Спосіб підвищення біосинтезу антибіотиків моеноміцинового ряду
Номер патенту: 71729
Опубліковано: 25.07.2012
Автори: Федоренко Віктор Олександрович, Осташ Богдан Омелянович, Рабик Марія Василівна





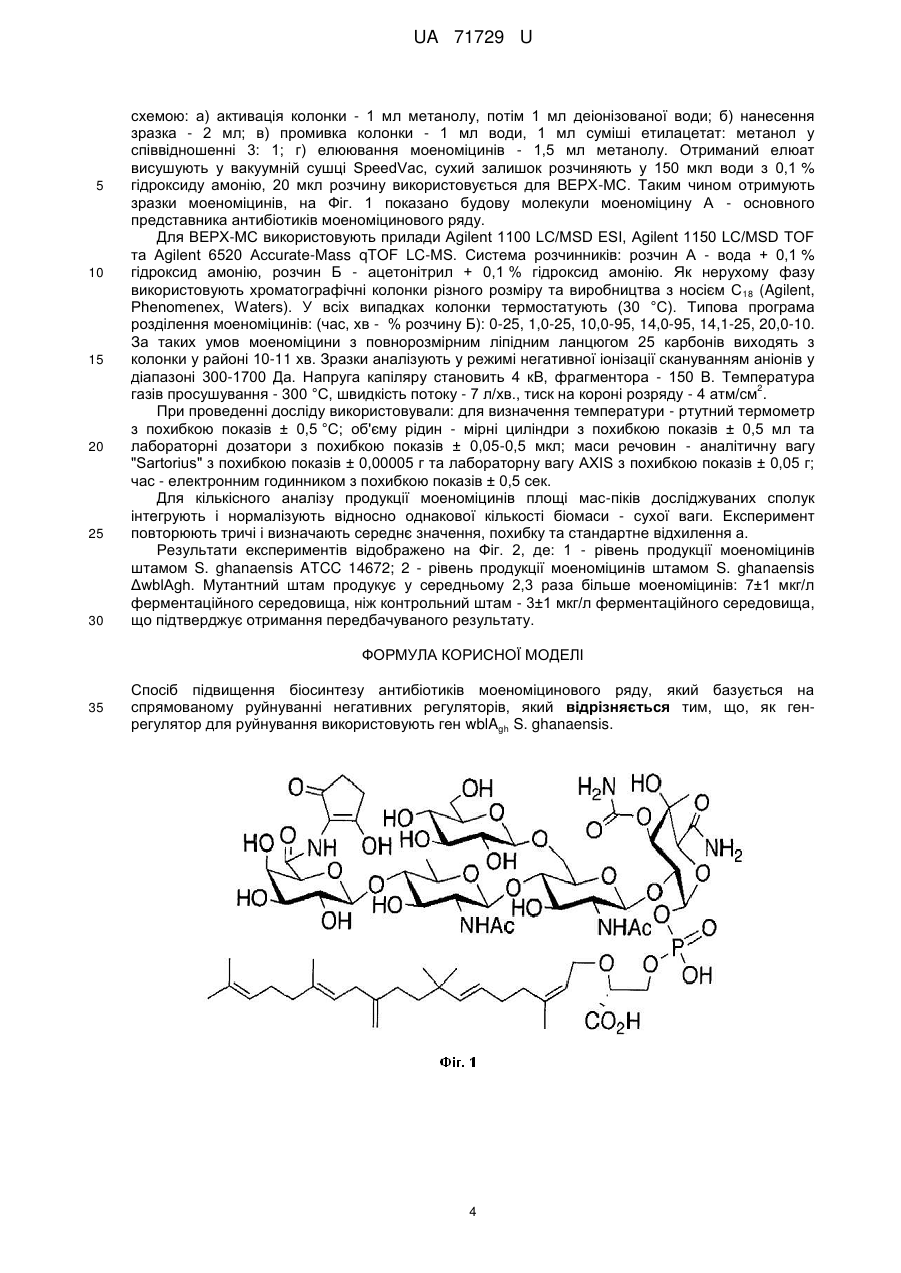
Формула / Реферат
Спосіб підвищення біосинтезу антибіотиків моеноміцинового ряду, який базується на спрямованому руйнуванні негативних регуляторів, який відрізняється тим, що, як ген-регулятор для руйнування використовують ген wblAgh S. ghanaensis.
