Застосування фторвмісного аналога g-аміномасляної кислоти із замісником b-cf3-b-он, для збільшення початкової швидкості накопичення та зниження стимульованого деполяризацією плазматичної мембрани вивільнення g-
Номер патенту: 114259
Опубліковано: 10.03.2017
Автори: Борисова Тетяна Олександрівна, Шайтанова Олена Миколаївна, Кухар Валерій Павлович, Дударенко Марина Володимирівна, Герус Ігор Іванович, Позднякова Наталія Георгіївна






Формула / Реферат
Застосування фторвмісного аналога g-аміномасляної кислоти з замісником b-CF3-b-OH, для збільшення початкової швидкості накопичення та зниження стимульованого деполяризацією плазматичної мембрани вивільнення g-аміномасляної кислоти нервовими терміналями головного мозку щурів.
Текст
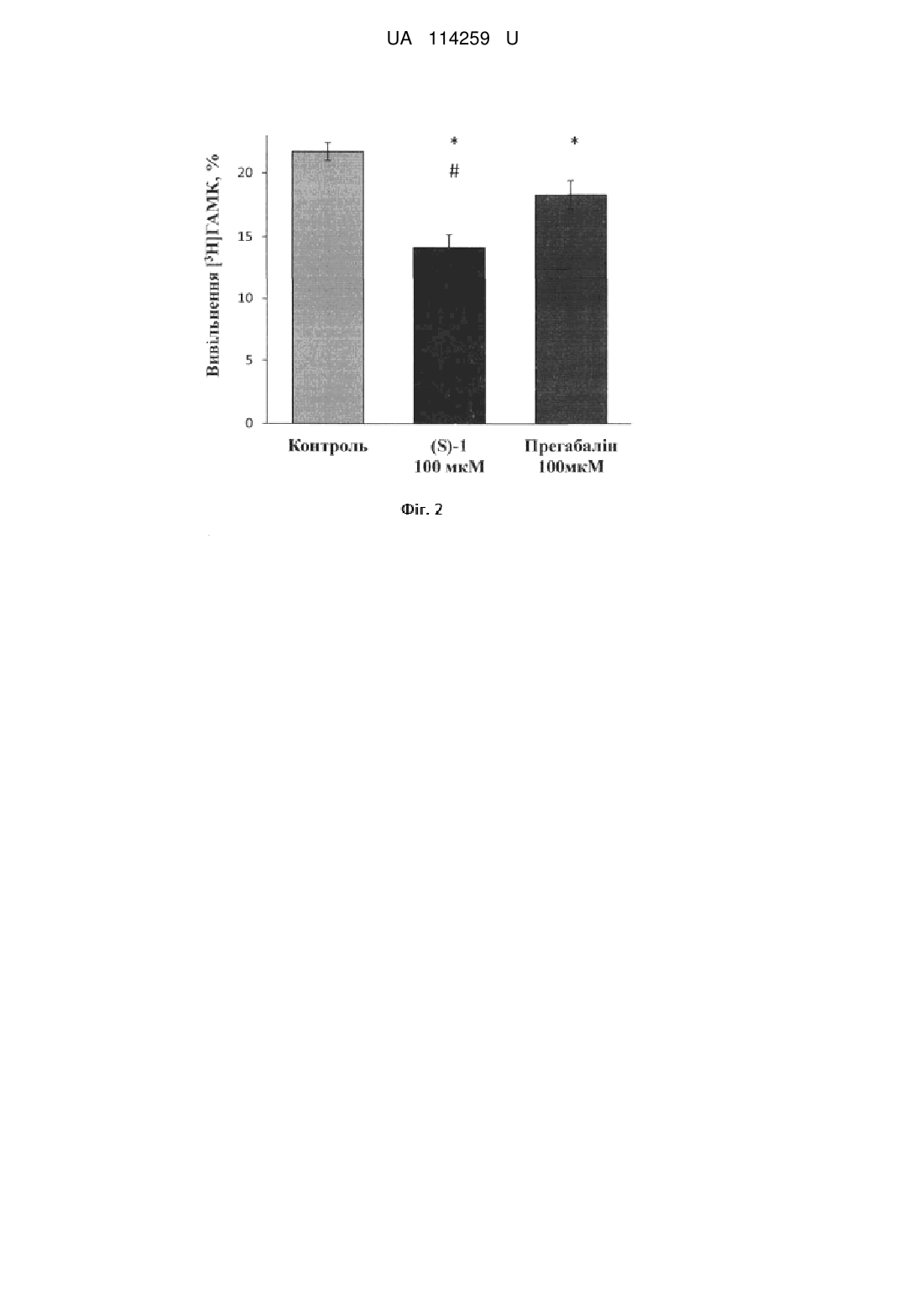
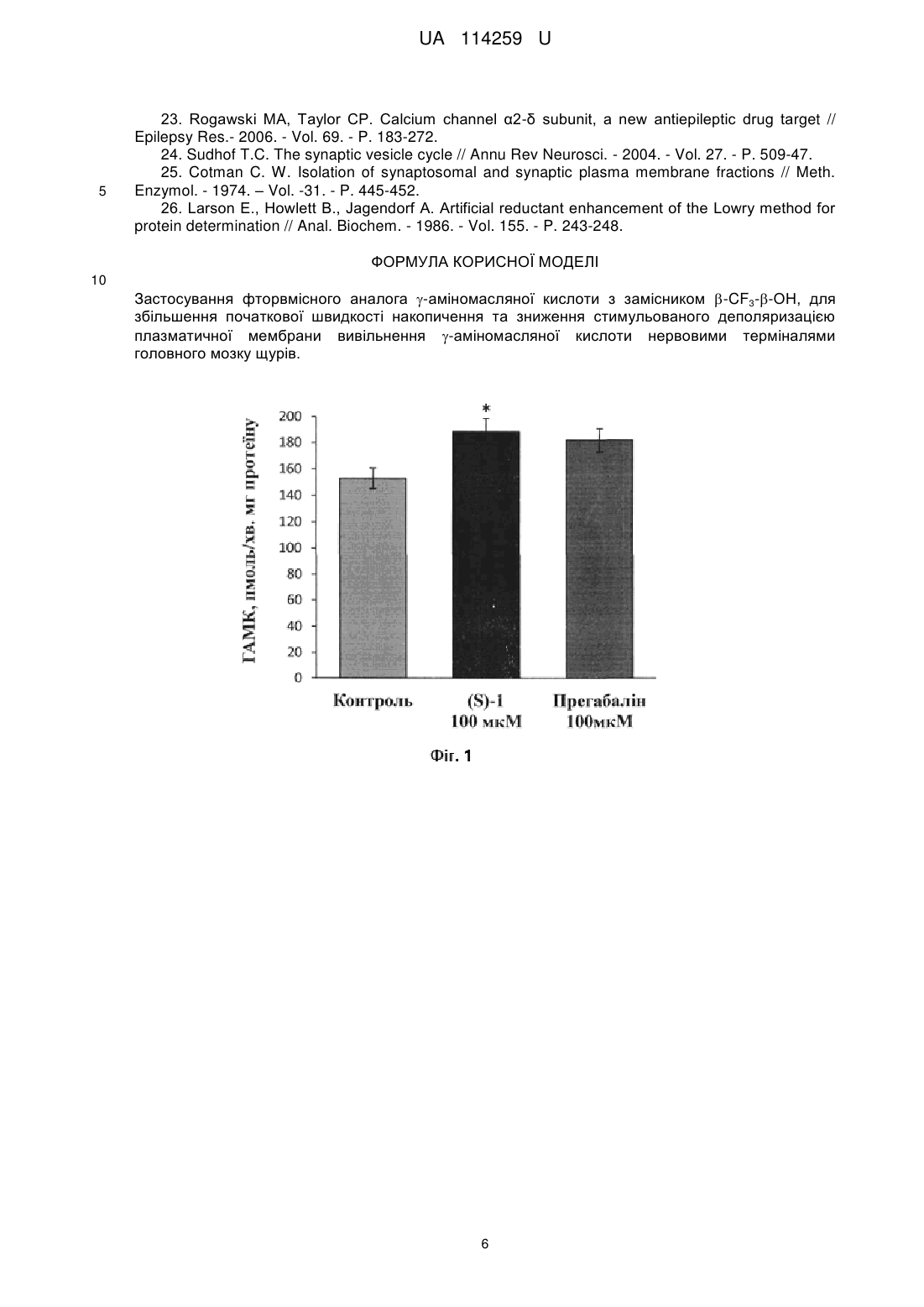
Реферат: Застосування фторвмісного аналога -аміномасляної кислоти з замісником -CF3--OH, для збільшення початкової швидкості накопичення та зниження стимульованого деполяризацією плазматичної мембрани вивільнення -аміномасляної кислоти нервовими терміналями головного мозку щурів. UA 114259 U (12) UA 114259 U UA 114259 U 5 10 15 20 25 30 35 40 45 Корисна модель належить до медицини, а саме до фармакології. Задачею корисної моделі є виявлення дії фторвмісного аналога γ-аміномасляної кислоти (ГАМК) із замісником β-CF3-β-OH ((S)-1) на початкову швидкість накопичення та стимульоване деполяризацією плазматичної мембрани вивільнення γ-аміномасляної кислоти нервовими терміналями головного мозку щурів (синаптосом). Вивчення дії фторвмісного аналога γ-аміномасляної кислоти із замісником β-CF3-β-OH ((S)1) на початкову швидкість накопичення та стимульоване деполяризацією плазматичної мембрани вивільнення γ-аміномасляної кислоти нервовими терміналями головного мозку щурів проведено у відділі нейрохімії Інституту біохімії ім. О.В. Палладіна НАН України. Фторвмісний аналог γ-аміномасляної кислоти із замісником β-CF3-β-OH ((S)-1) синтезовано у відділі тонкого органічного синтезу Інституту біоорганічної хімії та нафтохімії НАН України. Гальмівна ГАМКергічна система грає ключову роль у контролі нейрональної збудливості і підтримці балансу між процесами гальмування і збудження. Основним механізмом вивільнення γ-аміномасляної кислоти з пресинаптичних нервових терміналей є стимульований 2+ деполяризацією плазматичної мембрани Са -залежний екзоцитоз - злиття синаптичних везикул з плазматичною мембраною. ГАМК реалізує свою сигнальну функцію, діючи на іонотропні (ГАМК-А та ГАМК-С) та метаботропні (ГАМК-Б) рецептори, розташовані на поверхні клітин, що експресують їх. Тому концентрація ГАМК в позаклітинному середовищі визначає ступінь стимуляції рецепторів. Позаклітинний рівень ГАМК встановлюється як баланс між її транспортер-опосередкованим накопиченням і тонічним нестимульованим вивільненням. Позаклітинний рівень ГАМК може підтримуватися лише за допомогою транспортування її в клітину, тому що метаболізм цієї сполуки не відбувається в позаклітинному середовищі. Поглинання ГАМК нервовими клітинами забезпечується роботою високоафінних натрійзалежних транспортерів ГАМК, які локалізовані у плазматичній мембрані нейронів і гліальних клітин [1-3]. Сполуки, що впливають на процес гальмування, найчастіше входять до арсеналу протисудомних медичних препаратів, а дія ефективних антиконвульсантів найчастіше обумовлена їх впливом на функціонування транспортерів ГАМК, що має результатом підвищення рівня позаклітинної ГАМК. Серед інгібіторів ГАМК транспортерів