Нові похідні стафілокінази, спосіб їх одержання та фармацевтична композиція








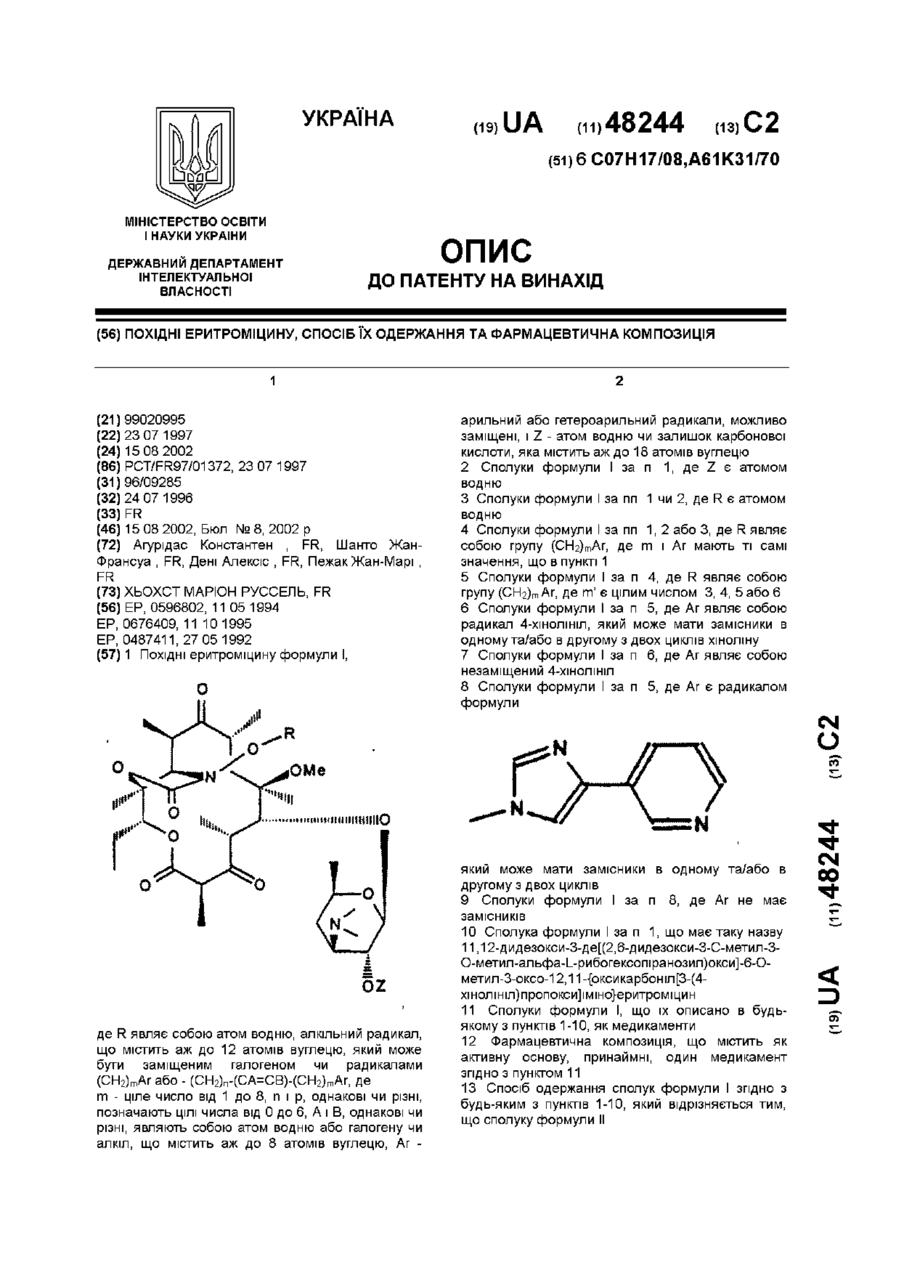
Формула / Реферат
1. Похідні стафілокінази, що проявляють знижену імуногенність порівняно зі стафілокіназою дикого типу, які мають по суті амінокислотну послідовність, зображену на фігурі 1:
в якій одна або більше амінокислот в одному чи більше з підкреслених кластерів були замінені іншою амінокислотою з руйнуванням, таким чином, відповідного(их) епітопу(ів).
2. Похідні стафілокінази за пунктом 1, що мають по суті амінокислотну послідовність, яка представлена на фігурі 1, в якій одна або більше амінокислот в одному чи більше з підкреслених кластерів замінені аланіном з руйнуванням, таким чином, відповідного(их) епітопу(ів).
3. Похідні стафілокінази за пунктом 1 або 2, що мають по суті амінокислотну послідовність, яка представлена на фігурі 1, в якій одна або більше амінокислот замінені аланіном зі зниженням, таким чином, реактивності цих похідних з панеллю моноклональних антитіл, що складається, зокрема, з антитіл 17G11, 26А2, 30А2, 2В12 і 3G10, спрямованих на епітопний кластер I.
4. Похідні стафілокінази за пунктом 1 або 2, що мають по суті амінокислотну послідовність, яка представлена на фігурі 1, в якій одна або більше амінокислот замінені аланіном зі зниженням, таким чином, реактивності цих похідних з панеллю моноклональних антитіл, що складається, зокрема, з антитіл 7Н11, 25Е1, 40С8, 24С4 і 1А10, спрямованих на епітопний кластер III.
5. Похідні стафілокінази за пунктом 1 або 2, що мають по суті амінокислотну послідовність, яка представлена на фігурі 1, в якій одна або більше амінокислот замінені аланіном зі зниженням, таким чином, реактивності цієї похідної з панелями моноклональних антитіл, спрямованих на епітопні кластери І та III.
6. Похідні стафілокінази за будь-яким одним з пунктів 1-5 для використання при лікуванні тромбозу артерій.
7. Похідна стафілокінази М8, що має амінокислотну послідовність, яка представлена на фігурі 1, в якій амінокислоти Lys у положенні 74, Glu у положенні 75 та Arq у положенні 77 у підкресленому кластері 8 замінені аланіном з руйнуванням, таким чином, відповідного епітопу.
8. Похідна стафілокінази за пунктом 7 для використання при лікуванні тромбозу артерій.
9. Похідна стафілокінази МЗ, що має амінокислотну послідовність, яка представлена на фігурі 1, в якій амінокислоти Lys у положенні 35 і Glu у положенні 38 у підкресленому кластері 3 замінені аланіном з руйнуванням, таким чином, відповідного епітопу.
10. Похідна стафілокінази за пунктом 9 для використання при лікуванні тромбозу артерій.
11. Похідна стафілокінази М9, що має амінокислотну послідовність, яка представлена на фігурі 1, в якій амінокислоти Glu у положенні 80 і Asp у положенні 82 у підкресленому кластері 9 замінені аланіном з руйнуванням, таким чином, відповідного епітопу.
12. Похідна стафілокінази за пунктом 11 для використання при лікуванні тромбозу артерій.
13. Похідна стафілокінази М3.8, що має амінокислотну послідовність, яка представлена на фігурі 1, в якій амінокислоти Lys у положенні 35 і Glu у положенні 38, Lys у положенні 74, Glu у положенні 75 і Arq у положенні 77 у підкреслених кластерах 3 і 8 замінені аланіном з руйнуванням, таким чином, відповідного епітопу.
14. Похідна стафілокінази за пунктом 13 для використання при лікуванні тромбозу артерій.
15. Похідна стафілокінази М8.9, що має амінокислотну послідовність, яка представлена на фігурі 1, в якій амінокислоти Lys у положенні 74, Glu у положенні 75, Arq у положенні 77, Glu у положенні 80 і Asp у положенні 82 у підкреслених кластерах 8 і 9 замінені аланіном з руйнуванням, таким чином, відповідного епітопу.
16. Похідна стафілокінази за пунктом 15 для використання при лікуванні тромбозу артерій.
17. Спосіб одержання похідних стафілокінази за будь-яким з пунктів 1-16, який включає стадії:
а) одержання фрагменту ДНК, що включає принаймні частину кодуючої послідовності стафілокінази, яка забезпечує її біологічну активність;
б) виконання in vitro сайт-спрямованого мутагенезу на фрагменті ДНК для заміни одного або більше кодонів амінокислот дикого типу кодоном іншої амінокислоти;
в) клонування мутантного фрагменту ДНК у придатний вектор;
г) трансформації або трансфекції придатної клітини-хазяїна цим вектором; і
д) культивування клітини-хазяїна в умовах, придатних для експресії фрагменту ДНК.
18. Спосіб за пунктом 17, в якому фрагмент ДНК являє собою EcoRI-HindIII фрагмент з 453 пар основ плазміди pMEX602SAK, сайт-спрямований мутагенез виконується in vitro за допомогою системи олігонуклеотид-спрямованого мутагенезу з використанням плазміди рМА/с і дефіцитного по репарації штаму E.coli WK6MutS, і мутантний фрагмент ДНК експресується в E.coli, штам WK6.
19. Фармацевтична композиція, що складається принаймні з похідних стафілокінази за будь-яким одним з пунктів 1-16 разом з придатним наповнювачем.
20. Фармацевтична композиція за пунктом 19 для лікування тромбозу артерій.
