Стимуляція тимозином генетичної імунізації






Формула / Реферат
1. Спосіб імунізації індивідуума, чутливого до інфекції, яка викликається вірусом гепатиту С, проти вказаної інфекції, який відрізняється тим, що індивідууму вводять полінуклеотид, який кодує один або декілька структурних пептидів або пептидів оболонки вірусу гепатиту С або їх фрагменти, які зберегли здатність вірусного протеїну викликати імунну відповідь, у сполученні з одним або декількома α-тимозинами, які мають активність тимозину α1.
2. Спосіб за п. 1, який відрізняється тим, що полінуклеотид кодує один або декілька протеїнів оболонки вірусу гепатиту С.
3. Спосіб за п. 1, який відрізняється тим, що полінуклеотид кодує один або декілька протеїнів вірусу гепатиту С, вибраних із групи, яка включає протеїн NS3, протеїн NS4A, протеїн NS4B, протеїн NS5A і протеїн NS5B.
4. Спосіб за п. 3, який відрізняється тим, що полінуклеотид кодує один або декілька протеїнів NS5.
5. Спосіб за будь-яким з пп. 1-4, який відрізняється тим, що α-тимозин являє собою тимозин α1.
6. Спосіб за будь-яким з пп. 1-4, який відрізняється тим, що тимозин вводять у дозі від 0,001 мг/кг ваги тіла/день до 10 мг/кг ваги тіла/день.
7. Спосіб за п. 6, який відрізняється тим, що тимозин вводять у дозі приблизно 0,02 мг/кг ваги тіла /день.
8. Застосування полінуклеотиду, який кодує один або декілька структурних пептидів або пептидів оболонки вірусу гепатиту С або їх фрагменти, які зберегли здатність вірусного протеїну викликати імунну відповідь, разом з одним або декількома α-тимозинами, які мають активність тимозину α1, для приготування фармацевтичної композиції для імунізації індивідуума, чутливого до інфекції, яка викликається вірусом гепатиту С, проти вказаної інфекції.
9. Застосування за п. 8, яке відрізняється тим, що полінуклеотид кодує один або декілька протеїнів оболонки вірусу гепатиту С.
10. Застосування за п. 8, яке відрізняється тим, що полінуклеотид кодує один або декілька протеїнів вірусу гепатиту С, вибраних із групи, яка включає протеїн NS3, протеїн NS4A, протеїн NS4B, протеїн NS5A і протеїн NS5B.
11. Застосування за п. 10, яке відрізняється тим, що полінуклеотид кодує один або декілька протеїнів NS5.
12. Застосування за будь-яким з пп. 8-11, яке відрізняється тим, що α-тимозин являє собою тимозин α1.
13. Застосування за будь-яким з пп. 8-11, яке відрізняється тим, що тимозин вводять у дозі від 0,001 мг/кг ваги тіла/день до 10 мг/кг ваги тіла/день.
14. Застосування за п. 13, яке відрізняється тим, що тимозин вводять у дозі приблизно 0,02 мг/кг ваги тіла /день.
15. Фармацевтична композиція, яка містить полінуклеотид, який кодує один або декілька структурних пептидів або пептидів оболонки вірусу гепатиту С або їх фрагменти, які зберегли здатність вірусного протеїну викликати імунну відповідь, разом з одним або декількома α-тимозинами, які мають активність тимозину α1.
16. Фармацевтична композиція за п. 15, яка відрізняється тим, що полінуклеотид кодує один або декілька протеїнів оболонки вірусу гепатиту С.
17. Фармацевтична композиція за п. 15, яка відрізняється тим, що полінуклеотид кодує один або декілька протеїнів вірусу гепатиту С, вибраних із групи, яка включає протеїн NS3, протеїн NS4A, протеїн NS4B, протеїн NS5A і протеїн NS5B.
18. Фармацевтична композиція за п. 17, яка відрізняється тим, що полінуклеотид кодує один або декілька протеїнів NS5.
19. Фармацевтична композиція за будь-яким з пп. 15-18, яка відрізняється тим, що α-тимозин являє собою тимозин α1.
20. Фармацевтична композиція за будь-яким з пп. 15-18, яка відрізняється тим, що тимозин вводять у дозі від 0,001 мг/кг ваги тіла/день до 10 мг/кг ваги тіла/день.
21. Фармацевтична композиція за п. 20, яка відрізняється тим, що тимозин вводять у дозі приблизно 0,02 мг/кг ваги тіла /день.
