Білковий комплекс феритин
Номер патенту: 93636
Опубліковано: 10.10.2014
Автори: Борисова Тетяна Олександрівна, Остапченко Людмила Іванівна, Крисанова Наталія Валеріївна, Романенко Олександр Вікторович, Борисов Арсеній Андрійович





Формула / Реферат
Білковий комплекс феритин, до складу якого входять феригідритні наночастинки, що збільшує позаклітинний рівень глутамату в нервових терміналах головного мозку.
Текст
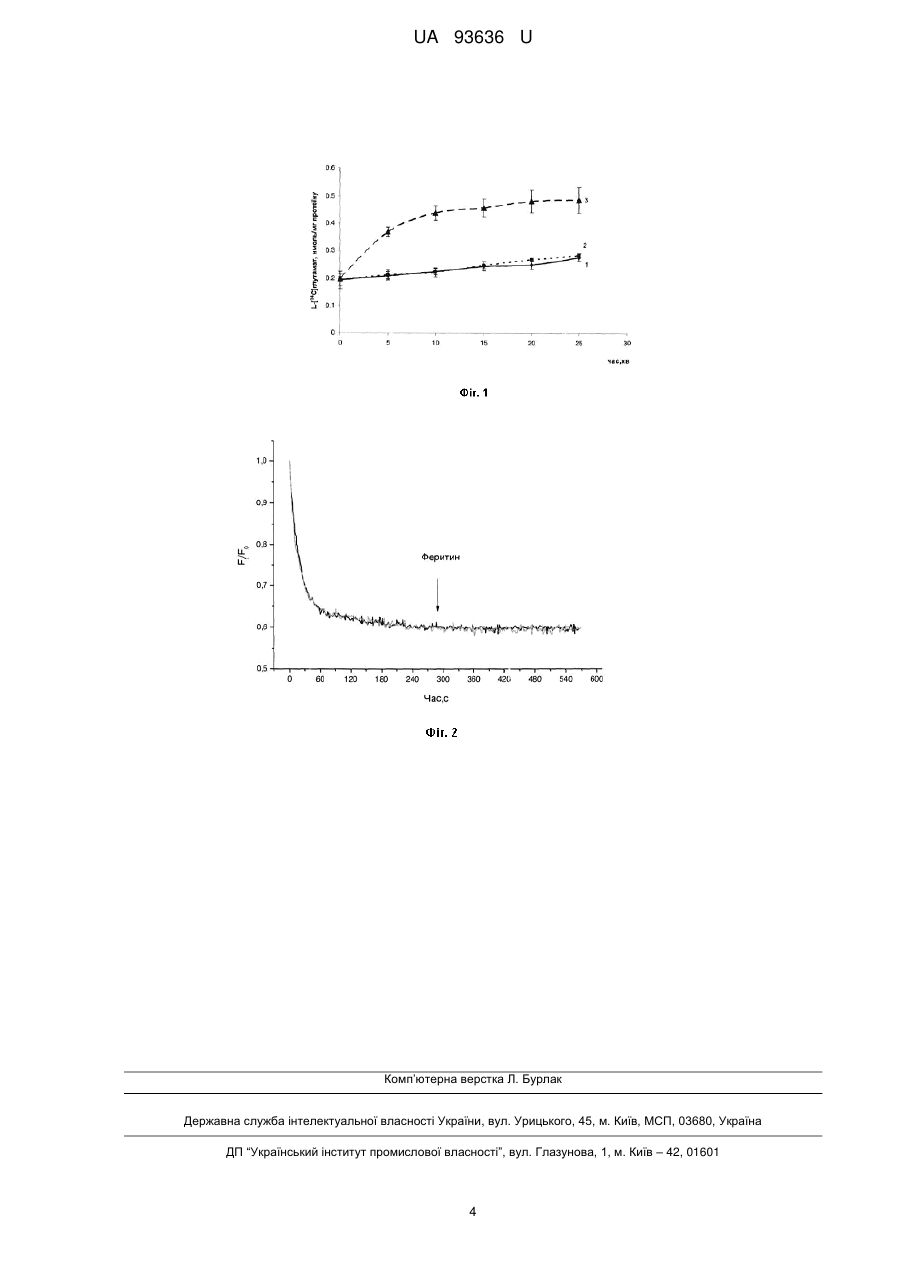
Реферат: Білковий комплекс феритин, до складу якого входять феригідритні наночастинки, що збільшує позаклітинний рівень глутамату в нервових терміналах головного мозку UA 93636 U (12) UA 93636 U UA 93636 U 5 10 15 20 25 30 35 40 45 50 55 60 Корисна модель належить до галузі біотехнології та медицини. Задачею корисної моделі є дослідження дії білкового комплексу феритину, до складу якого входять магнітні феригідритні наночастинки, на позаклітинний рівень глутамату та мембранний потенціал нервових терміналей головного мозку. Вивчення дії феритину на позаклітинний рівень глутамату та мембранний потенціал нервових терміналей головного мозку проводилось у відділі нейрохімії Інституту біохімії ім. О.В. Палладіна НАН України і на кафедрі біології Національного медичного університету імені О.О. Богомольця. Феритин складається з 24 субодиниць, які формують сферичну оболонку з великою порожниною, де розміщуються іони тривалентного заліза у формі компактних мінеральних кристалітів, що нагадують феригідрити. Феритинове ядро, діаметр якого змінюється в різних тканинах від 3,5 нм до 7,5 нм, виявляє суперпарамагнітні властивості, які характерні для магнітних наночастинок. Феритин забезпечує динамічний баланс заліза, захищаючи клітину від потенційного залізо-опосередкованого ушкодження вільними радикалами та вивільнюючи іони металу за потреби. В середині клітини феритин було знайдено в цитозолі, ядрах, ендолізосомальних компартментах та мітохондріях. Позаклітинний феритин знайдено в сироватці крові, синовіальній та цереброспінальній рі щнах. Внутрішньоклітинний та позаклітинний феритин може відігравати роль у між- та внутрішньоклітинному перерозподілі заліза. Нещодавно було висунуто гіпотезу, що однією з причин розвитку нейрологічних захворювань може бути порушення транспорту заліза у мозку [1]. Нині існує припущення що феритин може відігравати суттєву провокуючу роль у розвитку нейродегенерації [1]. Механізм, за допомогою якого феритин переміщується через гематоенцефатичний бар'єр є клатринзалежним, схожий на раніше виявлений для трансферину. Зв'язування екзогенного феритину з рецепторами клітинної поверхні розглядається як важливий шлях надходження заліза в мозок [2-4]. Порушення надходження заліза в мозок або регуляції цього процесу може спричинювати аномальний перерозподіл заліза, що сприяє розвитку цілої низки нейрологічних порушень. Передбачається використання феритину як моделі та прототипу синтезованих магнітних наночастинок, покритих полімерами [5]. Амінокислота глутамат в центральній нервовій системі ссавців є найбільш розповсюдженим збуджуючим медіатором і приймає участь в здійсненні багатьох функцій головного мозку, включаючи розпізнавання, пам'ять, навчання тощо. Найхарактернішою рисою патогенезу більшості неврологічних розладів та