Вакцина, що сприяє росту, яка базується на нейтралізуючому епітопі








Формула / Реферат
1. Ізольований пептид, що специфічно зв’язує анти-GDF8 антитіло, який складається з 50 або менше амінокислотних залишків, що включає
(а) амінокислотні залишки від 327 до 346 послідовності SEQ ID NO: 1, або
(b) амінокислотні залишки від 327 до 346 послідовності SEQ ID NO: 1, яка включає консервативну амінокислотну заміну; або
(с) амінокислотні залишки від 327 до 346 послідовності SEQ ID NO: 1, яка включає амінокислотні заміни у положеннях, вибраних з групи, яка складається з залишків 328, 329, 331, 333 та 335 та їх комбінацій, де
(i) амінокислотний залишок 328 являє собою His, Leu або Asn;
(ii) амінокислотний залишок 329 являє собою Gln або Lys;
(iii) амінокислотний залишок 331 являє собою Asn або Ser;
(iv) амінокислотний залишок 333 являє собою Arg або Lys; та/або
(v) амінокислотний залишок 335 являє собою Ser, Pro або Thr,
де у випадку (b) і (с) не більше, ніж п‘ять амінокислотних замін знаходяться між залишками амінокислот від 327 до 346; та пептид специфічно зв‘язується з моноклональним антитілом 788 пацюка.
2. Ізольований пептид за пунктом 1, який у випадку (а) включає амінокислотні залишки від 321 до 346 послідовності SEQ ID NO: 1.
3. Ізольований пептид за пунктом 2, який у випадку (а) включає амінокислотні залишки від 320 до 350 послідовності SEQ ID NO: 1.
4. Ізольований пептид за пунктом 3, який включає амінокислотні залишки від 312 до 361 послідовності SEQ ID NO: 1.
5. Ізольований пептид за пунктом 1, який у випадку (а) включає епітоп специфічної нейтралізації для анти-GDF8 антитіла.
6. Ізольований пептид за пунктом 5, в якому антитіло вибирають з групи, яка складається з анти-GDF8 моноклонального антитіла 788 пацюка та IgG фракції козячої анти-GDF8 поліклональної антисироватки.
7. Ізольований пептид за пунктом 1, який включає не більше, ніж одну аміноксилотну заміну між залишками 327 та 346 попередника GDF8, за умови, що пептид специфічно зв‘язується з моноклональним антитілом 788 пацюка.
8. Злитий білок, що специфічно зв‘язує анти-GDF8 антитіло, що включає пептид за пунктом 1, який складається з 50 або менше амінокислотних залишків, що включає амінокислотні залишки від 327 до 346 послідовності SEQ ID NO: 1 або антигенний субфрагмент цього пептиду, причому вищезгаданий пептид зв‘язаний за допомогою пептидного зв‘язку щонайменше з частиною іншого білка.
9. Молекула нуклеїнової кислоти, яка кодує пептид за пунктом 1, що специфічно зв‘язує анти-GDF8 антитіло.
10. Молекула нуклеїнової кислоти, яка кодує злитий білок за пунктом 8.
11. Молекула нуклеїнової кислоти за пунктом 9, яка включає послідовність нуклеїнової кислоти від нуклеотиду 1112 до нуклеотиду 1171 послідовності SEQ ID NO: 2.
12. Вектор експресії, що включає молекулу нуклеїнової кислоти за пунктом 9, яка кодує пептид за пунктом 1, який складається з 50 або менше амінокислотних залишків, що включає амінокислотні залишки від 327 до 346 послідовності SEQ ID NO: 1.
13. Клітина-хазяїн для одержання пептиду за п. 1, яка включає вектор експресії за пунктом 12.
14. Клітина-хазяїн за пунктом 13, яка є трансформованою вектором експресії за пунктом 12.
15. Клітина-хазяїн за пунктом 13, яка є еукаріотичною клітиною.
16. Спосіб одержання пептиду за пунктом 1, що включає етапи культивування клітини-хазяїна за пунктом 13 і експресії кодованого пептиду та виділення пептиду.
17. Вакцинна композиція для викликання анти-GDF8 імунної відповіді, що включає пептид за пунктом 1, який складається з 50 або менше амінокислотних залишків, що включає амінокислотні залишки від 327 до 346 послідовності SEQ ID NO: 1.
18. Вакцинна композиція для викликання анти-GDF8 імунної відповіді, що включає злитий білок за пунктом 8.
19. Вакцинна композиція за пунктом 17, що додатково включає ад‘ювант.
20. Спосіб викликання анти-GDF8 імунної відповіді у тварини, що включає введення тварині ефективної кількості вакцинної композиції за пунктом 17, яка включає пептид за пунктом 1, який складається з 50 або менше амінокислотних залишків, що включає амінокислотні залишки від 327 до 346 послідовності SEQ ID NO: 1.
21. Спосіб викликання анти-GDF8 імунної відповіді у тварини, що включає введення тварині ефективної кількості вакцинної композиції за пунктом 18.
22. Спосіб скринінгу на селекцію aнти-GDF8 aнтитіла або фрагмента антитіла серед множини антитіл або фрагментів антитіл, що мають відношення до антитіла анти-GDF8, що включає контактування пептиду за пунктом 1, який складається з 50 або менше амінокислотних залишків, що включає амінокислотні залишки від 327 до 346 послідовності SEQ ID NO: 1, зі зразком, який включає одне або множину антитіл або фрагментів антитіл, та визначення антитіла або фрагмента антитіла, що селективно зв‘язується з пептидом.
23. Спосіб знижувальної регуляції активності GDF8 у тварини, який включає введення антитіла або фрагмента антитіла тварині у кількості та протягом часу, що є ефективними для знижувальної регуляції активності GDF8 у тварини, де антитіло специфічно зв‘язується з пептидом за пунктом 1, який складається з 50 або менше амінокислотних залишків, що включає амінокислотні залишки від 327 до 346 послідовності SEQ ID NO: 1.
24. Спосіб знижувальної регуляції активності GDF8 у тварини, який включає імунізацію тварини за допомогою ефективної кількості вакцинної композиції за пунктом 17.
25. Спосіб знижувальної регуляції активності GDF8 у тварини, який включає імунізацію тварини за допомогою ефективної кількості вакцинної композиції за пунктом 18.
Текст
1. Ізольований пептид, що специфічно зв‟язує анти-GDF8 антитіло, який складається з 50 або менше амінокислотних залишків, що включає (а) амінокислотні залишки від 327 до 346 послідовності SEQ ID NO: 1, або (b) амінокислотні залишки від 327 до 346 послідовності SEQ ID NO: 1, яка включає консервативну амінокислотну заміну; або (с) амінокислотні залишки від 327 до 346 послідовності SEQ ID NO: 1, яка включає амінокислотні заміни у положеннях, вибраних з групи, яка складається з залишків 328, 329, 331, 333 та 335 та їх комбінацій, де (i) амінокислотний залишок 328 являє собою His, Leu або Asn; (ii) амінокислотний залишок 329 являє собою Gln або Lys; (iii) амінокислотний залишок 331 являє собою Asn або Ser; (iv) амінокислотний залишок 333 являє собою Arg або Lys; та/або (v) амінокислотний залишок 335 являє собою Ser, Pro або Thr, де у випадку (b) і (с) не більше, ніж п„ять амінокислотних замін знаходяться між залишками амінокислот від 327 до 346; та пептид специфічно зв„язується з моноклональним антитілом 788 пацюка. 2 (19) 1 3 93855 4 залишки від 327 до 346 послідовності SEQ ID NO: 1. 13. Клітина-хазяїн для одержання пептиду за п. 1, яка включає вектор експресії за пунктом 12. 14. Клітина-хазяїн за пунктом 13, яка є трансформованою вектором експресії за пунктом 12. 15. Клітина-хазяїн за пунктом 13, яка є еукаріотичною клітиною. 16. Спосіб одержання пептиду за пунктом 1, що включає етапи культивування клітини-хазяїна за пунктом 13 і експресії кодованого пептиду та виділення пептиду. 17. Вакцинна композиція для викликання антиGDF8 імунної відповіді, що включає пептид за пунктом 1, який складається з 50 або менше амінокислотних залишків, що включає амінокислотні залишки від 327 до 346 послідовності SEQ ID NO: 1. 18. Вакцинна композиція для викликання антиGDF8 імунної відповіді, що включає злитий білок за пунктом 8. 19. Вакцинна композиція за пунктом 17, що додатково включає ад„ювант. 20. Спосіб викликання анти-GDF8 імунної відповіді у тварини, що включає введення тварині ефективної кількості вакцинної композиції за пунктом 17, яка включає пептид за пунктом 1, який складається з 50 або менше амінокислотних залишків, що включає амінокислотні залишки від 327 до 346 послідовності SEQ ID NO: 1. 21. Спосіб викликання анти-GDF8 імунної відповіді у тварини, що включає введення тварині ефективної кількості вакцинної композиції за пунктом 18. 22. Спосіб скринінгу на селекцію aнти-GDF8 aнтитіла або фрагмента антитіла серед множини антитіл або фрагментів антитіл, що мають відношення до антитіла анти-GDF8, що включає контактування пептиду за пунктом 1, який складається з 50 або менше амінокислотних залишків, що включає амінокислотні залишки від 327 до 346 послідовності SEQ ID NO: 1, зі зразком, який включає одне або множину антитіл або фрагментів антитіл, та визначення антитіла або фрагмента антитіла, що селективно зв„язується з пептидом. 23. Спосіб знижувальної регуляції активності GDF8 у тварини, який включає введення антитіла або фрагмента антитіла тварині у кількості та протягом часу, що є ефективними для знижувальної регуляції активності GDF8 у тварини, де антитіло специфічно зв„язується з пептидом за пунктом 1, який складається з 50 або менше амінокислотних залишків, що включає амінокислотні залишки від 327 до 346 послідовності SEQ ID NO: 1. 24. Спосіб знижувальної регуляції активності GDF8 у тварини, який включає імунізацію тварини за допомогою ефективної кількості вакцинної композиції за пунктом 17. 25. Спосіб знижувальної регуляції активності GDF8 у тварини, який включає імунізацію тварини за допомогою ефективної кількості вакцинної композиції за пунктом 18. Перехресні посилання на заявки, пов'язані з даною Дана заявка представляє собою не попередню заявку, що заявляє пріоритет згідно з 35 U.S.C. § 119(е) попередньої заявки США з реєстраційним номером No. 60/533,719, що подана 31 грудня 2003 року, зміст яких введений у дану заявку як посилання у своїй цілісності. Галузь винаходу Винахід відноситься до білкового ростового фактору диференціації 8 та до фрагментів антигенного пептиду ростового фактору диференціації 8 та споріднених імуногенів, до вакцин ті способів лікування тварин для того, щоб модулювати активність ростового фактору диференціації 8. Передумови створення винаходу Ростовий фактор диференціації 8 представляє собою білок, який класифікується разом з надродиною трансформуючого фактора росту β («TGFβ»). Взагалі, білки надродини TGF-β спочатку експресуються як попередник (a/k/a прогормон), який піддається протеолітичному розщепленню у кластері основних залишків, що складається приблизно з 110-140 амінокислот С-термінального кінця білка-попередника. У кожному випадку активні або зрілівиди TGF-β, як вважають, представляють собою зв'язаний дисульфідним зв'язком димер Стермінальних ділянок розщепленого попередника білка. Ростовий фактор диференціації 8, далі GDF8, є також відомим як GDF-8 або міостатин. Гени, що кодують попередник GDF8 (далі називається «попередник GDF8»), були клоновані з широкого спектру організмів. Такі включають людський та мишачий попередник GDF8 [Nestor та ін., 1998, Рrос. Natl. Acad. Sci. 95: 14938-43; Патент США № 5,827,733, зміст яких введений у дану заявку як посилання у своїй цілісності]. Також було повідомлено про те, що імунореактивність GDF8 визначається у скелетній мускулатурі людини обох типів волокон 1 та 2. Антитіла та аналізи для визначення GDF8 є описаними наприклад, у Патенті США № 6,096,506. Було також показано, що GDF8 грає певну роль у знижувальній регуляції або інгібуванні росту та розвитку скелетної мускулатури, як підтверджено при використанні GDF8 кнок-аут мишей (McPherron та ін., 1997, Nature 387 83-90). З цієї причини були зроблені попередні спроби, зокрема, у галузі сільськогосподарського тваринництва, модулювати GDF8 активність у тварин за допомогою ряду способів з метою знижувальної регуляції активності GDF8 для того, щоб поліпшити ріст та/або відносну скелетну масу різноманітних м'ясо-молочних тварин. Наприклад, патент США № 6,399,312 описує промотор гену попередника GDF8 та аналіз з пропозицією, що цей аналіз можна використовувати для ідентифікації теоретичного інгібітору такого промотору. Патент США № 6,656,475 описує спосіб інгібування впливу GDF8 на клітину шляхом контакту клітини з продоменом GDF8, що конкурує 5 93855 за рецептор GDF8, у патенті також повідомляється, що С-термінальний кінець зрілого GDF8 може варіювати. Патент США № 6,004,937 описує застосування фолістатину як можливого антагоністу GDF8. Жоден з цих способів не приводить до якого-небудь практичного застосування у галузі сільськогосподарського тваринництва або при клінічному застосуванні (або у людини, або у ветеринарії). Були зроблені також інші спроби використати антитіла та вакцинну технологію для знижувальної регуляції функції GDF8. Наприклад, патент США № 6,369,201 [зміст якого введений у дану заявку як посилання у своїй цілісності], описує пептиди, тобто фрагменти білка GDF8, та вакцину, що викликає продукцію анти-GDF8 антитіл. Цей патент також повідомляє про точно не встановлений ступінь досягнення прибавки у рості