Спосіб ферментативного виробництва чотириатомних спиртів
Номер патенту: 103364
Опубліковано: 10.10.2013
Автори: Нагараджан Васанта, Флінт Денніс, Дональдсон Гейл К., Маггіо-Холл Лорі Енн, Еліот Ендрю С.






Формула / Реферат
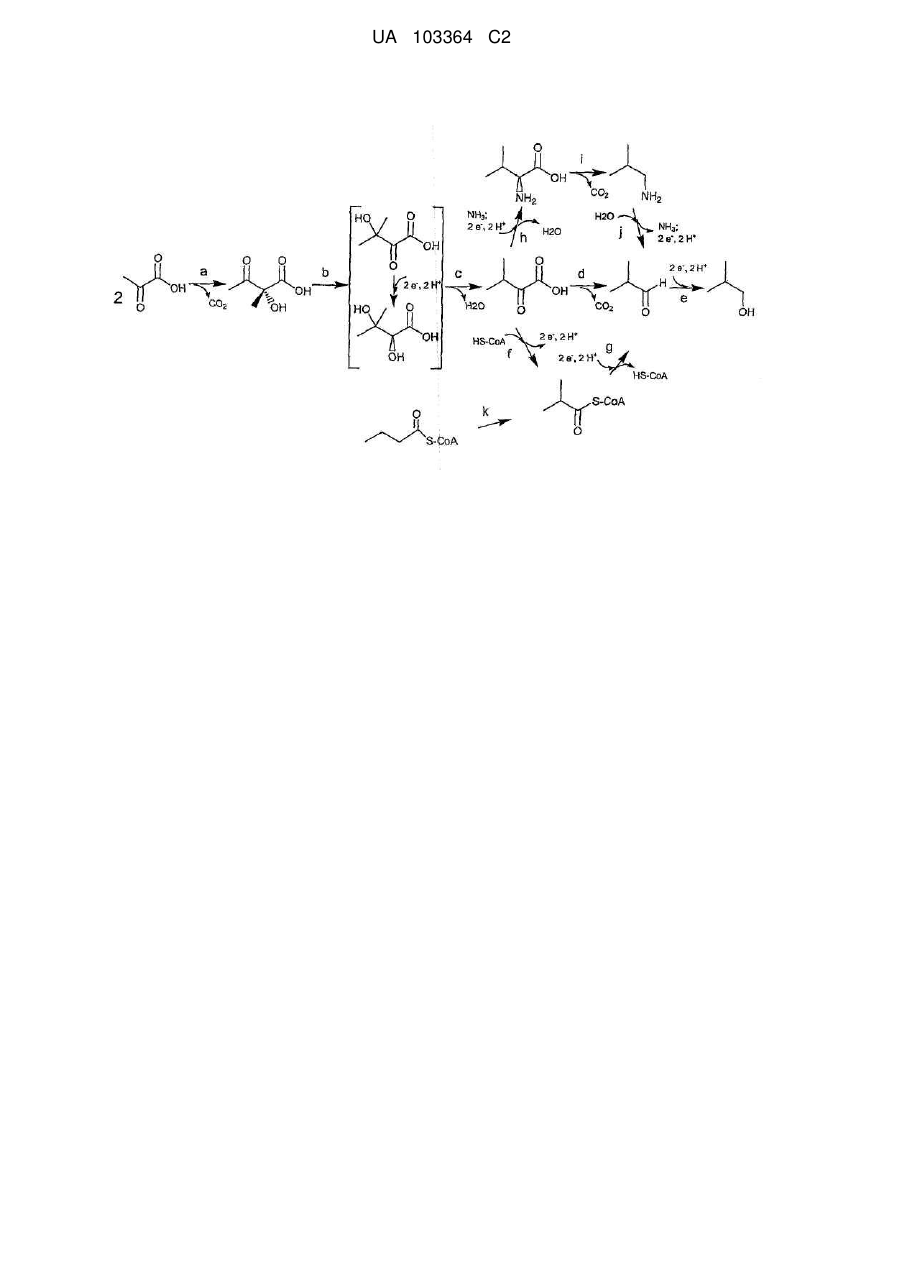
1. Рекомбінантна мікробна клітина-хазяїн, яка містить ферментативний шлях ізобутанолу, що включає молекули ДНК, які кодують набір поліпептидів, що каталізують наступні перетворення субстрату на продукт:
і) піруват на ацетолактат;
іі) ацетолактат на 2,3-дигідроксіізовалерат;
ііі) 2,3-дигідроксіізовалерат на α-кетоізовалерат;
iv) α-кетоізовалерат на валін;
v) валін на ізобутиламін;
vi) ізобутиламін на ізобутиральдегід; та
vii) ізобутиральдегід на ізобутанол,
де щонайменше одна молекула ДНК є гетерологічною для зазначеної мікробної клітини-хазяїна і де зазначена мікробна клітина-хазяїн продукує ізобутанол.
2. Клітина-хазяїн за п. 1, де поліпептид, що каталізує перетворення субстрату на продукт, піруват на ацетолактат, є ацетолактатсинтазою.
3. Клітина-хазяїн за п. 1, де поліпептид, що каталізує перетворення субстрату на продукт, ацетолактат на 2,3-дигідроксіізовалерат, є ізомероредуктазою ацетогідроксикислот.
4. Клітина-хазяїн за п. 1, де поліпептид, що каталізує перетворення субстрату на продукт, 2,3-дигідроксіізовалерат на α-кетоізовалерат, є дегідратазою ацетогідроксикислот.
5. Клітина-хазяїн за п. 1, де поліпептид, що каталізує перетворення субстрату на продукт, ізобутиральдегід на ізобутанол, є дегідрогеназою розгалужених спиртів.
6. Клітина-хазяїн за п. 1, де поліпептид, що каталізує перетворення субстрату на продукт, α-кетоізовалерат на валін, є трансаміназою.
7. Клітина-хазяїн за п. 1, де поліпептид, що каталізує перетворення субстрату на продукт, а-кетоізовалерат на валін, є валіндегідрогеназою.
8. Клітина-хазяїн за п. 1, де поліпептид, що каталізує перетворення субстрату на продукт, валін на ізобутиламін, є валіндекарбоксилазою.
9. Клітина-хазяїн за п. 1, де поліпептид, що каталізує перетворення субстрату на продукт, ізобутиламін на ізобутиральдегід, є омега-трансаміназою.
10. Клітина-хазяїн за п. 1, де клітину вибирають з групи, що складається з: бактерії, ціанобактерії, нитчастого гриба та дріжджів.
11. Клітина-хазяїн за п. 10, де клітина є членом роду, вибраного з групи, що складається з Clostridium, Zymomonas, Escherichia, Salmonella, Rhodococcus, Pseudomonas, Bacillus, Lactobacillus, Enterococcus, Alcaligenes, Klebsiella, Paenibacillus, Arthrobacter, Corynebacterium, Brevibacterium, Pichia, Candida, Hansenula та Saccharomyces.
12. Клітина-хазяїн за п. 11, де клітина є Escherichia coli.
13. Клітина-хазяїн за п. 11, де клітина є Alcaligenes eutrophus.
14. Клітина-хазяїн за п. 11, де клітина є Bacillus licheniformis.
15. Клітина-хазяїн за п. 11, де клітина є Paenibacillus macerans.
16. Клітина-хазяїн за п. 11, де клітина є Rhodococcus erythropolis.
17. Клітина-хазяїн за п. 11, де клітина є Pseudomonas putida.
18. Клітина-хазяїн за п. 11, де клітина є Bacillus subtilis.
19. Клітина-хазяїн за п. 11, де клітина є Lactobacillus plantarum.
20. Клітина-хазяїн за п. 11, де клітину вибирають з групи, що складається з Enterococcus faecium, Enterococcus gallinarium та Enterococcus faecalis.
21. Клітина-хазяїн за п. 11, де клітина є Saccharomyces cerevisiae.
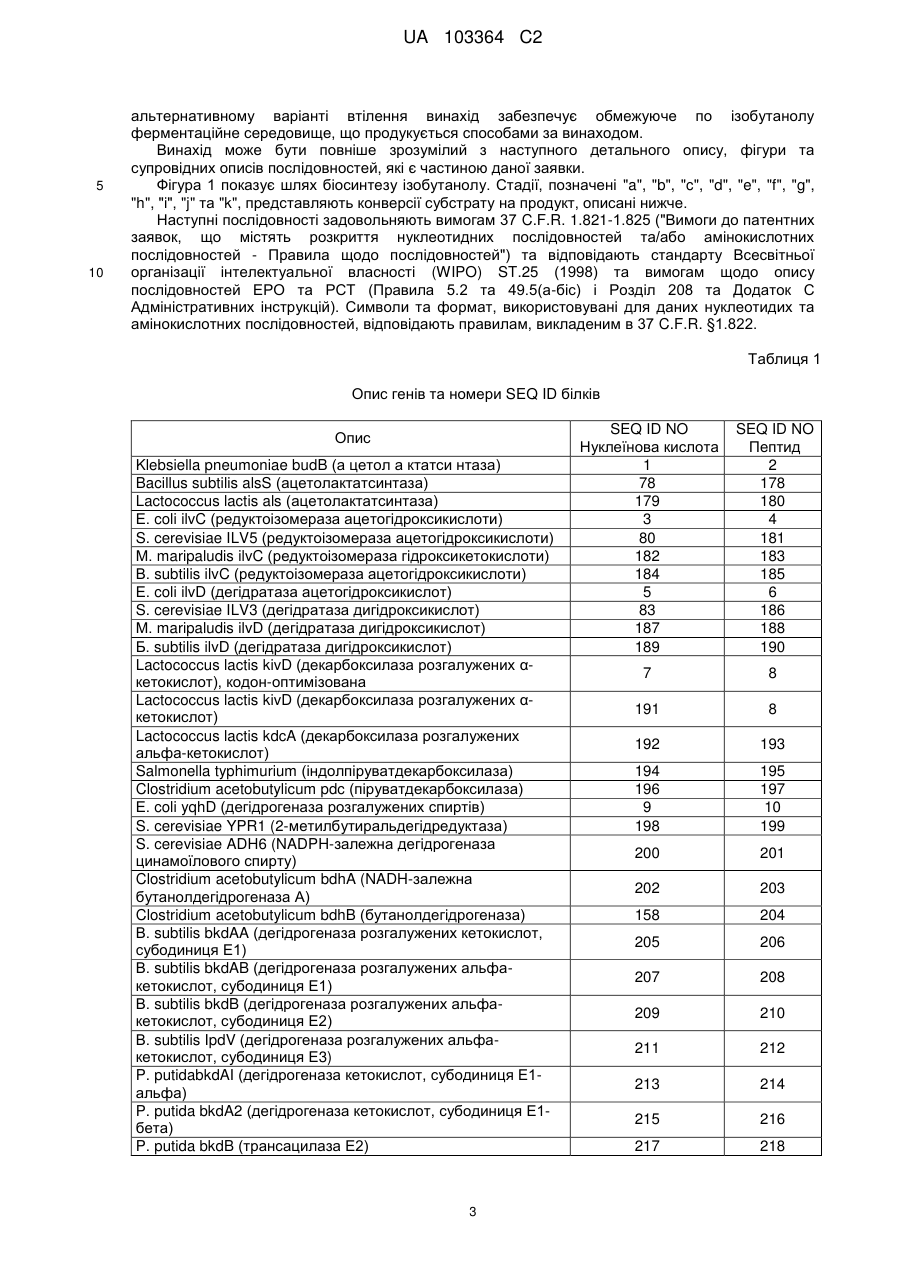
22. Клітина-хазяїн за п. 2, де ацетолактатсинтаза має амінокислотну послідовність, вибрану з групи, що складається з SEQ ID NO:2, SEQ ID NO:178 та SEQ ID NO:180.
23. Клітина-хазяїн за п. 3, де ізомероредуктаза ацетогідроксикислот має амінокислотну послідовність, вибрану з групи, що складається з SEQ ID NO:43, SEQ ID NO:181, SEQ ID NO:183 та SEQ IDNO:185.
24. Клітина-хазяїн за п. 4, де дегідратаза ацетогідроксикислот має амінокислотну послідовність, вибрану з групи, що складається з SEQ ID NO:6, SEQ ID NO:186, SEQ ID NO:188 та SEQ ID NO:190.
25. Клітина-хазяїн за п. 5, де дегідрогеназа розгалужених спиртів має амінокислотну послідовність, вибрану з групи, що складається з SEQ ID NO:10, SEQ ID NO:199, SEQ ID NO:201, SEQ ID NO:203 та SEQ ID NO:204.
26. Клітина-хазяїн за п. 6, де трансаміназа має амінокислотну послідовність, вибрану з групи, що складається з SEQ ID NO:232, SEQ ID NO:234, SEQ ID NO:236, SEQ ID NO:238 та SEQ ID NO:240.
27. Клітина-хазяїн за п. 7, де валіндегідрогеназа має амінокислотну послідовність, вибрану з групи, що складається з SEQ ID NO:242 та SEQ ID NO:244.
28. Клітина-хазяїн за п. 8, де валіндекарбоксилаза має амінокислотну послідовність, описану в SEQ ID NO:246.
29. Клітина-хазяїн за п. 9, де омега-трансаміназа має амінокислотну послідовність, вибрану з групи, що складається з SEQ ID NO:248, SEQ ID NO:250, SEQ ID NO:252 та SEQ ID NO:254.
30. Клітина-хазяїн за п. 1, де клітина-хазяїн є факультативним анаеробом.
31. Спосіб одержання ізобутанолу, який включає:
1) одержання рекомбінантної мікробної клітини-хазяїна, яка містить ферментативний шлях ізобутанолу, що включає молекули ДНК, які кодують набір поліпептидів, що каталізують наступні перетворення субстрату на продукт:
і) піруват на ацетолактат;
іі) ацетолактат на 2,3-дигідроксіізовалерат;
ііі) 2,3-дигідроксіізовалерат на α-кетоізовалерат;
iv) α-кетоізовалерат на валін;
v) валін на ізобутиламін;
vi) ізобутиламін на ізобутиральдегід; та
vi) ізобутиральдегід на ізобутанол,
де щонайменше одна молекула ДНК є гетерологічною для зазначеної мікробної клітини-хазяїна; та
2) контактування клітини-хазяїна (1) з ферментованим вуглецевим субстратом у ферментаційному середовищі в умовах, при яких продукується ізобутанол.
32. Спосіб за п. 31, де ферментований вуглецевий субстрат вибирають з групи, що складається з моносахаридів, олігосахаридів та полісахаридів.
33. Спосіб за п. 31, де вуглецевий субстрат вибирають з групи, що складається з глюкози, сахарози та фруктози.
34. Спосіб за п. 31, де умови, при яких продукується ізобутанол, є анаеробними.
35. Спосіб за п. 31, де умови, при яких продукується ізобутанол, є мікроаеробними.
36. Спосіб за п. 31, де клітина-хазяїн контактує з вуглецевим субстратом у мінімальному середовищі.
37. Спосіб за п. 31, де поліпептид, що каталізує перетворення субстрату на продукт, піруват на ацетолактат, є ацетолактатсинтазою.
38. Спосіб за п. 31, де поліпептид, що каталізує перетворення субстрату на продукт, ацетолактат на 2,3-дигідроксіізовалерат, є ізомероредуктазою ацетогідроксикислот.
39. Спосіб за п. 31, де поліпептид, що каталізує перетворення субстрату на продукт, 2,3-дигідроксіізовалерат на α-кетоізовалерат, є дегідратазою ацетогідроксикислот.
40. Спосіб за п. 31, де поліпептид, що каталізує перетворення субстрату на продукт, ізобутиральдегід на ізобутанол, є дегідрогеназою розгалужених спиртів.
41. Спосіб за п. 31, де поліпептид, що каталізує перетворення субстрату на продукт, α-кетоізовалерат на валін, є трансаміназою.
42. Спосіб за п. 31, де поліпептид, що каталізує перетворення субстрату на продукт, α-кетоізовалерат на валін, є валіндегідрогеназою.
43. Спосіб за п. 31, де поліпептид, що каталізує перетворення субстрату на продукт, валін на ізобутиламін, є валіндекарбоксилазою.
44. Спосіб за п. 31, де поліпептид, що каталізує перетворення субстрату на продукт, ізобутиламін на ізобутиральдегід, є омега-трансаміназою.
45. Спосіб за п. 31, де клітину-хазяїна вибирають з групи, що складається з: бактерії, ціанобактерії, нитчастого гриба та дріжджів.
46. Спосіб за п. 45, де клітина-хазяїн є членом роду, вибраного з групи, що складається з Clostridium, Zymomonas, Escherichia, Salmonella, Rhodococcus, Pseudomonas, Bacillus, Lactobacillus, Enterococcus, Alcaligenes, Klebsiella, Paenibacillus, Arthrobacter, Corynebacterium, Brevibacterium, Pichia, Candida, Hansenula та Saccharomyces.
47. Спосіб за п. 46, де клітина-хазяїн є Escherichia coli.
48. Спосіб за п. 46, де клітина-хазяїн є Alcaligenes eutrophus.
49. Спосіб за п. 46, де клітина-хазяїн є Bacillus licheniformis.
50. Спосіб за п. 46, де клітина-хазяїн є Paenibacillus macerans.
51. Спосіб за п. 46, де клітина-хазяїн є Rhodococcus erythropolis.
52. Спосіб за п. 46, де клітина-хазяїн є Pseudomonas putida.
53. Спосіб за п. 46, де клітина-хазяїн є Bacillus subtilis.
54. Спосіб за п. 46, де клітина-хазяїн є Lactobacillus plantarum.
55. Спосіб за п. 46, де клітину-хазяїна вибирають з групи, що складається з Enterococcus faecium, Enterococcus gatlinarium та Enterococcus faecalis.
56. Спосіб за п. 46, де клітина-хазяїн є Saccharomyces cerevisiae.
57. Спосіб за п. 37, де ацетолактатсинтаза має амінокислотну послідовність, вибрану з групи, що складається з SEQ ID NO:2, SEQ ID NO:178 та SEQ ID NO:180.
58. Спосіб за п. 38, де ізомероредуктаза ацетогідроксикислот має амінокислотну послідовність, вибрану з групи, що складається з SEQ ID NO:43, SEQ ID N0:181, SEQ ID NO:183 та SE0 ID NO:185.
59. Спосіб за п. 39, де дегідратаза ацетогідроксикислот має амінокислотну послідовність, вибрану з групи, що складається з SEQ ID NO:6, SEQ ID NO:186, SEQ ID NO:188 та SEQ ID NO:190.
60. Спосіб за п. 40, де дегідрогеназа розгалужених спиртів має амінокислотну послідовність, вибрану з групи, що складається з SEQ ID NO:10, SEQ ID NO:199, SEQ ID NO:201, SEQ ID NO:203 та SEQ ID NO:204.
61. Спосіб за п. 41, де трансаміназа має амінокислотну послідовність, вибрану з групи, що складається з SEQ ID NO:232, SEQ ID NO:234, SEQ ID NO:236, SEQ ID NO:238 та SEQ ID NO:240.
62. Спосіб за п. 42, де валіндегідрогеназа має амінокислотну послідовність, вибрану з групи, що складається з SEQ ID NO:242 та SEQ ID NO:244.
63. Спосіб за п. 43, де валіндекарбоксилаза має амінокислотну послідовність, описану в SEQ ID NO:246.
64. Спосіб за п. 44, де омега-трансаміназа має амінокислотну послідовність, вибрану з групи, що складається з SEQ ID NO:248, SEQ ID NO:250, SEQ ID NO:252 та SEQ ID NO:254.
65. Спосіб за п. 31, де клітина-хазяїн є факультативним анаеробом.
66. Ферментаційне середовище, що містить ізобутанол, яке одержане за способом за п. 31.
