Процес ферментації за участю генетично модифікованих дріжджів та спосіб здійснення ферментації







Формула / Реферат
1. Процес ферментації, який відрізняється тим, що мікроорганізм, яким є генетично модифіковані дріжджі, що мають розрив нативного шляху метаболізму PDC, перетворює шляхом ферментації ферментаційний субстрат, а питому швидкість поглинання кисню контролюють протягом виробничої фази цього процесу ферментації і принаймні один робочий параметр регулюють у відповідності з виміряною швидкістю поглинання кисню, причому концентрацію розчиненого кисню підтримують на рівні менше 1 % від рівня насичення.
2. Процес за п. 1, який відрізняється тим, що робочим параметром є один або більше параметрів, вибраних із групи, що складається із швидкості аерації, швидкості перемішування і складу аераційного газу.
3. Процес за п. 1, який відрізняється тим, що мікроорганізм має принаймні один функціональний екзогенний ген, який дозволяє клітині виробляти бажаний продукт ферментації.
4. Процес за п. 3, який відрізняється тим, що екзогенним геном є лактатдегідрогеназний ген.
5. Процес за п. 4, який відрізняється тим, що мікроорганізм є роду Candida або Kluyveromуces.
6. Спосіб здійснення ферментації у ферментаційному середовищі, що містить ферментуючий мікроорганізм, субстрат, що піддається ферментації цим мікроорганізмом, який (спосіб) відрізняється тим, що ферментаційний бульйон має певну кількість розчиненого кисню (DO), а ферментація показує питоме поглинання кисню (OUR), і який включає у себе:
а) вимірювання OUR протягом виробничої фази ферментації;
b) регулювання умов аерації таким чином, що OUR підтримується в межах заданого діапазону при підтриманні DO на рівні нижче 1 % насичення протягом виробничої фази ферментації,
причому OUR на стадії b) підтримують у межах приблизно від 0,8 до 3,0 ммоль О2/г. с.м./год., а DO підтримують на рівні нижче 10 мкмоль О2/л, та
де зазначеним мікроорганізмом є дріжджова клітина, що виказує Крабтри-негативний фенотип, причому дріжджова клітина є роду Kluyveromуces або Candida та має розірваний PDC-шлях метаболізму і принаймні один функціональний екзогенний ген, що дозволяє клітині виробляти бажаний продукт ферментації, причому екзогенним геном є лактатдегідрогеназний ген.
7. Спосіб за п. 6, який відрізняється тим, що субстрат включає гексозний цукор.
8. Спосіб за п. 7, який відрізняється тим, що гексозний цукор включає глюкозу.
Текст
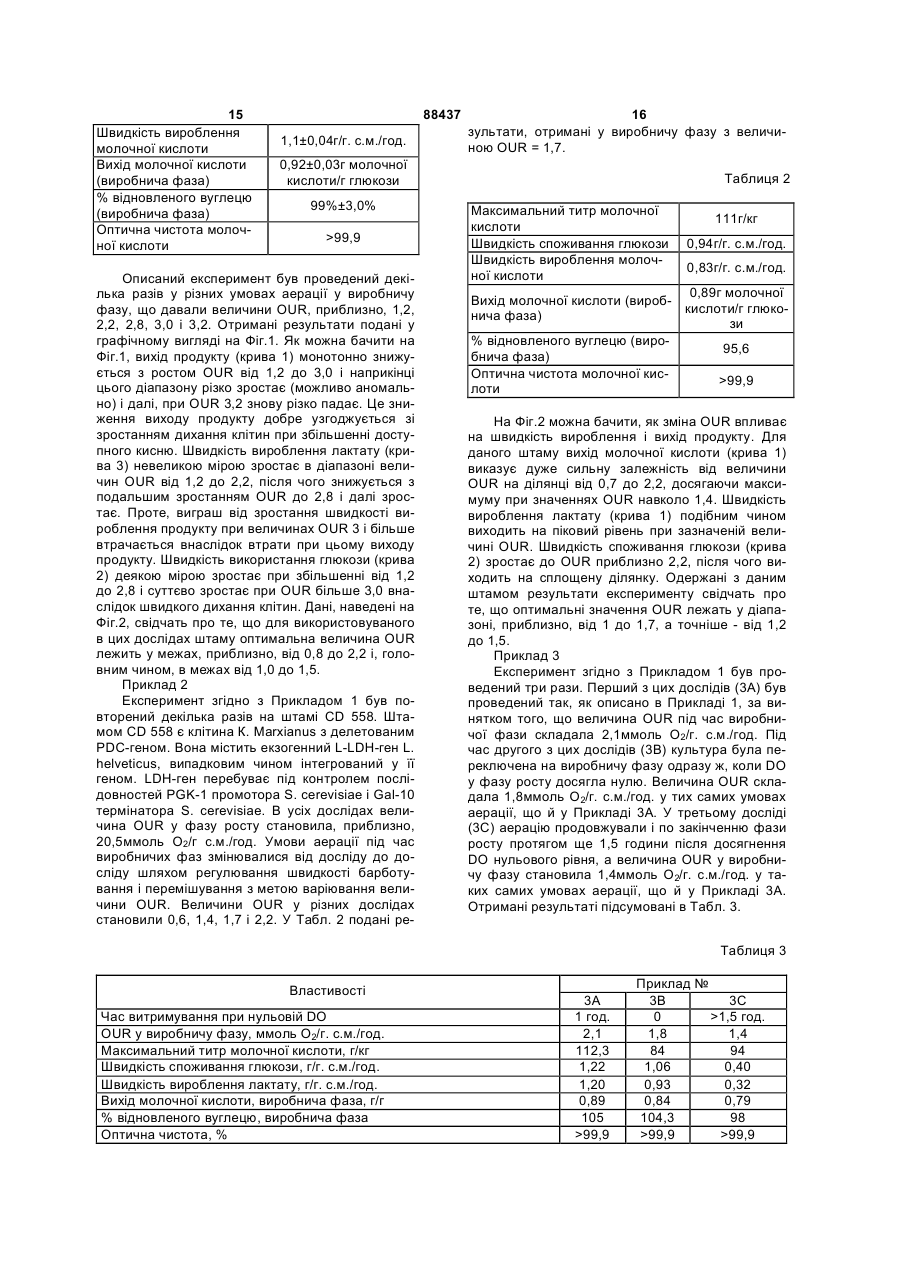
1. Процес ферментації, який відрізняється тим, що мікроорганізм, яким є генетично модифіковані дріжджі, що мають розрив нативного шляху метаболізму PDC, перетворює шляхом ферментації ферментаційний субстрат, а питому швидкість поглинання кисню контролюють протягом виробничої фази цього процесу ферментації і принаймні один робочий параметр регулюють у відповідності з виміряною швидкістю поглинання кисню, причому концентрацію розчиненого кисню підтримують на рівні менше 1 % від рівня насичення. 2. Процес за п. 1, який відрізняється тим, що робочим параметром є один або більше параметрів, вибраних із групи, що складається із швидкості аерації, швидкості перемішування і складу аераційного газу. 3. Процес за п. 1, який відрізняється тим, що мікроорганізм має принаймні один функціональний 2 (19) 1 3 Дана заявка претендує на пріоритет попередньої заявки США №60/384,333, поданої 30 травня 2002 р. Даний винахід був створений за підтримки уряду США згідно з Контрактом №DE-FC-3600G010598 від Департаменту енергетики. Уряд США має в даному винаході певну частину прав. Молочна кислота широко застосовується в промисловості і, в тому числі, в хімічній переробці та синтезі, косметиці, фармацевтичному виробництві, виробленні пластмас і в харчовій промисловості. У найбільших промислових масштабах молочну кислоту одержують за допомогою процесів ферментації. У таких процесах використовувалися найрізноманітніші бактерії-продуценти молочної кислоти. Останнім часом проводилися дослідження з використання рекомбінантних дріжджових штамів у процесах ферментації молочної кислоти. Рекомбінантні дріжджі мають потенційну спроможність надати ряд переваг порівняно з бактеріальною ферментацією. Деякі дріжджові штами є більш стійкими до високих температур. Це могло б дозволити ферментацію при вищих температурах, а отже і на більші швидкості ферментації. Краща толерантність до високих температур полегшує задачу очищання ферментаційного середовища від мікробів, що його забруднюють, оскільки це середовище можна в такому разі просто нагрівати до температури, при котрій небажані види гинуть, а бажані залишаються живими. Бактеріїпродуценти молочної кислоти, такі, як Lactobacilli, потребують для підвищення ефективності виробничого процесу більш складного ферментаційного середовища. Складність ферментаційного середовища підвищує вартість сировинних матеріалів і робить більш складним і дорогим відділяння молочної кислоти від ферментаційного середовища. Використання ж рекомбінантних дріжджів дає можливість зменшувати виробничі витрати завдяки застосуванню спрощеного ферментаційного середовища. Поро (Porro) зі співробітниками спробували за допомогою методів генної інженерії вивести дріжджі, що продукують молочну кислоту, шляхом вставляння екзогенного LDH-гена (лактатдегідрогенази) в клітини дріжджів видів S. cerevisiae, К. lactic, Т. delbrueckii та Z Bailii і розривання природного метаболічного шляху пірувату цих клітин [Porro et аl., "Development of metabolically engineered Saccharomyces cerevisiae cells for the production of lactic acid", Biotechnol. Prog. 1995 May-Jun; 11(3): 294-8; Porro et al., "Replacement of a metabolic pathway for large-scale production of lactic acid from engineered yeasts", App. Environ. Microbiol. 1999 Sep: 65(9):4211-5; Bianchi et al., "Efficient homolactic fermentation by Kluyveromyces lactis strains defective in pyruvate utilization and transformed with the heterologous LDH gene", App. Environ. Microbiol. 