Рослини, які мають підвищений врожай, та спосіб їх одержання























Формула / Реферат
1. Спосіб підвищення врожаю рослин, який включає введення у рослину нуклеїнової кислоти цикліну А2, яка кодує білок цикліну А2, при цьому нуклеїнова кислота цикліну А2 є оперативно зв'язаною з промотором, специфічним для насіння.
2. Спосіб згідно з пунктом 1, в якому вказаний врожай рослини є вибраним з однієї або більше наступних характеристик: підвищеної ваги насіння, підвищеної кількості насіння, збільшеного розміру насіння, підвищеного індексу збору врожаю, збільшеної ваги тисячі зернин та модифікованого складу насіння, кожна характеристика у порівнянні з контрольними рослинами.
3. Спосіб згідно з пунктами 1 або 2, в якому вказаний білок цикліну А2 включає фрагмент, який складається з W L V/I Е V S/A D/E D/E Y K/R/T L.
4. Спосіб згідно з будь-яким з пунктів 1-3, в якому вказана нуклеїнова кислота цикліну А2 вибрана з цикліну А2;1, цикліну А2;2, цикліну А2;3 та цикліну А2;4.
5. Спосіб згідно з пунктом 4, в якому вказаний циклін А2 включає фрагмент, який складається з W L V/I Е V S/A D/E D/E Y K/R/T L, та фрагмент, який складається з Е L Т LV/ I /T/M D/E/M Y T/S/H/P/G F R/L L/R/K/N F L Р S.
6. Спосіб згідно з будь-яким з пунктів 1-5, в якому вказаний циклін А2 являє собою варіант послідовності цикліну А2, вибраної з:
(і) функціональних частин нуклеїнової кислоти цикліну А2;
(іі) послідовностей, здатних гібридизуватися з нуклеїновою кислотою/геном цикліну А2 за умов високої жорсткості, що включають 0,1-1 х SSC/0,1 % ваг./об. SDS при 60 °С протягом 1-3 годин;
та
(ііі) похідних та активних фрагментів білка цикліну А2.
7. Спосіб згідно з пунктом 6, в якому варіант цикліну А2 (і)-(іі) є здатним кодувати білок, який включає фрагмент, який складається з W L V/I E V S/A D/E D/E Y K/R/T L, та фрагмент, який складається з Е L Т L V/ I /T/M D/E/M Y T/S/H/P/G F R/L L/R/K/N F L Р S.
8. Спосіб згідно з пунктом 6, в якому варіант цикліну А2 (ііі) включає фрагмент, який складається з W L V/I Е V S/A D/E D/E Y K/R/T L, та фрагмент, який складається з Е L Т L V/ I /T/M D/E/M Y T/S/H/P/G F R/L L/R/K/N F L P S.
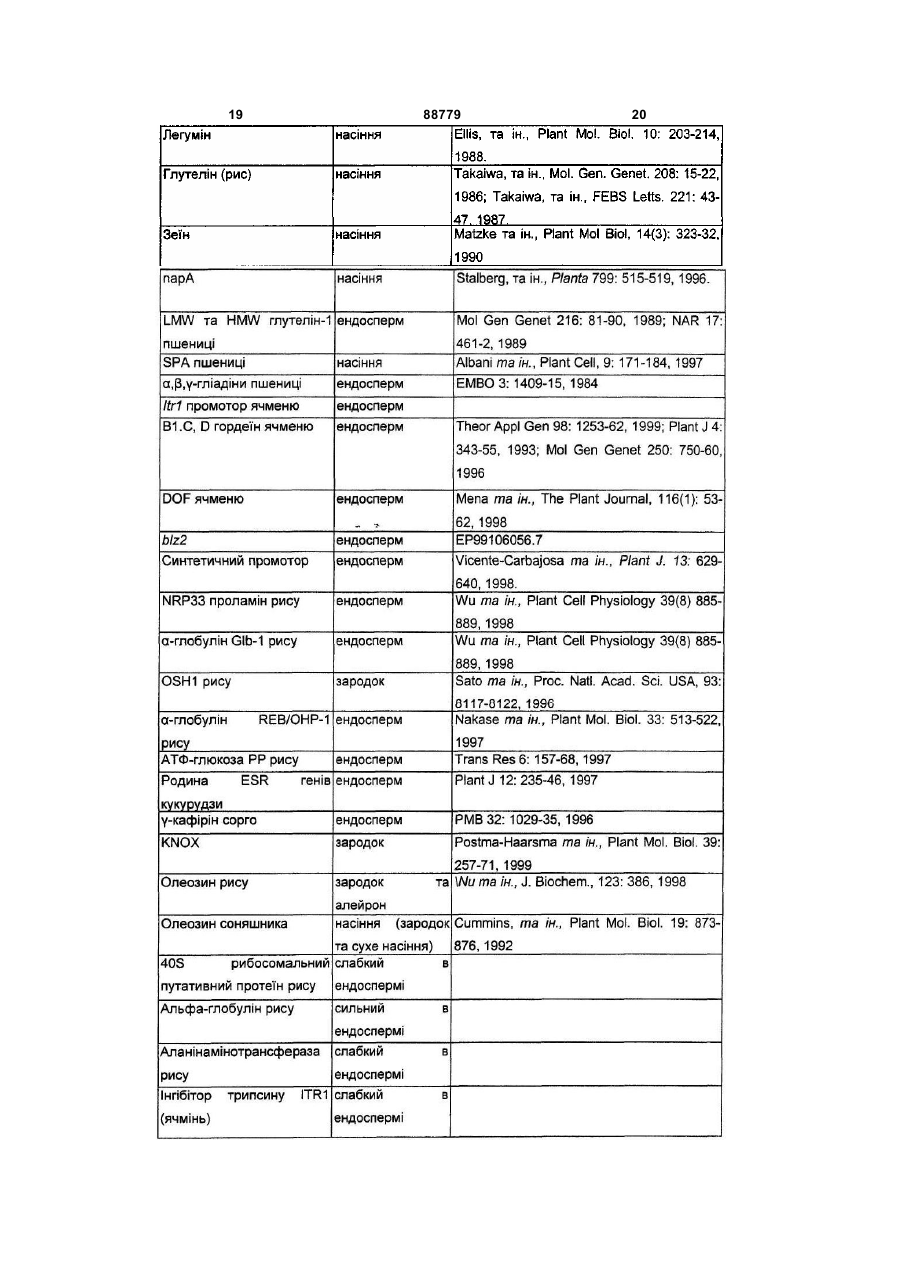
9. Спосіб згідно з будь-яким з пунктів 1-8, в якому промотор, специфічний для насіння, являє собою промотор, активний в ендоспермі.
10. Спосіб згідно з пунктом 9, в якому вказаний промотор являє собою проламіновий промотор.
11. Спосіб згідно з будь-яким з пунктів 1-10, в якому вказане підвищення врожаю досягається в оптимальних та субоптимальних умовах вирощування.
12. Спосіб згідно з пунктом 11, в якому вказані субоптимальні умови вирощування включають умови абіотичного стресу, такі, як сольовий стрес.
13. Спосіб згідно з будь-яким з пунктів 1-12, в якому вказана рослина є вибраною з рису, кукурудзи, пшениці, соєвих бобів, соняшнику, каноли, цукрової тростини, люцерни, проса, ячменю, рапсу, сорго та бавовнику.
14. Спосіб одержання трансгенних рослин, які мають підвищений врожай у порівнянні з контрольними рослинами, який включає введення у рослину нуклеїнової кислоти цикліну А2, що оперативно зв'язана з промотором, специфічним для насіння.
15. Конструкція, що включає:
(і) нуклеїнову кислоту, яка кодує білок, що включає фрагмент, який складається з W L V/I Е V S/A D/E D/E Y K/R/T L, та необов'язково, у доповнення, фрагмент, який складається з Е L Т L V/l/T/M D/E/M Y T/S/H/P/G F R/L L/R/K/N F L P S;
(іі) промотор, специфічний для насіння, та необов'язково
(ііі) послідовність термінації транскрипції.
16. Конструкція згідно з пунктом 15, в якій вказаний промотор, специфічний для насіння, являє собою промотор, активний в ендоспермі.
17. Конструкція згідно з пунктом 16, в якій вказаний промотор являє собою проламіновий промотор.
18. Рослина, яка експресує циклін А2 під контролем промотора, специфічного для насіння, де вказаний циклін А2 включає фрагмент, який складається з W L V/I Е V S/A D/E D/E Y K/R/T L, та необов'язково, у доповнення, фрагмент, який складається з Е L Т L V/ I /T/M D/E/M Y T/S/H/P/G F R/L L/R/K/N F L Р S, при цьому рослини мають підвищений врожай у порівнянні з відповідними рослинами дикого типу та у порівнянні з трансгенними рослинами, які конститутивно експресують циклін А2.
19. Рослина згідно з пунктом 18, в якій вказаний промотор, специфічний для насіння, являє собою промотор, активний в ендоспермі.
20. Рослина згідно з пунктом 19, в якій вказаний промотор являє собою проламіновий промотор.
