Спосіб гальмування активності аденілсукцинатсинтетази в метилтіоаденозинфосфорилаза-дефіцитних клітинах, спосіб гальмування активності аденілсукцинатсинтетази в клітинах ссавців-хазяїв
Номер патенту: 47465
Опубліковано: 15.07.2002
Автори: Коттам Говард Б., Ноборі Цутому, Каррера Карлос Дж., Карсон Денніс А.








Формула / Реферат
1. Спосіб гальмування активності аденілсукцинатсинтетази в метилтіоаденозинфосфорилаза-дефіцитних клітинах, який включає визначення того, чи є популяція клітин, отримана з клітин ссавців, метилтіоаденозинфосфорилаза-дефіцитною, введення терапевтично ефективної кількості інгібітора аденілсукцинатсинтетази в організм хазяїна, звільнення в результаті зазначеного контакту метилтіоаденозинфосфорилаза-дефіцитних клітин хазяїна від аденозин 5'-монофосфату.
2. Спосіб за п. 1, який відрізняється тим, що перший етап складається з отримання зразка клітин, які можуть бути метилтіоаденозинфосфорилаза-дефіцитними, додавання до зразка олігонуклеотидних проб, які будуть специфічно гібридизуватися з будь-якою присутньою у зразку нуклеїновою кислотою, яка кодує метилтіоаденозинфосфорилазу, за умов, які дозволятимуть пробам гібридизуватися до будь-якої нуклеїнової кислоти, присутньої у зразку, та визначення присутності у зразку нуклеїнової кислоти, яка кодує метилтіоаденозинфосфорилазу, де присутність вищезгаданої нуклеїнової кислоти виявляє присутність протеїну метилтіоаденозинфосфорилази у зразку клітин.
3. Спосіб за п. 1, який відрізняється тим, що інгібітором аденілсукцинатсинтетази є L-аланозин.
4. Спосіб за п. 1, який відрізняється тим, що хазяїном є людина.
5. Спосіб за п. 1, який відрізняється тим, що метилтіоаденозинфосфорилаза-дефіцитні клітини є клітинами первинної пухлини, вибраними з групи, яка складається з клітин недрібноклітинної карциноми легенів, клітин гострого лімфобластомного лейкозу, клітин гліоми та клітин пухлини оболонки сечовивідних органів.
6. Спосіб за п. 1, який відрізняється тим, що інгібітор аденілсукцинатсинтетази вводять в організм хазяїна дрібними дозами щоденно або шляхом тривалої інфузії.
7. Спосіб за п. 1, який відрізняється тим, що субстрат для внутрішньоклітинного продукування АМФ вводять в організм хазяїна після лікування згідно з другим етапом.
8. Спосіб гальмування активності аденілсукцинатсинтетази в клітинах ссавців-хазяїв, про які відомо, що вони є метилтіоаденозинфосфорилаза-дефіцитними, який відрізняється тим, що складається з введення терапевтично ефективної кількості інгібітора аденілсукцинатсинтетази в організм хазяїна, що звільнює метилтіоаденозинфосфорилаза-дефіцитні клітини хазяїна від аденозин 5'-монофосфату.
9. Спосіб за п. 8, який відрізняється тим, що інгібітором аденілсукцинатсинтетази є L-аланозин.
10. Спосіб за п. 8, який відрізняється тим, що хазяїном є людина.
11. Спосіб за п. 8, який відрізняється тим, що метилтіоаденозинфосфорилаза-дефіцитні клітини хазяїна є клітинами немілкоклітинної карциноми легенів, клітинами гострого лімфобластомного лейкозу, клітинами гліоми.
12. Спосіб за п. 8, який відрізняється тим, що інгібітор аденілсукцинатсинтетази вводять в організм хазяїна дрібними дозами щоденно або шляхом тривалої інфузії.
13. Спосіб за п. 8, який відрізняється тим, що субстрат для внутрішньоклітинного вироблення АМФ вводиться в організм хазяїна після введення йому інгібітора аденілсукцинатсинтетази.
Текст
1 Спосіб гальмування аісгивності аденілсукцинатсинтетази в метилтюаденозинфосфорилазадефіцитних клітинах, який включає визначення того, чи є популяція клітин, отримана з клітин ссавців, метилтюаденозинфосфорилаза-дефіцитною, введення терапевтично ефективної КІЛЬКОСТІ інпбітора аденілсукцинатсинтетази в організм хазяїна, звільнення в результаті зазначеного контакту метилтюаденозинфосфорилаза-дефіцитних клітин хазяїна від аденозин 5'-монофосфату 2 Спосіб за п 1, який відрізняється тим, що перший етап складається з отримання зразка клітин, які можуть бути метилтюаденозинфосфорилазадефіцитними, додавання до зразка олігонуклеотидних проб, які будуть специфічно гібридизуватися 3 будь-якою присутньою у зразку нуклеїновою кислотою, яка кодує метилтюаденозинфосфорилазу, за умов, які дозволятимуть пробам гібридизуватися до будь-якої нуклеїнової кислоти, присутньої у зразку, та визначення присутності у зразку нуклеїнової кислоти, яка кодує метилтюаденозинфосфорилазу, де присутність вищезгаданої нуклеїнової кислоти виявляє присутність протеїну метилтюаденозинфосфорилази у зразку клітин 3 Спосіб за п 1, який відрізняється тим, що інгібітором аденілсукцинатсинтетази є L-аланозин 4 Спосіб за п 1, який відрізняється тим, що хазяїном є людина 5 Спосіб за п 1, який відрізняється тим, що метилтюаденозинфосфорилаза-дефіцитні клітини є клітинами первинної пухлини, вибраними з групи, яка складається з клітин недрібноклітинної карциноми легенів, клітин гострого лімфобластомного лейкозу, клітин гліоми та клітин пухлини оболонки сечовивідних органів 6 Спосіб за п 1, який відрізняється тим, що інгібітор аденілсукцинатсинтетази вводять в організм хазяїна дрібними дозами щоденно або шляхом тривалої інфузм 7 Спосіб за п 1, який відрізняється тим, що субстрат для внутрішньоклітинного продукування АМФ вводять в організм хазяїна після лікування згідно з другим етапом 8 Спосіб гальмування активності аденілсукцинатсинтетази в клітинах ссавців-хазяїв, про які відомо, що вони є метилтюаденозинфосфорилазадефіцитними, який відрізняється тим, що складається з введення терапевтично ефективної КІЛЬКОСТІ шпбітора аденілсукцинатсинтетази в організм хазяїна, що звільнює метилтюаденозинфосфорилаза-дефіцитні клітини хазяїна від аденозин 5'монофосфату 9 Спосіб за п 8, який відрізняється тим, що інгібітором аденілсукцинатсинтетази є L-аланозин 10 Спосіб за п 8, який відрізняється тим, що хазяїном є людина 11 Спосіб за п 8, який відрізняється тим, що метилтюаденозинфосфорилаза-дефіцитні клітини хазяїна є клітинами немілкоклітинної карциноми легенів, клітинами гострого лімфобластомного лейкозу, клітинами гліоми 12 Спосіб за п 8, який відрізняється тим, що інгібітор аденілсукцинатсинтетази вводять в організм хазяїна дрібними дозами щоденно або шляхом тривалої інфузи 13 Спосіб за п 8, який відрізняється тим, що субстрат для внутрішньоклітинного вироблення АМФ вводиться в організм хазяїна після введення йому шпбітора аденілсукцинатсинтетази О ю (О 47465 