Спосіб одержання l-амінокислот з їх рацемічних n-ацетил-d, l-похідних за допомогою ферментативного розщеплення рацемата за допомогою виділених, рекомбінантних ферментів



Формула / Реферат
1. Спосіб одержання протеїногенних або непротеїногенних L-амінокислот з їх рацемічних N-ацетил-D, L-похідних, який відрізняється тим, що
(a) шляхом ферментативного розщеплення рацемата за допомогою виділених рекомбінантних гіпурат-гідролаз селективно деацетилюються N-ацетил-L-похідні відповідних L-амінокислот, в той час як N-ацетил-D-похідні відповідних D-амінокислот не деацетилюються,
(b) рацемічні N-ацетил- D,L-похідні знаходяться у концентрації від більше 50 мМ до 1500 мМ,
(с) одержані деацетильовані L-амінокислоти препаративно відділяють від недеацетильованих N-ацетил-D-похідних і/або не повністю деацетильованих N-ацетил-L-похідних.
2. Спосіб за пунктом 1, який відрізняється тим, що призначений для одержання L-фосфінотрицину (L-PPT) з N-ацетил-D, L-фосфінотрицину.
3. Спосіб за пунктом 1, який відрізняється тим, що призначений для одержання L-глутамінової кислоти, L-гістидину, L-лейцину, L-глутаміну і/або L-фенілаланіну з їх відповідних N-ацетил-D, L-похідних.
4. Спосіб за будь-яким з пунктів 1-3, який відрізняється тим, що здійснюють енантіоселективне одержання однієї або декількох L-амінокислот з їх відповідних рацемічних N-ацетил-D, L-похідних з ферментами deac 1 з Stenotrophomonas sp. і/або deac 2 з Comamonas acidovorans.
5. Спосіб за будь-яким з пунктів 1-3, який відрізняється тим, що здійснюють енантіоселективне одержання L-фосфінотрицину, L-глутамінової кислоти, L-гістидину, L-лейцину, L-глутаміну і/або L-фенілаланіну з їх відповідних рацемічних N-ацетил-D, L-похідних з ферментами deac 1 з Stenotrophomonas sp. і/або deac 1 з Comamonas acidovorans.
6. Спосіб за будь-яким з пунктів 1-5, який відрізняється тим, що одержані рекомбінантні гіпурат-гідролази використовують в іммобілізованій формі.
7. Спосіб за будь-яким з пунктів 1-6, який відрізняється тим, що температура реакції складає приблизно від 25°С до 65°С.
8. Застосування однієї або декількох гіпурат-гідролаз в способі за будь-яким з пунктів 1-7 для енантіоселективного одержання однієї або декількох L-амінокислот з їх відповідних рацемічних N-ацетил-D, L-похідних.
9. Застосування за п. 8, яке відрізняється тим, що як L-амінокислоти енантіоселективно одержують L-фосфінотрицину, L-глутамінової кислоти, L-гістидину, L-лейцину, L-глутаміну і/або L-фенілаланіну з їх відповідних рацемічних N-ацетил-D, L-похідних.
10. Застосування deac 1 з Stenotrophomonas sp. і/або deac 2 з Comamonas acidovorans в способі за будь-яким з пунктів 1-7 для енантіоселективного одержання однієї або декількох L-амінокислот з їх відповідних рацемічних N-ацетил-D, L-похідних.
11. Застосування за п. 10, яке відрізняється тим, що як L-амінокислоти енантіоселективно одержують L-фосфінотрицину, L-глутамінової кислоти, L-гістидину, L-лейцину, L-глутаміну, L-фенілаланіну з їх відповідних рацемічних N-ацетил-D, L-похідних.
Текст
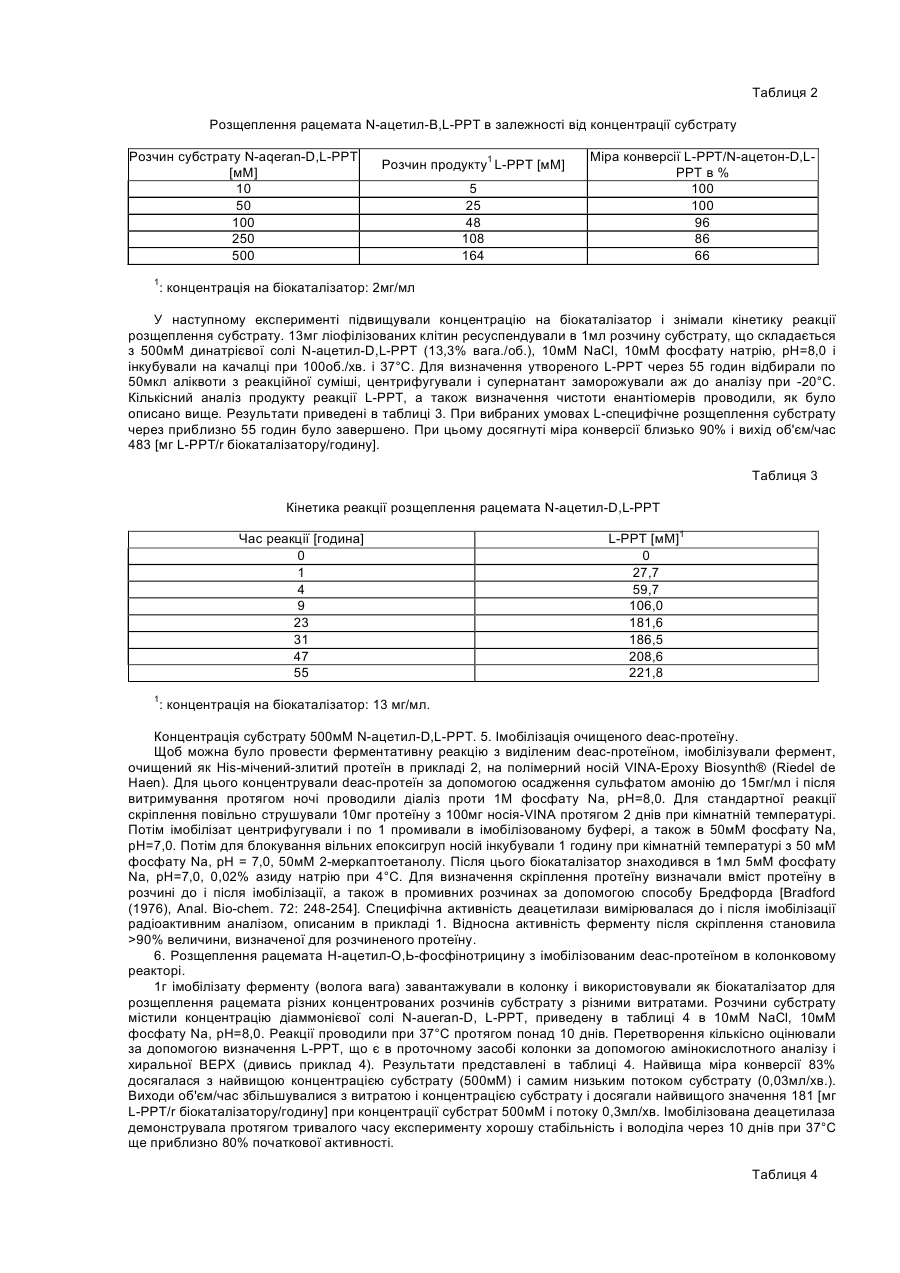
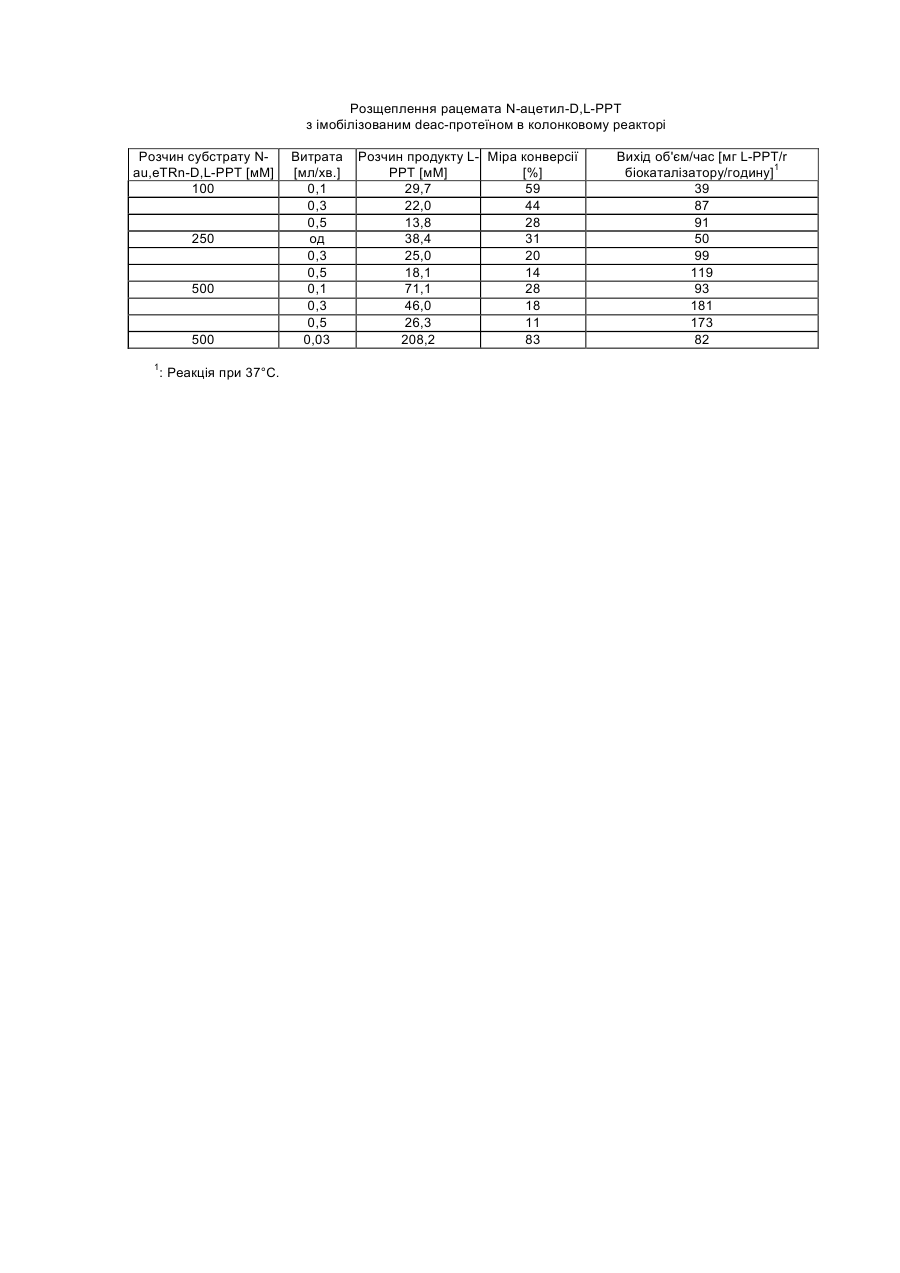
Одержання непротеїногенної амінокислоти L-PPT (L-фосфінотрицин) за допомогою розщеплення рацемата з високою чистотою і виходом досі було описане тільки за допомогою розщеплення фенацетилфосфінотрицину за допомогою пеніцилін-G-ацилазою з Escherichia coli (DE-A-3048612). Але синтез фенацетил-РРТ в порівнянні з N-Ac-PPT є більш трудомістким і дорогим. Однак С-ацилаза не володіє специфічністю до аліфатичних ацильних залишків і, тим самим, також до N-Ac-PPT. Інші відомі ацилази також не мають або мають лише незначну специфічність субстрату до N-Ac-PPT і досі використовувалися тільки в мікробних біотрансформаціях без очищення ферментів (як, наприклад, описано в заявці