Текст
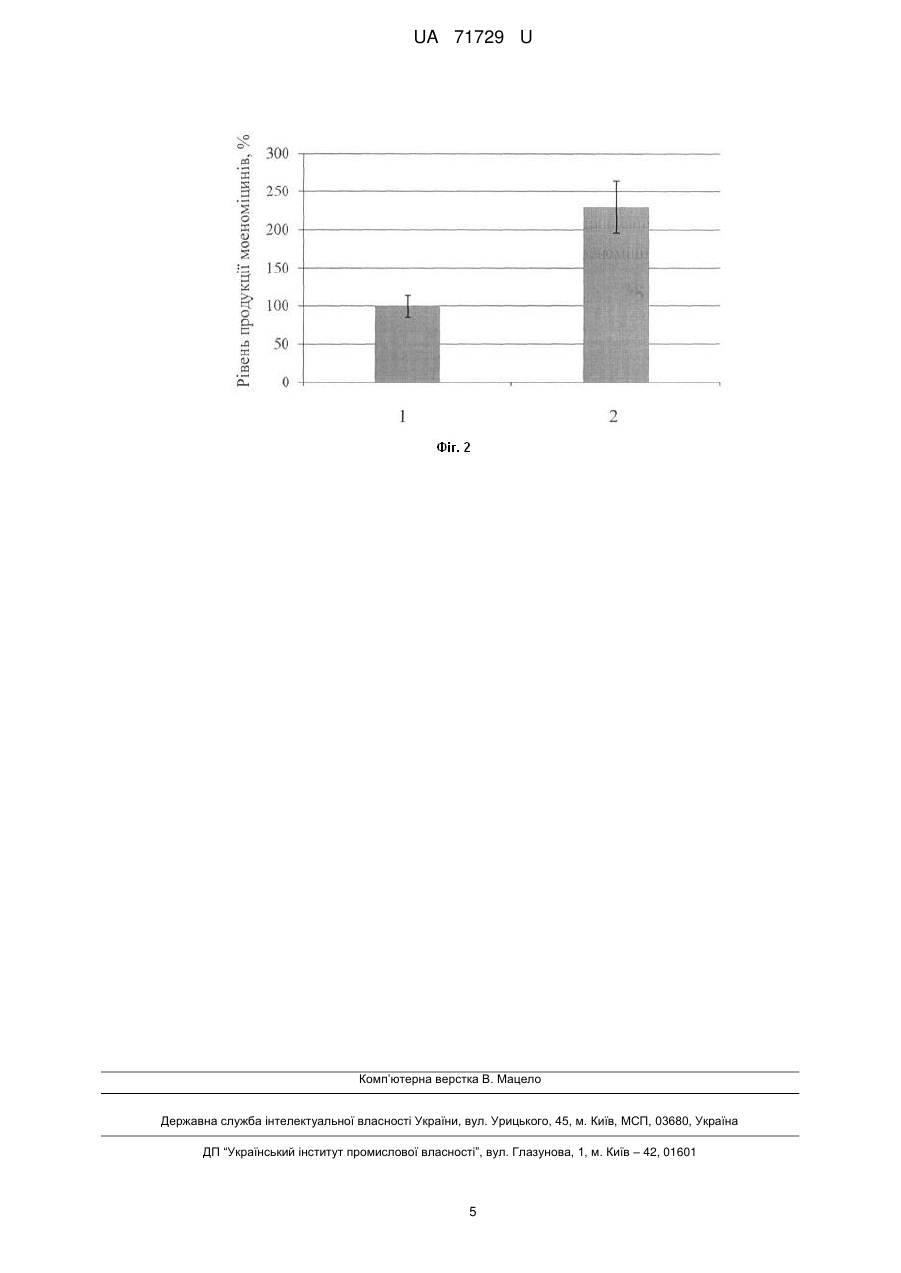
Реферат: Спосіб підвищення біосинтезу антибіотиків моеноміцинового ряду базується на спрямованому руйнуванні негативних регуляторів. Крім того, як ген-регулятор для руйнування використовують ген wblAgh S. ghanaensis. UA 71729 U (54) СПОСІБ ПІДВИЩЕННЯ БІОСИНТЕЗУ АНТИБІОТИКІВ МОЕНОМІЦИНОВОГО РЯДУ UA 71729 U UA 71729 U 5 10 15 20 25 30 35 40 45 50 55 Корисна модель належить до генетики бактерій та біотехнології і може бути використана для створення штамів актиноміцетів-надпродуцентів антибіотиків моеноміцинового ряду та їхніх похідних. Відомий спосіб отримання надпродуцентів моеноміцинів, згідно з яким спорову суспензію Streptomyces ghanaensis - продуцента моеноміцинів - опромінюють ультрафіолетом чи обробляють хімічним мутагеном, розсівають її на поживному агаризованому середовищі та відбирають клони із підвищеним рівнем біосинтезу антибіотика [Subramaniam-Niehaus В., Schneider Т., Metzger J.W., Wohlleben W. Isolation and analysis of moenomycin and its intermediates from Streptomyces ghanaensis (ATCC14672) wildtype and selected mutants // Z. Naturforsch. - 1997. - vol.52. - P. 217-226; Schuricht U., Hennig L., Findeisen M., Endler K., Welzel P., Arigoni D. The biosynthesis of moenocinol, the lipid part of the moenomycin antibiotics // Tetrahedron Lett. - 2001. - vol. 42. - P. 3835-3837]. Отримані мутанти синтезують у 2-2,5 раза більше моеноміцинів порівняно з вихідним штамом. Проте зазначений спосіб є досить тривалим, включає застосування складних мікробіологічних процедур. Мутації, що накопичуються у штамах, є унікальними для кожного з них і тому їх неможливо відтворити в іншому штамі, що продукує структурно відмінний моеноміцин. Найближчим за технічною суттю прототипом, є спосіб підвищення рівня синтезу доксорубіцину та даунорубіцину штамом S. pencetius 01М за допомогою руйнування гена wblAspe, який кодує плейотропний регулятор біосинтезу антибіотиків та морфологічної диференціації [Noh J.-H., Kim S.-Н., Lee H.-N., Lee S.Y., Kim E.-S. Isolation and genetic manipulation of the antibiotic down-regulatory gene, wblA ortholog for doxorubicin-producing Streptomyces strain improvement // Appl. Microbiol. Biotechnol. - 2010. - vol. 86. - P. 1145-1153]. Косміду, яка містить зруйнований ген wblAspe введено у штам S. peucetius OIM за допомогою кон'югації з Escherichia coli ET12567 (pUZ8002). Після інкубування при 30 °C протягом 16 год., кожну чашку Петрі з кон'югаційною сумішшю заливали 1 мл стерильної води, яка містить 50 мкг/мл апраміцину та 25 мкг/мл налідиксової кислоти. Інкубацію продовжували до появи екскон'югантів. Мутанти, в яких відбулося заміщення гена wblAspe на ген стійкості до канаміцину, відбирали за ознаками стійкості до апраміцину та чутливості до канаміцину. Нокаут гена підтверджували за допомогою ПЛР. Транскон'юганти вирощували протягом двох днів у рідкому середовищі TSB, потім протягом 6 днів у середовищі NDYE. Доксорубіцин та даунорубіцин екстрагували сумішшю ізопропілового спирту та 30 % НСl (50:1), центрифугували 10 хв при 15 000 об./хв., отриманий супернатант аналізували за допомогою високоефективної рідинної хроматографії. Транскон'юганти продукували у 1,5 раза більше даунорубіцину та у 0,4 раза більше доксорубіцину, ніж контрольний штам. Проте, штам