Це часткове продовження патентної заявки США № 08/176,855, зареєстрованої 29 грудня 1993 року, яка розглядається зараз Винахід стосується фармацевтичних препаратів та способів їх використання при хіміотерапевтичному лікуванні раку Більш конкретно, винахід стосується способів визначення ракових клітин, які не можуть перетворити метилтюаденозинфосфорилазу на аденін під час відновлювального синтезу аденшових нуклеотидів, та використання Lаланозину для інгібування de novo синтезу аденозин 5'-монофосфату (АМФ) в таких ракових клітинах Метилтюаденозин (МТА) розщеплюється в здорових клітинах ссавців метилтюаденозинфосфорилазою (МТАза) на аденін та метилтюрибозу1-Ф, остання з яких є субстратом метаболічного синтезу метіоніну Аденін утилізується в клітинному пулі аденозин 5'-монофосфату (АМФ), з якого клітини одержують аденозин 5'-трифосфат (АТФ) для метаболічної енергії та 2'-дезоксиаденозин-5'трифосфат (дАТФ) для синтезу ДНК Спираючись на дослідження in vitro, L-ізомер бактеріального антибіотичного аланозину (одержаного з Streptomyces alanosmicus, далі "Lаланозин") є придатним для використання як противірусний та протипухлинний агент Зокрема вважається, що L-аланозин інгібує аденілосукцинатсинтетазне (АСС) перетворення інозин 5'монофосфату (ІМФ) на АМФ, таким чином звільнюючи клітини-мішені від АМФ та АТФ (у відсутності аденіну) Проте КЛІНІЧНІ дослідження терапевтичної ефективності L-аланозину у пацієнтів, що страждають на рак, не дали успішних результатів (див , наприклад, дані, наведені Tyagi and Cooney, Adv Pharmacol Chemoterapy, 20 69-120, 1984 [результати ВІДНОСНО ефективності L-аланозинів при лікуванні раку людини, отримані в той час, були оцінені як "короткочасна підтримка"], Creagan, et al, Cancer, 52 615-618, 1993 [на Етапі II досліджень загальний рівень ВІДПОВІДІ становив лише 4 % ] , Creagan, et al, Am J Clm OncoL, 7 543-544, 1984 [на Етапі II досліджень у пацієнтів з меланомою спостерігалася незначна терапевтична ВІДПОВІДЬ], VonHoff, et al , Invest, New Drugs, 9 87-88, 1991 [ніякої очевидної ВІДПОВІДІ у пацієнтів з раком грудної залози]) В результаті в усіх КЛІНІЧНИХ випадках лікування раку від використання L-аланозину відмовились Іншим відомим інгібітором АСС активності є хадацидин Проте хадацидин вважається більш токсичним для людини, ніж L-аланозин Крім того, активність інших інгібіторів синтезу пурину de novo (таких як метотрексат, 6-меркаптопурин, 6тюгуанш та d-ідеазатетрапдрофолат), які блокують синтез ІМФ (та, таким чином, теоретично видаляють ІМФ як джерело синтезу АМФ), було виявлено in vivo за допомогою гіпоксантину, якого багато у плазмі, що використовується як субстрат для вироблення ІМФ Отже, на СЬОГОДНІШНІЙ день характеристики агентів in vivo, які блокують метаболічний шлях аденіну для вироблення внутрішньоклітинного АМФ, є незадовільними Однак з розвитком дуже чутливих тестів для визначення гомозиготної делецм гена, яка кодує МТАзу в певних ракових клітинах людини (див , звичайно наведену основну заявку на патент США з серійним номером 08/176,855) стало відомо, що пухлини, які КЛІНІЧНО лікуються L-аланозином, виробляють МТАзу та, таким чином, здатні забезпечити достатню КІЛЬКІСТЬ аденіну для підтримування пулу АМФ, незважаючи на інгібування синтезу АМФ за допомогою ІМФ Цей винахід, таким чином, передбачає спосіб визначення клітин, які не мають МТАзи, та лікування таких клітин шляхом звільнення їх від АМФ Було виявлено, що клітини, з яких був видалений ген, що кодує протеїн МТАзи, та які не здатні перетворити МТА на аденін ("МТАза-дефіцитні клітини"), селективно знищуються in vivo при контакті з терапевтичне ефективною дозою шпбітора синтезу АМФ de novo, такого як L-аланозин Таким чином, незважаючи на те, що L-аланозин не є терапевтичне ефективним проти всіх ракових клітин, він є терапевтичне ефективним проти МТАзадефіцитних клітин В одному аспекті винахід передбачає спосіб визначення, чи є особливі ракові клітини МТАзадефіцитними, за допомогою тестів на присутність у клітинах протеїну МТАзи Кращим практичним тестом є визначення наявності у клітинах гомозиготних делецій гена, який кодує протеїн МТАзи У іншому аспекті, винахід передбачає спосіб лікування МТАза-дефіцитних ракових пухлин дією на МТАза-дефіцитні клітини терапевтичне ефективної КІЛЬКОСТІ інпбітора синтезу пурину de novo, який інгібує активність АСС, краще L-аланозина Інгібітори АСС винаходу можуть бути введені будь-яким КЛІНІЧНО прийнятним шляхом, але краще їх вводити шляхом тривалої інфузм при концентрації препарату нижче максимальних пристосованих доз для того, щоб подовжити бажану активність інпбітора та зменшити токсичність щодо тканин хазяїна Також запропоновані комплекти препаратів для використання при застосуванні вищезгаданих способів винаходу, які включають реагенти для виконання тесту винаходу на дефіцит МТАзи, а також фармацевтичні композиції шпбітора АСС, краще L-аланозину та/або його активного метаболіту, L-аланозинілу AICOR Фіг 1 - нуклеотидна ПОСЛІДОВНІСТЬ геномної МТАзи (Посл Ід № 1), де екзони підкреслені Фіг 2 - графік, що відображає вплив in vivo Lаланозину після тривалої інфузм на відновлені МТАза-компетентні та МТАза-дефіцитні великі клітини ксенотрансплантантів легеневих ракових пухлин людини у оголених мишей Фіг 3 - графік, що відображає in vivo вплив Lаланозину після тривалої інфузм на відновлені МТАза-дефіцитні клітини ксенотрансплантантів людини з гострим лімфобластомним лейкозом у оголених мишей Фіг 4 та 5 (А-В) - графіки, що відображають вплив різних екзогенних субстратів МТАзи у МТАза-компетентні або МТАза-дефіцитні КЛІТИННІ ЛІНІЇ ПІСЛЯ лікування L-аланозином Фіг 6 - схематичне зображення внутрішньоклітинних метаболічних шляхів вироблення АМФ Фіг 7 - графік, що відображає вплив in vivo L аланозину після щоденної інєкцм на відновлені МТАза-дефіцитні великі клітини ксенотрансплантантів легеневих ракових пухлин людини у оголених мишей I Метаболічні шляхи внутрішньоклітинного вироблення АМФ Зрозуміти винахід допоможе схема, наведена у Фіг 6, що зображує внутрішньоклітинні метаболічні шляхи вироблення АМФ Взагалі існує три принципових джерела субстратів для внутрішньоклітинного синтезу АМФ Першим є катаболізм метилтюаденозину до аденіну за допомогою МТАзи Цей шлях блокований у МТАза-дефіцитних клітинах Другим є перетворення ІМФ на АМФ шляхом активного впливу АСС або