DE-А-2939269). Внаслідок цього були досяжні тільки дуже незначні виходи об'єм/час. З заявки на патент ЕР-А-0382113 відомо L-специфічне розщеплення складних ефірів карбонових кислот N-Ac-PPT за допомогою ацилази І. Цей фермент також не володів специфічністю до вільних карбонових кислот, і тому як додаткова стадія синтезу при приготуванні субстрат була потрібна етерифікація. Заявка на патент DE-A-19652284 описує цільове виділення мікробних деатилаз з ґрунтових проб зі специфічністю до N-ацетил-амінокислот, переважно N-ацетил-фосфінотрицину (N-Ac-PPT), а також клонування відповідних генів з Stenotrophomonas sp. і Comamonas acidovorans. На основі виявленої гомології послідовності і проведених тестів специфічності субстрату змогли показати, що описані деацетилази відносяться до групи гіпурат-гідролаз (EC 3.5.1.32), природним субстратом яких є Nбензоїл-гліцин, похідне амінокислоти з однією ароматичною N-ацильною функціональною групою. Тому представляло інтерес те, що ці ферменти як субстрат також можуть зв'язувати N-ацетильовані амінокислоти, особливо N-ацетил-фосфінотрицин, причому мова йде про похідні амінокислот з аліфатичними N-ацильними функціональними групами. З заявок на патенти DE-A-2939269 і DE-A-2717440 відомо, що гербіцидна дія рацемічного фосфінотрицину викликається тільки його L-енантіомером (L-PPT). У зв'язку з цим представляло інтерес, що виявлені деацетилази розщеплюють виключно L-енантіомери Nацетил-РРТ, а також N-ацетил-похідні деяких протеїногенних амінокислот з високою специфічністю. Тому ці ферменти чудово підходять для одержання L-амінокислот, особливо гербіцидної біологічно активної речовини L-фосфінотрицину з його рацемічних N-ацетил-похідних за принципом розщеплення рацемата, як це описується, наприклад, в заявках на патенти ЕР-А-0304021, DE-A-2939269 і DE-A-3048612. Істотні недоліки способів, описаних у вище вказаних заявках однак полягають в тому, що (1) можуть використовуватися тільки незначні концентрації субстрату (близько 0,5% у випадку заявки DE-A-2939269), внаслідок чого технічна придатність оцінюється як незначна, що (2) працюють з не виділеними ферментами, внаслідок чого проблема побічних реакцій і подальших стадій очищення не вирішується або є такою, що дуже дорого коштує і що (3) одержання продукту вимагає великих матеріальних витрат (як у разі заявки на патент DE-A-3048612). Задача винаходу полягає в тому, щоб експресувати одну або більше нових деацетилаз (як вже охарактеризовано в DE-A-19652284) у відповідних формі і кількості, і за допомогою цих ферментів забезпечити одержання L-PPT, а також деяких протеїногенних L-амінокислот з хімічно дуже легко доступних рацемічних N-ацетил-D,L-похідних за допомогою розщеплення рацемата з високим виходом і чистотою енантіомерів. У випадку, якщо ферментативна активність вже приблизно відома, і якщо вже сталося клонування послідовності нуклеїнової кислоти, що кодує фермент, перша дія полягає в тому, що відповідний фрагмент нуклеїнової кислоти переводять у відповідний вектор, для того, щоб потім або здійснити надпродукування в придатному штамі бактерії, і/або щоб виділити протеїн після надпродукування. Виділений таким чином фермент може або безпосередньо використовуватися для ферментативної реакції, або через відповідні зв'язуючі групи фіксуватися на матриці. Перший ферментативний тест може бути рекомендований для того, щоб одержати основне підтвердження, чи протікає бажана реакція. Всі подальші заходи служать для оптимізації параметрів відносно специфічності реакції (по відношенню до початкової речовини і продукту), швидкості реакції, ефективності реакції, періоду напіврозпаду ферменту, що використовується, а також можливих концентрацій субстрату. У випадку якщо окремі параметри не точно відповідають вимогам, потрібно виробити