Текст
Реферат: Винахід належить до способу ферментативного продукування спиртів з чотирма атомами вуглецю, який полягає в ферментативному вирощуванні рекомбінантної бактерії, що експресує метаболічний шлях біосинтезу ізобутанолу. UA 103364 C2 (12) UA 103364 C2 UA 103364 C2 5 10 15 20 25 30 35 40 45 50 55 60 Перехресне посилання на споріднену заявку Ця заявка претендує згідно з 35 U.S.C. §119 на пріоритет тимчасової заявки США № 60/721677, поданої 29 вересня 2005 р., та тимчасової заявки США № 60/814470, поданої 16 червня 2006 р. Винахід належить до області промислової мікробіології та продукування спиртів. Більш конкретно, ізобутанол продукують шляхом промислової ферментації рекомбінантного мікроорганізму. Бутанол є важливим технічним хімікатом, використовуваним як домішка до палив, як сировий хімічний реагент в промисловості пластмас та як харчовий екстрагент в промисловості харчових продуктів та смакових речовин. Кожного року ΙΟΙ 2 мільярдів фунтів бутанолу виробляють нафтохімічними методами і потреба в цьому товарному хімікаті буде імовірно зростати. Способи хімічного синтезу ізобутанолу є відомими, такі як процес Оксо (Охо Process), процес Реппе (Reppe Process), та гідрування кротональдегіду (Ullmann's Encyclopedia of th Industrial Chemistry, 6 edition, 2003, Wiley-VCH Verlag GmbH та Co., Weinheim, Germany, Vol. 5, pp. 716-719). Ці процеси використовують вихідні матеріали, одержувані з нафтопродуктів і є звичайно дорогими та екологічно шкідливими. Продукування ізобутанолу з сирових матеріалів рослинного походження буде мінімізувати емісію парникових газів та буде удосконаленням відомого рівня техніки. Ізобутанол продукують біологічно як побічний продукт дріжджового бродіння. Він є компонентом "сивушного масла", що утворюється в результаті неповного метаболізму амінокислот цією групою грибів. Ізобутанол специфічно продукується при катаболізмі L-валіну. Після споживання групи аміну L-валіну як джерела азоту, утворювана α-кетокислота декарбоксилюється та відновлюється до ізобутанолу ферментами так званого метаболічного шляху Ерліха (Dickinson et al., J. Biol. Chem. 273(40): 25752-25756 (1998)). Виходи сивушного масла та/або його компонентів, що досягаються при бродінні напоїв, є типово низькими. Наприклад, повідомляється, що концентрація ізобутанолу, продукованого при пивному бродінні, становить менш ніж 16 частин на мільйон (Garcia et al., Process Biochemistry 29:303-309 (1994)). Додавання екзогенного L-валіну до ферментації підвищує вихід ізобутанолу, як описано Dickinson et al., supra, де повідомляється, що вихід ізобутанолу у 3 г/л одержують шляхом забезпечення L-валіну в концентрації 20 г/л у ферментації. Однак, використання валіну як сировини буде економічно неприпустимим для виробництва ізобутанолу в промисловому масштабі. Біосинтез ізобутанолу безпосередньо з цукрів буде економічно виправданим та стане прогресом у рівні техніки. Досі не повідомлялося про рекомбінантний мікроорганізм, призначений для продукування ізобутанолу. Таким чином, існує потреба в екологічно безпечному, економічно ефективному способі продукування ізобутанолу як єдиного продукту. Даний винахід задовольняє цю потребу завдяки відкриттю рекомбінантного мікроорганізму-хазяїна продукування, який експресує метаболічний шлях біосинтезу ізобутанолу. Винахід пропонує рекомбінантний мікроорганізм, який має сконструйований метаболічний шлях біосинтезу ізобутанолу. Сконструйований мікроорганізм може бути використаний для комерційного продукування ізобутанолу. Відповідно, в одному варіанті втілення винахід пропонує рекомбінантну мікробну клітину-хазяїна, яка включає щонайменше одну молекулу ДНК, що кодує поліпептид, який каталізує субстрат до продукування конверсії, вибраної з групи, що складається з: і) піруват на ацетолактат (стадія метаболічного шляху а) іі) ацетолактат на 2,3-дигідроксіізовалерат (стадія метаболічного шляху b) ііі) 2,3-дигідроксіізовалерат на α-кетоізовалерат (стадія метаболічного шляху с) iv) α-кетоізовалерат на ізобутиральдегід (стадія метаболічного шляху d), і ν) ізобутиральдегід на ізобутанол (стадія метаболічного шляху e) , де щонайменше одна молекула ДНК є гетерологічною для зазначеної мікробної клітинихазяїна і де зазначена мікробна клітина-хазяїн продукує ізобутанол. В іншому варіанті втілення, винахід забезпечує рекомбінантну мікробну клітину-хазяїна, яка включає щонайменше одну молекулу ДНК, що кодує поліпептид, який каталізує конверсію субстрату на продукт, вибрану з групи, що складається з: і) піруват на ацетолактат (стадія метаболічного шляху а) іі) ацетолактат на 2,3-дигідроксіізовалерат (стадія метаболічного шляху b) ііі) 2,3-дигідроксіізовалерат на α-кетоізовалерат (стадія метаболічного шляху с) iv) α-кетоізовалерат на ізобутирил-СоА (стадія метаболічного шляху f) ν) ізобутирил-CoA на ізобутиральдегід (стадія метаболічного шляху g) і 1 UA 103364 C2 5 10 15 20 25 30 35 40 45 50 55 60 vi) ізобутиральдегід на ізобутанол (стадія метаболічного шляху e), де щонайменше одна молекула ДНК є гетерологічною для зазначеної мікробної клітинихазяїна і де зазначена мікробна клітина-хазяїн продукує ізобутанол. В іншому варіанті втілення, винахід забезпечує рекомбінантну мікробну клітину-хазяїна, яка включає щонайменше одну молекулу ДНК, що кодує поліпептид, який каталізує конверсію субстрату на продукт, вибрану з групи, що складається з: і) піруват на ацетолактат (стадія метаболічного шляху а) іі) ацетолактат на 2,3-дигідроксіізовалерат (стадія метаболічного шляху b) ііі) 2,3-дигідроксіізовалерат на α-кетоізовалерат (стадія метаболічного шляху с) iv) α-кетоізовалерат to валін (стадія метаболічного шляху h) ν) валін на ізобутиламін (стадія метаболічного шляху і) vi) ізобутиламін на ізобутиральдегід (стадія метаболічного шляху j) і vii) ізобутиральдегід на ізобутанол (стадія метаболічного шляху e), де щонайменше одна молекула ДНК є гетерологічною для зазначеної мікробної клітинихазяїна і де зазначена мікробна клітина-хазяїн продукує ізобутанол. В іншому варіанті втілення, винахід забезпечує спосіб продукування ізобутанолу, який включає: 1) забезпечення рекомбінантної мікробної клітини-хазяїна, яка включає щонайменше одну молекулу ДНК, що кодує поліпептид, який каталізує конверсію субстрату на продукт, вибрану з групи, що складається з: і) піруват на ацетолактат (стадія метаболічного шляху а) іі) ацетолактат на 2,3-дигідроксіізовалерат (стадія метаболічного шляху b) ііі) 2,3-дигідроксіізовалерат на α-кетоізовалерат (стадія метаболічного шляху с) iv) α-кетоізовалерат на ізобутиральдегід (стадія метаболічного шляху d) і ν) ізобутиральдегід на ізобутанол (стадія метаболічного шляху e), де щонайменше одна молекула ДНК є гетерологічною для зазначеної мікробної клітинихазяїна; і 2) введення в контакт клітини-хазяїна за (і) з ферментованим вуглецевим субстратом у ферментаційному середовищі в умовах, при яких відбувається продукування ізобутанолу. В іншому варіанті втілення, винахід забезпечує спосіб продукування ізобутанолу, який включає: 1) забезпечення рекомбінантної мікробної клітини-хазяїна, яка включає щонайменше одну молекулу ДНК, що кодує поліпептид, який каталізує конверсію субстрату на продукт, вибрану з групи, що складається з: і) піруват на ацетолактат (стадія метаболічного шляху а) іі) ацетолактат на 2,3-дигідроксіізовалерат (стадія метаболічного шляху b) ііі) 2,3-дигідроксіізовалерат на α-кетоізовалерат (стадія метаболічного шляху с) iv) α-кетоізовалерат на ізобутирил-СоА (стадія метаболічного шляху f) ν) ізобутирил-СоА на ізобутиральдегід (стадія метаболічного шляху д), і vi) ізобутиральдегід на ізобутанол (стадія метаболічного шляху e), де щонайменше одна молекула ДНК є гетерологічною для зазначеної мікробної клітинихазяїна; і 2) введення в контакт клітини-хазяїна за (і) з ферментованим вуглецевим субстратом у ферментаційному середовищі в умовах, при яких відбувається продукування ізобутанолу. В іншому варіанті втілення, винахід забезпечує спосіб продукування ізобутанолу, який включає: 1) забезпечення рекомбінантної мікробної клітини-хазяїна, яка включає щонайменше одну молекулу ДНК, що кодує поліпептид, який каталізує конверсію субстрату на продукт, вибрану з групи, що складається з: і) піруват на ацетолактат (стадія метаболічного шляху а) іі) ацетолактат на 2,3-дигідроксіізовалерат (стадія метаболічного шляху b) ііі) 2,3-дигідроксіізовалерат на α-кетоізовалерат (стадія метаболічного шляху с) iv) α-кетоізовалерат на валін (стадія метаболічного шляху h) ν) валін на ізобутиламін (стадія метаболічного шляху і) vi) ізобутиламін на ізобутиральдегід (стадія метаболічного шляху j), і vii) ізобутиральдегід на ізобутанол (стадія метаболічного шляху e), де щонайменше одна молекула ДНК є гетерологічною для зазначеної мікробної клітинихазяїна; і 2) введення в контакт клітини-хазяїна за (і) з ферментованим вуглецевим субстратом у ферментаційному середовищі в умовах, при яких відбувається продукування ізобутанолу. В 2 UA 103364 C2 5 10 альтернативному варіанті втілення винахід забезпечує обмежуюче по ізобутанолу ферментаційне середовище, що продукується способами за винаходом. Винахід може бути повніше зрозумілий з наступного детального опису, фігури та супровідних описів послідовностей, які є частиною даної заявки. Фігура 1 показує шлях біосинтезу ізобутанолу. Стадії, позначені "a", "b", "c", "d", "e", "f", "g", "h", "і", "j" та "k", представляють конверсії субстрату на продукт, описані нижче. Наступні послідовності задовольняють вимогам 37 C.F.R. 1.821-1.825 ("Вимоги до патентних заявок, що містять розкриття нуклеотидних послідовностей та/або амінокислотних послідовностей - Правила щодо послідовностей") та відповідають стандарту Всесвітньої організації інтелектуальної власності (WIPO) ST.25 (1998) та вимогам щодо опису послідовностей ЕРО та РСТ (Правила 5.2 та 49.5(а-біс) і Розділ 208 та Додаток С Адміністративних інструкцій). Символи та формат, використовувані для даних нуклеотидих та амінокислотних послідовностей, відповідають правилам, викладеним в 37 C.F.R. §1.822. Таблиця 1 Опис генів та номери SEQ ID білків Опис Klebsiella pneumoniae budB (а цетол а ктатси нтаза) Bacillus subtilis alsS (ацетолактатсинтаза) Lactococcus lactis als (ацетолактатсинтаза) Ε. coli ilvC (редуктоізомераза ацетогідроксикислоти) S. cerevisiae ILV5 (редуктоізомераза ацетогідроксикислоти) М. maripaludis ilvC (редуктоізомераза гідроксикетокислоти) В. subtilis ilvC (редуктоізомераза ацетогідроксикислоти) Е. coli ilvD (дегідратаза ацетогідроксикислот) S. cerevisiae ILV3 (дегідратаза дигідроксикислот) М. maripaludis ilvD (дегідратаза дигідроксикислот) Б. subtilis ilvD (дегідратаза дигідроксикислот) Lactococcus lactis kivD (декарбоксилаза розгалужених αкетокислот), кодон-оптимізована Lactococcus lactis kivD (декарбоксилаза розгалужених αкетокислот) Lactococcus lactis kdcA (декарбоксилаза розгалужених альфа-кетокислот) Salmonella typhimurium (індолпіруватдекарбоксилаза) Clostridium acetobutylicum pdc (піруватдекарбоксилаза) Ε. coli yqhD (дегідрогеназа розгалужених спиртів) S. cerevisiae YPR1 (2-метилбутиральдегідредуктаза) S. cerevisiae ADH6 (NADPH-залежна дегідрогеназа цинамоїлового спирту) Clostridium acetobutylicum bdhA (NADH-залежна бутанолдегідрогеназа А) Clostridium acetobutylicum bdhB (бутанолдегідрогеназа) В. subtilis bkdAA (дегідрогеназа розгалужених кетокислот, субодиниця Е1) В. subtilis bkdAB (дегідрогеназа розгалужених альфакетокислот, субодиниця Е1) В. subtilis bkdB (дегідрогеназа розгалужених альфакетокислот, субодиниця Е2) В. subtilis IpdV (дегідрогеназа розгалужених альфакетокислот, субодиниця Е3) P. putidabkdAI (дегідрогеназа кетокислот, субодиниця Е1альфа) P. putida bkdA2 (дегідрогеназа кетокислот, субодиниця Е1бета) P. putida bkdB (трансацилаза Е2) 3 SEQ ID NO Нуклеїнова кислота 1 78 179 3 80 182 184 5 83 187 189 SEQ ID NO Пептид 2 178 180 4 181 183 185 6 186 188 190 7 8 191 8 192 193 194 196 9 198 195 197 10 199 200 201 202 203 158 204 205 206 207 208 209 210 211 212 213 214 215 216 217 218 UA 103364 C2 P. putida 1pdV (ліпоаміддегідрогеназа) С. beijerinckii aid (кофермент А ацилююча альдегіддегідрогеназа) С. acetobutylicum adhel (альдегіддегідрогеназа) С. acetobutylicum adhe (алкоголь-альдегіддегідрогеназа) P. putida nahO (ацетальдегіддегідрогеназа) Т. thermophilus (ацетальдегіддегідрогеназа) Ε. coli avtA (валін-піруваттрансаміназа) β. licheniformis avtA (валін-піруваттрансаміназа) Ε. coli HvE (амінотрансфераза розгалужених амінокислот) S. cerevisiae BAT2 (амінотрансфераза розгалужених амінокислот) Μ. thermoautotrophicum (амінотрансфераза розгалужених амінокислот) S. coelicolor (валіндегідрогеназа) β. subtilis bed (лейциндегідрогеназа) S. viridifaciens (валіндекарбоксилаза) A. denitrificans aptA (омегаамінокислота:піруваттрансаміназа) R. eutropha (аланін-піруваттрансаміназа) S. oneidensis (бета-аланін-піруваттрансаміназа) P. putida (бета-аланін-піруваттрансаміназа) S. cinnamonensis icm (ізобутирил-СоА мутаза) S. cinnamonensis істВ (ізобутирил-СоА мутаза) S. coelicolor SC05415 (ізобутирил-СоА мутаза) S. coelicolor SCO4800 (ізобутирил-СоА мутаза) S. avermitilis icm А (ізобутирил-СоА мутаза) S. avermitilis істВ (ізобутирил-СоА мутаза) 5 10 15 20 25 30 219 220 221 222 -223 225 227 229 231 233 235 224 226 228 230 232 234 236 237 238 239 240 241 243 245 242 244 246 247 248 249 251 253 255 257 259 261 263 265 250 252 254 256 258 260 262 264 266 SEQ ID NOs:11-38, 40-69, 72-75, 85-138, 144, 145, 147-157, 159-176 є нуклеотидними послідовностями олігонуклеотидних праймерів клонування, скринінгу або секвенування, використовуваних в описаних тут Прикладах. SEQ ID NO:39 є нуклеотидною послідовністю генного кластера cscBKA, описаного в Прикладі 16. SEQ ID NO:70 є нуклеотидною послідовністю промотора глюкозоізомерази 1.6GI, описаного в Прикладі 13. SEQ ID NO:71 є нуклеотидною послідовністю промотора 1.5GI, описаного в Прикладі 13. SEQ ID NO:76 є нуклеотидною послідовністю промотора GPD, описаного в Прикладі 17. SEQ ID NO:77 є нуклеотидною послідовністю термінатора CYC1, описаного в Прикладі 17. SEQ ID NO:79 є нуклеотидною послідовністю промотора FBA, описаного в Прикладі 17. SEQ ID NO:81 є нуклеотидною послідовністю промотора ADH1, описаного в Прикладі 17. SEQ ID NO:82 є нуклеотидною послідовністю термінатора ADH1, описаного в Прикладі 17. SEQ ID NO:84 є нуклеотидною послідовністю промотора GPM, описаного в Прикладі 17. SEQ ID NO:139 є амінокислотною послідовністю сахарозогідролази (CscA). SEQ ID NO:140 є амінокислотною послідовністю D-фруктокінази (CscK). SEQ ID NG:141 є амінокислотною послідовністю сахарозопермеази (CscB). SEQ !D NO:142 є нуклеотидною послідовністю плазміди pFP988DssPspac, описаної в Прикладі 20. SEQ ID NO:143 є нуклеотидною послідовністю плазміди pFP988DssPgroE, описаної в Прикладі 20. SEQ ID NO:146 є нуклеотидною послідовністю фрагмента вектора pFP988Dss, описаного в Прикладі 20. SEQ ID NO:177 є нуклеотидною послідовністю інтеграційного вектора pFP988, описаного в Прикладі 21. SEQ ID NO:267 є нуклеотидною послідовністю плазміди рС194, описаної в Прикладі 21. Даний винахід стосується способів продукування ізобутанолу з використанням рекомбінантних мікроорганізмів. Даний винахід задовольняє ряд комерційних та промислових потреб. Бутанол є важливою промисловою хімічною речовиною з різноманітними застосуваннями, особливо важливими з яких є його потенціал як палива або домішки до 4 UA 103364 C2 5 10 15 20 25 30 35 40 45 50 55 60 палива. Хоча він є спиртом з лише чотирма атомами вуглецю, бутанол має енерговміст, близький до бензину і може бути змішаний з будь-яким викопним паливом. Бутанол має переваги при використанні як паливо або домішка до палива, тому що при його сгорянні в стандартному двигуні внутрішнього згоряння утворюється лише СO2 і незначна кількість або зовсім не утворюються SOx чи NOx. Додатково, бутанол є менш корозійним, ніж етанол, найкраща домішка до палива на сьогодні. На додаток до своєї придатності як біопаливо або домішка до палива, бутанол має потенціал впливу на вирішення проблем постачання водню у промисловості паливних елементів, що розвивається. Паливні елементи зараз переслідує занепокоєння щодо безпеки, асоційоване з транспортуванням та розповсюдженням водню. Бутанол може бути легко підданий крекінгу для видобутку його вмісту водню і може бути розповсюджений через існуючі автозаправні станції з чистотою, потрібною для паливних елементів або автомобілів. Зрештою, даний винахід продукує бутанол з джерел вуглецю рослинного походження, уникаючи негативного впливу на навколишнє середовище, асоційованого зі стандартними нафтохімічними процесами продукування бутанолу. Наступні визначення та абревіатури мають використовуватися для інтерпретації формули та опису винаходу. Термін "винахід" або "даний винахід" в тому значенні, що використовується тут, є необмежувальним терміном і не має стосуватися будь-якого окремого варіанта втілення певного винаходу, але охоплює усі можливі варіанти втілення, описані в описі та формулі винаходу. Термін "метаболічний шлях біосинтезу ізобутанолу" стосується ферментативних метаболічних шляхів продукування ізобутанолу. Терміни "ацетолактатсинтаза" та "ацетолактатсинтетаза" використовуються тут взаємозамінно для позначення ферменту, який каталізує конверсію пірувату на ацетолактат та СО2. Кращі ацетолактатсинтази відомі під номером EC 2.2.1.69 (Enzyme Nomenclature 1992, Academic Press, San Diego). Ці ферменти є доступними з ряду джерел, включаючи, але без обмеження, Bacillus subtilis (GenBank Nos: CAB15618 (SEQ ID NO:178), Z99122 (SEQ ID NO:78), NCBI (National Center for Biotechnology Information) амінокислотна послідовність, NCBI нуклеотидна послідовність, відповідно), Klebsiella pneumoniae [GenBank Nos: AAA25079 (SEQ ID NO:2), M73842 (SEQ ID NO:1)) і Lactococcus lactis (GenBank Nos: AAA25161 (SEQ ID NO:180), L16975 SEQ IDNO:179)). Терміни "ізомероредуктаза ацетогідроксикислот" та "редуктоізомераза ацетогідроксикислот" використовуються тут взаємозамінно для позначення ферменту, який каталізує конверсію ацетолактату на 2,3-дигідроксіізовалерат з використанням NADPH (відновлений нікотинамідаденіндинуклеотидфосфат) як електронного донора. Кращі ізомероредуктази ацетогідроксикислот відомі під номером EC 1.1.1.86, а послідовності є доступними від величезної кількості мікроорганізмів, включаючи, без обмеження, Escherichia со//(GenBank Nos: NP_418222 (SEQ ID N0:4), NC_000913 (SEQ ID NO:3)), Saccharomyces cerevisiae (GenBank Nos: NP_013459 (SEQ ID NO:181), NC_001144 (SEQ ID NO:80)), Methanococcus maripaludis (GenBank Nos: CAF30210 (SEQ ID NO:183), BX957220 (SEQ ID NO:182)) і Bacillus subtilis (GenBank Nos: CAB14789 (SEQ ID NO:185), Z99118 (SEQ ID NO:184)). Термін "дегідратаза ацетогідроксикислот" стосується ферменту, який каталізує конверсію 2,3-дигідроксіізовалерату на α-кетоізовалерат. Кращі дегідратази ацетогідроксикислот відомі під номером EC 4.2.1.9. Ці ферменти є доступними від величезної кількості мікроорганізмів, включаючи, без обмеження, Е. coli (GenBank Nos: YPJ326248 (SEQ ID NO:6), NC_000913 (SEQ ID NO:5)), S. cerevisiae (GenBank Nos: NP_012550 (SEQ ID NO:186), NC_001142 (SEQ ID NO:83)), M. maripaludis (GenBank Nos: CAF29874 (SEQ ID N0:188), BX957219 (SEQ ID NO: 187)) і В. subtilis (GenBank Nos: CAB14105 (SEQ ID NO:190), Z99115 (SEQ ID ΝΟ.