Текст
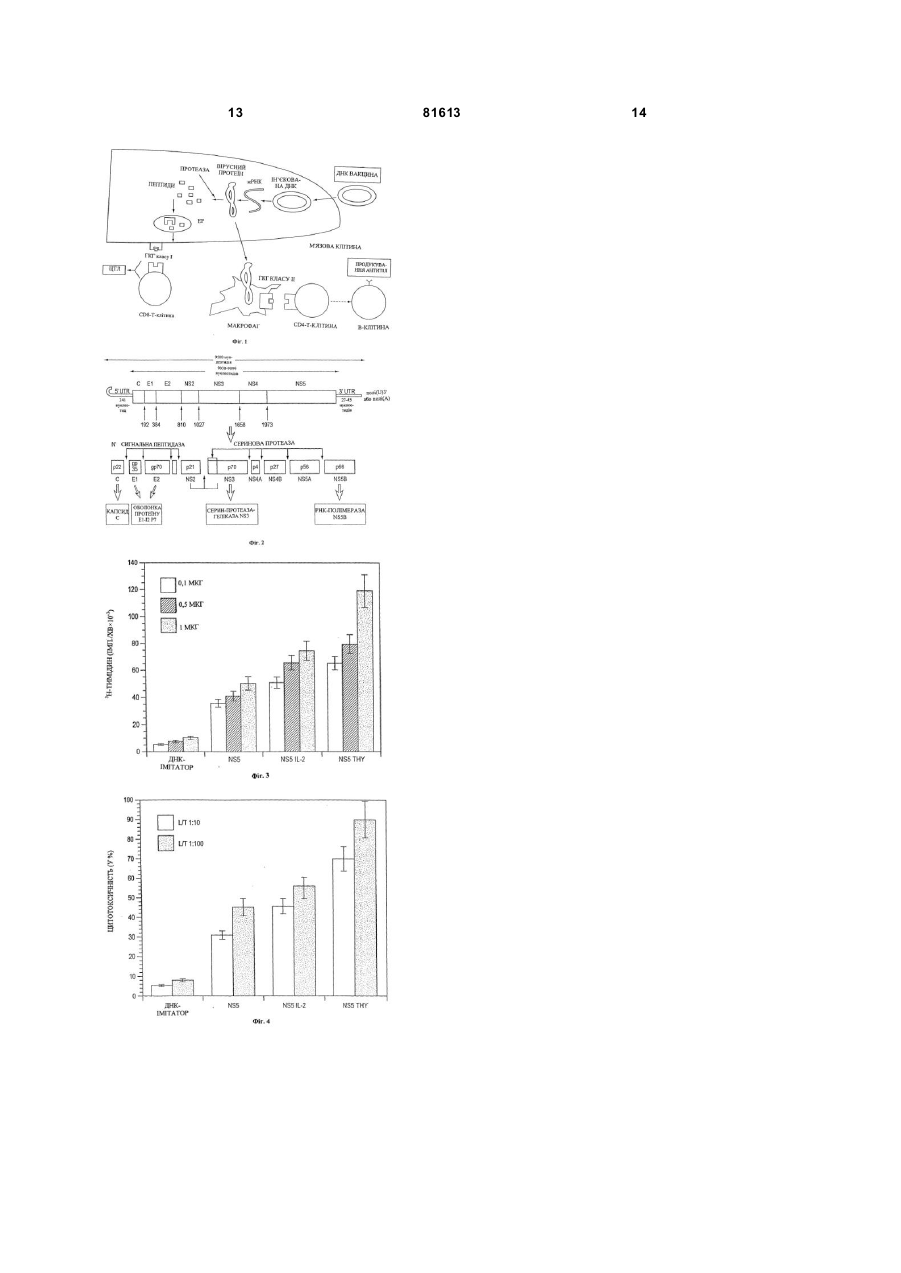
1. Спосіб імунізації індивідуума, чутливого до інфекції, яка викликається вірусом гепатиту С, проти вказаної інфекції, який відрізняється тим, що індивідууму вводять полінуклеотид, який кодує один або декілька структурних пептидів або пептидів оболонки вірусу гепатиту С або їх фрагменти, які зберегли здатність вірусного протеїну викликати імунну відповідь, у сполученні з одним або декількома α-тимозинами, які мають активність тимозину α1. 2. Спосіб за п. 1, який відрізняється тим, що полінуклеотид кодує один або декілька протеїнів оболонки вірусу гепатиту С. 3. Спосіб за п. 1, який відрізняється тим, що полінуклеотид кодує один або декілька протеїнів вірусу гепатиту С, вибраних із групи, яка включає протеїн NS3, протеїн NS4A, протеїн NS4B, протеїн NS5A і протеїн NS5B. 4. Спосіб за п. 3, який відрізняється тим, що полінуклеотид кодує один або декілька протеїнів NS5. 5. Спосіб за будь-яким з пп. 1-4, який відрізняє ться тим, що α-тимозин являє собою тимозин α1. 6. Спосіб за будь-яким з пп. 1-4, який відрізняє ться тим, що тимозин вводять у дозі від 0,001 мг/кг ваги тіла/день до 10 мг/кг ваги тіла/день. 7. Спосіб за п. 6, який відрізняється тим, що тимозин вводять у дозі приблизно 0,02 мг/кг ваги тіла /день. 8. Застосування полінуклеотиду, який кодує один або декілька структурних пептидів або пептидів оболонки вірусу гепатиту С або їх фрагменти, які зберегли здатність вірусного протеїну викликати імунну відповідь, разом з одним або декількома α 2 (19) 1 3 81613 4 19. Фармацевтична композиція за будь-яким з пп. 15-18, яка відрізняється тим, що α-тимозин являє собою тимозин α1. 20. Фармацевтична композиція за будь-яким з пп. 15-18, яка відрізняється тим, що тимозин вводять у дозі від 0,001 мг/кг ваги тіла/день до 10 мг/кг ваги тіла/день. 21. Фармацевтична композиція за п. 20, яка відрізняє ться тим, що тимозин вводять у дозі приблизно 0,02 мг/кг ваги тіла /день. За даною заявкою заявляється пріоритет відповідно до попередньої заявки на патент 60/330638, поданої 26 жовтня 2001 р. Хронічний гепатит, який викликається вірусом гепатиту С (HCV), являє собою дуже серйозну медичну проблему і приводить до хронічного захворювання печінки, цирозу і печінковоклітинного раку. Передбачається, що в більшості випадків вірус гепатиту С є збудником набутого після перфузії гепатиту. Незважаючи на поліпшення якості депо донорської крові і проведення тестування крові донорів, після перфузії усе ще часто зустрічаються випадки гострої інфекції в людей. Хронічний гепатит розвивається принаймні в половини пацієнтів, які страждають від гострої форми гепатиту, який викликається HCV (приблизно в 90% пацієнтів, які страждають від гепатиту ні А, ні В (ВГНАНБ)), а цироз розвивається принаймні в 20% представників цієї групи. Для призупинення або сповільнення розвитку захворювань, пов'язаних з HCV, був вивчений цілий ряд лікарських засобів. Сучасне стандартне лікування передбачає застосування інтерферону альфа і рибавірину. Однак значна кількість пацієнтів нечутливі до такого лікування. Було встановлено, що генетична імунізація стимулює широкий спектр клітинних імунних відповідей на структурні і неструктурні протеши HCV. Однак за допомогою декількох моделей з використанням тварин було встановлено, що різні конструкції, які несуть HCV, мають невисоку біологічну активність [Tokushige і ін. (1996)]. Генетична імунізація є новою альтернативою застосуванню традиційних заснованих на використанні антигенів вакцин, таких як вакцини, які включають ослаблений вірус або субодиниці протеїнів. При генетичній імунізації для імунізації реципієнта застосовують «оголену» ДНК. ДІЖ називають «оголеною» тому, що вона не містить ніякого носія для введення збудника інфекції, який може полегшувати проникнення в клітину, такого як вірусні частинки. Коли вводять «оголену» ДНК, організм реципієнта може експресувати протеши, що кодуються плазмідною ДНК, які потім можуть стимулювати специфічну імунн у відповідь, у якій беруть участь цитотоксичні Т-клітини, Т-клітинихелпери й антитіла. Застосування вакцин на основі ДНК усуває необхідність в очищенні патогену або імунозахисного антигену для вакцинації й усуває можливість реверсії вірулентності, оскільки ДНК кодує індивідуальний вірусний протеїн. Застосування для імунізації полінуклеотидів має численні переваги. Наприклад, імунізацію клітин можна здійснювати з використанням будь якого антигену, який кодується полінуклеотидом. Крім того, антигени, які кодуються полінуклеотидами, експресуються як «чисті» антигени в їх нативному стані і піддані звичайним модифікаціям у клітині-хазяїні. Полінуклеотиди також легко піддавати не дорогим маніпуляціям, і вони стабільні в складі безводного продукту або в розчині в широкому діапазоні температур. Таким чином, ця технологія має велике значення для розробки високоефективних субодиничних вакцин. Гуморальна і/або клітинно-опосередкована (клітинна) імунна відповідь, яка викликається, може забезпечувати захист або захисний імунітет проти інфекції, яка викликається патогенними агентами, такими як бактерії, віруси і еукаріотичні організми (наприклад, паразити). Потім