необхідні зміни в послідовності нуклеїнової кислоти, що кодує фермент, які спричинять подальшу оптимізацію природного ферменту, що є. Клонування окремих деацетилаз, що використовуються вже описувалося в заявці DE-A-19652284. Фрагменти нуклеїнової кислоти, що кодують відповідні деацетилази були переклоновані, як нижче приведено в прикладі 1, у відповідні вектори експресії, для того щоб таким чином забезпечити необхідну міру певної специфічності субстрату і відсутність побічних реакцій. Так, в заявці DE-A-19652284 змогли показати, що Nацетил-D-РРТ деацетилюється, однак не змогли показати, що N-ацетил-D-PPT не деацетилюється. Виняткове перетворення N-ацетил-L-амінокислот і N-ацетил-L-РРТ являє собою, однак, елементарну попередню умову для технічного застосування, що розглядається тут. Винахід відноситься до способу одержання протеїногенних або непротеїногенних L-амінокислот з їх рацемічних N-ацетил-D,L-похідних, який відрізняється тим, що а) за допомогою ферментативного розщеплення рацемата за допомогою виділених рекомбінантних ферментів селективно деацетилюють N-ацетил-L-похідні відповідних L-амінокислот, в той час як N-ацетил-Dпохідні відповідних D-амінокислот не деацетилюються, і b) отримані деацетильовані L-амінокислоти препаративно відділяють від не деацетильованих N-ацетил-Dпохідних і/або не повністю деацетильованих N-ацетил-L-похідних. Винахід відноситься, особливо, до способу одержання L-фосфінотрицину (L-РРТ) з N-ацетил-D,Lфосфінотрицину за допомогою ферментативного розщеплення рацемата за допомогою виділених рекомбінантних ферментів. Далі, винахід стосується способу одержання L-глутамінової кислоти, L-гістидину, L-лейцину, L-глутаміну і/або L-фенілаланіну з їх відповідних N-ацетил-D,L-похідних за допомогою ферментативного розщеплення рацемата. Далі, винахід стосується способу, який відрізняється тим, що здійснюють енантіоселективне одержання однієї або декількох L-амінокислот з їх відповідних рацемічних N-ацетил-D,L-похідних з однією або декількома деацетилазами з групи гіпурат-гідролаз, причому особливо здійснюють енантіоселективне одержання Lфосфінотрицину, L-глутамінової кислоти, L-гістидину, L-лейцину, L-глутаміну і/або L-фенілаланіну з їх відповідних рацемічних N-ацетил-D,L-похідних з однією або декількома деацетилазами з групи гіпуратгідролаз. Особливо винахід стосується способу, який відрізняється тим, що здійснюють енантіоселективне одержання однієї або декількох L-амінокислот з їх відповідних рацемічних N-ацетил-D,L-похідних з ферментами deac 1 з Stenotrophomonas sp. і/або deac 2 з Comamonas acidovorans (обидві деацетилази описані в заявці DE-A-19652284), і при цьому особливо стосується способів, які відрізняються тим, що здійснюють енантіоселективне одержання L-фосфінотрицину, L-глутамінової кислоти, L-гістидину, L-лейцину, L-глутаміну і/або L-фенілаланіну з їх відповідних рацемічних N-ацетил-О, L-похідних з ферментами deac 1 з Stenotrophomonas sp. і/або deac 2 з Comamonas acidovorans (обидві деацетилази описані в заявці DE-A19652284). Обидва ферменти характеризуються значною гомологією послідовності, яка забезпечує ідентичну або, принаймні, схожу функцію при розщепленні N-ацетил-О, L-похідних. Далі, винахід стосується застосування одержаних рекомбінантних деацетилаз як біокаталізаторів, які дозволяють проводити відповідні реакції з високими концентраціями субстрату і/або забезпечують високі виходи об'єм/час, особливо при застосуванні рекомбінантних деацетилаз в імобілізованій формі. Далі, винахід стосується проведення вище описаних способів при температурі реакції від близько 25°С до 65°С, переважно при температурі від близько 30°С до 45°С і особливо переважно при температурі від близько 35°С до 40°С. Далі, винахід стосується проведення вище описаних способів при