Спосіб ферментації галактози
Номер патенту: 108853
Опубліковано: 25.06.2015
Автори: Брурс Ніколете Ясмейн, Сейлеком ван Гейсбердіна Пітернела, Гілесен Біанка Елісабет Марія, Лат де Вілхелмус Теодорус Антоніус Марія, Класен Пол, Відеман Беате







Формула / Реферат
1. Спосіб отримання одного або більше продуктів ферментації з композиції цукрів, який передбачає наступні стадії:
a) ферментацію композиції цукрів у присутності дріжджів, які належать до роду Saccharomyces, Kluyveromyces, Candida, Pichia, Schizosaccharomyces, Hansenula, Kloeckera, Schwanniomyces або Yarrowia, і
b) вилучення продукту ферментації,
і в якому дріжджі містять гени аrаА, аrаВ і araD, а композиція цукрів включає глюкозу, галактозу і арабінозу.
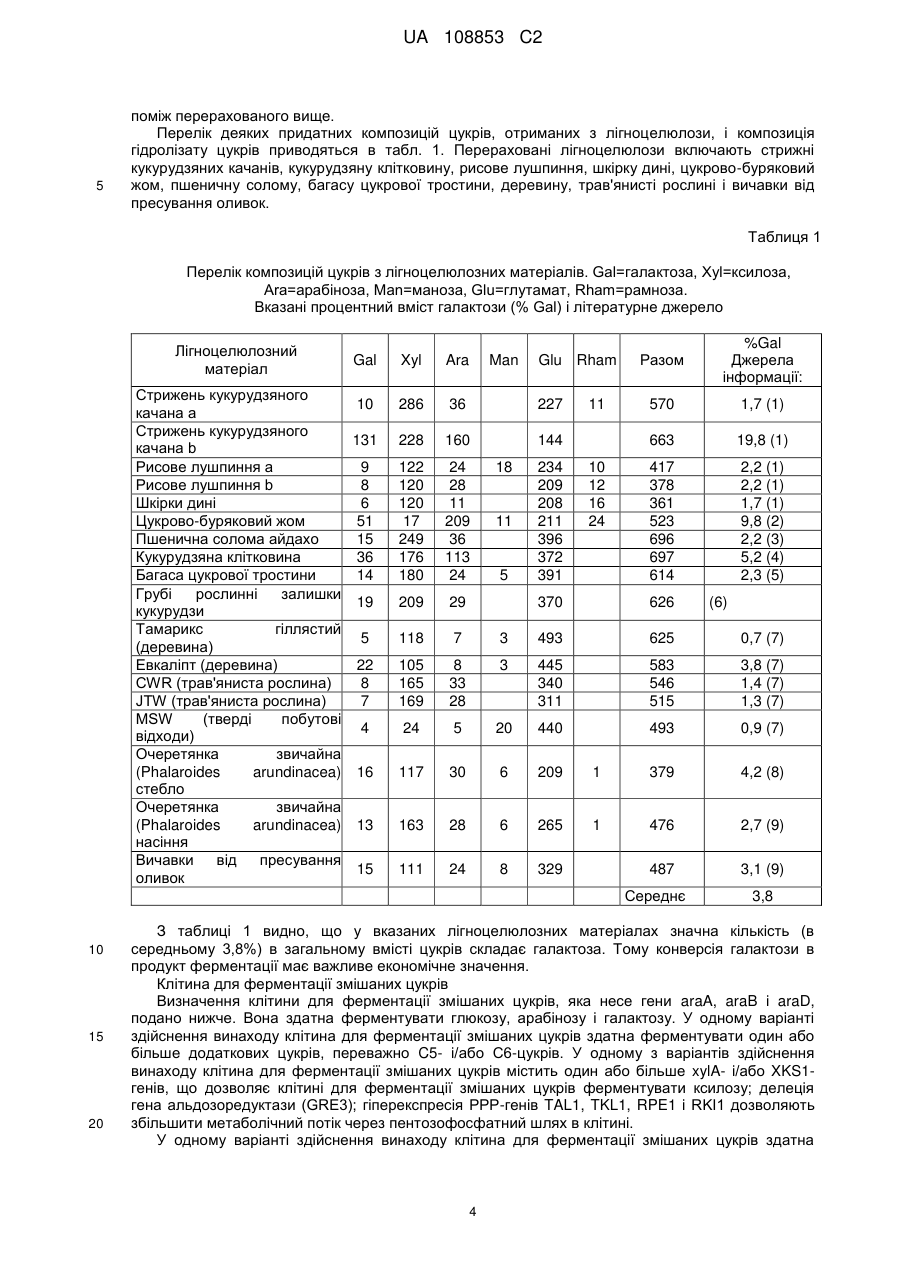
2. Спосіб за п. 1, який відрізняється тим, що цукри глюкоза, галактоза і арабіноза піддаються конверсії в продукт ферментації.
3. Спосіб за п. 1 або 2, який відрізняється тим, що одним або більше продуктом(тами) ферментації є етанол.
4. Спосіб за будь-яким з пп. 1-3, який відрізняється тим, що композиція цукрів отримана з лігноцелюлозного матеріалу шляхом:
a) передобробки одного або більше лігноцелюлозних матеріалів для отримання заздалегідь обробленого лігноцелюлозного матеріалу;
b) ферментативної обробки заздалегідь обробленого лігноцелюлозного матеріалу для отримання композиції цукрів.
5. Спосіб за будь-яким з пп. 1-4, який відрізняється тим, що клітина для ферментації змішаних цукрів є клітиною дріжджів роду Saccharomyces.
6. Спосіб за п. 5, який відрізняється тим, що клітина для ферментації змішаних цукрів є клітиною дріжджів Saccharomyces cerevisiae.
7. Спосіб за будь-яким з пп. 1-6, який відрізняється тим, що клітина для ферментації змішаних цукрів містить делецію гена альдозоредуктази.
8. Спосіб за будь-яким з пп. 1-7, який відрізняється тим, що ферментація проводиться за анаеробних умов або за умов обмеженої кількості кисню.
9. Спосіб за будь-яким з пп. 1-8, який відрізняється тим, що клітина для ферментації змішаних цукрів містить гіперекспресовані РРР-гени TAL1, TKL1, RPE1 і RKI1.
10. Спосіб за будь-яким з пп. 1-9, який відрізняється тим, що клітина для ферментації змішаних цукрів містить ху1А-ген і/або XKS1-ген.
11. Спосіб за будь-яким з пп. 1-10, який відрізняється тим, що гени були введені в клітину для ферментації змішаних цукрів шляхом введення в клітину-хазяїна:
a) кластера, який складається з РРР-генів TAL1, TKL1, RPE1 і RKI1, під контролем сильних промоторів,
b) кластера, який складається з xy1A-гена і XKS1-гена, обидва під контролем конститутивних промоторів,
c) кластера, що складається з генів аrаА, аrаВ і araD, і/або кластера з ху1А-гена і XKS1-гена; і
d) делеції гена альдозоредуктази;
і адаптивної еволюції конструкта для змішаних цукрів для отримання клітини для ферментації змішаних цукрів.
12. Спосіб за п. 11, який відрізняється тим, що клітина-хазяїн є стійкою до інгібіторів клітиною.
13. Спосіб за п. 11 або п. 12, який відрізняється тим, що клітина-хазяїн є промисловим штамом.
14. Спосіб за будь-яким з пп. 1-13, який відрізняється тим, що продукт ферментації вибраний з групи, яка складається з етанолу, n-бутанолу, ізобутанолу, молочної кислоти, 3-гідроксипропіонової кислоти, акрилової кислоти, оцтової кислоти, бурштинової кислоти, фумарової кислоти, яблучної кислоти, ітаконової кислоти, малеїнової кислоти, лимонної кислоти, адипінової кислоти, амінокислоти, такої як лізин, метіонін, триптофан, треонін і аспарагінова кислота; 1,3-пропандіолу, етилену, гліцерину, b-лактамного антибіотика і цефалоспорину; вітамінів, фармацевтичних засобів, кормових добавок для тварин, хімічних засобів спеціального призначення, хімічної сировини, пластмас, розчинників, палива, включаючи біопаливо і біогаз, або органічних полімерів; і промислового ферменту, такого як протеаза, целюлаза, амілаза, глюканаза, лактаза, ліпаза, ліаза, оксидоредуктаза, трансфераза або ксиланаза.
15. Застосування генів аrаА, аrаВ і araD для надання шляхом експресії цих генів дріжджам, які зброджують (ферментують) глюкозу і належать до роду Saccharomyces, Kluyveromyces, Candida, Pichia, Schizosaccharomyces, Hansenula, Kloeckera, Schwanniomyces або Yarrowia, здатності зброджувати (ферментувати) за анаеробних умов галактозу у присутності арабінози після виснаження глюкози.
Текст
Реферат: Винахід належить до способу отримання одного або більше продуктів ферментації з композиції цукрів, який передбачає ферментацію композиції цукрів у присутності дріжджів, що належать до роду Saccharomyces, Kluyveromyces, Candida, Pichia, Schizosaccharomyces, Hansenula, Kloeckera, Schwanniomyces або Yarrowia, та вилучення продукту ферментації в якому дріжджі містять гени аrаА, аrаВ і araD, а композиція цукрів включає глюкозу, галактозу і арабінозу. UA 108853 C2 (12) UA 108853 C2 UA 108853 C2 5 10 15 20 25 30 35 40 45 50 55 60 Галузь техніки, до якої відноситься винахід Винахід відноситься до ферментації змішаних цукрів, зокрема, до ферментації композиції цукрів, яка включає глюкозу, галактозу і арабінозу. Композиція цукрів може бути отримана з лігноцелюлозного матеріалу. Попередній рівень техніки Велику частину етанолу як альтернативи рідким паливам отримують в даний час ферментацією кукурудзяного крохмалю і сахарози цукрової тростини. Для досягнення поставлених амбітних цілей у виробництві поновлюваних палив розробляються нові технології перетворення (конверсії) нехарчової біомаси в продукти ферментації, такі як етанол. Дріжджі Saccharomyces cerevisiae є тими організмами, які були вибрані промисловістю з виробництва етанолу, але ці дріжджі не здатні утилізувати п'ятивуглецеві цукри, такі, що містяться в геміцелюлозному компоненті біомаси-сировини. Геміцелюлоза може складати до 20%-30% біомаси, причому C5-цукрами, які превалюють в ній є ксилоза і арабіноза. Гетерологічна експресія ксилоізомерази (XI) є одним з методів, які сприяють метаболізму дріжджових клітин і ферментації ними ксилози. До того ж експресія бактерійних генів araA, araB і araD в штамах S. cerevisiae призводить до утилізації і ефективного спиртового бродіння арабінози. Галактоза є C6-цукром і до того ж цукром, який часто присутній в лігноцелюлозі, часто в кількостях (~4% загальної маси цукрів), якими недоцільно нехтувати з економічних причин. J. van den Brink et al., Microbiology (2009) 155, 1340-1350, указує, що глюкоза є найбільш бажаним джерелом вуглецю для Saccharomyces cerevisiae і що при переході від ферментації в умовах обмеженої кількості глюкози до ферментації з надлишком галактози за анаеробних умов галактоза не споживається. До теперішнього часу не розроблений спосіб конверсії галактози в продукт ферментації разом з глюкозою і одним або більше C5-цукром. Тому метою винаходу є забезпечення способу конверсії галактози в продукт ферментації, в одному процесі з глюкозою і одним або більше C5цукром. Короткий виклад суті винаходу Даний винахід пропонує спосіб виробництва одного або більше продуктів ферментації з композиції цукрів, який передбачає наступні стадії: a) ферментація композиції цукрів у присутності дріжджів, які відносяться до роду Saccharomyces, Kluyveromyces, Candida, Pichia, Schizosaccharomyces, Hansenula, Kloeckera, Schwanniomyces або Yarrowia, і b) вилучення продукту ферментації і в якому дріжджі містять гени araA, araB і araD, а композиція цукрів включає глюкозу, галактозу і арабінозу. Переважно цукри – глюкоза, галактоза і арабіноза – перетворюються на продукт ферментації. Бажано клітина для ферментації змішаних цукрів відноситься до роду Saccharomyces, краще – до Saccharomyces cerevisiae. Крім того, винахід відноситься до використання генів araA, araB і araD для надання (шляхом експресії цих генів) штаму, який ферментує глюкозу, здатності до анаеробної ферментації галактози у присутності арабінози. Короткий опис фігур Фіг. 1 представляє фізичну карту плазміди pPWT006. Фіг. 2 представляє фізичну карту плазміди pPWT018. Фіг. 3 представляє авторадіограму саузерн-блота. Хромосомну ДНК штаму дикого типу CEN.PK113-7D (доріжка 1) і BIE104A2 (доріжка 2) розщеплювали EcoRI і HindIII. Блот гібридизували із специфічним SIT2-зондом. Фіг. 4 представляє фізичні карти SIT2-локуса дикого типу (панель а) і після введення araгенів інтеграцією плазміди pPWT018 з подальшою внутрішньомолекулярною рекомбінацією, яка веде до втрати вектора і послідовностей селектованих маркерів (панель b). Вказана гібридизація зонда. Фіг. 5 представляє фізичну карту плазміди pPWT080, послідовність якої дана в SEQ ID no. 4. Фіг. 6 представляє фізичну карту GRE3-локуса дикого типу (панель а) і однокопійну інтеграцію PWT080 в GRE3-локус (панель b, яка показує, де зв'язуються праймери, і панель С, яка показує, де зв'язується RKI1-зонд). Фіг. 7 представляє фізичну карту GRE3-локуса, в якій кодуюча область GRE3-гена замінена інтеграцією PPP-генів TAL1, TKL1, RKI1 і RPE1. Панель а показує, де зв'язуються праймери SEQ ID 5 і 6, панель b показує, де зв'язується RKI1-зонд. Фіг. 8 показує криві зростання BIE104P1A2 за аеробних умов на різних середовищах. Штам 1 UA 108853 C2 5 10 15 20 25 30 35 40 45 50 55 60 BIE104P1A2 заздалегідь вирощували на YNB з 2% галактози. Крива зростання починається на YNB з 2% галактози і 1 % арабінози з подією, яка відбувалася потім, і яка позначеною на графіці цифрою (1), – перенесенням на YNB з 2% арабінози як єдиного джерела вуглецю. Після досягнення OD600 (оптична щільність культури при довжині хвилі 600 нм), рівної >1, культуру переносили на свіже середовище з початковою OD600, рівною 0,2. Після трьох перенесень на чисте арабінозне середовище отриманий штам був позначений як BIE104P1A2c. Фіг. 9 показує криві зростання BIE104P1A2c за анаеробних умов на середовищі YNB з 2% арабінози як єдиного джерела вуглецю. Після досягнення OD600 більше 1 культуру переносили на свіже середовище з початковою OD600, яка дорівнювала 0,2. Після декількох перенесень отриманий штам отримав назву BIE104P1A2d (= BIE201). Фіг. 10 показує конверсію цукрів і утворення продукту штамом BIE104 на синтетичному модельному середовищі з кукурудзяними волокнами. Продукування CO 2 вимірювали постійно. Моніторинг зростання вели до подальшої оптичної щільності культури. Передкультуру вирощували на 2% глюкози. Фіг. 11 показує конверсію цукрів і утворення продукту штамом BIE104P1A2c на синтетичному модельному середовищі з кукурудзяними волокнами. Продукування CO 2 вимірювали постійно. Моніторинг зростання вели до подальшої оптичної щільності культури. Передкультуру вирощували на 2% глюкози. Фіг. 12 показує конверсію цукрів і утворення продукту штамом BIE201 на синтетичному модельному середовищі з кукурудзяними волокнами. Продукування CO 2 вимірювали постійно. Моніторинг зростання вели до подальшої оптичної щільності культури. Передкультуру вирощували на 2% глюкози. Фіг. 13 показує конверсію цукрів і утворення продукту штамом BIE104A2 на синтетичному модельному середовищі з кукурудзяними волокнами. Продукування CO 2 вимірювали постійно. Моніторинг зростання вели до подальшої оптичної щільності культури. Передкультуру вирощували на 2% глюкози. Фіг. 14 показує конверсію цукрів і утворення продукту штамом BIE105A2 на синтетичному модельному середовищі з кукурудзяними волокнами. Продукування CO 2 вимірювали постійно. Моніторинг зростання вели до подальшої оптичної щільності культури. Передкультуру вирощували на 2% глюкози. Фіг. 15 представляє фізичну карту плазміди pPWT007. Фіг. 16 представляє фізичну карту плазміди pPWT042. Фіг. 17 представляє фізичну карту SIT4-локуса дикого типу (панель а) і однокопійну інтеграцію PWT080 в SIT4-локус (панель b, яка показує, де зв'язуються праймери). Фіг. 18 показує графічне представлення кривих зростання штаму BIE104A2P1Y9 на різних середовищах. Панель а: штам BIE104A2P1Y9, вирощений на глюкозі, з наступними подіями, позначеними на графіці цифрами (1), – перенесення на (1% арабінози + 1% ксилози) і (2) – перенесення на (2% ксилози + 0,2% арабінози). Панель b: штам BIE104A2P1Y9, вирощений на галактозі, з подальшим (1) перенесенням на (1% арабінози + 1% ксилози) і (2) перенесенням на (2% ксилози + 0,2% арабінози). Фіг. 19 показує зростання штаму BIE104A2P1Y9 на Verduyn-середовищі з додаванням 2 % ксилози. Були протестовані дві автономні колонії. Після досягнення OD600, рівної 2, штами переносили на свіже середовище, на якому вони відразу починали знов рости на ксилозі. Фіг. 20 представляє фізичну карту плазміди pGBS416ARAABD. Короткий опис списку послідовностей SEQ ID NO: 1 представляє послідовність ксилоізомерази з штаму дикого типу Bacteroides uniformis ATCC 8492. Genbank accession no. AAYH02000036. SEQ ID NO: 2 представляє кодон-оптимізовану послідовність з SEQ ID NO: 1. SEQ ID NO: 3 представляє амінокислотну послідовність ксилоізомерази з Bacteroides uniformis ATCC 8492. SEQ ID NO: 4 представляє послідовність плазміди pPWT080. SEQ ID NO: 5 представляє послідовність прямого праймера. SEQ ID NO: 6 представляє послідовність зворотного праймера. SEQ ID NO: 7 представляє послідовність прямого багатофункціонального праймера для ПЛР-діагностики. SEQ ID NO: 8 представляє послідовність зворотного багатофункціонального праймера для ПЛР-діагностики. SEQ ID NO: 9 представляє послідовність RKI1-зонда прямого праймера. SEQ ID NO: 10 представляє послідовність RKI1-зонда зворотного праймера. SEQ ID NO: 11 представляє послідовність kanMX-касети прямого праймера. 2 UA 108853 C2 5 10 15 20 25 30 35 40 45 50 55 60 SEQ ID NO: 12 представляє послідовність kanMX-касети зворотного праймера. SEQ ID NO: 13 представляє послідовність прямого праймера. SEQ ID NO: 14 представляє послідовність зворотного праймера. SEQ ID NO: 15 представляє послідовність прямого багатофункціонального праймера для ПЛР-діагностики. SEQ ID NO: 16 представляє послідовність зворотного багатофункціонального праймера для ПЛР-діагностики. SEQ ID NO: 17 представляє послідовність плазміди pPWT018. SEQ ID NO: 18 представляє послідовність інтегруючої плазміди pPWT018 прямого праймера. SEQ ID NO: 19 представляє послідовність інтегруючої плазміди pPWT018 зворотного праймера. SEQ ID NO: 20 представляє послідовність SIT2-зонда прямого праймера. SEQ ID NO: 21 представляє послідовність SIT2-зонда зворотного праймера. SEQ ID NO: 22 представляє послідовність прямого праймера для ампліфікації касети експресії araABD. SEQ ID NO: 23 представляє послідовність зворотного праймера для ампліфікації касети експресії araABD. Докладний опис винаходу У даному описі і доданій формулі винаходу слова – «містити» і «включати» та, а також їхні похідні, такі як «включає», «містить», «який включає, «який містить», слід трактувати як «які утримують в собі». Тобто ці слова вживаються для вираження можливого включення інших елементів або цілих чисел, не вказаних конкретно там, де це дозволяє контекст. Слова, які використовуються в однині також позначають множину. Наприклад, елемент може означати один елемент або більше ніж один елемент. Різні варіанти здійснення винаходу, описані тут, можна перехресно комбінувати один з одним. Композиція цукрів Композиція цукрів відповідно до винаходу включає глюкозу, арабінозу і галактозу. У способі запропонованому винаходом цукри глюкоза, арабіноза і галактоза переважно перетворюються (конвертуються) на продукт ферментації. У винаході може використовуватися будь-яка композиція цукрів, яка задовольняє вказаним критеріям. У одному з кращих варіантів здійснення винаходу композиція цукрів є гідролізатом одного або більше лігноцелюлозних матеріалів. Лігноцелюлоза в контексті опису включає геміцелюлозу і геміцелюлозні частини біомаси. Лігноцелюлоза включає також лігноцелюлозні фракції біомаси. Придатні лігноцелюлозні матеріали можна знайти в наступному переліку: відходи фруктових садів, чапарель (субтропічна твердолиста чагарникова рослинність), відходи мукомельного виробництва, міські деревні відходи, побутові відходи, лісосічні відходи, відходи від проріджування лісів, деревні культури з короткими періодами ротації, промислові відходи, пшенична солома, вівсяна солома, рисова солома, ячмінна солома, житня солома, льняна солома, оболонки соєвих бобів, рисове лушпиння, рисова солома, кукурудзяний глютеновий корм (кукурудзяна мезга), вівсяне лушпиння, цукрова тростина, грубі рослинні залишки від прибирання кукурудзи, кукурудзяні стебла, стрижні кукурудзяних качанів, листя, які обгортають кукурудзяні качани, просо прутовидне, міскантус (ряд багаторічних трав'янистих рослин сімейства злаків), сорго цукрове, стебла каноли, стебла сої, високотравна рослинність прерій, гамаграс (трипсакум щитовидний; дикий родич