та вазі у порівняні з контролями у гризунів, імунізованих деякими з описаних пептидних фрагментів GDF8. Були також описані інші фізіологічні ролі GDF8. Наприклад, патент США № 6,368,597 [зміст якого введений у дану заявку як посилання у своїй цілісності] передбачає, що інгібування функції GDF8 є корисним при лікуванні діабету типу II, наприклад, шляхом введення aнти-GDF8 антитіла або анти-GDF8 вакцини пацієнтові, який має такий стан. Проте залишається стара потреба у галузі техніки щодо поліпшення антигенів та імуногенів для того, щоб викликати aнти-GDF8 імунну відповідь, а 327 V Val 328 Η His 329 Q Gin 330 A Ala 331 N Asn 332 Ρ Pro 333 R Arg 334 G Gly 335 S Ser 336 A Ala У додатковому втіленні винаходу GDF8 пептид необов'язково включає консервативні заміни однієї амінокислоти. Тільки з метою прикладу, такі можуть бути вибрані з одного з, принаймні, п'яти положень амінокислот у пептиді. В окремому втіленні існує, принаймні, одна консервативна амінокислотна заміна, наприклад, між залишками 327 та 346 GDF8. В іншому втіленні GDF8 пептид включає консервативні амінокислотні заміни, зроблені не більше, ніж у п'яти амінокислотних положеннях, у межах пептиду. Ще в одному втіленні існують дві консервативні амінокислотні заміни між залишками 327 та 346 GDF8. Ще в одному втіленні існують три консервативні амінокислотні заміни між залишками 327 та 346 GDF8. Ще в одному втіленні існують чотири консервативні амінокислотні заміни між залишками 327 та 346 GDF8. Бажано, коли заміни амінокислотних залишків знаходяться в одному або більше положеннях стосовно природного людського попередника GDF8 (SEQ ID NO: 1), які позначені варіаціями амінокислот внутрішньовидового порівняння Фіг. 2. Такі знаходяться у положеннях залишків 328, 329, 331, 333 та 335 та їх комбінаціях, де (a) амінокислотний залишок 328 представляє собою His, Leu або Asn; (b) амінокислотний залишок 329 представляє собою Gin або Lys; 6 також щодо поліпшення GDF8 антитіл, здатних до високоспецифічного зв'язування з GDF8. Наведення будь-якого посилання у даній заявці не слід тлумачити як припущення, що таке посилання є доступним як «Рівень техніки» стосовно даної заявки. Короткий виклад суті винаходу Даний винахід вирішує ці та інші недоліки у галузі техніки шляхом забезпечення пептидів GDF8 (наприклад, пептидних фрагментів GDF8 розміром 50 залишків або менше), що включають специфічний нейтралізуючий епітоп для GDF8. В одному втіленні винаходу забезпечуються пептиди GDF8, які включають, наприклад, ізольований пептид, що містить частину від приблизно 312 залишку до приблизно 361 залишку природного людського попередника GDF8 (SEQ ID NO: 1). Переважно, GDF8 пептид, що є предметом винаходу, включає частину від приблизно 320 залишку до приблизно 350 залишку, більш переважно включає частину від приблизно 321 залишку до приблизно 346 залишку, та найбільш бажано включає частину від приблизно 327 залишку до приблизно 346 залишку природного людського попередника GDF8. Проілюстрований GDF8 пептид, який позначений як DJ5 у даній заявці нижче, продемонстрований так, як представлено нижче, згідно з однобуквеним кодом та згідно з трьохбуквеним кодом у відповідності з нумерацією залишків на основі попередника GDF8 послідовності SEQ ID NO: 1, для зручності. DJ5(SEQ ID NO: 8) 337 G Gly 338 Ρ Pro 339 С Cys 340 С Cys 341 Τ Thr 342 Ρ Pro 343 Τ Thr 344 К Lys 345 Μ Met 346 S Ser (c) амінокислотний залишок 331 представляє собою Asn або Ser; (d) амінокислотний залишок 333 представляє собою Arg або Lys; та/або (e) амінокислотний залишок 333 представляє собою Ser, Pro або Thr. Бажано, коли такі модифіковані пептиди GDF8 включають специфічний нейтралізуючий епітоп для анти-GDF8 антитіла та, таким чином, зберігають властивість специфічного зв'язування з антиGDF8 антитілом, де антитіло представляє собою МонАТ 788 та/або IgG фракцію цапиної анти-GDF8 поліклональної сироватки, як проілюстровано в даній заявці нижче. Ще в одному втіленні винаходу забезпечуються молекули нуклеїнової кислоти тобто полінуклеотиди, які кодують згадані вище пептиди GDF8. Бажано, коли молекули нуклеїнової кислоти включають частину існуючого в природі гену людського попередника GDF8, від приблизно нуклеотиду 1112 до приблизно нуклеотиду 1171 (депозитний номер Genebank NM_005259, людський ген GDF8, SEQ ID NO:2). Ця частина кодує пептид DJ5, як описано вище. Слід зазначити, що пропис NM_005259 включає значну частину послідовності, що фланкує власне кодуючу ділянку. Середній спеціаліст у даній галузі також зможе оцінити, що 7 93855 DJ5 відповідає нуклеотидам 979-1038 власне кодуючої ділянки прогормону GDF8. Здатні до реплікації клонуючі вектори та прокаріотичні або еукаріотичні хазяйські клітини, які включають молекули нуклеїнової кислоти, також забезпечуються разом зі способами для одержання GDF8 пептиду, що включає етапи культивування хазяйської(их) клітини(клітин), яка(які) експресують кодований пептид, та відновлення пептиду. Середній спеціаліст у даній галузі зможе оцінити, що GDF8 пептид також легко одержують за допомогою стандартного, добре відомого спеціалістові у галузі хімічного синтезу способу. Ще в одному додатковому втіленні винаходу також забезпечується вакцинна композиція, що включає GDF8 пептид згідно з винаходом (або його білок злиття) наприклад, така, що бажано включає один або більше ад'ювантів та інші загальноприйняті у галузі техніки елементи вакцинної композиції на основі пептиду або на основі білка. Також забезпечуються способи для забезпечення анти-GDF8 імунної відповіді у тварини, що включають введення тварині ефективної кількості вакцинної композиції. В іншому додатковому втіленні забезпечується спосіб скринінгу для селекції анти-GDF8 антитіла або фрагменту антитіла серед множини антитіл або фрагментів антитіл, який включає контакт пептиду з одним або множиною антитіл або фрагментів антитіла, та визначення антитіла або фрагменту антитіла, що селективно зв'язується з пептидом. Ще в одному аспекті даний винахід забезпечує спосіб знижувальної регуляції активності GDF8 у тварини. В одному такому втіленні спосіб включає введення антитіла або фрагменту антитіла тварині у кількості та протягом часу ефективних для зниAnas platyrhynchos (качка) AAL35275 Anser anser (гуска) AAL35276 Anser anser (гуска) AAR18246 Bos taurus (корова) ААВ86687 Canis familiaris (собака) AAR14343 Capra hircus (коза) AAR12161 Columba livia (голуб) AAL35277 Coturnix chinensis (перепел) AAL35278 Danio rerio (смугаста гірелла) ААВ86693 Equus caballus (кінь) ВАВ16046 Gallus gallus (курка) ААК18000 Gallus gallus (курка) AAR18244 Homo sapiens (людина) NP_005250 I. punctatus (сом) ААК84666 Lepus capensis (заєць) AAN8789 Macaca fascicularis (мавпа) AAL17640 Meleagris gallopavo (індик) ААВ86692 Morone chrysops (білий американський окунь) ААК28707 Mus musculus (домашня миша) ААС53167 O. mykiss (форель) ААК71707 Ovis aries (вівця) ААВ86689 Рарio hamadryas (бабуїн) ААВ86686 Rattus norvegicus (пацюк) ААВ86691 Salmo salar (лосось) САС19541 Spams aurata (морський карась) AAL05943 Sus scrofa (свиня) ААС08035 Sus scrofa (свиня) AAR18245 8 жувальної регуляції активності GDF8 у тварини, де антитіло специфічно зв‟язується з пептидом, або шляхом імунізації тварини за допомогою вакцинної композиції, як описано в даній заявці. Тварина бажано представляє собою хребетну тварину та більш бажано ссавця, птаха або рибу. Бажано, коли ссавець представляє собою домашню тварину (наприклад, таку що використовується у тваринництві або альтернативно домашню тварину), але може необов'язково бути людиною, яка потребує такої знижувальної регуляції. Винахід також розглядає злиті білки, що включають GDF8 пептиди згідно з винаходом. Злиті білки можуть включати домени, які є сигнальними пептидами для поліпшеної секреції або експресії поверхнею клітини GDF8 злитого білка та/або для того, щоб дозволити проводити очистку за допомогою селективної зв'язувальної системи. Крім того, злиті білки розглядаються як такі, що зв'язуються з одним або більше доменами пептиду GDF8. В окремому втіленні злитий білок згідно з даним винаходом включає GDF8 пептид, який складається з 50 або менше амінокислотних залишків, що включає амінокислотні залишки від 327 до 346 послідовності SEQ ID NO:1. У спорідненому втіленні такого типу злитий білок включає антигенний субфрагмент GDF8 пептиду, наприклад, GDF8 пептид, що включає приблизно 10 суміжних амінокислотних залишків DJ5 (SEQ ID NO: 8). Короткий опис фігур Фігура 1 ілюструє пептиди, які перекриваються, для DJ1 - DJ7, в активній ділянці GDF8 (тобто у зрілому GDF8), яка знаходиться між залишками 266 та 375 послідовності попередника GDF8. Порівнювані пептиди мають наступні ідентифікаційні номери послідовностей (SEQ ID NO :11) (SEQ ID NO :12) (SEQ ID NO :13) (SEQ ID NO :14) (SEQ ID NO :15) (SEQ ID NO :16) (SEQ ID NO :17) (SEQ ID NO :18) (SEQ ID NO :19) (SEQ ID NO :20) (SEQ ID NO :21) (SEQ ID NO :22) (SEQ ID NO :8) (SEQ ID NO :23) (SEQ ID NO :24) (SEQ ID NO :25) (SEQ ID NO :26) (SEQ ID NO :27) (SEQ ID NO :28) (SEQ ID NO :29) (SEQ ID NO :30) (SEQ ID NO :31) (SEQ ID NO :32) (SEQ ID NO :33) (SEQ ID NO :34) (SEQ ID NO :35) (SEQ ID NO :36) 9 Детальний опис винаходу Таким чином, даний винахід ідентифікує GDF8 домени, що служать як специфічні епітопи нейтралізації для поліклональної анти-GDF8 цапиної антисироватки та для певних інших специфічних aнти-GDF8 антитіл. Ці епітопи також служать для забезпечення фрагментів GDF8 білка, які є корисними для забезпечення активної та специфічної імунної відповіді стосовно GDF8 білків, як in vitro, наприклад, для визначення GDF8 білка, так й in vivo, для знижувальної регуляції активності GDF8. Ці фрагменти у загальному випадку згадуються у даній заявці як GDF8 пептиди або пептидні фрагменти. Використання цих GDF8 пептидів включає використання імуногенів для того, щоб викликати анти-GDF8 імунну відповідь у тварин, та використання специфічних зв'язувальних антитіло мішеней у аналізах, пов'язаних з GDF8. Специфічні зв'язувальні епітопи GDF8 були ідентифіковані шляхом контакту анти-GDF8 антисироватки з рядом GDF8 пептидів, що перекриваються, та визначення ступеня зв'язувальної активності між пептидами та IgG антитілами анти сироватки. GDF8 антисироватку одержували від цапа, імунізованого за допомогою попередника білка GDF8, що має структуру, оптимізовану для експресії та антигенності. Для того, щоб більш повно оцінити даний винахід, забезпечуються наступні визначення. Застосування термінів в описі в однині для зручності ніяким чином не призначене для обмеження. Таким чином, наприклад, посилання на те, що композиція містить «поліпептид», включає посилання на один або більше таких поліпептидів. Як такий, що використовується у даній заявці, термін «приблизно» використовується почергово з терміном «близько» та означає, що значення знаходиться в межах двадцяти процентів від зазначеного значення, тобто, пептид, який містить «приблизно» 50 амінокислотних залишків, може містити від 40 до 60 амінокислотних залишків. Також є зрозумілим, що цей винахід не є обмеженим окремими конфігураціями, етапами процесу та матеріалами, розкритими у даній заявці як такі, конфігурації, етапи процесу та матеріали можуть до деякої міри варіювати. При цьому також зрозуміло, що використовувана термінологія застосовується тільки з метою опису окремих втілень, але не призначена для обмеження, оскільки об'єм даного винаходу буде обмежуватися тільки прикладеними пунктами та їх еквівалентами. Як використовується в даній заявці, термін «поліпептид» використовується почергово з терміном «білок» та означає полімер, який включає дві або більше амінокислоти, зв'язані пептидними зв'язками. Бажано, коли (якщо не вказано інше) термін «поліпептид» відрізняється від терміну «пептид», як використовується в даній заявці, розміром або довжиною ланцюга, при цьому «пептид» відноситься до полімерного ланцюга, що містить приблизно п'ятдесят або менше амінокислот, терміни «поліпептид» або «білок» відноситься до полімерного ланцюга, що включає більше, ніж приблизно п'ятдесят амінокислот. Необов'язково, 93855 10 пептид або поліпептид може не мати певних амінокислотних залишків, що кодуються геном або мРНК. Наприклад, ген або молекула мРНК можуть кодувати послідовність амінокислотних залишків на N-термінальному кінці поліпептиду (тобто сигнальну послідовність), яка відщеплюється від заключного білка та таким чином, може не бути його частиною. «GDF8 пептид» згідно з даним винаходом представляє собою відносно короткий фрагмент, який має походження від GDF8 білка. Навіть субфрагменти таких пептидів можуть називатися GDF8 пептидами. Не маючи на меті обмежувати максимальний розмір GDF8 пептиду згідно