концентраціях субстрату від близько 10мМ до 1500мМ, переважно при концентраціях субстрату більше ніж 50мМ, особливо переважно при концентраціях більше ніж 250мМ і вкрай переважно при концентраціях більше ніж 500мМ. Далі, винахід стосується відділення L-амінокислот або L-PTC, отриманих за допомогою вище описаних прийомів, від відповідних N-ацетил-D,L-похідних, які або, як у випадку N-ацетил-D-похідного, не зазнавали перетворення, або, як у випадку N-ацетил-L-похідних, не повністю зазнавали перетворення. Придатними способами при цьому є використання іонообмінної хроматографії на кислому іонообміннику або екстракція N-ацетил-D,L-похідних за допомогою органічних розчинників, як, наприклад, метилізобутилкетон, причому вище згадані отримані L-амінокислоти або L-PTC переходять у водну фазу, з якої потім концентруються висушуванням. Приклади: 1. Одержання deac-ферменту як рекомбінантного протеїну в Escherichia coli: Ген структури deac 1 з Stenotrophomonas sp., що кодує N-Ac-PPT-специфічну деацетилазу, клонували у вигляді фрагмента 1,4-kb Bam HI/Sall (описані в DE-A-19652284) в сайт Bam Hl/Sal 1 His-міченого вектора експресії pQE30 фірми Quiagen [Quiaexpress Kit, Type IV, Katalog Nr. 32149, Stuber і співр. (1990), Immunological Methods, Lefkovits, I. and Pernis, В., т IV, Academic Press, New York, стор.121-152]. Всі молекулярно-біологічні роботи проводили по стандартних методиках, як, наприклад, описано: Ausubel і співр. (1995), Current protocols in molecular biology, John Wiley and Sons, New York. Утворений фрагмент трансформували в штам бактерії Е.Соlli М15, рекомендованому виробником (Qiagen), і рекомбінантні клони відбирали в 100мкг/мл ампіциліну і 25мкг/мл канаміцину. Експресія рекомбінантного злитого протеїну індукувалася за допомогою додання 1мМ інозинфосфотригліцериду (IPTG). Клітини відбирали через 4-5 годин після індукції і зберігали для розділення протеїну при -20°С. Завдяки додецилсульфат натрію (SDS)/поліакриламідному електрофорезу аліквоти клітин індукованих клонів змогли виявити His-мічений – deas1-злитий протеїн у вигляді додаткової смуги 49-кДа. Доказ ферментативної активності злитого протеїна-деацетилази здійснювали радіоактивним аналізом з [14С]-N-ацетил-L-РРТ як субстрату. Для цього здійснювали проникнення 200мкл індукованих культур 0,5% толуолом, 0,5% етанолом протягом 30хв. при 37°С. Потім грануляти клітин ресуспендували в 25мкл 0,1мМ [14С]-N-ацетил-L-PPT, 10мМ NaCl, 10мМ фосфату натрію, рН=7,5 і інкубували 1 годину при 37°С. Для якісного визначення утвореного [14C]-L-PPT аналізували по 5мкл аліквоти суміші, що тестується за допомогою тонкошарової хроматографії на целюлозних пластинах ВЕТРХ (Merck) з розчинником: н-пропанол: 25% аміак=3:2. Радіоактивні зв'язки виявляли на рентгенівській плівці за допомогою радіографії. Для кількісного визначення перетворення ферментів суміші, що тестується вимірювали радіо-ВЕРХ з Spherisorb® SAX як розділовою колонкою (розчинник: 5мМ КН2РО4, 10% метанол, рН=1,92, витрата 0,5мл/хв.). У цих умовах [14C]-L-РРТ елюювався через 4,5хв. і [14C]-N-ацетил-L-PPT через 6,5хв. Клон експресії продукував протеїн-деацетилазу з дуже високої N-ацетил-L-РРТ специфічною активністю деацетилази, згідно з кількісним визначенням із вмістом близько 10% на загальний протеїн і використовувався для очищення ферменту і подальших описаних біотрансформацій. 