Текст
1. Спосіб підвищення врожаю рослин, який включає введення у рослину нуклеїнової кислоти цикліну А2, яка кодує білок цикліну А2, при цьому нуклеїнова кислота цикліну А2 є оперативно зв'язаною з промотором, специфічним для насіння. 2. Спосіб згідно з пунктом 1, в якому вказаний врожай рослини є вибраним з однієї або більше наступних характеристик: підвищеної ваги насіння, 2 (19) 1 3 88779 4 10. Спосіб згідно з пунктом 9, в якому вказаний (іі) промотор, специфічний для насіння, та необопромотор являє собою проламіновий промотор. в'язково 11. Спосіб згідно з будь-яким з пунктів 1-10, в яко(ііі) послідовність термінації транскрипції. му вказане підвищення врожаю досягається в оп16. Конструкція згідно з пунктом 15, в якій вказаний тимальних та субоптимальних умовах вирощуванпромотор, специфічний для насіння, являє собою ня. промотор, активний в ендоспермі. 12. Спосіб згідно з пунктом 11, в якому вказані 17. Конструкція згідно з пунктом 16, в якій вказаний субоптимальні умови вирощування включають промотор являє собою проламіновий промотор. умови абіотичного стресу, такі, як сольовий стрес. 18. Рослина, яка експресує циклін А2 під контро13. Спосіб згідно з будь-яким з пунктів 1-12, в яколем промотора, специфічного для насіння, де вкаму вказана рослина є вибраною з рису, кукурудзи, заний циклін А2 включає фрагмент, який складапшениці, соєвих бобів, соняшнику, каноли, цукроється з W L V/I Е V S/A D/E D/E Y K/R/T L, та вої тростини, люцерни, проса, ячменю, рапсу, сорнеобов'язково, у доповнення, фрагмент, який го та бавовнику. складається з Е L Т L V/ I /T/M D/E/M Y T/S/H/P/G F 14. Спосіб одержання трансгенних рослин, які маR/L L/R/K/N F L Р S, при цьому рослини мають ють підвищений врожай у порівнянні з контрольпідвищений врожай у порівнянні з відповідними ними рослинами, який включає введення у рослирослинами дикого типу та у порівнянні з трансгенну нуклеїнової кислоти цикліну А2, що оперативно ними рослинами, які конститутивно експресують зв'язана з промотором, специфічним для насіння. циклін А2. 15. Конструкція, що включає: 19. Рослина згідно з пунктом 18, в якій вказаний (і) нуклеїнову кислоту, яка кодує білок, що включає промотор, специфічний для насіння, являє собою фрагмент, який складається з W L V/I Е V S/A D/E промотор, активний в ендоспермі. D/E Y K/R/T L, та необов'язково, у доповнення, 20. Рослина згідно з пунктом 19, в якій вказаний фрагмент, який складається з Е L Т L V/l/T/M промотор являє собою проламіновий промотор. D/E/M Y T/S/H/P/G F R/L L/R/K/N F L P S; Даний винахід, в основному, відноситься до області молекулярної біології та стосується способу підвищення врожаю рослин. Зокрема, даний винахід стосується способу підвищення врожаю рослин шляхом введення у рослину нуклеїнової кислоти цикліну А, переважно такої, що кодує білок цикліну А, при цьому вказана нуклеїнова кислота є оперативно зв'язаною з промотором, який є переважним для насіння. Даний винахід також стосується рослин, які мають підвищену експресію нуклеїнової кислоти цикліну А у тканині насіння рослин та/або модульовану активність та/або рівні білка цикліну А у тканині насіння рослин, при цьому рослини мають підвищений врожай у порівнянні з рослинами дикого типу та у порівнянні з відповідними трансгенними рослинами, в яких циклін А конститутивно експресується. Населення світу, що постійно росте, та виснаженість запасів наявних орних земель, придатних для сільського господарства, штовхають сільськогосподарські дослідження до підвищення ефективності сільського господарства. Традиційні засоби для поліпшення сільськогосподарських та садових культур використовують методики селекційного розведення для ідентифікації рослин, які мають бажані характеристики. Проте такі способи селекційного розведення мають деякі недоліки, зокрема, такі, що ці методики типово вимагають великих затрат праці та приводять до одержання рослин, які часто містять гетерогенні генетичні компоненти, які можуть не завжди приводити до одержання бажаної характеристики, яка передається від батьківських рослин. Успіхи молекулярної біології дозволили людству модифікувати зародкову плазму тварин та рослин. Генетична інженерія рослин передбачає проведення ізоляції та маніпуляції з генетичним матеріалом (типово, у формі ДНК або РНК) та подальше введення такого генетичного матеріалу у рослину. Така технологія має здатність забезпечення сільськогосподарських культур або рослин, які володіють різноманітними поліпшеними економічними, агрономічними або садовими ознаками. Ознака, яка представляє особливий економічний інтерес, являє собою врожай. Врожай звичайно визначають як одержання здатної до вимірювання економічної оцінки сільськогосподарської культури. Він може бути визначений в значеннях кількості та/або якості. Врожай безпосередньо залежить від декількох факторів, наприклад, кількості та розміру органів, будови рослини (наприклад, кількість гілок), продукції насіння, тощо. Розвиток кореневої системи, споживання поживних речовин та толерантність до стресу також є важливими факторами у визначенні врожаю. Врожай сільськогосподарської культури може бути підвищений шляхом оптимізації одного зі згаданих вище факторів, що може бути зроблено шляхом модифікації спадкових ростових механізмів рослини. Спадкові ростові механізми рослини здійснюються за допомогою високо упорядкованої послідовності подій, які разом є відомими як «клітинний цикл». Проходження через клітинний цикл є фундаментальним для росту та розвитку усіх багатоклітинних організмів та є ключовим для клітинної проліферації. Основні компоненти клітинного циклу є високо консервативними у дріжджах, ссавців та рослин. Клітинний цикл типово розділяється на наступні послідовні фази: G0-G1-S-G2-Μ. Реплікація ДНК або синтез звичайно здійснюють під час S фази («S» означає синтез ДНК), а мітотична сегрегація хромосом відбувається під час фази Μ («Μ» означає мітоз) з проміжними «геп» фазами, G1 (під час якої клітини ростуть перед реплікацією ДНК) та G2 (період після реплікації ДНК, під час якого клітина готується до ділення). Клітинний поділ заве 5 88779 6 умов навколишнього середовища (таких, як посуршується після цитокінезу, останнього етапу Мха, солі, температура, відсутність елементів живфази. Про клітини, що вийшли з клітинного циклу лення), підвищена толерантність до рослинних та стали такими, що знаходяться у стані спокою, патогенів, що порушують клітинний цикл, або мокажуть, що вони знаходяться у фазі G0. Клітини у цій фазі можуть бути стимульовані для повторного жуть служити як мішені для поліпшення ідентифікації інгібіторів або активаторів ССР, які можуть вступу клітинного циклу у фазу G1. Буква «G» в G1, G2 та G0 означають «геп». Завершення пробути корисними як гербіциди або медіатори росту цесу клітинного циклу дозволяє кожній дочірній рослин. Врожай може бути підвищений багатьма шляклітині під час клітинного поділу одержати повну копію батьківського геному. хами, деякі з яких є несподіваними. Наприклад, основний фактор, що зробив свій внесок у підвиКлітинний поділ контролюється двома принциповими подіями клітинного циклу, зокрема, синтещення врожаю пшениці та рису в 1960-их роках (так звана зелена революція), представляв собою зом ДНК та ініціацією мітозу. Кожний перехід до кожної з цих ключових подій контролюється контзниження висоти рослин (Sakamotota Matsuoka, Current Opinion in Biotechnology 2004, 15: 144-147). рольною точкою, представленою специфічними білковими комплексами (втягнені у ДНК реплікацію При використанні великих кількостей азотних добрив традиційні сорти того часу виростали надлишта поділ). Експресія генів, необхідних для синтезу ково високими та падали, що приводило до значДНК на межі G1/S регулюється E2F родиною транскрипційних факторів у клітинах ссавців та них втрат врожаю. На противагу до цього, завдяки своєму невисокому росту напівкарликові сорти рослин (La Thangue, 1994; Muller та ін., 2001; De Veylder та ін., 2002). Вступ у клітинний цикл регузеленої революції були стійкими, що приводило до підвищення врожаю в два рази. люється/запускається за допомогою E2F/Rb комплексу, що інтегрує сигнали та активує транскрипЗараз було несподівано виявлено, що врожай рослин може бути підвищений шляхом введення у цію генів клітини. Перехід між двома фазами рослину нуклеїнової кислоти цикліну А, переважно клітинного циклу і, таким чином, проходження четакої, що кодує білок цикліну А, при цьому нуклеїрез клітинний цикл, управляється шляхом утвонова кислота цикліну А є оперативно зв'язаною з рення та активації різних гетеродимерних серин/треонін протеїнкіназ, що звичайно промотором, переважним для насіння. Експресія називаються циклінзалежними кіназами (CDK). А нуклеїнової кислоти цикліну А під контролем проНеобхідною умовою для активності цих кіназ є мотора, переважного для насіння, приводить до фізична асоціація зі специфічним цикліном, синбільшого врожаю, ніж той, що одержують при ексхронізація активації сильно залежить від експресії пресії цикліну А, який конститутивно експресується у рослині. цикліну. Зв'язування цикліну індукує конформаційТаким чином, згідно з одним втіленням даного ні зміни в N-термінальній частині сполучної CDKта винаходу забезпечується спосіб для підвищення робить свій внесок у локалізацію та субстратну врожаю рослин, який включає введення у рослину специфічність комплексу. Мономерні CDK активуються, коли вони зв'язуються з циклінами і, таким нуклеїнової кислоти цикліну А, переважно такої, чином, володіють кіназною активністю. Рівні білка що кодує білок цикліну А, при цьому нуклеїнова цикліну коливаються протягом клітинного циклу і, кислота цикліну А є оперативно зв'язаною з протаким чином, представляють собою основний фамотором, бажаним для насіння. ктор у визначенні часу активації. Періодична актиЗдійснення способу згідно з даним винаходом вація цих комплексів, що містять цикліни та CDK, приводить до підвищеного врожаю рослини. Термін «підвищений врожай», як визначено в даній під час клітинного циклу опосередковує часову заявці, охоплює збільшення біомаси (ваги) однієї регуляцію переходів на різні фази клітинного циклу або більше частин рослини у порівнянні з біома(контрольні точки). Інші фактори, які регулюють сою контрольних рослин. Термін також охоплює активність CDK, включають інгібітори CDK (СКІ або ІСK, ΚIΡ, CIP, INK), CDK активуючі кінази підвищення врожаю насіння, що включає збіль(САK), CDK фосфатази (Cdc25) та CDK субодинишення біомаси насіння (вага насіння) та/або збіцю (CKS) (Mironov та ін. 1999; Reed 1996). льшення кількості (виповненого) насіння, та/або Були описані різні підкласи циклінів Α-типу збільшення розміру насіння, та/або збільшення Arabidopsis (А1, А2 та A3) (включають 10 циклінів). об'єму насіння у порівнянні з контрольними рослиДва гени А1-типу (CYCA1;1 та CYCA1;2), чотири нами. Збільшення розміру насіння та/або збільгени А2-типу (CYCA2;1, CYCA2;2, CYCA2;3 та шення об'єму насіння може також впливати на CYCA2;4) та чотири гени А3-типу (CYCA3;1, склад насіння. Збільшення врожаю насіння може CYCA3;2, CYCA3;3 та CYCA3;4) були показані у бути наслідком збільшення кількості та/або розміVanderpoele та ін. (The Plant Cell, том. 14, 903-916, ру квітів. Збільшення врожаю може також підвиквітень, 2002). щувати коефіцієнт збору врожаю, який виражаєтьМіжнародна заявка WO 01/85946 описує декіся як співвідношення врожаю частин рослин, які лька білків клітинного циклу, у тому числі цикліни збираються, таких, як насіння, до загальної біомаА. Згадується, що білки клітинного циклу можуть си. Збільшення врожаю може також передбачати використовуватися у сільському господарстві для збільшення ваги тисячі зернин (TKW), яку екстраполіпшення ростових характеристик рослини, таполюють з підрахованої кількості виповненого наких, як швидкість росту або розмір специфічних сіння та його загальної ваги. тканин або органів, будова або морфологія рослиБеручи кукурудзу як приклад, можна сказати, ни, підвищений врожай сільськогосподарської що підвищення врожаю може виявлятися як одна культури, підвищена толерантність до стресових або більше наступних подій: збільшення кількості 7 88779 8 рослин на гектар або акр, збільшення кількості Оскільки трансгенні рослини згідно з даним колосків на рослину, збільшення кількості рядів винаходом мають підвищений врожай, ймовірно, зернин, збільшення кількості зернин у ряді, вага що ці рослини демонструють підвищену