Білковий комплекс феритин
Номер патенту: 93637
Опубліковано: 10.10.2014
Автори: Борисова Тетяна Олександрівна, Борисов Арсеній Андрійович, Романенко Олександр Вікторович, Остапченко Людмила Іванівна, Крисанова Наталія Валеріївна





Формула / Реферат
Білковий комплекс феритин, до складу якого входять залізні наночастинки, що викликає дисипацію протонного градієнта синаптичних везикул у нервових терміналях головного мозку.
Текст
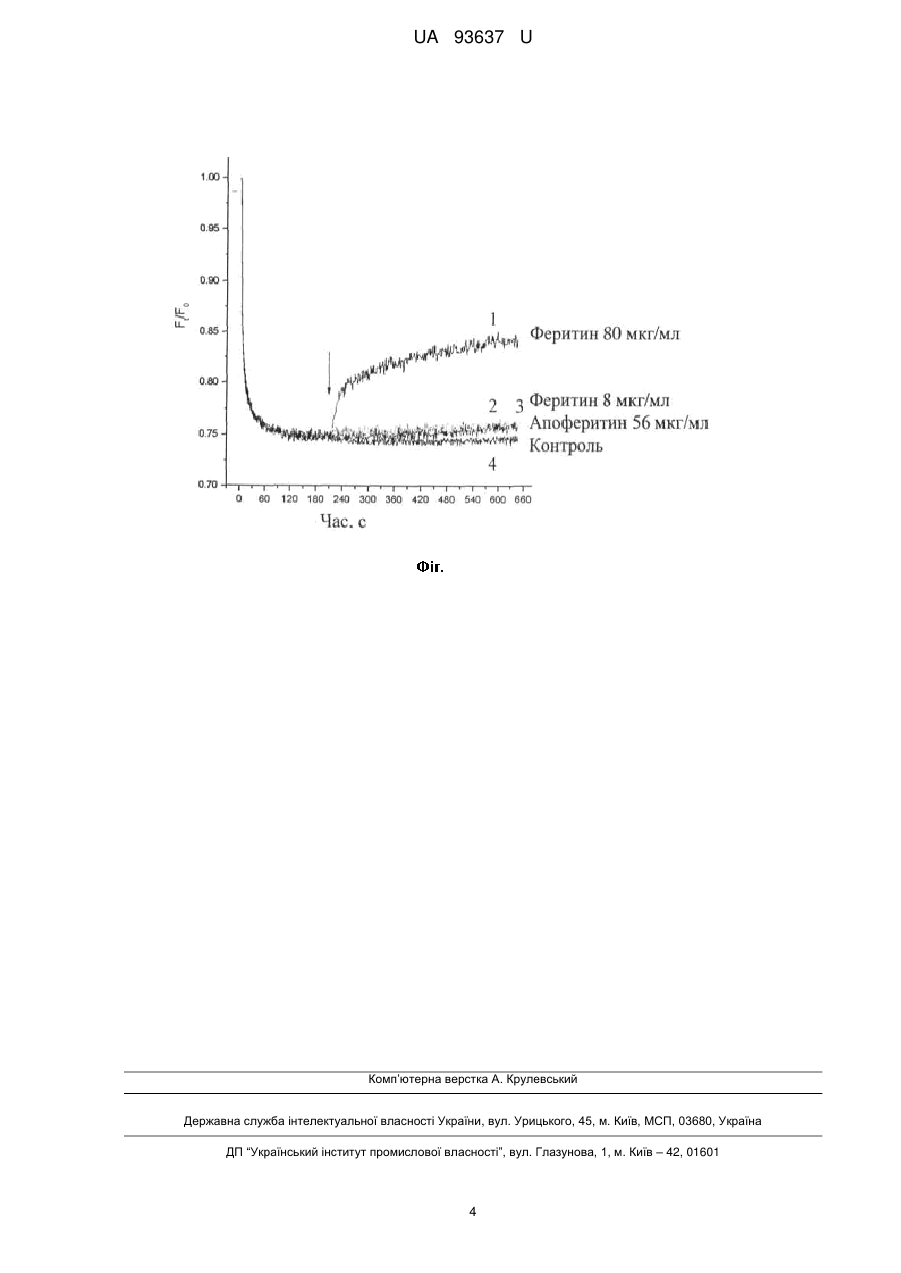
Реферат: Білковий комплекс феритин, до складу якого входять залізні наночастинки, що викликає дисипацію протонного градієнта синаптичних везикул у нервових терміналях головного мозку UA 93637 U (12) UA 93637 U UA 93637 U 5 10 15 20 25 30 35 40 45 50 55 Корисна модель належить до галузі біотехнології та медицини. Задачею корисної моделі є дослідження дії білкового комплексу феритину, до складу якого входять магнітні залізні наночастинки на протонний градієнт синаптичних везикул в нервових терміналях головного мозку. Вивчення дії феритину на протонний градієнт синаптичних везикул в нервових терміналях головного мозку проводилось у відділі нейрохімії Інституту біохімії ім. О.В. Палладіна НАН України і на кафедрі біології Національного медичного університету імені О.О. Богомольця. Одним з ключових етапів синаптичної трансмісії є екзоцитоз - вивільнення нейромедіаторів із синаптичних везикул шляхом їх злиття з плазматичною мембраною нервових закінчень. Синаптичні везикули є кислими компартментами нервових закінчень, які зберігають нейротрансмітери і вивільнюють їх шляхом екзоцитозу при стимуляції. Накопичення ацетилхоліну, моноамінів, -аміномасляної кислоти, гліцину і глутамату в синаптичні везикули здійснюється специфічними везикулярними транспортерами, функціонування яких залежить від + протонного електрохімічного градієнта ΔH , який створюється V-АТРазою [1, 2]. У цьому контексті закислення синаптичних везикул є загальною важливою характеристикою, яка забезпечує акумуляцію нейромедіаторів [3]. Оскільки протонний градієнт синаптичних везикул є важливим для акумуляції нейромедіаторів, очевидно, що його зменшення може призвести до екзоцитозу спустошених везикул у відповідь на деполяризацію плазматичної мембрани. Було показано, що неповне заповнення синаптичних везикул нейромедіатором призводить до зниження вивільнення шляхом екзоцитозу, що не супроводжується змінами в злитті синаптичних везикул і/або інших стадіях екзоцитозу [4-6]. Оскільки вивільнений з нервових закінчень в синаптичну щілину нейромедіатор зв'язується з мембранними рецепторами на постсинаптичній мембрані та активує відповідні сигнальні шляхи, то зменшення сигналу може викликати значні порушення процесу синаптичної передачі. Нещодавно було висунуто гіпотезу, що однією з причин розвитку нейрологічних захворювань може бути порушення транспорту заліза у мозку, що здійснюється за допомогою глобулярного білкового комплексу феритину, до складу якого входять магнітні залізні наночастинки [7]. Основною функцією феритину, який було знайдено в багатьох прокаріотичних та еукаріотичних організмах, є вилучення надлишку іонів заліза та переведення їх в безпечну мінеральну