аденілсукцинатліази (АСЛ) На наш час не існує інгібіторів АСЛ активності Однак з втратою АСС активності ІМФ АМФ перетворення зникає Третім є утилізація гіпоксантину у АМФ Проте, завдяки тому, що ІМФ АМФ перетворення відбувається окремо від утилізації гіпоксантину, інгібування АСС катаболізму ІМФ АМФ блокує шлях утилізації гіпоксантину, II Спосіб визначення МТАза-дефіцитних клітин у тестовому зразку пухлин ссавців А Полінуклеотидні реагенти, що використовуються для визначення МТАза-дефіцитних клітин Кращим способом визначення, чи є особлива популяція клітин МТАза-дефіцитною, є тест на гібридизацію для виявлення гомозиготної делецм гена, який кодує МТАзу клітин Процедури скринінгу, які основані на гібридизації нуклеїнових кислот, роблять можливим визначення будь-якої полінуклеотидної ПОСЛІДОВНОСТІ (та, як наслідок, будьякого протеїну, що кодується такою полінуклеотидною ПОСЛІДОВНІСТЮ) у будь-якому організмі, з якого можна взяти відповідну пробу Повний опис способів гібридизації, придатних для використання у винаході, а також опис гена, який кодує МТАзу, сформульовані у звичайно наведеній заявці на патент США з серійним номером 08/176,855, що також розглядається (зареєстровано 29 грудня 1993 року), опис якої включено сюди шляхом посилання разом з будь-якими поправками, які можуть бути тут зроблені Для спрощення посилання полінуклеотидну ПОСЛІДОВНІСТЬ гена, що кодує МТАзу, описано тут як Посл Ід №1 у Списку послідовностей та на Ф і г 1 , де кодуючи ділянки гена підкреслені Геномний полінуклеотид МТАзи розташований на хромосомі 9 у ДІЛЯНЦІ р21 Цікаво, що більшість клітин (дуже велика їх частина у відсотках), які мають гомозиготну делецію гена, що кодує пухлинний супресор ріб, мають також гомозиготну делецію гена, що кодує МТАзу Таким чином, альтернативними шляхами виявлення гомозиготної делецм вищевказаного гена (МТАза) є виявлення гомозиготної делецм попереднього гена (ріб) Для подальшого посилання, див звичайно наведену заявку на патент США з серійним номером 08/227,800, що також знаходиться в стадії розгляду, опис якої включено сюди шляхом посилання Штам Е соїі, який містить повну геномну ДНК для МТАзи щурів, був внесений в Американський банк типових культур, Роквілль, штат Мариленд, 47465 поштовим переказом до 29 грудня 1993 року та був ВІДПОВІДНО помічений по групам Позначками № 55536, 55537, 55538, 55539 та 55540 Не існує гарантій, що цей внесок не може бути, навмисне або випадково, кимось використаний для здійснення винаходу Однак внесок буде підтриманий у життєздатній формі протягом будь-якого періоду, під час якого він може вимагатися патентними законами, згідно з якими розглядається цей опис Для визначення, чи був ген МТАзи гомозиготно делетований з клітин, тестований зразок клітин отримують з організму хазяїна Такий зразок може містити рідину або клітини, наприклад, з тканини хазяїна або його пухлини Ці зразки отримують, застосовуючи способи, ВІДОМІ у КЛІНІЧНИЙ галузі, наприклад клітини пухлин можуть бути отримані за допомогою біопсії або хірургічної резекції Краще, якщо зразки клітин істотно ВІЛЬНІ ВІД "забруднювачів", тобто інших клітин, протеїнів та схожих компонентів, які могли б фальсифікувати тест Наприклад, у випадку, коли джерелом для тестованого клітинного зразка є тверді пухлини, нормальні неЗЛОЯКІСНІ КЛІТИНІ та МТАза, яка може бути виділена з нормальних клітин, отримані під час процедури отримання біологічного зразка, будуть вважатися забруднювачами Такі забруднювачі можуть бути видалені загальновживаними способами очистки, наприклад афінною хроматографією з анти-МТАза антитілом Завдяки тому, що винахід направлений на визначення присутності або відсутності цього гена у зразку клітин, які вважаються МТАза-негативними, нуклеїнові кислоти у зразку краще ампліфікувати для підвищення чутливості способу Ця ампліфікація краще виконується за допомогою полімеразноі ланцюгової реакції (ПЛР), хоча ланцюгова реакція на етапі полімеризації не є абсолютно необхідною Нуклеїнова кислота, ампліфікована у зразку, буде складатися з геномної ДНК або ДНК природного типу, що, як очікується, буде нормально кодувати МТАзу, якщо вона присутня у зразку (див Посл Ід № 1) Вважається, що ДНК, яка кодує МТАзу (далі - "ДНК-мішень"), є присутньою у всіх нормальних клітинах ссавців, включаючи клітини людини ДНК, що кодує геномну МТАзу та буде використовуватися як контроль або джерело олігонуклеотидних проб та праймерів, може бути виділена згідно зі способом, відомим у галузі як спосіб, описаний Mamatis, et al , Molecular Cloning A Laboratory Manual (Cold Spring Harbor Laboratory, 1982) Приклад, наведений тут, демонструє виділення клону гена геномної МТАзи людини, в якому бібліотека генів космід піддається скринінгу за допомогою олігонуклеотидної проби кДНК МТАзи (див Приклад III) Як відомо фахівцям у цій галузі, для одержання ДНК, яка кодує МТАзу, можуть бути використані ІНШІ придатні способи Наприклад, вважається, що бібліотека кДНК, що містить певний полінуклеотид, може піддаватися скринінгу ІН'ЄКЦІЄЮ різних мРНК, що походять від кДНК ооцитів, залишаючи достатньо часу для експресії генних продуктів кДНК, та тестуванню на присутність бажаного кДНК-експресійного продукту, використовуючи, наприклад, специфічні антиті ла для пептиду, який кодується певним полінуклеотидом, або проби для повторного аналізу та тканинної експресії головних характеристик пептиду, що кодується певним полінуклеотидом В альтернативному варіанті бібліотека кДНК може побічно піддаватися скринінгу для експресії певних пептидів (наприклад, протеїну МТАзи), що мають принаймні один епітоп, використовуючи для цього антитіла, специфічні для таких пептидів Такі антитіла можуть бути отримані політонально або моноклонально та використані для визначення експреси продукту, що вказує на присутність певної кДНК (див посилання на Maniatis, et al , Molecular Cloning A Laboratory Manual (Cold Spring Harbor Lab, New York, 1982)) Полінуклеотиди, які використовуються як контроль, проби або праймери у контексті цього винаходу, також можуть бути синтезовані способами та на устаткуванні для синтезу нуклеїнових кислот, які добре ВІДОМІ у галузі Як посилання див Ausubel, et al , Current Protocols in Molecular Biology, Chs 2 та 4 (Wiley Interscience, 1989) Б Ампліфікація геномної ДНК, яка кодує МТАзу, та тест на гібридизацію Для підвищення чутливості