Ί89)). Термін "декарбоксилаза розгалужених α-кетокислот" стосується ферменту, який каталізує конверсію α-кетоізовалерату на ізобутиральдегід та СО2. Кращі декарбоксилази розгалужених α-кетокислот відомі під номером EC 4.1.1.72 та є доступними з ряду джерел, включаючи, але без обмеження, Lactococcus lactis (GenBank Nos: AAS49166 (SEQ ID NO: 193), AY548760 (SEQ ID N0:192); CAG34226 (SEQ ID N0:8), AJ746364 (SEQ ID N0:191), Salmonella typhimurium (GenBank Nos: NP_461346 (SEQ ID N0:195), MCJ303197 (SEQ ID N0:194)) і Clostridium acetobutylicum (GenBank Nos: NPJ49189 (SEQ ID N0:197), NCJD01988 (SEQ ID N0:196)). Термін "дегідрогеназа розгалужених спиртів" стосується ферменту, який каталізує конверсію ізобутиральдегіду на ізобутанол. Кращі дегідрогенази розгалужених спиртівв відомі під номером EC 1.1.1.265, але можуть також бути класифіковані як інші алкогольдегідрогенази (конкретніше, EC 1.1.1.1 або 1.1.1.2). Ці ферменти використовують NADH (відновлений 5 UA 103364 C2 5 10 15 20 25 30 35 40 45 50 55 60 нікотинамідаденіндинуклеотид) та/або NADPH як електронний донор та є доступними з ряду джерел, включаючи, але без обмеження, S. cerevisiae (GenBank Nos: NP_010656 (SEQ ID NO:199), NC_001136 (SEQ ID NO:198); NPJ314051 (SEQ ID NO:201) NC_001145 (SEQ ID NO:200)), E. coll (GenBank Nos: NP_417484 (SEQ ID N0:10), NC_000913 (SEQ ID N0:9)) і С. acetobutylicum (GenBank Nos: NP_349892 (SEQ ID NO:203), NCJD03030 (SEQ ID N0:202); NP_349891 (SEQ ID N0:204), NCJD03030 (SEQ ID N0:158)). Термін "дегідрогеназа розгалужених кетокислот" стосується ферменту, який каталізує конверсію α-кетоізовалерату на ізобутирил-СоА (ізобутирил-кофермент А), використовуючи + NAD (нікотинамідаденіндинуклеотид) як акцептор електронів. Кращі дегідрогенази розгалужених кетокислот відомі під номером ЕС 1.2.4.4. Ці дегідрогенази розгалужених кетокислот складаються з чотирьох субодиниць та послідовності від усіх субодиниць є доступними від величезної кількості мікроорганізмів, включаючи, але без обмеження, В. subtilis (GenBank Nos: CAB14336 (SEQ ID N0:206), Z99116 (SEQ ID NO:205); CAB14335 (SEQ ID NO:208), Z99116 (SEQ ID NO:207); CAB14334 (SEQ ID N0:210), Z99116 (SEQ ID NO:209); і САВ14337 (SEQ IDNO:212), Z99116 (SEQ ID NO:211)) та Pseudomonas putida (GenBank Nos: AAA65614 (SEQ ID NO:214), M57613 (SEQ ID NO:213); AAA65615 (SEQ ID NO:216), M57613 (SEQ ID NO:215); AAA65617 (SEQ ID NO:218), M57613 (SEQ ID NO:217); та ААА65618 (SEQ ID NO:220), M57613 (SEQ ID NO:219)). Термін "ацилююча альдегіддегідрогеназа" стосується ферменту, який каталізує конверсію ізобутирил-СоА на ізобутиральдегід, з використанням NADH або NADPH як електронного донора. Кращі ацилюючі альдегіддегідрогенази відомі під номерами EC 1.2.1.10 та 1.2.1.57. Ці ферменти є доступними з численних джерел, включаючи, але без обмеження, Clostridium beijerinckii (GenBank Nos: AAD31841 (SEQ ID NO:222), AF157306 (SEQ ID NO:221)), С acetobutylicum (GenBank Nos: NPJ49325 (SEQ ID NO:224), NC_001988 (SEQ ID NO:223); NPJ49199 (SEQ ID NO:226), NC_001988 (SEQ ID NO:225)), P. putida (GenBank Nos: AAA89106 (SEQ ID NO:228), U13232 (SEQ ID NO:227)) і Thermus thermophilus (GenBank Nos: YPJ45486 (SEQ ID NO:230), NC_006461 (SEQ ID NO:229)). Термін "трансаміназа" стосується ферменту, який каталізує конверсію а-кетоізовалерату на L-валін, з використанням аланіну або глутамату як донора аміну. Кращі трансамінази відомі під номерами EC 2.6.1.42 та 2.6.1.66. Ці ферменти є доступними з ряду джерел. Приклади джерел аланін-залежних ферментів включають, без обмеження, Е. coli (GenBank Nos: YP_026231 (SEQ ID NO:232), NC_000913 (SEQ ID NO:231)) та Bacillus licheniformis (GenBank Nos: YP_093743 (SEQ ID NO:234), NC_006322 (SEQ ID N0:233)). Приклади джерел глутамат-залежних ферментів включають, без обмеження, Е. coli (GenBank Nos: YP_026247 (SEQ ID NO:236), NC_000913 (SEQ ID NO:235)), S. cerevisiae (GenBank Nos: NP_012682 (SEQ ID NO:238), NC_001142 (SEQ ID NO:237)) та Methanobacterium thermoautotrophicum (GenBank Nos: NP_276546 (SEQ ID NO:240), NC_000916 (SEQ ID NO:239)). Термін "валіндегідрогеназа" стосується ферменту, який каталізує конверсію акетоізовалерату на L-валін, з використанням NAD(P)H як електронного донора та аміаку як донора аміну. Кращі валіндегідрогенази відомі під номерами EC 1.4.1.8 та 1.4.1.9 і є доступними з ряду джерел, включаючи, без обмеження, Streptomyces coelicolor (GenBank Nos: NP_628270 (SEQ ID NO:242), NC_003888 (SEQ ID NO:241)) та В. subtilis (GenBank Nos: CAB14339 (SEQ ID NO:244), Z99116 (SEQ ID NO:243)). Термін "валіндекарбоксилаза" стосується ферменту, який каталізує конверсію L-валіну на ізобутиламін та СОг. Кращі валіндекарбоксилази відомі під номером EC 4.1.1.14. Ці ферменти знайдені в Streptomycetes, таких як, наприклад, Streptomyces viridifaciens (GenBank Nos: AAN10242 (SEQ ID NO:246), AY116644 (SEQ ID NO:245)). Термін "омега-трансаміназа" стосується ферменту, який каталізує конверсію ізобутиламіну на ізобутиральдегід з використанням придатної амінокислоти як донора аміну. Кращі омегатрансамінази відомі під номером EC 2.6.1.18 та є доступними з ряду джерел, включаючи, але без обмеження, Alcaligenes denitrificans (AAP92672 (SEQ ID NO:248), AY330220 (SEQ ID N0:247)), Ralstonia eutropha (GenBank Nos: YP_294474 (SEQ ID NO:250), NC_007347 (SEQ ID NO:249)), Shewanella oneidensis (GenBank Nos: NP_719046 (SEQ ID NO:252), NC_004347 (SEQ ID NO:251)) і P. putida (GenBank Nos: AAN66223 (SEQ ID NO-.254), AE016776 (SEQ ID NO.253)). Термін "ізобутирил-СоА мутаза" стосується ферменту, який каталізує конверсію бутирилСоА на ізобутирил-СоА. Цей фермент використовує кофермент Ві2 як кофактор. Кращі ізобутирил-СоА мутази відомі під номером ЕС 5.4.99.13. Ці ферменти знайдені в ряді Streptomycetes, включаючи, але без обмеження, Streptomyces cinnamonensis (GenBank Nos: AAC08713 (SEQ ID NO:256), U67612 (SEQ ID NO:255); CAB59633 (SEQ ID NO:258), AJ246005 (SEQ ID NO:257)), S. coelicolor (GenBank Nos: CAB70645 (SEQ ID NO:260), AL939123 (SEQ ID 6 UA 103364 C2 5 10 15 20 25 30 35 40 45 50 55 60 NO:259); CAB92663 (SEQ ID NO:262), AL939121 (SEQ ID NO:261)) і Streptomyces avermitilis (GenBank Nos: NP_824008 (SEQ ID NO:264), NC_003155 (SEQ ID NO:263); NP_824637 (SEQ ID NO:266), NC_003155 (SEQ ID NO:265)). Термін "факультативний анаероб" стосується мікроорганізму, який може рости як в аеробному, так і в анаеробному середовищі. Термін "вуглецевий субстрат" або "ферментований вуглецевий субстрат" стосується джерела вуглецю, здатного метаболізуватися організмами-хазяїнами за даним винаходом, і особливо джерел вуглецю, вибраних з групи, що складається з моносахаридів, олігосахаридів, полісахаридів та субстратів з одним атомом вуглецю або їх сумішей. Термін "ген" стосується фрагмента нуклеїнової кислоти який є здатним експресуватися як специфічний білок, необов'язково включаючи регуляторні послідовності, розташовані перед (5'некодуючі послідовності) та після (3'-некодуючі послідовності) кодуючої послідовності. "Нативний ген" стосується гена, як він існує в природі з його власними регуляторними послідовностями. "Хімерний ген" стосується будь-якого гена, що не є нативним геном, який включає регуляторні та кодуючі послідовності, що не зустрічаються разом у природі. Відповідно, хімерний ген може включати регуляторні послідовності та кодуючі послідовності, одержані з різних джерел, або регуляторні послідовності та кодуючі послідовності, одержані з одного джерела, але розташовані у спосіб, відмінний від того, як вони зустрічаються у природі. "Ендогенний ген" стосується нативного гена в його природному місцезнаходженні в геномі організму. "Чужорідний" або "гетерологічний ген" стосується гена, звичайно не присутнього в організмі-хазяїні, але введеного в організм-хазяїн перенесенням гена. Чужорідні гени можуть включати нативні гени, введені в ненативний організм, або хімерні гени. "Трансген" є геном, який був введений в геном за процедурою трансформації. В тому значенні, що використовується тут, термін "кодуюча послідовність" стосується послідовності ДНК, яка кодує специфічну амінокислотну послідовність. "Придатні регуляторні послідовності" стосуються нуклеотидних послідовностей, розташованих перед (5'-некодуючі послідовності), усередині або після (3'-некодуючі послідовності) кодуючої послідовності, які впливають на транскрипцію, процесинг чи стабільність РНК або трансляцію асоційованої кодуючої послідовності. Регуляторні послідовності можуть включати промотори, лідерні послідовності трансляції, інтрони, послідовності розпізнавання поліаденілювання, сайт процесингу РНК, сайт зв'язування ефектора та структуру стебло-петля. Термін "промотор" стосується послідовності ДНК, здатної контролювати експресію кодуючої послідовності або функціональної РНК. Загалом, кодуюча послідовність розташована з 3'-боку від промоторної послідовності. Промотори можуть бути цілком виділені з нативного гена або складатися з різних елементів, одержаних з різних промоторів, що зустрічаються в природі, або навіть включати синтетичні сегменти ДНК. Фахівцям в цій області техніки зрозуміло, що різні промотори можуть керувати експресією гена в різних тканинах чи типах клітин або на різних стадіях розвитку або у відповідь на різні навколишні чи фізіологічні умови. Промотори, які спричинюють експресію гена в більшості типів клітин у більшості випадків, звичайно називаються "конститутивні промотори". Далі зрозуміло, що оскільки у більшості випадків точні межі регуляторних послідовностей не були повністю визначені, фрагменти ДНК різної довжини можуть мати ідентичну промоторну активність. Термін "функціонально зв'язаний" стосується асоціації послідовностей нуклеїнових кислот на окремому фрагменті нуклеїнової кислоти таким чином, що функція одного впливає на інший. Наприклад, промотор є функціонально зв'язаним з кодуючою послідовністю, якщо він є здатним впливати на експресію цієї кодуючої послідовності (тобто, якщо кодуюча послідовність перебуває під транскрипційним контролем промотора). Кодуючі послідовності можуть бути функціонально зв'язаними з регуляторними послідовностями у смисловій чи антисмисловій орієнтації. Термін "експресія", в тому значенні, що використовується тут, стосується транскрипції та стабільного накопичення смислової (мРНК) або антисмислової РНК, похідної з фрагмента нуклеїнової кислоти за винаходом. Експресія може також стосуватися трансляції мРНК в поліпептид. В тому значенні, що використовується тут, термін "трансформація" стосується перенесення фрагмента нуклеїнової кислоти в організм-хазяїн, приводячи до генетично стабільної спадковості. Організми-хазяїни, що містять трансформовані фрагменти нуклеїнової кислоти, називаються "трансгенними" або "рекомбінантними" або "трансформованими" організмами. Терміни "плазміда", "вектор" та "касета" стосуються позахромосомального елемента, який часто несе гени, що не є частиною центрального метаболізму клітини, і звичайно мають форму кругових дволанцюгових фрагментів ДНК. Такі елементи можуть бути послідовностями, що 7 UA 103364 C2 5 10 15 20 25 30 35 40 45 50 55 60 автономно реплікуються, геном-інтегруючими послідовностями, фаговими або нуклеотидними послідовностями, лінійними або круговими, одно- або дволанцюгових ДНК або РНК, одержаних з будь-якого джерела, у якому ряд нуклеотидних послідовностей були з'єднані або рекомбіновані в унікальну конструкцію, що є здатною вводити промоторний фрагмент та послідовність ДНК для вибраного генного продукта разом з відповідною 3'-нетрансльованою послідовністю в клітину. "Касета трансформації" стосується специфічного вектора, що містить чужорідний ген та має елементи на додаток до чужорідного гена, які сприяють трансформації конкретної клітини-хазяїна. "Касета експресії" стосується специфічного вектора, що містить чужорідний ген та має елементи на додаток до чужорідного гена, які забезпечують посилену експресію цього гена в чужорідному хазяїні. В тому значенні, що використовується тут, термін "виродженість кодону" стосується природи генетичного коду, що дозволяє варіації нуклеотидної послідовності без впливу на амінокислотну послідовність кодованого поліпептиду. Кваліфікованому фахівцю добре відомо про "неоднозначність кодона", яку виявляє специфічна клітина-хазяїн при використанні нуклеотидних кодонів для визначення даної амінокислоти. Тому, при синтезі гена для поліпшеної експресії в клітині-хазяїні, бажано конструювати ген таким чином, щоб його частота використання кодона наближалася до кращої частоти використання кодона клітини-хазяїна. Термін "кодон-оптимізований", коли це стосується генів або кодуючих ділянок молекул нуклеїнової кислоти для трансформації різних хазяїнів, стосується альтерації кодонів у гені або кодуючих ділянках молекул нуклеїнової кислоти для відображення типового використання кодона організму-хазяїна без зміни поліпептиду, кодованого ДНК. Стандартні рекомбінантні методики клонування ДНК та молекулярного клонування, використовувані тут, є добре відомими фахівцям та описані Sambrook, J., Fritsch, E.F. and Maniatis, Т., Molecular Cloning: A Laboratory Manual, Second Edition, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY (1989) (тут та надалі "Maniatis"); та Silhavy, T.J., Bennan, M.L. and Enquist, L. W., Experiments with Gene Fusions, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY (1984); та Ausubel, F.M. et al., Current Protocols in Molecular Biology, опублікована Greene Publishing Assoc. та Wiley-Interscience (1987). Метаболічні шляхи біосинтезу ізобутанолу Мікроорганізми, що засвоюють вуглеводи, використовують метаболічний шлях ЕмбденаМейерхофа-Памаса (Embden-Meyerhof-Pamas, EMP), метаболічний шлях Ентнера-Дудороффа (Entner-Doudoroff) та пентозофосфатний цикл як головні метаболічні шляхи забезпечення енергії та клітинних прекурсорів для росту та підтримання. Ці шляхи мають спільним те, що проміжний гліцеральдегід-3-фосфат і, в остаточному підсумку, піруват утворюється безпосередньо або в комбінації з ЕМР шляхом. Згодом, піруват трансформується на ацетил-кофермент А (ацетил-СоА) різноманітними способами. Ацетил-СоА служить ключовим проміжним продуктом, наприклад, при утворенні жирних кислот, амінокислот та вторинних метаболітів. Об'єднані реакції конверсії цукрів на піруват продукують енергію (наприклад, аденозин-5'-трифосфат, АТР) та відновні еквіваленти (наприклад, відновлений нікотинамідаденіндинуклеотид, NADH, та відновлений нікотинамідаденіндинуклеотидфосфат, + + NADPH). NADH та NADPH мають бути рециркульовані в їх окиснені форми (NAD та NADP , 2відповідно). В присутності неорганічних акцепторів електронів (наприклад, Ог, NO3 та SO4 ), відновлювальні еквіваленти можуть бути використані для збільшення енергетичного пула; альтернативно, може бути утворений відновлений вуглецевий побічний продукт. Даний винахід дозволяє продукувати ізобутанол з вуглеводних джерел за допомогою рекомбінантних мікроорганізмів шляхом забезпечення чотирьох повних реакційних шляхів, як показано на Фігурі 1. Три з метаболічних шляхів включають конверсію пірувату на ізобутанол через ряд ферментативних стадій. Кращий метаболічний шлях ізобутанолу (Фігура 1, стадії а-е) включає такі конверсії субстрату на продукт: a) піруват на ацетолактат, каталізована, наприклад, ацетолактатсинтазою, b) ацетолактат на 2,3-дигідроксіізовалерат, каталізована, наприклад, ізомероредуктазою ацетогідроксикислот, c) 2,3-дигідроксіізовалерат на α-кетоізовалерат, каталізована, наприклад, дегідратазою ацетогідроксикислот, d) α-кетоізовалерат на ізобутиральдегід, каталізована, наприклад, декарбоксилазою розгалужених кетокислот, і e) ізобутиральдегід на ізобутанол, каталізована, наприклад, дегідрогеназою розгалужених спиртів. Цей метаболічний шлях об'єднує ферменти, що, як відомо, беруть участь в добре охарактеризованих метаболічних шляхах біосинтезу валіну (піруват на α-кетоізовалерат) та 8 UA 103364 C2 5 10 15 20 25 30 35 40 45 50 55 катаболізму валіну (α-кетоізовалерат на ізобутанол). Оскільки багато ферментів біосинтезу валіну також каталізують аналогічні реакції у метаболічному шляху біосинтезу ізолейцину, специфічність щодо субстрату є важливим міркуванням у виборі генних джерел. З цієї причини, первинними цікавими генами для ферменту ацетолактатсинтази є гени з Bacillus (alsS) та Klebsiella (budB). Відомо, що ці ацетолактатсинтази беруть участь у ферментації бутандіолу в цих організмах та виявляють підвищену спорідненість до пірувату у порівнянні з кетобутиратом (Gollop et al., J. Bacteriol. 