форму [8-11]. Феритин складається з 24 субодиниць, які формують сферичну оболонку з великою порожниною, де може розміщуватися до 4500 іонів тривалентного заліза у формі компактних мінеральних кристалітів, що нагадують феригідрити. Феритинове ядро виявляє суперпарамагнітні властивості, які характерні для магнітних наночастинок. Іони заліза входять до ядра феритину крізь міжсубодиничні канали, при цьому в середньому діаметр феритинового ядра змінюється в різних тканинах від 3,5 нм до 7,5 нм. Феритин забезпечує динамічний баланс заліза, захищаючи клітину від потенційного залізо-опосередкованого ушкодження вільними радикалами та вивільнюючи іони металу за потреби. В середині клітини феритин було знайдено в цитозолі, ядрах, ендолізосомальних компартментах та мітохондріях. Позаклітинний феритин знайдено в сироватці крові, синовіальній та цереброспінальній рідинах. Експерименти з клітинами кишечнику Сасо-2 показали, що на ентероцитах знаходиться феритиновий рецептор і вони можуть поглинати феритин рецептор-опосередкованим шляхом [10]. Також феритиновий рецептор присутній на плацентарних мембранах. У комах та червів, феритин належить до класичних білків-транспортерів заліза, що секретуються. Внутрішньоклітинний та позаклітинний феритин може відігравати роль у між- та внутрішньоклітинному перерозподілі заліза. Нині існує припущення, що феритин може відігравати суттєву роль при нейродегенерації [7]. Механізм, за допомогою якого феритин переміщується через гематоенцефалічний бар'єр є клатрин-залежним, схожий на раніше виявлений для трансферину. Зв'язування екзогенного феритину з рецепторами клітинної поверхні розглядається як важливий шлях надходження заліза в мозок [11, 12]. Порушення надходження заліза в мозок або регуляції цього процесу може спричинювати аномальний перерозподіл заліза, що сприяє розвитку цілої низки нейрологічних порушень. У рамках даної роботи був проаналізований вплив феритину на протонний градієнт синаптичних везикул у ізольованих нервових терміналях головного мозку. Ізольовані нервові терміналі (синаптосоми), виділені з головного мозку щурів, зберігають усі властивості інтактного нервового закінчення, що забезпечують процес передачі нервового імпульсу, а саме здатність накопичувати та вивільнювати нейромедіатори, підтримувати мембранний потенціал та функціональний стан синаптичних везикул. Беручи до уваги дані, що наведені вище, доцільним є аналіз впливу феритину на протонний градієнт синаптичних 1 UA 93637 U 5 10 15 20 25 30 35 40 45 50 55 везикул у синаптосомах, що матиме значення для використання феритину у галузі біотехнології та медицині для модуляції транспорту нейромедіаторів у нервових терміналях головного мозку. В основу корисної моделі поставлено задачу того, що відкриті нові властивості природного білкового комплексу феритину, до складу якого входять магнітні залізні наночастинки, щодо дії на протонний градієнт синаптичних везикул в нервових терміналях головного мозку. Показано, що феритин викликає дисипацію протонного градієнта синаптичних везикул в препараті ізольованих нервових терміналей головного мозку щурів. Синаптосоми попередньо навантажували