Штам дріжджів candida famata imb y-5033 – стабільний продуцент рибофлавіну (вітаміну в2)
Номер патенту: 90741
Опубліковано: 25.05.2010
Автори: Дмитрук Костянтин Васильович, Яцишин Валентина Юріївна, Федорович Дарія Василівна, Сибірний Андрій Андрійович, Вороновський Андрій Ярославович



Формула / Реферат
Штам дріжджів Candida famata IMB Y-5033 - продуцент рибофлавіну.
Текст
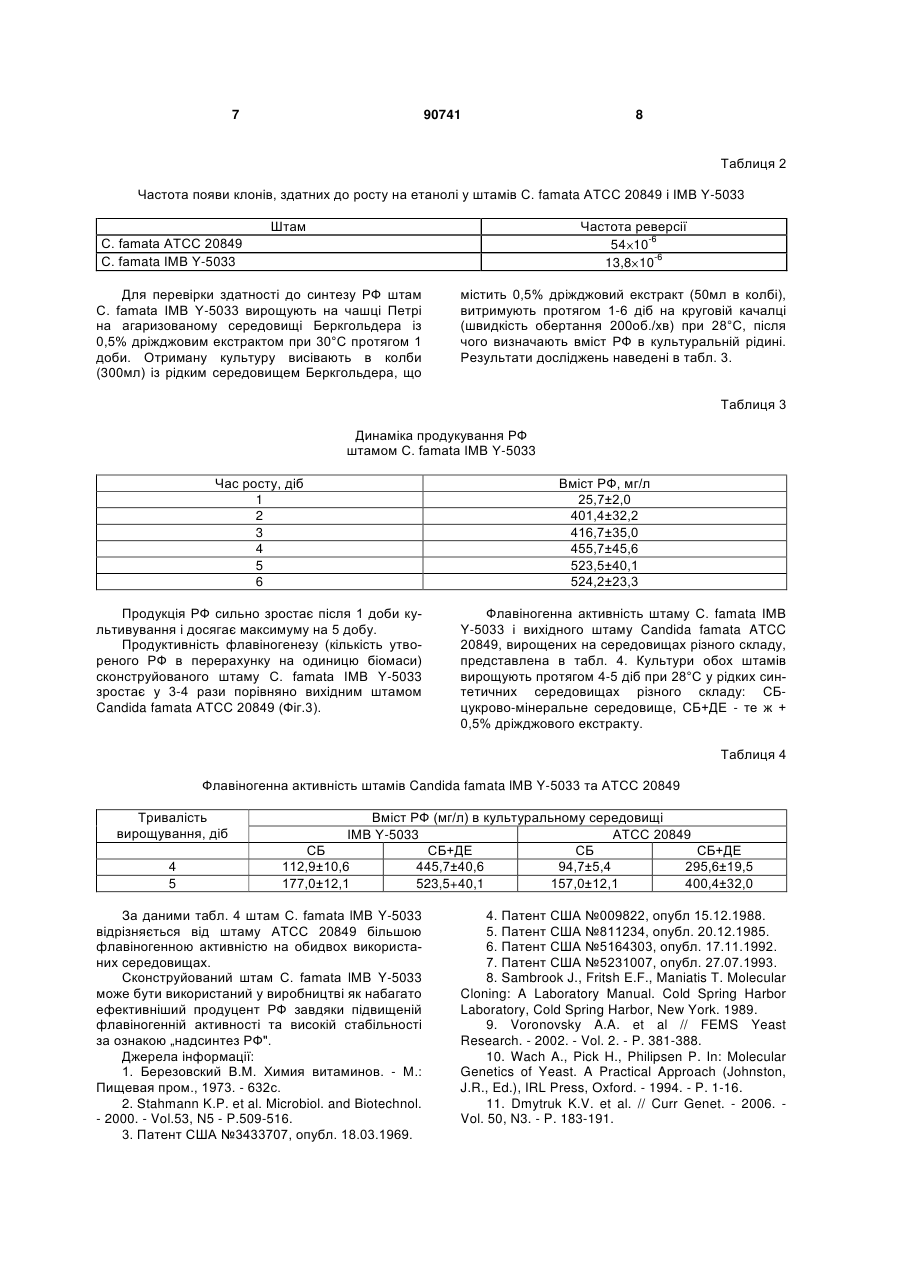
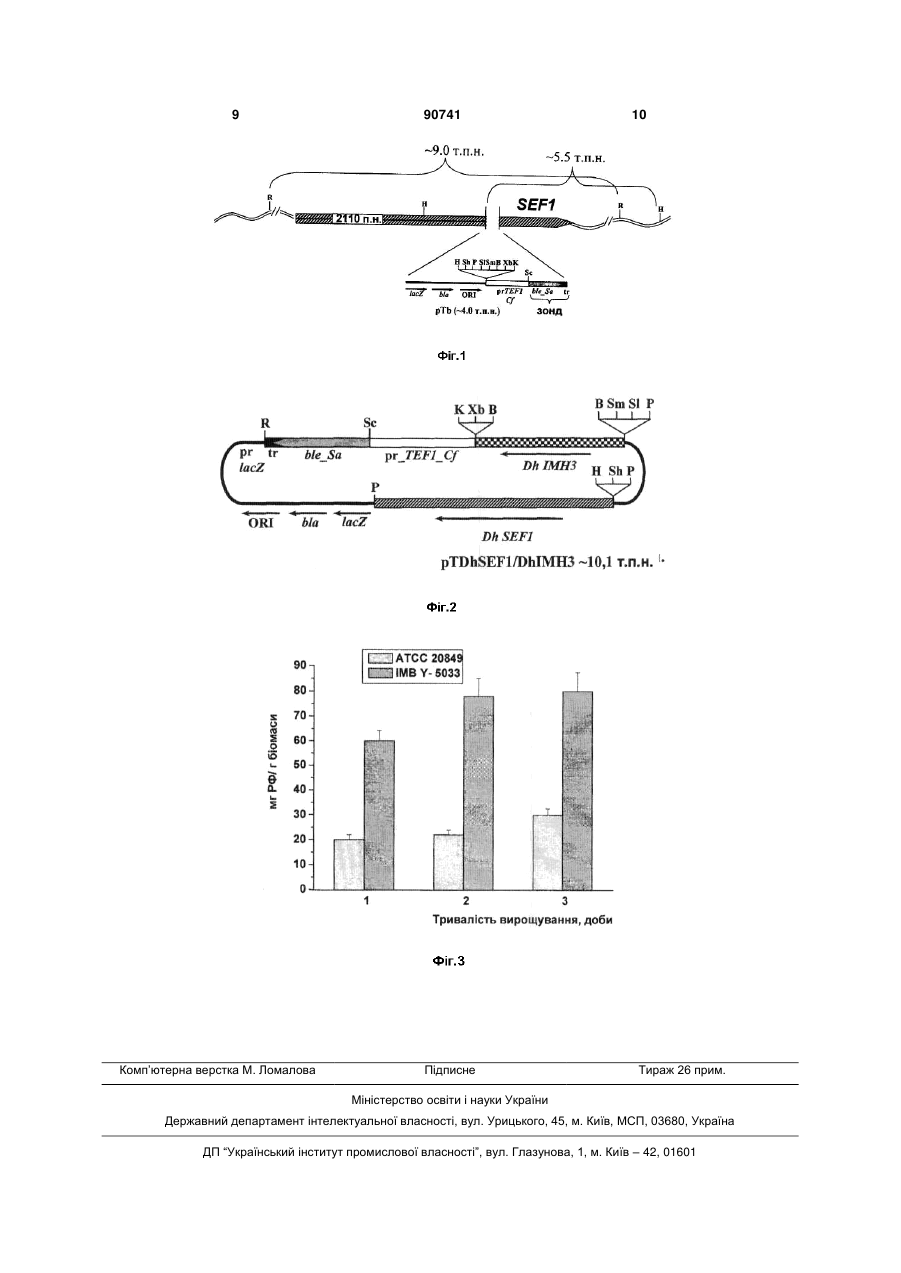
Штам дріжджів Candida famata IMB Y-5033 продуцент рибофлавіну. (19) (21) a200803778 (22) 25.03.2008 (24) 25.05.2010 (46) 25.05.2010, Бюл.№ 10, 2010 р. (72) ВОРОНОВСЬКИЙ АНДРІЙ ЯРОСЛАВОВИЧ, ДМИТРУК КОСТЯНТИН ВАСИЛЬОВИЧ, СИБІР 3 Штам дріжджів С. famata lMB Y-5033 зберігається в депозитарії Інституту мікробіології та вірусології НАН України. Культурально-морфологічні та фізіологобіохімічні особливості штаму ІМВ Y-5033. Вегетативні клітини штаму, вирощені в рідкому пивному суслі концентрацією 7°Б при 30°С, мають овальну форму, розміром 1,3-7,6 3,1-8,4мкм, містяться поодиноко, парами, іноді короткими ланцюжками чи невеликими гронами. Іноді утворюють псевдоміцелій. На агаризованому середовищі колонії віком 7 діб округлі, діаметром 2-4мм, профіль припіднятий, краї нерівні складчасті, поверхня гладка блискуча, жовтуватого кольору. На скошеному агарі 3-х добовий штрих гладкий однорідний блискучий, жовтуватого забарвлення. Клітини штаму С. famata ІMB Y-5033 асимілюють сахарозу, глюкозу, фруктозу, мальтозу, арабінозу, целобіозу, рафінозу, меліцитозу, трегалозу, сорбозу, крохмаль, глутамінову кислоту, сорбітол, маннозу, манніт, гліцерин, дульцит, інозит; слабо ростуть на ксилозі, рамнозі, ксиліті, лактозі, лактаті, галактозі, етанолі; проте не використовують яблучну і янтарну кислоту. Клітини запропонованого штаму не ферментують сахарозу, глюкозу, галактозу, ксилозу, лактозу, крохмаль, трегалозу, рафінозу, целобіозу та меліцитозу. Як джерело азоту клітини використовують сульфат амонію, диамоній фосфат, сечовину, але не нітрат натрію. Штам С. famata ІMB Y-5033 належить до облігатних аеробів. Штам росте при температурі 2532°С, оптимум температури становить 30°С; оптимум рН становить 6,0-9,0. За умов культивування у колбах на круговій качалці (200об/хв) у середовищі, що містить 2% глюкози, 1% пептону та 0,5% дріжджового екстракту протягом 4 діб запропонований штам нагромаджує близько 400мг РФ/л. Штам С.famata ІMB Y-5033 зберігається на агаризованому сусло-агарі в холодильнику. Клонується