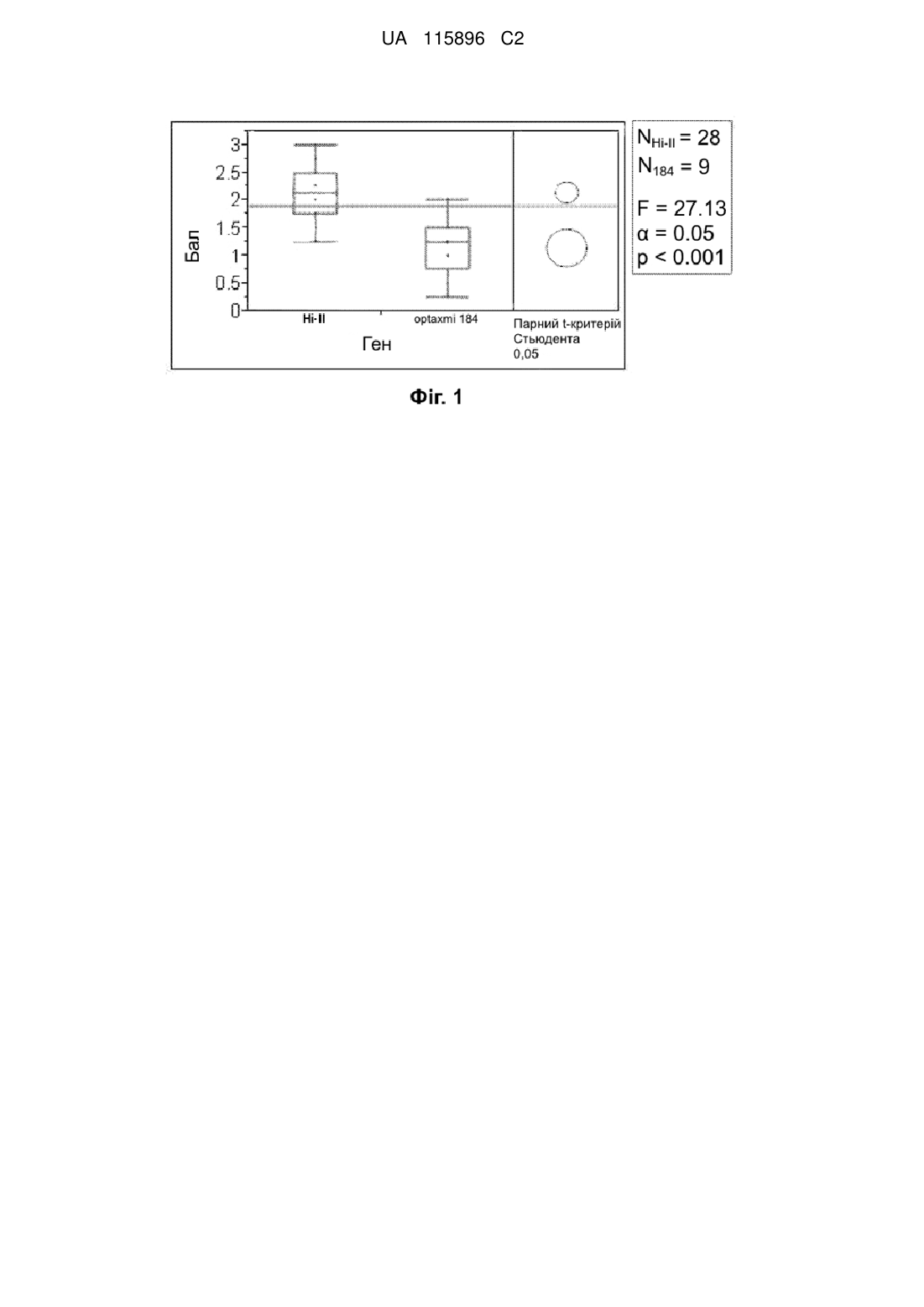
Застосування токсину axmi184 для контролю кукурудзяного жука







Формула / Реферат
1. Спосіб знищення або контролю популяції шкідника-кукурудзяного жука, що включає приведення зазначеної популяції у контакт з пестицидно ефективною кількістю поліпептиду, що містить амінокислотну послідовність, яка щонайменше на 95 % ідентична амінокислотній послідовності з SEQ ID NO: 3.
2. Спосіб захисту рослини кукурудзи від шкідника-кукурудзяного жука, що включає експресію у рослині кукурудзи або її клітині нуклеотидної послідовності, функціонально пов'язаної з промотором, здатним керувати експресією нуклеотидної послідовності у рослинній клітині, де зазначена нуклеотидна послідовність вибрана з групи, що складається з:
a) нуклеотидної послідовності, викладеної у SEQ ID NO: 1 або 2; та
b) нуклеотидної послідовності, яка кодує поліпептид, що містить амінокислотну послідовність, яка щонайменше на 95 % ідентична амінокислотній послідовності з SEQ ID NO: 3, де зазначений поліпептид характеризується пестицидною активністю проти шкідника-кукурудзяного жука.
3. Спосіб підвищення врожайності рослини кукурудзи, що включає вирощування в полі рослини кукурудзи або її насінини, які мають стабільно вбудовану в їхній геном ДНК-конструкцію, що містить нуклеотидну послідовність, функціонально пов'язану з промотором, здатним керувати експресією нуклеотидної послідовності в рослинній клітині, де зазначена нуклеотидна послідовність вибрана з групи, що складається з:
a) нуклеотидної послідовності, викладеної у SEQ ID NO: 1 або 2; та
b) нуклеотидної послідовності, яка кодує поліпептид, що містить амінокислотну послідовність, яка щонайменше на 95 % ідентична амінокислотній послідовності з SEQ ID NO: 3, де зазначений поліпептид характеризується пестицидною активністю проти шкідника-кукурудзяного жука; де зазначене поле заражене шкідником-кукурудзяним жуком.
4. Спосіб за пп. 1-3, де зазначений шкідник є шкідником-західним кукурудзяним жуком.
5. Спосіб за п. 2 або п. 3, де зазначена рослина додатково містить одну або декілька нуклеотидних послідовностей, що кодують один або декілька додаткових токсинів для комах.
6. Застосування рослини за п. 2 або п. 3 для контролю шкідника-кукурудзяного жука.
Текст
Реферат: Винахід належить до способу застосування токсину AXMI184 для знищення або контролю кукурудзяного кореневого жука, шляхом приведення шкідника у контакт з пестицидно ефективною кількістю поліпептиду, що містить вказаний токсин. Також винахід належить до способу підвищення врожайності рослин шляхом експресії токсину за даним винаходом. UA 115896 C2 (12) UA 115896 C2 UA 115896 C2 5 10 15 20 25 30 35 40 45 50 55 60 Посилання на перелік послідовностей Офіційна копія переліку послідовностей надається у електронній формі за допомогою EFSWeb як відформатований за ASCII перелік послідовностей у файлі з назвою "APA136001SEQLIST.txt", створеному 16 січня 2013, що має розмір 14 кілобайт і подається одночасно з даним описом. Перелік послідовностей, що міститься у цьому відформатованому за ASCII документі, є частиною даного опису і включений в даний документ за допомогою посилання у всій своїй повноті. Галузь винаходу Даний винахід стосується галузі молекулярної біології. Передбачені нові гени, які кодують пестицидні білки. Ці білки та послідовності нуклеїнових кислот, які їх кодують, є застосовними в одержанні пестицидних складів і в одержанні трансгенних рослин, стійких до шкідників. Передумови створення винаходу Кукурудзяний жук є важливим сільськогосподарським шкідником. Західний кукурудзяний жук, Diabrotica virgifera virgifera, є одним з видів кукурудзяного жука у Північній Америці, що спричиняє найбільші спустошення, особливо в областях вирощування кукурудзи на Середньому Заході, таких як Айова. Як личинки, так і дорослі кукурудзяні жуки можуть пошкоджувати рослини кукурудзи. Личинки, що нещодавно вивелися, живляться, переважно, кореневими волосками та зовнішньою тканиною кореня. Зі зростанням личинок зростають їхні потреби у їжі, тому для живлення вони проникають всередину коренів. Зазвичай, пошкодження личинками є найбільш серйозними після того, як добре розвинеться вторинна коренева система і розвиваються опорні корені. Кінчики коренів будуть ставати коричневими та часто є пронизаними ходами всередині та відгризені до стебла рослини. Можна знайти личинок, що роблять ходи всередині більш великих коренів, а у деяких випадках у надземних частинах рослини. У спробах контролювати їхні популяції були застосовані сівозміна, стратегії раннього висаджування, інсектициди та трансгенні рослини, які експресують токсини проти кукурудзяного жука. Трансгенні сільськогосподарські культури, розроблені для продукування інсектицидних токсинів, що одержані з бактерії Bacillus thuringiensis (Bt), у 2010 році були посаджені на більш ніж 58 мільйонах гектарів по всьому світі (James C (2010) Global Status of Commercialized Biotech/GM Crops: 2010. ISAAA Brief No. 42. Ithaca, New York: ISAAA). Переваги Btсільськогосподарських культур включають знижене застосування шкідливих інсектицидів та регіональне пригнічення деяких основних сільськогосподарських шкідників (Carrière et al. (2003) Proc Natl Acad Sci USA 100: 1519-1523; Cattaneo et al. (2006) Proc Natl Acad Sci U S A 103: 75717576; Huang et al. (2005) Science 308: 688-690; Wu (2008) Science 321: 1676-1678; та Hutchison et al. (2010) Science 330: 222-225). Західний кукурудзяний жук Diabrotica virgifera virgifera LeConte (Coleoptera: Chrysomelidae) є одним з найбільш важливих шкідників маїсу у Сполучених Штатах, при цьому живлення личинок на коренях маїсу призводить до більшості втрат врожаю від цього шкідника. Починаючи з 2003 року, Bt-маїс був введений у комерційних оборот для контролю личинок західного кукурудзяного жука, і він був швидко прийнятий фермерами, так що становив понад 45 % врожаю маїсу в Сполучених Штатах у 2009 році (James C (2009) Global Status of Commercialized Biotech/GM Crops: 2009. ISAAA Brief No. 41. Ithaca, New York: ISAAA; Environmental Protection Agency (2003) Biopesticides registration action document: event MON863 Bacillus thuringiensis Cry3Bbl Com). Стратегію рефугіумів застосовують у Сполучених Штатах та інших країнах для затримки розвитку стійкості шкідників до Bt-сільськогосподарських культур (Gould F (1998) Annu Rev Entomol 43: 701-72). У цій стратегії застосовуються рослини-хазяїни, що не містять токсини Bt, як рефугіум для генотипів, сприйнятливих до Bt. Однак, західний кукурудзяний жук неодноразово демонстрував свою здатність адаптуватися до стратегій боротьби із шкідниками (Gray et al. (2009) Annu Rev Entomol 54: 303-321). Приклади включають еволюцію стійкості до традиційних інсектицидів та агротехнічного прийому сівозміни (Meinke et al. (1998) J Econ Entomol 91: 594-600; Parimi et al. (2006) J Econ Entomol 25: 269-274; Levine et al. (2002) Am Entomol 48: 94-107). Крім того, Gassmann et al. (2011, PLoS ONE 6(7): e22629) нещодавно повідомив, що поля, визначені фермерами як такі, що мають серйозні пошкодження Bt-маїсу внаслідок живлення кукурудзяного жука, містили популяції західного кукурудзяного жука, що демонстрували значно вищу здатність до виживання на Cry3Bbl-маїсі у лабораторних біологічниханалізах, ніж західний кукурудзяний жук з полів, не пов'язаних з таким пошкодженням внаслідок живлення. Короткий опис винаходу Передбачені композиції та способи надання пестицидної активності бактеріям, рослинам, рослинним клітинам, тканинам та насінню. Зокрема, передбачені способи знищення або 1 UA 115896 C2 5 10 15 20 25 30 35 40 45 50 55 60 контролю популяції шкідника-твердокрилої комахи, зокрема, популяції шкідника-кукурудзяного жука. Способи включають приведення шкідника-твердокрилої комахи у контакт з пестицидно ефективною кількістю поліпептиду, що містить токсин до шкідника-твердокрилої комахи, зокрема, токсин до кукурудзяного жука. У різних варіантах здійснення токсин до шкідникатвердокрилої комахи включає амінокислотну послідовність з SEQ ID NO:3, або її пестицидно ефективні варіанти або фрагменти. У деяких варіантах здійснення спосіб захисту рослини або її клітини від популяції шкідника-твердокрилої комахи, зокрема, шкідника-кукурудзяного жука, включає експресію в рослині або її клітині нуклеїнової кислоти, що кодує SEQ ID NO: 3 або її варіант чи фрагмент, де нуклеїнова кислота функціонально пов'язана з промотором, здатним керувати експресією нуклеїнової кислоти в рослинній клітині. Додатково включені способи підвищення врожайності рослини, які включають вирощування на полі рослини або її насінини, що мають стабільно вбудовану в їхній геном ДНК-конструкцію, яка містить нуклеїнову кислоту, функціонально пов'язану з промотором, здатним керувати експресією нуклеїнової кислоти у рослинній клітині, де нуклеїнова кислота кодує SEQ ID NO: 3 або її пестицидно ефективний варіант або фрагмент. Композиції та способи за даним винаходом є застосовними для одержання організмів з підвищеною стійкістю або толерантністю до шкідника. Ці організми та композиції, що включають організми, є придатними для сільськогосподарських цілей. Короткий опис графічних матеріалів На фігурі 1 показана ефективність Axmi184 у зниженні пошкодження коренів західним кукурудзяним жуком. Детальний опис Даний винахід відноситься до способів регулювання стійкості або толерантності до шкідника в організмів, особливо рослинах або рослинних клітинах. Під виразом "стійкість" мається на увазі, що шкідник (наприклад, комаха) гине після прийому всередину або іншому контакті з поліпептидами за даним винаходом. Під виразом "толерантність" мається на увазі погіршення або зниження руху, живлення, розмноження або інших функцій шкідника. Способи передбачають трансформування організмів за допомогою нуклеотидної послідовності, що кодує пестицидний білок за даним винаходом. Зокрема, нуклеотидні послідовності за даним винаходом є застосовними для одержання рослин і мікроорганізмів, які мають пестицидну активність. Описані у даному документі способи є застосовними для контролю або знищення популяцій шкідника-твердокрилої комахи та отримання композицій з пестицидною активністю проти шкідників-твердокрилих комах. Під виразом "пестицидний токсин" або "пестицидний білок" мається на увазі токсин, який має токсичну активність проти одного або декількох шкідників-твердокрилих комах, в тому числі, але без обмежень, шкідників західного кукурудзяного жука (наприклад, Diabrotica virgifera virgifera), північного кукурудзяного жука (наприклад, Diabrotica barberi) та південного кукурудзяного жука (наприклад, Diabrotica undecimpunctata howardi), або білок, який має гомологію з таким білком. Пестицидні білки включають амінокислотні послідовності, виведені з нуклеотидних послідовностей повної довжини, розкритих у даному документі, та амінокислотні послідовності, які є коротшими за послідовності повної довжини або через застосування альтернативного сайту ініціювання, що розташований нижче, або через обробку, яка дає більш короткий білок, що має пестицидну активність. Обробка може відбуватися в організмі, у якому експресується цей білок, або в шкіднику після прийому всередину даного білка. В конкретних варіантах здійснення пестицидний білок включає білок Axmi184, викладений у SEQ ID NO: 3, а також його варіанти та фрагменти. Таким чином, у даному документі передбачені способи знищення або контролю популяції шкідника-твердокрилої комахи, які включають приведення шкідника у контакт з композицією, або піддавання шкідника впливу композиції, що містить пестицидний токсин за даним винаходом. Виділені молекули нуклеїнових кислот та їхні варіанти та фрагменти Один аспект даного винаходу стосується виділених або рекомбінантних молекул нуклеїнових кислот, що включають в себе нуклеотидні послідовності, які кодують пестицидні білки та поліпептиди або їхні біологічно активні частини, а також молекул нуклеїнових кислот, яких є придатними для застосування як гібридизаційні зонди для визначення молекул нуклеїнових кислот, що кодують білки з ділянками з гомологією послідовності. Також даним документом охоплені нуклеотидні послідовності, здатні до гібридизації з нуклеотидними послідовностями за даним винаходом в жорстких умовах, як визначено у іншому розділі даного документа. Як застосовується у даному документі, вираз "молекула нуклеїнової кислоти", як 2 UA 115896 C2 5 10 15 20 25 30 35 40 45 50 55 60 передбачається, включає в себе молекули ДНК (наприклад, рекомбінантну ДНК, кДНК або геномну ДНК) і молекули РНК (наприклад, мРНК), а також аналоги ДНК або РНК, отримані із застосуванням нуклеотидних аналогів. Молекула нуклеїнової кислоти може бути одноланцюговою або дволанцюговою, але переважно є дволанцюговою ДНК. Вираз "ізольована" або "рекомбінантна" послідовність нуклеїнової кислоти (або ДНК) застосовується у даному документі для позначення послідовності нуклеїнової кислоти (або ДНК), яка вже не перебуває у своєму природному оточенні, наприклад, знаходиться в умовах in vitro або в рекомбінантній бактерії або рослинній клітині-хазяїні. У деяких варіантах здійснення виділена або рекомбінантна нуклеїнова кислота не містить послідовності (переважно послідовності, які кодують білок), що у природному стані фланкують нуклеїнову кислоту (тобто послідовності, які розташовані на 5'- та 3'-кінцях нуклеїнової кислоти) у геномній ДНК організму, з якого нуклеїнова кислота отримана. В контексті даного винаходу при застосуванні виразу "виділений" стосовно молекул нуклеїнової кислоти, він не включає виділені хромосоми. Наприклад, в різних варіантах здійснення ізольована молекула нуклеїнової кислоти, яка кодує дельта-ендотоксин, може містити менше приблизно 5 т.о., 4 т.о., 3 т.о., 2 т.о., 1 т.о., 0,5 т.о. або 0,1 т.