швидкість зернини, вага тисячі зернин, довжина/діаметр коросту (під час, принаймні, частини їх життєвого лосу, серед інших. Беручи рис як приклад, можна циклу) у порівнянні зі швидкістю росту контрольних сказати, що підвищення врожаю може проявлятирослин на відповідному етапі їх життєвого циклу. ся як збільшення однієї або більше наступних Підвищена швидкість росту може бути специфічознак: кількість рослин на гектар або акр, кількість ною для однієї або більше частин рослини (вклюволотей на рослину, кількість вторинних колосків чаючи насіння) або може стосуватися суттєво усієї на волоть, кількість квітів на волоті, збільшення рослини. У випадку насіння, особливо такого зершвидкості виповнення зернин, збільшення ваги нових культур, зрілість насіння може бути пов'язатисячі насінин, серед інших. на з вмістом вологи у цьому насінні, що є інтактВрожай може бути додатково підвищений або ним на рослині. Вміст вологи, який дає свідчення врожай може бути додатково оцінений у гібридних зрілості насіння, буде також, таким чином, давати рослинах. Сільськогосподарська культура, така, як вказівку на швидкість росту насіння у порівнянні з кукурудза, є типово комерціалізованою як гібрид. контрольними рослинами. Спеціаліст в даній галуМетою польової селекції сільськогосподарської зі для даних видів рослин буде добре знати вміст культури є поєднання різноманітних бажаних влавологи, що є показовим для готовності насіння до стивостей в одному сорті або гібриді. Селекцію збору. Вміст вологи може бути виміряний при виздійснюють за допомогою різноманітних методик, користанні відомих методик. що користуються перевагою способу рослинного Крім того, підвищення швидкості росту може запилення (самозапилення, як у випадку рису, або мати місце на одній або більше стадіях циклу житперехресне запилення, як у випадку кукурудзи). тя рослини або під час усього циклу життя рослиСелекція зернових культур часто втягує етапи сани. Підвищена швидкість росту під час ранніх стамозапилення та перехресного запилення. Беручи дій життєвого циклу рослини відображає кукурудзи як приклад, можна сказати, що одерпідвищену енергію. жання нових сортів часто передбачає розвиток, Підвищення швидкості росту може змінювати селекцію та одержання інбредних батьківських цикл визрівання врожаю рослини, що дозволяє ліній, які послідовно використовуються для одерпізніше висівати рослини та/або збирати їх пізніше, жання гібридної кукурудзи з певними бажаними ніж це є можливим в іншому випадку. Термін «цикл характеристиками. Тому розробка гібридів кукурувизрівання врожаю», як визначено в даній заявці, дзи вимагає розвитку гомозиготних інбредних ліє взятим для визначення часу між посівом та збоній, схрещування цих ліній та оцінки результатів ром рослини. Якщо швидкість росту є достатньо таких схрещувань (гібрид). Введення бажаних хапідвищеною, то це може дозволяти проводити рактеристик може потім проводитися за допомопосів додаткового насіння тих самих видів рослин гою схрещування (генетичне введення) або шля(наприклад, посів та збір рослин рису, після чого хом молекулярного введення за допомогою проводять посів та збір додаткових рослин рису, і методик трансформації. Для визначення польової усе це здійснюють в межах одного традиційного поведінки продукту оцінка нової зернової культури періоду вирощування). Подібно до цього, якщо може бути здійснена на гомогенній популяції гомошвидкість росту є достатньо підвищеною, то це зиготних інбредних рослин, або на гібриді, одернадає можливість посіву додаткового насіння різжаному при схрещуванні двох гомозиготних інбрених інших видів рослин (наприклад, посів та збір дних ліній. Згадані вище методики є добре рослин рису, після чого, наприклад, проводять відомими у рівні техніки. посів та необов'язковий збір сої, картоплі або Зокрема, підвищений врожай виявляється як будь-якої іншої прийнятної рослини). Збір з того одна або більше наступних подій: підвищена вага самого кореневища може у випадку деяких рослин насіння, підвищена кількість виповненого насіння, проходити в додатковий період року. Можливість підвищена кількість насіння, збільшений розмір змінювати цикл визрівання врожаю рослини може насіння, підвищений індекс збору врожаю, підвиприводити до підвищення річної продукції біомаси щена вага тисячі насінин та модифікований склад на акр (завдяки збільшенню числа разів (на рік), насіння, кожна характеристика береться у порівпротягом яких окрема рослина може вирощуватинянні з контрольними рослинами. Таким чином, ся та збиратися). Підвищення швидкості росту згідно з даним винаходом забезпечується спосіб може також дозволити культивувати трансгенні підвищення врожаю рослини, в якому підвищений рослини у більш широкій географічній зоні, ніж їх врожай рослини є вибраним з однієї або більше аналоги дикого типу, оскільки територіальне обхарактеристик: підвищена вага насіння, підвищена меження для вирощування сільськогосподарської кількість виповненого насіння, підвищена кількість культури часто визначається шкідливими умовами насіння, збільшений розмір насіння, підвищений навколишнього середовища, або на час висаджуіндекс збору врожаю, підвищена вага тисячі насівання (ранній період проведення цієї операції), нин та модифікований склад насіння, кожна харакабо на час збору (пізній період проведення цієї теристика береться у порівнянні з контрольними операції). Такі шкідливі умови можуть бути усунені, рослинами, при цьому вказаний спосіб включає якщо цикл визрівання врожаю є вкороченим. введення у рослину нуклеїнової кислоти цикліну А, Швидкість росту може бути визначена шляхом переважно такої, що кодує білок цикліну А, при виведення різноманітних параметрів з кривих росцьому нуклеїнова кислота цикліну А є оперативно ту, які побудовані під час проведення ростових зв'язаною з промотором, переважним для насіння. експериментів, такими параметрами можуть бути: 9 88779 10 T-Mid (час, який займає у рослини досягнення 50% підкреслений вище) дозволяє ідентифікувати цю свого максимального розміру) та Т-90 (час, який послідовність як таку цикліну типу А2, більш перезаймає у рослини досягнення 90% свого максимаважно, ніж будь-якого іншого цикліну А. Тире (-) льного розміру), серед інших. вище представляє собою амінокислотні залишки, де одне тире відповідає амінокислотному залишку Згідно з даним винаходом здійснення способів згідно з винаходом приводить до одержання росу відповідному положенні у фрагменті 2. Термін «нуклеїнова кислота цикліну А2;2», як лин, які мають модифіковану швидкість росту. Тавизначено в даній заявці, представляє будь-яку ким чином, згідно з даним способом забезпечується спосіб підвищення швидкості росту рослин, де нуклеїнову кислоту цикліну А, що кодує білок, який має, у порядку бажаності, має принаймні, 65%, вказаний спосіб включає введення у рослини нуклеїнової кислоти цикліну А, переважно такої, що 70%, 75%, 80%, 85%, 90% або 95% ідентичності або подібності послідовності до амінокислотної кодує білок цикліну А, при цьому нуклеїнова кислота цикліну А є оперативно зв'язаною з промотопослідовності, що представлена SEQ ID NO: 2. Термін «амінокислотна послідовність цикліну ром, переважним для насіння. Підвищення врожаю та/або швидкості росту А» або «білок цикліну А», як визначено в даній заявці, узятий для розуміння амінокислотної потакож охоплює кращу поведінку рослини при нестресових умовах, а також при стресових умовах, слідовності, яка у своїй нативній формі включає у порівнянні з контрольними рослинами. Рослини фрагмент 1, який представлений як: W L V/I Ε V S/A D/E D/E Y K/R/T L (фрагмент 1), де символ (/) типово відповідають на вплив умов стресу шляхом більш повільного росту. В умовах жорсткого стресу означає «або», тобто «V/I» означає V або І. Присутність фрагменту 1 в амінокислотній послідовності рослина може повністю припинити ріст. Слабкий стрес, з Іншого боку, визначений у даній заявці як дозволяє ідентифікувати цю послідовність як таку цикліну А, більш переважно, ніж будь-якого іншого будь-який стрес, при якому рослина не припиняє ріст повністю. Завдяки успіхам сільського госпотипу цикліну. дарства (іригація, використання добрив, обробка Термін «амінокислотна послідовність цикліну пестицидами) тяжкі стреси не часто зустрічаються А» або «білок цикліну А», як визначено в даній серед культивованих сільськогосподарських росзаявці, представляє собою будь-яку амінокислотну лин. Як наслідок, порушення росту, індуковані послідовність, яка у своїй нативній формі включає слабким стресом, часто представляють собою фрагмент 1, як визначено вище, та додатково небажану характеристику для сільського госпофрагмент 2, який представлений як: Ε L Τ L V/l/T/M D/E/M Υ T/S/H/P/G F R/L L/R/K/N F L P S (фрагмент дарства. Слабкі стреси Е типовими стресами, які 2), де присутність, принаймні, двох ідентифіковаможуть впливати на рослини. Такі стреси можуть них залишків (-Т-----F--F---) (та підкреслений вище) бути щоденними біотичними та/або абіотичними дозволяє ідентифікувати цю послідовність як таку стресами (стресами навколишнього середовища), цикліну типу А2, більш переважно, ніж будь-якого яким піддається рослина. Типові абіотичні стреси іншого цикліну А. Тире (-) вище представляє соабо стреси навколишнього середовища включабою амінокислотні залишки, де одне тире відповіють температурні стреси, спричинені нетипово дає амінокислотному залишку у відповідному повисокими або низькими температураложенні у фрагменті 2. ми/температурами заморожуванння, сольовий Термін «нуклеїнова кислота цикліну А2;2», як стрес; водний стрес (недостатність або надлишок визначено в даній заявці, представляє будь-яку води). Абіотичні стреси можуть також бути спричинуклеїнову кислоту цикліну А, що кодує білок, який нені хімічними речовинами. Біотичні стреси предмає, у порядку бажаності, принаймні, 65%, 70%, ставляють совою типово такі стреси, які спричине75%, 80%, 85%, 90% або 95% ідентичності або ні патогенами, такими, як бактерії, віруси, гриби та подібності послідовності до амінокислотної послікомахи. довності, що представлена SEQ ID NO: 2. В одному втіленні даного винаходу циклін А, що вводиться у рослину, представляє собою цикНуклеїнова кислота цикліну А, що вводиться у рослину, може мати походження від будь-якого лін тилу А2, переважно циклін А2;2 джерела, за умови, що ця нуклеїнова кислота, коТермін «нуклеїнова кислота цикліну А», як визначено в даній заявці, узятий для розуміння нукли вона понадекпресується у тканині насіння рослин, приводить до підвищеного врожаю рослин. леїнової кислоти, яка кодує білок, що у своїй натиНуклеїнова кислота цикліну А, що вводиться у ровній формі включає фрагмент 1, який слину, може бути ізольована з мікробного джерепредставлений як. W L V/l Е V S/A D/E D/E Y K/R/T L (фрагмент 1), де символ (/) означає «або», тобто ла, такого, як бактерії, дріжджі або грибки, або з рослини, водорості або тваринного джерела. Ця «V/I» означає V або І. Присутність фрагменту 1 в нуклеїнова кислота може бути суттєво модифікоамінокислотній послідовності дозволяє ідентифівана, виходячи з її нативної форми у композиції кувати цю послідовність як таку цикліну А, більш переважно, ніж будь-якого іншого типу цикліну. та/або у геномному оточенні, за допомогою спеціальних маніпуляцій людини. Послідовність нуклеїТермін «нуклеїнова кислота цикліну А2», як нової кислоти бажано є гомологічною послідовнісвизначено в даній заявці, представляє собою нуктю нуклеїнової кислоти, тобто, послідовністю леїнову кислоту, яка кодує білок» що у своїй натинуклеїнової кислоти, що одержана з рослини або вній формі включає фрагмент 1, як визначено витого самого виду рослини, або іншого. Послідовще та додатково фрагмент 2, який представлений ність нуклеїнової кислоти може бути ізольована з як: Е L Т L V/I/T/M D/E/M Y T/S/H/P/G F R/L L/R/K/N однодольних або дводольних видів рослин, бажаF L Р S (фрагмент 2), де присутність, принаймні, но з родини Brassicaceae, більш бажано з двох ідентифікованих залишків (-Т-----F--F---) (та 11 88779 12 Arabidopsis thaliana. Найбільш бажано, коли циклін ним, щоб частина представляла частину нуклеїноА представляє собою циклін типу А2, такий, як вої кислоти цикліну А, як представлено послідовніциклін А2;1, А2;2, А2;3 або А2;4. В особливо бажастю SEQ ID NO: 1. ному втіленні циклін А2;2 представлений послідоТаким чином, згідно з іншим втіленням даного вністю SEQ ID NO: 1 та SEQ ID NO: 2. винаходу забезпечується