Людський цитокін як ліганд зальфа рецептора і його використання
Номер патенту: 84387
Опубліковано: 27.10.2008
Автори: Спрічер Сінді А., Новак Джулія Е., Преснелл Скотт Р., Гросс Джейн А., Джонстон Жанет В., Хеммонд Анджела К., Фостер Дональд К., Нелсон Ендрю Дж., Діллон Стейсі Р., Холлі Річард Д.







Формула / Реферат
1. Виділений поліпептид, який має послідовність амінокислотних залишків, які щонайменше на 90 % ідентичні залишкам від 41 (Gln) до 148 (Ile), що показані послідовністю SEQ ID NО: 2, причому залишком у позиції 44 є Asp, залишком у позиції 47 є Asp, а залишком у позиції 135 є Glu, і який зв'язує рецептор, показаний послідовністю SEQ ID NO: 115.
2. Виділений поліпептид за п. 1, в якому амінокислотними залишками 71, 78, 122 та 125 є цистеїн.
3. Виділений поліпептид, який має послідовність амінокислотних залишків, які щонайменше на 95 % ідентичні залишкам від 41 (Gln) до 148 (Ilе), що показані послідовністю SEQ ID NО: 2, і який зв'язує рецептор, показаний послідовністю SEQ ID NO: 115.
4. Виділений поліпептид, який має послідовність амінокислотних залишків, які ідентичні залишкам від 41 (Gln) до 148 (Ilе), що показані послідовністю SEQ ID NО: 2, і який зв'язує рецептор, показаний послідовністю SEQ ID NО: 115.
5. Виділений поліпептид, який має послідовність амінокислотних залишків, які щонайменше на 90 % ідентичні залишкам від 32 (Gln) до 148 (Ilе), що показані послідовністю SEQ ID NО: 2.
6. Виділений поліпептид, який має послідовність амінокислотних залишків, які щонайменше на 95 % ідентичні залишкам від 32 (Gln) до 148 (Ilе), що показані послідовністю SEQ ID NО: 2.
7. Виділений полінуклеотид, який має послідовність нуклеїнової кислоти, яка кодує поліпептиди, охарактеризовані в пп. 1, 3, 4, 5 або 6.
8. Експресійний вектор, що містить такі оперативно зв'язані елементи:
промотер транскрипції;
сегмент ДНК, охарактеризований в п. 1, і
термінатор транскрипції.
9. Культивована клітина, що містить експресійний вектор, охарактеризований в п. 8.
10. Виділений поліпептид, що має послідовність амінокислотних залишків, яку показано послідовністю SEQ ID NO: 2 від залишку 32 (Gln) до залишку 162 (Ser) або послідовністю SEQ ID NO: 56 (миші) від залишку 23 (Gln) до залишку 146 (Ser).
11. Виділений поліпептид за п. 10, в якому послідовність амінокислотних залишків, показана послідовністю SEQ ID NО: 2, складається з залишків від 1 (Met) до 162 (Ser), a показана послідовністю SEQ ID NО: 56 (миші), складається з залишків від 1 (Met) до 146 (Ser).
12. Виділений поліпептид, що містить щонайменше 10 суміжних амінокислотних залишків послідовності SEQ ID NО: 2.
13. Виділений поліпептид за п. 12, в якому амінокислотні залишки вибрані з групи, яка складається з:
амінокислотних залишків 41-56 послідовності SEQ ID NО: 2; амінокислотних залишків 69-84 послідовності SEQ ID NO: 2; амінокислотних залишків 92-105 послідовності SEQ ID NO: 2 та амінокислотних залишків 135-148 послідовності SEQ ID NO: 2.
14. Конденсований білок, що включає щонайменше чотири поліпептиди, причому порядок поліпептидів від N-кінця до С-кінця такий:
перший поліпептид, що включає послідовність амінокислотних залишків від 41 до 56 послідовності SEQ ID NO: 2;
перший спейсер з амінокислотних залишків 6-27;
другий поліпептид, що включає послідовність амінокислотних залишків, вибрану з групи, що складається з:
залишків IL-2-спіралі В 53-75 послідовності SEQ ID NO: 111;
залишків IL-4-спіралі В 65-83 послідовності SEQ ID NO: 112;
залишків IL-15-спіралі В 84-101 послідовності SEQ ID NO: 113;
залишків GMCSF-спіралі В 72-81 послідовності SEQ ID NO: 114 та
амінокислотних залишків 69-84 послідовності SEQ ID NО: 2;
другий спейсер з амінокислотних залишків 5-11;
третій поліпептид, що включає послідовність амінокислотних залишків, вибраних з групи, що складається з:
залишків IL-2-спіралі С 87-99 послідовності SEQ ID NО: 111;
залишків IL-4-спіралі С 95-118 послідовності SEQ ID NО: 112;
залишків IL-15-спіралі С 107-119 послідовності SEQ ID NO: 113;
залишків GMCSF-спіралі С 91-102 послідовності SEQ ID NO: 114 та
амінокислотних залишків 92-105 послідовності SEQ ID NO: 2;
третій спейсер з амінокислотних залишків 3-29; та
четвертий поліпептид, що включає послідовність амінокислотних залишків, вибрану з групи, що складається з:
залишків IL-2-спіралі D 103-121 послідовності SEQ ID NО: 111;
залишків IL-15-спіралі D 134-157 послідовності SEQ ID NО: 112;
залишків IL-4-спіралі D 134-160 послідовності SEQ ID NО: 113;
залишків GMCSF-спіралі D 120-131 послідовності SEQ ID NО: 114 та
амінокислотних залишків 135-148 послідовності SEQ ID NО: 2.
15. Виділена полінуклеотидна молекула, що має послідовність нуклеотидів, які кодують поліпептид, охарактеризований в п. 1.
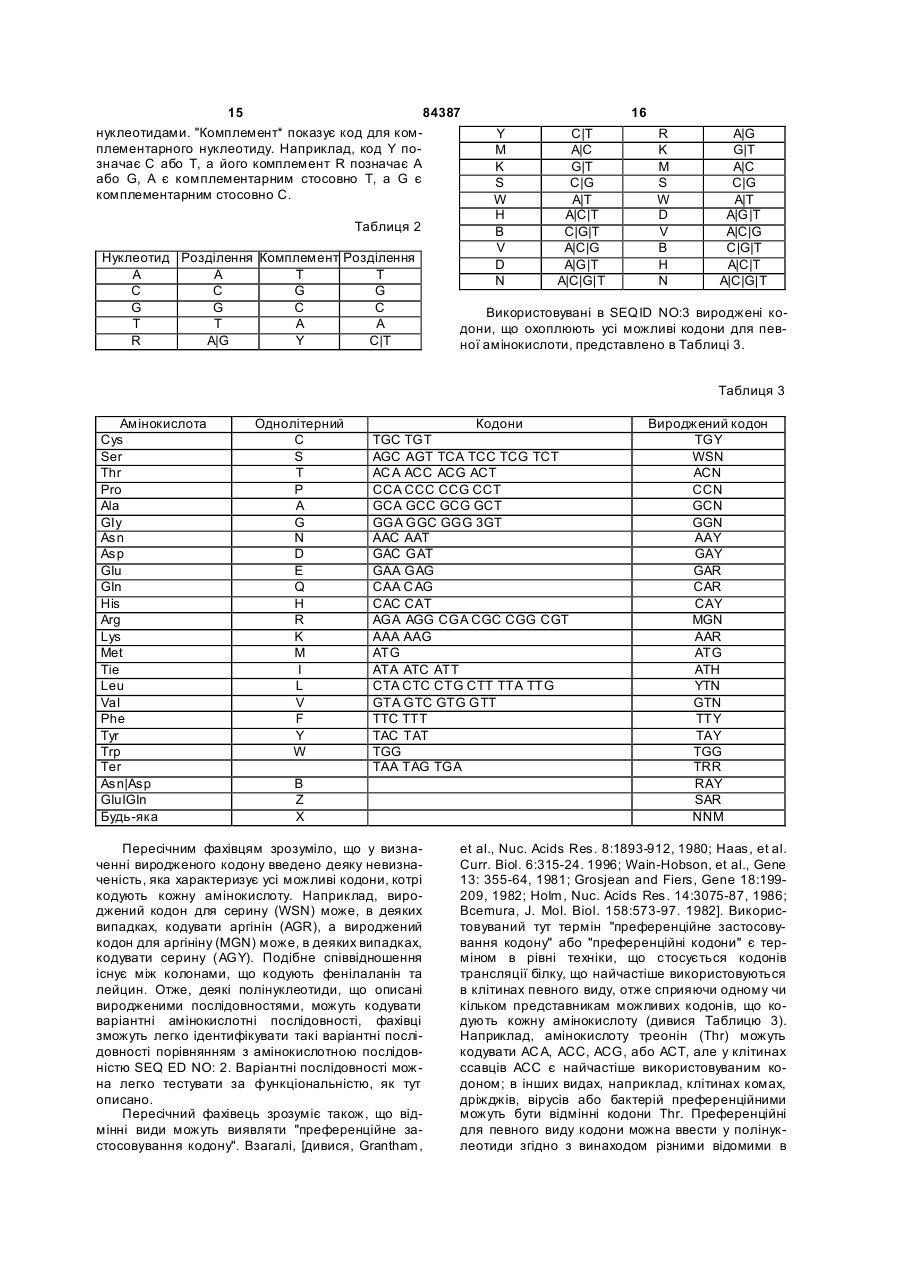
16. Виділена полінуклеотидна молекула за п. 15, в якій нуклеотиди показано послідовністю SEQ ID NО: 1 від нуклеотиду 167 до нуклеотиду 490 або послідовністю SEQ ID NО: 3 від нуклеотиду 121 до нуклеотиду 444.
17. Виділена полінуклеотидна молекула, що має послідовність нуклеотидів, які кодують поліпептид, охарактеризований в п. 13.
18. Виділена полінуклеотидна молекула, що має послідовність нуклеотидів, які кодують поліпептид, який є таким, що показаний послідовністю SEQ ID NО: 2 від залишку 32 до залишку 162 або послідовністю SEQ ID NО: 56 від залишку 23 до залишку 146.
19. Виділена полінуклеотидна молекула за п. 18, в якій нуклеотиди є такими, що показані послідовністю SEQ ID NО: 1 від нуклеотиду 140 до нуклеотиду 532 або послідовністю SEQ ID NO: 3 від нуклеотиду 94 до нуклеотиду 486.
20. Виділена полінуклеотидна молекула, що має послідовність нуклеотидів, які кодують поліпептид, який є таким, що показаний послідовністю SEQ ID NO: 2 від залишку 1 до залишку 162 або послідовністю SEQ ID NО: 56 від залишку 1 до залишку 146.
21. Виділена полінуклеотидна молекула за п. 20, в якій нуклеотиди є такими, що показані послідовністю SEQ ID NО: 1 від нуклеотиду 47 до нуклеотиду 532 або послідовністю SEQ ID NO: 3 від нуклеотиду 1 до нуклеотиду 486.
22. Спосіб виготовлення антитіла до поліпептиду ліганду zalphal 1, що включає:
щеплення тварини поліпептидом, вибраним з групи, що включає:
поліпептид, який складається з 9 до 131 амінокислоти, причому поліпептид є ідентичним суміжній послідовності з амінокислотних залишків SEQ ID NО: 2 від амінокислоти за номером 32 (Gln) до амінокислоти за номером 162 (Ser);
поліпептид, охарактеризований в п. 1;
поліпептид, що має амінокислотну послідовність з SEQ ID NO: 2 від амінокислоти за номером 41 (Gln) до амінокислоти за номером 148 (Ilе);
поліпептид, що має амінокислотну послідовність з SEQ ID NО: 2 від амінокислоти за номером 41 (Gln) до амінокислоти за номером 56 (Val);
поліпептид, що має амінокислотну послідовність з SEQ ID NO: 2 від амінокислоти за номером 69 (Thr) до амінокислоти за номером 84 (Leu);
поліпептид, що має амінокислотну послідовність з SEQ ID NО: 2 від амінокислоти за номером 92 (Asn) до амінокислоти за номером 105 (Arg);
поліпептид, що має амінокислотну послідовність з SEQ ID NО: 2 від амінокислоти за номером 135 (Glu) до амінокислоти за номером 148 (Ilе);
поліпептид, що має амінокислотну послідовність SEQ ID NО: 72;
поліпептид, що має амінокислотну послідовність SEQ ID NО: 73;
поліпептид, що має амінокислотну послідовність з SEQ ID NО: 2 від амінокислоти за номером 32 (Gln) до амінокислоти за номером 162 (Ser);
поліпептид, що має амінокислотну послідовність з SEQ ID NО: 2 від амінокислоти за номером 1 (Met) до амінокислоти за номером 162 (Ser);
поліпептид, що має амінокислотну послідовність з SEQ ID NO: 2 від амінокислоти за номером 1 14 до амінокислоти за номером 119;
поліпептид, що має амінокислотну послідовність з SEQ ID NО: 2 від амінокислоти за номером 101 до амінокислоти за номером 105;
поліпептид, що має амінокислотну послідовність з SEQ ID NО: 2 від амінокислоти за номером 126 до амінокислоти за номером 131;
поліпептид, що має амінокислотну послідовність з SEQ ID NО: 2 від амінокислоти за номером 113 до амінокислоти за номером 118;
поліпептид, що має амінокислотну послідовність з SEQ ID NО: 2 від амінокислоти за номером 158 до амінокислоти за номером 162; і
при цьому поліпептид викликає імунну реакцію у тварин для продукування антитіл; та виділення антитіла з тварини.
23. Антитіло, продуковане способом за п. 22, яке специфічно зв'язується з поліпептидом ліганду zalphal 1.
Текст
1. Виділений поліпептид, який має послідовність амінокислотних залишків, які щонайменше на 90 % ідентичні залишкам від 41 (Gln) до 148 (Ile), що показані послідовністю SEQ ID NО: 2 , причому залишком у позиції 44 є Asp, залишком у позиції 47 є Asp, а залишком у позиції 135 є Glu, і який зв'язує рецептор, показаний послідовністю SEQ ID NO: 115. 2. Виділений поліпептид за п. 1, в якому амінокислотними залишками 71, 78, 122 та 125 є цистеїн. 3. Виділений поліпептид, який має послідовність амінокислотних залишків, які щонайменше на 95 % ідентичні залишкам від 41 (Gln) до 148 (Ilе), що показані послідовністю SEQ ID NО: 2, і який зв'язує 2 (19) 1 3 84387 4 105 послідовності SEQ ID NO: 2 та амінокислотних послідовністю SEQ ID NО: 56 від залишку 23 до залишків 135-148 послідовності SEQ ID NO: 2. залишку 146. 14. Конденсований білок, що включає щонаймен19. Виділена полінуклеотидна молекула за п. 18, в ше чотири поліпептиди, причому порядок поліпепякій нуклеотиди є такими, що показані послідовністидів від N-кінця до С-кінця такий: тю SEQ ID NО: 1 від нуклеотиду 140 до нуклеотиду перший поліпептид, що включає послідовність 532 або послідовністю SEQ ID NO: 3 від нуклеотиду амінокислотних залишків від 41 до 56 послідовно94 до нуклеотиду 486. сті SEQ ID NO: 2; 20. Виділена полінуклеотидна молекула, що має перший спейсер з амінокислотних залишків 6-27; послідовність нуклеотидів, які кодують поліпептид, другий поліпептид, що включає послідовність аміякий є таким, що показаний послідовністю SEQ ID нокислотних залишків, вибрану з групи, що складаNO: 2 від залишку 1 до залишку 162 або послідовється з: ністю SEQ ID NО: 56 від залишку 1 до залишку залишків IL-2-спіралі В 53-75 послідо вності SEQ 146. ID NO: 111; 21. Виділена полінуклеотидна молекула за п. 20, в залишків IL-4-спіралі В 65-83 послідовності SEQ ID якій нуклеотиди є такими, що показані послідовнісNO: 112; тю SEQ ID NО: 1 від н уклеотиду 47 до нуклеотиду залишків IL -15-спіралі В 84-101 послідо вності 532 або послідовністю SEQ ID NO: 3 від нуклеотиду SEQ ID N O: 113; 1 до нуклеотиду 486. залишків GMCSF-спіралі В 72-81 послідовності 22. Спосіб виготовлення антитіла до поліпептиду SEQ ID NO: 114 та ліганду zalphal 1, що включає: амінокислотних залишків 69-84 послідовності SEQ щеплення тварини поліпептидом, вибраним з груID NО: 2; пи, що включає: другий спейсер з амінокислотних залишків 5-11; поліпептид, який складається з 9 до 131 амінокистретій поліпептид, що включає послідовність амінолоти, причому поліпептид є ідентичним суміжній кислотних залишків, вибраних з групи, що складапослідовності з амінокислотних залишків SEQ ID ється з: NО: 2 від амінокислоти за номером 32 (Gln) до залишків IL-2-спіралі С 87-99 послідовності SEQ амінокислоти за номером 162 (Ser); ID NО: 111; поліпептид, охарактеризований в п. 1; залишків IL-4-спіралі С 95-118 послідовності SEQ поліпептид, що має амінокислотну послідовність з ID NО: 112; SEQ ID NO: 2 від амінокислоти за номером 41 залишків IL-15-спіралі С 107-119 послідовності SEQ (Gln) до амінокислоти за номером 148 (Ilе); ID NO: 113; поліпептид, що має амінокислотну послідовність з залишків GMCSF-спіралі С 91-102 послідовності SEQ ID NО: 2 від амінокислоти за номером 41 SEQ ID NO: 114 та (Gln) до амінокислоти за номером 56 (Val); амінокислотних залишків 92-105 послідовності SEQ поліпептид, що має амінокислотну послідовність з ID NO: 2; SEQ ID NO: 2 від амінокислоти за номером 69 третій спейсер з амінокислотних залишків 3-29; та (Thr) до амінокислоти за номером 84 (Leu); четвертий поліпептид, що включає послідовність поліпептид, що має амінокислотну послідовність з амінокислотних залишків, вибрану з групи, що SEQ ID NО: 2 від амінокислоти за номером 92 складається з: (Asn) до амінокислоти за номером 105 (Arg); залишків IL-2-спіралі D 103-121 послідовності SEQ поліпептид, що має амінокислотну послідовність ID NО: 111; з SEQ ID NО: 2 від амінокислоти за номером 135 залишків IL-15-спіралі D 134-157 послідовності (Glu) до амінокислоти за номером 148 (Ilе); SEQ ID N О: 112; поліпептид, що має амінокислотну послідовність залишків IL-4-спіралі D 134-160 послідовності SEQ SEQ ID NО: 72; ID NО: 113; поліпептид, що має амінокислотну послідовність залишків GMCSF-спіралі D 120-131 послідовності SEQ ID NО: 73; SEQ ID NО: 114 та поліпептид, що має амінокислотну послідовність з амінокислотних залишків 135-148 послідовності SEQ ID NО: 2 від амінокислоти за номером 32 SEQ ID NО: 2. (Gln) до амінокислоти за номером 162 (Ser); 15. Виділена полінуклеотидна молекула, що має поліпептид, що має амінокислотну послідовність з послідовність нуклеотидів, які кодують поліпепSEQ ID NО: 2 від амінокислоти за номером 1 (Met) тид, о характеризований в п. 1. до амінокислоти за номером 162 (Ser); 16. Виділена полінуклеотидна молекула за п. 15, в поліпептид, що має амінокислотну послідовність з якій нуклеотиди показано послідовністю SEQ ID SEQ ID NO: 2 від амінокислоти за номером 1 14 до NО: 1 від нуклеотиду 167 до нуклеотиду 490 або амінокислоти за номером 119; послідовністю SEQ ID NО: 3 від нуклеотиду 121 до поліпептид, що має амінокислотну послідовність з нуклеотиду 444. SEQ ID NО: 2 від амінокислоти за номером 101 до 17. Виділена полінуклеотидна молекула, що має амінокислоти за номером 105; послідовність нуклеотидів, які кодують поліпептид, поліпептид, що має амінокислотну послідовність з охарактеризований в п. 13. SEQ ID NО: 2 від амінокислоти за номером 126 до 18. Виділена полінуклеотидна молекула, що має амінокислоти за номером 131; послідовність нуклеотидів, які кодують поліпепполіпептид, що має амінокислотну послідовність тид, який є таким, що показаний послідовністю з SEQ ID NО: 2 від амінокислоти за номером 113 SEQ ID NО: 2 від залишку 32 до залишку 162 або до амінокислоти за номером 118; 5 84387 6 поліпептид, що має амінокислотну послідовність з 23. Антитіло, продуковане способом за п. 22, яке SEQ ID NО: 2 від амінокислоти за номером 158 до специфічно зв'язується з поліпептидом ліганду амінокислоти за номером 162; і zalphal 1. при цьому поліпептид викликає імунну реакцію у тварин для продукування антитіл; та виділення антитіла з тварини. Проліферація та диференціація клітин багатоклітинних організмів регулюються гормонами та поліпептидними факторами росту. Ці здатні до дифузії молекули дозволяють клітинам спілкуватися одна з одною та діяти у згоді з утворенням клітин, тканин та органів та відновлення пошкодженої тканини. Приклади гормонів та факторів росту включають стероїдні гормони (наприклад, естроген, тестостерон), паратиреоїдний гормон, стимулюючий фолікули гормон, інтерлейкіни, похідний з тромбоцитів фактор росту (PDGF), епідермальний фактор росту (EGF), стимулюючий колонії гранулоцитів-макрофагів фактор (GM-CSF), еритропоетин (ЕРО) та кальцитонін. Гормони та фактори росту впливають на метаболізм клітин приєднанням до рецепторів. Рецептори можуть бути інтегрованими з мембраною білками, що зв'язані з шляхом сигналізації у клітині, як-то вторинні системи переносу. Іншими класами рецепторів є розчинні молекули, як-то фактори транскрипції. Цитокіни загалом стимулюють проліферацію або диференціацію клітин кровотворної лінії диференціювання або приймаючих участь у механізмах імунної та запальної реакцій організму. Прикладами цитокінів, що впливають на кровотворення, є еритропоетин (ЕРО), що стимулює розвиток червоних клітин крові, тромбопоетин (TPO), що стимулює розвиток клітин лінії диференціювання мегакаріоцитів; та стимулюючий колонії гранулоцитів-макрофагів фактор (G-CSF), що стимулює розвиток нейтрофілів. Ці цитокіни є корисними при відновленні нормальних рівнів клітин крові у пацієнтів, що потерпають від анемії, тромбоцитопенії та нейтропенії чи застосуванні хіміотерапії проти раку. Інтерлейкіни є родиною цитокінів, що опосередковують імунні реакції, включаючи запалення. Інтерлейкіни опосередковують множинність запальних патологій. Центральними в імунній реакції є Т-клітини, що продукують багато цитокінів та адаптивний імунітет до антигенів. Цитокіни, продуковані Т-клітинами, класифіковані як типи 1 та 2 [Kelso, A. Immun. Cell ВіоІ. 76:300-317, 1998]. Цитокіни типу 1 включають IL-2, IFN-g, LT-a, та є включеними у реакції запалення, імунітет до вір усів, імунітет до внутрішньоклітинних паразитів та відторгнення алотрансплантатів. Цитокіни типу 2 включають IL4, IL-5, IL-6, IL-10 та IL-13, та є включеними у гуморальні реакції, імунітет до глистів та алергічну реакцію. Цитокіни, що належать до типів 1 та 2 включають IL-3, GM-CSF та TNF-a. Є деякі свідоцтва, що означають, що тип 1 та тип 2, які продукують популяції Т-клітин, преференційно мігрують до запалених тканин різних типів. Зрілі Т-клітини можуть бути активованими, наприклад, антигеном чи іншими стимулами для продукування, наприклад, цитокінів, біохімічних сигнальних молекул або рецепторів, що крім того впливають на долю популяції T-клітин. В-клітини можуть бути активованими через рецептори на поверхні клітин, включаючи рецептор В-клітин та інші допоміжні молекули для виконання інших допоміжних функцій клітин, як-то продукування цитокінів. Нативні кілерні (NK) клітини мають спільний клітинний попередник з Т-клітинами та Вклітинами і грають роль в імунному нагляді. NKклітини, що включають до 15% лімфоцитів крові, не експресують антигенні рецептори, а тому не використовують MHC-розпізнавання, яке потрібне для приєднання до клітин-мішеней. NK-клітини є включеними у розпізнавання та знищення клітин деяких пухлин та інфікованих вірусами клітин. In vi vo, NK-клітини, можна думати, потребують активації, однак, in vitro, NK-клітини виявили здатність знищувати деякі типи клітин пухлин без активації. Продемонстрована in vivo активність родини цитокінів ілюструє величезний клінічний потенціал та необхідність у інших цитокінах, агоністах цитокінів, та антагоністах цитокінів. Представлений винахід спрямований на цю потребу забезпеченням нового цитокіну, що стимулює клітини кровотворної лінії диференціювання клітин, а також відповідних композицій та способів. Згідно з винаходом запропоновано такі поліпептиди для цього та інших використань, що фа хівці повинні зрозуміти з наведених тут пояснень Фігура є ілюстрацією складової укладки IL-2 людини, IL-15 людини, ліганду zalpha11 (SEQ ID NO: 2), IL-4 людини, IL-4 миші, GM-CSF людини та GM-CSF миші. До детального опису винаходу може бути корисним для розуміння визначити наступні терміни: Термін "афінна мітка" використано для позначення пептидного сегменту, що може бути приєднаним до поліпептиду для забезпечення очистки чи детектування поліпептиду, або забезпечення сайтів для приєднання поліпептиду до субстрату. В принципі, будь-який пептид або білок, для яких є антитіло або інший специфічний зв'язувальний засіб, можна використовувати як афінну мітку. Афінні мітки включають полігістидинову ділянку, протеїн A [Nilsson et al., EMBO J., 4:1075, 1985; Nilsson et al, Methods Enzymol. 198:3, 1991], глутатіон-8-трансферазу [Smith and Johnson, Gene 67:31, 1988], субстанцію Р, пептид FlagÔ [Hopp et al., Biotechnology 6: 1204-10, 1988; доступний від Eastman Kodak Co., New Haven, CT], зв'язуючий стрептавідин пептид, або інший антигенний епітоп 7 84387 8 або зв'язувальний домен. [Дивися взагалі, Ford et Термін "експресійний вектор" використано тут al., Protein Expression and Purification 2: 95-107, для позначення молекули ДНК, лінійної чи цикліч1991]. ДНК, що кодують афінні мітки, доступні від ної, що містить сегмент, який кодує потрібний покомерційних постачальників (наприклад, ліпептид, що оперативно зв'язаний з додатковими Pharmacia Biotech, Piscataway, NJ). сегментами, які забезпечують її транскрипцію. Такі Термін "алельна варіація" використано для пододаткові сегменти можуть включати промоторні значення будь-якого з двох чи більше альтернатита термінаторні послідовності та, як варіант, один вних форм гена, що займає той же самий хромочи більше початків реплікації, один чи більше присомний локус. Алельна варіація в природі датних для селекції маркерів, енхансер, поліадевідбувається внаслідок мутацій і може призводити нілаційний сигнал тощо. Експресійні вектори звидо фенотипового поліморфізму в популяції. Генні чайно походять з плазмід чи вірусної ДНК, або мутації можуть бути мовчазними (тобто без зміни можуть містити елементи обох. кодованого поліпептиду), або можуть кодувати Термін "виділений" при застосуванні до поліполіпептид, що має змінену амінокислотну послінуклеотиду, позначає, що полінуклео-тид видаледовність. Термін "алельний варіант" використано но з його природного генетичного середовища, і тут також для позначення кодованого алельним тому він позбавлений інших чужинних чи небажаваріантом гена білку. Включено також такий же них кодуючих послідовностей та існує у формі, що білок від того ж виду, який відрізняється від посипридатна для використання в генно-інженерній лальної амінокислотної послідовності внаслідок системі продукування білку. Такі виділені молекуалельної варіації. Алельна варіація стосується ли є тими, що видалено з їх природного середоприродно існуючих відмінностей серед осіб в гевища, і включають кДНК та геномні клони. Виділені нах, що кодують даний білок молекули ДНК згідно з винаходом позбавлені інТерміни "аміно-закінчення" та "карбоксиших генів, з якими вони звичайно спряжені, але закінчення" використано тут для позначення позиможуть включати природно існуючі 5' та 3' нетранцій у поліпептиді та білку. Коли дозволяє контекст, сльовані регіони, як-то промотери та термінатори. ці терміни використовують з посиланням на конкІдентифікація спряжених регіонів зрозуміла переретну послідовність або частину поліпептиду чи січним фахівцям [дивися, наприклад, Dynan and білку для позначення наближеності чи відносної Tijan, Nature 316:774-78. 