о. з нуклеотидних послідовностей, які у природному стані фланкують молекулу нуклеїнової кислоти в геномній ДНК клітини, з якої нуклеїнова кислота одержана. В різних варіантах здійснення білок дельта-ендотоксину, який практично не містить клітинний матеріал, включає в себе препарати білка, що мають менше приблизно 30 %, 20 %, 10 % або 5 % (за сухою вагою) білка, що не є дельта-ендотоксином (у даному документі також позначений як "забруднюючий білок"). Нуклеотидні послідовності, що кодують білки за даним винаходом, включають в себе послідовність, викладену у SEQ ID NO: 1-2, та її варіанти, фрагменти або комплементарні до неї послідовності. Під виразом "комплементарна послідовність" мається на увазі нуклеотидна послідовність, яка є достатньо комплементарною до даної нуклеотидної послідовності, що вона може гібридизуватися з даною нуклеотидною послідовністю з утворенням, таким чином, стабільного дуплекса. Відповідні амінокислотні послідовності пестицидних білків, закодовані цими нуклеотидними послідовностями, викладені у SEQ ID NO: 3. Молекули нуклеїнових кислот, які є фрагментами цих нуклеотидних послідовностей, що кодують пестицидні білки, також охоплюються даним винаходом. Під виразом "фрагмент" мається на увазі частина нуклеотидної послідовності, що кодує пестицидний білок. Фрагмент нуклеотидної послідовності може кодувати біологічно активну частину пестицидного білка, або він може являти собою фрагмент, який може бути застосований як гібридизаційний зонд або ПЛР-праймер із застосуванням способів, розкритих нижче. Залежно від передбаченого застосування, молекули нуклеїнових кислот, які є фрагментами нуклеотидної послідовності, що кодує пестицидний білок, становлять щонайменше приблизно 50, 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1100, 1200, 1300, 1350, 1400 суміжних нуклеотидів або аж до числа нуклеотидів, присутніх в нуклеотидній послідовності повної довжини, що кодує пестицидний білок, розкритий у даному документі. Під виразом "суміжні" нуклеотиди маються на увазі нуклеотидні залишки, які безпосередньо примикають один до одного. Фрагменти нуклеотидних послідовностей за даним винаходом будуть кодувати фрагменти білка, які зберігають біологічну активність пестицидного білка і, отже, зберігають пестицидну активність. Таким чином, також охоплюються біологічно активні фрагменти поліпептидів, розкритих у даному документі. Під виразом "зберігає активність" мається на увазі, що фрагмент буде характеризуватися щонайменше приблизно 30 %, щонайменше приблизно 50 %, щонайменше приблизно 70 %, 80 %, 90 %, 95 % або більшою пестицидною активністю пестицидного білка. У одному варіанті здійснення пестицидна активність являє собою активність, спрямовану на знищення твердокрилих комах. У іншому варіанті здійснення пестицидна активність являє собою активність, спрямовану на знищення лускокрилих комах. У іншому варіанті здійснення пестицидна активність являє собою активність, спрямовану на знищення нематод. У іншому варіанті здійснення пестицидна активність являє собою активність, спрямовану на знищення двокрилих комах. У іншому варіанті здійснення пестицидна активність являє собою активність, спрямовану на знищення напівтвердокрилих комах. Способи вимірювання пестицидної активності добре відомі в даній галузі техніки. Див., наприклад, Czapla and Lang (1990) J. Econ. Entomol. 83:2480-2485; Andrews et al. (1988) Biochem. J. 252:199-206; Marrone et al. (1985) J. of Economic Entomology 78:290-293; та патент США № 5743477, усі з яких включені в даний документ за допомогою посилання у всій своїй повноті. Фрагмент нуклеотидної послідовності, що кодує пестицидний білок, який кодує біологічно активну частину білка за даним винаходом, буде кодувати щонайменше приблизно 15, 25, 30, 50, 75, 100, 125, 150, 175, 200, 250, 300, 350, 400, 450 суміжних амінокислот або аж до загальної 3 UA 115896 C2 5 10 15 20 25 30 35 40 45 50 55 60 кількості амінокислот, присутніх у пестицидному білку повної довжини за даним винаходом. У деяких варіантах здійснення фрагмент являє собою фрагмент, одержаний внаслідок протеолітичного розщеплення. Наприклад, фрагмент, одержаний внаслідок протеолітичного розщеплення, може характеризуватися усіченням на N-кінці або С-кінці, що становить щонайменше приблизно 100 амінокислот, приблизно 120, приблизно 130, приблизно 140, приблизно 150 або приблизно 160 амінокислот, відносно SEQ ID NO: 3. У деяких варіантах здійснення фрагменти, що охоплені даним документом, одержують внаслідок видалення Скінцевого домену кристалізації, наприклад, шляхом протеолізу або шляхом вставки стоп-кодону в кодуючу послідовність. Переважні пестицидні білки за даним винаходом кодуються нуклеотидною послідовністю, яка є достатньо ідентичною з нуклеотидною послідовністю з SEQ ID NO: 1-2, або пестицидні білки є достатньо ідентичними з амінокислотною послідовністю, викладеною у SEQ ID NO: 3. Під виразом "достатньо ідентичний" мається на увазі амінокислотна або нуклеотидна послідовність, яка характеризується щонайменше приблизно 60 % або 65 % ідентичністю послідовності, приблизно 70 % або 75 % ідентичністю послідовності, приблизно 80 % або 85 % ідентичністю послідовності, приблизно 90 %, 91 %, 92 %, 93 %, 94 %, 95 %, 96 %, 97 %, 98 %, 99 % або більшою ідентичністю послідовності порівняно з еталонною послідовністю із застосуванням однієї з програм вирівнювання, описаних у даному документі, із застосуванням стандартних параметрів. Фахівцю в даній галузі буде зрозуміло, що ці значення можна відповідним чином регулювати для визначення відповідної ідентичності білків, закодованих двома нуклеотидними послідовностями, з урахуванням виродженості кодонів, амінокислотної подібності, розташування рамки зчитування тощо. Для визначення відсотка ідентичності двох амінокислотних послідовностей або двох послідовностей нуклеїнових кислот, для цілей оптимального порівняння послідовності вирівнюють. Відсоток ідентичності між двома послідовностями являє собою функцію кількості ідентичних положень, що є загальними для послідовностей (тобто відсоток ідентичності = кількість ідентичних положень/загальна кількість положень (наприклад, положення, що перекриваються) × 100). У одному варіанті здійснення дві послідовності мають однакову довжину. У іншому варіанті здійснення відсоток ідентичності розраховують в межах усієї еталонної послідовності (тобто послідовності, розкритої у даному документі як будь-яка з SEQ ID NO: 1-3). Відсоток ідентичності між двома послідовностями можна визначати із застосуванням методик, подібних до описаних нижче, з введенням гепів або без них. Зазвичай, під час розрахунку відсотка ідентичності враховуються точні співпадіння. Геп, тобто положення у вирівнюванні, де залишок присутній в одній послідовності, але відсутній в іншій, вважають положенням з неідентичними залишками. Визначення відсотка ідентичності між двома послідовностями може бути досягнуто із застосуванням математичного алгоритму. Необмежуючим прикладом математичного алгоритму, що використовується для порівняння двох послідовностей, є алгоритм з Karlin and Altschul (1990) Proc. Natl. Acad. Sci. USA 87: 2 264, модифікований як у Karlin and Altschul (1993) Proc Natl. Acad. Sci. USA 90:5873-5877. Такий алгоритм включений в програми BLASTN та BLASTX з Altschul et al. (1990) J. Mol. Biol. 215:403. Нуклеотидні пошуки BLAST можуть бути виконані за допомогою програми BLASTN, бал = 100, довжина слова = 12, з одержанням нуклеотидних послідовностей, що є гомологічними до пестицидоподібних молекул нуклеїнових кислот за даним винаходом. Білкові пошуки BLAST можуть бути виконані за допомогою програми BLASTX, бал = 50, довжина слова = 3, з одержанням амінокислотних послідовностей, що є гомологічними до молекул пестицидного білка за даним винаходом. Для одержання вирівнювань з гепами для цілей порівняння може використовуватись програма Gapped BLAST (в BLAST 2.0), як описано в Altschul et al. (1997) Nucleic Acids Res. 25:3389. Альтернативно, для виконання ітераційного пошуку, який виявляє віддалені взаємозв'язки між молекулами, може бути застосований PSI-Blast. Див. Altschul et al. (1997) вище. При використанні програм BLAST, Gapped BLAST та PSI-BLAST можуть бути застосовані параметри відповідних програм (наприклад, BLASTX та BLASTN) за замовчуванням. Вирівнювання також можна виконувати вручну шляхом огляду. Іншим необмежуючим прикладом математичного алгоритму, що застосовується для порівняння послідовностей, є алгоритм ClustalW (Higgins et al. (1994) Nucleic Acids Res. 22:46734680). ClustalW порівнює послідовності і вирівнює всю амінокислотну послідовність або послідовність ДНК, і, таким чином, може надавати дані про консервативність послідовності усієї амінокислотної послідовності. Алгоритм ClustalW застосовується в декількох комерційно доступних програмних пакетах аналізу ДНК/амінокислот, таких як ALIGNX, що є модулем програми Vector NTI Suite (Invitrogen Corporation, Карлсбад, Каліфорнія). Після вирівнювання 4 UA 115896 C2 5 10 15 20 25 30 35 40 45 50 55 60 амінокислотних послідовностей за допомогою ClustalW можна оцінювати відсоток ідентичності амінокислотних послідовностей. Необмежуючим прикладом програмного забезпечення, застосовного для аналізу вирівнювання ClustalW є GENEDOC™. GENEDOC™ (Karl Nicholas) створює можливість оцінки подібності амінокислотної послідовності (або ДНК) та ідентичність між декількома білками. Іншим необмежуючим прикладом математичного алгоритму, що використовується для порівняння послідовностей, є алгоритм з Myers and Miller (1988) CABIOS 4:11-17. Такий алгоритм включений в програму ALIGN (версія 2.0), яка є частиною пакету програмного забезпечення GCG Wisconsin Genetics, версія 10 (доступна від Accelrys, Inc., 9685 Scranton Rd., Сан-Дієго, Каліфорнія, США). При використанні програми ALIGN для порівняння амінокислотних послідовностей можна застосовувати таблицю ваги заміни залишку PAM 120, штраф за подовження гепу 12 та штраф за введення гепу 4. Якщо не вказане інше, GAP версії 10, яка застосовує алгоритм з Needleman and Wunsch (1970) J. Mol. Biol. 48 (3): 443-453, буде застосовуватись для визначення ідентичності або подібності послідовності із застосуванням наступних параметрів: % ідентичності та % подібності для нуклеотидної послідовності із застосуванням штрафу за введення гепу 50 і штрафу за подовження гепу 3, та матриці замін nwsgapdna.cmp; % ідентичності або % подібності амінокислотної послідовності із застосуванням штрафу за введення гепу 8 і штрафу за подовження гепу 2, та матрицю замін BLOSUM62. Також можуть бути застосовані еквівалентні програми. Під виразом "еквівалентна програма" мається на увазі будь-яка програма порівняння послідовності, яка, для будь-яких двох послідовностей, які розглядаються, створює вирівнювання, що характеризується ідентичними співпадіннями нуклеотидних залишків та ідентичним відсотком ідентичності послідовності порівняно з відповідним вирівнюванням, виконаним за допомогою GAP версії 10. Даний винахід також охоплює варіанти молекул нуклеїнових кислот. "Варіанти" нуклеотидних послідовностей, що кодують пестицидний білок, включають ті послідовності, що кодують пестицидні білки розкриті у даному документі, але які відрізняються за консервативними замінами внаслідок виродженості генетичного коду, а також ті, які є достатньо ідентичними, як обговорюється вище. Алельні варіанти, що зустрічаються у природі, можна визначати із застосуванням добре відомих методик молекулярної біології, таких як полімеразна ланцюгова реакція (ПЛР) та методики гібридизації, як вказано нижче. Варіанти нуклеотидних послідовностей також включають одержані синтетичним шляхом нуклеотидні послідовності, які були створені, наприклад, із застосуванням сайт-спрямованого мутагенезу, але які все ще кодують пестицидні білки, розкриті в даному винаході, як обговорюється нижче. Варіанти білків, охоплених даним винаходом, є біологічно активними, тобто вони продовжують характеризуватися потрібною біологічною активністю нативного білка, тобто пестицидною активністю. Під виразом "зберігає активність" мається на увазі, що варіант буде характеризуватися щонайменше приблизно 30 %, щонайменше приблизно 50 %, щонайменше приблизно 70 %, щонайменше приблизно 80 % пестицидною активністю нативного білка. Способи вимірювання пестицидної активності добре відомі в даній галузі техніки. Див., наприклад, Czapla and Lang (1990) J. Econ. Entomol. 