S. peucetius не продукує моеноміцинів і, тому цей штам і клонований ген wblAspe не можна використати для підвищення продукції останніх. В основу корисної моделі поставлено задачу удосконалити спосіб підвищення біосинтезу антибіотиків моеноміцинового ряду шляхом руйнування гену негативного регулятора wblAgh, у актиноміцетній культурі S. ghanaensis, що дасть змогу підвищити вихід продукції антибіотика. Поставлена задача вирішується тим, що у способі підвищення біосинтезу антибіотиків моеноміцинового ряду, який базується на спрямованому руйнуванні негативних регуляторів, при чому, як ген-регулятор для руйнування використовують ген wblAgh S. ghanaensis. Моеноміцини - це фосфогліколіпідні антибіотики, активні проти широкого спектра грампозитивних бактерій. Ці сполуки виявляють інгібуючу дію за наномолярних концетрацій, є значно активніші, ніж, зокрема, ванкоміцин [Ostash В., Saghatelian A., Walker S. A streamlined metabolic pathway for the biosynthesis of moenomycin A // Chem. Biol. - 2007. - vol. 14. - P. 257267]. Відомо, що моеноміцини - єдині відомі сполуки, які інгібують ріст бактерій шляхом безпосереднього зв'язування з пептидоглікан-глікозилтрансферазами - ферментами, які формують гліканові ланцюги бактерійної клітинної стінки із дисахаридних попередників [Ostash В., Walker S. Moenomycin family antibiotics: chemical synthesis, biosynthesis, biological activity // Nat. Prod. Rep. - 2010. - vol. 27. - P. 1594-1617]. Проте, існуючі штами-продуценти моеноміцинів синтезують малі кількості антибіотика, що є перешкодою для дослідження антибіотика та його похідних. Авторами вперше запропоновано використати нокаут гена wblAgh для підвищення виходу продукції моеноміцинових антибіотиків. Ген wblAgh кодує плейотропний регулятор синтезу антибіотиків та задіяний у морфогенезі. 1 UA 71729 U 5 10 15 20 25 30 35 40 45 50 55 60 Оскільки, делеції генів негативних регуляторів дають змогу швидко отримувати штами, які продукують збільшену кількість антибіотиків, то це призведе до підвищення виходу продукції антибіотиків моеноміцинового ряду. Фіг. 1 Хімічна будова моеноміцину А, основного представника антибіотиків моеноміцинового ряду. Фіг. 2 Результати аналізу продукції моеноміцинів штамами за допомогою високоефективної рідинної хроматографії, спряженої з мас-спектрометрією (ВЕРХ-МС), де: 1 - рівень продукції моеноміцинів штамом S. ghanaensis ATCC 14672; 2 - рівень продукції моеноміцинів штамом S. ghanaensis ΔwblAgh. Спосіб можна проілюструвати прикладами: Конструюють плазміду pKCwblAgh-neo. Спершу, для конструювання плазміди pUCwblAgh, фрагмент ДНК із хромосоми S. ghanaensis, який містить ген wblAgh ампліфіковують за допомогою праймерів wblA-gh-up (5'-AAATCTAGACACGTAGTCGCAGAAGAATC-3') та wblA-ghrev (5'-AAAGAATTCGACATCGATCCACTGATCGA-3'). Отриманий, фрагмент розміром 3,5 т.п.н. клонують у вектор pUC57, який обробляють ендонуклеазою рестрикції EcoRV та лігують з вищеописаним ампліфікованим фрагментом. Лігазною сумішшю трансформують штам Е. coli DH5α та відбирають клони, що несуть рекомбінантні плазміди на середовищі LA з 25 мкг/мл апраміцину, 65 мкг/мл 5-бром-4-хлор-3-індоліл-β-D-галактозиду та 30 мкг/мл ізопропілтіогалактозиду. Плазмідну ДНК із трансформантів аналізують рестрикційним картуванням ферментами HindIII та EcoRI [Гловер Д. Молекулярное клонирование ДНК. Методы. М., Мир - 1989 - т. 1 - 374 с.]