1985]. позиції. Наприклад, певна послідовність, що роз"Виділений" поліпептид або білок є поліпептиташована карбокси-термінально стосовно посиладом або білком, що отримано у відмінному від льної послідовності у білку, розташована поблизу природного середовищі, як-то поза кров'ю та ткакарбокси-закінчення посилальної послідовності, ниною тварини. У кращій формі виділений поліпеале не обов'язково на карбокси-закінченні повного птид по суті позбавлений інших поліпептидів, зокбілку. рема інших поліпептидів тваринного походження. Термін "комплементарна/антиКраще забезпечувати поліпептиди у високоочищекомплементарна пара" використано тут для познаній формі, тобто з чистотою більше 95%, краще з чення неідентичних груп, що утворюють нековалечистотою більше 99%. При використанні в цьому нтно асоційовану стабільну пару в прийнятних контексті термін "виділений" не виключає присутумовах. Наприклад, біотин та авідин(або стрептаність такого поліпептиду в альтернативних фізичвідин) є прототиповими членами комплементарних формах, як-то димери чи альтернативно гліконої/анти-комплементарної пари. Інші приклади зиловані або дериватизовані форми. комплементарних/анти-комплементарних пар Термін "стосуються пухлин", при застосуванні включають пари рецептор/ліганд, пари антитідо клітин, означає клітини, що піддалися новій та ло/антиген (або гаптен чи епітоп), пари сенсоанормальній проліферації, зокрема у тканині, де вих/антисенсових полінуклеотидів тощо. Коли бапроліферація є неконтрольованою та прогресуюжана наступна дисоціація комплементарної/античою, що призводить до новоутворення. Клітини, комплементарної пари, комплементарна/антищо стосуються пухлин, можуть бути злоякісними, комплементарна пара переважно має споріднетобто інвазивними та метастатичними, або доброність до зв'язування менше 10-9M. якісними. Термін "комплемента полінуклеотидних молеТермін "оперативно зв'язаний" при застосукул" використано тут для позначення полінуклеованні до нуклеотидних сегментів свідчить, що сегтидної молекули, що має комплементарну посліменти розташовано так, що вони функціонують у довність основ та зворотну орієнтацію у порівнянні злагоді для їх цільового призначення, наприклад, з посилальною послідовністю, наприклад, послітранскрипція починається на промотері та прододовність 5' ATGCACGGG 3' є комплементарною вжується через кодуючий сегмент до термінатора. стосовно 5' CCCGTGCAT 3'. Термін "ортолог" позначає поліпептид або біТермін, "вироджена нуклеотидна послідовлок, що отримані від одного виду, який є функціоність" використано тут для позначення послідовнальним аналогом поліпептиду або білку від відності нуклеотидів, що включає один чи більше вимінних видів. Відмінності послідовностей серед роджених кодонів (у порівнянні з посилальною ортологів є результатом видоутворення. полінуклеотидною молекулою, що кодує поліпеп"Паралоги" є відмінними, але структурно спотид). Вироджені кодони включають відмінні трирідненими білками, що виробляються організмом. плети нуклеотидів, але кодують той же самий аміПаралоги є, можна думати, виникають внаслідок нокислотний залишок (тобто, триплети GAU та подвоєння гена. Наприклад, a-глобін, b-глобін, та GAC, кожний, кодують Asp). міоглобін є паралогами один одного. 9 84387 10 Полінуклеотид - одно- чи дволанцюговий поPDGF, рецептор гормону росту, рецептор IL-3, лімер дезоксирибонуклеотидних чи рибонуклеотирецептор GM-CSF, рецептор G-CSF, рецептор дних основ при зчитуванні від 5' до 3' кінця. Поліеритропоетину та рецептор IL-6). нуклеотиди включають РНК та ДНК і можуть бути Термін "секреторна сигнальна послідовність" виділеним з природних джерел, синтезованими in позначає послідовність ДНК, що кодує поліпептид vitro, або виготовленими з комбінації природних та ("секреторний пептид"), який як компонент більшосинтетичних молекул. Розмір полінуклеотидів виго поліпептиду спрямовує більший поліпептид черажають як число пар основ (скорочено "по"), нукрез секреторний шлях клітини, в якій його синтелеотидів ("нт"), або кілооснов ("ко"). Коли дозволяє зовано. Більший поліпептид звичайно контекст, останні два терміни можуть описувати розщеплюють для видалення секреторного пептиполінуклеотиди, що є одно- чи дволанцюговими. ду протягом транзиту через секреторний шлях. Коли термін застосовано до дволанцюгових молеТермін "сплайс-варіація" використано тут для кул, його використовують для позначення повної позначення альтернативних форм транскрибовадовжини і зрозуміло, що він еквівалентний терміну ної з гена РНК. Сплайс-варіація виникає природно "пара основ". Фахівцям зрозуміло, що два ланцюга внаслідок використання альтернативних сайтів дволанцюгового полінуклеотиду можуть трохи сплайсингу у транскрибованій молекулі РНК, або відрізнятися за довжиною і що їх кінці можуть бути рідше між окремо транскрибованими молекулами розташовані ступінчасто в результаті розщепленРНК, та може призводити у деяких мРНК, трансня ферментами, отже не всі нуклеотиди у дволанкрибованих з того ж самого гена. Сплайс-варіації цюговому полінуклеотиді можуть бути спареними. можуть кодувати поліпептиди, що мають змінену "Поліпептид" є полімером з'єднаних пептидамінокислотну послідовність. Термін сплайсними зв'язками амінокислотних залишків, що проваріації використано тут також для позначення дукований природно чи синтетично. Поліпептиди білку, кодованого сплайс-варіацією мРНК, трансменше приблизно 10 амінокислотних залишків крибованої з гена. звичайно позначають як "пептиди". Молекулярні маси та довжини полімерів, що Термін "промотер" використано тут для познавизначали приблизними аналітичними способами чення частини гена, що має послідовність ДНК, (наприклад, гель-електрофорезом) слід розуміти яка забезпечує приєднання РНК-полімерази та як приблизні величини. Коли таку величину вирапочаток транскрипції. Промоторна послідовність жають як "приблизно" X, визначена величина X, звичайно, але не завжди, знаходиться у 5' некодуслід розуміти, визначена з похибкою ±10%. ючих регіонах генів. Усі цитовані тут посилання вставлені як нотат"Білок" є макромолекулою, що містить один ки у своїй цілісності. поліпептидний ланцюг чи більше. Білок може Представлений винахід оснований, зокрема, включати також непептидні компоненти, як-то вугна відкритті нової послідовності ДНК, що кодує леводні групи. Вуглеводи та інші непептидні замісбілок, який має структуру цитокіну з чотирьохники можуть бути доданими до білку клітиною, в спіральним жмутом. У процесі клонування, досліякій продукується білок, та варіюють в залежності джень проліферації та вивчення приєднання, що від типу клітин. Білки позначено тут в термінах детально описано тут, полінуклеотидну послідовструктури їх амінокислотного скелету; такі замісниність, що кодує поліпептиди нового ліганду, було ки, як вуглеводні групи взагалі не уточнюють однак ідентифіковано як ліганд з високою специфічністю вони можуть бути присутніми. стосовно попередньо ізольованого рецептору Термін "рецептор" позначає асоційований з zalpha11. Цей поліпептидний ліганд, позначений як клітиною білок, або поліпептидний елемент такого ліганд zalpha11, виділяли з бібліотеки кДНК, ствобілку, що приєднує біоактивну молекулу ("ліганд") реної з активованих клітин периферійної крові люта опосередковує дію ліганду на клітину. Зв'язані з дини (hPBC), які селектували стосовно CDS. CDS мембраною рецептори характеризуються мультиє маркером клітинної поверхні, унікальним стосовпептидною структурою, що містить екстрацелюляно клітин лімфоїдного походження, особливо Трний ліганд-звязуючий домен та інтрацелюлярний клітин. ефекторний домен, що звичайно залучений у пеУ наведених далі прикладах лінія клітин, що є редачу сигналу. Приєднання ліганду до рецептору залежною від зв'язаного з шляхом обміну ізольопризводить до конформаційних змін в рецепторі, ваного рецептору zalpha11 для виживання та росщо призводить до взаємодії між ефекторним доту у відсутності інши х факторів росту, використоменом рецептору та іншими молекулами в клітині. вували для скринінгу на джерело кДНК, що кодує Ці взаємодії в свою чергу призводять до змін меліганд zalpha11. Кращою лінією залежних від фактаболізму клітини. Пов'язані з взаємодіями рецептору росту клітин, що використовували для транстор-ліганд метаболічні перетворення включають фекції та експресії рецептору zalpha11 була BaF3 транскрипцію генів, фосфорилування, дефосфо[Palacios та Steinmetz, Cell 41: 727-734, 1985; рилування, проліферацію клітин, посилення проMathey-Pre vot et al., Mol. Cell. Biol. 6: 4133-4135, дукування циклічного AMP, мобілізацію клітинного 1986]. Однак, придатними для цього є інші лінії кальцію, мобілізацію мембранних ліпідів, адгезію залежних від фактору росту клітин, як-то FDC-P1 клітин, гідроліз інозитних ліпідів та гідроліз фос[Hapel et al. Blood 64: 786-790, 1984], та МО7е [Kiss фоліпідів. Взагалі, рецептори можуть бути зв'язаet al., Leukemia 7: 235-240, 1993]. ними з мембраною, цитозольними або ядерними; Амінокислотна послідовність рецептору мономерними (наприклад, рецептор тироїдного zalphal 1 свідчить, що кодований рецептор налестимулюючого гормону, бета-адренергічний рецежить до класу І надродини рецепторів цитокіну, що птор) або мультимерними (наприклад, рецептор включає, але без обмеження, рецептори JL-2, IL-4, 11 84387 12 IL-5. IL-15, ЕРО, ТРО, GM-CSF та G-CSF [для ліотеку кДНК CD3+-відібраних, стимульованих огляду дивися, Cosman, "The Hematopoietin PMA та іономіцином первинних клітин периферійReceptor Superfamily" (Надродина рецепторів геної крові людини ділили на об'єднання, що містять матопоетину) у Cytokine 5(2): 95-106, 1993]. Реценабір молекул кДНК, та трансфектували у лінію птор zalpha11 повністю описано у загальнодоступклітин хазяїна, наприклад, клітин BHK 570 (ATCC ній [Патентній заявці РСТ №US99/22149]. Аналіз №10314). Трансфектовані клітини хазяїна культитканинного розподілу мРНК рецептору zalpha11 вували у середовищі, що не містить екзогенних виявив експресію у лімфатичному вузлі, лейкоцифакторів росту та збирали кондиційоване середотах периферійної крові (PBL), кістковому мозку та вище. Кондиційовані середовища досліджували на тимусі. Більш того, мРНК було багато у лінії клітин здатність стимулювати проліферацію клітин BaF3, Раджі (АТСС №CCL-86), похідних з лімфоми Буртрансфектованих рецептором zalpha11. Об'єднанкітта. Тканинний розподіл рецептору свідчить, що ня кДНК, що продукують кондиційоване середометою для передбачуваного ліганду zalpha11 є вище, що стимульоване рецепторними клітинами лінія диференціювання кровотворних клітин, зокBaF3/zalpha11, ідентифікували. Цю об'єднану пларема попередників лімфоїдних клітин та лімфоїдзмідну кДНК електропорували в E. coli. кДНК видіних клітин. Інші відомі цитокіни з чотирьохляли з одиничних колоній та трансфектували спіральним жмутом, що діють на лімфоїдні клітиокремо у клітини BHK 570. Позитивні клони іденни, включають IL-2, IL-4, IL-1, та IL-15. Для огляду тифікували позитивним результатом у проліферацитокінів з чотирьох-спіральним жмутом , [дивися, ційному дослідженні рецептору BaFS/zalpha11, а Nicola et al. Advances in Protein Chemistry 52:1-65. специфічність визначали нейтралізацією проліфе1999 та Kelso. A., Immunol. Cell Biol. 76:300-317, рації, використовуючи розчинний рецептор 1998]. zalpha11. Кондиційовані середовища (CM) з СD3+Позитивний клон виділяли, і аналіз послідоввідібраних, стимульованих РМА/іономіцином кліності виявив, що полінуклеотидна послідовність, тин периферійної крові людини підтримували ріст яка міститься в плазмідній ДНК, була новою. Секклітин BaF3, що експресували рецептор zalpha11, реторна сигнальна послідовність включає аміноа в іншому відношенні були залежними від IL-3. кислотні залишки від 1 (Met) до 31 (GIy), а цілком Кондиційовані середовища з клітин, що не були: 1) розвинений поліпептид включає амінокислотні стимульованими РМА/іономіцином; або не були: 2) залишки від 32 (GIn) до 162 (Ser) (як показано поСD3-відібраними (зі стимуляцією РМА/іономіцином слідовністю SEQ ID NO: 2). чи без неї), не підтримували ріст рецепторних кліЗагалом прогнозовано, що цитокіни мають тин BaF3/zalpha11. Контрольні експерименти проструктур у з чотирьох a-спіралей зі спіралями A, C демонстрували, що цю проліферативну активність та D, що є найважливішими у взаємодіях лігандне можна віднести до інших відомих факторів росрецептор, та є більш високо-збереженими серед ту, і що здатність таких кондиційованих середовищ членів родини. Відносячи до ліганду zalpha11 люстимулювати проліферацію експресуючих рецепдини амінокислотну послідовність, показану у SEQ тор zalpha11 клітин можна було б нейтралізувати ID NO:2, груп ування амінокислотних послідовносрозчинною формою рецептору. тей ліганду zalpha11 людини, IL-15 людини, IL-4 Проліферацію експресуючих рецептор людини, та GM-CSF людини, прогнозовано, що zalpha11 клітин BaF3, підданих дії CM, з СD3+спіраль А ліганду zalpha11 обмежена амінокислотвибраних, стимульованих РМА/іономіцином клітин ними залишками 41-56; спіраль В амінокислотнипериферійної крові людини ідентифікували візуами залишками 69-84; спіраль C амінокислотними льно дослідженням культур та/або проліферацією. залишками 92-105; а спіраль D амінокислотними Багато придатних досліджень проліферації відомі залишками 135-148; як показано послідовністю в рівні техніки та включають дослідження редукції SEQ ID NO: 2. Структурним аналізом визначено, барвнику, як-то аламар-блакитний (alamarBlueÔ) що петля А/В є довгою, петля В/С є короткою, а (Accu Med International. Inc. Westlake, Ohio), бромід петля C/D є паралельно довгою. Ця структура з 3-(4,5-диметилтіазол-2-іл)-2,5петлями призводить до спіральної будови уверхдифенілтетразоліуму [Mosman, J. Immunol. Meth. увер х-униз-униз. Цистеїнові залишки є абсолютно 65: 55-63, 1983]; гідроксид 3-(4,5-диметилтіазол-2збереженими між лігандом zalpha11 та IL-15, як іл)-5-3-карбоксиметоксифеніл-2Н-тетразоліуму; показано на фігурі. Цистеїнові залишки, що є збе2,3-біс(2-метокси-4-нітро-5-сульфофеніл)-5реженими між IL-15 та лігандом zalpha11, відпові[(феніламіно)карбоніл]-2Н-тетразоліуму; та хлорид дають амінокислотним залишкам 71, 78, 122 та ціанодитоліл-тетразоліуму (що є комерційно до125 of SEQ ID NO: 2. Збереження деяких з цистеїступними від Polisciences, Inc., Warrington, PA); нових залишків, як також виявлено, у IL-2. IL-4, дослідження мітогенезису, як-то вимір включення GM-CSF та ліганді zalpha11 відповідає амінокис3 H-тимідину; дослідження ексклюзії барвнику, вилотним залишкам 78 та 125 послідовності SEQ ID користовуючи, наприклад, нафталін-чорний або NO:2, якпоказано на Фігурі. Узгоджене розміщення трипан-блакитний; поглинання барвнику, викорисцистеїну є подальшим підтвердженням структури з товуючи діацетилфлуоресцеїн; та вивільнення чотирьох-спіральним жмутом . Також високозбехрому. [Дивися, загалом, Freshney, Culture of реженою у родині, що містить IL-15, IL-2, IL-4, GMAnimal Cells: A Manual of Basic Technique, 3rd ed., CSF та ліганд zalpha11 є послідовність Glu-PheWiley-Liss, 1994, що включені як посилання]. Leu, яку показано на послідовності SEQ ID NO: 2 Бібліотеку кДНК виготовили з СD3+на залишках 136-138, як на Фігурі. відібраних, стимульованих PMA та іономіцином Подальший аналіз ліганду zalpha11, що базупервинних клітин периферійної крові людини. Бібється на множинності групувань (як показано на 13 84387 14 Фігурі), прогнозує, що амінокислотні залишки 44, онально значущим. D спіралі ліганду zalpha11 лю47 та 135 (як показано послідовністю SEQ ID NO: дини та миші є також високозбереженими. 2) грають важливу роль у приєднанні ліганду Антагоністи рецептору zalpha11 можуть бути ствоzalpha11 до його спорідненого рецептору. Більш реними мутаціями у спіралі D ліганду zalpha11. того, передбачувана амінокислотна послідовність Вони можуть включати усікання білку від залишку ліганду zalpha11 миші виявляє 57% ідентичності Gln145 (SEQ ID NO: 2). або мутацію заміною Gln145, стосовно передбачуваного білку людини. На базі або lle148 (of SEQ ID NO: 2; що відповідає Туr124 у порівняння між послідовностями лігандів zalpha11 IL-4 людини) на залишки, як-то AIa або Asp. Будьлюдини та миші добре збережені залишки знайяка мутація, що розщеплює спіральну структуру шли у регіонах, прогнозованих як кодуючі альфаліганду zalpha11 може анулювати приєднання до спіралі А та D. Відповідні полінуклеотиди, що койого рецептору, а тому інгібує передачу сигналу. дують поліпептидні регіони, домени, сигнальні Цитокіни з чотирьох-спіральним жмутом також послідовності, залишки та послідовності ліганду класифікують за довжиною спіралей їх компоненzalpha11, що описано тут, показані послідовністю тів. "Довго-спіральна" форма цитокінів загалом SEQ ID NO:1. складається зі спіралі з 24-30 залишків та включає Для IL-4 та IL-2, що є високо-спорідненими з IL-6, війковий нейротропний фактор (CNTF), лейлігандом zalpha11, проведено детальний мутаційкемічний інгібітор-ний фактор (LIF) та гормон росту ний аналіз. Аналіз IL-2 миші [Zurawski et al, EMBO людини (hGH). "Коротко-спіральна" форма цитокіJ. 12:5113-5119, 1993] виявляє, що залишки у спінів загалом складається зі спіралі з 18-20 залишків ралях А та C є важливими для приєднання до ILта включає IL-2, IL-4 та GM-CSF. Ліганд zalpha11 є, можна вважати, новим членом коротко-спіральної 2Rb; критичними залишками є Asp34, Asn99, та форми групи цитокінів. Дослідження, використоАsn103. Бага то залишків у петлі А/В та спіралі В ILвуючи CNTF та IL-6, продемонструвало, що спі2 миші є важливими для приєднання IL-2Ra, при раль CNTF можна замінити на еквівалентну спітому, що тільки одиничний залишок, Gln141 у спіраль у IL-6, порівнюючи властивості приєднання ралі D, є суттєвим для приєднання до IL-2Ra. Подо CTNF з химерою. Отже, виявлено, що функціодібно, спіралі А та C є сайтами взаємодії між IL-4 нальні домени чотирьох-спіральних цитокінів вита IL-4Ra (структурно подібним до IL-2Ra), а зазначають на основі структурної гомології, незалелишки у спіралі D є суттєвими для взаємодії з ILжно від ідентичності послідовностей, і вони можуть 2Ra [Wang et al, Proc. Natl. Acad. Sci. USA 94:1657підтримувати функціональну цілісність химери 1662, 1997; Kruse et al, EMBQ J. 11:3237-3244, [Kallen et al., J. Biol. Chem. 274:11859-11867, 1999]. 1992]. Зокрема, мутація з заміною Туr124 на Asp у Відтак, спіральні домени ліганду zalpha11 будуть IL-4 людини створює антагоніст, який приєднуєтькорисними для виготовлення химерних конденсося до IL-4Ra, але не до IL-2Ra, а тому не може ваних молекул, особливо з іншою короткосигналізувати [Kruse et al. вище, 1992]. спіральною формою цитокінів для визначення та При тому, що спіраль А є відносно добре збемодулювання специфічності приєднання рецептореженою для лігандів zalpha11 людини та миші, ру. Особливий інтерес представляє конденсоваспіраль C є більш дивергентною. При тому, що ний білок, сконструйований зі спіраллю А та/або обидва види мають кислі амінокислоти домінуюспіраллю D, та конденсований білок, що комбінує чими у цьому регіоні, відмінності можна пояснюваспіральні та петельні домени від інших коротких ти специфічністю видів у взаємодії між лігандом форм цитокінів, як-то IL-2, IL-4, IL-15 та GM-CSF. zalpha11 та його рецептором zalpha11 типу "бета". Амінокислотні залишки, які містять спіралі А, В, C, Петля А/В та спіраль В ліганду zalpha11 є добре та D, та петлі А/В, В/С та C/D для ліганду zalpha11, збереженою для виду; і хоча ще не ідентифіковано IL-2, IL-4, IL-15 та GM-CSF показані у таблиці 1. субелементів рецептору, відповідних IL-2Ra, збереження у цьому регіоні означає, що він є функціТаблиця 1 Спіраль А А/В Петля Спіраль В В/С Петля Спіраль C C/D Петля Спіраль D Залишки ліганду zalpha11 Залишки IL-2 Залишки IL-4 Залишки IL-15 Залишки GM-CSF 41-56 57-68 69-84 85-91 92-105 106-134 135-148 SEQ ID NO: 2 36-46 29-43 45-68 30-44 47-52 44-64 69-83 45-71 53-75 65-83 84-101 72-81 76-86 84-94 102-106 82-90 87-99 95-118 107-119 91-102 100-102 119-133 120-133 103-119 103-121 134-151 134-160 120-131 SEQ ID NO: 111 SEQ ID NO: 112 SEQ ID NO: 113 SEQ ID NO: 114 Згідно з винаходом запропоновано полінуклеотидні молекули, включаючи молекули ДНК та РНК, що кодують розкриті тут поліпептиди ліганду zalpha11. Фахівці легко здогадаються, що з точки зору виродженості генетичного коду серед цих поліпептидних молекул можливі значні варіації послідовностей. SEQ ID NO:3 є виродженою послідовністю ДНК, яка охоплює усі ДНК, котрі кодують поліпептиди ліганду zalpha11 з послідовністю SEQ ID NO:2. Фахівці здогадаються, що виродже на послідовність SEQ ID NO:3 забезпечує також усі послідовності РНК, котрі кодують SEQ ID NO:2, заміною U на T. Отже, полінуклеотиди, що кодують поліпептиди ліганду zalpha11, що містять нуклеотиди від 1 або 97 до нуклеотиду 486 послідовності SEQ ID NO:3 та їх РНК-еквіваленти, розглянуто у представленому винаході. Таблиця 2 представляє використовувані в SEQ ID NO:3 однолітерні коди для позначення вироджених нуклеотидних позицій. "Розділення" є позначеними літерним кодом 15 84387 нуклеотидами. "Комплемент* показує код для комплементарного нуклеотиду. Наприклад, код Y позначає C або T, а його комплемент R позначає А або G, А є комплементарним стосовно T, a G є комплементарним стосовно C. Таблиця 2 Нуклеотид Розділення Комплемент Розділення А А T T C C G G G G C C T T А А R A|G Y С|Т 16 Y M K S W H В V D N C|T А|С G|T C|G А|Т А|С|Т C|G|T A|C|G A|G|T A|C|G|T R K M S W D V В H N A|G G|T А|С C|G A|T A|G|T A|C|G С|G|T А|С|Т A|C|G|T Використовувані в SEQID NO:3 вироджені кодони, що охоплюють усі можливі кодони для певної амінокислоти, представлено в Таблиці 3. Таблиця 3 Амінокислота Cys Ser Thr Pro Ala GIy Asn Asp GIu GIn His Arg Lys Met Tie Leu VaI Phe Tyr Trp Ter Asn|Asp GIuIGIn Будь-яка Однолітерний C S T P А G N D E Q H R K M I L V F Y W Кодони TGC TGT AGC AGT TCA TCC TCG TCT АС А ACC ACG ACT CCA CCC CCG CCT GCA GCC GCG GCT GGA GGC GGG 3GT AAC AAT GAC GAT GAA GAG CAA C AG CAC CAT AGA AGG CGA CGC CGG CGT AAA AAG ATG ATA ATC ATT СТА CTC CTG CTT TTA TTG GTA GTC GTG GTT TTC TTT TAC TAT TGG TAA TAG TGA B Z X Пересічним фахівцям зрозуміло, що у визначенні виродженого кодону введено деяку невизначеність, яка характеризує усі можливі кодони, котрі кодують кожну амінокислоту. Наприклад, вироджений кодон для серину (WSN) може, в деяких випадках, кодувати аргінін (AGR), а вироджений кодон для аргініну (MGN) може, в деяких випадках, кодувати серину (AGY). Подібне співвідношення існує між колонами, що кодують фенілаланін та лейцин. Отже, деякі полінуклеотиди, що описані виродженими послідовностями, можуть кодувати варіантні амінокислотні послідовності, фахівці зможуть легко ідентифікувати такі варіантні послідовності порівнянням з амінокислотною послідовністю SEQ ED NO: 2. Варіантні послідовності мож на легко тестувати за функціональністю, як тут описано. Пересічний фахівець зрозуміє також, що відмінні види можуть виявляти "преференційне застосовування кодону". Взагалі, [дивися, Grantham, Вироджений кодон TGY WSN ACN CCN GCN GGN AAY GAY GAR CAR CAY MGN AAR ATG ATH YTN GTN TTY TAY TGG TRR RAY SAR NNM et al., Nuc. Acids Res. 8:1893-912, 1980; Haas, et al. Curr. Biol. 6:315-24. 1996; Wain-Hobson, et al., Gene 13: 355-64, 1981; Grosjean and Fiers, Gene 18:199209, 1982; Holm, Nuc. Acids Res. 14:3075-87, 1986; Bcemura, J. MoI. Biol. 158:573-97. 