Система експресії гетерологічних і гомологічних целюлаз

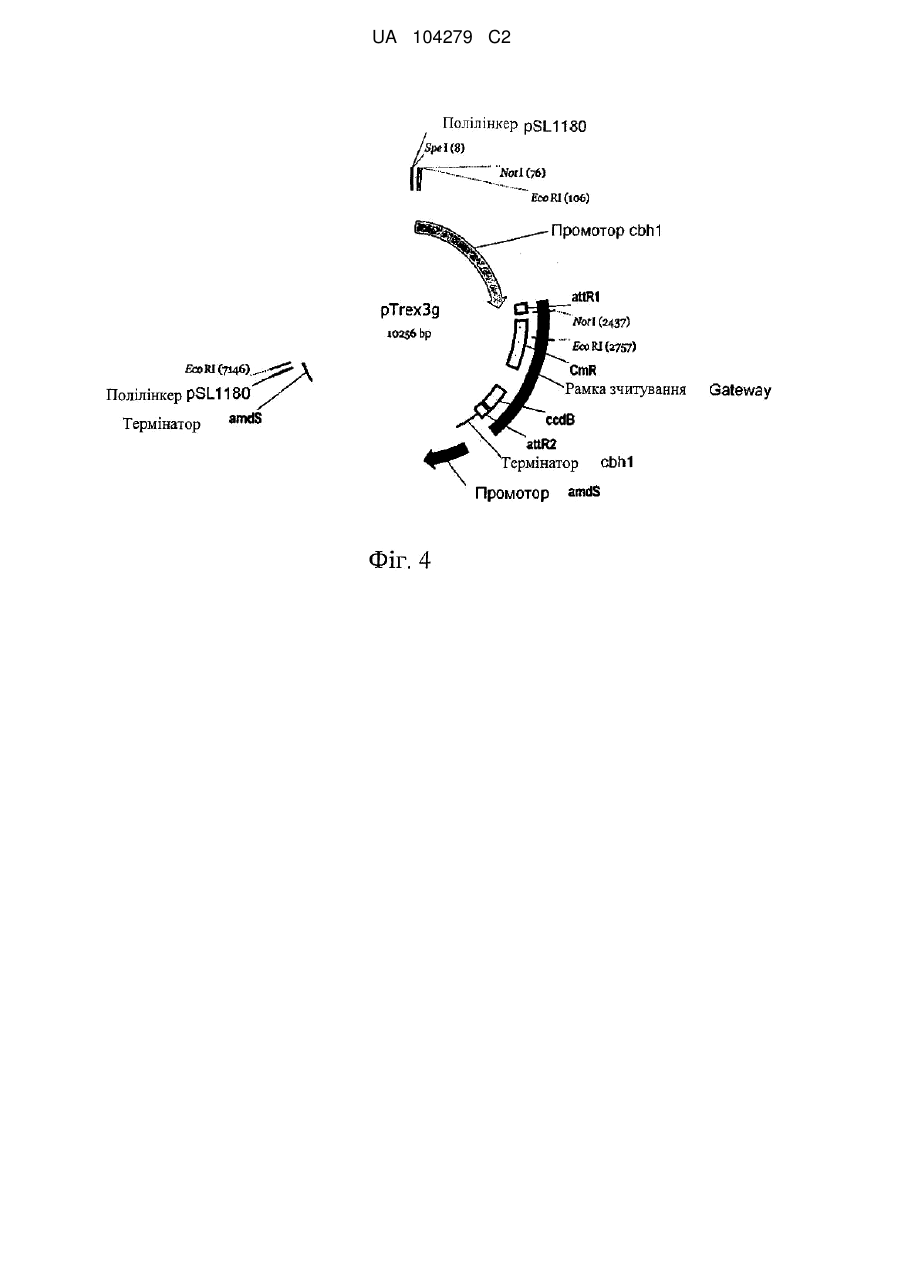






Формула / Реферат
1. Ниткоподібний гриб, який містить:
перший полінуклеотид, що кодує перший гетерологічний поліпептид,
другий полінуклеотид, що кодує другий гетерологічний поліпептид, і
третій полінуклеотид, що кодує гомологічний поліпептид,
при цьому вказаний ниткоподібний гриб може експресувати перший і другий гетерологічні поліпептиди і гомологічний поліпептид; і перший і другий гетерологічні поліпептиди і гомологічний поліпептид утворюють функціонально активну суміш целюлаз.
2. Ниткоподібний гриб за п. 1, в якому перший полінуклеотид функціонально зв'язаний з першим промотором.
3. Ниткоподібний гриб за п. 1, в якому другий полінуклеотид злитий з третім полінуклеотидом, при цьому другий і третій полінуклеотиди функціонально зв'язані з другим промотором.
4. Ниткоподібний гриб за п. 1, в якому перший полінуклеотид функціонально зв'язаний з промотором, нативним для гена, що кодує гомологічний поліпептид.
5. Ниткоподібний гриб за п. 1, в якому другий полінуклеотид злитий з третім полінуклеотидом, при цьому третій полінуклеотид функціонально зв'язаний з промотором гена, що кодує гомологічний поліпептид.
6. Ниткоподібний гриб за п. 1, в якому другий полінуклеотид злитий з третім полінуклеотидом з утворенням полінуклеотиду, що кодує злитий білок, при цьому злитий білок включає другий гетерологічний поліпептид і гомологічний поліпептид, розділені лінкером.
7. Ниткоподібний гриб за п. 6, в якому злитий білок далі включає сайт розщеплення.
8. Ниткоподібний гриб за п. 1, який далі включає четвертий полінуклеотид, що кодує маркер, який селектується.
9. Ниткоподібний гриб за п. 1, який далі включає четвертий полінуклеотид, що кодує третій гетерологічний поліпептид, при цьому вказаний ниткоподібний гриб може експресувати третій гетерологічний поліпептид.
10. Ниткоподібний гриб за п. 1, в якому перший гетерологічний поліпептид є модифікованим гомологічним поліпептидом.
11. Ниткоподібний гриб за п. 1, який далі включає четвертий полінуклеотид, що кодує третій гетерологічний поліпептид, при цьому перший і другий гетерологічні поліпептиди є модифікованими гомологічними поліпептидами.
12. Ниткоподібний гриб за п. 1, в якому перший гетерологічний поліпептид, другий гетерологічний поліпептид або гомологічний поліпептид є ферментом.
13. Ниткоподібний гриб за п. 1, в якому перший гетерологічний поліпептид, другий гетерологічний поліпептид або гомологічний поліпептид є целюлазою.
14. Ниткоподібний гриб за п. 1, в якому функціонально активна суміш є сумішшю целюлаз.
15. Ниткоподібний гриб за п. 1, в якому перший гетерологічний поліпептид, другий гетерологічний поліпептид або гомологічний поліпептид є целюлазою, яку вибирають з групи, що складається з екзоцелобіогідролаз, ендоглюканаз і бета-глюкозидаз.
16. Ниткоподібний гриб за п. 1, в якому перший гетерологічний поліпептид є екзоцелобіогідролазою і другий гетерологічний поліпептид є ендоглюканазою.
17. Ниткоподібний гриб за п. 1, в якому перший гетерологічний поліпептид є екзоцелобіогідролазою, яка вибирається з групи, що складається з сімейства GH 5, 6, 7, 9 і 48, і другий гетерологічний поліпептид є ендоглюканазою, яка вибирається з групи, що складається з сімейства GH 5, 6, 7, 8, 9, 12, 17, 31, 44, 45, 48, 51, 61, 64, 74 і 81.
18. Ниткоподібний гриб за п. 1, в якому перший гетерологічний поліпептид є екзоцелобіогідролазою, другий гетерологічний поліпептид є ендоглюканазою і гомологічний поліпептид є екзоцелобіогідролазою.
19. Ниткоподібний гриб за п. 1, в якому перший гетерологічний поліпептид є першою екзоцелобіогідролазою, другий гетерологічний поліпептид є ендоглюканазою і гомологічний поліпептид є другою екзоцелобіогідролазою, при цьому перша екзоцелобіогідролаза і друга екзоцелобіогідролаза є однаковими целобіогідролазами.
20. Ниткоподібний гриб за п. 1, який вибраний з групи, що складається з Aspergillus, Acremonium, Aureobasidium, Beauveria, Cеphalosporium, Ceriporiopsis, Chaetomium, Paecilomyces, Chrysosporium, Claviceps, Cochiobolus, Cryptococcus, Cyathus, Endothia, Fusarium, Gilocladium, Humicola, Magnaporthe, Myceliophthora, Myrothecium, Mucor, Neurospora, Phanerochaete, Podospora, Paecilomyces, Penicillium, Pyricularia, Rhizomucor, Rhizopus, Schizophylum, Stagonospora, Talaromyces, Trichoderma, Thermomyces, Thermoascus, Thielavia, Tolypocladium, Trichophyton, Trametes і Pleurotus.
21. Ниткоподібний гриб за п. 1, який є Т. reesei і в якому перший гетерологічний поліпептид є CBHI Humicola grisea, другий гетерологічний поліпептид є ендоглюканазою 1 Acidothermus cellulolyticus і гомологічний поліпептид є CBHI Trichoderma reesei.
22. Ниткоподібний гриб за п. 1, який є Т. reesei і в якому перший гетерологічний поліпептид або другий гетерологічний поліпептид вибраний з групи, що складається з целобіогідролази CBHI Penicillium funiculosum, ендоглюканаз Е3 Thermobifida, ендоглюканаз Е5 Thermobifida, GH74-core і GH48 Acidothermus cellulolyticus.
23. Ниткоподібний гриб за п. 1, який далі включає четвертий полінуклеотид, що кодує третій гетерологічний поліпептид, при цьому перший поліпептид є модифікованою CBHI Trichoderma reesei, другий гетерологічний поліпептид є модифікованою CBHII Trichoderma reesei, третій гетерологічний поліпептид є ендоглюканазою 1 Acidothermus cellulolyticus і гомологічний поліпептид є CBHI Trichoderma reesei.
24. Ниткоподібний гриб за п. 1, в якому перший гетерологічний поліпептид є екзоцелобіогідролазою, другий гетерологічний поліпептид є ендоглюканазою і гомологічний поліпептид є екзоцелобіогідролазою, при цьому експресія першого гетерологічного поліпептиду, другого гетерологічного поліпептиду і гомологічного поліпептиду утворює суміш термостійких целюлаз.
25. Ниткоподібний гриб за п. 1, в якому третій полінуклеотид є позахромосомним полінуклеотидом.
26. Ниткоподібний гриб за п. 1, в якому перший, другий і третій полінуклеотиди є позахромосомними полінуклеотидами.
27. Культуральне середовище, що включає популяцію ниткоподібного гриба за п. 1.
28. Функціональна суміш целюлаз, що включає перший гетерологічний поліпептид, другий гетерологічний поліпептид і гомологічний поліпептид, одержані з ниткоподібного гриба за п. 1.
29. Поліпептидна суміш за п. 28, яка є сумішшю целюлаз.
30. Спосіб одержання суміші целюлаз, який включає одержання поліпептидної суміші з ниткоподібного гриба за п. 1, при цьому поліпептидна суміш включає перший гетерологічний поліпептид, другий гетерологічний поліпептид і гомологічний поліпептид.
31. Спосіб одержання суміші целюлаз, який включає одержання поліпептидної суміші з ниткоподібного гриба за п. 1, при цьому поліпептидна суміш включає перший гетерологічний поліпептид, другий гетерологічний поліпептид і гомологічний поліпептид, з яких перший гетерологічний поліпептид є екзоцелобіогідролазою, другий гетерологічний поліпептид є ендоглюканазою і гомологічний поліпептид є екзоцелобіогідролазою.
32. Спосіб одержання суміші целюлаз, який включає одержання поліпептидної суміші з ниткоподібного гриба за п. 1, при цьому поліпептидна суміш включає перший гетерологічний поліпептид, другий гетерологічний поліпептид і гомологічний поліпептид, з яких перший гетерологічний поліпептид є першою екзоцелобіогідролазою, другий гетерологічний поліпептид є ендоглюканазою і гомологічний поліпептид є другою екзоцелобіогідролазою, де перша екзоцелобіогідролаза і друга екзоцелобіогідролаза є однаковими целобіогідролазами.
33. Спосіб одержання суміші целюлаз, який включає одержання поліпептидної суміші з ниткоподібного гриба за п. 1, при цьому поліпептидна суміш включає перший гетерологічний поліпептид, другий гетерологічний поліпептид і гомологічний поліпептид, де ниткоподібний гриб є Т. reesei, перший гетерологічний поліпептид є CBHI Humicola grisea, другий гетерологічний поліпептид є ендоглюканазою 1 Acidothermus cellulolyticus і гомологічний поліпептид є CBHI Trichoderma reesei.
34. Спосіб одержання суміші целюлаз, який включає одержання поліпептидної суміші з ниткоподібного гриба за п. 23, при цьому поліпептидна суміш включає перший гетерологічний поліпептид, другий гетерологічний поліпептид, третій гетерологічний поліпептид і гомологічний поліпептид.
35. Ниткоподібний гриб за п. 1, де перший гетерологічний поліпептид, другий гетерологічний поліпептид або гомологічний поліпептид є ксиланазою.
36. Ниткоподібний гриб за п. 1, де перший гетерологічний поліпептид, другий гетерологічний поліпептид або гомологічний поліпептид є ендоглюканазою.
37. Ниткоподібний гриб за п. 1, де ниткоподібний гриб експресує член сімейства GH61.
38. Ниткоподібний гриб за п. 1, де перший гетерологічний поліпептид, другий гетерологічний поліпептид або гомологічний поліпептид кожний є целюлазою, і де кожний поліпептид незалежно вибраний з групи, яка складається з екзоцелобіогідролази, ендоглюканази та бета-глюкозідази.
39. Ниткоподібний гриб за п. 1, де перший гетерологічний поліпептид є CHBI Trichoderma reesei, другий гетерологічний поліпептид є CHBII Trichoderma reesei та гомологічний поліпептид є CHBI Trichoderma reesei.
40. Поліпептидна суміш за п. 28, де поліпептидна суміш є функціональною сумішшю.