Текст
Цей винахід стосується нових похідних стафілокінази зі зниженою імуногенністю, їх виробництва та використання при лікуванні тромбозу артерій та для виготовлення фармацевтичної композиції для лікування тромбозу артерій. Більш конкретно, він стосується використання сконструйованих похідних стафілокінази для виготовлення фармацевтичної композиції для лікування інфаркту міокарду. Тромбозні ускладнення серцево-судинних захворювань є основною причиною смерті та непрацездатності, і отже, тромболіз (тобто розчинення за допомогою лікарських засобів згустку крові) могло б сприятливо впливати на кінець таких загрозливих життю захворювань, як інфаркт міокарду, тромбоз судин мозку та венозна тромбоемболія. Тромболітичні засоби є активаторами плазміногену, які перетворюють плазміноген, неактивний профермент фібринолітичної системи крові, в протеолітичний фермент плазмін. Плазмін розчиняє фібрин згустку крові, але може також руйнувати нормальні компоненти гемостатичної системи та викликати так званий "літичний стан". Фізіологічний фібриноліз, проте, є фібриноорієнтованим у результаті конкретних молекулярних взаємодій між тканинним активатором плазміногену, фібрином, плазміном (плазміногеном) та a2- антиплазміном (1,2). У даний час шість тромболітичних препаратів або утверджені для клінічного застосування або знаходяться на клінічних випробуваннях при лікуванні пацієнтів з гострим інфарктом міокарду. Ці препарати включають стрептокіназу, урокіназу, рекомбінантний активатор тканинного плазміногену (рт-АП) або його похідні, анізоільований плазміногено-стрептокіназний активаторний комплекс (АПСАК), рекомбінантний одноланцюговий урокіназний активатор плазміногену (роцу-АП, рекомбинантна проурокіназа) та рекомбінантна стафілокіназа (Sak) (2,3). Після лікування стрептокіназою, рт-АП або АПСАК у пацієнтів з гострим інфарктом міокарду спостерігалось зниження розміру області інфаркту, збереження функції шлуночків та зниження смертності (2). Одним з тромболітичних засобів, що використовуються у даний час для терапії, є стрептокіназа, білок з Мr 45000, секретуємий b-гемолітичними стрептококами. Її введення, однак, пов'язано з інтенсивним руйнуванням системного фібриногену, та її ефективність щодо коронарного тромболізу у хворих при гострому інфаркті міокарду обмежена, сягаючи приблизно 50-відсоткової реканалізації коронарних артерій протягом 90 хвилин (2). Крім того, введення стрептокінази викликає алергічні реакції у приблизно 5 відсотків пацієнтів, що лікувалися, і відповідно викликає утворення специфічних антитіл, що виключає її повторне застосування протягом місяців або років. Стафілокіназа - білок, що продукується деякими штамами Staphylococcus aureus, який має профібринолітичні властивості, що було показано більше 4 десятків років тому (5-7), також, видимо, представляє сильний тромболітичний засіб у хворих з гострим інфарктом міокарду (8). Ген стафілокінази був клонований з бактеріофагів sakÆC (9) та sak42D (10), а також з геномної ДНК (sakSTAR) лізогенного штаму Staphylococcus aureus (11). Він був експресований під контролем промотору lPR та його власних трансляційних сигналів в Escherichia coli, а також під контролем його природного промотору та трансляційних сигналів в Bacillus subtilis або Escherichia coli, що приводило до накопичення продукту гену в періплазматичному просторі або в культуральному середовищі, відповідно (10-13). Стафілокіназний ген кодує білок з 163 амінокислот, з амінокислотою 28, що відповідає NH2-кінцевому залишку зрілої стафілокінази повної довжини (10, 14, 15). Білкова послідовність варіанту дикого типу SakSTAR (15) представлена на фігурі 1. У кодуючих ділянках генів sakÆC, sak42D та SakSTAR було виявлено різницю лише чотирьох нуклеотидів, один з яких створював мовчащу мутацію (10, 14, 15). Було виділено та очищено декілька молекулярних різновидів стафілокінази з трохи різними Mr (1650018000 по електрофорезу в поліакриламідному гелі з додецилсульфатом натрію - ДСН-ЕПАГ) та ізоелектричними точками (11-13). Були одержані похідні зрілої стафілокінази з більш низьким Mr з відсутністю 6 (Sak-D6) або 10 (Sak-D10) NH2 - кінцевих амінокислот. При взаємодії з плазміном (плазміногеном) у буферному середовищі зріла стафілокіназа (з NH2 - кінцем Ser-Ser-Ser) швидко та кількісно перетворюється в Sak-D10 (NH2- кінець Lys-Gly-Asp-). Зріла стафілокіназа та Sak-D10, як було показано, мають однакову фібринолітичну активність (11, 12). Амінокислота в положенні 26 видимо має вирішальне значення для активації плазміногену стафілокіназою. Дійсно, заміна єдиного залишку Met в положенні 26 або Arg або Val приводить до втрати функціональної активності, тоді як заміна Leu або Cys має слабкий вплив на активність або зовсім не впливає на неї. Оскільки жодна з замін єдиної амінокислоти не викликає значних змін структури мутантних білків в розчині, механізм цієї диференцюючої поведінки залишається невідомим. В середовищі плазми стафілокіназа здатна розчиняти фібринові згустки без руйнування зв'язаного фібриногену (17-19). Ця фібриноспецифічність стафілокінази є результатом зниженого приглушення a2антиплазміном плазмін-стафілокіназного комплексу, зв'язаного з фібрином, рециркуляції стафілокінази від плазмін- стафілокіназного комплексу після приглушення a2-антиплазміном та запобігання перетворення циркулюючої плазміноген-стафілокінази в плазмін-стафілокіназу a2-антиплазміном (20-22). На декількох експериментальних моделях тварин стафілокіназа, видимо, проявляє однакову активність зі стрептокіназою щодо розчинення згустків цільної крові або плазми, але значно більш активна щодо розчинення багатих на тромбоцити або стиснених тромбів (23, 24). Результати, що подають надію, одержані зі стафілокіназою на моделях тромбозу на тваринах сформували основу для її оцінки в масштабі попереднього дослідження у хворих з гострим інфарктом міокарду (3, 25). У 4 з 5 хворих з гострим інфарктом міокарду 10мг рекомбінантної стафілокінази (SakSTAR), введені внутрівенно протягом 30хв., як було виявлено, викликають ангіографічно підтверджену реканалізацію коронарних артерій протягом 40хв. На рівні фібриногену плазми та a2-антиплазміну впливу не здійснювалося (залишкові рівні через 40хв. становили 90-95% від початкового), та алергічні реакції не спостерігались (3). У другій групі з 5 хворих з гострою оклюзією (закупорюванням) коронарних артерій внутрівенне введення 10мг стафілокінази (SakSTAR) протягом 30 хвилин приводило до реканалізації у всіх хворих протягом 20 хвилин без деградації зв'язаного фібриногену (25). Контрольна ангіографія через 24 години показала, що реканалізація залишалась. Імуногенність стафілокінази (SakSTAR) порівняно з стрептокіназою вивчали на собаках (23) та бабуїнах (24). В цілому ці дані, одержані на експериментальних тваринах, свідчать про більш низьку імуногенність стафілокінази порівняно зі стрептокіназою. Однак, у перших 5 пацієнтів з гострим інфарктом міокарду, що отримали внутрівенне вливання 10мг стафілокінази протягом 30 хвилин, титри нейтралізуючих антитіл до стафілокінази (SakSTAR) були низькими на початковий момент та до 6 днів після вливання, але високі титри (титри нейтралізуючих стафілокіназу антитіл, рівні 12-42мкг/мл плазми) постійно виявлялись у плазмі через 14-35 днів (3). Ці дослідження були повністю підтверджені у другому попередньому випробуванні у 5 пацієнтів (25). Так, що стосується імуногенності, первісні спостереження на людях не були такими обнадійливими, як дані, отримані на експериментальних тваринах. Таким чином, подібно стрептокіназі, застосування стафілокінази повинно бути обмежено єдиними введенням. Але, відсутність перехресної реактивності індуцьованих антитіл до стафілокінази та стрептокінази (26, 27) приводить до думки, що введення обох речовин не буде взаємовиключним. Імуногенність, властива стрептокіназі та стафілокіназі, явно перешкоджає їх необмеженому використанню. Не тільки хворі з попередніми високими титрами антитіл будуть несприйнятливі до тромболітичної дії цих речовин, але можуть з'являтися побічні алергічні ефекти та у рідких випадках загрозлива для життя анафілаксія (28). Оскільки як стрептокіназа, так і стафілокіназа є гетерологічними білками, не очевидно, що їх імуногенність могла б бути знижена інженерією білків. Дійсно, не повідомлялось про успішні спроби створення активних фрагментів з низькою молекулярною вагою з стрептокінази. У стафілокінази делеція NH2-кінцевих 17 амінокислот або СООН-кінцевих 2 амінокислот інактивує молекулу, яка, крім того, дуже чутлива до інактивації за допомогою сайт-специфічного мутагенезу (25, 29). Але, ми несподівано виявили, що варіант стафілокінази дикого типу (SakSTAR) (8, 15) містить три імунодомінантних епітопи, що неперекриваються, принаймні, два з яких можуть бути еліміновані за допомогою специфічного сайт-направленого мутагенезу без інактивації молекули. Ці сконструйовані варіанти стафілокінази є менш реактивними з антитілами, що виявляються у хворих, які лікувалися стафілокіназою дикого типу, та значно