з винаходом, є бажаним, коли максимальний розмір пептиду складає приблизно 50 залишків, більш бажано, коли максимальний розмір пептиду складає приблизно 30 залишків, та ще більш бажано, коли максимальний розмір пептиду складає приблизно 25 залишків. У загальному випадку GDF8 пептид бажано має розмір, який коливається у межах від приблизно 10 до приблизно 50 амінокислотних залишків у довжину, більш переважно від приблизно 15 до приблизно 30 амінокислотних залишків, та, зокрема, складає 20 амінокислотних залишків у довжину. GDF8 пептиди, які є меншими субфрагментами інших GDF8 пептидів згідно з даним винаходом, наприклад, GDF8 пептид, що включає приблизно 10 суміжних амінокислотних залишків з DJ5 (SEQ ID NO:8), переважно включає антигенну частину (наприклад, епітоп) з більшого GDF8 пептиду. В окремому втіленні GDF8 пептид включає пептидний домен, який має ступінь гомології, що коливається від приблизно 50% до 100% гомологічності з пептидом, який визначається номерами залишків 327-346 (SEQ ID NO:8) існуючого в природі попередника GDF8 (SEQ ID NO:1). Варіації у гомології зазначені вище, є переважно консервативними замінами та/або варіаціями, що зберігають антигенну структуру згідно з винаходом, яка специфічно впізнається певними специфічними анти-GDF8 антитілами. Ці консервативні заміни представляють собою заміни, що показані за допомогою порівняння на внутрішньовидову гомологію (див , наприклад, Фіг. 2) для збереження функції GDF8 та/або представляють заміни залишків між амінокислотами аналогічних хімічних (наприклад, фізичних) та електронних структур, зберігаючи, таким чином, та/або оптимізуючи зв'язувальну специфічність GDF8 пептидів згідно з винаходом. Приклади таких консервативних амінокислотних замін включають заміну одного гідрофобного залишку іншим гідрофобним залишком, таким як ізолейцин, валін або метіонін, або заміну одного полярного залишку еквівалентного заряду на інший, наприклад, заміну лізину аргініном, аспарагінової кислоти глутаміновою кислотою, або аспарагіну глутаміном. Зокрема, GDF8 пептид згідно з винаходом буде також включати специфічний епітоп нейтралізації для анти-GDF8 антитіла, тобто епітоп або антигенний домен, який буде специфічно зв'язуватися з PGA анти-GDF8 IgG поліклонального анти 11 тіла, описаного у Прикладах, представлених нижче, та/або такий, що специфічно зв'язується з комерційно доступним моноклональним антитілом (монАТ) Кат. № МАВ788 (R&D Systems Inc., Minneapolis, MN). Терміни «очищений» та «ізольований», які використовується в даній заявці, відносяться до матеріалів, розділених за умов, які знижують або усувають присутність небажаних матеріалів, тобто, забруднюючих речовин, включаючи нативні матеріали, з яких цей матеріал одержаний. Наприклад, очищений або ізольований білок бажано є вільним від інших білків або нуклеїнових кислот, з якими він може бути знайдений у клітині. Очищений матеріал може містити менше, ніж приблизно 50%, бажано менше, ніж приблизно 75%, та найбільш бажано менше, ніж приблизно 90%, клітинних компонентів, з якими він за походженням асоційований. Чистота може бути оцінена за допомогою хроматографії, гель-електрофорезу, імуноаналізу, композиційного аналізу, біологічного аналізу та інших способів, відомих у галузі техніки. З функціонального аспекту ізольований GDF8 пептид згідно з винаходом представляє собою такий, який в достатній мірі є відокремленим від інших матеріалів, включаючи попередник GDF8 білка та/або зрілий GDF8 білок, так, що є здатним викликати імунну відповідь, що є специфічною для GDF8 пептиду. Способи очистки є добре відомими в області техніки. Наприклад, нуклеїнові кислоти можуть бути очищені за допомогою преципітації, хроматографії, ультрацентрифугування та інших способів. Білки та поліпептиди, а також пептиди, можуть бути очищені за допомогою різноманітних способів, включаючи без обмеження, препаративний диск-гель, електрофорез, ізоелектричне фокусування, ВЕРХ, ВЕРХ зі зворотною фазою, гельфільтрацію, іонообмінну хроматографію та роздільну хроматографію, преципітацію та висолювальну хроматографію, екстракцію та протитоковий розподіл. Для деяких цілей є бажаним одержувати поліпептид у рекомбінантній системі, в якій білок містить додаткову послідовність мітки, що сприяє очищенню, таку, як, але без обмеження, послідовність полігістидину або послідовність, яка специфічно зв'язується з антитілом, таку, як FLAG® та GST. Поліпептид може бути потім очищений з сирового лізату або хазяйської клітини за допомогою хроматографії на прийнятному твердофазному матриксі. Альтернативно, антитіла, або його зв'язувальні фрагменти, одержані проти поліпептиду, можуть використовуватися як реагенти очистки. Термін «суттєво чистий» означає найвищий ступінь чистоти, який може бути досягнений при використанні традиційних методик очистки, які відомі в області техніки, та означає нуклеїнову кислоту, поліпептид, пептид або інший матеріал, що є вільним від інших забруднюючих білків, нуклеїнових кислот та інших біологічних речовин, які мають походження від організму початкового джерела або системи для експресії рекомбінантної ДНК. Суттєва чистота може бути оцінена за допомогою стандартних способів та буде типово перевищувати, принаймні, приблизно 75%, бажано, 93855 12 принаймні приблизно 90%, більш бажано, принаймні, приблизно 95% та найбільш бажано, принаймні, приблизно 99% чистоти. Оцінка чистоти може бути здійснена на масовій або молярній основі. «Полінуклеотид» або «молекула нуклеїнової кислоти» представляє собою молекулу, яка включає нуклеотиди, включаючи, але не обмежуючись такими, послідовності РНК кДНК, геномної ДНК та навіть синтетичної ДНК. Ці терміни також призначені для охоплення молекул нуклеїнових кислот, що включають будь-які відомі в області техніки аналоги ДНК та РНК. «Вектор» або «вектор реплікації» представляє собою реплікон, такий, як плазміда, фаг або косміда, до яких сегмент ДНК може бути приєднаний, або в який він може бути вбудований, так, щоб викликати реплікацію приєднаного сегменту. Термін також включає вбудований або приєднаний сегмент ДНК, що представляє інтерес. Вектори, які можуть використовуватися у цьому винаході, включають мікробні плазміди, віруси, бактеріофаги, здатні до інтеграції фрагменти ДНК та інші носії, що можуть поліпшити інтеграцію нуклеїнових кислот у геном хазяїна. Плазміди представляють собою форму вектора, що найбільш часто використовується, але усі інші форми векторів, які виконують еквівалентну функцію та які існують або є відомими в області техніки, є прийнятними для використання у даній заявці. Див., наприклад, Pouwels та ін., Клонуючі вектори: лабораторний практикум, 1985 та Додаток, Elsevier, Ν. Υ., та Rodriguez та ін (ред), Вектори: огляд молекулярних клонуючих векторів та їх застосування, 1988, Buttersworth, Boston, MA. Вбудовування ДНК, що кодує GDF8 пептид(и) згідно з винаходом, у вектор легко здійснити, коли обидва кінці ДНК та вектора включають сумісні рестрикційні сайти. Якщо це не можна здійснити, то може бути необхідно модифікувати кінці ДНК та/або вектора шляхом зворотного перетравлювання одноланцюгових виступаючих кінців ДНК, одержаних шляхом розщеплення рестрикційною ендонуклеазою для одержання тупих кінців, або шляхом досягнення того самого результату за допомогою заповнення одноланцюгових кінців прийнятною ДНК полімеразою. Альтернативно, бажані сайти можуть бути одержані, наприклад, шляхом лігування нуклеотидних послідовностей (лінкерів) у термінальні кінці. Такі лінкери можуть включати специфічні олігонуклеотидні послідовності, які визначають бажані рестрикційні сайти. Рестрикційні сайти можуть також бути одержані при використанні полімеразної ланцюгової реакції (ПЛР). Див., наприклад, Saiki та ін., Science 239 487 (1988). Розщеплений вектор та фрагменти ДНК можуть також бути модифіковані, якщо це є бажаним, шляхом гомополімерного розтягування. Рекомбінантні експресійні вектори, що використовуються у цьому винаході, є типово конструкціями ДНК або РНК, що самореплікуються, та які включають нуклеїнові кислоти, що кодують один або більше GDF8 пептид(ів) згідно з винаходом, що звичайно оперативно зв'язані з прийнятними елементами генетичного контролю, що є здатними до регуляції експресії нуклеїнових кислот у прийн 13 ятних хазяйських клітинах. Елементи генетичного контролю можуть включати прокаріотичну промоторну систему або еукаріотичну систему контролю експресії та типово включають транскрипційний промотор, необов'язковий оператор для контролю початку транскрипції, енхансери транскрипції для підвищення рівня експресії мРНК, послідовність, що кодує прийнятний сайт зв‟язування з рибосомою та послідовності термі нації, транскрипції та трансляції. Експресійні вектори можуть також містити джерело реплікації, що дозволяє вектору незалежно реплікуватися у хазяйські клітини. Експресія нуклеїнових кислот, які кодують GDF8 пептид(и) згідно з винаходом, може проводитися за допомогою традиційних способів або у прокаріотичних, або в еукаріотичних клітинах. «Кодуючі послідовності» ДНК або «послідовність, що кодує» певний білок або пептид, представляє собою послідовність ДНК, яка транскрибується та транслюється у поліпептид in vitro або in vivo, коли розміщується під контролем прийнятних регуляторних елементів. Межі кодуючої послідовності визначаються стартовим кодоном на 5'термінальному кінці та трансляційним стопкодоном на 3'-термінальному кінці. Кодуюча послідовність може включати, але не обмежена, прокаріотичні послідовності, кДНК, одержані з еукаріотичної мРНК, послідовності геномної ДНК з еукаріотичних (наприклад ссавцевих) ДНК та навіть синтетичні послідовності ДНК. Послідовність термінації транскрипції буде звичайно розміщуватися 3' стосовно кодуючої послідовності. Як використовується в даній заявці, терміни «злитий білок» та «злитий пептид» використовуються почергово та охоплюють «химерні білки та/або химерні пептиди» та злиті «інтеїнові білки/пептиди». Злитий білок включає, принаймні, частину GDF8 пептиду згідно з даним винаходом, зв'язану за допомогою пептидного зв'язку з частиною іншого білка. Наприклад, злиті білки можуть включати маркерний білок або пептид або білок або пептид, які сприяють ізоляції та/або очищенню та/або антигенності GDF8 пептиду згідно з даним винаходом. GDF8 злитий білок може включати, принаймні, частину білка, відмінного від GDF8, зв'язану за допомогою пептидного зв'язку з, принаймні, частиною GDF8 поліпептиду. У бажаних втіленнях GDF8 є функціональним, тобто зберігає антигенність. He-GDF8 послідовності можуть бути аміно- або карбокситермінальними по відношенню до послідовностей GDF8. Рекомбінантна молекула ДНК, яка кодує такий злитий білок, включає послідовність, яка кодує, принаймні, частину He-GDF8 білка, сполученого у рамці з кодуючою послідовністю GDF8, та може кодувати сайт розщеплення для специфічної протеази, наприклад, тромбіну або фактору Ха, бажано у місці сполучення або поблизу місця сполучення між послідовністю GDF8 та послідовністю, відмінною від GDF8. У специфічному втіленні злитий білок експресується в клітині СНО. Такий злитий білок може використовуватися для ізоляції GDF8 пептидів згідно з даним винаходом при використанні афінної колонки, що є специфічною для білка та/або мітки, злитої з GDF8 пептидом. Очи 93855 14 щений GDF8 пептид, наприклад, може потім вивільнятися зі злитого білка при використанні протеолітичного ферменту та сайту розщеплення, такого, який був згаданий вище. В одному такому втіленні химерний GDF8 пептид може бути одержаний, наприклад, у вигляді білка, злитого з глутатіон-S-трансферазою (GST), злитого з білком, зв'язуючим мальтозу (МВР), або білка, міченого полігістидином, для експресії у будь-якій клітині або, альтернативно, у безклітинній системі. Наприклад, GST зв'язує глутатіон, кон'югований з твердим матриксом носія, МВР зв'язується з мальтозним матриксом, а полігістидинові хелати з Νі-хелатіоном матриксу носія. Злитий білок може бути елюйований зі специфічного матриксу за допомогою прийнятних буферів або шляхом обробки протеазою, специфічною для сайту розщеплення, який звичайно створюють між GDF8 пептидом та партнером злиття (наприклад, GST, МВР, FLAG®) як проілюстровано нижче, або полі-His, як описано вище. «Гетерологічна нуклеотидна послідовність», як використовується у даній заявці, представляє собою нуклеотидну послідовність, що додається до нуклеотидної послідовності згідно з даним винаходом за допомогою рекомбінантних способів, з утворенням нуклеїнової кислоти, яка звичайно не утворюється у природі. Такі нуклеїнові кислоти можуть кодувати злиті (наприклад, химерні) білки. Таким чином, гетерологічна нуклеотидна послідовність може кодувати пептиди