захворювань (хвороба Альцгеймера, епілепсія, шизофренія та інші) є порушення транспорту глутамату [6]. У нервовій системі надлишковий позаклітинний глутамат має токсичні властивості, оскільки надмірна активація глутаматних рецепторів та масивний вхід кальцію може призвести до загибелі нейронів. Підтримання низького рівня позаклітинного глутамату є дуже важливим для певної спонтанної активності мозку та нормальної синаптичної передачі, причому навіть незначні зміни рівня позаклітинного глутамату здатні призводити до суттєвих порушень у функціонуванні центральної нервової системи. Термінація глутаматергічної нейропередачі відбувається шляхом видалення нейромедіатора з синаптичної щілини за рахунок його поглинання всередину нервової клітини. Поглинання глутамату нервовими клітинами забезпечується роботою високоафінних натрій-залежних глутаматних транспортерів, які локалізовані у плазматичній мембрані нейронів і гліальних клітин + + [6]. Рушійною силою для функціонування цих транспортерів є Na /K градієнт плазматичної мембрани. Вивільнений в синаптичну щілину шляхом екзоцитозу глутамат зв'язується з мембранними рецепторами та активує відповідні сигнальні шляхи, що забезпечує подальшу передачу сигналу. За відсутності стимуляції нервових закінчень концентрація позаклітинного глутамату є однією з основних характеристик, що визначає спонтанну активність мозку. Рівень глутамату у синаптичній щілині встановлюється балансом між накопиченням глутамату та його вивільненням. За нормальних умов концентрація глутамату в синаптичній щілині підтримується на достатньо низькому рівні, що становить менше ніж 1 мкМ. Завдяки цьому унеможливлюється надмірна активація іонотропних та метаботропних глутаматних пост- та пресинптичних рецепторів. В той же час надзвичайно важливим для нормального функціонування мозку є процес тонічної активації цих рецепторів позаклітинним глутаматом. Підвищена концентрація глутамату у синаптичній щілині порушує синаптичну передачу та створює підґрунтя для розвитку нейрологічних розладів. Ізольовані нервові терміналі (синаптосоми), виділені з головного мозку щурів, зберігають усі властивості інтактного нервового закінчення, що забезпечують процес передачі нервового імпульсу, а саме, здатність накопичувати та вивільнювати нейромедіатори, підтримувати мембранний потенціал та функціональний стан синаптичних везикул. Беручи до уваги наведені вище дані, є доцільним проаналізувати вплив феритину на позаклітинний рівень глутамату та 1 UA 93636 U 5 10 15 20 25 30 35 40 45 50 55 60 потенціал плазматичної мембрани в нервових закінченнях головного мозку щурів, що матиме значення для використання феритину у галузі біотехнології та медицині для модуляції транспорту нейромедіаторів у нервових терміналах головного мозку. В основу корисної моделі поставлено задачу того, що феритин викликає збільшення 14 позаклітинного рівню радіоактивно міченого L-[ C]глутамату в попередньо навантажених ним препаратах ізольованих нервових терміналей (синаптосом) головного мозку щурів. 14 Позаклітинний рівень L-[ C] глутамату в синаптосомах збільшувався в присутності феритину у концентрації 80 мкг/мл у середовищі інкубації приблизно у два рази у порівнянні з контролем. 