. Отриману плазміду позначають pUCwblAgh. Плазмідою pUCwblAgh трансформують штам Е. coli BW25113, що містить плазміду pIJ790 з red-генами фага λ під контролем індуцибельного промотора PBAD [Gust В., Chandra G., Jakimowicz D., Yuqing Т., Bruton C.J., Chater K.F. Lambda red-mediated genetic manipulation of antibiotic-producing Streptomyces // Adv. Appl. Microbiol. - 2004. - vol. 54. - P. 107-128]. З плазміди pKD4 [Datsenko K., Wanner B.L.A. One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products // Proc. Natl. Acad. Sci. USA. - 2000. - vol. 97. - P. 66406645] за допомогою мегапраймерів wblA-gh-p1 (5'-TTCGTTCAGGGAGCAGCGCAGAACAGG GCCAAGGCGGTGGTGTAGGCTGGAGCTGCTTC-3') та wblA-gh-p2 (5'GACCCCCGCGGGTGACCGAGGACCCCTGAGGAACCCTCACATATGAA TATCCTCCTTAG-3') ампліфікують фрагмент ДНК розміром 1.6 т.п.н., що містить ген стійкості до канаміцину neo. Праймери підбирають так, щоб вони складалися з двох частин: перші 39 п.н. гомологічні до послідовності ДНК, що оточує ген wblAgh та містять старт- і стоп-кодони гена, наступні 20 – ділянки P1I та P2 - до гена канаміцин-стійкості neo. В результаті ампліфікований фрагмент ДНК містить ген neo, 5'- і 3'-кінці якого містять по 39 п.н. гомологічних до послідовностей, які фланкують ген wblAgh. Цей лінійний фрагмент ДНК трансформують за допомогою електропорації у клітини Е. coli BW25113 (pIJ790; pUCwblAgh). Для цього Е. coli BW25113 (pIJ790; pUCwblAgh) вирощують протягом 18 год. при 30 °C в 10 мл середовища 2xYT [Hanahan D. Studies on transformation of Escherichia coli with plasmids // J. Моl. Biol. - 1983. - vol. 166. - P. 557-580], яке містить 25 мкг/мл хлорамфеніколу та 100 мкг/мл ампіциліну. 150 мкл культури Е. coli BW25113 (pIJ790; pUCwblAgh), вирощеної вночі протягом 12 год., інокулюють у 15 мл середовища 2xTY з 25 мкг/мл хлорамфеніколу та 100 мкг/мл ампіциліну, додають 150 мкл 1 М L-арабінози. Культуру вирощують до оптичної густини OD 600≈0,4 протягом 3-4 год., переносять у пробірки з об'ємом 15 мл та осаджують центрифугуванням при 4 тис. об./хв протягом 10 хв. Зливають супернантант та клітини ресуспендують в 1 мл охолодженої дистильованої води. Центифугують клітини протягом 30 сек., та повторюють промивання клітин холодною водою ще два рази. На четвертий раз клітини промивають у 1 мл охолодженого 10 % гліцеролу та ресуспендують у 100 мкл 10 % гліцеролу. 50 мкл отриманої суспензії клітин змішують із 1-2 мкл ПЛР продукту (ген пео) та переносять у кювету для елетропорації з відстанню між електродами 1мм. Елетропорацію здійснюють на приладі ВТХ Electroporator ECM 630 із такими параметрами: опір - 200 Ом, ємність - 25 мкФ, напруга - 1,8 кВ. Після електричного розряду негайно додають 1 мл охолодженого середовища 2xTY до кювети і інкубують її протягом 3 год. при 37 °C та висівають на чашки з середовищем LA з 100 мкг/мл ампіциліну та 50 мкг/мл канаміцину. Чашки інкубують при 37 °C 16 год. після чого трансформанти пересівають на свіже середовище LA з 50 мкг/мл канаміцину. Плазмідну ДНК із трансформантів - аналізують рестрикційним картуванням ферментом PstI. Отриману плазміну, позначену pUCwblAgh-neo, обробляють ендонуклеазами рестрикції HindIII та EcoRI. Вектор рКС1139 [Kieser Т., Bibb M.J., Buttner M.J., Chater K.F., Hopwood D.A. Practical Streptomyces genetics // John Innes Foundation, Norwich, United Kingdom. 