кукурудзи), лисохвіст (рід багаторічних лугових трав сімейства злаків); цукрово-буряковий жом, вичавки цитрусових плодів, оболонки насіння, целюлозні відходи при утриманні тварин, скошена трава газонів, бавовник, морські водорості, дерева, дерева з м'якою деревиною, дерева з твердою деревиною, тополі, сосни, чагарники, злакові трави, пшениця, пшенична солома, багаса цукрової тростини, кукурудза, листя, які обгортають кукурудзяні качани, стрижні кукурудзяних качанів, насіння кукурудзи, волокно з насіння, продукти і побічні продукти мокрого або сухого способу помелу зерна, тверді побутові відходи, паперові відходи, відходи скотних дворів, трав'янистий матеріал, сільськогосподарські залишки, лісосічні залишки, тверді побутові відходи, паперові відходи, пульпа, відходи паперових фабрик, гілки, чагарники, очерет, кукурудза, листя, які обгортають кукурудзяні качани, енергетична культура, лісокультура, плоди, квітки, зерно, трава, трав'яниста культура, листя, вторинна кора, глиця, колоди, кореневища, молоді пагони, чагарникові рослини, просо прутовидне, дерево, овочеві культури, шкірка плодів, в'юнкі рослини, цукрово-буряковий жом, пшенична крупа, вівсяне лушпиння, тверда або м'яка деревина, органічні відходи від переробки сільськогосподарської сировини, лісосічні відходи або комбінація будь-яких двох або більше з 3 UA 108853 C2 5 поміж перерахованого вище. Перелік деяких придатних композицій цукрів, отриманих з лігноцелюлози, і композиція гідролізату цукрів приводяться в табл. 1. Перераховані лігноцелюлози включають стрижні кукурудзяних качанів, кукурудзяну клітковину, рисове лушпиння, шкірку дині, цукрово-буряковий жом, пшеничну солому, багасу цукрової тростини, деревину, трав'янисті рослині і вичавки від пресування оливок. Таблиця 1 Перелік композицій цукрів з лігноцелюлозних матеріалів. Gal=галактоза, Xyl=ксилоза, Ara=арабіноза, Man=маноза, Glu=глутамат, Rham=рамноза. Вказані процентний вміст галактози (% Gal) і літературне джерело Лігноцелюлозний матеріал Gal Стрижень кукурудзяного 10 качана а Стрижень кукурудзяного 131 качана b Рисове лушпиння а 9 Рисове лушпиння b 8 Шкірки дині 6 Цукрово-буряковий жом 51 Пшенична солома айдахо 15 Кукурудзяна клітковина 36 Багаса цукрової тростини 14 Грубі рослинні залишки 19 кукурудзи Тамарикс гіллястий 5 (деревина) Евкаліпт (деревина) 22 CWR (трав'яниста рослина) 8 JTW (трав'яниста рослина) 7 MSW (тверді побутові 4 відходи) Очеретянка звичайна (Phalaroides arundinacea) 16 стебло Очеретянка звичайна (Phalaroides arundinacea) 13 насіння Вичавки від пресування 15 оливок Xyl Ara Man 286 36 227 228 160 144 122 120 120 17 249 176 180 24 28 11 209 36 113 24 209 29 118 7 105 165 169 18 11 5 234 209 208 211 396 372 391 11 10 12 16 24 Разом %Gal Джерела інформації: 570 1,7 (1) 663 Glu Rham 19,8 (1) 417 378 361 523 696 697 614 2,2 (1) 2,2 (1) 1,7 (1) 9,8 (2) 2,2 (3) 5,2 (4) 2,3 (5) 370 626 3 493 625 0,7 (7) 8 33 28 3 445 340 311 583 546 515 3,8 (7) 1,4 (7) 1,3 (7) 24 5 20 440 493 0,9 (7) 117 30 6 209 1 379 4,2 (8) 163 28 6 265 1 476 2,7 (9) 111 24 8 329 487 3,1 (9) Середнє 10 15 20 (6) 3,8 З таблиці 1 видно, що у вказаних лігноцелюлозних матеріалах значна кількість (в середньому 3,8%) в загальному вмісті цукрів складає галактоза. Тому конверсія галактози в продукт ферментації має важливе економічне значення. Клітина для ферментації змішаних цукрів Визначення клітини для ферментації змішаних цукрів, яка несе гени araA, araB і araD, подано нижче. Вона здатна ферментувати глюкозу, арабінозу і галактозу. У одному варіанті здійснення винаходу клітина для ферментації змішаних цукрів здатна ферментувати один або більше додаткових цукрів, переважно C5- і/або C6-цукрів. У одному з варіантів здійснення винаходу клітина для ферментації змішаних цукрів містить один або більше xylA- і/або XKS1генів, що дозволяє клітині для ферментації змішаних цукрів ферментувати ксилозу; делеція гена альдозоредуктази (GRE3); гіперекспресія PPP-генів TAL1, TKL1, RPE1 і RKI1 дозволяють збільшити метаболічний потік через пентозофосфатний шлях в клітині. У одному варіанті здійснення винаходу клітина для ферментації змішаних цукрів здатна 4 UA 108853 C2 5 10 15 20 25 30 35 40 45 50 55 60 ферментувати один або більше додаткових цукрів, переважно C5- і/або C6-цукрів. У одному з варіантів здійснення винаходу клітина для ферментації змішаних цукрів має один або більше xylA-, XYL1- і XYL2-генів і/або XKS1-генів, що дозволяє клітині для ферментації змішаних цукрів ферментувати ксилозу; делеція гена альдозоредуктази (GRE3); гіперекспресія PPP-генів TAL1, TKL1, RPE1 і RKI1 дозволяють збільшити метаболічний потік через пентозофосфатний шлях в клітині. У одному з варіантів клітина для ферментації змішаних цукрів є промисловою клітиною, краще – промисловими дріжджами. Промисловій клітині і дріжджовій промисловій клітині можна дати наступне визначення. Умови життя (дріжджових) клітин в промислових процесах істотно відрізняються від умов життя цих клітин в лабораторії. Промислові дріжджові клітини повинні бути здатні добре працювати за самих різних умов зовнішнього середовища, які можуть змінюватися в ході процесу. Такі зміни включають зміни джерела поживних речовин, pH, концентрації етанолу, температури, концентрації кисню тощо, які в своїй сукупності потенційно впливають на зростання клітин і продукування етанолу дріжджами Saccharomyces cerevisiae. За несприятливих промислових умов толерантні до зовнішнього середовища штами повинні демонструвати стійке зростання і продуктивність. Промислові штами дріжджів в більшості випадків стійкіші до перерахованих вище змін умов навколишнього середовища, які можуть відбуватися в місцях використання цих штамів, наприклад, в хлібопекарській промисловості, пивоварній промисловості, виноробстві і виробництві етанолу. У одному варіанті здійснення винаходу промислова клітина для ферментації змішаних цукрів конструюється на основі промислової клітини-хазяїна, причому це конструювання проводиться таким чином, як це описано нижче. Прикладами промислових ® ® ® дріжджів (S. cerevisiae) є Ethanol Red (Fermentis), Fermiol (DSM) і Thermosacc (Lallemand). У одному з варіантів здійснення винаходу клітина для ферментації змішаних цукрів є толерантною до інгібіторів. Толерантність до інгібіторів – це стійкість до інгібуючих сполук. Присутність і рівень вмісту інгібуючих сполук в лігноцелюлозі можуть змінюватись в широких межах залежно від сировини, способу попередньої обробки, процесу гідролізу. Прикладами категорій інгібіторів є карбонові кислоти, фуран і/або фенольні сполуки. Прикладами карбонових кислот є молочна кислота, оцтова кислота або мурашина кислота. Прикладами фурану є фурфурол і гідроксиметилфурфурол. Прикладами фенольних сполук є ванілін, бузкова кислота, ферулова кислота і кумарова кислота. Типові кількості інгібіторів, таких як карбонові кислоти, складають від декількох грамів/літр до 20 грамів/літр або більше, залежно від сировини, попередньої обробки і умов гідролізу. Для фурану ці кількості складають від декількох сотень міліграмів/літр до декількох грамів/літр залежно від сировини, попередньої обробки і умов гідролізу. Для фенольних сполук ці кількості складають від декількох десятків міліграмів/літр до 1 грама/літр залежно від сировини, попередньої обробки і умов гідролізу. Штами для ферментації змішаних цукрів відповідно до винаходу є стійкими до інгібіторів, тобто вони можуть протистояти звичайним інгібіторам при концентрації, яка досягається в типових випадках за загальноприйнятих умов попередньої обробки і гідролізу, так що штами для ферментації змішаних цукрів можуть знайти широке застосування, тобто вони показують широкий спектр можливостей застосування для різних видів сировини, різних способів попередньої обробки і різних умов гідролізу. У одному варіанті здійснення винаходу промислова клітина для ферментації змішаних цукрів конструюється на основі толерантної до інгібіторів клітини-хазяїна, причому конструювання клітини проводиться, як це описано нижче. Толерантні до інгібіторів клітинихазяїні можуть підбиратися скринінгом штамів, які вирощують на матеріалах, що містять інгібітори, таких як представлені Kadar et al. у Appl. Biochem. Biotechnol. (2007), Vol. 136-140, 847-858; авторами був відібраний толерантний до інгібіторів штам S. cerevisiae ATCC 26602. У одному з варіантів здійснення винаходу клітина для ферментації змішаних цукрів не містить маркера. У контексті опису термін «маркер» відноситься до гена, який кодує ознаку або фенотип, який робить можливим відбір або скринінг з метою відбору клітини-хазяїна, що містить маркер. «Не містить маркера» означає, що в клітині для ферментації змішаних цукрів маркери переважно відсутні. Відсутність маркера є особливо вигідним в тому випадку, якщо при конструюванні клітини для ферментації змішаних цукрів використовувалися антибіотикові маркери, які згодом були видалені. Видалення маркерів може здійснюватися будь-яким придатним методом попереднього рівня техніки, наприклад, внутрішньомолекулярною рекомбінацією. Придатний метод видалення маркерів описаний в прикладах. Клітина для ферментації змішаних цукрів може проявляти здатність до конверсії рослинної біомаси, целюлози, геміцелюлоз, пектинів, рамнози, галактози, фруктози, мальтози, мальтодекстринів, рибози, рибулози або крохмалю, похідних крохмалю, сахарози, лактози і 5 UA 108853 C2 5 10 15 20 25 30 35 40 45 50 55 60 гліцерину, наприклад, у придатні для ферментації цукри. Отже, клітина для ферментації змішаних цукрів може експресувати один або більше ферментів, таких як целюлаза (ендоцелюлаза або екзоцелюлаза) і геміцелюлаза (ендо- або екзоксиланаза або арабіназа), необхідна для конверсії целюлози в мономери глюкози і геміцелюлози в мономери ксилози і арабінози; пектиназа, здатна конвертувати пектини в глюкуронову кислоту і галактуронову кислоту, або амілаза, здатна конвертувати крохмаль в мономери глюкози. Крім того, клітина для ферментації змішаних цукрів може проявляти ферментативну активність, потрібну для конверсії пірувату в бажаний продукт ферментації, такий як етанол, бутанол, молочна кислота, 3-гідроксипропіонова кислота, акрилова кислота, оцтова кислота, бурштинова кислота, лимонна кислота, фумарова кислота, яблучна кислота, ітаконова кислота, амінокислота, 1,3-пропандіол, етилен, гліцерин, -лактамний антибіотик або цефалоспорин. У одному з варіантів здійснення винаходу клітина для ферментації змішаних цукрів є клітиною, яка за своєю природою здатна викликати спиртове бродіння, бажано – анаеробне спиртове бродіння. Клітина для ферментації змішаних цукрів бажано проявляє високу стійкість до етанолу, високу стійкість до низького pH (тобто здатна до зростання при pH нижче приблизно 5, приблизно 4, приблизно 3 або приблизно 2,5) і органіки і/або високої стійкості до підвищених температур. Будь-яка з вищеперелічених характеристик або активностей клітини для ферментації змішаних цукрів може від природи бути присутньою в клітині або може бути введена в клітину або змінена за допомогою генетичної модифікації. Конструювання штаму для ферментації змішаних цукрів Гени можуть вводитися в клітину для ферментації змішаних цукрів шляхом введення в клітину-хазяїна: a) кластера, який складається з PPP-генів TAL1, TKL1, RPE1 і RKI1, під контролем сильних промоторів b) кластера, який складається з xylA-гена і XKS1-гена, обидва під контролем конститутивних промоторів c) кластера, який складається з генів araA, araB і araD, і/або кластера з xylA-гена і/або XKS1гена і d) делеції гена альдозоредуктази і адаптивної еволюції для отримання клітини для ферментації змішаних цукрів. Вказана клітина може бути сконструйована методами рекомбінантної експресії. Рекомбінантна експресія Клітина винаходу є рекомбінантною клітиною. Тобто клітина запропонована винаходом містить або трансформується або генетично модифікується нуклеотидною послідовністю, яка від природи не зустрічається в даній клітині. Методи рекомбінантної експресії ферментів в клітині, а також додаткових генетичних модифікацій клітини запропонованої винаходом добре відомі фахівцям, кваліфікованим в даній галузі техніки. У типових випадках такі методи включають трансформацію клітини нуклеїнокислотним конструктом, який містить релевантну послідовність. Такі методи відомі, наприклад, із стандартних довідників, таких як Sambrook & Russel (2001) «Molecular Cloning: A Laboratory Manual» (3rd edition), Cold Spring Harbor Laboratory, Cold Spring Harbor Laboratory Press, або F. Ausubel et al., eds., «Current protocols in molecular biology», Green Publishing and Wiley Interscience, New York (1987). Методи трансформації і генетичної модифікації грибних клітин-хазяїнів відомі, наприклад, з EP-A-0635574, WO 98/46772, WO 99/60102, WO 00/37671, WO90/14423, EP-A-0481008, EP-A-0635574 і US 6 265 186. У типових випадках нуклеїнокислотний конструкт може бути плазмідою, наприклад, малокопійною плазмідою або мультикопійною плазмідою. Клітина запропонована даним винаходом може містити одну або багато копій нуклеотидної послідовності, яка кодує фермент, наприклад, за рахунок багатьох копій нуклеотидного конструкта або за рахунок використання конструкта, який містить велику кількість копій послідовності, що кодує фермент. Нуклеїнокислотний конструкт може підтримуватися епісомально і, отже, містити послідовність для автономної реплікації, таку як аутосомно-реплікована послідовність. Придатний епісомний нуклеїнокислотний конструкт може мати як основу, наприклад, дріжджові 2µ- або pKD1-плазміди (Gleer et al., 1991, Biotechnology 9: 968-975), або AMAплазміди (Fierro et al., 1995, Curr Genet. 29: 482-489). Альтернативно, кожен нуклеїнокислотний конструкт може інтегруватися в одній або більше копіях в геном клітини. Інтеграція в геном клітини може відбуватися випадково шляхом негомологічної рекомбінації, але бажано нуклеїнокислотний конструкт може інтегруватися в геном клітини гомологічною рекомбінацією, що є добре відомим з рівня техніки (див., напр., WO90/14423, EP-A-0481008, EP-A-0635574 і US 6 UA 108853 C2 5 10 15 20 25 30 35 40 45 50 55 60 6 265 186). Більшість епісомальних плазмід або 2µ-плазмід є відносно нестабільними і втрачаються -2 приблизно в 10 або більше клітинах в кожному поколінні. Навіть за умов селективного зростання тільки від 60% до 95% клітин зберігають епісомальну плазміду. Копійність більшості + епісомних плазмід коливається від 10 до 40 на клітину cir штамів-хазяїнів. Проте плазміди нерівномірно розподіляються серед клітин, і спостерігається значна варіабельність в кількості копій на клітину в популяціях. Штами, трансформовані інтеграційними плазмідами, є високостабільними, навіть за відсутності селективного тиску. Проте втрата плазмід може -3 -4 відбуватися приблизно з 10 – 10 частотою в результаті гомологічної рекомбінації між тандемною повтореною ДНК, приводячи до випетлювання послідовності вектора. Переважно конструкція вектора у випадку стабільної інтеграції така, що після втрати селективних маркерних генів (яка також відбувається в результаті внутрішньомолекулярної гомологічної рекомбінації) вказане випетлювання інтегрованого конструкта стає неможливим. Тому гени переважно інтегруються стабільно. Стабільна інтеграція в контексті опису визначається як інтеграція в геном, в якій випетлювання інтегрованого конструкта стає неможливим. Бажано селективні маркери відсутні. У типових випадках послідовність, яка кодує фермент, функціонально зв'язуватиметься з однією або більше нуклеїнокислотними послідовностями, здатними забезпечити або полегшити транскрипцію і/або трансляцію послідовності, яка кодує фермент. Термін «функціонально зв'язуватися» відноситься до положення контакту, в якому описані компоненти знаходяться у взаємодії, що дозволяє їм функціонувати властивим їм чином. Наприклад, промотор або енхансер функціонально пов'язаний з кодуючою послідовністю, при цьому вказаний промотор або енхансер впливає на транскрипцію кодуючої послідовності. У контексті опису термін «промотор» відноситься до фрагмента нуклеїнової кислоти, який виконує функцію контролю транскрипції одного або більше генів, які локалізуються перед (за напрямком транскрипції) сайтом ініціації транскрипції гена, і структурно ідентифікується присутністю сайту скріплення ДНК-залежної РНК-полімерази, сайтів ініціації транскрипції та іншої ДНК-послідовності, відомих фахівцеві в даній галузі техніки. «Конститутивний» промотор – це промотор, який є активним за більшості умов зовнішнього середовища і умов розвитку. «Індуцибельний» промотор – це промотор, який є активним при регулюванні умов зовнішнього середовища або умов