мутації в APP, пов'язані з хворобою Альцгеймера, виникають поблизу сайта β- або γ-секретази або усередини Аβ. На 24 приклад, положення 717 є сусіднім до сайта розщеплення APP γ-секретазою при його переробці на Аβ а положення 670/671 є сусідніми до сайта розщеплення β-секретазою. Вважається, що ці мутації викликають хворобу Альцгеймера шляхом взаємодії з реакціями розщеплення, в результаті яких утворюється пептид Аβ збільшуючи таким чином кількість амінокислотної форми 42/43 генерованого пептиду Аβ. Пептид Аβ має ту незвичайну властивість, що він може закріплювати й активувати як класичні, так і альтернативні шляхи активації комплемента. Зокрема, він зв'язується з CIq і, кінець кінцем, з С3bі. Це об'єднання полегшує зв'язування з макрофагами, що веде до активації В клітин. Крім того, С3bі розщеплюється далі, і після цього зв'язується з CR2 на В-клітинах залежним від Т-клітин чином, у результаті чого відбувається 10000кратне збільшення активації цих клітин. Цей механізм спонукає пептид Аβ до генерування імунної відповіді у більшій кількості порівняно з іншими антигенами. Пептид Аβ має декілька природних форм. Людські форми пептиду Аβ позначаються як Аβ39, Аβ40, Аβ41, Аβ42 і Аβ43. Послідовністі цих пептидів та їх взаємозв'язок з APP попередником ілюстровані на Фіг.1 із [Hardy et al., TINS 20,155-158 (1997)]. Наприклад, Аβ42 має таку послідовність: H2N-Asp-AIa-Glu-Phe-Arg-His-Asp-Ser-Gly-TyrGlu-Val-HisHis-Gln-Lys-Leu-Val-Phe-Phe-Ala-Glu-Asp-ValGly-Ser-AsnLys-Gly-Ala-lle-lle-Gly-Leu-Met-Val-Gly-Gly-ValVal-lleAIa-OH (SEQ ID NO. 21). Форми Аβ41, Аβ40 і Аβ39 відрізняються від Аβ42 пропуском залишків Ala, AIa-IIe і AIa-IIe-VaI, відповідно, від С-кінця. Аβ43 відрізняються від Аβ42 наявністю треонінового залишку на С-кінці. Пептидні імуногени, що є фрагментами пептиду Аβ мають для застосування в запропонованих процесах декілька переваг у порівнянні з інтактними молекулами. По-перше, оскільки корисну імуногенну відповідь для лікування хвороби Альцгеймера індукують лише деякі антигенні детермінанти в пептиді Аβ, однакові масові дози їх фрагментів, що містять такі антигенні детермінанти, дають більшу молярну концентрацію корисних імуногенних детермінант, ніж інтактний Аβ. По-друге, деякі пептидні імуногени пептиду Аβ генерують імуногенну відповідь проти амілоїдних депозитів, але не дають суттєвої імуногенної відповіді проти білка APP, із якого виведений пептид Аβ. По-третє, пептидні імуногени пептиду Аβ завдяки їхньому меншому розміру є більш простими у виготовленні, ніж інтактний Аβ. По-четверте, пептидні імуногени Аβ не агрегуються таким самим чином, як інтактний Аβ спрощуючи цим приготування кон'югатів з носіями. Деякі пептидні імуногени Аβ мають послідовності, щонайменше, із 2, 3, 5, 6, 10 або 20 суміжних амінокислот із природного пептиду. Деякі пептидні імуногени мають не більше, ніж 10, 9, 8, 7, 5 або 3 суміжні залишки із Аβ. У кращому варіанті для одержання кон'югатів використовуються пептидні імуногени із N-кінцевої половини пептиду Аβ. 25 84728 До числа кращих пептидних імуногенів належать Аβ1-5, 1-6, 1-7, 1-10, 1-11, 3-7, 1-3 і 1-4. Наприклад, позначення Аβ1-5 стосується N-кінцевого фрагмента, включаючи залишки 1-5 пептиду Аβ Особливо підходящими є Аβ фрагменти, що починаються на N-кінці і закінчуються одним із залишків ділянки 7-11 пептиду Аβ Можна використовувати також фрагмент Аβ1-12, але він є менш підходящим. У деяких процесах таким фрагментом є N-кінцевий фрагмент, відмінний від Аβ1-10. Прийнятними є також фрагменти Аβ13-28, 15-24, 1-28, 25-35, 35-40, 35-42 та інші внутрішні і С-кінцеві фрагменти. Деякі пептиди Аβ за даним винаходом є імуногенними, і після їх введення людини або тварині вони генерують антитіла, що специфічно зв'язуються з однією чи більше антигенними детермінантами на ділянці між залишками 16 і 25 пептиду Аβ. Кра щими при цьому є фрагменти Аβ16-22, 1623, 17-23, 17-24, 18-24 і 18-25. Антитіла, що специфічно зв'язуються з антигенними детермінантами на ділянці між залишками 16 і 25, специфічно зв'язуються з розчинним пептидом Аβ але не зв'язуються з бляшками пептиду Аβ Анти тіла цих типів можуть специфічно зв'язуватися з розчинним Аβ у кровообігу людини або тварини, не зв'язуючись специфічно з бляшковими відкладеннями пептиду Аβ в мозку людини або тварини. Специфічне зв'язування антитіла з розчинним Аβ інгібує вбудовування Аβ у бляшки, становлячись, таким чином, перешкодою для розвинення бляшок у пацієнта та гальмуючи подальше зростання розміру і частоти бляшок, якщо останні до початку лікування вже досягли помітного рівня розвитку. Бажано, щоб уведений фрагмент пептиду Аβ не мав антигенної детермінанти, здатної генерувати Т-клітинну імунну реакцію на цей фрагмент. У загальному випадку Т-клітинні антигенні детермінанти мають довжину більше 10 суміжних амінокислот. Отже, кращі фрагменти пептиду Аβ мають розмір 5-10, краще 7-10 суміжних амінокислот, а найкраще 7 суміжних амінокислот, тобто довжину, достатню для генерування гуморальної імунної відповіді, не викликаючи Т-клітинної імунної реакції. Умова відсутності Т-клітинних антигенних детермінант зумовлена тим, що ці детермінанти не є потрібними для імуногенної активності фрагментів і можуть викликати небажану запальну реакцію у певних підгруп пацієнтів [Anderson et al., (2002) J. Immunol. 168, 3697-3701; Senior (2002) Lancet Neurol. 1,3]. Фрагмент Аβ15-25 і субфрагменти із 7-8 його суміжних амінокислот є кращими, оскільки ці пептиди, безумовно, генерують високу імуногенну відповідь на пептид Аβ. До числа цих фрагментів належать Аβ16-22, Аβ16-23, Аβ16-24, Аβ17-23, Аβ17-24, Аβ18-24 і Аβ18-25. Особливо підходящими субфрагментами Аβ15-25 є такі, що мають довжину із 7 суміжних амінокислот. Наприклад, записом Аβ15-21 позначається фрагмент, який включає у себе залишки 15-21 пептиду Аβ і не має інших залишків цього пептиду. У кращому варіанті такі фрагменти мають довжину із 7-10 суміжних амінокислот. Такі фрагменти можуть генерувати гуморальну імунну відповідь, яка включає антитіла 26 з кінцевою специфічністю. Пептидні імуногени пептидів Аβ потрібно добирати за їхньою активністю в очищенні або відвертанні амілоїдних відкладень, див. заявку [WO 00/72880], включену тут в усій її повноті і для всіх цілей шляхом посилання. Введення N-кінцевих фрагментів пептиду Аβ викликає вироблення антитіл, які розпізнають Аβ-відкладення in vivo та in vitro. У деяких процесах використовуються фрагменти, які не містять щонайменше однієї, а іноді щонайменше 5 або 10 С-кінцевих амінокислот, наявних у природних формах пептиду Аβ. Наприклад, фрагмент, що не має 5 амінокислот від Скінця пептиду Аβ43, містить перші 38 амінокислот від N-кінця пептиду Аβ. Якщо не зазначено іншого, то позначенням Аβ охоплюються всі згадані вище людські амінокислотні послідовності, а також їх аналоги, включаючи алельні, видові та індуковані варіанти. Аналоги, звичайно, відрізняються від пептидів природного походження за одним, двома або кількома положеннями, часто в силу консервативних заміщень. Аналоги, звичайно, демонструють щонайменше 80 або 90% ідентичність послідовністей з натуральними пептидами. Деякі аналоги містять також штучні амінокислоти або модифікації N- чи С-кінцевих амінокислот в одному, двох або декількох положеннях. Наприклад, залишок неприродної аспартанової кислоти в положеннях 1 і/або 7 пептиду Аβ може бути заміщений на ізоаспартанову кислоту. Амінокислотами неприродного походження можуть бути, наприклад, D, альфа, альфадвозаміщені амінокислоти, N-алкільні амінокислоти, молочна кислота, 4-гідроксипролін, гаммакарбоксиглутамат, епсилон-N,N,N-триметиллізин, епсилон-N-ацетиллізин, О-фосфосерин, Nасетилсерин, N-формілметіонін, 3-метилгістидин, 5-гідроксилізин, омега-N-метиларгінін, β-аланін, орнітин, норлейцин, норвалін, гідроксипролін, тироксин, гамма-аміномасляна кислота, гомосерин, цитрулін та ізоаспартанова кислота. До числа імуногенних пептидів належать також аналоги пептиду Аβ та їх фрагменти. Деякими терапевтичними засобами згідно з даним винаходом є повністю-Dпептиди, наприклад, повністю-D-пептид Аβ, фрагмент повністю-D-пептиду Аβ, або аналоги повністю-D-пептиду Аβ або фрагмента повністю-Опептиду Аβ. Фрагменти та аналоги можуть добиратися за профілактичною або терапевтичною ефективністю шляхом випробувань на трансгенних тваринних моделях у порівнянні з необробленими або контрольними плацебо-зразками, як описано в заявці WO 00/72880. До числа пептидних імуногенів належать також поліпептиди більшої довжини, наприклад, імуногенний поліпептид пептиду Аβ разом з іншими амінокислотами. Кращими імуногенними пептидами є, наприклад, злиті білки, що містять сегмент пептиду Аβ приєднаний до гетерологічної амінокислотної послідовністі, що індукує Т-хелперклітинну відповідь проти гетерологічної амінокислотної послідовності і цим самим гуморальну імунну відповідь проти сегмента Аβ Такі поліпептиди можуть добиратися за профілактичною або терапевтичною ефективністю шляхом випробувань на 27 84728 трансгенних тваринних моделях у порівнянні з необробленими або контрольними плацебозразками, як описано в заявці WO 00/72880. Пептид Аβ його аналог, імуногенний фрагмент або інший поліпептид може вводитися пацієнту як у дезагрегатованій, так і в агрегатованій формі. Дезагрегатований пептид Аβ або його фрагменти є мономерними пептидними блоками. Дезагрегатований пептид Аβ або його фрагменти у загальному випадку є розчинними і здатними самоагрегатуватися з утворенням розчинних олігомерів, протофібрилів і ADDL. Олігомери пептиду Аβ та їх фрагменти зазвичай є розчинними й існують переважно у формі альфа-спіралей або випадкових клубків. Агрегованим пептидом Аβ або його фрагменти є олігомери пептиду Аβ або його фрагментів, що зібралися в нерозчинні бета-листові скупчення. Агрегованим пептидом Аβ або його фрагментами є також фібрилярні полімери. Фібрили зазвичай є нерозчинними. Деякі антитіла зв'язують або розчинний пептид Аβ чи його фрагменти, або і агрегований пептид Аβ чи його фрагменти. Деякі антитіла зв'язують як розчинний пептид Аβ чи його фрагменти, так і агрегований пептид Аβ чи його фрагменти. До числа імуногенних пептидів належать також мультимери мономерних імуногенних пептидів. Імуногенні пептиди, відмінні від пептидів Аβ, повинні індукувати імуногенну відповідь проти одного чи більше із вищеперелічених кращих фрагментів пептиду Аβ (наприклад, Аβ1-3, 1-7, 1-10 і 37). Імуногенні пептиди за даним винаходом