тільки тіагабін селективний інгібітор транспортера GAT1 використовується в клінічній практиці при лікуванні судом, хоча інгібітори накопичення ГАМК є дуже перспективними агентами з їх потенційним застосуванням в лікуванні епілепсії, болю, наркоманії, порушення сну та ін. [3-9]. Одним з найбільш ефективних методичних підходів до розробки нових регуляторів внутрішньоклітинної/позаклітинної концентрації ГАМК та позаклітинного ГАМК гомеостазу в центральній нервовій системі є структурна модифікація ГАМК. Різні похідні ГАМК та її аналоги широко відомі в якості лікарських засобів, наприклад, вігабатрин [10], баклофен (Lioresal® and Baclon®) [11], габапентин (GBP, Neurontin®) [12], прегабалін (Lyrica®, β-і-Bu-GABA) [13, 14]. Фторвмісні аналоги природних речовин відіграють важливу роль у розробці нових біологічно активних сполук [15]. Пошук ефективних препаратів на основі молекули ГАМК показав, що більш ліпофільні аналоги здатні легко подолати гематоенцефалічний бар'єр [16]. Цей факт вказує на перспективу відповідних досліджень в галузі дизайну протисудомних препаратів [16, 17]. Введення гідроксильної групи в С-3 положення в молекулі ГАМК сприяє підвищенню антиконвульсивної активності речовини. Хіральність молекули також має важливу роль. Введення об'ємних ліпофільних замісників в β-положення структурного ядра ГАМК також є один з важливих чинників, що призводить до отримання нових ефективних регуляторів. У рамках даної роботи був синтезований фторвмісний аналог γ-аміномасляної кислоти з двома замісниками у β-положенні: β-CF3-β-OH ((S)-1) [18, 19], хімічну структуру та 3D-модель якої наведено нижче: 50 1 UA 114259 U . 5 10 15 20 25 30 35 40 45 Проведено порівняльний аналіз впливу фторвмісного аналога ГАМК з існуючим комерційним медичним препаратом Прегабаліном (Lyrica®), [(S)-(+)-3-ізобутил-ГАМК]. Синтезована сполука є структурним, але не функціональним аналогом ГАМК. При первинній оцінці біологічної дії (S)-1 в експериментах на мишах не було виявлено токсичного ефекту [18]. Прегабалін схвалений в США і Європі для додаткової терапії парціальних епілептичних нападів у дорослих, для лікування болю від діабетичної нейропатії або постгерпетичної невралгії у дорослих, фіброміалгії, а останнім часом і для лікування тривожних розладів [20, 21]. Механізми, що лежать в основі різних дій цього медичного препарату в мозку остаточно не з'ясовані, незважаючи на його широке клінічне застосування [22]. Фармакологічні властивості Прегабаліну в значній мірі обумовлені його високою афінністю до α2-δ-субодиниці потенціалзалежних кальцієвих каналів. Модуляція кальцієвого струму всередину нейрона під дією Прегабаліну призводить до зменшення вивільнення нейротрансмітерів з пресинаптичних нервових терміналей. Було висловлене також припущення, що Прегабалін модулює вивільнення або накопичення ГАМК нервовими закінченнями, зокрема перерозподіл транспортерів ГАМК GAT1 типу [22, 23]. Дослідження проводять на ізольованих нервових терміналях головного мозку синаптосомах, виділених з головного мозку щурів. Синаптосоми зберігають усі властивості інтактного нервового закінчення щодо забезпечення процесу передачі нервового імпульсу, а саме, здатність накопичувати та вивільнювати нейромедіатори, підтримувати мембранний потенціал та функціональний стан синаптичних везикул. Синаптосоми вважаються однією з найкращих систем для дослідження взаємозв'язку між структурою білків, їх біохімічними і клітинно-біологічними властивостями та фізіологічними функціями [24]. Беручи до