розвитку. Промотор, який може використовуватися для досягнення експресії нуклеотидної послідовності, яка кодує фермент згідно даному винаходу, може не бути нативним промотором експресованої нуклеотидної послідовності, яка кодує фермент, тобто промотором, гетерологічним до нуклеотидної послідовності (кодуючої послідовності), з якою він функціонально зв'язаний. Проте промотор може бути гомологічним, тобто ендогенним, по відношенню до клітини-хазяїна. Промотори є широко доступними і відомими кваліфікованому фахівцеві. Відповідні приклади таких промоторів включають, наприклад, промотори з гліколітичних генів, такі як промотори фосфофруктокінази (PFK), триозо-фосфатізомерази (TPI), гліцеральдегід-3фосфатдегідрогенази (GPD, TDH3 або GAPDH), піруваткінази (PYK), фосфогліцераткінази (PGK) з дріжджів або міцеліальних грибів; докладнішу інформацію про такі промотори з дріжджів можна знайти в (WO 93/03159). Іншими корисними промоторами є промотори генів, які кодують рибосомальні білки, промотор гена лактази (LAC4), промотори алкогольдегідрогенази (ADHl, ADH4 і ін.) і промотор енолази (ENO). Інші промотори, як конститутивні, так і індуцибельні, і енхансери або розташовані вище активуючі послідовності відомі кваліфікованому в даній галузі техніки фахівцеві. Промотори, які використовуються в клітинаххазяїнах винаходу, можуть бути за необхідності піддані модифікації з метою впливу на їх функцію (характеристики) контролю. Придатні в даному контексті промотори включають як конститутивні, так і індуцибельні природні промотори, а також сконструйовані промотори, які добре відомі кваліфікованому в даній галузі техніки фахівцеві. Придатними промоторами в еукаріотичних клітинах-хазяїнах можуть бути GAL7, GAL10 або GAL1, CYC1, HIS3, ADH1, PGL, PH05, GAPDH, ADC1, TRP1, URA3, LEU2, ENO1, TPI1 і AOX1. Інші придатні промотори включають PDC1, GPD1, PGK1, TEF1 і TDH3. У клітині запропонованій винаходом 3'-кінець послідовності нуклеотидів, яка кодує фермент, бажано пов'язаний функціонально з послідовністю термінатора транскрипції. Бажано щоб послідовність термінатора функціонувала у відібраній клітині-хазяїні, такій як, наприклад, відібрані види дріжджів. У будь-якому випадку вибір термінатора не є критичним; він може бути отриманий, наприклад, з будь-якого дріжджового гена, хоча іноді термінатори можуть працювати, навіть якщо вони отримані не з гена дріжджів, а з гена іншого еукаріота. Зазвичай 7 UA 108853 C2 5 10 15 20 25 30 35 40 45 50 55 60 послідовність нуклеотидів, яка кодує фермент, містить термінатор. Бажано такі термінатори комбінуються з мутаціями щоб уникнути антисмислового розпаду мРНК в клітині-хазяїні винаходу (див., наприклад, Shirley et al., 2002, Genetics 161:1465-1482). Послідовність, яка термінує транскрипцію, переважно включає також сигнал поліаденилювання. У нуклеїнокислотному конструкті, придатному для використання у винаході, необов'язково може бути присутнім селектований маркер. У контексті опису термін «маркер» відноситься до гена, який кодує ознаку або фенотип, який дозволяє проводити відбір або скринінг на клітинухазяїна, яка містить маркер. Маркерний ген може бути геном стійкості до антибіотиків, причому відповідний антибіотик може використовуватися для відбору трансформованих клітин серед нетрансформованих клітин. Приклади придатних маркерів стійкості до антибіотиків включають, наприклад, дигідрофолатредуктазу, гігроміцин-B-фосфотрансферазу, 3'-O-фосфотрансферазу II (стійкість до канаміцину, неоміцину і G418). Маркери стійкості до антибіотику можуть бути найбільш зручними для трансформації поліплоїдних клітин-хазяїнів. Можуть використовуватися і маркери нестійкості до антибіотиків, такі як ауксотрофні маркери (URA3, TRPl, LEU2) або TPI ген з S. pombe (описаний Russell P.R., 1985, Gene 40: 125-130). У одному з кращих варіантів здійснення винаходу клітини-хазяїни, трансформовані нуклеїнокислотними конструктами, не містять маркерних генів. Методи конструювання рекомбінантних мікробних клітин-хазяїнів, які не містять маркерних генів, розкриваються в EP-A-O 635 574 і ґрунтуються на використанні двонаправлених маркерів, таких як ген amdS (ацетамідази) з A. nidulans або гени дріжджів URA3 і LYS2. Альтернативно, скринований маркер, такий як зелений флуоресціюючий білок, lacL, люцифераза, хлорамфенікол-ацетилтрансфераза, бета-глюкуронідаза, може вводитися в нуклеїнокислотні конструкти винаходу, що дозволяє проводити скринінг на трансформовані клітини. Необов'язкові додаткові елементи, які можуть бути присутніми в нуклеїнокислотних конструктах, придатних для використання у винаході, включають, але весь перелік не обмежується тільки названими тут, одну або більше з лідерних послідовностей, енхансерів, чинників інтеграції і/або репортерних генів, інтронів, центромер, теломер і/або послідовностей ділянок прикріплення до ядерного матриксу (MAR). Крім цього, нуклеїнокислотні конструкти запропоновані винаходом можуть включати послідовність для автономної реплікації, таку як ARS-послідовність. Таким чином, процес рекомбінації може здійснюватися відомими методами рекомбінації. Кваліфікованим в даній галузі техніки фахівцям відомі різні прийоми для експресії і гіперекспресії ферментів в клітині запропонованій винаходом. Зокрема, може бути здійснена гіперекспресія ферменту шляхом збільшення числа копій гена, який кодує фермент в клітиніхазяїні, наприклад, шляхом інтеграції додаткових копій гена в геном клітини-хазяїна, шляхом експресії гена з вектора епісомальної мультикопійної експресії або шляхом введення вектора епісомальної експресії, який містить множинні копії гена. Альтернативно, гіперекспресія ферментів в клітинах-хазяїнах винаходу може досягатися за допомогою промотору, який не є нативним послідовності, що кодує гіперекспресуємий фермент, тобто промотору, який є гетерологічним кодуючої послідовності, з якою він функціонально зв'язаний. Хоча промотор бажано є гетерологічним щодо кодуючої послідовності, з якою він функціонально зв'язаний, але краще також, якщо промотор є гомологічним, тобто ендогенним, щодо клітини-хазяїна. Переважно гетерологічний промотор здатний проводити підвищений стійкий рівень транскрипту, що включає кодуючу послідовність (або здатний проводити більше молекул транскрипту, тобто молекул мРНК, в одиницю часу), чим промотор, що є нативним кодуючої послідовності. Придатні в даному контексті промотори включають як конститутивні, так і індуцибельні природні промотори, а також сконструйовані промотори. У одному з варіантів здійснення винаходу клітина для ферментації змішаних цукрів не містить маркерів, що означає, що ауксотрофні або домінантні маркери, зокрема, маркери стійкості до антибіотику не присутні в геномі клітини або поза хромосомами. Кодуюча послідовність, яка використовується для гіперекспресії вищезазначених ферментів, бажано може бути гомологічною до клітини-хазяїна за винаходом. Проте можуть також використовуватися кодуючі послідовності, які є гетерологічними щодо клітини-хазяїна. Гіперекспресія ферменту в контексті продукування цього ферменту в генетично модифікованій клітині означає, що фермент продукується за підвищеного рівня специфічної ферментативної активності в порівнянні з немодифікованою клітиною-хазяїном за ідентичних умов. Зазвичай це означає, що ферментативний активний білок (або білки у випадку мультисубодиничних ферментів) продукується в підвищених кількостях або швидше за підвищеного стійкого рівня у порівнянні з немодифікованою клітиною-хазяїном за ідентичних 8 UA 108853 C2 5 10 15 20 25 30 35 40 45 50 55 60 умов. Так само, це зазвичай означає, що мРНК, яка кодує ферментативний активний білок, утворюється в підвищених кількостях або швидше за підвищеного стійкого рівня в порівнянні з немодифікованою клітиною-хазяїном за ідентичних умов. Бажано в клітині-хазяїні запропонованій винаходом яка піддається гіперекспресії фермент гіперекспресується, щонайменше, приблизно в 1,1, приблизно в 1,2, приблизно в 1,5, приблизно в 2, приблизно в 5, приблизно в 10 або приблизно в 20 разів у порівнянні з штамом, який є генетично ідентичним, за винятком генетичної модифікації, яка викликає гіперекспресію. Саме собою зрозуміло, що ці рівні гіперекспресії можна застосувати і до стійкого рівня ферментної активності, до стійкого рівня ферментного білка, а також до стійкого рівня транскрипту, який кодує фермент. Адаптація Адаптація – це еволюційний процес, в ході якого популяція стає краще пристосованого (адаптованою) до умов середовища або місць існування. Цей процес має місце протягом від декількох до багатьох генерацій і є одним з основних явищ в біології. Термін «адаптація» може також відноситися до специфічної ознаки, яка є особливо важливою для виживання організму. Такого роду адаптації приводять до мінливості популяції за рахунок появи краще пристосованих форм, здатних успішніше народжувати потомство, за рахунок природного відбору. Зміну умов навколишнього середовища змінюють результати природного відбору, які впливають на селективні переваги подальших адаптацій, які покращують пристосовність організму до нових умов. У випадку екстремальних змін навколишнього середовища виникнення і закріплення сприятливих адаптацій можуть бути дуже важливими для виживання. Велике число різних чинників, таких як, наприклад, доступність поживних речовин, температура, доступність кисню тощо, може стимулювати адаптивну еволюцію. Пристосованість Існує чіткий взаємозв'язок між здатністю до адаптації (здатністю організму виживати і народжувати потомство за даної сукупності умов місця існування) і пристосованістю. Пристосованість – це оцінка та інструмент прогнозування швидкості природного відбору. В результаті природного відбору відносна частота альтернативних фенотипів варіюватиме в часі, якщо вони успадковуватимуться. Генетичні зміни Якщо природний відбір впливає на генетичну варіабельність популяції, то генетичні зміни є основоположним механізмом його впливу. Під цим мають на увазі, що популяція генетично адаптується до умов навколишнього середовища. Генетичні зміни можуть виявлятися у видимих структурах або можуть регулювати фізіологічну активність організму так, щоб останній пристосувався до зміненого місця існування. Адаптивна еволюція Клітини для ферментації змішаних цукрів в процесі свого отримання піддаються адаптивній еволюції. Клітина запропонована винаходом може бути адаптована до утилізації цукрів шляхом відбору мутантів, що або виникають спонтанно, або є індукованими (наприклад, радіацією або хімікатами), для зростання на бажаному цукрі, бажано як єдиному джерелі вуглецю, краще – за анаеробних умов. Відбір мутантів може здійснюватися методами, які включають послідовне перенесення культур, як описано, наприклад, в Kuyper et al. (2004, FEMS Yeast Res. 4: 655-664), або культивуванням за селективного тиску в хемостатній культурі. Наприклад, в кращій клітиніхазяїні запропонованій винаходом, щонайменше, одна з описаних вище генетичних модифікацій, включаючи модифікації, отримані відбором мутантів, надає клітині-хазяїну здатності до зростання на ксилозі як джерелі вуглецю, бажано – як єдиному джерелі вуглецю і краще – за анаеробних умов. Бажано клітина не продукує, в основному, ксилітолу, наприклад, продукує ксилітол в кількостях, нижчих від межі виявлення, або, наприклад, в кількостях менше приблизно 5, менше 2, менше 1, менше 0,5 або менше 0,3% спожитого вуглецю в перерахунку на молярну основу. Адаптивна еволюція описана також, наприклад, в Wisselink H.W. et al., Applied and Environmental Microbiology, Aug. 2007, р. 4881–4891. У одному варіанті адаптивної еволюції застосовується режим, який включає повторне періодичне культивування з повторними циклами послідовного зростання на різних середовищах, наприклад, на трьох середовищах різного складу (глюкоза, ксилоза і арабіноза; ксилоза і арабіноза). Див. Wisselink et al. (2009) Applied and Environmental Microbiology, Feb. 2009, р. 907–914. Трансформація дріжджів і генетична стабільність Початок генної інженерії, тобто трансформації дріжджових клітин рекомбінантною ДНК, вперше було здійснено в 1978 р. [Beggs, 1978; Hinnen et al., 1978]. З того часу метод 9 UA 108853 C2 5 10 15 20 25 30 35 40 45 50 55 60 рекомбінантних ДНК в дріжджах став загальновизнаним. На даний час доступними є багато різних векторних конструктів. В більшості своїй це плазмідні вектори, які називаються човниковими векторами, містять генетичний матеріал з векторів E.coli, що складаються з ориджину реплікації (точки початку реплікації) і селектованого маркера (часто гена β-lactamase – ampR), що робить можливою їх реплікацію в E.coli перед трансформацією в дріжджові клітини. До того ж човникові вектори містять селектований маркер для селекції в дріжджах. Маркерами можуть бути гени, які кодують ферменти для синтезу конкретної амінокислоти або нуклеотиду, так що клітини, які несуть відповідну геномну делецію (або мутацію) для створення ауксотрофії або автотрофії, комплементуються. Альтернативно, ці вектори містять гетерологічні домінантні маркери стійкості, які надають рекомбінантним дріжджовим клітинам (тобто клітинам, до яких введена рекомбінантна ДНК і які експресують маркерний ген) стійкості до певних антибіотиків, таких як g418 (генетицин), гігроміцин B або флеоміцин. На додачу до цього, вказані вектори можуть містити послідовність (комбінованих) сайтів рестрикції (сайт множинного клонування або MCS), яка дозволяє клонувати чужорідну ДНК в цих сайтах, хоча існують і альтернативні методи. Традиційний чотири типи човникових векторів можуть розрізнятися за відсутністю або наявністю додаткових генетичних елементів. - Інтеграційні плазміди (YIp), які в результаті гомологічної рекомбінації інтегруються в геномі хазяїна в локус маркера або іншого гена, коли останній відкривається рестрикцією, і лінеаризована ДНК використовується для трансформації дріжджових клітин. В більшості випадків це призводить до присутності однієї копії чужорідної ДНК, інсеркованої в цей конкретний сайт в геномі. - Епісомальні плазміди (YEp), які несуть частину ДНК-послідовності плазміди 2 µ, необхідну для автономної реплікації в дріжджових клітинах. Множинні копії трансформованої плазміди розмножуються в дріжджовій клітині і підтримуються як епісоми. - Плазміди, які автономно реплікуються (YRp), які несуть ориджин реплікації дріжджів (ARS = послідовність, яка автономно реплікується), що дозволяє трансформованим плазмідам розмножуватися в декілька сотень разів. - CEN плазміди (YCp), які на додачу до ARS-послідовності несуть ще центромерну послідовність (отриману з однієї з ядерних хромосом), яка зазвичай гарантує стабільну мітотичну сегрегацію і зазвичай скорочує число копій самореплікувальної плазміди до однієї. Ці плазміди вводяться в дріжджові клітини трансформацією. Трансформація дріжджових клітин може досягатися декількома різними методами, такими як методи пермеабілізації клітин ацетатом літію (Ito et al., 1983) і електропорації. При промисловому застосуванні рекомбінантних мікроорганізмів найбільш важливою проблемою є нестабільність плазмід. Нестабільність – це схильність трансформованих клітин до втрати своїх сконструйованих властивостей унаслідок змін або втрати плазмід. Ця проблема детально обговорюється в Zhang et al., (Plasmid stability in recombinant Saccharomyces cerevisiae. Biotechnology Advances, Vol. 14, No. 4, pp. 401-435, 1996). Штами, трансформовані інтеграційними плазмідами, є високостабільними навіть за відсутності селективного тиску (Sherman, F. http://dbb.urmc.rochester.edu/labs/sherman_f/yeast/9.html і див. Джерела інформації: в описі). Гетерологічна ДНК зазвичай вводиться в організм у формі позахромосомних плазмід (YEp, YCp і YRp). На жаль, було встановлено як у випадку бактерій, так і дріжджів, що нові характеристики не можуть зберігатися, особливо за відсутності постійного селективного тиску. Причиною цього є нестабільність сегрегації гібридної плазміди, яка виникає, коли рекомбінантні клітини ростуть протягом тривалого періоду часу. Це веде до гетерогенності популяції і клональної варіабельності і зрештою – до популяції клітин, в якій більшість клітин втратили свої властивості, отримані при трансформації. Якщо використовуються вектори з ауксотрофними маркерами, то культивування в збагачених середовищах часто призводить до швидкої втрати вектора, оскільки вектор утримується тільки в мінімальних середовищах. Альтернативний варіант з використанням домінантних маркерів стійкості до антибіотику часто не сумісний з виробничими процесами. Застосування антибіотиків може бути небажаним з погляду реєстрації (вірогідність того, що слідові кількості антибіотиків опиняються в кінцевому продукті) або із економічних причин (витрати на використання антибіотиків в промисловому масштабі). Втрата векторів створює проблеми у великомасштабних виробничих ситуаціях. Існують і альтернативні методи введення ДНК в дріжджові клітини, такі як використання інтегруючих плазмід (YIp). ДНК інтегрується в геном хазяїна рекомбінацією, що приводить до високої стабільності. (Caunt, P. Stability of recombinant plasmids in yeast. Journal of Biotechnology 9(1988) 173 – 192). Авторами даного винаходу встановлено, що метод інтеграції, який використовує 10 UA 108853 C2 5 10 15 20 25 30 35 40 45 50 55 60 транспозони хазяїна, є хорошою альтернативою. Транспозони У одному з варіантів здійснення винаходу клітина може містити більше від однієї копії бажаного гена (генів). Наприклад, два або більше гени ксилоізомерази або генів ксилозоредуктази і ксилітолдегідрогенази можуть інтегруватися в геном клітини для ферментації змішаних цукрів. Це може здійснюватися будь-яким методом, відомим з рівня техніки, який призводить до введення генів. У кращому варіанті це може здійснюватися за допомогою вектора, який містить частини, гомологічні до послідовностей (транспозон) клітинихазяїна, які повторюються. Якщо клітина-хазяїн є дріжджовою клітиною, то відповіднимипослідовностями, що повторюються, є довгі кінцеві повтори (LTR) Ty-елемента, відомі як дельта-послідовність. Елементи Ty підрозділяються на дві вельми схожих підродини, які називаються Ty1 і Ty2. Ці елементи мають довжину близько 6 тис. пар основ-нуклеотидів (т.