2001 Dec; 67(12) 5621-5]. Поро зі співроб. спромоглися виробити рекомбінантні дріжджі, що продукували молочну кислоту, але ці штами працювали недостатньо добре для того, щоб їх можна було застосовувати у промислових процесах. Для ви 88437 4 знання штаму придатним до промислового застосування він повинен давати достатньо високий вихід молочної кислоти (тобто забезпечувати високе перетворення субстрату на молочну кислоту) і мати високу продуктивність (тобто швидке метаболічне перетворення субстрату на молочну кислоту). Такі дріжджі повинні також витримувати середовище з високим титром молочної кислоти. Нещодавно Раджгарія (Rajgarhia) зі співроб. створили рекомбінатні дріжджі, які демонстрували більш високі вихід продукту і продуктивність, ніж дріжджі Поро, див., наприклад, [WO 00/71738, WO 2/42471 і PCT/US02/16223]. У роботі Раджгарія [WO 00/71738] була здійснена спроба використати переваги так званого "Крабтри-негативного" фенотипу, що проявляється деякими видами дріжджів. Ефект Крабтри (Crabtree) є відомим як явище ферментативного метаболізму в аеробних умовах, викликаний інгібуванням споживання кисню мікроорганізмом, що культивується з високими питомими швидкостями росту (довготривалий ефект) або при наявності високих концентрацій глюкози (короткотривалий ефект). Крабтринегативні фенотипи цього ефекту не виказують і, таким чином, є здатними споживати кисень навіть при наявності високих концентрацій глюкози або при високих швидкостях росту. Таким чином, культури Крабтри-негативних мікроорганізмів, принаймні теоретично, можна переводити із фази культивування у фазу ферментації (виробничу фазу) шляхом регулювання постачання кисню. В умовах значної аерації мікроорганізми ростуть, продукуючи біоматеріал і CO2, у той час як в анаеробних умовах клітини замість цього піддають ферментації доступний субстрат, виробляючи молочну кислоту або інші продукти зброджування. Проте авторами даного винаходу було знайдено, що ферментація деякими штамами здійснюється не так ефективно, як було б бажано мати в строгих анаеробних умовах. Це спостерігається, наприклад, у генетично модифікованих дріжджових штамах, у котрих шлях метаболізму піруватдекарбоксилази (PDC) є делетований або розірваний. Але одержувані таким способом види було б дуже бажано використовувати у ферментації молочної кислоти (а також інших продуктів, що не є етанолом), оскільки розривання шляху PDC зменшує кількість етанолу, що продукується. Отже очевидною стає потреба у створенні поліпшеного процесу ферментації, в котрому штам, що у строгих анаеробних умовах зброджує неефективно, є здатним виробляти бажаний продукт ферментації із забезпеченням високої економічності процесу. На Фіг.1 показаний графік, що ілюструє вплив OUR на споживання глюкози, продукування молочної кислоти і на корисний вихід для деяких генетично модифікованих видів К. marxianus. На Фіг.2 показаний графік, що ілюструє вплив OUR на споживання глюкози, продукування молочної кислоти і на корисний вихід для інших генетично модифікованих видів К. marxianus. Даним винаходом в одній із його ознак є процес ферментації, в котрому питому швидкість 5 88437 6 поглинання кисню контролюють протягом виробдоброю швидкістю виробничого процесу. Виміничої фази цього процесу ферментації і, принайрювання OUR можуть використовуватися для мні, один робочий параметр регулюють відповідвстановлення і регулювання деяких параметрів но до виміряної швидкості поглинання кисню. процесу ферментації з метою підтримання оптиУ другій його ознаці даним винаходом є спомальних умов у виробничій фазі цього процесу. сіб здійснення процесу ферментації у ферментаПараметр OUR являє собою швидкість споційному середовищі, що містить ферментуючий живання кисню (O2) на одиницю сухої маси мікмікроорганізм, субстрат, що піддається ферменроорганізму за одиницю часу. OUR можна вигідно тації цим мікроорганізмом, де ферментаційне визначати, виходячи з кількості кисню, що спожисередовище показує кількість розчиненого кисню вається за одиницю часу, і з маси клітин протя(DO: dissolved oxygen) і питоме поглинання кисню гом цього часу. Споживання кисню визначають (OUR) під час ферментації, який включає у себе: шляхом вимірювання кількості кисню, введеного a) вимірювання OUR протягом виробничої у ферментаційну посудину і виведеного із неї за фази ферментації; одиницю часу. Отже величина OUR отримується b) регулювання умов аерації так, щоб підтриділенням кількості спожитого кисню на суху масу мувати OUR у заданих межах, підтримуючи при біоречовини в бульйоні. Маса біоречовини може цьому DO на рівні нижче 1% від рівня насичення бути визначена шляхом відбирання зразка, виміпід час виробничої фази ферментації. рювання концентрації в ньому клітин (маса/об'єм) У третій його ознаці даним винаходом є проі множення цієї концентрації на загальний об'єм цес, який включає у себе: бульйону. Кількість уведеного кисню може бути a) визначення оптимального діапазону веливиміряна безпосередньо за даними моніторингу чин OUR, при котрих ферментуючий мікрооргашвидкості аерації. Кількість кисню, виведеного із нізм перетворює вуглевод у бажаний продукт ферментаційної посудини, може бути виміряна за ферментації; допомогою будь-якого відомого методу і, зокреb) культивування мікроорганізму в середовима, за допомогою мас-спектроскопії, що є в дащі, що містить вуглеводень, який піддається меному випадку особливо підходящим. Різниця між таболічному перетворенню клітиною, і одну чи кількістю введеного кисню і кількістю видаленого більше живильних речовин, при аерації цього кисню показує, таким чином, кількість кисню, середовища таким чином, що коли клітини росспожитого клітинами. Величину OUR обчислюють туть і репродукуються, DO в середовищі зменшушляхом ділення кількості спожитого кисню на ється до рівня менше 1% від величини насиченсуху масу клітин і тривалість часу спостережень. ня, і клітини показують питому швидкість Виражають OUR у мілімолях O2 на грам (сухої поглинання кисню не менше 10ммоль O2/грам маси) клітин за годину. сухої маси клітин за годину (ммоль O2/г с.