захисні гуморальні і/або клітинні імунні відповіді впливають на інфективність або активність патогену або обмежують його поширення або ріст, що приводить до захисту від наступного зараження патогеном. Імунна відповідь може також забезпечувати захист від захворювань або порушень, які пов'язані з клітинами, що продукують специфічні протеши. При оцінці в дослідах на різних тваринах встановлено, що генетична імунізація за допомогою внутрішньом'язової ін'єкції є ефективною у відношенні багатьох вірусів (наприклад, імунізація морських свинок проти інфекції, яка викликається вірусом герпесу простого (HSV) типу 2 [Bourne і ін. (1996)]; імунізація мишей проти вірусу грипу [Fu і ін. (1997)]; вакцинація курчат проти вірусів грипу [Kodihalli і ін. (1997)]; і ссавців та птахів проти ротавірусів [Herrmann і ін. U.S. 5620896)]. Daheshia зі співавторами (1997) описали, що одна обробка рогівки тварин, ураженої герпетичним стромальним кератитом, «оголеною» ДНК, яка кодує IL-1, приводила до зменшення ушкоджень у тварин, викликаючи ремісію ушкоджень. Обговорення генетичної імунізації в цілому і її застосування див., наприклад, у [U.S. 5830876 (Weiner і ін.)]. Було встановлено, що представники класу поліпептидів-імуномодуляторів тимусного походження, тобто тимозини, запускають процеси, пов'язані з дозріванням лімфоцитів, стимулюють Т-клітинну функцію і підсилюють відновлення імунних дефектів. Тимозин α1 (ΤΗΝα1) являє собою кислотний поліпептид, який складається з 28 амінокислот, з молекулярною масою 3100, який має виражену імунологічну активність, таку як стимуляція виробництва а- і γ-інтерферонів, підвищення виробництва фактора, який інгібує міграцію макрофагів, індукція експресії Т-клітинних маркерів, рецепторів IL-2 і підвищення активності 5 81613 Т-клітин-хелперів. Виділення, характеристики і застосування ΤΗΝαί описані, наприклад, у [U.S. 4079127]. Для лікування хронічного гепатиту, який викликається HCV, як монотерапія застосовують різні антивірусні агенти, такі як ацикловір, відарабін і аденінарабінозид. Монотерапія з використанням вказаних антивірусних агентів, як правило, є неефективною або через високу токсичність агента, або через те, що агент спочатку викликає деяке інгібування реплікації вірусу, але не може підтримувати інгібування реплікації вірусу протягом тривалого часу [див, наприклад, Alexander і ін. (1988)]. Таким чином, зберігається необхідність у розробці лікування захворювань, які опосередковуються HCV, яке сполучило б ефективність у відношенні вірусу з меншими побічними діями, викликало модуляцію відповідей імунної системи і приводило до зниження частоти рецидивів. У винаході описане застосування α-тимозину для стимуляції CD4+ - і СО8+-клітинами імунних відповідей на імуногенні пептиди HCV, переважно на протеїн NS5 HCV. Стимуляція клітинних імунних відповідей на вірусні і клітинні протеїни являє собою важку задачу. У винаході описане застосування in vivo α-тимозину для різкого посилення цитотоксичної і проліферативної Тклітинних відповідей на епітопи протеїнів HCV. У переважному варіанті здійснення винаходу тимозин являє собою тимозин α1. Самі по собі вакцини, основою яких є ДНК, являють собою нову технологію і тому відрізняються від раніше відомих типів вакцинації (так, усі вакцини, основою яких є протеїни, пептиди або вбиті віруси, викликають гуморальний або опосередковуваний антитілом імунітет, у той час як вакцини на основі ДНК обумовлюють клітинний імунітет), а той факт, що α-тимозин підвищує ефективність вакцини, забезпечує нову і несподівану перевагу при застосуванні вакцин на основі ДНК для лікування і профілактики захворювань, пов'язаних з HCV. Короткий опис креслень На кресленнях показано: на фіг.1 - схематичне зображення механізму імунізації з використанням вакцини на основі ДНК; на фіг.2 - схематичне зображення геному вірусу гепатиту С (HC V); на фіг.3 - вплив спільного введення 5мкг тимозину α1 (і.р.) і полінуклеотиду, який кодує NS5, на Т-клітинну проліферацію при використанні трьох концентрацій для контрольного зараження (0,1,0,5 і 1мкг); на фіг.4 - вплив спільного введення 5 мкг тимозину α1 (і.р.) і полінуклеотиду, який кодує NS5, на цитотоксичну реакцію при двох співвідношеннях лімфоцит-ефектор/мішень (L/T). У винаході описане застосування α-тимозину в сполученні з заснованою на використанні ДНК або «генетичною» імунізацією для різкого посилення клітинних імунних відповідей на HCV. Переважним варіантом здійснення винаходу є стимуляція вакцин на основі ДНК за допомогою тимозину а! (тималфасин). Винахід може знайти широке 6 застосування для розробки профілактичних і терапевтичних вакцин і для значного посилення клітинних імунних відповідей на вірусні і клітинні протеїни після заснованої на застосуванні ДНК імунізації. Полінуклеотид, який кодує імуногенний пептид, поліпептид або протеїн HCV вводять безпосередньо тварині in vivo у сполученні з одним або декількома α-тимозинами. Полінуклеотид кодує поліпептид, який несе принаймні один епітоп імуногенного протеїну HCV, який є мішенню. Полінуклеотид експресують з використанням клітин індивідуума для одержання імуногенних протеїнів-мішеней, які