менш імуногенні, ніж стафілокіназа дикого типу, що показано на моделях на кроликах та бабуїнах та у хворих з закупоркою периферичних артерій. Даний винахід, таким чином, стосується похідних стафілокінази, що проявляють знижену імуногенність порівняно зі стафілокіназою дикого типу. Ці похідні мають по суті амінокислотну послідовність стафілокінази дикого типу або її модифіковані версії, але, принаймні, один імунодомінантний епітоп елімінований без порушення біологічної активності похідних. В одному здійсненні цього винаходу похідні мають по суті амінокислотну послідовність, яка зображена на фігурі 1, на якій одна або більше амінокислот в одному або більше підкреслених кластерів мають заміну іншою амінокислотою, таким чином, змінюючи відповідний(і) ізотоп(и). Переважно, амінокислоти заміняються аланіном. За допомогою зміни епітопу(ів) реактивність похідних з панеллю моноклональних антитіл до одного або більше з трьох епітопних кластерів І, II та III знижена. Це показує, що за допомогою заміни амінокислот дикого типу аланіном антигенність стафілокінази знижується. Цей винахід, зокрема, стосується похідного стафілокінази М8, що має амінокислотну послідовність, яка представлена на фігурі 1, в якій амінокислоти Lys в положенні 74, Glu в положенні 75 та Arg в положенні 77 у підкресленому кластері 8 були замінені аланіном, змінюючи таким чином відповідний еіпітоп, похідного стафілокінази М3, що має амінокислотну послідовність, яка зображена на фігурі 1, в якій амінокислоти Lys в положенні 35 та Glu в положенні 38 в підкресленому кластері 3 були замінені аланіном, змінюючи таким чином відповідний епітоп, похідного стафілокінази М9, що має амінокислотну послідовність, яка представлена на фігурі 1, в якій амінокислоти Glu в положенні 80 та Asp в положенні 82 в підкресленому кластері 9 були замінені аланіном, змінюючи таким чином відповідний епітоп, похідного стафілокінази М3.8, що має амінокислотну послідовність, яка представлена на фігурі 1, в якій амінокислоти Lys в положенні 35, Glu в положенні 38, Lys в положенні 74, Glu в положенні 75 та Arg в положенні 77 в підкреслених кластерах 3 або 8 були замінені аланіном, змінюючи таким чином відповідні епітопи, та похідного стафілокінази М8.9, що має амінокислотну послідовність, яка представлена на фігурі 1, в якій амінокислоти Lys в положенні 74, Glu в положенні 75, та А в положенні 77, Glu в положенні 80 та А в положенні 82 в підкреслених кластерах 8 і 9 були замінені аланіном, змінюючи таким чином відповідні епітопи. Таким чином М3.8 та М8.9 є подвійними мутантами, що мають два зруйнованих епітопи. Цей винахід демонструє, що сконструйовані варіанти стафілокінази зі зниженою імуногенністю можуть бути практичними альтернативними стрептокіназі або стафілокіназі дикого типу тромболітичними препаратами. Цей винахід також стосується способу продукції похідних цього винаходу шляхом одержання фрагменту ДНК, що містить, принаймні, частину кодуючої послідовності стафілокінази, яка забезпечує її біологічну активність; здійснення in vitro сайт-направленого мутагенезу у фрагменті ДНК для заміни одного або більше кодонів для амінокислот дикого типу кодоном для іншої амінокислоти; клонування фрагменту мутантної ДНК в підхожий вектор; трансформації або трансфекції придатної клітини-хазяїна цим вектором; та культивування клітин-хазяїнів в умовах, придатних для експресії фрагменту ДНК. Переважно, фрагмент ДНК є фрагментом Ecori-HindlD з 453 п.о. з плазміди pMEX602SAK, сайт-направлений мутагенез виконується за допомогою системи олігонуклеотиднаправленого мутагенезу з використанням плазміди рМа/с і дефіцитного по репарації штаму E:/COLi WK6Muts, та фрагмент мутантної ДНК клонується в Е.Соlі, штам WK6. Цей винахід також стосується фармацевтичних композицій, що містять, принаймні, одне з похідних стафілокінази по цьому винаходу разом з придатним носієм, для лікування артеріального тромбозу. Фармацевтичні композиції, що мають менш імуногенні варіанти стафілокінази в якості активного інгредієнту, для лікування тромбозу артерій у людей або у ветеринарній практиці можуть мати форму порошків або розчинів і можуть застосовуватись для внутрівенного чи внутріартеріального введення. Такі композиції можуть бути виготовлені шляхом об'єднання (наприклад, змішування, розчинення тощо) активної речовини з фармацевтично прийнятними наповнювачами нейтрального характеру (такими як водні або неводні розчинники, стабілізатори, емульгатори, детергенти та добавки), і крім того, якщо необхідно, барвниками. Концентрація активного інгредієнту у терапевтичній композиції може широко мінятися в інтервалі від 0,1% до 100%, залежно від характеру захворювання та способу введення. До того ж доза активного інгредієнту, яку потрібно застосувати, може змінюватися у межах від 0,05мг до 1,0мг на кг ваги тіла. Крім того, цей винахід стосується способу застосування похідних стафілокінази для лікування тромбозу артерій, зокрема, інфаркту міокарду, та використання похідних стафілокінази для виготовлення фармацевтичної композиції для лікування тромбозу артерій, зокрема, інфаркту міокарду. Вище і далі терміни "похідні", "мутанти", та "варіанти" використовуються, замінюючи один одного. Даний винахід буде показано більш детально в наступних прикладах, які, однак, не призначені для обмеження обсягу винаходу. Для досвідченого спеціаліста будуть очевидні декілька варіантів та удосконалень на основі даного винаходу. Таким чином, випадковий мутагенез, починаючи з комбінаційного мутанту 3.8 та з комбінаційного мутанту 8.9 повинен дати альтернативні мутанти зі зниженою імуногенністю та, можливо, підвищеною функціональною активністю, тоді як альтернативний мутагенез в епітоп-нейтралізуючих кластерах буде давати інші варіанти зі зниженою імуногенністю. Приклад 1 Картирування епітопу стафілокінази дикого типу Епітопна специфічність панелі з 17 мишиних моноклональних антитіл, індукованих стафілокіназою дикого типу (варіант SakSTAR), визначали шляхом аналізу біоспецифічної взаємодії у реальному часі (ВІА) з використанням прибору ВІАсоrеä (Pharmacia, Biosensor AB, Uppsala, Sweden). Моноклональні антитіла до SakSTAR продукувались за допомогою методу Galfre та Milstein (30). Мишей BALB/c Імунізували шляхом підшкірної ін'єкції 10мкг SakSTAR у повному ад'юванті Фрейнда, за якою через два тижні йшло внутрішньочеревинне введення 10мкг SakSTAR у повному ад'юванті Фрейнда. Після проміжку, рівного принаймні 6 тижням, миші одержували внутрішньочеревинно повторну ін'єкцію 10мкг SakSTAR у фізіологічному розчині на 4 день та за два дні до злиття клітин. Клітини селезінки виділяли та виробляли злиття з клітинами мієломи P3X63-Ag.8-6.5.3 (отриманими від д-ра О. Schönherr, Organon, Oss, Netherlands) пo Fazekas de St. Groth and Scheidegger (31). Після селекції у гіпоксантин-аміноптеринтімідиновому середовищі супернатанти відбирали по продукції специфічних антитіл за допомогою односайтового неконкурентного мікро-ELISA з використанням мікротитровальних плат, покритих стафілокіназою. Зв'язані імуноглобуліни визначали за допомогою кролячого протимишиного IgG, кон'югованого з пероксидазою хрону (32). Позитивні клони використовували для продукції асцитної рідини у приміруваних пристаном мишей BALB/c (33). Фракцію IgG моноклональних антитіл вичищували від асцитної рідини за допомогою аффінної хроматографії на протеїн А-сефарозі (34). Ця техніка аналізу біоспецифічної взаємодії, основана на поверхневому плазмовому резонансі (ППР) дає можливість прямого вимірювання взаємодії у реальний строк без використання міток (35). Стафілокіназа була імобілізована на поверхні сенсорного чипу -Sensor Chip CM5 з використанням набору для зв'язування амінів - Amine Coupling kit (Pharmacia Biosensor AB), як рекомендовано виробником. Ця процедура зв'язує первинні аміногрупи у ліганд до карбоксиметилованої декстранової поверхні сенсорного чипу (36). Імобілізацію виконували з розчинів білка з концентрацією 10мкг/мл в 10мМ ацетаті натрію з pH 5,0 при потоці зі швидкістю 5мкл/хв протягом 6хв. Це приводить до ковалентного зв'язування 1000-1500 РО (резонансних одиниць) стафілокіназних частин (що відповідають приблизно 0,07пмоль/мм2 (37). Другий взаємодіючий компонент (аналізуєма речовина: тобто моноклональні антитіла) вводили в розчин над сенсором. Концентрація вільної аналізуємої речовини підтримувалась постійною по безперервному потоку розчину при 20°С по поверхні сенсора. Вводили, принаймні, чотири концентрації кожної аналізуємої речовини (в інтервалі 0-400нм або 0-50мкМ) в 10мМ HEPES, 3,4мМ ЕДТК, 0,15 М NaCl та 0,005% сурфактанті Р20, pH 7,2 при швидкості потоку, що дорівнює 5мкл/хв у фазі зв'язування. Потім зразок заміняли буфером також при швидкості потоку, рівною 5мкл/хв протягом від 6 до 30 хвилин. Після кожного циклу поверхню сенсорного чипу регенирували за допомогою введення 5мкл 15мМ НС1. Константи швидкості зв'язування (kасс ) та дислокації (kдисс) виводили з сенсограм, як докладно описано в іншому місці (38). Константи рівноважного зв'язування (Кс), розраховані як відношення kасс та kдисс, для зв'язування стафілокінази дикого типу на панелі з 17 вивчаємих моноклональних антитіл змінювались в інтервалі між 0,6 та > 25х109 М-1 (середнє значення 1010 М-1) (таблиця 1). В таблиці 1 колонка, позначена "ID" вміщує різні стафілокіназні похідні. Позначення "17G11", "26А2" тощо стосуються моноклональних антитіл, що зв'язуються з вказаними епітопними кластерами І, II і III. В колонці "варіант" показані мутантні амінокислоти та їх положення в однолітерному коді для амінокислот. Епітопний кластер І розпинається антитілами 17П11, 26А2, 30А2, 2В12 і 3G10, тоді як епітопний кластер II розпізнається антитілами 29С1, 18F12, 14Н5, 28Н4, 20D6, 32В2 і 7F10 і епітопний кластер III - антитілами 7Н11, 25Е1, 40С8, 24С4 та 1А10. Моноклональні антитіла, направлені проти окремих епітопів, будуть зв'язуватися незалежно один від одного, тоді як моноклональні антитіла, направлені проти близькородинних епітопів будуть взаємодіяти кожний з місцем зв'язування іншого. Тому епітопна специфічність панелі моноклональних антитіл найбільш легко визначається шляхом визначення здатності пари моноклональних антитіл одночасно зв'язуватися з антигеном. Аналіз біоспецифічної взаємодії у реальний час (АБВ=ВІА ) може використовуватися для вимірювання конкурентного зв'язування пар моноклональних антитіл зі стафілокіназою, зв'язаною з поверхнею сенсорного чипу. Аналіз проводили, як описано в Application Note 101 (Pharmacia Biosensor AB). Випробування парного зв'язування розділили 17 моноклональних антитіл на 3 групи, що представляють З епітопи на антигені, що неперекриваються, як показано на фігурі 2. Незалежність цих епітопів була підтверджена прямою демонстрацією додаткового зв'язування моноклональних антитіл 26А2, 28Н4 і 24С4. Антитіла об'єднували за їх епітопною специфічністю, що показано в таблиці 1. Приклад 2 Конструювання та епітопне картирування варіантів стафілокінази "зapяджeний-клacтep-для-aлaнiнy"(chaгged-cluster-to-alanцle") При ретельному вивченні "заряджений-кластер-для-аланіну" намічали кластери гідрофільних заряджених амінокислот. Стафілокіназа (SakSTAR) містить 45 заряджених амінокислот (2 His, 14 Glu, 8 Asp, 1 Arg i Lys). Ці заряджені залишки піддавали мутагенезу на Ala в кластерах з двох або трьох амінокислот, що підсумовано на фігурі 1. Всього був створений 21 мутант, в яких підкреслені заряджені амінокислоти були замінені аланіном. Амінокислоти, які повинні бути замінені аланіном, показані за допомогою невеликої вертикальної лінії у кластері. Мутанти одержували шляхом сайт-направленого мутагенезу та експресували в Е.соlі, що детально показано нижче. Ферменти рестрикції були закуплені у Pharmacia Uppsala, Sweden or Boehringer Mannheim (Mannheim, Germany). ДНК-лігаза Т4, фрагмент Кленова ДНК-полімерази 1 Е.соlі та лужна фосфатаза були одержані від Boehringer Mannheim. Система олігонуклеотид-направленого мутагенезу та плазміди рМА/с були люб'язно надані Corvas (Ghent, Belgium) (39). Вектор експресії pMEX602SakB був люб'язно наданий Інститутом молекулярної біотехнології, Йена, Німеччина (25), Хелперний фаг М123КО7 був закуплений у Promega (Лейден, Нідерланди). Бульйонне ростове середовище Луріа було закуплено у Life Technologies (Merelbeke, Belgium). Плазміноген виділяли та очищували з людської плазми, як описано раніше (40). Ферментативні реакції виконували, використовуючи умови, які пропонувались постачальниками. Плазмідну ДНК виділяли, використовуючи методику очищення QIAGEN (надану Westburg, Leusden, Нідерланди). Трансформацію Е.соlі виконували з застосуванням кальцій-фосфатної методики. Секвенування ДНК виконували, використовуючи метод реакції термінації дідезокси-ланцюга та автоматичного лазерного флюоресцентного A.L.F.ä (Pharmacia). Сайт-направлений мутагенез для мутантів D5, К6 (М20) до К86, Е88, (М10) проводився з використанням рМа/с, з застосуванням дефіцитного по репарації штаму Е.соlі WK6Muts. Розмноження плазмід рМа/с або похідних, одержання однониткової ДНК і експресію проводили в Е/соlі WK6 (39). Мутанти D93, К94 (МИ) по К134, К135, К136 (М19) були сконструйовані в Інституті молекулярної біотехнології, Йєна, Німеччина, як описано раніше (16). Хромогенний субстрат (S2403) L-піроглютаміл-L-фенілаланіл-L-лізин-п-нітроаналін гідрохлорид був закуплений у Chromogenix. Фібриноген, мічений 125І, був закуплений у Amersham. Фрагмент EcoRI-HindІІІ з 453 пар основ, що містить всю кодуючу область для SakSTAR, вирізали з плазміди pMEX602SakB (стійкість до ампіциліну) і клонували в сайти EcoRI-HindІІІ плазміди рМс5-8 (стійкість до хлорамфеніколу), одержуючи рМс-STAR. Для сайт-направленого мутагенезу in vitro однониткову ДНК цієї конструкції одержували шляхом трансформації конструкції pMc-STAR в Е.соlі та введення в нічну культуру хелперного фагу М13КО7. Через чотири години після ін'єкції клітини відокремлювали від середовища за допомогою преципітації ПЕГ та екстракцією фенолом-хлороформом. Потім однониткову pMc-STAR гібридизували з однонитковою рМа (EcoRI-HindIII) векторної ДНК та відповідним синтетичним олігонуклеотидом з 28-44 основ з мовчащою мутацією, що створює або делегує рестрикційний сайт. Реакції подовження проводили з фрагментом Кленова ДНК-полімерази, якописано. Після трансформації Е.соlі WK6Muts та селекції на ампіцилінову рецистентність колонії підрощували на нітро-целюлозних мембранах, денатурували in situ, і ДНК гібридизували протягом ночі при кімнатній температурі з використанням відповідних радіоактивно мічених олігонуклеотидів (1,5x108 імп/хв [g32Р]-АТФ, використаної для мічення полінуклеотидкіназою Т4 20-30нг олігонуклеотиду). Фільтри промивали при 42°С, використовуючи розчини, що містять 0,1% ДСН та 2xSSC, 1xSSC, 0,2xSSC, 0,1xSSC. Плаз мідну ДНК екстрагували з 10мл бактеріальних культур, одержаних з кожного позитивного клону та аналізували шляхом розщеплення ферментом рестрикції. Бажані мутації підтверджували шляхом секвенування повної кодуючої послідовності з використанням A.L.F.ä. Мутований фрагмент Hindlll-EcoRI потім лігірували зворотно в вектор експресії pMEX602SakB, що містить промотор pTag (39). Мутантні білки продукувались внутріклітинно і у розчинній формі у клітинах Е.соlі WK6, трансформованих цим вектором. Мутанти виділяли та очищали зі зруйнованих ультразвуком бактеріальних екстрактів з використанням катіонообмінної та з гіброфобною взаємодією хроматографії (25). Мутанти SakSTAR були одержані з виходом у інтервалі від 10 до 80мг/л, що представляє виділення від 15 до 88% вихідного матеріалу. Очищений матеріал був чистим, що показано за допомогою електрофорезу на нередукованих 10-15% градієнтних гелях (не показано). Аналіз NH2-кінцевих амінокислот підтверджував послідовність Ser-Ser-Ser-Phe-Asp зрілої стафілокінази. Більш детальні біохімічні характеристики цих стафілокіназних мутантів повідомлялись раніше (41). Концентрації білка визначали по Bradford (42). Фібринолітичну активність розчинів SakSTAR визначали за допомогою дослідження з хромогенним субстратом, що проводиться в мікротитровальних платах з використанням суміші з 80мкл розчину SakSTAR та 100мкл розчину Glu-плазміногену (кінцева концентрація 0,5мМ). Після інкубації протягом 30хв. при 37°С генерований плазмід визначали кількісно шляхом додавання 30мкл S2403 (кінцева концентрація 1мкМ) та вимірювання поглинання при 405нм. Активність виражали в умовних одиницях (УО) шляхом порівняння з місцевим стандартом (сер. STAN5), який встановлював активність, рівну 100000 УО на мг білку, що визначалось по складу амінокислот (11). ДСН-ЕПАГ виконували за допомогою Phast Systemä (Pharmacia, Uppsala, Sweden), використовуючи 1015% градієнтні гелі та забарвлювання брильянтовим синім Coomassie. Відновлення зразків проводили шляхом нагрівання при 100°С протягом 3хв. у присутності 1% ДСН і 1% дитіоеритритолу. Конструювання, продукцію та очистку мутантів М3.8 та М8.9 проводили, як описано детально нижче. Олігонуклеотиди, використані для конструювання, 5'-ATAGCAATGCATTTCCTGCACTATCAAC-3' (М3), 5'-CTAATTCAACTACTGCAAACGCTGCATATGCTGTCGCATC-3’ (М8) і 5'-TTTGCGCTTGGCGCCAATGCAACTACTCTAAACTCTTTATAT-3' (М9) були синтезовані на замовлення в Pharmacia Biotech. При конструюванні мутанту М3.8 використовували однониткові pMc-STAR та фрагмент EcoRI-Hindlll рМа5-8 для одержання дуплексної молекули ДНК, що містить пробіли, яку гібридизували з синтетичним олігонуклеотидом М3 з 28 основ (що містить сайт рестрикції Nsil). Реакції подовження проводили за допомогою фрагменту Кленова ДНК-полімерази та лігази, як описано. Після переносу E.coli WK6MutS та селекції на середовищі з ампіциліном 81 колонію підрощували на нітроцелюлозних мембранах, денатурували in situ, і ДНК гібридизували протягом ночі при кімнатній температурі з використанням радіоактивно-міченого олігонуклеотиду М3 (1,5х108 імп/хв [g32Р]-АТФ на мічення полінуклеотидкіназою Т4 20-30нг олігонуклеотиду). Фільтри промивали при 42°С, використовуючи розчини, що містять 0,1% ДСН та 2xSSC, 1xSSC, 0,2xSSC, O,1xSSC. ДНК з 1 відібраного клону з 16 позитивних одержували з 150мл бактеріальних культур та аналізували за допомогою розщеплення рестрикційними ферментами Nsil і Pvul. Мутацію М3 (pMa-STAR3) підтверджували шляхом секвенування повної кодуючої області з використанням A.L.F.