14 Позаклітинний рівень L-[ С]глутамату становив за відсутності феритину - 0,200±0,015 нмоль/мг білка, в присутності феритину в середовищі інкубації синаптосом упродовж 5 хв. в концентрації 80 мкг/мл - 0,368±0,016 нмоль/мг білка. Позаклітинний рівень глутамату залишається суттєво підвищеним протягом 25 хв. Показано також, що феритин не викликає зміни мембранного потенціалу синаптосом. Ці факти є важливими для використання феритину у галузі біотехнології для модуляції транспорту глутамату у нервових терміналях головного мозку. Ознаки способу. Методика виділення нервових закінчень (синаптосом) з головного мозку щурів та 14 визначення позаклітинного рівня L-[ С]глутамату в синаптосомах Виділення синаптосом з головного мозку щурів Щурів-самців лінії Wistar масою 100-120 г декапітували, великі півкулі головного мозку швидко переносили в розчин, що містив 0,32 Μ сахарози, 5 мМ Hepes-NaOH (pH 7,4) та 0,2 мМ етилендіамінтетраоцтової кислоти (ЕДТА). Усі операції проводилися при 4 °C. Синаптосоми виділяли з гомогенату мозку диференційним центрифугуванням і центрифугуванням в градієнті щільності фіколу, застосовуючи метод Котмана [7] у такій модифікації: розчин сахарози для приготування градієнту фіколу містив 5 мМ Hepes-NaOH (pH 7,4) і 0,2 мМ ЕДТА. Синаптосомальну фракцію, отриману при центрифугуванні гомогенату головного мозку в градієнті щільності фіколлу, розводили 10 об'ємами 0,32 Μ сахарози, 5 мМ Hepes-NaOH (pH 7,4) і 0,2 мМ ЕДТА та центрифугували при 20000 g упродовж 20 хв. Отриманий осад повільно суспендували в 4 мл оксигенованого холодного середовища, що містило (в мМ): NaCl-126, КСl5, MgCl2-1,4, NaH2PO4-1,0, HEPES-20, СаСl2-2, d-глюкозу - 10 (pH 7,4). При цьому кінцева концентрація білка становила 4 мг/мл. Синаптосоми використовували в експериментах упродовж 2-4 годин після отримання. Концентрацію білка визначали за методом Ларсона [8]. 14 Визначення позаклітинного рівня L-[ С] глутамату в синаптосомах Суспензія синаптосом розводилася стандартним сольовим розчином так, що містила 1 мг 14 білка/мл, і після 10 хв. преінкубації при 37 °C навантажувалася L-[ С]глутаматом (500 нМ, 238 мСі/ммол) (Amersham, Великобританія) в кальцієвому стандартному сольовому розчині упродовж 10 хв. Після цього суспензія синаптосом відмивалася 10 об'ємами льодяного стандартного сольового розчину і розводилася до концентрації 1 мг білка/мл і відразу використовувалася I4 для визначення вивільнення L-[ C] глутамату з синаптосом. 14 Аліквоти (120 мкл синаптосом, навантажених L-[ С]глу там атом, 25-30 мкг білка), 14 преінкубували 10 хв при 37 °C. Вимірювання позаклітинного рівня L-[ C] глутамату в синаптосомах у кальційвмісному середовищі відбувалося 1-25 хв. Через визначений проміжок часу суспензію синаптосом швидко осаджували в мікроцентрифузі, 10000 g протягом 20 с Аліквоти надосаду (90 мкл) та солюбілізованого додецилсульфатом натрію осаду (90 мкл) змішували з синтиляційною рідиною ACS (Amersham, Великобританія) (1,5 мл) та визначали радіоактивність за допомогою синтиляційного лічильника Тгасог Analytic Delta 300 (США). Загальний вміст радіоактивності визначали як суму радіоактивності у аліквоті надосаду та у аліквоті солюбілізованого осаду. Згідно з протоколом дослідження позаклітинного рівня глутамату в ізольованих нервових закінченнях головного