та/або білки, що володіють регуляторними та/або структурними властивостями. В іншому такому втіленні гетерологічна нуклеотидна послідовність може кодувати білок або пептид, що функціонує як засіб для визначення білка або пептиду, який кодується нуклеотидною послідовністю згідно з даним винаходом після того, як рекомбінантна нуклеїнова кислота експресується. Ще в одному втіленні гетерологічна нуклеотидна послідовність може функціонувати як засіб для визначення нуклеотидної послідовності згідно з даним винаходом. Гетерологічна нуклеотидна послідовність може включати некодуючі послідовності, включаючи рестрикційні сайти, регуляторні сайти, промотори, тощо. Термін «хазяйська клітина» означає клітину, яка містить або яка є здатною для вміщення та експресії екзогенної молекули нуклеїнової кислоти, або транзієнтно або постійно. Клітина є «трансформованою» за допомогою екзогенної ДНК, коли така екзогенна ДНК була введена через клітинну мембрану. Екзогенна ДНК може або не може бути інтегрована (ковалентно зв'язана) у хромосомну ДНК, комплектуючи геном клітини. У прокаріотах та дріжджах, наприклад, екзогенна ДНК може підтримуватися як епісомальний елемент, такий, як плазміда. Стосовно еукаріотичних клітин стабільна трансформована клітина представляє собою таку, в яку екзогенна ДНК була інтегрована у хромосому так, що вона успадковується дочірніми клітинами шляхом хромосомної реплікації. Ця стабільність демонструється здатністю еукаріотичної клітини до утворення клітинних ліній або клонів, які включають популяцію дочірніх клітин, що містять екзогенну ДНК. 15 Прокаріоти включають як грам-негативні, так і грам-позитивні організми, наприклад, Е. соlli та В. subtilis. Вищі еукаріоти включають відомі лінії клітин, одержані з клітин тварин, які мають походження як від видів тварин, відмінних від ссавців, наприклад, клітин комах, та птахів, так і ссавців, наприклад, людини, приматів та гризунів. Прокаріотичні системи хазяїн-вектор включають широку різноманітність векторів для багатьох різноманітних видів. Як використовується в даній заявці, Е. соli та вектори на її основі будуть використовуватися в загальному випадку для включення в це поняття еквівалентних векторів, які використовуються в інших прокаріотичних організмах. Типовий вектор для ампліфікації ДНК представляє собою pBR322 або багато його похідних. Вектори, що можуть використовуватися для експресії GDF8 та/або GDF8 пептидів, включають, але не обмежені такими, що містять Іас промотор (pUC-серій); trp промотор (pBR322-trp); Ірр промотор (plN-серій); лямбда-рР або pR промотори (pOTS); або гібридні промотори, такі, як ptac (pDR540). Див. Brosius та ін., «Експресійні вектори, що використовують промотори, які мають походження від лямбда, trp, lac Ірр», в Rodriguez та Denhardt (ред.). Вектори: огляд молекулярних клонуючих векторів та їх застосування, 1988, Buttersworth, Boston, стор. 205-236. Дріжджі, а також клітини тканинної культури вищих еукаріот представляють собою бажані хазяйські організми для рекомбінантного одержання GDF8 пептидів згідно з винаходом, та/або антиGDF8 антитіл та/або фрагментів цих антитіл. Незважаючи на те, що можуть використовуватися будь-які клітини тканинної культури вищих еукаріот, включаючи бакуловірусні експресійні системи комах, клітини ссавців є переважними. Трансформація або трансфекція та розмноження таких клітин стало звичайною процедурою. Приклади корисних клітинних ліній включають клітини HeLa, клітинні лінії яєчника китайського хом'яка (СНО), клітинні лінії нирки дитинчати пацюка (BRK), клітинні лінії комах, клітинні лінії птахів та клітинні лінії мавпи (COS). Експресійні вектори для таких клітинних ліній включають, наприклад, джерело реплікації, промотор, сайт ініціації трансляції, сайт сплайсингу РНК (якщо використовується геномна ДНК), сайт поліаденілування та сайт термінації транскрипції. Ці вектори також звичайно містять селективний ген або ген для ампліфікації. Прийнятні експресійні вектори можуть бути плазмідами, вірусами або ретровірусами, що несуть промотори, які мають походження, наприклад, від таких джерел, як аденовіруси, SV40, парвовіруси, вірус вакцини або цитомегаловірус. Характерні приклади прийнятних експресійних векторів включають pCR®3.1, pCDNA1, pCD [Okayama та ін., Моl. Cell Biol. 5: 1136 (1985)], pMCIneo Poly-A [Thomas та ін., Cell 51: 503 (1987)], pUC19, pREP8, pSVSPORT та їх похідні, а також бакуловірусні вектори, такі, як рАС 373 або рАС 610. Прокаріотичні послідовності контролю експресії, що типово використовуються, включають промотори, у тому числі такі, що мають походження 93855 16 від β-лактамази, та системи на основі лактозного промотору [Chang та ін., Nature, 198: 1056 (1977)], системи на основі триптофанового (trp) промотору [Goeddel та ін., Nucleic Acids Res. 8: 4057 (1980)], системи на основі лямбда PL промотору [Shimatake та ін., Nature, 292: 128 (1981)] та tac промотору [De Boer та ін., Proc. Natl. Acad. Sci. USA 292: 128 (1983)]. Багаточисленні експресійні вектори, що містять такі контрольні послідовніості, є відомими в області техніки та є комерційно доступними. «Оперативно зв'язаний» відноситься до розміщення елементів, де компоненти, описані таким чином, розміщені так, щоб виконувати свою звичайну функцію. Таким чином, контрольні елементи, оперативно зв'язані з кодуючою послідовністю, є здатними впливати на експресію кодуючої послідовності. Контрольні елементи повинні бути суміжними з кодуючою послідовністю, оскільки вони функціонують для направлення її експресії. Таким чином, наприклад, перехідні послідовності, які не транслюються, але транскрибуються, можуть бути присутніми між промотором та кодуючою послідовністю, а промотор може все ще вважатися «оперативно зв'язаним» з кодуючою послідовністю. Винахід також включає поліклональні та моноклональні (монАТ) антитіла, що специфічно зв'язуються з GDF8 пептидами згідно з винаходом. Як використовується в даній заявці, термін «антитіло» відноситься до імуноглобуліну та/або його фрагментів. Існуючий в природі імуноглобулін складається з одного або більше поліпептидів, які суттєво кодуються генами імуноглобуліну. Визнані імуноглобулінові гени включають гени каппа, лямбда, альфа, гамма, дельта, епсілон та мю константних ділянок, а також міріади генів варіабельних ділянок імуноглобуліну. Антитіло або антитіла згідно з винаходом також охоплюють фрагменти антитіла, тобто, фрагменти, що зв'язують антиген, наприклад, Fv, Fab та F(ab')2, сконструйовані одноланцюгові зв'язувальні білки, (наприклад, Huston та ін., Рrос. Natl. Acad. Sci. U.S.A., 85, 5879-5883 (1988) та Bird та ін., Science, 242, 423-426 (1988), зміст яких введений в дану заявку як посилання у своїй цілісності), а також біфункціональні гібридні антитіла (наприклад, Lanzavecchia та ін., Eur. J. Immunol. 17, 105 (1987)). [Див., для загального огляду Hood та ін., Immunology, Benjamin, Ν.Υ., 2-е вид. (1984), Harlow та Lane, Антитіла. Лабораторний практикум. Cold Spring Harbor Laboratory (1988) та Hunkapiller та Hood, Nature, 323, 15-16 (1986), зміст яких введений в дану заявку як посилання у своїй цілісності]. Наприклад, сироватка, яка одержана від тварин, імунізованих за допомогою GDF8 пептидів згідно з винаходом при використанні стандартних способів, може використовуватися безпосередньо, або IgG фракція може відокремлюватися від сироватки при використанні стандартних способів, таких, як плазмафорез або абсорбційна хроматографія з lgG-специфічними адсорбентами, такими, як імобілізований Протеїн А або Протеїн G. Альтернативно, можуть бути одержані моноклональні антитіла та, необов'язково, зв'язувальні фрагменти або рекомбінантні зв'язувальні білки, які мають 17 походження від таких монАТ. Такі монАТ або їх фрагменти можуть необов'язково бути гуманізовані за допомогою способів, які відомі в галузі техніки. Гібридоми, що продукують монАТ, які селективно зв'язуються GDF8 пептидами згідно з винаходом, одержують за допомогою добре відомих методик. Звичайно процес втягує злиття іморталізованої клітинної лінії з B-лімфоцитом, який виробляє бажане антитіло. Альтернативно, можуть використовуватися методики, які не передбачають злиття, для одержання безсмертних ліній клітин, які продукують антитіла, наприклад, трансформація, індукована вірусами [Casali та ін., Science 234: 476 (1986)]. Іморталізовані клітинні лінії звичайно представляють собою трансформовані клітини ссавців, зокрема, клітини мієломи гризунів, корів та такі, що мають людське походження. Найбільш часто мієломні клітинні лінії пацюків або мишей використовуються як такі, що є зручними та доступними. Методики для одержання лімфоцитів, які продукують антитіла, одержані від ссавців, яким введено антигени, є добре відомими. В загальному випадку використовуються лімфоцити периферичної крові (PBL), якщо застосовують клітини людського походження, а клітини селезінки або лімфатичних вузлів використовують з джерел ссавців, які мають походження, відмінне від людини. Хазяйську тварину піддають ін'єкції за допомогою повторюваних дозувань очищеного антигену (людські клітини сенсибілізують in vitro), і тварина виробляє бажані клітини, які продукують антитіло перед тим, як їх піддають збору для злиття з іморталізованою клітинною лінією. Методики для злиття є також добре відомим у галузі техніки, вони включають перемішування клітин з агентом злиття, таким, як поліетиленгліколь. Гібридоми відбирають за допомогою стандартних процедур, таких, як HAT (гіпоксантинаміноптерин-тимідин) селекції. Такі, що секретують бажане антитіло, відбирають при використанні стандартного імуноаналізу, такого, як Вестернблоттінг, ELISA (твердофазний імуноферментний аналіз), RIA (радіоімуноаналіз) або за допомогою інших. Антитіла відновлюють із середовища при використанні стандартних методик для очистки білка [Tijssen, Практика та теорія ферментного імуноаналізу (Elsevier, Amsterdam, 1985)]. Багато посилань є доступними для забезпечення керівництва для застосування вказаних вище методик [Kohler та ін., Гібридомні технології (Cold Spring Harbor Laboratory, New York, 1980); Tijssen, Практика та теорія ферментного імуноаналізу (Elsevier, Amsterdam, 1985); Campbell, Технологія моноклональних антитіл (Elsevier, Amsterdam, 1984); Hurrell, Моноклональні гібридомні антитіла: методики та застосування (CRC Press, Boca Raton, FL, 1982)]. Моноклональні антитіла можуть також бути одержані при використанні добре відомих систем на основі фагових бібліотек. Див., наприклад, Huse, та ін., Science 246: 1275 (1989); Ward, та ін., Nature, 341: 544 (1989). Антитіла, які продукуються, або поліклональні, або моноклональні, можуть використовуватися, 93855 18 наприклад, в імобілізованій формі, у зв'язаному з твердим носієм вигляді за допомогою добре відомих способів для очищення GDF8 пептидів шляхом імуноафінної хроматографії. Антитіла проти GDF8 пептидів можуть також використовуватися в неміченій або міченій формі при використанні стандартних способів, як основа для проведення імуноаналізів для визначення або кількісного визначення GDF8. Специфічна мітка, яка використовується, буде залежати від типу імуноаналізу. Приклади міток, що можуть використовуватися, включають, але не обмежені, радіоакти32 125 3 14 вні мітки, такі, як Р, l, Н та С; флуоресцентні мітки, такі, як флуоресцеїн та його похідні, родамін та його похідні, дансил та умбеліферон, хемілюмінісцентні речовини, такі як люциферія та 2,3дигідрофталазиндіони та ферменти, такі, як пероксидаза хрону, лужна фосфатаза, лізоцим та глюкоза-6-фосфатдегідрогеназа. Антитіла можуть бути мічені такими мітками за допомогою відомих способів. Наприклад, агенти злиття, такі, як альдегіди, карбодііміди, дималеїміди, імідати, сукцинімідати, бісдіазотизований бензадин та подібні їм, можуть використовуватися для мічення антитіл за допомогою флуоресцентних, хемілюмінісцентних або ферментативних міток. Загальні способи, які використовуються, є добре відомими в області техніки та є описаними, наприклад, в роботі Імуноаналіз: практичне керівництво, 1987, Chan (ред.), Academic Press, Inc., Orlando, FL. Такі імуноаналізи можуть проводитися, наприклад, на фракціях, одержаних під час очистки рецепторів. Антитіла згідно з даним винаходом можуть також використовуватися для ідентифікації окремих клонів кДНК, які експресують споріднені з GDF8 поліпептиди в експресійних клонуючих системах. Нейтралізуючі антитіла, які є специфічними для ліганд-зв'язувального сайту рецептора, також можуть використовуватися як антагоністи (інгібітори) для блокування або знижувальної регуляції функції GDF8. Такі нейтралізуючі антитіла можуть бути легко ідентифіковані за допомогою звичайних експериментів, як проілюстровано