ä. Однониткову ДНК одержували шляхом трансформації pMa-STAR3 в E.coli, як описано вище. Однониткову pMa-STAR3 гібридизували з рМс (EcoRI-Hindlll) та з синтетичним олігонуклеотидом М8 з 40 основ, який містить сайт рестрикції Ndel, як описано вище. Після трансформації E.coli WK6MutS і селекції на середовищі з хлорамфеніколом підрощували 100 колоній на нітроцелюлозних мембранах і гібридизували з міченим олігонуклеотидом М8. Підрощували позитивні клони (2 з 7) та аналізували за допомогою розщеплення рестрикційним ферментом (Nsil-Hindlll і Ndel), що давало один позитивний клон. Подвійний мутант М3.8 потім лігірували зворотно в вектор експресії pMEX602SakB. З 12 мініпрепаратів ДНК 6 мали правильну рестрикційну карту. Один з цих клонів (pMEXSakSTAR.M38) секвенували та використовували для одержання мутанту М3.8 під контролем індуцьованого ІПТГ tac промотору та двох послідовностей Шайна-Дальгарно (Shine-Dalgarno), розташованими одна за одною. 10мкл суспензії клітин Е.соlі WK6, трансформовані за допомогою рекомбінантної плазміди pMEXSakSTAR.M38, інкубували в 100мл середовища LB (Gibco/BRL), що містить 100мкг/мл ампіциліну. Суміш інкубували протягом ночі при 37°С з перемішуванням при 200об/хв, що давало в результаті щільність клітин, рівну приблизно 5 одиницям поглинання при 600нм. Аліквоти у 20мл переносили в 2л об'єми (в 5л колбах) середовища LB, що містить 100мкл/мл ампіциліну. Суміші інкубували протягом 3 годин при 37°С при перемішуванні перед додаванням 200мкМ ІПТГ для індукції експресії М3.8, якій давали розвиватися протягом 4 годин. Клітини осаджували центрифугуванням при 4000об/хв протягом 20хв, ресуспендували в 1/10 об'єму 0,01М фосфатного буферу, pH 6,5, та руйнували ультразвуком при 0°С. Залишки клітин видаляли центрифугуванням протягом 30хв при 20000об/хв, і супернатант зберігали при 20°С до використання. У поєднаних просвітлених клітинних лізатів (2-х літрові об'єми) з 20-30 літрових бактеріальних культур pH доводили до 5,9, їх стерилізували фільтруванням через 0,22мкм фільтр Сарторіуса та наносили на 5x25см колонку сефарози, попередньо оброблену 0,5М NaOH і стерильним 0,01М фосфатним буфером, при швидкості потоку, рівній 12мл/хв при 4°С в ламінарному потоці. Колонку промивали 2-3 літрами буферу та елюювали за допомогою градієнту солі від 0 до 1М понад 500мл і від 1М до 2М понад 250мл при швидкості потоку, рівній 10мл/хв і при 4°С. Фракції, що містять М3.8, локалізовані за допомогою електрофорезу в гелі з ДСН, поєднували (приблизно 200мл) та діалізували проти 15л стерильного 0,01М фосфатного буфера, pH 8,0 при 4°С. Матеріал, що пройшов діаліз, центрифугували при 4000об/хв протягом 30хв, знов стерилізували фільтрацією та наносили на колонку 2,5x12см Q-сефарози з швидким струмом, попередньо оброблену 0,5М NaOH та стерильним 0,01М фосфатним буфером, pH 8,0, при швидкості потоку, рівній 3мл/хв при 4°С. Колонку промивали приблизно 600мл 0,01М фосфатного буферу, pH 8,0, при швидкості потоку, рівній 8мл/хв та елюювали розчином солі з градієнтом концентрації від 0 до 0,17М понад 30мл, від 0,17 до 0,2М понад 100мл та від 0,2М до 1,5М понад 200мл, при швидкості потоку, рівній 4мл/хв. Фракції, що містять М3.8, локалізовані за допомогою електрофорезу в гелі з ДСН, поєднували, концентрацію білка доводили до 1мг/мл та матеріал стерилізували шляхом фільтрації через 0,22мкм-фільтр Мілліпор. Три препарати М3.8 давали 80±25мг чистого білка (середнє±СО) зі специфічною активністю, рівною 45000±5200 УО/мг. При конструюванні мутанту М8.9 для одержання дуплексної молекули ДНК, що містить пробіли, використовували однониткову pMc-STAR і фрагмент EcoRI-Hincffll рМа5-8, які гібридизували з синтетичним олігонуклеотидом М9 з 42 основ, що містить сайт рестрикції Narl. Реакції подовження проводили за допомогою фрагменту Кленова ДНК-полімерази і лігази, як описано. Після трансформації E.coli WK6MutS та селекції на середовищі з ампіциліном, колонії підрощували на нітроцелюлозних мембранах, денатурували in situ та ДНК гібридизували протягом ночі при кімнатній температурі з використанням радіоактивно міченого олігонуклеотиду (1,5х108імп/хв [g32Р]-АТФ на мічення полінуклеотидкіназою Т4 2030нг олігонуклеотиду). Фільтри промивали при 42°С, використовуючи розчини, що містять 0,1% ДСН та 2х SSC, 1хSSC, 0,2хSSC, 0,1хSSC. Одержували ДНК з 2 відібраних клонів з 4 позитивних та 1 з них характеризували за допомогою аналізу нуклеотидної послідовності з використанням A.L.F.ä. Вставку EcoRI-HindІІІ з pMa-STAR9 потім лігірували зворотно в вектор експресії pMEX602SakB. Клони (58) сортували шляхом гібридизації in situ з радіоактивно міченим олігонуклеотидом М9 в якості проби. Один клон pMEXSakSTAR.M9 характеризували за допомогою аналізу нуклеотидної послідовності і потім використовування для конструювання мутанту М8.9. Щоб сконструювати М8.9 в М9 вводили мутацію 8 за допомогою полімеразної ланцюгової реакції (ПЛР). ПЛР проводили в загальному об'ємі, рівному 100мкл з використанням 5 од ферменту та 1мкг кожного з наступних праймерів: олігонуклеотиду ІІ=5' -CAGGAAACAGAATTCAGGAG, олігонуклеотиду ІІІ=5'TATATAATATTCGACATAGTATTCAATTTTT-3', олігонуклеотиду, ІУ=5'TATCCCGGGCATTAGATGCGAGATGCGACAGCATATGCAGCGTTTGCAGTA-3', та олігонуклеотиду V=5'CAAAACAGCCAAGCTTCATTCATTCAGC-3’. Концентрації dATP, dCTP, dGTP та dTTP складали 200мкМ. Денатурацію проводили протягом одної хвилини при 94°С, з ренатурацією протягом 2хв при 55°С та подовженням протягом 1,5хв при 72°С. Після 30 циклів зразки інкубували протягом 10 хвилин при 72°С та охолоджували до 4°С. У першій реакції ПЛР 2нг pMEXSakSTAR.M9 ампліфікували з використанням олігонуклеотидів IV і V як праймерів. Амплікон ПЛР розщеплювали за допомогою Smal і HindІІІ та очищували після електрофорезу в 1,5% агарозному гелі при використанні набору Prep-A-gene (Bio-Rad Laboratories, Hercules, CA, USA). Одержаний в результаті фрагмент клонували в сайти SmaI-HindІІІ pUC18 (Pharmacia BioTech, Uppsala, Sweden) з використанням набору для швидкого лігірування ДНК (Boeringer Mannheim). Після трансформації в клітини Е.соlі WK6, одержували ДНК з 12 колоній, і всі 12 генерували фрагмент в приблизно 230 пар основ при розщеплені EcoRI та HindІІІ. Одну з цих ДНК (pUC18-M89D) використовували для клонування другого продукту ПЛР (див. нижче). Другу реакцію ПЛР виконували на 2нг pMEXSakSTAR.M9 з використанням олігонуклеотидів II і III як праймерів. Продукт реакції ПЛР розщеплювали за допомогою Sspl та EcoRI та додатково очищували, як описано вище. Одержаний в результаті фрагмент лігірували в сайти Smal-EcoRI pUC18-M89 А. Після трансформації у клітини Е.соlі WK6 відбирали 6 клонів для одержання ДНК. П'ять з 6 генерували фрагмент з приблизно 453 пар основ після розщеплення за допомогою EcoRI та HindІІІ. Цей фрагмент, кодуючий весь мутант М8.9, клонували у сайти EcoRI-HindІІІ вектора експресії pMEX602SakB. Після трансформації клітин Е.соlі WK6 ДНК з 6 колоній аналізували за допомогою розщеплення EcoRI та HindІІІ, що дає фрагмент з приблизно 453 пар основ у всіх випадках. Одну з цих ДНК, крім того, характеризували шляхом аналізу нуклеотидної послідовності. 100мл середовища LB (GIBCO/BRL), що містить 100мкг/мл ампіциліну засівали 100мкл суспензії клітин Е.соlі WK6, трансформованих рекомбінантною плазмідою pMEXSakSTAR.M89. Культуру інкубували протягом ночі при 37°С з перемішуванням при 140об/хв до щільності клітин, рівної приблизно 5 одиницям поглинання при 600нм. Аліквоти по 4мл використовували для інокуляції 2-літрових культур (в колбах з перегородкою SL) в середовищі «Terrific Broth», що містить 150мкл/мл ампіциліну. Культури інкубували протягом приблизно 20 годин при 30°С та при 140об/хв, одержуючи в результаті кінцеву щільність клітин, рівну приблизно 4x109клітин/мл. Клітини осаджували центрифугуванням при 4000об/хв протягом 20хв, ресуспендували в 1/5 об'єму 0,01М фосфатного буферу, pH 6,5 та руйнували ультразвуком при 0°С. Потім pH доводили до 5,8 та залишки клітин видаляли 30-хвилиннм центрифугуванням при 20000об/хв. Супернатант зберігали при -20°С до наступної обробки. В об'єднаному просвітленому лізаті клітин (1800мл) pH доводили до 5,8 і наносили його на колонку 2,5x20см з SP-сефарози, попередньо обробленої 0,5М NaOH та свіжим буфером з 0,01М фосфату, 2,5М NaCl, pH 7,5 при швидкості потоку, що дорівнює 2мл/хв при 4°С. Колонку промивали 500мкл буферу та елюювали градієнтом солі від 0 до 1 М понад 200мл при швидкості потоку, що дорівнює 6мл/хв. Об'єднані фракції М8.9, ідентифіковані за допомогою електрофорезу з ДСН, доводили до 2,5М твердим NaCl та піддавали хроматографії гідрофобної взаємодії на колонці 2,5x20см феніл-сефароза, попередньо обробленої 0,5М NaOH та свіжим буфером з 0,01М фосфату, 2,5М NaCl з pH 7,5 при швидкості потоку, рівній 2мл/хв і 4°С. Колонку промивали приблизно 500мл