2000. - 613 pages] обробляють ендонуклеазами рестрикції Hindlll та EcoRI та лігують з вищеописаним фрагментом, що містить зруйнований ген wblAgh. Лігазною сумішшю 2 UA 71729 U 5 10 15 20 25 30 35 40 45 50 55 60 трансформують Е. coli DH5α та відбирають клони, що несуть рекомбінантні плазміди на середовищі LA з 25 мкг/мл апраміцину, 65 мкг/мл 5-бром-4-хлор-3-індоліл-β-D-галактозиду та 30 мкг/мл ізопропілтіогалактозиду. Плазмідну ДНК із трансформантів аналізують рестрикційним картуванням ферментами HindIII та EcoRI. Отриманою плазмідою pKCwblAgh-neo трансформують штам Е. coli ЕТ12567 (pUB307), який за рахунок tra-генів плазміди pUB307 забезпечує кон'югативне перенесення корезидентних плазмід [Flett F., Mersinias V., Smith C.P. High efficiency intergeneric conjugal transfer of plasmid DNA from Escherichia coli to methyl-DNA-restricting Streptomycetes // FEMS Microbiol. Lett. - 1997. vol. 155. - P. 223-229]. Одну колонію культури E. coli ET12567 (pUB307), вирощеної вночі протягом 12 год., засівають у 10 мл середовища LB з 50 мкг/мл канаміцину. Культуру вирощують до оптичної густини OD600≈0,1, переносять у мікропробірки з об'ємом 1,5 мл та осаджують центрифугуванням при 8 тис. об./хв протягом 1 хв. Зливають супернантант та клітини ресуспендують у 50 мкл середовища LB. Отриману суспензію клітин охолоджують у льоді 5 хв, додають 1 мл 0,1М розчину СаСl2 та інкубують у льоді 1 год. Клітини осаджують центрифугуванням при 8 тис. об./хв протягом 1 хв, зливають надосадову рідину та ресуспендують у 100 мкл 0.1М розчину СаСl 2. Інкубують 1 год. у льоді та додають розчин плазмідної ДНК. Інкубують 1 год. у льоді, після чого клітини піддають тепловому шоку протягом 1 хв при 40 °C, охолоджують та додають 1 мл середовища LB. Інкубують 2 год. при 37 °C для індукції експресії генів стійкості та висівають на чашки з середовищем LA з 25 мкг/мл апраміцину та 50 мкг/мл канаміцину. Чашки інкубують при 37 °C 16 год. після чого трансформанти пересівають на свіже середовище LA з 25 мкг/мл апраміцину та 50 мкг/мл канаміцину. Плазміду pKCwblAgh-neo в S. ghanaensis переносять шляхом міжродової кон'югації з відповідним штамом Е. coli ET12567 (pUB307; pKCwblAgh-neo). Суспензію спор штаму S. ghanaensis висівають на середовище ОМ г/л: вівсяне борошно - 30, агар - 18, вода водопровідна - до 1 л, рН до стерилізації - 7,0 та вирощують 7 діб при 28 °C для отримання спорової суспензії для кон'югаційних схрещувань. Штам Е. coli ET12567 (pUB307; pKCwblAghneo) висівають на чашку з середовищем LA з 25 мкг/мл апраміцину та 50 мкг/мл канаміцину та вирощують 18 год. при 37 °C. Готують суспензію клітин в 1 мл середовища LB. Одночасно готують суспензію спор штаму S. ghanaensis в 1 мл стерильної води. Спори піддають тепловому шоку протягом 10 хв. при 50 °C. Суспензію клітин та спор стрептоміцетів осаджують центрифугуванням 2 хв. при 8 тис. об./хв., зливають надосадову рідину, розчиняють у 100 мкл середовища LB, змішують між собою та висівають на чашки з середовищем ОМ. Чашки Інкубують 20 год. при 28 °C та заливають 1 мл водного розчину 25 мкг апраміцину та 50 мкг налідиксової кислоти. Отриманий штам S. ghanaensis (pKCwblAgh-neo) перевіряють на наявність