двічі на рік, відсівають 30-40 клонів. На 4-5 день вирощування відбирають клони за маркерною ознакою „здатність до надсинтезу рибофлавіну" Для конструювання штаму використовують такі методи: полімеразна ланцюгова реакція (ПЛР), конструювання рекомбінантних плазмід, виділення плазмід з Escherichia coli, рестрикційний аналіз, електрофорез в агарозному гелі, трансформація Е. coli методом електропорації, описані в [8]. Трансформацію С. famata проводять, як описано в [9]. Виділення сумарної ДНК з трансформантів С. famata проводять як для S. cerevisiae [10]. Вміст РФ визначають флуориметрично на приладі ЕФ3М. Ідентифікація гену, що забезпечує стабільність за ознакою „надсинтез рибофлавіну", та отримання стабільного надсинтетика цього вітаміну ілюструється графічними матеріалами. На Фіг.1 зображена лінійна схема плазміди рТb (4.0 т.п.н.), де відкриту рамку трансляції гену ble Staphylococcus aureus позначено товстою сі 90741 4 рою лінією; промотор власного гену TEF1 - товстим білим відрізком; бактерійна послідовність pUC57 - тонкою лінією; хвилястою лінією позначено хромосомну ДНК інсерційного штаму, товстою посмугованою лінією позначено ген SEF1; скорочення сайтів рестрикції: Н, Hindlll; Sp, Sphl; P, Pstl; SI, Sall; Xb, Xbal В, BamHl; Sm, Smal; K, Kpnl; Sc, Sacl; R, EcoRl. На Фіг.2 зображено лінійну схему плазміди pTDhSEFl/DhlМН3 (13.1т.п.н.), де відкриту рамку трансляції гену ble S. aureus позначено товстою сірою лінією; промотор гену TEFl - товстим білим відрізком; товстою шахованою лінією позначено послідовність гену ІМН3 D. hansenii; товстою посмугованою лінією - ген позитивної регуляції біосинтезу РФ SEFl D. hansenii; послідовність pUC57 тонкою лінією; скорочення сайтів рестрикції: Н, Hindlll; Sp, Sphl; P, Pstl; SI, Sall; Xb, Xbal; B, BamHl; Sm, Smal; K, Kpnl; Sc, Sacl; R, EcoRl. На Фіг.3 зображено зміни протягом 3 днів вирощування продуктивності флавіногенезу вихідного штаму С. famata ATCC 20849 та сконструйованого штаму lМВ Y-5033, де на осі абсцис відкладений час вирощування, а на осі ординат продуктивність флавіногенезу (кількість утвореного РФ в перерахунку на мг біомаси). Штам дріжджів С. famata lMB Y-5033 - стабільний мутант-надсинтетик РФ отримують у декілька етапів. Етап 1. Ідентифікація регуляторних генів, що спричинюють нестабільність штаму С. famata ATCC 20849. Як інсерційну касету використовують інтегративну плазміду з геном ble Staphylococcus aureus, що забезпечує резистентність до флеоміцину (Фіг.1) [11]. Перед трансформацією інсерційну касету лінеаризують рестриктазою ЕсоRl. Трансформацію проводять методом електропорації. Для позитивної селекції трансформанти штаму С. famata ATCC 20849 висівають на середовище з флеоміцином (5мг/л) та етанолом (1%) як джерелом вуглецю. Втрата здатності до надсинтезу РФ у ревертантів штаму АТСС 20849 завжди корелює з появою здатності рости в середовищі з етанолом, в той час як вихідний штам АТСС 20849 надсинтезує РФ та не росте на етанолі як єдиному джерелі вуглецю. Після серії трансформацій відбирають штам, що росте у середовищі з етанолом та виявляє резистентність до флеоміцину. Кількість копій касети в геномі відібраного штаму визначають за допомогою гібридизації за Саузерном. Для виділення касети з фланкуючими ділянками хромосомну ДНК інсерційного штаму обробляють рестриктазою Hindlll, здійснюють самолігування та трансформують в Е. соlі. З бактерійних трансформантів, що з'являються на середовищі з антибіотиком ампіциліном, виділяють плазмідну ДНК. Плазміда складається з вихідного вектора pUC57 та фрагменту геномної ДНК, що фланкує інсерційну касету. Послідовність нуклеотидів фланкуючої ділянки секвенують, використовуючи стандартні праймери (М13/pUC sequencing primer та M13/pUC reverse sequencing primer). Встановлено, що в ізольованого штаму касета розірвала ген SEFl, що кодує потенційний транскрипційний фак 5 тор (Фіг.1). Аналіз послідовності амінокислот виявив консервативний мотив, що відповідає за формування ДНК зв'язуючого домену Zn(2)-Cys(6), так званий цинковий палець. Він ідентифікований у багатьох дріжджових транскрипційних факторів. З метою отримання доказів інсерції касети саме в локус SEFl проводять комплементацію мутації відповідним геном дикого типу. Для цього на основі плазміди pTDhSEFl [11] сконструйовано плазміду pTDhSEFl/DhlMH3 розміром 10.1т.п.н., яка містить новий домінантний селекційний маркер для С. famata - ген ІМН3 Debaryomyces hansenii, 90741 6 що кодує інозинмонофосфат дегідрогеназу і визначає стійкість до мікофенольної кислоти (Фіг.2). Сконструйовану плазміду, що містить новий селекційний маркер (ген lМН3) та ген SEF1D. hansenii, використовують для комплементації мутації інсерційного штаму АТСС 20849. У трансформантів інсерційного штаму АТСС 20849 плазмідою pTDhSEFI/DhIMH3 втрачається здатність рости в середовищі з етанолом (Табл.1), натомість відновлюється здатність до надсинтезу РФ, що свідчить про те, що ген SEFl комплементує мутантний фенотип