1982]. Використовуваний тут термін "преференційне застосовування кодону" або "преференційні кодони" є терміном в рівні техніки, що стосується кодонів трансляції білку, що найчастіше використовуються в клітинах певного виду, отже сприяючи одному чи кільком представникам можливих кодонів, що кодують кожну амінокислоту (дивися Таблицю 3). Наприклад, амінокислоту треонін (Thr) можуть кодувати АС А, ACC, ACG, або ACT, але у клітинах ссавців ACC є найчастіше використовуваним кодоном; в інших видах, наприклад, клітинах комах, дріжджів, вірусів або бактерій преференційними можуть бути відмінні кодони Thr. Преференційні для певного виду кодони можна ввести у полінуклеотиди згідно з винаходом різними відомими в 17 84387 18 рівні техніки способами. Введення послідовностей гена ліганду zalpha11 у клітині змінюють вставкою преференційних кодонів у рекомбінантну ДНК, у локус ліганду zalpha11 констракту ДНК, що міснаприклад, посилює продукування білку здійснентить щонайменше націлюючу послідовність, регуням трансляції білку ефективнішою в певних типах ляторну послідовність, екзон та неспарений чи видах клітин. Тому, розкриті в SEQ ID NO:3 посплайс-донорний сайт. Націлююча послідовність є слідовності вироджених кодонів слугують темпланекодуючою послідовністю ліганду zalpha11, що тами для оптимізації експресії полінуклеотидів в дозволяє гомологічну рекомбінацію констракту з різних типах та видах клітин, що звичайно викориендогенним локусом ліганду zalpha11, причому стовують в рівні техніки і розкриті тут. Послідовнопослідовності у констракті стають оперативно сті, що містять преференційні кодони можна тесз'єднаними з кодуючою послідовністю ендогенного тувати та оптимізувати для експресії в різних ліганду zalpha11. Ha цьому шля ху, ендогенний видах, та тестувати на функціональність, як розпро-мотер ліганду zalpha11 може бути заміненим крито тут. або доповненим іншою регуляторною послідовнісЯк вищезазначено, виділені полінуклеотиди тю для забезпечення посиленої, тканинозгідно з винаходом включають ДНК та РНК. Споспецифічної, або інакше регульованої експресії. соби виділення ДНК та РНК добре відомі в рівні Згідно з винаходом запропоновано також анатехніки. Загалом РНК виділяють з тканини або логові поліпептиди та полінуклеотиди від іншого клітин, що продукують велику кількість ліганду виду (ортологи). Ці види включають, але без обzalpha11 РНК. Such тканини та клітини ідентифімеження, ссавців, птиць, амфібій, рептилій, риб, кують норзерн-блотингом [Thomas, Proc. Natl Acad. комах та інших членистоногих і нечленистоногих Sci. USA 77:5201, 1980], або скринінгом кондиційовидів. Особливо цікавими є поліпептиди ліганду ваного середовище з різних типів клітин на активzalpha11 від інших видів ссавців, включаючи поліність на цільових клітинах або тканині. Як тільки пептиди мишей, свиней, овець, корів, собак, кішок, активність або РНК, що продукують клітини або коней та інших приматів. Ортологи ліганду тканина, ідентифіковано, загальну РНК можна виzalpha11 людини можна клонувати, використовуюготовити, екстракцію гуанідином-НСІ з наступним чи інформацію та композиції згідно з представлевідділенням центрифугуванням в градієнтному ним винаходом у комбінації зі звичайними спосоCsCI [Chirgwin et al., Biochemistry 18:52-94, 1979]. бами клонування. Наприклад, кДНК можна + Полі-(А) -РНК виготовляють з загальної РНК, виклонувати, використовуючи мРНК, що отримано з користовуючи спосіб Авіва та Ледера [Proc.Natl. типів тканин або клітин, які експресують ліганд Acad. Sci. USA, 69:1408-12, 1972]. Комплементарzalpha11, як тут розкрито. Придатні джерела мРНК ну ДНК (кДНК) виготовляють з полі-(А)+-РНК, викоможна ідентифікувати зондуванням норзернристовуючи відомі способи. Полінуклеотиди, що блотів створеними з розкритих тут послідовностей кодують ліганд zalpha11, далі ідентифікують та зондами. Бібліотеку далі виготовляють з мРНК виділяють, наприклад, гібридизацією або PCR. позитивної лінії тканин або клітин. кДНК, що кодує Клон повної довжини, що кодує ліганд ліганд zalpha11, можна далі виділити багатьма zalpha11, можна отримати звичайним клонуспособами, як-то зондуванням повною чи часткованням. Клони комплементарної ДНК (кДНК) є вою кДНК людини або одним чи більше комплеккращими, хоча для деяких застосувань (напритами вироджених зондів на основі розкритих поклад, експресії у трансгенній тварині) може бути слідовностей. кДНК можна також клонувати, кращим використовувати геном-ний клон, або мовикористовуючи по-лімеразну ланцюгову реакцію, дифікувати клон кДНК для включення щонайменабо PCR [Mullis, Патент США №4683202], викорисше одного геномного інтрону. Способи виготовтовуючи праймери, створені з розкритої тут послілення кДНК та геномних клонів добре відомі та довності, що репрезентує ліганд zalpha11 людини. знані пересічними фахівцями і включають викориЗа додатковим способом, бібліотеку кДНК можна стання розкритої тут послідовності, або її частини використовува ти для трансформації або трансфедля зондування або праймуванням бібліотеки. кції клітини хазяїна, а експресію потрібної кДНК Експресійні бібліотеки можна зондувати антитіламожна детектувати антитілом до поліпептиду лігами до фрагментів рецептору zalpha11, або іншими нду zalpha11 дослідженням приєднання, або доспецифічно зв'язуючими партнерами. слідженням активності. Подібні способи можна Полінуклеотидні послідовності розкритого тут також застосовувати для виділення геномних клоліганду zalpha11 можна також використовувати як нів. зонди або праймери для клонування 5' некодуюПолінуклеотидна послідовність мишачого орчих регіонів гена ліганду zalpha11. З точки зору тологу ліганду zalpha11 ідентифіковано та показаспостереженої для ліганду zalpha11 тканиноно як SEQ ID NO: 55, а відповідну амінокислотну специфічної експресії цей регіон гена є очікуваним послідовність показано як SEQ ID NO: 56. Виявледля забезпечення кровотворно- та лімфоїдноно 62% ідентичності між послідовностями миші та специфічної експресії. Промотерні елементи з гена людини у регіоні зі 124 амінокислот, що відповідає ліганду zalpha11 тому можна було б використовузалишкам 30-153 у SEQ ID NO: 2 та залишкам 23вати для спрямування тканино-специфічної екс146 у SEQ ID NO: 56 ліганду zalpha11. Розвинена пресії гетерологічних генів у, наприклад, трансгенпослідовність для ліганду zalpha11 миші прогнозоній тварині або лікуємих генною терапією вано починається на His 18 (як показано послідовніпацієнтах. Клонування 5' фланкуючих послідовностю SEQ ID NO: 56), що відповідає His 23 (як покастей також полегшує продукування білку ліганду зано послідовністю SEQ ID NO: 2) у послідовності zalpha11 "активацією гена" як розкрито у [патенті людини. Оскільки усічена форма поліпептиду люСША №5641670]. Коротше, експресію ендогенного дини є активною, ймовірно, що еквівалентний по 19 84387 20 ліпептид ліганду zalpha11 миші (тобто без залишном; та 3) перехресно орієнтовану модель органіків His 18-Pro22 послідовності SEQ ID NO: 56) є акзмів, як-то мишей, що може допомагати при витивним також. Аналіз тканин виявив, що експресію значенні, яку функцію може мати певний ген. ліганду zalpha11 миші знаходять у яєчках, селезінЯк попередньо встановлено, генні залишки ліці та тимусі. ганду zalpha11 людини поблизу гена IL-2, що знаФахівці здогадаються, що розкрита як SEQ ID ходиться у регіоні хромосоми 4q, яку було виявлеNO: 1 послідовність репрезентує одиничний алель но як маючу зв'язок зі схильністю до запальних гена людини, та що очікуються алельні варіації та внутрішніх захворювань (IBD) (включаючи хворобу альтернативний сплайсинг. Алельні варіації цієї Крона (CD) та виразковий коліт) у деяких родинах послідовності можна клонувати зондуванням кДНК [Наmре et al. Am. J. Hum. Genet. 64: 808-816, 1999; чи геномних бібліотек від різних осіб чи тканин Cho et al. Proc. Natl. Acad. Sci. 95: 7502-7507, стандартними відомими в рівні техніки способами. 1998]. На додаток, ген рецептору zalpha11 картоАлельні варіанти показаної SEQ ID NO: 1 послідовано до 16рll, іншого геномного регіону, що асоцівності, включаючи ті, що мають мовчазні мутації, йований зі схильністю до CD [Hugot et al. Nature та ті, в яких мутації призводять до змін в амінокис379: 821-823, 1996: Ohmen et al., Hum. Mol. Genet.: лотній послідовності, включено до рамок винахо5: 1679-1683, 1996]. CD є хронічним запаленням ду, як і білки, що є алельними варіантами SEQ ID кишки, з частим системним залученням; при цьому NO: 2. Створені з альтернативно сплайсованих точна етіологія невідома, імунорегуляторна дисмРНК кДНК, які зберігають властивості поліпептифункція, що включає нестачу толерантності до ду лі ганду zalpha11, включено в рамки представзвичайних кишкових антигенів, є головним комполеного винаходу, як і поліпептиди, що кодовані нентом [для огляду, дивися (Braegger et al. Annals такими кДНК та мРНК. Алельні варіанти та сплайсAllergy 72: 135-141, 1994; Sartor. Am. J. варіанти цих послідовностей можна клонувати Gastroenterol. 92: 5S-11S, 1997)]. Кількома дослізондуванням кДНК чи геномних бібліотек від різних дженнями виявлено ненормальну активність NK у осіб чи тканин стандартними відомими в рівні техпацієнтів з CD [дивися, наприклад, (Egawa et. al., ніки способами. J. Clin. Lab. Immunol. 20: 187-192, 1986; AparicioГен ліганду zalpha11 картовано до структурноPages et al. J. Clin. Lab. Immunol. 29: 119-124, 1989; го маркеру IL-2 SHGC-12342, позиціонуючи ліганд van ToI et al., Scand. J. Gastroenterol. 27: 999-1005, zalpha11 приблизно на 180 ко від маркеру EL-2. 1992)], а також було показано дефектне утворення Використання оточуючих маркери позицій гена В-клітин пам'яті [Brogan et al., J. Clin. Lab. Immunol. ліганду zalpha11 у регіоні маркеру 4q27 на інтегро24: 69-74, 1987]. Оскільки ліганд zalpha11 грає ваній карті LDB хромосоми 4 (The Genetic Location роль в імунній регуляції, та оскільки гени рецептоDatabase (База даних розташування генів)), ру та ліганду лежать у регіонах схильності до CD, University of Southhampton,). Згідно з винаходом рецептор та ліганд є кандидатними генами генетизапропоновано також реагенти, що знайдуть викочної схильності до хвороби Крона. ристання при діагностиці. Наприклад, ген ліганду Визначення залучення рецептору zalpha11 zalpha11, зонд, що містить ДНК або РНК ліганду та/або ліганду zalpha11 у патології IBD можна виzalpha11 або їх субпослідовність, можна викорисконати кількома способами. Секвенсуванням екзотовувати для визначення, чи присутній ген ліганду нів з геномної ДНК можна виявити мутації кодуzalpha11 у хромосомі людини, як-то хромосомі 4, вання (включаючи мутації хибно-сенсові, або чи відбувається мутація гена. На базі анотації несенсові та зміщення рамки), як-то секвенсуванфрагменту геномної ДНК людини, що містить часням кДНК. Додатковою перевагою секвенсування з тину геномної ДНК ліганду zalpha11 (Genbank геномної ДНК є те, що сплайс-з'єднання також є Accession №АС007458), ліганд zalpha11 локалізунаявними у секвенсованих фрагментах, і можна ють у регіоні 4q27 хромосоми 4. Виявлювані хровиявити аномалії сплайсингу, які можна було б не мосомні аберації на локусі гена ліганду zalpha11 виявити у зразках кДНК, якщо наприклад, хибно включають, але без обмеження, анеуплоїдію, змісплайсовані РНК швидко вироджувалися. Визнани числа копій гена, втрату гетерогенності (LOH), чено геномну структуру ліганду zalpha11. Інші спотранслокації, вставки, делеції, зміни рестрикційних соби аналізу ліганду та рецептору zalpha11 у пацісайтів та перегрупування. Такі аберації можна деєнтів з IBD включають: (1) оцінку продукування тектувати , використовуючи полінуклеотиди предліганду з активованих Т-клітини від пацієнтів у поставленого винаходу, застосуванням техніки морівнянні з нормальними контрольними особами лекулярної генетики, як-то аналіз поліморфізму (тобто біодослідженням); (2) гібридизацію in situ довжини рестрикційних фрагментів (RFLP), аналіз РНК рецептору zalpha11 або ліганду zalpha11 з коротких тандемних повторів (STR), застосовуючи сегментами запаленого кишечнику від пацієнтів з техніку PCR та інши х аналізів зчеплення, що відоIBD у порівнянні з подібними сегментами від нормі в рівні техніки [Sambrook et al., вище; Ausubel et. мальних контрольних осіб; (3) імуногістохімію на al., вище; Marian, Chest 108: 255-65, 1995]. сегментах запаленого кишечнику від пацієнтів з Точні відомості про позицію гена можуть бути IBD у порівнянні з подібними сегментами від норкорисними для багатьох цілей, включаючи: 1) вимальних контрольних осіб; та (4) оцінку швидкості значення, чи є послідовність частиною існуючого реагування периферійних В-клітини пацієнтів з контигу, та отримання додаткових оточуючи х генелігандом zalpha11, як вимірювали дослідженням тичних послідовностей у різних формах, як-то кломітогенезису. нів YAC, ВАС або кДНК; T) забезпечення можливоДіагностика могла би допомогти лікарям при го кандидатного гена спадкового захворювання, визначенні типу захворювання та прийнятної теякий виявляє зв'язок з тим же хромосомним регіорапії, або могла би допомогти у генетичних висно 21 84387 22 вках. Як такі, антитіла проти ліганду zalpha11, пофрагменту, довжині продуктів PCR, довжині полінуклеотиди, та поліпептиди згідно з винаходом вторюваних послідовностей на генному локусі ліможна використовувати для визначення поліпепганду zalpha11t, тощо, є показником генної аноматиду, мРНК ліганду zalpha11, або антитіл проти лії, генетичної аберації, або алельної відмінності у ліганду zalpha11, отже вони можуть слугувати мапорівнянні з нормальним контролем. Контролі моркерами та безпосередньо використовува тися для жуть бути від неуражених членів родини, або невизначення генетичних захворювань або раку, як споріднених осіб, залежно від тесту та придатності тут описано, використовуючи відомі в рівні техніки зразків. Генетичні зразки для використання в рамта описані тут способи. Далі, полінуклеотидні зонках представленого винаходу включають геномну ди ліганду zalpha11 можна використовувати для ДНК, мРНК, та кДНК, виділені з будь-якої тканини визначення аномалії, що включає хромосому 4q27, або іншого біологічного зразку від пацієнту, як-то, як тут описано. Ці аномалії можна пов'язувати з але без обмеження, крові, слини, сперми, ембріозахворюваннями людини, або генезисом пухлин, нальних клітини, амніотичної рідини тощо. Полінуспонтанними абортами або іншими генетичними клеотидний зонд або праймер може бути РНК або розладами. Отже, полінуклеотидні зонди ліганду ДНК, та включатиме частину послідовності SEQ ID zalpha11 можна використовувати для визначення NO: 1, комплемент послідовності SEQ ID NO: 1, аномалій або генотипів, що пов'язані з цими дефеабо їх РНК-еквіваленти. Такі способи аналізу виктами. явлення генетичного зчіплювання стосовно феноЯк обговорено вище, дефекти у гені ліганду типів захворювань людини добре відомі в рівні zalpha11 самі можуть бути результатом спадковотехніки. Для посилання на способи на основі PCR го захворювання людини. Молекули представлеу діагностиці [дивися, загалом, Mathew (ed.), ного винаходу, як-то поліпептиди, антагоністи, Protocols in Human Molecular Genetics (Humana агоністи, полінуклеотиди та антитіла представлеPress, Inc. 1991), White (ed.), PCR Protocols: ного винаходу могли б допомогти при визначенні, Current Methods and Applications (Humana Press, діагностичному попередженні та лікуванні захвоInc. 1993), Cotter (ed.). Molecular Diagnosis of рювань, пов'язаних з дефектом гена ліганду Cancer (Humana Press, Inc. 1996), Hanausek та zalpha11tic. Ha додаток, полінуклеотидні зонди Walaszek (eds.), Tumor Marker Protocols (Humana ліганду zalpha11 можна використовувати для виPress, Inc. 1998), Lo (ed.). Clinical Applications of значення алельних відмінностей між хворими або PCR (Humana Press, Inc. 1998), та Meltzer (ed.), здоровими особами на хромосомному локусі лігаPCR in Bioanalisis (Humana Press, Inc. 1998))]. нду zalpha11. Як таку, послідовності ліганду Мутації, пов'язані з локусом ліганду zalpha11, zalpha11 можна використовува ти як діагностичні у можна детектувати, використовуючи молекули судово-діагностичному профілюванні ДНК. нуклеїнової кислоти представленого винаходу Загалом способи діагностики, використовувані застосуванням стандартних способів безпосереду аналізі генетичного зчіплювання, для визначення нього аналізу мутацій, як-то аналіз поліморфізму генної аномалії або аберації у пацієнта, відомі в довжини рестрикційних фрагментів, аналіз коротрівні техніки. Більшість способів діагностики вклюких тандемних повторень із застосуванням спосочає етапи (і) отримання генетичного зразку від бів PCR, аналіз непокірливих стосовно ампліфікапотенційно хворого пацієнта, хворого пацієнта або ції мутаційних систем, визначення потенційно здорового носія рецесивної алелі заодноланцюгового конформаційного поліморфізму, хворювання; (іі) продукування першого продукту способи розщеплення РНазою, гель-електрофорез реакції інкубуванням генетичного зразку з полінукз виродженим градієнтом, аналіз невідповідності леотидним зондом ліганду zalpha11, причому поза допомогою флуоресценції та інші відомі в рівні лінуклеотид гібридизуватиметься з комплементартехніки способи генетичного аналізу [дивися, наною полінуклеотидною послідовністю, як-то у приклад, Mathew.(ed.). Protocols in Human RFLP-аналізі, або інкубуванням генетичного зразку Molecular Genetics (Humana Press, Inc. 1991), з сенсовими та антисенсовими праймерами у реаMarian, Chest 108: 255 (1995), Coleman та кції PCR у прийнятних умовах реакції PCR; (Ні) Tsongalis. Molecular Diagnostics (Humana Press, візуалізацію першого продукту реакції гельInc. 1996), Elles (ed.) Molecular Diagnosis of Genetic електрофорезом та/або іншим відомим способом, Diseases(Humana Press, Inc. 1996), Landegren як-то візуалізація першого продукту реакції поліну(ed.), Laboratory Protocols for Mutation Detection клеотидним зондом ліганду zalpha11, причому (Oxford University Press 1996). Birren et al. (eds.). полінуклеотид гібридизуватиметься з комплеменGenome Analisis, Vol. 2: Detecting Genes (Cold тарною полінуклеотидною послідовністю з першої Spring Harbor Laboratory Press 1998). Dracopoli et реакції; та (iv) порівняння візуалізованого першого al. (eds.), Current Protocols in Human Genetics (John продукту реакції з другим продуктом контрольної Wiley & Sons 1998), та Richards та Ward, " реакції генетичного зразку від нормальної або конMolecular Diagnostics Testing" in Principles of трольної особи. Відмінність між першим продуктом Molecular Medicine, pages 83-88 (Humana Press, реакції та продуктом контрольної реакції є показInc. 1998)]. Прямий аналіз гена ліганду zalpha11 на ником генної аномалії у потенційно хворого чи мутації можна провести, використовуючи геномну хворого пацієнта, або наявність фенотипу гетероДНК осіб. Способи ампліфікації геномної ДНК, зиготного рецесивного носія для здорового пацієнотриманої наприклад з периферійної лімфоцитів та, або наявність генетичного дефекту у пухлині крові, добре відомі фахівцям, [дивися, наприклад, від хворого пацієнта, або наявність генної аномалії Dracopoli et al. (eds.), Current Protocols in Human в утробному плоді чи ембріоні для імплантації. Genetics, сторінки 7,1,6-7,1,7 (John Wiley & Sons Наприклад, відмінність у стр уктурі рестрикційного 1998)]. 23 84387 24 Позиції інтронів у гені ліганду zalpha11 визназонду, іонну силу гібридизаційних розчинів, та начали ідентифікацією геномних клонів, з наступним явність дестабілізуючих засобів у гібридизаційних секвенсуванням стиків інтрон/екзон. Перший інрозчинах. Для розрахунку Тm в рівні те хніки відомі трон знаходиться між амінокислотними залишками різні рівняння, що є специфічними для гібридів 56 (Leu) та 57 (VaI) у послідовності SEQ ID NO: 2, ДНК, РНК та ДНК-РНК і послідовностей полінуклета має 115 пар основ у довжину. Др угий інтрон є отидних зондів змінної довжини [дивися, напринайбільшим, маючи 4,4 кілооснов, та знаходиться клад, Sambrook et al. Molecular Cloning: A між амінокислотними залишками 68 (GIu) та 69 Laboratory Manual, Second Edition (Cold Spring (Thr) у послідовності SEQ ID NO: 2. Третій інтрон Harbor Press 1989); Ausubel et al, (eds.). Current має-2,6 кілооснов, та знаходиться між амінокислоProtocols in Molecular Biology (John Wiley та Sons, тними залишками 120 (Leu) та 121 (Thr) у послідоInc. 1987); Berger та KLimmel (eds.), Guide to вності SEQ ID NO: 2. Кінцевий інтрон, розміром 89 Molecular Cloning Techniques, (Academic Press, Inc. пар основ, знаходиться між амінокислотними за1987); та Wetmur, Crii. Rev. Biochem. MoI. Biol. 26: лишками 146 (Lys) та 147 (Met) у послідовності 111 (1990)]. Програмне забезпечення секвенсуSEQ ID NO: 2. Повна протяжність гена складає вання як-то OLIGO 6,0 (LSR; Long Lake, MN) та приблизно 8 ко. Primer Premier 4,0 (Premier Biosoft International; Структура гена ліганду zalpha11 є подібною до Palo Alto, CA), а також сайта в Інтернеті, є придатгена IL-2 [Fujita et al. Proc. Natl. Acad. Sci. 80: 7437ними інструментами для аналізу певної послідов7441, 1983], хоча ген ліганду zalpha11 містить один ності та розрахунку Tm на основі визначених кридодатковий інтрон (інтрон-4). Структура короткого теріїв користувача. Такі програми можуть також першого інтрону та довгого другого та третього аналізувати певну послідовність у визначених інтронів є збереженою для обох генів, хоча ген IL-2 умовах та ідентифікувати придатні послідовності є, взагалі, тро хи меншим (приблизно на 6 ко). Ген зонду. Звичайно гібридизацію довших полінуклеоIL-15, з іншого боку, складається з 8 екзонів та тидних послідовностей, більше 50 пар основ, пропростягається щонайменше на 34 ко [Anderson et водять при температурах приблизно на 20-25°C al. Genomics 25: 701-706, 1995]. Отже ген ліганду нижче розрахованої Тm. Для менших зондів, менzalpha11 є подібнішим за структурою до гена IL-2, ше 50 пар основ, гібридизацію звичайно проводять ніж до гена IL-15. при Tm або на 5-10°C нижче розрахованої Тm. Це За втіленнями згідно з винаходом виділені кодає максимальну швидкість гібридизації для гібридуючі ліганд zalpha11 молекули нуклеїнової кислодів ДНК-ДНК та ДНК-РНК. ти можуть гібридизуватися у суворих умовах з моПісля гібридизації, молекули нуклеїнової кислекулами нуклеїнової кислоти, що мають лоти можна промити для видалення негібридизонуклеотидну послідовність SEQ ID NO: 1, з молеваних молекул нуклеїнової кислоти у суворих умокулами нуклеїнової кислоти, що мають нуклеотидвах, або у дуже суворих умовах. Типові суворі ну послідовність з нуклеотидів 47-532 послідовноумови промивки включають промивку у розчині сті SEQ ID NO: 1, або з молекулами нуклеїнової 0,5-2хSSC (розчин хлориду та цитрату натрію) з кислоти, що мають нуклеотидну послідовність, 0,1% додецилсульфатом натрію (SDS) при 55комплементарну SEQ ID NO: 1. Загалом суворі 65°С. Іншою мовою, молекули нуклеїнової кислоумови вибирають приблизно на 5°С нижче темпети, що кодують варіацію поліпептидів ліганду ратури плавлення (Tm) певної послідовності при zalpha11, гібридизуються з молекулою нуклеїнової визначених іонній силі та рН. кислоти, що має нуклеотидну послідовність SEQ Tm є температурою (при визначених іонній силі ID NO: 1 (або її комплемент) у сувори х умовах та рН), при якій 50% цільової послідовності гібрипромивки, при яких суворість промивки є еквіваледизує з цілковито підігнаним зондом. нтною 0,5-2хSSC з 0,1% SDS при 55-65°С, вклюСпарені молекули нуклеїнової кислоти, як-то чаючи 0,5хSSC 0,1% SDS при 55°С, або 2хSSC з ДНК-ДНК, РНК-РНК та ДНК-РНК, можуть гібриди0,1% SDS при 65°C. Пересічний фахівець може зуватися, якщо нуклеотидні послідовності мають легко визначити еквівалентні умови, наприклад, деякий ступінь комплементарності. Гібриди можуть заміщенням SSC на SSPE у розчині для промивки. допускати невідповідність пари основ у подвійній Типові дуже суворі умови промивки включають спіралі, але стабільність гібриду залежить від ступромивку у розчині 0,1-0,2хSSC з 0,1% додецилпеню невідповідності. Tm невідповідного гібриду сульфатом натрію (SDS) при 50-65°C. Іншою мозменшується на 1°С для кожних 1-1,5% невідповівою, молекули нуклеїнової кислоти, що кодують дності пар основ. Варіювання суворості умов гібваріацію поліпептидів ліганду zalpha11, гібридизуридизації дозволяє регулювати ступінь невідповідються з молекулою нуклеїнової кислоти, що має ності, що буде наявною у гібриді. Ступінь суворості нуклеотидну послідовність SEQ ID NO: 1 (або її збільшується при збільшенні температури гібридикомплемент) у дуже суворих умовах промивки, в зації та зменшенні іонної сили гібридизаційного яких суворість промивки є еквівалентною 0,1буферу. 