плазміди. Вирощують у 15 мл рідкого середовища TSB та виділяють сумарну ДНК, зразками якої трансформують штам Е. coli DH5a [Kieser Т., Bibb M.J., Buttner M.J., Chater K.F., Hopwood D.A. Practical Streptomyces genetics // John Innes Foundation, Norwich, United Kingdom. - 2000. - 613 pages]. Плазмідну ДНК з трансформантів аналізують рестрикційним картуванням ферментами PstI, HindIII, EcoRI. Отриманий апраміцин-стійкий транскон'югант S. ghanaensis (pKCwblAgh-neo) містить цю плазміду в автономному стані. Для того, щоб плазміда інтегрувалася за гомологією у хромосому S. ghanaensis, вирощують рідку культуру штаму S. ghanaensis (pKCwblAgh-neo) без антибіотика у середовищі TSB при температурі 40 °C протягом 6 діб. Потім 100 мкл рідкої культури транскон'юганта висівають на середовище LA з 50 мкг/мл канаміцином. Відбирають один апраміцин-стійкий та канаміцин-стійкий клон і здійснюють його триразовий пересів за неселективних умов у середовищі TSB для стимуляції проходження вторинного кросинговеру. 100 мкл рідкої культури після пересівів висівають на чашки з середовищем ОМ та за допомогою методу реплік відбирають клони з вторинним кросинговером за ознакою чутливості до апраміцину, як наслідок втрати послідовностей вектора. Один із таких клонів позначено S. ghanaensis ΔwblAgh і використано для дальшого аналізу. Перевіряють рівень продукції моеноміцинів мутантним штамом S. ghanaensis ΔwblAgh. Штам висівають у 15 мл середовища TSB та вирощують протягом 2 діб при 30 °C. 1 мл прекультури інокулюють у 50 мл середовища TSB та вирощують протягом 5 діб при 30 °С. Біомасу з 50 мл середовища концентрують центрифугуванням і промивають дистильованою водою. До біомаси додають 0,5 мл води та 7-8 мл метанолу. Суміш ретельно гомогенізують та інкубують при температурі 37 °C протягом 12 год. Для отримання напівочищених зразків моеноміцинів для ВЕРХ-МС сирцевий екстракт очищують за допомогою твердофазних колонок Alltech С18 - 100 мг смоли-носія - згідно з такою 3 UA 71729 U 5 10 15 20 25 30 схемою: а) активація колонки - 1 мл метанолу, потім 1 мл деіонізованої води; б) нанесення зразка - 2 мл; в) промивка колонки - 1 мл води, 1 мл суміші етилацетат: метанол у співвідношенні 3: 1; г) елюювання моеноміцинів - 1,5 мл метанолу. Отриманий елюат висушують у вакуумній сушці SpeedVac, сухий залишок розчиняють у 150 мкл води з 0,1 % гідроксиду амонію, 20 мкл розчину використовується для ВЕРХ-МС. Таким чином отримують зразки моеноміцинів, на Фіг. 1 показано будову молекули моеноміцину А - основного представника антибіотиків моеноміцинового ряду. Для ВЕРХ-МС використовують прилади Agilent 1100 LC/MSD ESI, Agilent 1150 LC/MSD TOF та Agilent 6520 Accurate-Mass qTOF LC-MS. Система розчинників: розчин А - вода + 0,1 % гідроксид амонію, розчин Б - ацетонітрил + 0,1 % гідроксид амонію. Як нерухому фазу використовують хроматографічні колонки різного розміру та виробництва з носієм C 18 (Agilent, Phenomenex, Waters). У всіх випадках колонки термостатують (30 °С). Типова програма розділення моеноміцинів: (час, хв - % розчину Б): 0-25, 1,0-25, 10,0-95, 14,0-95, 14,1-25, 20,0-10. За таких умов моеноміцини з повнорозмірним ліпідним ланцюгом 25 карбонів виходять з колонки у районі 10-11 хв. Зразки аналізують у режимі негативної іонізації скануванням аніонів у діапазоні 