рН-чутливим флуоресцентним зондом акридиновим оранжевим, який акумулюється в кислих компартментах синаптосом, а саме синаптичних везикулах. Додавання феритину у концентрації 80 мкг/мл миттєво викликає вивільнення флуоресцентного зонда з синаптосом. Цей факт є важливим для використання феритину у галузі біотехнології та медицини для модуляції транспорту глутамату у нервових терміналях головного мозку. Методика виділення пресинаптичних нервових закінчень головного мозку щурів та визначення рівня закислення синаптичних везикул Виділення синаптосом з головного мозку щурів Щурів-самців лінії Wistar масою 100-120 г декапітували, великі півкулі головного мозку швидко переносили в льодяний розчин 0,32 М сахарози, 5 мМ Hepes-NaOH (рН 7,4) та 0,2 мМ етилендіамінтетраоцтової кислоти (ЕДТА). Усі операції проводилися при 4 °С. Синаптосоми виділяли з гомогенату мозку диференційним центрифугуванням і центрифугуванням в градієнті фіколу, застосовуючи метод Котмана [14] з модифікацією: розчин сахарози для приготування градієнта фіколу містив 5 мМ Hepes-NaOH (рН 7,4) і 0,2 мМ ЕДТА. Синаптосомальну фракцію, отриману при фракціонуванні в градієнті фіколу, розводили 10 об'ємами 0,32 М сахарози, 5 мМ Hepes-NaOH (рН 7,4) та центрифугували при 20000 g протягом 20 хв. Отриманий осад повільно суспендували в 4 мл оксигенованого холодного середовища, що містило (в мМ): NaCl - 126, КСl - 5, MgCl2 - 1,4, NaH2PO4 - 1,0, HEPES - 20, СаСl2 - 2 і d-глюкозу - 10 (рН 7,4), (кінцева концентрація білка становила біля 4 мг/мл), та використовували в експериментах протягом 2-4 годин після виділення. Концентрацію білка визначали за методом Larson [15]. Визначення рівня закислення синаптичних везикул Для дослідження закислення синаптичних везикул був використаний рН-чутливий флуоресцентний барвник акридиновий оранжевий, що, як показав Zoccarato (3), селективно накопичується у клітинних компартментах з кислими значенням рН (зокрема в синаптичних везикулах) [16]. Акридиновий оранжевий - це ліпофільний амін; непротонована форма якого вільно проникає в мембрану. Він стає непроникаючим після протонування в кислому просторі органел, що мають всередині позитивний мембранний потенціал. Розподіл акридинового оранжевого залежить від рН. Таким чином, флюоресценція акридинового оранжевого, акумульованого в органелах, пропорційна рН органели і якісно може віддзеркалювати рівень рушійної сили для наповнення синаптичних везикул нейромедіатором. Зміни інтенсивності флуоресценції реєстрували на спектрофлуориметрі Hitachi MPF-4 (Японія), на довжинах хвиль збудження та емісії 490 та 530 нм, відповідно, (ширина щілин по 5 нм). Реакція починалася після додавання акридинового оранжевого (кінцева концентрація 5 мкМ) до суспензії синаптосом (кінцева концентрація білка 0,2 мг/мл), при постійному перемішуванні та після преінкубації синаптосомальної суспензії протягом 10 хв. при 37 °С. Флуоресценцію визначали згідно формули: F Ft / F0 де F0 та Ft - інтенсивності флуоресценції акридинового оранжевого, відповідно, за відсутності та в присутності синаптосом. Значення F0 розраховувалось шляхом екстраполяції функції експоненційного затухання до t = 0. В наших