172(6):3444-3449 (1990); Holtzclaw et al., J. Bacteriol. 121(3):917-922 (1975)). Друга та третя стадії метаболічного шляху каталізуються редуктоізомеразою ацетогідроксикислот та дегідратазою, відповідно. Ці ферменти були охарактеризовані з ряду джерел, таких як, наприклад, Е. coli (Chunduru et al., Biochemistry 28(2):486-493 (1989); Flint et al., J. Biol. Chem. 268(29):14732-14742 (1993)). Кінцеві дві стадії кращого метаболічного шляху ізобутанолу, як відомо, спостерігаються у дріжджах, які можуть використовувати валін як джерело азоту і, у процесі, секретують ізобутанол α-Кетоізовалерат може бути перетворений на ізобутиральдегід за допомогою ряду ферментів декарбоксилази кетокислот, таких як, наприклад, піруватдекарбоксилаза. Для запобігання використанню пірувату не для продукування ізобутанолу, бажаною є декарбоксилаза зі зниженою спорідненістю до пірувату. Досі, у відомому рівні техніки існують два таких ферменти (Smit et al., Appl. Environ. Microbiol. 71(1):303-311 (2005); de la Plaza et al., FEMS Microbiol. Lett. 238(2):367-374 (2004)). Обидва ферменти походять зі штамів Lactococcus lactis та виявляють 50-200-кратну перевагу до кетоізовалерату у порівнянні з піруватом. Зрештою, ряд альдегідредуктаз був ідентифікований у дріжджах, багато з них з перекривною субстратною специфічністю. Ті, що виявляють перевагу до розгалужених субстратів у порівнянні з ацетальдегідом, включають, без обмеження, алкогольдегідрогеназу VI (ADH6) та Yprip (Larroy et al., Biochem. J. 361 (Pt 1): 163-172 (2002); Ford et al., Yeast 19(12): 1087-1096 (2002)), які обидва використовують NADPH як електронний донор. NADPH-залежна редуктаза, YqhD, активна відносно до розгалужених субстратів, також була недавно ідентифікована в Е. coli (Sulzenbacheretal., J. Мої. Biol. 342(2):489-502 [2004)). Інший метаболічний шлях для конверсії пірувату на ізобутанол включає такі конверсії субстрату на продукт (Фігура 1, стадії a,b,c,f,g,e): a) піруват на ацетолактат, каталізована, наприклад, ацетолактатсинтазою, b) ацетолактат на 2,3-дигідроксіізовалерат, каталізована, наприклад, ізомероредуктазою ацетогідроксикислот, c) 2,3-дигідроксіізовалерат на α-кетоізовалерат, каталізована, наприклад, дегідратазою ацетогідроксикислот, f) α-кетоізовалерат на ізобутирил-СоА, каталізована, наприклад, дегідрогеназою розгалужених кетокислот, g) ізобутирил-СоА на ізобутиральдегід, каталізована, наприклад, ацилюючою альдегіддегідрогеназою, і e) ізобутиральдегід на ізобутанол, каталізована, наприклад, дегідрогеназою розгалужених спиртів. Перші три стадії в цьому метаболічному шляху (а,b,с) є такими саме, як описані вище. αкетоізовалерат перетворюється на ізобутирил-СоА під дією дегідрогенази розгалужених кетокислот. Якщо дріжджі можуть лише використовувати валін як джерело азоту, багато інших організмів (як еукаріоти, так і прокаріоти) можуть також використовувати валін як джерело вуглецю. Ці організми мають дегідрогеназу розгалужених кетокислот (Sokatch et al. J. Bacteriol. 148(2):647-652 (1981)), яка генерує ізобутирил-СоА. Ізобутирил-СоА може бути перетворений на ізобутиральдегід за допомогою ацилюючої альдегіддегідрогенази. Дегідрогенази, активні по відношенню до розгалужених субстратів, були описані, але не клоновані, в Leuconostoc та Propionibaderium (Kazahaya et al., J. Gen. Appl. Microbiol. 18:43-55 (1972); Hosoi et al., J. Ferment. Technol. 57:418-427 (1979)). Однак, можливо також, що ацилюючі альдегіддегідрогенази, що функціонують для лінійних ацил-СоА (наприклад, бутирил-СоА), можуть також працювати з ізобутирил-СоА. Ізобутиральдегід потім перетворюється на ізобутанол дегідрогеназою розгалужених спиртів, як описано вище для першого метаболічного шляху. Інший метаболічний шлях для конверсії пірувату на ізобутанол включає такі конверсії субстрату на продукт (Фігура 1, стадії i,b,c,h,i,j,e): a) піруват на ацетолактат, каталізована, наприклад, ацетолактатсинтазою, b) ацетолактат на 2,3-дигідроксіізовалерат, каталізована, наприклад, ізомероредуктазою ацетогідроксикислот, c) 2,3-дигідроксіізовалерат на α-кетоізовалерат, каталізована, наприклад, дегідратазою ацетогідроксикислот, 9 UA 103364 C2 5 10 15 20 25 30 35 40 h) α-кетоізовалерат на валін, каталізована, наприклад, валіндегідрогеназою або трансаміназою, і) валін на ізобутиламін, каталізована, наприклад, валіндекарбоксилазою, j) ізобутиламін на ізобутиральдегід, каталізована, наприклад, омега-трансаміназою, і e) ізобутиральдегід на ізобутанол, каталізована, наприклад, дегідрогеназою розгалужених спиртів. Перші три стадії в цьому метаболічному шляху (а,b,с) є такими саме, як описані вище. Цей метаболічний шлях потребує додавання валіндегідрогенази або придатної трансамінази. Валін (та/або лейцин) дегідрогеназа каталізує відновне амінування та використовує аміак; значення К т для аміаку перебувають в мілімолярному діапазоні (Priestly et al., Biochem J. 261(3):853-861 (1989); Vancura et al., J. Gen. Microbiol. 134(12):3213-3219 (1938) Zink et al., Arch. Biochem. Biophys. 99:72-77 (1962); Sekimoto et al. J. Biochem (Japan) 116(1):176-182 (1994)). Трансамінази типово використовують глутамат або аланін як донори аміногруп і були охарактеризовані для ряду організмів (Lee-Peng et al.,. J. Bacteriol. 139(2):339-345 (1979); Berg et al., J. Bacteriol. 155(3):1009-1014 (1983)). Аланін-специфічний фермент може бути бажаним, оскільки генерування пірувату з цієї стадії може бути спряжене зі споживанням пірувату пізніше у метаболічному шляху, коли група аміну видаляється (див. нижче). Наступною стадією є декарбоксилювання валіну, реакція, що відбувається при біосинтезі валаніміцину в Streptomyces (Garg et al., Мої. Microbiol. 46(2):505-517 (2002)). Утворюваний ізобутиламін може бути перетворений на ізобутиральдегід шляхом піридоксаль-5'-фосфат-залежної реакції за допомогою, наприклад, ферменту із сімейства омега-амінотрансфераз. Такий фермент з Vibrio fluvialis демонстрував активність з ізобутиламіном (Shin et al., Biotechnol. Bioeng. 65(2):206-211 (1999)). Інша омега-амінотрансфераза з Alcaligenes denitrificans була клонована та виявляє деяку активність відносно до бутиламіну (Yun et al., Appl. Environ. Microbiol. 70(4):2529-2534 (2004)). В цьому напрямку, ці ферменти використовують піруват як акцептор аміногруп, утворюючи аланін. Як згадувалося вище, небажаному впливу на піруватний пул можна запобігти шляхом використання піруват-продукуючої трансамінази ранше у метаболічному шляху. Ізобутиральдегід потім перетворюється на ізобутанол дегідрогеназою розгалужених спиртів, як описано вище для першого метаболічного шляху. Четвертий метаболічний шлях біосинтезу ізобутанолу включає конверсії субстрату на продукт, зображені як стадії k,g,e на Фігурі 1. Відомо, що ряд організмів продукують бутират та/або бутанол через проміжний бутирил-СоА (Durre et al., FEMS Microbiol. Rev. 17(3):251-262 (1995); Abbad-Andaloussi et al., Microbiology 142(5):1149-1158 (1996)). Продукування ізобутанолу може бути викликане в цих організмах шляхом додавання мутази, здатної перетворювати бутирил-СоА на ізобутирил-СоА (Фігура 1, стадія к). Гени для обох субодиниць ізобутирил-СоА мутази, кофермент Віг-залежного ферменту, були клоновані з Streptomycete (Ratnatilleke et al., J. Biol. Chem. 274(44):31679-31685 (1999)). Ізобутирил-СоА перетворюється на ізобутиральдегід (стадія g на Фігурі 1), який перетворюється на ізобутанол (стадія є на Фігурі 1). Таким чином, при забезпеченні численних рекомбінантних метаболічних шляхів від пірувату до ізобутанолу, існує ряд можливостей виконання індивідуальних стадій конверсії, і фахівець в цій області техніки зможе використати публічно доступні послідовності для конструювання відповідних метаболічних шляхів. Перелік ряду типових генів, відомих фахівцям, та придатних для конструювання метаболічних шляхів біосинтезу ізобутанолу наведений нижче у Таблиці 2. 10 UA 103364 C2 Таблиця 2 Джерела генів метаболічного шляху біосинтезу ізобутанолу Ген Посилання GenBank Z99122, Bacillus subtilis повний геном (секція 19 з 21): від 3608981 до 3809670 gi|32468830|emb|Z99122.2|BSUB0019[32468830] М73842, Klebsiella pneumoniae ген ацетолактатсинтази (Ник), повні cds (кодуючі послідовності) gi|149210|gb|M73842.1|KPNILUK[149210] L16975, Lactococcus lactis ген альфа-ацетолактатсинтази (als), повні cds gi|473900|gb|L16975.1|LACALS[473900] NC_000913, Escherichia coli K12, повний геном gi|49175990|ref|NC_000913.2Ц49175990] NC_001144, Saccharomyces cerevisiae хромосома XII, повна хромосомна послідовність gi|42742286[ref|NC_001144.3|[42742286] ВХ957220, Methanococcus maripaludis S2 повний геном; сегмент 2/5 gi|44920669|emb|BX957220.1|[44920669] Z99118, Bacillus subtilis повний геном (секція 15 з 21): від 2812801 до 3013507 gi|32468802|emb|Z99118.2|BSUB0015[32468802] NC_000913, Escherichia coli K12, повний геном gi|49175990|ref|NC_000913.2|[49175990] NC_001142, Saccharomyces cerevisiae хромосома X, повна хромосомна послідовність gi|42742252|ref|NC_001142.5|[42742252] ВХ957219, Methanococcus maripaludis S2 повний геном; сегмент 1/5 gi|45047123|emb|BX957219.1|[45047123] Z99115, Bacillus subtilis повний геном (секція 12 з 21): від 2207806 до 2409180 gi|32468778|emb|Z99115.2|BSUB0012[32468778] ацетолактатсинтаза ізомероредуктаза ацетогідроксикислот дегідратаза ацетогідроксикислот декарбоксилаза розгалужених кетокислот α дегідрогеназа розгалужених спиртів дегідрогеназа розгалужених кетокислот ацилююча альдегіддегідрогеназ а ΑΥ548760, Lactococcus lactis ген декарбоксилази розгалужених альфакетокислот (kdcA), повні cds gi|44921616|gb|AY548760.11[44921616] AJ746364, Lactococcus lactis subsp. lactis kivd ген альфакетоізовалератдекарбоксилази, штам IFPL730 gi|51870501|emb|AJ746364.1|[51870501] NCJ303197, Salmonella typhimurium LT2, повний геном gi|16763390|ref|NC_003197.1|[16763390] NC_001988, Clostridium acetobutylicum ATCC 824 плазміда pSOL1, повна послідовність gi|15004705|ref|NC_001988.2|[15004705] NC_001136, Saccharomyces cerevisiae хромосома IV, повна хромосомна послідовність gi|50593138|ref|NC_00113B.6|[50593138] NC_001145, Saccharomyces cerevisiae хромосома XIII, повна хромосомна послідовність gi|44829554|ref|NC_001145.2][44829554] NC_000913, Escherichia coli K12, повний геном gi|49175990|ref|NC_000913.2|[49175990] NC_003030, Clostridium acetobutylicum ATCC 824, повний геном gi|15893298|ref|NC_003030.1|[15893298] Z99116, Bacillus subtilis повний геном (секція 13 з 21): від 2409151 до 2613687 gi|32468787|emb|Z99116.2|BSUB0013[32468787} М57613, Pseudomonas putida оперон дегідрогенази розгалужених кетокислот (bkdA1, bkdA1 та bkdA2), гени трансацилази Е2 (bkdB), bkdR та ліпоаміддепдрогенази (IpdV), повні cds gi|790512|gb|M57613.1|PSEBKDPPG2[790512] AF157306, Clostridium beijerinckii штам NRRL B593 гіпотетичний білок, гени кофермент А ацилюючої альдегіддегідрогенази (aid), ацетоацетат:бутират/ацетат кофермент А трансферази (ctfA), ацетоацетат:бутират/ацетат кофермент А трансферази (ctfB) і ацетоацетатдекарбоксилази (adc), повні cds gi|47422980|gb|AF157306.2|[47422980] 11 UA 103364 C2 трансаміназа валіндегідрогеназа валіндекарбоксилаза омега-трансаміназа ізобутирил-СоА мутаза 5 10 NC_001988, Clostridium acetobutylicum ATCC 824 плазміда pSOL1, повна послідовність gi|15004705|ref|NC_001988.2|[15004705] U13232, Pseudomonas putida NCIB9816 гени ацетальдегіддегідрогенази (nahO) та 4-гідрокси-2-оксовалератальдолази (nahM), повні cds, та гени 4-оксалокротонатдекарбоксилази (nahK) та 2-оксопент-4еноатгідратази (nahL), часткові cds gi|595671|gb|U13232.1|PPU13232[595671] NC_000913, Escherichia coli K12, повний геном gi|49175990|ref|NC_000913.2|[49175990] NC_006322, Bacillus licheniformis ATCC 14580, повний геном gi|52783855lref|NC_006322.1|[52783855] NC_001142, Saccharomyces cerevisiae хромосома X, повна хромосомна послідовність gi|42742252|ref|NC_001142.5|[42742252] NC_000916, Methanothermobacterthermautotrophicus str. Delta H, повний геном gi|15678031 |ref|NC_000916.11[15678031 ] NC_003888, Streptomyces coelicolorA3(2), повний геном gi|32141095|ref|NC_003888.3|[32141095] Z99116, Bacillus subtilis повний геном (секція 13 з 21): від 2409151 до 2613687 gi|32468787|emb|Z99116.2|BSUB0013[32468787] AY116644, Streptomyces viridifaciens ген амінотрансферази амінокислот, часткові cds; редуктоізомераза гідроксикетокислоти, мала субодиниця ацетолактатсинтетази, велика субодиниця ацетолактатсинтетази, повні cds; генний кластер азоксіантибіотика валаніміцину, повна послідовність; та гени гаданої трансферази і гаданого секретованого білка, повні cds gi|27777548|gb|AY116644.11[27777548] AY330220, Achromobacter denitrificans ген омегаамінокислота:піруваттрансамінази (aptA), повні cds gi|33086797|gb|AY330220.1|[33086797] NC_007347, Ralstonia eutropha JMP134 хромосома 1, повна послідовність gi|73539706|ref|NC_007347.1|[73539706] NC_0004347, Shewanella oneidensis MR-1, повний геном gi|24371600|ref|NC_004347.1|[24371600] NZ_AAAG02000002, Rhodospirillum rubrum RrubO2_2, повна геномна послідовність, визначена методом "пострілу з дробової рушниці" gi|48764549|ref|NZ_AAAG02000002.1|[48764549] АЕ016776, Pseudomonas putida KT2440 секція 3 з 21 повного геному gi|26557019|gb|AE016776.1|[26557019] U67612, Streptomyces cinnamonensis ген кофермент В12-залежної ізобутирил-СоА мутази (іст), повні cds gi|3002491|gb|U67612.1|SCU67612[3002491] AJ246005, Streptomyces cinnamonensis icmB ген ізобутирил-СоА мутази, мала субодиниця gi|6137076|emb|AJ246005.1 |SCI246005[6137076] AL939123, Streptomyces coelicolor A3(2) повний геном; сегмент 20/29 gi|24430032|emb|AL939123.1|SCO939123[24430032] AL9939121, Streptomyces coelicolor A3(2) повний геном; сегмент 18/29 gi|24429533|emb|AL939121.1|SC0939121[24429533] NC_003155, Streptomyces avermitilis MA-4680, повний геном gi|57833846|ref|NC_003155.3|[57833846] Мікробні хазяїни продукування ізобутанолу можуть бути вибрані з бактерій, ціанобактерій, нитчастих грибів та дріжджів. Мікробний хазяїн, використовуваний для продукування ізобутанолу, є краще толерантним до ізобутанолу, так що вихід не обмежений токсичністю бутанолу. Мікроби, метаболічно активні при високих рівнях титру ізобутанолу, не є добре відомими фахівцям. Хоч бутанол-толерантні мутанти були виділені з сольвентогенних Clostridia, мало інформації доступно щодо толерантності до бутанолу інших потенційно корисних бактеріальних штамів. Більшість досліджень з порівняння толерантності до спирту у бактерій дозволяють припустити, що бутанол є більш токсичним, ніж етанол (de Cavalho et al., Microsc. Res. Tech. 64:215-22 (2004) та Kabelitz et al., FEMS Microbiol. Lett. 220:223-227 (2003)). Tomas et 12 UA 103364 C2 5 10 15 20 25 30 35 40 45 50 55 60 al. (J. Bacteriol. 186:2006-2018 (2004)) повідомляють, що вихід 1-бутанолу при ферментації в Clostridium acetobutylicum може бути обмежений токсичністю 1-бутанолу. Первинним ефектом 1бутанолу на Clostridium acetobutylicum є порушення мембранних функцій (Hermann et al., Appl. Environ. Microbiol. 