м./год.); У найбільш загальному варіанті здійснення і після цього винаходу проводять вимірювання OUR протягом c) культивування мікроорганізму в буферновиробничої фази ферментації і регулюють, приму середовищі в умовах ферментації, включаючи наймні, один параметр процесу ферментації відумови мікроаерації, достатні для забезпечення повідно до виміряних величин OUR. Параметром, культури питомою швидкістю поглинання кисню котрий регулюють у відповідністю з виміряною (OUR) в оптимальному діапазоні. величиною OUR і звичайно пов'язується з аераВ іншій його ознаці даний винахід являє соцією ферментаційного бульйону, є, наприклад, бою процес ферментації, що включає у себе: швидкість аерації, швидкість перемішування, а) культивування генетично модифікованих склад аераційного газу (наприклад, збільшення дріжджових клітин, що мають розірваний шлях або зменшення концентрації кисню в газі), або ж метаболізму PDC і екзогенний ген, що дозволяє інший параметр, що впливає на швидкість, з якою клітині виробляти бажаний продукт ферментації у мікроорганізми в бульйоні споживають кисень. середовищі, котре містить вуглевод, що піддаУ кращому варіанті здійснення винаходу феється метаболічному перетворенню цією клітирментуючі клітини культивуються в різноманітних ною, при аерації цього середовища таким чином, умовах аерації з метою емпіричного встановленщо коли клітини ростуть і репродукуються, кільня оптимального діапазону величин OUR для кість розчиненого кисню в середовищі знижується конкретного типу клітин. Оптимальний діапазон до рівня менше 1% від величини насичення, і OUR у загальному випадку залежить від декільклітини показують питому швидкість поглинання кох чинників, серед яких найголовнішими є три кисню менше 10ммоль O2/г сухої маси клітин за такі: вихід бажаного продукту ферментації із фегодину (ммоль О2/г с.м./год.); і після цього рментаційного субстрату (звичайно виражений у b) культивування клітин у буферному сереграмах продукту на грам витраченого субстрату), довищі в умовах ферментації, включаючи умови питома продуктивність одержання бажаного промікроаерації, достатні для забезпечення культури дукту ферментації (звичайно виражається як мапитомою швидкістю поглинання кисню (OUR) в са продукту, поділена на суху масу клітин і на межах, приблизно, від 0,8 до-3,0ммоль O2/г час) і швидкість споживання субстрату (що вирас.м./год. жається, звичайно, як маса субстрату, спожитого Авторами було знайдено, що використання за одиницю часу). Ці чинники не завжди є всі опумов мікроаерації та швидкості поглинання кисню тимізовані в одних і тих самих умовах. Наприяк параметра для регулювання процесу дозволяє клад, швидкість споживання субстрату іноді зросоптимізувати процес ферментації, врівноважуючи тає зі зростанням OUR, але вихід при цьому має високий вихід бажаного продукту ферментації з тенденцію до зниження, протидіючи таким чином 7 88437 8 підвищенню швидкості виробничого процесу зі с.м./год. Клітини К. marxianus, що мають розрив збільшенням втрат виходу продукту і погіршуючи PDC-шляху й екзогенний LDH-ген, виказують здаекономічні показники процесу. Встановлення оптність давати максимальну величину OUR в метимального діапазону величин OUR у загальному жах, приблизно, 20-22ммоль O2/г с.м./год. випадку спричиняє врівноваження швидкостей з Рівень DO може бути і в кращому варіанті є виходом продукту для оптимізації усіх економічвище нуля протягом більшої частини фази росту них показників процесу. Якщо встановлюється за умови, що він знижується, приблизно, до нуля бажаний діапазон величин OUR, то умови фернаприкінці цієї фази. Отже, надлишок кисню біментації вибирають у виробничу фазу ферменльше цього рівня, що потребується для підтритації для встановлення і підтримання OUR в цих мання необхідної величини OUR, може вводитимежах. Як і раніше, величини OUR виміряють під ся в середовище на протязі більшої частини фази час виробничого процесу і підтримують у визнаросту і особливо на протязі періоду, під час якого ченому діапазоні шляхом регулювання одного чи клітини ростуть за експонентою. Оскільки загальбільше параметрів ферментації. не поглинання кисню під час фази росту збільшуУ виробничу фазу концентрацію розчиненого ється разом з репродукуванням клітин і накопикисню підтримують приблизно на нульовому рівченням біоматеріалу, загальна кількість кисню, ні, що відбиває умову того, що клітини при цьому що потребується для підтримання постійного споживають кисень приблизно з такою самою рівня OUR, буде зростати. Проте, оскільки DO швидкістю, з якою він розчиняється у ферментанаприкінці фази росту може відігравати позитивційному бульйоні. Концентрація розчиненого кисну роль, аерацію краще проводити таким чином, ню під час виробничої фази в загальному випадщоб кисень постачався з надлишком на початку і ку є нижчою 1% від рівня насичення (тобто від протягом експоненціальної стадії фази росту. Зі максимальної кількості кисню, яка може бути роззбільшенням накопиченого біоматеріалу загальна чинена в даному бульйоні в даних умовах темпекількість кисню, спожитого клітинами, зростає до ратури і тиску). Звичайно, підходящим є вміст рівня, на якому кількість постаченого кисню майрозчиненого кисню на рівні менше ніж, приблизже дорівнює кількості кисню, спожитого клітинано, 10мкмоль/л і краще - на рівні менше ніж, прими, внаслідок DO чого падає. Отже, постійні умоблизно, 5мкмоль/л. У найкращому варіанті вміст ви аерації можуть використовуватися у фазу розчиненого кисню є практично нульовим. Концеросту, якщо вони вибираються таким чином, що нтрацію розчиненого кисню звичайно виміряють DO падає до нуля разом з потрібною величиною за допомогою кисневого електроду з газопроникOUR, коли кількість виробленої маси досягає ною мембраною (електроду Кларка), який випусзаданого рівня. В альтернативному варіанті умокається, наприклад, фірмою "lngold" і розповсюви аерації у фазу росту можуть змінюватися за джується фірмою "В Braum" під номерами умови, що величина OUR підтримується на зада33182418 і 33182400. Здійснення виробничого ному рівні, a DO наприкінці фази росту стає припроцесу з рівнями розчиненого кисню нижче межі близно нульовою. виявлення таких вимірювальних інструментів Як тільки DO падає приблизно до нуля, а вивважається таким, що відповідає приблизно нуроблений біоматеріал досягає бажаної кількості, льовій концентрації розчиненого кисню згідно з ці умови аерації (OUR принаймні 10ммоль О2/г цілями даного винаходу. с.