сполучаються з носієм у відповідності із запропонованим процесом, утворюючи кон'югат. Для створення кон'югата імуногенний пептид може бути приєднаний до носія своїм амінним кінцем, карбоксильним кінцем або обома кінцями. У разі потреби в кон'югаті можуть бути наявними множинні повтори імуногенного пептиду. N-кінцевий фрагмент пептиду Аβ може бути приєднаний його С-кінцем до носійного пептиду з утворенням кон'югата. У таких кон'югатів Nкінцевий залишок фрагмента пептиду Аβ утворює собою N-кінцевий залишок кон'югата. Завдяки цьому такі кон'югати є ефективними в індукуванні антитіл, що зв'язуються з антигенною детермінантою, котрій потребується N-кінцевий залишок пептиду Aβ для того, щоб бути у вільній формі. Деякі імуногенні пептиди за даним винаходом містять множину повторів N-кінцевого сегмента пептиду Аβ, зв'язаного на С-кінці з однією чи більше копіями носійного пептиду, утворюючи кон'югат. Nкінцевий фрагмент пептиду Аβ, вбудованого в такі кон'югати, іноді починається на Аβ1-3 і кінцях на Аβ7-11. Кращими N-кінцевими фрагментами пептиду Аβ є Аβ1-7, 1-3, 1-4, 1-5 і 3-7. Деякі кон'югати містять різні N-кінцеві сегменти пептиду Аβ в тандемі. Наприклад, той чи інший кон'югат може містити Аβ1-7, за яким йде Aβ1-3, приєднаний до носія. У деяких кон'югатів N-кінцевий сегмент пептиду Aβ приєднаний до носійного пептиду принаймні на його N-кінці. Таке саме різноманіття N-кінцевих сегментів пептиду Аβ може використовуватися в 28 С-кінцевому зв'язку. Деякі кон'югати містять носійний пептид, приєднаний до N-кінця N-кінцевого сегмента пептиду Аβ який, у свою чергу, приєднаний до одного чи більше додаткових N-кінцевих сегментів пептиду Аβ у тандемі. У кращому варіанті такі імуногенні Аβ-фрагменти, кон'юговані з відповідним носієм, індукують імуногенну відповідь, специфічно спрямовану на даний Аβ-фрагмент, не спрямовуючись при цьому на інші фрагменти пептиду Аβ. Імуногенними пептидами за даним винаходом можуть бути також імуногенні гетеролопчні пептиди. У деяких імуногенних пептидів фрагмент пептиду Аβ приєднаний до носія, утворюючи імуногенний гетерологічний пептид. Цей гетерологічний пептид приєднується до носія у відповідності з процесом за даним винаходом і утворює кон'югат. Деякі з таких імуногенних гетерологічних пептидів містять фрагменти пептиду Аβ, зв'язані з такими антигенними детермінантами правцевого токсоїду, як описано в [US 5,196,512, EP 378,881 і EP 427,347]. В разі потреби імуногенний пептид може бути з'єднаний з однією чи багатьма копіями носія, наприклад, обома кінцями - N і C носія, утворюючи імуногенний гетерологічний пептид. Інші з таких імуногенних гетерологічних пептидів містять фрагменти пептиду Аβ з'єднані з носійними пептидами, описаними в [US 5,736,142]. Наприклад, імуногенний гетерологічний пептид може містити, Аβ1-7, за ним - Aβ1-3, а за ним - носій. Як приклади таких імуногенних гетерологічних пептидів можна назвати: Aβ1-7/правцевий токсоїд 830-844+947-967 у лінійній конфігурації DAEFRHDQYIKANSKFIGITELFNNFTVSFWLRVPKVSASHLE (SEQ ID NO.. 24) Пептиди, описані в [US 5,736,142] (усі в лінійних конфігураціях): PADRE/Aβ1-7: AKXVAAWTLKAAA-DAEFRHD (SEQ ID NO.:26) Aβ1-7´3/PADRE: DAEFRHD-DAEFRHD-DAEFRHDAKXVAAWTLKAAA (SEQ ID NO. 27) PADRE/Aβ1-7´3: AKXVAAWTLKAAA-D AEFRHD-DAEFRHDDAEFRHD (SEQ ID NO.:28) Aβ1-7/PADRE: DAEFRHD-AKXVAAWTLKAAA (SEQ ID NO.: 29) Аβ1-7/фрагмент альбуміну: DAEFRHD-ISQAVHAAH AEINEAGR (SEQ ID NO.:30) Aβ4-10/фрагмент альбуміну: FRHDSGY-ISQAVH AAH AEINEAGR (SEQ ID NO.:31) Аβ3-9/фрагмент альбуміну: EFRHDSG-ISQAVH AAH AEINEAGR (SEQ ID NO.:32) HA307-319/Aβ1-7´3: PKYVKQNTLKL AT-D AEFRHD-DAEFRHDDAEFRHD (SEQ ID NO.:33) Аβ1-7/HA307-319/Aβ1-7: DAEFRHD-PKYVKQNTLKL AT-D AEFRHD (SEQ ID NO.:34) Aβ1-7´3/H A307-319: 29 84728 DAEFRHD-DAEFRHD-DAEFRHDPKYVKQNTLKL AT (SEQ ID NO.:35) Aβ1-7´2/H A307-319: DAEFRHD-DAEFRHD-PKYVKQNTLKL AT (SEQ ID NO.:36) Aβ1-7/HA307-319/CS малярії/ТТ830-844/ТТ947967 /Аβ1-7 DAEFRHD-PKYVKQNTLKL ATEKKIAKMEKASSVFNV-QYIKANSKFIGITElFNNFTVSFWLRVPKVSASHLE-DAEFRHD (SEQ ID NO.:37) Aβ1-7´3/TT830-844/C/TT947-967 DAEFRHD-DAEFRHD-DAEFRHDQYIKANSKFIGITEL-CFNNFTVSFWLRVPKVSASHLE (SEQ ID NO.: 38) Aβ1-7/TT 830-844C/TT 947-967 DAEFRHDQYIKANSKFIGITELCFNNFTVSFWLR VPKVSASHLE (SEQ ID NO.:39) Aβ1-7/TT 830-844/C/TT947-967/Aβ1-7 DAEFRHD-QYIKANSKFIGITEL-CFNNFTVSFWLRVPKVSASHLE-DAEFRHD (SEQ ID NO.:20) Деякі імуногенні гетерологічні пептиди містять мультимер із імуногенних пептидів, що описуються формулою 2х, де х є ціле число від 1 до 5. У кращому варіанті число х дорівнює 1, 2 або 3, серед яких число 2 є найкращим. Якщо х є два, то такий мультимер має чотири імуногенні пептиди, зв'язані у кращій конфігурації під назвою МАР4 [US 5,229,490]. Такі імуногенні пептиди з'єднуються далі з носієм у відповідності з процесом за даним винаходом, утворюючи кон'югат. МАР4 конфігурація показана нижче. У цій конфігурації розгалужені структури одержуються шляхом ініціації пептидного синтезу як на N-кінці, так і на амінах лізину бічних ланцюгів. Залежно від того, скільки разів лізин вбудовується в цю послідовність і дозволяє на розгалуження, стільки N-кінців має утворювана в результаті структура. В ілюстрованому тут прикладі утворено чотири ідентичні N-кінці на розгалуженому лізинвмісному центрі. Така множинність значно підсилює реактивність спорідненої B-клітини. Де Peptide - пептид Імуногенними гетерологічними пептидами такого типу є, наприклад: Аβ1-7/правцевий токсоїд 830-844 у конфігурації МАР4: DAEFRHD-QYIKANSKFIGITEL (SEQ ID NO.:22) Аβ1-7/правцевий токсоїд 947-967 у конфігурації МАР4: DAEFRHD-FNNFTVSFWLRVPKVSASHLE (SEQ ID NO.:23) Аβ3-9/правцевий токсоїд 830-844 у конфігурації МАР4: EFRHDSG-QYIKANSKFIGITEL (SEQ ID NO.:25) DAEFRHD-QYIKANSKFIGITEL у смолі з 2 бічними ланцюгами 30 Пептид Аβ його аналог, активний фрагмент або інший поліпептид може вводитися пацієнту у сполученій чи мультимерній формі, або у дисоційованій формі. Терапевтичними засобами можуть бути також мультимери із мономерних імуногенних засобів. Засоби, відмінні від пептидів Аβ повинні викликати імуногенну відповідь проти одного чи більше із кращих фрагментів пептиду Аβ, перелічених вище (наприклад, 1-10, 1-7, 1-3 і 3-7), і можуть бути також кон'юговані з носієм у відповідності з процесом за даним винаходом. У кращому варіанті здійснення винаходу такі засоби, кон'юговані з відповідним носієм, індукують імуногенну відповідь, що є специфічно спрямованою на один із цих фрагментів, не будучи при цьому спрямованою на інші фрагменти пептиду Аβ. Для полегшення кон'югації пептидного імуногена з носієм можна до кінців антигенних детермінант приєднувати додаткові амінокислоти. Такі додаткові залишки можуть також використовуватися для модифікації фізичних або хімічних властивостей пептидного імуногена. Амінокислоти, наприклад, тирозин, цистеїн, лізин, глутамінова або аспартанова кислота і т.п.можуть приєднуватися до C- або N-кінця пептидного імуногена. Крім того, можуть вводитися також пептиді лінкери, що містять такі амінокислоти, як гліцин та аланін. Антигенні детермінанти можуть відрізнятися від природної послідовністі і бути модифікованими ацилюванням їхніх кінцевих NH2-груп, наприклад, шляхом ацетилювання алканоїлом (C1-C20) або тіогліколілом, амідуванням карбоксильного кінця, наприклад, аміаком, метиламіном, тощо. У деяких випадках ці модифікації можуть давати сайти для зв'язування з основою або іншою молекулою. Пептидні імуногени для створення кон'югатів згідно з даним винаходом за допомогою описаного тут процесу можуть об'єднуватися в комбінації шляхом зв'язування з утворенням полімерів (мультимерів), або в композиції без зв'язування, наприклад, шляхом змішування. Там, де пептид з'єднується з ідентичним пептидом у гомополімер, наявними є численні повтори блоків антигенних детермінант. Наприклад, за допомогою методу множинних антигенних пептидів (МАР), можна конструювати полімери, що містять як CTL, так і гуморальні пептиди і поліпептиди. "CTL антигенною детермінантою" є детермінанта, виведена із добраних детермінантних ділянок потенційних антигенів-мішеней. Якщо пептиди є різними, наприклад, у суміші із різних вірусних субтипів, то для цього передбачені різні антигенні детермінанти в межах субтипу, різні унікальні специфічності (контрольовані унікальною специфічністю головного комплексу гістосумісності (HLA)), або пептиди, що містять Т-хелпер-клітинні антигенні детермінанти, гетерополімери з повторами блоків. Окрім ковалентних зв'язків, передбачаються також нековалентні зв'язки, здатні утворювати міжмолекулярні та внутрішньоструктурні з'єднання. Такого типу пептидні імуногени та їх аналоги виготовляються шляхом твердофазного пептидного синтезу або рекомбінантної експресії, або ж 31 84728 одержуються із природних джерел. Промисловістю і, зокрема, фірмою Biosystems, м. Фостер, шт. Каліфорнія, виробляються підходящі для цього автоматичні синтезатори пептидів. Рекомбінантна експресія може здійснюватися в бактеріальних клітинах (наприклад, E. соlі), дріжджах, клітинах комах або клітинах ссавців. Методи рекомбінантної експресії описані, наприклад, у [Sambrook et al., Molecular Cloning: A Laboratory Manual (Cold Spring Harbor Press, NY, 2nd ed., 1989)]. Деякі імуногенні пептиди можуть бути придбані також комерційним шляхом (від таких виробників, наприклад, як American PeptidesCompany, Inc., Sunnyvale, CA, і California Peptide Research, Inc., Napa, CA). Підходящі пептидні імуногени можуть бути добрані також шляхом скринінгу статистични х бібліотек пептидів або інших сполук. Для багатьох типів сполук, які можуть синтезуватися в поступових процесах, можна створювати комбінаторні бібліотекі. До таких сполук належать поліпептиди, бетавиткові міметики, гормони, олігомерні N-заміщені гліцини, олігокарбамати і т.п.