300-1700 Да. Напруга капіляру становить 4 кВ, фрагментора - 150 В. Температура 2 газів просушування - 300 °C, швидкість потоку - 7 л/хв., тиск на короні розряду - 4 атм/см . При проведенні досліду використовували: для визначення температури - ртутний термометр з похибкою показів ± 0,5 °C; об'єму рідин - мірні циліндри з похибкою показів ± 0,5 мл та лабораторні дозатори з похибкою показів ± 0,05-0,5 мкл; маси речовин - аналітичну вагу "Sartorius" з похибкою показів ± 0,00005 г та лабораторну вагу AXIS з похибкою показів ± 0,05 г; час - електронним годинником з похибкою показів ± 0,5 сек. Для кількісного аналізу продукції моеноміцинів площі мас-піків досліджуваних сполук інтегрують і нормалізують відносно однакової кількості біомаси - сухої ваги. Експеримент повторюють тричі і визначають середнє значення, похибку та стандартне відхилення а. Результати експериментів відображено на Фіг. 2, де: 1 - рівень продукції моеноміцинів штамом S. ghanaensis ATCC 14672; 2 - рівень продукції моеноміцинів штамом S. ghanaensis ΔwblAgh. Мутантний штам продукує у середньому 2,3 раза більше моеноміцинів: 7±1 мкг/л ферментаційного середовища, ніж контрольний штам - 3±1 мкг/л ферментаційного середовища, що підтверджує отримання передбачуваного результату. ФОРМУЛА КОРИСНОЇ МОДЕЛІ 35 Спосіб підвищення біосинтезу антибіотиків моеноміцинового ряду, який базується на спрямованому руйнуванні негативних регуляторів, який відрізняється тим, що, як генрегулятор для руйнування використовують ген wblAgh S. ghanaensis. 4 UA 71729 U Комп’ютерна верстка В. Мацело Державна служба інтелектуальної власності України, вул. Урицького, 45, м. Київ, МСП, 03680, Україна ДП “Український інститут промислової власності”, вул. Глазунова, 1, м. Київ – 42, 01601 5
ДивитисяДодаткова інформація
Назва патенту англійськоюMethod for biosynthesis enhancement of moenomicine antibiotics biosynthesis
Автори англійськоюRabyk Mariia Vasylivna, Ostash Bohdan Omelianovych, Fedorenko Viktor Oleksandrovych
Назва патенту російськоюСпособ повышения биосинтеза антибиотиков моеномицинового ряда
Автори російськоюРабик Мария Васильевна, Осташ Богдан Емельянович, Федоренко Виктор Александрович
МПК / Мітки
МПК: C12Q 1/04, C12Q 1/00, C12Q 1/02
Мітки: моеноміцинового, ряду, спосіб, антибіотиків, підвищення, біосинтезу
Код посилання
<a href="https://ua.patents.su/7-71729-sposib-pidvishhennya-biosintezu-antibiotikiv-moenomicinovogo-ryadu.html" target="_blank" rel="follow" title="База патентів України">Спосіб підвищення біосинтезу антибіотиків моеноміцинового ряду</a>
Спосіб підвищення біосинтезу фосфогліколіпідних антибіотиків

Номер патенту: 63820
Опубліковано: 25.10.2011
Автори: Уокер-Кане С'юзен, Громико Олександр Миколайович, Федоренко Віктор Олександрович, Осташ Богдан Омелянович
МПК: C12Q 1/02, C12Q 1/04, C12Q 1/00
Мітки: підвищення, біосинтезу, спосіб, антибіотиків, фосфогліколіпідних
Формула / Реферат:
Спосіб підвищення біосинтезу фосфогліколіпідних антибіотиків, який базується на надекспресії генів плейотропних регуляторів у актиноміцетах-продуцентах цих антибіотиків, який відрізняється тим, що як регуляторний елемент використовують ген лейцил-тРНКUUА S. lividans, клонований у складі висококопійної плазміди pIJ584 у штамах S. ghanaensis ATCC14672, S. lividans ТК24, S. albus J1074.