83: 2480-2485; Andrews et al. (1988) Biochem. J. 252:199-206; Marrone et al. (1985) J. of Economic Entomology 78:290-293; та патент США № 5743477, усі з яких включені в даний документ за допомогою посилання у всій своїй повноті. Крім того, фахівцеві в даній галузі буде зрозуміло, що зміни можна вводити шляхом здійснення мутацій у нуклеотидних послідовностях за даним винаходом, які, таким чином, призводять до змін у амінокислотній послідовності закодованих пестицидних білків без зміни біологічної активності білків. Таким чином, варіанти виділених молекул нуклеїнових кислот можна створювати шляхом введення однієї або декількох нуклеотидних замін, додавань або делецій у відповідну нуклеотидну послідовність, що розкрита у даному документі, отже, одна або декілька амінокислотних замін, додавань або делецій вводяться у закодований білок. Мутації можна вводити за допомогою стандартних методик, таких як сайт-спрямований мутагенез та мутагенез, опосередкований ПЛР. Такі варіанти нуклеотидних послідовностей також охоплені даним винаходом. Наприклад, консервативні амінокислотні заміни можна робити за одним або декількома, передбаченими, несуттєвими амінокислотними залишками. Вираз "несуттєвий" амінокислотний залишок позначає залишок, який може бути змінений у послідовності пестицидного білка дикого типу без зміни біологічної активності, тоді як "суттєвий" амінокислотний залишок є потрібним для біологічної активності. Вираз "консервативна амінокислотна заміна" являє собою заміну, за якої амінокислотний залишок заміняється на амінокислотний залишок, який має подібний бічний ланцюг. Сімейства амінокислотних залишків, які мають подібні бічні ланцюги, були визначені в 5 UA 115896 C2 5 10 15 20 25 30 35 40 45 50 55 60 даній галузі техніки. Ці сімейства включають амінокислоти з основними бічними ланцюгами (наприклад, лізин, аргінін, гістидин), кислими бічними ланцюгами (наприклад, аспарагінова кислота, глутамінова кислота), незарядженими полярними бічними ланцюгами (наприклад, гліцин, аспарагін, глутамін, серин, треонін, тирозин, цистеїн), неполярними бічними ланцюгами (наприклад, аланін, валін, лейцин, ізолейцин, пролін, фенілаланін, метіонін, триптофан), бетарозгалуженими бічними ланцюгами (наприклад, треонін, валін, ізолейцин) та ароматичними бічними ланцюгами (наприклад, тирозин, фенілаланін, триптофан, гістидин). Амінокислотні заміни можна робити у неконсервативних ділянках зі збереженням функції. Як правило, такі заміни не будуть робити у консервативних амінокислотних залишках або амінокислотних залишках, розташованих у межах консервативного мотиву, де такі залишки є суттєвими для активності білка. Приклади залишків, які є консервативними та можуть бути суттєвими для активності білка включають, наприклад, залишки, які ідентичні в усіх білків, що містяться у вирівнюванні подібних або споріднених токсинів, та у послідовностях за даним винаходом (наприклад, залишки, які є ідентичними у вирівнюванні гомологічних білків). Приклади залишків, які є консервативними, але які можуть дозволяти консервативні амінокислотні заміни та все ще зберігати активність, включають, наприклад, залишки, які характеризуються лише консервативними замінами в усіх білках, що містяться у вирівнювання подібних або споріднених токсинів, та у послідовностях за даним винаходом (наприклад, залишки, які характеризуються лише консервативними замінами в усіх білках, що включені у вирівнювання гомологічних білків). Однак, фахівцеві у даній галузі буде зрозуміло, що функціональні варіанти можуть мати незначні консервативні або неконсервативні зміни у консервативних залишках. Альтернативно, варіанти нуклеотидних послідовностей можна отримати шляхом введення мутацій випадковим чином уздовж всієї або частини кодуючої послідовності, наприклад, шляхом насичуючого мутагенезу, та отриманих мутантів можна піддавати скринінгу щодо здатності надавати пестицидну активність для визначення мутантів, які зберігають активність. Після мутагенезу, закодований білок може експресуватися рекомбінантним шляхом та активність цього білка можна визначати із застосуванням стандартних методик аналізу. Із застосуванням способів, таких як ПЛР, гібридизація тощо, можна визначити відповідні пестицидні послідовності, при цьому такі послідовності характеризуються суттєвою ідентичністю з послідовностями за даним винаходом. Див., наприклад, Sambrook and Russell (2001) Molecular Cloning: A Laboratory Manual. (Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY), та Innis, et al. (1990) PCR Protocols: A Guide to Methods and Applications (Academic Press, NY). У способі гібридизації, всю або частину пестицидної нуклеотидної послідовності можна застосовувати для скринінгу кДНК або геномних бібліотек. Способи конструювання таких кДНК та геномних бібліотек, як правило, відомі в даній галузі техніки та розкриті у Sambrook and Russell, 2001, вище. Так звані гібридизаційні зонди можуть являти собою фрагменти геномної ДНК, фрагменти кДНК, фрагменти РНК або інші олігонуклеотиди, та їх можна мітити детектовною групою, такою як 32p, або будь-яким іншим детектовним маркером, таким як інші радіоізотопи, флуоресцентна сполука, фермент або кофактор ферменту. Зонди для гібридизації можна одержувати шляхом мічення синтетичних олігонуклеотидів, основаних на відомій нуклеотидній послідовності, що кодує пестицидний білок, розкритий у даному документі. Додатково можна застосовувати вироджені праймери, що розроблені на основі консервативних нуклеотидів або амінокислотних залишків у нуклеотидній послідовності або закодованій амінокислотній послідовності. Зазвичай, зонд містить ділянку нуклеотидної послідовності, яка гібридизується за жорстких умов з щонайменше приблизно 12, щонайменше приблизно 25, щонайменше приблизно 50, 75, 100, 125, 150, 175 або 200 послідовними нуклеотидами нуклеотидної послідовності, що кодує пестицидний білок за даним винаходом або його фрагмент або варіант. Способи одержання зондів для гібридизації загалом відомі в даній галузі техніки та розкриті в Sambrook and Russell, 2001, вище у даному документі, включеному за допомогою посилання. Наприклад, усю пестицидну послідовність, розкриту у даному документі, або одну або декілька її частин, можна застосовувати як зонд, здатний до специфічної гібридизації з відповідними послідовностями, подібними до пестицидного білка, та з матричними РНК. Для досягнення специфічної гібридизації за різних умов, такі зонди включають в себе послідовності, які є унікальними і становлять переважно щонайменше приблизно 10 нуклеотидів у довжину або щонайменше приблизно 20 нуклеотидів у довжину. Такі зонди можна застосовувати для ампліфікації відповідних пестицидних послідовностей від вибраного організму за допомогою ПЛР. Цю методику можна застосовувати для виділення додаткових кодуючих послідовностей з потрібного організму або як діагностичний аналіз для визначення наявності кодуючих 6 UA 115896 C2 5 10 15 20 25 30 35 40 45 50 55 60 послідовностей в організмі. Методики гібридизації включають в себе гібридизаційний скринінг висіяних ДНК-бібліотек (або бляшок, або колоній; див., наприклад, Sambrook et al. (1989) Molecular Cloning: A Laboratory Manual (2d ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York). Таким чином, даний винахід охоплює зонди для гібридизації, а також нуклеотидні послідовності, здатні до гібридизації з усією або частиною нуклеотидної послідовності за даним винаходом (наприклад, з щонайменше приблизно 300, щонайменше приблизно 400, щонайменше приблизно 500, 1000, 1200, 1500, 2000, 2500, 3000, 3500 нуклеотидами або аж до нуклеотидної послідовності повної довжини, розкритої у даному документі). Гібридизацію таких послідовностей можна здійснювати за жорстких умов. Під виразом "жорсткі умови" або "жорсткі умови гібридизації" маються на увазі умови, за яких зонд буде гібридизуватися з своєю цільовою послідовністю з помітно більш високим ступенем, ніж з іншими послідовностями (наприклад, щонайменше у 2 рази більше за фон). Жорсткі умови залежать від послідовності і будуть відрізнятися за різних обставин. Шляхом контролю жорсткості умов гібридизації та/або відмивання, можна виявляти цільові послідовності, які на 100 % комплементарні до зонда (гомологічне зондування). Альтернативно, умови жорсткості можна регулювати для створення можливості утворення деяких помилкових спарювань у послідовності, щоб виявляти більш низькі ступені подібності (гетерологічне зондування). Як правило, зонд становить менше приблизно 1000 нуклеотидів в довжину, переважно менше 500 нуклеотидів в довжину. Зазвичай, жорсткі умови будуть такими, за яких концентрація солі становить менше приблизно 1,5 М іонів Na, зазвичай приблизно 0,01-1,0 М концентрацію іонів Na (або інших солей) з рН від 7,0 до 8,3 та температурою щонайменше приблизно 30 °C для коротких зондів (наприклад, від 10 до 50 нуклеотидів) і щонайменше приблизно 60 °C для довгих зондів (наприклад, більше 50 нуклеотидів). Жорсткі умови можуть бути також досягнуті за допомогою додавання дестабілізуючих засобів, таких як формамід. Ілюстративні умови низької жорсткості включають гібридизацію у буферному розчині, що містить 30-35 % формаміду, 1 M NaCl, 1 % SDS (додецилсульфат натрію) при 37 °C, та відмивання в 1X-2Х SSC (20X SSC=3,0 М NaCl/0,3 М тринатрійцитрат) при 50-55 °C. Ілюстративні умови помірної жорсткості включають гібридизацію у розчині, що містить 40-45 % формаміду, 1,0 М NaCl, 1 % SDS при 37 °C, та відмивання в 0,5X-1X SSC при 55-60 °C. Ілюстративні умови високої жорсткості включають гібридизацію у розчині, що містить 50 % формаміду, 1 M NaCl, 1 % SDS при 37 °C, та відмивання у 0,1X SSC при 60-65 °C. Необов'язково, буфери для відмивання можуть включати від приблизно 0,1 % до приблизно 1 % SDS. Тривалість гібридизації, як правило, становить менше приблизно 24 години, звичайно, від приблизно 4 до приблизно 12 годин. Специфічність, зазвичай, залежить від відмивання після гібридизації, при цьому критичними факторами є іонна сила та температура кінцевого розчину для відмивання. Для гібридів ДНКДНК Tm можна приблизно вирахувати за допомогою рівняння Meinkoth та Wahl (1984) Anal. Biochem. 138:267-284: Tm=81,5 °C + 16,6 (log М) + 0,41 (% GC) - 0,61 (% form) - 500 (L); де М являє собою молярність одновалентних катіонів, % GC являє собою відсоткову частку нуклеотидів гуанозину та цитозину в ДНК, % form являє собою відсоткову частку формаміду в розчині для гібридизації, та L являє собою довжину гібрида у парах основ. T m являє собою температуру (за певної іонної сили та рН), при якій 50 % комплементарної цільової послідовності гібридизується з відмінно співпадаючим зондом. Кожне зниження T m на приблизно 1 °C веде до 1 % помилкового спарювання; таким чином, T m, умови гібридизації та/або відмивання можна регулювати для гібридизації з послідовностями з потрібною ідентичністю. Наприклад, якщо шукають послідовності з ≥90 % ідентичністю, Tm може бути знижена на 10 °C. Як правило, жорсткі умови вибирають таким чином, щоб вони були приблизно на 5 °C нижчі за точку плавлення (Tm) для конкретної послідовності та комплементарної до неї послідовності за певної іонної сили та рН. Однак, за надзвичайно жорстких умов можна використовувати гібридизацію та/або відмивання за температури, що на 1, 2, 3 або 4 °C нижча за температуру точки плавлення (Tm); за помірно жорстких умов можна використовувати гібридизацію та/або відмивання за температури, що на 6, 7, 8, 9 або 10 °C нижча за температуру точки плавлення (Tm); за умов низької жорсткості можна використовувати гібридизацію та/або відмивання за температури, що на 11, 12, 13, 14, 15 або 20 °C нижча за температуру точки плавлення (T m). Із застосуванням рівняння, композиції розчинів для гібридизації та відмивання та потрібної T m, фахівцям буде зрозуміло, що варіації в умовах жорсткості розчинів для гібридизації та/або відмивання описані за своєю суттю. Якщо потрібний ступінь помилкового спарювання веде до Tm меншої за 45 °C (водний розчин) або 32 °C (розчин формаміду), переважним є збільшення концентрації SSC, аби можна було застосовувати більш високі температури. Докладний посібник з гібридизації нуклеїнових кислот наведений у (1993) Laboratory Techniques in 7 UA 115896 C2 5 10 15 20 25 30 35 40 45 50 55 60 Biochemistry and Molecular Biology — Hybridization with Nucleic Acid Probes, Part I, Chapter 2 (Elsevier, New York); та Ausubel et al., eds. (1995) Current Protocols in Molecular Biology, Chapter 2 (Greene Publishing and Wiley-Interscience, New York). Див. Sambrook et al. (1989) Molecular Cloning: A Laboratory Manual (2d ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York). Виділені білки та їхні варіанти та фрагменти Пестицидні білки також охоплені в межах даного винаходу. Під виразом "пестицидний білок" мається на увазі білок, що має амінокислотну послідовність, викладену у SEQ ID NO: 3. Також передбачені фрагменти, біологічно активні частини та їхні варіанти, які можуть застосовуватися при здійсненні способів за даним винаходом на практиці. Вираз "виділений білок" або "рекомбінантний білок" застосовується для позначення білка, який більше не перебуває у своєму природному оточенні, наприклад, знаходиться в умовах in vitro або в рекомбінантній бактерії або рослинній клітині хазяїні. Вираз "фрагменти" або "біологічно активні частини" включає в себе поліпептидні фрагменти, що містять амінокислотні послідовності, які є достатньо ідентичними з амінокислотною послідовністю, викладеною у SEQ ID NO: 3, та які проявляють пестицидну активність. Біологічно активна частина пестицидного білка може являти собою поліпептид, який становить, наприклад, 10, 25, 50, 100, 150, 200, 250, 300, 350, 400, 450, 500, 550, 600, 650, 700, 750, 800, 850, 900, 950, 1000, 1050, 1100, 1150, 1200, 1250, 1300, 1350 або більше амінокислот у довжину. Такі біологічно активні частини можна одержувати за допомогою методик рекомбінації та оцінювати щодо пестицидної активності. Способи вимірювання пестицидної активності добре відомі в даній галузі техніки. Див., наприклад, Czapla and Lang (1990) J. Econ. Entomol. 83:2480-2485; Andrews et al. (1988) Biochem. J. 252:199-206; Marrone et al. (1985) J. of Economic Entomology 78:290-293; та патент США № 5743477, усі з яких включені в даний документ за допомогою посилання у всій своїй повноті. Як застосовується у даному документі, фрагмент містить щонайменше 8 суміжних амінокислот з SEQ ID NO: 3. Проте, винахід охоплює інші фрагменти, такі як будь-який фрагмент у білку, що становить більше приблизно 10, 20, 30, 50, 100, 150, 200, 250, 300, 350, 400, 450, 500, 550, 600, 650, 700, 750 або більше амінокислот у довжину. Під виразом "варіанти" маються на увазі білки або поліпептиди, що мають амінокислотну послідовність, яка щонайменше на приблизно 60 %, 65 %, приблизно на 70 %, 75 %, приблизно 80 %, 85 %, приблизно 90 %, 91 %, 92 %, 93 %, 94 %, 95 %, 96 %, 97 %, 98 % або 99 % ідентична амінокислотній послідовності з SEQ ID NO: 3. Варіанти також включають поліпептиди, закодовані молекулою нуклеїнової кислоти, яка гібридизується з молекулою нуклеїнової кислоти з SEQ ID NO: 1-2 або комплементарної до неї послідовності за жорстких умов. Варіанти включають поліпептиди, які відрізняються за амінокислотною послідовністю внаслідок мутагенезу. Варіанти білків, охоплених даним винаходом, є біологічно активними, тобто вони продовжують характеризуватися потрібною біологічною активністю нативного білка, тобто, зберігають пестицидну активність. У деяких варіантах здійснення варіанти мають поліпшену активністю відносно нативного білка. Способи вимірювання пестицидної активності добре відомі в даній галузі техніки. Див., наприклад, Czapla and Lang (1990) J. Econ. Entomol. 