буферу та елюювали 0,01М фосфатним буфером, pH 6,5. Фракції, що містять М8.9, локалізовані за допомогою електрофорезу в гелі з ДСН, об'єднували та піддавали діалізу в 2 літрах 0,01М фосфатного буферу, pH 9,0. Діалізований матеріал центрифугували при 4000об/хв протягом 30хв і наносили на 1,6x5см колонку Q-сефарози з швидким потоком, попередньо оброблену 0,5М NaOH та свіжим 0,01М фосфатним буфером, pH 9,0, при швидкості потоку, що дорівнює 2мл/хв і 4°С. Колонку промивали приблизно 150мл 0,01М фосфатного буферу, pH 9,0, та елюювали градієнтним розчином солі від 0 до 1М NaCl вище 100мл при швидкості потоку, що дорівнює 4 мл/хв. Колонку промивали приблизно 150мл 0,01М фосфатного буферу, pH 9,0, та елюювали градієнтом солі від 0 до 1М NaCl понад 100мл при швидкості потоку, що дорівнює 4мл/хв. Фракції, що містять М8.9, локалізовані за допомогою електрофорезу на гелі з ДСН, об'єднували і концентрацію білка доводили до 1мг/мл, і матеріал стерилізували фільтруванням через 0,22мкМ фільтр Мілліпор. Ці препарати М8.9 давали 73±17мг чистого білка зі специфічною активністю, рівною 51000±3500УО/мг. Результати по фібринолітичній активності різних мутантів SakSTAR, визначеній за допомогою дослідження з хромогенним субстратом, узагальнені в таблиці 1. З 21 мутанту, сконструйованого, як показано на фігурі 1, Е99, E100 (М13) і Е99, Е100, Е102 (М14) можна було одержати в очищеній формі, тоді як К11, D13, D14 (М1), Е46, Л50 (М4) і Е65, D69 (М7) були неактивними. Шістнадцять мутантів, зведених в таблицю 1, були вивчені детально разом з SakSTAR дикого типу. З цих мутантів D5, К6 (M20), K8, К10 (М21), D33, К35 (М2), К57, Е58, К59 (М5), Е61, Е65 (М6), К86, Е88 (M10), D93, К94 (М11), К96, К97, К98 (М12), Е108, К109 (М15), D115, Е118, Н119 (М16), Н119, К121 (М17), К130 (М18) і Е134, К135, К136 (М19) реагували з панеллю моноклональних антитіл також, як і SakSTAR. Однак К35, Е38 (М3) і Е80, D82 (М9) слабо реагували з антитільними кластерами 7Н11, 25Е1, 40С8, тоді як К74, Е75, R77 (М8) слабо реагували кластером 26А2, ЗОА2, 2В12 і 3G10. Адитивність елімінації епітопу була встановлена з мутантами К35, Е38ЛС74, Е75, R77 (М3.8) і К74, Е75, R77/E80, D82 (М8.9), які поєднували знижену реактивність з моноклональними антитілами обох батьківських молекул. Приклад 3 Адсорбція з варіантами антитіл до стафілокінази дикого типу і «заряджений-кластер-на-аланін», виявлених у хворих при лікуванні SakSTAR Щоб одержати інформацію по епітопній специфічності індукованих антитіл, виявлених у хворих з гострим інфарктом міокарду після лікування SakSTAR, зразки плазми від 16 хворих абсорбували за допомогою молярного надлишку (над нейтралізуючою активністю стафілокінази) одного або поєднаних мутантів «заряджений-кластер-для-аланіну» протягом 10 хвилин перед визначенням залишкового зв'язування з SakSTAR за допомогою аналізу біоспецифічної взаємодії. Нейтралізуючу активність стафілокінази у цих зразків визначали таким чином. Підвищені концентрації варіанту дикого типу або SakSTAR (об'єми 50мкл, що містять 0,2-1000мкг/іл) додавали до суміші 300мкл цитратної людської плазми та 50мкл буферу або досліджуваної плазми, відразу після цього додавали 100мкл суміші, що містить тромбін (50NIH од/мл) та СаСl (25мМ.) Час лізису згустків плазми вимірювали та наносили на графік залежності від концентрації SakSTAR складової. По цій кривій визначали концентрацію активатора плазміногену, яка викликала повний лізис згустку через 20 хвилин. Титр нейтралізуючої активності визначали як різницю між значеннями для досліджуваної плазми та буферу і виражали в мкг на мл досліджуваної плазми. Результати підсумовані в таблиці 2. В той час як SakSTAR дикого типу абсорбував більше 90 відсотків зв'язуючих антитіл з всіх зразків, з мутантом К35, Е38 (М3) у 4 хворих, з мутантом К74, Е75, R77 (М8) у 12 хворих та з мутантом Е80, D82 (М9) у 5 хворих спостерігалась неповна абсорбція. Абсорбція з поєднаними мутантами К35, Е38/К74, Е75, R77 (М3.8) та К74, Е75, R77/E80, D82 (М8.9) видаляла менше 90% антитіл у 13 хворих (середнє значення з 68 та 65 відсотків, відповідно, від 16 хворих), тоді як очікувалось, суміш вихідних молекул поєднаних мутантів (М8 і МЗ або М9) постійно абсорбували понад 90% антитіл. Приклад 4 Імуногенність варіантів стафілокінази “заряджений-кластер-на-аланін)” у кролів, імунізованих стафілокіназою (SakSTAR) дикого типу, мутантами К35. Е3.8 (М3), К74, Е75, R77(M8) і Е80, D82 (М9) та поєднаними мутантами К35, Е38/К74, Е75, R77 (М3.8) і К74, Е75, R77/E80, D82, (М8.9) Порівняну імуногенність SakSTAR щодо кожного з варіантів SakSTAR, М3, М8, М9, М3.8 та М8.9, вивчали після імунізації підшкірним введенням у групах з 4 або 8 кролів, призначених для введення SakSTAR, і в групах з 8 кролів, призначених для введення певного варіанту. Імунізацію проводили шляхом внутрівенного вливання 400мкг/кг SakSTAR та 200-1000мкг/кг мутантів на тиждень 0 (для визначення вихідної здатності лізису згустків) з наступною підшкірною ін'єкцією 400мкг тієї ж самої речовини у повному ад'юванті Фрейнда на 2-й тиждень та в неповному ад'юванті Фрейнда на 3 і 5 тижні. Імуногенність оцінювали кількісно через 6 тижнів шляхом визначення активності, що нейтралізує стафілокіназу, в плазмі та залишкової тромболітичної здатності, як описано детально нижче. Коротко, активність по нейтралізації стафілокінази в плазмі визначали шляхом додавання підвищених концентрацій SakSTAR дикого типу або мутанту SakSTAR (50мкл об'єми, що містять 0,2-1000мкг/мл) до суміші 300мкл цитратної людської плазми та 50мкл буферу або кролячої плазми з негайним наступним додаванням 100мкл суміші, що містить тромбін (50NIH од/мл) і СаСl2 (25мМ). Визначали час лізису згустку плазми та наносили його на графік залежності від концентрації SakSTAR або варіанту. По цій кривій визначали концентрацію активатору плазміногену, яка давала повний лізис згустку через 20 хвилин. Титр нейтралізуючої активності визначали як різницю між значеннями для кролячої плазми та для буферу і виражали в мкг на мл кролячої плазми. Тромболітичні властивості вивчали, використовуючи 0,3мл згустки кролячої плазми, збідненої тромбоцитами, мічені 125І-фібрином, поміщені в екстракорпоральні артеріовенозні петлі. Виділену стегнову артерію для цього катетеризували катетером Френча 4 (Portex White, Portex, Hythe, UK) і з'єднували через два підшкірних шприца з катетеризованою вушною веною. Потік крові через екстракорполярну петлю підтримували на рівні 10млн/хв за допомогою перистальтичного насосу. Згустки плазми з міченими 125Іфібрином вводили в кожний з двох шприців, вставлених в петлю. Згустки плазми одержували шляхом змішуванням 0,3мл бідної тромбоцитами плазми з невеликою кількістю (приблизно 1,5мкКю) розчину міченого 125І-людського фібриногену (Amersham, Buckinghamshire, UK) і 0,5М СаСl2 з наступною інкубацією протягом 30хв при 37°С. За тридцять хвилин до початку інфузії вводили 7,5мг/кг рідогреля (поєднані інгібітор тромбоксансінтази і антагоніст простагландін-ендопероксидних рецепторів) (43) у вигляді швидкої внутрівенної ін'єкції для попередження осадження тромбоцитів в екстракорпоральній петлі. Тваринам для антикоагуляції вводили гепарин (300од/кг з наступним безперервним вливанням 200од/кг/годину під час експерименту) та розподіляли по випадковій виборці на вливання 400мкг/кг SakSTAR (4-8 кролів) або на вливання 200-1000мкг/кг варіанту SakSTAR (8 кролів). Через 6 тижнів половині кролів призначених для варіанту SakSTAR знову вводили той же самий варіант SakSTAR, a другій половині вводили SakSTAR дикого типу, крім того кролям, імунізованим SakSTAR, вводили або варіант SakSTAR (якщо ця контрольна група складалась з 4 кролів) або по випадковій виборці - SakSTAR дикого типу або варіант SakSTAR (якщо ця контрольна група складалась з 8 кролів). Тромболітичні засоби вводили внутрівенно у вигляді ударного 10% або 90% вливання протягом 1 години. Хід з часом лізису згустку контролювали безперервно шляхом зовнішньої оцінки з гамма-випромінювання з використанням двох 3x0,5 дюймових кристалів йодиду натрію/талію (Bicron, Newbury, ОН), поміщених над екстракорпоральними петлями, Сцинтиляційні кристали з'єднували з пристосованою системою Canberra-SlOO (Canberra-Packard, Meriden, CT) і дані були проаналізовані, як описано раніше (44). У кінці експерименту залишкові згустки також видалялися з шприців для визначення вмісту в них радіоізотопу. Експерименти на тваринах проводили у погодженні з керівними принципами Американського фізіологічного товариства та Міжнародного комітету по тромбозу та гемостазу (45). Імуногенність SakSTAR і відповідних одиночних мутантів (М3, М8 і М9) порівнюється в таблиці 3А. Результати виражені як середнє значення ±СКО. У 8 кролів випадок вибраних для мутанту М3 початкова нейтралізуюча активність складала 0,0±0,0 мкг/мл як щодо SakSTAR, так і М3. Внутрівенне вливання 200мкг/кг М3 викликало 76±23-відсотковий лізис згустку. Ці 8 кролів потім імунізували М3, суспендованим у 500мкл повного ад'юванту Фрейнду через 2 тижні та в 500мкл неповного ад'юванту