Великі комбінаторні бібліотекі сполук можуть бути сконструйовані методом кодованих синтетичних бібліотек (ESL), описаним у заявках [WO 95/12608, WO 93/06121, WO 94/08051, WO 95/35503 і WO 95/30642], кожна з яких включена тут для всі х цілей шляхом посилання. Пептидні бібліотеки можуть бути виготовлені також методами відображення фагів, як описано, наприклад, у [Devlin, WO 91/18980]. Дериватизація і кон'югація імуногенного пептиду для білкового носія Сайт приєднання пептидного імуногена до білкового або поліпептидного носія і природа зшивального агента, використовуваного для приєднання пептидного імуногена до носія є обидва важливими факторами специфічності генерованих проти нього антитіл. Для правильного розпізнавання пептидний імуноген повинен бути з'єднаний з носієм у відповідній орієнтації. Для наступного за цим розпізнавання антитілом вільних пептидних імуногенів без носія кон'югат пептидного імуногена з білковим або поліпептидним носієм повинен представляти пептидні імуногени у відкритій і доступній формі. Оптимальна орієнтація часто досягається шляхом спрямування реакції зшивання на специфічні сайти пептидних імуногенів. Одним зі шляхів для досягнення цього пептидним імуногеном є приєднання кінцевого цистеїнового залишку під час пептидного синтезу. У результаті створюється сульфгідрильна група на одному кінці пептиду для кон'югації його з носієм. Зшиванням через цю групу забезпечується приєднання пептидного імуногена лише на одному кінці і, отже, його узгоджена орієнтація. Метою кон'югації пептидного імуногена з носієм є не підримання природного стану або стабільності носія, а представлення гаптена імунній системі якомога кращим способом. Для здійснення цього використовують хімію кон'югації, яка дозволяє визначати отримувані в результаті титр, а фінність і специфічність антитіл, вироблених проти цього гаптена. У деяких випадках може бути важливим провести правильний добір зшивального 32 агента, що містить достатньо довгу спейсерну групу для представлення даного антигена нерестриктованим чином. Може бути важливим також регулювати густин у пептидного імуногена на поверхні носія. Заміщення занадто малим пептидним імуногеном може призвести до слабкої відповіді і навіть до її відсутності. З іншого боку, занадто велика густина пептидного імуногена фактично може викликати імунологічну супресію і зниження відповіді. Крім того, зшивальний агент сам може викликати небажану імунну відповідь. Ці наслідки потрібно враховувати при доборі не тільки відповідних зшивальних реагентів, але також відповідного кількісного співвідношення між білковим або поліпептидним носієм і пептидним імуногеном. Для приєднання білкових або пептидних носіїв до пептидних імуногенів можуть використовуватися найрізноманітніші засоби. Іонні взаємодії є можливими через кінці або через є-аміногрупи лізину Можливим є також водневий зв'язок між бічними групами залишків і пептидним імуногеном. І нарешті, підви щити стабільність приєднання можуть конформаційні взаємодії між білковими або пептидними носіями й імуногенним пептидом. Кон'югати "пептидний імуноген - носій" з успіхом створювалися при використанні різноманітних зшивальних агентів, включаючи зшивальні агенти нульової довжини, гомобіфункціональні та гетеробіфункціональні зшивальні агенти. Найменшими серед доступних реагентів для біокон'югації є так звані "крос-лінкери нульової довжини". Ці сполуки опосередковують кон'югацію двох молекул шляхом формування зв'язку, що не містить додаткових атомів. Таким чином, один атом молекули є спейсером. У багатьох схемах кон'югації у кінцевому комплексі складові зв'язуються між собою завдяки хімічним компонентам, які додають чужорідну структур у речовинам, що зшиваються. У деяких випадках практичного застосування наявність цих доданих лінкерів може ставати шкідливим чинником для цільового використання. Наприклад, при одержанні кон'югатів "пептидний імуноген - носій" цей комплекс утворюється з метою генерації імунної відповіді на приєднаний гаптен. Іноді частина антитіла, виробленого цією відповіддю, має специфічність до зшивального агента, використовуваного в кон'югації. Зшивальні агенти нульової довжини виключають можливість виникнення перехресної реактивності цього типу завдяки тому, що вони опосередковують прямий зв'язок між двома речовинами. Гомобіфункціональні реагенти, які були першими зшивальними агентами, що використовувалися для модифікування і кон'югації макромолекул, складалися із біреактивних сполук, що містили однакові функціональні групи на обох кінцях [Hartman and WoId, 1966]. Ці реагент могли зв'язувати один білок з іншим шляхом ковалентної реакції між однаковими загальними групами в обох молекулах. Отже, є-аміни лізину або N-кінцеві аміни одного білка могли зшиватися з однаковими функціональними групами в обох білках просто шляхом змішування їх дво х разом при наявності гомобіфункціонального реагента. Гетеробіфункціональні реагенти кон'югації міс 33 84728 тять дві різні реактивні групи, які можуть з'єднуватися з двома різними функціональними мішенями в білках та інши х макромолекулах. Наприклад, одна частина крос-лінкера може містити амінну реактивну гр упу, у той час як інша його частина може складатися із сульфгідрильної реактивної групи. У результаті виникає можливість спрямовувати реакцію зшивання на цільові частини молекул-мішеней і, таким чином, краще керувати процесом кон'югації. Гетеробіфункціональні реагенти використовуються для зшивання білків та інших молекул у двоабо триступінчастому процесі, який обмежує ступінь полімеризації, що часто виникає при використанні гомобіфункціональних крос-лінкерів. Сьогодні відомо багато методів зв'язування пептидних імуногенів з білковими або поліпептидними носіями за допомогою гомобіфункціональних або гетеробіфункціональних крос-лінкерів нульової довжини. Більшість з цих методів спрямована на створення амінних, амідних, уретанових, ізотіосечовинних та дисульфідних зв'язків або в деяких випадках - тіоетерів. У більш загальному методі св'язування білків або пептидів з пептидами використовуються біфункціональні зшивальні реагенти. Це є малі спейсер-молекули, які мають активні групи на кожному кінці. Ці спейсер-молекули можуть мати на своїх кінцях ідентичні або різні активні групи. Найбільш загальними активними функціональними групами, зв'язувальними групами й утворюваними зв'язками є: 1. Альдегід-аміно®вторинний амін 2. Малеімід-сульфгідрил®тіоетер З.Сукцинімід-аміно®амід 4. Імідатні естери-аміно®амід б.Фенілазиди-аміно®феніламін 6. Галоїдацил-сульфгідрил®тіоетер 7. Піридилдисульфідисульфгідрил®дисульфід 8. Ізотіоціанат-аміно®ізотіосечовина. Реактивність даного носійного білка з погляду його здатності до модифікації зшивальним агентом таким чином, щоб він був кон'югований з пептидним імуногеном, визначається його амінокислотним складом і місцем індивідуальних амінокислот у послідовності у тримірній структурі цих молекул, а також амінокислотним складом пептидного імуногена. [0109] У тих випадках, коли лінкери ("L") розташовуються між білковими або пептидними носіями та іншими пептидами (наприклад, між білковими або пептидними носіями і пептидним імуногеном), спейсери, звичайно, вибирають серед Ala, GIy або інших нейтральних спейсерів неполярних амінокислот або нейтральних полярних амінокислот. У деяких варіантах здійснення винаходу нейтральним спейсером є AIa. Цілком зрозуміло, що наявному, в разі потреби, спейсеру не обов'язково складатися із таких самих залишків і, таким чином, ним може бути гетеро- або гомоолігомер. Типовими спейсерами є, наприклад, гомоолігомери аланіну (AIa). У разі наявності спейсера він, зазвичай, складається принаймні з одного чи двох, а частіше - із трьох-шести залишків. В інших варіантах здійснення винаходу білковий або полі 34 пептидний носій є кон'югований з пептидним імуногеном таким чином, що носій займає положення на амінному кінці. Пептид може сполучатися за посередництвом нейтрального лінкера, наприклад, AIa-AIa-AIa і т п., а в кращому варіанті, крім того, містить ліпідний залишок, наприклад, пальмітинову кислоту і т.п., що приєднується до альфа- й епсилон-аміногруп Lys-залишку ((PAM)2L yS), приєднаного до амінного кінця даного пептидного кон'югата, звичайно, Ser-Ser містком і т.п. Відповідно до деяких аспектів даного винаходу пептидним імуногеном є фрагмент пептиду Аβ, вибраний із групи, що складається із Аβ1-5-L, Аβ17-L, Аβ1-9-L і Аβ1-12-L Відповідно до деяких аспектів даного винаходу містком є GAGA (SEQ ID NO:10). Для полегшення кон'югації пептидного імуногена з носієм можуть добавлятися додаткові амінокислоти на кінцях антигенних детермінант. Додаткові залишки можуть використовуватися також для модифікації фізичних або хімічних властивостей пептидного імуногена. Такі амінокислоти, як тирозин, цистеїн, лізин, глутамінова або аспартанова кислота і т.п., можуть приєднуватися до Cабо N-кінця пептидного імуногена. Крім того, вводитися можуть також пептидні лінкери, що містять таку амінокислоту, як гліцин та аланін. У додаток до вищезазначеного, антигенні детермінанти можуть відрізнятися від природної послідовності модифікаціями шляхом ацилювання кінцевих NH2груп, наприклад, алканоїлом (С1-С20) або тіогліколілом, амідування карбоксильних кінців, наприклад, аміаком, метиламіном, і т.д. У деяких випадках ці модифікації можуть надавати сайти для зв'язування з основою або іншою молекулою. Відповідно до деяких аспектів даного винаходу пептидним імуногеном є фрагмент пептиду Аβ, вибраний із групи, що складається із Аβ1-5-С, Аβ17-С, Аβ1-9-С і Аβ1-12-С, де C є цистеїновий амінокислотний залишок. Відповідно до деяких аспектів даного винаходу пептидним імуногеном є фрагмент пептиду Аβ вибраний із групи, що складається із Аβ1-5-L-C, Аβ1-7-L-C, Аβ1-9-L-C і Аβ1-12-L-C. Пептидний імуноген приєднується до білкового або пептидного носія або безпосередньо або за посередництвом лінкера на амінному юнці чи на карбоксильному кінці пептидного імуногена. Амінний кінець пептидного імуногена або білкового чи пептидного носія може бути ацетильований. Крім того, кон'югат пептидного імуногена з білковим чи пептидним носієм може бути з'єднаний з певними алканоільними (C1-C20) ліпідами за посередництвом одного чи більше зшивальних залишків, наприклад, GIy, GIy-GIy, Ser, Ser-Ser, як описано нижче. Як ліпідна частина використовуватися можуть також холестерин, жирні кислоти і т п. Пептидні імуногени можуть з'єднуватися з носієм шляхом хімічного зшивання. Зшивання імуногена з носієм може здійснюватися методом утворення дисульфідних містків, використовуючи Nсукцинімідил-3-(2-піридил-тіо)пропіонат (SPDP) [Carlsson, J. et аl. (1978) Biochem. J., 173: 723] і сукцинімідил 4-(N-малеімідметил)циклогексан-1карбоксилат (SMCC) (якщо пептид не має сульфгідрильної групи, вона може бути створена шляхом 35 84728 додавання цистеїнового залишку до гаптена). Ці реагенти створюють дисульфідний місток між собою і цистеїновими залишками пептиду на одному білку й амідний місток через є-аміногрупу лізину або іншу вільну аміногрупу іншої амінокислоти. Різноманітні реагенти, що дозволяють створювати такі дисульфідні та амідні сполучення, описані в [Immune. Rev. 62: 85 (1982)]. Інші біфункціональні зв'язувальні агенти утврюють тіоетерні, а не дисульфідні містки. До числа таких агентів належать реактивний естер 6-малеімідкапронової кислоти, 2-бромоцтової кислоти і 2-йодооцтової кислоти, 4(N-малеімідометил)циклогексан-1-карбонова кислота. Карбоксильні групи можуть активуватися шляхом об'єднання їх із сукцинімідом або 1гідроксил-2-нітро-4-сульфоновою кислотою, сіллю натрію. Найчастіше лізинові залишки є найбільш численними серед амінокислотних залишків, що складають носійні білки. Отже, саме ці залишки модифікуються за допомогою зшивальних реагентів для створення нуклеофільних сайтів, які після цього сполучаються з гаптеном. Це сполучання здійснюється через будь-який із гідрофільних бічних ланцюгів у гаптенових молекулах, що є хімічно активними. До їх числа входять, принаймні, гуанідильна група аргініну, (карбоксильні групи глутамату й аспартанової кислоти, сульфгідрильна група цистеїну і е-аміногрупа лізину. Моди фікування білків таким чинм, щоб вони могли сполучатися з іншими частинами, здійснюється за допомогою зшивальних реагентів, які реагують з одним із бічних ланцюгів у молекулах білкового носія або гаптена. У відповідності з одним із аспектів даного винаходу носійний білок з молекулою лінкера або без неї функціоналізують (дериватизують) реагентом, який уводить реактивні сайти у молекули носійного білка, що піддаються подальшому модифікуванню з метою введення нуклеофільних груп.