викликають імунну відповідь широкого спектра проти HCV. Геном HCV кодує два протеїни оболонки (Е1 і Е2) і шість стр уктурних протеїнів (NS2, NS3, NS4A, NS4B, NS5A і NS5B) (фіг.2). Згідно із даним винаходом можна використовувати полінуклеотиди, які кодують будь-який з цих вірусних протеїнів або їх комбінації або фрагменти. Під обсяг винаходу підпадають нативний (тобто який зустрічається в природних умовах) α-тимозин, а також синтетичний α-тимозин і рекомбінантний α-тимозин, які мають амінокислотну послідовність нативних α-тимозинів, амінокислотні послідовності практично аналогічні до неї, або отримані з неї укорочені послідовності, і їх біологічно активні аналоги, що мають послідовності, які несуть заміни, делеції, подовження, заміщення або інші модифікації, і активність яких практично аналогічна до активності нативного α-тимозину. Переважний тимозин являє собою тимозин α1. Виділення, характеристики і застосування αтимозину описані, наприклад, у [U.S. 4079127, U.S. 4353821, U.S. 4148788 і U.S.4116951]. Кількість αтимозину, необхідна для того, щоб викликати потрібний ступінь стимуляції дії вакцини на основі ДНК, можна визначати за допомогою стандартних експериментів за типом «доза-титрування». Було встановлено, що α-тимозин безпечний для людей при введенні в дозах до 16 мг/кг ваги тіла/день. Переважна доза α-тимозину становить від 0,001 до 10мг/кг ваги тіла/день, причому найбільш переважною є доза приблизно 0,02 мг/кг ваги тіла/день. Полінуклеотидні послідовності, запропоновані у винаході, являють собою послідовності ДНК або РНК, які кодують антигенні/імуногенні поліпептиди HCV, функціонально зв'язані з послідовностями, які регулюють транскрипцію. Ці послідовності можна застосовувати в сполученні з іншими полінуклеотидними послідовностями, які кодують регуляторні протеши, що контролюють експресію цих поліпептидів. Регуляторний протеїн може діяти шляхом зв'язування з геномною ДНК таким чином, щоб регулювати її транскрипцію; в іншому варіанті він може діяти шляхом зв'язування з матричною РНК, підвищуючи або знижуючи її стабільність або ефективність трансляції. Поняття «функціонально зв'язані з послідовностями, які регулюють транскрипцію і трансляцію» означає, що послідовність, яка кодує поліпептид, і мінімальні послідовності, які контролюють транскрипцію і трансляцію, зв'язані 7 81613 таким чином, щоб забезпечувати експресію поліпептиду, коли відповідні молекули (наприклад, протеїни-активатори транскрипції) зв'язані з регуляторною(ими) послідовністю(ями). Полінуклеотид функціонально кодує поліпептид, коли він несе всю генетичну інформацію, необхідну для експресії в клітині-мішені, таку як промотори і т.п. Поняття «промотор» або «промоторна послідовність» у контексті даного опису стосується мінімальної послідовності, достатньої для контролю транскрипції. Полінуклеотидна послідовність ДНК, як правило, обмежена сайтом ініціації і сайтом термінації, тобто являє собою одиницю транскрипції ДНК, і транскрибується з утворенням первинного транскрипту. Полінуклеотидний продукт, введений у клітини in vivo, може мати будь-яку кількість форм, і даний винахід не обмежений яким-небудь конкретним полінуклеотидом, який кодує який-небудь конкретний поліпептид HCV, хоча полінуклеотиди, які кодують протеїн NS5 або його фрагмент, є переважними. Плазміди, що містять гени, які кодують антигени або імуногени HCV, описані в літературі і фахівці в даній галузі легко можуть їх одержувати [див., наприклад, Tokushige і ін., 1996]. Полінуклеотид може кодувати один або декілька антигенів, таких як антигени з двох або більшої кількості різних вірусних протеїнів. В альтернативному варіанті полінуклеотид може містити дві або більшу кількість різних послідовностей ДНК, при цьому одна послідовність кодує антиген, а інша(і) кодує(ють) поліпептиди, що можуть мати або можуть не мати антигенні властивості. Наприклад, вектор може кодувати два (або більшу кількість) антигени HCV. Відповідно до іншого варіанта здійснення винаходу інший(і) поліпептид(и) може(жуть) сприяти посиленню імунної відповіді проти HCV (наприклад, епітопи-хелпери, цитокіни, поліпептиди-носії, субодиниці холерного токсину або інші імуностимулятори). Полінуклеотид додатково може бути вбудований у вектор, який несе послідовності для експресії полінуклеотиду. Коли в одному векторі присутні два або більша кількість послідовностей ДНК, які кодують поліпептиди, то транскрипція кожної послідовності ДНК, яка кодує антиген, може знаходитися під контролем її власного промотору для експресії двох або більшої кількості не злитих поліпептидів. В іншому варіанті один промотор може контролювати експресію двох або більшої кількості послідовностей ДНК, які кодують антиген, злитих у рамці зчитування один з одною, для експресії злитого протеїну. Застосовувані в цих методах полінуклеотиди можуть являти собою послідовності, не інтегровані в геном клітинихазяїна. Вони можуть являти собою послідовності ДНК, які не реплікуються, або специфічні послідовності, які реплікуються, сконструйовані за допомогою методів генної інженерії таким чином, щоб у них була відсутня здатність до інтеграції в геном. Полінуклеотид можна