м./год. і DO = 0) слід підтримувати протягом В особливо кращому варіанті здійснення випевного часу перш, ніж переходити до виробнинаходу мікроорганізм культивують у різних умочої фази. Цей час складає приблизно від 15 хвивах росту і виробничого процесу. У фазу росту лин до 2 годин і в кращому варіанті - від 30 до 90 мікроорганізм перебуває в аеробних умовах. Кліхвилин. Найкращою є тривалість цього проміжку тини вирощуються в середовище що містить вочасу, приблизно, від 45 до 75 хвилин. Якщо мікду, вуглевод, який вони здатні переробляти шляроорганізм переключають на виробничу фазу хом метаболізму як у фазу росту, так і у надто швидко, то клітини будуть давати низьку виробничу фазу, і різноманітні живильні речовишвидкість продукування. Якщо ж зазначені умови ни, які більш повною мірою описані нижче. Умови аерації тривають занадто довго, то клітини буаерації вибирають таким чином, щоб (1) клітини дуть давати низький вихід бажаного продукту показували питому швидкість поглинання кисню ферментації, а також низьку швидкість продуку(OUR) не менше 10ммоль O2/г с.м./год. і (2) навання. прикінці фази росту концентрація розчиненого Культуру переключають на виробничу фазу кисню (DO) в середовищі знижувалася до рівня шляхом змінювання умов аерації. У виробничу менше, ніж 1% від рівня насичення при підтрифазу умови аерації добирають таким чином, щоб манні OUR на рівні, принаймні, 10ммоль O2/г величина OUR підтримувалася у заданому діапас.м./год. Величина OUR у кращому варіанті склазоні, як це зазначалося вище. Деякі мікроорганіздає, принаймні, 15, а в більш кращому - принайми виказують потребу у метаболічному перетвомні 18ммоль O2/г с.м./год. У фазу росту величина ренні малої кількості кисню для підтримання OUR в найкращому варіанті є такою високою, яку загальної життєздатності і здоров'я клітини. Татільки можуть дати вживані клітини. Отже максикими мікроорганізмами є, наприклад, генетично мальна величина OUR залежать певною мірою модифіковані дріжджі, що мають розрив PDCвід використовуваних дріжджових клітин, виведешляху і, зокрема, мікроорганізми, що мають екзоних шляхом генної інженерії. У загальному випагенний ген, який дозволяє їм продукувати певний дку очікується, що максимальна величина OUR продукт ферментації. У цілком анаеробних умобуде складати, приблизно, 20-30ммоль O2/г вах швидкість споживання субстрату і швидкість 9 88437 10 вироблення продукту ферментації, які демондосягнення бажаних величин OUR. струють ці клітини, звичайно є дуже низькими. У тому випадку, коли продуктом ферментації Крім того, втрат зазнає при цьому також вихід є кислота, середовище під час виробничої фази бажаного продукту ферментації. У мікроаеробних буферизують таким чином, щоб його рН підтриумовах при певних величинах OUR, що залежать мати в межах, приблизно, від 5,0 до 9,0, а в кравід конкретного штаму, клітини є здатними метащому варіанті - від 5,5 до 7,0. Підходящими для болічно перетворювати субстрат з набагато біцього буферними агентами є основні матеріали, льшою швидкістю. Результатом цього є зростанщо нейтралізують молочну кислоту при її утвоня швидкостей споживання субстрату і ренні. Це є, наприклад, гідроксид кальцію, карбовироблення бажаного продукту ферментації. Але нат кальцію, гідроксид натрію, гідроксид калію, як тільки споживання кисню, зростаючи, перевикарбонат калію, карбонат натрію, карбонат амощує певний рівень, вихід бажаного продукту зменію, аміак, гідроксид амонію і т.д. У загальному ншується, оскільки більша частина субстрату випадку використовуватися можуть також буферперетворюється на двоокис вуглецю. Крім того, ні агенти, які застосовуються у звичайних процеколи величина OUR перевищує певний рівень, сах ферментації. крива швидкості вироблення продукту виходить Інші умови ферментації, такі, як температура, на сплощену ділянку або навіть знижується, і густина клітин, субстрат або субстрати, живильні втрати виходу продукту більше не компенсуються речовини тощо, не вважаються критичними для підвищенням швидкості його вироблення. Отже даного винаходу і можуть добиратися з точки підтримування величини OUR у певних межах під зору економічності процесу. Температури як у час виробничої фази дозволяє досягати економіфазу росту, так і у виробничу фазу можуть варіючно оптимального врівноваження між виходом вати від температури заморожування ферментапродукту і швидкістю його вироблення. ційного середовища до температури вище 50°С, Оптимальна величина OUR деякою мірою хоча оптимальна температура звичайно певною залежить від використовуваного мікроорганізму, мірою залежить від конкретного мікроорганізму, хоча у загальному випадку діапазон OUR лежить що використовується. У кращому варіанті темпеу межах, приблизно, від 0,8 до 3ммоль О2/г ратура процесу особливо на виробничій фазі с.м./год. Оптимальні величини OUR для конкретлежить у межах 30-45°С. Так, наприклад, вивеного мікроорганізму легко визначаються емпіричдені за допомогою методів генної інженерії клітиним шляхом. Нижній край кращого діапазону ни К. Marxianus, витримують відносно високі темOUR визначається величиною, приблизно, ператури (приблизно від 40 до 50°C, і особливо 1,0ммоль О2/г с.м./год., а в ще кращому варіанті до 45°С). Інший, підходящий вид клітин, C. 1,2ммоль О2/г с.м./год. Верхній край кращого діаSonorensis здатний витримувати температури до пазону OUR визначається величиною, приблиз40°С. Цей температурний діапазон дає можлино, 2,5ммоль О2/г с.м./год., а в ще кращому варівість проводити ферментацію при таких підвищеанті - 2,0ммоль О2/г с.м./год. них температурах (знижуючи тим самим витрати Величина OUR, отримувана від культури, вена охолодження) без значних втрат продуктивноликою мірою залежить від самого мікроорганізму сті. Іншою перевагою, що дає здатність мікроорта умов аерації. Умови аерації впливають на кіганізму витримувати високі температури, є те, що лькість кисню, що розчиняється в культуральнов разі забруднення ферментаційного середовища му середовищі і, таким чином, стає доступним небажаними мікроорганізмами, останні можуть для цього мікроорганізму. Для даного мікрооргабути селективно вбиті нагрівом ферментаційного нізму OUR підвищенню сприяє (1) зростання середовища до 40°C і вище і, зокрема, до 45°C і швидкості постачання кисню і (2) утворення дрібвище, не завдаючи значної шкоди бажаним кліних кисневих пухирців (для поліпшення масопетинам за даним винаходом. реносу молекул кисню в рідку фазу). Утворення У виробничу фазу концентрація клітин у фепухирців можна легко здійснювати шляхом баррментаційному середовищі звичайно лежить у