В одному з варіантів здійснення винаходу проводять реакцію носія з галоїдацетилувальним реагентом, який реагує переважно з множиною таких функціональних груп в амінокислотних залишках білків, як сульфгідрильна група цистеїну, первинна єамінна група лізинового залишку, а-кінець а-амінів, тіоетер метіоніну та обидва азоти імідазоїльних бічних ланцюгів гістидину [Gurd, 1967]. У кращому варіанті первинні 6-амінні групи лізинових залишків носійного білка є дериватизованими b Nгідроксисукцинімідилбромацетатом для створення бромацетильованого носія. Кон'югацію пептидного імуногена з активованим білковим носієм проводили шляхом повільного додавання активованого носія до розчину, що містив пептидний імуноген. За допомогою процесу згідно з даним винаходом пептидні імуногени, розглянуті вище, в розділі В, можуть сполучатися в кон'югатах з будь-яким із носіїв, розглянутих вище, в розділі А. Кон'югати, вироблені за допомогою процесу згідно з винаходом, можуть використовува тися як імуногени для генерування антитіла проти Аβ як у пасивній, так і в активній імунотерапії. Крім того, пептид Аβ або його фрагмент, з'єднані з носієм, можуть вводитися лабораторній тварині для вироблення монокло 36 нальних антитіл проти пептиду Аβ. У відповідності з одним із аспектів даного винаходу запропонований кон'югат вибирають із групи, що складається із Аβ1-7-CRM197, (Аβ17´3)CRM197 і (Аβ1-7´5)-CRM197. У відповідності з одним із аспектів даного винаходу запропонований кон'югат вибирають із групи, що складається із CRM197-Аβ1-5, CRM197- Аβ1-7, CRM197-Аβ1-9 і CRM197. У відповідності з іншим аспектом даного винаходу запропонований кон'югат вибирають із групи, що складається із Аβ1-5-C-CRM197 , Аβ1-7-CCRM197, Аβ1-9-C-CRM197 і Аβ1-12-C-CRM197, Аβ1623-C-CRM197, Аβ17-24-C-CRM197 , Аβ18-25-CCRM197, CR M197-C-Аβ16-23, CRM197-C-Aβ17-24, CRM197-С-Аβ18-25, Аβ16-22-C-CRM197, Аβ17-23-CGRM197 , Аβ18-24-C-CRM197, CRM197-C-Aβ16-22, CRM197-C-Аβ17-23 і CRM197-C-Aβ18-24. У відповідності з ще одним аспектом даного винаходу запропонований кон'югат вибирають із групи, що складається із Аβ1-5-L-C-CRM197, Aβ1-7-L-CCRM197, Аβ1-9-L-C-CRM197 і Аβ1-12-L-C-CRM197. Кепінг Недоліком застосування реагентів сполучання, що вводять хімічно активні сайти у бічні ланцюги хімічно активних молекул амінокислот носія і/або молекул гаптена, є те, що хімічно активні сайти, якщо вони не нейтралізовані, можуть вільно реагувати з будь-якими небажаними молекулами як in vitro, так і in vivo. Згідно з процесом за даним винаходом, кепінг непрореагованих функціональних груп здійснюють шляхом реакції кон'югатів, що містять вільні реактивні групи, з реагентами, які дезактивують тобто "кепують" хімічно активні групи. До числа типових реагентів дезактивації або кепінгу для застосування в кон'югації згідно з даним винаходом є цистеїнамін, Nацетилцистеїнамін, та етаноламін. Іншим чином кепінг проводять за допомогою реакції з аміаком або бікарбонатом амонію, які є здатними перетворювати галоїдацетильні групи на аміноацетильні групи. Кепінг здійснюють також при лужній кислотності рН (9,0-9,8) за допомогою гідроксиду натрію або карбонату натрію, котрі перетворюють галоїдацетильні групи на гідроксіацетильні групи. Однією із потенційних переваг перетворення галоїдацетильних гр уп на аміноацетильні або гідроксіацетильні групи, на противагу реакції з цистеїнаміновими сполуками, етаноламіном і т.п., є можливість уводити функціональні групи відносно малих розмірів шляхом реакції з аміаком або гідроксидом чи карбонатом. Отримувані в результаті таких перетворень кеповані функціональні групи, наприклад аміноацетил або гідроксіацетил, привносять відносно мало збурень у білкову частину носія кон'югата. Кепований кон'югат "пептидний імуноген - носійний білок" очищають у відповідності з потребою відомими методами, включаючи хроматографію (гель-фільтраційну, іонообмінну, на основі гідрофобної взаємодії або афінності), діаліз, надфільтрацію-діафільтрацію, селективну преципітацію сульфатом амоню або спиртом, і т.п. Імуногенні кон'югати і склади Кеповані кон'югати "пептидний імуноген - носійний білок" вводять в імуногенному складі ссавцям і, зокрема, людям у профілактичних і/або те 37 84728 рапевтичних цілях. Кон'югати за даним винаходом використовуються для викликання і/або підсилення імунних відповідей проти імуногенів Наприклад, кон'югати CTL-носій використовуються у лікуванні і/або профілактиці вірусноінфекційних захворювань, амілоїдогенних захворювань, раку і т.д. Як альтернатива використовуються також кон'югати типу "поліпептидний імуноген - носій", які викликають гуморальні імунні відповіді. При терапевтичному застосуванні кон'югат за даним винаходом уводять пацієнту, що вже страждає на амілоїдогенну хворобу, наприклад, на хворобу Альцгеймера. Пацієнтів стан яких є у фазі інкубації або в гострій фазі захворювання, можна, в разі потреби, лікувати за допомогою запропонованих кон'югатів як відокремлено, так і водночас з іншими курсами лікування. При терапевтичному застосуванні композицію за даним винаходом уводять пацієнту в кількості, достатній для ефективної CTL (ЦТЛ) відповіді або гуморальної відповіді на мікроорганізм, амілоїдну бляшку або на пухлинний антиген, експресований на раковій клітині, і для лікування або, щонайменше, часткового припинення розвитку хвороби, симптомів і/або ускладнень. Кількість композиції, потрібна для здійснення цього, зветься "терапевтично ефективною дозою". Ефективні кількості частково залежать від складу пептиду, способу введення, стадії розвитку і тяжкості даного захворювання, маси тіла і загального стану здоров'я пацієнта, а також від власної думки лікаря, що призначає лікування. Терапевтично ефективні кількості імуногенних композицій за даним винаходом у загальному випадку лежать в інтервалі, що відповідає початковій імунізації для терапевтичного або профілактичного призначення, тобто приблизно від 0,1мкг до 10000мкг пептиду для пацієнта з масою тіла 70 кг, зазвичай приблизно від 0,1 до 8000мкг, краще приблизно від 0,1 до 5000мкг, і найкраще - приблизно від 0,1 до 1000мкг. Слідом за цими дозами призначаються бустер-дози приблизно від 0,1мкг до 1000мкг пептиду відповідно до режиму ревакцінації протягом курсу лікування від декількох тижнів до декількох місяців, залежно від реакції пацієнта та його стану за даними вимірювань специфічних імунних відповідей. У профілактичних цілях даний винахід використовується для відвертання і/або поліпшення амілоїдогенного захворювання. Ефективні кількості при цьому є такими, як зазначено вище. Крім того, для фахівця в даній галузі не буде складним вирішити питання регулювання або пристосування профілактичного лікування до конкретних умов, наприклад, шляхом ревакцинації і регулювання доз та режимів їх прийому. Терапевтичний курс можна починати при перших ознаках хвороби. Слідом за цим застосовують бустер-дози до тих пір, поки розвинення хвороби не буде зупинено або повернено назад, або поки симптоми не будуть ґрунтовно послаблені, і протягом певного періоду часу після цього. Імуногенні композиції за даним винаходом у терапевтичних або профілактичних цілях можуть уводитися пацієнту за допомогою засобів парен 38 терального, місцевого, внутрішньовенного, перорального, підшкірного, внутрішньоартеріального, інтракраніального, інтраперитонеального, інтраназального або внутрішньом'язового введення для профілактичного і/або терапевтичного лікування. Одним із типових шляхів уведення імуногенного агента є підшкірний, хоча так само ефективними можуть бути також інші способи. Іншим поширеним способом уведення є внутрішньом'язові ін'єкції. Ін'єкції цього типу найчастише роблять у м'язи рук або ніг. У деяких способах, наприклад, інтракраніальні ін'єкції лікувальних засобів роблять безпосередньо в тканину, де акумулювалося відкладення. Внутрішньом'язові ін'єкції або внутрішньовенні вливання є кращими для введення антитіл. У деяких способах терапевтичні антитіла спеціального призначення вводять шляхом ін'єкцій безпосередньо в череп.Відмінна легкість введення імуногенних композицій за даним винаходом робить їх особливо підходящими для перорального способу введення. Винаходом передбачені також імуногенні композиції для парентерального введення, які містять розчини пептидів або кон'югатів, розчинених або суспендованих у підходящому носії, яким у кращому варіанті є вода. У композиціях за даним винаходом можуть використовуватися найрізноманітніші розчинники, ексципієнти і буфери, включаючи, наприклад, воду, буферизовану воду, буферизований фосфатом фізіологічний розчин, 0,3% гліцин, гіалуронову кислоту і т.п.