в Прикладах, які приведені нижче. Антагонізм GDF8 активності може бути здійснений при використанні повних молекул антитіла або за допомогою добре відомих фрагментів, що зв'язують антиген, таких, як Fab, Fc, F(ab)2 та Fv фрагментів. Визначення таких фрагментів може бути знайдене в даній заявці вище, або, наприклад так, як описано у Klein, Імунологія (John Wiley, New York, 1982); Parham, Розділ 14, у Weir, ред. Імунохімія, 4-е вид. (Blackwell Scientific Publishers, Oxford, 1986). Застосування та одержання фрагментів антитіла було також описане, наприклад, Fab фрагментів [Tijssen, Практика та теорія імуноаналізів (Elsevier, Amsterdam, 1985)], Fv фрагментів [Hochman та ін., Biochemistry 72: 1130 (1973); Sharon та ін., Biochemistry 75: 1591 (1976); Ehrlich та ін., патент США № 4,355,023] половинок молекул антитіла (Auditore-Hargreaves, патент США № 4,470,925). Способи одержання рекомбінантних фрагментів Fv на основі відомих послідовностей варіабельних ділянок важких та легких ланцюгів 19 антитіла були також описані, наприклад, Moore та ін. (патент США №4,642,334) та Pluckthun [Biotechnology 9: 545 (1991)]. Альтернативно, вони можуть бути синтезовані за допомогою стандартних способів. Даний винахід також охоплює анти-ідіотипічні антитіла, як поліклональні, так і моноклональні, які одержують при використанні описаних вище антитіл як антигенів. Ці антитіла є корисними, оскільки вони можуть імітувати структури лігандів. Пептидний синтез Оскільки GDF8 пептиди згідно з винаходом, наприклад, DJ5 пептид, проілюстрований в даній заявці нижче, є відносно короткими (наприклад, переважно 50 амінокислотних залишків або менше), вони можуть бути одержані за допомогою відомих у галузі техніки способів пептидного синтезу. Синтетичні пептиди або поліпептиди, одержані при використанні добре відомих методик твердої фази, рідкої фази або методик пептидної конденсації, або будь-якої їх комбінації, можуть включати природні та неприродні амінокислоти. Амінокислоти, що використовуються для пептидного синтезу, можуть бути стандартними Вос (Nальфа-аміно захищений Nальфа-т-бутилоксикарбоніл) амінокислотними смолами зі стандартними прописами для зняття захисту, нейтралізації, конденсації та промивання вихідної процедури твердофазного синтезу Merrifield [J. Am. Chem. Soc, 85: 2149-2154 (1963)], або нестабільних до дії основи Nальфа-аміно захищених 9флуоренілметоксикарбоніл (Fmoc) амінокислот, які вперше були описані Саrріnо та Han [J. Org. Chem., 37: 3403-3409 (1972)]. Як Fmoc, так і Вос Nальфа-аміно захищені амінокислоти можуть бути одержані від Fluka, Bachem, Advanced Chemtech, Sigma, Cambridge Research Biochemical, Bachem, або Peninsula Labs, або інших хімічних компаній, добре відомих тим, хто працює в даній області техніки. У доповнення до цього, спосіб згідно з винаходом може використовуватися при застосуванні інших Nальфа-захисних груп, які добре відомі тим, хто працює в даній області техніки. Твердофазний пептидний синтез може проводитися за допомогою методик, знайомих тим, хто працює в цій галузі техніки та які забезпечуються, наприклад, у Stewart та Young, Твердофазний синтез, Друге видання, Pierce Chemical Co., Rockford, III. (1984); Fields та Noble, Int. J. Pept. Protein Res. 35: 161-214 (1990), або при використанні автоматичних синтезаторів, таких, як ті, що продаються ABS [Applied Biosystems, 850 Lincoln Centre Drive, Foster City, CA 94404 USA]. Таким чином, GDF8 пептиди згідно з винаходом можуть включати Dамінокислоти, комбінацію D-та L-амінокислот та різноманітні «сконструйовані» амінокислоти (наприклад, бета-метил амінокислоти, Сальфа-метил амінокислоти та Nальфа-метил амінокислоти, тощо) для передачі спеціальних властивостей. Синтетичні амінокислоти включають орнітин для лізину, фторфенілаланін для фенілаланіну та норлейцин для лейцину або ізолейцину. У доповнення до цього, за допомогою використання специфічних амінокислот на специфічних етапах конденсації мо 93855 20 жуть бути одержані, альфа-спіралі, бета-витки, бета-складки, гамма-витки та циклічні пептиди. Анти-GDF8 антисироватка Способи згідно з винаходом включали процес скринінгу GDF8 пептидів проти поліклональної анти-GDF8 антисироватки. За допомогою цього процесу ідентифікували епітоп, з яким певні антиGDF8 антитіла зв'язуються з високим ступенем специфічності. Анти-GDF8 антисироватку одержували шляхом імунізації тварин за допомогою попередника GDF8. Ген попередника GDF8 модифікували для забезпечення форми, оптимізованої для експресії та імуногенності. Наприклад, послідовність природного GDF8 прогормону (SEQ ID NO: 2) оптимізували для експресії в експресійних системах ссавців та вірусів. Крім того, вірусний хазяїн відключав механізми, які втягують транскрипційний контроль, РНК стабільність (сплайсинг) та подібні. Ці зміни роблять нуклеїнову кислоту менш подібною до хазяїна та більш подібною до вірусу. Крім того, послідовність ДНК переважно конструювали як таку, що настільки відрізняється від послідовності нуклеїнової кислоти ссавців, наскільки це є можливим. Наприклад, амінокислотну послідовність попередника GDF8 піддавали зворотній трансляції при використанні кодонів, переважних для дріжджів. Одержану послідовність досліджували на кодони, що зберігають свою гомологію до послідовності нуклеїнової кислоти GDF8 людини. Там, де це було можливим, ці кодони замінювали наступними найбільш переважними дріжджовими кодонами, які кодують ту саму амінокислоту. Одержаний оптимізований ген (SEQ ID NO: 3) може бути експресований у будь-якій прийнятній системі хазяїна, включаючи, наприклад, відомі в області техніки експресійні системи комах, ссавців, бактерій, вірусів та дріжджів. Наприклад, експресійні системи на основі клітин комах, такі, як бакуловірусні системи, є добре відомими та описані, наприклад, Summers та Smith, Texas Agricultural Experiment Station Bulletin No. 1555 (1987). Матеріали та способи для експресійних систем на основі бакуловірусів/клітин комах є комерційно доступними у формі наборів від, серед інших, Invitrogen, San Diego Calif. («MaxBac» набір). Подібно до цього бактеріальні експресійні системи та експресійні системи ссавців є добре відомими в області техніки та описані, наприклад, Sambrook та ін. (Молекулярне клонування: лабораторний практикум; клонування ДНК, Том. І та II; D. N. Glover ред.). Дріжджові експресійні системи є також відомими в області техніки та описані, наприклад, в роботі Генетична інженерія на основі дріжджів (Ваrr та ін., ред., 1989) Butterworths, London. Багато інших таких експресійних систем є відомим в області техніки та є доступними комерційно у формі наборів. Як проілюстровано в даній заявці, модифікований ген попередника GDF8 (SEQ ID NO: 3) експресували в Flp-ln™ CHO експресійній системі (Invitrogen, Carlsbad, СА) так, як описано більш детально у Прикладі 1, нижче. Як попередник GDF8 білка, так і пептиди зрілого GDF8 білка, можуть бути введені у будь-яку 21 вакцинну композицію, сумісну з білками або пептидами. Такі вакцинні композиції є добре відомими в області техніки та включають, наприклад, фізіологічно сумісні буфери та сольові розчини, тощо, а також ад'юванти, як описано більш детально в даній заявці нижче. Вакцинна композиція, яка включає попередник GDF8 білка, використовується для одержання антисироватки для скринінгу та ідентифікації специфічного нейтралізуючого епітопу для анти-GDF8 антитіла. Як проілюстровано в даній заявці, очищений попередник GDF8 білка, експресований за допомогою вектора, що включає SEQ ID NO:3, вводили цапу у складі вакцинної композиції, що включає 1 г попередника GDF8 білка, емульгованого у повному ад'юванті Фрейнда (CFA). Вакцинну композицію переважно вводили підшкірно (SC), глибше шкіри цапа. Подальші бустерні імунізації є бажаними. Такі можуть вводитися при прийнятних додаткових інтервалах при використанні такої самої або зниженої дози білка, наприклад, при інтервалах, які коливаються від 2 до 5 тижнів після початкової ін'єкції. Починаючи приблизно з двох тижнів після початкової ін'єкції, але бажано, починаючи з більш тривалого періоду часу, наприклад, від трьох до п'ятнадцяти тижнів, або більше, сироватку збирали, у разі необхідності, від імунізованої тварини. Зібрану сироватку потім переважно очищали та/або фракціонували за допомогою традиційних процедур очистки імуноглобуліну, таких, як, наприклад, хроматографія на основі протеїну А - Сефарози, протеїну G - агарози, гідроксилапатиту, гельелектрофорезу, діалізу або афінної хроматографії з прийнятним лігандом. Як проілюстровано в даній заявці, IgG фракція потім є доступною для скринінгу проти ряду пептидів зрілого GDF8, як описано більш детально за допомогою Прикладу 3 у даній заявці нижче. Епітопи, що зв'язуються з GDF8 Прийнятні анти-GDF8 моноклональні або поліклональні антитіла приводили у контакт з GDF8 білком протягом періоду часу, достатнього для того, щоб антитіло селективно зв'язалося з білком. Потім за допомогою GDF8 біоаналізів підтверджували, що антитіло нейтралізувало суттєво усю активність GDF8 білка. З цією метою може використовуватися будь-який GDF8 біоаналіз, хоча, як представлено в даній заявці нижче у Прикладі 3, in vitro аналіз транскрипційної активності згідно з Thies та ін., 2001, (Growth Factors 18, 251) є бажаним. В загальному випадку GDF8 пептид, який є корисним як антиген або зв'язувальний епітоп згідно з винаходом, включає послідовність від приблизно 312 залишку до приблизно 361 залишку GDF8 (SEQ ID NO: 1). Зокрема, пептид згідно з винаходом включає послідовність від приблизно 320 залишку до приблизно 350 залишку GDF8 (SEQ ID NO: 1). Пептид бажано включає послідовність від 327 до приблизно 346 залишку GDF8 (SEQ ID NO: 1). Спеціаліст в даній галузі зможе оцінити, що GDF8 пептид згідно з винаходом може бути легко модифікований для включення, принаймні, однієї 93855 22 консервативної заміни амінокислоти у будь-якому положенні. Бажано, коли пептид специфічно зв'язується з моноклональним антитілом 788 пацюка, як проілюстровано в даній заявці нижче. Такі консервативні заміни можуть включати, наприклад варіації у залишках 328, 329 та 335, та їх комбінації, де амінокислотний залишок у положенні 328 представляє собою His, Leu, Asn або Val; амінокислотний залишок у положенні 329 представляє собою Lys або Leu; a амінокислотний залишок у положенні 335 представляє собою Ser або Pro, або Thr. Залишки 328, 329 та 335 попередника GDF8 варіюють у межах послідовності GDF8 білка серед різних видів, як проілюстровано на Фіг.2, проте зрілий GDF8 залишається функціональним. Фіг. 1 ілюструє карту активної ділянки GDF8 (що формує зрілий білок) у контексті його білкового попередника. Накладеними на карту активної ділянки GDF8 є положення семи пептидів, що перекриваються. Ці пептиди, що перекриваються, були сконструйовані для того, щоб забезпечити мішені для ідентифікації епітопу, що зв'язує антитіло або епітопів GDF8. Пептид, позначений як DJ5, було ідентифіковано за допомогою скринінгу з IgG фракцією представленої цапиної анти-GDF8 антисироватки як тільки один значущий зв'язувальний епітоп GDF8 для проілюстрованої антисироватки. Цей пептид мав послідовність (SEQ ID NO: 8), що відповідає залишкам від 327 до 346 попередника GDF8 (SEQ ID NO: 1). Вакцинна композиція на основі GDF8 пептиду GDF8 пептиди, описані вище, переважно рецептували у вакцинні композиції. Ці вакцинні композиції можуть використовуватися для імунізації тварини для того, щоб викликати специфічну антиGDF8 імунну відповідь. Результат імунізації буде представляти собою знижувальну регуляцію функції GDF8 в імунізованій тварині. Такі вакцинні композиції є добре відомим в області техніки та включають, наприклад, фізіологічно сумісні буфери, консерванти та сольовий розчин, тощо, а також ад'юванти. «Ад'юванти» представляють собою агенти, які неспецифічно підвищують імунну відповідь на певний антиген, знижуючи, таким чином, кількість антигену, що є необхідною у будь-якій даній вакцині, та/або частоту проведення ін'єкцій, яка необхідна для того, щоб одержати адекватну імунну відповідь на антиген, який представляє інтерес. Прийнятні ад'юванти для вакцинації тварин включають, але не обмежені, Ад'ювант 65 (що містить олію земляного горіху, манідмоноолеат та алюмініймоностеарат); повний або неповний ад'ювант Фрейнда; мінеральні гелі, такі, як гідроксид алюмінію, фосфат алюмінію та галуни; сурфактанти, такі, як гексадециламін, октадециламін, лізолецитин, бромід диметилдіоктадециламонію, N,Nдіоктадецил-N',N'-біс(2-гідроксиметил)пропандіамін, метоксигексадецилгліцерин та поліоли плюроніка; поліаніони, такі, як піран, декстрансульфат, полі lС, поліакрилову кислоту та карбопол; пептиди, такі, як мурамілдипептид, диметилгліцин