0,2хSSC з 0;1% SDS при 50-65°C, включаючи В рамках компетенції фахівців добре адапту0,1хSSC з 0,1% SDS при 50°C, або 0,2хSSC 0,1% вати ці умови для використання з певним полінукSDS при 65°C. леотидним гібридом. Tm для певної цільової посліЗгідно з винаходом запропоновано також видідовності є температурою (при визначених умовах), лені поліпептиди ліганду zalpha11, що мають по при якій 50% цієї послідовності гібридизуватисуті подібну природу послідовності поліпептидів меться з цілковито підігнаною послідовністю зонстосовно послідовності SEQ ID NO: 2, або їх ортоду. Ці умови, які впливають на Tm, включають, велоги. Термін "по суті подібну природу послідовносличину та склад пар основ полінуклеотидного ті" використано тут для позначення поліпептидів, 25 84387 26 що мають щонайменше 70%, щонайменше 80%, амінокислотною послідовністю та амінокислотною щонайменше 90%, щонайменше 95%, або більше послідовністю передбачуваної варіації ліганду 95% ідентичності послідовності стосовно послідоzalpha11. Алгоритм FASTA описано [Pearson and вностей, показаних у SEQ ID NO: 2, або їх ортолоLipman, Proc. Natl. Acad. Sci. USA 85: 2444 (1988), гів. Представлений винахід також включає поліпета Pearson, Meth. En zymol. 183: 63 (1990)]. птиди, що включають амінокислотну послідовність, Коротше, FASTA, по-перше, характеризує пощо має щонайменше 70%, щонайменше 80%, щодібність послідовності ідентичними регіонами, спінайменше 90%, щонайменше 95% або більше 95% льно використовуваними з'ясованою послідовнісідентичності послідовності стосовно послідовності тю (наприклад, SEQ ID NO: 2) та досліджуваною амінокислотних залишків 1-162 або 33-162 посліпослідовністю, що мають або найвищу густину довності SEQ ID NO: 2. Представлений винахід ідентичних ділянок (якщо величина ktup є 1) або крім того включає молекули нуклеїнової кислоти, пари ідентичних ділянок (якщо ktup=2), без огляду що кодують такі поліпептиди. Способи визначення на заміни, вставки, або делеції консервативних проценту ідентичності описано нижче. амінокислот. Десять регіонів з найвищою густиною Представлений винахід також охоплює різні ідентичних ділянок реєструють далі порівнянням молекули нуклеїнової кислоти ліганду zalpha11, подібності усіх спарених амінокислот, використощо можна ідентифікувати, використовуючи два вуючи матрицю амінокислотних замін, а кінці регікритерії: визначення подібності між кодованим онів "підрізають" для включення тільки тих залишполіпептидом та амінокислотною послідовністю ків, що сприяють найвищій оцінці. Якщо є кілька SEQ ID NO: 2, та/або гібридизаційним дослідженрегіонів з оцінкою більше "граничної" величини ням, як описано вище. Такі варіації ліганду (розрахованої за попередньо визначеною формуzalpha11 включають молекули нуклеїнової кислолою на основі довжини послідовності та величини ти: (1) що гібридизуються з молекулою нуклеїнової ktup), далі підрізані початкові регіони оцінюють для кислоти, що має нуклеотидну послідовність послівизначення, чи можуть регіони поєднуватися для довності SEQ ID NO: 1 (або її комплемент) у сувоутворення приблизного суміщення з гепами. Під рих умовах промивки, де суворість промивки є кінець, найвище оцінені регіони двох амінокислотеквівалентною 0,5-2хSSC з 0,1% SDS при 55-65°С; них послідовностей суміщають, використовуючи або (2), що кодують поліпептид, що має щонаймемодифікацію алгоритму Needleman-Wunsch-Scllers нше 70%, щонайменше 80%, щонайменше 90%, [Needleman та Wunsch, J. Mol. Вюі. 48: 444 (1970); щонайменше 95% або більше 95% ідентичності Sellers, SIAM J. Appl. Math. 26: 787 (1974)], який послідовності стосовно амінокислотної послідовдозволяє амінокислотні вставки та делеції. Краності SEQ ID NO: 2. Альтернативно, варіації ліганщими параметрами для аналізу FASTA є: ktup=l, ду zalpha11 можна характеризувати як молекули пенальті початку гепу 10, пенальті подовження нуклеїнової кислоти: (1), що гібридизуються з могепу 1 та матрицю замін=BLOSUM62. Ці параметлекулою нуклеїнової кислоти, що має нуклеотидну ри можна уводити у програму FASTA модифікаціпослідовність SEQ ID NO: 1 (або її комплемент) у єю файлу матриці підрахунку ("SMATRlX"), як подуже суворих умовах промивки, де суворість прояснено у [додатку 2 у Pearson. Meth. Enzymol. 183: мивки є еквівалентною 0,1-0,2хSSC з 0,1% SDS 63 (1990)]. при 50-65°C; та (2), що кодують поліпептид, який FASTA можна також використовувати для вимає щонайменше 70%, щонайменше 80%, щонайзначення ідентичності послідовності молекул нукменше 90%, щонайменше 95% або більше 95% леїнової кислоти, використовуючи розкрите вище ідентичності послідовності стосовно амінокислотспіввідношення. Для порівнянь нуклеотидних поної послідовності SEQ ID NO: 2. слідовностей, величина ktup може бути від 1 до 6, краще 3-6. найкраще 3, з установкою інших параметрів за умовчуванням. Різні поліпептиди ліганду zalpha11 або поліпептиди з подібною по суті природою послідовності характеризуються як такі, що мають одну чи більше замін, делецій або вставок амінокислот. Ці зміни мають переважно другорядну природу, що є консервативними замінами амінокислот (дивися Таблицю 5) та іншими замінами, що помітно не впливають на укладку або активність поліпептидів; невеликими делеціями, звичайно 1-30 амінокислот; та аміно- або карбокси-кінцевими подовженнями, як-то аміно-кінцевим залишком метіоніну, невеликим лінкерним пептидом з приблизно 20-25 залишків, або афінною міткою. Представлений винахід, отже, включає поліпептиди з приблизно 108-216 амінокислотних залишків, що включають Фахівці здогадуються, що є багато встановлепослідовність, що є щонайменше на 70%, краще них, придатних для суміщення двох амінокислотщонайменше на 90%, ще краще на 95% або більних послідовностей алгоритмів. Алгоритм "FASTA" ше ідентична відповідному регіону послідовності дослідження подібності Пірсона та Ліпмана SEQ ID NO: 2. Поліпептиди, що містять афінні міт(Pearson та Lipman) є придатним способом суміки, можуть також включати сайт протеолітичного щення білків для виявлення кількості ідентичних розщеплення між поліпептидом ліганду zalpha11 ділянок, спільно використовуваних розкритою тут 27 84387 28 та афінною міткою. Кращі такі сайти включають Наприклад, можна зробити порівняння цистеїнової сайти розщеплення тромбіну та сайти розщепленструктури у варіантних та стандартних молекулах. ня фактору Xa. Mac-спектрометрія та хімічна модифікація, використовуючи відновлення та алкілування, забезпеТаблиця 5 чує способи визначення цистеїнових залишків, що пов'язані дисульфідними зв'язками або є вільними Заміщення консервативних амінокислот від такого поєднання [Bean et al. Anal. Biochem. 201: 216-226,1992; Gray, Protein Sci. 2: 1732-1748, 1993; та Patterson et al., Anal. Chem. 66: 3727-3732, Аргінін 1994]. Взагалі можна вважати, що якщо модифікоОсновні Лізин вана молекула не має такої цистеїнової структури, Гістидин як стандартна молекула, упаковка може бути ураГлутамінова кислота Кислі женою. Іншим добре відомим та доступним спосоАспаргінова кислота бом виміру укладки є циркулярний дихроїзм(СО). Глутамін Полярні Вимір та порівняння спектрів CD, генерованих моАспаргін дифікованою молекулою та стандартною молекуЛейцин лою є звичайними [Johnson, Proteins 7: 205-214. Гідрофобні Ізолейцин 1990]. Кристалографія є іншим добре відомим Валін способом аналізу укладки та структури. Ядерний Фенілаланін магнітний резонанс (NMR), картування пептидним Ароматичні Триптофан розщепленням та епітопне картування є також Тирозин добре відомими способами аналізу укладки та Гліцин структурної подібності між білками та поліпептиАланін дами [Schaanan et al. Science 257: 961-964, 1992]. Малі Серин Можна створювати профіль гідрофільності за Треонін Хоппом/Вудсом послідовності білку ліганду Метіонін zalpha11, яку показано послідовністю SEQ ID NO: 2 [Норр et al., Proc. Natl. Acad.Sci, 78: 3824-3828, Можна визначити амінокислотні залишки, що 1981; Норр, J. Immun. Meth. 88: 1-18, 1986 та включають регіони або домени, які є критичними Triquier et al., Protein Engineering 11: 153-169, для підтримування структурної цілісності. У цих 1998]. Профіль оснований на ковзному шестирегіонах можна визначити специфічні залишки, що залишковому вікні. Внутрішні залишки G, S, та T та будуть більш чи менш толерантними до зміни та відкриті залишки H, Y, та W ігнорували. Наприпідтримуватимуть загальну третинну стр уктуру клад, у ліганді zalpha11 гідрофільні регіони вклюмолекули. Способи аналізу стр уктури послідовночають амінокислотні залишки 114-119 послідовностей включають, але без обмеження, суміщення сті SEQ ID NO: 2, амінокислотні залишки 101-105 багатьох послідовностей з високою амінокислотпослідовності SEQ ID NO: 2, амінокислотні залишною або нуклеотидною ідентичністю, схильність ки 126-131 послідовності SEQ ID NO: 2, амінокисвторинних структур, бінарні структури, комплеменлотні залишки 113-118 послідовності SEQ ID NO: тарну упаковку та вн утрішні полярні взаємодії 2, та амінокислотні залишки 158-162 послідовності [Barton, Current Opin.Struct. Biol. 5: 372-376, 1995 SEQ ID NO: 2. та Cordes et al., Current Qpin. Struct. Biol. 6: 3-10, Фахівці здогадаються, що гідрофільність або 1996]. Взагалі, при проектуванні модифікації молегідрофобність треба брати до уваги при конструюкул або ідентифікуванні специфічних фрагментів ванні модифікацій у амінокислотній послідовності визначення структури супроводжуватиметься оціполіпептиду ліганду zalpha11, щоб не порушити нюванням активності модифікованих молекул. загальний структурний та біологічний профіль. Зміни амінокислотної послідовності здійснюОсобливо цікавими для заміни є гідрофобні залиють у поліпептидах ліганду zalpha11 так, щоб мінішки, вибрані з групи, що складається з VaI, Leu та мізувати порушення структур вищого порядку, що IIе або з групи, що складається з Met, GIy, Ser, Ala, є суттєвими для біологічної активності. Наприклад, Туr та Тrр. Наприклад, залишки толерантні до затам, де поліпептид ліганду zalpha11 включає одну мін можливо включають залишки 100 та 103, як чи більше спіралей, зміни у амінокислотних залипоказано послідовністю SEQ ID NO: 2. Цистеїнові шках будуть зроблені так, щоб не порушити спіразалишки у позиціях 71, 78, 122 та 125 послідовносльну геометрію та інші компоненти молекули, де ті SEQ ID NO: 2, будуть відносно нетолерантними зміни у конформації послаблюють деякі критичні до замін. функції, наприклад, приєднання молекули до її Ідентичність за незамінними амінокислотами партнерів по зв'язуванню, наприклад, спіралей А також можна визначити з аналізу подібності посліта D, залишків 44, 47 та 135 послідовності SEQ ID довностей IL-15, IL-2, IL-4 та GM-CSF стосовно NO: 2. Вплив зміни амінокислотної послідовності ліганду zalpha11, використовуючи способи, як-то можна прогнозувати, наприклад, комп'ютерним аналіз "FASTA", що описано попередньо, регіони моделюванням, як розкрито вище, або визначити високої подібності ідентифікують у родині білків та аналізом кристалічної структури [дивися, напривикористовують для аналізу амінокислотної посліклад, Lapmorn et al., Nat. Struct. Biol. 2: 266-268, довності збережених регіонів. Альтернативний 1995]. Інші способи, що добре відомі в рівні техніпідхід для ідентифікації варіації поліпептиду ліганки, порівнюють укладку варіантного білку зі станду zalpha11 на основі структури призначений для дартною молекулою (наприклад, нативного білку). визначення, чи може молекула нуклеїнової кисло 29 84387 30 ти, що кодує потенційну варіацію гена ліганду вності SEQ ID NO: 111; (b) спіралі В IL-4 zalpha11, гібридизуватися з молекулою нуклеїноамінокислотні залишки 65-83 послідовності SEQ ID вої кислоти, що має нуклеотидну послідовність NO: 112: (с) спіралі В IL-15 - амінокислотні залишSEQ ID NO: 1, яку обговорено вище. ки 84-101 послідовності SEQ ID NO: 113; (d) спіраІнші способи ідентифікування незамінних амілі В GMCSF -амінокислотні залишки 72-81 послінокислоти у поліпептидах представленого винаходовності SEQ ID NO: 114; та (e) - амінокислотні ду відомі в рівні техніки, як-то сайт-спрямований залишки 69-84 послідовності SEQ ID NO: 2; другий мутагенез або аланін-сканувальний мутагенез спейсер - амінокислотні залишки 5-11; третій полі[Cunningham та Wells. Science 244: 1081 (1989), пептид, що включає послідовність амінокислотних Bass et al., Proc. Nati Acad. Sci. USA 88: 4498 залишків, що вибрана з групи, яка складається з: (1991), Coombs та Corey, "Site-Directed (а) спіралі C IL-2 - залишки 87-99 послідовності Mutagenesis та Protein Engineering," у Protein: SEQ ID NO: 111; (b) спіралі C IL-4 - залишки 95-118 Analysis та Design. Angeletti (ed.), стор.259-311 послідовності SEQ ID NO: 112; (с) спіралі C IL-15 (Academic Press, Inc. 1998)]. За останнім спосозалишки 107-119 послідовності SEQ ID NO: 113; бом, одиничні аланінові мутації уводять на кожно(d) спіралі C GMCSF - залишки 91-102 послідовному залишку у молекулі, а утворені мутантні молесті SEQ ID NO: 114; та (e) - амінокислотні залишки кули досліджують на біологічну або біохімічну 92-105 послідовності SEQ ID NO: 2; третій спейсер активність, як розкрито нижче для ідентифікації - амінокислотні залишки 3-29; та четвертий поліамінокислотних залишків, що є критичними стосопептид, що включає амінокислотні залишки, вибвно активності молекули. [Ди вися також, Hilton et рані з групи, що складається з: (а) спіралі D EL-2 al., J. Biol. Chem. 277: 4699 (1996)]. амінокислотні залишки 103-121 послідовності SEQ. Представлений винахід також включає функID NO: 111; (b) спіралі D IL-15 - амінокислотні заціональні фрагменти поліпептиду ліганду zalpha11 лишки 134-157 послідовності SEQ ED NO: 112; (с) та молекули нуклеїнової кислоти, що кодують такі спіралі D IL-4 - амінокислотні залишки 134-160 функціональні фрагменти. "Функціональний" ліганд послідовності SEQ ID NO: 113; (d) спіралі D zalpha11 або його фрагмент, що визначено тут, GMCSF - амінокислотні залишки 120-131 послідохарактеризуються їх проліферативною або дифевності SEQ ID NO: 114; та (e) амінокислотні залиренціювальною активністю, їх здатністю індукува ти шки 135-148 послідовності SEQ ID NO: 2, причому або інгібувати спеціалізовані функції клітин, або їх щонайменше один з 4-х поліпептидів є похідним здатністю приєднуватися специфічно до антитіл від ліганду zalpha11. Згідно з іншим втіленням випроти ліганду zalpha11, або рецептору zalpha11 находу спейсерні пептиди вибиратимуть з петель (розчинного або іммобілізованого). Як попередньо ліганду zalpha11, IL-2, EL-4, EL-15 або GM-CSF описано, ліганд zalpha11 характеризується струкА/В, В/С та CfD, як показано у таблиці 1. турою з чотирьох -спіральним жмутом, що містить Стандартні делеційні аналізи молекули нуклеспіраль А (амінокислотні залишки 41-56), спіраль В їнової кислоти можна провести для отримання (амінокислотні залишки 69-84), спіраль C (амінокифункціональних фрагментів молекули нуклеїнової слотні залишки 92-105) та спіраль D (амінокислотні кислоти, що кодує поліпептид ліганду zalpha11. В залишки 135-148), як показано послідовністю SEQ якості ілюстрації, молекули ДНК, що мають нуклеID NO: 2. Отже, згідно з винаходом запропоновано отидну послідовність SEQ ID NO: 1 або її фрагметакож конденсовані білки, що включають: (а) полінти, можна розщепити нуклеазою Bal31 для отрипептидні молекули, що містять одну чи більше мання серії згрупованих делецій. Ці фрагменти описаних вище спіралей; та (b) функціональні ДНК далі вставляють в експресійні вектори у нафрагменти, що містять одну чи більше з цих спілежній рамці зчитування та виділяють і досліджуралей. Іншій частині поліпептиду конденсованого ють експресовані поліпептиди на активність ліганбілку може сприяти інший цитокін з чотирьохду zalpha11, або на здатність приєднувати спіральним жмутом, як-то IL-15, EL-2, IL-4 та GMантитіла проти ліганду zalpha11 або рецептор CSF, або штучний та/або неспоріднений секреторzalpha11. Одною альтернативою розщепленню ний сигнальний пептид, що полегшує секрецію екзонуклеазою є використання олігонуклеотидконденсованого білку. спрямованого мутагенезу для введення делецій Отже, згідно з винаходом запропоновано конабо стоп-кодонів для специфічного продукування денсовані білки, що містять щонайменше чотири потрібного фрагменту ліганду zalpha11. Альтернаполіпептиди, причому порядок поліпептидів від Nтивно, певні фрагменти гена ліганду zalpha11 мозакінчення до C-закінчення такий: перший поліпепжна синтезувати, використовуючи полімеразну тид включає амінокислоти, вибрані з групи, що ланцюгову реакцію. складається з: (а) спіралі A IL-2 - амінокислотні Стандартні способи ідентифікування функціозалишки 36-46 послідовності SEQ ID NO: 111; (b) нальних доменів добре відомі фахівцям. Наприспіралі A IL-15 - амінокислотні залишки 29-43 поклад, вивчення на усіченому одному з двох чи слідовності SEQ ID NO: 112; (с) спіралі A IL-4 обох закінченнях інтерферонів підсумовано амінокислотні залишки 45-68 послідовності SEQ ID [Horisberger and Di Marco, Pharmac. Ther. 66: 507 NO: 113; (d) спіралі А GMCSF - амінокислотні за(1995)]. Більш того, стандартні способи функціолишки 30-44 послідовності SEQ ID NO: 114: та (e) нального аналізу білків описано, наприклад, амінокислотні залишки 41-56 послідовності SEQ ID [Treuter et al., Molec. Gen-Genet. 240: 113 (1993); NO: 2; перший спейсер - амінокислоти 6-27; та Content et al., "Expression та preliminary deletion другий поліпептид, що включає амінокислотні заanalysis of the 42kDa 2-5A synthetase induced by лишки, вибрані з групи, що складається з: (а) спіhuman interferon," (Експресія та попередній делералі В IL-2 - амінокислотні залишки 53-75 послідоційний аналіз індукованої інтерфероном людини 31 84387 32 синтетази 2-5А у 42 кДа) у Biological Interferon спіралей з ліганду zalpha11 можна поєднувати з Systems, Proceedings of ISIR-TNO Meeting on іншими цитокінами для посилення їх біологічних Interferon Systems, Cantell (ed.), стор.65-72 (Nijhoff властивостей чи ефективності продукування. 1987); Herschman, "The EGF Рецептор," у Control Згідно з винаходом запропоновано тому серію of Animal Cell Proliferation L, Boynton et al., (eds.) нових гібридних молекул, в яких сегмент, що місстор.169-199 (Academic Press 1985); Coumailleau et тить одну чи більше спіралей з ліганду zalpha11, al., J. Biol. Chem. 270: 29270 (1995); Fukunaga et конденсований з іншим поліпептидом. Конденсаal., J. Biol. Chem. 270: 25291 (1995); Yamaguchi et цію краще здійснювати сплайсингом на рівні ДНК al., Bi-ochem. Pharmacol. 50: 1295 (1995); та Meisel для дозволу експресії химерних молекул у систеet al., Plant Molec. Biol. 30: 1 (1996)]. мах продукування рекомбінантів. Утворені молеБагато амінокислотних замін можна здійснити кули далі досліджують на такі властивості як пота дослідити, використовуючи відомі способи муліпшена розчинність, поліпшена стабільність, тагенезу та скринінгу, як-то розкриті Reidhaarподовжений період напіввидалення, поліпшені Olson та Sauer [Science 241: 53 (1988)) або Bowie рівні експресії та секреції, та фармакодинаміку. та Sauer (Proc. Natl. Acad. Sci. USA 86:2152 Такі гібридні молекули можуть крім того включати (1989)]. Коротше, ці автори описують способи оддодаткові амінокислотні залишки (наприклад, поночасної рандомізації двох чи більше позицій у ліпептидний лінкер) між компонентними білками поліпептиді, селекції на функціональні поліпептиабо поліпептидами. ди, а потім секвенсування мутагенізованих поліпеНеіснуючі в природі амінокислоти включають, птидів для визначення спектру дозволених замін в без обмеження, транс-3-метилпролін, 2,4кожній позиції. Інші способи, що можна використометанпролін, цис-4-гідроксипролін, транс-4вувати, включають фагове виявлення [наприклад, гідроксипролін, N-метилгліцин, аllо-треонін, метилLowman et al, Biochem. 30: 10832 (1991), Ladner et треонін, гідроксіетилцистеїн, гідроксіетилгомоцисal., Патент CШA №5223409, Huse, міжнародна теїн, нітроглутамін, гомоглутамін, піпеколінову публікація № WO 92/06204], та регіонкислоту, тіазолідинкарбонову кислоту, дегідропроспрямований мутагенез [Derbyshire et al. Gene 46: лін, 3- та 4-метилпролін. 3,3-диметилпролін, трет145 (1986), та Ner et al., DNA 7; 127, (1988)]. лейцин, норвалін, 2-азафенілаланін, 3Варіації розглянутих послідовностей нуклеоазафенілаланін, 4-азафенілаланін, та 4тиду та поліпептиду ліганду zalpha11 можна також флуорфенілаланін. В рівні техніки відомі кілька створювати перемішуванням ДНК, як розкрито способів вставки неіснуючих в природі амінокисло[Stemmer, Nature 370: 389 (1994), Stemmer, Proc. тних залишків у білки. Наприклад, можна застосоNatl Acad. Sci. USA 91: 10747 (1994) та у міжнаровувати систему in vitro, причому несенсові мутації дній публікації № WO 97/20078]. Коротше, варіації пригнічують, використовуючи хімічно аміноациломолекули ДНК створюють гомологічною рекомбівані супресорні тРНК. Способи синтезу амінокиснацією in vitro випадковою фрагментацією вихідної лот та аміноацилуючих тРНК відомі в рівні техніки. ДНК з наступним реконструюванням, використоТранскрипцію та трансляцію плазмід, що містять вуючи PCR, результатом чого є випадково уведені несенсові мутації, звичайно проводять у позбавмісцеві мутації. Ці способи можна модифікувати, леній клітин системі, що містить екстракт E. coli використовуючи родину вихідної молекули ДНК, S30 та комерційно доступні ферменти та інші реаяк-то алельні варіанти або молекули ДНК від різгенти. Білки очищають хроматографією. [Дивися, них видів, для введення додаткової варіабельності наприклад, Robertson et al., J. Am. Chem. Soc. 113: у процес. Селекція або скринінг на потрібну актив2722 (1991), Ellman et al., Methods EnzvmoL 202: ність з наступними додатковими повтореннями 301 (1991), Chung et al., Science 259: 806 (1993), та мутагенезу та дослідження забезпечує швидку Chung et al., Proc. Natl. Acad. Sci. USA 90: 10145 "еволюцію" послідовностей селекцією на потрібні (1993)]. мутації, одночасно селектуючи проти шкідливих За другим способом, трансляцію проводять в змін. ооцитах Xenopus мікроін'єкцією мутованої мРНК та Способи мутагенезу, які тут розкрито, можна хімічно аміноацилованих супресорних тРНК комбінувати з високопродуктивними автоматизо[Turcatti et al., J. Biol. Chem. 271: 19991 (1996)]. За ваними способами скринінгу для визначення актитретім способом, клітини E. coli культивують при вності клонованих, мутагені-зованих поліпептидів у відсутності природної амінокислоти, яку треба заклітинах хазяїна. Мутагенізовані молекули ДНК, мінити (наприклад, фенілаланіну), та у присутності що кодують біологічно активні поліпептиди, або потрібних неіснуючих в природі амінокислот (наполіпептиди, що приєднуються до антитіл проти приклад, 2-азафенілаланіну, 3-азафенілаланіну, 4ліганду zalpha11 або розчинного рецептору азафенілаланіну, або 4-флуорфенілаланіну). Неzalpha11, можна вилучити з клітин хазяїна та швиіснуючу в природі амінокислоту вставляють у білок дко секвенсувати, використовуючи сучасне обладна місце її природного еквівалента. [Дивися, Koide нання. Ці способи дозволяють швидке визначення et al., Biochem. 