відібраного інсерційного штаму. Таблиця 1 Ріст на етанолі та здатність до утворення РФ різними штамами С. famata Штам АТСС 20849 sefl Ревертанти АТСС 20849 І II III IV Трансформанти sefl/SEFl Ревертанти АТСС 20849/ SEFl трансформанти І II III IV Додатково отримано трансформанти з геном SEF1 D. hansenii чотирьох незалежних спонтанних ревертантів АТСС 20849 (І, II, III, IV), які з'являються з високою частотою на середовищі з етанолом. У відповідних трансформантів спостерігається аналогічний фенотип: відновлення здатності до надсинтезу РФ, а також втрата здатності рости в середовищі з етанолом (Табл.1). Таким чином, можна стверджувати, що ген SEF1 залучений у регуляцію біосинтезу РФ. Пошкодження цього гену у штаму АТСС 20849 призводить до втрати здатності надсинтезувати вітамін В2. Етап 2. Одержання рекомбінантних штамів С. famata з додатковою копією гему SEF1. Стабілізацію штаму продуцента РФ С. famata ATCC 20849 здійснюють шляхом введення в його геном додаткової копії гену SEF1 D. hansenii. З цією метою його трансформують плазмідою pTDhSEF1 (Фіг.2). Стабільність штамів оцінюють за нездатністю до росту на середовищі з етанолом. Рекомбінантні штами, отримані шляхом трансформації спонтанних ревертантів (І, II, III, IV) С. famata ATCC 20849, та отриманого інсерційного штаму з розірваним геном SEF1 плазмідою pTDh SEF1/DhIMH3, не здатні до росту на середовищі, Ріст на етанолі ± + Утворення РФ + + + + + + + + + + що містить етанол як єдине джерело вуглецю (Табл.1). Штам дріжджів С. famata ІMB Y-5033 пройшов лабораторне випробування. Етап 3. Перевірка стабільності сконструйованого штаму та його здатності до надсинтезу РФ. Для перевірки стабільності штами висівають на чашки з середовищем YPD, через 1 добу засівають у 50мл цього ж середовища в колби об'ємом 100мл. Після вирощування протягом 1,5 доби клітини осаджують центрифугуванням, двічі промивають стерильною дистильованою водою і висівають на синтетичне середовище Беркгольдера, що містить 1% етанол як джерело вуглецю, в кількості 15 106 клітин на чашку. Через 10-14 днів підраховують кількість колоній на чашці і розраховують частоту реверсії. Відсутність росту на середовищі з етанолом чітко корелює зі здатністю до надсинтезу РФ. Як видно з таблиці 2, сконструйований штам С. famata ІMB Y-5033 з додатковою копією гену SEF1, характеризується підвищенням стабільності приблизно в 3-4 рази порівняно з штамом АТСС 20849. 