та туфтсин; та масляні емульсії. Білок або пептиди можуть також вводитися після введення у ліпоосоми або інші мікроносії. Інформація, яка стосу 23 93855 ється ад'ювантів та різних аспектів імуноаналізів, розкрита, наприклад, у роботах Р. Tijssen, Практика та теорія ферментного імуноаналізу, 3-є вид., 1987, Elsevier, New York, що введено в дану заявку як посилання. Вакцинна композиція включає достатню кількість бажаного імуногену, такого, як GDF8 пептиди згідно з винаходом, для того, щоб викликати імунну відповідь. Кількість, що вводиться, може коливатися від приблизно 0,0001 г/кг до приблизно 1,0 г/кг, по відношенню до маси тварини. Будь-яка прийнятна хребетна тварина легко використовується для одержання поліклональної антисироватки. Бажано, коли тварина представляє собою ссавця, що включає таких представників, як миші, пацюки, кролі, конячі, собачі, котячі, коров'ячі, овечі, наприклад, кози та вівці, примати, наприклад, мавпи, людиноподібні мавпи та люди, тощо. Вакцинна композиція легко вводиться будьяким стандартним способом, включаючи внутрішньовенний, внутрішньом'язовий, підшкірний, інтраперитонеальний, in ovo (зокрема, для птахів), та/або перорально. Для видів риби способи введення вакцинної композиції або імуногенної композиції включають наведені вище, а також занурення риби у воду, яка включає антигенну концентрацію пептиду, піддання риби обробці аерозолями, що містять антигенну концентрацію пептиду, при швидкому видаленні риби з води, тощо. Спеціаліст у даній галузі зможе оцінити, що вакцинна композиція переважно рецептується прийнятним чином для кожного типу реципієнтної тварини та способу введення. Прийнятні тварини можуть включати диких тварин, худобу (наприклад, таку, що вирощується для одержання м'яса, молока, масла, яєць, хутра, шкіри та/або вовни), в'ючних тварин, дослідних тварин, домашніх тварин, а також таких, що вироТаксономічна назва Coregonus clupeaformis Coregonus hoyi Oncorhynchus keta Oncorhynchus gorbuscha Oncorhynchus kisutch Oncorhynchus masou Oncorhynchus nerka Oncorhynchus tshawytscha Prosopium cylindraceum Oncorhynchus clarki Oncorhynchus mykiss Salmo salar Salmo trutta Salmo trutta X S. fontinalis Salvelinus alpinus Salvelinus confluentus Salvelinus fontinalis Salvelinus leucomaenis Saivelinus malma Salvelinus namaycush Thymallus thymallus 24 щуються для/у зоопарку, диких умовах та/або природних ареалах. В окремому втіленні тварина представляє собою людиноподібну мавпу, таку, як горила, або людину. В одному переважному втіленні тварина представляє собою тварину, що «забезпечує продукти харчування», та результат імунізації полягає у досягненні прибавки у вазі тварини, зокрема, м'язової маси, у порівнянні з неімунізованою твариною. Для цілей даного винаходу термін «тварина, що забезпечує продукти харчування» слід розуміти як такий, що включає усіх тварин, які вирощуються для споживання людиною та/або іншими тваринами, або для одержання товарів споживання, таких, як яйця або молоко. Необмежувальний список таких тварин включає птахів (наприклад, курей, індичок, качок, гусей, страусів), корів (наприклад, яловичої/телячої худоби, молочних корів, розведення бичків, буйволів), свиней (наприклад, кабанів або поросят), овечих (наприклад, кіз або овець), конячих (наприклад, коней), а також водних тварин, включаючи молюсків та рибу, таку, як форель або лосось та інші види, які вирощуються або виловлюються для споживання людиною. Для цілей згідно з даним винаходом термін «риба» слід розуміти для включення, без обмеження класифікаційної групи риб Teleosti, тобто телеостів. Як загін Salmoniformes (що включає родину Salmonidae), так і загін Perciformes (що включає родину Centrarchidae) містяться у класифікаційній групі Teleosti. Приклади потенціальних рибних реципієнтів включають родину Salmonidae, родину Serranidae, родину Sparidae, родину Cichlidae, родину Centrarchidae, трьохлінійну ронку (Parapristipoma trilineatum), блакитноокого плекостомуса (Plecostomus spp). Родина Salmonidae Загальна назва Сиг озерний Оселедець Кета Горбуша Кичуж Сима Нерка Чавича Коник Лосось Кларка Райдужна форель Атлантичний лосось Річкова форель Тигрова гібридна форель Арктичний голець Великоголовий голець Кумжа Японський голець (голець з білими плямами) Мальма (голець міябі) Озерна форель Харіус 25 93855 26 Деякі члени родини Serranidae Таксономічна назва Centropristis ocyurus Centropristis philadelphicus Centropristis striata Diplectrum bivittatum Diplectrum formosum Epinephelus flavolimbatus Epinephelus morio Serranus phoebe Serranus tortugarum Загальна назва Морський береговий окунь Морський кам'яний окунь Чорноморський окунь Карликовий піщаний окунь Піщаний окунь Морський жовтий каймистий окунь Червоний морський окунь Татлер Вапняковий окунь Деякі члени родини Sparidae Таксономічна назва Archosargus probatocephalus Archosargus rhomboidalis Calamus penna Lagodon rhomboides Pagrus Major Spams aurata Stenotomus chrysops Загальна назва Кейпкодський карась Червоний морський карась Центральноамериканський карась Колюча чопа Морський карась Красного моря Золотистий спар Скап Деякі члени родини Cichlidae Таксономічна назва Aequidens latifrons Cichlisoma nigrofasciatum Crenichichla sp. Pterophyllum scalare Tilapia mossambica Oreochromis sp. Sarotherodon aurea Загальна назва Красивий пельматохроміс Конголезька цихліда Щукоподібна цихліда Риба-ангел Мозамбікська тилапія Тилапія Золота тилапія Деякі члени родини Centrarchidae Таксономічна назва Ambloplites rupestris Centrarchus macropterus Elassom everglades Elassoma okefenokee Elassoma zonatum Enneacanthus gloriosus Enneacanthus obesus Lepomis auritus Lepomis cyanellus Lepomis cyanellus X L gibbosu Lepomis gibbosus Lepomis gulosus Lepomis humilis Lepomis macrochirus Lepomis megalotis Micropterus coosae Micropterus dolomieui Micropterus punctulatus Micropterus salmoides Pomoxis annularis Pomoxis nigromaculatus Загальна назва Американський кам'яний окунь Вухастий окунь Еверглейдський карликовий сонячний окунь Окефенокський карликовий сонячний окунь Каймистий карликовий сонячний окунь Синьоплямистий сонячний окунь Каймистий сонячний окунь Червоногрудий сонячний окунь Зелений сонячний окунь Зелений сонячний окунь Звичайна сонячна риба Звичайна сонячна риба Муловий морський окунь Оранжевоплямиста сонячна риба Синьозяберна сонячна риба Рожева сонячна риба Стайова сонячна риба Малоротий окунь Плямистий морський окунь Великоротий окунь Біла краппі Чорна краппі 27 У додатковому бажаному втіленні тварина представляє собою домашніх тварин або людину, а вакцину вводять для забезпечення довгострокової знижувальної регуляції GDF8 для будь-яких ветеринарних або медичних цілей, які пов'язані з такою знижувальною регуляцією GDF8. Для цілей даного винаходу під терміном «домашні тварини» слід розуміти усіх тварин - коней (конячих), котів (котячих), собак (собачих), гризунів (включаючи мишей, пацюків, морських свинок), видів кролів та птахів, таких, як голуби, папуги, тощо. Інші птахи, які одержують таку вакцинацію або антитіла, можуть бути пов'язані або з комерційним або некомерційним птахівництвом. Такі включають, наприклад, Anatidae, такі, як лебеді, Columbidae, наприклад, голуби, такі, як домашні голуби, Phasianidae, наприклад, сірі куріпки та шотландські куріпки, Thesienidae, Psittacines, наприклад, довгохвості папуги, папуги ара та папуги, наприклад, які вирощуються як домашні улюбленці або для колекціонування, та членів родини Ratite. В іншому бажаному втіленні будь-яка із вказаних вище тварин (переважно відмінних від людини) піддається імунізації для того, щоб одержати анти-GDF8 антитіла, що специфічно зв'язуються з пептидами згідно з винаходом, та утворені антитіла збирають для використання в аналізах та/або у ветеринарії та медицині, наприклад, для забезпечення знижувальної регуляції GDF8 для будь-яких ветеринарних або медичних цілей, які пов'язані з такою знижувальною регуляцією GDF8. Даний винахід можна краще зрозуміти шляхом посилання на наступні необмежувальні приклади, які забезпечуються як ілюстрація винаходу. Наступні приклади є присутніми для того, щоб більш повно проілюструвати втілення винаходу, але вони ніяким чином не можуть вважатися такими, що обмежують широкий об'єм даного винаходу. Приклади Приклад 1 Матеріали та способи А. Експресія та очистка попередника GDF8 (GDF8 прогормону) Природну послідовність ДНК попередника GDF8 або прогормону (SEQ ID NO: 2) оптимізували для експресії в експресійних системах ссавців та вірусів. Для уникнення негативних ефектів вірусного вимикання механізмів хазяїна, послідовність ДНК була призначена бути настільки дивергентною стосовно послідовностей нуклеїнової кислоти ссавців, наскільки це можливо. Для здійснення цього амінокислотну послідовність прогормону GDF8 піддавали зворотній трансляції при використанні кодонів, переважних для дріжджів. Одержану послідовність досліджували на кодони, що зберігають свою гомологію до послідовності нуклеїнової кислоти GDF8 людини. Там, де це було можливим, ці кодони замінювали наступними найбільш переважними дріжджовими кодонами, які кодують ту саму амінокислоту. Одержану молекулу нуклеїнової кислоти (SEQ ID NO: 3) комерційно синтезували для вбудовування у прийнятні експресійні вектори. Flp-ln™ CHO експресійну систему (Invitrogen, Carlsbad, CA) використовували для експресії оп 93855 28 тимізованого GDF8 прогормону. Коротко, конструювали конструкцію GDF8 прогормону, що містить С-термінальний FLAG® (Sigma-Aldrich Corp., St. Louis, MO) злитий епітоп одержували шляхом вбудовування гену, що кодує модифікований GDF8 прогормон, у плазміду pCMVtag4B (Siratagene, San Diego, CA.). FLAG® злита мітка сприяє розділенню FLAG® злитих білків на анти-FLAG® гелевій колонці. ПЛР ДНК фрагмент, що містить модифікований ген GDF8 прогормону-FLAG®, потім клонували в плазмідний експресійний вектор pcDNA5/FRT (Invitrogen, Carlsbad, CA). Одержання Flp-ln™ CHO клітинної лінії, що експресує GDF8 прогормонFLAG® злитий білок, здійснювали шляхом сумісної трансфекції Flp-ln™ CHO лінії клітин за допомогою Flp-ln™ експресійного вектора, що містить ген GDF8-FLAG® та Flp рекомбіназну експресійну плазміду, POG44. FIp рекомбіназа опосередковує вбудовування Flp-ln експресійної касети в геном при інтегрованому FRT сайті шляхом сайтспецифічної рекомбінації ДНК. Стабільну клітинну лінію, що експресує та секретує GDF8 прогормон, що містить FLAG® епітоп, одержували при використанні селекції за допомогою гігроміцину В. Стабільну лінію клітин СНО, що експресує GDF8 прогормон, яка містить FLAG® мітку, адаптували до суспензійної культури у середовищі, вільному від сироватки, при використанні стандартних методик. Кондиціоноване середовище, що містить секретований GDF8 прогормон, одержували при використанні WAVE біореакторної системи (WAVE Biotech LLC, Bridgewater, NJ). Очищення міченого FLAG® GDF8 прогормону проводили за допомогою очистки при використанні афінної хроматографії, застосовуючи анти-FLAG® М2 афінний гель (Sigma-Aldrich Corp., St. Louis, MO). В. Очищення DJ5 специфічного антитіла DJ5 (SEQ ID NO: 8; див. Таблицю 2, нижче) специфічну фракцію антитіл очищали за допомогою афінної хроматографії. Афінну колонку готували шляхом конденсації 10 мг DJ5 синтетичного пептиду з 0,8 г Сефарози 4В, активованої ціаногенбромідом (Sigma Genosys, Woodlands, TX). Колонку промивали та врівноважували за допомогою PBS. Використовували приблизно 11 мл цапиної IgG фракції (10 мг/мл) для 25 мл PBS. Фракції по 1,0 мл збирали та досліджували на абсорбцію при довжині хвилі 280 нм. Зв'язаний матеріал елюювали за допомогою приблизно 10 мл 0,2 Μ гліцину (pH 1,85). Фракції 1,0 мл збирали та нейтралізували за допомогою 0,25 мл 0,5 Μ фосфату натрію, 0,75 Μ NaCI, pH 7,4. Аліквоти розміром приблизно 25 мкл зв'язаних фракцій 1-10 та зв'язаних фракцій 25-35 піддавали аналізу на ELISA реактивність стосовно DJ5 пептиду. Незв'язані фракції виявляли як такі, що були негативними щодо DJ5 реактивності. Зв'язані фракції демонстрували сильний пік реактивності до DJ5 пептиду. Незв'язані фракції 111 та зв'язані фракції 26-34 розділяли на пули. Розділені на пули зразки концентрували та замінювали їх буферний розчин на фосфатнобуферний сольовий розчин (PBS), як вказано нижче. Концентрації зразків визначали за допомогою способу на основі оптичної густини OD при 280 нм (Сучасні прописи в імунології. 2,7.3, John Wiley & 29 Sons, Inc.). Незв'язані зразки доводили до концентрації 10 мг/мл, а зв'язані зразки доводили до концентрації 1 мг/мл для подальшого використання. Приклад 2 Цапина aнти-GDF8 поліклональна IqG сироватка Цапиний анти-попередник GDF8 IgG одержували від імунізованого цапа за допомогою наступних способів. A. Імунізація цапа Цапа породи Саанен (молочна порода) (вік приблизно 2 роки, самець) імунізували за допомогою очищеного рекомбінантного GDF8 прогормону (одержаний так, як описано у Прикладі 1, вище) так, як описано нижче. 