33: 7470 (1994)]. Існуючі в природі важливості окремих амінокислотних залишків у амінокислотні залишки можна перетворити у неіспотрібному поліпептиді і можуть застосовуватися нуючі в природі види хімічною модифікацією in до поліпептидів невідомої структури. vitro. Хімічну модифікацію можна комбінувати з На додаток, білки представленого винаходу сайт-спрямованим мутагенезом для подальшого (або фрагменти їх поліпептидів) можна поєднувати вирощування ступеню заміни [Wynn and Richards, з іншими біоактивними молекулами, зокрема інProtein Sci. 2: 395 (1993)]. Може бути кращим сташими цитокінами, для забезпечення багатофункбілізувати ліганд zalpha11 для збільшення періоду ціональних молекул. Наприклад, одну чи більше напіввидалення молекул, зокрема для збільшення 33 84387 34 тривалості метаболічної дії у активному стані. Для (The Humana Press, Inc. 1992); Price, "Production та досягнення збільшеного періоду напіввидалення Characterization of Sinthetic Peptide-Derived молекули ліганду zalpha11 можна хімічно модифіAntibodies," у Monoclonal Antibodies: Production, кувати, використовуючи способи, що описано тут. Engineering та Clinical Application, Ritter та Ladyman Обробка PEG є одним звичайно використовуваним (eds.), стор.60-84 Cambridge University Press 1995), способом, що продемонстровано збільшенням та Coligan et al. (eds.). Current Protocols in періоду напіввидалення у плазмі, збільшенням Immunology, стор.9-3,1 - 9,3,5 та стор.9,4,1 - 9,4,11 розчинності, та зменшенням антигенності та імуно(John Wiley & Sons 1997)]. генності [Nucci et al., Advanced Drug Delivery Незалежно від окремої нуклеотидної послідоReviews 6: 133-155, 1991 та Lu et al., Int. J. Peptide вності варіації поліпептиду ліганду zalpha11, поліProtein Res, 43: 127-138, 1994]. нуклеотид кодує поліпептид, що характеризується Амінокислотні залишки ліганду zalpha11 можйого проліферативною або диференціювальною на замінити обмеженим числом некон-сервативних активністю, здатністю індукувати або інгібувати амінокислот, амінокислот, що не кодовані генетичспеціалізовані клітинні функції, або здатністю спеним кодом, неіснуючих в природі амінокислоти, та цифічно приєднуватися до антитіла проти ліганду протиприродних амінокислот. zalpha11 та рецептору zalpha11. Точніше, варіації Згідно з винаходом запропоновано також фраполінуклеотидів ліганду zalpha11, кодуватимуть гменти поліпептидів або пептиди, що містять часполіпептиди, які виявляють щонайменше 50%, а тину описаного тут поліпептиду ліганду zalpha11, краще, більше 70%, 80% або 90%, активності поліщо несе епітоп. Такі фрагменти або пептиди мопептиду, який показано послідовністю SEQ ID NO: жуть включати "імуногенний епітоп," який є части2. ною білку, що викликає реакцію антитіл, коли як Для будь-якого поліпептиду ліганду zalpha11, імуноген використовують суцільний білок. Імуновключаючи варіації та конденсовані білки, пересічгенні пептиди, що несуть епітоп, можна ідентифіний фахівець може легко створити повністю вирокувати, використовуючи стандартні способи [дивиджену полінуклеотидну послідовність, що кодує цю ся, наприклад, Geysen et al., Proc. Natl. Acad. Sci. варіацію, використовуючи інформацію з вищенаUSA 8J; 3998 (1983)]. ведених таблиць 1 та 2. За контрастом, поліпептидні фрагменти або Згідно з винаходом запропоновано також ряд пептиди можуть включати "антигенний епітоп," що інших поліпептидних конденсатів (та споріднених є регіоном молекули білку, до якого може специфімультимерних білків, що містять один чи більше чно приєднуватися антитіло. Деякі епітопи склаполіпептидних конденсатів). Наприклад, поліпепдаються з лінійної або суміжної ділянки амінокистиди ліганду zalpha11 можна виготовити конденлоти, і антигенність такого епітопу не знищується сованими у диме-ризований білок, як розкрито у денатуруючими засобами. В рівні техніки відомі [патентах США №5155027 та 5567584]. Кращі дивідносно короткі синтетичні пептиди, що можуть меризовані білки в цьому відношенні включають імітувати епітопи білку, які можна використовувати домени постійного регіону імуноглобуліну. Кондендля стимулювання продукування антитіл проти сати імуноглобулін-поліпептид ліганду zalpha11 білку [дивися, наприклад, Sutcliffe et al., Science можна експресувати у генетично сконструйованих 219: 660 (1983)]. Відповідно, антигенні пептиди та клітинах (для продукування ряду мультимерних поліпептиди представленого винаходу, що несуть аналогів ліганду zalpha11). З поліпептидами ліганепітоп, є корисними для генерування антитіл, що ду zalpha11 для спрямування їх до специфічних приєднуються до описаних тут поліпептидів. Для клітин, тканин, або макромолекул можна конденвизначення регіонів, що мають найвищий антигенсува ти допоміжні домени. Наприклад, поліпептид ний потенціал, можна використовувати профіль або білок ліганду zalpha11 можна спрямувати до гідрофільності за Хоппом/Вудсом [Норр et al., попередньо визначеного типу клітин конденсуван1981, вище, та Норр, 1986, вище]. У ліганді ням поліпептиду ліганду zalpha11 з лігандом, що zalpha11 ці регіони включають: амінокислотні заспецифічно приєднується до рецептору таких цілишки 114-119, 101-105, 126-131, 113-118, та 158льових клітин. Таким чином поліпептиди та білки 162 послідовності SEQ ID NO: 2. можна спрямовувати на терапевтичні або діагносАнтигенні пептиди та поліпептиди, що несуть тичні призначення. Поліпептид ліганду zalpha11 епітоп, містять переважно щонайменше 4-10 аміможна конденсувати з двома чи більше частинанокислот, щонайменше 10-14 амінокислот, або ми, як-то афінною міткою для очистки та спрямоприблизно 14-30 амінокислот послідовностей SEQ вуючим доменом. Поліпептидні конденсати можуть ID NO: 2 або SEQ ID NO: 56. Такі пептиди та політакож включати один чи більше сайтів розщепленпептиди, що несуть епітоп, можна продукувати ня, зокрема між доменами. [Дивися, Tuan et al. фрагментуванням поліпептиду ліганду zalpha11, Connective Tissue Research 34: 1-9,1996]. або хімічним пептидним синтезом, як тут описано. Використовуючи обговорені тут способи, переБільш того, епітопи можна селектувати фаговим січний фахівець може ідентифікувати та/або виговиявленням випадкових пептидних бібліотек [дитовити ряд поліпептидів, що мають по суті подібну вися, наприклад. Lane та Stephen, Curr. Opin. природу послідовності стосовно залишків 1-162 Immunol. 5: 268 (1993); та Cortese et al., Curr. Opin. або 33-162 послідовності SEQ ED NO: 2, або їх Biotechnol. 7: 616 (1996)]. Стандартні способи ідефункціональні фрагменти та конденсати, причому нтифікування епітопів та продукування антитіл з такі поліпептиди або фрагменти чи конденсати малих пептидів, що включають епітоп, описано, залишають властивості нативного білку, як-то зданаприклад, [Mole, "Epitope Mapping," у Methods in тність стимулювати проліферацію, диференціацію, Molecular Biology. T. 10, Manson (ed.), стор.105-116 індукувати спеціалізовану клітинну функцію або 35 84387 36 приєднуватися до рецептору zalpha11 або антитіл тиди можна зробити такими, що секреторна сигнадо ліганду zalpha11. льна послідовність, похідна від амінокислотних Поліпептиди ліганду zalpha11 представленого залишків 1-31 послідовності SEQ ID NO: 2, буде винаходу, включаючи поліпептиди повної довжиоперативно зв'язаною з послідовністю ДНК, що ни, функціональні фрагменти та конденсовані покодує інший поліпептид, використовуючи способи, ліпептиди, можна продукувати у генетично сконстщо відомі в рівні техніки та розкритітут. Секреторруйованих клітинах хазяїна звичайними на сигнальна послідовність, що міститься у конспособами. Придатними клітинами хазяїна є клітиденсованих поліпептидах представленого винахони такого типу, що можна трансформувати або ду, переважно конденсована з додатковим трансфектувати з екзогенною ДНК та вирощува ти пептидом для спрямування додаткового пептиду у у культурі, та включають клітини бактерій, грибів, секреторний шлях обміну амінотермінально. Такі та культивовані клітини вищих евкаріотів. Клітини констракти мають ряд застосувань, що відомі в евкаріотів, зокрема культивовані клітини багаторівні техніки. Наприклад, ці конденсовані констракклітинних організмів, є кращими. Способи обробки ти нової секреторної сигнальної послідовності моклонованих молекул ДНК та введення екзогенної жуть викликати секрецію активного компоненту ДНК у ряд клітин хазяїна розкрито [Sambrook et al. звичайно несекретованого білку. Такі конденсати Molecular Cloning: A Laboratory Manual, 2nd ed., можна використовувати in vivo або in vitro для Cold Spring Harbor Laboratory Press, Cold Spring спрямування пептидів через секреторний шлях Harbor, NY, 1989, та Ausubel et al., eds. Current обміну. Protocols in Methods in Molecular BjoIorv, John Wiley Культивовані клітини ссавців є придатними ха& Sons, Inc., NY, 1987]. зяями в рамках представленого винаходу. СпосоЗагалом послідовність ДНК, що кодує поліпепби уведення екзогенної ДНК у клітини ссавцятид ліганду zalpha11 оперативно зв'язана з іншими хазяїна включають опосередковану фосфатом генетичними елементами, що потрібні для її екскальцію трансфекцію [Wigler et al., Cell 14:725, пресії, включаючи у екс-пресійному векторі, зви1978; Corsaro та Pearson, Somatic Cell Genetics чайно, промотер та термінатор транскрипції. Век7:603. 1981; Graham and Van der Eb, Virology тор також звичайно містить один чи більше 52:456, 1973], електропорацію [Neumann et al., придатних для селекції маркерів та один чи більше EMBO J.1:841-45, 1982], опосередковану DEAEініціаторів реплікації, хоча фахівці здогадаються, декстраном [Ausubel et al., вище] та опосередкощо у деяких системах придатні для селекції марвану ліпосомами трансфекщю [Hawley-Nelson et кери можна запровадити на окремих векторах, а al., Focus 15:73, 1993; Ciccarone et al., Focus 15:80. реплікацію екзогенної ДНК можна запровадити 1993] та вектори вірусів [Miller та Rosman, інтегруванням у геном клітини хазяїна. Селекція BioTechniqoues 7: 980-90. 1989: Wang та Finer, промотерів, термінаторів, придатних для селекції Nature Med. 2: 714-6, 1996]. Продукування рекоммаркерів, векторів та інших елементів є предмебінантного поліпептиду у к ультивованих клітинах том стандартного конструювання, знаного пересіссавців розкрито, наприклад, [Levinson et al., Пачними фахівцями. Багато таких елементів описано тент США №4713339; Hagen et al., Патент США в літературі та доступно від комерційних постача№4784950; Palmiter et al., Патент США №4579821; льників. та Ringold, Патент США №4656134]. Придатні Для спрямування поліпептиду ліганду zalpha11 культивовані клітини ссавців включають COS 1 у секреторний шлях обміну клітин хазяїна у екс(ATCC №. CRL 1650), COS-7 (ATCC №. CRL 1651), пресійний вектор запроваджують секреторну сигBHK (ATCC №. CRL 1632), BHK 570 (ATCC №. CRL нальну послідовність, (також відому як лідерну 10314), 293 (ATCC №. CRL 1573; [Graham et al., J. послідовність, препропослідовність або препосліGen. Virol. 36:59-72. 1977]), Jurkat (ATCC №. CRLдовність). Секреторна сигнальна послідовність 8129), BaF3 (лінія похідних з кісткового мозку миші може бути від ліганду zalpha11 або похідною з залежних від інтерлейкіну-3 прелімфоїдних клітин. іншого секретова-ного білку (наприклад, t-PA) або [Дивися, Palacios та Steinmetz, Cell 41: 727-34, синтезованою de novo. Секреторна сигнальна по1985; Mathey-Prevot et al., Mol. Cell. Biol. 6: 4133-5, слідовність оперативно зв'язана з послідовністю 1986] та лінії клітин яєчнику китайського хом'яка ДНК ліганду zalpha11, тобто, дві послідовності об'(наприклад. CHO-Kl; ATCC №. CCL 61). Додаткові єднують у коректній рамці зчитування та позиціопридатні лінії клітин відомі в рівні техніки та достунують для спрямування наново синтезованого пні з загальнодоступних депозитаріїв, як-то поліпептиду у секреторний шлях обміну клітин American Type Culture Collection (Американська хазяїна. Секреторні сигнальні послідовності звиколекція типів культур), Rockville, Maryland. Взагачайно 5'-позиціонують стосовно послідовності лі, кращими є такі сильні промотери транскрипції, ДНК, що кодує потрібні поліпептиди, хоча деякі як промотери від SV-40 або цитомегаловірусу. секреторні сигнальні послідовності можна позиціо[Дивися, наприклад, Патент США №4956288]. Інші нувати в іншому місці потрібної послідовності ДНК придатні промотери включають промотери з мета[дивися, наприклад, Welch et al., патент США лотіонеїнових генів [Патенти США №№. 4579821 №5037743; Holland et al., патент США №5143830]. та 4601978] та головний пізній промотер аденовіАльтернативно, секреторну сигнальну послірусу. довність, що міститься у поліпептидах представСелекцію ліками звичайно використовують для леного винаходу, використовують для спрямуванвибору культивованих клітин ссавців в які уведено ня інших поліпептидів у секреторний шлях обміну. чужинну ДНК. Такі клітини звичайно позначають як Згідно з винаходом запропоновано такі конденсо"трансфектанти". Клітини, що культивовані у. привані поліпептиди. Сигнальні конденсовані поліпепсутності засобу селекції та здатні передавати по 37 84387 38 трібний ген своєму потомству, позначають як "стаекспресії секрето-ваних білків. [Дивися, Hill-Perkins більні трансфектанти". Кращим придатним до сета Possee, J. Gen. Virol. 71:971-6, 1990; Bonning, et лекції маркером є ген, що кодує резистентність до al., L Gen. Virol. 75:1551-6. 1994; та Chazenbalk and антибіотику неоміцин. Селекцію проводять у приRapoport, J. Biol. Chern. 270:1543-9, 1995]. У таких сутності ліків типу неоміцину, як-то G-418 або поконстрактах вектору переносу можна використовудібних. Селекційні системи можна також викорисвати короткі або довгі варіанти основного промотовувати для збільшення рівня експресії теру білку. Більш того, вектори переносу можна потрібного гена, процес позначають як "ампліфікасконструювати так, щоб замінити нативну секретоцію." Ампліфікацію проводять культивуванням рну сигнальну послідовність ліганду zalpha11 сектрансфектантів у присутності низької кількості зареторною сигнальною послідовністю, що походить собу, а потім підвищують кількість засобу селекції від білку комах. Наприклад, секреторну сигнальну для вибору клітин, що продукують високі рівні пропослідовність з екдистероїд-глюкозилтрансферази дукту уведеного гена Кращим придатним для амп(EGT), мелітин з бджолиного меду (Melittin, ліфікації та селекції маркером є дигідрофолатInvitrogen, Carlsbad, CA), або бакуловірус gp67 редуктаза, яка надає резистентності до метатрек(PharMingen, San Diego, CA) можна використовусату. Інші гени резистентності до ліків (наприклад, вати у констрактах для заміни нативної секреторрезистентності до гігроміцину, резистентності до ної сигнальної послідовності ліганду zalpha11. На багатьох ліків, пуроміцин-ацетилтрансферазу) додаток, вектори переносу можуть включати в можна використовувати також. Альтернативні марамці генетичного коду конденсат з ДНК, що кодує ркери, що представляють змінений фенотип, білок епітопну мітку на C- або N- закінченні поліпептиду зеленої флуоресценції, або білки поверхні клітин, ліганду zalpha11, наприклад, епітопну мітку Gluяк-то CD4, CD8, класу І MHC, плацентарну лужну Glu [Grussenmeyer et al., Proc. Natl. Acad. Sci. фосфа тазу, можна використовува ти для сорту82:7952-4, 1985]. Використовуючи відомі в рівні вання трансфектованих клітин від нетрансфектотехніки способи, вектор переносу, що містить ліваних клітин такими засобами, як FACSганд zalpha11 трансформують в E. coli, та скринусортування або спосіб сепарації магнітними кульють на басміди, що містять переривчастий ген lacZ ками. як свідоцтво рекомбінантного бакуловірусу. БасміМожна використовувати як хазяїв також клітидну ДНК, що міститься в рекомбінантному бакулони інших вищи х евкаріотів, включаючи клітини ровірусному геномі, виділяють, використовуючи звислин, клітини комах та клітини птиць. Використанчайні способи, та використовують для трансфекції ня Agrobacterium rhizogenes як вектору для клітин Spodoptera frugiperda, наприклад, клітин експресії гена в клітинах рослин обговорено Sf9. Далі продукується рекомбінантний вірус, що [Sinkar et al.. J. Biosci. (Bangalore) 11:47-58. 1987]. експресує ліганд zalpha11. Рекомбінантні вірусні Трансформація клітин комах та продукування ништами виробляють звичайно використовуваними в ми чужинного поліпептиду розкрито [Guarino et al., рівні техніки способами. Патент США №5162222 та публікація WIPO WO Рекомбінантний вірус використовують для ін94/06463]. Клітини комах можна інфікувати рекомфікування клітин хазяїна, звичайно похідних від бінантним бакуловірусом, похідним, звичайно, від осінніх маршових хробаків, Spodoptera frugriperda, Autographa californica nuclear polyhedrosis virus лінії клітин. [Дивися, взагалі, Click та Pasternak, (AcNPV). [Ди вися, King L.A. & Possee R.D., The Molecular Biotechnology: Principles and Applications Baculovirus Expression System: A Laboratory Guide, of Recombinant ДНК, ASM Press, Washington, D.C, London, Chapman & Hall; O'Reilly et al., Baculovirus 1994. Іншими придатними лініями клітин є лінія Expression Vectors: A Laboratory Manual. New York, клітин High FiveOÔ (Invifcrogen), що походить від Oxford University Press., 1994; та Richardson, Ed., Trichoplusia nі (Патент США №5300435)]. Baculovirus Expression Protocols. Methods in Клітини грибів, включаючи клітини дріжджів, Molecular Biology. Totowa. NJ, Humana Press, можна також використовувати згідно з винаходом. 1995]. Другий спосіб створення рекомбінантного Особливо цікаві, в цьому плані види дріжджів бакуловірусу використовує систему на базі трансвключають Saccharomyces cerevisiae, Pichia позону, що описана Луковом [Luckow, et al., J pastoris, та Pichia methanolica. Способи трансфорVirol., 67:4566-79, 1993]. Ця система, що застосомації клітин S. cerevisiae екзогенною ДНК та провує вектори переносу, продається в комплекті Bacдукування з неї рекомбінантного поліпептиду розto-BacÔ [Life Technologies, Rockville, MD]. Ця сискрито, наприклад, [Kawasaki, Патент США №4599311; Kawasaki et al., Патент США тема застосовує вектор переносу, pFastBadÔ [Life №4931373; Brake, Патент США №4870008; Welch Technologies], що містить транс-позон Тn7, для et al., Патент США №5037743; та Murray et al., Папереносу ДНК, що кодує поліпептид ліганду тент США №4845075]. Трансформовані клітини zalpha11, у геном бакуловірусу, що міститься в E. селектують за визначеним придатним для селекції соli, як велика плазміда під назвою "басміда". Векмаркером фенотипом, звичайно резистентності до тор переносу, pFastBadÔ використовує поліедриліків або здатності до вирощування за відсутності новий промотер AcNPV для запуску експресії попевної живильної речовини (наприклад, лейцину). трібного гена, в цьому випадку ліганду zalpha11. Переважною векторною системою для викорисОднак, pFastBadÔ можна значною мірою модифітання у Saccharomyces cerevisiae є розкрита кувати. Поліедриновий промотер можна видалити [Kawasaki et al. (Патент США №4931373)] векторна та замінити основним промотером білку бакуловісистема РОТІ, яка дозволяє селектувати трансфорусу (також відомий як промотер Pсоr, р6,9 або рмовані клітини вирощуванням у середовищі з MP), що експресований раніше у бакуловірусній вмістом глюкози. Придатні промотери та термінаінфекції, та має, як було показано, переваги при 39 84387 40 тори для використання у дріжджах включають ностей ДНК добре відомі в рівні техніки [дивися, промотери та термінатори генів гліколітичних фенаприклад, Sambrook et al., вище]. При експресії рментів [дивися, наприклад. Kawasaki, Патент поліпептиду ліганду zalpha11 у таких бактеріях, як США №4599311; Kingsman et al., Патент США E. coli, поліпептид може залишатися у цитоплазмі, №4615974; та Bitter, Патент США №4977092] та звичайно як нерозчинні гранули, або може бути генів алкоголь-дегідрогенази. [Дивися також Патеспрямованим у периплазматичні проміжки послінти США №№. 