спосіб підвищення вроНезважаючи на те, що даний винахід було жаю рослин, який включає введення у рослину проілюстровано з нуклеїновою кислотою, предстафункціональної частини нуклеїнової кислоти циквленою SEQ ID NO: 1, та амінокислотною послідоліну А, бажано такої, як SEQ ID NO: 1, при цьому вністю, представленою SEQ ID NO: 2, способи функціональна частина є оперативно зв'язаною з можуть також бути здійснені при використанні вапромотором, переважним для насіння. ріанту амінокислотної послідовності цикліну А та Послідовності, здатні до гібридизації з нуклеїваріанту послідовності нуклеїнової кислоти цикліну новою кислотою цикліну А, такою, як представлено А. SEQ ID NO: 1, можуть також бути корисними у Варіанти послідовностей нуклеїнової кислоти здійсненні способів згідно з даним винаходом. Тета варіанти амінокислотних послідовностей, які є рмін «гібридизація», як визначено в даній заявці, корисними у практиці здійснення способів згідно з представляє собою процес, в якому суттєво гомовинаходом, включають: логічні комплементарні нуклеотидні послідовності (і) Функціональні частини нуклеїнової кислоти відпалюються одна від іншої. Процес гібридизації цикліну А; може проходити повністю у розчині, тобто коли (іі) Послідовності, здатні гібридизуватися з нуобидві комплементарні нуклеїнові кислоти перебуклеїновою кислотою/геном цикліну А вають у розчині. Засоби у молекулярній біології, (ііі) Альтернативні варіанти сплайсингу нуклеїякі базуються на такому процесі, включають полінової кислоти/гену цикліну А; меразну ланцюгову реакцію (ПЛР, та усі способи, (iv) Алельні варіанти сплайсингу нуклеїнової що грунтуються на ній), субтрактивну гібридизакислоти/гену цикліну А; цію, рендомне подоження праймера, картування (ν) Варіанти, одержані завдяки виродженню за допомогою нуклеази S1, подовження праймера, генетичного коду; та зворотну транскрипцію, синтез кДНК, диференціа(vi) Гомологи, похідні та активні фрагменти більне дисплейне визначення послідовності РНК та лка цикліну А. ДНК. Процес гібридизації може, крім того, здійснюТермін «нуклеїнова кислота», як такий, що виватися з однією з комплементарних нуклеїнових користовується в даній заявці охоплює комплемекислот, імобілізованих на твердій основі, такій, як нтарні ланцюги та відповідні РНК, ДНК, кДНК та магнітні кульки, кульки Сефарози або будь-які інші геномну ДНК. Нуклеїнова кислота може бути двосмоли. Засоби у молекулярній біології, які ґрунтуланцюговою або одноланцюговою. ються на такому процесі, включають ізоляцію поДля спеціаліста в даній області техніки зрозулі(А+) мРНК. Процес гібридизації може також здійміло, що послідовність ДНК повної довжини цикліснюватися з однією з комплементарних ну А не єнеобхідною умовою для здійснення спонуклеїнових кислот, імобілізованих на твердій оссобів згідно з винаходом, але означає, що нові, такій, як нітроцелюлоза або нейлонова мемфункціональні частини нуклеїнової кислоти цикліну брана, або імобілізованих за допомогою, наприА також можуть використовуватися. Функціональклад, фотолітографії, на, наприклад, носіях на на частина відноситься до частини ДНК, що має основі кремнеземвмісного скла (останні є відомипоходження від або одержана з вихідної (більшої) ми як матриці для нуклеїнової кислоти або мікромолекули ДНК, частина ДНК якої при введенні та матриці, чіпи нуклеїнової кислоти). Засоби у молеекспресії у рослині, забезпечує рослини, який макулярній біології, які ґрунтуються на такому ють підвищений врожай. Частина може включати процесі, включають гелевий аналіз блотів РНК та багато генів, які містять або не містять додаткові ДНК, гібридизацію колоній мікроорганізмів, бляшконтрольні елементи, або може містити тільки покову гібридизацію, гібридизацію in situ та гібридислідовності спейсера. Частини, прийнятні для визацію мікроматриць. Для того, щоб провести гібкористання у способах згідно з даним винаходом, ридизацію, молекули нуклеїнової кислоти можуть бути легко визначені при використанні звизвичайно піддають термальній або хімічній деначайних методик. Наприклад, у послідовності нуктурації для розплавлення подвійного ланцюга на леїнової кислоти SEQ ID NO: 1 можуть бути зробдва одиничні ланцюги та/або видаляють шпильки лені одна або більше делецій та/або відрізання або інші вторинні структури з молекул одноланцючастин без впливу на її здатність приймати участь гових нуклеїнових кислот. На жорсткість гібридиу способах згідно з даним винаходом. Частини, зації впливають умови, такі, як температура, конприйнятні для використання у способах згідно з центрація солі та склад буфера для гібридизації. винаходом, можуть бути легко визначені при викоУмови високої жорсткості включають високу темристанні традиційних методик, таких, як шляхом пературу та/або низьку концентрацію солі (солі аналізу активності цикліну А та/або за допомогою включають NaCI та Na3-цитрат) та/або включення способів, описаних у розділі Приклади, шляхом формаміду у гібридизаційний буфер та/або знипростої заміни послідовності, що використовується ження концентрації сполук, таких, як SDS (детеру даному прикладі частиною, яка аналізується стогент) у гібридизаційному буфері та/або виключенсовно її функціональності. Бажана частина для ня сполук, таких, як декстрансульфат або застосування у способах згідно з винаходом є здаполіетиленгліколь (сприяння молекулярному ущітною кодувати білок, який включає фрагмент 1 та льненню) з гібридизаційного буфера. Традиційні бажано додатково фрагмент 2. Крім того, є бажаумови гібридизації є описаними, наприклад, у 13 88779 14 Sambrook (2001) Молекулярне клонування: лабофункціональних сегментів білка, який кодується раторний практикум, 3-є видання, Cold Spring нуклеїновою кислотою. Такі сплайсингові варіанти Harbor Laboratory Press, CSH, New York, але спеможуть бути виявлені у природі або можуть бути ціаліст у даній галузі зможе оцінити, що багаточистворені людиною. Способи для одержання таких сленні умови гібридизації можуть бути змодельосплайсингових варіантів є добре відомими в облавані при використанні функції відомої або сті техніки. Бажані сплайсингові варіанти кодують очікуваної гомології та/або довжини послідовності білок, який включає фрагмент 1 та бажано додатнуклеїнової кислоти. Умови гібридизації низької ково фрагмент 2. Сплайсингові варіанти нуклеїножорсткості є особливо бажаними для ізоляції гетевої кислоти цикліну А, прийнятні для застосування рологічних нуклеїнових кислот з послідовностями у способах згідно з даним винаходом, можуть бути ДНК згідно з винаходом, як визначено вище. Прилегко визначені при використанні традиційних споклад умов низької жорсткості представляє собою собів, таких, як аналіз на активність цикліну А 4-6xSSC/0,1-0,5% ваг./об. SDS при 37-45°С протята/або при використанні наступних способів, опигом 2-3 годин. В залежності від джерела та концесаних у розділі Приклади, шляхом простої заміни нтрації нуклеїнової кислоти, що втягнені у гібридипослідовності, що використовується у цих приклазацію, можуть використовуватися альтернативні дах, сплайсинговим варіантом, який досліджується умови жорсткості такі як середні умови жорсткості. на функціональність. Приклади середніх умов жорсткості включають 1Таким чином, згідно з іншим втіленням даного 4xSSC/0,25% ваг./об. SDS при 45°С протягом 2-3 винаходу забезпечується спосіб для підвищення годин. Приклад умов високої жорсткості включає врожаю рослин, який включає введення у рослину 0,1-1xSSC/0,1% ваг./об. SDS при 60°С протягом 1сплайсингового варіанту нуклеїнової кислоти цик3 годин. Спеціаліст у даній галузі зможе оцінити ліну А, бажано сплайсингового варіанту послідоврізноманітні параметри, які можуть змінюватися ності нуклеїнової кислоти, як представлено SEQ ID під час гібридизації та промивання, та які будуть NO: 1, де сплайсинговий варіант є оперативно або підтримувати або змінювати умови жорсткості. зв'язаним з промотором, переважним для насіння. Елементи, які роблять свій внесок у гетерологічЄ бажаним, коли способи згідно з даним винаність, включають алелізм, виродженість генетичходом можуть також здійснюватися на практиці ного коду та відмінності у бажаності використання при використанні алельних варіантів нуклеїнової кодону. кислоти цикліну А, бажано алельних варіантів нукБажані послідовності, здатні до гібридизації з леїнової кислоти цикліну А, як представлено SEQ нуклеїновою кислотою цикліну А, такою, як предID NO: 1. Алельні варіанти існують у природі, та ставлено SEQ ID NO: 1, представляють собою такі охоплені даним винаходом способи використовупослідовності, які гібридизуються та які здатні коють ці природні алелі. Алельні варіанти охоплють дувати білок, який включає фрагмент 1 та бажано поліморфізм одного нуклеотиду (SNP), а також додатково фрагмент 2. Послідовності, що гібридиполіморфізм невеликої інсерції/делеції (INDEL). зуються, які є прийнятними для застосування у Розмір INDEL є звичайно меншим, ніж 100п.о. SNP способах згідно з даним винаходом, можуть бути та INDEL утворюють більші набори варіантів полегко визначені при використанні традиційних спослідовності існуючих в природі поліморфних штасобів, таких, як аналіз на активність цикліну А мів більшості організмів. Бажані алельні варіанти та/або наступні способи, які описані у розділі Прикодують білок, який включає фрагмент 1 та бажаклади, шляхом простої заміни послідовності, що но додатково фрагмент 2. Алельні варіанти нуклевикористовується у даному прикладі, послідовнісїнової кислоти цикліну А, прийнятні для використю, що гібридизується та піддається аналізу на тання у способах згідно з даним винаходом, функціональність. можуть бути легко визначені при використанні звиТаким чином, згідно з іншим втіленням даного чайних методик, описаних у розділі Приклади, винаходу забезпечується спосіб підвищення врошляхом простої заміни послідовності, що викорисжаю рослин, який включає введення у рослину товується у цих прикладах, алельним варіантом, нуклеїнової кислоти, здатної гібридизуватися з який досліджується на функціональність. нуклеїновою кислотою цикліну А, як визначено в Таким чином, згідно з іншим аспектом даного даній заявці вище, бажано з нуклеїновою кисловинаходу забезпечується спосіб для підвищення тою цикліну А, як представлено у SEQ ID NO: 1, врожаю рослин, який включає введення у рослину при цьому послідовність, що гібридизується, є алельного варіанту нуклеїнової кислоти цикліну А, оперативно зв'язаною з промотором, який є перебажано алельного варіанту послідовності нуклеїважним для насіння. нової кислоти, як представлено SEQ ID NO: 1, де Способи згідно з даним винаходом можуть таалельний варіант є оперативно зв'язаним з промокож використовуватися при застосуванні альтертором, переважним для насіння. нативного сплайсингового варіанту нуклеїнової Приклади варіантних амінокислотних послідокислоти цикліну А, так, як представлено послідоввностей цикліну А включають гомологи, похідні та ністю SEQ ID NO: 1. Термін «альтернативні сплайактивні фрагменти цикліну А, представлені SEQ ID сингові варіанти», як використовується в даній NO: 2. заявці, охоплює варіанти послідовностей нуклеї«Гомологи» білка цикліну А охоплюють пептинової кислоти, в яких вибрані інтрони та/або екзоди, олігопептиди, поліпептиди, білки та ферменти, ни були вирізані, заміщені або додані. Такі варіанякі мають амінокислотні заміни, делеції та/або інти будуть такими, в яких біологічна активність серції стосовно немодифікованого даного білка та білка залишається незмінною, та які можуть бути мають подібну біологічну та функціональну активодержані за допомогою селективного збереження ність, що й немодифікований білок, з якого вони 15 88779 16 одержані. Для одержання таких гомологів амінокиТермін «варіанти білка, одержані шляхом заслоти білка можуть бути замінені іншими амінокиміни» є такими, в яких, принаймні, один залишок в слотами, які мають подібні властивості (такі, як амінокислотній послідовності був видалений, та подібна гідрофобність, гідрофільність, антигенвідмінний залишок було вбудовано на його місце. Амінокислотні заміни є типово замінами одного ність, здатність утворювати або розкладати aзалишку, але вони можуть бути кластиризовані в спіральні структури або b-пластиночні структури). залежності від функціонального обмеження поліТаблиці консервативних замін є добре відомими в пептиду, інсерції будуть звичайно лежати у межах області техніки (див. для прикладу Creighton від