гібридизаційних тестів винаходу при тестуванні клітинного зразка краще створювати такі умови, які сприяють селективній ампліфікації нуклеїнової кислоти-мішені Дуже добре, якщо нуклеїнова кислота-мішень є полінуклеотидною частиною гена, який кодує МТАзу (наприклад, "полінуклеотид-мішень") Кращим засобом ампліфікування полінуклеотидумішені є ПЛР ПЛР являє собою спосіб ферментативного синтезу in vitro специфічних послідовностей ДНК або РНК, використовуючи олігонуклеотидні праймери, що гібридизуються з ПОСЛІДОВНІСТЮ специфічної нуклеїнової кислоти, та розташовуються на кінцях певної ділянки нуклеїнової кислоті-мішені Повторювані сери циклів матричної денатурації, гібридизації праймера та ферментативного видовження гібридизованих праймерів приводять до експоненційного накопичення фрагментів специфічної нуклеїнової кислоти, які можна визначити по праймерам на 5' КІНЦІ Кінцеві продукти (ПЛР-продукти), що були синтезовані в одному циклі, є матрицями для слідуючих циклів, отже, КІЛЬКІСТЬ копій нуклеїнових кислот-мішеней приблизно подвоюється у кожному циклі Основні способи проведення ПЛР описані у Патенті США 4,683,195 та 4,683,202 у Mulhs, et al , описи яких включені сюди як приклади загальноприйнятих способів для виконання ПЛР Проте винахід не обмежується використанням способів ПЛР, які розглядаються у патенті '202 Mulhs, et al Этого часу, як почали застосовувати спосіб Mulhs, et al , було розроблено багато тестів, основаних на ПЛР, при виконанні яких використовують модифікації цього способу Ці модифікації добре ВІДОМІ у галузі та не будуть, таким чином, описані тут в деталях Проте, з метою зображення обсягу способів у цій галузі, кілька цих модифікацій описані нижче Способи ПЛР, які забезпечують внутрішній ампліфікаційний стандарт за допомогою конкурентної матриці, яка відрізняється від нуклеїнової кис 47465 8 лоти-мішені ПОСЛІДОВНІСТЮ та розмірами, описані в Proc Natl Acad Sci USA (1990) 87 2725-2729 (Gilhland, et al , authors) Інший спосіб для виконання "конкурентної" ПЛР, для якого використовуються матриці, що відрізняються ПОСЛІДОВНІСТЮ, а не розміром, описано в Nuc Acids Res , 21 34693472, (1993), (Kohsaka, et al , authors) Цей спосіб є найкращим способом для використання у технологи фермент-зв'язаного імуносорбентного тесту (ELISA) для аналізу ампліфікованої нуклеїнової кислоти (кислот) Неконкурентний спосіб ПЛР, який використовує сайт-специфічні олігонуклеотиди для визначення мутації або поліморфізму генів, який може також бути застосований до способу винаходу, описано в Proc Natl Acad Sci USA (1989) 86 6230-6234 (Saiki, et al , authors) Кожний з цих способів має переваги у використанні гібридизаційних проб, які допомагають уникнути фальшивих позитивних результатів, отриманих від будьякої неспецифічної ампліфікації, яка може відбуватися протягом ПЛР Для більш глибшого ознайомлення з вказаною темою фахівці у цій галузі можуть за бажанням звернутися до Inms, et al, "Optimization PCR's", PCR Protocols A Guide to Methods and Applications (Acad Press, 1990) Ця публікація являє собою підсумок способів впливу на специфічність, точність та вихід бажаних продуктів ПЛР Вибираються олігонуклеотидні праймери (принаймні одна пара праймерів), які будуть специфічно гібридизуватися до невеликої ділянки пар основ на кожному КІНЦІ (тобто 5' та 3') полінуклеотидумішені МТАзи (тобто "кінцевих послідовностей") Фахівці у цій галузі можуть вибрати придатні праймери без зайвих експериментів, спираючись на інформацію про полінуклеотидні ПОСЛІДОВНОСТІ, визначені у Списку послідовностей, що зазначений тутякПосл Ід № 1 , т а н а Ф і г 1 Щодо конструювання праймера, важливо, щоб праймери не мали в своєму складі такі комплементарні основи, які можуть гібридизуватися самі з собою Для того, щоб уникнути ампліфікації будьякого забруднювача, який може бути присутній у зразку, праймери краще конструювати так, щоб з'єднувати екзони (які для гена МТАзи підкреслені на Фіг 1) Як відмічено вище, для того, щоб адекватно ампліфікувати нуклеїнову кислоту у зразку, зовсім необов'язково використовувати ланцюгову реакцію на цьому етапі полімеризації Наприклад, якщо вищеописаний спосіб Kohsaka, et al , реалізований так, що спочатку виконується етап полімеризації на твердій фазі, а потім гібридизація зі специфічною пробою полінуклеотиду-мішені, чутливість тесту буде така, що достатньо буде простої полімеризації полінуклеотиду-мішені Після етапу ампліфікації ПЛР-продукти тестують на присутність у зразку гена, який кодує МТАзу Краще, якщо дволанцюгові ПЛР-продукти будуть зв'язані з твердою фазою, тоді їх ланцюги можуть бути відділені денатурацією, дозволяючи, таким чином, пробам із специфічними послідовностями гібридизуватися із зв'язаним "невизначеним" ланцюгом ПЛР-продукту для остаточного визначення гена, як це описано вище у Kohsaka, et а/ В альтернативному варіанті ПЛР-продукти мо 47465 жуть бути вилучені з реакційного середовища та відділені від ампліфікаційної суміші перед додаванням проб для гібридизації з дволанцюговими ПЛР-продуктами При останньому ПІДХОДІ ПЛРпродукти ВІДДІЛЯЮТЬ від ампліфікаційної суміші згідно зі способами, відомими у галузі, що відповідають особливому способу визначення гена, застосовують, наприклад, гелеву ексклюзію, електрофорез або афінну хроматографію Виявити ампліфікований продукт можна за допомогою гібридизаційних проб, які стабільно зв'язані з міткою Мітка є речовиною, яка може бути ковалентно приєднана або міцно зв'язана з пробою нуклеїнової кислоти, що дасть можливість виявити пробу Наприклад, міткою може бути радіоізотоп, ферментний субстрат або інгібітор, фермент, контрастна речовина (включаючи колоїдні метали), флуоресценти, хемілюмінесцентні молекули, ліпосоми, що містять будь-які вищевказані мітки, або специфічно зв'язані парні елементи Придатна мітка не втрачає своїх властивостей під час ампліфікації Вченим-діагностам добре ВІДОМІ придатні мітки, які легко можна визначити при виконанні тестів in vitro Наприклад, придатні радіоізотопи для ви3 125 131 32 14 користання in vitro включають Н, І, І, Р, С, 35 S Ампліфіковані фрагменти, мічені радіоізотопом, можуть бути визначені безпосередньо гамалічильником або авторадіографічною денситометрією, Саузерн-блоттингом ампліфікованих фрагментів, поєднаний з денсітометрією Прикладами придатних хемілюмінесцентних молекул є акридини або люміноли