4990446; 5063154; 5139936 та довністю бактеріальної секреції. В попередньому 4661454]. Відомими в рівні техніки є трансформавипадку, клітини лізують, а гранули відбирають і ційні системи для інших дріжджів, включаючи денатурують, використовуючи, наприклад, ізотіоHansenula polymorpha, Schizosaccharomyces ціанат гуанідину або сечовину. Денатурований pombe, Kluyveromyces Iactis, Kluyveromyces fragills, поліпептид можна далі переукласти та димеризуUstilago maydis, Pichia paskons, Pichia methanolica, вати розбавленням денатуруючого засобу, як-то Pichia gnillerinondii та Candida maltosa. [Дивися, діалізом проти розчину сечовини та комбінації віднаприклад, Gleeson et al., J. Gen. Microbiol. новленого та окисненого глутатіону, з наступним 132:3459-65, 1986 та Cregg, Патент США діалізом проти буферованого сольового розчину. №4882279]. Згідно зі способом [McKnight et al., В останньому випадку поліпептид можна виділити Патент США №4935349, можна використовувати з периплазматичного проміжку у розчинній та функлітини Aspergillus]. Способи трансформації кціональній формі розщепленням клітин (наприAcremoniuin chrysogenum розкрито [Sumino et al., клад, обробкою ультразвуком або осмотичним Патент США №5162228]. Способи трансформації шоком) для вивільнення вмісту периплазматичноNeurospora розкрито [Lambowitz, Патент США го проміжку та витягу білку, таким чином уникаючи №4486533]. необхідності денатурації та переукладки. Використання Pichia methanolica як хазяїна Трансформовані та трансфектовані клітини для продукування рекомбінантного білку розкрито культивують звичайними способами в культивав [інтернаціональних публікаціях №№. WO ційному середовищі, що містить живильні речови97/17450, WO 97/17451, WO 98/02536 та WO ни та інші потрібні для вирощування вибраних клі98/02565]. Молекули ДНК для використання у тин хазяїна компоненти. В рівні техніки відомо трансформації P. methanolica виготовляють звибагато придатних середовищ, включаючи стандачайно як дволанцюгові циклічні плазміди, які перертні та комплексні середовища, вони звичайно важно перед трансформацією лінеаризують. Для включають джерело карбону, джерело нітрогену, продукування поліпептиду у P. methanolica, краще, незамінні амінокислоти, вітаміни та мінерали. Сещоб промотер та термінатор були промотером та редовища можуть також містити такі компоненти термінатором з гена P. methanolica, як-то гена P. як фактори росту або сироватки, за потребою. methanolica утилізації спирту {AUGl або AUG2}. Середовища для вирощування звичайно вибираІнші корисні промотери включають про-мотери ють для клітин, що містять екзогенно додану ДНК, генів дигідроксіацетон-синтази (DHAS), форміатнаприклад, селекцією за допомогою ліків або недегідрогенази (FMD), та каталази (CAT). Для полестачі необхідних живильних речовин, які доповнені гшення інтеграції ДНК у хромосому хазяїна краще придатним для селекції маркером, який входить в мати суцільний фланкований з обох кінців посліекспресійний вектор або є співтрансфектованим у довностями ДНК хазяїна експресійний сегмент клітину хазяїна. Клітини P. methanolica культивуплазміди. Кращим придатним для селекції маркеють у середовищі, що містить, адекватні джерела ром для використання у Pichia methanolica є ген P. карбону, нітрогену та слідови х живильних речовин, methanolica, ADE2, що кодує фосфорибозил-5при температурі приблизно 25-35°С. Рідкі культури аміноімідазол-карбоксилазу (AIRC; EC 4.1.1.21), забезпечують достатньою аерацією звичайними що дозволяє клітинам хазяїна ade2 рости при відзасобами, як-то збовтування невеликих флаконів сутності аденіну. Для великомасштабного промисабо барботаж фер-менторів. Кращим культиваційлового процесу, де бажано мінімізувати викорисним середовищем для P. methanolica є YEPD (2% тання метанолу, краще використовувати клітини D-глюкози, 2% Пептону BactoÔ (Difco Laboratories, хазяїна, в яких обидва гени утилізації метанолу Detroit, MI), 1% дріжджового екстракту BactoÔ (AUG1 та AUG2) є видаленими. Для продукування (Difco Laboratories), 0,004% аденіну та 0.006% Lсекреторного білку кращими є позбавлені генів лейцину). вакуоля-рної протеази клітини хазяїна (РЕР4 та Краще очищати поліпептиди згідно з винахоPRB1). Для полегшення уведення плазміди з вмісдом до чистоти більше 80%, краще до чистоти том ДНК, що кодує потрібний поліпептид, у клітини більше 90%, ще краще до чистоти більше 95%, а P. methanolica використовують електропорацію. найкраще у фармацевтично чистому стані, тобто з Краще трансформувати клітини P. methanolica чистотою більше 99,9%, з огляду на забруднюючі електропорацією, використовуючи експоненціальмакромолекули, зокрема інші білки та нуклеїнові но згасаюче, пульсуюче електричне поле, що має кислоти, та позбавляти від інфекційних та пірогеннапруженість поля 2,5-4,5кВ/см, переважно приних агентів. Переважно, очищений поліпептид є по близно 3,75кВ/см, константу часу (t) 1-40 мілісесуті позбавленим інших поліпептидів, особливо кунд, найкраще приблизно 20 мілісекунд. інших поліпептидів тваринного походження. Прокаріотні клітини хазяїна, включаючи штами Експресовані рекомбінантні поліпептиди лігабактерій Escherichia coli, Bacillus та інших видів, в нду zalpha11 (чи химерні поліпептиди ліганду рамках винаходу також є корисними клітинами zalpha11) можна очищати, використовуючи фракхазяїна. Способи трансформації цих хазяїв та ексціонування та/або звичайні способи та середовипресування клонованих в них чужинних послідовща для очистки. Для фракціонування зразків мож 41 84387 42 на використовувати осадження сульфатом амонію поліпептиду та афінної мітки (наприклад, зв'язуюта кислотну або хаотропну екстракцію. Приклади чого мальтозу білку, імуногло-булінового домену). стадій очистки можуть включати гідроксіапатитну, Більш того, використовуючи способи що опивитіснення за розміром, FPLC (рідинну екс-прессано в області техніки, поліпептидні конденсати, хроматографію білків) та високоефективну рідинну або гібридні білки ліганду zalpha11, конструюють, хроматографію з оберненою фазою. Придатні анівикористовуючи регіони або домени ліганду онообмінні середовища включають дериватизоваzalpha11 згідно з винаходом у комбінації з регіоні декстрани, агарозу, целюлозу, поліакриламід, нами або доменами білків іншої родини цитокінів спеціалізовані види діоксиду силіцію тощо. Кралюдини (наприклад, інтерлейкінів або GM-CSF), щими є похідні PEl, DEAE, QAE та Q, а краще або гетерологічних білків [Sambrook et al., вище, усього DEAE Fast-Flow Sepharose (Pharmacia, Altschul et al., вище, Picard, Cur. Opin. Biology, 5: Piscataway, NJ). Приклади хроматографічних се511-5, 1994, та посилання у них]. Ці способи доредовищ включають дериватизовані фенілом, бузволяють визначення біологічної важливості більтилом, або октилом середовища, як-то Phenylших доменів або регіонів у потрібному поліпептиді. Sepharose FF (Pharmacia), Toyopearl butyl 650 Такі гібриди можуть змінювати кінетику реакцій, (Toso Haas, Montgomeryville, PA), Octyl-Sepharose зв'язування, звуження чи розширення специфічно(Pharmacia) тощо; або такі поліакрилові смоли, як сті субстрату, або змінювататканинну та клітинну Amberchrom CG 71 (Toso Haas) тощо. Придатні локалізацію поліпептиду, та можуть бути застосотверді підкладки включають скляні кульки, смоли ваними до поліпептидів невідомої структури. на базі діоксиду силіцію, целюлозні смоли, агарозКонденсовані білки можна виготовити відомині кульки, кульки з перехресно зшитої агарози, ми фахівцям способами виготовленням кожного полістирольні кульки, перехресно зшиті поліакрикомпоненту конденсованого білку та їх хімічним ламідні смоли тощо, які є нерозчинними в умовах сполученням. Альтернативно, полінуклеотид, що використання. Ці підкладки можна модифікувати кодує обидва компоненти конденсованого білку у реакційними групами, що дозволяють приєднувати підхожій рамці зчитування можна створювати, вибілки через аміногрупи, карбоксигрупи, сульфгідкористовуючи відомі способи та експресувати спорильні групи, гідроксигрупи та/або вуглеводні угрусобами, що тут описано. Наприклад, частину або пування. Приклади хімічних способів конденсації усі зі спіралей, що надають біологічну функцію, включають ціаногенбромідну активацію, Nможна обмінювати між лігандом zalpha11 предстагідроксисукцинамідну активацію, епоксидну активленого винаходу з функціонально еквівалентнивацію, суль фгідрильну активацію, гідра-зидну акми спіралями від іншого члену родини, як-то IL-15, тивацію, та карбокси- та амінопохідні для хімічних DL-2, IL-4 або GM-CSF. Такі компоненти включаспособів карбодіімідної конденсації. Ці та інші твеють, але без обмеження, секреторну сигнальну рді середовища добре відомі і широко використопослідовність; спіралі А, В, C, D; петлі А/В, В/С, вуються в рівні техніки та доступні від комерційних C/D цитокінів з чотирьох-спіральним жмутом. постачальників. Способи приєднання рецепторноТакі конденсовані білки, можна чекати, матиго поліпептиду до середовища підкладки добре муть біологічний функціональний профіль, що є відомі в рівні техніки. Вибір конкретних способів є таким же або подібним до поліпептидів представпредметом рутинного розрахунку та частково виленого винаходу або інших відомих білків родини значається властивостями вибраної підкладки. цитокінів з чотирьох-спіральним жмутом, залежно [Дивися, наприклад, Affinity Chromatography: від сконструйованого конденсату. Більш того, як Principles & Methods, Pharmacia LKB Biotechnology, тут розкрито, такі конденсовані білки можуть виявUppsala, Sweden, 1988]. ляти інші властивості. Поліпептиди згідно з винаходом можна видіСтандартні молекулярно-біологічні способи та лити на базі їх фізичних чи біохімічних властивосспособи клонування можна використовувати для тей. Наприклад, для очистки збагачених гістидиобміну еквівалентних доменів між поліпептидом ном білків включаючи ті, що містять ліганду zalpha11 та тими поліпептидами з якими полігістидинову мітку, можна використовувати вони конденсовані. Загалом, сегмент ДНК, що кохроматографію на базі адсорбції на іммобілізовадує потрібний домен, наприклад, спіралі A-D ліганих іонах металу (IMAC). Коротше, гель спочатку нду zalpha11, або інший описаний тут домен, опенасичують іонами двовалентного металу з утворативно зв'язаний у рамці зчитування ренням хелату [Sulkowski, Trends in Biochem., 3:1щонайменше з одним іншим сегментом ДНК, що 7, 1985]. Збагачені гістидином білки адсорбують на кодує додатковий поліпептид (наприклад домен цих матрицях з різною спорідненістю залежно від або регіон з іншого цитокіну, як-то EL-2, або подібвикористовуваного іону металу та елююють конкуного), та вставлений у прийнятний експресійний рентним елююванням, зниженням рН, або викоривектор, як описано тут. Загалом констракти ДНК станням сильних хелатуючи х засобів. Інші способи роблять такими, щоб кілька сегментів ДНК, що очистки включають очистку глікозилованих білків кодують відповідні регіони поліпептиду, що операхроматографією на базі спорідненості до лецитину тивно зв'язані у рамці зчитування, утворювали та іонообмінною хроматографією [Methods in одиничний констракт, що кодує суцільний конденEnzymol., Vol.182, "Guide to Protein Purification" сований білок, або його функціональну частину. (Посібник з очистки білків), M. Deutscher, (ed.), Наприклад, констракт ДНК міг би кодувати від NAcad. Press, San Diego, 1990, pp.529-39]. Згідно з закінчення до С-закінчення конденсований білок, додатковим втіленням винаходу для полегшення що містить сигнальні поліпептиди з наступним очистки можна створити конденсати потрібного розвиненим конденсованим білком цитокіну з чотирьох-спіральним жмутом, що містить спіраль А, 43 84387 44 далі спіраль В, далі спіраль C і далі спіраль D. Такі Дослідження з виміру диференціації включають, конденсовані білки можна експресувати, виділяти, наприклад, визначення маркерів поверхні клітин, та досліджувати на активність як тут описано. пов'язаних з фазо-специфічною експресією тканиПоліпептиди ліганду zalpha11 або їх фрагменни, ферментною активністю, функціональною акти можна також виготовити хімічним синтезом, тивністю або морфологічними змінами [Watt, поліпептиди ліганду zalpha11 можуть бути моноFASEB, 5: 281-284, 1991; Francis, Differentiation 57: мерами або мультимерами; глікозилованими або 63-75, 1994; Raes, Adv. Anim. Cell Biol. Technol. неглікозилованими; обробленими PEG або необBioprocesses, 161-171, 1989; усі представлені тут робленими PEG; та можуть включати чи не вклюяк посилання]. чати початковий метіоніновий амінокислотний заМолекули представленого винаходу можна лишок. Наприклад, поліпептиди можна виготовити досліджувати in vivo, використовуючи вірусні систвердофазним пептидним синтезом, наприклад, як теми доставки. Приклади вірусів для цієї мети описано [Merrifield, J. Am. Chem. Soc. 85: 2149, включають аденовірус, вірус герпесу, ретровіруси, 1963]. вірус коров'ячої віспи, та адено-асоційований вірус Активність молекул представленого винаходу (AAV). Аденовірус, вірус з дволанцюговою ДНК, є можна вимірювати, використовуючи ряд дослізараз найкраще дослідженим вектором переносу джень, що вимірюють проліферацію клітин та/або гену для доставки гетерологічних нуклеїнових кисзв'язування з клітинами, що експресують рецептор лот [для огляду, дивися Т.С. Becker et al., Meth. zalpha11. Особливо цікавими є заміни у залежних Cell Biol. 43: 161-89, 1994; та J.T. Douglas & D.T. від ліганду zalpha11 клітинах. Придатні лінії клітин, C.Uriel, Science & Medicine 4: 44-53, 1997]. що сконструйовані як залежні від ліганду включаЯк ліганду, активність поліпептиду ліганду ють залежну від IL-3 лінію клітин BaF3 [Palacios та zalpha11 можна вимірювати біосенсор-ним мікроSteinmetz, Cell 4J.: 727-734, 1985; Mathey-Prevot et фізіометром на базі силіцію, який вимірює асоціal., Mol. Cell. Biol. 6: 4133-4135, 1986], FDC-P1 йовані зі зв'язуванням рецептору та наступною [Hapel et al,. Blood 64: 786-790, 1984], та MO7e фізіологічною реакцією клітини величини екстра[Kiss et al., Leukemia 7: 235-240, 1993]. Залежні від целюлярної ацидифіка-ції або виділення протонів. фактору росту лінії клітин можна встановити згідно Прикладом пристрою є мікрофізіометр з опублікованими способами [наприклад, CytosensorÔ , що виготовлений Molecular devices, Greenberger et al., Leukemia Res. 8: 363-375, 1984; Sunnyvale, CA. Багато клітинних реакцій, як-то Dexter et al., у Baum et al. Eds., Experimental проліферація клітин, транспорт іонів, продукуванHematology Today, 8th Ann. Mtg. Int. Soc. Exp. ня енергії, реакція запалення, регуляторна та реHematol. 1979, 145-156, 1980]. цепторна активація тощо, можна виміряти цим Білки представленого винаходу є корисними способом. [Дивися, наприклад, McConnell et al., для стимулювання проліферації, активації, дифеScience 257:1906-12, 1992; Pitchford et al., Meth. ренціації та/або індукції або інгібування спеціаліEnzymol. 228:84-108, 1997; Arimilli et al., J. Immunol. зованої клітинної функції клітини, задіяної у гомеоMeth. 212:49-59, 1998; Van Liefde et al., Eur. J. стазі кровотворення та імунної функції. Зокрема, Pharmacol. 346:87-95, 1998]. поліпептиди ліганду zalpha11 є корисними для Більш того, ліганд zalpha11 можна використостимулювання проліферації, активації, диференцівувати для ідентифікації клітин, тканин чи ліній ації, індукції або інгібування спеціалізованої кліклітин, що відповідають стимульованому лігандом тинної функції клітин кровотворних ліній диференzalpha11 шляху обміну. Вищеописаний мікрофізіоціювання, включаючи, але без обмеження, Тметр можна використовувати для швидкої ідентиклітини, В-клітини, NK-клітини, дендритні клітини, фікації чутливи х до ліганду клітин, як-то клітин, моноцити, та макрофаги, а також епітеліальні клічутливи х до ліганду zalpha11 представленого витини. Проліферацію та/або диференціацію кровонаходу. Клітини можна культивувати у присутності творних клітин можна вимірювати in vitro, викорисабо відсутності поліпептидів ліганду zalpha11. Ті товуючи культивовані клітини, або застосуванням клітини, що викликають вимірну зміну у екстрацеin vivo молекул згідно з винаходом до прийнятної люлярній ацидифікації у присутності ліганду тваринної моделі. Дослідження з виміру проліфеzalpha11, є чутливими до ліганду zalpha11. Такі рації або диференціації клітин добре відомі в рівні клітини або лінії клітин можна використовувати техніки. Наприклад, дослідження з виміру пролідля ідентифікації антагоністів та агоністів описаноферації включають такий аналіз, як хемочутлиго вище поліпептиду ліганду zalpha11. вість до барвника нейтральний червоний З точки зору розподілу лігандів по тканинам, [Cavanaugh et al., Investigational New Drugs 8: 347що спостерігають для рецептору zalpha11 агоністи 354, 1990, представлено тут як посилання], вклю(включаючи нативний ліганд чення радіоактивно мічених нуклеотидів [Cook et zalpha11/субстрат/кофактор/тощо) та/або антагоal. Analytical Biochem. 179: 1-7. 1989, представлено ністи мають величезний потенціал при застосутут як посилання], включення 5-бром-2'ванняхin vitro та in viva. Сполуки, що ідентифікодезоксіуридину (BrdU) у ДНК проліферуючих клівані як агоністи ліганду zalpha11, є корисними для тин [Porstmann et al., J. Immunol. Methods 82: 169вирощування, проліферації, активації, диференці179, 1985, представлено тут як посилання], та виації, та/або індукції або інгібування спеціалізовакористання солі тетразоліуму [Mosmann, J. них клітинних функцій клітини, задіяної у гомеоImmunol. Methods 65: 55-63, 1983; Alley et al. Рак стазі кровотворення та імунної функції. Наприклад, Res. 48: 589-601. 1988; Marshall et al., Growth Reg. ліганд zalpha11 та сполуки агоністів є корисними 5:69-84, 1995; та Scudiero et al. Рак Res. 48: 4827як компоненти визначених клітинних культивацій4833, 1988; усі представлені тут як посилання]. них середовищ, і їх можна використовувати по 45 84387 46 одинці або у комбінації з іншими цитокінами та ропні засоби (гуанідин-НСІ), або зміни рН, щоб гормонами для заміни сироватки, що є звичайно порушити приєднання ліганду до рецептору. використовуваною у культурі клітин. Агоністи є Переважно можна застосовувати систему дотому корисними при специфічному стимулюванні слідження, що використовує рецептор (або антитіросту та/або розвитку Т-клітин, В-клітин, NK-клітин, ло, один член пари комплемент-антикомплемент), цитотоксичних лімфоциті, та інши х клітин лімщо зв'язує ліганд, або його зв'язуючий фрагмент, фоїдних та мієлоїдних ліній диференціювання у та комерційно доступний біосенсорний прилад культурі. (BIAcore, Pharmacia Biosensor, Piscataway, NJ). Антагоністи також є корисними як реагенти Такий рецептор, антитіло, член пари комплементпри дослідженнях для характеризування сайтів антикомплемент або фрагмент іммобілізують на взаємодії ліганд-рецептор. Антагоністи є корисниповерхні рецепторного чіпа. Використання цього ми для інгібування вирощування, проліферації, приладу розкрито [Karlsson, J. Immunol. Methods активації, диференціації та/або диференціації клі145: 229-40, 1991 та Cunningham та Wells, J. Mol. тин, що залучено у регулюванні кровотворення. Biol. 234: 554-63, 1993]. Рецептор, антитіло, член Інгібітори активності ліганду zalpha11 (антагоністи або фрагмент є приєднують ковалентно, викорисліганду zalpha11) включають антитіла проти лігантовуючи хімію амінів або сульфгідрильну хімію, на ду zalpha11 та розчинні рецептори ліганду декстранові волокна, які нанесено на плівку золота zalpha11, а також інші пептидні та непептидні агев проточній комірці. Тест-зразок пропускають ченти (включаючи рибосоми), ліганд zalpha11 можна рез комірку. Якщо у зразку наявні ліганд, епітоп, також використовувати для ідентифікації інгібіторів або протилежний член пари комплемент(антагоністів) їх активності. Тест-сполуки додають антикомплемент, він зв'язується з іммобілізованим до досліджених розкритих тут для ідентифікації рецептором, антитілом або членом, викликаючи, сполук, що інгібують активність ліганду zalpha11. відповідно, зміну коефіцієнту рефракції середовиHa додаток до цих досліджень, розкритих тут, зраща, яке детектується як зміна резонансу поверхзки можна досліджувати на інгібування активності невого плазмону золотої плівки. Ця система доліганду zalpha11 у ряді досліджень, призначених зволяє визначення верхньої та нижньої меж для виміру приєднання рецептору, стимулявеличини, за якою можна розрахувати споріднеції/інгібування залежних від ліганду zalpha11 кліність до зв'язування, та оцінити стехіометрію зв'ятинних реакцій або проліферації експресуючих зування. Альтернативно, зв'язування ліганду з розчинний рецептор zalpha11 клітин. рецептором можна аналізувати, використовуючи Поліпептид ліганду zalpha11 можна експресуспосіб SELDI(TM) (Ciphergen, Inc., Palo Alto, C A). вати як конденсат з постійним регіоном важкого Ліганд-зв'язуючий рецепторний поліпептид ланцюга імуноглобуліну, звичайно фрагменту Fc, можна також використовувати в інши х відомих в що має два домени постійного регіону та втрачерівні техніки досліджуваних системах. Такі системи ний змінний регіон. Способи виготовлення таких включають аналіз за Скат-чардом (Scatchard) на конденсатів розкрито у [патентах США №№. визначення спорідненості до зв'язування [дивися, 5155027 та 5567584]. Такі конденсати звичайно Scatchard, Ann. NY Acad. Sci. 51: 660-72, 1949] та секретуються як мультимерні молекули причому калориметричний аналіз [Cunningham et al., частини Fc зв'язані одна з одною дисульфідними Science 253:545-48, 1991; Cunningham et al., зв'язками, а два поліпептиди не-Ig є розташовані Science 245:821-25, 1991]. поблизу один від одного. Конденсати цього типу Поліпептиди ліганду zalpha11 можна також виможна використовувати, наприклад, для димерикористовувати для виготовлення антитіл, що призації, збільшуючи стабільність та період напіввиєднуються до епітопів, пептидів або поліпептидів далення in vivo, для а фінної очистки ліганду, як ліганду zalpha11. Поліпептид ліганду zalpha11 або інструмент дослідження in vitro, або антагоніст. його фрагмент слугує антигеном (імуногеном) для Для використання у дослідженнях, химери приєдщеплення тварин та виклику імунної реакції. Перенують до підкладки регіоном Fc та використовують січний фахівець зрозуміє, що антигенні поліпептиу форматі ELISA. ди, що несуть епітоп, мають послідовність щонайПоліпептиди, що зв'язують ліганд zalpha11, менше з 6, краще щонайменше з 9, та ще краще можна також використовувати для очистки ліганду. приблизно з 15-30 суміжних амінокислотних залиПоліпептиди іммобілізують на твердій підкладці, шків поліпептиду ліганду zalpha11 (наприклад, як-то кульках агарози, пе- -рехресно-зв'язаній агаSEQ ID NO: 2). Включеними є поліпептиди, що розі, склі,-целюлозних смолах, смолах на базі симістять більшу частину поліпептиду ліганду ліцію, полістиролі, перехресно-зв'язаному поліакzalpha11, тобто, від 30-100 залишків до амінокисриламіді, або подібних матеріалах, що є стійкими в лотної послідовності повної довжини. Антигени умовах використання. Способи приєднання поліабо імуногенні епітопи можуть також включати пептидів до твердих підкладок відомі в рівні техніприєднані мітки, ад'юванти та носії, як тут описано. ки та включають хімію амінів, ціанобромідну актиПридатні антигени включають поліпептид ліганду вацію, N-гідроксисукцинімідну активацію, zalpha11, що зашифрований послідовністю SEQ ID епоксидну активацію, сульфгідрильну активацію, NO: 2 від амінокислоти №32 до амінокислоти та гідразиду активацію. Утворене середовище вза№162, або їх суміжних амінокислотних фрагментів галі оформлюють у формі колонки, а рідини, що 9-131. Інші придатні антигени включають, ліганд містять ліганд пропускають через колонку один чи повної довжини та розвинений ліганд zalpha11, більше разів, щоб дозволити ліганду приєднатися спіралі A-D, та окремі чи багато спіралей А, В, C, до рецепторного поліпептиду. Ліганд далі елююта D стр уктури ліганду zalpha11 з чотирьохють, використовуючи зміни концентрації солі, хаотспіральним жмутом, як тут описано. Кращими пеп 47 84387 48 тидами для використання в якості антигенів є гідWIPO WO 98/24893]. Краще, коли ендогенні гени рофільні пептиди, як-то ті, що спрогнозує пересічімуноглобуліну у цій тварині є інактивованими або ний фахівець за графіком гідрофобності, як тут видаленими, як-то гомологічною рекомбінацією. описано, наприклад, амінокислотні залишки 114Антитіла позначають як специфічно зв'язуючі, 119, 101-105, 126-131, 113-118, та 158-162 посліякщо: 1) вони виявляють пороговий рівень активдовності SEQ ID NO: 2. ності до зв'язування, та/або 2) вони не виявляють Антитіла від імунної реакції, створені щепленпомітних перехресних реакцій з молекулами споням тварини цими антигенами можна виділити та рідненого поліпептиду. Пороговий рівень зв'язуочистити як тут описано. Способи для виготовленвання визначають, якщо антитіла проти ліганду ня та виділення поліклональних та моноклональzalpha11 специфічно приєднуються до поліпептиних антитіл добре відомі в рівні техніки. [Дивися, ду, пептиду або епітопу ліганду zalpha11 зі споріднаприклад. Current Protocols in Immunology, неністю щонайменше у 10 разів більшою спорідCooligan, et al. (eds.), National Institutes of Health, неності до приєднання контрольних поліпептидів John Wiley та Sons. Inc., 1995; Sambrook et al., (не з ліганду zalpha11). Methods in Molecular Cloning: A Laboratory Manual, Краще, щоб антитіла виявляли спорідненість Second Edition. Cold Spring Harbor, NY, 1989; та до приєднання (Kа) 106моль -1 або більше, переваHurrell, J. G. R., Ed., Monoclonal Hvbridoma жно 107моль -1 або більше, краще 108моль -1 або Antibodies: Techniques та Applications. CRC Press, більше, а найкраще 109моль-1 або більше. СпорідInc., Boca Raton, FL, 1982]. неність до приєднання антитіла може легко визнаЯк повинен розуміти пересічний фахівець, почити пересічний фахівець, наприклад, аналізом ліклональні антитіла можна створювати щепленСкатчарда [Scatchard, Ann, NY Acad. Sci. 51: 660ням ряду теплокровних тварин як-то коней, корів, 672, 1949]. кіз, овець, собак, курей, кролів, мишей та щурів Чи не виявляють антитіла проти ліганду поліпептидом ліганду zalpha11 або його фрагменzalpha11 помітних перехресних реакцій з молекутом. Імуногенний поліпептид ліганду zalpha11 молами спорідненого поліпептиду, показано, наприжна посилювати використанням ад'юванту, як-то клад, детектуванням антитілами поліпептиду лігаалюм (гідроксид алюмінію) або повний або неповнду zalpha11 але не відомих споріднених ний ад'ювант Фрейнда. Поліпептиди, що корисні поліпептидів, використовуючи стандартний аналіз для імунізації також включають конденсовані поліВестерн-блотингом [Ausubel et al., вище]. Приклапептиди, як-то конденсати ліганду zalpha11 або ди відомих споріднених поліпептидів розкриті в його частини з імуноглобуліновим поліпептидом попередньому рівні техніки, як-то відомі ортологи, або зі зв'язуючим мальтозу білком. Поліпептидний паралоги, та подібні відомі члени білку родин. імуноген може бути молекулою повної довжини Скринінг можна також здійснити, використовуючи або її частиною. Якщо частина поліпептиду є "гапліганд zalpha11 не від людини, та мутантні поліпетеноподібною," таку частину можна переважно птиди ліганду zalpha11. Більш того, антитіла можоб'єднувати чи .