Фрейнду через 3 і 5 тижнів. Через 6 тижнів нейтралізуюча активність плазми збільшилася до 11±6,7мкг/мл щодо SakSTAR і до 11±7.2мкг/мл щодо М3. Через 6 тижнів вливання 400мкг SakSTAR у 4 з цих кролів, вибраних випадковим чином, викликало 18±27-відсотковий лізис згустків, тоді як вливання 200мкг/кг М3 у 4 інших кролів викликало 16±17-вісотковий лізис. У 4 кролів, призначених для введення SakSTAR, вихідна нейтралізуюча активність склала 0,2±0,2мкг/мл щодо SakSTAR і 0,0±0,0мкг/мл щодо М3. Внутрівенне вливання 400мкг/кг SakSTAR продукувало 89±8,6відсотковий вихідний лізис. Цих чотирьох кролів потім імунізували 400мкг SakSTAR, суспендованими в або 500мкг повного (на другому тижні) або неповного (на 3 і 5 тижні) ад'юванту Фрейнду. На 6 тижні нейтралізуюча активність плазми збільшилася до 35±23 мкг/мл щодо SakSTAR і до 19±13мкг/мл щодо М3. Внутрівенне вливання 200мкг/кг SakSTAR. МЗ у цих кролів на 6 тижні індукувало 9,3±8,2-відсотковий лізис. У 8 кролів, призначених для мутанту М8, вихідна нейтралізуюча активність у плазмі складала 1,4±0,2мкг/мл щодо SakSTAR та 0,6±0,5мкг/мл щодо М8. Внутрівенне вливання 1000мкг/мл М8 продукувало 41±13-відсотковий лізис. Цих кролів імунізували 400мкг М8, суспендованого в повному ад'юванті Фрейнду на 2 тижні та тією ж самою кількістю в неповному ад'юванті Фрейнду на 3 та 5 тижні. На 6 тижні нейтралізуюча активність плазми підвищувалась до 3,8±1,8мкг/мл щодо SakSTAR і до 5,9±2,7мкг/мл щодо М8. Вливання 400мкг/кг SakSTAR у 4 з цих кролів давало 49±28-відсотковий лізис, тоді як вливання 1000мкг/кг М8 у 4 інших кролів продукувало 24±11-відсотковий лізис. У 8 кролів, призначених для групи SakSTAR вихідна нейтралізуюча активність в плазмі складала 0,9±0,6мкг/мл щодо SakSTAR і 0,6±0,3мкг/мл щодо М8. Внутрівенне вливання 400мкг/кг SakSTAR продукувало 68±18-відсотковий лізис. Цих кролів потім імунізували підшкірно 400мкг SakSTAR, суспендованого в повному ад'юванті Фрейнду на 2 тижні та тією ж самою кількістю в неповному ад'юванті Фрейнду на 3 та 5 тижні. На 6 тиждень нейтралізуюча активність плазми збільшувалась до 59±47мкг/мл щодо SakSTAR і до 22±16мкг/мл щодо М8, тоді як залишкова тромболітична здатність 400мкг/кг SakSTAR знижувалась до 7,5±2,4 відсотків і 1000мкг/кг М8 - до 4,1±4,8 відсотків. У 8 кролів, призначених для мутанту М9, вихідна нейтралізуюча активність у плазмі складала 0,2±0,05 мкг/мл щодоSakSTAR і 0,03±0,05мкг/мл щодо М9. Внутрівенне вливання 400мкг/кг М9 продукувало 72±11 відсотковий лізис згустку. Цих кролів потім імунізували 400мкг М9, суспендованими в повному ад'юванті Фрейнду на другому тижні та тією ж кількістю в неповному ад'юванті Фрейнду на 3 та 5 тижні. На 6 тижні нейтралізуюча активність плазми підвищувалась до 8,0±4,6мкг/мл щодо SakSTAR і до 3,5±2,6мкг/мл щодо М9. На 6 тижні вливання 400мкг/кг SakSTAR у 4 цих кролів продукувало 53±11-відсотковий лізис згустків, тоді як вливання 400мкг/кг М9 у 4 інших кролів давало 40±7,8 відсотковий лізис. У 4 контрольних кролів, призначених для SakSTAR, вихідна нейтралізуюча активність плазми складала 0,1±0,05мкг/мл щодо SakSTAR і 0,05±0,06мкг/мл щодо М9. Внутрівенне вливання 400мкг/кг SakSTAR давало 78±13-відсотковий лізис. Цих кролів потім імунізували 400мг SakSTAR, суспендованим у повному (2-й тиждень) і неповному (3 та 5 тиждень) ад'юванті Фрейнда, відповідно. На 6 тижні нейтралізуюча активність у плазмі підвищувалась до 16±5,0мкг/мл щодо SakSTAR і до 12±9,1мкг/мл щодо М9, тоді як тромболітична здатність М9 знижувалася до 24±33 відсотків. Імуногенність SakSTAR порівняно з подвійними мутантами М3.8 і М8.9 порівнюється у таблиці 3В. У 8 кролів, призначених для групи М3.8, вихідна нейтралізуюча активність у плазмі складала 0,6±0,3мкг/мл щодо SakSTAR і 3,5±2,0мкг/мл щодо М3.8. Внутрівенне вливання 1000мкг/кг М3.8 продукувало 53±13% лізис. Цих кролів потім імунізували 400мкг М3.8, суспендованими у повному ад'юванті Фрейнда, на 2 тижні та тією ж самою кількістю у неповному ад'юванті Фрейнда на 3 і 5 тижні. На 6 тижні нейтралізуюча активність плазми підвищувалась тільки до 1,7±0,7мг/мл щодо SakSTAR і до 6,1±3,0мкг/мл щодо М3.8. Вливання 400мкг/мл SakSTAR у 4 з цих кролів продукувало 77±18 відсотковий лізис згустку, тоді як вливання 1000мг/кг М3.8 у 4 інших кролів давало 59±25-відсотковий лізис. У 8 кролів, призначених для групи SakSTAR, вихідна нейтралізуюча активність у плазмі складала 0,6±0,4мкг/мл щодо SakSTAR і 2,0±2,0мкг/мл щодо М3.8. Внутрівенне вливання 400мкг/мл SakSTAR продукувало 80±10 відсотковий лізис. Цих кролів потім імунізували підшкірно 400мкг SakSTAR, суспендованими у повному ад'юванті Фрейнда, на 2 тижні та тією ж самою кількістю у неповному ад'юванті Фрейнда на 3 та 5 тижні. На 6 тижні нейтралізуюча активність у плазмі збільшувалась до 20±15мкг/мл щодо SakSTAR і до 21±22мкг/мл щодо М3.8, тоді як залишкова тромболітична здатність 400мкг/кг SakSTAR знижувалась до 8,5±5,7 відсотків, а 1000мкг/кг М3.8 до 30±29 відсотків. У 8 кролів призначених для групи М8.9 вихідна нейтралізуюча активність у плазмі складала 0,3±0,2мкг/мл щодо SakSTAR і 1,6±0,5мкг/мл щодо М8.9. Внутрівенне вливання 800мкг/кг М8.9 продукувало 39±13-відсотковий лізис згустку в вихідний момент. Цих 8 кролів потім імунізували 400мкг М8.9, суспендованими у повному (2 тижні) або в неповному (тижні 3 і 5) ад'юванті Фрейнду. Через 6 тижнів нейтралізуюча активність плазми збільшувалась тільки до 2,5±1,5мкг/мл щодо SakSTAR і до 4,9±1,3мкг/мл щодо М8.9. На 6 тижні вливання 400мкг/кг SakSTAR у 4 з цих кролів продукувало 51±35 відсотковий лізис згустку, в той час як вливання 800мкг/кг М8.9 і у інших кролів давало 39±12 відсотковий лізис. У 4 контрольних кролів, призначених для SakSTAR, нейтралізуюча активність до введення препаратів становила 0,2±0,1мкг/мл щодо SakSTAR і 0,7±0,3мкг/л щодо М8.9 Внутрівенне вливання 400мкг/кг SakSTAR викликало 67±19 відсотковий лізис згустку. Цих 4 кролів потім імунізували 400мкг SakSTAR, суспендованими у повному (тиждень 2) або неповному (тиждень 3 і 5) ад'юванті Фрейнда. На 6 тижні нейтралізуюча активність у плазмі підвищувалась до 20±15 мкг/мл щодо SakSTAR і до 18±15мкг/мл щодо М8.9, тоді як залишкова тромболітична дія М8.9 знижувалась тільки до 31±30-відсоткового лізису. Ці результати показують, що в цьому прямому порівняльному дослідженні SakSTAR і вибраних варіантів, особливо подвійні мутанти (М3.8 та М8.9) індукують значно меншу нейтралізуючу активність, пов'язану з антитілами, та стійкість до лізису, ніж SakSTAR. Приклад 5 Порівняна імуногенність SakSTAR і М3.8 у бабуїнів Порівняну імуногенність SakSTAR і М3.8 щодо індукції нейтралізуючих антитіл і рефрактерності до тромболізу при повторному введенні вивчали на бабуїнах. Дослідження на тваринах виконували відповідно до керівних принципів Американського фізіологічного товариства і Міжнародного комітету з тромбозів та гемостазу (45). У анестезованих та інтубованих бабуїнів створювали екстракорпоральний артеріовенозний кровопотік шляхом з'єднання через зовнішню поліетиленову петлю катетеризованої великогомілкової або плечової артерії з периферичною веною. Перистальтичний насос спрямовував і підтримував безперервно контрольований потік крові через екстракорпоральну петлю, в якій були вставлені два пристосованих шприца для підшкірного введення, кожний з яких мав один свіжий 0,3мл згусток спільної плазми бабуїнів з міченим 125І фібрином. Хід лізису згустку під час вливання SakSTAR або варіанту М3.8 безперервно контролювали шляхом зовнішнього вимірювання гамма-випромінення над шприцами. В якості альтернативи підраховували баланс визначення ізотопу шляхом порівняння суми підрахування радіоактивності у всій крові в кінці експерименту (помноженої на фактор 3 для корекції на екстраваскулярне розподілення) плюс радіоактивність у повернених тромбах, разом з тою, яка початково була присутня у згустках. Перед кожним експериментом тромболізу бабуїнам попередньо вводили внутрівенно ударну дозу рідогрелю 3мг/кг для запобігання осіданню тромбоцитів у екстракорпоральній системі. Протягом експериментів тромболізу внутрівенно вводили гепарин у вигляді ударної дози 300ME/кг з наступним вливанням 200МЕ/кг/год. Дванадцять дорослих самців-бабуїнів (Раріо Hamadryas) по випадковій виборці на початку експерименту (тиждень 0) розподіляли на введення 50мкг/кг SakSTAR (група 1) або варіанту М3.8 (група 2), що вливаються внутрівенно протягом одної години шляхом 10% ударної дози, та вихідну тромболітичну здатність оцінювали шляхом спостереження за зникненням радіоактивності зі згустків протягом 2 годин. Потім бабуїнів імунізували підшкірним введенням 500мкг або SakSTAR (група 1) або варіанту М3.8 (група 2), суспендованих у повному ад'юванті Фрейнду, через 2 тижні та в неповному ад'юванті Фрейнда через 3 і 5 тижнів. Через 6 тижнів тромболітичну дію оцінювали кількісно за допомогою моделі екстракорпорального тромболізу протягом 4 годин: по випадковій виборці відбирали 3 з 6 бабуїнів з групи 1, яким вводили на початку експерименту SakSTAR, а потім їх імунізували, і 3 з 6 бабуїнів групи 2, яким на початку експерименту вводили М3.8 і яких потім ним імунізували, та вводили їм спочатку 50мкг/кг SakSTAR шляхом внутрівенного вливання протягом одної години одної ударної 10% ін'єкції, а потім через 2 години від початку вливання SakSTAR давали М3.8 у тому ж режимі (групи 1А і 2А, відповідно). Інші 6 бабуїнів одержували ті ж препарати, але у зворотному порядку: спочатку М3.8, потім SakSTAR (групи 1В і 2В для бабуїнів, попередньо імунізованих SakSTAR і М3.8 відповідно). Через 18 тижнів тромболітичну активність оцінювали втретє шляхом контролю зникнення радіоактивності з фібринових згустків протягом 3 годин: 3 з 6 бабуїнів, імунізованих SakSTAR, вводили 250мкг/кг SakSTAR у вигляді внутрівенної ударної ін'єкції протягом 2,5 хвилин (група 1 А), в той час як 3 інші бабуїни, імунізовані SakSTAR, одержували ту ж саму кількість М3.8 (група 1В). З 6 бабуїнів імунізованих М3.8, 3 одержували внутрівенну ударну ін'єкцію 250мкг/кг SakSTAR (група 2А) та 3 - ту ж саму кількість М3. 