Стерилізація запропонованих композицій може здійснюватися за допомогою звичайних, добре відомих методів і засобів або стерилізаційних фільтрів. Отримувані в результаті такої обробки водні розчини можуть пакуватися для застосування в їх незмінному стані або піддаватися ліофілізації, а ліофілізовані препарати перед їх уведенням об'єднуватися зі стерильним розчином. Композиції можуть містити фармацевтично прийнятні допоміжні речовини, що дозволяють у відповідності з потребою наблизитися до фізіологічних умов. Такими речовинами можуть бути буферні агенти, агенти регулювання тонусу, змочувальні агенти і т.п., наприклад, ацетат натрію, лактат натрію, хлорид натрію, хлорид калію, хлорид кальцію, монолаурат сорбіту, триетаноламінолеат і т.д. У твердих композиціях можуть використовуватися звичайні нетоксичні тверді носії, а саме, наприклад, фармацевтичні сорти маніту, лактози, крохмалю, стеарату магнію, натрійсахарин, тальк, целюлоза, глюкоза, сахароза, карбонат магнію і т.п.Для перорального введення фармацевтично прийнятну нетоксичну композицію готують шляхом об'єднання будь-яких звичайно використовуваних ексципієнтів, наприклад, вищеперелічених носіїв, і в загальному випадку 10-95% активного інгредієнта, тобто одного чи більше кон'югатів за даним винаходом, концентрація яких у кращому варіанті становить 25-75%. Концентрація імуногенних композицій за даним винаходом фармацевтичних препаратах може варіювати в широких межах, тобто від менш, ніж приблизно 0,1%(мас), зазвичай, щонайменше, від приблизно 2%(мас.) до 20%(мас.) - 50%(мас.) або 39 84728 більше і вибирається у першу чергу залежно від об'єму рідини, в'язкості і т.д. у відповідності з вибраним способом введення. Кон'югати за даним винаходом можуть уводитися також за посередництвом ліпосом, які постачають кон'югати в цільові тканини, наприклад, лімфоїдну тканину або селективно в інфіковані клітини, а також збільшують період напіввиведення пептидного складу. Ліпосоми можуть мати форму емульсій, пінки, міцел, нерозчинних моношарів, рідких кристалів, фосфоліпідних дисперсій, ламелярних шарів і т.п.У ци х препаратах призначений для постачання склад вбудований у формі частини ліпосом відокремлено або у сполученні з молекулою, що зв'язується, наприклад, з рецептором, який превалює в лімфоїдних клітинах. Такими молекулами можуть бути моноклональні антитіла, що зв'язуються з антигеном CD45 або з іншими терапевтичними чи імуногенними складами. Отже, ліпосоми наповнені бажаним складом згідно з даним винаходом можуть бути спрямовані в місце розташування лімфоїдних клітин, куди ліпосоми після цього постачають розміщені в них терапевтичні та імуногенні пептидні склади. Ліпосоми для застосування в даному винаході готують зі стандартних ліпідів, що утворюють везикули, тобто в загальному випадку - із нейтральних і негативно заряджених фосфоліпідів і стерину, наприклад, холестерину. При виборі ліпіду в загальному випадку керуються міркуваннями розміру ліпосом, кислотостійкості і стабільності ліпосом у кровотоку. Для готування ліпосом існують численні методи, описані, наприклад, у публікаціях [Szoka, et al., Ann. Rev. Biophys. Bioeng. 9: 467 (1980), U.S. Pat №№4,235,871, 4,501,728, 4,837,028, 5,019,369], включених тут шля хом посилання. Для аерозольного введення композиції за даним винаходом постачаються переважним чином у тонко розподіленій формі разом з поверхневоактивною речовиною і пропелентом. Типовий вміст композиції в аерозольній суміші лежить в інтервалі 0,01-20%(мас), а в кращому варіанті 1-10%. Зрозуміло, що поверхнево-активна речовина повинна бути нетоксичною і, бажано, розчинною в пропеленті. Типовими матеріалами для таких агентів є естери або часткові естери жирних кислот, що містять від 6 до 22 атомів вуглецю, тобто таких, як капронова, октанова, лауринова, пальмітинова, стеаринова, лінолева, ліноленова, олестеринова та олеїнова кислоти з аліфатичним полі гідридним спиртом або його циклічним ангідридом. Можуть використовува тися змішані естери, наприклад, змішані або природні гліцериди. Поверхневоактивна речовина може становити 0,1-20%(мас), а в кращому варіанті 0,25-5% складу. Рештою композиції зазвичай є пропелент. У разі потреби до композиції може входити також носій, наприклад, лецитин для інтраназального постачання. Кон'югати за даним винаходом, в разі потреби, можуть уводитися в комбінації з іншими засобами, які є принаймні частково ефективними в лікуванні і/або поліпшенні хворого стану і/або його симптомів. У випадках хвороби Альцгеймера і синдрому Дауна, що супроводжуються амілоїдними відкладеннями в мозку, кон'югати за даним винаходом 40 можуть уводитися пацієнту в сполученні з іншими засобами, що збільшують проходження засобів за даним винаходом через гематоенцефалічний бар'єр. Імуногенна композиція згідно з винаходом, звичайно, містить ад'ювант. Ад’ювантом є речовина, що підсилює імунну відповідь при введенні її разом з імуногеном або антигеном. Було показано, що чимало цитокінів або лімфокінів володіють імуномодуляційною активністю і, таким чином, можуть використовува тися як ад’юванти; до них належать, наприклад, інтерлейкіни 1-α, 1-β, 2, 4, 5, 6, 7, 8, 10, 12 див., наприклад, [U.S. Patent No.5,723,127], 13, 14, 15, 16, 17 і 18 (та їхні мутантні форми), інтерферони α, β і γ, колонієстимулювальний фактор гранулоцитів і макрофагів, див, наприклад, [U.S Patent No.5,078,996], колонієстимулювальний фактор макрофагів, колонієстимулювальний фактор гранулоцитів (GSF), а також фактор α і β некрозу пухлин. Серед інших ад’ювантів, що можуть використовуватися в даному винаходу, слід згадати хемокіни, включаючи без обмеження, МСР-1, MIP1α, МІР-1β і RANTES. У ролі ад’ювантів можуть використовува тися також молекули адгезії, наприклад, селектин, а саме L-селектин, P-селектин і Еселектин. Серед інших корисних ад’ювантів можна назвати, без обмеження, муциноподібні молекули, наприклад, CD34, GlyCAM-1 і MadCAM-1, членів сімейства інтегринів, наприклад, LFA-1, VLA-1, Мас-1 і р150.95, членів надсімейства імуноглобулінів, наприклад, PECAM, ICAM, зокрема, ІСАМ-1, ІСАМ-2 та ІС АМ-3, CD2 і LFA-3, співстимуляторні молекули, наприклад, CD40 і CD40L, фактори росту, включаючи судинний фактор росту, фактор росту нервів, фактор росту фібробластів, епідермальний фактор росту, В7.2, PDGF, BL-1 та судинний ендотеліальний фактор росту, рецепторні молекули, включаючи Fas, TNF рецептор, Fit, Apo1, p55, WSL-1, DR3, TRAMP, Apo-3, AIR, L ARD, NGRF, DR4, DR5, KILLER, TRAIL-R2, TRICK2 і DR6. Крім того, молекулою ад’юванту може служити також каспаза (ICE). Див. також [International Patent Publication №№WO98/17799, WO99/43839], включені тут в усій їхній повноті і для всіх цілей шляхом посилання. Ад’ювантами, підходящими для підсилення імунної відповіді, є, наприклад, MPL™ (3-Одеацетильований монофосфорильний ліпід A; Corixa, Hamilton, MT), описаний у [U.S. Patent №4,912,094], включений тут для всіх цілей шляхом посилання. Підходящими для застосування як ад’юванти є також синтетичні аналоги ліпіду А або аміноалкілглюкозамінофосфатні сполуки (AGP), їх похідні та аналоги, що виробляються фірмою Corixa (Hamilton, MT) і описані в [U.S. Patent №6,113,918], включеному тут шляхом посилання. Однією з таких AGP сполук є 2-[(R)-3тетрадеканоїлокситетрадканоїламіно]етил-2деокси-4-О-фосфон-3-O-[(S)-3тетрадеканоїлокситетрадканоїл]-2[(глікопіранозид, відомий також під позначенням 529 (а також під маркою RC529; Соrіха). Ад’ювант 529 виробляють у формі водного розчину (529 AF) або стабільної емульсії (529 SE). До числа підходящих ад’ювантів входять та 41 84728 кож мінеральні масла і водні емульсії, солі кальцію, наприклад, фосфат кальцію, солі алюмінію, наприклад, гідроксид алюмінію, фосфат алюмінію і т.д., Амфіген, Авридин, L121/сквален, D-лактидполілактид/глікозид, плюронові кислоти, поліоли, дипептид мурамілу, умертвлена Bordetella, сапоніни, наприклад, Stimulon™ QS-21 (Antigenics, Framingham, MA), описані в [U.S. Patent №5,057,540], включеному тут шляхом посилання, і створені з них частки, наприклад, ISCOMS (імуностимуляційні комплекси), Mycobacterium tuberculosis, бактеріальні ліпополісахариди, синтетичні полінуклеотиди, наприклад, олігонуклеотиди, що містять мотив CpG [U.S. Patent №6,207,646], включений тут шля хом посилання, токсин коклюшу (PT), або термолабільний токсин E. coli (LT), зокрема LT-K63, LT-R72, PT-K9/G129; див., наприклад, [International Patent Publication №№WO 93/13302 і WO 92/19265], включені тут для всіх цілей шляхом посилання. Корисними для даного винаходу ад’ювантами є також токсини холери та їх мутанти, включаючи описані в публікації міжнародної заявки [International Patent Application №WO 00/18434] (де глутамінова кислота в положенні 29 амінокислотної послідовності заміщена на іншу амінокислоту (відмінну від аспартанової кислоти, у кращому варіанті на гістидин). Подібні CT токсини та їх мутанти описані в публікації міжнародної заявки [International Patent Application №WO 02/098368] (де ізолейцин у положенні 16 амінокислотної послідовності заміщений на іншу амінокислоту відокремлено або в комбінації із заміщенням серину в положенні 68 на іншу амінокислоту; і/або де валін заміщений на іншу амінокислоту в положенні 72 амінокислотної послідовності) Інші варіанти CT токсинів описані в публікації міжнародної заявки [International Patent Application number WO 02/098369 (де аргінін заміщений на іншу амінокислоту у положенні 25 амінокислотної послідовності; і/або амінокислота вставлена в положення 49 амінокислотної послідовності; і/або де дві амінокислоти вставлені в положення 35 і 36 амінокислотної послідовності). Цілком зрозуміло, що зроблені в даному описі посилання на ту чи іншу теорію для пояснення поданих результатів не мають метою обмежити об'єм винаходу. Незалежно від способу застосування даного винаходу описані тут результати і переваги можуть бути досягнуті при втіленні винаходу, в тому числі, відповідно до розглянутих нижче прикладів його здійснення. Для фахівця в даній галузі цілком зрозуміло, що поданий опис не виключає змін і модифікацій, які можуть бути внесені в нього, не виходячи за межі ідеї та об'єму, визначені доданою Формулою винаходу. Усі цитовані тут публікації, патенти і патентні заявки включені в даний опис в усій їхній повноті і для всіх цілей шляхом посилання в тій самій мірі, як у разі індивідуального позначення їх включеними в даний опис шляхом посилання. Приклад 1 Кон'югація CRM197 з пептидом Aβ Кон'югацію гаптенів та антигенних пептидів здійснювали за допомогою реакції активованого 42 носія CRM197, який має тридцять дев'ять лізинових залишків, з гаптеном або антигенним пептидом, що мають вільну бічну тіолову груп у, дотримуючись при цьому процесу, описаного нижче (Фіг.1). Усі пептиди Аβ містили цистеїновий залишок на карбоксильному кінці для полегшення їх кон'югування через цистеїніл-сульфгідрильну груп у з носійним білком. Ці пептиди були одержані шляхом твердофазного синтезу. I. Активація Вільні аміногрупи носія CRM197 піддавали бромацетилюванню шляхом їх реакції з узятим з надлишком N-гідроксисукцинімідним естером бромоцтової кислоти (Sigma Chemical Co., St. Louis, MO) [Bematowicz і Matsueda, 1986]. До охолодженого на льоду розчину CRM197 (приблизно 15мг), додали 10%(об.) 1,0м NaHCO3 (рН8,4). NГідроксисукцинімідний естер бромоцтової кислоти, взятий у такій самій масовій кількості, що і використовуваний носій CRM197 , розчинили у 200мкл диметилформаміду (DMF), повільно додали до CRM197 і обережно перемішали при кімнатній температурі в темноті протягом 1 години. Утворений бромацетильований (активований) білок промили перепусканням через знесолювальну колонку (P6DG), використовуючи як елюент фізіологічний розчин з фосфатним буфером (PBS) і 1 ммоль етилендіамінтетраоцтової кислоти (EDTA) (рН7,0). Після очистки були зібрані фракції, що відповідали активованому CRM197, і проведена оцінка концентрації білка шляхом BCA аналізу на білок. Аміногрупи білка як перед обробкою N-гідроксисукцинімідним естером бромоцтової кислоти, так і після неї, приводилися у хімічну взаємодію з 2,4,6тринітробензолсульфокислотою (TNBSA), яка служила індикатором бромацетилювання [Means et al., 1972]. II. Кон'югування Перед кон'югуванням пептиди піддавалися реакції з 5,5'-дитіо-біс(2-нітробензойною кислотою) (реагентом Ельмана (Ellman)) для перевірки вмісту вільної SH групи (знижений на 62-88%). В експерименті з першими чотирма Аβ-пептидами (амінокислоти 1-7 без лінкера, амінокислоти 1-12 з GAGAC-лінкером, амінокислоти 1-9 з GAGACлінкером і амінокислоти 1-7 з GAGAC-лінкером) приблизно 8,0 - 10,0мг пептиду розчиняли у стерильній дистильованій воді до концентрації приблизно 20мг/мл. До охолодженого активованого CRM197 повільно додавали пептид у масовому співвідношенні 1:1, і рН суміші встановлювали приблизно на 7,0-7,2 додаванням 20-36мкл 1н. розчину NaOH. Утворену суміш повільно перемішували протягом ночі при температурі 4°С в темноті, після цього проводили її діаліз в темноті при двох 1л замінах PBS, рН7,2. В експерименті з наступними чотирма Аβ-пептидами (амінокислоти 15 без лінкера, амінокислоти 1-9 без лінкера, амінокислоти 1-12 без лінкера й амінокислоти 1-5 з лінкером) для перевірки вільних SH-груп застосовувалися реакції з реагентом Ельмана (Ellman). CRM197 був підданий бромацетилюванню, очищанню і реакції з TNBSA, як описано вище. Величину рН кожного пептиду відрегульовували на рівень 7,0 додаванням 0,1моль NaPO4 (рН8,5) при 43 84728 2,2 об'єми розчиненого пептиду. Пептид повільно додавали до охолодженого активованого CRM197 у співвідношенні 1:1, і суміш залишали реагува ти протягом ночі при температурі 4°С в темноті. Утворений у результаті матеріал піддавали діалізу. Остаточний контрольний пептид (1-12-мер у зворотній орієнтації) кон'югували з CRM197, як зазначено вище, а після цього модифікували. Замість регулювання рН пептиду на рівень 7,0, рН активованого носія CRM197 устанавлювали приблизно на 7,5 додаванням 20%(об.) 0,5моль NaPO4 (рН8,0). Після діалізу кожний із кон'югатів переносили до стерильної 15мл поліпропіленової пробірки, обгортали алюмінієвою фольгою і зберігали при температурі 4°С. Після цього послідовно, за допомогою мас-спектрометрії, перевіряли активацію хімічно активних амінних залишків носія. Кон'югат Аβ1-5-C-CRM197 Аβ1-7-C-CRM197 Аβ1-9-C-CRM197 Аβ1-12-C-CRM197 Аβ1-5-L-C-CRM197 Аβ1-7-L-C-CRM197 Аβ1-9-L-C-CRM197 Імуногенний пептид DAEFR-C (SEQ. ID. NO.: 1) DAEFRHD-C (SEQ. ID NO. :2) DAEFRHDSG-C (SEQ ID N0:3) DAEFRHDSGYEV-C (SEQ ID N0:4) DAEFR-GAGA-C (SEQ ID NO.:5) DAEFRHD-GAGA-C (SEQ ID NO.:6) DAEFRHDSG-GAGA-C (SEQ ID NO.:7) DAEFRHDSGYEV-GAGA-C (SEQ ID N0.:8) VEYGSDHRFEAD- C (SEQ ID NO.: 9) Аβ1-12-L-CCRM197 Аβ12-1 -C-CRM197 (-VE Контроль) L=лінкер (GAGA) (SEQ ID NO.: 10) Приклад 2 Одержання кон'югата "пептид Аβ-CRMiS7" і очистка його надфільтрацією Бромацетилювання носія CRM197 Носій CRM197 (100мг) у 0,01M фосфатнатрієвому буфері, 0,9% NaCI, рН7,0, піддавали реакції з N-гідроксисукцинімідним естером бромоцтової кислоти (розчиненим до 20мг/мл у DMSO) у масовому співвідношення 1:1 в атмосфері аргону. Реакційну суміш в разі потреби титрували для підтримання величини рН7,0. Суміш перемішували в темноті протягом 1,5 години при кімнатній температурі. Реакційну суміш перепускали через 1,2мкм фільтр у резервуар для зберігання системи UF/DF (МіІІіроге Labscale TFF, Billerica, MA). Очистк у проводили, використовуючи 10K або 30K UF мембрану, шляхом діафільтрації (30-кратно) проти 0,01M фосфа тнатрієвого буфера/0,9% NaCI, рН7,0. Бромацетильований CRM197 фільтрували перепусканням через 0,2мкм фільтр. Був визначений ступінь бромацетилювання шляхом реакції активованого CRM197 з цистеїном, після чого проводили амінокислотний аналіз та кількісну оцінку одержаного в результаті карбоксиметилцистеїну (CMC). Кон'югування пептиду Аβ з бромацетильованим CRM197 і кепінг N-ацетилцистеїнаміном Бромацетильований CRM197 (50мг) переносили до реакційної посудини. До перемішуваного розчину, який підтримували при температурі 28°C, додавали 1M карбонат/бікарбонату натрію. Суміш титрували до досягнення цільової рН 9,0 в атмосфері аргону. Окремо наважували 50мг пептиду Аβ і розчиняли у воді для ін'єкцій (WFI) до 44 20мг/мл. До цього розчину додавали 1M карбонат/бікарбонату натрію до досягнення рН9,0. Цей пептидний розчин додавали до бромацетильованого розчину CRM197, і суміш перемішували при температурі 2-8°С протягом 14-18 годин. Решту бромацетильних груп кепували взятим з 20кратним молярним надлишком Nацетилцистеїнаміном протягом 3-6 годин при 28°C. Реакційну суміш перепускали через 1,2 мкм фільтр у резервуар для зберігання системи UF/DF (МіІІіроге XL), і кон'югат промивали при кімнатній температурі шляхом 30-кратної діафільтрації на 10К або 30K MWCO мембрані (МіІІіроrе) діафільтруванням проти 0,01M фосфа тнатрієвого буфера/0,9% NaCI, рН7,0. Утримуваний продукт збирали, перепускали через 0,2мкм фільтр та аналізували на вміст білка (колориметричний аналіз методом Лоурі (Lowry) або Micro-ВСА), піддавали SDS-PAGE аналізу, амінокислотному аналізу та аналізу на імуногенність у мишей. Приклад 3 Перетворення шляхом кепінгу непрореагованих бромацетильованих груп на аміноацетильні групи Бромацетильований CRM197 (50мг), приготований так, як описано вище, у Прикладі 2, перенесли до реакційної посудини. До перемішуваного розчину, який підтримували при температурі 28°С, додали 1M карбонат/бікарбонат натрію Суміш титрували до досягнення цільової рН9,0 в атмосфері аргону. Окремо наважили 50мг пептиду Аβ і розчинили у воді для ін'єкцій до 20мг/мл. До цього розчину додали 1M карбонат/бікарбонат натрію до встановлення рН9,0. Цей пептидний розчин додали до розчину бромацетильованого CRM197, і суміш перемішували при 2-8°С протягом 14-18 годин. Решту бромацетильних груп кепували, використовуючи 8% розчин бікарбонату амонію, протягом 4 годин при температурі 2-8°C. Реакційну суміш перепустили через 1,2 мкм фільтр у резервуар для зберігання системи UF/DF (Millipore XL), і кон'югат очистили при кімнатній температурі шляхом 30-кратної діафільтрації на 10К і 30K MWCO мембрані діафільтруванням проти 0,01M фосфатнатрієвого буфера/0,9% NaCI, рН7,0. Фільтрат зібрали, перепустили через 0,2мкм фільтр і піддали аналізу на вміст білка (колориметричний аналіз методом Лоурі (Lowry) або Micro-BCA), SDS-PAGE аналізу, амінокислотному аналізу та аналізу на імуногенність у мишей. Приклад 4 Кількісний аналіз S-карбоксиметилцистеїну і Sкарбоксиметилцистеїнаміну для оцінки ступеню кон'югації і кепінгу кон'югатів "пептидний імуногенбілок/поліпептид" Шляхом кислотного гідролізу білок-пептидних кон'югатів, створених за допомогою хімічного процесу активації бромацетилу, були одержані кислотостійкий S-карбоксиметилцистеїн (CMC) із цистеїнів у кон'югованих сайтах і кислотостійкий Sкарбоксиметилцистеїнамін (CMCA) із цистеїнаміну в кепованих сайтах (Фіг.2). Усі кон'юговані і кеповані лізини були піддані зворотному перетворенню на лізин і проаналізовані в цьому стані. Усі інші 45 84728 амінокислоти були піддані зворотному гідролізу на вільні амінокислоти, за винятком триптофану і цистеїну, які були зруйновані умовами гідролізу. Аспарагін і глутамін були перетворені, відповідно, на аспартанову кислоту і глутамінову кислоту. Зразки кон'югатів розбавили деіонізованою водою до загальної концентрації білка менше 1мг/мл. Дві 10мкг аліквоти кожного кон'югата просушили і повторно суспендували в 100мкл 6н. розчину HCI [Pierce], 5мкл розплавленого фенолу [Sigma-Aldrich] і 1мкл 2-меркаптоетанолу [SigmaAldrich]. Після цього зразки витримували в термостаті під вакуумом (100мТ) при 110°C протягом 22 годин. Отримані в результаті гідролізати просушили, повторно суспендували у 250мкл Na-S цитратнатрієвого буфера Бекмана (Beckman) для розбавляння зразків (рН2,2) [Beckman Instruments, lnc , Fullerton, CA] і профільтрували через 0,2мкм нейлонові фільтри Ватмана (Whatman) кінцівок шприців і 1мл шприци. Після цього кожний зі зразків завантажували в петлю зразка амінокислотного аналізатора Бекмана Beckman 6300 і поміщали в аналізатор. Амінокислоти кожного гідролізованого зразка і контрольного зразка розділяли за допомогою іонообмінної хроматографії і наступної реакції з розчином нінгідрину NinRX Бекмана (Beckman) при температурі 135°С. Далі дериватизовані амінокислоти виявляли у видимому діапазоні спектра на хвиля х 570нм і 440нм (Таблиця 1). У кожній групі аналізу разом з експериментальними і контрольними зразками через аналізатор перепускали стандартний комплект амінокислот [Pierce Amino Acid Standard H] що містив 500пікомоль кожної амінокислоти. До стандарту додавали Sкарбоксиметилцистеїн (Sigma-Aldrich). Таблиця 1 Час в итримув ання амінокислот за Градієнтною програмою 1 в амінокислотному аналізаторі Бекмана "Beckman 6300" Час в иДовжина трихв илі при Амінокислоти мування, де текхв . тув анні 8,3 Карбоксиметилцистеїн CMC 570 9,6 Аспартанова кислота й ас- Asx 570 парагін 11,3 Треонін Thr 570 12,2 Серин Ser 570 15,8 Глутамінова кислота і глу- GIx 570 і 440 тамін 18,5 Пролін Pro 440 21,8 Гліцин GIy 570 23,3 Аланін Ala 570 29,0 Валін VaI 570 32,8 Метіонін Met 570 35,5 Ізолейцин He 570 