уваги дані, що наведені вище, доцільним є аналіз впливу фторвмісного аналога ГАМК із замісником β-CF3-β-OH ((S)-1) на початкову швидкість накопичення та стимульоване 3 деполяризацією плазматичної мембрани вивільнення радіоактивно міченої [ Н]ГАМК нервовими терміналями головного мозку щурів (синаптосомами), що матиме значення для використання цієї сполуки у галузі фармакології для модуляції транспорту ГАМК у нервових терміналях головного мозку. В основу корисної моделі поставлено задачу застосування фторвмісного аналога ГАМК із замісником β-CF3-β-OH ((S)-1), що викликає збільшення початкової швидкості накопичення та зниження стимульованого деполяризацією плазматичної мембрани вивільнення радіоактивно 3 міченої [ Н]ГАМК нервовими терміналями головного мозку щурів. Фторвмісний аналог ГАМК із замісником β-CF3-β-OH ((S)-1) викликає збільшення початкової 3 швидкості накопичення ГАМК, яка реєструється як накопичення [ Н]ГАМК за першу хвилину процесу в синаптосомах. Вплив (S)-1 (100 мкМ) оцінюється після його попередньої інкубації з 3 синаптосомами протягом 20 хв. Початкова швидкість накопичення [ Н]ГАМК, яка в контролі складає 153,0±7,7 нмоль/хв. мг протеїну, в присутності (S)-1 підвищується на 23 % і складає 188,6±10,1 нмоль/хв. мг протеїну (Р0,05; t-тест Стьюдента, n=6). Прегабалін (100 мкМ), так само, як і (S)-1, після попередньої інкубації з синаптосомами 3 протягом 20 хв. підвищує початкову швидкість накопичення [ Н]ГАМК, яка дорівнює 182,1±8,9 нмоль/хв. мг протеїну. Фторвмісний аналог ГАМК із замісником β-CF3-β-OH ((S)-1) викликає зниження стимульованого калієвою деполяризацією (15 мМ КСl) плазматичної мембрани вивільнення 3 [ Н]ГАМК шляхом екзоцитозу з синаптосом головного мозку щурів. У контролі вивільнення 3 [ Н]ГАМК з синаптосом складає 21,7±0,7 %, а за присутності (S)-1 (100 мкМ) знижується на 35 % і складає 14,1±1,0 % від загальної кількості накопиченої синаптосомами мітки (Р0,05; т-тест Стьюдента, n=6). Прегабалін (100 мкМ) спричиняє незначне, але статистично достовірне 2 UA 114259 U 3 5 10 зниження вивільнення [ Н]ГАМК (на 15 %), яке складає 18,3±1,1 %, від загальної кількості накопиченої синаптосомами мітки (Р0,05; t-тест Стьюдента, n=6). (S)-1 виявився ефективнішим 3 у модуляції вивільнення [ Н]ГАМК за існуючий комерційний медичний препарат Прегабалін (Lyrica®). Цей факт є важливим для використання синтезованої сполуки у галузі фармакології для модуляції транспорту ГАМК у нервових терміналях головного мозку. Приклад 1. Синтез β-гідрокси-β-трифторметил-ГАМК ((S)-1. Схема синтезу включає такі основні стадії (які є звичайними для органічної хімії): приєднання триметилсилілціаніду до карбонільної групи доступного енону 2 з наступним відновленням ціаногрупи алюмогідридом літію, розкристалізацією діастереомерних винних солей, захистом аміногрупи (з утворенням фталімідної групи), послідовним гідролізом етоксивінільного фрагменту, окисненням альдегіду (S)-4, звільненням амінофункції та відділенням амінокислоти (S)-1 за допомогою іонообмінної смоли [18, 19]. Схема синтезу β-гідрокси-β-трифторметил-ГАМК 15 20 25 30 35 40 45 де: і - Me3SiCN, Et3N; ii-LiAlH4; iii-D-винна кислота, кристалізація; iv - фталевий ангідрид, піридин; v - окиснення за Джонсом, водн. 10 % НСl, 80 °C; vi - водн. 10 % НСl, СН3СООН, кип'ятіння; vii-Amberlite IRA-120. Приклад 2. Виділення синаптосом з головного мозку щурів. Щурів-самців лінії Wistar масою 100-120 г декапітують, великі півкулі головного мозку швидко переносять в розчин, що містить 0,32 М