Послідовності-мішені, гібридизовані з пробами, які є похідними акридинового естеру, захищені від гідролізу штеркаляцією Прикладами придатних флуоресцентів є флуоресцеїн, фікобіліпротеш, рідкі земельні хелати, дансил або родамін Прикладами придатних ферментних субстратів або інгібіторів є сполуки, які специфічно зв'язуються з пероксидазою хріну, глюкозоксидазою, глюкозо-6-фосфатдепдрогеназою, ргалактозидазою, піруваткіназою або лужною фосфатазою ацетилхолінестерази Прикладами контрастних речовин є колоїдне золото або магнитні частинки Специфічна зв'язуюча пара складається з двох різних молекул, де одна з молекул має таку ділянку на поверхні або в порах, що специфічно зв'язується з певною площинною та полярною структурою іншої молекули Члени специфічної зв'язуючої пари часто відносяться як ліганд та рецептор, або ліганд та анти-ліганд Наприклад, якщо рецептор є антитілом, ліганд є ВІДПОВІДНИМ антигеном Інші специфічні зв'язуючі пари включають гормон-рецепторні пари, ферментсубстратні пари, бютин-авідинові пари та глікопротеш-рецепторні пари Включення являють собою фрагменти та частини специфічних зв'язуючих пар, які підтримують зв'язуючу специфічність, наприклад фрагменти імуноглобулінів, включаючи Fab-фрагменти та подібні до них Антитіла можуть бути або моноклональні, або політональні Якщо елемент специфічної зв'язуючої пари використовується як мітка, кращою процедурою виділення буде афінна хроматографія 10 Якщо ніякий ампліфікований продукт не може бути визначений за допомогою тесту, описаного вище, це є показником присутності у зразку МТАза-дефіцитих клітин Завдяки тому, що нормальні (тобто незлоякісні) клітини завжди мають ген, який кодує МТАзу, у визначних кількостях (навіть із втратою однієї алелі), можна зробити висновок, що якщо в клітинах не вистачає гена, який кодує МТАзу (тобто існує гомозиготна делеція гена), тоді в цих клітинах також не вистачає як каталітично активної, так і каталітично неактивної МТАзи Проте, якщо потрібно, зразок може бути підданий прескринінгу на каталітичну активність МТАзи, використовуючи спосіб, описаний Seidenfeld, et al , Biochem Biophys Res Coimnun , 95 1861-1866 (1980), див Також Приклад 1, нижче) Тест винаходу буде тоді використаний для визначення, чи є ген, який кодує МТАзу, присутнім у клітинному зразку Зразок також може бути тестованим у присутності каталітично активного або неактивного протеїну для скринінгу клітинних забруднювачів у зразку, тобто незлоякісних клітин Придатний імунотест для альтернативного використання (тобто замість гібридизаційного тесту), описаного у Nobon, et al , Cancer Res 53 10981101 (1991) та сформульовані в звичайно наведеній основній Патентній заявці США, серія № 08/177,855, що також розглядається, зареєстрованій 29 грудня 1993 року, опис якої включено як посилання, В МТАза-дефіцитні клітини Використовуючи способи тестування, описані вище, такі наступні первинні пухлини людини були визначені як МТАза-дефіцитні Зрозуміло, що цей список є ілюстративним, але не виключним, щодо типів раку, які можуть бути визначені як МТАзадефіцитні, використовуючи способи, що застосовуються при виконанні описаних тестів - Гострий лімфобластомний лейкоз (приблизно 80% випадків) - Гліома (приблизно 67% випадків) -Злоякісна пгантоклітинна пухлина легенів (приблизно 16% випадків) - Пухлини оболонки сечовивідних органів (наприклад, рак сечового міхура, сфера розповсюдження залежить від типу пухлин) Спираючись на ці дані, МТАза-дефіцит варто очікувати від пацієнтів, що страждають на вказані захворювання Таким чином, КЛІТИННІ зразки, взяті у таких пацієнтів, повинні бути протестовані на МТАза-дефіцитність для визначення ефективності терапевтичного лікування за способом цього винаходу КЛІТИННІ зразки, одержані від інших пацієнтів, що страждають на рак, повинні бути протестовані на МТАза-дефіцитність для КЛІНІЧНОГО підтвердження діагнозу Наприклад, зразки первинних пухлин виявились МТАза-дефіцитними (тобто рак є "МТАза-компетентним") у пацієнтів, що страждають на - Рак грудей - Рак товстої кишки - Рак голови та шиї - Меланому - Рак нирок - Нелімфобластомний лейкоз дорослих 12 11 47465 - Деякі гострі лейкози (дорослих та дітей) клітин, через це зменшується токсичний вплив на неушкоджені клітини У КЛІНІЧНИХ випадках використовували Lаланозин для лікування вище-перелічених МТАзаНезлоякісні МТАза-компетентні клітини також компетентних злоякісних пухлин, але без особлиможуть бути захищені від будь-яких ефектів дії вого успіху АСС-шпбіторів через введення МТА або аналогічного придатного субстрату для використання у III Метод лікування МТАза-дефіцитних клітин синтезі аденіну До придатних сполук належать А Фармакологічні та ТОКСИКОЛОГІЧНІ характериМТА, 2'-5'дидезоксіаденозин, 5'-дезоксіаденозин, стики для L-аланозину 2'-дезокси-5-дезокси-5'метилтюаденозин (див, У приматів приблизно 75% L-аланозину видінаприклад, Приклад II) Проте треба зазначити, що ляється з сечею протягом 24 годин, головним чиМТАза-компетентні клітини здатні виробляти аденом у вигляді нуклеозидних форм L-аланозинілунін в результаті метаболізму метилтюаденозину ІМФ та L-anaHO3HHiny-AICOR У людини вивільдля поповнення клітинного пулу АМФ та, таким нення з плазми після внутрішньовенного введення = чином, не очікується зменшення АМФ до такого ж є двофазним, з t-i/2 а 14 хвилин та t-i/2 p = 99 хви11 рівня, як і в МТАза-дефіцитних клітинах лин (де "ti/2 - це період напіввиведення, та час (t) дається наближено) Винахід повністю описаний, його способи ілюстровані прикладами, наведеними нижче ЗрозуУ попередніх КЛІНІЧНИХ випадках токсичність міло, що приклади не обмежують обсяг винаходу, обмежувалася дозою і проявлялася впливом на який визначено формулою, яка додається У принирки, стоматитом, езофаптом та, з меншою часкладах використані стандартні абревіатури, такі як тотою, мієлосупресія, головний біль, нудота та "мл" (мілілітр), "год" (година) та "мг" (міліграм) ппо- або гіпертензія Ниркова токсичність зустрічається при введенні одиничної дози у вигляді боПриклад І люсу більше ніж 4г/м2 ваги тіла Крім того, обидва Вплив L-аланозину in vivo на МТАза-дефіцити і педіатричні пацієнти, які отримали найвищі дози, та МТАза-компетентні пухлинні ксенотрансплантатобто біля 350мг/м2 ваги тіла на день у окремих нти людини після тривалої інфузм дозах, піддалися ушкодженню печінки Стоматит Для