36,8 Лейцин Leu 570 40,5 Тирозин Tyr 570 42,3 Фенілаланін Phe 570 45.4 Карбоксиметилцистеїнамін CMCA 570 48,8 Гістидин His 570 53,6 Лізин Lys 570 70,8 Аргінін Arg 570 46 Площа кожного стандартного піка приймалася за кількісний еквівалент для пропорційної оцінки кожного зразка. Пролін був визначений із довжини хвилі 440нм і перетворений на еквівалентні йому 570нм, використовуючи глутамінову кислоту, тобто найближчу амінокислоту. Кожне з цих значень в пікомолях було перетворене на молярне співвідношення амінокислотних залишків шляхом порівняння значень у пікомолях лізину з теоретичною величиною для лізину, наявного в даному білку. Лізин був вибраний для цієї оцінки з погляду його ковалентного зв'язування з цистеїном і цистешаміном, а також очікуваного подібного гідролізу. Отримані кількості молей амінокислот були приведені у порівняння з амінокислотним складом білка і подані разом з величинами для CMC і CMCA. Величина для CMC використовувалася безпосередньо для оцінки ступеню кон'югації, а величина для CMCA використовувалася безпосередньо для оцінки ступеню кепінгу. Приклад 5 Характеризація й оптимізація пептидних кон'югатів Ag-CRM197 Для контролю кон'югації всі кон'югати пептидСРМ197 піддавалися амінокислотному аналізу і мас-спектрометричному аналізу методом MALDITOF (matrix-assisted laser desorption ionization-time of flight) матричної мас-спектрометрії з лазерною десорбцією і часо-пролітною іонізацією Для кожного кон'югата кількість молей пептиду, кон'югованих з кожним молем CRM197, визначали за допомогою амінокислотного аналізу (за кількістю Sкарбоксиметилцистеїнових залишків) і масспектрометрії методом MALDI-TOF. Величини, отримані тим та іншим методом, в цілому збігалися. І. Гель-хроматографія Зразки партій концентрату видобували із резервуара для зберігання і залишали відігріватися до кімнатної температури. Зразок Аβ-пептидного кон'югата повільно перемішували до одержання гомогенного препарату. Зразок Аβ-пептидного кон'югата піддавали центрифугуванню в мікроцентрифузі Епендорфа (Eppendorf) для видалення з нього всіх часток. Супернатант видобували і піддавали гель-хроматографії на колонці TosoHaas TSK-GeI G3000SW (TosoHaas, Stuttgart, Germany). Колонку TosoHaas TSK-GeI G3000SW сполучали з HPLC-хроматографом (рідинної хроматографії високої розрізнювальної спроможності), і встановлювали межу тиску на рівень 1,4МПа. Колонку зрівноважували щонайменше 30мл PBS буфера (10ммоль фосфату натрію, 150мМ Nad, рН7,2±0,1) при витраті 0,75мл/хв Зразок Аβ-пептидного кон'югата завантажували в колонку TosoHaas TSK-GeI G3000SW при таких параметрах: Концентрація зразка Аβ-пептидного кон'югата sample: 1,5±1,0мг/мл Витрата: 0,75мл/хв Об'єм зразка: 0,1мл Час утримування: 30 хвилин Моніторинг поглинання проводили як на 280нм, так і на 210нм. У випадках тривалого збері 47 84728 гання колонку TosoHaas TSK-GeI G3000SW зрівноважували щонайменше 50мл 20% етанолу при витраті 0,5-1,0мл/хв. II. PAGE (електрофорез у поліакриламідному гелі) Активований (бромацетильований) CRM197 і Aβ-пептид-CRM197 кон'югати досліджувалися в гелях з додецилсульфатом натрію (SDS) методом NuPAGE Bis-Tris електрофорезу (Novex, Frankfurt, Germany) при нейтральній рН у системі передформованого поліакриламідного мінігеля і NuPAGE MES SDS Running Buffer. Аліквоту 8мкг кожного активованого CRM або кон'югата змішували з відновним буфером для зразка і гріли при 100°C протягом 5 хвилин. Кон'югати і стандарти молекулярної маси (MW) (Invitrogen, Carlsbad, CA) поміщали на 10% (мас/об., акриламід) NuPage гель (Novex) на основі буферизованої Bis-Tris-HCI системи і піддавали електрофорезу з розділяльним буфером MES SDS Running Buffer-PAGE (Laemmli). Після SDS-PAGE електрофорезу гель забарвлювали блакитним Pierce Gel Code Blue (Pierce, Rockford, IL). Кон'югат Aβ-пептид-CRM197 був представлений головною смугою навколо 66кДа, вище смуги природного CRM і смуги димеру навколо 120кДа, разом з несуттєвими смужками мультимерів (не показані). III. Mac-спектрометричний аналіз пептидCRM197 кон'югатів методом MALDI-TOF Для негайної апроксимації ступеню кон'югування використовували мас-спектрометрію. Відповідні аліквоти зразків активованогоCRM197 і кон'югата піддавали мас-спектрометричному аналізу методом MALDI-TOF, використовуючи як матрицю 3,5-диметокси-4-гідроксикоричну кислоту (синапінову кислоту). Молекулярна маса активованого CRM197, визначена за допомогою MALDI-TOF масспектрометрії на приладі (Finnigan MAT Lasermat 2000 Mass Spectrometer, Ringoes, NY), була розподілена навколо 60,5кДа, а у кон'югатів вона лежала в інтервалі від 65кДа до 74кДа залежно від ступеню кон'югування (результати аналізу не подані). До 22 лізинів (~50%) у CRM197 виявилися модифікованими у співвідношенні 1:1. IV. Експерименти з оптимізації Ступені активації і кон'югації є функцією співвідношення реагент: білок, температури реакції і величини рН реакційного буфера. Нижче наведено декілька прикладів, що ілюструють оптимальні умови кон'югації, здійснених для визначення оптимального рН з метою отримання відтворюваних параметрів реакцій кон'югації. Одержані результати (Фіг.3) показали, що реакція кон'югації з Аβ 5 48 мером (DAEFRC) (SEQ ID NO: 1), а також із Аβ 7мером (DAEFRHDC) (SEQ ID NО:2) залежить від рН і дає більший ступінь модифікації та кон'югації, коли рН реакції підвищується Використовуючи сіль TFA 5-мерних і 7-мерних пептидів, був оцінений ступінь кон'югаці при рН9,0 і різних кількостях завантаженого пептиду (Фіг.4). Із цих результатів з очевидністю випливає, що пептидні кон'югати з визначеною кількістю пептидних копій на молекулу CRM можуть створюватися шляхом варіювання співвідношення пептид:активований CRM під час процесу кон'югації. Аналогічні експерименти були проведені при використанні ацетату 7-мерного пептиду Аβ. Стосовно кон'югації Aβ1-7/CRM кепінг-процес був оцінений шляхом порівняння кількості молей CMCA на моль CRM з кількістю молей CMC на моль CRM. Оскільки загальна кількість CMC і CMCA була постійною при кожному співвідношенні пептид:CRM у даному тесті, вважалося, що кепінгпроцес відбувався повністю (Фіг.5). Загальна модифікація в кон'югаті лежала в межах від 19 до 21, що були порівняними з кількістю бромацетильованих лізинів (Фіг.5). Ці експерименти проводилися з TFA як з протиіоном для даного пептиду. Кон'югацію Aβ1-7/CRM повторювали, використовуючи замість солі TFA ацетат пептиду; результати цього експерименту показані на Фіг.5 і 6. Кепінг-процес демонстрував своє повне завершення і давав загальні кількості CMC і CMCA по кожній точці в інтервалі від 20 до 22 Умови реакції Aβ-CRM кон'югації були оптимізовані при рН9,0, а ступінь кон'югації регулювали співвідношенням між пептидом і CRM у реакційному середовищі. Шляхом варіювання цього співвідношення в межах від 0,1 до 1,5 можна було змінювати ступінь кон'югаці (Фіг.6). Ступені активації та кон'югації є функцією співвідношення реагент:білок, температури реакції і рН реакційного буфера. Ступінь модифікації (кон'югації) для кожного кон'югата обчислювали відніманням значення маси активованого CRM197 із значень маси кожного кон'югата і діленням отриманої різниці на масу пептиду, використовуваного в одержанні кон'югата. Значення ступеня модифікації (кон'югації) для всіх кон'югатів подані в Таблиці 2. Було проведено також порівняння ступеня кон'югації з величинами, отриманими при оцінці кількості утворених S-карбоксиметилцистеїнових залишків на моль CRM197 (також наведені в Таблиці 2). Таблиця 2 Ступінь модифікації: порів няння даних мас-спектрометрії методом MALDI-TOF і даних амінокислотного аналізу Зразок CRM197 BrAc-CRM Aβ1-7/CRM Aβ1-7/CRM Aβ1-5/CRM Aβ1-5/CRM Дальтон и (за даними масспектрометрії) 58408 60752 74463 72375 75425 71690 Ступінь кон'югації (за даними мас-спектрометри*) 19 14 12 20 15 Ступінь кон'югації (за даними CMCамінокислотного аналізу) 15 14 21 18 49 84728 Приклад 6 Дослідження імуногенності Аβ-пептидних кон'югатів Пептиди, замкнені між N-кінцевими залишками 1-5, 1-7, 1-9 і 1-12 пептиду Аβ (з лінкерною послідовністю GAGAC і без неї) і пептид, що відповідав N-кінцю пептиду Aβ у зворотній послідовності від дванадцятої амінокислоти до першої амінокислоти (1-12-мер у зворотній послідовності), кожний з яких був кон'югований з CRM197, використовувалися для імунізації мишей разом з некон'югованим Аβ 112-мерним пептидом у препараті зі Стимулоном (STIMULON™ QS-21). Мишей усі х гр уп імунізували підшкірними ін'єкціями з дозами 30мкг або 5мкг зразків, складених з 20мкг ад'юванту STIMULON™ QS-21, на початку досліджень (тиждень 0) і після цього - на 3-й і 6-й тижні. Протокол досліджень поданий у Таблиці 3. Як показано в Таблиці 3, пептиди, замкнені між N-кінцевими залишками 1-5, 1-7, 1-9 і 1-12 пептиду Аβ (з лінкерною послідовністю GAGAC і без неї), і пептид, що відповідав N-кінцю пептиду Аβ у зворотній послідовності від дванадцятої амінокислоти до першої амінокислоти (1-12-мер у зворотній послідовності), кон'юговані з CRM197 , використовувалися для імунізації мишей разом з некон'югованим Аβ 1-12-мерним пептидом у препараті з QS-21. Мишей усі х груп імунізували підшкірними ін'єкціями з дозами 30мкг або 5мкг зразків, складених з 20мкг ад'юванту QS-21, на початку досліджень (тиждень 0) і після цього - на 3-й і 6-й тижні. В дослідженнях використовувалися миші породи Швейцарського Вебстера (Swiss Webster), пo 5 тварин у кожній групі. Об'єм ін'єкції становив 100мкл; В= кровопускання; V= вакцинація; E= знекровлення. Титри антитіл проти Аβ виміряли шляхом 50 твердофазного імуноферментного аналізу (ELISA) проти Aβ і CRM197, як описано нижче. Коротко, 96лункові планшети Costar (№3591) протягом ночі при кімнатній температурі покривали 2мкг/мл Аβ142 у стерильному карбонат/бікарбонатному буфері, рН9,6. Планшети спорожнювали і блокували протягом двох годин при кімнатній температурі 200мкл/лунку 0,05% розчину альбуміну бичачої сироватки (BSA) в 1´ PBS/0,05% Tween 20. Блоковані планшети спорожнювали і промивали засобом для мийки планшетів, що містив TBS, 0,1% промивного буфера Brij-35 (без азиду). Усі первинні антисироватки послідовно промивали 0,05% розчином BSA в 1´ PBS (забуференому фосфатом фізіологічному розчині), що містив 0,05% Tween 20/0,02% азиду. Після цього по 100мкл кожного розчину переносили у відповідні лунки й інкубували при кімнатній температурі протягом 2 годин. Далі планшети спорожнювали і промивали, як описано вище. Кон'юговане з лужною фосфатазою