приблизно 1 до 10 амінокислотних залишків, а (1984) Proteins. W.H. Freemanand Company). Баделеції будуть коливатися від приблизно 1 до 20 жано, коли гомологи мають ідентичність послідовзалишків. Бажано, коли амінокислотні заміни ності (функціональну ідентичність) у порядку зросвключають консервативні амінокислотні заміни. тання, принаймні, 40%, 45%, 50%, 55%, 60%, 65%, Інсерційні варіанти білка є такими, в яких один 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% або більше амінокислотних залишків вбудовані у або 99% або більше до цикліну, що представлений попередньо визначений сайт білка. Інсерції моSEQ ID NO: 2. Гомологи, які мають, принаймні, жуть включати амінотермінальні та/або карбокси40% ідентичності послідовності, охоплюють циклітермінальні злиття, а також інсерції усередині пони А без перекривання будь-якого іншого класу слідовності однієї або декількох амінокислот. цикліну. Взагалі, інсерції усередині амінокислотної посліДві спеціальні форми гомології, ортологи (надовності будуть меншими, ніж аміно- або карбокприклад, копії одного гена у видів, утворених внаситермінальні злиття, та містять приблизно від 1 слідок дивергенції одного виду) та паралоги (гомодо 10 залишків. Приклади аміно- або карбокситерлогія внаслідок дуплікації) є еволюційними мінальних злитих білків включають зв'язувальний поняттями, що використовуються для описування домен або домен активації транскрипційного актиспадкових взаємовідносин генів. Термін «параловатора, як використовується у дріжджовій двогібгічний» відноситься до дуплікацій генів у геномі ридній системі, білки оболонки фагу, (гістидин)6видів, які приводять до утворення паралогічних мітку, мітку на основі глутатіон S-трансферази, генів. Термін «ортологічний» відноситься до гомопротеїн А, протеїн, що зв'язує мальтозу, дигідрологічних генів у різних організмах завдяки спадкофолатредуктазу, Таg-100 епітоп, с-mус епітоп, вим взаємодіям. FLAGÒ-епітоп, lacZ, CMP (кальмодулінзв'язувальОртологічність, наприклад, у однодольних виній пептид), НА епітоп, епітоп протеїна С та VSV дів рослин може бути легко знайдена шляхом епітоп. здійснення так званого реципрокного пошуку блас«Делеційні варіанти» білка характеризуються тів. Це може бути здійснено за допомогою першого видаленням однієї або більше амінокислот з білка. бласту, що втягує бластування послідовності, яка Амінокислотні варіанти білка можуть бути легко представляє інтерес (SEQ ID NO: 1 або SEQ ID одержані при використанні методик для синтезу NO: 2) проти будь-якої послідовності бази даних, пептидів, які добре відомі в області техніки, такі, як такої як відома база даних NCBI, яка може бути пептидний синтез на твердій фазі, та подібні, або знайдена на http://www.ncbi.nlm.nih.gov. Якщо здійметодики на основі рекомбінантної ДНК. Наприснюють пошук на ортологи у рису, то дану посліклад, методики для здійснення мутацій на основі довність необхідно піддавати бластуванню проти, замін у попередньо визначених сайтах в ДНК є наприклад, клонів кДНК повної довжини (28,469) з добре відомими спеціалісту в даній області техніки Oryza sativa Nipponbare, що доступні від NCBI. та включають М13 мутагенез, T7-Gen in vitro мутаBLASTn може використовуватися, виходячи з нукгенез (USB, Clevel та ОН), трансформуючий сайтлеотидів або TBLASTX, якщо виходять з білка, направлений мутагенез (Stratagene, San Diego, який має відсутні стандартні значення (передбаCA), ПЛР-опосередкований сайт-направлений мучення 10, вирівнювання 50). Бластні результати тагенез або інші прописи для сайт-направленого можуть бути відфільтровані. Послідовності повної мутагенезу. довжини, відфільтровані результати або невідфіСпособи для пошуку та ідентифікації гомологів льтровані результатів потім піддають зворотному цикліну знаходяться у сфері знань спеціаліста у бластуванню (другий бласт) проти даної послідовданій області техніки. Способи для порівняння є ності (SEQ ID NO: 1 або 2). Результати першого та добре відомими в області техніки, такі способи другого бластів порівнюють. У випадку великих включають GAP, BESTFIT, BLAST, FASTA та родин, використовують ClustalW, після чого застоTFASTA. GAP використовує алгоритм Нідлмана та совують дерево, що поєднує сусідів для того, що Ванча (J. Mol. ВіоІ. 48: 443-453, 1970) для проведопомогти візуалізувати кластери. Приклади ортодення порівняльного аналізу двох повних послідолопв цикліну А включають послідовності, які задевностей, що максимізує кількість пар та мінімізує поновані під наступними депозитними номерами: кількість генів. Алгоритм BLAST підраховує проортолог рису, задепонований під депозитним ноцент ідентичності послідовності та проводить стамером білка АK106653 (циклін типу А2), ортолог тистичний аналіз на подібність між двома послідорису, задепонований під депозитним номером білвностями. Програмне забезпечення для ка ВАА86628 (циклін типу А2) та ортолог кукурупроведення BLAST аналізу є доступним через Надзи, задепонований під депозитним номером ціональний Центр з біотехнологічної інформації. ААС50013. Термін «похідні» відноситься до пептидів, оліТермін «гомолог», як використовується в даній гопептидів, поліпептидів, білків та ферментів, які заявці, також охоплює паралоги та ортологи білків, можуть включати заміни, делеції або додання ісякі є корисними згідно з даним винаходом. 17 88779 18 нуючих у природі або неприродних амінокислотних гена, який представляє інтерес, так, що промоторзалишків у порівнянні з амінокислотною послідовна послідовність є здатною ініціювати транскрипністю цикліну А, такою, як представлено SEQ ID цію гена, який представляє інтерес. NO: 2. Термін «похідні білка цикліну А» охоплює Вищезгаданими термінами охоплюються попептиди, олігопептиди, поліпептиди, білки та ферслідовності транскрипційних регуляторних елемементи, які можуть включати існуючі в природі, змінтів, що мають походження від класичного еукарінені, глікозильовані, ацильовані або неіснуючі в отичного гену (включаючи ТАТА бокс, який є природі амінокислотні залишки у порівнянні з амінеобхідним для точної ініціації транскрипції, з або нокислотною послідовністю існуючої в природі без послідовності боксу ССААТ) та додаткові реформи поліпептиду. Похідна може також включати гуляторні елементи (тобто розміщені вище актиодну або більше амінокислотних замін у порівнянні вуючі послідовності, енхансери та послідовності, з амінокислотною послідовністю, з якої вона похоякі глушать), що змінюють генну експресію у відподить, наприклад, репортерну молекулу або інший відь на стимули розвитку та/або зовнішні стимули, ліганд, ковалентно або нековалентно, зв'язаний з або специфічним для тканини чином. У вказані амінокислотною послідовністю, такою, як напритерміни також включається послідовність регуляції клад, репортерна молекула, з якою вона зв'язана транскрипції класичного прокаріотичного гену, у для поліпшення її визначення, та неіснуючі в прицьому випадку вона може включати -35 послідовроді амінокислотні залишки стосовно амінокислотність боксу та/або -10 послідовність боксу регуляної послідовності існуючого в природі білка. ції транскрипції. Термін «регуляторний елемент» «Активні фрагменти» білка цикліну А включатакож охоплює синтетичну злиту молекулу або ють, принаймні, фрагмент 1 та бажано додатково похідну, яка забезпечує, активує або сприяє ексфрагмент 2 та зберігають подібну біологічну пресії молекули нуклеїнової кислоти у клітині, тката/або функціональну активність з існуючим у принині або органі. роді білком. Бажано, коли способи згідно з винаходом моРослини трансформують за допомогою вектожуть бути здійснені при використанні промотора, ра, при цьому такий вектор включає послідовність, переважного для насіння. Промотор, переважний яка представляє інтерес (тобто нуклеїнову кислоту для насіння, у контексті даного винаходу предстацикліну А), послідовність якого є оперативно зв'являє собою промотор, який переважно є активним заною з промотором, переважним для насіння. у тканині насіння, але не обов'язково виключно Таким чином, згідно з іншим втіленням даного активний у тканині насіння. Тканина насіння вклювинаходу забезпечується конструкція, яка включає будь-яку частину насіння, включаючи оболонку чає: насіння, алейроновий шар, ендосперм (для одно(і) нуклеїнову кислоту цикліну А; дольних та ендоспермних дводольних), зародок (іі) промотор переважний для насіння; та не(скутелум, епібласт, первинна листова брунька, обов'язково зародковий корінь для однодольних; сім'ядолі, (ііі) послідовність термінації транскрипції. гіпокотиль та зародковий корінь для дводольних). Нуклеїнова кислота цикліну А може бути будьБажаний промотор для здійснення способу згідно якою згаданою вище послідовністю цикліну А, що з винаходом представляє собою такий, що є активключає варіант послідовності цикліну А. Прийнявним в ендоспермі, такий, як альфа-глобуліновий тні промоторі, переважні для насіння, включають промотор для рису, глобуліновий промотор для такі, як визначено в даній заявці нижче. вівса, глутеліновий промотор для рису або пшениТермін «регуляторний елемент», «контрольна ці, blz2, транскрипційний фактор рису RISBZ1. послідовність» та «промотор» усі використовуютьОсобливо бажаним є промотор, активний в ендося в даній заявці почергово та узяті у широкому спермі, при цьому цей промотор є переважно акконтексті для позначення регуляторних послідовтивним під час проростання, такий, наприклад, ностей нуклеїнових кислот, які здатні впливати на представляє собою проламіновий промотор з риекспресію послідовностей, до яких вони ліговані. су. Термін «оперативно зв'язаний», як використовуПриклади прийнятних промоторів для здійсється в даній заявці, відноситься до функціональнення способів згідно з даним винаходом забезпеного сполучення промоторної послідовності та чуються у Таблиці 1, що представлена нижче. 19 88779 20 21 Необов'язково одна або більше послідовностей термінації можуть також використовуватися у конструкції, що вводиться у рослину. Термін «термінатор» охоплює контрольну послідовність, яка представляє собою ДНК послідовність на кінці транскрипційної одиниці, яка посилає сигнал 3'процесингу, поліаденілування первинного транскрипту та термінації транскрипції. Додаткові регуляторні елементи можуть включати енхансери транскрипції та трансляції. Спеціаліст в даній області зможе оцінити послідовності термінації та енхансери, які можуть бути прийнятними для використання при здійсненні винаходу. Такі послідовності будуть відомим або можуть бути легко одержані спеціалістом в даній галузі. Генетичні конструкції згідно з винаходом можуть додатково включати послідовність джерела реплікації, яка є необхідною для підтримання та/або реплікації специфічного типу клітин. Один приклад представляє собою випадок, коли генетична конструкція необхідна для підтримання у бактеріальній клітині як епісомальний генетичний елемент (наприклад, у вигляді молекули плазміди або косміди). Бажані джерела реплікації включають, але не обмежені, f1-оrі та соІЕ1. Генетична конструкція може необов'язково включати селективний маркерний ген. Як використовується в даній заявці, термін «селективний маркерний ген» включає будь-який ген, який забезпечує фенотип у клітини, в якій він експресується, для поліпшення ідентифікації та/або селекції клітин, які були трансфіковані або трансформовані за допомогою конструкції нуклеїнової кислоти згідно з винаходом. Прийнятні маркери можуть бути вибрані з маркерів, які забезпечують стійкість до антибіотиків або гербіцидів, або таких, що вводять нову метаболічну властивість, або які дозволяють проводити візуальну селекцію. Клітини, які містять рекомбінантну ДНК, будуть, таким чином, здатні виживати у присутності концентрацій антибіотика або гербіциду, які вбивають не трансформовані клітини. Приклади селективних маркерних генів включають ген bar, який забезпечує стійкість до гербіциду Basta; ген npt, який надає стійкості до антибіотика канаміцину, ген hpt, який надає стійкості до гігроміцину. Візуальні маркери, такі, як зелений флуоресцентний білок (GFP, Haseloff та ін., 1997), р-глюкуронідаза (GUS) або люцифераза, також можуть використовуватися як селективні маркери. Конструкції, корисні у методах згідно з даним винаходом, можуть бути сконструйовані при використанні технології рекомбінантної ДНК, що добре знайома спеціалістам у даній області. Генні конструкції можуть бути вбудовані у вектори, які можуть бути комерційно доступні, прийнятні для трансформації у рослини та прийнятні для експресії гена, який представляє інтерес, у трансформованих клітинах. 