83:2480-2485; Andrews et al. (1988) Biochem. J. 252:199-206; Marrone et al. (1985) J. of Economic Entomology 78:290-293; та патент США № 5743477, усі з яких включені в даний документ за допомогою посилання у всій своїй повноті. Бактеріальні гени, такі як гени axmi за даним винаходом, доволі часто мають декілька метіонінових кодонів ініціації в безпосередній близькості від старт-кодону відкритої рамки зчитування. Часто ініціація трансляції з одного або декількох цих старт-кодонів веде до утворення функціонального білка. Ці старт-кодони можуть включати ATG-кодони. Однак, бактерії, такі як Bacillus sp. також розпізнають кодон GTG як старт-кодон, і білки, трансляція яких ініціювалася з GTG-кодонів, містять метіонін на місці першої амінокислоти. У рідкісних випадках, трансляція в бактеріальних системах може ініціюватися з TTG-кодону, хоча в цьому випадку TTG кодує метіонін. Крім того, не часто визначається заздалегідь, який з цих кодонів застосовується природним чином у бактерії. Таким чином, слід розуміти, що застосування одного з альтернативних кодонів метіоніну може також вести до отримання пестицидних білків. Ці пестицидні білки охоплені даним винаходом і їх можна застосовувати в способах за даним винаходом. Буде зрозуміло, що при експресії в рослинах, необхідно змінювати альтернативний старт-кодон на ATG для правильної трансляції. У різних варіантах здійснення даного винаходу пестицидні білки включають в себе амінокислотні послідовності, виведені з нуклеотидних послідовностей повної довжини, розкритих у даному документі, та амінокислотні послідовності, які є коротшими за послідовності повної довжини внаслідок використання альтернативного стартового сайту, що розташований 8 UA 115896 C2 5 10 15 20 25 30 35 40 45 50 55 60 нижче по ланцюгу. Таким чином, нуклеотидна послідовність за даним винаходом та/або вектори, клітини-хазяїни та рослини, що містять нуклеотидну послідовність за даним винаходом (і способи одержання та застосування нуклеотидної послідовності за даним винаходом) може містити нуклеотидну послідовність, що кодує амінокислотну послідовність, яка відповідає залишкам 3-790 з SEQ ID NO: 3, залишкам 36-790 з SEQ ID NO: 3 або залишкам 38-790 з SEQ ID NO: 3. Також охоплюються антитіла до поліпептидів за даним винаходом або до їхніх варіантів або фрагментів. Способи одержання антитіл добре відомі в даній області техніки (див., наприклад, Harlow and Lane (1988) Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor, NY; патент США № 4196265). Таким чином, один аспект даного винаходу стосується антитіл, молекул, що зв'язують одноланцюговий антиген, або інших білків, які специфічно зв'язуються з однією або декількома білковими або пептидними молекулами за даним винаходом та їхніми гомологами, молекулами злиття або фрагментами. В особливо переважному варіанті здійснення антитіло специфічно зв'язується з білком, який має амінокислотну послідовність, викладену у SEQ ID NO: 3, або з його фрагментом. У іншому варіанті здійснення антитіло специфічно зв'язується зі злитим білком, що містить амінокислотну послідовність, вибрану з амінокислотної послідовності, викладеної у SEQ ID NO: 3, або його фрагментом. Антитіла за даним винаходом можна застосовувати для кількісного або якісного виявлення білкових або пептидних молекул за даним винаходом, або для виявлення посттрансляційних модифікацій білків. Як застосовується у даному документі, кажуть, що антитіло або пептид "специфічно зв'язуються" з білковою або пептидною молекулою за даним винаходом, якщо таке зв'язування не інгібується конкурентно внаслідок наявності неспоріднених молекул. Змінені або поліпшені варіанти Слід зазначити, що послідовності ДНК пестицидного білка можна змінювати різними способами, і що ці зміни можуть приводити до послідовностей ДНК, що кодують білки з амінокислотними послідовностями, які відрізняються від тих, що кодують пестицидний білок за даним винаходом. Цей білок можна змінювати різними шляхами, в тому числі за допомогою амінокислотних замін, делецій, усічення та вставок однієї або декількох амінокислот з SEQ ID NO: 3, в тому числі до приблизно 2, приблизно 3, приблизно 4, приблизно 5, приблизно 6, приблизно 7, приблизно 8, приблизно 9, приблизно 10, приблизно 15, приблизно 20, приблизно 25, приблизно 30, приблизно 35, приблизно 40, приблизно 45, приблизно 50, приблизно 55, приблизно 60, приблизно 65, приблизно 70, приблизно 75, приблизно 80, приблизно 85, приблизно 90, приблизно 100, приблизно 105, приблизно 110, приблизно 115, приблизно 120, приблизно 125, приблизно 130, приблизно 135, приблизно 140, приблизно 145, приблизно 150, приблизно 155 або більшої кількості амінокислотних замін, делецій або вставок. Способи таких маніпуляцій, у цілому, відомі в даній галузі техніки. Наприклад, варіанти амінокислотної послідовності пестицидного білка можна одержувати за допомогою здійснення мутацій в ДНК. Цього також можна досягати за допомогою однієї з декількох форм мутагенезу та/або спрямованою еволюцією. У деяких аспектах, зміни, закодовані в амінокислотної послідовності, не будуть суттєвого впливати на функцію білка. Такі варіанти будуть характеризуватися потрібною пестицидною активністю. Однак, зрозуміло, що здатність пестицидного білка забезпечувати пестицидну активність, можна поліпшувати за допомогою застосування таких методик щодо композицій за даним винаходом. Наприклад, можна експресувати пестицидний білок в клітинах-хазяїнах, які проявляють високі рівні помилкового вбудовування основ під час реплікації ДНК, таких як XL-1 Red (Stratagene, Ла-Хойя, Каліфорнія). Після вирощування у таких штамах можна виділяти ДНК (наприклад, шляхом одержання плазмідної ДНК, або шляхом ампліфікації за допомогою ПЛР і клонування одержаного ПЛР-фрагмента в вектор), культивувати пестицидний білок з мутаціями в штамах, що не спричиняють мутації, та визначати мутовані гени з пестицидною активністю, наприклад, шляхом виконання аналізу для тестування пестицидної активності. Як правило, білок змішують і застосовують в аналізах живлення. Див., наприклад, Marrone et al. (1985) J. of Economic Entomology 78:290-293. Такі аналізи можуть включати приведення рослини у контакт з одним або декількома шкідниками та визначення здатності рослин виживати та/або спричиняти загибель шкідників. Приклади мутацій, які ведуть до підвищеної токсичності, розглянуті в Schnepf et al. (1998) Microbiol. Mol. Biol. Rev. 62:775-806. Альтернативно, зміни можна робити у білковій послідовності багатьох білків на аміно- або карбокси-кінцях без здійснення суттєвого впливу на активність. Такі зміни можуть включати вставки, делеції або зміни, які введені за допомогою сучасних способів молекулярної біології, таких як ПЛР, в тому числі ПЛР-ампліфікації, які змінюють або розширюють білок-кодуючу 9 UA 115896 C2 5 10 15 20 25 30 35 40 45 50 55 60 послідовність шляхом включення послідовностей, що кодують амінокислоту, в олігонуклеотиди, які використовуються у ПЛР-ампліфікації. Альтернативно, додані білкові послідовності можуть включати в себе цілі білок-кодуючі послідовності, які звичайно застосовуються у даній галузі техніки для одержання злитих білків. Такі злиті білки часто застосовують для (1) збільшення експресії білка, що становить інтерес, (2) введення домену зв'язування, ферментативної активності або епітопу для полегшення або очищення білка, виявлення білка, або інших експериментальних застосувань, відомих у даній галузі техніки, (3) цілеспрямованої секреції або трансляції білка в субклітинній органелі, такій як периплазматичний простір грамнегативних бактерій або ендоплазматичний ретикулум еукаріотичних клітин, останнє з яких часто веде до глікозилювання білка. Варіанти нуклеотидних та амінокислотних послідовностей за даним винаходом також охоплюють послідовності, одержані за допомогою процедур мутагенезу та рекомбінації, таких як ДНК-шафлінг. За допомогою такої процедури одну або декілька різних ділянок, що кодують пестицидний білок, можна застосовувати для створення нового пестицидного білка, що характеризується потрібними властивостями. Таким чином, одержують бібліотеки рекомбінантних полінуклеотидів з популяції полінуклеотів зі спорідненими послідовностями, які містять ділянки послідовності, які характеризуються суттєвою ідентичністю послідовності та можуть гомологічно рекомбінуватися in vitro або in vivo. Наприклад, із застосуванням цього підходу мотиви послідовності, які кодують домен, що становить інтерес, можна піддавати шафлінгу між пестицидним геном за даним винаходом та іншими відомими пестицидними генами з одержанням нового гена, що кодує білок з покращеною властивістю, що становить інтерес, наприклад, такою як підвищена інсектицидна активність. Стратегії такого ДНК-шафлінгу відомі в даній галузі техніки. Див., наприклад, Stemmer (1994) Proc. Natl. Acad. Sci. USA 91:10747-10751; Stemmer (1994) Nature 370:389-391; Crameri et al. (1997) Nature Biotech. 15:436438; Moore et al. (1997) J. Mol. Biol. 272:336-347; Zhang et al. (1997) Proc. Natl. Acad. Sci. USA 94:4504-4509; Crameri et al. (1998) Nature 391:288-291; та патенти США №№ 5605793 та 5837458. Заміна або шафлінг доменів являє собою інший механізм для одержання змінених пестицидних білків. Домени можна заміняти між пестицидними білками, що веде до гібридних або химерних токсинів з підвищеною пестицидною активністю або спектром мішеней. Способи одержання рекомбінантних білків та їхнього тестування щодо пестицидної активності добре відомі в даній галузі техніки (див., наприклад, Naimov et al. (2001) Appl. Environ. Microbiol. 67:5328-5330; de Maagd et al. (1996) Appl. Environ. Microbiol. 62:1537-1543; Ge et al. (1991) J. Biol. Chem. 266:17954-17958; Schnepf et al. (1990) J. Biol. Chem. 265:20923-20930; Rang et al. 91999) Appl. Environ. Microbiol. 65:2918-2925). Вектори Пестицидна послідовність за даним винаходом може передбачатися у касеті експресії для експресії в рослині, що становить інтерес. Під виразом "рослинна касета експресії" мається на увазі ДНК-конструкція, яка здатна приводити до експресії білка з відкритої рамки зчитування в рослинній клітині. Зазвичай вона містить промотор та кодуючу послідовність. Часто такі конструкції також будуть містити 3'-нетрансльовану ділянку. Такі конструкції можуть містити "сигнальну послідовність" або "лідерну послідовність" для полегшення котрансляційного або посттрансляційного транспорту пептиду до деяких внутрішньоклітинних структур, таких як хлоропласт (або інша пластида), ендоплазматичний ретикулум або комплекс Ґольджі. Під виразом "сигнальна послідовність" мається на увазі послідовність, що, як відомо або передбачається, веде до котрансляціонного або посттрансляційного транспорту пептиду через клітинну мембрану. У еукаріотів це, звичайно, передбачає секрецію у комплекс Ґольджі, де відбувається деякий ступінь глікозилювання. Інсектицидні токсини бактерій часто синтезуються як протоксини, які протеолітично активуються у кишці цільового шкідника (Chang (1987) Methods Enzymol. 153:507-516). У деяких варіантах здійснення даного винаходу сигнальна послідовність розташована у нативній послідовності або її можна одержувати з послідовності за даним винаходом. Під виразом "лідерна послідовність" мається на увазі будь-яка послідовність, яка при трансляції дає амінокислотну послідовність, достатню для запуску котрансляційного транспорту пептидного ланцюга до субклітинної органели. Таким чином, це передбачає лідерні послідовності, які спрямовують транспорт та/або глікозилювання шляхом переходу в ендоплазматичний ретикулум, переходу у вакуолі, пластиди, в тому числі хлоропласти, мітохондрії тощо. Під виразом "вектор трансформації рослини" мається на увазі молекула ДНК, яка необхідна для ефективної трансформації рослинної клітини. Така молекула може складатися з однієї або декількох рослинних касет експресії, та вона може бути організована в дві або більше молекул 10 UA 115896 C2 5 10 15 20 25 30 35 40 45 50 55 60 ДНК-"вектора". Наприклад, бінарні вектори являють собою вектори трансформації рослин, в яких використовують два несуміжні ДНК-вектори для кодування усіх необхідних функцій, що діють у цис- та транс-положенні, для трансформації рослинних клітин (Hellens and Mullineaux (2000) Trends in Plant Science 5:446-451). Вираз "вектор" стосується конструкції нуклеїнової кислоти, розробленої для перенесення між різними клітинами-хазяїнами. Вираз "вектор експресії" стосується вектору, який має здатність вбудовувати, інтегрувати та експресувати гетерологічні послідовності або фрагменти ДНК у чужорідній клітині. Касета буде включати 5'та/або 3'-регуляторні послідовності, функціонально пов'язані з послідовністю за даним винаходом. Під виразом "функціонально пов'язаний" мається на увазі функціональний зв'язок між промотором та другою послідовністю, де промоторна послідовність ініціює та опосередковує транскрипцію послідовності ДНК, що відповідає другій послідовності. Як правило, функціонально пов'язаний означає, що зв'язані послідовності нуклеїнової кислоти є суміжними та, якщо необхідно поєднати дві білок-кодуючі ділянки, суміжні та містяться в одній і тій самій рамці зчитування. Касета може додатково містити щонайменше один додатковий ген, який має бути котрансформованим у організм. Альтернативно, додатковий ген(и) можуть знаходитись на декількох касетах експресії. У різних варіантах здійснення нуклеотидна послідовність за даним винаходом функціонально пов'язана з промотором, наприклад, рослинним промотором. Вираз "промотор" стосується послідовності нуклеїнової кислоти, яка функціонує для керування транскрипцією кодуючої послідовності, що розташована нижче по ланцюгу. Промотор разом з іншими послідовностями нуклеїнових кислот, що регулюють транскрипцію та трансляцію (що також називають "контролюючі послідовності"), необхідні для експресії послідовності ДНК, що становить інтерес. Таку касету експресії забезпечують великою кількістю сайтів рестрикції для вставки пестицидної послідовності, яка має перебувати під транскрипційним контролем регуляторних ділянок. Касета експресії буде включати в себе, у 5'-3' напрямку транскрипції, ділянку ініціації транскрипції та трансляції (тобто промотор), послідовність ДНК за даним винаходом та ділянку термінації трансляції та транскрипції (тобто ділянку термінації), які функціонують в рослинах. Промотор може бути нативним або аналогічним, чужорідним або гетерологічним щодо рослинихазяїна та/або щодо послідовності ДНК за даним винаходом. Крім того, промотор може являти собою послідовність природного походження або, альтернативно, синтетичну послідовність. Якщо промотор є "нативним" або "гомологічним" щодо рослини-хазяїна, мається на увазі, що цей промотор міститься у нативній рослині, у яку вводять цей промотор. Якщо промотор є "чужорідним" або "гетерологічним" щодо послідовності ДНК за даним винаходом, мається на увазі, що промотор не є нативним або природним промотором для функціонально пов'язаної послідовності ДНК за даним винаходом. Ділянка термінації може бути нативною з ділянкою ініціації транскрипції, може бути нативною з функціонально пов'язаною послідовністю ДНК, що становить інтерес, може бути нативною з рослиною-хазяїном або може бути одержана з іншого джерела (наприклад, бути чужорідною або гетерологічною щодо промотора, послідовності ДНК, що становить інтерес, рослини-хазяїна або будь-якого їх комбінації). Придатні ділянки термінації доступні з Ti-плазміди A. tumefaciens, такі як ділянки термінації октопінсінтази та нопалінсинтази. Див. також Guerineau et al. (1991) Mol. Gen. Genet. 