оцінювання впливу L-аланозину in vivo на та езофагіт трапляються після тривалого застосуВІДОМІ МТАза-дефіцитні пухлини людини після вання болюсних доз Інші реакції були специфічтривалої інфузм у порівнянні з впливом цих ЛІКІВ на ними для кожного пацієнта ВІДОМІ МТАза-компетентні пухлини людини 2 х 106 МТАза-дефіцитних Н292 NSCLC клітин та МТАзаПід час першого етапу двофазного тесту докомпетентних Calu-6 клітин пухлин (у 0,3мл носія з рослим, що страждають на гострий нелімфоблас50% MATRIGEL™) вводили у правий та лівий бік томний лейкоз, призначали тривалу інфузію преВІДПОВІДНО 8 Balb/C оголеним мишам з видаленим парата у дозі приблизно 125мг/м2 ваги тіла тимусом MATRIGEL™ приймає форму твердої протягом 5 днів Дозообмежуючою токсичністю матриці in vivo, яку вимірювали у кожної миші в було запалення слизової оболонки перший день інфузм для контролю за розміром Б Введення АСС інгібіторів до організму хазяпухлини (яка ресорбується через 14 діб) Для поїна дальшого порівняння 107 СЕМ T-ALL (МТАзаСсавців, наприклад людей, що страждають від дефіцитний гострий лімфобластомний лейкоз) раку, яких можна віднести до МТАза-дефіцитних клітин у носи з 50% MATRIGEI™ вводили у правий згідно з тестом на дефіцит МТАзи винаходу, лікубік 8 інших оголених мишей Багато клітин були ють терапевтичне ефективною дозою АССотримані з комерційне доступних клітинних ЛІНІЙ шпбітора, такого як L-аланозин, L-аланозинілвід АТСС, придбаних згідно з АТСС Accession Nos AICOR або хадацидин, краще першим Так "тераCRL-1848TaHTB-56 певтичне ефективна доза" викликає об'єктивну пухлинну ВІДПОВІДЬ у пацієнтів, де пухлинна ВІДПОВІДЬ є зупинкою або регресією у рості, що визначається за КЛІНІЧНО прийнятими стандартами (див , наприклад, Eagan, et al , Cancer, 44 1125-1128, 1979 [опис якого включено сюди шляхом посилання] та Публічні звіти про результати проведених (особливо у Eagan, et al) в КЛІНІЧНИХ умовах тестів під IND#14,247 (Food and Drug Administration)) Використовуючи ці дані, визначення терапевтичне ефективної дози АСС-шпбіторів, що використовуються у винаході для зменшення КІЛЬКОСТІ внутрішньоклітинної АМФ, може бути легко зроблено звичайними фахівцями в ОНКОЛОГІЧНІЙ галузі Взагалі щоденне введення або тривала інфузія АСС-шпбіторів в дозах, менших ніж ті, що викликають негативну реакцію, є найкращим терапевтичним призначенням для антиметабол ічних ЛІКІВ Можна сподіватися, що завдяки особливій чутливості МТАза-дефіцитних клітин до цього способу лікування, будуть потрібні менші дози, ніж ті, які КЛІНІЧНО тестувалися при застосуванні Lаланозину для лікування МТАза-компетентних Наступного дня 4 мишам з двобічними NSCLC та 4 мишам з T-ALL щепленнями імплантували під шкіру ALZET™ 1007D за допомогою осмотичної шфузюнної помпи у спину на відстані від місця введення клітин пухлин Помпи заповнювали Lаланозином у концентрації бОмг/кг для щоденної тривалої інфузм Оскільки після 7 днів при дозі бОмг/кг ніякої токсичності не спостерігалося, помпи видаляли та замінювали помпами, які містили Lаланозин у дозі 90мг/кг на день з розрахунку на 7 додаткових днів Як показано на Фіг 2 та на Фіг 3, введений Lаланозин викликав зменшення сталих МТАзадефіцитних NSCLC ("H292 L-alan Тх", Фіг 2) та ТALL ("Піддослідна миша", Фіг 3) прищепленого ксенотрансплантанту у імунодефіцити их хазяївмишей та запобігання росту МТАза-дефіцитних NSCLC або T-ALL ксенотрансплантантів у імунодефіцитних хазяїв-мишей у порівнянні з ПІДДОСЛІДНИМИ МТАза-компетентними NSCLC клітинами та контрольними МТАза-дефіцитними клітинами Зокрема, сталі МТАза-компетентні пухлини ростуть 14 13 47465 швидко, незважаючи на L-аланозин, як у випадку з МТАза-компетентні (Calu-6, ATCC Designation No контрольними МТАР-дефіцитними NSCLC або ТНТВ-56) та МТАза-дефіцитні клітини (А549, АТСС ALL ксенотрансплантантами Designation No CCL-185) Геномну ДНК виділяли, як описано у Прикладі III, та 1 мікрограм и викориПриклад II стовували у ПЛР Захист МТАза-компетентних здорових клітин субстрата МТАзи Ампліфікацію проводили протягом 40 циклів, як описано вище, кожний цикл складався з денаВибіркова нездатність МТАза-дефіцитних турації (92°С, 1хв), гібридизації (55°С, 1хв) та виNSCLC клітин проліферувати у присутності Lдовження (72°С, 1/2хв) ПЛР-продукти аналізували аланозину (у 40мкмоль), незважаючи на додаванна 1,5% агарозному гелі У КЛІТИННІЙ ЛІНІЇ, яка віня до кожної культури субстрату МТАзи метилтюдома як МТАза-дефіцитна, не було визначено аденозину (МТА), була підтвержена порівнянням МТАзи, в той час як у МТАза-компетентній КЛІТИНдвох клітинних ЛІНІЙ, МТАза-компетентних Calu-6 НІЙ ЛІНІЇ МТАзу було визначено та МТАза-дефіцитних Н292 (Фіг 4) Контрольні культури, які містять аденін (tAde), проліферують Приклад V незважаючи на L-аланозин, підтверджуючи, що Інгібування L-аланозину МТАР-дефіцитних пувибіркова токсичність пояснюється нездатністю хлин після щоденної ін'єкції оголеним мишам із МТАза-дефіцитних клітин перетворювати МТА на сталим пухлинним ксенотрансплантантом аденін 107 А549 або А427 NSCLC клітин уводили підшкірно 6-тижневим старим Balb/c оголеним мишам При подальшому порівнянні додавання МТА з видаленим тимусом Коли пухлини досягали або МТА-субстратного аналога S'приблизно 0,4см у діаметрі, лікування починалося дезоксіаденозину призводить до зменшення росту з L-аланозину або фізіологічного розчину інтрапелише для МТАза-компетентних А427 клітин та ритональною ІН'ЄКЦІЄЮ КОЖНІ 12 годин MOLT-4 клітин, де ріст МТАза-дефіцитних А549 та СЕМ клітин залишається інгібованим (Фіг5А та Миші отримували 15мг/кг на ІН'ЄКЦІЮ протягом 5В) Завдяки тому, що МТА є зворотним інгібіто7 днів, потім 22,5мг/кг протягом 5 днів Розмір пухром синтетази сперми, деякі КЛІТИННІ ЛІНІЇ, такі як лини вимірювали по перпендикулярним діаметMOLT-4, інгібуються при високих концентраціях рам Результати для ПІДДОСЛІДНИХ мишей (п = 6 Це призводить до підвищення концентрації МТА у [розмір зразка] для кожного типу клітин) проти кондвофазній кривій відновлення росту (Фіг 5А) тролю (п = 4 [розмір зразка] для кожного клітинного типу) показано на Фіг 7 Приклад III Клонування та часткова характеристика геноПри будь-якій дозі ніякої токсичності не спомного клону МТАзи стерігали (ПІДДОСЛІДНІ мали стабільну вагу протягом досліду) L-аланозин, уведений подвійною щоГеномний клон МТАзи людини виділяли так денною болюсною ІН'ЄКЦІЄЮ, призвів до