50:1238-1243 (1985)). Мікробні хазяїни, вибрані для продукування ізобутанолу, є краще толерантними до ізобутанолу і здатними перетворювати вуглеводи на ізобутанол. Критерії вибору придатних мікробних хазяїнів включають таке: природжена толерантність до ізобутанолу, високі показники утилізації глюкози, наявність генетичних засобів для генної маніпуляції та здатність генерувати стабільні хромосомальні альтерації. Придатні штами-хазяїни з толерантністю до ізобутанолу можуть бути ідентифіковані шляхом скринінгу на основі природженої толерантності штаму. Природжена толерантність мікробів до ізобутанолу може бути виміряна шляхом визначення концентрації ізобутанолу, яка забезпечує 50 % інгібування швидкості росту (ІС50) при вирощуванні у мінімальному середовищі. Значення ІС50 можуть бути визначені за допомогою способів, відомих фахівцям. Наприклад, мікроби, що представляють інтерес, можуть вирощуватися в присутності різних кількостей ізобутанолу, а швидкість росту контролюють шляхом вимірювання оптичної густини при 600 нанометрах. Час подвоєння може бути обчислений з логарифмічної частини кривої росту і використаний як міра швидкості росту. Концентрація ізобутанолу, яка спричинює 50 % інгібування росту, може бути визначена з графіка залежності процента інгібування росту від концентрацій ізобутанолу. Краще, штам-хазяїн повинен мати ІС50 для ізобутанолу більше приблизно 0,5%. Мікробний хазяїн для продукування ізобутанолу повинен також утилізувати глюкозу з високою швидкістю. Більшість мікробів є здатними утилізувати вуглеводи. Однак, певні мікроби навколишнього середовища не можуть утилізувати вуглеводи з високою ефективністю і тому будуть непридатними хазяїнами. Здатність до генетичної модифікації хазяїна є суттєвою для продукування будь-яких рекомбінантних мікроорганізмів. Метод, використовуваний у технології перенесення гена, може бути трансформацією шляхом електропораціїї, кон'югації, трансдукції або природної трансформації. Широкий спектр хазяїн-кон'югативних плазмід та маркери резистентності до лікарських засобів є доступними. Клонуючі вектори пристосовують до організмів-хазяїнів на основі природи маркерів резистентності до антибіотиків, що можуть функціонувати в цьому хазяїні. Мікробний хазяїн також має бути підданий маніпуляціям з метою інактивації конкурентних метаболічних шляхів руху вуглецю шляхом делеції різних генів. Це вимагає наявності транспозонів для керування інактивацією або векторів хромосомальної інтеграції. Додатково, хазяїн продукування повинен бути сприйнятливим до хімічного мутагенезу таким чином, щоб можна було одержати мутації для поліпшення природженої толерантності до ізобутанолу. На основі описаних вище критеріїв, придатні мікробні хазяїни продукування ізобутанолу включають, без обмеження, членів родів Clostridium, Zymomonas, Escherichia, Salmonella, Rhodococcus, Pseudomonas, Bacillus, Lactobacillus, Enterococcus, Alcaligenes, Klebsiella, Paenibacillus, Arthrobacter, Corynebacterium, Brevibacterium, Pichia, Candida, Hansenula та Saccharomyces. Кращі хазяїни включають: Escherichia coli, Alcaligenes eutrophus, Bacillus licheniformis, Paenibacillus macerans, Rhodococcus erythropolis, Pseudomonas putida, Lactobacillus plantarum, Enterococcus faecium, Enterococcus gallinarium, Enterococcus faecalis, Bacillus subtilis та Saccharomyces cerevisiae. Рекомбінантні організми, що містять необхідні гени, які кодують ферментативний метаболічний шлях конверсії ферментованого вуглецевого субстрату на ізобутанол, можуть бути сконструйовані з використанням методик, добре відомих фахівцям. В даному винаході, гени, що кодують ферменти одного з метаболічних шляхів біосинтезу ізобутанолу за винаходом, наприклад, ацетолактатсинтазу, ізомероредуктазу ацетогідроксикислот, дегідратазу ацетогідроксикислот, декарбоксилазу розгалужених α-кетокислот та дегідрогеназу розгалужених спиртів, можуть бути ізольовані з різних джерел, як описано вище. Способи одержання бажаних генів з бактеріального геному є поширеними та добре відомими фахівцям з молекулярної біології. Наприклад, якщо послідовність гена є відомою, придатні геномні бібліотеки можуть бути створені шляхом гідролізу за допомогою ендонуклеази рестрикції і можуть бути піддані скринінгу зондами, комплементарними до бажаної генної послідовності. Після ізоляції послідовності, ДНК може бути ампліфікована за допомогою стандартних способів ампліфікації з використанням праймерів, таких як полімеразна ланцюгова реакція (Mullis, патент США № 4683202) для одержання кількості ДНК, придатної для трансформації з використанням відповідних векторів. Засоби оптимізації кодонів для експресії в гетерологічному хазяїні є легко доступними. Деякі засоби оптимізації кодонів є доступними на 13 UA 103364 C2 основі вмісту GC організму-хазяїна. Вміст GC деяких типових мікробних хазяїнів наведений в Таблиці 3. Таблиця 3 Вміст GC мікробних хазяїнів Штам %GC 46 42 37 50 61 61 51 62 50 50 В. licheniformis В. subtilis С. acetobutylicum Ε. coli P. putida A. eutrophus Paenibacillus macerans Rhodococcus erythropolis Brevibacillus Paenibaoillus polymyxa 5 10 15 20 25 30 35 40 Після того, як застосовні гени метаболічного шляху будуть ідентифіковані та ізольовані, вони можуть бути трансформовані в придатних хазяїнів експресії засобами, добре відомими фахівцям. Вектори або касети, придатні для трансформації різноманітних клітин-хазяїнів є звичайними та комерційно доступними від таких компаній, як EPICENTRE® (Madison, Wl), Invitrogen Corp. (Carlsbad, CA), Stratagene (La Jolla, CA) та New England Biolabs, Inc. (Beverly, MA). Типово, вектор або касета містить послідовності, що керують транскрипцією та трансляцією відповідного гена, селектований маркер та послідовності, що дозволяють автономну реплікацію або хромосомальну інтеграцію. Придатні вектори включають ділянку 5' гена, яка включає контроль ініціації транскрипції, та ділянку 3' фрагмента ДНК, яка контролює термінацію транскрипції. Обидві контрольні ділянки можуть бути похідними з генів, гомологічних для трансформованої клітини-хазяїна, хоча слід розуміти, що такі контрольні ділянки можуть також бути одержані з генів, які не є нативними для конкретних видів, вибраних як хазяїн продукування. Контрольні ділянки ініціації або промотори, які є придатними для керування експресією відповідних кодуючих ділянок метаболічного шляху в бажаній клітині-хазяїні, є численними та відомі фахівцям в цій області техніки. Практично будь-який промотор, здатний керувати цими генетичними елементами, є придатними для даного винаходу, включаючи, без обмеження, CYC1, HIS3, GAL1, GAL10, ADH1, PGK, PHO5, GAPDH, ADC1, TRP1, URA3, LEU2, ENO, ТРІ, CUP1, FBA, GPD, та GPM (придатні для експресії в Saccharomyces); A0X1 (придатні для експресії в Pichia); та Іас, ага, tet, trp, IPL, IPR, T7, tac та trc (придатні для експресії в Escherichia coli, Alcaligenes та Pseudomonas); промотори ату, арг, прг та різні фагові промотори, придатні для експресії в Bacillus subtilis, Bacillus licheniformis та Paenibacillus macerans; nisA (придатний для експресії в грампозитивних бактеріях, Eichenbaum et al. Appl. Environ. Microbiol. 64(8):27632769 (1998)); та синтетичний промотор Р11 (придатний для експресії в Lactobacillus plantarum, Rud et al., Microbiology 152:1011-1019 (2006)). Ділянки контролю термінації можуть також бути одержані з різних генів, нативних для кращих хазяїнів. Необов'язково, сайт термінації може бути непотрібним, однак, він найкраще включений. Певні вектори є здатними реплікуватися у широкому спектрі бактерій-хазяїнів і можуть бути перенесені шляхом кон'югації. Повна та анотована послідовність pRK404 та три споріднені вектори - pRK437, pRK442 та pRK442(H) є доступними. Ці похідні виявилися цінними інструментами генетичних маніпуляцій у грамнегативних бактерій [Scott et al., Plasmid 50(1):7479 (2003)). Кілька плазмід, похідних від Inc P4 плазміди з широким спектром хазяїнів RSF1010, є також доступними, з промоторами, що можуть функціонувати у ряді грамнегативних бактерій. Плазміди pAYC36 та pAYC37 мають активні промотори разом з численними сайтами клонування, що дозволяє експресію гетерологічного гена в грамнегативних бактеріях. Інструменти заміщення хромосомальних генів також є широко доступними, наприклад, термочутливий варіант реплікону з широким спектром хазяїнів pVW101 був модифікований для конструювання плазміди pVE60'02, яка може бути використана для проведення заміщення гена у різних грампозитивних бактерій (Maguin et al., J. Bacteriol. 174(17):5633-5638 (1992)). 14 UA 103364 C2 5 10 15 20 25 30 35 40 45 50 55 Додатково, є доступними in vitro транспозоми для створення випадкових мутацій у різноманітних геномах з комерційних джерел, таких як EPICENTRE®. Експресія метаболічного шляху біосинтезу ізобутанолу в різних кращих мікробних хазяїнах описана більш детально нижче. Експресія метаболічного шляху біосинтезу ізобутанолу в Е. соlі. Вектори або касети, придатні для трансформації Е. соїі, є поширеними та комерційно доступними від компаній, зазначених вище. Наприклад, гени метаболічного шляху біосинтезу ізобутанолу можуть бути виділені з різних джерел, клонованв і модифікований вектор pUC19 та трансформовані в Е. соїі NM522, як описано в Прикладах 6 та 7. Експресія метаболічного шляху біосинтезу ізобутанолу в Rhodococcus ervthropolis Ряд Ε. coli-Rhodococcus шаттл-векторів є доступними для експресії в R. erythropolis, включаючи, без обмеження, pRhBR17 та pDA71 (Kostichka et al., Appl. Microbiol. Biotechnol. 62:61-68(2003)). Додатково, ряд промоторів є доступними для експресії гетерологічних генів в R. erythropolis (див., наприклад, Nakashima et al., Appl. Environ. Microbiol. 70:5557-5568 (2004) і Tao et al., Appl. Microbiol. Biotechnol. 2005, DOI 10.1007/s00253-005-0064). Розрив по цільовому гену у хромосомальних генах R. erythropolis може бути створений за методом, описаним Tao et al., supra та Brans et al. (Appl. Environ. Microbiol. 66: 2029-2036 (2000)). Гетерологічні гени, потрібні для продукування ізобутанолу, як описано вище, можуть бути клоновані спочатку в pDA71 або pRhBR71 та трансформовані в Е. соїі. Вектори можуть потім бути трансформовані в R. erythropolis шляхом електропорації, як описано Kostichka et al., supra. Рекомбінанти можуть бути вирощені в синтетичному середовищі, що містить глюкозу, а продукування ізобутанолу можна спостерігати з використанням способів, відомих фахівцям. Експресія метаболічного шляху біосинтезу ізобутанолу в В. subtilis Способи генної експресії та створення мутацій у В. subtilis є також добре відомими фахівцям. Наприклад, гени метаболічного шляху біосинтезу ізобутанолу можуть бути виділені з різних джерел, клоновані в модифікований вектор pUC19 та трансформовані в Bacillus subtilis ΒΕ10Ί0, як описано у Прикладі 8. Додатково, п'ять генів метаболічного шляху біосинтезу ізобутанолу можуть бути розділені на два оперони для експресії, як описано у Прикладі 20. Три гени метаболічного шляху (ЬиЬВ, ilvD та kivD) були інтегровані в хромосому Bacillus subtilis BE 1010 (Payne and Jackson, J. Bacteriol. 173:2278-2282 (1991)). Решта два гени (HvC та bdhB) були клоновані в вектор експресії та трансформовані в штам Bacillus, який несе інтегровані гени ізобутанолу. Експресія метаболічного шляху біосинтезу ізобутанолу в В. licheniformis Більшість плазмід та шаттл-векторів, що реплікуються в В. subtilis, можуть бути використані для трансформації В. licheniformis шляхом трансформації протопласта або електропорації. Гени, потрібні для продукування ізобутанолу, можуть бути клоновані в похідні плазмід рВЕ20 або рВЕбО (Nagarajan et al., Gene 114:121-126 (1992)). Способи трансформації В. licheniformis відомі фахівцям (наприклад, див. Fleming et al., Appl. Environ. Microbiol., 61 (11):3775-3780 (1995)). Плазміди, сконструйовані для експресії в В. subtilis, можуть бути трансформовані в В. licheniformis для одержання рекомбінантного мікробного хазяїна, який продукує ізобутанол. Експресія метаболічного шляху біосинтезу ізобутанолу в Paenibacillus macerans Плазміди можуть бути сконструйовані, як описано вище, для експресії в В. subtilis та використані для трансформації Paenibacillus macerans шляхом трансформації протопласта для створення рекомбінантного мікробного хазяїна, який продукує ізобутанол. Експресія метаболічного шляху біосинтезу ізобутанолу в Alcaligenes (Ralstonia) eutrophus Способи генної експресії та створення мутацій в Alcaligenes eutrophus відомі фахівцям (див., наприклад, Taghavi et al., Appl. Environ. Microbiol., 60(10):3585-3591 (1994)). Гени метаболічного шляху біосинтезу ізобутанолу можуть бути клоновані в будь-який з широкого спектра векторівхазяїнів, описаних вище, та електропоровані для створення рекомбінантів, які продукують ізобутанол. Метаболічний шлях полі(гідроксибутирату) в Alcaligenes був детально описаний, разноманітні генетичні методики модифікації геному Alcaligenes eutrophus є відомими, і ці інструменти можуть бути застосовані для створення метаболічного шляху біосинтезу ізобутанолу. Експресія метаболічного шляху біосинтезу ізобутанолу в Pseudomonas putida Способи генної експресії в Pseudomonas putida відомі фахівцям (див., наприклад, BenBassat et al., патент США № 6586229, який включений сюди за посиланням). Гени метаболічного шляху бутанолу можуть бути вставлені в pPCU18 та ця лігована ДНК може бути електропорована в електрокомпетентні клітини Pseudomonas putida DOT-T1 C5aAR1 для створення рекомбінантів, які продукують ізобутанол. Експресія метаболічного шляху біосинтезу ізобутанолу в Saccharomyces cerevisiae 15 UA 103364 C2 5 10 15 20 25 30 35 40 45 50 55 Способи генної експресії в Saccharomyces cerevisiae відомі фахівцям (див., наприклад, Methods in Enzymology, Volume 194, Guide to Yeast Genetics and Molecular and Cell Biology (Part A, 2004, Christine Guthrie and Gerald R. Fink (Eds.), Elsevier Academic Press, San Diego, CA). Експресія генів у дріжджах типово потребує промотора, за яким йде потрібний ген та термінатор транскрипції. Ряд дріжджових промоторів можуть бути використані при конструюванні касет експресії для генів, що кодують метаболічний шлях біосинтезу ізобутанолу, включаючи, без обмеження, конститутивні промотори FBA, GPD, ADH1 та GPM та індуковані промотори GAL1, GAL10 та CUP1. Придатні термінатори транскрипції включають, без обмеження FBAt, GPDt, 3PMt, ERGIOt, GAL1t, CYC1 та ADH1. Наприклад, придатні промотори, термінатори транскрипції та гени метаболічного шляху біосинтезу ізобутанолу можуть бути клоновані в Е. со//-дріжджові шаттл-вектори, як описано у Прикладі 17. Експресія метаболічого шляху біосинтезу ізобутанолу в Lactobacillus plantarum Рід Lactobacillus належить до сімейства Lactobacillales і багато плазмід та векторів, використовуваних для трансформації Bacillus subtilis та Streptococcus, можуть бути використані для Lactobacillus. Необмежувальні приклади придатних векторів включають ρΑΜβΙ та його похідні (Renault et al., Gene 183:175-182 (1996); та O'Sullivan et al., Gene 137:227-231 (1993)); pMBB1 та pHW800, похідне pMBB1 (Wyckoff et al., Appl. Environ. Microbiol. 62:1481-1486 (1996)); pMG1, кон'югативна плазміда (Tanimoto et al., J. Bacteriol. 184:5800-5804 (2002)); pNZ9520 (Kleerebezem et al., Appl. Environ. Microbiol. 63:4581-4584 (1997)); pAM401 (Fujimoto et al., Appl. Environ. Microbiol. 67:1262-1267 (2001)); та pAT392 (Arthur et al., Antimicrob. Agents Chemother. 38:1899-1903 (1994)). Кілька плазмід з Lactobacillus plantarum також були описані (наприклад, van Kranenburg R, Golic Ν, Bongers R, Leer RJ, de Vos WM, Siezen RJ, Kleerebezem M. Appl. Environ. Microbiol. 2005 Mar; 71(3): 1223-1230). Наприклад, експресія метаболічного шляху біосинтезу ізобутанолу в Lactobacillus plantarum описана в Прикладі 21. Експресія метаболічного шляху біосинтезу ізобутанолу в Enterococcus faecium, Enterococcus pallinarium та Enterococcus faecalis Рід Enterococcus належить до сімейство Lactobacillales і багато плазмід та векторів, використовуваних для трансформації Lactobacillus, Bacillus subtilis та Streptococcus, можуть бути використані для Enterococcus. Необмежувальні приклади придатних векторів включають ρΑΜβΙ та його похідні (Renault et al., Gene 183:175-182 (1996); та O'Sullivan et al., Gene 137:227-231 (1993)); pMBB1 та pHW800, похідне рМВВ1 (Wyckoff et al. Appl. Environ. Microbiol. 62:1481-1486 (1996)); pMG1, кон'югативну плазміду (Tanimoto et al., J. Bacteriol. 184:5800-5804 (2002)); pNZ9520 (Kleerebezem et al., Appl. Environ. Microbiol. 63:4581-4584 1997)); pAM401 (Fujimoto et al., Appl. Environ. Microbiol. 67:1262-1267 2001)); та рАТ392 (Arthur et al., Antimicrob. Agents Chemother. 18:1899-1903 (1994)). Вектори експресії для Е. faecalis, що використовують ген nisA з Lactococcus, також можуть бути використані (Eichenbaum et al., Appl. Environ. Microbiol. 64:2763-2769 (1998)). Додатково, можуть бути використані вектори для заміщення гена у хромосомі Е. faecium (Nallaapareddy et al., Appl. Environ. Microbiol. 72:334-345 (2006)). Наприклад, експресія метаболічного шляху біосинтезу ізобутанолу в Enterococcus faecalis описана в Прикладі 22. Ферментаційні середовища в даному винаході повинні містити придатні вуглецеві субстрати. Придатні субстрати можуть включати, без обмеження, моносахариди, такі як глюкоза та фруктоза, олігосахариди, такі як лактоза або сахароза, полісахариди, такі як крохмаль чи целюлоза, або їх суміші та неочищені суміші відновлюваної сировини, такої як пермеат сироварної сироватки, рідкий кукурудзяний екстракт, меляса з цукрового буряка та ячмінний солод. Додатково, вуглецевий субстрат також може бути субстратами з одним атомом вуглецю, такими як діоксид вуглецю або метанол, для яких було продемонстровано метаболічну конверсію на ключові біохімічні проміжні речовини. На додаток до субстратів з одним та двома атомами вуглецю, відомо, що метилотрофні організми також утилізують ряд інших вуглецевмісних сполук, таких як метиламін, глюкозамін та різні амінокислоти для метаболічної активності. Наприклад, відомо, що метилотрофні дріжджі утилізують вуглець з метиламіну для утворення трегалози або гліцерину (Bellion et al., Microb. Growth C1 Compd., [Int. Symp.], 7th (1993), 415-32. Editor(s): Murrell, J. Collin; Kelly, Don P. Publisher: Intercept, Andover, UK). Аналогічно, різні види Candida будуть метаболізувати аланін або олеїнову кислоту (Suiter et al., Arch. Microbiol. 153:485-489 (1990)). Тому передбачається, що джерело вуглецю, утилізованого в даному винаході, може охоплювати різноманітні вуглецевмісні субстрати і буде обмежуватися лише вибором організму. Хоча передбачається, що всі вищезгадані вуглецеві субстрати та їх суміші є придатними в даному винаході, кращими вуглецевими субстратами є глюкоза, фруктоза та сахароза. 