2. Очищення деацетилази за допомогою афінної хроматографії. Виділення протеїну-деацетилази з наново названого pQEDEAC штаму експресії після клонування, описаного в прикладі 1, відбувалося по методиці Quiagen (Qiaexpress-Kit) за допомогою афінної хроматографії на нікель-нітрилтриацетатній матриці (Ni-NTA) в нативних умовах. Для цього 800мл культури штаму експресії pQEDEAC ферментувати, як описано в прикладі 1 і індукували. Зібраний гранулят клітин (4г) ре-суспендували в 20мл лізинового буферу і обробляли ультразвуком. Після центрифугування змішували прозорий протеїнолізат (120мг загального протеїну) з 4мл 50%-ної суспензії Ni-NTA для скріплення His-міченого злитого протешу і потім матеріал завантажували в ферментативну колонку. Розділову матрицю промивали 10 об'ємами водного буфера і потім елюювали за допомогою 4мл елюатного буфера. Для перевірки чистоти розділення аналізували аліквоту оброблених ультразвуком клітин, промивного розчину і елюату за допомогою додецилсульфат натрію /поліакриламідного гель-електрофорезу. Активність деацетилази різних протеїнвмісних фракцій визначали радіоактивним аналізом, описаним в прикладі 1 (реакційні суміші: 9мкл розчину протеїну + 1мкл 1мМ [14С]-N-ацетил-L-РРТ, умови реакції описані вище). Для кількісного визначення ферменту інкубували протеїнові фракції з 50мМ N-ацетил-D,L-РРТ (9мкл розчину протеїну + 1мкл 500мМ Nацетил-D,L-PPT) протягом 30хв. при 37°С. РРТ, що утворився потім вимірювали в амінокислотному аналізаторі (Biotronic LC 5001). Протеїн в сирому екстракті після обробки клітин мав специфічну активність відносно деацетилази 3нмоль субстрат/сек./мг протешу. З 800мл культури штаму експресії змогли виділити за допомогою афінної хроматографії 10мг His-міченого –deas-злитого протеїну зі специфічною активністю 20нмоль субстрат/сек./мг протеїну і чистотою близько 80%. 3. Специфічність субстрату і стереоселективність деацетилази. Для дослідження специфічності субстрату і стереоспецифічності deacl-протеїну інкубували по 5мкг очищеного ферменту в 20мкл сумішей 10мМ NaCl, 10мМ фосфату натрію, рН=7,5 і 25мМ N-ацетил-Lамінокислот або відповідних D-енантіомерів, вказаних в таблиці 1. Для порівняння використали гіпурову кислоту (N-бензоїлгліцин), природний субстрат деацетилази. Реакційні суміші інкубували 1 годину при 37°С, і потім визначали на утворення вільних амінокислот в амінокислотному аналізаторі (Biotronic LC 5001). У таблиці 1 вказана відносна активність у відношенні деацетилази для L-енантіомерів різних субстратів. Крім гіпурової кислоти і N-Ac-L-PPT також deac1-фермент розщеплює ще N-ацетил-похідні ряду природних амінокислот, особливо N-Ac-L-глутамінову кислоту, N-Ac-гліцин, N-Ac-L-гістидин, N-Ac-L-орнітин, N-Ac-Lлейцин, N-Ac-L-глутамін і N-Ac-L-фенілаланін. У всіх випадках фермент був виключно активний з Lенантіомерами, в той час як відповідні D-сполуки ні в одному випадку не могли зазнавати перетворення. У всіх інших випадках, що тестуються (дивись таблицю 1) перетворення N-ацетил-L-похідних мало місце тільки в дуже незначній мірі або взагалі було відсутнім. Таблиця 1 Специфічність субстрат deac-ферменту Субстрат1 Гіпурова кислота (N-бензоїлгліцин) N-Ac-L-фосфінотрицин N-Ac-L-орнітин N-Ac-L-метіонін N-Ac-L-триптофан N-Ac-L-феніл аланін N-Ac-L-тирозин N-Ac-L-глутамінова кислота Ν-Ac-L-глутамін Ν-Ac - гліцин Ν-Ac-L-гістидин N-Ac-L-лейцин N-Ac-L-валін Ν-Ac-L-серин N-Ac-L-пролін Відносн. Активність 2 [%] 100 43 37 0 0,4 24 11 100 30 59 43 33 2 4 0 1 : для всіх сполук мова йде про L-енантіомери : специфічну активність, виміряну з гіпуровою кислотою [нмоль гліцину/хв./мг протеїну] приймали за 100%, і інші значення співвідносилися до цієї величини. 