262:141-144; Proudfoot (1991) Cell 64:671-674; Sanfacon et al. (1991) Genes Dev. 5:141-149; Mogen et al. (1990) Plant Cell 2:1261-1272; Munroe et al. (1990) Gene 91:151-158; Balias et al. (1989) Nucleic Acids Res. 17:7891-7903; та Joshi et al. (1987) Nucleic Acid Res. 15:9627-9639. У разі необхідності, ген(и) можна оптимізувати для підвищеної експресії в трансформованій клітині-хазяїні. Тобто, для підвищеної експресії гени можна синтезувати із застосуванням кодонів, переважних для клітини-хазяїна, або можна синтезувати із застосуванням кодонів з частотою застосування кодонів, яка є переважною у хазяїні. Як правило, вміст GC у гені буде збільшено. Див., наприклад, Campbell and Gowri (1990) Plant Physiol. 92: 1-11 для обговорення застосування кодонів, яке є переважним у хазяїні. З рівня техніки доступні способи синтезу генів, переважних для рослин. Див., наприклад, патенти США №№ 5380831 та 5436391, публікацію заявки на патент США № 20090137409 та Murray et al. (1989) Nucleic Acids Res. 17: 477-498, включені в даний документ за допомогою посилання. В одному варіанті здійснення пестицидний білок спрямовується до хлоропласту для експресії. Таким чином, якщо пестицидний білок не вводять безпосередньо в хлоропласт, касета експресії буде додатково містити нуклеїнову кислоту, що кодує транзитний пептид для спрямовування пестицидного білка до хлоропластів. Такі транзитні білки відомі в даній галузі 11 UA 115896 C2 5 10 15 20 25 30 35 40 45 50 55 60 техніки. Див., наприклад, Von Heijne et al. (1991) Plant Mol. Biol. Rep. 9:104-126; Clark et al. (1989) J. Biol. Chem. 264:17544-17550; Della-Cioppa et al. (1987) Plant Physiol. 84:965-968; Romer et al. (1993) Biochem. Biophys. Res. Commun. 196:1414-1421; та Shah et al. (1986) Science 233:478-481. Пестицидний ген, який підлягає спрямуванню до хлоропласту, може бути оптимізований для експресії у хлоропласті з урахуванням різниці застосування кодонів між ядром і цієї органелою рослини. Таким чином, нуклеїнові кислоти, що становлять інтерес, можна синтезувати із застосуванням кодонів, переважних для хлоропластів. Див., наприклад, патент США № 5380831, включений у даний документ за допомогою посилання Трансформація рослини Способи за даним винаходом передбачають введення нуклеотидної конструкції в рослину. Під виразом "введення" мається на увазі презентування рослині нуклеотидної конструкції таким чином, аби ця конструкція проникла всередину клітини рослини. Способи за даним винаходом не вимагають застосування конкретного способу введення нуклеотидної конструкції в рослину, а вимагають лише, щоб нуклеотидна конструкція проникла всередину щонайменше однієї клітини рослини. Способи введення нуклеотидних конструкцій в рослини є відомими в даній галузі техніки, в тому числі без обмежень, способи стабільної трансформації, способи транзієнтної трансформації і способи трансформації, опосередкованої вірусом. Під виразом "рослина" маються на увазі цілі рослини, органи рослин (наприклад, листя, стебла, коріння тощо), насіння, рослинні клітини, матеріал для вегетативного розмноження, зародки та їхні нащадки. Рослинні клітини можуть бути диференційованими або недиференційованими (наприклад, калюс, клітини суспензійної культури, протопласти, клітини листя, клітини кореня, клітини флоеми, пилок). Вираз "трансгенні рослини", або "трансформовані рослини", або "стабільно трансформовані" рослини, або клітини, або тканини стосується рослин, у яких у рослинну клітину вбудовані або інтегровані екзогенні послідовності нуклеїнових кислот або фрагменти ДНК. Ці послідовності нуклеїнових кислот включають в себе такі, які є екзогенними або не присутні у нетрансформованій рослинній клітині, а також такі, які можуть бути ендогенними або присутні у нетрансформованій рослинній клітині. Вираз "гетерологічний", як правило, стосується послідовностей нуклеїнових кислот, які не є ендогенними щодо клітини або частини нативного геному, у якому вони присутні, а були додані до клітини шляхом інфекції, трансфекції, мікроін'єкції, електропорації, бомбардування мікрочастинками тощо. Трансгенні рослини за даним винаходом експресують один або декілька нових послідовностей токсину, розкритих у даному документі. У різних варіантах здійснення трансгенна рослина, крім того, містить один або декілька додаткових генів, що забезпечують стійкість до комах (наприклад, Cry1, такий як представники сімейств Cry1A, Cry 1B, Cry1C, Cry1D, Cry1E та Cry1F; Cry2, такий як представник сімейства Cry2A; Cry9, такий як представники сімейства Cry9A, Cry9B, Cry9C, Cry9D, Cry9E та Cry9F; тощо). Фахівцеві у даній галузі буде зрозуміло, що трансгенна рослина може містити будь-який ген, що забезпечує агрономічну ознаку, що становить інтерес. Трансформації рослинних клітин можна досягати за допомогою однієї з декількох методик, відомих в даній галузі техніки. Пестицидний ген за даним винаходом можна модифікувати для отримання або посилення експресії в рослинних клітинах. Як правило, конструкція, яка експресує такий білок, буде містити промотор для керування транскрипцією гена, а також 3' нетрансльовану ділянку для створення можливості термінації транскрипції та поліаденилювання. Будова таких конструкцій добре відома в даній галузі техніки. У деяких випадках може бути корисною розробка гена таким чином, що одержаний пептид секретується або іншим чином спрямовується всередині рослинної клітини. Наприклад, ген можна розроблений таким чином, щоб він містив сигнальний білок для полегшення переносу пептиду в ендоплазматичний ретикулум. Також, переважною може бути розробка рослинної касети експресії, що містить інтрон, так що для експресії потрібен процесинг мРНК з видаленням інтрону. Як правило, ця "рослинна касета експресії" буде введена до "вектору трансформації рослини". Цей вектор трансформації рослини може складатися з одного або декількох векторів ДНК, необхідних для досягнення трансформації рослини. Наприклад, використання векторів трансформації рослини, які складаються з більш ніж одного суміжного сегмента ДНК, є звичайною практикою в даній галузі техніки. Ці вектори часто називають в даній галузі техніки "бінарними векторами". Бінарні вектори, а також вектори з плазмідами-помічниками найчастіше застосовуються для трансформації, опосередкованої Agrobacterium, де розмір і складність сегментів ДНК, необхідних для досягнення ефективної трансформації, досить великі та переважним є розділення функціональних елементів на окремих молекулах ДНК. Бінарні 12 UA 115896 C2 5 10 15 20 25 30 35 40 45 50 55 60 вектори, зазвичай, містять плазмідний вектор, що містить послідовності, що діють у цисположенні, які потрібні для перенесення Т-ДНК (такі як ліва гранична та права гранична послідовності), селектовний маркер, який розроблений таким чином, щоб бути здатним до експресії в рослинній клітині, та "ген, що становить інтерес" (ген, розроблений таким чином, щоб бути здатним до експресії в рослинній клітині, для якої потрібно одержання трансгенних рослин). Також цьому плазмідному векторі присутні послідовності, які потрібні для реплікації у бактеріях. Послідовності, що діють у цис-положенні, розташовані в певному порядку, щоб створювати можливість ефективного переносу в рослинні клітини та експресії у них. Наприклад, ген селектовного маркера та пестицидний ген розташовані між лівою та правою гранічними послідовностями. Часто другий плазмідний вектор містить фактори, що діють у трансположенні, які опосередковують перенос Т-ДНК з Agrobacterium до рослинних клітин. Ця плазміда часто містить функціональні елементи, що визначають вірулентність (гени Vir), які створюють можливість для Agrobacterium інфікувати рослинні клітини та переносити ДНК шляхом розщеплення гранічних послідовностей та перенесення ДНК, опосередкованого vir, які відомі в даній галузі техніки (Hellens and Mullineaux (2000) Trends in Plant Science 5:446-451). Для трансформації рослин можна застосовувати декілька типів штамів Agrobacterium (наприклад LBA4404, GV3101, EHA101, EHA105 тощо). Другий плазмідний вектор не є необхідним у випадку трансформації рослин за допомогою інших способів, таких як бомбардування мікрочастинками, мікроін'єкція, електропорація, застосування поліетиленгліколю тощо. У цілому, способи трансформації рослин передбачають перенесення гетерологичної ДНК в цільові рослинні клітини (наприклад, незрілі або зрілі зародки, суспензійні культури, недиференційований калюс, протопласти тощо), з подальшим застосуванням максимального порогового рівня відповідного добору (залежно від гена селектовного маркера) для відновлення трансформованих рослинних клітин з групи нетрансформованої клітинної маси. Як правило, експлантатів переносять у свіжу порцію такого самого середовища та культивують за стандартних умов. Згодом, після вміщення на середовище для регенерації, доповнене максимальним пороговим рівнем засобу для відбору трансформовані клітини диференціюються в пагони. Потім пагони переносять до селективного середовища вкорінення для утворення пагона з коренями або паростка. Потім трансгенний паросток виростає у зрілу рослину та продукує плідне насіння (наприклад, Hiei et al. (1994) The Plant Journal 6:271-282; Ishida et al. (1996) Nature Biotechnology 14:745-750). Як правило, експлантатів переносять у свіжу порцію такого самого середовища та культивують за стандартних умов. Загальний опис методик та способів одержання трансгенних рослин знаходиться в Ayres and Park (1994) Critical Reviews in Plant Science 13:219-239, та Bommineni and Jauhar (1997) Maydica 42:107-120. Оскільки трансформований матеріал містить багато клітин, як трансформовані, так і нетрансформовані клітини присутні в будь-якій частці підданого обробці цільового калюсу, або тканини, або групи клітин. Внаслідок можливості забезпечення загибелі нетрансформованих клітин та проліферації трансформованих клітин одержують культури трансформовані рослин. Часто, можливість видалення нетрансформованих клітин являє собою обмеження для швидкого відновлення трансформованих рослинних клітин та успішного одержання трансгенних рослин. Протоколи трансформації, а також протоколи для введення нуклеотидних послідовностей у рослини можуть змінюватися залежно від типу рослини або рослинної клітини, тобто однодольних або дводольних рослин, на які спрямована трансформація. Створення трансгенних рослин можна виконувати за допомогою одного з декількох способів, в тому числі без обмежень мікроін'єкції, електропорації, прямого перенесення генів, введення гетерологичної ДНК в рослинні клітини за допомогою Agrobacterium (трансформація, опосередкована Agrobacterium), бомбардування рослинних клітин гетерологичною чужорідною ДНК, що приєднана до частинок, балістичного прискорення частинок, трансформації із застосуванням аерозольного пучкового інжектора (опублікована заявка на патент США № 20010026941; патент США № 4945050; публікація міжнародної заявки на патент № WO 91/00915; опублікована заявка на патент США № 2002015066), Lec1-трансформація, а також різні інші способи прямого перенесення ДНК без застосуваннячастинок. Способи трансформації хлоропластів добре відомі в даній галузі техніки. Див., наприклад, Svab et al. (1990) Proc. Natl. Acad. Sci. USA 87:8526-8530; Svab and Maliga (1993) Proc. Natl. Acad. Sci. USA 90:913-917; Svab and Maliga (1993) EMBO J. 12:601-606. Спосіб залежить від доставки ДНК, що містить селектовний маркер, з використанням генної гармати та спрямування ДНК в геном пластиди шляхом гомологічної рекомбінації. Крім того, трансформації пластиди можна досягати шляхом трансактивації сплячого трансгена, що міститься у пластиді, завдяки експресії, переважно у певній тканині, РНК-полімерази, яка кодується ядерною ДНК та 13 UA 115896 C2 5 10 15 20 25 30 35 40 45 50 55 60 спрямовується до пластиди. Про таку систему повідомляли McBride et al. (1994) Proc. Natl. Acad. Sci. USA 91:7301-7305. Після інтеграції гетерологічної чужорідної ДНК в рослинні клітині у середовищі застосовують максимальний пороговий рівень відповідного засобу для відбору, аби забезпечити загибель нетрансформованих клітин, а також відокремлення та розмноження імовірно трансформованих клітин, які виживають після цієї обробки засобом для відбору, за допомогою регулярного перенесення до свіжого середовища. Шляхом безперервного пересівання та стимулювання відповідним засобом для добору виявляють та розмножують клітини, що трансформовані плазмідним вектором. Потім для підтвердження наявності інтегрованого гетерологічного гена, що становить інтерес, в геномі трансгенної рослини можна застосовувати молекулярні та біохімічні методи. Клітини, які були трансформовані, можна вирощувати у рослини згідно з традиційними методами. Див., наприклад, McCormick et al. (1986) Plant Cell Reports 5:81-84. Потім ці рослини можна вирощувати та запилювати за допомогою або тієї самої трансформованої лінії, або інших ліній, та одержаний гібрид матиме конститутивну експресію визначеної потрібної фенотипічної ознаки. Щоб переконатися, що експресія потрібної фенотипічної ознаки стабільно підтримується та успадковується, можна вирощувати два або більше поколінь, після чого збирають насіння, щоб переконатися у тому, що експресія потрібної фенотипічної ознаки була досягнута. Таким чином, у даному винаході передбачене трансформоване насіння (яке також називають "трансгенне насіння"), що має нуклеотидну конструкцію за даним винаходом, наприклад, касету експресії за даним винаходом, яка стабільно вбудована в його геном. Оцінка трансформації рослини Після введення гетерологичної чужорідної ДНК в рослинні клітини трансформацію або інтеграцію гетерологічного гена в геномі рослини підтверджують за допомогою різних способів, таких як аналіз нуклеїнових кислот, білків та метаболітів, пов'язаних з інтегрованим геном. ПЛР аналіз являє собою швидкий спосіб скринінгу трансформованих клітин, тканин або пагонів на наявність вбудованого гена на ранній стадії, до пересадки у ґрунт (Sambrook and Russell (2001) Molecular Cloning: A Laboratory Manual. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY). ПЛР проводять із застосуванням олігонуклеотидних праймерів, специфічних щодо гена, що становить інтерес, або векторної основи Agrobacterium тощо. Трансформацію рослини можна підтверджувати за допомогою саузерн-блот аналізу геномної ДНК (Sambrook and Russell, 2001, вище). У цілому, загальну ДНК екстрагують з трансформанта, розщеплюють за допомогою відповідних рестрикційних ферментів, фракціонують в агарозному гелі та переносять на нітроцелюлозну або найлонову мембрану. Потім мембрану або "блот" зондують, наприклад, за допомогою цільового фрагмента ДНК з радіоактивною міткою Р, щоб підтвердити інтеграцію введеного гена в геном рослини згідно зі стандартними методиками (Sambrook and Russell, 2001, вище). У нозерн-блот аналізі РНК виділяють з конкретних тканин трансформанта, фракціонують у формальдегід-агарозному гелі та переносять на найлоновий фільтр згідно зі стандартними процедурами, які зазвичай застосовуються в даній галузі техніки (Sambrook and Russell, 2001, вище). Потім перевіряють експресію РНК, закодованої пестицидним геном, шляхом гібридизації фільтра з радіоактивним зондом, отриманим з пестицидного гена, за допомогою способів, відомих в даній галузі техніки (Sambrook and Russell, 2001, вище). Вестерн-блот, біохімічні аналізи та подібне можна виконувати на трансгенних рослинах, щоб підтвердити наявність білка, що кодується пестицидним геном, за допомогою стандартних процедур (Sambrook and Russell, 2001, вище) із застосуванням антитіл, які зв'язуються з одним або декількома епітопами, присутніми на пестицидному білку. Пестицидна активність в рослинах В іншому аспекті даного винаходу можна одержати трансгенні рослини, що експресують пестицидний білок, який характеризується пестицидною активністю. Способи, описані вище як приклад, можна використовувати для одержання трансгенних рослин, але спосіб, в який одержують трансгенні рослинні клітини, не має вирішального значення для цього винаходу. За рішенням експериментатора можуть застосовуватися відомі або описані в даній галузі техніки способи, такі як трансформація, опосередкована Agrobacterium, біолістична трансформація та способи, не опосередковані застосуванням частинок. Рослини, які експресують пестицидний білок, можна виділяти за допомогою звичайних способів, описаних в даній галузі техніки, наприклад, шляхом трансформації калюсу, відбору трансформованого калюсу та регенерації плідних рослин з такого трансгенного калюсу. У такому процесі будь-який ген можна застосовувати як селектовний маркер, за умови що його експресія в рослинних клітинах надає можливість визначити або відібрати трансформовані клітини. 14 UA 115896 C2 5 10 15 20 25 30 35 40 45 50 55 60 Була розроблена низка маркерів для застосування в рослинних клітинах, таких, які надають стійкість до хлорамфеніколу, аміноглікозиду G418, гігроміціну тощо. Інші гени, що кодують продукт, який бере участь у метаболізмі хлоропластів, також можна застосовувати як селектовні маркери. Наприклад, гени, що забезпечують стійкість до рослинних гербіцидів, таких як гліфосат, бромоксиніл або імідазолінон, можуть бути особливо застосовними. По такі гени повідомляли (Stalker et al. (1985) J. Biol. Chem. 263: 6310-6314 (ген нітрилази, що забезпечує стійкість до бромоксинілу); та Sathasivan et al. (1990) Nucl. Acids Res. 18: 2188 (ген AHAS, що забезпечує стійкість до імідазолінону). Крім того, гени, розкриті в даному документі, застосовні як маркери для оцінки трансформації бактерій або рослинних клітин. Способи виявлення наявності трансгена у рослині, органі рослини (наприклад, листі, стеблах, коренях тощо), насінні, рослинній клітині, матеріалі для вегетативного розмноження, зародку або його нащадках, добре відомі в даній галузі техніки. У одному варіанті здійснення наявність трансгена виявляють шляхом тестування щодо пестицидної активності. Плідні рослини, що експресують пестицидний білок, можна тестувати щодо пестицидної активності, та рослин, що демонструють оптимальну активність, відбирають для подальшого розведення. В даній галузі техніки є доступними способи аналізу активності проти шкідника. Як правило, білок змішують і застосовують в аналізах живлення. Див., наприклад, Marrone et al. (1985) J. of Economic Entomology 78:290-293. Даний винахід можна застосовувати для трансформації будь-якого виду рослин, в тому числі без обмежень однодольних та дводольних рослин. Приклади рослин, що становлять інтерес, включають без обмежень кукурудзу (маїс), сорго, пшеницю, соняшник, томат, хрестоцвіті, різновиди перцю, картоплю, бавовник, рис, сою, цукровий буряк, цукрову тростину, тютюн, ячмінь та масличний ріпак, Brassica sp., люцерну, жито, просо, сафлор, різновиди арахісу, солодку картоплю, маніоку, каву, кокос, ананас, цитрусові дерева, какао, чай, банани, авокадо, інжир, гуаву, манго, маслину, папайю, кеш'ю, макадамію, мигдаль, овес, овочі, декоративні рослини та хвойні. Овочеві культури включають без обмежень різновиди томату, салат, зелені боби, лімські боби, різновиди гороху та представників роду Curcumis, таких як огірок, канталупа та мускусна диня. Декоративні культури включають без обмежень азалію, гортензію, гібіскус, троянди, різновиди тюльпанів, нарцисів, петунії, гвоздику, пуансетію та хризантеми. Переважно рослини за даним винаходом являють собою сільськогосподарські культури (наприклад, маїс, сорго, пшениця, соняшник, томат, хрестоцвіті, різновиди перцю, картопля, бавовник, рис, соя, цукровий буряк, цукрова тростина, тютюн, масличний ріпак тощо). Застосування в пестицидному контролі В даній галузі техніки є відомими загальні способи застосування ліній, що містять нуклеотидну послідовність за даним винаходом або її варіант, у контролі шкідників або при розробці інших організмів як пестицидні засоби. Див., наприклад, патент США № 5039523 та патентний документ EP № 0480762A2. Штами Bacillus, які містять нуклеотидну послідовність за даним винаходом або її варіант, або мікроорганізми, які були генетично змінені, щоб містити пестицидний ген за даним винаходом та білок, можна застосовувати для захисту сільськогосподарських культур та продуктів від шкідників. В одному аспекті даного винаходу цілі, тобто не піддані лізису, клітини організму, що виробляє токсин (пестицид), обробляють реагентами, які подовжують активність токсину, що виробляється у клітині, коли клітину вносять в середовище цільового шкідника(ів). Альтернативно, пестицид виробляється при введенні пестицидного гена в клітину-хазяїна. Експресія пестицидного гена веде, безпосередньо або опосередковано, до внутрішньоклітинної продукції та зберігання пестициду. В одному аспекті даного винаходу потім ці клітини обробляють в таких умовах, які подовжують активність токсину, що виробляється у клітині, коли клітину вносять в середовище цільового шкідника(ів). Одержаний продукт зберігає токсичність токсину. Ці інкапсульовані природнім шляхом пестициди потім можна складати згідно з традиційними методиками внесення в середовище, де мешкає цільовий шкідник, наприклад, в ґрунт, воду та на листя рослин. Див., наприклад патентний документ EPA 0192319 та процитовані у ньому посилання. Альтернативно, можна складати клітини, які експресують ген за даним винаходом таким чином, щоб створити можливість застосування одержаного матеріалу у вигляді пестициду. Активні інгредієнти за даним винаходом, як правило, застосовують у вигляді композицій, і їх можна вносити у місця зростання сільськогосподарської культури або на рослину, що підлягають обробці, одночасно або послідовно з іншими сполуками. Ці сполуки можуть являти собою добрива, засоби боротьби з бур'янами, кріопротектори, поверхнево-активні речовини, мийні засоби, пестицидні мила, масла для обробки рослин у стані спокою, полімери та/або 15 UA 115896 C2 5 10 15 20 25 30 35 40 45 50 55 склади з повільним вивільненням або носіями, що піддаються біорозкладанню, які роблять можливим довгострокову дозовану обробку цільової області після одноразового внесення складу. Вони також можуть являти собою селективні гербіциди, хімічні інсектициди, віруліциди, мікробоциди, амебоциди, пестициди, фунгіциди, бактерициди, нематоциди, моллюскоциди або суміші декількох з цих препаратів, за необхідності, разом з додатковими прийнятними для сільського господарства носіями, поверхнево-активними речовинами або допоміжними речовинами, що сприяють внесенню, які, зазвичай, використовуються в галузі техніки одержання складів. Придатні носії та допоміжні речовини можуть бути твердими або рідкими та являють собою речовини, що звичайно використовуються в технології одержання складів, наприклад, природні або регенеровані мінеральні речовини, розчинники, диспергувальні засоби, змочувальні засоби, засоби, що надають клейкість, зв'язувальні речовини або добрива. Подібним чином, склади можна готувати як їстівні "приманки" або надавати форму "пасток" для шкідника, щоб зробити можливим живлення або прийом всередину пестицидного складу цільовим шкідником. Способи внесення активного інгредієнта за даним винаходом або агрохімічної композиції за даним винаходом, які містять щонайменше один пестицидний білок, що продукується бактеріальними штамами за даним винаходом, включають нанесення на лист, покриття насіння та внесення в ґрунт. Кількість нанесень та швидкість внесення залежать від інтенсивності зараження відповідним шкідником. Композицію можна складати у вигляді порошку, дусту, пелети, гранули, аерозолю, емульсії, колоїду, розчину або подібних до них, і її можна одержувати за допомогою таких традиційних способів, як сушіння, ліофілізація, гомогенізація, екстракція, фільтрація, центрифугування, осадження або концентрація з культури клітин, що містять поліпептид. У всіх таких композиціях, які містять щонайменше один пестицидний поліпептид, такий поліпептид може бути присутнім у концентрації від приблизно 1 % до приблизно 99 % за вагою. Шкідників-твердокрилих комах можна знищувати або їхню кількість можна знижувати у визначеній ділянці за допомогою способів за даним винаходом, або ці способи можна застосовувати профілактично до ділянки навколишнього середовища для запобігання зараженню вразливим шкідником. Переважно шкідник приймає всередину або контактує з пестицидно ефективною кількістю поліпептиду. Під виразом "пестицидно ефективна кількість" мається на увазі кількість пестициду, яка здатна призвести до загибелі щонайменше одного шкідника або помітно послабляти ріст, живлення або нормальний фізіологічний розвиток шкідника. Ця кількість буде змінюватись залежно від таких факторів, як, наприклад, конкретні цільові шкідники, що підлягають контролю, специфічне середовище, розташування, рослина, сільськогосподарська культура або сільськогосподарська ділянка, що підлягають обробці, умови навколишнього середовища та спосіб, норма внесення, концентрація, стабільність та кількість внесень пестицидно ефективноїполіпептидної композиції. Склади також можуть змінюватися відповідно до кліматичних умов, міркувань, пов'язаних із забрудненням довкілля, та/або частоти внесення та/або тяжкості зараження шкідником. Описані пестицидні композиції можна отримувати за допомогою складання або бактеріальної клітини, кристалу та/або суспензії спор, або виділеного білкового компонента з потрібним прийнятним для сільського господарства носієм. Композиції можна складати перед застосуванням за допомогою відповідних засобів, як, наприклад, ліофілізовані, висушені виморожуванням, висушені, або за допомогою водного носія, середовища або придатного розріджувача, такого як сольовий розчин або інший буфер. Складені композиції можуть перебувати у формі дусту або гранульованого матеріалу, або суспензії в олії (рослинній або мінеральній), або водної емульсії або емульсії масло у воді, або у вигляді порошку, що змочується, або в комбінації з будь-яким іншим матеріалом-носієм, придатним для сільськогосподарського застосування. Придатні для сільського господарства носії можуть бути твердими або рідкими, та вони добре відомі в даній галузі техніки. Вираз "прийнятний для сільського господарства носій" охоплює усі допоміжні речовини, інертні сполуки, диспергувальні засоби, поверхнево-активні речовини, засоби, що надають клейкість, зв'язувальні речовини тощо, які зазвичай застосовують в технології складання пестицидів; при цьому вони добре відомі фахівцям у галузі складання пестицидів. Склади можна змішувати з однією або декількома твердими або рідкими допоміжними речовинами та одержувати різними способами, наприклад, шляхом гомогенного перемішування, змішування та/або подрібнення пестицидної композиції з відповідними допоміжними речовинами із застосуванням традиційних методик складання. Придатні склади і способи внесення описані у патенті США № 6468523, який включений у даний документ за допомогою посилання. 