7 90741 8 Таблиця 2 Частота появи клонів, здатних до росту на етанолі у штамів С. famata ATCC 20849 і ІMB Y-5033 Штам Частота реверсії 54 10-6 13,8 10-6 С. famata АТСС 20849 С. famata ІMB Y-5033 Для перевірки здатності до синтезу РФ штам С. famata ІMB Y-5033 вирощують на чашці Петрі на агаризованому середовищі Беркгольдера із 0,5% дріжджовим екстрактом при 30°С протягом 1 доби. Отриману культуру висівають в колби (300мл) із рідким середовищем Беркгольдера, що містить 0,5% дріжджовий екстракт (50мл в колбі), витримують протягом 1-6 діб на круговій качалці (швидкість обертання 200об./хв) при 28°С, після чого визначають вміст РФ в культуральній рідині. Результати досліджень наведені в табл. 3. Таблиця 3 Динаміка продукування РФ штамом С. famata ІMB Y-5033 Час росту, діб 1 2 3 4 5 6 Продукція РФ сильно зростає після 1 доби культивування і досягає максимуму на 5 добу. Продуктивність флавіногенезу (кількість утвореного РФ в перерахунку на одиницю біомаси) сконструйованого штаму С. famata ІMB Y-5033 зростає у 3-4 рази порівняно вихідним штамом Candida famata ATCC 20849 (Фіг.3). Вміст РФ, мг/л 25,7±2,0 401,4±32,2 416,7±35,0 455,7±45,6 523,5±40,1 524,2±23,3 Флавіногенна активність штаму С. famata ІMB Y-5033 і вихідного штаму Candida famata ATCC 20849, вирощених на середовищах різного складу, представлена в табл. 4. Культури обох штамів вирощують протягом 4-5 діб при 28°С у рідких синтетичних середовищах різного складу: СБцукрово-мінеральне середовище, СБ+ДЕ - те ж + 0,5% дріжджового екстракту. Таблиця 4 Флавіногенна активність штамів Candida famata lMB Y-5033 та ATCC 20849 Тривалість вирощування, діб 4 5 Вміст РФ (мг/л) в культуральному середовищі ІМВ Y-5033 АТСС 20849 СБ СБ+ДЕ СБ СБ+ДЕ 112,9±10,6 445,7±40,6 94,7±5,4 295,6±19,5 177,0±12,1 523,5+40,1 157,0±12,1 400,4±32,0 За даними табл. 4 штам С. famata lMB Y-5033 відрізняється від штаму АТСС 20849 більшою флавіногенною активністю на обидвох використаних середовищах. Сконструйований штам С. famata lMB Y-5033 може бути використаний у виробництві як набагато ефективніший продуцент РФ завдяки підвищеній флавіногенній активності та високій стабільності за ознакою „надсинтез РФ". Джерела інформації: 1. Березовский В.М. Химия витаминов. - М.: Пищевая пром., 1973. - 632с. 2. Stahmann K.P. et al. Microbiol. and Biotechnol. - 2000. - Vol.53, N5 - P.509-516. 3. Патент США №3433707, опубл. 18.03.1969. 4. Патент США №009822, опубл 15.12.1988. 5. Патент США №811234, опубл. 20.12.1985. 6. Патент США №5164303, опубл. 17.11.1992. 7. Патент США №5231007, опубл. 