2 4. Розщеплення рацемата N-ацетил-D,L-фосфінотрицину за допомогою біотрансформації клітинами штаму-виробника - deac: 400мл культури штаму експресії pQEDEAC ферментувати як описано в прикладі 1 і індукували. Вирощені клітини потім промивали в 10мМ NaCl, 10мМ фосфату натрію, рН=7,5 і після витримування протягом ночі ліофілізували. Оброблені таким чином клітини могли зберігатися понад декількох тижнів при 4°С, зберігаючи високу специфічну активність у відношенні деацетилази, і служили в подальших дослідах як біокаталізатор; 2мг ліофілізованих клітин ресуспендували в 1мл наступних розчинів субстрату різної концентрації: (1) 10мМ динатрієвої солі N-aneraraD,L-PPT (0,3%) (2) 50мМ " " " " " " " " " " (1,3%) (3) 100мМ " " " " " " " " " " (2,6%) (4) 250мМ " " " " " " " " " " (6,7%) (5) 500мМ " " " " " " " " " " (13,3%) Суміші містили як реакційний буфер 10мМ NaCl, 10мМ фосфату натрію, рН=8,0 і були інкубовані 48 годин на качалці 100об./хв. і 37°С. Вільний РРТ, утворений при реакції гідролізу, визначали в амінокислотному аналізаторі (Biotronic LC 5001). Результати приведені в таблиці 2. Чистоту енантіомерів продукту реакції аналізували за допомогою хиральної ВЕРХ з розділовою колонкою Chirex® (D) Penicillamin (Phenomenex). Як розчинник використали 2мМ CuSO4, 10% метанол з витратою 0,5мл/хв. Визначення відбувалося фотометрично при 254нм. У цих умовах елююються еталонні розчини L-PPT через 17хв. і D-PPT через 21хв. У всіх пробах, що тестуються змогли довести виключно наявність L-PPT як продукту реакції. Досягнуті міри конверсії [продукований L-PPT/N-ацетил-L-PPT в субстраті x 100] знаходилися між 100% при низьких концентраціях субстрату (10мМ, 50MM) і 66% при самій високій концентрації субстрату (500мМ). Таблиця 2 Розщеплення рацемата N-ацетил-В,L-PPT в залежності від концентрації субстрату Розчин субстрату N-aqeran-D,L-PPT [мМ] 10 50 100 250 500 1 Розчин продукту1 L-PPT [мМ] 5 25 48 108 164 Міра конверсії L-PPT/N-ацетон-D,LPPT в % 100 100 96 86 66 : концентрація на біокаталізатор: 2мг/мл У наступному експерименті підвищували концентрацію на біокаталізатор і знімали кінетику реакції розщеплення субстрату. 13мг ліофілізованих клітин ресуспендували в 1мл розчину субстрату, що складається з 500мМ динатрієвої солі N-ацетил-D,L-PPT (13,3% вага./об.), 10мМ NaCl, 10мМ фосфату натрію, рН=8,0 і інкубували на качалці при 100об./хв. і 37°С. Для визначення утвореного L-РРТ через 55 годин відбирали по 50мкл аліквоти з реакційної суміші, центрифугували і супернатант заморожували аж до аналізу при -20°С. Кількісний аналіз продукту реакції L-PPT, а також визначення чистоти енантіомерів проводили, як було описано вище. Результати приведені в таблиці 3. При вибраних умовах L-специфічне розщеплення субстрату через приблизно 55 годин було завершено. При цьому досягнуті міра конверсії близько 90% і вихід об'єм/час 483 [мг L-PPT/r біокаталізатору/годину]. Таблиця 3 Кінетика реакції розщеплення рацемата N-ацетил-D,L-PPT Час реакції [година] 0 1 4 9 23 31 47 55 1 L-PPT [мМ]1 0 27,7 59,7 106,0 181,6 186,5 208,6 221,8 : концентрація на біокаталізатор: 13 мг/мл. Концентрація субстрату 500мМ N-ацетил-D,L-РРТ. 5. Імобілізація очищеного dеас-протеїну. Щоб можна було провести ферментативну реакцію з виділеним deac-протеїном, імобілізували фермент, очищений як His-мічений-злитий протеїн в прикладі 2, на полімерний носій VINA-Epoxy Biosynth® (Riedel de Haen). Для цього концентрували dеас-протеїн за допомогою осадження сульфатом амонію до 15мг/мл і після витримування протягом ночі проводили діаліз проти 1Μ фосфату Na, pH=8,0. Для стандартної реакції скріплення повільно струшували 10мг протеїну з 100мг носія-VINA протягом 2 днів при кімнатній температурі. Потім імобілізат центрифугували і по 1 промивали в імобілізованому буфері, а також в 50мМ фосфату Na, pH=7,0. Потім для блокування вільних епоксигруп носій інкубували 1 годину при кімнатній температурі з 50 мМ фосфату Na, pH = 7,0, 50мМ 2-меркаптоетанолу. Після цього біокаталізатор знаходився в 1мл 5мМ фосфату Na, pH=7,0, 0,02% азиду натрію при 4°С. Для визначення скріплення протеїну визначали вміст протеїну в розчині до і після імобілізації, а також в промивних розчинах за допомогою способу Бредфорда [Bradford (1976), Anal. Bio-chem. 72: 248-254]. Специфічна активність деацетилази вимірювалася до і після імобілізації радіоактивним аналізом, описаним в прикладі 1. Відносна активність ферменту після скріплення становила >90% величини, визначеної для розчиненого протеїну. 