ботування і/або перемішування. межах, приблизно, 1-150, у кращому варіанті - у Швидкість аерації у фазу росту може складамежах, приблизно, 3-10, а в ще кращому - в мети, наприклад, приблизно від принаймні 0,2 об'жах, приблизно, 3-6г сухих клітин на літр фермеєму повітря/об'єм ферментаційного середовинтаційного середовища. ща/хв. (об./об./хв.), у кращому варіанті Добір вуглеводу для ферментаційного сереприблизно від 0,3 до 2об./об./хв., а в ще кращому довища залежить від використовуваної клітини- приблизно від 0,4 до 1об./об./хв. У виробничу хазяїна і від того, чи розрахована дана клітинафазу цей інтервал лежить у межах, приблизно, хазяїн, що була виведена із застосуванням генної від 0,01 до 0,1об./об./хв., краще - від 0,02 до інженерії, на метаболічне перетворення цього 0,75об./об./хв. і найкраще - від 0,02 до вуглеводу на піруват. Кращими вуглеводами при 0,5об./об./хв. У тих випадках коли як аераційний цьому є гексозні цукри, такі, як глюкоза, фруктоза, газ використовується кисень, зазначені об'єми глюкозні олігомери, такі, як мальтоза, ізомальтопропорційно зменшуються.. Аерацію краще здійза, мальтотриоза, крохмаль і мальтодекстрини. У снювати в умовах, наприклад, барботування, що випадку олігомерних цукрів може потребуватися сприяють утворенню дрібних пухирців газу. Позидодавання до ферментаційного бульйону фертивним чинником при цьому є перемішування і, ментів з метою перетравлення цих олігомерів на особливо, коли потребуються високі величини мономерний цукор. Підходящим пентозним цукOUR. Звичайно швидкість аерації і перемішуванром є, наприклад, ксилоза. Найкращою при цьоня добирають невід'ємно одна від одної з метою му є глюкоза. 11 88437 12 При буферній ферментації кислотні продукти стала зменшеною, принаймні, на 90%. Розрив ферментації, такі, як молочна кислота, при їх може досягатися змінюванням шляху метаболізутворенні нейтралізуються у відповідний лактат. му (або одного чи більше генів, зв'язаних зі шляОтже відновлення кислоти спричиняє регенерахом метаболізму) таким чином, що функція шляцію вільної кислоти. Звичайно це здійснюється ху зменшується або вилучається, або ж шляхом видалення клітин і підкислення ферменвидаленням одного чи більше генів, потрібних таційного бульйону сильною кислотою, якою модля функціонування даного шляху. Кращою кліже бути, наприклад, сірчана кислота. Утворювана тиною є така, що має делецію РРС-гена. побічна сіль (якою в тих випадках, коли як нейтКращим екзогенним геном є лактатдегідрогералізатор використовується сіль кальцію, а як назний (LDH) ген. Цей ген у кращому варіанті підкислювач - сірчана кислота, є сульфат кальвбудовується в геном клітини. Генетично модифіцію) відокремлюється від кислоти. Після цього кована дріжджова клітина може мати одну копію кислоту відновлюють за допомогою таких метоабо множину копій екзогенного LDH-гена. Вона дів, як екстракція рідких речовин рідинами, дисможе містити два чи більше різних LDH-генів. У тиляція, абсорбція та інші, описані, наприклад, у ще кращому варіанті здійснення винаходу цей [Т.В. Vickroy, Vol. 3, Chapter 38 of Comprehensive ген інтегрується в геном клітини на місце нативBiotechnology, (ed. M. Moo-Young), Pergamon, ного PDC-гена, який делетується. В особливо Oxford, 1985; R. Datta, et al., FEMS Microbiol. Rev., кращому варіанті здійснення винаходу LDH-ген 1995; 16:221-231; U.S. Patent Nos. 4,275,234, перебуває під функціональним керуванням з боку 4,771,001, 5,132,456, 5,420,304, 5,510,526, оперативних промоторних і термінаторних послі5,641,406, 5,831,122, і Міжнародна патентна задовностей, що є принаймні на 90% гомологічними явка №: WO 93/00440]. промоторним і термінаторним послідовностям Ферментаційне середовище звичайно містить (зокрема, промоторним і термінаторним послідоживильні речовини, потрібні для даних клітин, вностям PDC), що є нативними для даної клітини. включаючи джерело азоту (наприклад, амінокисЗазначені кращі й особливо кращі клітини описані лоти, білки, неорганічні джерела азоту, такі, як більш докладно в попередній патентній заявці аміак або солі амонію, тощо), різноманітні вітаміСША №60/384,333 від 30 травня 2002 p., включени, мінерали і т.п. ній тут шляхом посилання. Процес за даним винаходом може здійснюШтамами, що мають підходящі Lватися у безперервному режимі, періодичному лактатдегідрогеназні гени, котрі можуть бути клорежимі або в комбінованому режимі того та іншоновані для застосування у виробленні технологіго. чних дріжджів, є Lactobacillus helveticus, Мікроорганізмом для використання в процесі Pediococcus acidolactid, Lactobacillus casei, Kluyза даним винаходом може бути будь-який мікроveromyces thermotolerans, Torulaspora delbrueckii, організм, котрий (1) перетворює шляхом фермеSchizosaccharomyces pombii і В. megaterium. нтації вуглевод на бажаний продукт і (2) здійснює Кращими серед них є два L-лактатдегідрогеназні ферментацію більш ефективно у мікроаеробних гени - L. helveticus і В. Megaterium Lумовах, ніж у жорстких анаеробних умовах. Осолактатдегідрогеназа. Штамами, що мають підхобливо прийнятними для цього є деякі генетично дящі D-лактатдегідрогенази, які можуть бути кломодифіковані дріжджові клітини, які відрізняються новані для використання в модифікованих дріжтим, що вони мають (1) розірваний PDC-шлях і джах, є Lactobacillus helveticus, Lactobacillus (2) принаймні один функціональний екзогенний johnsonii, Lactobacillus bulgaricus, Lactobacillus ген, що дозволяє клітині виробляти бажаний проdelbruecki, Lactobacillus plantarum і Lactobacillus дукт ферментації. Такого роду підходящі дріжpentosus. Кращим серед них Dджові клітини описані, наприклад, у [Porro et al., лактатдегідрогеназним геном є L. helveticus D"Development of metabolically engineered лактатдегідрогеназа. Saccharomyces cerevisiae cells for the production of Для можливості промислового застосування lactic acid", Biotechnol Prog. 1995 May-Jim; 11(3): генетично модифікована клітина повинна мати 294-8; Porro et al., "Replacement of a metabolic декілька таких властивостей. Дріжджі повинні pathway for large-scale production of lactic acid from перетворювати значну частину вуглеводу на баengineered yeasts", App. Environ. Microbiol. 