0,5 мг білка емульгували у повному ад'юванті Фрейнда (CFA) та вводили підшкірно (SC) цапу. Наступні бустерні імунізації, що містили 0,3 мг білка, емульгованого в неповному ад'юванті Фрейнда (IFA), проводили підшкірно на третьому, шостому та десятому тижнях. Збирали кров з яремної вени при використанні шприца та поміщали у термос. Кров збирали у пробірки, що містять антикоагулянт та центрифугували при 2500 RPM протягом 20 хвилин для видалення еритроцитів. Плазму повторно піддавали видаленню кальцію для одержання сироватки. Зразок сироватки, який збирали через 15 тижнів після початкової імунізації, використовували для подальшого аналізу. B. Одержання та очистка поліклонального IgG Сироватку одержували від цапа через 15 тижнів, та IgG фракцію очищали з цієї сироватки так, як описано нижче. lgG фракцію цапиної сироватки очищали на колонці з Протеїн G агарозою згідно з прописом виробника (Kirkegaard and Perry Laboratories, Inc., Gaithersburg, MD). Елюйовані фракції розділяли на пули, концентрували та замінювали буфер фосфатно-буферним сольовим розчином (PBS), використовуючи Centriprep центрифужні фільтри (Centriprep YM-10, Millipore Corporation, Billerica, ΜΑ). Концентрації зразків визначали за допомогою способу визначення оптичної густини OD при 280 нм (Сучасні прописи в імунології, див. вище) та доводили концентрацію до 10 мг/мл. Приклад 3 Характеристика цапиної антисироватки Цапину антисироватку, яка забезпечувалася у Прикладі 2, приведеному вище, позначали як PGA. Очікувалося, що PGA IgG фракція містить антитіла, направлені проти різних епітопів у молекулі GDF8 прогормону. PGA антисироватку характеризували за допомогою in vitro аналізу активації транскрипції так, як описано нижче. In vitro аналіз активації транскрипції, який використовували для кількісного вимірювання GDF8 біонейтралізації, суттєво представляє собою такий Thies та ін. (Growth Factors 18, 251 (2001)). Вміст 96 комірок планшетів для аналізу культури тканин (PerkinElmer Life and Analytical Sciences, Inc., Boston, MA), оброблених за допомогою люмінометру ViewPlate™, висівали при концентрації 1,0 5 10 клітин/комірка клітин А204 рабдосаркоми (АТСС НТВ-82) та інкубували при 37°С, 5% СО2, у зволоженій камері. Повне А204 культуральне се 93855 30 редовище складалося з середовища McCoy 5A, 10% фетальної телячої сироватки, 2% L-глутаміну та 1% пеніциліну/стрептоміцину. При досягненні більше, ніж 80% злиття, клітини транзієнтно трансфікували за допомогою суміші плазміди рDРС4люцицераза та HCMV ΙΕ-lacZ при використанні пропису, рекомендованого виробником реагенту для трансфекції FUGENE (Roche Diagnostics Corporation, Indianapolis, IN) та інкубували протягом 16 годин при 37°С, 5% СО2, у зволоженій камері. Плазміда рDРС4-люцифераза містила чотири копії CAGA боксу, що має походження від інгібітору активатора плазміногену людини (РАl-1), що надає GDF8 чутливості до гетерологічної репортерної конструкції промотору. Плазміда HCMV ΙΕ-lacZ містила ген бетагалактозидази під контролем конститутивного найбільш раннього промотору людського цитомегаловірусу. Цей ген додається як контроль для нормалізації ефективності трансфекції. Клітини потім обробляли за допомогою GDF8 білка у концентрації 100 нг/комірка (R&D Systems Inc., Minneapolis, MN) та інкубували протягом додаткових 16 годин при 37°С, 5% СО2, у зволоженій камері. Люциферазу та бета-галактозидазу кількісно оцінювали в оброблених клітинах при використанні Dual-Light люциферазного аналізу (Tropix, Applied Biosystems, Foster City, CA). Кожний зразок розганяли у двох повторностях (2 комірки). Сигнал для кожної комірки підраховували як сигнал люциферази, розділений на сигнал бета-галактозидази, та помножений на 100. Сигнал зразка підраховували як середнє значення для двох комірок. Для аналізу біонейтралізаційної активності зразка антитіла різні концентрації очищених IgG фракцій інкубували з GDF8 білком (приблизно протягом 16 годин при 4°С) перед обробкою клітин. Процент інгібування підраховували як 100 (100 X сигнал зразка) / (сигнал тільки з GDF8 - сигнал без додання GDF8). Результати in vitro аналізу активації транскрипції підсумовані у Таблиці, що представлена нижче. Таблиця 1 Титри нейтралізації GDF8 для цапиної сироватки PGA Зразок (мкг IgG) % інгібування активності GDF8 Цапина - нормальна (250) 0 Цапина - PGA (250) 95 Цапина –PGA (125) 86 Цапина - PGA (63) 62 Цапина-PGA (31) 22 Цапина-PGA (16) 3 Аналіз нейтралізації підтвердив, що фракція IgG зібраної цапиної сироватки містить антитіла, здатні до нейтралізації, принаймні, 95% GDF8, що використовувався в аналізі. Приклад 4 Цапині поліклональні антитіла визначають специфічний епітоп нейтралізації білка GDF8 31 93855 Для того, щоб визначити специфічність нейтралізуючої імунної відповіді, PGA IgG фракцію аналізували на її реактивність з набором семи пептидів, що перекриваються (DJ 1-7 див. Таблицю 2 та Фігуру 1), які охоплюють повну кодуючу ділянку активного білка GDF8. Реактивність цапиного PGA IgG стосовно кожного індивідуального пептиду визначали за допомогою твердофазного імуно-ферментного аналізу (ELISA). ΕLISA для GDF8 пептиду здійснювали суттєво так, як описано в наборі Protein Detector™ ELISA Kit HRP, ABTS System (Kirkegaard and Perry Laboratories, Inc., Gaithersburg, MD). Використовували наступні модифікації в аналізі. Синтетичні пептиди DJ 1-7 (див. Таблицю 2, нижче) традиційно синтезували керуючись інструкціями ProSci, Inc. (Poway, CA). 32 Планшети покривали синтетичними пептидами при концентрації 500 нг на комірку та очищали GDF8 прогормон при концентрації 250 нг на комірку. Первинні антитіла представляли собою IgG фракції з різних зразків. Вторинні антитіла використовували при розведенні 1:2000. Для зразків цапиних первинних антитіл вторинне антитіло представляло собою мічене пероксидазою кроляче антитіло до цапиного IgG. Для зразків первинного антитіла пацюка вторинне антитіло представляло собою мічене пероксидазою цапине антитіло до IgG пацюків. OD при 405 нм зчитували протягом 15 хвилин при використанні пристрою для зчитування ELISA. ELISA реактивність підраховували як OD при 405 нм на хвилину, помножене на 1000. Таблиця 2 Активна щодо GDF8 ділянка пептидів Назва DJ1 DJ2 DJ3 DJ4 DJ5 DJ6 DJ7 Координати* 267-286 282-301 297-316 312-331 327-346 342-361 357-375 Амінокислотна послідовність DFGLDCDEHSTESRCCRYPL CRYPLTVDFEAFGWDWIIAP WIIAPKRYKANYCSGECEFV ECEFVFLQKYPHTHLVHQAN VHQANPRGSAGPCCTPTKMS PTKMSPINMLYFNGKEQIIY EQIIYGKIPAMWDRCGCS SEQ ID NO:4 SEQ ID NO:5 SEQ ID NO:6 SEQ ID NO: 7 SEQ ID NO: 8 SEQ ID NO:9 SEQ ID NO: 10 * по відношенню до людського GDF8 прогормону (депозитний номер Genebank NP_005250) Результати ELISA підсумовані у Таблиці 3, що представлена нижче. Таблиця 3 ELISA реактивність PGA IgG (10 мг/мл) до пептидів активної ділянки GDF8 Антиген DJ1* DJ2* DJ3* DJ4* DJ5* DJ6* DJ7* проGDF8** OD 405/хвилинах 1000 1:20 1:40 1:80 23 10 1 3 0 0 0 10 0 3 0 0 121 37 27 3 0 0 10 1 0 194 196 199 * пептид, ** прогормон PGA IgG фракція реагувала специфічно як з очищеним GDF8 прогормоном, так і з DJ5 пептидом. Серед активних ділянок пептидів GDF8 IgG фракція реагувала виключно з DJ5 пептидом. Це є чіткою вказівкою на те, що нейтралізуюча здатність цієї сироватки направлена на епітоп, який визначається пептидом DJ5. Для того, щоб підтвердити цю гіпотезу, очищали DJ5 специфічну фракцію PGA антитіла. Це здійснювали за допомогою афінної хроматографії так, як описано у розділі Матеріали та способи. PGA антитіла розділяли на зв'язані та незв'язані фракції DJ5 пептиду. Обидві фракції аналізували на нейтралізаційну активність проти білка GDF8. Результати, представлені у Таблиці 4, показують, що основна частина нейтралізаційної здатності GDF8 належить антитілу, яке специфічно зв'язується з DJ5 пептидом. Це ясно демонструє, що DJ5 пептид визначає нейтралізаційний епітоп білка GDF8. Таблиця 4 Нейтралізаційна активність стосовно GDF8 DJ5 специфічних антитіл Зразок (мкг lgG) Цапина - нормальна (250) ** DJ5 незв'язаний IgG (250) DJ5 зв'язаний IgG (25) % інгібування активності GDF8 7 26 90 ** Нормальний цапиний IgG представляв собою негативний контроль, очищений від сироватки неімунізованого цапа (комерційно закуповується) Цікаво, що у попередньому експерименті нейтралізуючі GDF8 антитіла не одержували, коли 33 93855 двом кролям вводили людський антиген DJ5, кон'югований з гемоціаніном фісурелли. Як можна побачити на Фігурі 2, амінокислотні послідовності, які відповідають DJ5 для кролячого та людського GDF8, є ідентичними, у той час, як амінокислотна послідовність цапиного DJ5 є відмінною. Таким чином, з огляду на дані, які забезпечуються вище для цапа, попередні дані для кроля дають змогу запропонувати, що може бути вигідно використовувати DJ5 антиген, що включає амінокислотну послідовність, яка є відмінною від такої для відповідної ділянки/частини GDF8 тварини-хазяїна. Таким чином, у цьому випадку здатність рекомбінантного прогормону GDF8 людини індукувати біонейтралізуючі антитіла у цапа може відбуватися завдяки, принаймні, частково, тому факту, що використовуваний антиген, включає амінокислотну послідовність, яка відрізняється від такої послідовності хазяїна однією заміною амінокислоти у DJ5 ділянці/частині GDF8 [див., амінокислотний залишок 333 на Фігурі 2]. Зокрема, як представлено на Фігурі 2, Аrg333 у послідовності людини замінюється залишком Lys333 у цапиній послідовності. Ця єдина консервативна амінокислотна заміна може представляти собою зміну, що є достатньо значущою для надання білку ознаки «чужерідності» для цапиної системи імунологічного контролю. Приклад 5 Нейтралізуюче GDF8 монАТ 788 пацюка визначає специфічний епітоп нейтралізації білка GDF8 Моноклональне антитіло пацюка 788 показане як таке, що нейтралізує GDF8 біоактивність миші (R&D Systems Inc., Кат. № МАВ788, Minneapolis, MN). Для того, щоб підтвердити цей результат ми аналізували моноклональне антитіло на нейтралізуючу активність проти білка GDF8. Антитіло було охарактеризоване так, як описано у Прикладі 2, що приведений вище. Результати цього аналізу підсумовані у Таблиці 5, як представлено нижче. Таблиця 5 Титри нейтралізації GDF8 для моноклонального антитіла 788 Зразок (мкг IgG) МонАТ-788 (12,5) МонАТ-788 (6,3) МонАТ-788(3,1) МонАТ-788(1,8) МонАТ-788 (0,1) % інгібування активності GDF8 47 17 7 0 0 34 Таблиця 5 підтверджує, що антитіло є здатним до нейтралізації активності GDF8 білка. Для того, щоб визначити цю нейтралізуючу імунну відповідь моноклональне антитіло пацюка аналізували на його реактивність з набором семи пептидів, що перекриваються (DJ 1-7 див. Таблицю 2 та Фігуру 1), які охоплюють повну кодуючу ділянку активного білка GDF8. Реактивність моноклонального антитіла стосовно кожного індивідуального пептиду визначали за допомогою аналізу ELISA (див. розділ матеріали та методи). Таблиця 6 ELISA реактивність МонАТ 788 пацюка (10 мг/мл) до пептидів активної ділянки GDF8 Антиген DJ1* DJ2* DJ3* DJ4* DJ5* DJ6* DJ7* проGDF8** OD 405/хвилинах 1000 1:20 1:40 1:80 4 0 0 0 0 0 0 0 0 0 0 0 133 118 102 0 0 0 0 0 0 132 127 132 * пептид, ** прогормон Моноклональне антитіло пацюка специфічно реагувало як з очищеним прогормоном GDF8, так і з DJ5 пептидом. Типово, моноклональне антитіло мало специфічність до одного епітопу. Серед пептидів активної ділянки GDF8 це моноклональне антитіло специфічно та виключно реагує з DJ5 пептидом. Цей результат забезпечує додаткове незалежне підтвердження того факту, що DJ5 пептид визначає нейтралізуючий епітоп білка GDF8. Багато модифікацій та варіацій винаходу можуть бути зроблені без відходу від духу та об'єму винаходу, як це буде зрозумілим для спеціаліста в даній галузі техніки. Специфічні втілення, описані в даній заявці, пропонуються тільки як приклади, а винахід обмежується тільки рамками прикладених пунктів формули разом з об'ємом еквівалентів, до яких ці пункти відносяться. В описі наведені багаточисленні посилання, включаючи депозитні номери Genebank опублікованих та/або викладених в Інтернеті послідовностей нуклеїнових кислот та послідовностей поліпептидів/білка, розкриття яких введено в дану заявку як посилання у їх цілісності. 35 93855 36 37 93855 38 39 93855 40 41 93855 42 43 93855 44 45 93855 46 47 93855 48 49 93855 50 51 93855 52 53 93855 54 55 93855 56 57 93855 58 59 93855 60
ДивитисяДодаткова інформація
Назва патенту англійськоюNeutralizing epitope-based growth enhancing vaccine
Автори англійськоюJunker David E., Cochran Mark D.