16 UA 103364 C2 5 10 15 20 25 30 35 40 45 50 55 На додаток до придатного джерела вуглецю, ферментаційні середовища повинні містити придатні мінеральні речовини, солі, кофактори, буфери та інші компоненти, відомі фахівцям в цій області техніки, придатні для вирощування культур та промотування ферментативного метаболічного шляху, необхідного для продукування ізобутанолу. Типово клітини вирощують при температурі в інтервалі значень від приблизно 25 °С до приблизно 40 °С в придатному середовищі. Придатними середовищами для вирощування в даному винаході є звичайні комерційно виготовлені середовища, такі як середовище ЛуріаБертані (LB), декстрозне середовище Сабуро (SD) або дріжджове середовище (YM). Інші визначені або синтетичні середовища для вирощування також можуть бути використані і придатне середовище для вирощування конкретного мікроорганізму буде відомим фахівцю в області мікробіології або науки про ферментацію. Використання агентів, що модулюють катаболітну репресію прямо чи опосередковано, наприклад, циклічний аденозин-2':3'монофосфат, також може бути включене в ферментаційне середовище. Придатний інтервал значень рН для ферментації становить від рН 5,0 до рН 9,0, причому інтервал від рН 6,0 до рН 8,0 є кращим як початкові умови. Ферментації можуть проводитися в аеробних чи анаеробних умовах, причому анаеробні або мікроаеробні умови є кращими. Кількість ізобутанолу, продукованого в ферментаційному середовищі, може бути визначена з використанням ряду способів, відомих фахівцям, наприклад, високоефективної рідинної хроматографії (ВЕРХ) або газової хроматографії (ГХ). Процес за даним винаходом використовує періодичний спосіб ферментації. Класична періодична ферментація є закритою системою, де склад середовища встановлюється на початку ферментації і не зазнає штучних змін під час ферментації. Таким чином, на початку ферментації середовище інокулюють бажаним організмом або організмами і ферментації дозволяють протікати, нічого не додаючи до системи. Типово, однак, "періодична" ферментація є періодичною відносно до додавання джерела вуглецю і часто робляться спроби контролю таких факторів, як рН та концентрація кисню. В періодичних системах склад метаболітів та біомаси системи постійно змінюються до моменту припинення ферментації. В періодичних культурах клітини проходять від статичної лаг-фази до фази швидкого логарифмічного росту і зрештою до стаціонарної фази, у якій швидкість росту зменшується чи зупиняється. Якщо нічого не робити, клітини у стаціонарній фазі поступово загинуть. Клітини у лог-фазі загалом відповідають за основну частку продукування кінцевого або проміжного продукту. Варіантом стандартно періодичної системи є система з періодичним підживленням. Ферментаційні процеси з періодичним підживленням є також придатними в даному винаході та включають типову періодичну систему за винятком того, що субстрат додають порціями в ході ферментації. Системи з періодичним підживленням є корисними, коли катаболітна репресія є здатною інгібувати метаболізм клітин і де бажано мати обмежену кількість субстрату у середовищі. Вимірювання фактичної концентрації субстрату в системах з періодичним підживленням є складним і тому обчислюється на основі змін вимірних факторів, таких як рН, розчинений кисень та парціальний тиск відхідних газів, таких як СОг. Періодична ферментація та ферментація з періодичним підживленням є звичайними та добре відомі фахівцям, і приклади можна знайти в Thomas D. Brock, Biotechnology: A Textbook of Industrial Microbiology, Second Edition (1989) Sinauer Associates, Inc., Sunderland, MA., або Deshpande, Mukund V., Appl. Biochem. Biotechnol., 36:227, (1992), включені сюди за посиланням. Хоча даний винахід здійснюється в періодичному режимі, передбачається, що спосіб може бути адаптованим до способів безперервної ферментації. Безперервна ферментація є відкритою системою, у якій ферментаційне середовище визначеного складу безперервно додають в біореактор і рівну кількість кондиціонованого середовища одночасно видаляють для переробки. Безперервна ферментація звичайно підтримує культури при постійній високій щільності, де клітини переважно находяться в лог-фазі росту. Безперервна ферментація дозволяє модулювати один фактор або будь-яке число факторів, що впливають на клітинний ріст або концентрацію кінцевого продукту. Наприклад, в одному способі рівень обмежувальної поживної речовини, такої як джерело вуглецю або азоту, буде підтримуватися на постійному значенні, дозволяючи всім іншим параметрам змінюватися. В інших системах ряд факторів, що впливають на ріст, можуть змінюватися безперервно, у той час як концентрація клітин, вимірювана за мутністю середовища, підтримується постійною. Безперервні системи намагаються підтримувати умови усталеного росту і, таким чином, втрата клітин внаслідок відбирання середовища має врівноважуватися швидкістю клітинного росту у ферментації. Способи модуляції поживних речовин та фактори росту процесів безперервної 17 UA 103364 C2 5 10 15 20 25 30 35 40 45 50 55 ферментації, а також методики максимізації швидкості утворення продукту, є добре відомими фахівцям з промислової мікробіології, і разноманітні способи детально описані у Brock, supra. Передбачається, що даний винахід може практикуватися з використанням періодичних, з періодичним підживленням або безперервних процесів, і що будь-який відомий спосіб ферментації буде придатним. Додатково, передбачається, що клітини можуть бути іммобілізовані на субстраті як клітинні каталізатори та піддані умовам ферментація для продукування ізобутанолу. Біопродукований ізобутанол може бути виділений з ферментаційного середовища з використанням способів, відомих фахівцям. Наприклад, тверді речовини можуть бути видалені з ферментаційного середовища шляхом центрифугування, фільтрації, декантації тощо. Потім, ізобутанол може бути виділений з ферментаційного середовища, яке було оброблене з метою видалення твердих речовин, як описано вище, з використанням таких способів, як дистиляція, рідинної екстракції або мембранної сепарації. Оскільки ізобутанол утворює низькокиплячу азеотропну суміш з водою, дистиляція може бути використана лише для розділення суміші до її азеотропного складу. Дистиляція може бути використана в комбінації з іншим способом розділення для забезпечення сепарації азеротропу. Способи, що можуть бути використані в комбінації з дистиляцією для виділення та очищення ізобутанолу включають, без обмеження, декантацію, рідинну екстракцію, адсорбцію та мембранні методики. Додатково, ізобутанол може бути виділений за допомогою азеотропної дистиляції з використанням азеотропоутворювача (див., наприклад, Doherty and Malone, Conceptual Design of Distillation Systems, McGraw Hill, New York, 2001). Суміш ізобутанол-вода утворює гетерогенний азеотроп, так що дистиляція може бути використана в комбінації з декантацією для виділення та очищення ізобутанолу. В цьому способі, ізобутанол, що містить ферментаційне середовище, дистилюють до майже азеотропного складу. Потім, азеотропну суміш конденсують і ізобутанол виділяють з ферментаційного середовища шляхом декантації. Декантована водна фаза може бути повернена в першу дистиляційну колону як зрошення. Декантована органічна фаза з підвищеним вмістом ізобутанолу може бути далі очищена дистиляцією у другій дистиляційній колоні. Ізобутанол також може бути виділений з ферментаційного середовища за допомогою рідинної екстракції в комбінації з дистиляцією. В цьому способі, ізобутанол екстрагують з ферментаційного середовища з використанням рідинної екстракції з придатним розчинником. Органічну фазу, що містить ізобутанол, потім дистилюють для виділення ізобутанолу з розчинника. Дистиляція в комбінації з адсорбцією також може бути використана для виділення ізобутанолу з ферментаційного середовища. В цьому способі, ферментаційне середовище, що містить ізобутанол, дистилюють до майже азеотропного складу і потім решту води видаляють з використанням адсорбента, такого як молекулярні сита (Aden et al. Lignocellulosic Biomass to Ethanol Process Design and Economics Utilizing Co-Current Dilute Acid Prehydrolysis and Enzymatic Hydrolysis for Corn Stover, Report NREUTP-510-32438, National Renewable Energy Laboratory, June 2002). Додатково, дистиляція в комбінації з дифузійним випаровуванням може бути використана для виділення та очищення ізобутанолу з ферментаційного середовища. В цьому способі, ферментаційне середовище, що містить ізобутанол, дистилюють до майже азеотропного складу і потім решту води видаляють дифузійним випаровуванням через гідрофільну мембрану (Guo et al., J. Membr. Sci. 245, 199-210(2004)). Приклади Даний винахід далі описується у наведених Прикладах. Слід розуміти, що ці Приклади, що вказують кращ варіанти втілення винаходу, наведені лише для ілюстрації. На основі наведеного вище обговорення та цих прикладів, фахівець в даній області зможе уяснити суттєві характеристики даного винаходу і, не виходячи за межі його суті та обсягу, зробити різні зміни та модифікації винаходу з метою його адаптації до різних галузей застосування та умов. Стандартні методики рекомбінантних ДНК та молекулярного клонування, використовувані в Прикладах, є добре відомими фахівцям та описані у Sambrook, J., Fritsch, E.F. and Maniatis, Т. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY (1989) (Maniatis) та Т. J. Silhavy, M. L. Bennan and L.W. Enquist, Experiments with Gene Fusions, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, ΝΎ. (1984) та Ausubel, F.M. et al, Current Protocols in Molecular Biology, опубл. Greene Publishing Assoc та Wiley-lnterscience (1987). 18 UA 103364 C2 5 10 Матеріали та способи, придатні для підтримання та росту бактеріальних культур, є добре відомими фахівцям. Методики, придатні для використання в таких Прикладах, можна знайти в Manual of Methods for General Bacteriology (Phillipp Gerhardt, R.G.E. Murray, Ralph N. Costilow, Eugene W. Nester, Willis A. Wood, Noel R. Krieg та G. Briggs Phillips, eds.), American Society for Microbiology, Washington, DC. (1994)) або Thomas D. Brock e Biotechnology: A Textbook of Industrial Microbiology, Second Edition, Sinauer Associates, Inc., Sunderland, MA (1989). Усі реагенти, рестрикційні ферменти та матеріали, використовуван для вирощування та підтримання бактеріальних клітин, були одержані від Aldrich Chemicals (Milwaukee, Wl), BD Diagnostic Systems (Sparks, MD), Life Technologies (Rockville, MD) або Sigma Chemical Company (St. Louis, MO) якщо не зазначено інше. Мікробні штами були одержані від The American Type Culture Collection (ATCC), Manassas, VA, якщо не зазначено інше. Олігонуклеотидні праймери для використання в таких Прикладах наведені у Таблиці 4. Всі олігонуклеотидні праймери синтезовані Sigma-Genosys (Woodlands, TX). 15 Таблиця 4 Праймери для клонування, скринінгу та секвенування олігонуклеотидів Назва N80 N81 N100 N101 N102 N103 N104 N105 N110 N111 N112 N113 N114 N115 N116 N117 N118 N119 BenNF BenASR N110.2 N111.2 N112.2 N113.2 N114.2 Послідовність CACCATGGACAAACAGTATCCGGT ACGCC CGAAGGGCGAIAGCTTTACCAATCC CACCATGGCTAACTACTTCAATACA CTGA CCAGGAGAAGGCCTTGAGTGTTTT CTCC CACCATGCCTAAGTACCGTTCCGC CACCA CGCAGCACTGCTCTTAAATATTCGG С CACCATGAACAACTTTAATCTGCAC ACCC GCTTAGCGGGCGGCTTCGTATATA CGGC GCATGCCTTAAGAAAGGAGGGGGG TCACATGGACAAACAGTATCC ATGCATTTAATTAATTACAGAATCTG ACTCAGATGCAGC GTCGACGCTAGCAAAGGAGGGAAT CACCATGGCTAACTACTTCAA TCTAGATTAACCCGCAACAGCAATA CGTTTC TCTAGAAAAGGAGGAATAAAGTATG CCTAAGTACCGTTC GGATCCTTATTAACCCCCCAGTTTC GATTTA GGATCCAAAGGAGGCTAGACATAT GTATACTGTGGGGGA GAGCTCTTAGCTTTTATTTTGCTCC GCAAAC GAGCTCAAAGGAGGAGCAAGTAAT GAACAACTTTAATCT GAATTCACTAGTCCTAGGTTAGCGG GCGGCTTCGTATATACGG CAACATTAGCGATTTTCITTTCTCT CATGAAGCTTACTAGTGGGCTTAAG TTTTGAAAATAATGAAAACT GAGCTCACTAGTCAATTGTAAGTAA GTAAAAGGAGGTGGGTCACATGGA CAAACAGTATCC GGATCCGATCGACTTAAGCCTCAG CTTACAGAATCTGACTCAGATGCAG С GAGCTCCTTAAGAAGGAGGTAATC ACCATGGCTAACTACTTCAA GGATCCGATCGAGCTAGCGCGGCC GCTTAACCCGCAACAGCAATACGTT TC GAGCTCGCTAGCAAGGAGGTATAA 19 SEQ ID NO: budB прямий 11 budB зворотний 12 ilvC прямий 13 ilvC зворотний 14 ilvD прямий 15 ilvD зворотний 16 yqhD прямий 17 yqhD зворотний 18 Опис budB прямий 19 budB зворотний 20 ilvC прямий 21 ilvC зворотний 22 ilvD прямий 23 ilvD зворотний 24 kivD прямий 25 kivD зворотний 26 yqhD прямий 27 yqhD зворотний 28 Npr прямий 29 Npr зворотний 30 bud В прямий 31 budB зворотний 32 ilvC прямий 33 NvC зворотний 34 ilvD прямий 35 UA 103364 C2 N115.2 N116.2 N117.2 AGTATGCCTAAGTACCGTTC GGATCCGATCGATTAATTAACCTAA GGTTATTAACCCCCCAGTTTCGATT ТА GAGCTCTTAATTAAAAGGAGGTTAG ACATATGTATACTGTGGGGGA GGATCCAGATCTCCTAGGACATGTT TAGCTTTTATTTTGCTCCGCAAAC N130SeqF1 TGTTCCAACCTGATCACCG N130SeqF2 GGAAAACAGCAAGGCGCT N130SeqF3 CAGCTGAACCAGTTTGCC N130SeqF4 AAAATACCAGCGCCTGTCC N130SeqR1 TGAATGGCCACCATGTTG N130SeqR2 GAGGATCTCCGCCGCCTG N130SeqR3 AGGCCGAGCAGGAAGATC N130SeqR4 TGATCAGGTTGGAACAGCC N131SeqF1 AAGAACTGATCCCACAGGC N131SeqF2 ATCCTGTGCGGTATGTTGC N131SeqF3 ATTGCGATGGTGAAAGCG N131SeqR1 ATGGTGTTGGCAATCAGCG N131SeqR2 GIGCTTCGGIGAIGGTTT N131SeqR3 TTGAAACCGTGCGAGTAGC N132SeqF1 TATTCACTGCCATCTCGCG N132SeqF2 CCGTAAGCAGCTGTTCCT N132SeqF3 GCTGGAACAATACGACGTTA N132SeqF4 TGCTCTACCCAACCAGCTTC N132SeqR1 ATGGAAAGACCAGAGGTGCC N132SeqR2 TGCCTGTGTGGTACGAAT N132SeqR3 TATTACGCGGCAGTGCACT N132SeqR4 GGTGATTTTGTCGCAGTTAGAG N133SeqF1 TCGAAATTGTTGGGTCGC N133SeqF2 GGTCACGCAGTTCATTTCTAAG N133SeqF3 TGTGGCAAGCCGTAGAAA N133SeqF4 AGGATCGCGTGGTGAGTAA 20 ilvD зворотний 36 kivD прямий 37 kivD зворотний 38 праймер секвенування праймер секвенування праймер секвенування праймер секвенування праймер секвенування праймер секвенування праймер секвенування праймер секвенування праймер секвенування праймер секвенування праймер секвенування праймер секвенування праймер секвенування праймер секвенування праймер секвенування праймер секвенування праймер секвенування праймер секвенування праймер секвенування праймер секвенування праймер секвенування праймер секвенування праймер секвенування праймер секвенування праймер секвенування праймер секвенування 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 .58 59 60 61 62 63 64 65 UA 103364 C2 праймер секвенування праймер секвенування праймер секвенування праймер секвенування праймер секвенування праймер секвенування праймер секвенування праймер секвенування N133SeqR1 GTAGCCGTCGTTATTGATGA N133SeqR2 GCAGCGAACTAATCAGAGATTC N133SeqR3 TGGTCCGATGTATTGGAGG N133SeqR4 TCTGCCATATAGCTCGCGT Scr1 CCTTTCTTTGIGAAICGG Scr2 AGAAACAGGGTGTGATCC Scr3 AGTGATCATCACCTGTTGCC Scr4 AGCACGGCGAGAGTCGACGG T-budB (BamHI) B-kivD (BamHI) AGATAGATGGATCCGGAGGTGGGT CACATGGACAAACAGT budB прямий 144 CTCTAGAGGATCCAGACTCCTAGG ACATG kivD зворотний 145 AGATAGATCTCGAGAGCTATTGTAA CATAATCGGTACGGGGGTG ATTATGTCAGGATCCACTAGTTTCC TCCTTTAATTGGGAATTGTTATCCG С AGCTATTGTAACATAATCGGTACGG GGGTG ACATTGATGGATCCCATAACAAGGG AGAGATTGAAATGGTAAAAG TAGACAACGGATCCACTAGTTTAAT TTTGCGCAACGGAGACCACCGC TTACCGTGGACTCACCGAGTGGGT AACTAGCCTCGCCGGAAAGAGCG TCACAGTTAAGACACCTGGTGCCG TTAATGCGCCATGACAGCCATGAT ACAGATAGATCACCAGGTGCAAGC TAATTCCGGTGGAAACGAGGTCAT С ACAGTACGATACACGGGGTGTCAC TGCCCGCTTTCCAGTCGGGAAACC TCGGATTACGCACCCCGTGAGCTA TTGTAACATAATCGGTACGGGGGT G CTGCTGATCTCACACCGTGTGTTAA TTTTGCGCAACGGAGACCACCGC TCGATAGCATACACACGGTGGTTAA CAAAGGAGGGGTTAAAATGGTTGA TTTCG ATCTACGCACTCGGTGATAAAACGA AAGGCCCAGTCTTTCGACTGAGCC TTTCGTTTTATCTTACACAGATTTTT TGAATATTTGTAGGAC GACGTCATGACCACCCGCCGATCC CTTTI GATATCCAACACCAGCGACCGACG TATTAC ATTTAAATCTCGAGTAGAGGATCCC AACAAACGAAAATTGGATAAAG ACGCGTTATTATAAAAGCCAGTCAT TAGG CCTAGCGCTATAGTTGTTGACAGAA TGGACATACTATGATATATTGTTGC TATAGCGA CTAGTCGCTATAGCAACAATATATC ATAGTATGTCCATTCTGTCAACAAC TATAGCGCTAGG AAGCTTGTCGACAAACCAACATTAT GACGTGTCTGGGC GGATCCTCATCCTCTCGTAGTGAAA ATT PgroE прямий 147 T-groE (Xhol) B-groEL (Spel, BamHI) T-groEL T-ilvCB.s. (BamHI) B-ilvCB.s. (SpelBamHI) T-BD64 (DralH) B-BD64 (Dralll) T-laclq (Dralll) B-laclq (Dralll) T-groE (Dralll) B-B.s.ilvC (Dralll) T-bdhB (Dralll) B-bdhB (ГГПВТ1 Dralll) LDH EcoRV F LDH AatllR CmF Cm R P11 F-Stul P11 R-Spel PldhL F-Hindlll PldhL R 21 PgroE зворотний PgroE прямий 66 67 68 69 72 73 74 75 148 149 ilvC прямий 150 ilvC зворотний 151 pBD64 прямий 152 pBD64 зворотний 153 laclq прямий 154 laclq зворотний 155 PgroE прямий 156 ilvC зворотний 157 bdhB прямий 159 bdhB зворотний 160 IdhL прямий IdhL зворотний 161 162 Cm прямий 163 Cm зворотний 164 P11 промотор 165 прямий P11 промотор 166 зворотний IdhL прямий 167 IdhL зворотний 168 UA 103364 C2 BamHI F-bdhB-Avrll R-bdhBBamHI F-ilvC (B.s-)Aflll R-ilvC (B.s.)Notl F-PnisA (Hindlll) R-PnisA (Spel BamHI) 5 10 15 20 25 30 35 40 45 TTCCTAGGAAGGAGGTGGTTAAAAT GGTTGATTTCG bdhB прямий TTGGAICCTTACACAGATTTTTTGAA TAT bdhB зворотний 170 AACTTAAGAAGGAGGTGATTGAAAT GGTAAAAGTATATT ilvC прямий 171 AAGCGGCCGCTTAATTTTGCGCAA CGGAGACC ivIC зворотний 172 169 nisA промотор 173 прямий nisA промотор TTGGATCCAAACTAGTATAATTTATT TTGTAGTTCCTTC 174 зворотний TTAAGCTTGACATACTTGAATGACC TAGTC Способи визначення концентрації ізобутанолу в культуральному середовищі Концентрація ізобутанолу в культуральному середовищі може бути визначена рядом способів, відомих фахівцям. Наприклад, спосіб специфічної високоефективної рідинної хроматографії (ВЕРХ) з використанням колонки Shodex SH-1011 із Shodex SH-G захисною колонкою, обидві придбані у Waters Corporation (Milford, MA), з детектуванням за показником заломлення (RI). Хроматографічне розділення досягалося з використанням 0,01 Μ H2SO4 як рухомої фази при об'ємній витраті 0,5 мл/хв та температурі колонки 50 °С. Ізобутанол мав час утримання 46,6 хв. в умовах, що використовувалися. Альтернативно, є доступними способи газової хроматографії (ГХ). Наприклад, конкретний ГХ спосіб використовує HP-INNOWax колонку (30 м χ 0,53 мм внутрішній діаметр (id), товщина плівки 1 мкм, Agilent Technologies, Wilmington, DE), з полум'яноіонізаційним детектором (FID). Газом-носієм був гелій з об'ємною витратою 4,5 мл/хв, вимірюваною при 150 °С при постійному напорному тиску; розділення на інжекторі становило 1:25 при 200 °С; температура печі становила 45 °С протягом 1 хв., від 45 до 220 °С при 10 °С/хв. та 220 °С протягом 5 min; і детектування FID використовувалося при 240 °С при 26 мл/хв гелію як газу підживлення. Час утримання ізобутанолу становив 4,5 хв. Абревіатури мають такі значення: "с" позначає секунди, "хв." позначає хвилини, "год." позначає години, "psi" позначає фунти на квадратний дюйм (1 psi = 6,895 кПа), "ни" позначає нанометри, "д" позначає дні, "мкл" позначає мікролітри, "мл" позначає мілілітри, "л" позначає літри, "мм" позначає міліметри, "нм" позначає нанометри, "мМ" позначає мілімолі, "мкМ" позначає мікромолі, "М" позначає молі, "ммоль" позначає мілімолі, "мкмоль" позначає мікромолі", "г" позначає грами, "мкг" позначає мікрограми і "нг" позначає нанограми, "ПЛР" позначає полімеразну ланцюгову реакцію, "OD" позначає оптичну густину, "OD600" позначає оптичну густину, вимірювану на довжині хвилі 600 нм, "кДа" позначає кілодальтони, "g" позначає гравітаційну сталу, "п.