16 UA 115896 C2 5 10 15 20 25 30 35 40 45 50 55 60 Вираз "шкідник-твердокрила комаха" включає без обмежень шкідників кукурудзяних жуків, наприклад, західного кукурудзяного жука (наприклад, Diabrotica virgifera virgifera), північного кукурудзяного жука (наприклад, Diabrotica barberi) та південного кукурудзяного жука (наприклад, Diabrotica undecimpunctata howardi). Способи підвищення врожайності рослини Передбачені способи підвищення врожайності рослини. Способи включають одержання рослини або рослинної клітини, що експресують полінуклеотид, що кодує послідовність пестицидного поліпептиду, розкриту у даному документі, та вирощування рослини або її насінини у полі, зараженому (або сприйнятливого до зараження) шкідником, проти якого зазначений поліпептид характеризується пестицидною активністю. У деяких варіантах здійснення поліпептид Axmi184, описаний у даному документі, характеризується пестицидною активністю проти шкідника-твердокрилої комахи, та зазначене поле є зараженим зазначеним шкідником-твердокрилою комахою. У різних варіантах здійснення шкідником-твердокрилою комахою є шкідник-кукурудзяний жук, такий як західний кукурудзяний жук. Як визначено у даному документі, вираз "врожайність" рослини стосується якості та/або кількості біомаси, яку виробляє рослина. Під виразом "біомаса" мається на увазі будь-який вимірюваний рослинний продукт. Зростанням продукування біомаси є будь-яке поліпшення врожайності вимірюваного рослинного продукту. Зростання врожайності рослини має різні комерційні застосування. Наприклад, зростання біомаси листя рослини може збільшувати врожайність листових овочів для споживання людиною або твариною. Крім того, зростання біомаси листя можна застосовувати для зростання продукування фармацевтичних або промислових продуктів рослинного походження. Зростання врожайності може включати в себе будь-яке статистично значуще зростання, в тому числі без обмежень зростання щонайменше на 1 %, зростання щонайменше на 3 %, зростання щонайменше на 5 %, зростання щонайменше на 10 %, зростання щонайменше на 20 %, зростання щонайменше на 30 %, зростання щонайменше на 50 %, зростання щонайменше на 70 %, зростання щонайменше на 100 % або більш велике зростання врожайності порівняно з рослиною, що не експресує пестицидну послідовність. У конкретних способах, врожайність рослини зростає внаслідок підвищеної стійкості до шкідника у рослини, що експресує пестицидний білок, розкритий у даному документі. Експресія пестицидного білка веде до зниженої здатності шкідника до інфікування або живлення. Рослини також можна обробляти за допомогою однієї або декількоххімічних композицій, що включають один або декілька гербіцидів, інсектицидів або фунгіцидів. Ілюстративні хімічні композиції включають: гербіциди для захисту плодових/овочевих культур: атразин, бромацил, діурон, гліфосат, лінурон, метрибузин, симазин, трифлуралін, флуазифоп, глюфосинат, галосульфурон від Gowan, паракват, пропізамід, сетоксидим, бутафенацил, галосульфурон, індазифлам; інсектициди для захисту плодових/овочевих культур: альдикарб, Bacillus thuriengiensis, карбарил, карбофуран, хлорпірифос, циперметрин, дельтаметрин, абамектин, цифлутрин/бета-цифлутрин, есфенвалерат, лямбда-цигалотрин, ацеквіноцил, біфеназат, метоксифенозид, новалурон, хромафенозид, тіаклоприд, динотефуран, флуакрипірим, спіродиклофен, гамма-цигалотрин, спіромезифен, спіносад, ринаксипір, ціазипір, трифлумурон, спіротетрамат, імідаклоприд, флубендіамід, тіодикарб, метафлумізон, сульфоксафлор, цифлуметофен, ціанопірафен, клотіанідин, тіаметоксам, спіноторам, тіодикарб, флонікамід, метіокарб, емамектину бензоат, індоксакарб, фенаміфос, пірипроксифен, фенбутатин-оксид; фунгіциди для захисту плодових/овочевих культур: аметоктрадин, азоксистробін, бентіавалікарб, боскалід, каптан, карбендазим, хлороталоніл, мідь, ціазофамід, цифлуфенамід, цимоксаніл, ципроконазол, ципродиніл, дифеноконазол, диметоморф, дитіанон, фенамідон, фенгексамід, флуазинам, флудіоксоніл, флуопіколід, флуопірам, флуоксастробін, флуксапіроксад, фолпет, фосетил, іпродіон, іпровалікарб, ізопіразам, крезоксим-метил, манкозеб, мандипропамід, металаксил/мефеноксам, метирам, метрафенон, міклобутаніл, пенконазол, пентіопірад, пікоксистробін, пропамокарб, пропіконазол, пропінеб, проквіназид, протіоконазол, піраклостробін, піриметаніл, квіноксифен, спіроксамін, сірка, тебуконазол, тіофанат-метил, трифлоксистробін; гербіциди для захисту зернових культур: 2,4-D, амідосульфурон, бромоксиніл, карфентразон-Е, хлоротолурон, хлорсульфурон, клодинафоп-P, клопіралід, дикамба, диклофоп-М, дифлуфенікан, феноксапроп, флорасулам, флукарбазон-NA, флуфенацет, флупіросульфурон-М, флуроксипір, флуртамон, гліфосат, йодосульфурон, іоксиніл, ізопротурон, МСРА, мезосульфурон, метсульфурон, пендиметалін, піноксаден, пропоксикарбазон, просульфокарб, піроксулам, сульфосульфурон, тифенсульфурон, тралкоксидим, триасульфурон, трибенурон, трифлуралін, тритосульфурон; фунгіциди для захисту зернових культур: азоксистробін, біксафен, боскалід, карбендазим, хлороталоніл, цифлуфенамід, ципроконазол, ципродиніл, димоксистробін, епоксиконазол, фенпропідин, 17 UA 115896 C2 5 10 15 20 25 30 35 40 45 50 55 60 фенпропіморф, флуопірам, флуоксастробін, флуквінконазол, флуксапіроксад, ізопіразам, крезоксим-метил, метконазол, метрафенон, пентіопірад, пікоксистробін, прохлораз, пропіконазол, проквіназид, протіоконазол, піраклостробін, квіноксифен, спіроксамін, тебуконазол, тіофанат-метил, трифлоксистробін; інсектициди для захисту зернових культур: диметоат, лямбда-цигалотрин, дельтаметрин, альфа-циперметрин, ß-цифлутрин, біфентрин, імідаклоприд, клотіанідин, тіаметоксам, тіаклоприд, ацетаміприд, динетофуран, хлорпірифос, піримікарб, метіокарб, сульфоксафлор; гербіциди для захисту маїсу: атразин, алахлор, бромоксиніл, ацетохлор, дикамба, клопіралід, (S)-диметенамід, глуфосинат, гліфосат, ізоксафлутол, (S)-метолахлор, мезотріон, нікосульфурон, примісульфурон, римсульфурон, сулькотріон, форамсульфурон, топрамезон, темботріон, сафлуфенацил, тієнкарбазон, флуфенацет, піроксасульфон; інсектициди для захисту маїсу: карбофуран, хлорпірифос, біфентрин, фіпроніл, імідаклоприд, лямбда-цигалотрин, тефлутрин, тербуфос, тіаметоксам, клотіанідин, спіромезифен, флубендіамід, трифлумурон, ринаксипір, дельтаметрин, тіодикарб, ß-цифлутрин, циперметрин, біфентрин, люфенурон, тебупіримфос, етіпрол, ціазипір, тіаклоприд, ацетаміприд, динетофуран, авермектин; фунгіциди для захисту маїсу: азоксистробін, біксафен, боскалід, ципроконазол, димоксистробін, епоксиконазол, фенітропан, флуопірам, флуоксастробін, флуксапіроксад, ізопіразам, метконазол, пентіопірад, пікоксистробін, пропіконазол, протіоконазол, піраклостробін, тебуконазол, трифлоксистробін; гербіциди для захисту рису: бутахлор, пропаніл, азимсульфурон, бенсульфурон, цигалофоп, даімурон, фентразамід, імазосульфурон, мефенацет, оксазикломефон, піразосульфурон, пірибутикарб, квінклорак, тіобенкарб, інданофан, флуфенацет, фентразамід, галосульфурон, оксазикломефон, бензобіциклон, пірифталід, пеноксулам, біспірибак, оксадіаргіл, етоксисульфурон, претілахлор, мезотріон, тефурилтріон, оксадіазон, феноксапроп, піримісульфан; інсектициди для захисту рису: діазинон, фенобукарб, бенфуракарб, бупрофезин, динотефуран, фіпроніл, імідаклоприд, ізопрокарб, тіаклоприд, хромафенозид, клотіанідин, етипрол, флубендіамід, ринаксипір, дельтаметрин, ацетаміприд, тіаметоксам, ціазипір, спіносад, спіноторам, емамектину бензоат, циперметрин, хлорпірифос, етофенпрокс, карбофуран, бенфуракарб, сульфоксафлор; фунгіциди для захисту рису: азоксистробін, карбендазим, карпропамід, диклоцимет, дифеноконазол, едифенфос, феримзон, гентаміцин, гексаконазол, гімексазол, іпробенфос (IBP), ізопротіолан, ізотіаніл, касугаміцин, манкозеб, метоміностробін, орисастробін, пенцикурон, пробеназол, пропіконазол, пропінеб, піроквілон, тебуконазол, тіофанат-метил, тіадиніл, трициклазол, трифлоксистробін, валідаміцин; гербіциди для захисту бавовнику: діурон, флуометурон, MSMA, оксифлуорфен, прометрин, трифлуралін, карфентразон, клетодим, флуазифоп бутил, гліфосат, норфлуразон, пендиметалін, піритіобакнатрію, трифлоксисульфурон, тепралоксидим, глуфосинат, флуміоксазин, тидіазурон; інсектициди для захисту бавовнику: ацефат, альдикарб, хлорпірифос, циперметрин, дельтаметрин, абамектин, ацетаміприд, емамектину бензоат, імідаклоприд, індоксакарб, лямбда-цигалотрин, спіносад, тіодикарб, гамма-цигалотрин, спіромезифен, піридаліл, флонікамід флубендіамід, трифлумурон, ринаксипір, бета-цифлутрин, спіротетрамат, клотіанідин, тіаметоксам, тіаклоприд, динетофуран, флубендіамід, ціазипір, спіносад, спіноторам, гамма-цигалотрин, 4-[[(6-хлор-піридин-3-іл)метил(2,2-дифторетил)аміно]фуран2(5Н)-он, тіодикарб, авермектин, флонікамід, піридаліл, спіромезифен, сульфоксафлор; фунгіциди для захисту бавовнику: азоксистробін, біксафен, боскалід, карбендазим, хлороталоніл, мідь, ципроконазол, дифеноконазол, димоксистробін, епоксиконазол, фенамідон, флуазинам, флуопірам, флуоксастробін, флуксапіроксад, іпродіон, ізопіразам, ізотіаніл, манкозеб, манеб, метоміностробін, пентіопірад, пікоксистробін, пропінеб, протіоконазол, піраклостробін, квінтозен, тебуконазол, тетраконазол, тіофанат-метил, трифлоксистробін; гербіциди для захисту сої: алахлор, бентазон, трифлуралін, хлоримурон-етил, хлорансуламметил, феноксапроп, фомесафен, флуазифоп, гліфосат, імазамокс, імазахін, імазетапір (S)метолахлор, метрибузин, пендиметалін, тепралоксидим, глюфосинат; інсектициди для захисту сої: лямбда-цигалотрин, метоміл, імідаклоприд, клотіанідин, тіаметоксам, тіаклоприд, ацетаміприд, динетофуран, флубендіамід, ринаксипір, ціазипір, спіносад, спіноторам, емамектину бензоат, фіпроніл, етипрол, дельтаметрин, В-цифлутрин, гамма- та лямбдацигалотрин, 4-[[(6-хлор-піридин-3-іл)метил(2,2-дифторетил)аміно]фуран-2(5Н)-он, спіротетрамат, спінодиклофен, трифлумурон, флонікамід, тіодикарб, бета-цифлутрин; фунгіциди для захисту сої: азоксистробін, біксафен, боскалід, карбендазим, хлороталоніл, мідь, ципроконазол, дифеноконазол, димоксистробін, епоксиконазол, флуазинам, флуопірам, флуоксастробін, флутриафол, флуксапіроксад, ізопіразам, іпродіон, ізотіаніл, манкозеб, манеб, метконазол, метоміностробін, міклобутаніл, пентіопірад, пікоксистробін, пропіконазол, пропінеб, протіоконазол, піраклостробін, тебуконазол, тетраконазол, тіофанат-метил, трифлоксистробін; 18 UA 115896 C2 5 10 15 20 25 30 35 40 45 50 55 60 гербіциди для захисту цукрового буряку: хлоридазон, десмедифам, етофумезат, фенмедифам, триалат, клопіралід, флуазифоп, фенацил, метамітрон, квінмерак, циклоксидим, трифлусульфурон, тепралоксидим, квізалофоп; інсектициди для захисту цукрового буряку: імідаклоприд, клотіанідин, тіаметоксам, тіаклоприд, ацетаміприд, динетофуран, дельтаметрин, В-цифлутрин, гамма/лямбда-цигалотрин, 4-[[(6-хлорпіридин-3-іл)метил(2,2дифторетил)аміно]фуран-2(5Н)-он, тефлутрин, ринаксипір, ціаксипір, фіпроніл, карбофуран; гербіциди для захисту каноли: клопіралід, диклофоп, флуазифоп, глюфосинат, гліфосат, метазахлор, трифлуралін, етаметсульфурон, квінмерак, квізалофоп, клетодим, тепралоксидим; фунгіциди для захисту каноли: азоксистробін, біксафен, боскалід, карбендазим, ципроконазол, дифеноконазол, димоксистробін, епоксиконазол, флуазинам, флуопірам, флуоксастробін, флусилазол, флуксапіроксад, іпродіон, ізопіразам, мепікват-хлорид, метконазол, метоміностробін, паклобутразол, пентіопірад, пікоксистробін, прохлораз, протіоконазол, піраклостробін, тебуконазол, тіофанат-метил, трифлоксистробін, вінклозолін; інсектициди для захисту каноли: карбофуран, тіаклоприд, дельтаметрин, імідаклоприд, клотіанідин, тіаметоксам, ацетаміприд, динетофуран, В-цифлутрин, гамма- та лямбда-цигалотрин, тау-флувалеріат, етипрол, спіносад, спіноторам, флубендіамід, ринаксипір, ціазипір, 4-[[(6-хлорпіридин-3іл)метил(2,2-дифторетил)аміно]фуран-2(5Н)-он. Наступні приклади пропонуються як ілюстрація, а не як обмеження. Експериментальні приклади Приклад 1. Експресія та активність Axmi184 у Zea mays Ген Axmi184 (SEQ ID NO: 1) виділяли зі штаму ATX14775 Bacillus thuringiensis, як описано в публікації заявки на патент США № 20100004176. Амінокислотна послідовність AXMI184 (SEQ ID NO: 3) на 93 % ідентична послідовності Vip3Afl (№ доступу GenBank CAI43275). Було показано, що білки VIP3A є токсичними до таксономічно різноманітної групи Lepidoptera (Estruch et al. (1996) Proc Natl Acad Sci USA 93:5389-5394). Однак, не було жодних доказів, що якийнебудь ген, що має гомологію до токсинів класу VIP, є активним проти шкідників-твердокрилих комах. Трансгенні рослини маїсу, трансформовані за допомогою оптимізованої версії Axmi184 (optAxmi184, що викладена у даному документі як SEQ ID NO: 2), отримували шляхом трансформації, опосередкованої Agrobacterium. Рослини, які, як показано за ПЛР-аналізом, містили придатну конструкцію Axmi184, відбирали та переносили до контейнерів для саджанців. T0-рослини, що містять optAxmi184, пересаджували до контейнерів для саджанців і вирощували упродовж приблизно трьох тижнів. Потім кожну окрему рослину заражали ~125 яйцями західного кукурудзяного жука (Diabrotica virgifera), що не перебувають на стадії діапаузи. Рослини аналізували щодо експресію білка AXMI184 за допомогою вестерн-блот-аналізу із застосуванням антитіла до AXMI184. Рослин, що експресують AXMI184 з кількостями, що можна виявляти, відбирали для аналізу. Після п'ятнадцяти днів оцінювали показник пошкодження коріння у кожної рослини із застосуванням шкалу пошкодження вузлів (CRWNIS) (Oleson et al. 2005. J. Econ Entomol. 