88779 22 Білок цикліну А сам по собі та/або нуклеїнова кислота цикліну А сама по собі можуть бути введені безпосередньо у рослинну клітину або в саму рослину (включаючи введення у тканину, орган або будь-яку іншу частину рослини). Згідно з бажаною характеристикою даного винаходу нуклеїнова кислота переважно вводиться у рослину шляхом трансформації. Термін «трансформація», як згадується в даній заявці, охоплює перенос екзогенного полінуклеотиду у хазяйську клітину, незалежно від способу, який використовується для переносу. Рослинна тканина, здатна до подальшого клонального розмноження, або шляхом органогенезу, або шляхом ембріогенезу, може бути трансформована за допомогою генетичної конструкції згідно з даним винаходом, після чого цілу рослину регенерують з неї. Певна тканина, яку вибирають, буде варіювати в залежності від систем клонального розмноження, доступних для, та найкраще прийнятних для, окремого виду, який піддають трансформації. Приклади тканини, яка представляє собою мішень, включають листові диски, пилок, зародки, сім'ядолі, гіпокотилі, мегагаметофіти, калусну тканину, існуючу меристемну тканину (наприклад, меристему сім'ядолей та меристему гіпокотилю). Полінуклеотид може бути транзієнтно або стабільно введений у хазяйську клітину та може підтримуватися в неінтегрованій формі, наприклад, як плазміда. Альтернативно, він може бути інтегрований у хазяйський геном. Одержана трансформована рослинна клітина може потім використовуватися для регенерації трансформованої рослини за допомогою способу, відомого спеціалістам у даній галузі. Трансформація рослинних видів зараз представляє собою доволі звичайну методику. Переважно, будь-який з деяких способів трансформації може використовуватися для введення гену, який представляє інтерес у прийнятну батьківську клітину. Способи трансформації включають застосування ліпосом, електропорації, хімічних реагентів, що підвищують поглинання вільної ДНК, введення ДНК безпосередньо у рослину, бомбардування частинками, трансформація при використанні вірусів або пилку та мікроін'єкції. Способи можуть бути вибрані з кальцій/поліетиленгліколевого методу для протопластів (Krens, F.A. та ін., 1882, Nature 296, 72-74; Negrutiu І. та ін., June 1987, Plant Mol. Biol. 8, 363-373); електропорації протопластів (Shillito R.D. та ін, 1985 Bio/Technol 3, 1099-1102); мікроін'єкції у рослинний матеріал (Crossway А. та ін., 1986, Mol. Gen Genet 202, 179-185); бомбардування частинками, покритими ДНК або РНК (Klein Т.М. та ін., 1987, Nature 327, 70) інфекції за допомогою (неінтегративної) вірусами, тощо. Бажаний спосіб згідно з даним винаходом представляє собою пропис відповідно до Ніеі та ін. 1994 у випадку трансформації рису. Для трансформації кукурудзи 23 88779 24 способи включають трансформацію тканини кукурослинні клітини. Винахід також охоплює частини рудзи на основі Agrobacterium , що був описаний рослини, які піддаються збору, такі, як, але без раніше в ЕР0604662, ЕР0672752, ЕР0971578, обмеження, насіння, листя, плоди, квіти, культури ЕР0955371, ЕР0558676, тощо. Бажані способи для стебла, стебло, ризоми, коріння, клубні та цибулитрансформації з високою ефективністю представни. ляють собою прописи, представлені у Ishida та ін. Термін «рослина», як такий, що використову(Високоефективна трансформація кукурудзи (Zea ється в даній заявці, охоплює цілі рослини, батьmays L), опосередкована Agrobacterium ківські рослини та потомство рослин та частин tumefaciens. Nat Biotechnol. 1996 Jun; 14(6): 745рослин, включаючи насіння, пагони, стебла, корені 50) та описаний у Frame та ін. (Трансформація (включаючи бульби) та клітини рослин, тканини та зародків кукурудзи, опосередкована Agrobacterium органи. Термін «рослина» також, таким чином, tumefaciens, при використанні стандартної бінарної охоплює суспензійні культури, зародки, меристемвекторної системи. Plant Physiol. 2002 May; 129(1): ні ділянки, калусну тканину, листя, насіння, корені, 13-22). пагони, гаметофіти, спорофіти, пилок та мікроспоУ загальному випадку після трансформації рори. слинні клітини або групи клітин піддають селекції Рослини, які є особливо корисними у способах на присутність одного або більше маркерів, які згідно з винаходом, включають усі рослини, які кодуються здатними до експресії у рослині генами, належать до надродини Viridiplantae, зокрема, які сумісно трансфіковані з геном, що представляє включаючи однодольні та дводольні рослини, інтерес, після чого трансформований матеріал включаючи корм для худоби або корм на основі регенерують у цільну рослину. бобових, оздоблювальні рослини, рослини, що Після переносу ДНК та регенерації путативно забезпечують продукти харчування, дерева або трансформовані рослини можуть бути оцінені, накущі, вибрані зі списку, який включає Acacia spp., приклад, за допомогою Саузерн-аналізу, на присуAcer spp., Actinidia spp., Aesculus spp., Agathis тність гену, який представляє інтерес, на кількість australis, Albizia amara, Alsophila tricolor, копій та/або геномну організацію. Альтернативно Andropogon spp., Arachis spp., Areca catechu, або додатково рівні експресії нової введеної ДНК Astelia fragrans, Astragalus deer, Baikiaea plurijuga, можуть піддаватися моніторингу при використанні Betula spp., Brassica spp., Bruguiera gymnorrhiza, Нозерн-та/або Вестерн-аналізу, обидві методики є Burkea africana, Butea frondosa, Cadaba farinosa, добре відомим середнім спеціалістам у даній галуCalliandra spp., Camellia sinensis, Canna indica, зі техніки. Capsicum spp., Cassia spp., Centroema pubescens, Одержані трансформовані рослини можуть буChaenomeles spp., Cinnamomum cassia, Coffea ти розмножені за допомогою різноманітності споarabica, Colophospermum mopane, Coronillia varia, собів, таких, як клональне розмноження або клаCotoneaster serotina, Crataegus spp., Cucumis spp., сичні технології схрещування. Наприклад, перше Cupressus spp., Cyathea dealbata, Cydonia oblonga, покоління (або Т1) трансформованих рослин може Cryptomeria japonica, Cymbopogon spp., Cynthea піддаватися самозапиленню для одержання гомоdealbata, Cydonia oblonga, Dalbergia monetaria, зиготного другого покоління (або Т2) трансформаDavallia divaricata, Desmodium spp., Dicksonia нтів, а Т2 рослини також розмножують за допомоsquarosa, Diheteropogon amplectens, Dioclea spp., гою класичних способів схрещування. Dolichos spp., Dorycnium rectum, Echinochloa Одержані трансформовані організми можуть pyramidalis, Ehrartia spp., Eleusine coracana, набувати різноманітних форм. Наприклад, вони Eragrestis spp., Erythrina spp., Eucalyptus spp., можуть бути химерами трансформованих клітин та Euclea schimpeh, Eulalia villosa, Fagopyrum spp., нетрансформованих клітин; клональними трансFeijoa seilowiana, Fragaria spp., Flemingia spp., формантами (наприклад, коли усі трансформовані Freycinetia banksii, Geranium thunbergii, Ginkgo клітини містять експресійну касету); пересадженою biloba, Glycine javanica, Gliricidia spp., Gossypium тканиною трансформованих та нетрансформоваhirsutum, Grevillea spp., Guibourtia coleosperma, них тканин (наприклад, у рослинах трансформоHedysarum spp., Hemarthia altissima, Heteropogon вані кореневища, привиті до нетрансформованих contortus, Hordeum vulgare, Hyparrhenia rufa, пагонів). Hypericum erectum, Hyperthelia dissoluta, Indigo Даний винахід охоплює будь-яку рослинну кліincamata, Iris spp., Leptarrhena pyrolifolia, Lespediza тину або рослину, одержану за допомогою будьspp., Lettuca spp., Leucaena leucocephala, Loudetia якого зі способів, описаних в даній заявці, та усі simplex, Lotonus bainesii, Lotus spp., Macrotyloma частини рослин або їх паростки. Даний винахід axillare, Malus spp., Manihot esculenta, Medicago охоплює потомство первинно трансформованої sativa, Metasequoia glyptostroboides, Musa або трансфікованої клітини, тканину, орган або sapientum, Nicotianum spp., Onobrychis spp., цілої рослини, що було одержане згідно з будьOmithopus spp., Oryza spp., Peltophorum africanum, яким зі згаданих вище способів, єдина вимога, Pennisetum spp., Persea gratissima, Petunia spp., щоб потомство демонструвало той самий генотип Phaseolus spp., Phoenix canariensis, Phormium та/або фенотипічну(і) характеристику(и), що й ті, cookianum, Photinia spp., Picea glauca, Pinus spp., які притаманні батьківській рослині, при викорисPisum sativum, Podocarpus totara, Pogonarthria танні способів згідно з винаходом. Винахід також fieckii, Pogonarthria squarrosa, Populus spp., включає хазяйські клітини, що містять ізольовану Prosopis cineraria, Pseudotsuga menziesii, молекулу нуклеїнової кислоти цикліну А, бажано Pterolobium stellatum, Pyrus communis, Quercus таку, що кодує білок цикліну А. Бажані хазяйські spp., Rhaphiolepsis umbellata, Rhopalostylis sapida, клітини згідно з винаходом представляють собою Rhus natalensis, Ribes grossularia, Ribes spp., 25 88779 26 Robinia pseudoacacia, Rosa spp., Rubus spp., Salix повтор, LB Ті С58) та правою граничною послідовspp., Schyzachynum sanguineum, Sciadopitys ністю (RB повтор, RB Ті С58)). verticillata, Sequoia sempervirens, Sequoiadendron Фіг.3 представляє собою таблицю, яка показує giganteum, Sorghum bicolor, Spinacia spp., перехресні для видів консервативні фрагменти, які Sporobolus fimbriatus, Stiburus alopecuroides, знайдені у цикліні А. Фрагмент 1 може використоStylosanthos humilis, Tadehagi spp., Taxodium вуватися для розрізнення цикліну А2 від інших distichum, Themeda triandra, Trifolium spp., Triticum циклінів А типу. Як приклад, показаний фрагмент, spp., Tsuga heterophylla, Vaccinium spp., Vicia spp., знайдений у цикліні В; 1. Vitis vinifera, Watsonia pyramidata, Zantedeschia Фіг.4 представляє собою послідовності, які виaethiopica, Zea mays, амарант, артишок, спаржу, користовуються у способах згідно з винаходом. броколлі, брюсельську капусту, кочанну капусту, Опис послідовностей канолу, моркву, цвітну капусту, селеру, листову SEQ ID NO: 1 представляє нуклеїнову кислоту капусту, льон, кормову капусту, чечевицю, оліявцикліну А2;2, що використовується у способах згімісне насіння рапсу, окру, цибулю, картоплю, рис, дно з даним винаходом. Вона є ідентичною кодуюсоєві боби, полуницю, цукровий буряк, цукрову чій послідовності, що задепонована під депозиттростину, соняшник, томати, гарбуз городній, деним номером NIVM21168, за винятком двох замін, рева та водорості, серед інших. перша у положенні, 851, в якій С замінює G, а друЗгідно з бажаною характеристикою даного вига у положенні 1295, в якій С замінює Т. Ці зміни находу рослина представляє собою сільськогосне передбачаються як такі, що мають які-небудь подарську рослину, що включає томати, картоплю, наслідки. тютюн, жито, соєві боби, соняшник, канолу, люцеSEQ ID NO: 2 представляє амінокислотну порну, рапс або бавовник. Крім того, переважно росслідовність цикліну А2;2, що кодується нуклеїнолини згідно з даним винаходом представляють вою кислотою згідно з SEQ ID NO: 1. Вона є іденсобою однодольні рослини, такі, як цукрова тростичною послідовності, яка задепонована під тина. Найбільш бажана рослина представляє содепозитним номером NP_568248, за винятком бою злакову культуру, таку, як овес, жито, рис, того, що вона містить дві амінокислотні заміни, кукурудза, пшениця, просо, сорго або ячмінь. згідно з першою пролін у положенні 284 замінює Даний винахід також охоплює рослини, які моаргінін, а згідно з другою серии у положенні 432 жуть бути одержані згідно зі способами за даним замінює фенілаланін. Ці зміни не передбачаються винаходом. Даний винахід, таким чином, забезпеяк такі, що мають які-небудь наслідки. чує рослини, які можуть бути одержані згідно з SEQ ID NO: 3 представляє собою проламіноданим винаходом, при цьому такі рослини мають вий промотор з рису. підвищений врожай у порівняні з контрольними SEQ ID NO: 4 та SEQ ID NO: 5 представляють рослинами, де рослини також мають переважну собою послідовності праймерів, які використовуекспресію цикліну А у рослинній тканині насіння. ються для клонування генів. Згідно з іншим втіленням даного винаходу заПриклади безпечується спосіб для одержання трансгенних Даний винахід буде описаний з посиланням на рослин, які мають підвищений врожай у порівнянні наступні приклади, які представлені тільки з метою з контрольними рослинами, що включає введення ілюстрації. у рослину нуклеїнової кислоти цикліну А, що опеМаніпуляції з ДНК ративно зв'язана з промотором, переважним для Якщо не визначено інше, то методики рекомнасіння. Циклін А може характеризуватися нуклеїбінантної ДНК здійснюють згідно зі стандартними новою кислотою згідно з SEQ ID NO: 1 або може прописами, які описані у Sambrook (2001) Молекупредставляти собою будь-який один з варіантів лярне клонування: лабораторне керівництво, 3-є нуклеїнової кислоти цикліну А, як визначено вище, видання, Cold Spring Harbor Laboratory Press, CSH, або може представляти собою будь-яку нуклеїноNew York) або у томах 1 та 2 Ausubel та ін. (1984), ву кислоту, яка підпадає під визначення нуклеїноСучасні прописи в молекулярній біології, сучасні вої кислоти цикліну А, як вищначено вище. прописи. Стандартні матеріали та способи для Опис фігур молекулярних маніпуляцій з рослинами описані в Даний винахід буде описаний з посилання на Plant Molecular Biology Labfase (1993) R.D.D. Cray, наступні фігури, в яких: опубл. BIOS Scientific Publications Ltd (UK) та Фіг.1 представляє собою філогенетичне дереBlackwell Scientific Publications (UK). во, одержане шляхом порівняння деяких послідоПриклад 1: Клонування генів вностей повної довжини білка цикліну А (за винятЦиклін А2;2 Arabidopsis thaliana (внутрішнє поком 1 часткової послідовності рису). Порівняння силання CDS95) ампліфікували за допомогою проводили при використанні програми Clustal, виПЛР, використовуючи як матрицю бібліотеку кДНК користовуючи установки по умовчанню, а дані проростків Arabidopsis thaliana (Invitrogen, Paisley, представляли у вигляді філограми. Як показано на UK). Після зворотної транскрипції РНК, екстрагофігурі, послідовності згруповані у чотири основні ваної з проростків, кДНК клонували в pCMV Sport представлені групи. 