о." позначає пари основ, "т.п.о." позначає тисячі пар основ, "% мас/об." позначає масові/об'ємні проценти, "% об./об." позначає об'ємні/об'ємні проценти, "IPTG" позначає ізопропіл- -D-тіогалактопіранозид, "RBS" позначає рибосомальний сайт зв'язування, "ВЕРХ" позначає високоефективну рідинну хроматографію і "ГХ" позначає газову хроматографію. Термін "молярна селективність" позначає число молей продукту, вироблених на моль спожитого цукрового субстрату, і виражається в процентах. Приклад 1 Клонування та експресія ацетолактатсинтази Метою цього Прикладу було клонування гена budB з Klebsiella pneumoniae та його експресія в Е. coli BL21-AI. Ген budB був ампліфікований з геномної ДНК Klebsiella pneumoniae, штам АТСС 25955, з використанням ПЛР, з одержанням 1,8 т.п.о. продукту. Геномну ДНК готують, використовуючи набір Gentra Puregene (Gentra Systems, Inc., Minneapolis, MN; номер за каталогом D-5000A). Ген budB ампліфікують з геномної ДНК Klebsiella pneumoniae методом ПЛР з використанням праймерів N80 та N81 (див. Таблицю 2), наведених як SEQ ID NOs:11 та 12, відповідно. Інші реагенти для ПЛР ампліфікації постачалися в наборах від виробника, наприклад, Finnzymes Phusion™ High-Fidelity PCR Master Mix (New England Biolabs Inc., Beverly, MA; номер за каталогом F-531) та використовувалися згідно з протоколом виробника. Ампліфікацію проводили в DNA Thermocycler GeneAmp 9700 (ΡΕ Applied Biosystems, Foster City, CA). Для досліджень експресії використовували технологію клонування Gateway (Invitrogen Corp., Carlsbad, CA). Вхідний (entry) вектор pENTRSDD-TOPO дозволяв спрямоване клонування та забезпечував Shine-Dalgarno послідовність для потрібного гена. Вектор призначення pDEST14 використовув Т7 промотор для експресії гена без мітки. Прямий праймер включає чотири основи (САСС), безпосередньо прилеглі до старт-кодону транскрипції для забезпечення 22 UA 103364 C2 5 10 15 20 25 30 35 40 45 50 55 60 спрямованого клонування в pENTRSDD-ТОРО (Invitrogen) з утворенням плазміди pENTRSDDTOPObudB. Конструкт pENTR трансформували в Е. coli ТорЮ (Invitrogen) клітини та висіювали на чашки згідно з рекомендаціями виробника. Трансформанти вирощували протягом ночі та плазмідну ДНК готують, використовуючи QIAprep Spin Miniprep kit (Qiagen, Valencia, CA; номер за каталогом 27106) згідно з рекомендаціями виробника. Клони секвенували для підтвердження того, що гени вставлені з правильною орієнтацією, та для підтвердження послідовності. Нуклеотидна послідовність відкритої рамки зчитування (ORF) для цього гена та завбачена амінокислотна послідовність ферменту наведені як SEQ ID N0:1 та SEQ ID N0:2, відповідно. Для створення експресуючого клону, ген budB переносили до вектора pDEST 14 шляхом рекомбінації для створення pDEST14budB. Вектор pDEST14budB трансформували в клітини Е. coli BL21-A1 (Invitrogen). Трансформанти інокулювали в середовище Луріа-Бертані (LB) з добавкою 50 мкг/мл ампіциліну та вирощували протягом ночі. Аліквоту нічної культури використовували для інокуляції 50 мл LB з добавкою 50 мкг/мл ампіциліну. Культуру інкубують при 37 °С зі струшуванням, поки OD60o не досягне 0,6-0,8. Культуру розділяють на дві 25-мл культури та арабінозу додають в одну з колб до кінцевої концентрації 0,2% мас/об. Колбу негативного контролю не індукують арабінозою. Колби інкубують протягом 4 год. при 37 °С зі струшуванням. Клітини збирають центрифугуванням і клітинний осад ресуспендують в 50 мМ MOPS, буфер рН 7,0. Клітини руйнують озвучуванням або пропусканням через French Pressure Cell. Цільний клітинний лізат центрифугують, одержуючи супернатант або безклітинний екстракт та осад або нерозчинну фракцію. Аліквоту кожної фракції (цільний клітинний лізат, безклітинний екстракт та нерозчинна фракція) ресуспендують в SDS (MES) завантажувальному буфері (Invitrogen), нагрівають до 85 °С протягом 10 хв. та піддають аналізу методом SDS-PAGE (NuPAGE 4-12% Bis-Tris Gel, номер за каталогом NP0322Box, Invitrogen). Білок з очікуваною молекулярною вагою від приблизно 60 кДа, обчисленою за послідовністю нуклеїнової кислоти, був присутнім в індукованій культурі, але не в неіндукованому контролі. Активність ацетолактатсинтази в безклітинних екстрактах вимірюють у спосіб, описаний Bauerle et al. [Biochim. Biophys. Acta 92(1): 142-149(1964)). Приклад 2 (Передбачувальний) Клонування та експресія редуктоізомерази ацетогідроксикислот Задачею цього Прикладу є опис того, як клонувати ген HvC з Е. coli K12 та експресувати його в Е. coli BL21-AI. Ген HvC ампліфікують з геномної ДНК Е. coli за допомогою ΠЛР. Ген ilvC клонують та експресують у такий саме спосіб, як ген budB, описаний в Прикладі 1. Геномну ДНК з Ε. coli готують з використанням набору Gentra Puregene (Gentra Systems, Inc., Minneapolis, MN; номер за каталогом D-5000A). Ген ilvC ампліфікують методом ПЛР з використанням праймерів N100 та N101 (див. Таблицю 2), наведених як SEQ ID NOs:13 та 14, відповідно, одержуючи 1,5 т.п.о. продукт. Прямий праймер включає чотири основи (ССАС), безпосередньо прилеглі до старт-кодону трансляції, для забезпечення спрямованого клонування в pENTR/SD/D-TOPO (Invitrogen) для одержання плазміди pENTRSDD-TOPOilvC. Клони секвенують для підтвердження того, що гени вставлені в правильній орієнтації та для підтвердження послідовності. Нуклеотидна послідовність відкритої рамки зчитування (ORF) для цього гена та завбачена амінокислотна послідовність ферменту наведені як SEQ ID N0:3 та SEQ ID N0:4, відповідно. Для створення експресуючого клона, ген ilvC переносять в вектор pDEST 14 (Invitrogen) шляхом рекомбінації для створення pDEST14ilvC. Вектор pDEST14ilvC трансформують в клітини Е. coli BL21-AI та експресію з промотора Т7 індукують шляхом додавання арабінози. Білок з очікуваною молекулярною вагою від приблизно 54 кДа, обчисленою за послідовністю нуклеїнової кислоти, є присутнім в індукованій культурі, але не в неіндукованому контролі. Активність редуктоізомерази ацетогідроксикислоти в безклітинних екстрактах вимірюють у спосіб, описаний Arfin and Umbarger (J. Biol. Chem. 244(5): 1118-1127 (1969)). Приклад 3 (Передбачувальний) Клонування та експресія дегідратази ацетогідроксикислот Задачею цього передбачувального Прикладу є опис того, як клонувати ген ilvD з Е. coli K12 та експресувати його в Е. coli BL21-AI. Ген ilvD ампліфікують з геномної ДНК Е. coli за допомогою ПЛР. Ген ilvD клонують та експресують у такий саме спосіб, як ген budB, описаний в Прикладі 1. Геномну ДНК з Е. coli готують, використовуючи набір Gentra Puregene (Gentra Systems, Inc., Minneapolis, MN; номер за каталогом D-5000A). Ген ilvD ампліфікують методом ПЛР з використанням праймерів N102 та N103 (див. Таблицю 2), наведених як SEQ ID NOs:15 та 16, відповідно, одержуючи 1,9 т.п.о. продукт. Прямий праймер включає чотири основи (ССАС), 23 UA 103364 C2 5 10 15 20 25 30 35 40 45 50 55 60 безпосередньо прилеглі до старт-кодону трансляції, для забезпечення спрямованого клонування в pENTR/SD/D-TOPO (Invitrogen) для одержання плазміди pENTRSDD-TOPOilvD. Клони піддають секвенуванню для підтвердження того, що гени вставлені в правильній орієнтації та для підтвердження послідовності. Нуклеотидна послідовність відкритої рамки зчитування (ORF) для цього гена та завбачувана амінокислотна послідовність ферменту наведені як SEQ ID N0:5 та SEQ ID N0:6, відповідно. Для створення експресуючого клону, ген ilvD переносять в вектор pDEST 14 (Invitrogen) шляхом рекомбінації для створення pDEST14ilvD. Вектор pDEST14ilvD трансформують в клітини Е. coll BL21-AI та експресію з промотора Т7 індукують шляхом додавання арабінози. Білок з очікуваною молекулярною вагою від приблизно 66 кДа, обчисленою за послідовністю нуклеїнової кислоти, є присутнім в індукованій культурі, але не в неіндукованому контролі. Активність дегідратази ацетогідроксикислот в безклітинних екстрактах вимірюють у спосіб, описаний Flint et al. (J. Biol. Chem. 268(20):14732-14742(1993)). Приклад 4 (Передбачувальний) Клонування та експресія декарбоксилази розгалужених кетокислот Задачею цього передбачувального Прикладу є опис того, як клонувати ген kivD з Lacfococci/s lactis та експресувати його в Е. coli BL21-AI. ДНК послідовність, що кодує декарбоксилазу розгалужених кетокислот (kivD) з L lactis, одержують від GenScript (Piscataway, NJ). Одержана послідовність є кодон-оптимізованою для експресії як в Е. coli, так і в В. subtilis та клонується в pUC57, для утворення pUC57-kivD. Кодоноптимізована нуклеотидна послідовність відкритої рамки зчитування (ORF) для цього гена та завбачена амінокислотна послідовність ферменту наведені як SEQ ID N0:7 та SEQ ID N0:8, відповідно. Для створення експресуючого клону сайти рестрикції Ndel та ВатНІ використовують для клонування 1,7 т.п.о. kivD фрагмента з pUC57-kivD в вектор рЕТ-За (Novagen, Madison, WI). Це дає експресуючий клон рЕТ-За-kivD. Вектор pET-3a-kivD трансформують в клітини Ε coli BL21-AI та експресію з промотора Т7 індукують шляхом додавання арабінози. білок з очікуваною молекулярною вагою від приблизно 61 кДа, обчисленою за послідовністю нуклеїнової кислоти, є присутнім в індукованій культурі, але не в неіндукованому контролі. Активність декарбоксилази розгалужених кетокислот в безклітинних екстрактах вимірюють у спосіб, описаний Smit et al. (Appl. Microbiol. Biotechnol. 64:396-402 (2003)). Приклад 5 (Передбачувальний) Клонування та експресія дегідрогенази розгалужених спиртів Задачею цього передбачувального Прикладу є опис того, як клонувати ген yqhD з Е. coli K12 та експресувати його в Е. coli BL21-AL, Gene yqhD ампліфікують з геномної ДНК Ε. coli за допомогою ПЛР. Ген yqhD клонують та експресують у такий саме спосіб, як ген budB, описаний в Прикладі 1. Геномну ДНК з Е. coli готують, використовуючи набір Gentra Puregene (Gentra Systems, Inc., Minneapolis, MN; номер за каталогом D-5000A). Ген yqhD ампліфікують методом ПЛР з використанням праймерів N104 та N105 (див. Таблицю 2), наведених як SEQ ID NOs: 17 та 18, відповідно, одержуючи 1,2 т.п.о. продукт. Прямий праймер включає чотири основи (ССАС), безпосередньо прилеглі до старт-кодону транскрипції, для забезпечення спрямованого клонування в pENTR/SD/D-TOPO (Invitrogen) для одержання плазміди pENTRSDD-TOPOyqhD. Клони піддають секвенуванню для підтвердження того, що гени вставлені в правильній орієнтації та для підтвердження послідовності. Нуклеотидна послідовність відкритої рамки зчитування (ORF) для цього гена та завбачена амінокислотна послідовність ферменту наведені як SEQ ID NO 9 та SEQ ID N0:10, відповідно. Для створення експресуючого клону, ген yqhD переносять в вектор pDEST 14 (Invitrogen) шляхом рекомбінації для створення pDEST14yqhD. Вектор pDEST14ilvD трансформують в клітини Е. coli BL21-AI та експресію з промотора Т7 індукують шляхом додавання арабінози. Білок з очікуваною молекулярною вагою від приблизно 42 кДа, обчисленою за послідовністю нуклеїнової кислоти, є присутнім в індукованій культурі, але не в неіндукованому контролі. Активність дегідрогенази розгалужених спиртів в безклітинних екстрактах вимірюють у спосіб, описаний Sulzenbacher et al. (J. Мої. ВЫ., 342(2):489-502 (2004)). Приклад 6 (Передбачувальний) Конструювання вектора трансформації для генів метаболічного шляху біосинтезу ізобутанолу Задачею цього передбачувального Прикладу є опис того, як сконструювати вектор трансформації, що включає гени, які кодують п'ять стадій метаболічний шлях біосинтезу 24 UA 103364 C2 5 10 15 20 25 30 35 40 45 50 55 60 ізобутанолу. Всі гени поміщають в єдиний оперон під контролем єдиного промотора. Індивідуальні гени ампліфікують методом ПЛР з праймерами, що включають сайти рестрикції для подальшого клонування, а прямі праймери містять оптимізован Е. coli рибосомальний сайт зв'язування (AAAGGAGG). Продукти ПЛР ТОРО-клонують в вектор pCR 4Blunt-TOPO та трансформують в клітини Е. coli ТорЮ (Invitrogen). Плазмідну ДНК готують з ТОРО-клонів та послідовність генів верифікують. Рестрикційні ферменти та Т4 ДНК лігазу (New England Biolabs, Beverly, MA) використовують згідно з рекомендаціями виробника. Для експериментів з клонування, рестрикційні фрагменти очищають на гелі, використовуючи набір QIAquick Gel Extraction (Qiagen). Після підтвердження послідовності, гени субклонують в модифікований вектор pUC19 як платформу клонування. Вектор pUC19 модифікують шляхом гідролізу за допомогою Hindlll/Sapl, одержуючи pUC19dHS. Гідроліз видаляє Іас-промотор, прилеглий до MCS (сайт множинного клонування), запобігаючи транскрипції оперонів в вектор. Ген budB ампліфікують з геномної ДНК К. pneumoniae ATCC 25955 методом ПЛР, використовуючи пару праймерів N110 та N111 (див. Таблицю 2), наведених як SEQ ID NOs:19 та 20, відповідно, одержуючи 1,8 т.п.о. продукт. Прямий праймер включає сайти рестрикції Sphl та Aflll та рибосомальний сайт зв'язування (RBS). Зворотний праймер включає сайти рестрикції Pad та Nsil. Продукт ПЛР клонують в pCR4 Blunt-TOPO, одержуючи pCR4 Blunt-TOPO-budB. Плазмідну ДНК готують з ТОРО-клонів та послідовність гена верифікують. Ген ilvC ампліфікують з геномної ДНК Ε. coli K12 методом ПЛР, використовуючи пару праймерів N112 та N113 (див. Таблицю 2), наведених як SEQ ID N0s:21 та 22, відповідно, одержуючи 1,5 т.п.о. продукт. Прямий праймер включає сайти рестрикції Sail та Nhel та RBS. Зворотний праймер включає сайт рестрикції Xbal. ПЛР продукт клонують в pCR4 3lunt-TOPO, одержуючи pCR4 Blunt-TOPO-ilvC. Плазмідну ДНК готують з ТОРО-клонів та послідовність гена верифікують. Ген HvD ампліфікують з геномної ДНК Ε. coli K12 методом ПЛР, використовуючи пару праймерів N114 та N115 (див. Таблицю 2), наведених як SEQ ID NOs:23 та 24, відповідно, одержуючи 1,9 т.п.о. продукт. Прямий праймер включає сайт рестрикції Xbal та RBS. Зворотний праймер включає сайт рестрикції ВатНІ. ПЛР продукт клонують в pCR4 Blunt-TOPO, одержуючи pCR4 Blunt-TOPO-ilvD. Плазмідну ДНК готують з ТОРО-клонів та послідовність гена верифікують. Ген kivD ампліфікують з pUC57-kivD (описан у Прикладі 4) методом ПЛР, використовуючи пару праймерів N116 та N117 (див. Таблицю 2), наведених як SEQ ID NOs:25 та 26, відповідно, одержуючи 1,7 т.п.о. продукт. Прямий праймер включає сайт рестрикції ВатНІ та RBS. Зворотний праймер включає сайт рестрикції Sacl. ПЛР продукт клонують в pCR4 Blunt-TOPO, одержуючи pCR4 Blunt-TOPO-kivD. Плазмідну ДНК готують з ТОРО-клонів та послідовність гена верифікують. Ген yqhD ампліфікують з геномної ДНК Ε. coli K12 методом ПЛР, використовуючи пару праймерів N118 та N119 (див. Таблицю 2), наведених як SEQ ID NOs:27 та 28, відповідно, одержуючи 1,2 т.п.о. продукт. Прямий праймер включає сайт рестрикції Sacl. Зворотний праймер включає сайти рестрикції Spel та EcoRI. ПЛР продукт клонують в pCR4 Blunt-TOPO, одержуючи pCR4 Blunt-TOPO-yqhD. Плазмідну ДНК готують з ТОРО-клонів та послідовність гена верифікують. Для того, щоб сконструювати оперон метаболічного шляху ізобутанолу, ген yqhD вирізають з pCR4 Blunt-TOPO-yghD за допомогою Sad та EcoRI, одержуючи 1,2 т.п.о. фрагмент. Його лігують з pUC19dHS, що був попередньо гідролізований за допомогою Sad та EcoRI. Одержаний клон, pUC19dHS-yqhD, підтверджують шляхом рестрикційного аналізу. Далі, ген UvC вирізають з pCR4 Blunt-TOPO-ilvC за допомогою Sail та Xbal, одержуючи 1,5 т.п.о. фрагмент. Його лігують з pUC19dHS-yqhD, що був попередньо гідролізований за допомогою Sail та Xbal. Одержаний клон, pUC19dHS-ilvC-yqhD, підтверджують шляхом рестрикційного аналізу. Ген budB потім вирізають з pCR4 Blunt-TOPO-budB за допомогою Sphl та Nsil, одержуючи 1,8 т.п.о. фрагмент. pUC19dHS-HvC-yqhD гідролізують за допомогою Sphl та Pstl та лігують з фрагментом Sphl/Nsil budB (Nsil та Pstl утворюють сумісні кінці), одержуючи pUC19dHS-budBilvC-yqhD. Фрагмент 1,9 т.п.о., що містить ген ilvD, вирізають з pCR4 Blunt-TOPO-ilvD за допомогою Xbal та ВатНІ і лігують з pUC19dHS-budB-ilvC-yqhD, яки гідролізують цими самими ферментами, одержуючи pUC19dHS-budB-ilvC-ilvD-yqhD. Зрештою, kivD вирізають з pCR4 BluntTOPO-kivD за допомогою ВатНІ та Sad, одержуючи 1,7 т.п.о. фрагмент. Цей фрагмент лігують з pUC19dHS-budB-ilvC-ilvD-yqhD, що був попередньо гідролізований за допомогою ВатНІ та Sad, одержуючи pUC19dHS-budB-ilvC-ilvD-kivD-yqhD. Вектор pUC19dHS-budB-ilvC-ilvD-kivD-yqhD гідролізують за допомогою Aflll та Spel, вивільняючи 8,2 т.п.