мозку щурів вивчалося з використанням радіоактивно міченого L14 [ С]глутамату, яким попередньо були навантажені синаптосоми (див. методику). В наших експериментах спостерігається постійне нестимульоване (базальне) вивільнення L14 [ С]глутамату, що є однією з основних характеристик препарату синаптосом (Фіг. 1, суцільна 14 лінія). На теперішній час механізм, завдяки якому L-[ С]глутамат спонтанно вивільнюється з нервових закінчень у синаптичну щілину, остаточно не з'ясовано, але передбачається можливість залучення до цього процесу спонтанного екзоцитозу, аніоних каналів, цистеїн глутаматного обмінника та трансмембранної дифузії. Було продемонстровано, що додавання феритину до нервових закінчень призводило до збільшення позаклітинного рівня L14 [ C]глутамату у синаптосомах. На Фіг. 1 (пунктирна лінія 3) представлено динаміку зміни 14 позаклітинного рівня L-[ C]глутамату внаслідок його нестимульованого вивільнення з 2 UA 93636 U 5 10 15 20 25 30 35 40 45 синаптосом у часі за умови їх інкубації з феритином в концентрації 80 мкг/мл. Показано, що 14 феритин суттєво підвищує позаклітинний рівень L-[ C]глутамату в ізольованих нервових закінченнях упродовж 1-25 хв. (n=5; Р0,05). Додаткові контрольні експерименти проводились у присутності альбуміну (80 мкг / мл) Фіг.1 (пунктирна лінія 2). + + Na /K електрохімічний градієнт на плазматичній мембрані є рушійною силою для транспортер-залежного накопичення глутамату, зміни мембранного потенціалу можуть впливати на початкову швидкість цього процесу [9-11]. Як з'ясувалося, деполяризація плазмалеми 35 мм КС1 призводила до зниження початкової швидкості накопичення L14 [ C]глутамату удвічі. Вимірювання потенціалу плазматичної мембрани Em проводили з використанням катіонного потенціал-чутливого флуоресцентного барвника родаміну 6G (Molecular Probes, США). Показано, що додавання феритину до синаптосом не призводить до збільшення флуоресцентного сигналу зонду, що свідчить про відсутність деполяризації плазматичної мембрани нервових закінчень (Фіг.2). Таким чином, слід підкреслити наступне: 14 1. Феритин збільшує позаклітинний рівень L-[ C]глутамату в ізольованих нервових закінченнях (синаптосомах) головного мозку щурів. 2. Внесення феритину у концентрації 80 мкг/мл у середовище інкубації синаптосом 14 обумовлює збільшення позаклітинного рівня L-[ С]глутамату на 85 %. 3. Додавання феритину у концентрації 80 мкг/мл до синаптосом не призводить до сигналу потенціал-чутливого флуоресцентно флуоресцентного барвника родаміну 6G, що свідчить про відсутність деполяризації плазматичної мембрани нервових закінчень. Джерела інформації:: 1. Friedman Α., P. Arosio, D. Finazzi, D. Koziorowski, J. Galazka-Friedman, Ferritin as an important player in neurodegeneration, Parkinsonism and Related Disorders 2011 17 423-430. 2. Kidane T.Z., E. Sauble, M.C. Linder, Release of iron from ferritin requires lysosomal activity, Am. J. Physiol. Cell Physiol. 2006, V. 291. - C445-C455. 3. Kalgaonkar S., B. Lonnerdal Receptor-mediated uptake of ferritin-bound iron by human intestinal Caco-2 cells, J. Nutr. Biochem. 2009, V. 20-304-311. 4. Burdo J.R., D.A. Antonetti, E.B. Wolpert, J.R. Connor, Mechanisms and regulation of transferrin and iron transport in a model blood-brain barrier system, Neuroscience 2003, V. 121-883-890. 