41. Поліпептидна суміш за п. 40, яка не містить жодних бактеріальних ферментів у комбінації з білком-носієм ниткоподібного гриба і/або де функціональна суміш не утворює антитіла або функціонального фрагмента антитіла.
42. Поліпептидна суміш за п. 40, яка демонструє покращену целюлазну активність, активність оцукрювання, термостійкість, зміну значень pH, стабільну активність протягом більш тривалих періодів часу при постійній температурі.
43. Поліпептидна суміш за п. 42, де поліпептидна суміш є функціональною сумішшю, яка демонструє покращену целюлазну активність.
44. Поліпептидна суміш за п. 42, де поліпептидна суміш є функціональною сумішшю, яка демонструє покращену активність оцукрювання.
45. Ниткоподібний гриб, що містить два або більше гетерологічні поліпептиди і гомологічний поліпептид, де ниткоподібний гриб здатний експресувати гетерологічні поліпептиди та гомологічні поліпептиди, і де гетерологічні поліпептиди та гомологічний поліпептид утворюють функціональну суміш целюлаз.
46. Ниткоподібний гриб за п. 45, який не містить жодних бактеріальних ферментів у комбінації з білком-носієм ниткоподібного гриба і/або де функціональна суміш не утворює антитіла або функціонального фрагмента антитіла.
47. Рекомбінантний ниткоподібний гриб, який генетично модифікований таким чином, що експресує суміш гетерологічних і гомологічних поліпептидів, які утворюють функціонально активну суміш целюлаз.
48. Рекомбінантний ниткоподібний гриб за п. 47, який продукує функціональну суміш.
49. Рекомбінантний ниткоподібний гриб за п. 47, який генетично модифікований таким чином, що експресує два або більше гетерологічні поліпептиди і гомологічний поліпептид.
50. Рекомбінантний ниткоподібний гриб за п. 49, який продукує функціональну суміш.
51. Рекомбінантний ниткоподібний гриб за п. 50, де функціональна суміш є функціональною сумішшю целюлаз.
52. Рекомбінантний ниткоподібний гриб за п. 51, де функціональна суміш має функцію, яка походить від двох або трьох поліпептидів суміші.
53. Рекомбінантний ниткоподібний гриб за п. 49, який генетично модифікований таким чином, що експресує три або більше гетерологічні поліпептиди і гомологічний поліпептид.
54. Рекомбінантний ниткоподібний гриб за п. 53, який продукує функціональну суміш.
55. Рекомбінантний ниткоподібний гриб за п. 54, де функціональна суміш є функціональною сумішшю целюлаз.
56. Рекомбінантний ниткоподібний гриб за п. 55, де функціональна суміш функціонально отримана із двох або трьох поліпептидів суміші.
57. Рекомбінантний ниткоподібний гриб за п. 49, який генетично модифікований таким чином, що експресує чотири або більше гетерологічні поліпептиди і гомологічний поліпептид.
58. Рекомбінантний ниткоподібний гриб за п. 57, який продукує функціональну суміш.
59. Рекомбінантний ниткоподібний гриб за п. 58, де функціональна суміш є функціональною сумішшю целюлаз.
60. Рекомбінантний ниткоподібний гриб за п. 59, де функціональна суміш функціонально отримана із двох, трьох або чотирьох поліпептидів суміші.
61. Рекомбінантний ниткоподібний гриб за п. 49, де гетерологічні поліпептиди та гомологічний поліпептид є целюлазами.
62. Рекомбінантний ниткоподібний гриб за п. 61, де кожна целюлаза незалежно вибрана із групи, яка складається з екзоцелобіогідролази, ендоглюканази та бета-глюкозидази.
63. Рекомбінантний ниткоподібний гриб за п. 62, який генетично модифікований таким чином, що експресує екзоцелобіогідролазу.
64. Рекомбінантний ниткоподібний гриб за п. 63, де екзоцелобіогідролаза є ферментом CBHI-типу.
65. Рекомбінантний ниткоподібний гриб за п. 64, де фермент CBHI-типу є варіантом CBHІ H. jecorіna.
66. Рекомбінантний ниткоподібний гриб за п. 63, де екзоцелобіогідролаза є ферментом CBHII-типу.
67. Рекомбінантний ниткоподібний гриб за п. 66, де фермент CBHII-типу є варіантом СВНІІ H. jecorіna.
68. Рекомбінантний ниткоподібний гриб за п. 62, який генетично модифікований таким чином, що експресує ендоглюканазу.
69. Рекомбінантний ниткоподібний гриб за п. 62, який генетично модифікований таким чином, що експресує бета-глюкозидазу.
70. Рекомбінантний ниткоподібний гриб за п. 47, який генетично модифікований таким чином, що експресує гетерологічну екзоцелобіогідролазу та гетерологічну ендоглюканазу.
71. Рекомбінантний ниткоподібний гриб за п. 70, де екзоцелобіогідролазою є GH5, GH6, GH7, GH9 або GH48, і де ендоглюканазою є GH5, GH6, GH7, GH8, GH9, GH12, GH17, GH31, GH44, GH45, GH48, GH51, GH61, GH64, GH74 або GH81.
72. Рекомбінантний ниткоподібний гриб за п. 47, який генетично модифікований таким чином, що експресує функціональну суміш поліпептидів, вибраних з EGІ T. reeseі, EGІІ T. reeseі, EGІІІ T. reeseі, EGІІІ H. grіsea, E5 T. fusca, E3 T. reeseі, E1 A. cellulolytіcus і GH74 T. reeseі.
73. Рекомбінантний ниткоподібний гриб за п. 49, де гетерологічні поліпептиди є екзоцелобіогідролазою і ендоглюканазою, і де гомологічний поліпептид є екзоцелобіогідролазою.
74. Рекомбінантний ниткоподібний гриб за п. 49, де щонайменше один гетерологічний поліпептид і щонайменше один гомологічний поліпептид експресується як злитий поліпептид.
75. Рекомбінантний ниткоподібний гриб за п. 74, де вказаний гетерологічний поліпептид і вказаний гомологічний поліпептид розділені лінкером або лінкерною областю, де лінкер необов'язково є лінкером глюкоамілази Aspergіllus або лінкером CBHІ Trіchoderma.
76. Рекомбінантний ниткоподібний гриб за п. 74, де вказаний гетерологічний поліпептид і вказаний гомологічний поліпептид розділені лінкером або лінкерною областю та сайтом розщеплення, де сайт розщеплення необов'язково є сайтом розщеплення кексину, сайтом розпізнавання протеази трипсину або сайтом розпізнавання ендопротеїнази Lys-C.
77. Рекомбінантний ниткоподібний гриб за п. 45, який містить полінуклеотид, що кодує маркер, що селектується, де маркер, що селектується, необов'язково є маркером, що надає бактеріальну стійкість, pyr4 T. reeseі, ацетолактатсинтазою T. reeseі, hyg Streptomyces, amdS Aspergіllus nіdulans або pyrG Aspergіllus nіger.
78. Рекомбінантний ниткоподібний гриб за п. 49, де щонайменше один гетерологічний поліпептид і щонайменше один гомологічний поліпептид не експресуються у вигляді злитого поліпептиду.
79. Рекомбінантний ниткоподібний гриб за п. 47, де гетерологічні або гомологічні поліпептиди кодуються полінуклеотидами, які функціонально зв'язані з одним або декількома промоторами.
80. Рекомбінантний ниткоподібний гриб за п. 79, де полінуклеотиди функціонально зв'язані з одним або декількома промоторами, нативними для ниткоподібного гриба.
81. Рекомбінантний ниткоподібний гриб за п. 79, де полінуклеотиди функціонально зв'язані з одним або декількома гетерологічними промоторами.
82. Рекомбінантний ниткоподібний гриб за п. 79, де полінуклеотиди експресовані під контролем конститутивного промотору.
83. Рекомбінантний ниткоподібний гриб за п. 79, де полінуклеотиди експресуються під контролем індуцибельного промотору.
84. Рекомбінантний ниткоподібний гриб за п. 79, де один або декілька промоторів вибрані із промотору целюлази, промотору ксиланази та промотору 1818.
85. Рекомбінантний ниткоподібний гриб за п. 79, де один або більше промоторів є промотором целюлази ниткоподібного гриба.
86. Рекомбінантний ниткоподібний гриб за п. 85, де промотор целюлази є промотором екзоцелобіогідролази, промотором ендоглюканази або промотором бета-глюкозидази.
87. Рекомбінантний ниткоподібний гриб за п. 86, де промотор є промотором cbh1.
88. Рекомбінантний ниткоподібний гриб за п. 79, де один або декілька промоторів вибрані із групи промоторів cbh1, cbh2, egl1, egl2, egl3, egl4, egl5, pkі1, gpd1, xyn1 або xyn2.
89. Рекомбінантний ниткоподібний гриб за п. 47, який генетично модифікований таким чином, що експресує целюлазу, геміцелюлазу, ксиланазу або мананазу.
90. Рекомбінантний ниткоподібний гриб за п. 47, який генетично модифікований таким чином, що експресує члени сімейств GH5, GH6, GH7, GH9 або GH48.
91. Рекомбінантний ниткоподібний гриб за п. 47, який генетично модифікований таким чином, що експресує члени сімейства GH5, GH6, GH7, GH8, GH9, GH12, GH17, GH31, GH44, GH45, GH48, GH51, GH61, GH64, GH74 або GH81.
92. Рекомбінантний ниткоподібний гриб за п. 91, який генетично модифікований таким чином, що експресує член сімейства GH61.
93. Рекомбінантний ниткоподібний гриб за п. 47, який генетично модифікований таким чином, що експресує члени сімейств GH1, GH3, GH9 або GH48.
94. Рекомбінантний ниткоподібний гриб за будь-яким з пп. 47-93, який вибраний з Aspergіllus, Acremonіum, Aureobasіdіum, Beauverіa, Cephalosporіum, Cerіporіopsіs, Chaetomіum, Paecіlomyces, Chrysosporіum, Clavіceps, Cochіobolus, Cryptococcus, Cyathus, Endothіa, Fusarіum, Gіlocladіum, Humіcola, Magnaporthe, Mycelіophthora, Myrothecіum, Mucor, Neurospora, Phanerochaete, Podospora, Paecіlomyces, Penіcіllіum, Pyrіcularіa, Rhіzomucor, Rhіzopus, Schіzophylum, Stagonospora, Talaromyces, Trіchoderma, Thermomyces, Thermoascus, Thіelavіa, Tolypocladіum, Trіchophyton, Trametes і Pleurotus.
95. Спосіб одержання комбінації гетерологічних і гомологічних поліпептидів, що передбачає культивування рекомбінантного ниткоподібного гриба за п. 94.
96. Культуральне середовище, що містить ниткоподібний гриб за п. 94.
97. Функціональна суміш целюлаз, що містить гетерологічні та гомологічні поліпептиди, що експресуються рекомбінантним ниткоподібним грибом за будь-яким з пп. 48, 50, 54 і 58.
98. Функціональна суміш за п. 97, яка демонструє покращені властивості і/або активність, або де функція вказаної функціональної суміші покращена відносно активності, пов'язаної з ниткоподібним грибом або забезпечуваної ниткоподібним грибом.
99. Функціональна суміш за п. 98, де покращеними властивостями, активністю або функцією є покращена целюлазна активність, покращена активність оцукрювання, покращена термостійкість, зміна значень pH або стабільна активність протягом більш тривалих періодів часу при постійній температурі.
100. Функціональна суміш за п. 99, де покращеними властивостями, активністю або функцією є покращена целюлазна активність.
101. Функціональна суміш за п. 99, де покращеними властивостями, активністю або функцією є покращена активність оцукрювання.
102. Функціональна суміш за п. 99, яка містить целюлази, геміцелюлази, ксиланази та мананази.
103. Функціональна суміш за п. 99, яка містить целюлазу, геміцелюлазу, ксиланазу або мананазу.
104. Функціональна суміш за п. 99, яка є функціональною целюлазною сумішшю.