експериментах додавання синаптосомальної суспензії до розчину акридинового оранжевого супроводжувалось частковим гасінням флюоресценції внаслідок входу барвника в кислі компартменти синаптосом, а саме синаптичні везикули. Акумуляція акридинового оранжевого не є швидким процесом, стабілізація рівня флуоресцентного сигналу проходила впродовж декількох хвилин (Фіг.). Додавання феритину (80 мкг/мл) до синаптосом призводило до вивільнення акумульованого барвника і, як наслідок, до суттєвого (~ 40 %) збільшення флуоресцентного сигналу, що свідчить про миттєву дисипацію протонного градієнта синаптичних везикул, викликану феритином (Фіг.). Як було зазначено вище, молекула феритину складається з білкової оболонки, в середині якої розташована порожнина з мінеральною наночастинкою, що нагадує феригідрит. Для 2 UA 93637 U 5 10 15 20 25 30 35 40 45 50 дослідження ефектів, пов'язаних з протеїновим компонентом молекули феритину, було проаналізовано протонний градієнт синаптичних везикул в присутності апоферитину, який містить лише залишок слідів заліза. Слід зазначити, що в молекулі апоферитину (А3660, Sigma лот 061М6039) концентрація заліза складає 0,025 мг/мл (0,0025 % залишкового заліза), в той час як в молекулі феритину (F4503, Sigma, лот 079К7001) - 7,13 мг/мл (вміст заліза - 0,7 %). Апоферитин в концентрації 80 мкг/мл практично не впливав на протонний градієнт синаптичних везикул у синаптосомах. Отже, вплив феритину на протонний градієнт синаптичних везикул обумовлений неорганічним компонентом молекули, а саме феррігідритом (Фіг.). Таким чином, слід підкреслити наступне: 1. Феритин миттєво викликає дисипацію протонного градієнта синаптичних везикул в ізольованих нервових закінченнях головного мозку щурів. 2. Зниження протонного градієнта синаптичних везикул в ізольованих нервових закінченнях головного мозку щурів на 40 % відбувається при додаванні 80 мкг/мл феритину в середовище інкубації. 3. Вплив феритину на протонний градієнт синаптичних везикул обумовлений неорганічним компонентом молекули, а саме феригідритом. Література: 1. Moriyama Y, Maeda M, Futai M. The role of V-ATPase in neuronal and endocrine systems. J Exp Biol. 1992 Nov; 172:171. 2. Nelson N. Structure and function of V-ATPases in endocytic and secretory organelles. J Exp Biol. 1992 Nov; 172:149. 3. Zoccarato, F., Cavallini, L., Alexandre A. The pH-sensitive dye acridine orange as a tool to monitor exocytosis/endocytosis in synaptosomes. J. Neurochem. 1999, V. 72, P. 625-633. 4. Dröse S. and Altendorf K. Bafilomycins and concanamycins as inhibitors of V-ATPases and PATPases. J. Exp. Biol. 1997, V. 200, P. 1-8. 5. Floor E., Leventhal P.S., Schaeffer S.F. Partial purification and characterization of the vacuolar H(+)-ATPase of mammalian synaptic vesicles. J. Neurochem. 1990, V. 55, P. 1663-1670. 6. Zhou Q, Petersen С. С Н, and Nicoll RA Effects of reduced vesicular filling on synaptic transmission in rat hippocampal neurones J Physiol. 2000 May V. 15, P. 195-206. 7. Friedman A., P. Arosio, D. Finazzi, D. Koziorowski, J. Galazka-Friedman, Ferritin as an important player in neurodegeneration, Parkinsonism and Related Disorders 2011, V. 17, P. 423-430. 