сахарози, 5 мМ Hepes-NaOH (pH 7,4) та 0,2 мМ етилендіамінтетраоцтової кислоти (ЕДТА). Усі операції проводять при 4 °C. Синаптосоми виділяють з гомогенату мозку диференційним центрифугуванням і центрифугуванням в градієнті щільності фіколлу, застосовуючи метод Котмана [25] у такій модифікації: розчин сахарози для приготування градієнту фіколлу містить 5 мМ Hepes-NaOH (pH 7,4) і 0,2 мМ ЕДТА. Синаптосомальну фракцію, отриману при центрифугуванні гомогенату головного мозку в градієнті щільності фіколлу, розводять 10 об'ємами 0,32 М сахарози, 5 мМ Hepes-NaOH (pH 7,4) і 0,2 мМ ЕДТА та центрифугують при 20000 g упродовж 20 хв. Отриманий осад повільно суспендують в 4 мл оксигенованого холодного середовища, що містить (в мМ): NaCl-126, КСl-5, MgCl2-1,4, NaH2PO4-1,0, HEPES-20, СаСl2-2, d-глюкозу - 10 (pH 7,4). При цьому кінцева концентрація протеїну становить 4 мг/мл. Синаптосоми використовують в експериментах упродовж 2-4 годин після отримання. Концентрацію протеїну визначають за методом Ларсона [26]. 3 Приклад 3. Визначення накопичення [ Н]ГАМК синаптосомами. 3 3 В дослідах з накопичення [ Н]ГАМК (γ-[2,3- Н(N)]-аміномасляна кислота; Perkin Elmer, США) синаптосомами стандартний сольовий розчин містить 100 мкМ амінооксіоцтової кислоти, інгібітора ГАМК-трансамінази, для запобігання утворення метаболітів ГАМК. Концентрація протеїну синаптосом в пробі дорівнює 200 мкг/мл, об'єм проби дорівнює 0,6 мл. Синаптосоми преінкубують 20 хв. при 37 °C з (S)-1 (100 мкМ) або Прегабаліном (100 мкМ), після чого ініціюють процес накопичення внесенням суміші ГАМК (1 мкМ ГАМК та 50 nМ - 0,2 мкКі/мл 3 UA 114259 U 3 5 10 15 20 25 30 35 40 45 50 55 60 [ Н]ГАМК). Через 1 хв. аліквоти (0,5 мл) фільтрують через GF/C фільтри (Sigma, США). Фільтри двічі промивають охолодженим стандартним сольовим розчином, висушують та вимірюють рівень радіоактивності у сцинтиляційній рідині OCS (Amersham, Великобританія) (1 мл) в лічильнику Delta 300 ("Tracor Analytic", США). 3 Приклад 4. Вивільнення [ Н]ГАМК із синаптосом головного мозку щурів. Синаптосоми (2 мг протеїну/мл) в оксигенованому стандартному сольовому розчині, який -7 містить 10 мкМ амінооксіоцтової кислоти, інкубують 5 хв. при 37 °C в присутності 5 × 10 М (0,1 3 Кі/мл) [ Н]ГАМК. Після охолодження на льоду, суспензію втричі розводять охолодженим сольовим розчином і центрифугують при 4000g 5 хв. Осад суспендують при температурі 4 °C і концентрації протеїну 1 мг/мл в сольовому розчині, який містить 10 мкМ амінооксіоцтової кислоти. Синаптосоми (120 мкл суспензії) преінкубують 10 хв. при 37 °C, потім 20 хв. з (S)-1 (100 мкМ) або Прегабаліном (100 мкМ). 3 Вивільнення [ Н]ГАМК викликають деполяризацією синаптосом 15 мМ КСl. Зразки інкубують 5 хв., після чого центрифугують у мікроцентрифузі Eppendorf (10,000×g, 20 s). Рівень 3 радіоактивності вивільненої [ Н]ГАМК в аліквотах супернатанту (90 мкл) вимірюють в лічильнику Delta 300 ("Tracor Analytic", США), з використанням сцинтиляційної рідини ACS (1 мл на 1 аліквоту). Вміст міченої ГАМК у супернатантах виражають у відсотках від загального вмісту 3 [ Н]ГАМК в синаптосомах. 