Текст
Реферат: Даний винахід стосується ниткоподібних грибів, які коекспресують комбінацію гетерологічних і гомологічних поліпептидів, які утворюють функціонально-активну суміш целюлаз. Винахід також належить до функціональних сумішей целюлаз та способів одержання таких сумішей. UA 104279 C2 (12) UA 104279 C2 UA 104279 C2 5 10 15 20 25 30 35 40 45 50 55 1. Перехресне посилання на споріднену заявку За даною заявкою заявляється про пріоритет попередньої заявки на патент США № US 60/933894, поданої 8 червня 2007 р., яка повністю включена у даний опис винаходу як посилання. 2. Заява про фінансування наукової роботи на федеральному рівні Деякі частини даної наукової роботи фінансувалися Національною лабораторією відновлюваної енергії за субдоговором № ZC0-0-30017-01 на основі основного договору № DEAC36-99G010337, укладеного з Міністерством енергетики США. Таким чином, уряд США може мати певні права на винахід. 3. Вступ Целюлоза і геміцелюлоза є найбільш поширеними рослинними речовинами, які утворюються внаслідок фотосинтезу. Вказані речовини можуть розщеплюватися і використовуватися як джерело енергії багатьма мікроорганізмами, включаючи бактерії, дріжджі і гриби, які продукують позаклітинні ферменти, здатні гідролізувати полімерні субстрати у мономерні цукри. Целюлази є ферментами, які гідролізують целюлозу (бета-1,4-глюканові або бета-Dглікозидні зв'язки) з утворенням глюкози, целобіози, целоолігосахаридів і тому подібних. Целюлази традиційно розділені на три основних класи: ендоглюканази (ЄС 3.2.1.4) ("EG"), екзоглюканази або целобіогідролази (ЄС 3.2.1.91) ("CBH") і бета-глюкозидази ([бета]-Dглюкозид-глюкогідролаза; ЄС 3.2.1.21) ("BG"). Ендоглюканази діють в основному в аморфних частинах целюлозного волокна, у той час як целобіогідролази здатні також розщеплювати кристалічну целюлозу. Для ефективного перетворення кристалічної целюлози у глюкозу необхідна повна система целюлаз, що включає компоненти СВН, EG і BG, оскільки окремо виділені компоненти менш ефективно гідролізують кристалічну целюлозу (Filho et al., Can. J. Microbiol. 42:1-5, 1996). Досить перспективною є експресія вказаних багатокомпонентних систем целюлаз у ниткоподібних грибах для промислового виробництва целюлаз. 4. Суть винаходу Таким чином, даний винахід стосується ниткоподібних грибів, які експресують комбінацію гетерологічних і гомологічних поліпептидів, поліпептидних сумішей, що включають комбінацію гетерологічних і гомологічних поліпептидів, і способів одержання поліпептидних сумішей. Деякі варіанти здійснення даного винаходу стосуються ниткоподібного гриба, що включає два або більше полінуклеотидів, які кодують два або більше гетерологічних поліпептидів, і полінуклеотид, що кодує гомологічний поліпептид. Ниткоподібний гриб може експресувати гетерологічні і гомологічні поліпептиди, які разом утворюють функціонально активну суміш. Деякі варіанти здійснення даного винаходу стосуються культурального середовища, що включає популяцію ниткоподібного гриба за даним винаходом. Деякі варіанти здійснення даного винаходу стосуються поліпептидної суміші, яка включає два або більше гетерологічних поліпептидів і гомологічний поліпептид. Поліпептидна суміш може бути одержана з ниткоподібних грибів за даним винаходом. Деякі варіанти здійснення даного винаходу стосуються способу одержання суміші целюлаз. Вказаний спосіб включає одержання поліпептидної суміші, що містить два або більше гетерологічних поліпептидів і гомологічний поліпептид, з ниткоподібного гриба за даним винаходом. У деяких варіантах здійснення винаходу гетерологічні поліпептиди є екзоцелобіогідролазою і ендоглюканазою, і гомологічний поліпептид є екзоцелобіогідролазою. Гетерологічна екзоцелобіогідролаза і гомологічна екзоцелобіогідролаза можуть бути, але не обов'язково є однаковими екзоцелобіогідролазами. Нижче представлені згадані вище та інші ознаки даного винаходу. 5. Короткий опис креслень Фахівцю повинно бути зрозуміло, що креслення наведені тільки з метою ілюстрації. Представлені креслення жодним чином не обмежують обсяг даного винаходу. На фігурі 1 показана нуклеотидна послідовність (SEQ ID NO:1) злитої конструкції гетерологічної целюлази, що включає 2656 основ. На фігурі 2 показана амінокислотна послідовність (SEQ ID NO:2) злитого білка целюлази, яка прогнозується на основі послідовності нуклеїнової кислоти, зображеної на фігурі 1. На фігурах 3А-F показана нуклеотидна послідовність (SEQ ID NO:14) вектора pTrex4, що містить каталітичний домен Е1. На фігурі 4 показана плазмідна карта експресуючого вектора pTrex3g T.reesei. На фігурі 5А показаний експресуючий вектор pTrex3g-Hgrisea-cbh1, що використовується для одержання типового трьохкомпонентного штаму. 1 UA 104279 C2 5 10 15 20 25 30 35 40 45 50 55 60 На фігурах 5В-Е показана нуклеотидна послідовність (SEQ ID NO:7) експресуючого вектора, зображеного на фігурі 5А. На фігурі 6 показано три ДНК-експресуючих фрагменти, трансфіковані у штам з делецією cbh1 для створення чотирьохкомпонентного штаму. На фігурі 7А показана нуклеотидна послідовність (SEQ ID NO:8) від ініціюючого кодону до термінуючого кодону полінуклеотиду, що експресує генетично створений білок CBHI. На фігурі 7В показана послідовність генетично створеного білка CBHI (SEQ ID NO:9). Сигнальна послідовність CBHI підкреслена. На фігурі 8А показаний cbhl-експресуючий вектор pTrex3g-cbhl. На фігурах 8В-F показана нуклеотидна послідовність (SEQ ID NO:10) експресуючого вектора pTrex3g-cbhl. На фігурі 9А показана нуклеотидна послідовність (SEQ ID NO:11) від ініціюючого кодону до термінуючого кодону полінуклеотиду, що експресує генетично створений білок CBHI. На фігурі 9В показана амінокислотна послідовність генетично створеного білка CBНII (SEQ ID NO:12). Сигнальна послідовність підкреслена. На фігурі 10А показаний cbhII-експресуючий вектор pExp-cbhII. На фігурах 10В-G показана нуклеотидна послідовність (SEQ ID NO:13) експресуючого вектора pExp-cbhII. 6. Докладний опис різних варіантів здійснення винаходу Далі буде наданий докладний опис винаходу з використанням наведених нижче визначень термінів і прикладів. За винятком особливо обумовлених випадків, всі технічні і наукові терміни, використані у даному описі винаходу, мають загальноприйняті значення, відомі фахівцю у науковій галузі, до якої належить даний винахід. Числові інтервали охоплюють всі числа, вказані у даному інтервалі. Заголовки, наведені у даному описі винаходу, не обмежують різні об'єкти і варіанти здійснення винаходу і можуть розглядатися як такі, що стосуються опису винаходу загалом. Таким чином, наведені нижче терміни мають більш повне визначення, що стосується всього опису винаходу. Термін "поліпептид", що використовується у даному описі, означає сполуку, що складається з одного ланцюга амінокислотних залишків, зв'язаних пептидними зв'язками. Термін "білок", що використовується в даному описі, має взаємозамінне значення з терміном "поліпептид". Термін "нуклеїнова кислота" і "полінуклеотид" мають взаємозамінні значення і означають одноланцюжкову або двохланцюжкову ДНК, РНК, кДНК і їх хімічні модифікації. Оскільки генетичний код є виродженим, для кодування визначеної амінокислоти може бути використано декілька кодонів, і в обсяг даного винаходу входять всі полінуклеотиди, що кодують визначену амінокислотну послідовність. Термін "рекомбінантний", що використовується застосовно до клітини, нуклеїнової кислоти, білка або вектора, означає, що вказана клітина, нуклеїнова кислота, білок або вектор модифіковані шляхом введення гетерологічної нуклеїнової кислоти або білка, або зміни нативної нуклеїнової кислоти або білка, що дана клітина одержана з модифікованої таким чином клітини або що білок експресований у ненативному або генетично модифікованому середовищі, наприклад, в експресуючому векторі для прокаріотичної або еукаріотичної системи. Таким чином, наприклад, рекомбінантні клітини експресують нуклеїнові кислоти або поліпептиди, що не зустрічаються у природній (нерекомбінантній) формі клітини, або експресують нативні гени, які в іншому випадку були б анормально експресовані, недостатньо експресовані, надекспресовані або взагалі не експресовані. Термін "гетерологічний" застосовно до полінуклеотиду або поліпептиду означає полінуклеотид або поліпептид, що має послідовність, яка у природних умовах відсутня у клітиніхазяїні. В одних варіантах здійснення винаходу поліпептид є комерційно важливим промисловим білком, і в інших варіантах здійснення винаходу гетерологічний поліпептид є терапевтичним білком. Передбачається, що у визначення даного терміну входять білки, кодовані природними генами, мутованими генами і/або синтезованими генами. Термін "гомологічний" застосовно до полінуклеотиду або поліпептиду означає полінуклеотид або поліпептид, що має послідовність, яка природно присутня у клітині-хазяїні. У використовуваному у даному випадку значенні термін "злита нуклеїнова кислота" означає дві або більше число нуклеїнових кислот, функціонально зв'язаних одна з одною. Нуклеїнова кислота може бути ДНК, геномною ДНК і кДНК, РНК або гібридом РНК і ДНК. У конструкції послідовностей злитої нуклеїнової кислоти може бути використана нуклеїнова кислота, що кодує всю або частину послідовності поліпептиду. В одних варіантах здійснення винаходу використана нуклеїнова кислота, що кодує непроцесовані поліпептиди. В інших варіантах здійснення винаходу може бути використана нуклеїнова кислота, що кодує частину поліпептиду. 2 UA 104279 C2 5 10 15 20 25 30 35 40 45 50 55 Термін "злитий поліпептид" означає білок, що включає щонайменше дві окремих і різних області, які можуть бути одержані з одного і того ж білка або з різних білків. Наприклад, злитим поліпептидом або злитим білком може вважатися білок, в якому сигнальний пептид, пов'язаний з білком, що представляє інтерес, звичайно не асоційований з вказаним білком, що представляє інтерес. Терміни "виділений" і "відділений" мають взаємозамінні значення і означають білок, клітину, нуклеїнову кислоту, амінокислоту і т.д., видалені щонайменше з одного компонента, з яким вони асоційовані у природних умовах. У використовуваному у даному випадку значенні термін "ген" означає полінуклеотид (наприклад, сегмент ДНК), що бере участь в утворенні поліпептидного ланцюга, який може включати або не включати області, які передують або йдуть за кодуючою областю, наприклад, 5'-кінцеві нетрансльовані (5'UTR) або "лідерні" послідовності і 3'UTR або "трейлерні" послідовності, а також вставні послідовності (інтрони) між окремими кодуючими сегментами (екзонами). У використовуваному у даному випадку значенні термін "промотор" означає послідовність нуклеїнової кислоти, яка направляє транскрипцію гена нижньої області. Промотор звичайно повинен відповідати клітині-хазяїну, в якій експресований ген-мішень. Промотор разом з іншими послідовностями нуклеїнової кислоти, що регулюють транскрипцію і трансляцію (які іменують також "регуляторними послідовностями"), необхідний для експресії даного гена. Як правило, послідовності, що регулюють транскрипцію і трансляцію, включають, не обмежуючись ними, промоторні послідовності, сайти скріплення рибосом, послідовності, що ініціюють і термінують транскрипцію, послідовності, що ініціюють і термінують трансляцію, енхансерні або активаторні послідовності. У використовуваному у даному випадку значенні термін "функціонально зв'язаний" означає, що нуклеїнова кислота, що регулює транскрипцію, знаходиться по відношенню до кодуючих послідовностей у положенні, що забезпечує ініціацію транскрипції. Як правило, це означає, що промотор і послідовності, що ініціюють транскрипцію, розташовані у 5' напрямі відносно кодуючої області. Нуклеїнова кислота, що регулює транскрипцію, звичайно повинна відповідати клітині-хазяїну, що використовується для експресії вказаного білка. У даній галузі відомі відповідні експресуючі вектори і прийнятні регуляторні послідовності багатьох типів для різних клітин-хазяїнів. У використовуваному у даному випадку значенні термін "експресія" означає процес утворення поліпептиду на основі послідовності нуклеїнової кислоти гена. Вказаний процес включає транскрипцію і трансляцію. У використовуваному у даному випадку значенні термін "вектор" означає полінуклеотидну конструкцію, створену для введення нуклеїнових кислот у клітини одного або декількох типів. Вектори включають клонуючі вектори, експресуючі вектори, шатл-вектори, плазміди, кластери і тому подібні. У використовуваному у даному випадку значенні термін "експресуючий вектор" означає вектор, здатний включати і експресувати фрагменти гетерологічної ДНК у чужорідній клітині. Багато прокаріотичних і еукаріотичних експресуючих векторів є комерційно доступними. У використовуваному у даному випадку значенні терміни "векторна ДНК", "трансформуюча ДНК" і "експресуючий вектор" мають взаємозамінні значення і стосуються ДНК, що використовується для введення послідовностей у клітину-хазяїна або організм. Вказана ДНК може бути створена in vitro за допомогою полімеразної ланцюгової реакції (ПЛР) або будь-яких інших прийнятних методів, відомих у даній галузі, наприклад, за допомогою стандартних методів молекулярної біології, описаних у публікації