8. Munro H.N., M.C. Linder Ferritin: structure, biosynthesis, and role in iron metabolism, Physiol. Rev. 1978, V. 58, P. 317-396. 9. Andrews S.C., P. Arosio, W. Bottke, J.F. Briat, M. vonDarl, P.M. Harrison, J.P. Laulhere, S. Levi, S. Lobreaux, S.J. Yewdall Structure, function, and evolution of ferritins, J. Inorg. Biochem. 1992, V. 47, P. 161-174. 10. Kidane T.Z., E. Sauble, M.C. binder, Release of iron from ferritin requires lysosomal activity, Am. J. Physiol. Cell Physiol. 2006, V. 291, C. 445-C455. 11. Kalgaonkar S., B. Lonnerdal Receptor-mediated uptake of ferritin-bound iron by human intestinal Caco-2 cells, J. Nutr. Biochem. 2009, V. 20, P. 304-311. 12. Burdo J.R., D.A. Antonetti, E.B. Wolpert, J.R. Connor, Mechanisms and regulation of transferrin and iron transport in a model blood-brain barrier system, Neuroscience 2003, V. 121, P. 883-890. 13. Fisher J., K. Devraj, J. Ingram, B. Slagle-Webb, A.B. Madhankumar, X. Liu, M. Klinger, I.A. Simpson, J.R. Connor, Ferritin: a novel mechanism for delivery of iron to the brain and other organs, Am. J. Physiol. Cell Physiol. 2007, V. 293, P. 641-C649. 14. Cotman C.W. Isolation of synaptosomal and synaptic plasma membrane fractions // Meth. Enzymol. - 1974, V. 31, P. 445-452. 15. Larson E., Howlett В., Jagendorf A. Artificial reductant enhancement of the Lowry method for protein determination // Anal. Biochem. - 1986. - Vol. 155, № 2. - P. 243-248. 16. Zoccarato F., Cavallini L., Alexandre A. The pH-sensitive dye acridine orange as a tool to monitor exocytosis/endocytosis in synaptosomes // J. Neurochem. - 1999, V. 72, P. 625-633. ФОРМУЛА КОРИСНОЇ МОДЕЛІ 55 Білковий комплекс феритин, до складу якого входять залізні наночастинки, що викликає дисипацію протонного градієнта синаптичних везикул у нервових терміналях головного мозку. 3 UA 93637 U Комп’ютерна верстка А. Крулевський Державна служба інтелектуальної власності України, вул. Урицького, 45, м. Київ, МСП, 03680, Україна ДП “Український інститут промислової власності”, вул. Глазунова, 1, м. Київ – 42, 01601 4
ДивитисяДодаткова інформація
Автори англійськоюBorysova Tetiana Oleksandrivna, Romanenko Oleksandr Viktorovych, Krysanova Natalia Valeriivna, Borysov Arsenii Andriiovych, Ostapchenko Liudmyla Ivanivna
Автори російськоюБорисова Татьяна Александровна, Романенко Александр Викторович, Крысанова Наталия Валериевна, Борисов Арсений Андреевич, Остапченко Людмила Ивановна
МПК / Мітки
МПК: C07D 277/00
Мітки: феритин, комплекс, білковий
Код посилання
<a href="https://ua.patents.su/6-93637-bilkovijj-kompleks-feritin.html" target="_blank" rel="follow" title="База патентів України">Білковий комплекс феритин</a>
Білковий комплекс феритин

Номер патенту: 93636
Опубліковано: 10.10.2014
Автори: Романенко Олександр Вікторович, Борисов Арсеній Андрійович, Крисанова Наталія Валеріївна, Остапченко Людмила Іванівна, Борисова Тетяна Олександрівна
МПК: C07D 277/00
Мітки: комплекс, білковий, феритин
Формула / Реферат:
Білковий комплекс феритин, до складу якого входять феригідритні наночастинки, що збільшує позаклітинний рівень глутамату в нервових терміналах головного мозку.