о. фрагмент оперону, який клонують в pBenAS, шаттл-вектор E.coli-B. 25 UA 103364 C2 5 10 15 20 25 30 35 40 45 50 55 60 subtilis. Плазміду pBenAS створюють шляхом модифікації вектора рВЕ93, який описаний Nagarajan (WO 93/24631, Приклад 4). Для того, щоб зробити pBenAS промотор нейтральної протеази (NPR) Bacillus amyloliquefaciens, сигнальну послідовність та ген phoA видаляють шляхом гідролізу рВЕ93 за допомогою Ncol/Hindlll. Промотор NPR ампліфікують методом ПЛР з рВЕ93 праймерами BenNF та BenASR, наведеними як SEQ ID NOS:29 та 30, відповідно. Праймер BenASR включає сайти Aflll, Spel та Hindi 11 справа від промотора. ПЛР продукт гідролізують за допомогою Ncol та Hindlll і фрагмент клонують у відповідні сайти вектора, одержуючи pBenAS. Фрагмент оперона субклонують в сайти Aflll та Spel pBenAS, одержуючи pBen-budB-ilvC-ilvD-kivD-yqhD. Приклад 7 (Передбачувальний) Експресія метаболічного шляху біосинтезу ізобутанолу в Е. соlі Задачею цього передбачувального Прикладу є опис того, як експресувати метаболічний шлях біосинтезу ізобутанолу в Е. соїі. Плазміду pBen-budB-ilvC-ilvD-kivD-yqhD, сконструйовану, як описано у Прикладі 6, трансформують в Е. соїі NM522 (АТСС No. 47000), одержуючи Е. соїі штам NM522/pBen-budBilvC-ilvD-kivD-yqhD, та експресію генів в опероні контролюють методами аналізу SDS-PAGE, ферментного аналізу та вестерн-блот аналізу. Для вестерн-блот аналізів, антитіла до синтетичних пептидів вирощує Sigma-Genosys (The Woodlands, TX). Ε. coli штам NM522/pBen-budB-ilvC-ilvD-kivD-yqhD інокулюють в струшувану колбу на 250 мл, що містить 50 мл середовища, та струшують при 250 об/хв. та 35 °С. Середовище складається з: глюкози (5 г/л), MOPS (0,05 Μ), сульфату амонію (0,01 М), фосфату калію, однозаміщеного (0,005 М), суміші металів S10 (1% (об./об.)), дріжджового екстракту (0,1% (мас./об.)), казамінових кислот (0,1% (мас./об.)), тіаміну (0,1 мг/л), проліну (0,05 мг/л) і біотину (0,002 мг/л) і титрують до рН 7,0 за допомогою КОН. Суміш металів S10 містить: МдСІ2 (200 мМ), СаСІ2 (70 мМ), МпСІ2 (5 мМ), FeCI3 (0,1 мМ), ZnCI2 (0,1 мМ), тіамін гідрохлорид (0,2 мМ), CuSO4 (172 мкМ), СоСІ2 (253 мкМ) і Na2MoO4 (242 мкМ). Через 18 год., ізобутанол детектують за допомогою ВЕРХ або ГХ аналізу, використовуючи способи, що є добре відомими фахівцям, наприклад, як описано у розділі Загальні способи вище. Приклад 8 (Передбачувальний) Експресія метаболічного шляху біосинтезу ізобутанолу в Bacillus subtilis Задачею цього передбачувального прикладу є опис того, як експресувати метаболічний шлях біосинтезу ізобутанолу в Bacillus subtilis. Використовується такий саме підхід, як описано у Прикладі 7. Використовується плазміда pBen-budB-ilvC-ilvD-kivD-yqhD, сконструйована як описано у Прикладі 6. Цю плазміду трансформують в Bacillus subtilis BE1010 (J. Bacteriol. 173:2278-2282 (1991)), одержуючи В. subtilis штам ВЕ1010/pBen-budB-ilvC-ilvD-kivD-yqhD та експресію генів в в кожному опероні контролюють, як описано у Прикладі 7. В. subtilis штам ВЕЮЮ/pBen-budB-ilvC-ilvD-kivD-yqhD інокулюють в струшувану колбу на 250 мл, що містить 50 мл середовища, та струшують при 250 об/хв. та 35 °С протягом 18 год. Середовище складається з: декстрози (5 г/л), MOPS (0,05 Μ), глутамінової кислоти (0,02 М), сульфату амонію (0,01 М), фосфату калію, однозаміщеного буфера (0,005 М), суміші металів S10 (як описано у Прикладі 11,1 % (об./об.)), дріжджового екстракту (0,1 % (мас/об.)), казамінових кислот (0,1% (мас/об.)), триптофану (50 мг/л), метіоніну (50 мг/л) і лізину (50 мг/л) і титрують до рН 7,0 за допомогою КОН. Через 18 год., ізобутанол детектують за допомогою ВЕРХ або ГХ аналізу, використовуючи способи, що є добре відомими фахівцям, наприклад, як описано у розділі Загальні способи вище. Приклад 9 Клонування та експресія ацетолактатсинтази Для створення іншого експресуючого клону ацетолактатсинтази, ген budB клонують в вектор рТгс99А. Ген budB спочатку ампліфікують з pENTRSDD-TOPObudB (описаний у Прикладі 1) з використанням праймерів (N110.2 та N111.2, наведених як SEQ ID NOs:31 та 32, відповідно), які вводять сайти Sacl, Spel та Mfel на 5' кінці та сайти BbvC\, AflW та ВатН\ на 3' кінці. Одержаний 1,75 т.п.о. ПЛР продукт клонують в pCR4-Blunt ТОРО (Invitrogen) та ДНК послідовність підтверджують (використовуючи N130Seq праймери секвенування F1-F4 та R1-R4, наведені як SEQ ID NOs:40-47, відповідно). Ген budB потім вирізають з цього вектора за допомогою Sad та ВатНІ та клонують в pTrc99A (Amann et al., Gene 69(2):301-315 (1988)), одержуючи pTrc99A::budB. Вектор pTrc99A::budB трансформують в клітини Е. coli TOP10 та трансформанти інокулюють в LB середовище з добавкою 50 мкг/мл ампіциліну та вирощують протягом ночі при 37 °С. Аліквоту нічної культури використовують для інокуляції 50 мл LB середовища з добавкою 50 мкг/мл ампіциліну. Культуру інкубують при 37 °С зі струшуванням, поки OD600 не досягне 26 UA 103364 C2 5 10 15 20 25 30 35 40 45 50 55 60 значення від 0,6 до 0,8. Експресію budB з промотора Тгс потім індукують шляхом додавання 0,4 мМ IPTG. Готують також колби негативного контролю, які не індукують IPTG. Колби інкубують протягом 4 год. при 37 °С зі струшуванням. Безклітинні екстракти готують, як описано у Прикладі 1. Активність ацетолактатсинтази в безклітинних екстрактах вимірюють, як описано у Прикладі 1. через три години після індукування IPTG, було виявлено активність ацетолактатсинтази 8 одиниць/мг. Контрольний штам, що несе лише плазміду рТгс99А, мав активність ацетолактатсинтази 0,03 одиниць/мг. Приклад 10 Клонування та експресія редуктоізомерази ацетогідроксикислот Задачею цього Прикладу було клонувати ilvC з Е. coli K12 та експресувати його в Е. coli ТОР10. Ген ilvC ампліфікують з геномної ДНК Ε. coli K12 штаму FM5 (АТСС 53911) за допомогою ПЛР. Ген ilvC клонують та експресують у спосіб, подібний до описаного для клонування та експресії ilvC у Прикладі 2 вище. ПЛР використовують для ампліфікації /А/С з геному Е. coli FM5 з використанням праймерів N112.2 та N113.2 (SEQ ID NOs:33 та 34, відповідно). Праймери створювали сайти Sac\ та AfllW та оптимальний RBS на 5' кінці та сайти Not\, Nhe\ та ВатН\ на 3' кінці ilvC. 1,5 т.п.о. ПЛР продукт клонують в pCR4Blunt ТОРО згідно з протоколом виробника (Invitrogen), одержуючи pCR4Blunt TOPO::ilvC. Послідовність ПЛР продукту підтверджують, використовуючи праймери секвенування (NI31SeqF1-F3 та N131SeqR1-R3, наведені як SEQ ID NOs:48-53, відповідно). Для створення експресуючого клону, ген ilvC вирізають з pCR4Blunt TOPO::ilvC за допомогою Sad та ВатН\ та клонують в рТгс99А. Вектор pTrc99A::ilvC трансформують в клітини Е. coli TOP10 та експресію з промотора Тгс індукують шляхом додавання IPTG, як описано у прикладі 9. Безклітинні екстракти готують, як описано у Прикладі 1. Активність редуктоізомерази ацетогідроксикислоти в безклітинних екстрактах вимірюють, як описано у Прикладі 2. Через три години після індукування IPTG детектують активність редуктоізомерази ацетогідроксикислот 0,026 одиниць/мг. Контрольний штам, що несе лише плазміду рТгс99А, виявляв активність редуктоізомерази ацетогідроксикислот менш ніж 0,001 одиниць/мг. Приклад 11 Клонування та експресія дегідратази ацетогідроксикислот Метою цього Прикладу було клонування ilvD з Е. coli K12 та експресія його в Е. coli ТорЮ. Ген ilvD ампліфікують з геномної ДНК Ε. coli K12 штаму FM5 (АТСС 53911) за допомогою ПЛР. Ген ilvD клонують та експресують у спосіб, подібний до ilvC, описаний в Прикладі 10. ПЛР використовують для ампліфікації ilvD з геному Е. coli FM5 з використанням праймерів N114.2 та N115.2 (SEQ ID NOs:35 та 36, відповідно). Праймери створюють сайти Sad та Nhe\ та оптимальний RBS на 5' кінці та сайти Bsu36\, Рас\ та ВатН\ на 3' кінці ilvD. 1,9 т.п.о. ПЛР продукт клонують в pCR4Blunt TOPO згідно з протоколом виробника (Invitrogen), одержуючи pCR4Blunt TOPO::ilvD. Послідовність ПЛР продукту підтверджують (праймери секвенування N132SeqF1-F4 та N132SeqR1-R4, наведені як SEQ ID NOs:54-61, відповідно). Для створення експресуючого клону, ген ilvD вирізають з плазміди pCR4BluntTOPO::ilvD за допомогою Sad та ВатНІ та клонують в рТгс99А. Вектор pTrc99A::ilvD трансформують в клітини Е. coli 7ΌΡ10 та експресію з промотора Тгс індукують додаванням IPTG, як описано в Прикладі 9. Безклітинні екстракти готують, як описано у Прикладі 1. Активність дегідратази ацетогідроксикислот в безклітинних екстрактах вимірюють, як описано у Прикладі 3. Через три години після індукування IPTG детектують активність дегідратази ацетогідроксикислот 46 одиниць/мг. Контрольний штам, що несе лише плазміду, рТгс99А не виявляє детектованої активності дегідратази ацетогідроксикислот. Приклад 12 Клонування та експресія декарбоксилази розгалужених кетокислот Задачею цього Прикладу було клонування kivD з Lactococcus lactis та експресія його вЕ. coli TOP10. Ген kivD клонують та експресують у спосіб, подібний до описаного для ilvC у Прикладі 10 вище. ПЛР використовують для ампліфікації kivD з плазміди pUC57-kivD (див. Приклад 4, вище) з використанням праймерів N116.2 та N117.2 (SEQ ID NOs:37 та 38, відповідно). Праймери створюють сайти Sad та Расl та оптимальний RBS на 5' кінці та сайти Рсіl, Ανril, BglII та ВаmНl на 3' кінці kivD. 1.7 т.п.о. ПЛР продукт клонують в pCR4Blunt TOPO згідно з протоколом виробника (Invitrogen), одержуючи pCR4BluntTOP0::kivD. Послідовність ПЛР продукту підтверджують з використанням праймерів N133SeqF1-F4 та N133SeqR1-R4 (наведених як SEQ 27 UA 103364 C2 5 10 15 20 25 30 35 40 45 50 55 ID NOs:62-69, відповідно). Для створення експресуючого клону, ген kivD вирізають з плазміди pCR4Blunt TOPO::kivD за допомогою Sacl та ВаmНl та клонують в рТrс99А. Вектор pTrc99A::kivD трансформують в клітини Е. coli TOP10 та експресію з промотора Тгс індукують шляхом додавання IPTG, як описано у прикладі 9. Безклітинні екстракти готують, як описано у прикладі 1. Активність декарбоксилази розгалужених кетокислот в безклітинних екстрактах вимірюють, як описано у Прикладі 4, за винятком того, що реагент Purpald® (Aldrich, номер за каталогом 162892) використовують для детектування та кількісних вимірів альдегідних реакційних продуктів. Через три години після індукування IPTG, детектують активність декарбоксилази розгалужених кетокислот більш ніж 3,7 одиниць/мг. Контрольний штам, що несе лише плазміду рТгс99А, не виявляє детектованої активності декарбоксилази розгалужених кетокислот. Приклад 13 Експресія дегідрогенази розгалужених спиртів Е. coli містить нативний ген (yqhD), який був ідентифікований як 1,3пропандіолдегідрогеназа (патент США № 6514733). Білок YqhD має 40% ідентичність з AdhB (кодований adhB) з Clostridium, гаданою NADH-залежною бутанолдегідрогеназою. Ген yqhD поміщають під конститутивною експресією варіанта промотора глюкозоізомерази 1.6GI (SEQ ID NO. 70) в Ε. coli штам MG1655 1.6yqhD::Cm (WO 2004/033646) з використанням технології λ Red (Datsenko and Wanner, Proc. Natl. Acad. Sci. U.S.A. 97:6640 (2000)). MG1655 1.6yqhD::Cm містить касету FRT-CmR-FRT, так що маркер антибіотика може бути видалений. Аналогічно, нативний промотор був заміщений на промотор 1.5GI (WO 2003/089621) (SEQ ID NO. 71), з утворенням штаму MG1655 1.5GI-yqhD::Cm, тим самим, заміняючи промотор 1.6GI з MG1655 1.6yqhD::Cm на промотор 1.5GI. Штам MG 1655 1.5GI-yqhD::Cm вирощують в LB середовищі до середини лог-фази та безклітинний екстракти готують, як описано у Прикладі 1. Було знайдено, що цей штам має активність NADPH-залежної ізобутиральдегідредуктази при аналізі клітинних екстрактів шляхом відслідковування зменшення оптичного поглинання на 340 нм при рН 7,5 та 35 °С. Для створення другого експресуючого штаму, що містить 1.5GI yqhD::Cm, P1 лізат готують з MG1655 1.5GI yqhD::Cm та касету переносять в BL21 (DE3) (Invitrogen) шляхом трансдукції, створюючи BL21 (DE3) 1.5GI-yqhD::Cm. Приклад 14 Конструювання вектора трансформації перших чотирьох генів в метаболічному шляху біосинтезу . Задачею цього Прикладу було конструювання вектора трансформації, що включає перші чотири гени (тобто, budB, ilvC, ilvD та kivD) в метаболічному шляху біосинтезу ізобутанолу. Для конструювання вектора трансформації, спочатку, ген ilvC одержують з pTrc99A::ilvC (описаний у Прикладі 10) шляхом гідролізу за допомогою АШ та ВатНІ та клонують в pTrc99A::budB (описаний у Прикладі 9), який гідролізують задопомогою АШ та ВатНІ для продукування плазміди pTrc99A::budB-ilvC. Далі, гени ilvD та kivD одержують з pTrc99A::ilvD (описано у Прикладі 11) та pTrc99A::kivD (описано у Прикладі 12), відповідно, шляхом гідролізу за допомогою Nhe\ та Расі (ilvD) і Расі та ВатНІ (kivD). Ці гени вводять в pTrc99A::budB-ilvC, який спочатку гідролізують за допомогою Nhe\ та ВатН\, шляхом потрійного лігування. Присутність усіх чотирьох генів у кінцевій плазміді, pTrc99A::budB-ilvC-ilvD-kivD, підтверджують методами ПЛР скринінгу та рестрикційного гідролізу. Приклад 15 Експресія метаболічного шляху біосинтезу ізобутанолу в Е. соlі, вирощуваних на глюкозі Для створення штамів Е. соїі, що продукують ізобутанол, pTrc99A::budB-ilvC-ilvD-kivD (описано у прикладі 14) трансформують в Е. соїі MG1655 1.5GI yqhD::Cm та Е. соїі BL21 (DE3)1.5GI yqhD::Cm (описано у прикладі 13). Трансформанти спочатку спочатку вирощують в LB середовищі, що містить 50 мкг/мл канаміцину та 100 мкг/мл карбеніциліну. Клітини з цих культур використовують для інокуляції струшуваних колб (загальний об'єм приблизно 175 мл), що містять 50 або 170 мл середовища ТМЗа/глюкоза (з відповідними антибіотиками) для створення умов високого та низького вмісту кисню, відповідно. Середовище ТМЗа/глюкоза містить (на літр): глюкоза (10 г), КН2РО4 (13,6 г), лимонна кислота моногідрат (2,0 г), (NH4)2SO4 (3,0 г), MgSO4-7H2O (2,0 г), CaCI2-2H2O (0,2 г), цитрат заліза-амонію (0,33 г), тіамін-НСІ (1,0 мг), дріжджовий екстракт (0,50 г) і 10 мл розчину слідових елементів. рН доводять до 6,8 за допомогою NH4OH. Розчин слідових елементів містить: лимонна кислота-Н2О (4,0 г/л), MnSO4H2O (3,0 г/л), NaCI (1,0 г/л), FeSO4-7H2O (0,10 г/л), СоСІ2-6Н2О (0,10 г/л), ZnSO4-7H2O (0,10 г/л), CuSO4-5H2O (0,010 г/л), Н3ВО3 (0,010 г/л) і Na2MoO4-2H2O (0,010 г/л). 28
ДивитисяДодаткова інформація
Автори російськоюDonaldson Gail K., Eliot Andrew C., Flint Dennis, Maggio-Hall Lori Ann, Nagarajan Vasantha
МПК / Мітки
МПК: C12P 7/00, C12N 1/19, C12N 1/21, C12N 15/59
Мітки: спосіб, ферментативного, виробництва, чотириатомних, спиртів
Код посилання
<a href="https://ua.patents.su/232-103364-sposib-fermentativnogo-virobnictva-chotiriatomnikh-spirtiv.html" target="_blank" rel="follow" title="База патентів України">Спосіб ферментативного виробництва чотириатомних спиртів</a>
Спосіб ферментативного виробництва чотириатомних спиртів

Номер патенту: 103363
Опубліковано: 10.10.2013
Автори: Флінт Денніс, Маггіо-Холл Лорі Енн, Еліот Ендрю С., Дональдсон Гейл К., Нагараджан Васанта
МПК: C12N 15/52, C12N 1/21, C12N 1/19, C12P 7/16
Мітки: ферментативного, спосіб, виробництва, спиртів, чотириатомних
Формула / Реферат:
1. Рекомбінантна мікробна клітина-хазяїн, яка містить ферментативний шлях ізобутанолу, що включає молекули ДНК, які кодують набір поліпептидів, що каталізують наступні перетворення субстрату на продукт:і) піруват на ацетолактат;іі) ацетолактат на 2,3-дигідроксіізовалерат;ііі) 2,3-дигідроксіізовалерат на α-кетоізовалерат;iv) α-кетоізовалерат на ізобутирил-СоА;v) ізобутирил-СоА на ізобутиральдегід,...
Багатокомпонентна система для ферментативного окиснювання субстратів і спосіб ферментативного окиснювання

Номер патенту: 68421
Опубліковано: 16.08.2004
Автор: Кандусіо Антон
МПК: D21C 9/10, C12N 9/02, C11D 3/38, D21C 5/00, D21C 9/147
Мітки: спосіб, субстратів, ферментативного, багатокомпонентна, окиснювання, система
Формула / Реферат:
1. Багатокомпонентна система для опосередковуваного медіатором ферментативного окиснювання, що включає каталізатор окиснювання, окисник і медіатор, яка відрізняється тим, щоа) каталізатор окиснювання вибрано із групи марганцевих оксидаз,б) окисник вибрано із групи, куди входять кисень і кисневмісні сполуки,в) медіатор вибрано із групи сполук, що містять іони Мn.2. Багатокомпонентна система за п. 1, яка...
Спосіб одержання аліфатичних спиртів для виробництва моторного палива

Номер патенту: 4647
Опубліковано: 28.12.1994
Автори: Тараканов Анатолій Олексійович, Павлій Людмила Василівна, Францєнюк Іван Васільєвіч, Леонов Валерій Євгенович, Зубілін Іван Георгієвич, Успенський Сергій Константинович
МПК: C07C 31/00, C07C 29/151
Мітки: аліфатичних, спиртів, одержання, виробництва, моторного, палива, спосіб
Формула / Реферат:
1. Способ получения алифатических спиртов для производства моторных топлив из синтез-газа при повышенной температурена цинк-хром-калий-ванадиевом катализаторе, отличающийся тем, что в зону катализа вместе с синтез-газом вводят этиленсодержащий газ с концентрацией его в газовой фазе в пределах 0,05-3,00 об. %, причем соотношение Н2/СО в синтез-газе составляет 1,8-2,2, и процесс ведут при температуре 378-400°С.2. Способ по п. 1,...
Спосіб виробництва дубової клепки для витримки коньячних спиртів у великих резервуарах

Номер патенту: 14137
Опубліковано: 15.05.2006
Автори: Луканін Олександр Сергійович, Агафонов Михайло Федорович, Крайнікова Ірина Геннадіївна, Зражва Сергій Григорович, СИДОРЕНКО ОЛЕКСАНДР МИКОЛАЙОВИЧ
Мітки: коньячних, дубової, спосіб, резервуарах, виробництва, клепки, великих, витримки, спиртів
Формула / Реферат:
Спосіб виробництва дубової клепки, що використовується для витримки коньячних спиртів у великих резервуарах, який передбачає виробництво клепки II типу з деревини дуба шляхом радіального та тангенціального розпилювання за розмірами у відповідності до чинних технічних умов, відсортування клепки за такими нормативами вад:- сучки - допускаються без ознак гнилей;- торцеві тріщини - допускаються;- пластові тріщини -...
Спосіб виготовлення добавки для ферментативного розкладання мікотоксинів, добавка та її застосування

Номер патенту: 103335
Опубліковано: 10.10.2013
Автори: Шатцмайр Герд, Хартінгер Доріс, Молль Вульф-Дітер, Біндер Єва-Марія, Грісслер Карін
МПК: C12N 9/18, C12N 9/10, C12P 7/06, C12N 15/52, A23K 1/165
Мітки: мікотоксинів, добавка, застосування, добавки, спосіб, виготовлення, ферментативного, розкладання
Формула / Реферат:
1. Спосіб виготовлення добавки для ферментативного розкладання фумонізинів у рослинній сировині, який відрізняється тим, що забезпечують принаймні одну нуклеїново-кислотну послідовність генів, що відповідає послідовностям SEQ ID NOs:8, 18 і 24, експресують принаймні одну послідовність нуклеїнових кислот у прокаріотичних або еукаріотичних клітинах і використовують принаймні один приготований таким чином фермент, що відповідає послідовностям...
Попередній патент: Спосіб ферментативного виробництва чотириатомних спиртів
Наступний патент: Установка для отримання тепла або холоду (варіанти)
Випадковий патент: Спосіб керування фазами заряджання і розряджання опорного конденсатора носія даних і схемний пристрій для його здійснення