3 Вивільнення [ Н]ГАМК з синаптосом, преінкубованих без стимулюючого агенту (15 мМ КСl) 3 приймається за базальне вивільнення. Стимульоване вивільнення [ Н]ГАМК обчислюється як 3 різниця між кількістю [ Н]ГАМК, що вивільнюється під впливом стимулюючого агента, і базальним вивільненням [8, 9]. Згідно з методологічним протоколом, дослідження транспортер-залежного накопичення та стимульованого деполяризацією плазматичної мембрани вивільнення ГАМК ізольованими нервовими терміналями головного мозку щурів проводиться з використанням радіоактивно 3 міченої [ Н]ГАМК (методи 2, 3, 4). Фторвмісний аналог ГАМК із замісником β-CF3-β-OH ((S)-1) викликає збільшення початкової швидкості накопичення та зниження стимульованого деполяризацією плазматичної мембрани 3 вивільнення [ Н]ГАМК нервовими терміналями головного мозку щурів. 3 Початкова швидкість накопичення, що реєструється як накопичення [ Н]ГАМК за першу хвилину процесу, в контролі складає153,0±7,7 нмоль/хв. мг протеїну; у присутності (S)-1 підвищується на 23 % і складає 188,6±10,1 нмоль/хв. мг протеїну (Р0,05; t-тест Стьюдента, n=6). Прегабалін (100 мкМ), так само, як і (S)-1, після попередньої інкубації з синаптосомами 3 протягом 20 хв. підвищує початкову швидкість накопичення [ Н]ГАМК, яка дорівнює 182,1±8,9 нмоль/хв. мг протеїну. При підвищенні концентрацій (S)-1 і Прегабаліну до 200 мкМ не 3 виявляється ніяких змін у початковій швидкості накопичення [ Н]ГАМК синаптосомами у порівнянні з тим ефектом, який спостерігається при аплікації 100 мкМ (Фіг. 1). 3 Вивільнення [ Н]ГАМК шляхом екзоцитозу обчислюють як різницю між рівнем стимульованого калієвою деполяризацією (15 мМ КСl) вивільненням нейромедіатора за 5 хв. і базальним вивільненням. Деполяризацію викликають, додаючи КСl до синаптосом, попередньо 3 2+ 3 навантажених [ Н]ГАМК в середовищі, що містить Са . У контролі вивільнення [ Н]ГАМК з синаптосом складає 21,7±0,7 %, а за присутності (S)-1 (100 мкМ) знижується на 35 % і складає 14,1±1,0 % від загальної кількості накопиченої синаптосомами мітки (*, Р0,05; t-тест Стьюдента, n=6). Прегабалін (100 мкМ) спричиняє незначне, але статистично достовірне 3 зниження вивільнення [ Н]ГАМК (на 15 %), яке складає 18,3±1,1 %, від загальної кількості накопиченої синаптосомами мітки (*, Р0,05; t-тест Стьюдента, n=6). (S)-1 виявляється 3 ефективнішим за Прегабалін (#, Р0,05 відносно вивільнення [ Н]ГАМК у присутності Прегабаліну.) (Фіг. 2). Таким чином, наведені результати експериментів підтверджують досягнення наступного технічного результату при здійсненні корисної моделі: - фторвмісний аналог ГАМК із замісником β-CF3-β-OH ((S)-1) викликає збільшення 3 початкової швидкості накопичення [ Н]ГАМК нервовими терміналями головного мозку щурів (синаптосомами). - фторвмісний аналог ГАМК із замісником β-CF3-β-OH ((S)-1) викликає зниження 3 стимульованого деполяризацією плазматичної мембрани вивільнення [ Н]ГАМК нервовими терміналями головного мозку щурів. - концентрація фторвмісного аналога ГАМК із замісником β-CF3-β-OH ((S)-1), за якої було зафіксоване статистично достовірне збільшення початкової швидкості накопичення на 23 % та 3 зниження стимульованого деполяризацією плазматичної мембрани вивільнення [ Н]ГАМК на 35 % становить 100 мкМ. 4 UA 114259 U 5 10 15 20 25 30 35 40 45 50 55 - фторвмісний аналог ГАМК із замісником β-CF3-β-OH ((S)-1) виявляється ефективнішим у 3 модуляції вивільнення [ Н]ГАМК за існуючий комерційний медичний препарат Прегабалін (Lyrica®). Джерела інформації: 1. Borisova T, Sivko R, Borysov A, Krisanova N. Diverse presynaptic mechanisms underlying methyl-beta-cyclodextrin-mediated changes in glutamate transport // Cell Моl Neurobiol. - 2010. - Vol. 30. - P. 1013-23. 2. Krisanova NV, Trikash IO, Borisova ТА. Synaptopathy under conditions of altered gravity: Changes in synaptic vesiclefusion and glutamate release // Neurochem Int. - 2009. - Vol. 55. - P. 72431. 