6. Розщеплення рацемата Н-ацетил-О,Ь-фосфінотрицину з імобілізованим dеас-протеїном в колонковому реакторі. 1г імобілізату ферменту (волога вага) завантажували в колонку і використовували як біокаталізатор для розщеплення рацемата різних концентрованих розчинів субстрату з різними витратами. Розчини субстрату містили концентрацію діаммонієвої солі N-aueran-D, L-PPT, приведену в таблиці 4 в 10мМ NaCl, 10мМ фосфату Na, pH=8,0. Реакції проводили при 37°С протягом понад 10 днів. Перетворення кількісно оцінювали за допомогою визначення L-PPT, що є в проточному засобі колонки за допомогою амінокислотного аналізу і хиральної ВЕРХ (дивись приклад 4). Результати представлені в таблиці 4. Найвища міра конверсії 83% досягалася з найвищою концентрацією субстрату (500мМ) і самим низьким потоком субстрату (0,03мл/хв.). Виходи об'єм/час збільшувалися з витратою і концентрацією субстрату і досягали найвищого значення 181 [мг L-PPT/r біокаталізатору/годину] при концентрації субстрат 500мМ і потоку 0,3мл/хв. Імобілізована деацетилаза демонструвала протягом тривалого часу експерименту хорошу стабільність і володіла через 10 днів при 37°С ще приблизно 80% початкової активності. Таблиця 4 Розщеплення рацемата N-ацетил-D,L-PPT з імобілізованим dеaс-протеїном в колонковому реакторі Розчин субстрату Nau,eTRn-D,L-РРТ [мМ] 100 250 500 500 1 : Реакція при 37°С. Витрата [мл/хв.] 0,1 0,3 0,5 од 0,3 0,5 0,1 0,3 0,5 0,03 Розчин продукту L- Міра конверсії PPT [мМ] [%] 29,7 59 22,0 44 13,8 28 38,4 31 25,0 20 18,1 14 71,1 28 46,0 18 26,3 11 208,2 83 Вихід об'єм/час [мг L-PPT/r біокаталізатору/годину]1 39 87 91 50 99 119 93 181 173 82
ДивитисяДодаткова інформація
Назва патенту англійськоюA method for the production of l-amino acids from their racemic n-acetyl-d,l-derivatives using enzymatic racemate cleavage by means of isolated recombinant enzymes
Назва патенту російськоюСпособ получения l-аминокислот из их рацемических n-ацетил-d, l-производных при помощи ферментативного расщепления рацемата при помощи выделенных, рекомбинантных ферментов
МПК / Мітки
МПК: C12N 15/09, C12P 41/00, C12N 9/78, C12P 13/04, C12N 15/55
Мітки: виділених, l-амінокислот, спосіб, l-похідних, одержання, n-ацетил-d, рекомбінантних, розщеплення, допомогою, ферментів, рацемата, рацемічних, ферментативного
Код посилання
<a href="https://ua.patents.su/5-73743-sposib-oderzhannya-l-aminokislot-z-kh-racemichnikh-n-acetil-d-l-pokhidnikh-za-dopomogoyu-fermentativnogo-rozshheplennya-racemata-za-dopomogoyu-vidilenikh-rekombinantnikh-fermentiv.html" target="_blank" rel="follow" title="База патентів України">Спосіб одержання l-амінокислот з їх рацемічних n-ацетил-d, l-похідних за допомогою ферментативного розщеплення рацемата за допомогою виділених, рекомбінантних ферментів</a>
Спосіб одержання препарату похідних хімотрипсин-трипсинподібних протеолітичних ферментів для парентерального введення

Номер патенту: 50369
Опубліковано: 15.06.2005
Автори: Комісаренко Сергій Васильович, Волков Георгій Леонідович, Верьовка Сергій Вікторович
МПК: A61K 38/46
Мітки: ферментів, препарату, парентерального, протеолітичних, похідних, одержання, введення, хімотрипсин-трипсинподібних, спосіб
Формула / Реферат:
Лікеро-горілчаний напій, що включає водно-спиртову рідину з спирту етилового ректифікованого і води питної підготовленої, цукровий сироп 65,8%-ний, кислоту лимонну, ароматичну лимонну сировину, який відрізняється тим, що додатково містить ароматизатор "Пітахая", підсолоджувач Ацесульфам - К(Е950) та бензоат натрію, взятих у відповідному співвідношенні інгредієнтів на 1000 дал. напою: Ароматизатор харчовий...