Sambrook et al. Крім того, ДНК експресуючої конструкції може бути синтезована штучно, наприклад, хімічним шляхом. Векторна ДНК, трансформуюча ДНК або рекомбінантний експресуючий кластер можуть бути введені у плазміду, хромосому, позахромосомний елемент, мітохондріальну ДНК, плазмідну ДНК, вірус або фрагмент нуклеїнової кислоти. Рекомбінантний експресуючий кластер експресуючого вектора, векторної ДНК або трансформуючої ДНК звичайно включає нарівні з іншими послідовностями послідовність нуклеїнової кислоти, що транскрибується, і промотор. У переважних варіантах здійснення винаходу експресуючі вектори здатні вводити і експресувати фрагменти гетерологічної ДНК у клітині-хазяїні. Термін "введений" застосовно до введення послідовності нуклеїнової кислоти у клітину означає "трансфекцію", "трансформацію" або "трансдукцію" і стосується введення послідовності нуклеїнової кислоти в еукаріотичну або прокаріотичну клітину, в якій дана послідовність нуклеїнової кислоти може бути вбудована у геном клітини (наприклад, у хромосому, 3 UA 104279 C2 5 10 15 20 25 30 35 40 45 50 55 60 позахромосомний елемент, плазміду, пластиду або мітохондріальну ДНК), перетворена в автономний реплікон або тимчасово експресована (наприклад, трансфікована мРНК). Термін "клітина-хазяїн" означає клітину, яка містить вектор і підтримує реплікацію і/або транскрипцію, і трансляцію (експресію) експресуючої конструкції. У використовуваному у даному випадку значенні термін "культивування" означає вирощування популяції клітин у прийнятних умовах у рідкому, напівтвердому або густому середовищі. У використовуваному у даному випадку значенні терміни "заміщений" і "модифікований" мають взаємозамінні значення і стосуються послідовності, такої як амінокислотна послідовність або послідовність нуклеїнової кислоти, яка включає делецію, інсерцію, заміну або переривання природної послідовності. Часто у контексті даного винаходу заміщена послідовність включає заміну природного залишку. У використовуваному у даному випадку значенні термін "модифікований фермент" означає фермент, який включає делецію, інсерцію, заміну або переривання природної послідовності. Термін "варіант" означає область білка, що містить одну або декілька інших амінокислот у порівнянні з вихідним білком, наприклад, природним білком або білком дикого типу. Термін "целюлоза" означає категорію ферментів, здатних гідролізувати полімери целюлози (бета-1,4-глюканові або бета-D-глюкозидні зв'язки) з утворенням більш коротких целоолігосахаридних олігомерів, целобіози і/або глюкози. Термін "екзоцелобіогідролаза" (СВН) означає групу целюлазних ферментів, які класифікуються як ЄС 3.2.1.91, і/або ферментів у визначених сімействах GH, які включають, не обмежуючись ними, ферменти у сімействах GH 5, 6, 7, 9 або 48. Вказані ферменти відомі також як екзоглюканази або целобіогідролази. Ферменти СВН гідролізують целобіозу з кінця целюлози, що відновлюється або не відновлюється. Як правило, фермент типу CBHI переважно гідролізує целобіозу з кінця целюлози, що відновлюється, і фермент типу CBHII переважно гідролізує целобіозу з кінця целюлози, що не відновлюється. Термін "активність целобіогідролази" у використовуваному у даному випадку значенні означає активність 1,4-D-глюканцелобіогідролази, яка каталізує гідроліз 1,4-бета-D-глюкозидних зв'язків у целюлозі, целотетріозі або будь-якому полімері, що містить бета-1,4-зв'язану глюкозу, відщеплюючи целобіозу від кінців ланцюга. У використовуваному у даному випадку значенні активність целобіогідролази визначається вивільненням з целюлози водорозчинного цукру, що відновлюється, що вимірюється методом PHBAH, описаним у публікації Lever et al., 1972, Anal. Biochem. 47:273-279. Відмінність між впливом екзоглюканази, целобіогідролази і ендоглюканази визначають аналогічним чином на основі вивільнення цукру, що відновлюється, із заміщеної целюлози, такої як карбоксиметилцелюлоза або гідроксіетилцелюлоза (Ghose, 1987, Pure & Appl. Chem. 59:257-268). Справжня целобіогідролаза характеризується дуже високим рівнем активності відносно незаміщеної целюлози у порівнянні із заміщеною целюлозою (Bailey et al., 1993, Biotechnol. Appl. Biochem. 17:65-76). Термін "ендоглюканаза" (EG) означає групу целюлазних ферментів, які класифікуються як ЄС 3.2.1.4, і/або ферментів у визначених сімействах GH, які включають, не обмежуючись ними, ферменти у сімействах GH 5, 6, 7, 8, 9, 12, 17, 31, 44, 45, 48, 51, 61, 64, 74 або 81. Фермент EG гідролізує внутрішні бета-1,4-глюкозидні зв'язки целюлози. Термін "ендоглюканаза" у використовуваному у даному випадку значенні означає ендо-1,4-(1,3;1,4)-бета-D-глюкан-4глюканогідролазу, яка каталізує ендогідроліз 1,4-бета-D-глюкозидних зв'язків у целюлозі, похідних целюлози (наприклад, карбоксиметилцелюлозі), ліхеніні, бета-1,4-зв'язки у змішаних бета-1,3-глюканах, таких як бета-D-глюкани або ксилоглюкани злаків, і в інших рослинах, що містять целюлозні компоненти. У використовуваному у даному випадку значенні активність ендоглюканази визначають шляхом гідролізу карбоксиметилцелюлози (СМС) методом, описаним у публікації Ghose, 1987, Pure and Appl. Chem. 59:257-268. Термін "бета-глюкозидаза" у використовуваному у даному випадку значенні означає бета-Dглюкозид-глюкогідролазу, що класифікується як ЄС 3.2.1.21, і/або ферменти у визначених сімействах GH, які включають, не обмежуючись ними, ферменти у сімействах GH 1, 3, 9 або 48, що каталізують гідроліз целобіози з вивільненням бета-D-глюкози. У використовуваному у даному випадку значенні активність бета-глюкозидази може бути виміряна методами, відомими у даній галузі, наприклад, ВЕРХ. Термін "активність, що розкладає целюлозу" означає активність екзоглюканази, активність ендоглюканази або активність ферментів обох типів, а також активність бета-глюкозидази. Терміни "термічно стійкий" і "термостійкий" стосуються поліпептидів або ферментів за даним винаходом, які певною мірою зберігають біологічну, наприклад, ферментативну активність після впливу підвищеної температури, тобто температури, що вища кімнатної. У деяких варіантах 4 UA 104279 C2 5 10 15 20 25 30 35 40 45 50 55 здійснення винаходу поліпептид або фермент вважається термостійким при збереженні більше 50 %, 60 %, 70 %, 75 %, 80 %, 85 %, 90 %, 95 % або 98 % біологічної активності після впливу визначеної температури, наприклад, 40 °C, 45 °C, 50 °C, 55 °C, 60 °C, 65 °C, 70 °C, 75 °C або 80 °C, протягом 2, 5, 7, 10, 15, 20, 30, 40, 50 або 60 хвилин при рН, що дорівнює, наприклад, 4, 4,5, 5, 5,5, 6, 6,5, 7, 7,5 або 8. Термін "ниткоподібні гриби" означає будь-які, а також всі ниткоподібні гриби, відомі фахівцям у даній галузі. Як правило, ниткоподібні гриби є еукаріотичними мікроорганізмами і включають всі ниткоподібні форми підвідділу Eumycotina. Вказані гриби характеризуються наявністю вегетативного міцелію, клітинна оболонка якого складається з хітину, бета-глюкану та інших складних полісахаридів. У деяких варіантах здійснення винаходу ниткоподібні гриби за даним винаходом у морфологічному, фізіологічному і генетичному відношенні відрізняються від дріжджів. У деяких варіантах здійснення винаходу ниткоподібні гриби включають, не обмежуючись ними, наведені далі роди: Aspergillus, Acremonium, Aureobasidium, Beauveria, Cеphalosporium, Ceriporiopsis, Chaetomium paecilomyces, Chrysosporium, Claviceps, Cochiobolus, Cryptococcus, Cyathus, Endothia, Endotia mucor, Fusarium, Gilocladium, Humicola, Magnaporthe, Myceliophthora, Myrothecium, Mucor, Neurospora, Phanerochaete, Podospora, Paecilomyces, Penicillium, Pyricularia, Rhizomucor, Rhizopus, Schizophylum, Stagonospora, Talaromyces, Trichoderma, Thermomyces, Thermoascus, Thielavia, Tolypocladium, Trichophyton і Trametes pleurotus. У деяких варіантах здійснення винаходу ниткоподібні гриби включають, не обмежуючись ними, наведені далі види: А. nidulans, А. niger, А. awomari, наприклад, NRRL 3112, ATCC 22342 (NRRL 3112), ATCC 44733, ATCC 14331 і штам UVK 143f, А. oryzae, наприклад, АТСС 11490, N. crassa, Trichoderma reesei, наприклад, NRRL 15709, АТСС 13631, 56764, 56765, 56466, 56767, і Trichoderma viride, наприклад, АТСС 32098 і 32086. Термін "Trichoderma" або "вид Trichoderma" у використовуваному тут значенні означає будьякі гриби, які раніше були класифіковані як вид або штам Trichoderma і які у наш час класифіковані як вид або штам Trichoderma або як вид або штам Hypocrea. У деяких варіантах здійснення винаходу вказаний вид включає Trichoderma longibrachiatum, Trichoderma reesei, Trichoderma viride або Hypocrea jecorina. Як вихідний штам можуть бути також використані штами, які надпродукують целюлазу, такі як Т. longibrachiatum/reesei RL-P37 (Sheir-Neiss et al., Appl. Microbiol. Biotechnology, 20 (1984) pp. 46-53; Montenecourt B.S., Can., 1-20, 1987), і штам Rut-C30. У деяких варіантах здійснення винаходу продукування целюлаз у видах, призначених для поліпшення, строго регулюється і є чутливим до різних оточуючих умов. Даний винахід стосується ниткоподібного гриба, що включає два або більше полінуклеотидів, які кодують два або більше гетерологічних поліпептидів, і полінуклеотид, що кодує гомологічний поліпептид. Ниткоподібний гриб може експресувати гетерологічні і гомологічні поліпептиди, які утворюють функціонально активну суміш. У деяких варіантах здійснення винаходу ниткоподібний гриб містить перший полінуклеотид і другий полінуклеотид, що кодують відповідно перший гетерологічний поліпептид і другий гетерологічний поліпептид, а також третій полінуклеотид, що кодує гомологічний поліпептид. У деяких варіантах здійснення винаходу ниткоподібний гриб містить додатковий полінуклеотид, четвертий полінуклеотид, що кодує третій гетерологічний поліпептид. У деяких варіантах здійснення винаходу ниткоподібний гриб містить чотири або більше полінуклеотидів, що кодують чотири або більше гетерологічних поліпептидів, і один або декілька полінуклеотидів, що кодують один або декілька гомологічних поліпептидів. Відповідно до даного винаходу функціонально активна суміш включає будь-яку суміш поліпептидів за умови, що така суміш виконує щонайменше одну функцію, біологічну або якунебудь ще, яка є наслідком активності щонайменше двох або трьох поліпептидів у суміші. Іншими словами, щонайменше два або три поліпептиди у вказаній суміші забезпечують активність поліпептидної суміші на детектованому рівні. У деяких варіантах здійснення винаходу функціонально активна суміш включає щонайменше три поліпептиди і має функціональну активність, що забезпечується щонайменше двома або трьома поліпептидами у вказаній суміші. У деяких інших варіантах здійснення винаходу функціонально активна суміш включає щонайменше три поліпептиди і виконує ферментативну функцію, здійснювану щонайменше двома або трьома поліпептидами у вказаній суміші. У деяких варіантах здійснення винаходу функціонально активна суміш включає щонайменше три поліпептиди і виконує функцію целюлази, що забезпечується щонайменше двома або трьома поліпептидами у вказаній суміші. У деяких варіантах здійснення винаходу функціонально активна суміш включає чотири поліпептиди і виконує функцію, що забезпечується двома, трьома або чотирма поліпептидами у вказаній суміші. 5 UA 104279 C2 5 10 15 20 25 30 35 40 45 50 55 60 У деяких варіантах здійснення винаходу функціонально активна суміш виконує функцію, що відповідає поліпшенню будь-якої активності, наприклад, активності білка, що секретується, що включає без яких-небудь обмежень активність целюлази, активність оцукрювання або термостійкість, і асоційована з ниткоподібним грибом або забезпечується вказаним грибом. У деяких варіантах здійснення винаходу функціонально активна суміш виконує функцію, що забезпечується активністю екзоцелобіогідролаз, ендоглюканаз, бета-глюкозидаз або будь-якої їх комбінації. У деяких варіантах здійснення винаходу функціонально активна суміш не містить жодного бактеріального ферменту у комбінації з білком-носієм ниткоподібного гриба. У деяких варіантах здійснення винаходу функціонально активна суміш не утворює жодного антитіла або функціонально активних фрагментів антитіла, таких як Fab, одноланцюжкове антитіло і т.