1999 жаний продукт ферментації (тобто давати висоSep:65(9):4211-5; Bianchi et al., "Efficient кий вихід продукту). Вона повинна забезпечувати homolactic- fermentation by Метформин lactis високу питому продуктивність, тобто виробляти strains defective in pyruvate utilization and велику кількість продукту ферментації відносно transformed with the heterologous LDH gene", App. маси клітини за одиницю часу. Бажано також, Environ. Microbiol. 2001 Dec; 67(12)5621-5; WO щоб така клітина була толерантною до високих 00/71738, WO 02/42471, PCT/US02/16223, і попеконцентрацій продукту ферментації. Остання з редня заявка США №60/384,333, подана 30 травцих властивостей дозволяє у процесі ферментаня 2002 р.]. У кращому, варіанті дана клітина тації використовувати високі концентрації сировинкож виказує Крабтри-негативний фенотип, тобто ного вуглеводу. вона дихає і росте в умовах аерації, наявності У загальному випадку вважається за бажане, високих концентрацій глюкози і високих питомих щоб процес ферментації згідно з даним винахошвидкостей росту. дом мав деякі або всі перелічені нижче ознаки. Використовуваний тут термін "розірваний" А. Вихід продукту: принаймні 30, краще - приозначає, що нативний шлях метаболізму PDC був наймні 40, ще краще - принаймні 60 і ще краще змінений таким чином, що функція шляху PDC принаймні 75г продукту ферментації на грам суб 13 88437 14 страту. Теоретично бажаний вихід продукту у рації деяких варіантів практичного здійснення такому процесі становить 100%, але на практиці винаходу і жодною мірою не обмежують його об'він обмежується величиною, приблизно, 98%. єму та ідеї. Якщо не зазначено іншого, то всі виВ. Питома продуктивність: принаймні 0,1, кракористовувані тут співвідношення в частинах або ще - принаймні 0,3, ще краще - принаймні 0,4 і ще відсотках виражені в одиницях маси. краще - принаймні 0,5г продукту ферментації на Приклад 1 грам клітин за годину. Питому продуктивність Інокуляційний штам генетично модифіковабажано мати якомога більшою. них дріжджових клітин, позначений як CD 587, C. Титр (максимальна концентрація продукту був приготований у 250мл шейк-колбі, що містила ферментації): принаймні 15г на літр фермента100мл СаСО3-буферизованого (42г/л) дріжджоційного середовища, краще - принаймні 20г/л, ще вий екстракт (10г/л) - пептонового (20г/л) середокраще - принаймні 40г/л, ще краще - принаймні вища з 100г/л глюкози. При OD600 = 10 клітини 80г/л, до 150г/л, і краще - приблизно до 120г/л. У збиралися шляхом центрифугування і після цього випадку молочної кислоти температура ферменповторно суспендувалися в 15% (мас/об.) розчитаційного середовища деякою мірою впливає на ну гліцерину і зберігалися в 1,5мл аліквотах при верхній край титрів, що легко досягається, оскільтемпературі -80°С. ки висококонцентровані розчини молочної кислоКлітиною CD 587 є клітина К. Marxianus, у коти (тобто, близько 150г/л) мають схильність до трій PDC-ген делетований, а в її геном на місце того, щоб ставати дуже в'язкими або гелеподібделетованого PDC-гена вбудований екзогенний ними при температурах нижче, ніж приблизно D-LDH-ген L. helveticus під керуванням нативних 35°C. Застосування більш високих температур промоторних і термінаторних послідовностей ферментації, наприклад, у межах 35-50°C дозвоPDC. Клітина CD 587 та її одержання докладно ляє на більші титри без небезпеки гелеутворення описані в попередній заявці США №60/384,333 або небажаного підвищення в'язкості. від 30 травня 2002 р. Крім того, процес ферментації за даним виФаза росту розпочинається інокуляцією 3л находом у кращому варіанті досягає високої об'бродильного чана 1,5мл вихідним гліцериновим ємної продуктивності. Об'ємна продуктивність розчином, що дає початкову OD600 0,05. Фазу виражається в кількості продукту, виробленого на росту проводять в аеробних умовах при безпереодиницю об'єму ферментаційного середовища за рвному барботуванні повітрям з витратою одиницю часу, звичайно, в грамах продукту на 1,5л/хв. (0,5об./об./хв.) при безперервному перелітр середовища за годину. Бажаною є об'ємна мішуванні зі швидкістю 800об./хв. Ріст триває продуктивність, принаймні, 1,5г/л/год., краще доти, поки DO не знижується до 5% від рівня напринаймні, 2,0г/л/год. і ще краще - принаймні, сиченості повітрям. Це збігається з постійною 2,5г/л/год. В умовах кращої густини клітин до 3-6 концентрацією CO2 у газі на виході. Величину клітин на літр ферментаційного середовища макOUR виміряють під час моніторингу кількості ввесимальна продуктивність становить, приблизно, деного повітря й аналізу газів на виході на кисень 5,0г/л/год., а в більш типових випадках - приблизметодом мас-спектрометрії. Під час фази росту но, 4,0г/л/год. Ферментацію дуже бажано прововеличина OUR складає 20,8±2,5ммоль О2/г дити таким чином, щоб зазначені тут величини с.м./год. У цих умовах OUR обмежується здатнісоб'ємної продуктивності досягалися в умовах тю клітини метаболічно перетворювати доступзазначених вище величин рН і/або температури ний кисень. Кінцева густина клітин становить, середовища. приблизно, 4г/л. Молочна кислота, що виробляється згідно з Коли DO досягає нуля, умови аерації підтриданим винаходом, може використовуватися для муються протягом 1 год. до початку виробничої одержання лактиду, циклічного ангідриду двох фази. Виробничу фазу розпочинають швидким молекул молочної кислоти. Залежно від стереопереключенням витрати потоку повітря на ізомеру молочної кислоти таким лактидом може 0,1л/хв. (0,033об./об./хв.) і зменшення швидкості бути D-лактид (утворений з двох молекул Dперемішування до 500об./хв. Ця зміна умов аемолочної кислоти), L-лактид (утворений з двох рації приводить до встановлення величини OUR молекул L-молочної кислоти) або D-L-лактид на рівень 1,5±0,1ммоль О2/г с.м./год. і DO на ну(утворений із однієї L-молекули і однієї Dльовий рівень протягом виробничої фази. Фермолекули молочної кислоти). Підходящим спосоментація триває приблизно 60 годин. При цьому бом одержання лактиду із молочної кислоти є періодично, шляхом відбирання зразків для анаметод полімеризації-деполімеризації, описаний у лізу методами HPLC/IC/GC-MS, проводять виміпатенті США 5,142,023 Грабера зі співроб. рювання швидкості споживання глюкози, вироб(Gruber et al.). лення лактату і виходу лактату. Отримані при Лактид, у свою чергу, може частково викориздійсненні такого процесу результати виробничої стовуватися як мономер для одержання полілакфази наведені в Табл. 1. тидних полімерів (PLA) і співполімерів. Процес виготовлення таких полімерів також описаний у патенті США 5,142,023 Грабера зі співроб. Таблиця 1 (Gruber et al.). Кращими PLA-продуктами є стійкі щодо плавлення полімери, описані в патенті Максимальний титр мо106±3,1г/кг США 5,338,822 (Gruber et al.). PLA-матеріали молочної кислоти жуть бути напівкристалічними або аморфними. Швидкість споживання 1,2±0,05г/г. с.