Назва патенту російськоюВакцина, способствует росту, базирующейся ha нейтрализующем эпитопе
Автори російськоюДжанкер Девид Э., Коучран Марк Д.
МПК / Мітки
МПК: C07K 14/495, A61K 38/18, C12N 15/18, G01N 33/53, C12N 15/62
Мітки: росту, нейтралізуючому, яка, вакцина, епітопи, базується, сприяє
Код посилання
<a href="https://ua.patents.su/31-93855-vakcina-shho-spriyaeh-rostu-yaka-bazuehtsya-na-nejjtralizuyuchomu-epitopi.html" target="_blank" rel="follow" title="База патентів України">Вакцина, що сприяє росту, яка базується на нейтралізуючому епітопі</a>
Антагоністичні аналоги рилізинг-гормону гормону росту (gh-rh), що інгібують інсуліноподібні фактори росту (igf-i і -ii)

Номер патенту: 68407
Опубліковано: 16.08.2004
Автори: Варга Джозеф, Заранді Марта, Шеллі Ендрю В.
МПК: A61P 5/06, C07K 14/60, A61K 38/25, A61P 35/00
Мітки: аналоги, gh-rh, гормону, фактори, росту, антагоністичні, інсуліноподібні, інгібують, igf-i, рилізинг-гормону
Формула / Реферат:
1. Пептид, що є антагоністичним аналогом рилізинг-гормону гормону росту (GH-RH), що інгібує інсуліноподібні фактори росту (IGF-I і -II), вибраний з групи, що має формулу:X-Rl-R2-Asp-Ala-R5-R6-Thr-R8-R9-R10-Arg-R12-R13-R14-R15-R16-Leu-R18-R19-Arg-R21-R22-Leu-GIn-Asp-Ile-R27-R28-R29-NH2,де Х означає PhAc, IndAc або Nac,R1 означає Туr або His,R2 означає D-Arg,R5 означає Ile або Val,R6 означає Phe або...
Атенуйований рекомбінантний вірус герпесу великої рогатої худоби типу 1 (bhv-1), вакцина, що його містить, та спосіб виявлення тварин, яким була введена рекомбінантна вірусна вакцина

Номер патенту: 75035
Опубліковано: 15.03.2006
Автор: Чоудхарі Шефікл І.
МПК: A61P 31/22, C12N 15/869, G01N 33/573, G01N 33/53, A61P 37/04, C12N 7/04, C12N 7/01, A61K 39/245, C12N 15/09, C12Q 1/34
Мітки: атенуйований, рогатої, герпесу, bhv-1, була, тварин, рекомбінантна, яким, худоби, вірусна, великої, введена, містить, рекомбінантний, вірус, вакцина, типу, спосіб, виявлення
Формула / Реферат:
1. Атенуйований рекомбінантний вірус герпесу великої рогатої худоби типу І, який являє собою вірус герпесу великої рогатої худоби типу І, в якому частина нативної кодуючої ділянки глікопротеїну Е видалена і замінена генетичною вставкою, яка включає кодуючу ділянку стороннього гена -галактозидази і негайний ранній промотор цитомегаловірусу людини, і при цьому даний...
Нейротрофічний фактор росту людини (еновін), молекула нуклеїнової кислоти, що його кодує, вектор, плазміда, фармацевтична композиція (варіанти), антитіло, спосіб виявлення фактору росту, набір, спосіб ідентифік

Номер патенту: 73922
Опубліковано: 17.10.2005
Автори: Геертс Х'юго Альфонсо, Меерт Тео Франс, Сік Мірослав, Масуре Стефан Лео Джозеф, Вер Донк Люк Аугуст Лаурентіус
МПК: A61P 1/08, A61K 48/00, C07K 16/22, A01K 67/027, A61P 25/14, A61P 29/00, A61P 25/00, A61P 25/28, A61P 1/10, C12N 1/19, A61P 43/00, C12Q 1/02, C12N 1/21, C12N 5/10, C12N 1/15, A61K 31/711, A61K 45/00, A61P 1/12, C07K 14/48, C12N 15/09, G01N 33/53, A61P 25/16, A61K 38/00, A61P 1/00, C07K 14/475, G01N 33/68, A61P 25/04, A61P 9/00
Мітки: росту, композиція, вектор, фармацевтична, ідентифік, спосіб, нуклеїнової, людини, фактор, плазміда, кодує, виявлення, антитіло, фактору, варіанти, еновін, молекула, нейротрофічний, кислоти, набір
Формула / Реферат:
1. Виділена молекула нуклеїнової кислоти, яка кодує нейротрофічний фактор росту людини (еновін) і має нуклеотидну послідовність, що відповідає амінокислотній послідовності, що показана на фіг. 1, або кодує послідовність, що має щонайменше 70%, а переважно 80%, 90% або 95% гомології з амінокислотною послідовністю, що показана на фіг. 1.2. Молекула нуклеїнової кислоти за п.1, яка є молекулою ДНК.3. Молекула нуклеїнової кислоти за...
Похідні пептиду (варіанти), що здатні вивільнювати гормон росту, фармацевтична композиція (варіанти), спосіб стимулювання вивільнення гормону росту з гіпофіза, способи підвищення швидкості і темпу росту та виро
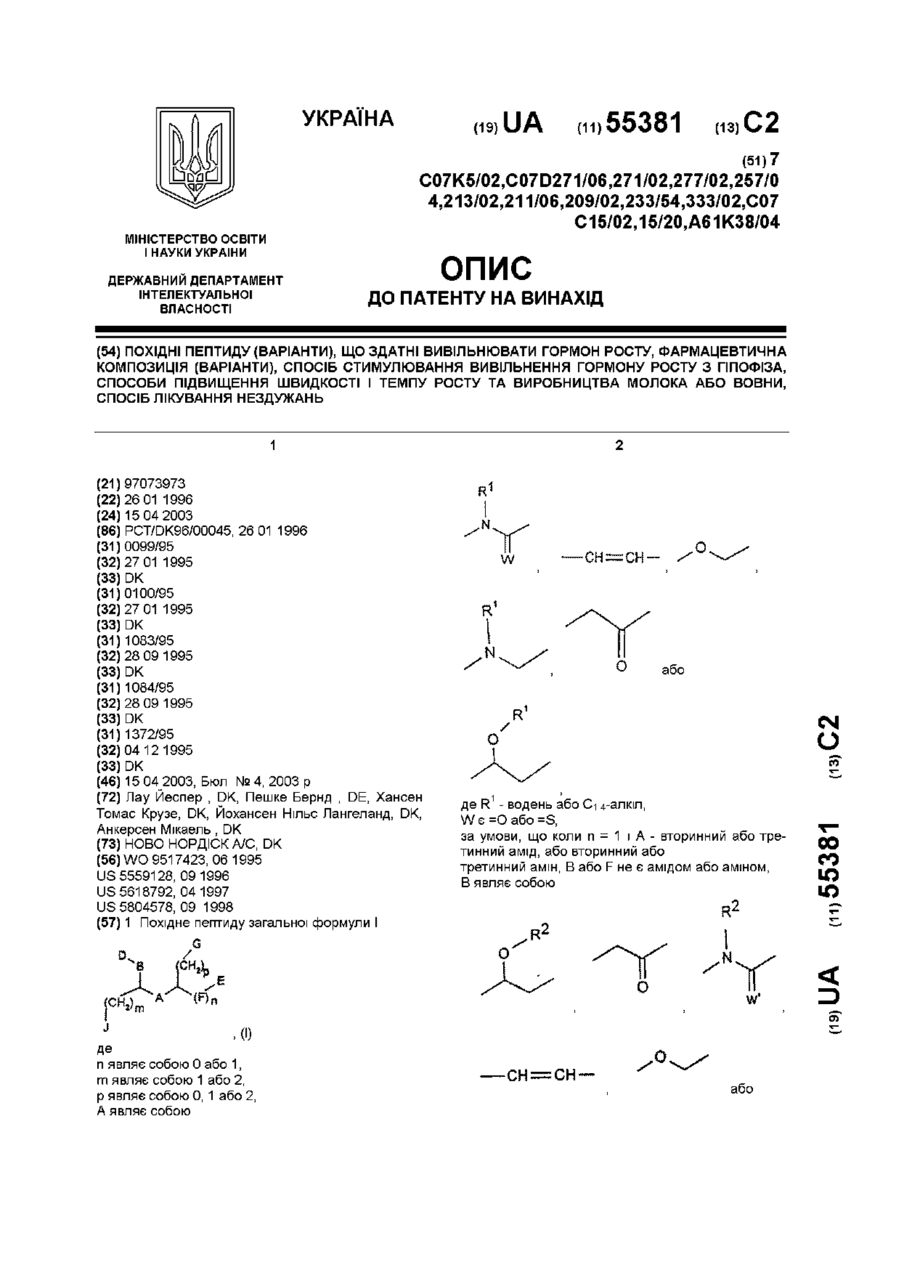
Номер патенту: 55381
Опубліковано: 15.04.2003
Автори: Анкерсен Мікаель, Пешке Бернд, ЙОХАНСЕН Нільс Лангеланд, ХАНСЕН Томас Крузе, Лау Йеспер
МПК: A61K 31/445, C07D 277/20, A61K 31/4427, A61K 31/41, C07D 211/60, A61K 31/165, C07D 257/00, C07D 417/12, A61K 31/4465, A61K 31/42, A61K 31/4245, C07D 211/62, C07D 277/56, C07K 5/065, C07D 271/10, C07C 237/22, A61K 31/425, A61K 31/27, A61K 38/00, C07D 413/12, A61K 31/415, C07C 233/38, C07D 271/06, C07K 14/60, C07D 401/12, C07C 237/32, A61P 43/00, A61K 31/426, C07C 237/36
Мітки: гормону, підвищення, здатні, стимулювання, фармацевтична, вивільнювати, композиція, гормон, виро, спосіб, способи, пептиду, темпу, гіпофіза, похідні, вивільнення, варіанти, росту, швидкості
Формула / Реферат:
1. Похідне пептиду загальної формули I, (I)де n являє собою 0 або 1,m являє собою 1 або 2,p являє собою 0, 1 або 2,А являє собою , , , , або , де R1 - водень або С1-4-алкіл,W є =O або =S,за умови, що коли n = 1 і А - вторинний або третинний амід, або вторинний...
Спосіб очищення забрудненої води, який базується на використанні цеолітів

Номер патенту: 83179
Опубліковано: 25.06.2008
Автори: Сісто Раффаелло, Бернарді Адріано, Віньола Родольфо, рілло Джованні
МПК: B01J 20/16, B01J 20/18, C02F 1/28
Мітки: спосіб, цеолітів, забрудненої, базується, використанні, води, очищення
Формула / Реферат:
1. Спосіб очищення води, забрудненої дихлоретиленом, вінілхлоридом, метил-третбутилетером або їх сумішами, який відрізняється тим, що очищенню піддають забруднену ґрунтову воду, пропускаючи її крізь проникний реактивний бар'єр, розміщений у відповідному місці перпендикулярно до потоку ґрунтової води, реактивний агент якого складається з цеоліту типу ZSM-5 або морденіту з відношенням діоксид силіцію/оксид алюмінію > 50 та структурними...
Попередній патент: Імуногенний кон’югат та спосіб його одержання
Наступний патент: Пристрій та спосіб для пакування виробів з мінеральної вати
Випадковий патент: Топка котла