5. Alekseenko A.V., Waseem T.V., Fedorovich S.V. Ferritin, a protein containing iron nanoparticles, induces reactive oxygen species formation and inhibits glutamate uptake in rat brain synaptosomes // Brain Res. - 2008. - V. 1241. - Р. 193-200. 6. Danbolt N. Glutamate uptake // Prog. Neurobiol. - 2001. - V. 65. - P. 1-105. 7. Cotman C. W. Isolation of synaptosomal and synaptic plasma membrane fractions // Meth. Enzymol.-1974.- V. 31. - P. 445-452. 8. Larson E., Howlett В., Jagendorf A. Artificial reductant enhancement of the Lowry method for protein determination // Anal. Biochem. - 1986. -Vol.155, № 2. - P. 243-248. 9. Borisova Т., S vko R., Borysov Α., Krisanova N. Diverse presynaptic mechanisms underlying methyl-beta-cyclodextrin-mediated changes in glutamate transport // Cell. Мої. Neurobiol. - 2010. - V. 30, № 7. - P. 1013-1023. 10. Borisova T. Krisanova N., Sivko R., Borysov A. Cholesterol depletion attenuates tonic release but increases the ambient level of glutamate in rat brain synaptosomes // Neurochem. Int. - 2010. - V. 56. - P. 466-478. 11. Krisanova N, Sivko R, Kasatkina L, Borisova T. Neuroprotection by lowering cholesterol: a decrease in membrane cholesterol content reduces transporter-mediated glutamate release from brain nerve terminals // Biochim Biophys Acta. - 2012. - V. 1822. - P. 1553-1561. ФОРМУЛА КОРИСНОЇ МОДЕЛІ 50 Білковий комплекс феритин, до складу якого входять феригідритні наночастинки, що збільшує позаклітинний рівень глутамату в нервових терміналах головного мозку. 3 UA 93636 U Комп’ютерна верстка Л. Бурлак Державна служба інтелектуальної власності України, вул. Урицького, 45, м. Київ, МСП, 03680, Україна ДП “Український інститут промислової власності”, вул. Глазунова, 1, м. Київ – 42, 01601 4
ДивитисяДодаткова інформація
Автори англійськоюBorysova Tetiana Oleksandrivna, Romanenko Oleksandr Viktorovych, Krysanova Natalia Valeriivna, Borysov Arsenii Andriiovych, Ostapchenko Liudmyla Ivanivna
Автори російськоюБорисова Татьяна Александровна, Романенко Александр Викторович, Крысанова Наталия Валериевна, Борисов Арсений Андреевич, Остапченко Людмила Ивановна
МПК / Мітки
МПК: C07D 277/00
Мітки: феритин, комплекс, білковий
Код посилання
<a href="https://ua.patents.su/6-93636-bilkovijj-kompleks-feritin.html" target="_blank" rel="follow" title="База патентів України">Білковий комплекс феритин</a>
Спосіб прогнозування чутливості до неоад’ювантної терапії у хворих на рак молочної залози

Номер патенту: 106148
Опубліковано: 25.07.2014
Автори: Лук'янова Наталія Юріївна, Антіпова Светлана Володимирівна, Чехун Василь Федорович, Шепіль Олександр Володимирович
МПК: G01N 33/49, A61B 10/00
Мітки: хворих, неоад'ювантної, рак, спосіб, прогнозування, залози, терапії, молочної, чутливості
Формула / Реферат:
Спосіб прогнозування чутливості до неоад'ювантної терапії у хворих на рак молочної залози з верифікованим діагнозом, який відрізняється тим, що перед проведенням першого курсу неоад'ювантної хіміотерапії у хворих на РМЗ визначають рівень феритину в сироватці крові, і при значеннях рівня феритину від 10 до 200 нг/мл прогнозують чутливість пухлини до неоад'ювантної хіміотерапії, а при значеннях рівня феритину ³ 200 нг/мл -...