27.07.1993. 8. Sambrook J., Fritsh E.F., Maniatis T. Molecular Cloning: A Laboratory Manual. Cold Spring Harbor Laboratory, Cold Spring Harbor, New York. 1989. 9. Voronovsky A.A. et al // FEMS Yeast Research. - 2002. - Vol. 2. - P. 381-388. 10. Wach A., Pick H., Philipsen P. In: Molecular Genetics of Yeast. A Practical Approach (Johnston, J.R., Ed.), IRL Press, Oxford. - 1994. - P. 1-16. 11. Dmytruk K.V. et al. // Curr Genet. - 2006. Vol. 50, N3. - P. 183-191. 9 Комп’ютерна верстка М. Ломалова 90741 Підписне 10 Тираж 26 прим. Міністерство освіти і науки України Державний департамент інтелектуальної власності, вул. Урицького, 45, м. Київ, МСП, 03680, Україна ДП “Український інститут промислової власності”, вул. Глазунова, 1, м. Київ – 42, 01601
ДивитисяДодаткова інформація
Назва патенту англійськоюYeast strain candida famata imb y-5033 - stable producer of riboflavin (vitamin b2)
Автори англійськоюVoronovskyi Andrii Yaroslavovych, Dmytruk Kostiantyn Vasyliovych, Sybirnyi Andrii Andriiovych, Fedorovych Dariia Vasylivna, Yatsyshyn Valentyna Yuriivna
Назва патенту російськоюШтамм дрожжей candida famata имв y-5033 - стабильный продуцент рибофлавина (витамина в2)
Автори російськоюВороновский Андрей Ярославович, Дмитрук Константин Васильевич, Сибирный Андрей Андреевич, Федорович Дария Васильевна, Яцишин Валентина Юрьевна
МПК / Мітки
МПК: C12P 25/00, C12N 1/19
Мітки: candida, штам, рибофлавіну, продуцент, стабільний, вітаміну, y-5033, famata, дріжджів
Код посилання
<a href="https://ua.patents.su/5-90741-shtam-drizhdzhiv-candida-famata-imb-y-5033-stabilnijj-producent-riboflavinu-vitaminu-v2.html" target="_blank" rel="follow" title="База патентів України">Штам дріжджів candida famata imb y-5033 – стабільний продуцент рибофлавіну (вітаміну в2)</a>
Штам дріжджів candida famata imb y-5033 – стабільний продуцент рибофлавіну (вітаміну b2)

Номер патенту: 34554
Опубліковано: 11.08.2008
Автори: Вороновський Андрій Ярославович, Яцишин Валентина Юріївна, Дмитрук Костянтин Васильович, Сибірний Андрій Андрійович, Федорович Дарія Василівна
МПК: C12N 1/19
Мітки: штам, продуцент, рибофлавіну, стабільний, вітаміну, candida, famata, дріжджів, y-5033
Формула / Реферат:
Штам дріжджів Candida famata - стабільний синтетик рибофлавіну (вітаміну В2) з підвищеною продуктивністю синтезу, депонований в Депозитарії Інституту мікробіології та вірусології НАН України за № IMB Y-5033, який призначений для мікробіологічного отримання вітаміну В2.