п.н.) і пов'язані з довгими кінцевими повторами (LTR) – послідовностями, які містять близько 335 пар основ (Boeke JD et al., The Saccharomyces cerevisiae Genome Contains Functional and Nonfunctional Copies of Transposon Ty1. Molecular and Cellular Biology, Apr. 1988, р. 1432-1442 Vol. 8, No. 4). У штамі з повністю секвенованим геномом S. cerevisiae S288c транспозонами, які найчастіше зустрічаються є Ty1 (31 копія) і Ty2 (13 копій) (Gabriel A, Dapprich J, Kunkel M, Gresham D, Pratt SC, et al., (2006) Global mapping of transposon location. PLoS Genet 2(12): e212.doi:10.1371 /journal.pgen.0020212). Ці транспозони складаються з двох відкритих рамок зчитування (ORF), які перекриваються і кожна з яких кодує декілька білків. Кодуючі області фланковані вищезазначеними приблизно ідентичними LTR. Інші більш несхожі Ty-елементи, які рідше зустрічаються в S. cerevisiae, включають Ty3, Ty4 і Ty5. Для кожної родини повнорозмірних Tyелементів існує порядок величини крупніших solo-елементів LTR, розсіяних по геному. Передбачається, що вони утворюються в результаті LTR–LTR рекомбінації повнорозмірних елементів з випетлюванням внутрішніх областей, які кодують білки. Механізм ретротранспозиції Ty-ретротранспозона використаний для інтеграції множинних копій по геному (Boeke et al., 1988; Jacobs et al., 1988). Довгі кінцеві повтори (LTR) Ty-елемента, відомі як дельта-послідовності, також є хорошими мішенями для інтеграції гомологічною рекомбінацією, оскільки вони зустрічаються приблизно в 150-200 копіях, які асоціюються або з Ty, або з solo-сайтами (Boeke, 1989; Kingsman and Kingsman, 1988). (Parekh R.N. (1996): An Integrating Vector for Tunable, High Copy, Stable Integration into the Dispersed Ty DELTA Sites of Saccharomyces cerevisiae. Biotechnol. Prog. 1996, 12, 16-21). Клітина-хазяїн Клітина-хазяїн може бути будь-якою клітиною-хазяїном, придатною для отримання корисного продукту. Клітина винаходу може бути будь-якою придатною клітиною, такою як прокаріотична клітина, наприклад, бактерія, або еукаріотична клітина. У типових випадках клітина винаходу є еукаріотичною клітиною, наприклад, клітиною дріжджів або міцеліального гриба. У контексті опису дріжджі визначаються як еукаріотичні мікроорганізми і включають всі види дріжджів підвідділу Eumycotina (Alexopoulos, C. J.,1962, In: Introductory Mycology, John Wiley & Sons, Inc., New York), які ростуть переважно в одноклітинній формі. Дріжджі можуть рости або брунькуванням одноклітинного талому, або можуть рости діленням організму. Кращі дріжджі як клітини винаходу можуть відноситися до роду Saccharomyces, Kluyveromyces, Candida, Pichia, Schizosaccharomyces, Hansenula, Kloeckera, Schwanniomyces або Yarrowia. Бажано дріжджами є дріжджі, здатні до анаеробної ферментації або до ферментації за умов обмеженої кількості кисню, краще – здатні до анаеробного спиртового бродіння. У контексті опису міцеліальні гриби визначаються як еукаріотичні мікроорганізми, які включають всі міцеліальні форми підвідділу Eumycotina. Ці гриби характеризуються вегетативним міцелієм, який складається з хітину, целюлози і інших складних полісахаридів. Міцеліальні гриби, придатні для застосування як клітина даного винаходу, морфологічно, фізіологічно і генетично відрізняються від дріжджів. Можуть використовуватися переважно клітини міцеліальних грибів, оскільки більшість грибів не вимагає стерильних умов для розмноження і є несприйнятливими до інфікування бактеріофагом. Вегетативне зростання міцеліальних грибів відбувається за рахунок подовження гифів, а вуглецевий катаболізм більшості міцеліальних грибів є облігатним аеробним. Кращі міцеліальні гриби як клітинихазяїни за даним винаходом можуть відноситися до роду Aspergillus, Trichoderma, Humicola, Acremoniurra, Fusarium або Penicillium. Краща клітина міцеліальних грибів може бути клітиною Aspergillus niger, Aspergillus oryzae, Penicillium chrysogenum або клітиною Rhizopus oryzae. 11 UA 108853 C2 5 10 15 20 25 30 35 40 45 50 55 60 У одному варіанті винаходу клітина-хазяїн може бути клітиною дріжджів. Бажано клітина-хазяїн є промисловою клітиною-хазяїном, краще – промисловою дріжджовою клітиною. Промислову клітину-хазяїна і промислову дріжджову клітину можна визначити таким чином. Умови життя дріжджових клітин в промислових процесах істотно відрізняються від умов їх існування в лабораторії. Промислові дріжджові клітини повинні бути здатними добре працювати за багатьох умов зовнішнього середовища, які можуть варіювати в ході процесу. Таке варіювання включає зміни джерел поживних речовин, pH, концентрації етанолу, температури, концентрації кисню тощо, які в своїй сукупності потенційно впливають на зростання клітин і продукування етанолу дріжджами Saccharomyces cerevisiae. У шкідливихпромислових умовах толерантні до зовнішнього середовища штами повинні показувати інтенсивне зростання і продуктивність. В більшості випадків промислові штами дріжджів є стійкішими до вказаних змін умов зовнішнього середовища, які можуть відбуватися в місцях використання цих штамів, наприклад, в хлібопекарській промисловості, пивоварній промисловості, виноробстві і у виробництві етанолу. Прикладами промислових дріжджів (S. cerevisiae) є Ethanol Red® (Fermentis), Fermiol® (DSM) і Thermosacc® (Lallemand). У одному з варіантів здійснення винаходу хазяїн є толерантним до інгібіторів. Толерантні до інгібіторів клітини-хазяїни можуть відбиратися шляхом скринінгу штамів на зростання на матеріалах, які містять інгібітори, такі як представлені в Kadar et al. у Appl. Biochem. Biotechnol. (2007), Vol. 136-140, 847-858, з яких був відібраний стійкий до інгібіторів штам S. cerevisiae ATCC 26602. Бажано клітина-хазяїн є промисловою і стійкою до інгібіторів. Гени araA, araB і araD Клітина винаходу здатна утилізувати арабінозу. Отже, клітина винаходу здатна конвертувати L-арабінозу в L-рибулозу і/або ксилулозо-5-фосфат, і/або в бажаний продукт ферментації, наприклад, в один із згаданих в описі. Організми, наприклад, штами S. cerevisiae, які здатні продукувати етанол з L-арабінози, можуть бути отримані модифікацією клітини шляхом введення до неї генів araA (Lарабінозоізомерази), araB (L-рибулокінази) і araD (L-рибулозо-5-P4-епімерази) з придатного джерела. Вказані гени можуть вводитися в клітину запропоновану винаходом для того, щоб надати їй здатності до утилізації арабінози. Такий підхід описаний в WO2003/095627. З цією метою можуть використовуватися гени araA, araB і araD з Lactobacillus plantarum, як описано в WO2008/041840. Також можуть використовуватися ген araA з Bacillus subtilis і гени araB і araD з Escherichia coli, як описано в EP1499708. У іншому варіанті здійснення винаходу гени araA, araB і araD можуть бути отримані, щонайменше, із одного з родів Clavibacter, Arthrobacter і/або Gramella, зокрема, одного з Clavibacter michiganensis, Arthrobacter aurescens і/або Gramella forsetii, як описано в WO 2009011591. PPP-гени Клітина винаходу може містити одну або більше генетичних модифікацій, які збільшують метаболічний потік через пентозофосфатний шлях. Зокрема, генетична модифікація(ї) може(можуть) призводити до збільшеного потоку через неокислювальну частину пентозофосфатного шляху. У контексті опису під «генетичною модифікацією», що викликає збільшений потік через неокислювальну частину пентозофосфатного шляху, розуміють модифікацію, яка збільшує потік, щонайменше, приблизно в 1,1, приблизно в 1,2, приблизно в 1,5, приблизно в 2, приблизно в 5, приблизно в 10 або приблизно в 20 разів в порівнянні з потоком в штамі, який є генетично ідентичним, за винятком генетичної модифікації, яка викликає збільшений потік. Потік через неокислювальну частину пентозофосфатного шляху може бути визначений шляхом вирощування модифікованого хазяїна на ксилозі як єдиному джерелі вуглецю, визначення питомої швидкості споживання ксилози і віднімання питомої швидкості продукування ксилітолу від питомої швидкості споживання ксилози, якщо ксилітол взагалі продукується. Проте потік через неокислювальну частину пентозофосфатного шляху пропорційний швидкості росту на ксилозі як єдиному джерелі вуглецю, бажано – швидкості анаеробного росту на ксилозі як єдиному джерелі вуглецю. Існує лінійна залежність між швидкістю росту на ксилозі як єдиному джерелі вуглецю (µmax) і потоком через неокислювальну частину пентозофосфатного шляху. Питома швидкість споживання ксилози (Q s) дорівнює швидкості росту (µ), розділеній на вихід біомаси на цукрі (Y xs), оскільки вихід біомаси на цукрі є постійною величиною (за заданої сукупності умов: анаеробний режим, поживне середовище, pH, генетичний фон штаму та ін. (тобто Q s = µ/Yxs). Таким чином, збільшений потік через неокислювальну частину пентозофосфатного шляху може бути виведений із збільшення максимальної швидкості росту за вказаних умов за відсутності транспорту (поглинання лімітоване). 12 UA 108853 C2 5 10 15 20 25 30 35 40 45 50 55 60 Одна або більше генетичних модифікацій, які збільшують потік через пентозофосфатний шлях, можуть вводитися в клітину-хазяїна різними шляхами. Ці шляхи включають, наприклад, досягнення підвищених стійких рівнів активності ксилулозокінази і/або одного або більше ферментів неокислювальної частини пентозофосфатного шляху, і/або зниженого стійкого рівня активності неспецифічної альдозоредуктази. Вказані зміни стійких рівнів активності можуть бути викликані підбором мутантів (спонтанних або індукованих хімічними речовинами або радіацією) і/або методом рекомбінантних ДНК, наприклад, гіперекспресією або інактивацією відповідно генів, які кодують ферменти або фактори регуляції цих генів. У кращій клітині-хазяїні генетична модифікація включає гіперекспресію, щонайменше, одного ферменту (неокислювальної частини) пентозофосфатного шляху. Бажано фермент вибирається з групи, яка складається з таких ферментів, як рибулозо-5-фосфатізомераза, рибулозо-5-фосфатепімераза, транскетолаза і трансальдолаза. Можлива гіперекспресія різних комбінацій ферментів (неокислювальної частини) пентозофосфатного шляху. Наприклад, ферментами, які здатні до гіперекспресуватися, можуть бути, щонайменше, ферменти рибулозо-5-фосфатізомераза і рибулозо-5-фосфатепімераза; або, щонайменше, ферменти рибулозо-5-фосфатізомераза і транскетолаза; або, щонайменше, ферменти рибулозо-5фосфатізомераза і трансальдолаза; або, щонайменше, ферменти рибулозо-5фосфатепімераза і транскетолаза; або, щонайменше, ферменти рибулозо-5-фосфатепімераза і трансальдолаза; або, щонайменше, ферменти транскетолаза і трансальдолаза; або, щонайменше, фермент рибулозо-5-фосфатепімераза, транскетолаза і трансальдолаза; або, щонайменше, ферменти рибулозо-5-фосфатізомераза, транскетолаза і трансальдолаза; або, щонайменше, ферменти рибулозо-5-фосфатізомераза, рибулозо-5-фосфатепімераза і трансальдолаза; або, щонайменше, ферменти рибулозо-5-фосфатізомераза, рибулозо-5фосфатепімераза і транскетолаза. У одному варіанті здійснення винаходу кожен з ферментів – рибулозо-5-фосфатізомераза, рибулозо-5-фосфатепімераза, транскетолаза і трансальдолаза – гіперекспресується в клітині-хазяїні. Кращою є клітина-хазяїн, в якій генетична модифікація включає, щонайменше, гіперекспресію обох ферментів, – транскетолази і трансальдолази, оскільки така клітина-хазяїн вже здатна до анаеробного зростання на ксилозі. Фактично за деяких умов клітини-хазяїни, які гіперекспресують тільки транскетолазу і трансальдолазу, вже демонструють таку ж швидкість анаеробного росту на ксилозі, що і клітини-хазяїни, які гіперекспресують всі чотири ферменти, тобто рибулозо-5-фосфатізомеразу, рибулозо-5фосфатепімеразу, транскетолазу і трансальдолазу. Більше того, клітини-хазяїни, які гіперекспресують обидва ферменти – рибулозо-5-фосфатізомеразу і рибулозо-5фосфатепімеразу – є більш бажаними, ніж клітини-хазяїни, які гіперекспресують тільки ізомеразу або тільки епімеразу, оскільки гіперекспресія тільки одного з цих ферментів може призвести до метаболічного дисбалансу. Фермент «рибулозо-5-фосфатепімераза» (ЄС 5.1.3.1) в контексті опису визначається як фермент, який каталізує епімеризацію D-ксилулозо-5-фосфату в D-рибулозо-5-фосфат і навпаки. Фермент відомий також як фосфорибулозоепімераза; еритрозо-4-фосфатізомераза; фосфокетопентозо-3-епімераза; ксилулозофосфат-3-епімераза; фосфокетопентозоепімераза; рибулозо-5-фосфат-3-епімераза; D-рибулозофосфат-3-епімераза; D-рибулозо-5фосфатепімераза; D-рибулозо-5-фосфат-3-епімераза; D-ксилулозо-5-фосфат-3-епімераза; пентозо-5-фосфат-3-епімераза або D-рибулозо-5-фосфат-3-епімераза. Рибулозо-5фосфатепімераза може додатково визначатися її амінокислотною послідовністю. Так само, рибулозо-5-фосфатепімераза може визначатися послідовністю нуклеотидів, яка кодує фермент, а також послідовністю нуклеотидів, яка гібридизується з референс-послідовністю нуклеотидів, яка кодує рибулозо-5-фосфатепімеразу. Послідовність нуклеотидів, яка кодує рибулозо-5фосфатепімеразу, позначена в описі як RPE1. Фермент «рибулозо-5-фосфатізомераза» (ЄС 5.3.1.6) визначається в контексті опису як фермент, який каталізує пряму ізомеризацію D-рибозо-5-фосфату в D-рибулозо-5-фосфат і навпаки. Фермент відомий також як фосфопентозоізомераза; фосфорибоізомераза; рибозофосфатізомераза; 5-фосфорибозоізомераза; D-рибозо-5-фосфатізомераза; D-рибозо-5фосфат-кетол-ізомераза або D-рибозо-5-фосфат-альдозо-кетозоізомераза. Рибулозо-5фосфатізомераза може додатково визначатися своєю амінокислотною послідовністю. Так само, рибулозо-5-фосфатізомераза може визначатися послідовністю нуклеотидів, яка кодує фермент, а також послідовністю нуклеотидів, яка гібридизується з референсною послідовністю нуклеотидів, яка кодує рибулозо-5-фосфатізомеразу. Послідовність нуклеотидів, яка кодує рибулозо-5-фосфатізомеразу, позначена в описі як RKI1. Фермент «транскетолаза» (ЄС 2.2.1.1) визначається в контексті опису як фермент, який каталізує реакцію: D-рибозо-5-фосфат + D-ксилулозо-5-фосфат седогептулозо-7-фосфат + 13 UA 108853 C2 5 10 15 20 25 30 35 40 45 50 55 60 D-гліцеральдегід-3-фосфат і навпаки. Фермент відомий також як глікольальдегідтрансфераза або седогептулозо-7-фосфат:D-гліцеральдегід-3-фосфат-глікольальдегідтрансфераза. Транскетолаза може також визначатися своєю амінокислотною послідовністю. Так само, транскетолаза може визначатися послідовністю нуклеотидів, яка кодує фермент, а також послідовністю нуклеотидів, яка гібридизується з референсною послідовністю нуклеотидів, яка кодує транскетолазу. Послідовність нуклеотидів, яка кодує транскетолазу, позначена в описі як TKL1. Фермент «трансальдолаза» (ЄС 2.2.1.2) визначається в контексті опису як фермент, який каталізує реакцію: седогептулозо-7-фосфат + D-гліцеральдегід-3-фосфат D-еритрозо-4фосфат + D-фруктозо-6-фосфат і навпаки. Фермент відомий також як дигідроксиацетонтрансфераза; дигідроксиацетонсинтаза; формальдегідтранскетолаза або седогептулозо-7-фосфат:D-гліцеральдегід-3-фосфат- гліцеронтрансфераза. Трансальдолаза може також визначатися своєю амінокислотною послідовністю. Так само, трансальдолаза може визначатися послідовністю