98(1): 1-8), де 0,00 означає відсутність пошкодження, а 3,00 означає, що усі корені підрізані в межах 1,5 дюйма від стебла на трьох повних вузлах. На фігурі 1 показано, що AXMI184 веде до зниження пошкодження коренів, порівняно з контрольними рослинами, зараженими у той самий спосіб. Приклад 2. Результати польових випробувань T1-рослини, що експресують optAxmi184, тестували у двох місцерозташуваннях у Айові. Кожна експериментальна одиниця складалася з однорядкової ділянки маїсу розміром приблизно 15 футів. Ділянки розташовували із застосуванням рандомізованого повноблокового плану з чотирма повторами. Ефективність проти західного кукурудзяного жука визначали шляхом оцінки до восьми коренів маїсу на ділянку. Інбредні лінії B110 та B116, CRW-чутливий комерційний гібрид та два стандарти для контролю CRW: B110 зі стандартним гранульованим інсектицидом (FORCE® 3G) та B110 з інсектицидною обробкою насіння (PONCHO® 1250), застосовували як контролі Поля готували для цього дослідження із застосуванням агротехнічних прийомів, подібні до тих, що застосовують на комерційних маїсових полях у Айові. Рослини, що не містять трансгенів для контролю CRW, виявляли та виключали з дослідження із застосуванням нанесення 16 унцій/акр гліфосату, коли рослини перебували на стадії VI листка. Через п'ять-десять днів фіксували кількість позитивних і негативних рослин у кожній ділянці. Потім негативні рослини вилучали, а позитивні рослини, що залишилися, проріджували до 8 рослин на рядок. Потім усі рослини заражали вручну яйцями західного кукурудзяного жука, які не знаходилися у стадії діапаузи. Яйця суспендували у розбавленому розчині агару та калібрували для доставки 750 яєць на рослину для усіх ділянок. 19 UA 115896 C2 Коріння відкопували, промивали і оцінювали на пошкодження CRW після періоду живлення личинок. Спостереження розвитку личинок на незахищеному маїсі (B110) застосовували для визначення оптимального часу для цієї оцінки. Пошкодження, спричинені кукурудзяним жуком, характеризували із застосуванням CRWNIS. Результати показані в таблиці 1. 5 Таблиця 1 Рослина B110 B110+Force B110+Poncho B116 Негативний гібрид optAxmi184 10 15 20 25 30 35 40 45 50 Кількість рослин Середній CRWNIS 58 30 31 56 32 175 1,53 0,30 0,08 1,52 0,83 0,12 Стандартна помилка 0,075 0,105 0,103 0,077 0,101 0,043 Приклад 3. Вбудовування генів у вектори для експресії у рослині Для експресії у рослинах кодуючі ділянки за даним винаходом з'єднували з придатним промоторною та термінаторною послідовностями. Такі послідовності добре відомі в даній галузі техніки та вони можуть включати актиновий промотор рису або убіквітиновий промотор маїсу для експресії в однодольних рослинах, промотор UBQ3 Arabidopsis або промотор 35S CaMV для експресії в дводольних рослинах та термінатори nos або PinII. Методики одержання та підтвердження конструкції промотор-ген-термінатор також добре відомі в даній галузі техніки. В одному аспекті даного винаходу розробляють та отримують синтетичні послідовності ДНК. Ці синтетичні послідовності мають змінену нуклеотидну послідовність відносно батьківської послідовності, але вони кодують білки, які є фактично ідентичними з батьківською послідовністю. В іншому аспекті даного винаходу модифіковані версії синтетичних генів розроблені таким чином, що одержаний білок спрямовується у рослинну органелу, таку як ендоплазматичний ретикулум або апопласт. В даній галузі техніки відомі пептидні послідовності, які, як це відомо, ведуть до спрямовування злитих білків до органел рослини. Наприклад, N-кінцева ділянка гена кислої фосфатази з білого люпину Lupinus albus (GENBANK® ID GE14276838, Miller et al. (2001) Plant Physiology 127: 594-606), як відомо в даній галузі техніки, веде до спрямовування гетерологічних білків до ендоплазматичного ретикулуму. Якщо одержаний злитий білок також містить послідовність, яка забезпечує його утримання в ендоплазматичному ретикулумі, що містить пептид N-кінець-лізин-аспарагінова кислота-глутамінова кислота-лейцин (тобто мотив "KDEL", SEQ ID NO: 4) на С-кінці, то злитий білок буде спрямовуватися до ендоплазматичного ретикулуму. Якщо злитий білок не містить на С-кінці послідовність, що спрямовує його до ендоплазматичного ретикулуму, то білок буде спрямовуватися до ендоплазматичного ретикулуму, але, в кінцевому рахунку, буде надходити до апопласту. Таким чином, цей ген кодує злитий білок, який містить тридцять одну амінокислоту з N-кінця гена кислої фосфатази з білого люпину Lupinus albus (GENBANK® ID GL14276838, Miller та ін., 2001, вище), злиті з N-кінцем амінокислотної послідовності за даним винаходом, а також послідовність KDEL (SEQ ID NO: 4) на С-кінці. Таким чином, як передбачається, одержаний білок буде спрямовуватися до ендоплазматичного ретикулуму рослини після експресії в рослинній клітині. Рослинні касети експресії, описані вище, поєднують з придатним селектовним маркером рослини для сприяння відбору трансформованих клітин і тканин, та лігують у вектори трансформації рослин. Вони можуть включати бінарні вектори для трансформації, опосередкованої Agrobacterium, або прості плазмідні вектори для аерозольної або біолістичної трансформації. Приклад 4. Трансформація генів за даним винаходом в рослинні клітини за допомогою трансформації, опосередкованої Agrobacterium Качани найкраще збирати через 8-12 днів після запилення. З качанів виділяють зародків, та такі зародки розміром 0,8-1,5 мм є переважними для застосування у трансформації. Зародків розміщують щитковою стороною догори на придатному середовищі для інкубації та інкубують протягом ночі при 25 °C в темряві. Однак, per se не є необхідним інкубувати ембріонів протягом ночі. Ембріонів приводять у контакт зі штамом Agrobacterium, який містить відповідні вектори для перенесення, опосередкованої Ti-плазмідою, протягом приблизно 5-10 хв., а потім вміщують на середовище для спільного культивування на період приблизно 3 дні (22 °C у 20 UA 115896 C2 5 10 15 темряві). Після спільного культивування експлантатів переносять у середовище періоду відновлення на 5-10 днів (при 25 °C у темряві). Експлантати інкубують в середовищі для відбору до восьми тижнів, залежно від природи та особливостей конкретного засобу для відбору, який використовують. Після періоду відбору одержаний калюс переносять до середовища дозрівання зародка до того часу, як буде спостерігатися утворення зрілих соматичних зародків. Потім одержаних зрілих соматичних зародків поміщають під слабке освітлення та ініціюють процес регенерації, як відомо в даній галузі техніки. Усі публікації та патентні заявки, згадані в даному описі, вказують на рівень кваліфікації фахівців у даній галузі техніки, до якої належить даний винахід. Усі публікації та патентні заявки включені в даний документ за допомогою посилання у тому самому об'ємі, якщо б кожна окрема публікація або патентна заявка була конкретно та індивідуально вказана як так, що включена за допомогою посилання. Хоча вищенаведений винахід було описано більш докладно з метою ілюстрації та прикладу для цілей ясності розуміння, буде очевидно, що певні зміни та модифікації можна здійснювати на практиці в рамках доданої формули винаходу. 21 UA 115896 C2 22 UA 115896 C2 23 UA 115896 C2 24 UA 115896 C2 25 UA 115896 C2 26 UA 115896 C2 ФОРМУЛА ВИНАХОДУ 5 1. Спосіб знищення або контролю популяції шкідника-кукурудзяного жука, що включає приведення зазначеної популяції у контакт з пестицидно ефективною кількістю поліпептиду, що 27 UA 115896 C2 5 10 15 20 25 містить амінокислотну послідовність, яка щонайменше на 95 % ідентична амінокислотній послідовності з SEQ ID NO: 3. 2. Спосіб захисту рослини кукурудзи від шкідника-кукурудзяного жука, що включає експресію у рослині кукурудзи або її клітині нуклеотидної послідовності, функціонально пов'язаної з промотором, здатним керувати експресією нуклеотидної послідовності у рослинній клітині, де зазначена нуклеотидна послідовність вибрана з групи, що складається з: a) нуклеотидної послідовності, викладеної у SEQ ID NO: 1 або 2; та b) нуклеотидної послідовності, яка кодує поліпептид, що містить амінокислотну послідовність, яка щонайменше на 95 % ідентична амінокислотній послідовності з SEQ ID NO: 3, де зазначений поліпептид характеризується пестицидною активністю проти шкідникакукурудзяного жука. 3. Спосіб підвищення врожайності рослини кукурудзи, що включає вирощування в полі рослини кукурудзи або її насінини, які мають стабільно вбудовану в їхній геном ДНК-конструкцію, що містить нуклеотидну послідовність, функціонально пов'язану з промотором, здатним керувати експресією нуклеотидної послідовності в рослинній клітині, де зазначена нуклеотидна послідовність вибрана з групи, що складається з: a) нуклеотидної послідовності, викладеної у SEQ ID NO: 1 або 2; та b) нуклеотидної послідовності, яка кодує поліпептид, що містить амінокислотну послідовність, яка щонайменше на 95 % ідентична амінокислотній послідовності з SEQ ID NO: 3, де зазначений поліпептид характеризується пестицидною активністю проти шкідникакукурудзяного жука; де зазначене поле заражене шкідником-кукурудзяним жуком. 4. Спосіб за пп. 1-3, де зазначений шкідник є шкідником-західним кукурудзяним жуком. 5. Спосіб за п. 2 або п. 3, де зазначена рослина додатково містить одну або декілька нуклеотидних послідовностей, що кодують один або декілька додаткових токсинів для комах. 6. Застосування рослини за п. 2 або п. 3 для контролю шкідника-кукурудзяного жука. Комп’ютерна верстка А. Крижанівський Міністерство економічного розвитку і торгівлі України, вул. М. Грушевського, 12/2, м. Київ, 01008, Україна ДП “Український інститут інтелектуальної власності”, вул. Глазунова, 1, м. Київ – 42, 01601 28
ДивитисяДодаткова інформація
Назва патенту англійськоюUse of axmi184 for the control of rootworm insects
Автори англійськоюMcNulty, Brian, Stauffer, Maria
Автори російськоюМакНалти Брайан, Стауффер Мария
МПК / Мітки
МПК: C12N 15/82, C07K 14/325, A01N 63/02
Мітки: застосування, жука, axmi184, кукурудзяного, контролю, токсину
Код посилання
<a href="https://ua.patents.su/30-115896-zastosuvannya-toksinu-axmi184-dlya-kontrolyu-kukurudzyanogo-zhuka.html" target="_blank" rel="follow" title="База патентів України">Застосування токсину axmi184 для контролю кукурудзяного жука</a>
Гібридні інсектицидні білки, що мають активність проти західного кукурудзяного кореневого жука або європейського кукурудзяного метелика

Номер патенту: 96187
Опубліковано: 10.10.2011
Автори: Харт Хоуп, Чень Жен С., Стейсі Черіл, Вальтерс Фредерік
МПК: C12N 15/82, A01H 5/00, C07K 14/325
Мітки: гібридні, західного, кукурудзяного, активність, мають, білки, жука, європейського, метелика, кореневого, інсектицидні
Формула / Реферат:
1. Сконструйований гібридний інсектицидний білок, що включає від N-кінця до С-кінця N-кінцеву ділянку першого Bacillus thuringiensis (Bt) Cry-білка, злитого із С-кінцевою ділянкою другого Bt Cry-білка, що відрізняється від першого Bt Cry-білка, причому щонайменше одна позиція кросинговера між першим і другим Bt Cry-білками розташована в консервативному блоці 2, консервативному блоці 3, варіабельній ділянці 4 або консервативному блоці 4, і що...
Спосіб підвищення стійкості кукурудзи до західного кукурудзяного жука

Номер патенту: 18041
Опубліковано: 16.10.2006
Автори: Фокін Андрій Володимирович, Адамчук Олександр Сергійович
МПК: A01M 3/00
Мітки: кукурудзи, кукурудзяного, стійкості, спосіб, жука, підвищення, західного
Формула / Реферат:
Спосіб підвищення стійкості кукурудзи до західного кукурудзяного жука, що включає штучний вплив на культуру, який відрізняється тим, що насіння обробляють мікробіологічним препаратом “Клепс” перед висівом у ґрунт в нормі 150 мл на 1 тонну.
Ген токсину axmi335 bacillus thuringiensis та спосіб його застосування

Номер патенту: 115235
Опубліковано: 10.10.2017
Автори: Лехтінен Дуан, Сампсон Кімберлі С., Тайєр Ребекка
МПК: C07K 14/32, C12N 15/82
Мітки: токсину, спосіб, thuringiensis, bacillus, axmi335, застосування, ген
Формула / Реферат:
1. Молекула рекомбінантної нуклеїнової кислоти, що містить нуклеотидну послідовність, яка кодує амінокислотну послідовність, що має пестицидну активність, яка відрізняється тим, що зазначена нуклеотидна послідовність вибрана з групи, яка складається з:a) нуклеотидної послідовності, викладеної в SEQ ID NO: 1 або 3;b) нуклеотидної послідовності, що кодує поліпептид, який містить амінокислотну послідовність з SEQ ID NO: 2 або...
Застосування ботулінового токсину для профілактики та лікування бромідрозу

Номер патенту: 78718
Опубліковано: 25.04.2007
Автор: Хекманн Марк
МПК: A61P 17/00, A61K 35/74, A61K 8/99
Мітки: токсину, лікування, застосування, профілактики, бромідрозу, ботулінового
Формула / Реферат:
1. Застосування ботулінового токсину для виробництва препаратів для профілактики та лікування бромідрозу.2. Застосування за п. 1, де ботуліновий токсин вибирають з ботулінових токсинів типу А, В, С, D, Е, F та G або їхніх похідних чи фрагментів.
Спосіб контролю оптичних характеристик кукурудзяного глютену
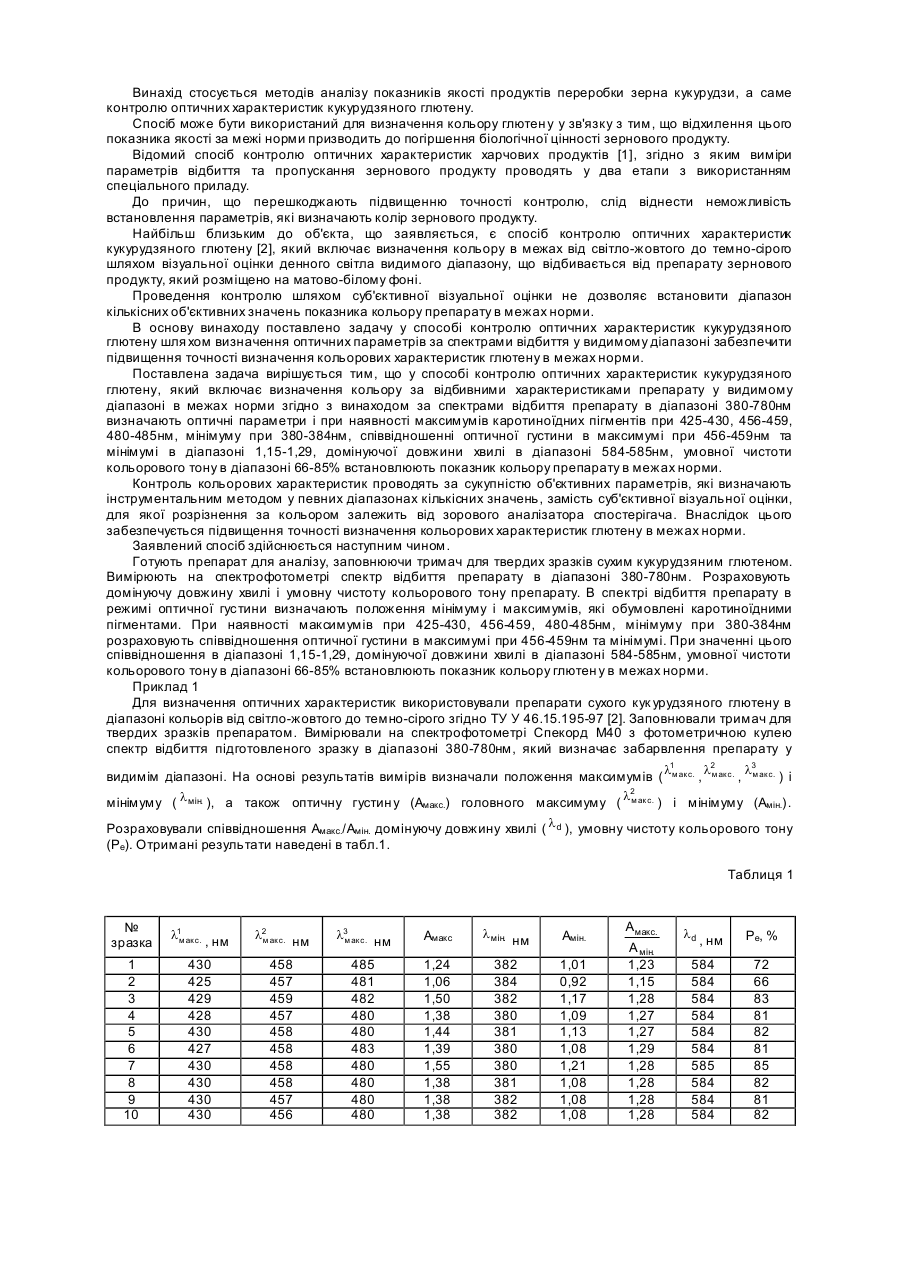
Номер патенту: 72158
Опубліковано: 17.01.2005
Автори: Феденко Володимир Савелійович, Стружко Віктор Степанович, Кияк Світлана Володимирівна
МПК: G01N 33/02
Мітки: кукурудзяного, глютену, контролю, характеристик, спосіб, оптичних
Формула / Реферат:
Спосіб контролю оптичних характеристик кукурудзяного глютену, що включає визначення кольору за відбивними характеристиками препарату у видимому діапазоні в межах норми, який відрізняється тим, що за спектрами відбиття в діапазоні 380-780 нм визначають оптичні параметри препарату і при наявності максимумів каротиноїдних пігментів при 425-430, 456-459, 480-485 нм, мінімуму при 380-384 нм, співвідношенні оптичної густини в максимумі при 456-459...
Попередній патент: Полівалентна вакцина проти грипу на основі гібридного білка
Наступний патент: Туковисівний апарат
Випадковий патент: Фотогальванічний модуль та спосіб його виготовлення