вводити пацієнту в присутності ад'ювантів або інших субстанцій, які можуть підвищувати здатність нуклеїнової кислоти 8 до проникнення або підвищувати рекрутмент клітин імунної системи в ділянці інокуляції. Слід відзначити, що сам полінуклеотид експресується в клітині-хазяїні за допомогою факторів транскрипції, які властиві клітині-хазяїну або одиниці транскрипції ДНК. Відповідно до способів, запропонованих у винаході, у клітини для одержання в них продукту трансляції у вигляді поліпептиду можна вводити як експресовану ДНК, так і мРНК. Якщо нуклеїнові кислоти містять відповідні контролюючі послідовності, то вони повинні забезпечувати синтез відносно великих кількостей кодованого протеїну. Дозу імуногенного поліпептиду легко може визначити практикуючий лікар або ветеринар з використанням як моделі тваринних або інших систем для тестування, які добре відомі в даній галузі. Композиції повинні містити ефективну кількість ДІЖ у водному розчині. Кількість ДНК, що підлягає введенню, залежить від таких факторів, як вік, вага і фізіологічний стан індивідуума, який підлягає вакцинації. Кількість ДНК залежить також від здатності імунної системи індивідуума синтезувати антитіла і від рівня потрібного захисту. Ефективні дози може легко визначити фа хівець у даній галузі за допомогою звичайних дослідів, необхідних для побудови графіків залежності відповіді від дози. Переважна доза становить від 1 мкг/кг ваги тіла до 1 мг/кг ваги тіла індивідуума. Індивідуума можна імунізувати шляхом однократного введення ДНК або за допомогою декількох введень. Декілька введень може знадобитися для підтримання стану імунітету індивідуума до конкретного патогену. Композиції гетерологічних полінуклеотидів і метод їх конструювання для успішної трансформації добре відомі фахівцю в даній галузі, і такі композиції і методи можна застосовувати для одержання полінуклеотидів, запропонованих у винаході. Конкретна композиція полінуклеотидів не має вирішального значення для даного винаходу, і винахід не залежить від композиції конкретних полінуклеотидів, які застосовуються для трансформації. Прийнятні компоненти полінуклеотиду, такі як промотори, послідовності для поліаденілування, сигнали термінали, сигнали сплайсингу, селектовані маркери, репортерні гени, енхансери, вірусні реплікони, інтрони і бактеріальні плазмідні послідовності, добре відомі в даній галузі. У Sambrook із співавторами (1989) описані прийнятні методи конструювання гетерологічних полінуклеотидів. Полінуклеотиди можна одержувати за допомогою цілого ряду відомих методів. Наприклад, ДНК, яка кодує попередньо відібраний антиген, можна вбудовува ти в експресійний вектор (див., наприклад, Sambrook і ін. (1989)). При наявності обладнання для автоматичного синтезу нуклеїнових кислот ДНК можна синтезувати безпосередньо, якщо нуклеотидна послідовність є відомою, або за допомогою сполучення методів ПЛР, клонування і ферментації. Крім того, якщо послідовність попередньо відібраного поліпептиду 9 81613 є відомою, то можна передбачити прийнятну кодувальну послідовність полінуклеотиду. Коли полінуклеотид являє собою мРНК, то її легко можна одержувати in vitro з відповідної ДНК. Наприклад, відповідно до загальноприйнятих методик фагові РНК-полімерази SP6, ТЗ або Т7 застосовують для одержання мРНК на ДНКматриці в присутності індивідуальних рибонуклеозидтрифосфатів. Відповідний промотор фага, такий як сайт ініціації реплікації фага Т7, поміщають у ДНК-матриці безпосередньо проти ходу транскрипції гена, який підлягає транскрибуванню. Системи, засновані на такому застосуванні фага Т7, добре відомі й описані в літературі. Ще одним об'єктом винаходу є імунологічна композиція, яка після її інтродукції в організм індивідуума, який має здатність або повинен індукувати імунну відповідь, індукує імунну відповідь у такого індивідуума на протеїн HCV, який кодується цією композицією, причому композиція містить полінуклеотид, який кодує і експресує один або декілька антигенів/імуногенів HCV, у сполученні з одним або декількома αтимозинами. Імунну відповідь можна застосовувати в терапевтичних або профілактичних цілях і можна застосовувати для формування зв'язаного з антитілами імунітету або клітинного імунітету, з умовленого, наприклад, ЦТЛ (цитотоксичні Т-лімфоцити) або CD4+-Т-клітинами. Приклад 1 Конструювання плазмід Як джерело вірусних генів плазміду, позначену як pBRTM/HCVl-ЗОП, яка несе повнорозмірну відкриту рамку зчитування (ВРЗ) HCV, застосовували для клонування в експресійних векторах [Grakoui і ін., 1993]. Конструкцію рАр031Ns5 клонували за допомогою ПЛР після вбудовування створених стартового і стоп-кодонів, а також сайтів, які розпізнаються рестриктазами, з використанням наступних праймерів для NS5:5'-T CAG ТСТ AGA ATG ТСС GGC ТСС TGG СТА AGG GA-3' (Abal) [SEQ ID No. 1] і 5'-А GCT ACG CGT TCA CCG GTT GGG GAG GAG GT-3' (Mlul) [SEQ ID No. 2]. Після ПЛР-ампліфікації з використанням ПЛР-системи, яка забезпечує високу точність, (фірма Boehringer Mannheim, Індіанаполіс, шт. Індіана), фрагменти кДНК вбудовували в плазмідний експресійний вектор рАр031, який містить енхансерний елемент вірусу саркоми Рауса і промотор