д. У деяких варіантах здійснення винаходу полінуклеотиди, що кодують гетерологічні або гомологічні поліпептиди, функціонально зв'язані з одним або декількома промоторами. Промотор може бути будь-яким прийнятним промотором, який відомий у даній галузі у наш час або буде відкритий у майбутньому. У деяких варіантах здійснення винаходу полінуклеотиди експресовані під впливом промотору, нативного для ниткоподібного гриба. У деяких варіантах здійснення винаходу полінуклеотиди експресовані під впливом гетерологічного промотору. У деяких варіантах здійснення винаходу полінуклеотиди експресовані під впливом конститутивного або індуцибельного промотору. Приклади прийнятних промоторів включають, не обмежуючись ними, промотор целюлази, промотор ксиланази, промотор 1818 (який раніше був ідентифікований як високоекспресований білок при картуванні EST Trichoderma). У деяких варіантах здійснення винаходу промотор є промотором целюлази ниткоподібного гриба. У деяких варіантах здійснення винаходу промотор є промотором екзоцелобіогідролази, ендоглюканази або бета-глюкозидази. У деяких варіантах здійснення винаходу промотор є промотором целобіогідролази I (cbh1). Необмежувальні приклади промоторів включають промотори cbh1, cbh2, egl1, egl2, egl3, egl4, egl5, pki1, gpd1, xyn1 і xyn2. Крім того, два або більше полінуклеотидів, що кодують гетерологічні або гомологічні поліпептиди або їх частини, можуть бути злиті один з одним з утворенням злитого полінуклеотиду. Злитий полінуклеотид може бути функціонально зв'язаний з будь-яким описаним вище прийнятним промотором. У деяких варіантах здійснення винаходу перший полінуклеотид, що кодує перший гетерологічний поліпептид, функціонально зв'язаний з першим промотором. Перший промотор може відрізнятися, хоча і не обов'язково, від одного або декількох промоторів, з якими функціонально зв'язаний другий або третій полінуклеотид. У деяких варіантах здійснення винаходу перший полінуклеотид функціонально зв'язаний з промотором гена, що кодує гомологічний поліпептид. У деяких варіантах здійснення винаходу полінуклеотид, наприклад, другий полінуклеотид, що кодує другий гетерологічний поліпептид, злитий з іншим полінуклеотидом, наприклад, з третім полінуклеотидом, що кодує гомологічний поліпептид, з утворенням злитого полінуклеотиду. Злитий полінуклеотид може бути функціонально зв'язаний з будь-яким прийнятним промотором, який включає, не обмежуючись ним, промотор гена, що кодує гомологічний поліпептид. Злитий полінуклеотид кодує злитий поліпептид або злитий білок, який включає два поліпептиди, домени або їх частини. Частини або домени поліпептидів можуть являти собою будь-яку частину або домен поліпептидів, які виконують щонайменше одну функцію, біологічну або яку-небудь ще, або стають функціонально активними при утворенні злитого поліпептиду або об'єднанні з іншими поліпептидами функціонально активної суміші. У деяких варіантах здійснення винаходу злитий білок включає другий гетерологічний поліпептид і гомологічний поліпептид. У деяких варіантах здійснення винаходу злитий полінуклеотид кодує злитий білок, що включає два поліпептиди, наприклад, другий гетерологічний поліпептид і гомологічний поліпептид, розділені лінкером або лінкерною областю. Лінкер може бути будь-яким прийнятним лінкером, що використовується для зв'язування двох поліпептидів. Лінкерна область звичайно утворює подовжений напівжорсткий спейсер між доменами пептидів з незалежно укладеним ланцюгом. Лінкерна область між поліпептидами злитого білка є більш переважною, роблячи можливим незалежне укладання ланцюга поліпептидів. У деяких варіантах здійснення винаходу лінкер одержують з глюкоамілази виду Aspergillus і лінкерів СВНI виду Tricoderma. У деяких варіантах здійснення винаходу лінкер може бути, хоча і не обов'язково, частиною поліпептидів, які утворюють злитий білок. У деяких варіантах здійснення винаходу поліпептиди злитого білка являють собою другий гетерологічний поліпептид і гомологічний поліпептид. У деяких варіантах здійснення винаходу злитий полінуклеотид кодує злитий білок, що включає два поліпептиди, розділених лінкером або лінкерною областю, і сайт розщеплення. У деяких варіантах здійснення винаходу поліпептиди злитого білка являють собою другий 6 UA 104279 C2 5 10 15 20 25 30 35 40 45 50 55 60 гетерологічний поліпептид і гомологічний поліпептид. Як правило, сайт розщеплення розташований у лінкерній області і розділяє послідовності, що межують з сайтом розщеплення. Сайт розщеплення може включати будь-яку послідовність, яка може бути розщеплена будьяким способом, який відомий у наш час або буде відкритий у майбутньому, і включає, не обмежуючись ним, розщеплення протеазою або під впливом визначених хімічних речовин. Приклади таких послідовностей включають, не обмежуючись ними, сайт розщеплення кексином, наприклад, сайт пізнавання КЕХ2, що містить кодони для амінокислот Lys і Arg, сайти пізнавання Lys і Arg протеази трипсину і сайт пізнавання і розщеплення Lys-C ендопротеїнази. У деяких варіантах здійснення винаходу ниткоподібний гриб за даним винаходом далі включає полінуклеотид, який кодує маркер, що селектується. Вказаний маркер може бути будьяким прийнятним маркером, що дозволяє проводити відбір трансформованих клітин-хазяїнів. Маркер, що селектується, звичайно є геном, що експресується у клітині-хазяїні, який полегшує відбір вказаних клітин-хазяїнів, що містять вектор. У використовуваному тут значенні вказаний термін звичайно стосується генів, які показують, що клітина-хазяїн поглинула ДНК, що представляє інтерес, або відбулася яка-небудь інша реакція. Маркери, що селектуються, звичайно є генами, які надають бактеріальну стійкість або метаболічну перевагу клітині-хазяїну, що дозволяє відрізнити клітини, які містять екзогенну ДНК, від клітин, які не одержали екзогенну послідовність у процесі трансформації. Приклади таких маркерів, що селектуються, включають, не обмежуючись ними, антибактеріальні засоби (наприклад, канаміцин, еритроміцин, актиноміцин, хлорамфенікол і тетрациклін). Додаткові приклади маркерів включають, не обмежуючись ними, ген pyr-4 Т. reesei, ацетолактат-синтазу, ген hyg Streptomyces, ген amdS Aspergillus nidulans і ген pyrG Asрergillus niger. У деяких варіантах здійснення винаходу ниткоподібний гриб за даним винаходом далі включає і може експресувати четвертий полінуклеотид, що кодує третій гетерологічний поліпептид. Гетерологічні або гомологічні поліпептиди можуть бути природними поліпептидами або їх варіантами. У деяких варіантах здійснення винаходу один або декілька гетерологічних поліпептидів можуть бути варіантами гомологічних поліпептидів. Наприклад, перший гетерологічний поліпептид може бути модифікованим гомологічним поліпептидом. У деяких варіантах здійснення винаходу перший і другий гетерологічні поліпептиди є модифікованими гомологічними поліпептидами. У деяких варіантах здійснення винаходу перший і другий гетерологічні поліпептиди є модифікованими гомологічними поліпептидами, і ниткоподібний гриб містить четвертий полінуклеотид, що кодує третій гетерологічний поліпептид. Третій гетерологічний поліпептид може бути або не бути модифікованим гомологічним поліпептидом. Гетерологічні і гомологічні поліпептиди за даним винаходом можуть бути будь-якими необхідними поліпептидами, які при змішуванні з іншими поліпептидами за даним винаходом утворюють функціонально активну суміш, яка виконує щонайменше одну функцію, біологічну або яку-небудь ще, що забезпечується щонайменше двома або трьома поліпептидами у суміші. У деяких варіантах здійснення винаходу суміш гетерологічних і гомологічних поліпептидів дозволяє функціонально активній суміші краще виконувати функцію, що відповідає активності ниткоподібного гриба або забезпечується вказаним грибом. У деяких варіантах здійснення винаходу вказана активність включає, не обмежуючись ними, кращу активність білка, що секретується, кращу активність оцукрювання або термостійкість, тобто стійкість при більш високих температурах або при змінених значеннях рН, і/або стабільну активність протягом більш тривалих періодів часу при постійній температурі. У деяких варіантах здійснення винаходу гетерологічні або гомологічні поліпептиди не включають жодного бактеріального ферменту у комбінації з білком-носієм ниткоподібного гриба. У деяких варіантах здійснення винаходу гетерологічні або гомологічні поліпептиди не утворюють жодного антитіла або функціонально активних фрагментів антитіла, таких як Fab, одноланцюжкове антитіло і т.д. У деяких варіантах здійснення винаходу один або декілька поліпептидів з першого або другого гетерологічного поліпептиду або гомологічного поліпептиду є ферментом або його частиною. У деяких варіантах здійснення винаходу перший або другий гетерологічний поліпептид або гомологічний поліпептид є целюлазою, геміцелюлазою, ксиналазою, мананазою, їх доменом або частиною. У деяких варіантах здійснення винаходу перший або другий гетерологічний поліпептид або гомологічний поліпептид є целюлазою або її частиною. У деяких варіантах здійснення винаходу перший і другий гетерологічні поліпептиди і гомологічний поліпептид разом утворюють функціонально активну суміш целюлаз. У деяких варіантах здійснення винаходу перший або другий гетерологічний поліпептид або гомологічний поліпептид є целюлазою, вибираною з групи, що включає екзоцелобіогідролази, ендоглюканази, бета-глюкозидази або їх частини. Перший або другий гетерологічний 7 UA 104279 C2 5 10 15 20 25 30 35 40 45 50 55 поліпептид, гомологічний поліпептид і, у випадку його наявності, третій гетерологічний поліпептид без яких-небудь обмежень можуть бути вибрані з групи, що включає екзоцелобіогідролази, ендоглюканази, бета-глюкозидази або їх домени. У деяких варіантах здійснення винаходу декілька гетерологічних або гомологічних поліпептидів можуть належати до одного і того ж класу або групи целюлаз. Наприклад, два або більше поліпептидів можуть належати до класу екзоцелобіогідролаз. У деяких варіантах здійснення винаходу один з гетерологічних поліпептидів належить до того ж класу целюлаз, що і гомологічний поліпептид. У деяких варіантах здійснення винаходу гетерологічні і гомологічні поліпептиди є однаковими членами одного класу, але мають послідовності різного походження. У деяких варіантах здійснення винаходу ниткоподібний гриб за даним винаходом містить перший полінуклеотид і другий полінуклеотид, що кодують відповідно перший гетерологічний поліпептид і другий гетерологічний поліпептид, при цьому перший гетерологічний поліпептид є екзоцелобіогідролазою і другий гетерологічний поліпептид є ендоглюканазою. У деяких варіантах здійснення винаходу перший гетерологічний поліпептид є екзоцелобіогідролазою, що класифікується як ЄС 3.2.1.91, і другий гетерологічний поліпептид є ендоглюканазою, що класифікується як ЄС 3.2.1.4. У деяких варіантах здійснення винаходу перший гетерологічний поліпептид є екзоцелобіогідролазою, вибираною з групи, що складається з сімейства GH 5, 6, 7, 9, 48, і другий гетерологічний поліпептид є ендоглюканазою, вибираною з групи, що складається з сімейства GH 5, 6, 7, 8, 9, 12, 17, 31, 44, 45, 48, 51, 61, 64, 74 і 81. Як було вказано вище, гетерологічні і гомологічні поліпептиди за даним винаходом без якихнебудь обмежень можуть бути вибрані з класів целюлазних ферментів. У даному описі винаходу наведені типові комбінації ферментів. У деяких варіантах здійснення винаходу перший гетерологічний поліпептид є екзоцелобіогідролазою, другий гетерологічний поліпептид є ендоглюканазою і гомологічний поліпептид є екзоцелобіогідролазою. У деяких варіантах здійснення винаходуперший гетерологічний поліпептид є першою екзоцелобіогідролазою, другий гетерологічний поліпептид є ендоглюканазою, гомологічний поліпептид є другою екзоцелобіогідролазою, при цьому перша екзоцелобіогідролаза і друга екзоцелобіогідролаза є однаковими целобіогідролазами, наприклад, перша і друга екзоцелобіогідролази є СВНI або обидві є CBHII. Ниткоподібні гриби за даним винаходом можуть бути будь-якими ниткоподібними грибами, відомими фахівцям у даній галузі. У деяких варіантах здійснення винаходу ниткоподібні гриби включають, не обмежуючись ними, наведені далі роди: Aspergillus, Acremonium, Aureobasidium, Beauveria, Cеphalosporium, Ceriporiopsis, Chaetomium paecilomyces, Chrysosporium, Claviceps, Cochiobolus, Cryptococcus, Cyathus, Endothia, Endotia mucor, Fusarium, Gilocladium, Humicola, Magnaporthe, Myceliophthora, Myrothecium, Mucor, Neurospora, Phanerochaete, Podospora, Paecilomyces, Penicillium, Pyricularia, Rhizomucor, Rhizopus, Schizophylum, Stagonospora, Talaromyces, Trichoderma, Thermomyces, Thermoascus, Thielavia, Tolypocladium, Trichophyton і Trametes pleurotus. У деяких варіантах здійснення винаходу ниткоподібні гриби включають, не обмежуючись ними, наведені далі види: А. nidulans, А. niger, А. awomari, наприклад, NRRL 3112, ATCC 22342 (NRRL 3112), ATCC 44733, ATCC 14331 і штам UVK 143f, А. oryzae, наприклад, АТСС 11490, N. crassa, Trichoderma reesei, наприклад, NRRL 15709, АТСС 13631, 56764, 56765, 56466, 56767, і Trichoderma viride, наприклад, АТСС 32098 і 32086. У деяких варіантах здійснення винаходу ниткоподібний гриб за даним винаходом є Trichoderma. У деяких варіантах здійснення винаходу ниткоподібний гриб за даним винаходом є Trichoderma reesei. У деяких варіантах здійснення винаходу гетерологічні поліпептиди можуть бути вибрані з групи, що включає Humicola grisea, Acidothermus cellulolyticus, Thermobifida fusca або Penicillium funiculosum. У деяких варіантах здійснення винаходу гетерологічні поліпептиди вибрані з Humicola grisea, Acidothermus cellulolyticus, Thermobifida, наприклад, Thermobifida fusca, або Penicillium funiculosum, і гомологічний поліпептид вибраний з Trichoderma reesei. У даному описі винаходу наведені типові комбінації гетерологічних і гомологічних поліпептидів. У деяких варіантах здійснення винаходу гетерологічні і гомологічні поліпептиди функціонально активної суміші можуть бути вибрані з групи, що складається з EGI, EGII, EGIII T. reesei (відповідно CEL7B, 5A, 12A), варіантів CEL12A, EGIII H. grisea, E5 і Е3 T. fusca і E1 і GH74 A. cellulolyticus. У деяких варіантах здійснення винаходу гетерологічні поліпептиди функціонально активної суміші можуть являти собою злиту конструкцію з екзо- і ендоцелюлаз. У деяких варіантах здійснення винаходу злитий білок має активність, що розкладає целюлозу, завдяки наявності каталітичного домену, виділеного з екзоцелобіогідролази грибів, і каталітичного домену, виділеного з ендоглюканази. Необмежувальні приклади наведені у публікації заявки на патент США № 20060057672. 