Спосіб виявлення антибіотиків тіострептонового ряду

Номер патенту: 91725
Опубліковано: 25.08.2010
Автори: Осташ Богдан Омелянович, Осташ Ірина Степанівна, Федоренко Віктор Олександрович, Мироновський Максим Леонідович
МПК: C12Q 1/04, C12Q 1/02, C12Q 1/00
Мітки: виявлення, ряду, тіострептонового, антибіотиків, спосіб
Формула / Реферат:
Спосіб виявлення антибіотиків тіострептонового ряду, що базується на індукції експресії гена канаміциностійкості nео у Streptomyces lividans, який відрізняється тим, що як векторну молекулу ДНК використовують інтегративну плазміду рIJ6902.
Спосіб підвищення специфічної активності антибіотиків

Номер патенту: 34206
Опубліковано: 11.08.2008
Автори: Дикий Ігор Леонідович, Манський Олександр Анатолійович, Домарьов Анатолій Павлович, Філімонова Наталія Ігорівна
МПК: A61L 2/08
Мітки: специфічно, антибіотиків, активності, підвищення, спосіб
Формула / Реферат:
Спосіб підвищення специфічної активності антибіотиків шляхом обробки останніх γ-опромінюванням, який відрізняється тим, що опромінювання проводять протягом понад 1,0 до 1,5 сек. до одержання дози понад 10 до 15 Грей.
Спосіб підвищення специфічної активності антибіотиків
Номер патенту: 73447
Опубліковано: 15.07.2005
Автори: Стрельников Леонід Семенович, Шевельова Наталія Єфимівна, Шевченко Михайло Іванович, Холупяк Ірина Юріївна, Ковальова Тетяна Іванівна, Дикий Ігор Леонідович, Чуєшов Владислав Іванович, Яковенко Володимир Дмитрович, Велика Марина Марківна
Мітки: спосіб, активності, антибіотиків, специфічно, підвищення
Формула / Реферат:
Способ повышения специфической активности антибиотиков, включающий добавление активаторов антибиотической активности, отличающийся тем, что, с целью снижения аллергенного и дисбиотического эффекта, в качестве активатора используют антигистаминные препараты.
Спосіб підвищення та відновлення чутливості патогенної мікрофлори до антибіотиків

Номер патенту: 47879
Опубліковано: 15.07.2002
Автори: Мартинець Петро Андрійович, Негара Вікторія Анатоліївна, Жалдаченко Вікторія Петрівна
МПК: A61K 31/715
Мітки: підвищення, мікрофлори, спосіб, відновлення, антибіотиків, патогенної, чутливості
Формула / Реферат:
Спосіб підвищення та відновлення чутливості патогенної мікрофлори до антибіотиків шляхом безпосереднього впливу медикаментозних препаратів на мікрофлору, який відрізняється тим, що хворому додатково проводять 5-7 інгаляцій натрієвої солі гепарину в дозі 5000 Од у 1мл 0,85% розчину хлориду натрію один раз на добу протягом 15 хвилин.
Попередній патент: Робочий орган скребкового транспортера-змішувача
Наступний патент: Спосіб лікування аритмій у хворих на цукровий діабет
Випадковий патент: Спосіб одержання оптично активних або рацемічних ефірів циклопропанкарбонових кислот