нуклеотидів, яка кодує фермент, а також послідовністю нуклеотидів, яка гібридизується з референсною послідовністю нуклеотидів, яка кодує трансальдолазу. Послідовність нуклеотидів, яка кодує трансальдолазу, позначена в описі як TAL1. Гени ксилоізомерази або ксилозоредуктази і ксилітолдегідрогенази Відповідно до винаходу одна, дві або більше копій одного або більше генів ксилоізомерази і/або одного або більше генів ксилозоредуктази і ксилітолдегідрогенази вводяться в геном клітини-хазяїна. Присутність цих двох або більше генетичних елементів надає клітині здатності до конверсії ксилози ізомеризацією або відновленням. У одному варіанті здійснення винаходу одна, дві або більше копії одного або більше генів ксилоізомерази вводяться в геном клітини-хазяїна. «Ксилозоізомераза» (ЄС 5.3.1.5) в контексті опису визначається як фермент, який каталізує пряму ізомеризацію D-ксилози в D-ксилулозу і/або навпаки. Фермент відомий також як Dксилозокетоізомераза. Ксилозоізомераза в контексті винаходу може бути також здатною каталізувати конверсію між D-глюкозою і D-фруктозою (і тому може бути віднесена відповідно до глюкозоізомерази). Ксилозоізомераза в контексті винаходу може вимагати присутності двовалентного катіона, такого як магній, марганець або кобальт, як кофактора. Таким чином, така клітина для ферментації змішаних цукрів здатна до ізомеризації ксилози в ксилулозу. Здатність до ізомеризації ксилози в ксилулозу надається клітині-хазяїну трансформацією клітини-хазяїна нуклеїнокислотним конструктом, який містить послідовність нуклеотидів, яка кодує певну ксилоізомеразу. Клітина для ферментації змішаних цукрів ізомеризує ксилозу в ксилулозу прямою ізомеризацією ксилози в ксилулозу. Одиницю (U) ксилоізомеразної активності в контексті опису можна визначити як кількість ферменту, яка продукує 1 нмоль ксилулози за хвилину за умов, описаних в Kuyper et al. (2003, FEMS Yeast Res. 4: 69-78). Ген ксилоізомерази може мати різне походження, наприклад, з Pyromyces sp., як описано в WO2006/009434. Інші придатними джерелами походження є Bacteroides, зокрема, Bacteroides uniformis, як описано в PCT/EP2009/52623; Bacillus, зокрема, Bacillus stearothermophilus, як описано в PCT/EP2009/052625. У іншому варіанті здійснення винаходу дві або більше копій одного або більше генів ксилозоредуктази і ксилітолдегідрогенази вводяться в геном клітини-хазяїна. У цьому варіанті конверсія ксилози здійснюється як двохстадійна конверсія ксилози в ксилулозу з утворенням ксилітолу як проміжного продукту, причому стадії конверсії каталізують відповідно ксилозоредуктазою і ксилітолдегідрогеназою. У одному з варіантів ксилозоредуктаза (XR), ксилітолдегідрогеназа (XDH) і ксилокіназа (XK) можуть гіперекспресуватися, і необов'язково має місце активація одного або більше генів, які кодують ферменти, які продукують NADPH (НАДФ = никотинамідаденіндинуклеотидфосфат), і одного або більше генів, які кодують ферменти, що споживають NADH (никотинамідаденіндинуклеотид, відновлена форма), як описано в WO 2004085627. Ген XKS1 Клітина винаходу може мати одну або більше генетичних модифікацій, які підвищують питому ксилулозокіназну активність. Бажано генетична модифікація або модифікації викликають гіперекспресію ксилулозокінази, наприклад, в результаті гіперекспресії послідовності нуклеотидів, яка кодує ксилулозокіназу. Ген, який кодує ксилулозокіназу, може бути ендогенним по відношенню до клітини-хазяїна або ксилулозокіназа може бути гетерологічною до клітинихазяїна. Послідовність нуклеотидів, яка використовується для гіперекспресії ксилулозокінази в клітині-хазяїні запропонованій винаходом, є послідовність нуклеотидів, яка кодує поліпептид з ксилулозокіназною активністю. Фермент «ксилулозокіназа» (ЄС 2.7.1.17) в контексті опису визначається як фермент, який 14 UA 108853 C2 5 10 15 20 25 30 35 40 45 50 55 60 каталізує реакцію: АТФ + D-ксилулоза = АДФ+D-ксилулозо-5-фосфат. Фермент відомий також як фосфорилююча ксилулокіназа, D-ксилулокіназа або АТФ:D-ксилулозо-5-фосфотрансфераза. Ксилулозокіназа винаходу може також визначатися своєю амінокислотною послідовністю. Так само, ксилулозокіназа може визначатися послідовністю нуклеотидів, яка кодує фермент, а також послідовністю нуклеотидів, яка гібридизується з референсною послідовністю нуклеотидів, яка кодує ксилулозокіназу. У клітині запропонованій винаходом генетична модифікація або модифікації, яка(і) підвищує(ють) питому ксилулозокіназну активність, може(можуть) комбінуватися з будь-якою з модифікацій, які збільшують потік через пентозофосфатний шлях, як описано вище. Проте це не істотно. Таким чином, клітина-хазяїн запропонована винаходом може містити тільки генетичну модифікацію або модифікації, яка(і) підвищує(ють) питому ксилулозокіназну активність. Відомі з рівня техніка різні методи для досягнення і аналізу гіперекспресії ксилулозокінази в клітинаххазяїнах відповідно до винаходу аналогічні до методів, описаних вище стосовно ферментів пентозофосфатного шляху. Бажано в клітинах-хазяїнах запропонованих винаходом, ксилулозокіназа, яка піддається гіперекспресії гіперекспресується щонайменше, приблизно в 1,1, приблизно в 1,2, приблизно в 1,5, приблизно в 2, приблизно в 5, приблизно в 10 або приблизно в 20 разів у порівнянні з штамом, який є генетично ідентичним, за винятком генетичної модифікації(й), що викликає(ють) гіперекспресію. Саме собою зрозуміло, що вказані рівні гіперекспресії можуть бути застосовними і до стійкого рівня ферментної активності, стійкого рівня ферментного білка, а також до стійкого рівня транскрипту, який кодує фермент. Делеція гена альдозоредуктази (GRE3) У варіанті здійснення винаходу, в якому XI використовується як ген конверсії ксилози, він здатний бажано знижувати активність альдозоредуктази. Тому клітина запропонована винаходом може містити одну або більше генетичних модифікацій, яка(і) знижує(ють) неспецифічну альдозоредуктазну активність в клітині-хазяїні. Бажано неспецифічна альдозоредуктазна активність в клітині-хазяїні знижується в результаті однієї або більше генетичних модифікацій, які знижують експресію або інактивують ген, який кодує неспецифічну альдозоредуктазу. Бажано генетична модифікація(ії) знижують або інактивують експресію кожної ендогенної копії гена, що кодує неспецифічну альдозоредуктазу в клітині-хазяїні (у описі це названо делецією GRE3). Клітини-хазяїни можуть містити множинні копії генів, що кодують неспецифічні альдозоредуктази як результат ди-, полі- або анеуплоїдії, і/або клітина-хазяїн може містити декілька різних (ізо)ферментів з альдозоредуктазною активністю, які розрізняються амінокислотною послідовністю і кожен з яких кодується різними генами. І в цих випадках бажано експресія кожного гена, який кодує неспецифічну альдозоредуктазу, знижується або інактивується. Бажано ген інактивується в результаті делеції, щонайменше, частини гена або в результаті руйнування гена, тому в даному контексті термін «ген» включає будь-яку не кодуючу послідовність справа або зліва від кодуючої послідовності, (часткова) делеція або інактивація якої призводить до зниження експресії неспецифічної альдозоредуктазної активності в клітині-хазяїні. Послідовність нуклеотидів, яка кодує альдозоредуктазу, активність якої може знижуватися в клітині-хазяїні запропонованій винаходом, є послідовністю нуклеотидів, яка кодує поліпептид з альдозоредуктазною активністю. Тому клітина-хазяїн запропонована винаходом, яка містить тільки генетичну модифікацію або модифікації, яка(і) знижує(ють) неспецифічну альдозоредуктазну активність в клітиніхазяїні, спеціально включена до складу винаходу. Фермент «альдозоредуктаза» (ЄС 1.1.1.21) в контексті опису визначається як фермент, який здатний відновлювати ксилозу або ксилулозу до ксилітолу (ксиліту). В контексті даного винаходу альдозоредуктаза може бути будь-якою неспецифічною альдозоредуктазою, яка є нативною (ендогенною) щодо клітини-хазяїна запропонованої винаходом і яка здатна відновлювати ксилозу або ксилулозу до ксилітолу (ксиліту). Неспецифічні альдозоредуктази + + каталізують реакцію: альдоза + NAD(P)H + H - альдитол (альдит) + NAD(P) (окислена форма НАДФ). Фермент проявляє широкий спектр специфічності і відомий також як альдозоредуктаза; + + поліолдегідрогеназа (NADP ); алдитол:NADP-оксидоредуктаза; алдитол:NADP -1оксидоредуктаза; NADPH-альдопентозоредуктаза або NADPH-альдозоредуктаза. Конкретний приклад такої неспецифічної альдозоредуктази, яка є ендогенною щодо S. cerevisiae і яка кодується геном GRE3, див. Traff et al., 2001, Appl. Environ. Microbiol. 67: 5668-74. Таким чином, альдозоредуктаза запропонована винаходом може також визначатися своєю амінокислотною послідовністю. Так само, альдозоредуктаза може визначатися послідовностями 15 UA 108853 C2 5 10 15 20 25 30 35 40 45 50 55 60 нуклеотидів, які кодують фермент, а також послідовністю нуклеотидів, яка гібридизується з референсною послідовністю нуклеотидів, яка кодує альдозоредуктазу. Ідентичність послідовностей Ідентичність послідовностей (або схожість послідовностей) в контексті опису визначається як «відповідність» між двома або більше амінокислотними (поліпептид або білок) послідовностями або між двома або більше нуклеїнокислотними (полінуклеотиди) послідовностями, яка визначається шляхом порівняння послідовностей. Зазвичай ідентичність або схожість послідовностей порівнюється в типових випадках за всією довжиною порівнюваних послідовностей. Проте послідовності можуть порівнюватися і за коротшими вікнами порівняння. У рівні техніки «ідентичність» означає також ступінь спорідненості між амінокислотними або нуклеїнокислотними послідовностями, яка визначається в деяких випадках за збігами між нитками таких послідовностей. Розроблені кращі методи визначення ідентичності з виявленням максимальної кількості збігів між аналізованими послідовностями. Методи визначення ідентичності і схожості кодифікують в вільно-доступних комп'ютерних програмах. Кращі методи визначення ідентичності і схожості між двома послідовностями з використанням комп'ютерних програм включають, наприклад, BestFit, BLASTP, BLASTN і FASTA (Altschul, S. F. et al., J. Mol. Biol. 215:403-410 (1990), вільно-доступні з NCBI та інших джерел (BLAST Manual, Altschul, S. et al., NCBI NLM NIH Bethesda, MD 20894). Кращими параметрами порівняння амінокислотних послідовностей за допомогою BLASTP є: штраф за відкриття пропуску 11,0, штраф за продовження пропуску 1, матриця Blosum 62. Кращими параметрами порівняння нуклеїнокислотних послідовностей за допомогою BLASTP є: штраф за відкриття пропуску 11,0, штраф за продовження пропуску 1, повнорозмірна ДНК-матриця (матриця ідентичності ДНК). Необов'язково при визначенні ступеня амінокислотної схожості кваліфікований фахівець може також брати до уваги так звані «консервативні» амінокислотні заміни, що стане зрозумілим в ході визначення. Консервативні амінокислотні заміни відносяться до взаємозамінюваності залишків, які мають схожі бічні ланцюги. Наприклад, група амінокислот, які мають аліфатичні бічні ланцюги, включає гліцин, аланін, валін, лейцин та ізолейцин; група амінокислот, які мають аліфатичнігідроксильні, бічні ланцюги, включає серин і треонін; група амінокислот, які мають амідовмісні бічні ланцюги, включає аспарагін і глутамін; група амінокислот, які мають ароматичні бічні ланцюги, включає фенілаланін, тирозин і триптофан; група амінокислот, які мають основні бічні ланцюги, включає лізин, аргінін і гістидін, і група амінокислот, які мають сірковмісні бічні ланцюги, включає цистеїн і метіонін. Кращими групами консервативних амінокислотних замін є: валін-лейцин-ізолейцин, фенілаланін-тирозин, лізин-аргінін, аланін-валін і аспарагін-глутамін. Варіантами замін в описаних тут амінокислотних послідовностях є такі варіанти, в яких, щонайменше, один залишок видалений з описаних послідовностей, а інший залишок, який відрізняється від видаленого, інсерцируваний на його місце. Бажано амінокислотна заміна є консервативною. Кращими консервативними замінами для кожної з амінокислот, які зустрічаються в природі, є наступні: Ala > ser; Arg > lys; Asn > gln або his; Asp > glu; Cys > ser або ala; GIn > asn; GIu > asp; GIy > pro; His > asn або gln; He > leu або val; Leu > ile або val; Lys > arg, gln або glu; Met > leu або ile; Phe > met, leu або tyr; Ser > thr; Thr > ser; Trp > tyr; Tyr > trp або phe і Val > ile або leu. Жорсткі умови гібридизації в контексті опису визначаються як умови, які роблять можливою гібридизацію нуклеїнокислотної послідовності, яка складається з, щонайменше, приблизно 25, краще – з приблизно 50, 75 або 100 нуклеотидів і найкраще – з приблизно 200 або більше нуклеотидів, за температури біля 65 °C в розчині, який містить приблизно 1 M солі, бажано 6 × SSC (хлорид натрію, цитрат натрію), або в будь-якому іншому розчині з співрозмірною іонною силою і відмивання при 65 °C в розчині, який містить приблизно 0,1 M солі або менше, бажано – 0,2 × SSC, або в будь-якому іншому розчині з співрозмірною іонною силою. Бажано гібридизація здійснюється протягом ночі, тобто протягом, щонайменше, 10 годин, а відмивання бажано проводиться протягом, щонайменше, однієї години, щонайменше, з двома замінами промивального розчину. Ці умови зазвичай є придатними для специфічної гібридизації послідовностей із ступенем ідентичності приблизно 90% або вище. Помірні умови в контексті опису визначаються як умови, які роблять можливою гібридизацію нуклеїнокислотних послідовностей, які складаються з, щонайменше, 50 нуклеотидів, краще – з приблизно 200 або більше нуклеотидів, за температури біля 45 °C в розчині, який містить приблизно 1 M солі, бажано 6 × SSC, або в будь-якому іншому розчині з співрозмірною іонною силою і відмивання за кімнатної температури в розчині, який містить приблизно 1 M солі, Бажано – 6 × SSC, або в будь-якому іншому розчині з співрозмірною іонною силою. Бажано 16 UA 108853 C2 5 10 15 20 25 30 35 40 45 50 55 60 гібридизація здійснюється протягом ночі, тобто протягом, щонайменше, 10 годин, а відмивання, бажано, проводиться протягом, щонайменше, однієї години, щонайменше, з двома замінами промивального розчину. Ці умови зазвичай є придатними для специфічної гібридизації послідовностей із ступенем ідентичності до 50%. Фахівець, кваліфікований в даній галузі техніки, може самостійно модифікувати вказані умови гібридизації для специфічної ідентифікації послідовностей із ступенем ідентичності від 50% до 90%. Щоб збільшити вірогідність того, що введений фермент експресується в активній формі в клітини запропоновані винаходом, можна адаптувати відповідну кодуючу послідовність нуклеотидів для оптимізації частоти використання її кодонів наблизивши її до частоти використання кодонів відібраної дріжджової клітини. У рівні техніки відомо декілька методів оптимізації кодонів. Кращим методом оптимізації частоти використання кодонів нуклеотидних послідовностей до частоти використання кодонів дріжджів є метод оптимізації пар кодонів, описаний в WO2006/077258 і/або WO2008/000632. WO2008/000632 відноситься до оптимізації пар кодонів. Оптимізація пар кодонів – це метод, в якому послідовності нуклеотидів, які кодують поліпептид, модифікуються за частотою використання кодонів, зокрема, за частотою використання пар кодонів, для досягнення покращеної експресії послідовності нуклеотидів, яка кодує поліпептид, і/або покращеного продукування кодованого поліпептиду. Пари кодонів визначаються як набір з двох послідовних триплетів (кодонів) в кодуючій послідовності. Індекс адаптації кодонів (CAI) використовується у винаході як проста міра експресії генів і ефективності трансляції, як описано в Xuhua Xia, Evolutionary Bioinformatics 2007: 3, 53-58. Індекс використовує стандартний набір високоекспресованих генів з одного виду для оцінки відносних показників кожного кодону, а оцінка у балах для гена розраховується, виходячи з частоти використання всіх кодонів в цьому гені. Індекс оцінює той ступінь, при якому відбір показав свою ефективність у формуванні патерну частоти використання кодонів. У зв'язку з цим, він може бути корисним для прогнозування рівня експресії гена, для оцінки адаптації вірусних генів до їх хазяїв і для проведення порівняльної оцінки частоти використання кодонів в різних організмах. Індекс може також дати наближену оцінку ймовірнісної ефективності гетерологічної експресії гена. У генах з оптимізованою парою кодонів згідно винаходу CAI складає 0,6 або вище, 0,7 або вище, 0,8 або вище, 0,85 або вище, 0,87 або вище, 0,90 або вище, 0,95 або вище або близько 1,0. Таким чином, клітина запропонована винаходом є клітиною, яка містить, тобто була трансформована ним, нуклеїнокислотний конструкт, який містить послідовність нуклеотидів, яка кодує гени araA, araB і araD, як вказано вище. Нуклеїнокислотний конструкт, який містить послідовність, яка кодує araA, бажано здатний експресувати гени araA в клітині-хазяїні. Бажано гени експресуються в цитозолі. Експресія в цитозолі може досягатися в результаті делеції або модифікації мітохондріального або пероксисомального цільового сигналу. Виробництво біопродуктів Протягом багатьох років виносилися на обговорення пропозиції про інтродукцію різних організмів для виробництва біоетанолу з цукрів сільськогосподарських культур. Проте на практиці у всіх основних способах виробництва етанолу як продуцент етанолу продовжували використовувати дріжджі роду Saccharomyces. Це пояснюється багатьма привабливими властивостями дріжджів виду Saccharomyces для промислових процесів, тобто їх високою стійкістю до кислот і етанолу, високою осмотолерантністю, здатністю до анаеробного зростання і, зрозуміло, їх високою здатністю до спиртового бродіння. Кращими видами дріжджів, які можуть бути клітинами-хазяїнами, є S. cerevisiae, S. bulderi, S. barnetti, S. exiguus, S. uvarum, S. diastaticus, K. lactis, K. marxianus або K fragilis. Клітина запропонована винаходом може бути здатною до конверсії рослинної біомаси, целюлози, геміцелюлоз, пектинів, рамнози, галактози, фруктози, мальтози, мальтодекстринів, рибози, рибулози або крохмалю, похідних крохмалю, сахарози, лактози і гліцерину, наприклад, у ферментовані цукри. Відповідно клітина запропонована винаходом може експресувати один або більше ферментів, таких як целюлаза (ендоцелюлаза або екзоцелюлаза); геміцелюлаза (ендо- або екзоксиланаза або арабіназа), необхідних для конверсії целюлози в мономери глюкози, а геміцелюлози – в мономери ксилози і арабінози; пектиназа, здатна перетворювати пектини на глюкуронову кислоту і галактуронову кислоту, або амілаза, яка перетворює крохмаль на мономери глюкози. Крім того, клітина бажано проявляє ферментативну активність, потрібну для конверсії пірувату в бажаний продукт ферментації, такий як етанол, бутанол, молочна кислота, 3гідроксипропіонова кислота, акрилова кислота, оцтова кислота, бурштинова кислота, лимонна кислота, фумарова кислота, яблучна кислота, ітаконова кислота, амінокислота, 1,3-пропандіол, етилен, гліцерин, β-лактамний антибіотик або цефалоспорин. 