3. Borden LA. GABA transporter heterogeneity: pharmacology and cellular localization // Neurochem Int. - 199. - Vol. 29. - P. 335-56. 4. Richerson GB, Wu Y. Dynamic equilibrium of neurotransmitter transporters: not just for reuptake anymore// J Neurophysiol. - 2003. - Vol. 90. - P. 1363-74. 5. Pozdnyakova N, Dudarenko M, Yatsenko L, Himmelreich N, Krupko O, Borisova T. Perinatal hypoxia: different effects of the inhibitors of GABA transporters GAT1 and GAT3 on the initial velocity 3 of [ H]GABA uptake by cortical, hippocampal, and thalamic nerve terminals // Croat Med J. - 2014. Vol. 55. - P. 250-58. doi: 10.3325/cmj.2014.55.250. 6. Dalby NO. Inhibition of gamma-aminobutyric acid uptake: anatomy, physiology and effects against epileptic seizures // Eur J Pharmacol. - 2003. - Vol. 479. - Vol. 127-37. 7. Richerson G. B, Wu Y. Role of GABA transporter in epilepsy // Adv Exp Med Biol. - 2004. - Vol. 548. - P. 76-91. 8. Pozdnyakova N, Yatsenko L, Parkhomenko N, Himmelreich N. Perinatal hypoxia induces longlasting increase in unstimulated GABA release in rat brain cortex and hippocampus. The protective effect of pyruvate // Neurochem Int. - 2011. - Vol. 58. - P. 14-21. 9. Yatsenko L, Pozdnyakova N, Dudarenko M, Himmelreich N. The dynamics of changes in hippocampal GABAergic system in rats exposed to early-life hypoxia-induced seizures // Neurosci Lett. - 2012. - Vol. 524. - P. 69-73. 10. Wei Z-Y, Knaus E.E. Asymmetric Synthesis of (S)-Vigabatrin. An approach using methionine as the chiral pool // Synlett 1993. - Vol. 4. - P. 295-6. DOI: 10.1055/S-1993-22438. 11. Felluga F, Gombac V, Pitacco G, Valentin E. A short and convenient chemoenzymatic synthesis of both enantiomers of 3-phenylGABA and 3-(4-chlorophenyl) GABA (Baclofen) // Tetrahedron: Asymmetry. - 2005. - Vol. 16. - P. 1341-45. DOI: 10.1016/j.tetasy.2005.02.019. 12. Bryans J.S, Wustrow D.J. 3-substituted GABA analogs with central nervous system activity: A review // Med Res Rev. - 1999. - Vol. 19. - P. 149-77. 13. Belliotti T.R, Capiris T, Ekhato I.V, Kinsora J.J, Field M.J, Heffner T.G, et al. Structure-activity relationships of pregabalin and analogues that target the α2-δ protein // J Med Chem. - 2005. - Vol. 48. - P. 2294-2307. 14. Silverman R.B. From basic science to blockbuster drug: The discovery of Lyrica // Angew Chem Int Ed. - 2008. - Vol. 47. - P. 3500-4. DOI: 10.1002/anie.200704280. 15. Begue J-P, Bonnet-Delpon D. Bioorganic and medicinal chemistry of fluorine // Wiley, 2008. 365 p. 16. Kirk K.L. Fluorine in medicinal chemistry: Recent therapeutic applications of fluorinated small molecules // J Fluor Chem 2006. - Vol. -127. - P. 1013-29. DOI: 10.1016/j.jfluchem.2006.06.007. 17. Filler R, Kobayashi Y, Yagupolskii LM. Organofluorine Compounds in Medicinal Chemistry and Biochemical Applications // Elsevier: Amsterdam, - 1993. 18. Shaitanova E.N, Gerus I.I, Metelitsa L.A, Charochkina L.L. Synthesis of β-polyfluoroalkyl containing γ-aminoacids and the preliminary study of the biological activity // Ukrainica Bioorganica Acta. - 2007. - -Vol. - 5. - P. 43-51. 19. Shaitanova E.N, Gerus I.I, Belik M.Yu, Kukhar V.P. Synthesis of optically active β-hydroxy-βpolyfluoromethyl GABAs // Tetrahedron: Asymmetry. - 2007. – Vol. 18. - P. 192-8. 20. Taylor C.P, Angelotti T, Fauman E. Pharmacology and mechanism of action of pregabalin: the calcium channel alpha2-delta (alpha2-delta) subunit as a target for antiepileptic drug discover // Epilepsy Res. - 2007. - Vol. 73. - P. 137-50. 