Спосіб одержання препарату похідних хімотрипсин-трипсин-подібних протеолітичних ферментів для парентерального введення

Номер патенту: 50369
Опубліковано: 15.10.2002
Автори: Верьовка Сергій Вікторович, Волков Георгій Леонідович, Комісаренко Сергій Васильович
МПК: A61K 38/46
Мітки: одержання, спосіб, похідних, препарату, протеолітичних, парентерального, хімотрипсин-трипсин-подібних, ферментів, введення
Формула / Реферат:
Лікеро-горілчаний напій, що включає водно-спиртову рідину з спирту етилового ректифікованого і води питної підготовленої, цукровий сироп 65,8%-ний, кислоту лимонну, ароматичну лимонну сировину, який відрізняється тим, що додатково містить ароматизатор "Пітахая", підсолоджувач Ацесульфам - К(Е950) та бензоат натрію, взятих у відповідному співвідношенні інгредієнтів на 1000 дал. напою: Ароматизатор харчовий...
Спосіб одержання циталопраму (варіанти), s-циталопраму, проміжні кетони та спосіб одержання рацемічних сполук

Номер патенту: 72238
Опубліковано: 15.02.2005
Автори: Петерсен Ханс, Рок Майкл Харольд, Еллегор Петер
МПК: C07C 253/30, C07C 255/56, C07D 307/87
Мітки: рацемічних, s-циталопраму, сполук, одержання, кетони, варіанти, циталопраму, проміжні, спосіб
Формула / Реферат:
1. Спосіб одержання циталопраму, згідно з яким здійснюють реакцію сполуки формули IV, IVде R являє собою ацил, з 3-(N,N-диметиламіно)пропілмагнійгалогенідом, переважно з 3-(N,N-диметиламіно)пропілмагнійхлоридом, з одержанням циталопраму формули I, Iякий виділяють у вигляді основи або її фармацевтично прийнятної солі.2. Спосіб за п. 1, який відрізняється тим, що проміжну сполуку формули IV одержують...
Похідні (2r, 3s, 4s, 5r, 6r, 10r, 11r)-2,4,6,8,10-пентаметил-11-ацетил-12,13-діоксабіцикло[8.2.1]тридец-8-ен-1-ону, лікарські засіб та спосіб одержання похідних
Номер патенту: 57030
Опубліковано: 16.06.2003
Автори: Пройшофф Улф, Еккхаут Крістіан, Жассеран Даніель
МПК: A61K 31/7048, A61K 31/7042, C07H 17/08, A61P 1/00, A61K 31/70, A61K 31/00, A61P 43/00, A61P 1/14
Мітки: похідних, 11r)-2,4,6,8,10-пентаметил-11-ацетил-12,13-діоксабіцикло[8.2.1]тридец-8-ен-1-ону, спосіб, одержання, засіб, лікарські, похідні
Формула / Реферат:
1. Похідні (2R, 3S, 4S, 5R, 6R, 10R, 11R)-2,4,6,8,10-пентаметил-11-ацетил-12,13-діоксабіцикло [8. 2.1] тридец-8-ен-1-ону загальної формули І,ІдеR1 позначає водень або метил іR2 означає водень або нижчий алканоїл,і їх фізіологічно прийнятні кислі адитивні солі.2. Сполуки згідно з п. 1, де R1 означає метил.3. Сполуки згідно з будь-яким із пп. 1 або 2, де R2 означає водень.4. Лікарський...
Спосіб отримання похідних бензаміду, або їх солей, або рацемічних сумішей, або стерєоізомерів

Номер патенту: 4753
Опубліковано: 28.12.1994
Автори: Геста Леннарт Фрорвалл, Свен Ове Егрен
МПК: A61K 9/48, C07C 51/347, A61P 25/18, C07D 207/09, C07C 67/00, C07C 65/00, C07C 51/00, A61K 9/20, A61K 31/40
Мітки: солей, рацемічних, бензаміду, отримання, стерєоізомерів, сумішей, спосіб, похідних
Формула / Реферат:
Способ получения производных бензамида общей формулыгде R1 - алкил с 1-3 С;R2 и R3 - одинаковые или различные и означают водород, хлор или бром,или их солей, или рацемических смесей, или сте-реоизмеров, отличающийся тем, что соединение общей формулыгде R1, R2 и R3 имеют вышеуказанные значения, подвергают взаимодействию с амином общей формулыв среде органического растворителя и целевой...
Попередній патент: Клейова композиція
Наступний патент: Спосіб отримання лікарського засобу седативної дії на основі рослинної сировини і лікарський засіб “флорісед”, отриманий цим способом
Випадковий патент: Прилад для вимірювання лінійних розмірів з цифровою індикацією і динамометром з лінійною індикацією