8 UA 104279 C2 5 10 15 20 25 30 35 40 45 50 55 60 У деяких варіантах здійснення винаходу гетерологічні поліпептиди функціонально активної суміші можуть бути варіантами CBHI H. jecorina, ферменту Cel7. У деяких варіантах здійснення винаходу целобіогідролази, які включають, не обмежуючись ними, целобіогідролази, описані у публікаціях заявок на патент США №№ 20050277172 і 20050054039, можуть характеризуватися поліпшеною термостійкістю і реверсивністю. У деяких варіантах здійснення винаходу гетерологічні поліпептиди функціонально активної суміші можуть бути варіантами CBH2 H. jecorina, ферменту Cel7. У деяких варіантах здійснення винаходу целобіогідролази, які включають, не обмежуючись ними, целобіогідролази, описані у публікації заявки на патент США № 20060205042, можуть характеризуватися поліпшеною термостійкістю і реверсивністю. У деяких варіантах здійснення винаходу ниткоподібний гриб є Т. reesei, перший гетерологічний поліпептид є CBHI Humicola grisea, другий гетерологічний поліпептид є ендоглюканазою 1 Acidothermus cellulolyticus і гомологічний поліпептид є CBHI Trichoderma reesei. У деяких варіантах здійснення винаходу ниткоподібний гриб є Т. reesei, перший гетерологічний поліпептид або другий гетерологічний поліпептид вибирають з групи, що складається з целобіогідролази CBHI Penicillium funiculosum, ендоглюканаз Е3 Thermobifida, ендоглюканаз Е5 Thermobifida, GH74-core і GH48 Acidorhermus cellulolyticus. У деяких варіантах здійснення винаходу ниткоподібний гриб включає четвертий полінуклеотид, що кодує третій гетерологічний поліпептид. У цьому випадку перший поліпептид є модифікованою CBHI T. reesei, другий гетерологічний поліпептид є модифікованою CBHII T. reesei, третій гетерологічний поліпептид є ендоглюканазою 1 Acidothermus cellulolyticus, і гомологічний поліпептид є CBHI T. reesei. Даний винахід стосується також функціонально активних сумішей з поліпшеними властивостями і/або активністю. У деяких варіантах здійснення винаходу перший гетерологічний поліпептид є екзоцелобіогідролазою, другий гетерологічний поліпептид є ендоглюканазою, і гомологічний поліпептид є екзоцелобіогідролазою. У цьому випадку перший гетерологічний поліпептид, другий гетерологічний поліпептид і гомологічний поліпептид утворюють суміш термостійких целюлаз. У деяких варіантах здійснення винаходу полінуклеотиди, що кодують гетерологічні і гомологічні поліпептиди, можуть бути позахромосомними, тобто можуть знаходитися у векторі або плазміді, або полінуклеотиди можуть бути вбудовані у хромосоми ниткоподібного грибахазяїна. У деяких варіантах здійснення винаходу ниткоподібний гриб-хазяїн містить щонайменше один полінуклеотид, що кодує перший, другий або третій гетерологічний поліпептид або гомологічний поліпептид, вбудований в його геном. У деяких варіантах здійснення винаходу ниткоподібний гриб-хазяїн містить щонайменше один полінуклеотид, що кодує перший, другий або третій гетерологічний поліпептид або гомологічний поліпептид, вбудований в його геном, і щонайменше один полінуклеотид, що кодує гетерологічний або гомологічний поліпептид, у стійкому векторі, трансфікованому у гриб-хазяїн. У деяких варіантах здійснення винаходу хазяїном є Т. reesei, що містить щонайменше один полінуклеотид, що кодує перший або другий гетерологічний поліпептид або гомологічний поліпептид, вбудований в його геном. У деяких варіантах здійснення винаходу хазяїном є Т. reesei, що містить два полінуклеотиди, вбудованих в його геном. Вказані полінуклеотиди кодують перший, другий або, у випадку його наявності, третій гетерологічний поліпептид або гомологічний поліпептид. У деяких варіантах здійснення винаходу один або декілька полінуклеотидів, що експресують гетерологічну або гомологічну екзоцелобіогідролазу, вбудовані у геном хазяїна, що є Т. reesei. У деяких варіантах здійснення винаходу полінуклеотид, що кодує гетерологічну ендоглюканазу, вбудований у геном хазяїна, що є Т. reesei. У деяких варіантах здійснення винаходу полінуклеотид, що кодує гетерологічну ендоглюканазу, і полінуклеотид, що кодує гетерологічну або гомологічну екзоцелобіогідролазу, вбудовані у геном хазяїна, що є Т. reesei. Очевидно, що, коли тільки один або два з трьох або чотирьох полінуклеотидів, що кодують поліпептиди функціонально активної суміші, вбудовані у геном хазяїна, інші полінуклеотиди трансфіковані у хазяїна і присутні у векторі або плазміді. У деяких варіантах здійснення винаходу ниткоподібний гриб містить перший полінуклеотид і другий полінуклеотид, що кодують відповідно перший гетерологічний поліпептид і другий гетерологічний поліпептид, і третій полінуклеотид, що кодує гомологічний поліпептид, при цьому всі три полінуклеотиди є позахромосомними. Даний винахід стосується також культурального середовища, що містить популяцію описаних вище ниткоподібних грибів. Культуральне середовище може бути щільним, напівтвердим або рідким, причому середовище вибирають в залежності від хазяїна, а також від поліпептидів, що експресуються. 9 UA 104279 C2 5 10 15 20 25 30 35 40 45 50 55 60 Даний винахід стосується також поліпептидної суміші, що включає перший гетерологічний поліпептид, другий гетерологічний поліпептид і гомологічний поліпептид, одержані з ниткоподібних грибів, розглянутих у даному описі винаходу. У деяких варіантах здійснення винаходу поліпептидна суміш є сумішшю ферментів або їх доменів. У деяких варіантах здійснення винаходу поліпептидна суміш є сумішшю целюлаз, геміцелюлаз, ксиланаз, мананаз або їх доменів. Крім того, даний винахід стосується способу одержання суміші поліпептидів, який включає одержання поліпептидної суміші з ниткоподібних грибів за даним винаходом. Поліпептидна суміш містить перший гетерологічний поліпептид, другий гетерологічний поліпептид і гомологічний поліпептид. У деяких варіантах здійснення винаходу суміш поліпептидів містить третій гетерологічний поліпептид. Як було вказано вище, суміш поліпептидів є функціонально активною сумішшю. У деяких варіантах здійснення винаходу суміш поліпептидів є сумішшю ферментів або їх доменів. У деяких варіантах здійснення винаходу суміш поліпептидів є сумішшю целюлаз, геміцелюлаз, ксиланаз, мананаз або їх доменів. У деяких варіантах здійснення винаходу суміш поліпептидів є сумішшю целюлаз, що включає перший гетерологічний поліпептид, який є екзоцелобіогідролазою, другий гетерологічний поліпептид, який є ендоглюканазою, і гомологічний поліпептид, який є екзоцелобіогідролазою. У деяких варіантах здійснення винаходу суміш целюлаз містить перший гетерологічний поліпептид, який є першою екзоцелобіогідролазою, другий гетерологічний поліпептид, який є ендоглюканазою, і гомологічний поліпептид, який є другою екзоцелобіогідролазою. У цьому випадку перша екзоцелобіогідролаза і друга екзоцелобіогідролаза є однаковими целобіогідролазами. У деяких варіантах здійснення винаходу перша і друга екзоцелобіогідролази є CBHI. У деяких варіантах здійснення винаходу перша і друга екзоцелобіогідролази є CBHII. Як повинно бути очевидно фахівцю у даній галузі, у ниткоподібних грибах за даним винаходом може бути експресовано декілька інших комбінацій гетерологічних і гомологічних поліпептидів. Інша типова суміш целюлаз включає перший гетерологічний поліпептид, який є CBHI Humicola grisea, другий гетерологічний поліпептид, який є ендоглюканазою 1 Acidothermus cellulolyticus, і гомологічний поліпептид, який є CBHI Trichoderma reesei. Об'єкти даного винаходу можуть бути краще зрозумілі при ознайомленні з наведеними нижче прикладами, які не обмежують обсяг даного винаходу. Фахівцям у даній галузі повинно бути очевидно, що у межах обсягу даного винаходу можуть бути використані численні модифікації як матеріалів, так і способів. 7. Приклади 7.1. Приклад 1. Створення трьохкомпонентного штаму Трьохкомпонентний штам складається з наведених далі трьох частин: (i) штаму, що продукує целюлазу Т. reesei; (ii) нуклеїнової кислоти, що включає ген cbh1 Humicola grisea у вказаному штамі; і (iii) гібриду екзо- і ендонуклеаз гена cbh1 Т. reesei з ендоглюканазою 1 Acidothermus cellulolyticus. Створення злитого вектора CBH1-E1 Злита конструкція CBH1-E1 включає промотор гена cbh1 Т. reesei; послідовність гена cbh1 Т. reesei від ініціюючого кодону до кінця лінкеру cbh1 і 12 додаткових основ від 5'-кінця ДНК до початку кодуючої послідовності ендоглюканази, кодуючу послідовність ендоглюканази, термінуючий кодон і термінатор cbh1 Т. reesei. Нуклеотидна послідовність (SEQ ID NO:1) злитої конструкції гетерологічної целюлази складається з 2656 основ (див. фігуру 1) і включає сигнальну послідовність cbh1 Т. reesei; каталітичний домен cbh1 Т. reesei; лінкерну послідовність cbh1 Т. reesei; сайт розщеплення кексином, що включає кодони для амінокислот SKR, і послідовність, що кодує каталітичний домен GH5A-E1 Acidothermus cellulolyticus. Прогнозована амінокислотна послідовність (SEQ ID NO:2) злитого білка целюлази, створена на основі послідовності нуклеїнової кислоти, показаної на фігурі 1, представлена на фігурі 2. 12 додаткових основ ДНК, ACTAGTAAGCGG (нуклеотиди 1565-1576 SEQ ID NO:1) кодують рестрикційну ендонуклеазу SpeI і амінокислоти Thr, Ser, Lys і Arg. Плазміда E1-pUC19, що містить відкриту рамку зчитування для локусу гена Е1, була використана як матрична ДНК при виконанні ПЛР. (Еквівалентні плазміди описані у патенті США № 5536655, в якому також розглянуте клонування гена Е1 з актиноміцету Acidothermus cellulolyticus АТСС 43068, Mohagheghi A. et al., 1986). Стандартні методи роботи з плазмідною ДНК і ампліфікації ДНК за допомогою полімеразної ланцюгової реакції (ПЛР) описані у публікації Sambrook, et al., 2001. Наведені нижче дві затравки були використані для ампліфікації кодуючої області каталітичного домену ендоглюканази Е1. Верхня затравка 1=EL-316 (що містить сайт Spe1): 10 UA 104279 C2 5 10 15 20 25 30 35 40 45 50 55 60 GCTTATACTAGTAAGCGCGCGGGCGGCGGCTATTGGCACAC (SEQ ID NO:3); нижня затравка 2=EL-317 (що містить сайт AscI і комплекс термінуючого кодону і нижнього комплементу): GCTTATGGCGCGCCTTAGACAGGATCGAAAATCGACGAC (SEQ ID NO:4). Обробку здійснювали у наведених нижче умовах реакції з використанням матеріалів з набору ДНК-полімераз PLATINUM Pfx (Invitrogen, Carlsbad, CA): 1 мкл основної суміші dNTP (кінцева концентрація 0,2 мМ); 1 мкл затравки 1 (кінцева концентрація 0,5 мкМ); 1 мкл затравки 2 (кінцева концентрація 0,5 мкМ); 2 мкл матричної ДНК (кінцева концентрація 50-200 нг); 1 мкл 50 мМ MgSO4 (кінцева концентрація 1 мМ); 5 мкл 10-кратної кількості буфера для ампліфікації Pfx; 5 мкл 10-кратної кількості розчину підсилювача ПЛР; 1 мкл ДНК-полімерази Platinum Pfx (всього 2,5 од.); 33 мкл води до досягнення загального об'єму реакційної суміші, що дорівнює 50 мкл. Параметри ампліфікації були наступними: стадія 1: 94 °C протягом 2 хвилин (1-ий цикл служить тільки для денатурації антитілозв'язаної полімерази); стадія 2: 94 °C протягом 45 секунд; стадія 3: 60 °C протягом 30 секунд; стадія 4: 68 °C протягом 2 хвилин; стадія 5: повторне виконанні стадії 2 протягом 24 циклів; і стадія 6: 68 °C протягом 4 хвилин. Продукт ПЛР відповідної величини клонували у вектор ТОРО Zero Blunt і трансфікували у хімічно компетентні клітини Top10 E. coli (Invitrogen Corp., Carlsbad, Calif.), висівали на відповідне вибірне середовище (середовище LA, що містить 50 частин на мільйон канаміцину) і вирощували протягом ночі при 37 °C. З середовища збирали декілька колоній і вирощували їх протягом ночі при 37 °C у вибірному середовищі (середовище LB, що містить 50 частин на мільйон канаміцину) у вигляді 5 мл культур, з яких одержували плазмідні мініпрепарати. Плазмідну ДНК з декількох клонів розщеплювали рестрикційними ферментами для підтвердження правильної величини вставки. Правильність послідовності підтверджували шляхом секвенування ДНК. Після перевірки послідовності з вектора ТОРО вирізали каталітичний домен Е1, розщеплюючи рестрикційними ферментами SрeI і AscI. Вказаний фрагмент лігували у вектор pTrex4, який був розщеплений рестрикційними ферментами SpeI і AscI, як показано на фігурі 3. Ліговану суміш трансфікували у компетентні клітини ММ294 E. coli, які висівали на відповідне вибірне середовище (середовище LA, що містить 50 частин на мільйон карбеніциліну) і вирощували протягом ночі при 37 °C. З середовища збирали декілька колоній і вирощували їх протягом ночі при 37 °C у вибірному середовищі (середовище LB, що містить 50 частин на мільйон карбеніциліну) у вигляді 5 мл культур, з яких одержували плазмідні мініпрепарати. Правильність лігування векторів, що містять злитий білок CBH1-E1, підтверджували шляхом розщеплення рестрикційними ферментами. Створення вектора, що експресує cbh1 Н. grisea Конструкція, що експресує cbh1 Н. grisea, включає промотор cbh1 R. reesei; послідовність гена cbh1 Н. grisea, термінатор сbhl Т. reesei і маркер, що