Застосування сполуки 3-децилоксикарбонілметил-4-метил-5-(2-гідроксіетил)тіазолій хлорид як сполуки, яка викликає зменшення протонного градієнта синаптичних везикул в ізольованих нервових закінченнях головного м

Номер патенту: 45943
Опубліковано: 25.11.2009
Автори: Касаткіна Людмила Олександрівна, Крисанова Наталія Валеріївна, Остапченко Людмила Іванівна, Вовк Андрій Іванович, Борисова Тетяна Олександрівна, Романенко Олександр Вікторович, Борисов Арсеній Андрійович
МПК: C07D 277/00
Мітки: протонного, викликає, закінченнях, синаптичних, зменшення, ізольованих, головного, везикул, застосування, хлорид, нервових, 3-децилоксикарбонілметил-4-метил-5-(2-гідроксіетил)тіазолій, яка, градієнта, сполуки
Формула / Реферат:
Застосування сполуки 3-децилоксикарбонілметил-4-метил-5-(2-гідроксіетил)тіазолій хлорид як сполуки, яка викликає зменшення протонного градієнта синаптичних везикул в ізольованих нервових закінченнях головного мозку щурів, загальної формули:.
Спосіб діагностики анемії при системному червоному вовчаку

Номер патенту: 73897
Опубліковано: 10.10.2012
Автори: Шевчук Сергій Вікторович, Куріленко Ірина Володимирівна
МПК: A61B 5/00, G01N 33/00
Мітки: анемії, системному, вовчаку, діагностики, червоному, спосіб
Формула / Реферат:
Спосіб діагностики анемії при системному червоному вовчаку, який включає проведення загального, біохімічного та імунологічного аналізів крові, загальних аналізів сечі, калу, УЗД внутрішніх органів, визначення концентрації заліза і рівня феритину в сироватці крові та вмісту заліза в аспіраті кісткового мозку, який відрізняється тим, що при відсутності заліза в кістковому мозку діагностують залізодефіцитну анемію.
Спосіб лікування синдрому “неспокійних ніг” шляхом введення композиції, що містить комплекс заліза

Номер патенту: 83483
Опубліковано: 25.07.2008
Автори: Геленек Мері Джейн, Токарс Марк Л., Лендж Ральф А., Олдхем Фред Б.
МПК: A61P 21/00, A61K 33/26
Мітки: неспокійних, спосіб, містить, синдрому, введення, ніг, композиції, комплекс, заліза, лікування, шляхом
Формула / Реферат:
1. Спосіб лікування cиндрому неспокійних ніг, який відрізняється тим, що залізо-комплекс, який вводять суб'єкту, має швидкість вивільнення заліза, більшу ніж IDI (декстран заліза для ін'єкцій), де швидкість вивільнення заліза визначається при концентрації щонайменше 2.000 мкг/дл.2. Спосіб за п. 1, який відрізняється тим, що засіб вводиться парентерально.3. Спосіб за п. 1 або 2, який відрізняється тим, що вказаний залізо-комплекс...
Білковий крем

Номер патенту: 88870
Опубліковано: 10.04.2014
Автори: Соколовська Ірина Олександрівна, Харкава Ольга Олександрівна, Камбулова Юлія Вікторівна
Формула / Реферат:
Білковий крем, що включає білок яєчний, цукор білий кристалічний, який відрізняється тим, що додатково вводиться комплекс структуроутворювачів - пектин низькоетерифікований амідований і альгінат натрію в наступному співвідношенні компонентів, %: Компоненти Рецептурна кількість,% білок яєчний сухий 6,85 цукор білий кристалічний 92,45 ...
Попередній патент: Білковий комплекс феритин
Наступний патент: Спосіб усунення перехресних похибок у надлишкових блоках двокомпонентних інерціальних датчиків
Випадковий патент: Електрична лампа розжарювання загального призначення