зменшення Банк космідних генів, побудованих з ДНК плаценти у розмірі сталих МТАР-дефіцитних пухлин, але людини (Clontech), піддавали скринінгу, викорисніяк не впливав на ріст МТАР-позитивних клітин товуючи генну пробу кДНК МТАзи, Not I/ECORI фрагмент з субклону МТАР-7 Трансформовані Огляд послідовностей клітини Е соїі з банку висівали у LB чашки, які місПОСЛІДОВНІСТЬ ІД № 1 Є геномним клоном для тили ампіцилін (50мг/л), з густиною колонії 1-2 х метилтюаденозинфосфорилази (МТАза) 4 10 /135 х 15мм чашку Список послідовністей (1) Загальна інформація Далі виконували такі процедури 3 ПІВМІЛЬЙОН(I) Подавець Магістри Каліфорнійського уніноі колони, виділяли одиничну позитивну колонію, верситету названу як МТАР-10, та частково знімали характе(II) Назва винаходу Метод інгібування аденілристики аналізом ПЛР та прямим визначенням сукцинатсинтетазної активності у злоякісних клітиПОСЛІДОВНОСТІ Два праймери, "визначений" олігонах з дефіцитом метилтюаденозинфосфорилази нуклеотид, розташований на 120 пар основ вище стоп-кодону, та "невизначений" олігонуклеотид, (III) Номер ПОСЛІДОВНОСТІ 1 розташований на 20 пар основ нижче стоп-кодону, (IV) Адреса для кореспонденції синтезували та використовували під час аналізу (A) Адресат Robbms, Berliner & Carson ПЛР ПЛР проводили протягом 25 циклів, кожний (B) Вулиця 201 N Figueroa Street, 5th Floor цикл складався з денатурації (92°С, 1хв), гібриди(C) Місто Лос-Анжелес зації (55°С, 2хв) та видовження (72°С, 5хв), ПЛР(D) Штат Каліфорнія продукти були відділені на 0,8% агарозному гелі (E) Країна США (F) Код 90012-2628 Розташування екзонів, визначене у генів МТА(V) Комп'ютерна форма зи вищеописаним способом, позначається на Фіг 1 (A) Тип носія Гнучкий диск підкресленням (B) Комп'ютер IBM PC-сумісний Приклад IV (C) Операційна система PC-DOS/MS-DOS Застосування послідовність-специфічних олі(D) Програмне забезпечення Patentm Release гонуклеотидів МТАзи до злоякісних клітинних ЛІНІЙ »1 0, Version #1 25 для визначення присутності або відсутності в них (VI) Дані про сучасну заявку МТАзи (A) Номер заявки Для визначення корисності олігонуклеотидних (B) Дата реєстрації проб, отриманих від геномного клону МТАзи так, (C) Класифікація як описано у Прикладі IV (Посл Ід № 1), ПЛР за(VIII) Інформація про повіреного/агента стосовували до кількох ракових легеневих клітин(А) Ім'я Berliner, Robert них ЛІНІЙ людини, про які відомо, що вони містять 15 (B) Репстраційний номер 20,121 (C) Номер посилання/виписки 5555-423 (IX) Телекомунікаційна інформація (A) Телефон (213)977-1001 (B) Телефакс (213)977-1003 (2) Інформація про Поел Ід № 1 16 47465 Т Ш М Л К С АДМЕСІТЛС TGCTCACISC CATACCTCAG ATAGGSICCA CAGAAIGGTC 1740 АОАЯАСССГС CAIftACCTGfL AGQIAAGIGC ACCCAIGGAC AAICAGGCAT GICIGIAGAC 18Q0 TCTCTAITGT СПСПГТСТ IACITGCAT1 TCACCTTIGG ІССГСАТйїА TTTTITGCCA Ш0 GCCTWATGI IITCAACAAG TIITTOTGAC ДТСТАСЇАСТ АССЙТАССАА CCACTTGTGA 1930 АйСІОАОІАС ТСІІАІТПС ITGGCTeGTA GtbCAGAHKH М Ш Н Н Ы Ш Ш NMAATAAACA 1980 ATAATCCAGG CTGCGCTGUI ATCGCHATAA GTGAT1ATCA CAACAATGET CTCAGATAAG Z100 AGTTCCAGAT CIGCTTAGTG ААСЇТСАААА TITTC1GIAG А А Т П А П Т А А А & Ш А Т С Т 2160 TTCCIGCGIC СІСАСП1СА ICTAGAAAAT CAAAATCIGT TTTTITTTJT AACAAACATC (A) Довжина 2763 пар основ (B) Тип нуклеїнова кислота (C) Ланцюг одиничний (D) Топологія лінійна (II) Тип молекули ДНК (геномна) (VII) Джерело (В) Клон Метиладенозинфосфатаза (IX) Властивість (A) Ім'я/ключ CDS (B) Знаходження 1 2763 2O4Q EATIAITARC CTCACtTTAC AGGAMGGGA eGTGAGGAAC CAACAGTTTA GAGTACCCGA (I) Характеристика ПОСЛІДОВНОСТІ 2£2G tCAGTAAUA CCCCAACATG TGAA7ATCAC T5CCTCCTH СТІССТГТСА GAATAtCUCC 2280 CACITTTC1G ТГТТАТІАСС AAGACATtAA AGIAGCATGG CICCCCAGGA GAAAAGAAGA 3340 CAIICTAAIT ССАЕІСАТП TGGGAATICC TGCTTAACIJ GAAAAAAATA TCGGAMGAC H400 ATGCAGCTIT CAIGCCCTTG CCIA7CAAAG AC1ATGTTGT AAGAAAGACA AGACAIIGTG 2460 IGTATAGAGA CTCCICAATG AIT1AGACAA CITCAAAA.TA CAEAAGAAAA ЄСАААТСАСТ HS20 ACTSACAiCT GGGAAAAUT АТТАСАПП AAGGGGGAAA AAAAACCCCA CCATTCTCI1 £5B0 CrCCCCCtAI TAA*ITTRCA ACAnTAAAGG 6IGKAG5GTA А Ї С Т С Ї А Ш ТССГАТАСТЕ і к и SEQUENCE ш с к ї Р П О К : sea и ТПАТАСАСА ЄСАЇ«МШ Ш М Ш С * ЕТАСЕЗТЕЇС ТСТВССШС ТДСАШПб АЛАСАбСШ ШГТТСГОШ ЛАЇССШС* ШЯШЗП 37DD GTACRGTATr TCTGGAGGGC AAIIIGGIAA ААГЕСАТСАА AAGACIIAAA AATACGGACG 60 ТАСТТІТИТ ЇЄСТСАСТСЕ 26Л0 CCAAAGAATS IGfiGGAAGAA AIGGGACIC! TIGGTfAITT ATIGATGCGA CTGTAAATtG п о п і 2760 120 ЛСТСббТТСЇ A G G C C CTSTGTtACT aGTCGTCAT T6CIASSAG* АГОІСТІЛЯТ E A AEC 180 ТТТТТСЇАС* СЇСІЛЄСАЄ* MSCMTTSG HiGIGTACTC АЇСАССЄ6Т) АШАТТШ 24В ІСЇЕТШТС cmSfiGAtG OAGGC&Ga СМШМТС СШШЛС6 TCAACTACCft 500 TGGUCntCA AECMGAECS СЇЄТДСДШ CICJtIAOICft ССЙСДШТС 350 ftQGGAGOASA ТТОМЙЖ&ВЕ СШАТТСТС ЛТТАТГШС ACTTOHTG* «В 201 TTTATACAGA їйсдтдттта TAGTnTAGT CGG T A T T GCT AAACAGTTGG G AGCG CT A A T A CG A T M A GG T CC CG GTATG T CA S G G TGA A T GGA T G T T CG 4В0 ІАВЕГТАТСС ACACCMTTG AETCTGCACT ЛАЛ£А£С£ХА ЛТАТТйДССІ ЛОМАААБТТ Ї40 ЫСГСАССДС СКТКСЛШ AIGGAMGST QOCCIFAAM ТМАЙСМАС *АМ*ССТТТ 400 301 G C T T fG G T C Ct A T G TC CG GT T AC CT A T A TA T AC C 6 T T A GGT CTAG6GTCTG TCTGCCACTC AATCCAGTGA ACCTAAATTT AGGAGACCCC CTGTGTTAGT TTTTTCTAGA CTCTA6GAGA ААСААТТГСТ TCTCTCCT1C ШШМСйІС Л Ш З С П Ж ШШШШІ---.СШМЖЖ ЄІСЙТШШЙ. AflTTCATTGA GCTnGTAAA AAAGACCCAA ATGGAAAGAT CANHNMMMNM CAATTGTCTT ATATTGACCT GGCCTTAAAA GTGTGTAGTC mCTSGMG TGGGCTTGGG GTGGCATCTG GCATTTGgrT CCATACCCTC ACTCAAGTrT GCTTTGTATT ATTTCCTGTT ССТААГМТТ TNMMNNNMNN TTAAGTTGTG CATGTGCTAG TATGTTTTGA TAGGTTCTTft ТДОАЙАГСТЙО ТДДОАЛВСТД 251 ЯЮШЮГШ ИАИЗТССАСВ ЕШЄСТТА* ШЇТВГШ СШГСТСЇТ ШЖШН ФІГ 1А G G T CC G GC T A СДССАТС 351 ГТТЕСТТї*! TTTGMGGAC CAaAtMW ССТЄ*ЕГССЇ ТСТЯТИТВС ААЄТатіСТ Ш 501 ТвТССаОЛС GMTGIGC» lATtCCWTG CCIiWGCCGT tTTKECOUl ЛАСЕНШІІУіЕ 720 551 ATTATTGATC GTATCGAfM AGTcrGGAGT CCCTCGGAGfi 601 TTTGCTTTAT ЇІЕТЄ!*СТС ITtCTSGAAG GTCTACC«SA AtAMTCAJE tCGSCITEGG GTGCOLTCIE MO ШУПТЙЕТГ М С С 6 сі2імтбса: аатксссТЕ и і с А д с т к т т а ш і П В АД вт 40! 