вторинне антитіло IgG кози проти миші від Southern Biotech розбавляли у співвідношенні 1:1000 0,05% розчином BSA у PBS, що містив 0,05% Tween 20/0,02% азид, додавали по 100мкл до кожної лунки та інкубували при кімнатній температурі протягом 1 години. Після цього планшети спорожняли, промивали, як описано вище, і наприкінець інкубували при кімнатній температурі протягом 1 години зі 100мкл/лунку 1мг/мл розчину р-нітрофенілфосфатного субстрату, приготованого в суміші діетаноламіну з MgCI2, рН9,8. Еволюцію кольору припиняли додаванням 50мкл/лунку 3н. розчину NaOH. Планшети зчитували на хвилі 405нм з 690нм еталоном. Кінцеві титри обчислювали при оптичній густині О.D. 0,1 довільних одиниць. Таблиця 3 Протокол досліджень з імунізації мишей Код групи АЕ488 АЕ489 АЕ490 АЕ491 АЕ492 АЕ493 АЕ494 АЕ495 АЕ496 АЕ497 АЕ498 АЕ499 АЕ500 АЕ501 АЕ502 AE503 AE504 AE505 AE506 AE507 AE508 AE509 AE510 AE511 Опис CRM/1-7 без лінкера CRM/1-12 з лінкером CRM/1-9 з лінкером CRM/1-7 з лінкером CRM/1-5 без лінкера CRM/1-9 без лінкера CRM/1-12 без лінкера CRM/1-5 з лінкером CRM/1-7 без лінкера CRM/1-12 з лінкером CRM/1-9 з лінкером CRM/1-7 з лінкером CRM/1-5 без лінкера CRM/1-9 без лінкера CRM/1-12 без лінкера CRM/1-5 3 линкером CRM197C1-6151 CRM197C1-6151 CRM/12-1-мер CRM/12-1-мер 1-12-мерний пептид 1-12-мерний пептид Ab Ab Доза,мкг 30 30 30 30 30 30 30 30 5 5 5 5 5 5 5 5 30 5 30 5 30 5 30 5 Тижд. 0 B,V B,V B,V B,V B,V B,V B,V B,V B,V B,V B,V B,V B,V B,V B,V B,V B,V B,V B,V B,V B,V B,V B,V B,V Тижд. 3 B,V B,V B,V B,V B,V B,V B,V B,V B,V B,V B,V B,V B,V B,V B,V B,V B,V B,V B,V B,V B,V B,V B,V B,V Тижд. 6 B,V B,V B,V B,V B,V B,V B,V B,V B,V B,V B,V B,V B,V B,V B,V B,V B,V B,V B,V B,V B,V B,V B,V B,V Тижд. 8 В В В В В В В В В В В В В В В В В В В В В В В В Тижд. 13 В В В В В В В В В В В В В В В В В В В В В В В В Тижд. 16 E E E E E E E E E E E E E E E E E E E E E E E E 51 84728 Твердофазний імуноферментний аналіз CRM197 Планшети Грейнера (Greiner) на 96 лунок (№650011) покривали при температурі 37°C протягом 90 хвилин з 5,0мкг/мл (100мкл/лунку) білка CRM197 у стерильному карбонат/бікарбонатному буфері, рН9,6. Планшети спорожнювали і промивали засобом для мийки планшетів, що містив 1´ TBS1 0,1% промивного буфера Brij-35. Усі первинні антисироватки послідовно промивали 1´ PBS1 що містив 0,3% Tween 20/EDTA. Після цього по 100мкл кожного розчину переносили у відповідні лунки й інкубували при 37°С протягом 1 години. Далі планшети спорожнювали і промивали, як описано вище. Кон'юговане з лужною фосфатазою вторинне антитіло IgG кози проти миші від Southern Biotech розбавляли у співвідношенні 1:1000 розчином 1´ PBS, що містив 0,05% Tween 20/0,02% азид, і по 100мкл додавали до кожної лунки та інкубували при 37°С протягом 1 години. Після цього планшети спорожнювали, промивали, як описано вище, і наприкінець інкубували при кімнатній температурі протягом 1 години зі 100мкл/лунку 1мг/мл розчину рнітрофенілфосфатного субстрату, приготованого в суміші діетаноламіну з MgCI2, рН9,8. Еволюцію кольору припиняли додаванням 50мкл/лунку 3 н. розчину NaOH. Планшети зчитували на хвилі 405нм з 690нм еталоном. Кінцеві титри обчислювали при оптичній густині O.D. 0,1 довільних одиниць У Таблицях 4-6 наведені визначені методом ELISA кінцеві титри проти Аβ. Після первинної імунізації всі вісім кон'югатів (за винятком негативного контролю) викликали відчутні IgG імунні відповіді проти Аβ. Проте, 30мкг доза, а не 5мкг доза, пептиду Аβ давала позитивну відповідь на 3-й тиждень після первинної імунізації. Серед усіх кон'югатів пептид Аβ 1-7, кон'югований без лінкера, демонстрував здатність викликати таку ж хорошу або навіть кращу відповідь, ніж інші досліджені кон'югати. При 5мкг дозі пептид Аβ 1-5С діяв краще в період 8-16 тижнів. Пептид Аβ 1-7С давав найкращі результати при дозі 30мкг. Аналіз титрів антитіл після другої та третьої імунізації як з дозою 5мкг, так і з дозою 30мкг показав, що максимальна імунна відповідь на Аβ для більшості кон'югатів спостерігалася після другої імунізації. Третя імунізація, принаймні у мишей, не супроводжувалася підсиленням імунної відповіді. Проте, пептид Аβ для досягнення максимальної імунної відповіді проти нього потребував проводити три імунізації з дозою 30мкг (Таблиця 5). Що стосується послаблення антитіл з плином часу, то рівень антитіл із групи тварин, імунізованих кон'югатами, знижувався у 2-3 рази порівняно з максимальним рівнем у цій групі. Окремі зразки 6-го і 8-го тижнів були проаналізовані для обчислення середньогеометричних (GMT) титрів проти Аβ по кожній із груп (Таблиця 6) і, таким чином, встановлення того, чи є серед груп, що отримували кон'югати, яка-небудь група, набагато краще інших. Статистичний аналіз титрів 6-го тижня від кон'югатів Аβ1 52 5С, Аβ1-7С і Аβ1-9С показав, що кон'югат Аβ 1-7 продемонстрував значно вищий титр. Із цього експерименту є очевидним також, що лінкерна послідовність GAGAC не зробила внеску в підсилення імунної відповіді на даний пептид. Таблиця 4 Група 1-5С
ДивитисяДодаткова інформація
Назва патенту англійськоюMethod of producing immunogenic conjugate, a-beta immunogenic peptide carrier conjugate and immunogenic composition containing thereof
Автори англійськоюArumugham Rasappa G, Prasad A Krishna
Назва патенту російськоюПроцесс образования иммуногенного конъюгата, иммуногенный конъюгат белка ab
Автори російськоюАрумугхам Расаппа Дж., Прасад A. Кришна
МПК / Мітки
МПК: C07K 1/06, A61K 39/39, C07K 1/04, A61P 25/28, A61K 38/17
Мітки: кон'югат, утворення, процес, кон'югату, білка, імуногенного, імуногенний
Код посилання
<a href="https://ua.patents.su/56-84728-proces-utvorennya-imunogennogo-konyugatu-imunogennijj-konyugat-bilka-ab.html" target="_blank" rel="follow" title="База патентів України">Процес утворення імуногенного кон’югату, імуногенний кон’югат білка ab</a>
Пептид, здатний нейтралізувати або модулювати утворення антитіл до мвр (мієлінового основного білка), фармацевтична композиція та спосіб лікування розсіяного склерозу

Номер патенту: 55375
Опубліковано: 15.04.2003
Автори: Уоррен Кеннет, Кац Інгрід
МПК: C07K 14/47
Мітки: склерозу, модулювати, фармацевтична, основного, нейтралізувати, здатний, антитіл, мвр, лікування, білка, композиція, розсіяного, утворення, спосіб, пептид, мієлінового
Формула / Реферат:
1. Пептид или его соли, способный нейтрализовать или модулировать образование антител к миелиновому основному белку, имеющий формулу R1-Val-His-Phe-Phe-Lys-Asn-Ile-R2,где R1 выбрано из группы, включающейAsn-Pro-Val,Рrо-Val,Val,Lys-Ser-His-Gly-Arg-Thr-Gln-Asp-Glu-Asn-Pro-Val,Thr-Gln-Asp-Glu-Asn-Pro-Val,водород игидрокси,R2 выбрано из...
Спосіб одержання полімерзв’язаних циклічних пептидів

Номер патенту: 72460
Опубліковано: 15.03.2005
Автори: Следескі Адам В., Менсел Джеймс Дж.
МПК: C07K 17/00, C07K 5/09, C07K 7/06, C07K 7/04, C07K 5/113, C07K 14/635, C07K 7/08, C07K 1/04, C07K 5/062, C07K 7/54
Мітки: спосіб, циклічних, пептидів, полімерзв'язаних, одержання
Формула / Реферат:
1. Спосіб одержання, ІІв якій J, L і М представляють лінійні пептидні фрагменти;К1 відсутній або представляє циклічний пептидний фрагментіК2 представляє циклічний пептидний фрагмент,який включає:(1) одержання послідовним приєднанням придатним чином заміщених амінокислотних залишків до полімеру з одержанням:, ІІІде представляє придатний синтетичний полімер для пептиду, і М...
Спосіб створення ліній та гібридів кукурудзи з підвищеним вмістом білка та олії і поліпшеною якістю білка

Номер патенту: 21548
Опубліковано: 15.03.2007
Автори: Ніколенко Ірина Анатоліївна, Тимчук Віктор Михайлович, Тимчук Сергій Михайлович
МПК: A01H 1/04
Мітки: якістю, створення, вмістом, олії, підвищеним, поліпшеною, гібридів, ліній, спосіб, білка, кукурудзи
Формула / Реферат:
Спосіб створення ліній та гібридів кукурудзи з підвищеним вмістом білка та олії і поліпшеною якістю білка, що включає використання неалельних взаємодій між генами структури ендосперму, який відрізняється тим, що для підвищення вмісту білка та олії в зерні і незамінних амінокислот в білку використовують біохімічний ефект генної рекомбінації o2sh2.
Імуногенний препарат, що викликає імунну відповідь проти парвовірусу та цирковірусу свиней, вакцина проти багатосистемного синдрому виснаження свиней (бсвс) та набір для вакцинації

Номер патенту: 72891
Опубліковано: 16.05.2005
Автори: Міхен Брайан Мартін, Елліс Джон Алберт, Аллан Гордон Мур, Одонне Жан-Крістоф Франсіс, Мак-Нейлі Френсіс, Краковка Джордж Стівен
МПК: A61P 31/12, A61K 39/295, C07K 14/01, C12N 7/00, A61P 31/22, A61P 37/04, A61P 43/00
Мітки: цирковірусу, викликає, відповідь, імунну, препарат, набір, багатосистемного, вакцинації, свиней, парвовірусу, вакцина, синдрому, виснаження, імуногенний, бсвс
Формула / Реферат:
1. Імуногенний препарат, що викликає імунну відповідь проти парвовірусу та цирковірусу свиней, що включає щонайменше один антиген цирковірусу свиней та щонайменше один антиген парвовірусу свиней у розчиннику або наповнювачі, прийнятному з ветеринарної точки зору.2. Імуногенний препарат за п.1, що включає антиген цирковірусу свиней типу II.3. Імуногенний препарат за п.2, у якому антиген цирковірусу свиней типу II вибраний з...
Кристал білка, зшитого багатофункціональним зшивальним агентом (варіанти), пристрій, що містить кристал білка та спосіб отримання аспартаму

Номер патенту: 27035
Опубліковано: 28.02.2000
Автори: Ст.Клер Ненсі Л., Нейвіа Мен'юел А.
МПК: A61K 38/46, C07K 5/072, C12N 11/00, C12Q 1/00, C12N 9/66, C12N 9/78, C12M 1/40, G01N 33/66, C12P 21/06, C12N 9/14, G01N 33/531, C12N 9/36, C12N 9/52, C12Q 1/37, C12M 1/34, C12P 21/02, C12N 9/18, A61K 38/43
Мітки: багатофункціональним, містить, кристал, агентом, зшитого, пристрій, варіанти, аспартаму, білка, отримання, зшивальним, спосіб
Формула / Реферат:
1. Кристалл белка, сшитого многофункциональным сшивающим агентом, отличающийся тем, что он обладает устойчивостью к экзогенному протеолизу, такой, что указанный белковый кристалл сохраняет, по меньшей мере, 91% его исходной активности после инкубирования в течение, по меньшей мере, трех часов в присутствии проназы (Pronase™) в концентрации, вызывающей у растворимой формы белка, кристаллизующейся с образованием этого белкового кристалла,...
Попередній патент: Шлунково-кишковий пристрій
Наступний патент: Різальна пластина різця для обробки циліндричних поверхонь
Випадковий патент: Верхня кришка для пральної машини