21. Arnold L, Mease P, Silverman S. Pregabalin: an alpha2-delta (alpha2-delta) ligand for the management of fibromyalgia // Am J Manag Care. - 2010. - Vol. 16. - P. 138-43. 22. Whitworth T.L, Quick M.W. Upregulation of gamma-aminobutyric acid transporter expression: role of alkylated gamma-aminobutyric acid derivatives // Biochem Soc Trans. - 2001. - Vol. 29. - P. 736-41. 5 UA 114259 U 5 23. Rogawski MA, Taylor CP. Calcium channel α2-δ subunit, a new antiepileptic drug target // Epilepsy Res.- 2006. - Vol. 69. - P. 183-272. 24. Sudhof T.C. The synaptic vesicle cycle // Annu Rev Neurosci. - 2004. - Vol. 27. - P. 509-47. 25. Cotman C. W. Isolation of synaptosomal and synaptic plasma membrane fractions // Meth. Enzymol. - 1974. – Vol. -31. - P. 445-452. 26. Larson E., Howlett В., Jagendorf A. Artificial reductant enhancement of the Lowry method for protein determination // Anal. Biochem. - 1986. - Vol. 155. - P. 243-248. ФОРМУЛА КОРИСНОЇ МОДЕЛІ 10 Застосування фторвмісного аналога -аміномасляної кислоти з замісником -CF3--OH, для збільшення початкової швидкості накопичення та зниження стимульованого деполяризацією плазматичної мембрани вивільнення -аміномасляної кислоти нервовими терміналями головного мозку щурів. 6 UA 114259 U Комп’ютерна верстка Л. Ціхановська Державна служба інтелектуальної власності України, вул. Василя Липківського, 45, м. Київ, МСП, 03680, Україна ДП “Український інститут інтелектуальної власності”, вул. Глазунова, 1, м. Київ – 42, 01601 7
ДивитисяДодаткова інформація
Автори англійськоюBorysova Tetiana Oleksandrivna, Kukhar Valerii Pavlovych, Herus ihor Ivanovych
Автори російськоюБорисова Татьяна Александровна, Кухарь Валерий Павлович, Герус Игорь иванович
МПК / Мітки
МПК: A61K 31/02, A61K 49/00, A61K 31/325, G01N 33/483
Мітки: швидкості, стимульованого, збільшення, фторвмісного, мембрани, плазматичної, аналога, накопичення, деполяризацією, вивільнення, зниження, замісником, b-cf3-b-он, початкової, застосування, кислоти, g-аміномасляної
Код посилання
<a href="https://ua.patents.su/9-114259-zastosuvannya-ftorvmisnogo-analoga-g-aminomaslyano-kisloti-iz-zamisnikom-b-cf3-b-on-dlya-zbilshennya-pochatkovo-shvidkosti-nakopichennya-ta-znizhennya-stimulovanogo-depolyarizacieh.html" target="_blank" rel="follow" title="База патентів України">Застосування фторвмісного аналога g-аміномасляної кислоти із замісником b-cf3-b-он, для збільшення початкової швидкості накопичення та зниження стимульованого деполяризацією плазматичної мембрани вивільнення g-</a>
Застосування фторвмісного аналога g-аміномасляної кислоти із замісником b-cf2cf2h, для збільшення початкової швидкості накопичення та зниження стимульованого деполяризацією плазматичної мембрани вивільнення g-а

Номер патенту: 114258
Опубліковано: 10.03.2017
Автори: Шайтанова Олена Миколаївна, Герус Ігор Іванович, Борисова Тетяна Олександрівна, Кухар Валерій Павлович, Дударенко Марина Володимирівна, Позднякова Наталія Георгіївна
МПК: A61K 31/325, A61K 49/00, G01N 33/483, A61K 31/02
Мітки: плазматичної, збільшення, аналога, швидкості, накопичення, мембрани, вивільнення, замісником, g-аміномасляної, застосування, кислоти, b-cf2cf2h, стимульованого, деполяризацією, початкової, зниження, фторвмісного
Формула / Реферат:
Застосування фторвмісного аналога g-аміномасляної кислоти із замісником b-CF2CF2H, для збільшення початкової швидкості накопичення та зниження стимульованого деполяризацією плазматичної мембрани вивільнення g-аміномасляної кислоти нервовими терміналями головного мозку щурів.