CMV. Конструкціями трансформували клітини лінії DH5a і плазмідну ДНК потім очищали або двократним центрифугуванням у хлориді цезію, або за допомогою набору Qiagen Giga з використанням буферної системи Ендофрі (Endofree) (Сайта Клара, шт. Каліфорнія). Правильність інсерції кДНК, які кодують неструктурні протеїни, підтверджували секвенуваннм з використанням стандартних методів. Для створення ліній клітин, що стабільно експресують NS5, які використовували як клітини-мішені для аналізів ЦТЛ, генні фрагменти, які кодують неструктурні протеїни, клонували також у експресійних 10 векторах pcDNA3 і pcDNA3.1/Zeo(-) (фірма Invitrogen, Сан-Дієго, шт. Каліфорнія) з використанням неоміцину як селектованого маркера. Хbаl- і М«І-фрагмент NS5 субклонували в сайті Nhel/Mlul вектора Litmus-38 (фірма New England Biolabs, Беверлі, шт. Масачусетс), розщеплювали за допомогою EcoRI i Sail і вбудовували шляхом лігування в множинний сайт клонування векторів EcoRI/Xhol pcDNA3 і pcDNA3, I/Zeo(-) відповідно. Hba-l і ЯшиНІ-фрагмент, який містить NS4, вбудовували шля хом лігування у вектор Litmus-29 (фірма New England Biolabs), повторно розщеплювали за допомогою Кргії і EcoRJ, і потім вбудовували шля хом лігування у вектор рс DNA3. Плазміди були позначені як pcDNA3.1/Zeo(-)-NS5. Плазміду, яка експресує ДНК, що кодує мишиний IL-2 (pcD/3-Ш), клонували й очищали за допомогою описаних раніше методів [Geissler і ін., 1997а, 19976, 1997в]. Генетична імунізація Для підвищення проникнення в клітину плазмідної ДНК у кілька ділянок на чотириголовому м'язі мишей лінії Balb/c ін'єкували всього 100мкл 0,25% гупівакаїну (hupivacaine). Через 4 дні плазмідні конструкції ін'єкували в цю ж ділянку у 5 різних місць у кінцевому об'ємі 100мкл 0,9% NaCl. Через 2 тижні мишей піддавали бустерін'єкції, вводячи в протилежну ногу плазмідну ДНК. Через 10 днів після останньої імунізації мишей умертвляли. Для досліду використовували три групи мишей по 5 особин у кожній. Група 1 - миші, яких імунізували 100мкг ДНК-імітатора; група 2 миші, яких імунізували 50мкг рАр031NS5 і 50мкг ДНК-імітатора; група 3 - мишей, яких імунізували 50мкг рАр031 NS5 і 50мкг ДНК-імітатора і яким вводили 5 мкг тимозину αϊ (і.р.) двічі в тиждень. Тварин імунізували три рази через 2 тижні і дослідження проводили через 10 днів після останньої імунізації. Аналіз Т-клітинної проліферації Мишей анестезували ізофлураном (фірма Аеггапе, Анаквест, шт. Нью-Джерсі), брали кров за допомогою ретробульбарної пункції і збирали клітини селезінки. Еритроцити видаляли інкубацією в 8,3% NH4Cl/O,17моля/л Трис (рН 7,4) протягом 10хв при 37°С. Клітини селезінки культивували в трьох повторах з використанням 96-лункових плоскодонних планшетів із щільністю 5´105 клітин на лунку в 100мкл повного середовища (модифікованого за способом Дульбекко середовища Ігла, фірма Mediatech, Вашингтон, федеральний округ Колумбія), що містить 10% ФТС. Клітки селезінки стимулювали рекомбінантним NS5 у різних концентраціях (1, 5, 10мкг/мл). І, нарешті, додавали 2-меркаптоетанол до кінцевої концентрації 50мкмолів/л. Як контроль антигенної специфічності ефекторні клітини стимулювали 10мкг/мл рекомбінантної βсубодиниці людського хоріонічного гонадотропіну, який секретується з клітини і, як було встановлено, є сильним Т-клітинним імуногеном (Geissler, і ін., 1997г). Клітини селезінки стимулювали протягом 3 днів. Після додавання [3Н]-тимідину (1 мкКі /лунку) клітки інкубували протягом 18год. Включення [3Н]тимідину в ДНК оцінювали після збирання (фіг.3). 11 81613 Включення радіоактивності коректували з урахуванням фонової активності (Δ срm (число імпульсів на хв)). Аналіз цитотоксичності Клітини селезінки з організму імунізованих мишей суспендували в повному середовищі DMEM, яке містить 10% ФТС і 50мкмолів/л 2меркаптоетанолу; потім через 5 днів після стимуляції in vitro клітки' аналізували у відношенні цитотоксичної активності. Додавали рекомбінантний мишиний IL-2 у вигляді однієї порції в концентрації 5од./мл і клітини-респондери (4x107) спільно культивували з 6,25´106 сингенних клітин, які стабільно експресують NS5 після обробки 20000 рад. Популяції цитотоксичних ефекторних лімфоцитів збирали після 5-денної інкубації. Протягом 4год у 96-лункових круглодонних планшетах здійснювали аналіз вивільнення 51Сr, використовуючи як клітинимішені мічену за допомогою 51Сr лінію SP2-NS. ЦТЛ-аналізи здійснювали при співвідношеннях лімфоцитів-ефекторів:клітин-мішеней (L/T) 1:10 і 1:100. Цитотоксичність (у відсотках) оцінювали в такий спосіб: Вивільнення в експерименті означає середню кількість імпульсів на хв, які вивільняються клітинами-мішенями в присутності клітинефекторів. Загальне вивільнення означає радіоактивність, яка вивільняється після загального лізису клітин-мішеней 5% TritonX-100 (фіг.4). Спонтанне вивільнення означає присутню в середовищі радіоактивність, отриману тільки з клітин-мішеней. Статистичний аналіз Для порівняння результатів, отриманих для різних груп, застосовували непараметричний Uкритерій Манна-Уітні. Значення Ρ
ДивитисяДодаткова інформація
Назва патенту англійськоюThymosin augmentation of genetic immunization
Автори англійськоюWands Jack R.