селектується, amdS А. nidulans. Вказані послідовності можуть бути зібрані рядом способів, відомих фахівцям у даній галузі, один з яких описаний нижче. Геномну ДНК екстрагували зі зразка міцелію Humicola grisea var. thermoidea (CBS 225.63). Геномна ДНК може бути виділена будь-яким методом, відомим у даній галузі. Може бути виконана наведена нижче процедура. Клітини вирощували при 45 °C у 20 мл декстрозокартопляного бульйону (PDB) протягом 24 годин. Клітини розводили у співвідношенні 1:20 у свіжому середовищі PDB і вирощували протягом ночі. Два мілілітри клітин центрифугували і клітинний дебрис промивали в 1 мл КС (60 г KCl, 2 г лимонної кислоти на один літр, рН доводили до 6,2, додаючи 1 М розчин КОН). Клітинний дебрис ресуспендували у 900 мкл КС. Додавали 100 мкл (20 мг/мл) новозиму, обережно перемішували і при 37 °C під мікроскопом слідкували за утворенням протопластів, доки не було утворено більше 90 % протопластів протягом максимум 2 годин. Клітини центрифугували зі швидкістю 1500 обертів/хв. (4600×G) протягом 10 хвилин. Додавали 200 мкл TES/SDS (10 мМ трис, 50 мМ EDTA, 150 мМ NaCl, 1 % SDS), змішували та інкубували при кімнатній температурі протягом 5 хвилин. ДНК виділяли, використовуючи набір для виділення мініпрепаратів Qiagen (Qiagen). Колонку елюювали 100 мкл води milli-Q і збирали ДНК. Альтернативним методом є метод FastPrep, що служить для виділення геномної ДНК з виду Н.grisea var thermoidea, що вирощується на планшетах PDA при 45 °C. Вказана система складається з пристрою FastPrep і набору FastPrep для виділення нуклеїнової кислоти. (Систему FastPrep можна придбати у компанії Qbiogene, MP Biomedicals, United States, 29525 Fountain Pkwy., Solon, OH 44139). Затравки для ампліфікації гена cbh1 Н. grisea методом ПЛР були створені на основі NCBI, номер доступу D63515. Вказані затравки були призначені для ампліфікації гена cbh1 Н. grisea 11 UA 104279 C2 5 10 15 20 25 30 35 40 45 50 55 від ініціюючого кодону до термінатора. Послідовність верхньої затравки включала 4 нуклеотиди САСС для полегшення клонування у вектор pENTR ТОРО з метою використання системи клонування Gateway (Invitrogen). Верхня затравка: 5' CACCATGCGTACCGCCAAGTTCGC 3' (SEQ ID NO:5). Нижня затравка: 5' TTACAGGCACTGAGAGTACCAG 3' (SEQ ID NO:6). Умови виконання ПЛР Продукт ПЛР клонували у вектор pENTR/D відповідно до процедури для системи Gateway компанії Invitrogen. Потім вектор трансфікували у хімічно компетентні клітини Top10 E.coli (Invitrogen), проводячи відбір клітин канаміцином. Плазмідну ДНК з декількох клонів розщеплювали рестрикційними ферментами для підтвердження правильної величини вставки і потім секвенували для підтвердження правильності послідовності. Плазмідну ДНК з одного клону додавали до реакційного середовища LR, що містить клоназу (система Gateway компанії Invitrogen) з ДНК цілеспрямованого вектора pTrex3g/amdS. Створення вектора pTrex3g У даному розділі описане створення основного вектора, що використовується для експресії генів, які представляють інтерес. Вектор pTrex3g був описаний раніше; див., наприклад, публікацію заявки на патент США № 20070015266. Вказаний вектор створений на основі вектора pSL1180 Е. coli (Pharmacia Inc., Piscataway, NJ, USA), який є вектором на основі фагоміду pUC118 (Brosius, J. (1989) DNA 8:759) з подовженим сайтом множинного клонування, що містить 64 шестичленних послідовності, пізнавані рестрикційними ферментами. Створений вектор є цілеспрямованим вектором системи Gateway (Hartley, J.L. et al., (2000) Genome Research 10: 1788-1795), який за допомогою методу Gateway (Invitrogen) дозволяє вставляти будь-яку необхідну відкриту рамку зчитування між промоторною і термінуючою областями гена cbh1 R. reesei. Ген amdS Aspergillus nidulans був вставлений для використання як маркера, що селектується, у процесі трансформації. Промотор і термінатор були розташовані з можливістю експресії гена, що представляє інтерес. Докладний опис вектора pTrex3g Вказаний вектор має довжину, що дорівнює 10,3 т.п.о. У полілінкерну область вектора pSL1180 вставлені наведені далі сегменти ДНК: (i) сегмент ДНК довжиною 2,2 п.о. з промоторної області гена cbh1 Т. reesei; (ii) кластер рамки зчитування А Gateway довжиною 1,7 т.п.о. компанії Invitrogen, який включає сайти рекомбінації attR1 і attR2 біля кінця, що фланкує ген стійкості до хлорамфеніколу (CmR) і ген ccdB; (iii) сегмент ДНК довжиною 336 п.о. з термінуючої області гена cbh1 Т. reesei; і (iv) фрагмент ДНК довжиною 2,7 т.п.о., що містить ген amdS Aspergillus nidulans з нативними промоторною і термінуючою областями. На фігурі 4 показана плазмідна карта експресуючого вектора pTrex3g Т. reesei. Клон гена cbh1 Н. grisea в описаному вище векторі pENTR використовували для рекомбінації цілеспрямованого вектора pTrex3g, що виконується у реакційному середовищі LR, яке містить клоназу, відповідно до інструкцій виробника (Invitrogen). Гени CmR і ccdB цілеспрямованого вектора pTrex3g були замінені геном cbh1 Н. grisea з вектора pENTR/D. У результаті рекомбінації ген cbh1 Н. grisea був вставлений безпосередньо між промотором cbh1 Т. reesei і термінатором cbh1 Т. reesei цілеспрямованого вектора. Внаслідок рекомбінації були одержані послідовності AttB довжиною 25 п.о., що фланкують гени cbh1 Н. grisea вгорі і внизу. Аліквоту реакційного середовища LR, що містить клоназу, трансфікували у хімічно компетентні клітини Top10 E. coli (Invitrogen), які вирощували протягом ночі, проводячи відбір карбеніциліном. Плазмідну ДНК з декількох клонів розщеплювали відповідними рестрикційними ферментами для підтвердження правильної величини вставки і потім секвенували для підтвердження правильності послідовності. Щоб одержати ДНК для трансформації, плазмідну ДНК з правильного клону розщеплювали ендонуклеазою Xba1 для вивільнення експресуючого фрагмента, що включає промотор cbh1 Т. reesei:ген cbh1 Н. grisea:термінатор cbh1 Т. reesei:ген amdS А. nidulans. Вказаний фрагмент довжиною 6,2 т.п.о. виділяли з ДНК E. coli шляхом екстракції в агарозному гелі за допомогою стандартних методів і трансфікували у штам Т. reesei, одержаний із загальнодоступного штаму QM6a відповідно до наведеного нижче опису. Експресуючий вектор, що включає два сайти XbaI, схематично зображений на фігурі 5А, і нуклеотидна послідовність (SEQ ID NO:7) експресуючого вектора показана на фігурі 5В. Котрансформація і ферментація Trichoderma reesei Як штам-хазяїн для трансформації конструкціями за даним винаходом використовували похідний штам-хазяїн RL-P37 T. reesei (Sheir-Neiss, et al., 1984), який був підданий цілому ряду стадій мутагенезу для збільшення продукування целюлази, включаючи делецію нативного гена cbh1 (Suominen, P.L. et al., 1993, Mol. Gen. Genet. 241:523-30). 12 UA 104279 C2 5 10 15 20 25 30 35 40 45 50 55 60 Біолітичну трансформацію Т. reesei за допомогою конструкції, що експресує cbh1 Н. grisea, і злитої конструкції cbh1 Т. reesei і Е1 A. cellulolyticus виконували описаним нижче способом. 8 Одержували завись спор (приблизно 3,5×10 спор/мл) зі штаму Т. reesei, що є похідним Р37. Приблизно 100-200 мкл вказаної зависі спор розподіляли у центрі планшетів з ацетамідним середовищем ММ. Ацетамідне середовище ММ має наведений далі склад: 0,6 г/л ацетаміду; 1,68 г/л CsСl; 20 г/л глюкози; 20 г/л КН2РО4; 0,6 г/л СаCl2.2H2O; 1 мл/л 1000-кратного розчину слідових елементів; 20 г/л нобль-агару; рН 5,5. 1000-кратний розчин слідових елементів містив 5,0 г/л FeSO4.7H2O, 1,6 г/л MnSO4.H2O, 1,4 г/л ZnSO4.7H2O і 1,0 г/л CoCl2.6H2O. Завись спор залишали сохнути на поверхні ацетамідного середовища ММ у стерильній витяжній шафі. Т. reesei трансформували за допомогою системи доставки частинок Biolistic® PDS-1000/He компанії Bio-Rad (Hercules, CA) відповідно до інструкцій виробника (Lorito, M. et al., 1993, Curr. Genet. 24:349-56). У мікроцентрифугову пробірку вводили 60 мг частинок вольфраму М10. Додавали 1 мл етанолу, суміш інтенсивно перемішували протягом короткого періоду часу і залишали відстоюватися протягом 15 хвилин. Частинки центрифугували зі швидкістю 15000 обертів/хв. протягом 15 хвилин. Етанол видаляли і частинки тричі промивали стерильною деіонізованою Н2О, після чого додавали 1 мл 50 % (об./об.) стерильного гліцерину. Суміш інтенсивно перемішували протягом десяти секунд для суспендування вольфраму, видаляли 25 мкл суспензії частинок вольфраму у гліцерині і вводили у мікроцентрифугову пробірку. При безперервному інтенсивному перемішуванні до 25 мкл суспензії частинок вольфраму у гліцерині додавали наведені нижче речовини у вказаному порядку та інкубували суміш між додаваннями протягом 5 хвилин: 2 мкл (100-300 нг/мкл) експресуючого вектора cbh1 Н. grisea (фрагмент, вирізаний XbaI), 2 мкл (100-300 нг/мкл) експресуючого вектора cbh1-E1 (фрагмент, вирізаний XbaI), 25 мкл 2,5М розчину СaCl2 і 10 мкл 0,1М розчину спермідину. Після додання спермідину частинки інкубували протягом 5 хвилин і центрифугували протягом 3 секунд. Супернатант видаляли, частинки промивали 200 мкл 70 % (об./об.) етанолу і центрифугували протягом 3 секунд. Супернатант видаляли, частинки промивали 200 мкл 100 % етанолу і центрифугували протягом 3 секунд. Супернатант видаляли, додавали 24 мкл 100 % етанолу і змішували піпетуванням. Пробірку вміщували у ванну для ультразвукового очищення приблизно на 15 секунд для ресуспендування частинок в етанолі. Під час знаходження пробірки у ванні для ультразвукового очищення видаляли 8 мкл аліквоти суспендованих частинок і вміщували у центр дисків макроносія, які встановлювали у сушильну шафу. Розчин вольфраму/ДНК після висушування на макроносії (приблизно 15 хвилин) вміщували у бомбардувальну камеру. Поряд вміщували планшет з ацетамідним середовищем ММ зі спорами, і проводили бомбардування, використовуючи розривні мембрани, що знаходяться під тиском 77 атм (1100 фунтів/кв.дюйм), відповідно до інструкцій виробника. Після бомбардування спор, що культивуються, частинками вольфраму/ДНК, планшети інкубували при 28 °C. Великі трансформовані колонії через 5 днів переносили на свіжі вторинні планшети з ацетамідним середовищем ММ (Penttila et al., (1987) Gene 61:155-164) та інкубували ще 3 дні при 28 °C. Колонії, які характеризувалися щільним, непрозорим ростом на вторинних планшетах, переносили на окремі планшети з ацетамідним середовищем ММ. Вказані колонії вирощували ще три дні, переносили на планшети з декстрозокартопляним агаром (PDA) та інкубували протягом 7-10 днів при 28 °C для споруляції. Експресію ферментів трансформантами оцінювали у двохкамерних струшуваних колбах. Трансформанти спочатку вирощували у струшуваній колбі для інокуляту, що містить наведене далі середовище: 22,5 г/л Proflo, 30 г/л а-лактози.Н2О, 6,5 г/л (NH4)2SO4, 2 г/л КН2РО4, 0,3 г/л MgSO4.7Н2О, 0,26 г/л CaCl2.2H2O, 0,72 г/л СаСО3, 2 мл 10 % твіну 80, 1 мл 1000-кратної кількості слідових солей TRI (1000-кратна кількість слідових солей TRI складається з: 5 г/л FeSO 4.7H2O, 1,6 г/л MnSO4.H2O, 1,4 г/л ZnSO4.7H2O). Умови вирощування були наступними: 50 мл середовища у 250-мл струшуваній колбі з 4 перегородками (Bellco Biotechnology, 340 Edrudo Road, Vineland, NJ 08360, USA), інкубація при 28 °C, швидкість струшування 225 обертів/хв. з діаметром обертання 2,5 см). Трансформанти інокулювали у струшуваних колбах для інокуляту, 2 переносячи шматочки PDA розміром 4 см , що містять міцелій і спори трансформанту. Після вирощування у колбі для інокуляту протягом 2 днів, 5 мл переносили у струшувану колбу для експресії, що містить 50 мл наведеного далі середовища: 5 г/л (NH4)2SO4, 33 г/л буфера PIPPS, 9 г/л кислот Bacto Casamino, 4,5 г/л КН 2РО4, 1,32 г/л CaCl2.2H2O, 1 г/л MgSO4.7Н2О, 5 мл антиспінювача Mazu DF204, 2,5 мл 400-кратної кількості слідових солей Т. reesei (400-кратна кількість слідових солей Т. reesei складається з: 175 г/л лимонної кислоти (безводної), 200 г/л FeSO 4.7H2O, 16 г/л ZnSO4.7H2O, 3,2 г/л CuSO4.5H2O, 1,4 г/л MnSO4.H2O, 0,8 г/л Н3ВО3, що додаються у вказаному порядку), рН доводили до 5,5, середовище стерилізували, здійснювали постстерилізацію і додавали 40 мл 40 % лактози. Культуру вирощували у 13 UA 104279 C2 5 10 15 20 25 30 35 40 45 50 55 60 струшуваній колбі для експресії у наведених далі умовах: 250-мл струшувана колба з 4 перегородками, інкубація при 28 °C, швидкість струшування 225 обертів/хв. Зразок видаляли через 5 днів, супернатант аналізували на білкових гелях для SDS-PAGE і забарвлювали кумасі. 