Застосування наноалмазів, отриманих методом детонаційного синтезу, для зниження транспортер-залежного накопичення глутамату і гамма-аміномасляної кислоти (гамк) нервовими терміналями головного мозку щурів

Номер патенту: 114255
Опубліковано: 10.03.2017
Автори: Ільницька Галина Дмитриївна, Пастухов Артем Олегович, Дударенко Марина Володимирівна, Лещенко Ольга Володимирівна, Борисов Арсеній Андрійович, Борисова Тетяна Олександрівна, Галкін Максим Олексійович, Позднякова Наталія Георгіївна
МПК: C01B 32/15, C01B 32/25, G09B 23/28
Мітки: щурів, нервовими, гамк, гамма-аміномасляної, накопичення, синтезу, головного, отриманих, наноалмазів, застосування, глутамату, методом, мозку, терміналями, зниження, детонаційного, кислоти, транспортер-залежного
Формула / Реферат:
Застосування наноалмазів, що отримані методом детонаційного синтезу, для зниження транспортер-залежного накопичення глутамату і гамма-аміномасляної кислоти (ГАМК) нервовими терміналями головного мозку щурів.
Застосування наноалмазів, отриманих методом детонаційного синтезу, для збільшення позаклітинного рівня глутамату і гамма-аміномасляної кислоти в препараті нервових терміналей головного мозку щурів

Номер патенту: 114254
Опубліковано: 10.03.2017
Автори: Лещенко Ольга Володимирівна, Борисов Арсеній Андрійович, Дударенко Марина Володимирівна, Пастухов Артем Олегович, Галкін Максим Олексійович, Позднякова Наталія Георгіївна, Борисова Тетяна Олександрівна, Ільницька Галина Дмитриївна
МПК: C01B 32/15, C01B 32/25, G09B 23/28
Мітки: позаклітинного, методом, отриманих, терміналей, детонаційного, препарати, збільшення, рівня, кислоти, головного, застосування, щурів, наноалмазів, гамма-аміномасляної, нервових, глутамату, мозку, синтезу
Формула / Реферат:
Застосування наноалмазів, що отримані методом детонаційного синтезу, для збільшення позаклітинного рівня глутамату і гамма-аміномасляної кислоти в препараті нервових терміналей головного мозку щурів.
Застосування сполуки 3-децилоксикарбонілметил-4-метил-5-(2-гідроксіетил)тіазолій хлорид як сполуки, яка викликає деполяризацію плазматичної мембрани ізольованих нервових закінчень головного мозку щурів

Номер патенту: 45941
Опубліковано: 25.11.2009
Автори: Крисанова Наталія Валеріївна, Касаткіна Людмила Олександрівна, Вовк Андрій Іванович, Борисов Арсеній Андрійович, Романенко Олександр Вікторович, Остапченко Людмила Іванівна, Борисова Тетяна Олександрівна
МПК: C07D 277/00
Мітки: хлорид, плазматичної, щурів, деполяризацію, мозку, яка, застосування, закінчень, 3-децилоксикарбонілметил-4-метил-5-(2-гідроксіетил)тіазолій, мембрани, нервових, ізольованих, сполуки, викликає, головного
Формула / Реферат:
Застосування сполуки 3-децилоксикарбонілметил-4-метил-5-(2-гідроксіетил)тіазолій хлорид як сполуки, яка викликає деполяризацію плазматичної мембрани ізольованих нервових закінчень головного мозку щурів, загальної формули:.
Застосування наноалмазів, отриманих методом детонаційного синтезу, для деполяризації плазматичної мембрани нервових терміналей головного мозку щурів

Номер патенту: 114257
Опубліковано: 10.03.2017
Автори: Галкін Максим Олексійович, Борисов Арсеній Андрійович, Позднякова Наталія Георгіївна, Борисова Тетяна Олександрівна, Ільницька Галина Дмитриївна, Лещенко Ольга Володимирівна, Дударенко Марина Володимирівна, Пастухов Артем Олегович
МПК: C01B 32/25, G09B 23/28, C01B 32/15
Мітки: методом, отриманих, деполяризації, мозку, терміналей, щурів, мембрани, головного, детонаційного, нервових, синтезу, застосування, наноалмазів, плазматичної
Формула / Реферат:
Застосування наноалмазів, що отримані методом детонаційного синтезу, для деполяризації плазматичної мембрани нервових терміналей головного мозку щурів.
Наступний патент: Спосіб виготовлення автомобільного запасного колеса методом намотування
Випадковий патент: Циркуляційний клапан