6,0. Середній розмір вставки складав 1,5 кілобаз, а Фіг.2 Бінарний вектор для експресії в Oryza вихідна кількість клонів становила 1,59x107 КУО. saliva гену цикліну А2;2 Arabidopsis thaliana під Вихідний титр, який визначали, становив 9,6x105 контролем промотора PROLAMIN. Вектор містить КУО/мл, а після першої ампліфікації він складав Т-ДНК, що має походження від Ті-плазміди, ДНК є 6x1011 КУО/мл. Після плазмідної екстракції 200нг обмеженою зліва граничною послідовністю (LB матриці використовували у 50мкл суміші ПЛР. Праймери prm582 (смисловий, стартовий кодон 27 88779 28 представлений жирним шрифтом, AttB1 сайт видізиготи) та приблизно 10 Т1 проростків, в яких нелений курсивом: 5' має трансгену (нулізиготи) ggggacaagfffgfacaaaaaagcaggcffcacaatgtattgctcttcttc (і) Вимірювання вегетативного росту: gatgc 3') та prm583 (зворотний, комплементарний, Відібрані Т1 рослини (приблизно 10 з трансгестоп-кодон представлений жирним шрифтом, ном та приблизно 10 без трансгену) переносили у AttB2 сайт виділений курсивом: 5' вегетаційні будиночки. Кожна рослина одержувала ggggaccacftfgfacaagaaagcfgggfgctfggfgtcatcttgagaat мітку зі штрих-кодом для однозначного пов'язуag 3'), які включають сайти AttB для рекомбінації вання фенотипічних даних з відповідною рослиGateway, використовували для ПЛР ампліфікації. ною. Відібрані Т1 рослини вирощували на ґрунті у ПЛР проводили при використанні Hifi Taq ДНК погорщиках з діаметром 10см при наступних умовах лімерази у стандартних умовах. ПЛР фрагмент навколишнього середовища: фотоперіод = 11,5 розміром 1311п.о. піддавали ампліфікації та очигодин, інтенсивність денного світла = 30,000 люкс щали при використанні стандартних способів. Поабо більше, денна температура = 28°С або вище, тім проводили перший етап процедури Gateway, нічна температура = 22°С, відносна вологість = 60BP реакцію, під час якої ПЛР фрагмент рекомбіно70%. Трансгенні рослини та відповідні нулізиготи вали in vivo з pDONR201 плазмідою для одержанвирощували разом при випадковому розміщенні. ня згідно з термінологією Gateway, «вхідного клоПочинаючи з етапу посіву до стадії зрілості кожну ну» р754. Плазміду pDONR201 закуповували від рослину декілька разів пропускали через камеру формування цифрового зображення для створенInvitrogen, як частину технології GatewayÒ . ня образів. У кожний момент часу цифрові зобраПриклад 2: Конструювання вектора ження (2048x1536 пікселів, 16 мільйонів кольорів) Вхідний клон р754, що містить послідовність знімали для кожної рослини під, принаймні, 6 різцикліну А2.2, послідовно використовували в LR ними кутами. Параметри, такі, як площа надземної реакції з цільовим вектором, здійснюючи трансфоповерхні, одержували автоматично з кожного цирмацію Oryza saliva. Цей вектор містив як функціофрового зображення для усіх рослин, використональні елементи у межах границь Т-ДНК. рослинвуючи програмне забезпечення для аналізу зоний селективний маркер, маркер для візуалізації бражень. та Gateway касету, призначену для LR in vivo ре(іі) Вимірювання параметрів, пов'язаних з накомбінації з послідовністю, що представляє інтесінням: рес, що вже була клонована у вхідний клон. ПроЗрілі первинні волоті збирали, поміщали у міламіновий промотор для понадекспресії (PRO90) шечки, мітили за допомогою штрих-коду, а потім розміщувався вище від Gateway касети. висушували протягом трьох днів у сушильній шафі Одержаний після етапу LR рекомбінації експри 37°С. Волоті потім обмолочували та усе насінпресійний вектор, показаний на Фігурі 2 (Циклін А ня збирали та підраховували. Наповнену обгортку 2;2: проламін - підвищувальна регуляція), тра та порожню обгортку відокремлювали одна від нсформували в A grobacterium та послідовно в одної за допомогою пристрою для продування рослини Oryza sativa. Трансформовані рослини повітрям. Порожню обгортку відкидали, а фракцію, рису вирощували, а потім перевіряли на параметщо залишилася, знову підраховували. Наповнену ри, описані у Прикладі 3. обгортку зважували на аналітичних вагах. Ця проПриклад 3: Оцінка та результати цедура приводила до одержання набору параметА. Статистичний аналіз: F-аналіз рів, пов'язаних з насінням, описаного нижче. Двофакторний ANOVA (аналіз варіантів) вико(а) Загальна кількість насіння ристовували як статистичну модель для загальної Загальну кількість насіння визначали шляхом оцінки фенотипічних характеристик рослини. Fпідрахунку кількості обгорток, зібраних з рослини. аналіз проводили для усіх виміряних параметрів Результати для загальної кількості насіння з росусіх рослин для усіх подій трансформації з геном лин, які експресують циклін А2;2 під контролем згідно з даним винаходом. F-аналіз здійснювали промотора, переважного для насіння (проламінодля перевірки впливу гену на усі події трансфорвий), показані нижче у Таблиці 2 мації та для перевірки на загальний вплив гену, що є відомим як «глобальний ефект гену». Якщо Таблиця 2 оцінка F-аналізу показувала, що дані є достовірними, то вона потім включалася як така, що має Загальна кількість насіння для рослин, «генний» ефект, це означає, що більше, ніж тільки що експресують циклін А2;2: промотор, бажаний присутність положення гену, спричинює відмінносдля насіння (проламіновий) ті у фенотипі. Поріг для достовірності для справді глобального генного ефекту гену встановлювався на рівні 5% достовірності для F-аналізу Загальна кількість насіння В. Пропис для оцінки % рОдержували приблизно від 15 до 20 незалежЛінія TR нульові різниця різзначення них ТО трансформантів рису. Первинні трансфорниці манти переносили з камер для тканинної культури Загальна 549,8 437,6 112,25 26 0,0397 до вегетаційних будиночків для вирощування та кількість збору врожаю Т1 насіння. Зберігали деякі події, при яких Т1 потомство сегрегує у співвідношенні Загальна кількість насіння для рослин, які екс3:1 стосовно присутності/відсутності трангену. Для пресують циклін А2;2 під контролем конститутивкожної з цих подій відбирали приблизно 10 Т1 ного промотора (дані не представлені) була менпроростків, що містять трансген (гетеро- та гомошою від загальної кількості насіння, одержаного 29 88779 30 від рослин, які експресують ген цикліну А2;2 під для кількості виповненого насіння для рослин, які контролем проламінового промотора (який, як поекспресують циклін А2;2 під контролем промотора, казано вище, дає значуще р-значення, одержане з переважного для насіння (проламіновий), показані F-аналізу, що свідчить про загальний вплив гену). нижче (Таблиця 3 для Т1 рослин та Таблиця 4 для (b) Кількість виповненого насіння Т2 рослин) проти результатів кількості виповненоКількість виповненого насіння визначали шляго насіння з рослин, які експресують циклін А2;2 хом підрахунку кількості наповнених обгорток, що під контролем конститутивного промотора (Таблизалишилися після етапу розділення. Результати ця 5). Таблиця 3 Т1 оцінка - кількість виповненого насіння для рослин, що експресують циклін А2;2: проламіновий промотор Лінія Загальна кількість Кількість виповненого насіння нульові різниця 322,9 89,98 TR 412,9 Результати показують значущий загальний генний ефект (зі значущим р-значенням, одержаним з F-аналізу). Т1 трансгенні рослини показують значне підвищення кількості виповненого насіння у порівнянні з контрольними рослинами. Таблиця 4 Т2 оцінка - кількість виповненого насіння для рослин, що експресують циклін А2;2: проламіновий промотор Лінія Загальна кількість Кількість виповненого насіння % рTR нульові різниця різзначення ниці 438,4 315,1 123,32 39 0,0139 Результати показують значущий загальний генний ефект (зі значущим р-значенням, одержаним з F-аналізу). Т2 трансгенні рослини показують значне підвищення кількості виповненого насіння у порівнянні з контрольними рослинами. Таблиця 5 Кількість виповненого насіння для рослин, які експресують циклін А2;2: pGOS2 % різниці 28 р-значення 0,0261 (с) Загальний врожай насіння Загальний врожай насіння на рослину вимірювали шляхом зважування усіх виповнених обгорток, зібраних з рослини. Результати загального врожаю насіння рослин, які експресують циклін А2;2 під контролем проламінового промотора (див Таблицю 6 для Т1 оцінки результатів та Таблицю 7 для результатів Т2 оцінки) проти загального врожаю насіння для рослин, які експресують циклін А2;2 під контролем pGOS2 (див Таблицю 8), представлені нижче. Таблиця 6 Т1 оцінка - загальний врожай насіння для рослин, які експресують циклін А2;2: проламіновий промотор Кількість виповненого насіння % рЛінія TR нульові різниця різзначення ниці Загальна 9 6,9 2,07 30 0,0211 кількість Результати Т1 оцінки показують загальний генний ефект зі значущим р-значенням, одержаним з F-аналізу. Таблиця 7 Лінія Загальна кількість Кількість виповненого насіння % рTR нульові різниця різзначення ниці 256,8 241,4 15,41 6 0,1007 Результати показують, що існує підвищення кількості виповненого насіння для рослин, які експресують циклін А2;2 під контролем конститутивного промотора, у порівнянні з контрольними рослинами, проте рослини які екпресують циклін А2;2 під контролем проламінового промотора, показують більшу кількість виповненого насіння (див. Таблицю 3 та Таблицю 4 вище). Т2 оцінка - загальний врожай насіння для рослин, які експресують циклін А2;2: проламіновий промотор Кількість виповненого насіння % рЛінія TR нульові різниця різзначення ниці Загальна 9,5 6,6 2,85 43 0,0128 кількість Результати Т2 оцінки показують загальний генний ефект зі значущим р-значенням, одержаним з F-аналізу. 