Штам дріжджів candida famata imb y-5034 – продуцент рибофлавіну (вітаміну в2)

Номер патенту: 36164
Опубліковано: 10.10.2008
Автори: Федорович Дарія Василівна, Дмитрук Костянтин Васильович, Сибірний Андрій Андрійович
МПК: C12N 1/19
Мітки: продуцент, дріжджів, famata, штам, вітаміну, candida, рибофлавіну, y-5034
Формула / Реферат:
Штам дріжджів Candida famata - продуцент рибофлавіну (вітаміну В2), депонований в Депозитарії Інституту мікробіології та вірусології НАН України за № ІMB Y-5034, який має високу флавіногенну активність і призначений для мікробіологічного отримання вітаміну В2.
Штам дріжджів candida famata imb y-5028 – продуцент флавінмононуклеотиду (5′-фмн)

Номер патенту: 42499
Опубліковано: 10.07.2009
Автори: Федорович Дарія Василівна, Іщук Олена Петрівна, Яцишин Валентина Юріївна, Сибірний Андрій Андрійович, Вороновський Андрій Ярославович
МПК: C12N 1/19
Мітки: штам, 5'-фмн, famata, дріжджів, продуцент, флавінмононуклеотиду, candida, y-5028
Формула / Реферат:
Штам дріжджів Candida famata - продуцент флавінмононуклеотиду (5'-ФМН), з підвищеною здатністю до синтезу цього нуклеотиду без додавання АТФ або його попередників і рибофлавіну у поживне середовище, депонований в Депозитарії Інституту мікробіології та вірусології НАН України за № IMB Y-5028, який призначений для мікробіологічного отримання флавінмононуклеотиду.
Термотолерантний штам дріжджів candida tropicalis ст-96 – продуцент біомаси на відходах спиртового виробництва – бразі

Номер патенту: 30185
Опубліковано: 15.11.2000
Автори: Федорович Дарія Василівна, Андрейко Степан Ілліч, Когут Олександра Григорівна, Сосницький Віталій Володимирович
Мітки: ст-96, бразі, продуцент, термотолерантний, спиртового, біомаси, tropicalis, candida, дріжджів, відходах, штам, виробництва
Формула / Реферат:
Термотолерантний штам дріжджів Candida tro-picalis CT-96, здатний рости при температурі 30-42°С і ефективно нагромаджувати біомасу з вмістом білка 35-45%, штам Candida tropicalis CT-96 зберігається в депозитарії Інституту мікробіології та вірусології НАН України, реєстраційний номер: Candida tropicalis ІMB У-5005.
Штам дріжджів рісніа guіllіеrмоndіі sy 6/5, що нагромаджує в клітинах великі кількості рибофлавіну при культивуванні в середовищах з промисловими субстратами

Номер патенту: 24911
Опубліковано: 06.10.1998
Автори: Шавловський Георгій Михайлович, Струговщикова Людмила Петрівна, Бабяк Любов Ярославівна, Сенюта Елеонора Зиновіївна
МПК: C12N 1/16, C12R 1/645, C12P 25/00
Мітки: нагромаджує, великі, штам, промисловими, guіllіеrмоndіі, середовищах, культивуванні, дріжджів, рісніа, кількості, рибофлавіну, клітинах, субстратами
Формула / Реферат:
Штам дріжджів Pichia guilliermondii SY 6/5 здатний до надсинтезу рибофлавіну (95,5 - 324,3мг/л) і нагромадження великих кількостей вітаміну B2 в клітинах (24,3 - 36,3мг/г сухої ваги), штам дріжджів Pichia guilliermondii SY 6/5 зберігається в депозитарії Інституту мікробіології та вірусології НАН України, реєстраційний номер: Pichia guilliermondii IMB Y-5006.
Попередній патент: Комплексна домішка для бетонних сумішей та будівельних розчинів
Наступний патент: Похідні піразолону
Випадковий патент: Кіоск з термоізоляційною ємністю для тимчасового зберігання та продажу питної артезіанської води