7.2. Приклад 2. Створення чотирьохкомпонентного штаму Був створений штам, що включає чотири частини: (i) штам-хазяїн, що складається з продукуючого штаму з делецією гена cbh1; (ii) послідовність нуклеїнової кислоти для експресії злитого гена cbh1-E1; (iii) послідовність нуклеїнової кислоти для експресії термостійкого гена cbh1 Т. reesei генетично створеного білка і (iv) послідовність нуклеїнової кислоти для експресії термостійкого гена cbhII Т. reesei генетично створеного білка. ДНК всіх трьох експресуючих фрагментів котрансфікували у продукуючий штам з делецією гена cbh1, як показано на фігурі 6. Трансформанти T. reesei досліджували у присутності всіх трьох експресуючих фрагментів, вбудованих у геном. Були створені пари затравок для ПЛР, призначені для ампліфікації кожного з трьох експресуючих фрагментів. Для ферментації у струшуваних колбах були вибрані 32 трансформанти, які за результатами ПЛР характеризувалися наявністю всіх трьох експресуючих фрагментів. Культури у струшуваних колбах вирощували протягом трьох днів, брали зразки супернатанту і досліджували у 8 % трис-гліцинових гелях NuPAGЕ (Invitrogen) (1 мм) у розділяючому буфері для трис-гліцинового гелю з додецилсульфатом натрію (SDS). Зразки інкубували при 100 °C протягом 7 хвилин, потім на льоді протягом 5 хвилин і завантажували у кількості 20 мкл/смуга за винятком особливо обумовлених випадків (8 мкл супернатанту + 2 мкл відновника + 10 мкл 2-кратної кількості буфера для трис-гліцинового гелю з SDS). Деякі з 32 зразків характеризувалися високим рівнем експресованих генів, про що свідчать смуги білка. ДНК, що кодує варіант амінокислотної послідовності генів cbhI і cbhII Т. reesei, можна одержати різними методами, відомими у даній галузі. Такі методи включають, не обмежуючись ними, синтез генів, сайтнаправлений мутагенез (або сайтнаправлений мутагенез з використанням олігонуклеотидів), мутагенез методом ПЛР і кластерний мутагенез раніше одержаної ДНК, що кодує послідовність кДНК Т. reesei. Вектор був створений на основі вектора pTrex3g, що експресує ген cbh1 Т. reesei генетично створеного ферменту, що кодує генетично створений білок з наведеними нижче мутаціями у зрілій амінокислотній послідовності: S8P+T41I+N49S+A68T+N89D+S92T+S113N+S196T+P227L+D249K+T255P+S278P+E295K+T296P +T322Y+V403D+S411F. Послідовність ДНК від ініціюючого кодону до термінуючого кодону мала 1545 основ (SEQ ID NO:8), як показано на фігурі 7А. Послідовність генетично створеного білка CBHI (SEQ ID NO:9) показана на фігурі 7В (сигнальна послідовність CBHI підкреслена). cbh1Експресуючий вектор pTrex3g-cbh1 схематично зображений на фігурі 8А. Послідовність ДНК експресуючого вектора pTrex3g-cbh1 мала 10145 основ (SEQ ID NO:10), як показано на фігурі 8В. Був створений вектор для експресії білка CBHII генетично створеного ферменту. Вказаний вектор включав промотор cbhII, генетично створений ген cbhII, термінатор cbhII, ацетамідазу А. nidulans (amdS) як маркер, що селектується, і додаткову 3'-кінцеву фланкуючу послідовність термінатора cbhII. Даний вектор був створений з використанням шатл-вектора pCR-XL-TOPO (Invitrogen). Експресуюча частина вектора була вирізана з шатл-вектора шляхом розщеплення плазміди спеціальними рестрикційними ендонуклеазами NotI і SrfI, що утворюють фрагмент довжиною приблизно 10,68 т.п.о., який був використаний для трансформації Т. reesei. Вказаний вектор експресував ген cbhII Т. reesei, що кодує генетично створений білок з наступними мутаціями в амінокислотній послідовності: P98L, M134V, T154A, I212V, S316P і S413Y. Послідовність ДНК від ініціюючого кодону до термінуючого кодону мала 1416 основ (SEQ ID NO:11), як показано на фігурі 9А. Амінокислотна послідовність (SEQ ID NO:12) показана на фігурі 9В (сигнальна послідовність підкреслена). Вектор, що експресує cbhII, схематично зображений на фігурі 10А. Послідовність ДНК всього cbhII-експресуючого вектора pExp-cbhII мала 14158 основ (SEQ ID NO:11), як показано на фігурі 10В. Котрансформацію штаму Т. reesei з делецією cbh1 виконували, використовуючи три фрагменти ДНК. Генетично створений cbhII-експресуючий фрагмент був вирізаний з плазміди pExp-cbhII ферментами NotI і SrfI. Генетично створений ген cbh1 в експресуючому векторі pTrex3g використовували як матрицю для ПЛР з метою створення лінійного фрагмента тільки промотору cbhI, генетично створеного гена cbh1 і термінатора cbh1 (без маркера amdS). Злитий фрагмент cbh1-E1, описаний у попередньому прикладі, був використаний як матриця для ПЛР з метою створення лінійного фрагмента, що складається з промотору cbh1, злитого гена cbh1-E1 і термінатора 14 UA 104279 C2 5 10 15 20 25 30 35 40 cbh1 (без маркера amdS). Три вказаних фрагменти були використані для покривання частинок вольфраму у процесі біолітичної котрансформації. Вказану процедуру виконували відповідно до опису, наведеного у попередньому прикладі. У процесі котрансформації кожний з трьох фрагментів 1, 2 і 3 додавали до частинок вольфраму в об'ємі 1,5 мкл (концентрація ДНК 100300 нг/мкл). Відбір трансформантів здійснювали в ацетамідному середовищі ММ відповідно до наведеного опису. 7.3. Приклад 3. Аналіз активності, що розкладає целюлозу, трансформованих клонів Trichoderma reesei Активність, що розкладає целюлозу, злитого білка CBHI-EI визначали за допомогою наведених нижче аналізів з використанням вказаних субстратів. Штами Tr-A і Tr-D Trichoderma reesei були одержані з RL-P37 шляхом мутагенезу. Попередня обробка кукурудзяної соломи (PCS): кукурудзяну солому заздалегідь обробляли 2 % мас./мас. Н2SO4 відповідно до опису, наведеного у публікації Schell, D. et al., J. Appl. Biochem. Biotechnol. 105:69-86 (2003), і потім декілька разів промивали деіонізованою водою до досягнення рН, що дорівнює 4,5. Додавали ацетат натрію до кінцевої концентрації, що дорівнює 50 мМ, і титрували до рН 5,0. Вимірювання загального білка: концентрації білка вимірювали методом на основі біцинхонінової кислоти, використовуючи бичачий сироватковий альбумін як еталон (Smith, P.K., et al. (1985) Anal. Biochem. 150:76-85). Конверсію целюлози (визначення розчинного цукру) оцінювали за допомогою ВЕРХ, методами, описаними у публікації Baker et al., Appl. Biоchem. Biotechnol. 70-72:395-403 (1998). При виконанні експериментів використовували стандартний аналіз конверсії целюлози. У даному аналізі фермент і забуферений субстрат вміщували у контейнери та інкубували при визначеній температурі протягом визначеного періоду часу. Реакцію гасили 100 мМ гліцину, рН 11,0, з метою доведення значення рН реакційної суміші щонайменше до рН 10. Після того як реакція була погашена, аліквоту реакційної суміші фільтрували через 0,2-мікронну фільтрувальну мембрану для видалення твердих речовин. Фільтрований розчин аналізували відносно розчинних цукрів описаним вище методом ВЕРХ. Концентрація целюлози у реакційній суміші дорівнювала приблизно 7 %. Фермент або суміші ферментів вводили у кількості 1-60 мг загального білка на грам целюлози. У наведеній нижче таблиці 1 підсумовані дані, що свідчать про більш високу питому продуктивність чотирьохкомпонентного штаму у порівнянні з модифікованим штамом Tr-D. мг/г Чотирьохкомпонентний штам Модифікований штам Tr-D 10 9,5 5,1 20 14,2 8,1 PCS (13 %), SSC, 20 годин, 65 °C. У наведеній нижче таблиці 2 підсумовані дані, що свідчать про більш високу питому продуктивність трьохкомпонентного штаму у порівнянні зі штамом Tr-A. мг/г Трьохкомпонентний штам Штам Tr-А 15 61 45 10 45 31 PCS (13 %), SSC, 72 години, 59 °C. Всі посилання і публікації, наведені у даному описі винаходу, повністю включені в опис винаходу як посилання. Потрібно зазначити, що існують альтернативні способи здійснення даного винаходу. Таким чином, наведені варіанти здійснення винаходу потрібно розглядати як ілюстративні і такі, що не обмежують обсяг винаходу, тому даний винахід не обмежується конкретними варіантами і може бути модифікований у межах обсягу і еквівалентів доданої формули винаходу. 15 UA 104279 C2 16 UA 104279 C2 17 UA 104279 C2 18 UA 104279 C2 19 UA 104279 C2 20 UA 104279 C2 21 UA 104279 C2 22 UA 104279 C2 23 UA 104279 C2 24 UA 104279 C2 25 UA 104279 C2 26 UA 104279 C2 27 UA 104279 C2 28
ДивитисяДодаткова інформація
Назва патенту англійськоюHeterologous and homologous cellulase expression system
Автори російськоюBower, Benjamin, S., Larenas, Edmund, A.
МПК / Мітки
МПК: C12N 15/80, C12N 1/15, C12P 21/02
Мітки: гомологічних, целюлаз, експресії, система, гетерологічних
Код посилання
<a href="https://ua.patents.su/71-104279-sistema-ekspresi-geterologichnikh-i-gomologichnikh-celyulaz.html" target="_blank" rel="follow" title="База патентів України">Система експресії гетерологічних і гомологічних целюлаз</a>
Людська гібридна клітина для експресії гену ссавців, клітинна лінія людини нкв11, спосіб отримання гібридних людських клітин, придатних для експресїї гетерологічного протеїну

Номер патенту: 77642
Опубліковано: 15.01.2007
Автор: Чо М'юнг-Сам
МПК: C12N 5/10, C12N 5/02, C12N 5/16, C12N 5/06
Мітки: людини, гетерологічного, нкв11, придатних, людська, лінія, клітинна, протеїну, отримання, експресії, клітин, ссавців, спосіб, людських, гібридна, гену, гібридних, експрес, клітина
Формула / Реферат:
1. Клітина, отримана злиттям клітини людської ембріональної нирки , що має походження від 293 та клітини, що має походження від лімфоми Беркітта і є клітиною 2В8 (депозитний номер АТСС: CRL-12569).2. Клітина за п. 1, в якій клітина людської ембріональної нирки є клітиною, що має походження від 293S.3. Клітина за п. 1, яка експресує гетерологічний протеїн.4. Клітина за п. 3, в якій гетерологічний протеїн вибрано з групи,...
Saf-поліпептид, фрагмент днк, що кодує saf-поліпептид, рекомбінантна плазмідна днк для експресії safполіпептиду (варіанти), штам грибів streptomyces lividans, спосіб експресії saf-поліпептиду

Номер патенту: 27696
Опубліковано: 16.10.2000
Автори: Мартін Хуан Франціско, Гарсіа Томас Вігаль, Жіль Хосе Антоніо, Ортега Антоніо Даза
МПК: C07K 14/41, C07K 14/37, C12N 15/31, C12P 21/02, C12N 15/09, C12N 1/21, C07K 14/00, C07K 14/36
Мітки: плазмідна, streptomyces, кодує, варіанти, safполіпептиду, рекомбінантна, штам, saf-поліпептид, lividans, saf-поліпептиду, грибів, спосіб, днк, експресії, фрагмент
Текст:
...от штамма АТСС 10137 S. griseus. 7.2 кб Bgl II фрагмент от pULADI и 1 кб Bgl II фрагмент от pULAD3 субклониовали раздельно в обеих ориентациях в ВамНI-переваренной pUC 19. Все эти плазмидные конструкции проверялись в клетках Е. coli M103 и затем использовались для трансформирования Е. coli E15 (Sartiy и др. J. Bacteriol, 145:288-292,1981) и Е. coli A 1046, в В-ХР не было обнаружено никаких синих колоний, позволяющих предположить, что...
Спосіб одержання гетерологічних білкових речовин з рослинного матеріалу

Номер патенту: 72552
Опубліковано: 15.03.2005
Автори: Рескі Ральф, Горр Гільберт
МПК: C12N 15/09, C12P 21/02, C12N 15/82, C07K 14/52
Мітки: матеріалу, речовин, рослинного, білкових, одержання, гетерологічних, спосіб
Формула / Реферат:
1. Спосіб одержання гетерологічних білкових речовин з рослинного матеріалу, який відрізняється тим що, як рослинний матеріал використовують протонему тканини моху, причому білкові речовини одержують з культурального середовища без пошкодження продукуючих тканин або клітин.2. Спосіб за п. 1, який відрізняється тим, що білкові речовини, що вивільняються в культуральне середовище, є біологічно активними.3. Спосіб за п. 1 або 2,...
Похідна інсуліну або її фізіологічно прийнятна сіль, попередник похідної інсуліну, днк-послідовність, що кодує попередник, вектор експресії, субстрат, трансформований вектором експресії, фармацевтична композиція

Номер патенту: 65529
Опубліковано: 15.04.2004
Автори: Хаберманн Пауль, Гайзен Карл, Ертль Йоханн, Зайпке Герхард
МПК: C12P 21/02, C12N 1/19, C12N 1/15, C12N 5/10, C12N 15/17, C12N 1/21, A61K 38/28, C12N 15/09, C07K 14/62
Мітки: попередник, трансформований, експресії, композиція, фармацевтична, сіль, прийнятна, вектор, інсуліну, похідної, кодує, фізіологічно, похідна, вектором, субстрат, днк-послідовність
Формула / Реферат:
1. Похідна інсуліну або її фізіологічно прийнятна сіль формули І, (I)в якій позначають: (А1-А5) амінокислотні залишки в положеннях А1-А5 ланцюга А людського (SEQ ID № 1) або тваринного інсуліну, (А12-А19) амінокислотні залишки в положеннях А12-А19 ланцюга А людського (SEQ ID № 2) або тваринного інсуліну, А21 Asn, Asp,...
Очищений поліпептид, що містить антиген вірусу гепатиту с, полінуклеотид, вектор, клітини, система експресії, моноклональне антитіло, препарат поліклональних антитіл, нуклеотидна проба, аналітичні набори, спосі

Номер патенту: 50829
Опубліковано: 15.11.2002
Автори: Куо Джордж, Чу Кві-Лім, Хоутон Майкл
МПК: A61K 31/70, C07H 21/04, A61K 39/395, C07K 7/06, C12N 15/09, A61P 31/12, C12N 15/51, C12P 21/08, C12N 7/00, G01N 33/15, C07K 16/08, C07K 7/08, G01N 33/531, G01N 33/577, G01N 33/569, C12Q 1/70, C07K 14/18, C07K 14/10, C12Q 1/68, A61K 48/00, A61K 39/29, G01N 33/53, G01N 33/576, C07K 19/00, C12N 15/02, C07K 14/00, A61K 39/00, C12N 1/19, C07K 14/02, C07K 16/00, C12N 5/10, C12P 21/02
Мітки: гепатиту, препарат, поліклональних, проба, система, антитіл, антитіло, моноклональне, антиген, поліпептид, вірусу, нуклеотидна, набори, містить, експресії, клітині, вектор, полінуклеотид, спосі, очищений, аналітичні
Формула / Реферат:
1. Очищенный полипептид, содержащий антиген вируса гепатита С (HCV), причем указанный антиген имеет следующую аминокислотную последовательность:или часть указанной аминокислотной последовательности полипептида, являющейся антигеном HCV, причем указанная часть состоит из по меньшей мере 8 или по меньшей мере 9, или по меньшей мере 10 аминокислот.2. Полипептид по п. 1, отличающийся тем, что указанная...
Попередній патент: Сушильна установка для продуктів подрібнення деревини
Наступний патент: Великогабаритний пакувальний контейнер для бітуму
Випадковий патент: Спосіб відбору молочних корів за їх будовою тіла