Застосування сполуки 3-децилоксикарбонілметил-4-метил-5-(2-гідроксіетил)тіазолій хлорид як сполуки, яка викликає збільшення нестимульованого вивільнення глутамату з нервових закінчень головного мозку щурів

Номер патенту: 45942
Опубліковано: 25.11.2009
Автори: Сівко Роман Віталійович, Вовк Андрій Іванович, Борисов Арсеній Андрійович, Борисова Тетяна Олександрівна, Романенко Олександр Вікторович, Крисанова Наталія Валеріївна
МПК: C07D 277/00
Мітки: застосування, викликає, глутамату, хлорид, головного, збільшення, закінчень, нестимульованого, 3-децилоксикарбонілметил-4-метил-5-(2-гідроксіетил)тіазолій, нервових, щурів, сполуки, мозку, яка, вивільнення
Формула / Реферат:
Застосування сполуки 3-децилоксикарбонілметил-4-метил-5-(2-гідроксіетил)тіазолій хлорид як сполуки, яка викликає збільшення нестимульованого вивільнення глутамату з нервових закінчень головного мозку щурів, загальної формули:.
Білковий крем

Номер патенту: 88870
Опубліковано: 10.04.2014
Автори: Соколовська Ірина Олександрівна, Харкава Ольга Олександрівна, Камбулова Юлія Вікторівна
Формула / Реферат:
Білковий крем, що включає білок яєчний, цукор білий кристалічний, який відрізняється тим, що додатково вводиться комплекс структуроутворювачів - пектин низькоетерифікований амідований і альгінат натрію в наступному співвідношенні компонентів, %: Компоненти Рецептурна кількість,% білок яєчний сухий 6,85 цукор білий кристалічний 92,45 ...
Комплекс безпечного отримання продукції від загазованих шахт будь-якої глибини “комплекс краснікова”

Номер патенту: 61907
Опубліковано: 10.08.2011
Автор: Красніков Евгеній Аркадійович
МПК: E21C 41/00
Мітки: отримання, шахт, комплекс, продукції, безпечного, глибини, загазованих, краснікова, будь-якої
Формула / Реферат:
Комплекс безпечного отримання продукції від загазованих шахт будь-якої глибини, що містить головний та аварійний стовбури, штреки, підйомно-транспортне обладнання, насосно-водне обладнання, електрокабелі, системи сигналізації: небезпечної концентрації горючих газів (СН4, H2S тощо) та рівня О2 у повітрі шахти, рівня ґрунтових вод, температури та вологості повітря, який відрізняється тим, що виконаний у вигляді системи забезпечення повної...
Вуглеводно-білковий харчовий продукт

Номер патенту: 82319
Опубліковано: 25.07.2013
Автори: Притульська Наталія Володимирівна, Мотузка Юлія Миколаївна, Вдовенко Наталія Володимирівна, Гуліч Марія Павлівна, Хробатенко Олександра Віталіївна
МПК: A23J 1/20, A23L 2/66, A23L 2/39
Мітки: вуглеводно-білковий, продукт, харчовий
Формула / Реферат:
1. Вуглеводно-білковий продукт, що спрямований на забезпечення організму необхідною кількістю макроергічних сполук, а також інтенсифікацію їх внутрішньоклітинного синтезу, який відрізняється тим, що містить у своєму складі АТФ-ліпідний комплекс, а також інші компоненти в наступному співвідношенні, %: концентрат білків молочної сироватки 39,6-48,4 глюкоза 22,5-27,5 ...
Попередній патент: Фармацевтична композиція для лікування хронічних обструктивних захворювань легень
Наступний патент: Білковий комплекс феритин
Випадковий патент: Спосіб консервування перги бурштиновою кислотою