м./год. Наведені нижче приклади служать для ілюстглюкози 15 Швидкість вироблення молочної кислоти Вихід молочної кислоти (виробнича фаза) % відновленого вуглецю (виробнича фаза) Оптична чистота молочної кислоти 88437 1,1±0,04г/г. с.м./год. 16 зультати, отримані у виробничу фазу з величиною OUR = 1,7. 0,92±0,03г молочної кислоти/г глюкози 99%±3,0% >99,9 Описаний експеримент був проведений декілька разів у різних умовах аерації у виробничу фазу, що давали величини OUR, приблизно, 1,2, 2,2, 2,8, 3,0 і 3,2. Отримані результати подані у графічному вигляді на Фіг.1. Як можна бачити на Фіг.1, вихід продукту (крива 1) монотонно знижується з ростом OUR від 1,2 до 3,0 і наприкінці цього діапазону різко зростає (можливо аномально) і далі, при OUR 3,2 знову різко падає. Це зниження виходу продукту добре узгоджується зі зростанням дихання клітин при збільшенні доступного кисню. Швидкість вироблення лактату (крива 3) невеликою мірою зростає в діапазоні величин OUR від 1,2 до 2,2, після чого знижується з подальшим зростанням OUR до 2,8 і далі зростає. Проте, виграш від зростання швидкості вироблення продукту при величинах OUR 3 і більше втрачається внаслідок втрати при цьому виходу продукту. Швидкість використання глюкози (крива 2) деякою мірою зростає при збільшенні від 1,2 до 2,8 і суттєво зростає при OUR більше 3,0 внаслідок швидкого дихання клітин. Дані, наведені на Фіг.2, свідчать про те, що для використовуваного в цих дослідах штаму оптимальна величина OUR лежить у межах, приблизно, від 0,8 до 2,2 і, головним чином, в межах від 1,0 до 1,5. Приклад 2 Експеримент згідно з Прикладом 1 був повторений декілька разів на штамі CD 558. Штамом CD 558 є клітина К. Marxianus з делетованим PDC-геном. Вона містить екзогенний L-LDH-ген L. helveticus, випадковим чином інтегрований у її геном. LDH-ген перебуває під контролем послідовностей PGK-1 промотора S. cerevisiaeі Gal-10 термінатора S. cerevisiae. В усіх дослідах величина OUR у фазу росту становила, приблизно, 20,5ммоль O2/г с.м./год. Умови аерації під час виробничих фаз змінювалися від досліду до досліду шляхом регулювання швидкості барботування і перемішування з метою варіювання величини OUR. Величини OUR у різних дослідах становили 0,6, 1,4, 1,7 і 2,2. У Табл. 2 подані ре Таблиця 2 Максимальний титр молочної кислоти Швидкість споживання глюкози Швидкість вироблення молочної кислоти 111г/кг 0,94г/г. с.м./год. 0,83г/г. с.м./год. 0,89г молочної Вихід молочної кислоти (виробкислоти/г глюконича фаза) зи % відновленого вуглецю (виро95,6 бнича фаза) Оптична чистота молочної кис>99,9 лоти На Фіг.2 можна бачити, як зміна OUR впливає на швидкість вироблення і вихід продукту. Для даного штаму вихід молочної кислоти (крива 1) виказує дуже сильну залежність від величини OUR на ділянці від 0,7 до 2,2, досягаючи максимуму при значеннях OUR навколо 1,4. Швидкість вироблення лактату (крива 1) подібним чином виходить на піковий рівень при зазначеній величині OUR. Швидкість споживання глюкози (крива 2) зростає до OUR приблизно 2,2, після чого виходить на сплощену ділянку. Одержані з даним штамом результати експерименту свідчать про те, що оптимальні значення OUR лежать у діапазоні, приблизно, від 1 до 1,7, а точніше - від 1,2 до 1,5. Приклад 3 Експеримент згідно з Прикладом 1 був проведений три рази. Перший з цих дослідів (3А) був проведений так, як описано в Прикладі 1, за винятком того, що величина OUR під час виробничої фази складала 2,1ммоль О2/г. с.м./год. Під час другого з цих дослідів (3B) культура була переключена на виробничу фазу одразу ж, коли DO у фазу росту досягла нулю. Величина OUR складала 1,8ммоль О2/г. с.м./год. у тих самих умовах аерації, що й у Прикладі 3А. У третьому досліді (3C) аерацію продовжували і по закінченню фази росту протягом ще 1,5 години після досягнення DO нульового рівня, а величина OUR у виробничу фазу становила 1,4ммоль О2/г. с.м./год. у таких самих умовах аерації, що й у Прикладі 3А. Отримані результаті підсумовані в Табл. 3. Таблиця 3 Властивості Час витримування при нульовій DO OUR у виробничу фазу, ммоль О2/г. с.м./год. Максимальний титр молочної кислоти, г/кг Швидкість споживання глюкози, г/г. с.м./год. Швидкість вироблення лактату, г/г. с.м./год. Вихід молочної кислоти, виробнича фаза, г/г % відновленого вуглецю, виробнича фаза Оптична чистота, % 3А 1 год. 2,1 112,3 1,22 1,20 0,89 105 >99,9 Приклад № 3В 3С 0 >1,5 год. 1,8 1,4 84 94 1,06 0,40 0,93 0,32 0,84 0,79 104,3 98 >99,9 >99,9 17 У даних умовах аерації під час виробничої фази величина OUR служила індикатором метаболічної активності мікроорганізму. Зниження OUR у тих випадках, коли час витримування дорівнював нулю або перевищував 1,5 год., (відносно того, що мало місце після 1 год. витримування) вказує на те, що мікроорганізм працював у цих умовах трохи гірше. Ця обставина відбивається також у зниженні споживання глюкози, продукування лактату і виходу продукту. Приклад 4 Інокуляційний штам дріжджової клітини CD 587 культивували в аеробних умовах у 14л лабораторному бродильному чані з буферним (рН 5,5) мінеральним середовищем, доповненим 160г/л глюкози. Аерація здійснювалася барботуванням повітря з витратою 5л/хв. при перемішуванні. Концентрація DO протягом цієї початкової фази росту складала спочатку 100% і знизилася до 20% протягом цього циклу. Величина OUR підтримувалася на рівні, приблизно, 20ммоль О2/г. с.м./год. Коли величина OD600 досягла 10,4 літри ферментаційного бульйону перенесли у 240л промисловий бродильний чан, що містив ще 220л культурально 88437 18 го середовища, доповненого 170г/л глюкози. У цьому чані клітини вирощувалися далі в аеробних умовах при температурі 42°С і рН 5,5 протягом, приблизно, 8 год. На протязі цього часу кількість DO знизилася від початкового рівня до нуля. Величина OUR підтримувалася на рівні 20ммоль О2/г. с.