31 88779 32 Таблиця 8 Таблиця 10 Загальний врожай насіння для рослин, які експресують циклін А2;2: pGOS2 TKW для рослин, які експресують циклін А2;2: проламіновий промотор Загальна вага насіння Лінія Загальна кількість % рTR нульові різниця різзначення ниці 6,3 5,7 0,6 11 0,0803 Як показано у Таблиці 8, загальний врожай насіння для рослин, які експресують циклін А2;2 конститутивно, не є таким добрим, як результати для експресії під контролем переважного для насіння промотора (див. Таблицю 6 та Таблицю 7 вище). (d) Індекс збору врожаю Індекс збору врожаю визначається в даній заявці як співвідношення між загальним врожаєм насіння та надземною площею рослини (мм2), помноженому на коефіцієнт 106. Індекс збору врожаю для рослин, які експресують циклін А2;2 під контролем промотора, переважного для рослин (проламіновий), показаний нижче у Таблиці 9. Таблиця 9 Індекс збору врожаю для рослин, які експресують циклін А2;2: проламіновий промотор Індекс збору врожаю Лінія TR Загальна кількість 103,7 % рнульові різниця різзначення ниці 83,4 20,29 24 0,0711 Індекс збору врожаю для рослин, які експресують циклін А2;2 під контролем прламінового промотора, показує, що існує загальний генний ефект (див. р-значення, одержані з F-аналізу). Індекс збору врожаю для рослин, які експресують циклін А2;2 під контролем конститутивного промотора (результати не показані), не є таким добрим, як результати, показані у Таблиці 9 вище. (e) Вага тисячі зернин (TKW) TKW виводять з кількості підрахованого виповненого насіння та його загальної ваги. Результати TKW для рослин, які експресують циклін А2;2 під контролем проламінового промотора (див. Таблицю 10) проти TKW для рослин, які експресують циклін А2;2 під контролем pGOS2 (див. Таблицю 11), показані нижче. TKW Лінія Загальна кількість % рTR нульові різниця різзначення ниці 21,1 19,9 1,26 6 0,108 Загальний генний ефект є очевидним з таблиці, що приведена вище, яка показує значуще рзначення, одержане з F-аналізу Таблиця 11 TKW для рослин, які експресують циклін А2;2 pGOS2 TKW Лінія Загальна кількість % рTR нульові різниця різзначення ниці 24 23,3 0,7 3 0,1509 Результати TKW для рослин, які експресують циклін А2;2 під контролем pGOS2, показують підвищення значення TKW, але підвищення не є таким великим, як підвищення, представлене у Таблиці 10 вище. (iiі) Стресова оцінка рослин, які експресують циклін А2;2: проламіновий промотор Висівали насіння та через 10 днів проростки пересаджували у горщики, діаметром 10 см, заповнені 1:1 сумішшю волого піску та вермікуліту. Горщики діаметром 10см ставили у горщики діаметром 12см з одним шаром пластикової сітки між двома горщиками для запобігання вилужування субстрату. Горщики потім поливали свіжою водою перед пересаджуванням. Через один після пересаджування проростки поміщали у сольові умови. Горщики зволожували 4 рази на день у 8, 12, 16 годин та о двадцять першій годині за допомогою поживного розчину, який індукує сольовий стрес та містить наступні елементи: - NPK поживна суміш, 20-20-20 Peters professional (Scotts) у концентрації 1кг/м3 - Хелат магнію Mg (BMS, Bornem, Belgium) при концентрації 333,33мл/м3 - Хелат заліза, Libfer (CIBA, Bradford, UK) при концентрації 21,67г/м3 - NaCl 1,425кг/м3 За концентрацією солі спостерігали кожного тижня, здійснюючи додання у разі необхідності. Рослини вирощували в умовах сольового стресу до моменту початку виповнення зерна. У першій точці часу їх переносили у різноманітні місця вегетаційного будиночка, де вони піддавалися щоденному поливу за допомогою свіжої води до збору насіння. Вимірювали та реєстрували наступні па 33 88779 раметри таким самим чином, як для рослин, які не вирощували в умовах сольового стресу так, як зазначено в (а)-(e) вище. Таблиця 12 34 Таблиця 15 Індекс збору врожаю для рослин, які експресують циклін А2;2: проламіновий промотор Загальна вага насіння для рослин, які експресують циклін А2;2: проламіновий промотор Лінія Загальна вага насіння Лінія Загальна кількість % рTR нульові різниця різзначення ниці 1,3 0,9 0,47 55 0,0217 Результати, представлені у Таблиці 12, показують значущий генний ефект, який є очевидним з р-значення F-аналізу для рослин, які експресують циклін А2;2 під контролем проламінового промотора в умовах стресу, як проілюстровано за допомогою сольового стресу Таблиця 13 Кількість виповненого насіння для рослин, які експресують циклін А2;2: проламіновий промотор Лінія Загальна кількість Кількість виповненого насіння % рTR нульові різниця різзначення ниці 70,8 48,5 22,33 46 0,0396 Результати, представлені Таблиці 13, показують значущий генний ефект, який є очевидним з рзначення F-аналізу для рослин, які експресують циклін А2;2 під контролем проламінового промотора в умовах стресу, як проілюстровано за допомогою сольового стресу. Таблиця 14 Загальна кількість насіння для рослин, що експресують циклін А2;2: проламіновий промотор Лінія Загальна кількість Загальна кількість насіння % рTR нульові різниця різзначення ниці 140,7 105,1 35,55 Індекс збору врожаю 34 0,0594 Результати, представлені у Таблиці 14, показуют збільшення загальної кількості насіння у порівнянні з контрольними рослинами. Загальна кількість % рTR нульові різниця різзначення ниці 62,1 30,4 31,64 104 0,3522 Результати, представлені у Таблиці 15, показують збільшення індексу збору врожаю у порівнянні з контрольними рослинами. Таблиця 16 TKW для рослин, які експресують циклін А2;2: проламіновий промотор TKW Лінія TR нульові різниця Загальна 17,4 кількість 0,98 6 % різрниці значення 0,0269 17,4 Результати, представлені у Таблиці 16, показують значущий генний ефект, який є очевидним з р-значення F-аналізу для рослин, які експресують циклін А2;2 під контролем проламінового промотора в умовах стресу, як проілюстровано за допомогою сольового стресу. Приклад 4: Застосування винаходу на кукурудзі Винахід, описаний в даній заявці, може також використовуватися на кукурудзі. Цикілн клонують під контролем бажаного для насіння промотора у трансформаційний вектор, прийнятний для здійснення опосередкованої Agrobacterium трансформації кукурудзи. Такі вектори та способи для трансформації кукурудзи, були описані в літературі (ЕР0604662, ЕР0672752, ЕР0971578, ЕР0955371, ЕР0558676, Ishida та ін. 1996; Frame та ін., 2002). Трансгенні рослини, одержані за допомогою цих способів, вирощували у вегетаційних будиночках для одержання насіння Т1. Здатність до успадковування та кількість копій трансгену перевіряли за допомогою кількісної ПЛР у реальному часі та Саузерн-блотінгу, а експресійні рівні трансгену визначали за допомогою зворотної ПЛР та Нозерн-аналізу. Трансгенні лінії з одноколійними інсерціями трансгену та з рівнями експресії трансгену, що варіюють, відбирали для одержання Т2 насіння. Насіння потомства розмножували та вирощували у вегетаційних будиночках в умовах, адаптованих для кукурудзи (16:8 фотоперіод, 2628°С денна температура та 22-24°С нічна температура) а також в умовах водного дефіциту, недостатності азоту та надлишку NaCI. У випадку самозапилення нульові сегреганти з однієї й тієї ж батьківської лінії, а також рослини дикого типу одного й того ж культивару використо 35 88779 36 вували як контролі. Рослини потомства, які були бування та схрещування при використанні маркеодержані в результаті самозапилення або перера, з метою перенесення трансгенних ознак, які хресного схрещування, оцінювали на різні парамемають польове значення, у комерційну зародкову три біомаси та ростові параметри, включаючи вагу плазму. Способи аналізу кукурудзи на ріст та парослин, товщина стовбура/стебла, кількість листя, раметри, пов'язані з врожаєм у польових умовах, є загальну надземну масу, зелене забарвлення лисдобре відомими у галузі техніки, як наприклад метя, час до досягнення зрілості, час до фази появи тодики для інтрогресії специфічних локусів (таких маточкових стовпчиків, час цвітіння, кількість колояк локуси, що містять трансген) з однієї зародкової сків, довжина колосу, кількість рядків у колоску, плазми в іншу. Такі також включають перенос кількість зернин, розмір зернин, вміст олії у зернах, ознаки(ознак), яка(які) представляє (представлязрілість зерна, час збору. Лінії, які є найбільше ють) інтерес з трансформованої інбредної лінії у поліпшені згідно з одним зі згаданих вище парамекомерційний гібрид з бажаною доданою агрономітрів, відбирали для подальшого польового випрочною або поживною або медичною цінністю. 37 88779 38 39 88779 40 41 88779 42 43 88779 44 45 88779 46 47 88779 48 49 Комп’ютерна верстка О. Гапоненко 88779 Підписне 50 Тираж 28 прим. Міністерство освіти і науки України Державний департамент інтелектуальної власності, вул. Урицького, 45, м. Київ, МСП, 03680, Україна ДП “Український інститут промислової власності”, вул. Глазунова, 1, м. Київ – 42, 01601
ДивитисяДодаткова інформація
Назва патенту англійськоюPlants having increased yield, and method for obtaining thereof
Автори англійськоюFRANKARD VALERIE, Mironov Vladimir
Назва патенту російськоюРастения, имеющие повышенный урожай, и способ их получения
Автори російськоюФранкард Валери, Миронов Владимир
МПК / Мітки
МПК: C12N 9/12, C12N 15/82, A01H 5/00, C12N 15/00
Мітки: спосіб, мають, рослини, підвищений, врожай, одержання
Код посилання
<a href="https://ua.patents.su/25-88779-roslini-yaki-mayut-pidvishhenijj-vrozhajj-ta-sposib-kh-oderzhannya.html" target="_blank" rel="follow" title="База патентів України">Рослини, які мають підвищений врожай, та спосіб їх одержання</a>
Плазміда pslc1-1/prd400, штам gv3101 agrobacterium tumefaciens, спосіб одержання трансгенної рослини з масловмісним насінням, спосіб виділення їстівної або неїстівної олії з масловмісного насіння рослини

Номер патенту: 49833
Опубліковано: 15.10.2002
Автори: Келлер Вілфред А., Маккензі Самюел Л., Катавік Весна, Зоу Джітао, Тейлор Девід С.
МПК: C12N 1/21, C12N 15/09, C12N 5/10, C11B 1/00, C12N 15/82, C12N 15/00, A01H 5/00, C11B 1/10, C12N 9/10
Мітки: рослини, agrobacterium, насінням, одержання, масловмісним, спосіб, неїстівної, їстівної, штам, масловмісного, олії, tumefaciens, виділення, насіння, трансгенної, gv3101, плазміда
Формула / Реферат:
1. Плазміда pSLC1-1/pRD400 (ATCC 97545).2. Штам GV3101 Agrobacterium tumefaciens, який відрізняється тим, що вищезгаданий штам модифікований введенням дріжджового гена SLC1-1, що має нуклеотидну послідовність SEQ ID NO: 1.3. Спосіб одержання трансгенної рослини з масловмісним насінням, який відрізняється тим, що в геном вищезгаданої рослини вводять дріжджовий ген SLC1-1, що здатен до експресії і який має нуклеотидну...
Продукт на основі насіння рослини і спосіб його одержання

Номер патенту: 77356
Опубліковано: 15.11.2006
Автори: Качановська Лідія Дмитрівна, Цигульов Олег Васильович
МПК: A23L 1/025, A23L 1/18, A23L 1/052
Мітки: основі, рослини, продукт, одержання, насіння, спосіб
Формула / Реферат:
1. Продукт на основі насіння рослини, що містить вуглевод і складові насіння, який відрізняється тим, що вуглевод присутній у вигляді рекомбінанта полісахариду загальної формули R•R1(H2O)n, де R - амілопектиновий радикал, R1 - радикал глюкозидного залишку (D-GIc D-Fru), n=4 або 8, а співвідношення компонентів складає, мас. %: рекомбінант полісахариду 10-80 ...
Спосіб одержання соняшникової рослини стійкої до вовчка

Номер патенту: 86569
Опубліковано: 12.05.2009
Автор: Хассан Ель Сайєд Абдел Рауф Садік
МПК: A01H 4/00, A01H 1/04, A01H 5/10, C12N 5/04, A01H 5/00
Мітки: стійкої, одержання, вовчка, спосіб, рослини, соняшникової
Формула / Реферат:
1. Спосіб одержання соняшникової рослини, стійкої до сортів вовчка (Orobanche сumana) поряд із сортами А-Е, який містить рослину, одержану методом, що включає:схрещення першої вихідної соняшникової рослини з другою вихідною соняшниковою рослиною та збір урожаю отриманого потомства насіння, в якому першою вихідною соняшниковою рослиною є рослина, вирощена з насіння лінії U00S9LM, представленої ATCC...
Похідні n-оксиду піридину, що мають цитокінінову активність, та спосіб їх одержання

Номер патенту: 65526
Опубліковано: 15.04.2004
Автори: Білогубова Олена Миколаївна, Дульнєв Петро Георгійович, Чернишенко Тетяна Володимирівна, Сидоров Володимир Анатолійович, Кондратенко Сергій Іванович
МПК: A01P 21/00, C07D 213/89, A01N 43/40
Мітки: n-оксиду, активність, одержання, цитокінінову, піридину, спосіб, мають, похідні
Формула / Реферат:
1. Похідні N-оксиду піридину загальної формули (І)що мають цитокінінову дію,де n = 1, 2,R = R’ = H абоR = H, R’ = CH3 в 2 або 3, або 4 положеннях, абоR = R’ = CH3 в 2, 6 положеннях,X = MnY2, де Y - галоїд або X = R”-COOH, де R” = алкіл, арил, гетероарил, -(СН2)m-COOH, -(CH=CH)m-COOH,де m = 1-4.2. Спосіб...
Сполуки, що мають ауксинову дію, та спосіб їх одержання

Номер патенту: 56161
Опубліковано: 15.05.2003
Автори: Дульнєв Петро Георгійович, Сидоров Володимир Анатолійович, Чернишенко Тетяна Володимирівна, Кондратенко Сергій Володимирович
МПК: C07D 213/89, C07F 3/00, A01N 43/40, A01P 21/00, A01N 55/02
Мітки: дію, мають, спосіб, сполуки, ауксинову, одержання
Формула / Реферат:
1. Сполуки загальної формули (І),де R = R1 = H, метил або R = H, R1 = метил у 2 або 3, або 4, або 6 положенні; n = 2; Х = Zn(галоген)2, де галоген - Cl, І, що мають ауксинову дію.2. Спосіб одержання сполук формули (І),де R = метил; R1 = H, метил у 2 або 3, або...
Попередній патент: Будівельний елемент, будівельна конструкція, спосіб виготовлення будівельного елемента і спосіб виготовлення будівельної конструкції
Наступний патент: Харчова добавка, що гальмує метаболізм алкоголю
Випадковий патент: Сонячний опріснювач