8 (група 2В). Зразки крові збирали у цитратні пробірки (кінцева концентрація 0,01М) на початку дослідження і через певні проміжки часу потім для вимірювання активованого парціального тромбопластинового (аПТВ), визначення фібриногену, a2-антиплазміна (на початку та в кінці кожного експерименту) та активності по нейтралізації SakSTAR і М3.8 (перед тромболітичним вливанням). Для цього до суміші 300мкл цитратної людської плазми та 50мкл буферу або досліджуваної плазми бабуїнів додавали кількості, що збільшуються, або a2-антиплазміна або М3.8 (50мкл об'єми, які містять від 0,2 до 1000мкг/мл) безпосередньо після додавання 100мкл суміші тромбіну (50 NTH Од/мл) та СаСl2 (25мМ). Час лізису згустку плазми вимірювали та наносили на графік залежності від концентрації SakSTAR або М3.8. По цій кривій визначали концентрацію активатора плазміногену, яка давала повний лізис згустку через 20 хвилин. Нейтралізуючу активність визначали як різницю між значеннями для досліджуваної плазми і для буфера і виражали в мкг/мл плазми. Системний фібриноген не руйнувався, як і не виснажувався a2-антиплазмін, що відображало загальну фібриноспецифічність обох препаратів. По закінченні 6 тижнів нейтралізуюча активність щодо SakSTAR у групи 1 була значно вище, ніж нейтралізуюча активність щодо М3.8 у групі 2. У групі 1 нейтралізуюча активність розвивалася скоріше та значно помітніше щодо SakSTAR, ніж щодо М3.8, тоді як активність нейтралізації М3.8 ніколи не перевищувала активність нейтралізації SakSTAR у групі 2 (таблиця 4). По закінченні 8 тижнів у групі 1 розвивалась значно більша нейтралізуюча активність як щодо SakSTAR, так і щодо М3.8, ніж у групі 2 (таблиця 4). На початку дослідження 50мкг/кг SakSTAR, що вливали внутрівенно протягом одної години, індукували 77±2,9% лізис згустку через 2 години у 6 бабуїнів (група 1) і 50мкг/кг М3.8, індукували 83±3,6% лізис згустку через 2 години у 6 інших бабуїнів (група 2) (середнє ±СКО), р=0,2. Через 6 тижнів лізируюча дія 50мкг/кг SakSTAR, що вливалися внутрівенно протягом 1 години, через 2 години знижувалась до 9,2±1,0% у 3 бабуїнів, імунізованих SakSTAR (група 1А) і до 8,5±3,2% у 3 бабуїнів, імунізованих М3.8 (група 2А), в той час як ефективність лізису 50мкг/кг М3.8 введених внутрівенно протягом 2 годин, знижувалась до 10±6,9% у трьох бабуїнів, імунізованих SakSTAR (група 1В) і до 11±7,4% у 3 бабуїнів, імунізованих М3.8 група 2В; р
ДивитисяДодаткова інформація
Назва патенту англійськоюNovel staphylokinase derivatives, a method for the preparation thereof and pharmaceutical composition
Назва патенту російськоюНовые производные стафилокиназы, способ их получения и фармацевтическая композиция
МПК / Мітки
МПК: A61K 38/48, A61K 38/16, C07K 14/31, C12N 15/31, A61K 39/02, C12N 9/52
Мітки: спосіб, композиція, одержання, фармацевтична, стафілокінази, похідні, нові
Код посилання
<a href="https://ua.patents.su/16-64693-novi-pokhidni-stafilokinazi-sposib-kh-oderzhannya-ta-farmacevtichna-kompoziciya.html" target="_blank" rel="follow" title="База патентів України">Нові похідні стафілокінази, спосіб їх одержання та фармацевтична композиція</a>
Нові проміжні продукти для одержання n,n’-місткових бісіндолілмалеімідів, спосіб отримання цих проміжних продуктів та фармацевтична композиція на їх основі

Номер патенту: 56145
Опубліковано: 15.05.2003
Автори: Фоул Маргарет Мері, Крамріх Крістін Енн, Уіннероскі Леонард Ларрі, мол.
МПК: A61P 25/28, A61P 25/00, A61P 9/00, A61P 3/00, A61K 31/407, A61K 31/00, A61P 29/00, A61P 17/00, C07D 498/22, A61P 9/10, A61P 3/10, A61K 31/40, A61P 35/00
Мітки: одержання, n,n'-місткових, продукти, продуктів, проміжних, проміжні, спосіб, цих, композиція, фармацевтична, отримання, основі, нові, бісіндолілмалеімідів
Формула / Реферат:
1. Сполука формули,де R1 - Вr, І або O-тозил.2. Сполука за п. 1 формули3. Сполука за п. 1 або п. 2, де R1 - Вr або І.4. Фармацевтична лікарська форма, до складу якої входить сполука за будь-яким з пп. 1-3 та один або більше фармацевтично прийнятних носіїв, розріджувачів або наповнювачів.5. Спосіб одержання амінозаміщеного N,N'-місточкового бісіндолілмалеіміду формули,де: R2 -...
Похідні еритроміцину, спосіб їх одержання та фармацевтична композиція
Номер патенту: 48244
Опубліковано: 15.08.2002
Автори: Дені Алексіс, Агурідас Константен, Пежак Жан-Марі, Шанто Жан-Франсуа
МПК: C07H 17/08, A61K 31/7048, A61P 31/04
Мітки: фармацевтична, похідні, спосіб, еритроміцину, композиція, одержання
Формула / Реферат:
1. Похідні еритроміцину формули І;,де R являє собою атом водню; алкільний радикал, що містить аж до 12 атомів вуглецю, який може бути заміщеним галогеном чи радикалами (СН2)mАr або - (СН2)n-(СA=СB)-(СН2)mАr, деm - ціле число від 1 до 8, n і p, однакові чи різні, позначають цілі числа від 0 до 6; А і В, однакові чи різні, являють собою атом водню або галогену чи алкіл, що містить аж до 8 атомів вуглецю; Аr - арильний або...
Похідні бензоїлгуанідину, спосіб їх одержання та фармацевтична композиція на їх основі

Номер патенту: 48214
Опубліковано: 15.08.2002
Автори: Роос Отто, Бюргер Еріх, Айкмайєр Крістіан
МПК: A61K 31/495, C07D 207/34, A61P 9/06, C07D 333/40, C07D 333/38, A61P 9/00, A61K 31/40, C07D 307/68, C07D 307/52, A61K 31/00, A61K 31/4025, A61K 31/496, C07D 295/192, A61P 35/00, A61P 9/10, C07D 295/155, C07D 295/185, C07D 295/12
Мітки: основі, фармацевтична, композиція, бензоїлгуанідину, похідні, спосіб, одержання
Формула / Реферат:
1. Похідні бензоїлгуанідину загальної формули (І): деR1 означає алкіл з 1-8 атомами вуглецю,А означає групу:В означає групу (-CH2-)a або (-СО-)b, деа означає ціле число від 0 до 8, переважно 1, 2, 3 або 4,b означає 0, 1 або 2, переважно 1,R2 означає незаміщений або заміщений алкіл з 1-8 атомами вуглецю, незаміщений феніл, NR3R4, де R3 і R4 кожен означає алкіл з...
Похідні карбапенему, їх фармацевтично прийнятні солі та ефіри, що гідролізують in vivo, спосіб їх одержання (варіанти), фармацевтична композиція, яка має антибактеріальну активність, похідні піролідин-4-іл-тіол

Номер патенту: 44885
Опубліковано: 15.03.2002
Автори: Дейвіс Гарет Морс, Свейн Майкл Лінгард, Беттс Майкл Джон
МПК: C07D 403/12, C07F 9/6574, C07D 207/16, A61K 31/397, C07D 477/00, C07F 9/568, A61K 31/40, C07F 9/572, A61P 31/04
Мітки: фармацевтично, гідролізують, композиція, прийнятні, фармацевтична, одержання, антибактеріальну, vivo, карбапенему, яка, варіанти, похідні, має, солі, активність, спосіб, піролідин-4-іл-тіол, ефіри
Формула / Реферат:
1. Производные карбапенема общей формулы I в которой R1 - оксиметил или 1-оксиэтил,R2 - водород или (С1-С4)-алкил,R3 - водород или (С1-С4)-алкил,R4 и R5, одинаковые или различные, представляют собой атом водорода, атом галогена, амино, цианогруппу, (С1-С4)-алкил, гидроксигруппу, карбоксигруппу, (С1-С4)алкоксигруппу, (С1-С4)алкоксикарбонил, карбамоил, (С1-С4)алкилкарбамоил,...
Похідні 2-(4-заміщеного)бензиламіно-2-метилпропанаміду, спосіб їх одержання та фармацевтична композиція

Номер патенту: 44331
Опубліковано: 15.02.2002
Автори: Бонсіньорі Альберто, Варазі Маріо, Амічі Рафаелла, Певарелло Паоло, Сальваті Патрісія
МПК: C07C 231/00, C07C 323/36, C07C 323/32, A61K 31/00, A61P 25/24, C07C 237/06, A61P 25/16, A61K 31/165, A61P 25/20
Мітки: композиція, спосіб, 2-(4-заміщеного)бензиламіно-2-метилпропанаміду, похідні, одержання, фармацевтична
Формула / Реферат:
1. Похідні 2-(4-заміщеного)бензиламіно-2-метилпропанаміду формули (І)де:n дорівнює нулю, 1, 2 або 3;X означає -О-,-S-, -СН2- або -NH-;R та R1 кожен незалежно означає водень, С1-С6-алкіл, галоген, гідрокси, С1-С4-алкокси або трифторметил;R2, R3 та R4 кожен незалежно означає водень, С1-С6-алкіл або С3-С7-циклоалкіл;або її фармацевтично прийнятна сіль як активний терапевтичний засіб.2....
Попередній патент: Ліофілізат ліпідного комплексу нерозчинного у воді камптотецину
Наступний патент: Еластомерна композиція, суміш для її виготовлення, спосіб її одержання, виріб на її основі
Випадковий патент: Спосіб дослідження фізико-механічних властивостей грунтів