17 UA 108853 C2 5 10 15 20 25 30 35 40 45 50 55 60 Кращою клітиною запропонованою винаходом є клітина, здатна за своєю природою до спиртового бродіння, бажано – до анаеробного спиртового бродіння. Клітина запропонована винаходом бажано характеризується високою стійкістю до етанолу, високою стійкістю до низького pH (тобто вона здатна до зростання при pH нижче приблизно 5, приблизно 4, приблизно 3 або приблизно 2,5) і до органічних кислот, таких як молочна кислота, оцтова кислота або мурашина кислота, і/або до продуктів розпаду цукрів, таких як фурфурол і гідроксиметилфурфурол, і/або високою стійкістю до підвищених температур. Будь-яка з вищеперелічених характеристик або активностей клітини запропонованої винаходом може бути присутньою в клітині від природи або може бути введена в неї або модифікована шляхом генетичної модифікацієї. Клітина запропонована винаходом може бути клітиною, придатною для отримання етанолу. Проте клітина запропонована винаходом може бути придатна і для отримання інших, крім етанолу, продуктів ферментації. Такі «не етанольні» продукти ферментації включають, в принципі, будь-яку з низьким ступенем чистоти або високочисту хімічну речовину, яка продукується еукаріотичним мікроорганізмом, таким як дріжджі або міцеліальний гриб. Такими продуктами ферментації можуть бути, наприклад, бутанол, молочна кислота, 3гідроксипропіонова кислота, акрилова кислота, оцтова кислота, бурштинова кислота, лимонна кислота, яблучна кислота, фумарова кислота, ітаконова кислота, амінокислота, 1,3-пропандіол, етилен, гліцерин, β-лактамний антибіотик або цефалоспорин. Краща клітина запропонована винаходом для отримання «не етанольних» продуктів ферментації є клітиною-хазяїном, яка містить генетичну модифікацію, що призводить до зниження алкогольдегідрогеназної активності. У наступному аспекті винахід відноситься до способів ферментації, в яких клітини запропоновані винаходом використовуються для ферментації джерела вуглецю, включаючи джерело ксилози, таке як ксилоза. На додачу до джерела ксилози джерело вуглецю в середовищі ферментації може також включати джерело глюкози. Джерело ксилози або глюкози може бути чистою ксилозою або чистою глюкозою або може бути будь-яким вуглеводневим оліго- або полімером, який містить одиниці ксилози або глюкози, таким як, наприклад, лігноцелюлоза, ксилани, целюлоза, крохмаль тощо. Для вивільнення одиниць ксилози або глюкози з таких вуглеводів відповідна карбогідраза (такі як ксиланази, глюканази, амілази та ін.) може додаватися до середовища ферментації або можуть продукуватися самою клітиною. У останньому випадку клітина може сконструйована шляхом генної інженерії з наданням їй здатності до продукування і виділення такої карбогідрази. Додаткова перевага від використання оліго- або полімерних джерел глюкози полягає в тому, що це робить можливою підтримку низької(зниженої) концентрації вільної глюкози в процесі ферментації, наприклад, за рахунок використання карбогідрази в кількостях, які обмежують швидкість. А це, у свою чергу, перешкоджатиме репресії систем, необхідних для метаболізму і транспорту інших цукрів (не глюкози), таких як ксилоза. У кращому способі клітина ферментує і ксилозу, і глюкозу, бажано одночасно; в цьому випадку бажано використовується клітина, яка є нечутливою до репресії глюкози, щоб уникнути диауксичного росту. На додачу до джерела ксилози (і глюкози) як джерело вуглецю середовище ферментації може містити також відповідний інгредієнт, потрібний для росту клітини. Склад середовищ ферментацій для зростання мікроорганізмів, таких як дріжджі, добре відомий в рівні техніки. Спосіб ферментації – це спосіб отримання продукту ферментації, такого як, наприклад, етанол, бутанол, молочна кислота, 3-гідроксипропіонова кислота, акрилова кислота, оцтова кислота, бурштинова кислота, лимонна кислота, яблучна кислота, фумарова кислота, ітаконова кислота, амінокислота, 1,3-пропандіол, етилен, гліцерин, β-лактамний антибіотик, такий як Пеніцилін G або Пеніцилін V і їх ферментативні похідні, і цефалоспорин. Лігноцелюлоза Лігноцелюлоза, яку можна розглядати як потенційно поновлювану промислову сировину, в більшості випадків містить полісахариди – целюлозу (глюкани) і геміцелюлози (ксилани, гетероксилани і ксилоглюкани). Додатково деякі геміцелюлози можуть бути присутніми у формі глюкомананів, наприклад, в деревній сировині. Ферментативний гідроліз цих полісахаридів до розчинних цукрів, включаючи як мономери, так і мультимери, наприклад, глюкозу, целобіозу, ксилозу, арабінозу, галактозу, фруктозу, манозу, рамнозу, рибозу, галактуронову кислоту, глюкуронову кислоту та інші гексози і пентози, відбувається під дією різних ферментів, які діють в сукупності. До того ж, пектини і інші пектинові речовини, такі як арабінани, можуть складати значну частку сухої маси стінок типових клітин з недеревних рослинних тканин (пектини можуть складати приблизно від ¼ до половини сухої маси). 18 UA 108853 C2 5 10 15 20 25 30 35 40 45 50 55 60 Передобробка Передобробка може бути бажаною за необхідності виділення цукрів, здатних ферментуватися згідно винаходу, з лігноцелюлозного (включаючи геміцелюлозний) матеріалу. Ця стадія може здійснюватися традиційними способами, таким як, наприклад Ферментативний гідроліз Ферментативний гідроліз може проводитися традиційними способами. Ферментація Спосіб ферментації може бути аеробним або анаеробним способом ферментації. Анаеробний спосіб ферментації в контексті опису визначається як спосіб ферментації, який протікає за відсутності кисню, або як спосіб, в якому кисень в основному не споживається або споживається в кількості бажано менше приблизно 5, приблизно 2,5 або приблизно 1 ммол/л/год., краще – 0 ммол/л/год. (тобто споживання кисню неможливо виявити), і в якому органічні молекули служать і донорами, і акцепторами електронів. За відсутності кисню NADH (відновлена форма никотинамідаденіндинуклеотиду), який утворюється в процесі гліколізу і утворення біомаси, не здатний окислюватися окислювальним фосфорилюванням. Щоб вирішити цю проблему, багато мікроорганізмів використовують піруват або одну з його похідних як акцептор електронів і водню, регенеруючи, тим самим, NAD+.(окислена форма никотинамідаденіндинуклеотиду). Таким чином, в кращому анаеробному способі ферментації піруват використовується як акцептор електронів (і водню) і відновлюється до продуктів ферментації, таких як етанол, бутанол, молочна кислота, 3-гідроксипропіонова кислота, акрилова кислота, оцтова кислота, бурштинова кислота, лимонна кислота, яблучна кислота, фумарова кислота, амінокислота, 1,3пропандіол, етилен, гліцерин, β-лактамний антибіотик або цефалоспорин. Спосіб ферментації бажано протікає за температури, яка є оптимальною для клітини. Отже, при використанні клітин-хазяїнів більшості дріжджів або міцеліальних грибів спосіб ферментації проводиться за температури нижче приблизно 42 °C, бажано – нижче приблизно 38 °C. У випадку клітин-хазяїнів дріжджів або міцеліального гриба спосіб ферментації проводиться за температури нижче приблизно 35 °C, приблизно 33 °C, приблизно 30 °C або приблизно 28 °C і за температури вищої від приблизно 20 °C, приблизно 22 °C або приблизно 25 °C. Вихід етанолу на ксилозі і/або глюкозі в способі бажано складає, щонайменше, близько 50%, близько 60%, близько 70%, близько 80%, близько 90%, близько 95% або близько 98%. У контексті опису вихід етанолу визначається як відсоток від теоретичного максимального виходу. Винахід відноситься також до способу отримання продукту ферментації. Способи ферментації можуть проводитися в періодичному, періодичному з підживленням або безперервному режимі. Він може також здійснюватися в режимі роздільного проведення процесів гідролізу і ферментації (SHF-спосіб) або в режимі одночасного проведення процесів зцукрення (цукроутворення) і ферментації (SSF-спосіб). Можлива також комбінація вказаних режимів способу ферментації для досягнення оптимальної продуктивності. Спосіб ферментації за даним винаходом може протікати в аеробних і анаеробних умовах. Бажано спосіб проводиться за мікроаерофільних умов або за умов обмеженої кількості кисню. Анаеробний спосіб ферментації в контексті опису визначається як спосіб ферментації, який протікає за відсутності кисню, або як спосіб, в якому кисень в основному не споживається або спожита кількість кисню бажано складає менше приблизно 5, приблизно 2,5 або приблизно 1 ммол/л/год. і в якому органічні молекули служать і донорами, і акцепторами електронів. Спосіб ферментації в умовах обмеженої кількості кисню – це спосіб, в якому споживання кисню обмежене перенесенням кисню з газу в рідину. Ступінь обмеження кисню визначається кількістю і складом вхідного газового потоку, а також фактичними показниками змішування/масопереносу у використовуваному устаткуванні ферментації. Бажано в способі за умов обмеженої кількості кисню швидкість споживання кисню становить, щонайменше, близько 5,5, краще, щонайменше, близько 6, наприклад, щонайменше 7 ммол/м/год. Спосіб запропонований винаходом передбачає вилучення продукту ферментації. У кращому варіанті способу клітина ферментує як ксилозу, так і глюкозу, бажано одночасно, тому в цьому випадку бажано використовується клітина, яка є нечутливою до репресії глюкози, щоб уникнути диауксичного зростання. На додачу до джерела ксилози (і глюкози) як джерело вуглецю середовище ферментації містить також відповідний інгредієнт, необхідний для зростання клітини. Склади середовищ ферментацій для зростання мікроорганізмів, таких як дріжджі, добре відомі з рівня техніки. Спосіб ферментації може здійснюватися в періодичному, періодичному з підживленням або безперервному режимі. Він може також здійснюватися в режимі роздільного проведення процесів гідролізу і ферментації (SHF-спосіб) або в режимі одночасного проведення процесів 19 UA 108853 C2 5 10 15 20 25 30 35 40 45 50 55 60 зцукрення (цукроутворення) і ферментації (SSF-спосіб). Можлива також комбінація вказаних режимів способу ферментації для досягнення оптимальної продуктивності. Детальніше вищезгадані способи описані нижче. SSF-режим У випадку режиму з одночасним проведенням процесів цукроутворення і ферментації (SSF) час реакції на стадії розрідження/гідролізу або на стадії попереднього цукроутворення залежить від часу реалізації бажаного виходу, тобто виходу від конверсії целюлози в глюкозу. Цей вихід бажано є високим, наскільки це можливо, і бажано складає 60% або вище, 65% або вище, 70% або вище, 75% або вище, 80% або вище, 85% або вище, 90% або вище, 95% або вище, 96% або вище, 97% або вище, 98% або вище, 99% або вище, навіть 99,5% або вище або 99,9% або вище. Згідно винаходу досягаються дуже високі концентрації цукрів в SHF-режимі і дуже високі концентрації продукту (наприклад, етанолу) в SSF-режимі. У SHF-режимі концентрація глюкози складає 25 г/л або більше, 30 г/л або більше, 35 г/л або більше, 40 г/л або більше, 45 г/л або більше, 50 г/л або більше, 55 г/л або більше, 60 г/л або більше, 65 г/л або більше, 70 г/л або більше, 75 г/л або більше, 80 г/л або більше, 85 г/л або більше, 90 г/л або більше, 95 г/л або більше, 100 г/л або більше, 110 г/л або більше, 120 г/л або більше, або може складати, наприклад, від 25 г/л до 250 г/л, від 30 г/л до 200 г/л, від 40 г/л до 200 г/л, від 50 г/л до 200 г/л, від 60 г/л до 200 г/л, від 70 г/л до 200 г/л, від 80 г/л до 200 г/л, 90 г/л, від 80 г/л до 200 г/л. Концентрація продукту в SSF-режимі У SSF-режимі концентрація продукту (г/л) залежить від кількості глюкози, що утворилася, але це практично не можна побачити, оскільки цукри конвертуються в продукт в SSF-режимі, і концентрації продукту можна пов'язати з відповідною концентрацією глюкози, лише помноживши на теоретичний максимальний вихід (Ypsmax в грамах продукту на грам глюкози). Теоретичний максимальний вихід (Ypsmax в грамах продукту на грам глюкози) продукту ферментації можна знайти в підручнику з біохімії. У випадку етанолу 1 моль глюкози (180 г) за умов нормального протікання ферментативного гліколізу в дріжджах дає 2 моля етанолу (=2 × 46 = 92 г етанолу). Тому теоретичний максимальний вихід етанолу на глюкозі складає 92/180 = 0,511 г етанолу/г глюкози. У випадку бутанолу (ММ 74 г/моль) або ізобутанолу теоретичний максимальний вихід складає 1 моль бутанолу намоль глюкози. Тому Ypsmax для (з) бутанолу = 74/180 = 0,411 г (з) бутанолу/г глюкози. Для молочної кислоти вихід при гомоферментативному молочнокислому бродінні складає 2 моля молочної кислоти (ММ = 90 г/моль) на моль глюкози. Відповідно до цієї стехіометрії, Yps max = 1 г молочної кислоти/г глюкози. Аналогічні розрахунки можуть бути проведені і для інших продуктів ферментації. SSF-режим У SSF-режимі концентрація продукту складає 25 * Yps г/л або більше, 30 * Yps г/л або більше, 35g * Yps г/л або більше, 40 * Yps г/л або більше, 45 * Yps г/л або більше, 50 * Yps г/л або більше, 55 * Yps г/л або більше, 60 * Yps г/л або більше, 65 * Yps г/л або більше, 70 * Yps г/л або більше, 75 * Yps г/л або більше, 80 * Yps г/л або більше, 85 * Yps г/л або більше, 90 * Yps г/л або більше, 95 * Yps г/л або більше, 100 * Yps г/л або більше, 110 * Yps г/л або більше, 120 г/л * Yps або більше або може складати, наприклад, від 25 * Yps г/л до 250 * Yps г/л, від 30 * Yps г/л до 200 * Yps г/л, від 40 * Yps г/л до 200 * Yps г/л, від 50 * Yps г/л до 200 * Yps г/л, від 60 * Yps г/л до 200 * Yps г/л, від 70 * Yps г/л до 200 * Yps г/л, від 80 * Yps г/л до 200 * Yps г/л, 90 * Yps г/л, від 80 * Yps г/л до 200 * Yps г/л. Таким чином, винахід пропонує спосіб отримання продукту ферментації, який передбачає: a. розщеплювання лігноцелюлози способом, описаним вище, і b. ферментацію отриманого матеріалу з отриманням продукту ферментації. Продукт ферментації Продукт ферментації за даним винаходом може бути будь-яким корисним продуктом. У одному варіанті здійснення винаходу продукт ферментації є продуктом, вибраним з групи, яка складається з етанолу, n-бутанолу, ізобутанолу, молочної кислоти, 3-гідроксипропіонової кислоти, акрилової кислоти, оцтової кислоти, бурштинової кислоти, фумарової кислоти, яблучної кислоти, ітаконової кислоти, малеїнової кислоти, лимонної кислоти, адипінової кислоти, амінокислоти (такої як лізин, метіонін, триптофан, треонін і аспарагінова кислота), 1,3пропандіолу, етилену, гліцерину, β-лактамного антибіотика і цефалоспорину, вітамінів, фармацевтичних засобів, добавок до корму для тварин, спеціальних хімічних засобів, хімічної промислової сировини, пластмас, розчинників, палива, включаючи біопаливо і біогаз, або органічних полімерів і промислового ферменту, такого як протеаза, целюлаза, амілаза, 20 UA 108853 C2 5 10 15 20 25 30 35 40 45 50 55 60 глюканаза, лактаза, ліпаза, ліаза, оксидоредуктаза, трансфераза або ксиланаза. Вилучення продукту ферментації Для вилучення продукту ферментації застосовуються існуючі технології. Для різних продуктів ферментації придатними є різні процеси вилучення. Існуючі технології вилучення етанолу з водних сумішей традиційно використовують методи фракціонування і адсорбції. Наприклад, для обробки ферментованого продукту, який містить етанол у водній суміші, можна використовувати брагоперегонний апарат для отримання багатої на етанол суміші, яку можна піддати подальшому фракціонуванню (наприклад, фракційній дистиляції або іншим аналогічним методам). Потім фракції, які містять максимальні концентрації етанолу, можна пропустити через адсорбер для видалення більшої частини (якщо не всієї) залишкової води з етанолу. Наступні приклади ілюструють винахід. Приклади Якщо не вказано що-небудь інше, то використані методи є стандартними біохімічними методами. Приклади відповідних довідників за загальною методологією включають Sambrook et al., Molecular Cloning, а Laboratory Manual (1989) і Ausubel et al., Current Protocols in Molecular Biology (1995), John Wiley & Sons, Inc. Трансформація S. cerevisiae Трансформацію S. cerevisiae проводили, як описано Gietz and Woods (2002; Transformation of the yeast by the LiAc/SS carrier DNA/PEG method. Methods in Enzymology, 350: 87-96). Колонії Ізолят одиничної колонії відбирали пластиковою зубочисткою і ресуспендували в 50 мкл milliQ-води. Зразок інкубували протягом 10 хвилин при 99 °C. 5 мкл інкубованого зразка ® використовували як матрицю для ПЛР реакції із застосуванням ДНК-полімерази Phusion (Finnzymes) відповідно до інструкції виробника. Умови ПЛР реакції: стадія 1 3 хв. 98 °C стадія 2 10 сек. 98 °C стадія 3 15 сек. 58 °C повторення стадій 2 - 4 протягом 30 циклів стадія 4 30 сек. 72 °C стадія 5 4 хв. 72 °C стадія 6 30 сек. 20 °C Склад поживного середовища Експерименти з вирощування: штами Saccharomyces cerevisiae вирощували на середовищі, яке має наступний склад: 0,67% (мас./об.) дріжджової азотистої основи або синтетичного середовища (Verduyn et al., Yeast 8:501-517, 1992) і або глюкоза, арабіноза, галактоза, або ксилоза або комбінація цих субстратів (див. нижче). Для чашок з агаром до середовища додавали 2% (мас./об.) бактеріологічного агару. Отримання етанолу: культивування проводили при 30 °C в 100 мл синтетичного модельного середовища (Verduyn-середовище [Verduyn et al., Yeast 8:501-517, 1992] з 5 % глюкози, 5 % ксилози, 3,5 % арабінози і 1%-1,5% галактози) в BAM (монітор біологічної активності, Halotec, The Netherlands). Перед стерилізацією pH середовища встановлювали на рівні pH 4,2 за допомогою 2 M NaOH/H2SO4. До синтетичного середовища для анаеробного культивування додавали 0,01 г/л ергостабілу (ергостерину) і 0,42 г/л Твін 80, розчиненого в етанолі (Andreasen and Stier. J. Cell Physiol. 41:23-36, 1953; and Andreasen and Stier. J. Cell Physiol. 43:271-281, 1954). Культури перемішували магнітною мішалкою. Анаеробні умови в ході ферментації досягалися швидко, оскільки культура не була аерована. Проводили безперервний моніторинг утворення CO2. Конверсію цукрів і утворення продуктів ферментації аналізували ЯМР-методом. Моніторинг зростання вели за необхідною оптичною щільністю культури при 600 нм на спектрофотометрі LKB Ultraspec K. Попередні культури отримували інокуляцією 25 мл Verduyn-середовища (Verduyn et al., Yeast 8:501-517, 1992) з додаванням 2 % глюкози замороженою початковою (вихідною) культурою або одиничною колонією з чашки з агаром в періодично струшуваній колбі на 100 мл. Після інкубації при 30 °C в орбітальному шейкері (200 об./хв.) протягом приблизно 24 годин проводили збір клітин з культури і використовували їх для інокуляції BAM при OD 600, рівною приблизно 2. Приклад 1 Введення генів araA, araB і araD в геном S. cerevisiae. 1.1 Конструювання вектора експресії, який містить гени для арабінозного шляху Плазміду pPWT018, представлену на Фіг. 2, конструювали таким чином: вектор pPWT006 (Фіг. 1), який складається з SIT2-локуса (Gottlin-Ninfa and Kaback (1986) Molecular and Cell 21 UA 108853 C2 5 10 15 20 25 30 35 40 45 50 55 60 Biology, vol. 6, no. 6, 2185-2197) і маркерів, що дозволяють проводити відбір трансформантів за стійкістю до антибіотика G418 і за здатністю до зростання на ацетаміді (див. вище), розщеплювали ферментами рестрикції BsiWI і MluI. kanMX-маркер, який надає стійкості до G418, був виділений з p427TEF (Dualsystems Biotech), а фрагмент, який містить amdS-маркер, описаний раніше в літературі (Swinkels, B.W., Noordermeer, A.C.M. and Renniers, A.C.H.M (1995) The use of the amdS cDNA of Aspergillus nidulans as а dominant, bidirectional selectable marker for yeast transformation. Yeast Volume 11, Issue 1995A, page S579; і US 6051431). Гени, які кодують арабінозоізомеразу (araA), L-рибулокіназу (araB) і L-рибулозо-5-фосфат-4-епімеразу (araD) з Lactobacillus plantarum, як описано в патентній заявці WO2008/041840, синтезували за допомогою BaseClear (Leiden, The Netherlands). Один великий фрагмент, який несе три вищезазначені ara-гени, був синтезований і знаходився під контролем сильних промоторів з S. cerevisiae (або був функціонально пов'язаний з промоторами), тобто TDH3-промотора, який контролює експресію araA-гена; ENO1-промотора, який контролює araB-ген, і PGI1-промотора, який контролює araD-ген. Цей фрагмент був оточений унікальними ферментами рестрикції Acc65I і MluI. Клонування цього фрагмента у векторі pPWT006, розщепленому ферментами рестрикції MluI і BsiWI, дозволило отримати плазміду pPWT018 (Фіг. 2). Послідовність плазміди pPWT018 представлена в SEQ ID 17. 1.2 Трансформація дріжджів CEN.PK113-7D (MATa URA3 HIS3 LEU2 TRP1 MAL2-8 SUC2) був трансформований плазмідою pPWT018, яка заздалегідь була лінеаризована з використанням SfiI (New England Biolabs) відповідно до інструкції виробника. Синтетичний SfiI-сайт був сконструйований в 5’фланку SIT2-гена (див. Фіг. 2). Трансформаційні суміші висівали на чашки з YPD-агаром (на літр: 10 грамів дріжджового екстракту, 20 грамів/літр пептону, 20 грамів/літр декстрози, 20 грамів агару), який містить 100 мкг G418 (Sigma Aldrich) /мл. Через дві – чотири добу на чашках з'явилися колонії, тоді як негативний контроль (тобто без додавання ДНК в експерименті із трансформації) показав відсутність зростання колоній на YPD/G418-чашках. Інтеграція плазміди pPWT018 націлена на SIT2-локус. Для характеристики трансформантів використовували ПЛРаналіз і метод Саузерн-блотінга. Реакції ПЛР, які служать ознакою коректної інтеграції однієї копії плазміди pPWT018, виконували з праймерами, представленими SEQ ID 18 і 15, 15 і 14 (див. Фіг. 4). Коректність інтеграції в SIT2-локусі контролювалася парою праймерів SEQ ID 18 і 15. Якщо плазміда pPWT018 інтегрується в множинні копії (інтеграція за типом голова-до-хвоста), то пара праймерів SEQ ID 15 і 14 дає один ПЛР- продукт. Відсутність такого ПЛР-продукту указує на інтеграцію однокопійної плазміди pPWT018. Штам, в якому одна копія плазміди pPWT018 була інтегрована в SIT2-локус, був позначений як BIE104R2. 1.3 Збереження маркера Щоб зробити можливою трансформацію дріжджового штаму іншими конструктами, які використовують ті ж самі селективні маркери, необхідно видалити селектовані маркери. Конструкція плазміди pPWT018 така, що при інтеграції pPWT018 в хромосому гомологічні послідовності знаходяться в безпосередній близькості одна з одною. Така конструкція забезпечує втрату селектованих маркерів в результаті спонтанної внутрішньомолекулярної рекомбінації цих гомологічних областей. При вегетативному зростанні внутрішньомолекулярнарекомбінація має місце, хоча і з низькою частотою. Частота цієї рекомбінації залежить від довжини гомології і локусу в геномі (неопубліковані результати). При послідовному перенесенні субфракції культури на свіже середовище внутріклітинні рекомбінанти з часом акумулюються. З цією метою штам BIE104R2 культивували в YPD-середовищі (на літр: 10 грамів дріжджового екстракту, 20 грамів/літр пептону, 20 грамів/літр декстрози), починаючи з ізоляту одиничної колонії. Для інокуляції свіжій YPD-середовища використовували 25 мкл культури, отриманої культивуванням штаму протягом ночі. Після, щонайменше, п'яти таких послідовних перенесень визначали оптичну щільність культури і клітини розводили до концентрації приблизно 5000 в 1 мл. 100 мкл клітинної суспензії висівали на чашки з середовищем Yeast Carbon Base (Difco), яке містить 30 мМ KPi (pH 6,8), 0,1% (NH4)2SO4, 40 мМ фторацетаміду (Amersham) і 1,8% агару (Difco). Клітини, ідентичні до клітин штаму BIE104R2, тобто без внутріклітинної рекомбінації, все ще містять amdS-ген. Для цих клітин фторацетамід токсичний. Ці клітини не здатні рости і не утворюють колоній на середовищі, яке містить фторацетамід. Проте, якщо внутрішньомолекулярна рекомбінація мала місце, то BIE104R2-варіанти, які втратили селектовані маркери, будуть здатні рости на середовищі, яке містить фторацетамід, оскільки вони не здатні до конверсії фторацетаміду в сполуки, які інгібують ріст. Ці клітини і формуватимуть колонії на вказаному вище агаровому середовищі. 22 UA 108853 C2 5 10 15 20 25 30 35 40 45 50 55 60 Отримані таким шляхом стійкі до фторацетаміду колонії піддавали ПЛР-аналізу з використанням праймерів SEQ ID 18 і 15, 14 і 19. Праймери SEQ ID 18 і 5 дають смугу в тому випадку, якщо рекомбінація селектованих маркерів відбулася, як і очікувалося. В результаті цього касета з генами araA, araB і araD під контролем сильних промоторів дріжджів інтегрувалася в SIT2-локус генома штаму-хазяїна. В цьому випадку ПЛР-реакція з використанням праймерів SEQ ID 14 і 19 не повинна призводити до утворення ПЛР-продукту, оскільки праймер 14 здатний праймувати в області, яка повинна бути втрачена в результаті рекомбінації. Якщо ж вказані праймери дають смугу, то це служить ознакою присутності повної плазміди pPWT018 в геномі, отже, рекомбінація не мала місця. Якщо праймери SEQ ID 18 і 15 не приводять до отримання ПЛР-продукту, то це означає, що рекомбінація відбулася, але таким шляхом, що повна плазміда pPWT018 рекомбінувалася поза геномом. Втрачені були не тільки селектовані маркери, але і гени арабінози. Фактично дріжджі переродилися в дріжджі дикого типу. Ізоляти, які показали результати ПЛР відповідно до інтеграції однієї копії pPWT018, піддавали аналізу Саузерн-блотінгом. Хромосомну ДНК штамів CEN.PK113-7D і коректні рекомбінанти розщеплювали за допомогою EcoRI і HindIII (подвійне розщеплення). Був сконструйований SIT2-зонд з праймерами SEQ ID 20 і 21 з використанням хромосомної ДНК штамів CEN.PK113-7D як матриці. Результат експерименту з гібридизації показаний на Фіг. 3. Очікуваний патерн гібридизації може бути виділеним з фізичних карт, представлених на Фіг. 4 (панелі а і b). У штамі дикого типу спостерігається смуга 2,35 т.п.н., яка відповідає передбачуваному розміру гена дикого типу (Фіг. 4, панель а). Після інтеграції і часткової втрати рекомбінацією плазміди pPWT018 очікувалася смуга в 1,06 т.п.н. (Фіг. 4, панель b). І ця смуга дійсно спостерігається, як показує Фіг. 3 (доріжка 2). Одним з штамів, які показали коректний патерн смуг на Саузерн-блоті (як можна бачити з Фіг. 3), є штам, позначений як BIE104A2. 1.4 Введення чотирьох конститутивно експресованих генів неокислювального пентозофосфатного шляху Saccharomyces cerevisiae BIE104A2, який конститутивно експресує гени araA, araB і araD, був трансформований плазмідою pPWT080 (Фіг. 5). Послідовність плазміди pPWT080 представлена в SEQ ID NO: 4. Процедура трансформації і селекції після відбору однокопійного трансформанта аналогічна до описаної вище в розділах 1.1, 1.2 і 1.3. Якщо говорити коротко, то BIE104A2 був трансформований SfiI-розщепленою плазмідою pPWT080. Трансформаційні суміші висівали на чашки з YPD-агаром (з розрахунку на літр: 10 грамів дріжджового екстракту, 20 грамів/літр пептону, 20 грамів/літр декстрози, 20 грамів агару), що містить 100 мкг G418 (Sigma Aldrich)/мл. Через дві - чотири доби на чашках з'явилися колонії, тоді як негативний контроль (тобто без додавання ДНК в експерименті по трансформації) не показав зростання на чашках з YPD/G418. Інтеграція плазміди pPWT080 націлена на GRE3-локус. Характеристики трансформантів визначали за допомогою ПЛР і Саузерн-блотінга. Трансформант, який показав коректну інтеграцію однієї копії плазміди pPWT080 відповідно до очікуваногопатерну гібридизації, був позначений як BIE104A2F1. Щоб зробити можливим введення генів, які кодують ксилоізомеразу і ксилулокіназу (Приклад 5), необхідно видалити селективні маркери, введені інтеграцією плазміди pPWT080. З цією метою штам BIE104A2F1 культивували в YPD-середовищі, починаючи з ізоляту колонії. 25 мкл культури, отриманої культивуванням протягом ночі, використовували для інокуляції свіжого YPD-середовища. Після п'яти послідовних перенесень визначали оптичну щільність культури, і клітини розбавляли до концентрації приблизно 5000 в 1 мл. 100 мкл клітинної суспензії висівали на чашки з середовищем Yeast Carbon Base (Difco), яке містить 30 мМ KPi (pH 6,8), 0,1% (NH4)2SO4, 40 мМ фторацетаміду (Amersham) і 1,8% агару (Difco). Стійкі до фторацетаміду колонії піддавали ПЛР-аналізу, і у випадку коректних ПЛР-профілів – Саузерн-блотінгу (розділ 1.3 Прикладу 1). Одним з штамів, які показали коректний патерн смуг на Саузерн-блоті, є штам, позначений як BIE104A2P1. Приклад 2 Адаптивна еволюція 2.1 Адаптивна еволюція (аеробні умови) Ізолят одиничної колонії штаму BIE104A2P1 використовували для інокуляції YNBсередовища (Difco), до якого додавали 2% галактози. Попередню культуру інкубували протягом приблизно 24 годин при 30 °C і 280 об./хв. Далі проводили збір клітин в культурі і інокуляцію YNB-середовища, яке містить 1% галактози і 1% арабінози при початковій OD 600, рівній 0,2 23 UA 108853 C2 5 10 15 20 25 30 35 40 45 50 (Фіг. 8). Клітини росли при 30 °C і 280 об./хв. Проводили регулярний моніторинг оптичної щільності при 600 нм. Коли оптична щільність досягла значення 5, аліквоту культури переносили на свіже YNBсередовище такого ж складу, який був вказаний вище. Кількість доданих клітин була такою, щоб початкова OD 600 культур складала 0,2. Після повторного досягнення OD 600, рівною 5, аліквоту культури переносили на YNB-середовище, яке містить 2% арабінози як єдине джерело вуглецю (подія, позначена як (1) на Фіг. 8). Після перенесення на YNB-середовище з 2% арабінози як єдиного джерела вуглецю зростання можна було спостерігати через приблизно два тижні. Коли оптична щільність при 600 нм досягала значення, щонайменше, 1, клітини переносили в періодично струшувану колбу зі свіжим YNB-середовищем, в яке додавали 2% арабінози, з початковою OD 600, рівною 0,2 (фіг. 8). Послідовне перенесення повторювали тричі, як показано на фіг. 8. Отриманий штам, який показав здатність до швидкого зростання на арабінозі, був позначений як BIE104A2P1c. 2.2 Адаптивна еволюція (анаеробні умови) Після адаптації до зростання на арабінозі за аеробних умов одиничну колонію штаму BIE104A2P1c використовували для інокуляції YNB-середовища, до якого додавали 2% глюкози. Попередню культуру інкубували протягом приблизно 24 годин при 30 °C і 280 об./хв. Далі проводили збір клітин в культурі і інокуляцію YNB-середовища, яке містить 2% арабінози, з оптичною щільністю OD 600, рівною 0,2. Колби закривали водним затвором з метою гарантованого підтримування анаеробних умов зростання після видалення кисню з середовища і вільного простору над середовищем в колбах. Після досягнення OD600 значення, рівного мінімум 3, аліквоту культури переносили на свіже YNB-середовище, яке містить 2% арабінози (Фіг. 9); кожна культура мала оптичну щільність OD 600, рівну 0,2. Отриманий після декількох пересадок штам був позначений як BIE104A2P1d (=BIE201). Приклад 3 Визначення ферментативної здатності Ізоляти одиничних колоній штамів BIE104, BIE104A2P1c і BIE201 використовували для інокуляції YNB-середовища (Difco), в яке додавали 2% глюкози. Попередні культури інкубували протягом приблизно 24 годин при 30 °C і 280 об./хв. Далі проводили збір клітин в культурі і інокуляцію синтетичного модельного середовища (Verduyn et al., Yeast 8:501-517, 1992; 5% глюкози, 5% ксилози, 3,5% арабінози, 1% галактози) з початковою OD 600, рівною приблизно 2, в BAM. Проводили постійний моніторинг продукування CO 2. Конверсію цукрів і утворення продуктів аналізували ЯМР-методом. Дані представляють залишкову кількість цукрів у вказаному середовищі (глюкоза, арабіноза, галактоза і ксилоза в грамах/літр) і утворення (побічних) продуктів (етанолу, гліцерину). Моніторинг росту вели за оптичною щільністю культури при 600 нм (Фіг. 10, 11, 12). Тривалість експерименту склала близько 140 годин. Експерименти чітко показують, що референс-штам BIE104 швидко конвертує глюкозу, але він не здатний конвертувати ні арабінозу, ні галактозу за 140 годин (Фіг. 10). Проте штами BIE104A2P1c і BIE201 здатні конвертувати арабінозу і галактозу (Фіг. 11 і 12, відповідно). Утилізація галактози і арабінози почалася відразу ж після виснаження глюкози опісля менше ніж 20 годин. Конверсія обох цукрів була одночасною. Проте штам BIE201, який було покращено для зростання на арабінозі за анаеробних умов, споживав обидва цукри набагато швидше (Фіг. 12). У всіх процесах ферментації наголошувалася утворення тільки гліцерину як побічного продукту. Дані ферментації штамом BIE201 приводяться нижче в табл. 2. Таблиця 2. Концентрації цукрів і концентрації етанолу (г/л) в процесі ферментації штамом BIE201, як показано на Фіг. 12. Максимальна концентрація етанолу розраховується шляхом множення концентрацій на 0,51 для кожного цукру і подальшого підсумовування. Концентрація етанолу в 136 год. (39,2 г/л) означає вихід етанолу – 0,45 етанолу/г цукру. Цей вихід показує, що всі цукри перетворилися на етанол. 