зв'язувати з макромолекулярним на "скринувати проти" відомих споріднених поліносієм (як-то гемоціанін ключового блюдечка пептидів для ізоляції популяції, що специфічно (KLH), альбумін коров'ячої сироватки (BSA) або приєднується до ліганду zalpha11 людини. Напритоксоїд тетануса) для імунізації. клад, антитіла, що створені на ліганд zalpha11, Використаний тут термін "антитіла" включає адсорбуються на споріднених поліпептидах, які поліклональні антитіла, афінно очищені поліклоприкріплені до нерозчинної матриці, антитіла, що нальні антитіла, моноклональні антитіла, та антиспецифічні до поліпептидів ліганду zalpha11 люген-зв'язуючі фрагменти, як-то протеолітичні фрадини, протікатимуть через матрицю в належних гменти F(ab')2 та Fab. Також є включеними умовах буферування. Такий скринінг дозволяє генетично сконструйовані суцільні антитіла або ізоляцію поліклональних та моноклональних антифрагменти, як-то химерні антитіла, Fv-фрагменти, тіл, що не виявляють перехресних реакцій з блиантитіла з одиничним ланцюгом тощо, а також зько спорідненими поліпептидами [Antibodies: A синтетичні антиген-зв'язуючі пептиди та поліпепLaboratory Manual, Harlow and Lane (eds.), Cold тиди. Ан титіла не від людини можна пристосувати Spring Harbor Laboratory Press, 1988; Current до людини прищепленням тільки CDR не від люProtocols in Immunology, Coohgan, et al. (eds.). дини до рамки зчитування та незмінених регіонів National Institutes of Health, Fohn Wiley and Sons, людини, або вставкою суцільних змінних доменів Inc., 1995]. Скринінг та виділення специфічних анне від людини (як варіант, "покриваючи" їх людититіл є добре відомими в рівні техніки [дивися, ноподібною поверхнею заміщенням оголених заFundamental Immunology, Paul (eds.), Raven Press, лишків, результатом чого є "фанероване" антиті1993; Getzoff et al., Adv. in Immunol. 43: 1-98, 1988; ло). В деяких випадках, пристосовані до людини Monoclonal Antibodies: Principles and Practice. антитіла можуть зберігати залишки не від людини Goding, J.W. (eds.). Academic Press Ltd. 1996; у доменах рамок змінних регіонів людини для поBenjamin et al., Ann. Rev. Immunol. 2: 67-101, 1984]. силення належних характеристик зв'язування. Специфічне зв'язування антитіла проти ліганду Пристосуванням антитіл до людини можна збільzalpha11 можна детектувати рядом розкритих в шити період їх біологічного напіввидалення, а морівні техніки та показаних нижче способів. жливість шкідливих імунних реакцій при застосуРяд досліджень, що відомі фахівцям, можна ванні до людей зменшується. Більш того, антитіла використовува ти, для визначення антитіл, що прилюдини можна продукувати у трансгенній, нелюєднуються до білків або поліпептидів ліганду дяній тварині, що сконструйована з вмістом генів zalpha11. Типові дослідження детально описано у імуноглобуліну людини, як розкрито у [публікації [довіднику Antibodies: A Laboratory Manual, Harlow 49 84387 50 та Lane (Eds.), Cold Spring Harbor Laboratory Press. пари комплемент-антикомплемент є пара біо1988]. Репрезентативні приклади таких аналізів тин/стрептавідин. включають: паралельний імуноелектрофорез, раЗв'язуючі поліпептиди можуть також діяти як діоімуноаналіз, радіоімунопреципітацію, імуносор"антагоністи" ліганду zalpha11 для блокування бен-тний аналіз на зв'язаних ферментах (ELISA), приєднання ліганду zalpha11 тапередачі сигналу in аналіз дот-блотингом або Вестерн-блотингом, інгіvitro та in vivo. Ці поліпептиди проти приєднання бування або конкурентний аналіз, та сендвічліганду zalpha11 могли б бути корисними для інгіаналіз. На додаток, антитіла можна скринувати на бування активності ліганду zalpha11 або зв'язуприєднання білків або поліпептидів ліганду вання білку. zalpha11-нативного типу проти мутантних білків Конденсовані білки полі пептид-токсин або або поліпептидів. конденсовані білки антитіло-токсин можна викориАнтитіла до ліганду zalpha11 можна викорисстовувати для інгібування або видалення намічетовувати для мічених клітин, що екс-пресують ліних клітин або тканин (наприклад, для обробки ганд zalpha11; для виділення ліганду zalpha11 ракових клітин або тканин). Альтернативно, якщо афінною очисткою; для діагностичних досліджень полі пептид має багато функціональних доменів при визначенні рівнів поліпептиду ліганду (тобто, домен активації або домен приєднання, zalpha11, що знаходиться в обігу; для виявлення рецептору, плюс цільовий домен), конденсований або кількісного визначення розчинного ліганду білок, що включає тільки цільовий домен, може zalpha11 як маркеру - патології.або захворювання, бути придатним для спрямування виявлюваної що лежить в основі; у аналітичних способах із замолекули, цитотоксичної молекули або комплеместосуванням FACS; для скринінгу експресійних нтарної молекули до клітин або тканин потрібного бібліотек; для створення анти-ідіотипових антитіл; типу. У випадках, коли домен конденсованого білта як нейтралізуючі антитіла або як антагоністи ку включає тільки комплементарну молекулу, антидля блокування активності ліганду zalpha11 in vitro комплементарна молекула може бути сполученою та in vivo. Придатні безпосередні мітки або мічені з виявлюваною або цитотоксичною молекулою. атоми включають радіонукліди, ферменти, субТакі конденсованибілки з доменомстрати, кофактори, інгібітори, флуоресцентні маркомплементарної молекули,.отже, репрезентують кери, хемілюмінесцентні маркери, магнітні частинспільний цільовий переносник для клітики тощо; непрямі мітки або мічені атоми можуть но/тканино-специфічної доставки спільних антиграти головну роль при використанні біотинукомплементарно-виявлюваних/цитотоксичних споавідину або іншої пари комплементлучень молекул. антикомплемент як інтермедіатів. Анти тіла .тут Конденсовані білки ліганду zalpha11 цитокіну можна також безпосередньо або небезпосередньо або конденсовані білки антитіло-цитокін можна сполучати з ліками, токсинами, радіонуклідами використовува ти для посилення знищення in vivo тощо, і ці сполучення використовувати для діагноцільових тканин (наприклад, раку крові та кістковостичного або терапевтичного застосування in vivo. го мозку), якщо поліпептид ліганду zalpha11 або Більш того, антитіла до ліганду zalpha11 або його антитіло проти ліганду zalpha11 націлене на гіперфрагментів можна використовувати in vitro для проліферативну клітину крові чи кісткового мозку визначення виродженого ліганду zalpha11 або йо[Дивися, загалом, Homick et al. Blood 89: 4437-47, го фрагментів у дослідженнях, наприклад. Вес1997]. Описані конденсовані білки здатні націлютерн-блотингом або іншими відомими в рівні технівати цитокін на потрібну ділянку дії, забезпечуючи ки способами. тим збільшену місцеву концентрацію цитокіну. Придатні виявлювані молекули можуть бути Придатні поліпептиди ліганду zalpha11 або антитібезпосередньо або небезпосередньо приєднанила проти ліганду zalpha11 націлені на небажану ми до поліпептидів або антитіл, та включають раклітину або тканину (тобто, п ухлини або лейкемії), діонукліди, ферменти, субстрати, кофактори, інгіі конденсований цитокін опосередковує посилений бітори, флуоресцентні маркери, лізис цільових клітин ефекторними клітинами. хемілюмінесцентні маркери, магнітні частинки тоПридатні для цього цитокіни включають, наприщо. Придатні цитотоксичні молекули можуть бути клад, інтерлейкін 2 та фактор стимулювання колобезпосередньо або небезпосередньо приєднаниній гранулоцитів-макрофагів (GM-CSF). ми до поліпептидів або антитіл, і включають бакДиференціація є поступовим та динамічним теріальні або рослинні токсини (наприклад, токсин процесом, що починається плюрипотен-тними дифтерії, сапорин, екзотоксин Pseudomonas, ристовбурними клітинами та закінчується у кінці дицин, абрин тощо), а також терапевтичні радіонукференційованими клітинами. Плюрипотентні стовліди, як-то йод-131, реній-188 або ітрій-90 (приєдбурні клітини, що можуть регенерувати комітуваннані до поліпептидів чи антитіл безпосередньо або ня до лінії диференціювання, експресують ряд приєднані небезпосередньо, наприклад, через маркерів диференціювання, що втрачається, коли хелатні групи). Поліпептиди або антитіла можуть відбувається комітування до лінії диференціювантакож бути сполученими з цитотоксичними ліками, ня клітин. Вихідні клітини експресують ряд маркеяк-то адріаміцин. Для небезпосереднього приєдрів диференціювання, що можуть або ні продовнання виявлюваної або цитотоксичної молекули, жувати експресуватися при тому, що клітини виявлювана або цитотоксична молекула може просунулися по шляху обміну клітин лінії диферебути сполученою з членом пари комплементнціювання протягом повного розвитку. Маркери антикомплемент, де інший член приєднаний до диференціювання, що експресуються виключно частин поліпептидів або антитіл. Прикладом цієї розвиненими клітинами, мають звичайно функціональні властивості, як-то клітинних продуктів, фе 51 84387 52 рментів для продукування клітинних продуктів та збільшення, чи втрати експресії ліганду zalpha11 у рецепторів. Етап диференціації популяції клітин передракових або ракових умовах у порівнянні з відслідковують ідентифікацією присутніх у клітиннормальною тканиною, можуть слугувати як діагній популяції маркерів. ностика для трансформації, інвазії та метастазів Виявлені свідоцтва означають, що фактори, при прогресуванні пухлин. Як так, знання стадії які стимулюють специфічні типи клітин на шляху прогресування пухлини або її метастазів допомообміну при кінцевій диференціаці або дедиференже лікарю при виборі найпридатнішої терапії, або ціації впливають на чисті популяції клітин, що посуворості лікування для індивідуального ракового ходять зі звичайних попередників чи стовбурних пацієнта. Способи виміру збільшення чи втрати клітин. Отже, згідно з винаходом запропоновано експресії (мРНК або білку) добре відомі в рівні стимулювання або інгібування проліферації лімтехніки та описано тут, і їх можна застосовувати до фоїдних клітин, кровотворних клітин та епітеліальекспресії ліганду zalpha11. Наприклад, виявлення них клітин. або зникнення поліпептидів, що регулюють рухоЛіганд zalpha11 виділяли з тканини, відомої як мість клітин можна використовувати для допомоги маючу важливу імунологічну функцію, яка містить лікарю діагностувати та прогнозувати рак простати клітини, що грають роль у імунній системі. Ліганд [Banyard, J. та Zetter, B.R., Рак та Metast. Rev. j_7: zalpha11 експресується у СD3+-відібраних, акти449-458, 1999]. Як ефектор рухомості клітин, лівованих клітинах периферійної крові, і показано, ганд zalpha11 збільшення чи втрати експресії мощо експресія ліганду zalpha11 збільшується після жуть слугувати як діагностика для лімфоїдного, Вактивації Т-клітин. Більш того, результати експеклітинного, епітеліального, гематопое-тичного та риментів, що описано у розділі Приклади, демоніншого раку. струють, що поліпептиди представленого винахоБільш того, активність та вплив ліганду ду впливають на зростання/вирощування та/або zalpha11 на прогресування пухлини або її метастан диференціації NK-клітин або NKстази можна вимірювати in vivo. Розроблено кілька попередників. Додаткове свідоцтво демонструє, сингенних мишачих моделей для дослідження що ліганд zalpha11 впливає на проліферацію впливу поліпептидів, сполук або інших видів лікута/або диференціацію Т-клітин та В-клітин in vivo. вання на прогресування пухлини. У цих моделях, Фактори, що стимулюють проліферацію кровотвоклітини пухлини з культури імплантують у мишей рних попередників та активують розвинені клітини тієї ж породи, що і донор пухлини. Клітини розвизагалом відомі. NK-клітини є чутливими до IL-2 ватимуться у пухлини, що мають подібні характесамого по собі, але проліферація та активація заристики у мишах-реципієнтах, а у деяких моделях галом потребують додаткових факторів росту. Наз'являтимуться також метастази. Прийнятні моделі приклад, показано, що IL-7 та фактор Steel (ліганд пухлин для нашого вивчення включають, серед c-kit) потребували для утворення колоній NKіншого, карциному легенів Л'юїса (ATCC №CRLпопередників. IL-15+IL-2 у комбінації з IL-7 та фак1642) та меланому В-16 (ATCC №CRL-323). Звитором Steel були ефективнішими [Mrozek et al. чайно використовують лінії сингенних стосовно Blood 87: 2632-2640, 1996]. Однак, для проліферамишей C57BL6/J пухлин, які легко культивува ти та ції специфічних підкласів NK-клітин та/або NKманіпулювати з ними in vitro. Утворені від імпланпопередників можуть бути необхідними неідентитації кожної з цих ліній клітин пухлини здатні до фіковані цитокіни [Robertson et. al. Blood 76: 2451метастазів у ле гені у мишей C57BL6/J. Модель 2438, 1990]. Композиція, що містить ліганд карциноми легенів Л'юїса нещодавно використано zalpha11 та IL-15, стимулює NK-попередники та на мишах для ідентифікації інгібітору ангіогенезу NK-клітини з очевидністю, що ця композиція є по[O'Reilly MS, et al. Cell 79: 315-328,1994]. Мишей тужнішою за попередньо описані фактори та комC57BL6/J обробляли експериментальним агентом бінації факторів. через кожну добу ін'єкцією рекомбінантного білку, Виміри диференціації включають, наприклад, агоністу чи антагоністу, або одноразовою ін'єкцією вимір клітинних маркерів, пов'язаних зі специфічрекомбінантного аденовірусу. Через 3 доби після ною стосовно етапу експресією тканини, ферментобробки 105-106 клітин імплантують під шкіру спиною активністю, функціональною активністю або ни. Альтернативно, самі клітини можна інфікувати морфологічними змінами [Watt, FASEB, 5: 281-284, рекомбінантним аденовірусом, як-то тим, що екс1991; Francis, Differentiation 57: 63-75, 1994; Raes, пресує ліганд zalpha11, перед імплантацією, так Ad v. Anim. Cell Biol. Technol. Bioprocesses, 161щоб білок синтезувався скоріше на ділянці пухли171, 1989; всі які надані тут як посилання]. Альтени або інтрацелюлярно, ніж у всьому організмі. У рнативно, поліпептид ліганду zalpha11 сам може мишей звичайно протягом 5 діб розвиваються вислугувати додатковим маркером поверхні клітини димі пухлини. Пухлинам дозволяють рости протяабо секретованим маркером, пов'язаним зі специгом до 3 діб, протягом оцього періоду вони можуть фічною стосовно етапу експресією тканини. Як так, досягти розміру 1500-1800мм 3 - у контрольній оббезпосередній вимір поліпептиду ліганду zalpha11, робленій групі. Величину пухлини та масу тіла або втрати його експресії у тканині як його дифеуважно відслідковують протягом експерименту. ренціатів, може слугувати маркером для дифереПід час умертвіння пухлин у видаляють та зважунціації тканин. ють разом з легенями та печінкою. Маса легенів Подібно, безпосередній вимір поліпептиду лівиявляє гарну кореляцію з масою метастатичної ганду zalpha11, або втрати його експресії у тканині пухлини. Як додатковий вимір, підраховують метаможна визначити у тканині або у клітинах як свідостази поверхні легенів. Вирізані пухлину, легені та цтво їх піддавання прогресуванню пухлини. Збіпечінку готують для гістопатологічного дослідженльшення інвазивності та рухливості клітини, або ня, імуногістохімії та гібридизації in situ, викорис 53 84387 54 товуючи відомі в рівні техніки та описані тут споможна лікувати лігандом zalpha11. Опосередковасоби. Отже, можна визначати вплив розглянутих на конденсованим токсином активація рецептору експресованих поліпептидів, наприклад, ліганду zalpha11 забезпечує два незалежні засоби для zalpha11, на здатність пухлини створювати судинінгібування росту цільових клітин, перший є іденну сітку та утворювати метастази. На додаток, за тичний розглянутій дії ліганду поодинці, а другий винятком використання аденовірусу, імплантовані обумовлений доставкою токсину через засвоєння клітини можна тимчасово трансфектувати ліганрецептору. Обмежений характер лімфоїдної ексдом zalpha11. Використання стабільних трансфекпресії рецептору zalpha11 означає, що сполучення тантів ліганду zalpha11, а також використання здаліганд-сапорин може бути толерантним для пацієтних до індукції промотерів для активації експресії нтів. ліганду zalpha11 in vivo є відомим в рівні техніки, і Коли лікування злоякісних включає транспланце можна використовувати в такій системі для дотацію алогенного кісткового мозку або стовбурних слідження індукції метастазів лігандом zalpha11. клітин, ліганд zalpha11 може бути корисним для Більш того, очищені ліганд zalpha11 або кондиціпосилення дії трансплантату на пухлин у. Ліганд йовані середовища ліганду zalpha11 можна безпоzalpha11 стимулює створення літичних NK-клітин з середньо ін'єктувати в таких мишачій моделі, а обмежених попередників та стимулює проліферавідтак використовувати в цій системі. Для загальцію Т-клітин після активації рецепторів антигену. ного посилання [дивися, O'Reilly MS, et al. Cell 79: Відтак, коли пацієнти сприймають алогенні транс315-328, 1994; та Rusciano D, et al. Murine Models плантати кісткового мозку, ліганд zalpha11 посиof Liver Metastasis (Мишачі моделі метастазів у люватиме генерацію анти-пухлинної реакції, з інпечінці), Invasion Metastasis 14: 349-361, 1995]. фузією донорних лімфоцитів або без неї. Ліганд zalpha11 буде корисним при лікуванні Тканинний розподіл рецептору даного цитокіну онкогенезу, а відтак міг би бути корисним при лікудає змогу чітко виявити можливі ділянки дії цього ванні раку. Ліганд zalpha11 інгібує стимульовану цитокіну. Норзерн-аналіз рецептору zalpha11 виEL-4 проліферацію стимульованих анти-IgM норявив транскрипти у селезінці, тимусі, лімфатичномальних В-клітин, і подібний вплив спостерігають у му вузлі, кістковому мозку, та лейкоцитах перифелініях B-клітин пухлин, означаючи, що може бути рійної крові людини. Специфічні типи клітин терапевтично корисно лікувати пацієнтів лігандом ідентифікували як експресуючі рецептор zalpha11, zalpha11 для індукування меншої проліферації і спостерігали сильні сигнали у реакції змішаних клітин В-клітинної пухлини. Ліганд можна було б лімфоцитів (MLR) та у лімфомі Буркітта Раджі. Дві застосовувати у комбінації з іншими засобами вже лінії моноцитів, THP-1 [Tsuchiya et al. Int. J. Cancer при використанні, включаючи звичайні хіміотера26: 171-176, 1980] та U937 [Sundstrom et al. Int. J. певтичні засоби, а також імунні модулятори, як-то Cancer 17: 565-577, 1976], були негативними. альфа-інтерферон. Показано, що альфаРецептор zalpha11 експресується у відносно інтерферон є ефективним при лікуванні деяких високих рівнях у MLR, в яких периферійні мононулейкемій та тваринних моделей захворювань, а клеарні клітини крові (PBMNC) від двох осіб змізростання інгібіторного впливу альфа-інтерферону шують, що призводить до двобічної активації. Вита ліганду zalpha11 є адитивним для щонайменше значення високих рівнів транскрипту у MLR, але не одної похідної від В-клітин пухлини лінії клітин. на відпочиваючих T- або В-клітинних популяціях, Згідно з винаходом запропоновано спосіб змеозначає, що експресію рецептору zalpha11 можна ншення проліферації В- або Т-клітин, що стосуіндукувати у одному чи більше типах клітин протяються пухлин, що включає застосування до ссавця гом активації. Активації виділених популяцій T- та з В- або Т-клітинним новоутворенням кількості В-клітин можна досягти штучно стимулюванням композиції ліганду zalpha11, що достатня для змеклітин PMA та іономіцином. Коли класифіковані ншення проліферації В- або Т-клітин, що стосуклітини піддавали таким умовам активації, рівні ються пухлин. У інших втіленнях, композиція може транскрипту рецептору zalpha11 зростали в обох включати щонайменше один інший цитокін, вибратипах клітин, підтверджуючи роль цього рецептору ний з групи, що складається з IL-2, IL-.15, IL-4, GMта ліганду zalpha11 в імунній реакції, особливо у CSF, ліганду Flt3 або фактору стовбурних клітин. експансії автокринних та паракринних T- та ВЗгідно з ще одним аспектом винаходу запроклітин протягом активації. Ліганд zalpha11 може поновано спосіб зменшення проліферації В- або Ттакож грати роль у експансії примітивніших попеклітин, що стосуються пухлин, що включає засторедників, що залучено у лімфоцитоутворення. сування до ссавця з В- або T-клітинним новоутвоПрисутність низького рівня рецептору zalpha11 ренням кількості композиції антагоністу ліганду знайшли у спочиваючих T- та В-клітинах, і він був zalpha11, що достатня для зменшення проліферавище регульованим при активації в обох типах ції В- або Т-клітин, що стосуються пухлин. У інших клітин. Цікаво, що В-клітини також регулюють інвтіленнях, композиція може включати щонайменформацію до кінця швидше, ніж Т-клітини, вказуюше один інший цитокін,. вибраний з групи, що чи, що амплітуда сигналу та час гасіння сигналу є складається з IL-2, IL-15, IL-4, GM-CSF, ліганду важливими для прийнятної регуляції реакцій ВFlt3 або фактору проліферації стовбурних клітин, клітин. що стосуються В- або Т-клітинних пухлин. Крім На додаток, велика частина власних клітин того, антагоніст ліганду zalpha11 може бути конкишкової пластини виявляє позитивні сигнали гібденсованим білком ліганд/токсин. ридизації з рецептором zalpha11. Ця тканина Конденсат токсину ліганд zalpha11-сапорин складається зі змішаної популяції лімфоїдних кліможна застосовувати проти подібного кола лейкетини, включаючи активовані СD4+-Т-клітини та мій та лімфом, розширюючи кількість лейкемій, які активовані В-клітини. Імунна дисфункція, зокрема, 55 84387 56 хронічна активація імунної реакції у слизовій обопериферійної крові у порівнянні з клітинами кістколонці, грає важливу роль у етіології хвороби Крового мозку або периферійної крові, культивованина; анормальна реакція на прозапальні цитокіни ми при відсутності ліганду zalpha11. Згідно з іншита/або продукування прозапальних цитокінів є тами втіленнями кровотворні клітини та попередники кож підозрілим фактором [Braegger et al., Annals кровотворних клітин є лімфоїдними клітинами. Allergy 72: 135-141, 1994; Sartor RB Am. J. Згідно з іншим втіленням лімфоїдні клітини є NKGastroenterol. 92: 5S-11S, 1997]. Ліганд zalpha11 у клітинами або цитотоксичними Т-клітинами. Крім взаємодії з IL-15 розвиває NK-клітини з поперетого, композиція може також включати щонаймендників з кісткового мозку та збагачує NK-клітинну ше один інший цитокін, вибраний з групи, що склаефекторну функцію, ліганд zalpha11 також співсдається з IL-2, IL-15, IL-4, GM-CSF, ліганду Flt3 та тимулюс розвинені В-клітини, що стимульовані фактору стовбурних клітин. антитілами проти CD40, але інгібує проліферацію Альтернативно, ліганд zalpha11 може активуВ-клітин на сигнали через IgM. Ліганд zalpha11 вати імунну систему, що могло би бути важливим у посилює проліферацію Т-клітин у взаємодії з сигпідтримці імунітету щодо інфекційних захворюналом через рецептор Т-клітин, а надекспресія у вань, лікуванні імунокомпро-місних пацієнтів, як-то трансгенних мишей призводить до лімфопенії та ВІЛ+ пацієнтів, або у посиленні вакцин. Зокрема, вирощування моноцитів та гранулоцитів. Ця плестимуляція ліганду zalpha11 або вирощування NKйотропна дія ліганду zalpha11 означає, що вона клітин, або їх попередників, могли б забезпечити може забезпечити терапевтичну корисність для терапевтичну корисність при лікуванні вірусної широкого кола захворювань, що виникають від інфекції та як анти-пухлинного фактору. NKдефектів імунної системи, включаючи (але без клітини, можна думати, грають головну роль у обмеження) системний червоний вовчак (SLE), знищенні метастатичних клітин пухлини, а пацієнревматоїдний артрит (RA), розсіяний склероз (MS), ти з метастазами та твердими пухлинами мають важку міастенію, та діабет. Важливо відмітити, що зменшені рівні активності NK-клітин [Whiteside et. ці захворювання є результатом комплексного ланal., Curr. Top. Microbiol. Immunol. 