Назва патенту російськоюСтимуляция тимозином генетической иммунизации
Автори російськоюВондс Джек Р.
МПК / Мітки
МПК: A61K 38/16, A61K 45/00, A61K 39/29
Мітки: тимозином, стимуляція, генетично, імунізації
Код посилання
<a href="https://ua.patents.su/7-81613-stimulyaciya-timozinom-genetichno-imunizaci.html" target="_blank" rel="follow" title="База патентів України">Стимуляція тимозином генетичної імунізації</a>
Спосіб лікування гліобластоми тимозином-a1

Номер патенту: 77999
Опубліковано: 15.02.2007
Автори: де ла Монте С'юзенн, Вондс Джек Р.
МПК: A61K 38/16, A61P 35/00, A61K 31/17
Мітки: гліобластоми, спосіб, тимозином-a1, лікування
Формула / Реферат:
1. Спосіб лікування гліобластоми, згідно з яким хлоретилнітрозосечовину вводять в комбінації з пептидом тимозин-1 (ТА1).2. Спосіб за п. 1, де хлоретилнітрозосечовина означає BCNU.3. Спосіб за п. 2, де BCNU вводять у дозі 150-200 мг/м2.4. Спосіб за п. 1, де пептид ТА1 означає тимозин-1...
Вакцина для імунізації свиней проти плевропневмонії, спосіб її приготування та спосіб імунізації свиней проти плевропневмонії

Номер патенту: 70287
Опубліковано: 15.10.2004
Автори: Інзана Томас Дж., Уорд Крістін
МПК: A61P 31/04, A61K 39/085, A61P 43/00, A61P 31/00, A61K 39/102, C07K 14/285, A61P 37/04, A61K 39/02, A61K 39/00
Мітки: приготування, імунізації, свиней, вакцина, плевропневмонії, спосіб
Формула / Реферат:
1. Вакцина для імунізації свиней проти плевропневмонії, яка відрізняється тим, що включає авірулентний некапсульований мікроорганізм Actinobacillus pleuropneomoniae, при цьому у вказаного мікроорганізму відсутні ДНК-послідовності, що кодують біосинтез капсули, та вказані послідовності розміщені вище від сайта гібридизації для BamHI-Xbal фрагмента pCW-1C або розміщені вище від гена капсулярного експорту вказаної бактерії.2. Вакцина за...
Застосування похідних n-азабіциклоаміду для лікування чи профілактики психотичних розладів, а також станів, коли корисною є стимуляція нікотинового рецептора a7
Номер патенту: 74172
Опубліковано: 15.11.2005
Автори: Маллен Джордж, Балестра Майкл, Шмізінг Ріхард, Філліпс Ейфіон
МПК: C07D 453/00, A61P 25/18, A61K 31/444, C07D 451/04, A61P 25/24, A61P 25/34, A61P 25/14, A61P 25/00, A61P 43/00, A61P 1/04, A61P 25/22, A61K 31/40, A61P 25/28, A61P 25/16, A61K 31/439, A61K 31/4375, A61P 29/00, A61K 31/46