24 UA 108853 C2 Концентрації в г/л Час (год.) 0 16 23 39 48 65 111 136 5 10 15 20 25 30 35 40 45 Glu 42,8 0,1 0,0 0,1 0,0 0,0 0,0 0,0 Xyl 50,2 54,1 49,2 52,8 52,5 55,1 48,8 49,6 Ara 31,6 35,8 31,3 16,3 8,9 4,3 0,5 0,2 Gal 12,9 10,8 8,4 0.7 0,2 0,3 0,3 0,3 ETOH 0,7 22,9 18,7 32,1 29,4 40,3 38,1 39,2 Максимальні концентрації етанолу (у г/л) з глюкози 21,8 арабінози 16,1 галактози 6,6 Всього 44,5 Вихід етанолу в експериментах 0,45 г етанолу/г цукру З цього розрахунку очевидно, що кожен з цукрів – глюкоза, галактоза і арабіноза – конвертувався в етанол. Приклад 4 Вплив PPP-генів на конверсію цукрів Для вивчення впливу PPP-генів на конверсію цукрів одиничні колонії штамів BIE104A2 і BIE105A2 використовували для інокуляції YNB-середовища (Difco), до якого додавали 2% глюкози. Обидва штами містять ara-гени і були еволюціонованими для зростання на арабінозі (як описано в Прикладі 2, розділ 2.1). Основою штаму BIE105A2 є промисловий штам. Проте він був трансформований такими ж методами і конструктами, які описані вище (Приклад 1, підрозділ 1.2). Попередні культури збирали і інокулювали в синтетичному модельному середовищі з кукурудзяними волокнами (Verduyn et al., Yeast 8:501-517, 1992; 5% глюкози, 5% ксилози, 3,5% арабінози, 1,5% галактози) з початковою OD 600, яка дорівнює приблизно 2, в BAM. Проводили постійний моніторинг продукування CO2. Конверсію цукрів і утворення продуктів аналізували ЯМР-методом. Дані представляють залишкову кількість цукрів у вказаному середовищі (глюкоза, арабіноза, галактоза і ксилоза в грамах/літр) і утворення (побічних) продуктів (етанолу, гліцерину). Моніторинг зростання вели за оптичною щільністю культури при 600 нм. Тривалість експерименту склала близько 160 годин. Експерименти показують, що обидва штами здатні конвертувати арабінозу і галактозу відразу після виснаження глюкози без гіперекспресії PPP-генів (Фіг. 13 і 14). Приклад 5 Введення конститутивно експресованих генів, які кодують ксилоізомеразу і ксилулокіназу 5.1 Трансформація дріжджів Штам BIE104A2P1 (MATa URA3 HIS3 LEU2 TRP1 MAL2-8 SUC2 SIT2:[TDH3-araA, ENO1araB, PGI1-araD] ΔGRE3:[TPI1p-TAL1, ADH1p-TKL1, PGI1p-RPE1, ENO1p-RKI1]) трансформували плазмідою pPWT042 (Фіг. 16). Плазміда pPWT042 була отримана з вектора pPWT007 (Фіг. 15). Вона містить оптимізовану парою кодонів ксилулокіназу з S. cerevisiae і оптимізовану парою кодонів ксилоізомеразу з Bacteroides uniformis (SEQ 2), як описано в патентній заявці PCT/EP2009/52623. Перед трансформацією штаму BIE104A2P1 плазміда pPWT042 була лінеаризованою з використанням ферменту рестрикції SfiI відповідно до інструкції виробника. Трансформаційні суміші висівали на чашки з YPD-агаром (на літр: 10 грамів дріжджового екстракту, 20 грамів/літр пептону, 20 грамів/літр декстрози, 20 грамів агару), який містить 100 мкг G418 (Sigma Aldrich) /мл. Через дві - чотири доби на чашках з'явилися колонії, тоді як негативний контроль (тобто без додавання ДНК в експерименті з трансформації) не показав зростання на чашках з YPD/G418. Після розщеплювання плазміди pPWT042 за допомогою SfiI її інтеграція націлена на SIT4локус (Gottlin-Ninfa and Kaback (1986) Molecular and Cellular Biology Vol. 6, No. 6, 2185-2197) в геномі (Фіг. 17). Характеристику трансформантів проводили методами ПЛР і Саузерн-блотінга, як описано в Прикладі 1 (розділ 1.2). Штам з однією копією плазміди pPWT042, інтегрованою в геном, був позначений як BIE104A2P1Y9. 25 UA 108853 C2 5 10 15 20 25 30 35 40 45 50 55 60 5.2 Експерименти із зростання Ізоляти одиничних колоній штамів BIE104A2P1Y9 використовували для інокуляції YNBсередовища (Difco), до якого додавали 2% глюкози або 2% галактози. Інокульоване середовище в колбах інкубували при 30 °C і 280 об./хв. до досягнення оптичної щільності при 600 нм значення, що дорівнює, щонайменше, 2,0. YNB-середовище з додаванням 1% арабінози і 1% ксилози інокулювали культурами, які інкубуються протягом ночі, при початковій OD 600, рівній 0,2. Клітини росли при 30 °C і 280 об./хв. Вели регулярний моніторинг оптичної щільності при 600 нм. Коли оптична щільність досягла значення вище 2,0, аліквоту культури переносили на свіже YNB-середовище, яке містить 2% ксилози і 0,2% арабіноз. Кількість доданих клітин була такою, щоб початкова OD 600 культур складала 0,2. Моніторинг оптичної щільності проводили регулярно. Результати представлені на Фіг. 18, панель а (попередні культури на галактозі) і панель b (попередні культури на глюкозі). Результати чітко показують, що штами здатні утилізувати глюкозу, галактозу, арабінозу і ксилозу. 5.3 Збереження маркера Для видалення селективного маркера, введеного інтеграцією плазміди pPWT042, штам BIE104A2P1Y9 культивували в YPD-середовищі, починаючи з ізоляту одиничної колонії. 25 мкл культури, отриманої культивуванням протягом ночі, використовували для інокуляції свіжого YPD-середовища. Після серії послідовних перенесень визначали оптичну щільність культури, і клітини розбавляли до концентрації приблизно 5000 в 1 мл. 100 мкл клітинної суспензії висівали на чашки з середовищем Yeast Carbon Base (Difco), що містить 30 мМ KPi (pH 6,8), 0,1% (NH4)2SO4, 40 мМ фторацетаміду (Amersham) і 1,8% агару (Difco). Стійкі до фторацетаміду колонії піддавали ПЛР-аналізу, а у випадку коректних ПЛР-профілів – Саузерн-блотінгу (підрозділ 1.3, Приклад 1). Одним з штамів, який показали коректний патерн смуг на Саузернблоті, є штам, який був позначений як BIE104A2P1X9. 5.4 Експерименти із зростання Ізоляти одиничних колоній штаму BIE104A2P1X9 (BIE104A2P1X9a1 і BIE104A2P1X9a2) використовували для інокуляції Verduyn-середовища (Difco), до якого додавали 2% глюкози. Колби з інокульованим середовищем інкубували при 30 °C і 280 об./хв. протягом приблизно 24 годин. Verduyn-середовище з додаванням 2% ксилози інокулювали культурами, отриманими інкубацією протягом ночі, з початковою OD 600, рівною 0,2. Клітини росли при 30 °C і 280 об./хв. Проводили регулярний моніторинг оптичної щільності при 600 нм. Результати показані на Фіг. 19. Результати чітко показують, що обидві автономні колонії штаму BIE104A2P1X9 все ще здатні утилізувати ксилозу після вилучення маркера. Як показано вище в Прикладі 3, штам здатний утилізувати глюкозу, арабінозу і галактозу (Фіг. 11 і Фіг. 12). Приклад 6 Трансформація S. cerevisiae для отримання бурштинової кислоти на арабінозі і галактозі 6.1. Експресійні конструкти Експресійний конструкт pGBS414PPK-3, який містить фосфоенол-піруват-карбоксикіназу PCKa (E.C. 4.1.1.49) з Actinobacillus succinogenes і глікосомальну фумаратредуктазу FRDg (E.C. 1.3.1.6) з Trypanosoma brucei, і експресійний конструкт pGBS415FUM3, який містить фумаразу (E.C. 4.2.1.2.) з Rhizopus oryzae і пероксисомальну малатдегідрогеназу MDH3 (E.C. 1.1.1.37), конструювали, як описано раніше в WO2009/065778 на стор. 19-20 і 22-30, включеній до даного опису у вигляді посилання разом з фігурами і списком послідовностей. Експресійний конструкт pGBS416ARAABD, який містить гени araA, araB і araD з Lactobacillus plantarum, конструювали шляхом клонування продукту ПЛР, який містить касету araABDекспресії з плазміди pPWT018, в плазміді pRS416. ПЛР-фрагмент отримували за допомогою ® ДНК-полімерази Phusion (Finnzymes) і ПЛР- праймерів, визначених тут як SEQ ID 22 і SEQ ID 23. ПЛР-продукт розщеплювали ферментами рестрикції SALI і NOTI, як і плазміду pRS416. Після лігування і трансформації E. coli TOP10 відбирали правильні рекомбінанти на основі рестрикційного аналізу. Фізична карта плазміди pGBS416ARAABD представлена на Фіг. 20. 6.2. Штами S. cerevisiae Плазміди pGBS414PPK-3, pGBS415-FUM-3 трансформували в штам S. cerevisiae CEN.PK113-6B (MATA ura3-52 leu2-112 trp1-289). На додачу до цього, у вказаний штам трансформували плазміду pGBS416ARAABD з метою створення прототрофних штамів дріжджів. Експресійні вектори трансформували в дріжджі електропорацією. Трансформаційну суміш висівали на чашки з середовищем Yeast Nitrogen Base (YNB) мас./об. AA (Difco) + 2% 26 UA 108853 C2 5 10 15 20 глюкози. Штами піддавали адаптивній еволюції (див. підрозділ 2) для зростання на арабінозі як єдиному джерелі вуглецю. 6.3. Експерименти із зростання і отримання бурштинової кислоти Трансформантами інокулювали 20 мл середовища для попередньої культури, яке складається з Verduyn-середовища (Verduyn et al., 1992, Yeast. July. 8(7): 501-17), яке містить 2% галактози (мас./об.); зростання трансформантів відбувалося за анаеробних умов в періодично струшуваних колбах на 100 мл в термостаті із струшуючим механізмом при 30 °C і 250 об./хв. Через приблизно 24 години клітини чотири рази переносили на свіже Verduynсередовище, яке містить або 2% глюкози, 2% галактози або 2% арабінози, або їх суміші. Дві колби інкубували за аеробних умов, дві – за анаеробних умов, наприклад, шляхом закривання колб водяним затвором або шляхом інкубації в анаеробному орбітальному шейкері. Зразки культур відбирали з інтервалами в часі. Зразки центрифугували 5 хвилин при 4750 об./хв. 1 мл супернатанту використовували для вимірювання рівнів бурштинової кислоти ВЕРХ-методом (методом високоефективної рідинної хроматографії), як описано в розділі 6.4. 6.4. ВЕРХ-аналіз ВЕРХ застосовували для визначення органічних кислот і цукрів. Принцип розділення на колонці Phenomenex Rezex-RHM-Monosaccharide заснований на розмірній ексклюзії за розміром, іонній ексклюзії і іонному обміні з використанням механізмів оберненої фази. Детектування проводили за допомогою диференціального детектора індексу рефракції і ультрафіолетового детектора. Джерела інформації: № з/п Джерело (1) Bioresource Technology 1994 Vol. 47, p. 283-284 (2) Micard, Enzyme Microbiol. Technology 1996 Vol. 19, p. 163-170 (3) DOE Radke, Idaho wheat straw composition (4) Grohman and Botast Process Biochemistry 1997 Vol. 32, No. 5, 405-415 (5) Saska B&B 1995, 517-523 (6) PCT/EP2009/52623 (7) Zheng Appl. Biochem. Microbiol. 2007 Vol. 136-140, p. 423-436 (8) Bradshaw Appl. Biochem. Microbiol. 2007 Vol. 136-140, p. 395-406 (9) Cara Appl. Biochem. Microbiol. 2007 Vol. 136-140, p. 379-394. 27 UA 108853 C2 28
ДивитисяДодаткова інформація
Назва патенту англійськоюFermentative production of ethanol from glucose, galactose and arabinose employing a recombinant yeast strain
Автори англійськоюKlaassen, Paul, Suylekom, Van, Gijsberdina, Pieterntlla, Gielesen, Bianca, Elisabeth, Maria, Broers, Nicolette, Jasmijn, Wiedemann, Beate, Laat, De, Wilhelmus, Theodorus, Antonius, Maria
Автори російськоюКласен Пол, Сейлеком Ван Гейсбердина Питернела, Гилесен Бианка Элисабет Мария, Брурс Николете Ясмейн, Видеман Беате, Лат Де Вилхелмус Теодорус Антониус Мария
МПК / Мітки
МПК: C12P 7/10
Мітки: ферментації, спосіб, галактози
Код посилання
<a href="https://ua.patents.su/68-108853-sposib-fermentaci-galaktozi.html" target="_blank" rel="follow" title="База патентів України">Спосіб ферментації галактози</a>
Спосіб керування процесом ферментації у виробництві біоетанолу

Номер патенту: 64618
Опубліковано: 10.11.2011
Автори: Ярощук Людмила Дем`янівна, Раухвергер Ірина Анатоліївна
МПК: C12M 1/36
Мітки: керування, процесом, біоетанолу, виробництві, ферментації, спосіб
Формула / Реферат:
Спосіб керування процесом ферментації у виробництві біоетанолу, що включає стабілізацію температури біомаси зміною витрати теплоносія в оболонці ферментера, стабілізацію рН біомаси зміною витрати аміачної води, стабілізацію рівня піни над біомасою уведенням у неї піногасника, стабілізацію тиску надлишкових газів над біомасою зміною їх витрати на виході з ферментера, який відрізняється тим, що вимірюють концентрації етанолу та цукрів у...
Процес ферментації за участю генетично модифікованих дріжджів та спосіб здійснення ферментації

Номер патенту: 88437
Опубліковано: 26.10.2009
Автори: Арістідоу Арістос, Ращ Бріан, ван Хоек Пім
МПК: C12P 7/40, C12N 1/19, C12N 1/16, C12N 15/81, C12P 17/02
Мітки: ферментації, генетично, здійснення, участю, модифікованих, дріжджів, спосіб, процес
Формула / Реферат:
1. Процес ферментації, який відрізняється тим, що мікроорганізм, яким є генетично модифіковані дріжджі, що мають розрив нативного шляху метаболізму PDC, перетворює шляхом ферментації ферментаційний субстрат, а питому швидкість поглинання кисню контролюють протягом виробничої фази цього процесу ферментації і принаймні один робочий параметр регулюють у відповідності з виміряною швидкістю поглинання кисню, причому концентрацію розчиненого кисню...
Спосіб ферментації баклажана

Номер патенту: 65370
Опубліковано: 12.12.2011
Автори: Зінченко Євгенія Василівна, Тернова Тетяна Андріївна
МПК: A23B 7/00
Мітки: ферментації, спосіб, баклажана
Формула / Реферат:
1. Спосіб ферментації баклажана, що включає підготовку сировини бланшуванням у солоній воді, додаванням кухонної солі, ферментуванням, зберіганням, який відрізняється тим, що плоди баклажана нарізані крупною соломкою, залиті киплячим розсолом, в який додають харчовий консервант, з доданням рецептурних компонентів.2. Спосіб за п. 1, який відрізняється тим, що рецептурним компонентом є бланшовані у воді гриби (печериці) з доданням...
Спосіб переробки осаду очисних споруд і ставкового мулу методом біологічної ферментації

Номер патенту: 52957
Опубліковано: 10.09.2010
Автори: Бунчак Олександр Миронович, Гнидюк Володимир Сергійович, Сендецький Володимир Миколайович, Колісник Наталія Михайлівна, Мельник Іван Панасович
МПК: C05F 7/00
Мітки: переробки, очисних, спосіб, біологічно, мулу, осаду, методом, ферментації, ставкового, споруд
Формула / Реферат:
Спосіб переробки осаду очисних споруд та ставкового мулу методом біологічної ферментації, який характеризується тим, що перед ферментацією готують компостну суміш компонентів розрахунковим методом за показниками вологи, азоту та вуглецю, на майданчику осад очисних споруд або ставковий мул змішують з пташиним послідом, гноєм великої рогатої худоби або свиней з додаванням торфу або подрібненої соломи, або очерету, або листя, мікробіологічного...
Спосіб застосування органічних добрив, одержаних методом біологічної ферментації (одуд) в технологіях вирощування картоплі

Номер патенту: 54152
Опубліковано: 25.10.2010
Автор: Бунчак Олександр Миронович
МПК: C05F 11/00
Мітки: застосування, органічних, ферментації, одуд, картоплі, вирощування, методом, технологіях, добрив, біологічно, одержаних, спосіб
Формула / Реферат:
Спосіб застосування органічних добрив, одержаних методом біологічної ферментації (ОДУД) в технології вирощування картоплі шляхом застосування органічного добрива, який відрізняється тим, що органічні добрива ОДУД застосовують як основне добриво, в дозі 5-15 т\га.
Попередній патент: Застосування одного або комбінації фітоканабіноїдів для лікування епілепсії
Наступний патент: Композиції з більш низьким вмістом іміквімоду та короткі режими дозування для лікування генітальних і періанальних бородавок
Випадковий патент: Екрануючий комплект