230: 221-244, цюга імунної дисфункції (SLE, наприклад, є вияв1998]. Подібно, стимуляція лігандом zalpha11 імуленням дефектів у T- та В-клітинах), та що імунні нної реакції проти вірусних та невірусних патогенклітини є залежними від взаємодії одна з одною них агентів (включаючи бактерії, найпростіших та для виклику можливої імунної реакції. Відтак, лігриби) забезпечуватимуть терапевтичну корисганд zalpha11 (або антагоніст ліганду), що можна ність при лікуванні такої інфекції інгібуванням росвикористовува ти для впливу на більше ніж один ту таких інфекційних агентів. Визначення безпосетип імунних клітин, є притягальним терапевтичним редньо або небезпосередньо рівнів патогену або кандидатом для впровадження на багатьох етапах антигену, як то клітини пухлини, присутніх у тілі, захворювання. можна досягти рядом відомих в рівні техніки та Поліпептиди та білки представленого винахоописаних тут способів. ду можна також використовувати ех vi vo, як-то у Представлений винахід включає спосіб стимуавтологічний культурі кісткового мозку. Коротше, ляції імунної реакції у підданого дії патогену або кістковий мозок видаляють з пацієнта до хіміотеантигену ссавця, який включає етапи: (1) визнарапії або трансплантації органу і обробляють лігачення безпосередньо або небезпосередньо рівнів ндом zalpha11, як варіант, у комбінації з одним чи патогену або антигену, присутні х у вказаному ссабільше іншими цитокінами. Оброблений кістковий вці, (2) застосування композиції, яка включає полімозок далі повертають пацієнту після хіміотерапії пептид ліганду zalpha11 у фармацевтично прийнядля прискорення відновлення кісткового мозку або тному наповнювачі, (3) визначення безпосередньо після трансплантації для пригнічення хвороби у або небезпосередньо рівнів патогену або антигену проти хазяїна. На додаток, білки представленого у вказаному ссавці, (4) порівняння рівнів патогену винаходу можна також використовувати для виабо антигену на етапі 1 з рівнем патогену або анрощування ex vi vo кісткового мозку або клітинтигену на етапі 3, при цьому зміна цього рівня є попередників периферійної крові (PBPC). До обропоказовою стосовно стимуляції імунної реакції. бки кістковий мозок можна стимулювати фактором Згідно з іншим втіленням винаходу композицію стовбурних клітин (SCF) для вивільнення ранніх ліганду zalpha11 застосовують повторно. Згідно з клітин-попередників у периферійний кровообіг. Ці іншими втіленнями винаходу антиген є Впопередники можна зібрати з периферійної крові клітинною пухлиною, вірусом, паразитом чи бактета концентрувати, а потім обробити у культурі лірією. гандом zalpha11, як варіант, у комбінації з одним Згідно з ще одним аспектом винаходу запрочи більше іншими цитокінами, включаючи але без поновано спосіб стимуляції імунної реакції у піддаобмеження SCF, IL-2, IL-4, IL-7 або IL-15, для диного дії патогену або антигену ссавця, який вклюференціації та проліферації у лімфоїдних к ультучає етапи: (1) визначення рівня специфічного рах, які далі можна повернути пацієнту після хіміостосовно патогену або антигену антитіла, (2) затерапії або трансплантації. стосування композиції, яка включає поліпептид Згідно з винаходом запропоновано спосіб виліганду zalpha11 у фармацевтично прийнятному рощування кровотворних клітин та попередників наповнювачі, (3) визначення після застосування кровотворних клітин, що включає культивування рівня специфічного стосовно патогену або антигекісткового мозку або клітин периферійної крові з ну антитіла, (4) порівняння рівнів антитіла на етапі композицією, що містить кількість ліганду zalpha11, 1 з рівнем антитіла на етапі 3, при цьому збільщо достатня для продукування збільшеного числа шення цього рівня є показовим стосовно стимулялімфоїдних клітин у клітинах кісткового мозку або ції імунної реакції. 57 84387 58 Полінуклеотиди, що кодують поліпептиди ліганою конденсацією, DEAE-декстраном, осадженням нду zalpha11, корисні при застосуванні в рамках фосфа том кальцію, використанням генного "постгенної терапії, де потрібно посилити інгібування рілу з дробовика "або використання транспортеру активності ліганду zalpha11. Якщо ссавець має ДНК-вектору. [Дивися, наприклад, Wu et al. J. Віоі. мутований чи відсутній ген ліганду zalpha11, ген Chem. 207: 963-7, 1992: Wu et al., J. Biol. Chem. ліганду zalpha11 можна ввести у клітини ссавця. 263: 14621-4, 1988]. Згідно з одним втіленням винаходу ген, що кодує Антисенсову методологію можна використовуполіпептид ліганду zalpha11, уводять in vivo у вірувати для інгібування транскрипції гену ліганду сному векторі. Такі вектори включають послаблеzalpha11, як-то інгібування проліферації клітин in ний чи дефективний ДНК-вірус, як-то, але без обvi vo. Полінуклеотиди, що є комплементарними до меження, вірус герпесу (HSV), вір ус папіломи, сегменту кодуючого ліганд zalpha11 полінуклеотивірус Епштейна-Барра (EBV), аденовірус, аденоаду (наприклад, полінуклеотиду, представленого соційований вірус (AAV), то що. Дефективні віруси, послідовністю SEQ ID NO: 1) призначені для прищо повністю чи частково втратили вір усні гени, є єднання до кодуючої ліганд zalpha11 мРНК та інгікращими. Дефективні віруси,після уведення у клібування трансляції такої мРНК. Такі антисенсові тину не є інфекційними. Використання векторів полінуклеотиди використовують для інгібування дефективних вірусів дозволяє застосування до експресії кодуючих поліпептид ліганду zalpha11 клітин у специфічних локалізованих ділянках без генів у к ультурі клітин або у суб'єкті. турботи, що вектор може інфікувати інші клітини. Можна створювати також мишей, що сконстПриклади окремих векторів включають, але без руйовані для експресії гена ліганду zalpha11 і виобмеження вектор дефективного вірусу герпесу 1 значені як "трансгенні миші", та мишей, що вияв(HSV1) [Kaplitt et al., Моіес. Cell. Neurosci. 2: 320ляють повну відсутність функції гена ліганду 30, 1991]; вектор послабленого аденовірусу, як-то zalpha11 та визначені як "нокаутні миші" вектор, що описано [Stratford-Perricaudet et al., J. [Snouwaert et al., Science 257: 1083, 1992; Lowell et Clin. Invest. 90: 626-30, 1992]; та вектор дефективal. Nature 366: 740-42, 1993; Capecchi, M.R., ного аденоасоційованого вірусу [Samulski et al., J. Science 244: 1288-1292, 1989; Palmiter, R.D. et al. Virol. 61: 3096-101, 1987; Samulski et al., J. Virol. 63: Annu Rev Genet. 20: 465-499, 1986]. Наприклад, 3 822-8, 1989]. трансгенні миші, що надекспресують ліганд Ген ліганду zalpha11 можна уводити у ретровіzalpha11, повсюдно або під впливом тканинорусний вектор, наприклад, як описано [Anderson et специфічного або тканино-обмежувального проal., патент США №5399346; Mann et al. Cell 33,: мотеру, можна використовувати для запиту, чи 153, 1983; Temin et al., патент США №4650764; впливає надекспресія на фенотип. Наприклад, Temin et al., патент США №4980289; Markowitz et надекспресія поліпептиду ліганду zalpha11 нативal., J. Virol. 62: 1120, 1988; Temin et al., патент ного типу, фрагменту поліпептиду або його мутанСША №5124263; Міжнародна патентна публікація ту може змінювати нормальні клітинні процеси, № WO 95/07358, опублікована 16 березня 1995р., приводячи до фенотипу, що іденти фікує тканини, в Dougherty et al.; and Kuo et al. Blood 82: 845, 1993]. яких експресія ліганду zalpha11 є функціонально Альтернативно, вектор можна уводити ліпофекцірелевантною і може вказувати терапевтичні мішеєю in vi vo, використовуючи ліпосоми. Синтетичні ні для ліганду zalpha11, його агоністів або антагокатіоногенні ліпіди можна використовувати для ністів. Наприклад, кращою трансгенною мишею виготовлення ліпосом для трансфекціі in vivo гена, для створення є така, що надекспресує ліганд що кодує маркер [Feigner et al., Proc. Natl. Acad. zalpha11 (амінокислотні залишки 32-162 послідовSci. USA 84: 7413-7, 1987; Mackey et al., Proc. Natl. ності SEQ ID NO: 2). Більш того, така надекспресія Acad. Sci. USA 85: 8027-31, 1988]. Використання може призвести до фенотипу, що виявляє подібліпофекції для введення екзогенних генів у спеність із захворюваннями людини. Подібно, нокаутцифічні органи in vivo має деякі практичні переваних стосовно ліганду zalpha11 мишей можна викоги. Молекулярне спрямування ліпосом до специристовувати для визначення, де ліганд zalpha11 є фічних клітин репрезентує одну область абсолютно потрібним in vivo. Можна мати фенотип корисності. Конкретніше, одну область корисності нокаутних мишей, що передбачає дію in vivo такорепрезентує спрямування транс-фекції до певних го антагоністу ліганду zalpha11, як той, що тут опиклітин. Наприклад, спрямування трансфекції до сано. Для створення нокаутної миші можна викопевних типів клітин буде зокрема переважним у ристовувати кДНК ліганду zalpha11 людини або тканині з клітинною гетерогенністю, як-то імунній миші. Цих мишей можна застосовувати для дослісистемі, підшлунковій залозі, печінці, нирках, та дження гена ліганду zalpha11 та кодованого у зв'ямозку. Ліпіди для спрямування можна хімічно спозку з цим білку у системі in vi vo і використовувати лучати з іншими молекулами. Спрямовані пептиди як моделі in vivo для відповідних захворювань лю(наприклад, гормони або нейротрансмітери), білки, дини. як-то антитіла, або непептидні молекули можна Більш того, експресію трансгенними мишами сполучати з ліпосомами хімічно. антисенсових полінуклеотидів або спрямованих Для видалення клітин-мішеней з тіла можливо: проти ліганду zalpha11 рибосом ліганду zalpha11, уводити вектор як оголену ДНК-плазміду, а потім що тут описано, можна застосовувати аналогічно реімплантувати трансформовані клітини у тіло. до описаних вище трансгенних мишей. Вивчення Оголені ДНК-вектори для генної терапії можна також можна проводити застосуванням очищеного уводити у потрібні клітини хазяїна відомими в рівні білку ліганду zalpha11. техніки способами, наприклад, трансфекцією, елеДля фармацевтичного використання, білки ктропорацією, мікроін'єкцією, трансдукцією, клітинпредставленого винаходу формують для паренте 59 84387 60 ральної, зокрема внутрішньовенної чи підшкірної ційні умови аналогічні вищезазначеним. Продукт доставки звичайними способами. Біоактивні споPCR проганяли на 1% агарозі з низкою температулучення поліпептидів або антитіл, що тут описано, рою плавлення (Boerhinger Mannheim) та фрагможна доставляти внутрішньовенно, внутрішньомент zalpha11 розміром приблизно 900 по виділяартеріально або внутрішньопроточно, або можна ли, використовуючи гель-екстракційний комплект уводити локально у призначеній ділянці дії. ВнутQiaquick за інструкціями виробника. рішньовенне застосування здійснюють звичайно Кожний з описаних вище виділених фрагменболюсною ін'єкцією або інфузією протягом звичайтів змішували при об'ємному співвідношенні 1:1 та ного періоду у одну чи кілька годин. Загалом фарвикористовували у реакції PCR, використовуючи мацевтичні композиції звичайно включають білок ZC17,212 (SEQ ID NO: 5) та ZC20,097 (SEQ ID NO: ліганду zalpha11 у комбінації з фармацевтично 9) для створення химери MPL-zalpha11. Реакційні прийнятним наповнювачем, як-то фізіологічний умови були такими: 95°C протягом 1 хвилини, 35 розчин, буферований фізіологічний розчин, 5% циклів при 95°С протягом 1 хвилини, 55°C протядекстроза у воді тощо. Композиції можуть включагом 1 хвилини. 72°С протягом 2 хвилин, а потім ти один чи більше ексципієнтів, консервантів, со72°С протягом 10 хвилин, і далі просочування при любілізаторів, буферних засобів, альбумін для 10°C. Чистий продукт PCR проганяли на 1% агаропопередження втрати білку на стінках склянки тозі з низкою температурою плавлення (Boehringer що. Способи формування добре відомі в рівні техMannheim) та виділяли фрагмент химери MPLніки та розкриті, наприклад, у [Remington: The zalpha11 розміром приблизно 2,4 ко, використовуScience та Practice of Pharmacy, Gennaro, ed. Mack ючи гель-екстракційний комплект QiaquickÔ Publishing Co., Easton, PA, 19th ed., 1995]. Терапе(Qiagen) за інструкціями виробника. Фрагмент хивтичні дози знаходитимуться загалом в межах 0,1мери MPL-zalpha11 розщеплювали за допомогою 100мкг/кг маси пацієнта на добу, краще 0,5EcoRI (BRL) та Xbal (Boerhinger Mannheim) за ін20мкг/кг на добу, з визначенням точної дози лікаструкціями виробника. Весь продукт розщеплення рем згідно з загальноприйнятними стандартами, проганяли на 1% агарозі з низкою температурою беручи до уваги природу та суворість лікуємого плавлення (Boehringer Mannheim) та розщеплену стану, характерні особливості пацієнта тощо. Вихимеру MPL-zalpha11 виділяли, використовуючи значення дози є у компетенції пересічних фахівців. гель-екстракційний комплект QiaquickÔ (Qiagen) Білки можна застосовувати при гострому лікуванні, за інструкціями виробника. Одержану розщеплену протягом тижня чи менше, часто протягом одноїхимеру MPL-zalpha11 вставляли у експресійний трьох діб, або можна використовувати при хронічвектор як описано нижче. ному лікуванні, протягом кількох місяців чи років. Реципієнтний експресійний вектор pZP-5N Загалом, терапевтично ефективна кількість ліганрозщеплювали за допомогою EcoRI (BRL) та ду zalpha11 є кількістю, що достатня для продукуHindIII (BRL) за інструкціями виробника, а гель вання клінічно помітних змін у кровотворній або очищали як описано вище. Цей фрагмент вектору імунній функції. комбінували з виділеною вище розщепленою Винахід далі ілюстровано наступними необEcoRI та Xbal химерою MPL-zalpha11 та межувальними прикладами. Xbal/Hindlll лінкерним фрагментом у реакції лігуПриклади вання. Лігування здійснювали, використовуючи Приклад 1 Конструювання поліпептидної хилігазу ФАГУ Т4 (BRL), при 15°C протягом ночі. мери MPL-zalpha11: екстрацелюлярний MPL та Зразок після лігування електропорували у електдомен TM конденсували з інтрацелюлярним сигрокомпетентні клітини E. coli DHlOB ElectroMAXÔ і налізаційним доменом zalpha11 (25мкФ, 200Ом, 2,3В). Трансформанти засівали на Екстрацелюлярний та трансмембранний допланшети LB+ампіцилін та скринували одиничні мени рецептору MPL миші виділяли з плазмід, що колонії за допомогою PCR для перевірки химери містять рецептор MPL миші (плазміда PHZ1/MPL), MPL-zalpha11, використовуючи ZCI7,212 (SEQ ID використовуючи PCR з праймерами ZC17,212 NO: 5) та ZC20,097 (SEQ ID NO: 9), використовую(SEQ ID NO: 5) та ZC19.914 (SEQ ID NO: 6). Реакчи описані вище умови PCR. ційні умови були такими: 95°С протягом 1 хвилини, Підтвердження послідовності химери MPL35 циклів при 95°С протягом 1 хвилини, 45°С проzalpha11 провели секвенсуванням, використовуютягом 1 хвилини, 72°C протягом 2 хвилин, а потім чи такі праймери: ZC 12,700 (SEQ ID NO: 10), 72°С протягом 10 хвилин, і далі просочування при ZC5,020 (SEQ ID NO: 11). ZC6,675 (SEQ ID NO: 10°C. Продукт PCR проганяли на 1% агарозі з низ12), ZC7,727 (SEQ ID NO: 13), ZC8,290 (SEQ ID кою температурою плавлення (Boerhinger NO: 14), ZC 19,572 (SEQ ID NO: 15), ZC6,622 (SEQ Mannheim, Indianapolis, IN) та виділяли фрагмент ID NO: 16), ZC7,736 (SEQ ID NO: 17) та ZC9,273 рецептору MPL розміром приблизно 1,5 ко, вико(SEQ ID NO: 18). Інсерція мала розмір приблизно ристовуючи гель-екстракційний комплект 2,4 ко, та була повної довжини. QiaquickÔ (Qiagen) за інструкціями виробника. Приклад 2 Проліферація на основі химери Інтрацелюлярні домени zalpha11 людини виMPL-zalpha11 у дослідженні BAF3 з використанням діляли з плазміди, що містить кДНК рецептору аламар-блакитного zalpha11, використовуючи PCR з праймерами ZC А. Констр уювання експресуючих химери MPL19,913 (SEQ ID NO: 8) та ZC20,097 (SEQ ID NO: 9). zalpha11 клітин BaF3 Полінуклеотидна послідовність, що відповідає до Залежну від інтерлейкіну-3 (IL-3) лінію предлікодуючій рецептор zalpha11 послідовності, покамфоїдних клітин BaF3, похідну від кісткового мозку зана як SEQ ID NO: 7, а відповідна амінокислотна миші [Palacios та Steinmetz, Cell 41: 727-734. 1985: послідовність показана як SEQ ID NO: 115. РеакMathey-Pre vot et al., Mol. Cell. Biol. 6: 4133-4135,
ДивитисяДодаткова інформація
Назва патенту англійськоюHuman cytokine as a ligand of zalpha receptor and use thereof
Автори англійськоюNOVAK JULIA E., Presnell Scott R., Sprecher Cindy A., Foster Donald K., HOLLY RICHARD D, GROSS JANE A, JOHNSTON JANET V, NELSON ANDREW J, Dillon Stacey R., HAMMOND ANGELA K
Назва патенту російськоюЧеловеческий цитокин как лиганд зальфа рецептора и его применение
Автори російськоюНовак Джулия Э., Преснелл Скотт Р., Спричер Синди А., Фостер Дональд К., Холли Ричард Д., Гросс Джейн А., Джонстон Жанет В., Нелсон Эндрю Дж., Диллон Стейси Р., Хеммонд Анджела К.
МПК / Мітки
МПК: C12N 15/19, A61K 38/00, C07K 14/52
Мітки: рецептора, цитокін, ліганд, зальфа, людський, використання
Код посилання
<a href="https://ua.patents.su/129-84387-lyudskijj-citokin-yak-ligand-zalfa-receptora-i-jjogo-vikoristannya.html" target="_blank" rel="follow" title="База патентів України">Людський цитокін як ліганд зальфа рецептора і його використання</a>
Способи використання розчинного рецептора br43x2

Номер патенту: 74330
Опубліковано: 15.12.2005
Автори: Ксу Венфанг, Ї Девід П., Гросс Джейн А., Медден Карен
МПК: C07K 14/705, A61P 7/02, A61P 25/28, A61P 37/00, A61P 11/06, A61P 31/04, A61P 31/18, A61P 19/02, C12N 1/19, C07K 14/715, C12N 1/15, A61P 11/00, A61P 3/10, A61P 37/02, C07K 19/00, A61P 7/06, A61K 38/00, A61K 39/395, A61P 43/00, A61P 35/02, A61P 29/00, A61P 21/04, A61P 13/12, A61P 9/12, A61P 37/06, C12N 5/10, C12P 21/02, C12N 1/21, C07K 16/28, C12P 21/08, C12N 15/09, A61K 38/17, A61P 35/00
Мітки: використання, br43x2, рецептора, способи, розчинного
Формула / Реферат:
1. Спосіб інгібування активності ztnf4 у ссавця, що включає призначення вказаному ссавцю ефективної кількості сполуки, яка вибрана з групи, що складається з:a) поліпептиду, що включає екстрацелюлярний домен BR43x2 (SEQ ID NO:2),b) розчинного поліпептиду, що включає екстрацелюлярний домен ТАСІ,c) поліпептиду, що включає екстрацелюлярний домен ВСМА,d) поліпептиду, що включає послідовність SEQ ID NO:10,e)...
Ліганди 5-нт рецептора та їх використання

Номер патенту: 74443
Опубліковано: 15.12.2005
Автори: Новомісле Уільям Альберт, Уелч Уіллард МакКован, Чіанг Юан-Чінг Фебе
МПК: A61P 25/06, A61P 25/18, A61P 1/14, A61K 31/506, A61P 1/00, A61P 25/24, C07D 417/12, A61P 25/14, C07D 239/46, A61P 25/34, A61P 25/22, A61P 1/04, A61P 43/00, A61P 3/04, A61P 25/08, A61P 15/12, A61P 25/20, A61P 9/00, C07D 241/20, A61P 15/00, A61P 13/02, A61P 15/10, C07D 403/04, A61P 25/00, A61P 9/10, C07D 401/12, C07D 405/12, A61P 25/28, C07D 409/12, A61K 45/00, A61K 31/497, A61P 3/10, C07D 239/42, A61P 25/32
Мітки: ліганди, використання, 5-нт, рецептора
Формула / Реферат:
1. Сполука формули (IA), (ІА)в якійX і Y означають CR і Z означає N, або X означає N, а Y і Z означають CR, де R в кожному випадку означає водень; W означає окси; принаймні один з R1a, R1b, R1d та R1e незалежно вибраний з групи, яка складається з галогену, нітро, аміно, ціано, -C(O)NH2 та (C1-C4)алкілу, або R1a та R1b, взяті разом,...
Похідні індолін-2-ону, спосіб їх виготовлення та використання як лігандів рецептора окситоцину

Номер патенту: 73163
Опубліковано: 15.06.2005
Автори: Серрадель-ле Галь Клодін, ВАЛЕТТ Жерар, Фулон Лоік, Гарсіа Жорж
МПК: A61P 17/02, A61K 31/55, C07D 401/04, A61K 31/496, A61P 15/18, A61K 31/5377, A61P 15/02, A61P 15/06, A61K 31/4439, C07D 405/10, A61P 5/10, A61P 25/18, A61P 3/04, C07D 403/12, A61P 3/06, A61P 9/12, A61P 13/02, A61P 15/14, A61K 31/454, A61P 13/08, A61P 25/22, A61P 15/10, C07D 405/12, A61P 25/28, A61P 25/24, A61P 9/04, C07D 209/34, A61K 31/404, C07D 401/12, A61P 43/00, A61P 3/12, A61P 9/10, A61K 45/00, C07D 209/38, C07D 403/10, A61P 25/02, A61P 15/00, A61P 25/30, A61P 15/08, A61P 35/00, C07D 401/10
Мітки: рецептора, індолін-2-ону, окситоцину, виготовлення, використання, похідні, спосіб, лігандів
Формула / Реферат:
1. Сполуки у формі чистого енантіомера чи суміші енантіомерів формули,де:- R0 - група, вибрана з;де:- Z1 - атом хлору, брому, йоду або флуору, або (С1-С4)алкіл, (С1-С4)алкоксил або трифлуорметил;- Z2 - атом гідрогену, хлору, брому, йоду або флуору, або...
Похідні амінотіазолу, спосіб їх одержання, фармацевтична композиція та їх використання як лігандів рецептора crf

Номер патенту: 73125
Опубліковано: 15.06.2005
Автори: Гюллі Даніель, Фонтен Евеллін, Жеслен Мішель, Роже П'єр, Прадін Антуан
МПК: A61P 25/20, A61P 17/06, A61P 9/00, A61P 25/08, A61P 25/18, A61P 1/12, A61P 31/00, A61P 15/06, A61P 43/00, A61P 25/24, A61K 31/426, A61P 3/10, A61P 5/04, C07D 417/12, A61P 25/02, C07D 277/20, A61P 25/16, A61P 25/22, A61P 21/00, A61P 37/00, A61P 29/00, A61P 9/14, A61P 25/32, A61P 25/00, A61P 25/14, A61P 27/02, A61P 1/00, A61P 19/02, A61P 25/28, A61P 15/00, A61P 25/06, A61P 37/06, A61P 25/30, A61P 3/04, A61P 35/00, C07D 277/42, C07B 53/00
Мітки: рецептора, похідні, одержання, фармацевтична, лігандів, спосіб, використання, амінотіазолу, композиція
Формула / Реферат:
1. Сполуки, у рацемічній формі або у формі чистого енантіомера, формули: , (I.2)деR1 та R2, що можуть бути однаковими або різними, кожний незалежно – атом галогену; гідроксі(C1-C5)алкіл; (C1-C5)алкіл; аралкіл, в якому арильною частиною є (C6-C8), а алкільною частиною -(C1-C4); (C1-C5)алкоксил; трифлуорметил; нітрогрупа; нітрильна група; група -SR,...
Хінуклідинові похідні та їх використання як лігандів мускаринового м3-рецептора

Номер патенту: 73509
Опубліковано: 15.08.2005
Автори: Буіл Альберо Марія Антонія, Фернандез Форнер Долорс, Прат Квінонез Марія
МПК: A61P 1/04, A61P 13/06, A61P 27/16, A61P 11/02, A61K 31/439, A61P 11/06, A61P 13/10, A61P 13/02, C07D 453/00, A61P 13/00, A61P 11/14, A61P 1/06, A61P 1/12, A61P 11/08, A61P 11/00, A61P 43/00, A61P 1/00
Мітки: використання, мускаринового, м3-рецептора, хінуклідинові, похідні, лігандів
Формула / Реферат:
1. Сполука формули (І) , (І)у якій© означає фенільне кільце, С4-С9-гетероароматичну групу, що містить один чи кілька гетероатомів, або нафталінільну, 5,6,7,8-тетрагідронафталінільну чи біфенільну групу,кожен з R1, R2 та R3 незалежно означає атом гідрогену чи атом галогену або гідроксильну групу, або феніл, групу -OR4, -SR4, -NR4R5, -NHCOR4, -CONR4R5, -CN, -NO2, -COOR4 чи -CF3, або пряму чи розгалужену нижчу...
Попередній патент: Спосіб виробництва перлового вина
Наступний патент: Змінений штам модифікованого вірусу віспи великої рогатої худоби ankara (mva)
Випадковий патент: Спосіб профілактики та симптоматичного лікування променевої хвороби сільськогосподарських та домашніх тварин