Мітки: психотичних, колії, n-азабіциклоаміду, рецептора, похідних, нікотинового, станів, стимуляція, розладів, лікування, також, корисною, застосування, профілактики
Формула / Реферат:
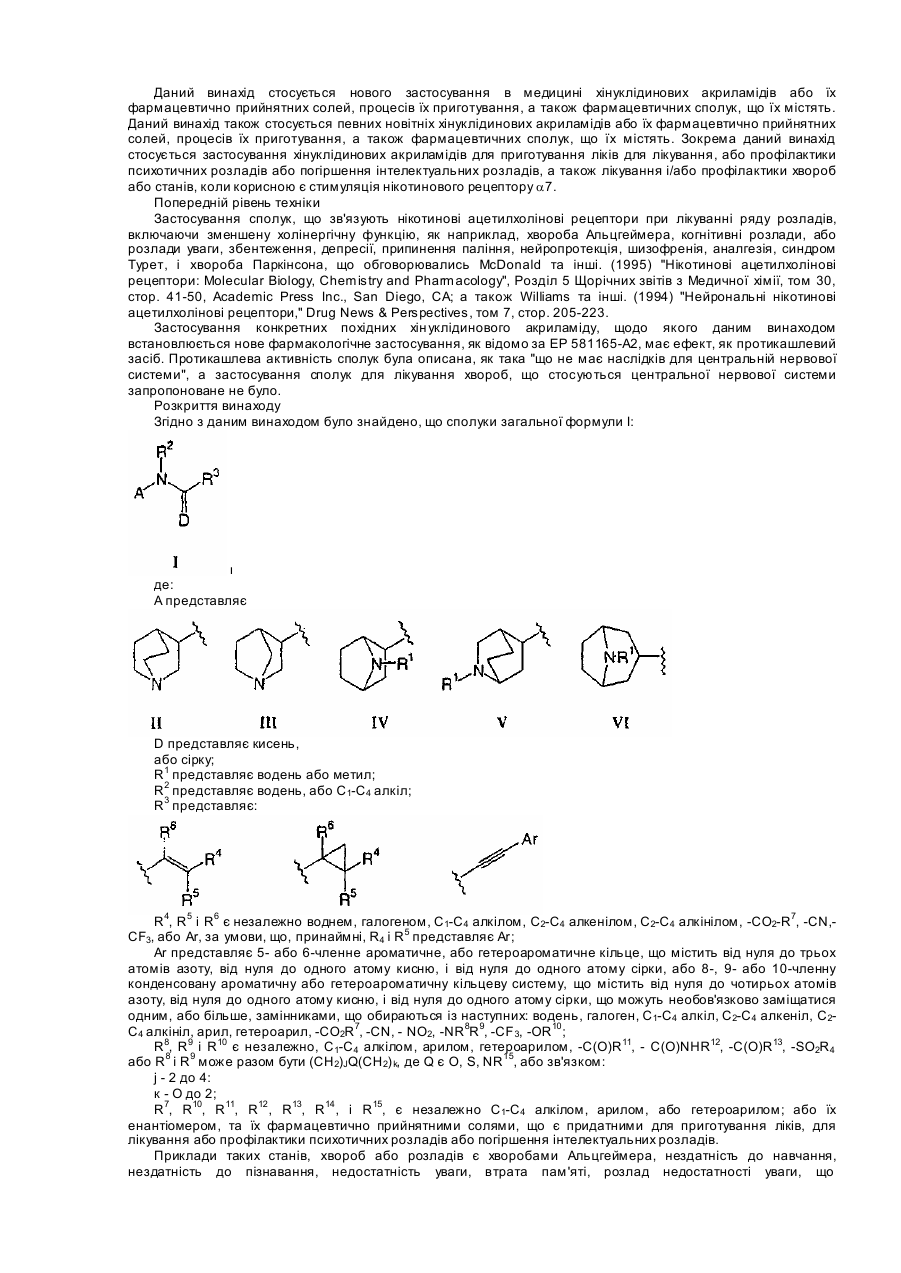
1. Застосування сполуки загальної формули І , IдеА являє собою: II III IV
Цитокіноподібний поліпептид-10 ссавців, полінуклеотид, що його кодує, антитіло та антиідіотипічне антитіло

Номер патенту: 73275
Опубліковано: 15.07.2005
Автори: Гросман Анжеліка, Хелдемен Бетті А., Конклін Деррел К.
МПК: C12N 15/19, A61P 35/00, A61P 11/00, C12N 15/09, A61P 43/00, C07K 14/52, A61P 37/02, A61P 1/02, A01K 67/027, A61K 48/00, C07K 16/24, A61K 38/00, C07K 16/42, A61P 17/00
Мітки: поліпептид-10, антитіло, антиідіотипічне, полінуклеотид, цитокіноподібний, ссавців, кодує
Формула / Реферат:
1. Виділений полінуклеотид, який кодує поліпептид, цей поліпептид щонайменше на 80% ідентичний одному чи більше з поліпептидів, які вибрані з групи, яка складається з ПОСЛІДОВН. №2, ПОСЛІДОВН. №4, ПОСЛІДОВН. №12, ПОСЛІДОВН. №13, ПОСЛІДОВН. №19, ПОСЛІДОВН. №20, ПОСЛІДОВН. №25, ПОСЛІДОВН. №26, ПОСЛІДОВН. №34 та ПОСЛІДОВН. №35, або виділений полінуклеотид, який кодує поліпептид, який вибраний з групи, яка складається з ПОСЛІДОВН. №14, ПОСЛІДОВН....
Вакцина, що гіперекспресує гомологічний антиген, та спосіб імунізації хребетного від бруцельозу

Номер патенту: 73469
Опубліковано: 15.08.2005
Автори: Корбейл Лінетт, Бойл Стівен М., Шуріг Герхардт Г., Вемулапаллі Рамеш, Шрірнаганатан Наммалвар, Краверо Сільвіо
МПК: A61K 39/02, A61K 39/00, A61K 39/10
Мітки: хребетного, гіперекспресує, спосіб, антиген, гомологічний, вакцина, імунізації, бруцельозу
Формула / Реферат:
1. Вакцина для імунізації, профілактики або лікування хребетного у випадку ризику або страждаючого від бруцельозу, яка відрізняється тим, що вказана вакцина включає атенуйований чи авірулентний штам іншої патогенної бактерії роду Brucella, причому зазначений штам надекспресує принаймні один гомологічний антиген, кодований принаймні одним геном з зазначеної бактерії, причому вказаний принаймні один антиген здатний індукувати захисну або...
Попередній патент: Розчин для ін’єкцій, який містить антагоніст lhrh
Наступний патент: Пристрій для виготовлення алмазів, вузол утримування зразка (варіанти) та спосіб виготовлення алмазів (варіанти)
Випадковий патент: Теплоізоляційна плита