м./год. шляхом аерації з витратою повітря 15л/хв. Культуру витримували з нульовою DO протягом 1 години. Після цього вона була переключена на виробничу фазу зниженням аерації, внаслідок чого величина OUR встановилася на рівні 1,51,7ммоль О2/г. с.м./год. Для підтримання рН на рівні 5,5±0,1 був доданий додатковий буферний агент Ca(OH)2. Під час виробничої фази кількість розчиненого кисню (DO) залишалася на рівні 0%. Споживання глюкози тривало 30 годин, що дало титр лактату 114г/кг. Середня питома швидкість споживання глюкози протягом виробничої фази становила 1,1г/г. с.м./год. Середня питома швидкість вироблення молочної кислоти складала 0,8г/г. с.м./год. при виході продукту 0,76г молочної кислоти на грам глюкози і загальному виході (включаючи фазу росту) 0,67г молочної кислоти на грам глюкози. 19 Комп’ютерна верстка А. Крулевський 88437 Підписне 20 Тираж 28 прим. Міністерство освіти і науки України Державний департамент інтелектуальної власності, вул. Урицького, 45, м. Київ, МСП, 03680, Україна ДП “Український інститут промислової власності”, вул. Глазунова, 1, м. Київ – 42, 01601
ДивитисяДодаткова інформація
Назва патенту англійськоюProcess for fermentaion with participation of genetically modified yeast and the method for realization of fermentation
Автори англійськоюvan HOEK PIM, ARISTIDOU ARISTOS, RUSH BRIAN
Назва патенту російськоюПроцесс ферментации при участии генетически модифицированых дрожжей и способ осуществления ферментации
Автори російськоюван Хоек Пим, Аристидоу Аристос, Ращ Бриан
МПК / Мітки
МПК: C12P 7/40, C12N 1/16, C12P 17/02, C12N 15/81, C12N 1/19
Мітки: модифікованих, генетично, ферментації, участю, дріжджів, здійснення, процес, спосіб
Код посилання
<a href="https://ua.patents.su/10-88437-proces-fermentaci-za-uchastyu-genetichno-modifikovanikh-drizhdzhiv-ta-sposib-zdijjsnennya-fermentaci.html" target="_blank" rel="follow" title="База патентів України">Процес ферментації за участю генетично модифікованих дріжджів та спосіб здійснення ферментації</a>
Спосіб ферментації меляси з одержанням етанолу та хлібопекарських дріжджів

Номер патенту: 21229
Опубліковано: 04.11.1997
Автори: Руда Валентина Володимирівна, Ільїна Лада Дмитрівна, Середа Лілія Оксентіївна, Вовнянко Євгенія Кононівна, Янчевський Віктор Казимирович, Ткаченко Алла Федосіївна, Рудніченко Людмила Вікторівна
МПК: C12N 1/38, C12P 7/08, C12N 1/18
Мітки: дріжджів, одержанням, спосіб, ферментації, хлібопекарських, етанолу, меляси
Формула / Реферат:
Способ ферментации мелассы с получением этанола и хлебопекарных дрожжей, предусматривающий антисептирование мелассного сусла, подкисление, разбавление водой, обогащение питательными солями, аэробный и анаэробный циклы ферментации с последующим выделением хлебопекарных дрожжей и этанола, отличающийся тем, что в мелассное сусло на стадии его подготовки вводят щелочной или термокислотный экстракт - отход дрожжевой биомассы производства...
Живильне середовище для глибинної ферментації штаму aspergillus terreus г-232 – продуцента ітаконової кислоти

Номер патенту: 32328
Опубліковано: 15.12.2000
Автор: Мигаль Сергій Іванович
МПК: C12P 7/40, C12N 1/14, C12P 1/02, C12R 1/66
Мітки: кислоти, ферментації, г-232, штаму, ітаконової, середовище, aspergillus, глибинної, продуцента, живильне, terreus
Текст:
...(по прототипу) до 6,0-6,3. Підвищення рН середовища при зменшенні кількості компонентів середовища і оптимальному їх співвідношенні дозволяє збільшити вихід ітаконової кислоти при глибинній ферментації штаму Aspergillus terreus Г-232. Живильне середовище готують наступним чином. В ємкість заливають 35% від загальної кількості води, додають харчовий цукор, механічно перемішують і підігрівають до повного його розчинення. В другу ємкість...
Спосіб переробки та ферментації тютюнової сировини

Номер патенту: 46119
Опубліковано: 15.05.2002
Автори: Матковський Анатолій Борисович, Федорчук Ємілія Борисівна, Немчина Неля Юхимівна, Гончарук Володимир Якович
МПК: A24B 3/00
Мітки: ферментації, переробки, спосіб, сировини, тютюнової
Формула / Реферат:
Спосіб переробки та ферментації тютюнової сировини, який містить сортування тютюнової сировини по сортотипам та вологості на пунктах первинної переробки тютюнової сировини, транспортування на пункт ферментації тютюнової сировини, сортування тютюнової сировини на три типи вологості: нормально-вологе, 16-19% вологості, вологе, більше 19%, и сухе, нижче 16%, кондиціювання сировини до нормальної вологості, ферментацію тютюнової сировини до...
Спосіб регульованої ферментації для виготовлення мевіноліну та спосіб одержання мевіноліну

Номер патенту: 73074
Опубліковано: 15.06.2005
Автори: Ола Антал, Серес Петер, Шеке Ласло, Балог Габор
МПК: C12P 17/02, C12P 7/62, C12P 7/40
Мітки: ферментації, спосіб, регульованої, виготовлення, одержання, мевіноліну
Формула / Реферат:
1. Спосіб ферментації для виготовлення мевіноліну з використанням штаму, що належить до роду Aspergillus, у глибинній культурі при pH в межах від 5,2 до 7,0, при температурі в межах від 24оС до 30оС, на живильному середовищі, яке містить засвоювані джерела вуглецю та азоту і мінеральні солі, який включає стадію посівної культури та основну стадію ферментації і в якому на основній стадії ферментації з метою підтримання культури у...
Штам дріжджів saccharomyces cerevisiae imb-у-5016 для одержання пресованих хлібопекарських дріжджів в двопродуктовому виробництві спирту і хлібопекарських дріжджів

Номер патенту: 47821
Опубліковано: 15.01.2004
Автори: Ткаченко Алла Феодосіївна, Коваль Катерина Олександрівна, Левандовський Леонід Вікторович, Хоменко Анатолій Іванович, Рудніченко Людмила Вікторівна, Бейко Наталія Євгенівна
МПК: C12R 1/865, C12N 1/18
Мітки: штам, спирту, saccharomyces, cerevisiae, хлібопекарських, дріжджів, двопродуктовому, виробництві, imb-у-5016, пресованих, одержання
Формула / Реферат:
Штам дріжджів Saccharomyces cerevisiae ІМВ-У-5016 для одержання хлібопекарських дріжджів в двопродуктовому виробництві спирту і хлібопекарських дріжджів.
Попередній патент: Літак
Наступний патент: Пристрій для керування газами,зокрема для випробувального стенда газотурбінного двигуна
Випадковий патент: Установка для буріння свердловин