451 701 *ТЙЙА0ТП АІС6АВ»ЙГГ ЯТІТЕЕТВЇТ Ш Ш * А Т Ї ІИМННКЖШІ Н Н Щ К Н НИ МН МО AAGlCCAGtt ТІААСІІСІС САГСТШІІЯЙ ГАТЕТТПЄ» AGTTTCTCGT ТПТСТТПС WB 851 901 ATAAATCATG CGGAGTGGCC ATGGAGAGTT AAGTGCAGCC 951 ТПТСТТТТС 751 80 f ГМСТІГТІА ТДЕ*«СТЕС ГДМАЛЕСГ* И^СГСЕЕСІ ШЗСТСШ SBStUCAATS 102] CIMCAATCG ACCCACCTCG ІТТіАССТСС СЙЗвЕАОААА ВСТТСАТЄП ЄСбСАССТІШ 10S0 СоетСЯМІС ТТАІСААСДТ еЛСЫСАВТ! CCSGACCTGa ТТСГТССТАА CQWSCtGGA 1U0 1051 *ITTGIT*CG СААЕТЛТШ: Е Х С С Е ШТАТРіСТ КІвЦАЛЕй» ИЖСАВЕМ ЛССАА 1ЇП0 GCWIf*GWS 61АТГСТІП СЇМПИСАІ *Т*СС*Т15И ТІТЕЇСЕЗШ СОШТМЕЗТ 1Й60 ISSS GTGTACGAGA AATTGGCAGA ATGCAAGTTT 1101 CTCTfMCIB ІТТБГТТСТА ТГАССТІДЕТ ПСШША6І СССГГТСІДС Мб&ГШ«А NNNNNNP1NNN GAG6TCGAC6 TAOCTTATCC AGAGGAATTG AGArAAAQTT GACTCACCAQ TAAAAGAAAC АААМССПТ ASITGHJAT ДИИСТ6ТА SHCCmcs МЕСТАМ»* CMWAIQW *ACAA«iWA 13K1 аШАШТП ТАСССМбОЛ Т€*ЕІ*ЕІСЯ АМГАСІДСА ЇТОІШМаї ИАШЄТбї 1МЄ ТМЕЫЙСАТ АСЇА*(»СІГ GGBECItAMK K№HKNHNH B N KW1 H ИІІЇАСССТАС S M I 1H 1S00 ATISWCATI CCCmCUX АСХТАЛАТТГ ШЗІВШСА МСЯТТІЛСЕ CTGIKQCAiW 1И.Э CCTEGAOCTC АОидААТБТ ИТ*Т6*САД ОСАЗІССЯА) ТТТАА6ТТСГ ACTJUCCTK 1630 ITTCTCTAES mcGGICGA CCGSQTETIA MGACXCIOA Д С А Л С АААСС 1680 1001 GGGGACAATG CGGGCAGAAA 1201 SCfiGTAGGTG 1251 C A T G G CAA G T 1301 GCCTTTCTAC 1351 AAGGTMGAA 1401 T A T G G CGA T A BTCftCAATCfi GCTTCATGTT CCGCACCTGG fiGGGCGGATG ТТДТСААСАТ GMTiCTirr GTCTTAACTG AAGGTTITG* CAAAGATCAA AAATAGTACA CTAAGCACAT TTTGTTTCTA AGTTGTTAAT AAGAAAGAAA TTGTAGGCAT ATAGCATGGG TTACGfTAGT ATTTTCTGTA GAGACACTTT GTAGATGTGT TTTCTG6GTG TTCAGAAAGT GTTCCATTGG TACCCAAGGA TGAGAATCAT Об'єм пухлини %, день 0 Об'єм пухлини %, день 0 Життєздатні клітини, % еід контролю C O -fe. J О) СП % Контрольних життєздатних клітин •в m 19 20 47465 ФІГ. 6 Екскреція сечової кислоти Кров Апанозии ФІГ. 7 А427 No їх 300 А427 і-АІап їх A54S No їх 250 А549 L~A!an їх 200150100 5000 12 3 4 5 6 7 День лікування 9 Ю U ДП «Український інститут промислової власності» (Укрпатент) вул Сім'ї Хохлових, 15, м Київ, 04119, Україна ( 0 4 4 ) 4 5 6 - 2 0 - 90 ТОВ "Міжнародний науковий комітет" вул Артема, 77, м Київ, 04050, Україна (044)216-32-71
ДивитисяДодаткова інформація
Назва патенту англійськоюMethod for suppressing the activity of adenylosuccinate synthetase in the methylthioadenosine phosphorylase-depleted cells, a method for suppressing the activity of adenylosuccinate synthetase in mammalian host-cells
Назва патенту російськоюСпособ подавления активности аденилсукцинатсинтетазы в метилтиоаденозинфосфорилаза-дефицитных клетках, способ подавления активности аденилсукцинатсинтетазы в клетках млекопитающих-хозяев
МПК / Мітки
МПК: C12N 15/09, A61K 31/198, A61K 45/00, A61P 35/00, A61K 31/00, G01N 33/50, A61K 31/195, C12Q 1/25
Мітки: клітинах, гальмування, метилтіоаденозинфосфорилаза-дефіцитних, активності, ссавців-хазяїв, аденілсукцинатсинтетази, спосіб
Код посилання
<a href="https://ua.patents.su/10-47465-sposib-galmuvannya-aktivnosti-adenilsukcinatsintetazi-v-metiltioadenozinfosforilaza-deficitnikh-klitinakh-sposib-galmuvannya-aktivnosti-adenilsukcinatsintetazi-v-klitinakh-ssavciv.html" target="_blank" rel="follow" title="База патентів України">Спосіб гальмування активності аденілсукцинатсинтетази в метилтіоаденозинфосфорилаза-дефіцитних клітинах, спосіб гальмування активності аденілсукцинатсинтетази в клітинах ссавців-хазяїв</a>
Індуктор інтерферону в клітинах ссавців, які культивуються
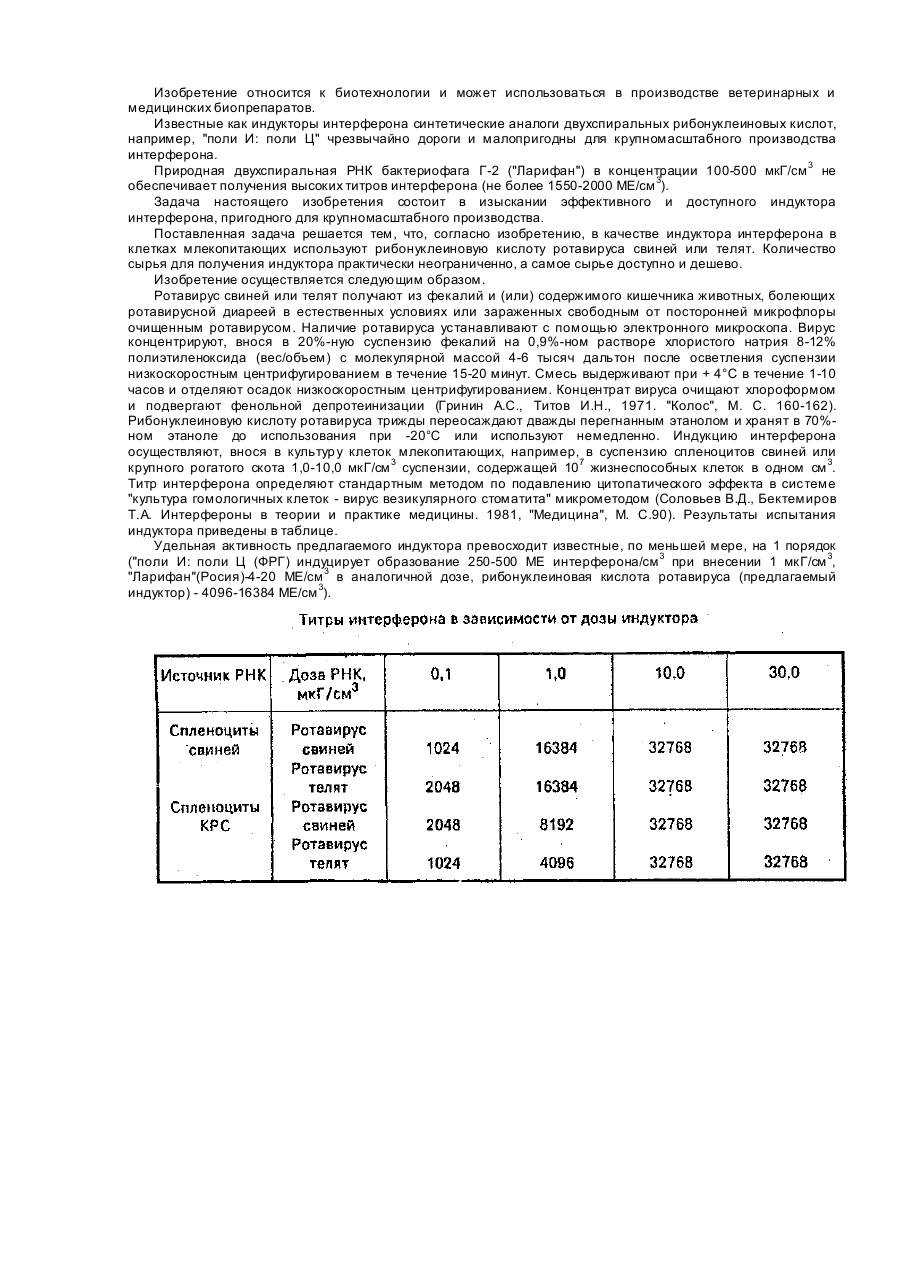
Номер патенту: 10461
Опубліковано: 25.12.1996
Автори: Купчинський Леонід Георгійович, Собко Анатолій Іванович, Карась Олег Якович, Співак Микола Якович, Лазарєв Олексій Павлович
МПК: A61K 38/21
Мітки: ссавців, індуктор, клітинах, інтерферону, культивуються
Формула / Реферат:
Применение рибонуклеиновой кислоты ротавируса свиней или телят в качестве индуктора интерферона в культивируемых клетках млекопитающих.
Спосіб визначення апоптозу в тестованих клітинах організму

Номер патенту: 31503
Опубліковано: 15.12.2000
Автори: Лісовська Наталія Юріївна, Кайряк Ольга Василівна
МПК: G01N 27/26, G01N 33/487
Мітки: тестованих, організму, клітинах, спосіб, визначення, апоптозу
Формула / Реферат:
Спосіб визначення апоптазу в тестованих клітинах організму, шляхом індикації фрагментів ДНК при гель-електрофоретичному аналізі, який відрізняється тим, що диференціює однонитяні фрагменти від двонитяних шляхом реєстрації різної флюоресценції фрагментів ДНК, забарвлених акридиновим оранжевим.
Спосіб оцінки інтенсивності вільнорадикальних реакцій в клітинах тварин

Номер патенту: 29908
Опубліковано: 15.11.2000
Автори: Сокирко Тетяна Олександрівна, Квачов Володимир Григорович, Попова Елеонора Михайлівна
МПК: A61K 31/515
Мітки: реакцій, спосіб, клітинах, вільнорадикальних, оцінки, тварин, інтенсивності
Формула / Реферат:
Спосіб оцінки інтенсивності вільнорадикальних реакцій в клітинах тварин, що включає визначення продуктів перекисного окислення ліпідів за допомогою перекису водню, азиду натрію та тіобарбітурової кислоти з фіксуванням кінцевого метаболіту перекисного окислення ліпідів за оптичною густиною забарвленого продукту реакції, який відрізняється тим, що перекис водню вводять в кількості 4 - 6мл, a оцінку вільнорадикальних реакцій проводять за...
Спосіб боротьби з нозематозом комах-хазяїв ентомофагів

Номер патенту: 29288
Опубліковано: 16.10.2000
Автор: Дрозда Валентин Федорович
МПК: A01K 67/00
Мітки: боротьби, ентомофагів, комах-хазяїв, спосіб, нозематозом
Формула / Реферат:
Спосіб боротьби з нозематозом комах-хозяїв ентомофагів, що включає дію на яйця зернової молі та млинової вогнівки, який відрізняється тим, що незаражену популяцію комах обробляють антигеном із живим збудником нозематозу сумісно з диметилсульфоксидом в період першої доби ембріонального розвитку.
Штам дріжджів рісніа guіllіеrмоndіі sy 6/5, що нагромаджує в клітинах великі кількості рибофлавіну при культивуванні в середовищах з промисловими субстратами

Номер патенту: 24911
Опубліковано: 06.10.1998
Автори: Сенюта Елеонора Зиновіївна, Шавловський Георгій Михайлович, Бабяк Любов Ярославівна, Струговщикова Людмила Петрівна
МПК: C12R 1/645, C12P 25/00, C12N 1/16
Мітки: кількості, дріжджів, рісніа, середовищах, штам, промисловими, клітинах, субстратами, великі, рибофлавіну, guіllіеrмоndіі, культивуванні, нагромаджує
Формула / Реферат:
Штам дріжджів Pichia guilliermondii SY 6/5 здатний до надсинтезу рибофлавіну (95,5 - 324,3мг/л) і нагромадження великих кількостей вітаміну B2 в клітинах (24,3 - 36,3мг/г сухої ваги), штам дріжджів Pichia guilliermondii SY 6/5 зберігається в депозитарії Інституту мікробіології та вірусології НАН України, реєстраційний номер: Pichia guilliermondii IMB Y-5006.
Попередній патент: Механічний дозатор сім’я
Наступний патент: Система контролю та реєстрації параметрів силової установки літального апарата
Випадковий патент: Похідні піридинкарбоксаміду як пестициди